
Dawn chorus interpretation differs when using songs or ... · using songs or calls: the Dupont’s...
17
Submitted 13 March 2018 Accepted 21 June 2018 Published 19 July 2018 Corresponding author Cristian Pérez-Granados, [email protected] Academic editor Jose Maria Cardoso da Silva Additional Information and Declarations can be found on page 12 DOI 10.7717/peerj.5241 Copyright 2018 Pérez-Granados et al. Distributed under Creative Commons CC-BY 4.0 OPEN ACCESS Dawn chorus interpretation differs when using songs or calls: the Dupont’s Lark Chersophilus duponti case Cristian Pérez-Granados 1 ,3 , Tomasz S. Osiejuk 2 and Germán M. López-Iborra 1 1 Ecology Department/Multidisciplinary Institute for Environmental Studies ‘‘Ramón Margalef’’, Universidad de Alicante, Alicante, Spain 2 Department of Behavioural Ecology, Institute of Environmental Biology, Faculty of Biology, Adam Mickiewicz University of Poznan, Poznan, Poland 3 Current affiiation: Terrestrial Ecology Group (TEG-UAM), Department of Ecology, Universidad Autónoma de Madrid, Madrid, Spain ABSTRACT Background. Territorial songbirds vocalise intensively before sunrise and then decrease their vocal activity. This creates a communication network that disseminates essential information for both males and females. The function of dawn chorus in birds has been frequently interpreted according to seasonal variation of singing as the breeding season advances, but potential differences in seasonal variation of song and calls for the same species have not been taken into account. Methods. We chose Dupont’s Lark as a model species to study whether the seasonal pattern of dawn chorus differs between singing and calling activity, because in this species most daily songs and calls are uttered at dawn. We registered vocal activity of Dupont’s Lark before and around dawn in three different populations, through repeated sampling over the entire breeding season of two consecutive years. Results. We found that dawn singing parameters remained constant or presented an increasing trend while dawn calling activity decreased as breeding season advanced. We also found different daily patterns for singing and calling, with birds calling mostly during the first 30 minutes of dawn choruses and singing peaking afterwards. Discussion. The different time patterns of songs and calls may indicate that they serve diverse functions at dawn in the Dupont’s Lark. Relaxation of dawn calling activity after the first month of the breeding season would suggest that dawn calling may be mainly related to mate attraction, while constant dawn singing throughout the breeding period would support a relationship of dawn singing to territorial defence. Our study highlights that the type of vocalisation used is an important factor to consider in further research on dawn choruses, since results may differ depending on whether calls or songs are analysed. Subjects Animal Behavior, Ecology Keywords Acoustic, Call, Mate attraction, Passerine, Song, Territorial defence, Vocalization INTRODUCTION Dawn chorus occurs when birds, usually males, begin to sing before sunrise and then cease or decrease song activity for the rest of the day (Mace, 1987; Catchpole & Slater, 2008, but see Garamszegi et al., 2006 and Webb et al., 2016 for female singing). It is a phenomenon How to cite this article Pérez-Granados et al. (2018), Dawn chorus interpretation differs when using songs or calls: the Dupont’s Lark Chersophilus duponti case. PeerJ 6:e5241; DOI 10.7717/peerj.5241
Transcript of Dawn chorus interpretation differs when using songs or ... · using songs or calls: the Dupont’s...

Submitted 13 March 2018Accepted 21 June 2018Published 19 July 2018
Corresponding authorCristian Peacuterez-Granadoscristianperezuaes
Academic editorJose Maria Cardoso da Silva
Additional Information andDeclarations can be found onpage 12
DOI 107717peerj5241
Copyright2018 Peacuterez-Granados et al
Distributed underCreative Commons CC-BY 40
OPEN ACCESS
Dawn chorus interpretation differs whenusing songs or calls the Dupontrsquos LarkChersophilus duponti caseCristian Peacuterez-Granados13 Tomasz S Osiejuk2 and Germaacuten M Loacutepez-Iborra1
1 Ecology DepartmentMultidisciplinary Institute for Environmental Studies lsquolsquoRamoacuten MargalefrsquorsquoUniversidad de Alicante Alicante Spain
2Department of Behavioural Ecology Institute of Environmental Biology Faculty of BiologyAdamMickiewicz University of Poznan Poznan Poland
3Current affiiation Terrestrial Ecology Group (TEG-UAM) Department of Ecology UniversidadAutoacutenoma de Madrid Madrid Spain
ABSTRACTBackground Territorial songbirds vocalise intensively before sunrise and thendecreasetheir vocal activity This creates a communication network that disseminates essentialinformation for bothmales and females The function of dawn chorus in birds has beenfrequently interpreted according to seasonal variation of singing as the breeding seasonadvances but potential differences in seasonal variation of song and calls for the samespecies have not been taken into accountMethods We chose Dupontrsquos Lark as a model species to study whether the seasonalpattern of dawn chorus differs between singing and calling activity because in thisspecies most daily songs and calls are uttered at dawn We registered vocal activity ofDupontrsquos Lark before and arounddawn in three different populations through repeatedsampling over the entire breeding season of two consecutive yearsResults We found that dawn singing parameters remained constant or presented anincreasing trend while dawn calling activity decreased as breeding season advancedWe also found different daily patterns for singing and calling with birds calling mostlyduring the first 30 minutes of dawn choruses and singing peaking afterwardsDiscussion The different time patterns of songs and calls may indicate that they servediverse functions at dawn in the Dupontrsquos Lark Relaxation of dawn calling activityafter the first month of the breeding season would suggest that dawn calling may bemainly related tomate attraction while constant dawn singing throughout the breedingperiod would support a relationship of dawn singing to territorial defence Our studyhighlights that the type of vocalisation used is an important factor to consider in furtherresearch on dawn choruses since results may differ depending onwhether calls or songsare analysed
Subjects Animal Behavior EcologyKeywords Acoustic Call Mate attraction Passerine Song Territorial defence Vocalization
INTRODUCTIONDawn chorus occurs when birds usually males begin to sing before sunrise and then ceaseor decrease song activity for the rest of the day (Mace 1987 Catchpole amp Slater 2008 butsee Garamszegi et al 2006 and Webb et al 2016 for female singing) It is a phenomenon
How to cite this article Peacuterez-Granados et al (2018) Dawn chorus interpretation differs when using songs or calls the Dupontrsquos LarkChersophilus duponti case PeerJ 6e5241 DOI 107717peerj5241
found in many bird species and has been studied since the late 19th and early 20th centuries(egWright 1912 Allen 1913) Previous works have suggested that the dawn chorus maybe regarded as a reliable signal of male quality (eg Grava Grava amp Otter 2009 Mariniet al 2017) To understand dawn chorus in territorial birds studies should account for(1) the reason of the concentration of vocal activity at dawn which has been explainedby female behaviour conditions for sound transmission and low foraging success atdawn among others (Kacelnik amp Krebs 1983 Kunc Amrhein amp Naguib 2005 Naguib etal 2016) and (2) the function of the vocalisations uttered at dawn which has been relatedto mate attraction (Eriksson amp Wallin 1986 Poesel et al 2006 Murphy et al 2008) andto territorial defence and mate guarding (Moslashller 1988 Moslashller 1991 Amrhein amp Erne2006) although the two functions can be complementary (Moslashller 1991 Slagsvold Dale ampSaeligtre 1994)
Despite the prevalence of dawn-singing in birds and the numerous studies focusedon that topic its origin evolution and causes remain unclear and seem to be context-dependent differing among species populations and studies (eg Staicer SpectorampHorn 1996 Catchpole amp Slater 2008 Zhang Celis-Murillo amp Ward 2016) This couldbe explained in part because researchers have indistinctly analysed or have even pooledcalls and songs (Poesel Foerster amp Kempenaers 2001 Grava et al 2013 Stanley et al 2016Lee MacGregor-Fors amp Yeh 2017) to elucidate dawn chorus function (but see LaZerte Otteramp Slabbekoorn 2017) Nonetheless the bulk of dawn chorus studies have been focusedon singing activity (eg Gil Graves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu2004) Although the distinction between calls and songs is not always clear-cut in practicea given vocalisation can usually be readily classified as either a song or a call and theseclassifications are commonly used in the birdsong literature (Zann 1990 Wang et al2012) In oscines hummingbirds and parrots songs are defined as long loud complexand learned spontaneous vocalisations whose function at dawn has usually been relatedto mate attraction but also to compete with members of the same sex (Slagsvold Dale ampSaeligtre 1994 Catchpole amp Slater 2008) Calls are usually described as shorter simpler lessspontaneous and genetically determined vocalisations uttered throughout the year and thatcan be produced for a variety of reasons includingmaintaining group contact and signallingabout food or danger such as in cases of fight threat or alarm (Marler 2004 Catchpole ampSlater 2008) The use of calls in dawn chorus has been little studied (but see LaZerte Otteramp Slabbekoorn 2017) and is not well understood In some species it has been suggested thatcalls are targeted to the current female (McCallum Grundel amp Dahlsten 1999 Stowell Gillamp Clayton 2016) Thus as the origin and evolution of the two types of vocalisations differthe interpretation of dawn chorus function could vary according to the type considered
