
Cyclic alternating pattern and spectral analysis of heart rate variability during normal sleep
6
Cyclic alternating pattern and spectral analysis of heart rate variability during normal sleep RAFFAELE FERRI 1,2 , LIBORIO PARRINO 3 , ARIANNA SMERIERI 3 , MARIO G. TERZANO 3 , MAURIZIO ELIA 2 , SEBASTIANO A. MUSUMECI 2 andSALVATOREPETTINATO 1 1 Sleep Research Center, 2 Department of Neurology, Oasi Institute for Research on Mental Retardation and Brain Aging (IRCCS), Troina, 3 Sleep Disorders Center, Department of Neurology, University of Parma, Parma, Italy Accepted in revised form 24 October 1999; received 23 December 1998 INTRODUCTION The natural electroencephalographic (EEG) arousal rhythm of non-rapid eye movement (NREM) sleep is known as the cyclic alternating pattern (CAP) (Terzano et al. 1985, 1988). CAP consists of arousal-related phasic events (Phase A) that interrupt, at intervals of 20–40 s, the tonic theta/delta activities of NREM sleep (Phase B). Functionally, CAP translates a condition of sustained arousal instability while the comple- mentary EEG pattern, i.e. non-CAP (NCAP), characterized by a rhythmic background activity with few, randomly distributed SUMMARY The natural arousal rhythm of non-rapid eye movement (NREM) sleep is known as the cyclic alternating pattern (CAP), which consists of arousal- related phasic events (Phase A) that periodically interrupt the tonic theta/ delta activities of NREM sleep (Phase B). The complementary condition, i.e. non-CAP (NCAP), consists of a rhythmic electroencephalogram background with few, randomly distributed arousal-related phasic events. Recently, some relation between CAP and autonomic function has been preliminarily reported during sleep in young adults by means of spectral analysis of heart rate variability (HRV). The present study was aimed at analysing the eects of CAP on HRV in a group of normal children and adolescents. Six normal children and adolescents (age range 10.0–17.5 y) were included in this study. All-night polygraphic recordings were performed after adaptation to the sleep laboratory. Six 5-min epochs were selected from sleep Stage 2 and six from Stages 3 and 4 (slow-wave sleep), both in CAP and NCAP conditions. From such epochs, a series of parameters describing HRV was then calculated, in both time and frequency domains, on the electrocardiographic R–R intervals. Statistical comparison between CAP and NCAP epochs revealed a significant dierence for most of the frequency domain parameters (increase of the low-frequency band, increase of the low-frequency/high-frequency ratio and decrease in the high-frequency band during CAP) both in Stage 2 and in slow-wave sleep. Our results demonstrate that the physiological fluctuations of arousal during sleep described as CAP are accompanied by subtle, but significant, changes in balance between the sympathetic and vagal components of the autonomic system. KEYWORDS autonomic function, cyclic alternating pattern, heart rate variability, sleep, spectral analysis Correspondence: Dr R. Ferri, Sleep Research Center, Oasi Institute, Via Conte Ruggero 73, 94018 Troina, Italy. Tel.: +39 935 936 111; fax: +39 935 653 327; e-mail: [email protected]. J. Sleep Res. (2000) 9, 13–18 Ó 2000 European Sleep Research Society 13
-
Upload
raffaele-ferri -
Category
Documents
-
view
213 -
download
1
Transcript of Cyclic alternating pattern and spectral analysis of heart rate variability during normal sleep

Cyclic alternating pattern and spectral analysis
of heart rate variability during normal sleep
RAFFAELE FERR I 1 , 2 , L I BOR IO PARR INO 3 , AR IANNA SMER IER I 3 ,
MAR IO G . TERZANO 3 , MAUR IZ IO EL IA 2 , S EBAST IANO A . MUSUMEC I 2
and SALVATORE PETT INATO 1
1Sleep Research Center, 2Department of Neurology, Oasi Institute for Research on Mental Retardation and Brain Aging (IRCCS), Troina,3Sleep Disorders Center, Department of Neurology, University of Parma, Parma, Italy
Accepted in revised form 24 October 1999; received 23 December 1998
INTRODUCTION
The natural electroencephalographic (EEG) arousal rhythm of
non-rapid eye movement (NREM) sleep is known as the cyclic
alternating pattern (CAP) (Terzano et al. 1985, 1988). CAP
consists of arousal-related phasic events (Phase A) that
interrupt, at intervals of 20±40 s, the tonic theta/delta activities
of NREM sleep (Phase B). Functionally, CAP translates a
condition of sustained arousal instability while the comple-
mentary EEG pattern, i.e. non-CAP (NCAP), characterized by
a rhythmic background activity with few, randomly distributed
SUMMARY The natural arousal rhythm of non-rapid eye movement (NREM) sleep is
known as the cyclic alternating pattern (CAP), which consists of arousal-
related phasic events (Phase A) that periodically interrupt the tonic theta/
delta activities of NREM sleep (Phase B). The complementary condition,
i.e. non-CAP (NCAP), consists of a rhythmic electroencephalogram
background with few, randomly distributed arousal-related phasic events.