In this study we selected the Dupontrsquos Lark (Chersophilus duponti) as a model to explorepotential differences in the interpretation of dawn chorus when analysing songs or callsWe chose this species because it is a diurnal passerine with high vocal activity concentratedat dawn and because it utters a large number of both types of vocalisations during dawnchoruses There are several studies published about the song and the territorial call of thisspecies which provide good background for deeper analysis (Laiolo amp Tella 2005 Laiolo ampTella 2006 Laiolo 2008 Laiolo et al 2008 Peacuterez-Granados Osiejuk amp Loacutepez-Iborra 2016)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 217
All of these studies have assumed that only males sing and produce territorial calls sincefemales do not appear to utter long range vocalisations (Laiolo et al 2007) OccasionallyDupontrsquos Lark males also produce warning calls (to repel intruders from their territoriesLaiolo et al 2005) and both sexes may utter distress calls an ultimate alarm signal givenby individuals that are in predator risk (Laiolo et al 2005 C Peacuterez-Granados pers obs2018) Although some female larks are known to sing such as the Eurasian Lark (Alaudaarvensis) or the Woodlark (Lullula arborea) (Garamszegi et al 2006 Odom amp Benedict2018) there is no evidence of Dupontrsquos Lark females singing or uttering territorial orwarning calls (Laiolo amp Tella 2005 Garamszegi et al 2006 Laiolo 2008 Laiolo et al 2005Laiolo et al 2007 Laiolo et al 2008 Barrero et al 2017 C Peacuterez-Granados pers obs2011ndash2012) despite the large number of bioacoustic studies focussed on the species andthe fact that hundreds of Dupontrsquos Larks have been ringed and later resighted singingTherefore in this study we considered that all monitored birds uttering songs or territorialcalls were males We are aware that females are able to utter distress calls in the nest andduring handling (C Peacuterez-Granados pers obs 2018) but this type of vocalisation as wellas warning calls were not included in our study and we will refer to territorial calls simplyas calls hereinafter
In this paper we aimed to investigate if singing and calling activity at dawn in theDupontrsquos Lark have different seasonal patterns and to explore the consequences of usingsongs or calls when interpreting dawn chorus function For this purpose we monitored thedawn singing and calling activity of the Dupontrsquos Lark during the breeding season in twoconsecutive years We hypothesised that if songs and calls have different functions theirseasonal pattern throughout the breeding period would differ (Celis-Murillo et al 2016)Vocalisation functions cannot be demonstrated without experimental or more detailedobservational analyses However we assumed that the relationship between function andseasonal patterns of vocalisations in the Dupontrsquos Lark is similar to that described for otherEuropean passerines Therefore we expected that Dupontrsquos Lark singing or calling activityat dawn would decrease after pairing if they are related to attracting females at that timeas described for other passerine species (eg Gil Graves amp Slater 1999 Amrhein Korner ampNaguib 2002 Liu 2004 Celis-Murillo et al 2016) but would remain constant if they arerelated to territorial defence (eg Olinkiewicz amp Osiejuk 2003 Kunc Amrhein amp Naguib2005 Amrhein amp Erne 2006 Liu amp Kroodsma 2007)
MATERIAL AND METHODSStudy areaWe conducted a field study from late March to early June in 2013 and 2014 in the threelargest Dupontrsquos Lark populations in Valencia province (eastern Spain Peacuterez-Granadosamp Loacutepez-Iborra 2013) where patch size and number of territorial males ranged between193ndash200 ha and 10ndash19 males respectively Study sites were located at 1000 m asl andthe vegetation was a shrub-steppe dominated by low shrub species (Thymus spp Genistascorpius and Rosmarinus officinalis) In all sites there were some dispersed trees especiallypines Pinus spp and junipers (Junipers oxycedrus and J communis) More information onthe study area can be found in Peacuterez-Granados Loacutepez-Iborra amp Seoane (2017a)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 317
Figure 1 Location of the sampling stations in the three studied sites and sampling station procedureSong activity was registered individually for each Dupontrsquos Lark male according to their different distanceand location to observer
Full-size DOI 107717peerj5241fig-1
Data collectionWe registered dawn singing and calling activity of Dupontrsquos Lark males in three sitesover two consecutive breeding seasons (years) Acoustic activity was measured throughrepeated sampling over the entire breeding season from one fixed counting station per siteand year Counting station location remained the same within a year but were separatedby at least 500 m among years (Fig 1) in order to monitor two different areas withineach site This is also expected to favour the monitoring of different males since in thestudy area adult Dupontrsquos Larks have been reported to move on average 100 m betweenbreeding seasons (Peacuterez-Granados amp Loacutepez-Iborra 2015) Likewise only about half of theadult males survive between successive years (Laiolo et al 2008) which would furtherdecrease the probability of detecting the same male in the two study years Songs and callswere assigned to a specific male based on differing direction and distance to the observer(Fig 1) We only considered birds located within a 250 m radius from a station locationthat did not change their location during the survey (estimated displacement less than50 m) to facilitate recognition Distance from observer to vocalising bird was determinedupon acoustic clues and based on previous experience counting the species in the studyarea (eg Peacuterez-Granados amp Loacutepez-Iborra 2013 Peacuterez-Granados Loacutepez-Iborra amp Seoane2017a) This wide radius was considered given that the speciesrsquo songs may be heard from
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 417
long distances (Laiolo et al 2007 Voumlgeli et al 2010) We monitored vocal activity of up toa maximum of four males per station and night with a mean (plusmnSD) number of 35 plusmn 05males monitored per stationnight
Surveys were labelled as lsquolsquo1 2 3 4 5 or 6rsquorsquo following the order in which they wereconducted In both years the first survey was carried out between the 26th and 31st ofMarch and successive surveys were carried out fortnightly from then until 14ndash16 of JuneThis period corresponds to most of the breeding period of the species (Herranz et al 1994Peacuterez-Granados et al 2017b) The first two surveys (between late March and early April)correspond to the first stage of the breeding time in the study area when most females arebuilding their nests and laying first clutches The third and fourth surveys (late AprilndashearlyMay) were carried out at the peak of the breeding activity of the species (Herranz et al1994 Peacuterez-Granados et al 2017b) During this time of the season breeding pairs arerearing their first clutch or in some cases second or replacement clutches The fifth andsixth surveys (late Mayndashearly June) were carried out at the end of the breeding season atime when some breeding pairs may have finished their breeding period and a relaxationin territory defence may occur (Peacuterez-Granados et al 2017b)
Dawn acoustic activity was always monitored by the same researcher (CPG) whostood at the station from 100-min before sunrise to ten minutes after dawn The observerstood for 5 min in silence at each station before the study began Daily times of sunriseat the geographic location of the study area were obtained from the Spanish Ministryof Development (httpastronomiaigneswebguesthora-salidas-y-puestas-de-sol) Wedivided the continuous survey into 5-min intervals and registered the number of songs andcalls uttered per male per 5-min period For each interval we calculated songcall outputas the total number of songscalls uttered per male We also monitored songcall start andend time (5-min interval in which the first and the last song or call were produced by anymale each night) and performance time measured as the total number of 5-min intervalsthat each bird was vocally active each night
The song of the Dupontrsquos Lark in the study area includes a mean of five discrete songtypes with no differences in male repertoire size among studied sites (Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra 2016) Song types are largely shared and repeated in the sameorder by neighbouring males and end with a common sequence of the speciesrsquo so-calledlsquowhee-ur-wheeeersquo (Cramp 1988) We considered the whole sequence of repeated songtypes as a unique song to facilitate data collection and used the definite lsquowhee-ur-wheeeersquoto define the boundary between one song and another A sonogram of a typical sequencerecorded in the study area can be found in Fig 2 and a song recorded in the study areamay be found in Peacuterez-Granados (XC216990 on httpwwwxeno-cantoorg) Sonogramsof the commonest song types in the three studied sites can be found in Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra (2016) Calls consist of discrete and short whistles (a completedescription and sonograms can be found in Laiolo et al 2007)
Surveys were conducted only during windless dawns with good weather conditions(cloud cover lt10 and no rain) to avoid potential biases in bird behaviour Dawn singingactivity of the Dupontrsquos Lark varies with moon phase (authorsrsquo own data) Thereforesurveys were carried out on the two days following or preceding full or new moons to
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 517
Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

found in many bird species and has been studied since the late 19th and early 20th centuries(egWright 1912 Allen 1913) Previous works have suggested that the dawn chorus maybe regarded as a reliable signal of male quality (eg Grava Grava amp Otter 2009 Mariniet al 2017) To understand dawn chorus in territorial birds studies should account for(1) the reason of the concentration of vocal activity at dawn which has been explainedby female behaviour conditions for sound transmission and low foraging success atdawn among others (Kacelnik amp Krebs 1983 Kunc Amrhein amp Naguib 2005 Naguib etal 2016) and (2) the function of the vocalisations uttered at dawn which has been relatedto mate attraction (Eriksson amp Wallin 1986 Poesel et al 2006 Murphy et al 2008) andto territorial defence and mate guarding (Moslashller 1988 Moslashller 1991 Amrhein amp Erne2006) although the two functions can be complementary (Moslashller 1991 Slagsvold Dale ampSaeligtre 1994)
Despite the prevalence of dawn-singing in birds and the numerous studies focusedon that topic its origin evolution and causes remain unclear and seem to be context-dependent differing among species populations and studies (eg Staicer SpectorampHorn 1996 Catchpole amp Slater 2008 Zhang Celis-Murillo amp Ward 2016) This couldbe explained in part because researchers have indistinctly analysed or have even pooledcalls and songs (Poesel Foerster amp Kempenaers 2001 Grava et al 2013 Stanley et al 2016Lee MacGregor-Fors amp Yeh 2017) to elucidate dawn chorus function (but see LaZerte Otteramp Slabbekoorn 2017) Nonetheless the bulk of dawn chorus studies have been focusedon singing activity (eg Gil Graves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu2004) Although the distinction between calls and songs is not always clear-cut in practicea given vocalisation can usually be readily classified as either a song or a call and theseclassifications are commonly used in the birdsong literature (Zann 1990 Wang et al2012) In oscines hummingbirds and parrots songs are defined as long loud complexand learned spontaneous vocalisations whose function at dawn has usually been relatedto mate attraction but also to compete with members of the same sex (Slagsvold Dale ampSaeligtre 1994 Catchpole amp Slater 2008) Calls are usually described as shorter simpler lessspontaneous and genetically determined vocalisations uttered throughout the year and thatcan be produced for a variety of reasons includingmaintaining group contact and signallingabout food or danger such as in cases of fight threat or alarm (Marler 2004 Catchpole ampSlater 2008) The use of calls in dawn chorus has been little studied (but see LaZerte Otteramp Slabbekoorn 2017) and is not well understood In some species it has been suggested thatcalls are targeted to the current female (McCallum Grundel amp Dahlsten 1999 Stowell Gillamp Clayton 2016) Thus as the origin and evolution of the two types of vocalisations differthe interpretation of dawn chorus function could vary according to the type considered
In this study we selected the Dupontrsquos Lark (Chersophilus duponti) as a model to explorepotential differences in the interpretation of dawn chorus when analysing songs or callsWe chose this species because it is a diurnal passerine with high vocal activity concentratedat dawn and because it utters a large number of both types of vocalisations during dawnchoruses There are several studies published about the song and the territorial call of thisspecies which provide good background for deeper analysis (Laiolo amp Tella 2005 Laiolo ampTella 2006 Laiolo 2008 Laiolo et al 2008 Peacuterez-Granados Osiejuk amp Loacutepez-Iborra 2016)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 217
All of these studies have assumed that only males sing and produce territorial calls sincefemales do not appear to utter long range vocalisations (Laiolo et al 2007) OccasionallyDupontrsquos Lark males also produce warning calls (to repel intruders from their territoriesLaiolo et al 2005) and both sexes may utter distress calls an ultimate alarm signal givenby individuals that are in predator risk (Laiolo et al 2005 C Peacuterez-Granados pers obs2018) Although some female larks are known to sing such as the Eurasian Lark (Alaudaarvensis) or the Woodlark (Lullula arborea) (Garamszegi et al 2006 Odom amp Benedict2018) there is no evidence of Dupontrsquos Lark females singing or uttering territorial orwarning calls (Laiolo amp Tella 2005 Garamszegi et al 2006 Laiolo 2008 Laiolo et al 2005Laiolo et al 2007 Laiolo et al 2008 Barrero et al 2017 C Peacuterez-Granados pers obs2011ndash2012) despite the large number of bioacoustic studies focussed on the species andthe fact that hundreds of Dupontrsquos Larks have been ringed and later resighted singingTherefore in this study we considered that all monitored birds uttering songs or territorialcalls were males We are aware that females are able to utter distress calls in the nest andduring handling (C Peacuterez-Granados pers obs 2018) but this type of vocalisation as wellas warning calls were not included in our study and we will refer to territorial calls simplyas calls hereinafter
In this paper we aimed to investigate if singing and calling activity at dawn in theDupontrsquos Lark have different seasonal patterns and to explore the consequences of usingsongs or calls when interpreting dawn chorus function For this purpose we monitored thedawn singing and calling activity of the Dupontrsquos Lark during the breeding season in twoconsecutive years We hypothesised that if songs and calls have different functions theirseasonal pattern throughout the breeding period would differ (Celis-Murillo et al 2016)Vocalisation functions cannot be demonstrated without experimental or more detailedobservational analyses However we assumed that the relationship between function andseasonal patterns of vocalisations in the Dupontrsquos Lark is similar to that described for otherEuropean passerines Therefore we expected that Dupontrsquos Lark singing or calling activityat dawn would decrease after pairing if they are related to attracting females at that timeas described for other passerine species (eg Gil Graves amp Slater 1999 Amrhein Korner ampNaguib 2002 Liu 2004 Celis-Murillo et al 2016) but would remain constant if they arerelated to territorial defence (eg Olinkiewicz amp Osiejuk 2003 Kunc Amrhein amp Naguib2005 Amrhein amp Erne 2006 Liu amp Kroodsma 2007)
MATERIAL AND METHODSStudy areaWe conducted a field study from late March to early June in 2013 and 2014 in the threelargest Dupontrsquos Lark populations in Valencia province (eastern Spain Peacuterez-Granadosamp Loacutepez-Iborra 2013) where patch size and number of territorial males ranged between193ndash200 ha and 10ndash19 males respectively Study sites were located at 1000 m asl andthe vegetation was a shrub-steppe dominated by low shrub species (Thymus spp Genistascorpius and Rosmarinus officinalis) In all sites there were some dispersed trees especiallypines Pinus spp and junipers (Junipers oxycedrus and J communis) More information onthe study area can be found in Peacuterez-Granados Loacutepez-Iborra amp Seoane (2017a)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 317
Figure 1 Location of the sampling stations in the three studied sites and sampling station procedureSong activity was registered individually for each Dupontrsquos Lark male according to their different distanceand location to observer
Full-size DOI 107717peerj5241fig-1
Data collectionWe registered dawn singing and calling activity of Dupontrsquos Lark males in three sitesover two consecutive breeding seasons (years) Acoustic activity was measured throughrepeated sampling over the entire breeding season from one fixed counting station per siteand year Counting station location remained the same within a year but were separatedby at least 500 m among years (Fig 1) in order to monitor two different areas withineach site This is also expected to favour the monitoring of different males since in thestudy area adult Dupontrsquos Larks have been reported to move on average 100 m betweenbreeding seasons (Peacuterez-Granados amp Loacutepez-Iborra 2015) Likewise only about half of theadult males survive between successive years (Laiolo et al 2008) which would furtherdecrease the probability of detecting the same male in the two study years Songs and callswere assigned to a specific male based on differing direction and distance to the observer(Fig 1) We only considered birds located within a 250 m radius from a station locationthat did not change their location during the survey (estimated displacement less than50 m) to facilitate recognition Distance from observer to vocalising bird was determinedupon acoustic clues and based on previous experience counting the species in the studyarea (eg Peacuterez-Granados amp Loacutepez-Iborra 2013 Peacuterez-Granados Loacutepez-Iborra amp Seoane2017a) This wide radius was considered given that the speciesrsquo songs may be heard from
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 417
long distances (Laiolo et al 2007 Voumlgeli et al 2010) We monitored vocal activity of up toa maximum of four males per station and night with a mean (plusmnSD) number of 35 plusmn 05males monitored per stationnight
Surveys were labelled as lsquolsquo1 2 3 4 5 or 6rsquorsquo following the order in which they wereconducted In both years the first survey was carried out between the 26th and 31st ofMarch and successive surveys were carried out fortnightly from then until 14ndash16 of JuneThis period corresponds to most of the breeding period of the species (Herranz et al 1994Peacuterez-Granados et al 2017b) The first two surveys (between late March and early April)correspond to the first stage of the breeding time in the study area when most females arebuilding their nests and laying first clutches The third and fourth surveys (late AprilndashearlyMay) were carried out at the peak of the breeding activity of the species (Herranz et al1994 Peacuterez-Granados et al 2017b) During this time of the season breeding pairs arerearing their first clutch or in some cases second or replacement clutches The fifth andsixth surveys (late Mayndashearly June) were carried out at the end of the breeding season atime when some breeding pairs may have finished their breeding period and a relaxationin territory defence may occur (Peacuterez-Granados et al 2017b)