Recently, some relation between CAP and autonomic function has been
preliminarily reported during sleep in young adults by means of spectral
analysis of heart rate variability (HRV). The present study was aimed at
analysing the e�ects of CAP on HRV in a group of normal children and
adolescents. Six normal children and adolescents (age range 10.0±17.5 y)
were included in this study. All-night polygraphic recordings were
performed after adaptation to the sleep laboratory. Six 5-min epochs were
selected from sleep Stage 2 and six from Stages 3 and 4 (slow-wave sleep),
both in CAP and NCAP conditions. From such epochs, a series of
parameters describing HRV was then calculated, in both time and
frequency domains, on the electrocardiographic R±R intervals. Statistical
comparison between CAP and NCAP epochs revealed a signi®cant
di�erence for most of the frequency domain parameters (increase of the
low-frequency band, increase of the low-frequency/high-frequency ratio
and decrease in the high-frequency band during CAP) both in Stage 2 and
in slow-wave sleep. Our results demonstrate that the physiological
¯uctuations of arousal during sleep described as CAP are accompanied
by subtle, but signi®cant, changes in balance between the sympathetic and
vagal components of the autonomic system.
KEYWORDS autonomic function, cyclic alternating pattern, heart rate
variability, sleep, spectral analysis
Correspondence: Dr R. Ferri, Sleep Research Center, Oasi Institute,
Via Conte Ruggero 73, 94018 Troina, Italy. Tel.: +39 935 936 111;
fax: +39 935 653 327; e-mail: [email protected].
J. Sleep Res. (2000) 9, 13±18
Ó 2000 European Sleep Research Society 13

arousal-related phasic events, re¯ects a condition of stable
arousal.
The CAP sequences, formed by the regular succession of
Phases A and B, constitute the microstructural component of
sleep that accompanies the dynamic shifts between the NREM
stages and seems to play a role in the transition from NREM
to REM sleep (Terzano et al. 1985). Instead, NCAP is the
EEG expression of consolidated sleep within the NREM
stages. The physiological balance between CAP and NCAP
varies across the life span (Parrino et al. 1998) and has been
found to be altered in a number of sleep-disturbed conditions,
such as insomnia (Terzano and Parrino 1993), epilepsy
(Terzano et al. 1989, 1992), parasomnias (Zucconi et al.
1995) and mood disorders (Parrino et al. 1994).
CAP and NCAP not only constitute the dynamic sca�olds
for the microstructural organization of sleep, but also have
repercussions for motor and vegetative activities, which
¯uctuate during CAP and remain quiescent during NCAP.
Recently, some relation between CAP/NCAP and autonomic
function has been preliminarily reported in young adults
(Ferini-Strambi et al. 1997) by means of spectral analysis of
heart rate variability (HRV) during sleep. Heart rate is under
the control of e�erent sympathetic and vagal activities directed
to the sinus node, which are modulated by central brainstem
and peripheral oscillators (Malliani et al. 1991). Spectral
analysis of HRV is a quantitative, reliable method for
analysing the modulatory e�ects of neural mechanisms on
the sinus node (Task Force of the European Society of
Cardiology and the North American Society of Pacing and
Electrophysiology 1996) and two main components are
currently considered, high-frequency (HF) and low-frequency
(LF). The vagal activity is the major contributor to the HF
component, while the LF component is considered by some
authors to be a marker of sympathetic modulation and by
others to be a parameter including both vagal and sympathetic
in¯uences.
The aim of this study was to analyse the e�ects of the CAP
and NCAP conditions on HRV during NREM sleep in a
group of normal children and adolescents.
METHODS
Subjects
Ten subjects (mean age 13.4 y, range 10.0±17.5 y) were
admitted to this study. The protocol was approved by our
ethical review committee and informed consent was obtained
from the families of all subjects. All individuals were carefully
evaluated from the neurological, otorrhino-laryngoiatric and
cardiovascular points of view and failed to show signi®cant
abnormalities.