Dawn acoustic activity was always monitored by the same researcher (CPG) whostood at the station from 100-min before sunrise to ten minutes after dawn The observerstood for 5 min in silence at each station before the study began Daily times of sunriseat the geographic location of the study area were obtained from the Spanish Ministryof Development (httpastronomiaigneswebguesthora-salidas-y-puestas-de-sol) Wedivided the continuous survey into 5-min intervals and registered the number of songs andcalls uttered per male per 5-min period For each interval we calculated songcall outputas the total number of songscalls uttered per male We also monitored songcall start andend time (5-min interval in which the first and the last song or call were produced by anymale each night) and performance time measured as the total number of 5-min intervalsthat each bird was vocally active each night
The song of the Dupontrsquos Lark in the study area includes a mean of five discrete songtypes with no differences in male repertoire size among studied sites (Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra 2016) Song types are largely shared and repeated in the sameorder by neighbouring males and end with a common sequence of the speciesrsquo so-calledlsquowhee-ur-wheeeersquo (Cramp 1988) We considered the whole sequence of repeated songtypes as a unique song to facilitate data collection and used the definite lsquowhee-ur-wheeeersquoto define the boundary between one song and another A sonogram of a typical sequencerecorded in the study area can be found in Fig 2 and a song recorded in the study areamay be found in Peacuterez-Granados (XC216990 on httpwwwxeno-cantoorg) Sonogramsof the commonest song types in the three studied sites can be found in Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra (2016) Calls consist of discrete and short whistles (a completedescription and sonograms can be found in Laiolo et al 2007)
Surveys were conducted only during windless dawns with good weather conditions(cloud cover lt10 and no rain) to avoid potential biases in bird behaviour Dawn singingactivity of the Dupontrsquos Lark varies with moon phase (authorsrsquo own data) Thereforesurveys were carried out on the two days following or preceding full or new moons to
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 517
Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

All of these studies have assumed that only males sing and produce territorial calls sincefemales do not appear to utter long range vocalisations (Laiolo et al 2007) OccasionallyDupontrsquos Lark males also produce warning calls (to repel intruders from their territoriesLaiolo et al 2005) and both sexes may utter distress calls an ultimate alarm signal givenby individuals that are in predator risk (Laiolo et al 2005 C Peacuterez-Granados pers obs2018) Although some female larks are known to sing such as the Eurasian Lark (Alaudaarvensis) or the Woodlark (Lullula arborea) (Garamszegi et al 2006 Odom amp Benedict2018) there is no evidence of Dupontrsquos Lark females singing or uttering territorial orwarning calls (Laiolo amp Tella 2005 Garamszegi et al 2006 Laiolo 2008 Laiolo et al 2005Laiolo et al 2007 Laiolo et al 2008 Barrero et al 2017 C Peacuterez-Granados pers obs2011ndash2012) despite the large number of bioacoustic studies focussed on the species andthe fact that hundreds of Dupontrsquos Larks have been ringed and later resighted singingTherefore in this study we considered that all monitored birds uttering songs or territorialcalls were males We are aware that females are able to utter distress calls in the nest andduring handling (C Peacuterez-Granados pers obs 2018) but this type of vocalisation as wellas warning calls were not included in our study and we will refer to territorial calls simplyas calls hereinafter
In this paper we aimed to investigate if singing and calling activity at dawn in theDupontrsquos Lark have different seasonal patterns and to explore the consequences of usingsongs or calls when interpreting dawn chorus function For this purpose we monitored thedawn singing and calling activity of the Dupontrsquos Lark during the breeding season in twoconsecutive years We hypothesised that if songs and calls have different functions theirseasonal pattern throughout the breeding period would differ (Celis-Murillo et al 2016)Vocalisation functions cannot be demonstrated without experimental or more detailedobservational analyses However we assumed that the relationship between function andseasonal patterns of vocalisations in the Dupontrsquos Lark is similar to that described for otherEuropean passerines Therefore we expected that Dupontrsquos Lark singing or calling activityat dawn would decrease after pairing if they are related to attracting females at that timeas described for other passerine species (eg Gil Graves amp Slater 1999 Amrhein Korner ampNaguib 2002 Liu 2004 Celis-Murillo et al 2016) but would remain constant if they arerelated to territorial defence (eg Olinkiewicz amp Osiejuk 2003 Kunc Amrhein amp Naguib2005 Amrhein amp Erne 2006 Liu amp Kroodsma 2007)
MATERIAL AND METHODSStudy areaWe conducted a field study from late March to early June in 2013 and 2014 in the threelargest Dupontrsquos Lark populations in Valencia province (eastern Spain Peacuterez-Granadosamp Loacutepez-Iborra 2013) where patch size and number of territorial males ranged between193ndash200 ha and 10ndash19 males respectively Study sites were located at 1000 m asl andthe vegetation was a shrub-steppe dominated by low shrub species (Thymus spp Genistascorpius and Rosmarinus officinalis) In all sites there were some dispersed trees especiallypines Pinus spp and junipers (Junipers oxycedrus and J communis) More information onthe study area can be found in Peacuterez-Granados Loacutepez-Iborra amp Seoane (2017a)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 317
Figure 1 Location of the sampling stations in the three studied sites and sampling station procedureSong activity was registered individually for each Dupontrsquos Lark male according to their different distanceand location to observer
Full-size DOI 107717peerj5241fig-1
Data collectionWe registered dawn singing and calling activity of Dupontrsquos Lark males in three sitesover two consecutive breeding seasons (years) Acoustic activity was measured throughrepeated sampling over the entire breeding season from one fixed counting station per siteand year Counting station location remained the same within a year but were separatedby at least 500 m among years (Fig 1) in order to monitor two different areas withineach site This is also expected to favour the monitoring of different males since in thestudy area adult Dupontrsquos Larks have been reported to move on average 100 m betweenbreeding seasons (Peacuterez-Granados amp Loacutepez-Iborra 2015) Likewise only about half of theadult males survive between successive years (Laiolo et al 2008) which would furtherdecrease the probability of detecting the same male in the two study years Songs and callswere assigned to a specific male based on differing direction and distance to the observer(Fig 1) We only considered birds located within a 250 m radius from a station locationthat did not change their location during the survey (estimated displacement less than50 m) to facilitate recognition Distance from observer to vocalising bird was determinedupon acoustic clues and based on previous experience counting the species in the studyarea (eg Peacuterez-Granados amp Loacutepez-Iborra 2013 Peacuterez-Granados Loacutepez-Iborra amp Seoane2017a) This wide radius was considered given that the speciesrsquo songs may be heard from
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 417
long distances (Laiolo et al 2007 Voumlgeli et al 2010) We monitored vocal activity of up toa maximum of four males per station and night with a mean (plusmnSD) number of 35 plusmn 05males monitored per stationnight
Surveys were labelled as lsquolsquo1 2 3 4 5 or 6rsquorsquo following the order in which they wereconducted In both years the first survey was carried out between the 26th and 31st ofMarch and successive surveys were carried out fortnightly from then until 14ndash16 of JuneThis period corresponds to most of the breeding period of the species (Herranz et al 1994Peacuterez-Granados et al 2017b) The first two surveys (between late March and early April)correspond to the first stage of the breeding time in the study area when most females arebuilding their nests and laying first clutches The third and fourth surveys (late AprilndashearlyMay) were carried out at the peak of the breeding activity of the species (Herranz et al1994 Peacuterez-Granados et al 2017b) During this time of the season breeding pairs arerearing their first clutch or in some cases second or replacement clutches The fifth andsixth surveys (late Mayndashearly June) were carried out at the end of the breeding season atime when some breeding pairs may have finished their breeding period and a relaxationin territory defence may occur (Peacuterez-Granados et al 2017b)
Dawn acoustic activity was always monitored by the same researcher (CPG) whostood at the station from 100-min before sunrise to ten minutes after dawn The observerstood for 5 min in silence at each station before the study began Daily times of sunriseat the geographic location of the study area were obtained from the Spanish Ministryof Development (httpastronomiaigneswebguesthora-salidas-y-puestas-de-sol) Wedivided the continuous survey into 5-min intervals and registered the number of songs andcalls uttered per male per 5-min period For each interval we calculated songcall outputas the total number of songscalls uttered per male We also monitored songcall start andend time (5-min interval in which the first and the last song or call were produced by anymale each night) and performance time measured as the total number of 5-min intervalsthat each bird was vocally active each night
The song of the Dupontrsquos Lark in the study area includes a mean of five discrete songtypes with no differences in male repertoire size among studied sites (Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra 2016) Song types are largely shared and repeated in the sameorder by neighbouring males and end with a common sequence of the speciesrsquo so-calledlsquowhee-ur-wheeeersquo (Cramp 1988) We considered the whole sequence of repeated songtypes as a unique song to facilitate data collection and used the definite lsquowhee-ur-wheeeersquoto define the boundary between one song and another A sonogram of a typical sequencerecorded in the study area can be found in Fig 2 and a song recorded in the study areamay be found in Peacuterez-Granados (XC216990 on httpwwwxeno-cantoorg) Sonogramsof the commonest song types in the three studied sites can be found in Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra (2016) Calls consist of discrete and short whistles (a completedescription and sonograms can be found in Laiolo et al 2007)
Surveys were conducted only during windless dawns with good weather conditions(cloud cover lt10 and no rain) to avoid potential biases in bird behaviour Dawn singingactivity of the Dupontrsquos Lark varies with moon phase (authorsrsquo own data) Thereforesurveys were carried out on the two days following or preceding full or new moons to
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 517
Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Figure 1 Location of the sampling stations in the three studied sites and sampling station procedureSong activity was registered individually for each Dupontrsquos Lark male according to their different distanceand location to observer
Full-size DOI 107717peerj5241fig-1
Data collectionWe registered dawn singing and calling activity of Dupontrsquos Lark males in three sitesover two consecutive breeding seasons (years) Acoustic activity was measured throughrepeated sampling over the entire breeding season from one fixed counting station per siteand year Counting station location remained the same within a year but were separatedby at least 500 m among years (Fig 1) in order to monitor two different areas withineach site This is also expected to favour the monitoring of different males since in thestudy area adult Dupontrsquos Larks have been reported to move on average 100 m betweenbreeding seasons (Peacuterez-Granados amp Loacutepez-Iborra 2015) Likewise only about half of theadult males survive between successive years (Laiolo et al 2008) which would furtherdecrease the probability of detecting the same male in the two study years Songs and callswere assigned to a specific male based on differing direction and distance to the observer(Fig 1) We only considered birds located within a 250 m radius from a station locationthat did not change their location during the survey (estimated displacement less than50 m) to facilitate recognition Distance from observer to vocalising bird was determinedupon acoustic clues and based on previous experience counting the species in the studyarea (eg Peacuterez-Granados amp Loacutepez-Iborra 2013 Peacuterez-Granados Loacutepez-Iborra amp Seoane2017a) This wide radius was considered given that the speciesrsquo songs may be heard from
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 417
long distances (Laiolo et al 2007 Voumlgeli et al 2010) We monitored vocal activity of up toa maximum of four males per station and night with a mean (plusmnSD) number of 35 plusmn 05males monitored per stationnight
Surveys were labelled as lsquolsquo1 2 3 4 5 or 6rsquorsquo following the order in which they wereconducted In both years the first survey was carried out between the 26th and 31st ofMarch and successive surveys were carried out fortnightly from then until 14ndash16 of JuneThis period corresponds to most of the breeding period of the species (Herranz et al 1994Peacuterez-Granados et al 2017b) The first two surveys (between late March and early April)correspond to the first stage of the breeding time in the study area when most females arebuilding their nests and laying first clutches The third and fourth surveys (late AprilndashearlyMay) were carried out at the peak of the breeding activity of the species (Herranz et al1994 Peacuterez-Granados et al 2017b) During this time of the season breeding pairs arerearing their first clutch or in some cases second or replacement clutches The fifth andsixth surveys (late Mayndashearly June) were carried out at the end of the breeding season atime when some breeding pairs may have finished their breeding period and a relaxationin territory defence may occur (Peacuterez-Granados et al 2017b)
Dawn acoustic activity was always monitored by the same researcher (CPG) whostood at the station from 100-min before sunrise to ten minutes after dawn The observerstood for 5 min in silence at each station before the study began Daily times of sunriseat the geographic location of the study area were obtained from the Spanish Ministryof Development (httpastronomiaigneswebguesthora-salidas-y-puestas-de-sol) Wedivided the continuous survey into 5-min intervals and registered the number of songs andcalls uttered per male per 5-min period For each interval we calculated songcall outputas the total number of songscalls uttered per male We also monitored songcall start andend time (5-min interval in which the first and the last song or call were produced by anymale each night) and performance time measured as the total number of 5-min intervalsthat each bird was vocally active each night
The song of the Dupontrsquos Lark in the study area includes a mean of five discrete songtypes with no differences in male repertoire size among studied sites (Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra 2016) Song types are largely shared and repeated in the sameorder by neighbouring males and end with a common sequence of the speciesrsquo so-calledlsquowhee-ur-wheeeersquo (Cramp 1988) We considered the whole sequence of repeated songtypes as a unique song to facilitate data collection and used the definite lsquowhee-ur-wheeeersquoto define the boundary between one song and another A sonogram of a typical sequencerecorded in the study area can be found in Fig 2 and a song recorded in the study areamay be found in Peacuterez-Granados (XC216990 on httpwwwxeno-cantoorg) Sonogramsof the commonest song types in the three studied sites can be found in Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra (2016) Calls consist of discrete and short whistles (a completedescription and sonograms can be found in Laiolo et al 2007)
Surveys were conducted only during windless dawns with good weather conditions(cloud cover lt10 and no rain) to avoid potential biases in bird behaviour Dawn singingactivity of the Dupontrsquos Lark varies with moon phase (authorsrsquo own data) Thereforesurveys were carried out on the two days following or preceding full or new moons to
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 517
Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

long distances (Laiolo et al 2007 Voumlgeli et al 2010) We monitored vocal activity of up toa maximum of four males per station and night with a mean (plusmnSD) number of 35 plusmn 05males monitored per stationnight
Surveys were labelled as lsquolsquo1 2 3 4 5 or 6rsquorsquo following the order in which they wereconducted In both years the first survey was carried out between the 26th and 31st ofMarch and successive surveys were carried out fortnightly from then until 14ndash16 of JuneThis period corresponds to most of the breeding period of the species (Herranz et al 1994Peacuterez-Granados et al 2017b) The first two surveys (between late March and early April)correspond to the first stage of the breeding time in the study area when most females arebuilding their nests and laying first clutches The third and fourth surveys (late AprilndashearlyMay) were carried out at the peak of the breeding activity of the species (Herranz et al1994 Peacuterez-Granados et al 2017b) During this time of the season breeding pairs arerearing their first clutch or in some cases second or replacement clutches The fifth andsixth surveys (late Mayndashearly June) were carried out at the end of the breeding season atime when some breeding pairs may have finished their breeding period and a relaxationin territory defence may occur (Peacuterez-Granados et al 2017b)
Dawn acoustic activity was always monitored by the same researcher (CPG) whostood at the station from 100-min before sunrise to ten minutes after dawn The observerstood for 5 min in silence at each station before the study began Daily times of sunriseat the geographic location of the study area were obtained from the Spanish Ministryof Development (httpastronomiaigneswebguesthora-salidas-y-puestas-de-sol) Wedivided the continuous survey into 5-min intervals and registered the number of songs andcalls uttered per male per 5-min period For each interval we calculated songcall outputas the total number of songscalls uttered per male We also monitored songcall start andend time (5-min interval in which the first and the last song or call were produced by anymale each night) and performance time measured as the total number of 5-min intervalsthat each bird was vocally active each night
The song of the Dupontrsquos Lark in the study area includes a mean of five discrete songtypes with no differences in male repertoire size among studied sites (Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra 2016) Song types are largely shared and repeated in the sameorder by neighbouring males and end with a common sequence of the speciesrsquo so-calledlsquowhee-ur-wheeeersquo (Cramp 1988) We considered the whole sequence of repeated songtypes as a unique song to facilitate data collection and used the definite lsquowhee-ur-wheeeersquoto define the boundary between one song and another A sonogram of a typical sequencerecorded in the study area can be found in Fig 2 and a song recorded in the study areamay be found in Peacuterez-Granados (XC216990 on httpwwwxeno-cantoorg) Sonogramsof the commonest song types in the three studied sites can be found in Peacuterez-GranadosOsiejuk amp Loacutepez-Iborra (2016) Calls consist of discrete and short whistles (a completedescription and sonograms can be found in Laiolo et al 2007)
Surveys were conducted only during windless dawns with good weather conditions(cloud cover lt10 and no rain) to avoid potential biases in bird behaviour Dawn singingactivity of the Dupontrsquos Lark varies with moon phase (authorsrsquo own data) Thereforesurveys were carried out on the two days following or preceding full or new moons to
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 517
Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Figure 2 Sonogram of a typical Dupontrsquos Lark song in the study areaFull-size DOI 107717peerj5241fig-2
control for the potential effect of moon phase in the analyses Moon phases were obtainedfrom the Spanish Ministry of Development (httpastronomiaigneswebguestagenda-astronomica)
Statistical analysesAll statistical analyses were performed with R v 341 (R Development Core Team 2014)and all results are expressed as mean plusmn SE To analyse the variation of dawn singing orcalling activity across the breeding season we fitted independent Linear Mixed Models forsongs and calls Start time end time performance time or total night output were used asresponse variables Survey as a continuous variable (lsquolsquo123456rsquorsquo) was included in modelsas a fixed effect and site (lsquolsquoHontanarLosarPinarrsquorsquo) moon phase (lsquolsquoFullNewrsquorsquo) and year(lsquolsquo20132014rsquorsquo) as random effects to control for variation owing to site lunar cycle andinter-annual variations Linear Mixed Models were fitted using the lsquolsquolmerrsquorsquo function inthe R package lsquolsquolme4rsquorsquo (Bates et al 2015) The degrees of freedom were calculated usingthe Kenward-Roger method (Kenward amp Roger 1997) We present models with p-valuesignificance for fixed effects calculated from type III F-tests using the lmerTest package(Kuznetsova Brockhoff amp Christensen 2014)
We also calculated for each survey (pooling sites and years) the Pearson correlationbetween the total number of songs and calls uttered per male to ascertain whether therewas a trade-off between the two types of vocalisations To evaluate similarity betweensurveys in the temporal pattern of vocalisations in relation to dawn we calculated aPearson correlation matrix between surveys for each type of vocalisation (mean numberof songs or calls uttered per male per 5-min period) Each correlation matrix was used asthe similarity matrix in a cluster analysis (Unweighted pair-group method with arithmeticaverages UPGMA method Sneath amp Sokal 1973) aimed at identifying groups of surveyswith similar dawn activity patterns for songs and calls
RESULTSThe only characteristic of singing activity related to the advance of the breeding seasonwas singing output since total number of songs uttered per male and night significantlyincreased in later surveys while singing performance and singing start or ending time didnot show a significant trend through surveys (Table 1 Fig 3) On the contrary callingactivity was strongly related to the advance of the breeding season since calling start timecalling performance period and calling output differed significantly among surveys while
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 617
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717
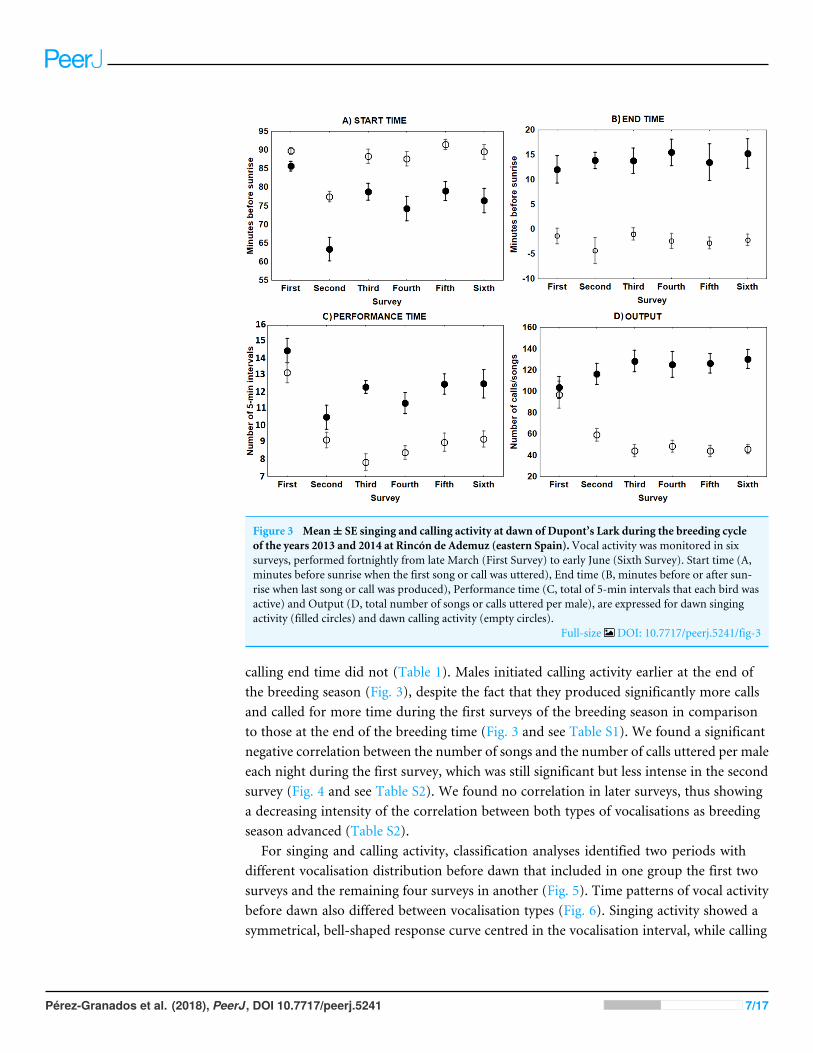
Figure 3 Meanplusmn SE singing and calling activity at dawn of Dupontrsquos Lark during the breeding cycleof the years 2013 and 2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in sixsurveys performed fortnightly from late March (First Survey) to early June (Sixth Survey) Start time (Aminutes before sunrise when the first song or call was uttered) End time (B minutes before or after sun-rise when last song or call was produced) Performance time (C total of 5-min intervals that each bird wasactive) and Output (D total number of songs or calls uttered per male) are expressed for dawn singingactivity (filled circles) and dawn calling activity (empty circles)
Full-size DOI 107717peerj5241fig-3
calling end time did not (Table 1) Males initiated calling activity earlier at the end ofthe breeding season (Fig 3) despite the fact that they produced significantly more callsand called for more time during the first surveys of the breeding season in comparisonto those at the end of the breeding time (Fig 3 and see Table S1) We found a significantnegative correlation between the number of songs and the number of calls uttered per maleeach night during the first survey which was still significant but less intense in the secondsurvey (Fig 4 and see Table S2) We found no correlation in later surveys thus showinga decreasing intensity of the correlation between both types of vocalisations as breedingseason advanced (Table S2)
For singing and calling activity classification analyses identified two periods withdifferent vocalisation distribution before dawn that included in one group the first twosurveys and the remaining four surveys in another (Fig 5) Time patterns of vocal activitybefore dawn also differed between vocalisation types (Fig 6) Singing activity showed asymmetrical bell-shaped response curve centred in the vocalisation interval while calling
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 717
Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Figure 4 Linear relationships between the number of songs and calls uttered per each Dupontrsquos Larkmale and survey (data of 2013 and 2014 pooled) at Rincoacuten de Ademuz (eastern Spain)Vocal activitywas monitored in six surveys performed fortnightly from late March (First Survey) to early June (SixthSurvey) Surveys with significant Pearsonrsquos correlations are represented with filled symbols and their R2
value can be found in legend (see complete analyses in Table S2)Full-size DOI 107717peerj5241fig-4
Table 1 Test of significance of the survey period as fixed effect in the GLMMs for different responsevariables describing singing and calling behaviour of the Dupontrsquos Lark at dawn
Songs Calls
Response variable Sum Sq F Pr(gtF) Sum Sq F Pr(gtF)
Start time 7265 132 0252 90246 4583 lt0001End time 10292 090 0343 165 0034 0854Performance time 138 022 0639 9628 1758 lt0001Output 67053 448 0036 25704 2170 lt0001
activity peaked between 85ndash75 min before dawn and decreased from that time Timepatterns of both vocalisation types were advanced with respect to dawn in the secondgroup of surveys and were remarkably similar within each group In addition callingpattern during the last phase of the breeding season presented a bimodal pattern with analmost total absence of calls between 30 and 45 min before dawn which was lacking in thefirst two surveys (Fig 6)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 817
Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Figure 5 Similarity in the distribution of each type of vocalisations (A songs B calls) duringDupontrsquos Lark dawn chorusesDendrogram resulted from hierarchical cluster analyses using completelinkage Pearsonrsquos correlation between surveys in the mean number of songs or calls uttered per male per5-min period was used as similarity measure Correlations were converted to distances computed as 10minus Pearson r
Full-size DOI 107717peerj5241fig-5
Figure 6 Dawn vocal activity pattern of Dupontrsquos Lark during the breeding cycle of the years 2013 and2014 at Rincoacuten de Ademuz (eastern Spain)Vocal activity was monitored in six surveys performed fort-nightly from late March (First Survey) to early June (Sixth Survey) Vocal activity is expressed as the meannumber of songs or calls uttered per male per 5-min interval Mean singing (filled line) and calling (dot-ted line) activity in the first (A) and the second (B) periods identified by cluster analysis (see Fig 4) Meandaily pattern of singing (C) and calling (D) activity for the surveys included in the periods defined in Fig4
Full-size DOI 107717peerj5241fig-6
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 917
DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

DISCUSSIONTo our knowledge this is the first study to analyse seasonal patterns of dawn chorus of apasserine using songs and calls with data collected at the same time (but see LaZerte Otteramp Slabbekoorn 2017) This is also the first study to quantitatively describe the dawn chorusof the Dupontrsquos Lark an endangered species We found that dawn seasonal patterns ofsinging and calling behaviour differed significantly dawn singing output increased withthe advance of the breeding period while dawn calling activity decreased and started earlieras the breeding season advanced The different seasonal patterns of calls and songs suggestthat they serve diverse functions at dawn (Celis-Murillo et al 2016) and therefore the useof one or the other vocalisation could lead to different conclusions about dawn chorusfunction in the studied species
In several passerine species a pattern of greater dawn singing activity at the beginningof the breeding period has been described followed by a relaxation after pair formationin agreement with the hypothesis that dawn song plays a role in mate attraction (eg GilGraves amp Slater 1999 Amrhein Korner amp Naguib 2002 Liu 2004) Conversely most ofthe dawn singing activity parameters analysed in the Dupontrsquos Lark remained constantthroughout the breeding season and even song output showed a seasonal increasing trendover time This temporal pattern does not agree with the patterns expected if the functionof songs was to attract females This suggests that dawn song could play a role in territorialdefence in the studied species due to constant dawn singing activity over time as hasbeen proposed in other passerines (Olinkiewicz amp Osiejuk 2003 Amrhein Kunc amp Naguib2004 Liu 2004 Kunc Amrhein amp Naguib 2005) Previous studies have assumed that dawnsinging behaviour in passerines can be used in both male and female interaction contextswith their functions difficult to ascertain (Slagsvold Dale amp Saeligtre 1994 Olinkiewicz ampOsiejuk 2003 Zhang Celis-Murillo amp Ward 2016) Both functions have been assignedespecially to continuous and loud singing species at dawn such as the Dupontrsquos Larkwhich are able to repel intruders while attracting potential mates or extra-pair mates(Moslashller 1991 Naguib et al 2011)