Recordings
All subjects slept in the laboratory for two consecutive nights.
The data were analysed during the second night. To determine
sleep stages, EEG (six channels), electro-oculogram (EOG)
and mental electromyogram were recorded by means of an
Oxford Medilog 9000-II recorder. Other physiological vari-
ables, such as electrocardiogram (CM4 derivation: anode in
position V4 and cathode attached to the manubrium of the
sternum), peripheral oxygen saturation, chest wall movement
by thoracic impedance and oro-nasal air¯ow with thermistors,
were recorded using an Oxford MPA-II recorder. All signals
were sampled at a rate of 128 Hz and were also reproduced on
paper by means of a Siemens Mingograf EEG 21 polygraph.
Sleep and HRV analysis
Sleep staging (macrostructure of sleep) was accomplished on
paper recordings, following standard criteria (Rechtscha�en
and Kales 1968). In particular, sleep staging was carried out by
visually analysing the EEG (C3-right earlobe derivation), EOG
(left and right outer canthi referred to the left earlobe) and
electromyogram (submentalis muscle). Body position was also
controlled by means of a video-camera; during all epochs
chosen for HRV analysis patients rested in a supine position.
In order to study sleep-stage-related HRV, a series of
5-min epochs was chosen from quiet wakefulness (W) and
the following stages during the ®rst two sleep cycles: sleep
Stage 1 comprising the preceding 2 min of quiet wakefulness
(W + S1), sleep Stage 2 (S2), sleep Stages 3 and/or 4 (slow-
wave sleep, SWS) and REM sleep. This ®rst-step selection
was carried out regardless of the ongoing microstructural
condition. Subsequently, CAP and NCAP sequences were
detected in each recording, during S2 and SWS, according to
the rules de®ned by Terzano et al. (1985, 1988). Therefore,
HRV was additionally studied on the recording night in
three di�erent 5-min epochs from CAP and in three di�erent
epochs from NCAP periods during both S2 and SWS. For
each subject, 12 epochs were selected (three from S2-CAP,
three from S2-NCAP, three from SWS-CAP and three from
SWS-NCAP).
In order to avoid gross e�ects on HRV, only CAP sequences
and NCAP periods without transient activation phases
(Schieber et al. 1971) or arousal (American Sleep Disorders
Association 1992) were selected. Moreover, because of the age
range of our subjects, children and adolescents, low amounts
of arousal were expected; in fact, the number of arousal events
shows a linear increase with age (Mathur and Douglas 1995;
Boselli et al. 1998). Within the structure of sleep, most of the
CAP sequences selected were identi®ed during the transition
from light to deep NREM sleep, in which the A phases are
basically composed of K-complexes or delta bursts (Ferrillo
et al. 1997), which are accompanied by less evident heart rate
changes. Finally, the eventual presence of apneas and hypop-
neas was also carefully controlled so that the epochs selected
for analysis were free of respiratory events.
In each 5-min epoch, ECG signals were analysed for
automatic detection of R waves using a self-made program
utilizing a simple threshold plus ®rst and second derivative
algorithms; however, careful visual inspection for possible
14 R. Ferri et al.
Ó 2000 European Sleep Research Society, J. Sleep Res., 9, 13±18
errors was performed on all epochs. In order to overcome the
problem of the low sampling rate of our recorders (128 Hz),
which might have caused a bias in the estimation of the
R-wave ®ducial point (Task Force of the European Society of
Cardiology and the North American Society of Pacing and
Electrophysiology 1996) and a consequent alteration of the
spectrum, a parabolic interpolation was used to re®ne its
evaluation (Merri et al. 1990; Bianchi et al. 1993). A series of
time domain measures was calculated: mean R±R-value,
standard deviation of all R±R intervals (SDNN), the square
root of the mean of the sum of the squares of di�erences
between adjacent R±R intervals (RMSSD), number of pairs of
adjacent R±R intervals di�ering by more than 50 ms in the
entire epoch (NN50) and percentage of NN50 among the total
R±R intervals (pNN50). The ®rst 256 R±R intervals from each
epoch were utilized for all subsequent analysis steps. The R±R
interval tachograms were processed by means of a FFT
algorithm and the following spectral parameters were
obtained: power in very low-frequency range, < 0.04 Hz
(VLF), power in low-frequency range, 0.04±0.15 Hz (LF),
power in high-frequency range, 0.15±0.4 Hz (HF), total power
(VLF + LF + HF), LF power in normalized units: LF/(total
power-VLF) ´ 100 (LF%), HF power in normalized units:
HF/(total power-VLF) ´ 100 (HF%), ratio of LF to HF (LF/
HF), frequency of highest peak in the VLF range (VLF peak),
frequency of highest peak in the LF range (LF peak) and
frequency of highest peak in the HF range (HF peak).
Statistical analysis
The comparison between HRV parameters obtained from
CAP and NCAP epochs, during each sleep stage considered in
this study, was performed by means of the Wilcoxon test for
paired data sets.
RESULTS
On average, the subjects included in this study slept for 8 h,
their sleep e�ciency index (total sleep time/time in bed) was
good (0.892) and the number of awakenings/h was very low
(0.770). A normal sleep macrostructure was observed in all
cases. The mean length of the CAP cycle during CAP
sequences was 25.2 s (Phase A 8.4 s and Phase B 16.8 s).
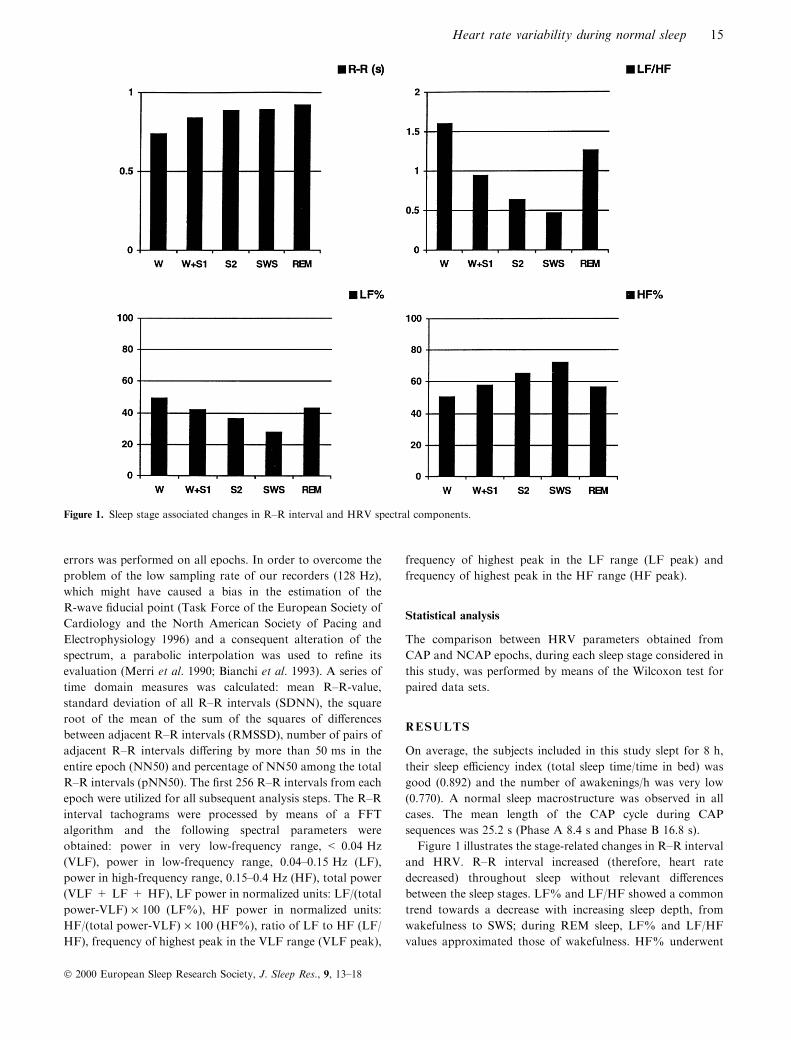
Figure 1 illustrates the stage-related changes in R±R interval
and HRV. R±R interval increased (therefore, heart rate
decreased) throughout sleep without relevant di�erences
between the sleep stages. LF% and LF/HF showed a common
trend towards a decrease with increasing sleep depth, from
wakefulness to SWS; during REM sleep, LF% and LF/HF
values approximated those of wakefulness. HF% underwent
Figure 1. Sleep stage associated changes in R±R interval and HRV spectral components.
Heart rate variability during normal sleep 15
Ó 2000 European Sleep Research Society, J. Sleep Res., 9, 13±18
complementary variations in wakefulness and in the di�erent
sleep stages.
The results of the comparison between HRV during CAP
and NCAP epochs in S2 and SWS, in the absence of sleep
apnea or arousal, are shown in Tables 1 and 2.