Contrary to dawn song most of the calling activity parameters analysed changed asthe breeding season advanced The number of calls and the amount of time dedicatedto calling decreased as breeding season progressed especially after the first two surveysLaiolo et al (2008) also found that Dupontrsquos Lark calling activity peaked in the first monthsof the breeding period This seasonal pattern is compatible with the pattern expected ifthis vocalisation serves to attract mates as has been proposed for other passerine birds(eg McCallum Grundel amp Dahlsten 1999 Gil Graves amp Slater 1999 Amrhein Korneramp Naguib 2002 Liu 2004) In multi-brooded species vocalisations that attract femalesare maintained albeit with reduced intensity This can be explained as a male response tomaintain their mates and quickly mate between successive clutches or if the nest is predatedor fails (Mace 1986 Paumlrt 1991) The same explanation could be valid for the DupontrsquosLark as this species has a long breeding season with up to three clutches per year (earlyMarchndashearly July) and high nest predation rates (Herranz et al 1994 Peacuterez-Granados etal 2017b)
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1017
Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Song start time did not vary over time while males tended to initiate calling activityearlier with the advance of the breeding season If calls were used for mate attraction thisresult may be related to acquiring extra-pair copulations which usually occur before dawna phenomenon well established among larks (Saacutenchez et al 2004 Hutchinson amp Griffith2008) Previous studies on passerines have found that males who began their daily dawnchorus the earliest were the most successful in obtaining extra-pair copulations (Poesel etal 2006 Dolan et al 2007) Time relative to dawn of the last call or song uttered did notdiffer throughout the breeding season likely because the end of dawn choruses is relatedto increasing light intensity as sunrise approaches (eg Da Silva et al 2014 York Youngamp Radford 2014) However this result may be influenced by the methodology employedsince on some occasions males continued calling after surveys ended which may haveinfluenced our results
The number of songs and calls uttered per bird each night were strong and negativelyrelated in the first survey but this relationship weakened with the advance of the breedingseason and was non-existent from the third survey onwards This pattern could be theconsequence of a trade-off between territorial defence and mate attraction (Hasselquist ampBensch 1991 Slagsvold Dale amp Saeligtre 1994) because males may utilise different acousticstrategies (calling or singing) when attracting mates or defending territories (ZhangCelis-Murillo amp Ward 2016) This trade-off may be enhanced in species like DupontrsquosLark that concentrate vocal activity in short periods in the daily cycle A larger proportionof unpaired males at the first stage of the breeding season would make this trade-off moreapparent at that time as thesemales would invest more time in calling for attracting femalesat the cost of reducing song output while after pairing more efforts would be devoted tosinging
We found different daily patterns at dawn for singing and calling activity In generalDupontrsquos Lark uttered calls mostly during the first 30 min of dawn choruses with songsbeing the commonest vocalisation onwards The mean daily pattern of singing and callingactivity were similar along the breeding season but that pattern was more advancedin the later surveys with birds singing and calling earlier in time This definite dailypattern together with seasonal differences found between vocalisation types support ourassumption that calls and songs may serve diverse functions in the studied species as hasbeen proposed for other passerines (Celis-Murillo et al 2016) According to hypotheticalfunctions assumed for dawn singing and dawn calling in the studied species our resultssuggest that males may dedicate more time to mate attraction at the beginning of dawnchoruses under low-light conditions and to territorial defence from that moment onThese different patterns may also reflect a trade-off between singing and calling withineach period before dawn as the peak in singing is associated with a decrease in calling Thetemporal pattern of calling becomes bimodal from the third survey onwards as callingactivity ceases completely between 45 and 35min before dawn just after the peak in signingSong output increases as season advances and particularly after the second survey andthus cessation of calling could be due to increasing efforts in singing
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1117
CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

CONCLUSIONSIn this study we found evidence that dawn singing and calling activity in Dupontrsquos Larkpresent different temporal patterns of activity both along the breeding season and in theperiod before dawn when most of the vocal activity of this species occurs These differentpatterns suggest that calls and songs in the Dupontrsquos Lark may have functions differentto those expected However experimental studies are needed to obtain conclusive resultsabout the concrete function of each vocalisation type in the studied species To unravel thefunctions of these vocalisations the next step should be to develop experimental studiestesting responses of males and females to songs and calls that would provide additionalinsight into dawn chorus signalling functions in the Dupontrsquos Lark Nevertheless ourresults highlight that the type of vocalisation used is an important factor to consider whenseasonal changes of vocal activity are studied and thus previous interpretations madefor birdsrsquo dawn choruses may need to be re-evaluated in light of the type of vocalisationanalysed We encourage researchers to clearly describe the type of vocalisation used infuture studies
ACKNOWLEDGEMENTSWe are grateful to three anonymous reviewers whose comments helped to improvethe manuscript We wish to thank the Servicio de Vida Silvestre of ConselleriadrsquoInfraestructures Territori i Medi Ambient (Generalitat Valenciana) and Juan Jimeacutenezfor supporting our work We thank Sarah Young for help with the written English
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis research was supported by the project lsquolsquoEstudio aplicado a la conservacioacuten de laconservacioacuten de las poblaciones de alondra ricotiacute (Chersophilus duponti) en el entornodel municipio de Vallancarsquorsquo funded by lsquoLevantina y Asociado de Minerales SArsquorsquo Thefunders had no role in study design data collection and analysis decision to publish orpreparation of the manuscript
Grant DisclosuresThe following grant information was disclosed by the authorsLevantina y Asociado de Minerales SA
Competing InterestsThe authors declare there are no competing interests
Author Contributionsbull Cristian Peacuterez-Granados conceived and designed the experiments performed theexperiments analyzed the data contributed reagentsmaterialsanalysis tools preparedfigures andor tables authored or reviewed drafts of the paper approved the final draft
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1217
bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

bull Tomasz S Osiejuk prepared figures andor tables materialsanalysis tools authored orreviewed drafts of the paper approved the final draft
bull Germaacuten M Loacutepez-Iborra conceived and designed the experiments analyzed the datacontributed reagentsmaterialsanalysis tools prepared figures andor tables authoredor reviewed drafts of the paper approved the final draft
Data AvailabilityThe following information was supplied regarding data availability
Peacuterez Granados Cristians (2018) Dawn chorus Dupontrsquos lark figshare Filesethttpsdoiorg106084m9figshare5924641v1
Supplemental InformationSupplemental information for this article can be found online at httpdxdoiorg107717peerj5241supplemental-information
REFERENCESAllen FH 1913More notes on the morning awakening Auk 30229ndash235
DOI 1023074071548Amrhein V Erne N 2006 Dawn singing reflects past territorial challenges in the winter
wren Animal Behaviour 71(5)1075ndash1080 DOI 101016janbehav200507023Amrhein V Korner P NaguibM 2002 Nocturnal and diurnal singing activity in the
nightingale correlations with mating status and breeding cycle Animal Behaviour64(6)939ndash944 DOI 101006anbe20021974
Amrhein V Kunc HP NaguibM 2004 Seasonal patterns of singing activity vary withtime of day in the nightingale (Luscinia megarhynchos) The Auk 121(1)110ndash117DOI 1016420004-8038(2004)121[0110SPOSAV]20CO2
Barrero A Goacutemez-Catasuacutes J Peacuterez-Granados C Abril-Coloacuten I Bustillo-de la RosaD Traba J 2017 Son los mejores machos los que maacutes defienden el territorio InComportamiento territorial de los machos de alondra ricotiacute durante el periodo reproduc-tor en la ZEPA Altos de Barahona Soria Poacutester Actas del XXIII Congreso Espantildeol deOrnitologiacutea pp 140 2ndash5 Noviembre Badajoz Espantildea
Bates D Maechler M Bolker BWalker S 2015 lme4 Linear mixed-effects models usingEigen and S4 R package version 11ndash7 2014 Institute for Statistics and Mathematicsof WU website Available at http cranr-projectorgpackage=lme4 (accessed on 18March)
Catchpole CK Slater PJ 2008 Bird song biological themes and variations 2nd editionCambridge Cambridge University Press
Celis-Murillo A Stodola KW Pappadopoli B Burton JMWardMP 2016 Seasonaland daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla) Journalof Ornithology 157(3)853ndash860 DOI 101007s10336-015-1318-y
Cramp S 1988 The birds of the Western Palearctic Vol 5 New York Oxford UniversityPress
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1317
Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Da Silva A Samplonius JM Schlicht E ValcuM Kempenaers B 2014 Artificial nightlighting rather than traffic noise affects the daily timing of dawn and dusk singing incommon European songbirds Behavioral Ecology 25(5)1037ndash1047
Dolan AC MurphyMT Redmond LJ Sexton K Duffield D 2007 Extrapair paternityand the opportunity for sexual selection in a socially monogamous passerineBehavioral Ecology 18(6)985ndash993 DOI 101093behecoarm068
Eriksson DWallin L 1986Male bird song attracts femalesmdasha field experimentBehavioral Ecology and Sociobiology 19(4)297ndash299 DOI 101007BF00300645
Garamszegi LZ Pavlova DZ Eens M Moslashller AP 2006 The evolution of song in femalebirds in Europe Behavioral Ecology 18(1)86ndash96