The mean R±R-values were very similar in both CAP and
NCAP conditions, with a limited interindividual variability.
During both S2 (Table 1) and SWS (Table 2) a signi®cant
di�erence between CAP and NCAP conditions emerged for
LF% and LF/HF ratio (higher in CAP than in NCAP) and for
HF% (lower in CAP than in NCAP); in SWS also VLF
showed values signi®cantly higher during CAP than during
NCAP states.
None of the time-domain parameters describing HRV
considered in this study (SDNN, RMSSD, NN50, pNN50)
showed any signi®cant di�erence between CAP and NCAP
epochs.
DISCUSSION
In the second century, Galen observed that sleep is accompan-
ied by a slowing in heart rate (Furley and Wilkie 1984);
however, it was necessary to wait for almost 2000 years in
order to achieve further insight into the sleep-related changes
of heart rate with the report by MacWilliam (1923) of increases
in heart rate and blood pressure with dreaming. Entering the
EEG and ECG era, sleep-stage-related changes in heart rate
started to be described in detail (Baust and Bonhert 1969;
Malpas and Purdie 1990; Cajochen et al. 1994; Pivik et al.
1996). More recently, spectral analysis of HRV has been used
to study ¯uctuations in the activity of the autonomic nervous
system during sleep. Using this technique, it has been reported
that LF shows a decrease during sleep, reaching minimal
values during SWS; in contrast, REM sleep is accompanied by
elevated LF values, similar to those of wakefulness, both in
children (Baharav et al. 1995) and young adults (Vaughn et al.
1995). In children, the opposite is true for HF, which increases
with sleep onset and reaches its highest values during SWS
(Baharav et al. 1995); in young adults HF has been reported to
be maximal during sleep Stage 2 (Vaughn et al. 1995). The
ratio between these two components of HRV (LF/HF) shows
changes similar to those of LF (Baharav et al. 1995).
Thus, spectral analysis of HRV has been considered to be a
reliable noninvasive method of quantifying changes in the
sustained tonic autonomic in¯uences on the heart during sleep.
The results obtained from the present analysis of sleep-stage
in¯uences on heart rate show good agreement with the ®ndings
of previous reports (Baharav et al. 1995; Vaughn et al. 1995).
Moreover, it must be emphasized that time-domain param-
eters were unable to disclose signi®cant CAP-related changes
in HRV, being less sensitive to subtle modi®cations of the
autonomic control of heart rate during sleep.
It must be noted that the age of the subjects included in this
study ranged over a period of life in which signi®cant changes
in sleep structure are to be expected. In particular, teenagers
show higher percentages of SWS and lower amounts of arousal
than older controls (Boselli et al. 1998; Parrino et al. 1998).
However, in this study we did not aim to evaluate develop-
mental aspects of HRV.
In addition to the correlation between depth of sleep and
variations of vegetative activity, there is also consolidated
evidence of transient changes in autonomic activity in
Table 1 Comparison between HR ®ndings during sleep Stage 2 CAP and NCAP epochs
S2±CAP S2±NCAPWilcoxon
Mean SD n Mean SD n P <
Mean R±R-value (s) 0.873 0.2368 10 0.902 0.2247 10 NS
SDNN 0.100 0.0585 10 0.090 0.0626 10 NS
RMSSD 0.089 0.0554 10 0.095 0.0929 10 NS
NN50 94.07 54.338 10 100.4 68.139 10 NS
pNN50 36.89 21.308 10 39.37 26.717 10 NS
VLF abs. (s2/beat) 2.699 3.4153 10 1.349 1.8085 10 NS
LF abs. (s2/beat) 1.001 1.0893 10 0.589 0.6959 10 NS
HF abs. (s2/beat) 1.224 1.2644 10 2.067 3.9133 10 NS
Total abs. (s2/beat) 4.925 5.3813 10 4.005 6.1430 10 NS
LF%* 41.67 5.6386 10 31.26 10.623 10 0.012
HF%* 58.33 5.6386 10 68.74 10.623 10 0.012
LF/HF 0.753 0.1945 10 0.513 0.2577 10 0.012
VLF peak (cycles/beat) 0.022 0.0088 10 0.016 0.0062 10 NS
LF peak (cycles/beat) 0.064 0.0177 10 0.071 0.0148 10 NS
HF peak (cycles/beat) 0.227 0.1797 10 0.220 0.0590 10 NS
SDNN, standard deviation of all R±R intervals, RMSSD, square root of the mean of the sum of the squares of differences between adjacent R±R
intervals, NN50, number of pairs of adjacent R±R intervals differing by more than 50 ms in the entire epoch, pNN50, NN50%, VLF, power in
very low-frequency range, <0.04 Hz, LF, power in low-frequency range, 0.04±0.15 Hz, HF, power in high-frequency range, 0.15±0.4 Hz, total
power, VLF + LF + HF, LF%, LF power in normalized units: LF/(total power-VLF) ´ 100, HF%, HF power in normalized units: HF/(total
power-VLF) ´ 100, LF/HF, ratio LF/HF, VLF peak, frequency of highest peak in the VLF range, LF peak, frequency of highest peak in the LF
range, HF peak, frequency of highest peak in the HF range. *Note that the value of SDs of LF% and HF% are identical because the sum
of these two parameters is always 100, in each subject.
16 R. Ferri et al.
Ó 2000 European Sleep Research Society, J. Sleep Res., 9, 13±18

association with spontaneous arousal-related phasic events.
Among such events, the most powerful e�ect on sympathetic
activity is exerted by arousal periods, which are actually
accompanied by robust increases in muscle sympathetic
activity, heart rate and blood pressure (Guilleminault and
Stoohs 1995). However, a less prominent, but still signi®cant,
acceleration of heart rate and rises in muscle sympathetic
activity and blood pressure are observed during NREM sleep
in the seconds that follow a K-complex (Hornyak et al. 1991)
or a delta burst (Church et al. 1978). In other words, all
arousal-related phasic events occurring during NREM sleep
are associated with short-lasting increases in sympathetic
activity and constitute a sort of continuum from weaker
(K-complexes and delta bursts) to powerful (arousal) EEG
features. In order to cast light exclusively on the impact of
the weaker events, in the present study, arousal periods were
excluded from the correlation between microstructural con-
ditions and HRV.
In a preliminary report, Ferini-Strambi et al. (1997) have
already shown that in young adults CAP and NCAP are
accompanied by signi®cant changes in HRV parameters.
However, these authors made no distinction between the
di�erent types of A phases included in their study and
probably also analysed epochs containing arousal or transient
activation phases. Our results indicate that such an e�ect is
also evident in children and adolescents, and that it reaches
statistical signi®cance even when the microanalysis of sleep is
cleansed of arousal. In particular, the increase in LF and LF/
HF ratio during S2±CAP and SWS±CAP (at the expense of a
complementary HF reduction) implies that during CAP the
sympatho-vagal balance is shifted towards sympathetic pre-
valence. Even if CAP-associated changes are of small ampli-
tude, they might be able to in¯uence the results of studies on
HRV during sleep in which they are not taken into account
due to the omitted scoring of CAP. No statistical comparison
was possible between epochs in which CAP was ignored and
only macrostructural analysis was accomplished, and those in
which CAP was evaluated. However, if we roughly compare
the HRV values expressed during the NREM sleep stages
(Fig. 1) and during the CAP/NCAP conditions (Tables 1 and
2), it can be seen that the CAP-related increase in sympathetic
activity is always higher than the mean value expressed by the
corresponding sleep stage as a whole. It is also interesting to
note that signi®cant autonomic di�erences between CAP and
NCAP were found both in Stage 2, which generally occupies
» 50% of the total sleep time in children and adolescents, and
in SWS, which generally occupies »25% of the total sleep time.
In other words, we are dealing with a microstructural
phenomenon that may have extensive HRV repercussions
throughout sleep. Therefore, the presence or absence of CAP
should be veri®ed, whenever possible, when studying HRV
during sleep.