Gil D Graves JA Slater PJ 1999 Seasonal patterns of singing in the willow war-bler evidence against the fertility announcement hypothesis Animal Behaviour58(5)995ndash1000 DOI 101006anbe19991211
Grava A Otter KA Grava T LaZerte SE Poesel A Rush AC 2013 Character dis-placement in dawn chorusing behaviour of sympatric mountain and black-cappedchickadees Animal Behaviour 86(1)177ndash187 DOI 101016janbehav201305009
Grava T Grava A Otter KA 2009 Supplemental feeding and dawn singing in black-capped chickadees The Condor 111560ndash564 DOI 101525cond2009080061
Hasselquist D Bensch S 1991 Trade-off between mate guarding and mate attrac-tion in the polygynous great reed warbler Behavioral Ecology and Sociobiology28(3)187ndash193
Herranz J Manrique J Yanes M Suaacuterez F 1994 The breeding biology of Dupontrsquos larkChersophilus duponti in Europe Avocetta 18(2)141ndash146
Hutchinson J Griffith SC 2008 Extra-pair paternity in the Skylark Alauda arvensis Ibis150(1)90ndash97
Kacelnik A Krebs JR 1983 The dawn chorus in the great tit (Parus major) proximateand ultimate causes Behaviour 83287ndash309 DOI 101163156853983X00200
KenwardMG Roger JH 1997 Small sample inference for fixed effects from restrictedmaximum likelihood Biometrics 53(3)983ndash997
Kunc HP Amrhein V NaguibM 2005 Seasonal variation in dawn song char-acteristics in the common nightingale Animal Behaviour 70(6)1265ndash1271DOI 101016janbehav200502010
Kuznetsova A Brockhoff PB Christensen RHB 2014 lmerTest tests for random andfixed effects for linear mixed effect models (lmer objects of lme4 package) R packageversion 20ndash11 Available at httpswwwCRANR-project orgpackage=lme4
Laiolo P 2008 Characterizing the spatial structure of songbird cultures EcologicalApplications 18(7)1774ndash1780 DOI 10189007-14331
Laiolo P Tella JL 2005Habitat fragmentation affects culture transmission patternsof song matching in Dupontrsquos lark Journal of Applied Ecology 42(6)1183ndash1193DOI 101111j1365-2664200501093x
Laiolo P Tella JL 2006 Landscape bioacoustics allow detection of the effectsof habitat patchiness on population structure Ecology 87(5)1203ndash1214DOI 1018900012-9658(2006)87[1203LBADOT]20CO2
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1417
Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Laiolo P Voumlgeli M Serrano D Tella JL 2005 Two new calls from the Dupontrsquos LarkChersophilus duponti and imitations by other Alaudids ecological and monitoringimplications Ardeola 52(1)167ndash172
Laiolo P Voumlgeli M Serrano D Tella JL 2007 Testing acoustic versus physical markingtwo complementary methods for individual-based monitoring of elusive speciesJournal of Avian Biology 38(6)672ndash681 DOI 101111j20070908-885704006x
Laiolo P Voumlgeli M Serrano D Tella JL 2008 Song diversity predicts the viability offragmented bird populations PLOS ONE 3(3)e1822DOI 101371journalpone0001822
LaZerte SE Otter KA Slabbekoorn H 2017Mountain chickadees adjust songs callsand chorus composition with increasing ambient and experimental anthropogenicnoise Urban Ecosystems 20(5)989ndash1000
Lee JGH MacGregor-Fors I Yeh PJ 2017 Sunrise in the city disentangling driversof the avian dawn chorus onset in urban greenspaces Journal of Avian Biology48(7)955ndash964 DOI 101111jav01042
LiuWC 2004 The effect of neighbours and females on dawn and daytime singingbehaviours by male chipping sparrows Animal Behaviour 68(1)39ndash44DOI 101016janbehav200306022
LiuWC Kroodsma DE 2007 Dawn and daytime singing behavior of chipping sparrows(Spizella passerina) Auk 12444ndash52DOI 1016420004-8038(2007)124[44DADSBO]20CO2
Mace R 1986 Importance of female behaviour in the dawn chorus Animal Behaviour34(2)621ndash622 DOI 101016S0003-3472(86)80139-7
Mace R 1987 The dawn chorus in the great tit Parus major is directly related to femalefertility Nature 330745ndash746 DOI 101038330745a0
Marini KL ReudinkMW LaZerte SE Otter KA 2017 Urban mountain chickadees(Poecile gambeli) begin vocalizing earlier and have greater dawn chorus output thanrural males Behaviour 154(12)1197ndash1214 DOI 1011631568539X-00003464
Marler P 2004 Bird calls their potential for behavioral neurobiology Annals of the NewYork Academy of Sciences 1016(1)31ndash44 DOI 101196annals1298034
McCallumDA Grundel R Dahlsten DL 1999 Mountain chickadee (Poecile gambeli)In Poole A ed The birds of North America online Ithaca Cornell Laboratory ofOrnithology
Moslashller AP 1988 Spatial and temporal distribution of song in the yellowhammerEmberiza citrinella Ethology 78(4)321ndash331
Moslashller AP 1991Why mated songbirds sing so much mate guarding and maleannouncement of mate fertility status American Naturalist 138(4)994ndash1014DOI 101086285264
MurphyMT Sexton K Dolan AC Redmond LJ 2008 Dawn song of the easternkingbird an honest signal of male quality Animal Behaviour 75(3)1075ndash1084DOI 101016janbehav200708020
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1517
NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

NaguibM Kunc HP Sprau P Roth T Amrhein V 2011 Communication networks andspatial ecology in Nightingales Advances in the Study of Behavior 43239ndash271DOI 101016B978-0-12-380896-700005-8
NaguibM Van Rooij EP Snijders L Van Oers K 2016 To sing or not to sing seasonalchanges in singing vary with personality in wild great tits Behavioral Ecology27(3)932ndash938 DOI 101093behecoarv235
OdomKJ Benedict L 2018 A call to document female bird songs applications fordiverse fields The Auk 135(2)314ndash325 DOI 101642AUK-17-1831
Olinkiewicz A Osiejuk TS 2003 Effect of time of season and neighbours on singingactivity in the Corn BuntingMiliaria calandra Acta Ornithologica 38117ndash122DOI 1031610680380208
Paumlrt T 1991 Is dawn singing related to paternity insurance The case of the collaredflycatcher Animal Behaviour 41(3)451ndash456 DOI 101016S0003-3472(05)80847-4
Peacuterez-Granados C Loacutepez-Iborra GM 2013 Census of breeding birds and popula-tion trends of the Dupontrsquos Lark Chersophilus duponti in Eastern Spain Ardeola60(1)143ndash150 DOI 1013157arla6012012143
Peacuterez-Granados C Loacutepez-Iborra GM 2015 Baja dispersioacuten adulta y baja tasa derecaptura juvenil de la alondra ricotiacute (Chersophilus duponti) en el Rincoacuten de Ademuz(Valencia) In XX Congreso de Anilladores 10 de octubre Ceuta Oral communicationGrupo de Anillamiento Chagra Ceuta
Peacuterez-Granados C Loacutepez-Iborra GM Garza V Traba J 2017b Breeding biology ofthe endangered Dupontrsquos Lark Chersophilus duponti in two separate Spanish shrub-steppes Bird Study 64(3)328ndash338 DOI 1010800006365720171359232
Peacuterez-Granados C Loacutepez-Iborra GM Seoane J 2017a A multi-scale analysisof habitat selection in peripheral populations of the endangered DupontrsquosLark Chersophilus duponti Bird Conservation International 27(3)398ndash413DOI 101017S0959270916000356
Peacuterez-Granados C Osiejuk T Loacutepez-Iborra GM 2016Habitat fragmentation effectsand variations in repertoire size and degree of song sharing among close DupontrsquosLark Chersophilus duponti populations Journal of Ornithology 157471ndash482DOI 101007s10336-015-1310-6
Poesel A Foerster K Kempenaers B 2001 The dawn song of the blue tit Parus caeruleusand its role in sexual selection Ethology 107(6)521ndash531DOI 101046j1439-0310200100680x
Poesel A Kunc HP Foerster K Johnsen A Kempenaers B 2006 Early birds are sexymale age dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus)caeruleus Animal Behaviour 72(3)531ndash538 DOI 101016janbehav200510022
RDevelopment Core Team 2014 R a language and environment for statistical com-puting Vienna R Foundation for Statistical Computing Available at httpswwwR-projectorg
Saacutenchez AM Herranz J Martiacutenez JG Suaacuterez F 2004 Extra-pair paternity in Short-toedand Lesser Short-toed Larks Calandrella brachydactyla and C rufescens Bird Study51(3)278ndash279 DOI 10108000063650409461364
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1617
Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717

Slagsvold T Dale S Saeligtre GP 1994 Dawn singing in the great tit (Parus major)mate attraction mate guarding or territorial defence Behaviour 131(1)115ndash138DOI 101163156853994X00244
Sneath PH Sokal RR 1973Numerical taxonomy San Francisco FreemanStaicer CA Spector DA Horn AG 1996 The dawn chorus and other diel patterns in
acoustic signaling In Kroodsma DE Miller EH eds Ecology and evolution of acousticcommunication in birds New York Cornell Univ Press 426ndash453
Stanley CQWalter MH VenkatramanMXWilkinson GS 2016 Insect noise avoid-ance in the dawn chorus of Neotropical birds Animal Behaviour 112255ndash265DOI 101016janbehav201512003
Stowell D Gill L Clayton D 2016 Detailed temporal structure of communica-tion networks in groups of songbirds Journal of the Royal Society Interface13(119)20160296 DOI 101098rsif20160296
Voumlgeli M Serrano D Pacios F Tella JL 2010 The relative importance of patch habitatquality and landscape attributes on a declining steppe-bird metapopulationBiological Conservation 143(5)1057ndash1067 DOI 101016jbiocon200912040
Wang G Harpole CE Trivedi AK Cassone VM 2012 Circadian regulation of birdsong call and locomotor behavior by pineal melatonin in the zebra finch Journalof Biological Rhythms 27(2)145ndash155 DOI 1011770748730411435965
WebbWH Brunton DH Aguirre JD Thomas DB ValcuM Dale J 2016 Femalesong occurs in songbirds with more elaborate female coloration and reduced sexualdichromatism Frontiers in Ecology and Evolution 422 DOI 103389fevo201600022
Wright HW 1912Morning awakening and even-song Auk 29307ndash327DOI 1023074071038
York JE Young AJ Radford AN 2014 Singing in the moonlight dawn song per-formance of a diurnal bird varies with lunar phase Biology Letters 1020130970DOI 101098rsbl20130970
Zann R 1990 Song and call learning in wild zebra finches in south-east Australia AnimalBehaviour 40(5)811ndash828 DOI 101016S0003-3472(05)80982-0
Zhang VY Celis-Murillo AWardMP 2016 Conveying information with one song typechanges in dawn song performance correspond to different female breeding stagesBioacoustics 25(1)19ndash28 DOI 1010800952462220151076348
Peacuterez-Granados et al (2018) PeerJ DOI 107717peerj5241 1717


