As a constituent manifestation of NREM sleep, CAP occurs
even in the absence of sleep-perturbing factors (e.g. noise, pain,
myoclonic jerks, respiratory events). Basically, internal and
environmental disturbances vary the physiological amount of
CAP, but have limited e�ects on the 20±40-s alternating
rhythm of Phases A and B. Previous investigation has
ascertained that this periodicity is involved in the modulation
of a number of repetitive phenomena, such as periodic limb
movements during sleep (Parrino et al. 1996), sleep bruxism
(Macaluso et al. 1998) and sleep-related respiratory events
(Terzano et al. 1996). With respect to the present study, it can
be suggested that the same cerebral mechanisms underlying the
Table 2 Comparison between HR ®ndings during slow-wave sleep CAP and NCAP epochs
SWS±CAP SWS±NCAPWilcoxon
Mean SD n Mean SD n P <
Mean R±R-value (s) 0.879 0.2320 10 0.895 0.2268 10 NS
SDNN 0.078 0.0602 10 0.069 0.0554 10 NS
RMSSD 0.082 0.0871 10 0.088 0.0824 10 NS
NN50 83.63 64.146 10 99.38 68.119 10 NS
pNN50 32.79 25.153 10 37.79 26.705 10 NS
VLF abs. (s2/beat) 0.840 1.2892 10 0.116 0.0582 10 0.04
LF abs. (s2/beat) 0.445 0.7289 10 0.274 0.3877 10 NS
HF abs. (s2/beat) 1.590 3.5618 10 1.798 3.5829 10 NS
Total abs. (s2/beat) 2.874 5.0441 10 2.189 4.0020 10 NS
LF% 32.84 12.855 10 22.95 14.721 10 0.008
HF% 67.16 12.855 10 77.05 14.721 10 0.008
LF/HF 0.567 0.3569 10 0.365 0.3735 10 0.008
VLF peak (cycles/beat) 0.022 0.0084 10 0.018 0.0078 10 NS
LF peak (cycles/beat) 0.067 0.0176 10 0.084 0.0376 10 NS
HF peak (cycles/beat) 0.236 0.0603 10 0.240 0.0577 10 NS
SDNN, standard deviation of all R±R intervals, RMSSD, square root of the mean of the sum of the squares of differences between adjacent R±R
intervals, NN50, number of pairs of adjacent R±R intervals differing by more than 50 ms in the entire epoch, pNN50, NN50%, VLF, power in
very low-frequency range, <0.04 Hz, LF, power in low-frequency range, 0.04±0.15 Hz, HF, power in high-frequency range, 0.15±0.4 Hz, total
power, VLF + LF + HF, LF%, LF power in normalized units: LF/(total power-VLF) ´ 100, HF%, HF power in normalized units: HF/(total
power-VLF) ´ 100, LF/HF, ratio LF/HF, VLF peak, frequency of highest peak in the VLF range, LF peak, frequency of highest peak in the LF
range, HF peak, frequency of highest peak in the HF range.
Heart rate variability during normal sleep 17
Ó 2000 European Sleep Research Society, J. Sleep Res., 9, 13±18

regulation of arousal instability might be involved in the
genesis and enhancement of the sympathetic activity, since the
two phenomena (CAP and LF) operate within a common
20±40-s scale.
In conclusion, our data are in tune with the idea that
CAP modulates not only EEG activities, but also a series of
other sleep-related phenomena including autonomic ¯uctua-
tions (Terzano and Parrino 1993; Terzano et al. 1996).
Further studies are needed to clarify the vegetative impact of
both spontaneous and evoked CAP sequences, including
those containing arousal, and to investigate the role of
CAP-related autonomic changes in di�erent pathological
conditions.
REFERENCES
American Sleep Disorders Association. Arousals. Scoring rules and
examples. A preliminary report from the sleep disorders atlas task
force of the American Sleep Disorders Association. Sleep, 1992, 15:
174±184.
Baharav, A., Kotagal, S., Gibbons, V., Rubin, B. K., Pratt, G., Karin,
J. and Akselrod, S. Fluctuations in autonomic nervous activity
during sleep displayed by power spectrum analysis of heart rate
variability. Neurology, 1995, 45: 1183±1187.
Baust, W. and Bonhert, B. The regulation of heart rate during sleep.
Exp. Brain Res., 1969, 7: 169±180.
Bianchi, A. M., Mainardi, L. T., Petrucci, E., Signorini, M. G.,
Mainardi, M. and Cerutti, S. Time-variant power spectrum analysis
for the detection of transient episodes in HRV signal. IEEE Trans.
Biomed. Eng., 1993, 40: 136±144.
Boselli, M., Parrino, L., Smerieri, A. and Terzano, M. G. E�ects of age
on EEG arousals in normal sleep. Sleep, 1998, 21: 351±357.
Cajochen, C., Pischke, J., Aeschbach, D. and Borbe ly, A. A. Heart rate
dynamics during human sleep. Physiol. Behav., 1994, 55: 769±774.
Church, M. W., Laverne, C. W. and Seales, D. M. Evoked K-
complexes and cardiovascular responses to spindle-synchronous and
spindle-asynchronous stimulus clicks during NREM sleep. Electro-
enceph. Clin. Neurophysiol., 1978, 45: 443±453.
Ferini-Strambi, L., Mattioli, S., Bianchi, A., Zucconi, M., Oldani, A.,
Castronovo, C. and Smirne, S. The impact of cyclic alternating
pattern (CAP) on the spectral analysis of heart rate variability
during sleep in normal subjects. Sleep Res., 1997, 26: 74.
Ferrillo, F., Gabarra, M., Nobili, L., Parrino, L., Schiavi, G.,
Stubinski, B. and Terzano, M. G. Comparison between visual
scoring of cyclic alternating pattern (CAP) and computerized
assessment of slow EEG oscillations in the transition from light to
deep non-REM sleep. J. Clin. Neurophysiol., 1997, 14: 210±216.
Furley, D. and Wilkie, J. S. (Eds). Galen on Respiration and the
Arteries. Princeton University Press, Princeton, NJ, 1984.
Guilleminault, C. and Stoohs, R. Arousal, increased respiratory
e�orts, blood pressure and obstructive sleep apnoea. J. Sleep.
Res., 1995, 4 (Suppl. 1): 117±124.
Hornyak, M., Cejnar, M., Elam, M., Matousek, M. and Wallin, G.
Sympathetic muscle nerve activity during sleep in man. Brain, 1991,
114: 1281±1295.
Macaluso, G., Guerra, P., Di Giovanni, G., Boselli, M., Parrino, L.
and Terzano, M. G. Sleep bruxism is a disorder related to periodic
arousal during sleep. J. Dental Res., 1998, 77: 565±573.
MacWilliam, J. A. Some applications of physiology to medicine. III.
Blood pressure and heart action in sleep and dreams: their relation
to haemorrhages, angina, and sudden death. Br. Med. J., 1923, 2:
1196±1200.
Malliani, A., Pagani, M., Lombardi, F. and Cerutti, S. Cardiovascular
neural regulation explored in the frequency domain. Circulation,
1991, 84: 1482±1492.
Malpas, S. C. and Purdie, G. L. Circadian variation of heart rate
variability. Cardiovasc. Res., 1990, 24: 210±213.
Mathur, R. and Douglas, N. J. Frequency of EEG arousal from
nocturnal sleep in normal subjects. Sleep, 1995, 18: 330±333.
Merri, M., Farden, D. C., Mottley, J. G. and Titlebaum, E. L.
Sampling frequency of the electrocardiogram for the spectral
analysis of heart rate variability. IEEE Trans. Biomed. Eng., 1990,
37: 99±106.
Parrino, L., Boselli, M., Buccino, G. P., Spaggiari, M. C., Di
Giovanni, G. and Terzano, M. G. The cyclic alternating pattern
plays a gate-control on periodic limb movements during non-rapid
eye movement sleep. J. Clin. Neurophysiol., 1996, 13: 314±323.
Parrino, L., Boselli, M., Spaggiari, M. C., Smerieri, A. and Terzano,
M. G. Cyclic alternating pattern (CAP) in normal sleep: polysom-
nographic parameters in di�erent age groups. Electroenceph. Clin.
Neurophysiol., 1998, 107: 439±450.
Parrino, L., Spaggiari, M.C., Boselli, M., Di Giovanni, G. and
Terzano, M.G. Clinical and polysomnographic e�ects of trazodone
CR in chronic insomnia associated with dysthymia. Psychopharma-
cology, 1994, 116: 389±395.
Pivik, R. T., Busby, K. A., Gill, E., Hunter, P. andNevins, R. Heart rate
variations during sleep in preadolescents. Sleep, 1996, 19: 117±135.
Rechtscha�en, A. and Kales, A. (Eds). A Manual of Standardized
Terminology, Techniques and Scoring System of Sleep Stages of
Human Subjects. Washington Public Health Service. US Govern-
ment Printing O�ce, Washington D.C., 1968.
Schieber, J. P., Muzet, A. and Ferriere, P. J. R. Les phases d'activation
transitoire spontane es au cours du sommeil chez l'homme. Arch.
Sci. Physiol., 1971, 25: 443±465.
Task Force of the European Society of Cardiology and the North
American Society of Pacing and Electrophysiology. Heart rate
variability. Standards of measurements, physiological interpretation
and clinical use. Circulation, 1996, 93: 1043±1065.
Terzano, M. G., Mancia, D., Salati, M. R., Costani, G., Decembrino,
A. and Parrino, L. The cyclic alternating pattern as a physiologic
component of normal NREM sleep. Sleep, 1985, 8: 137±145.
Terzano, M. G., Parrino, L., Anelli, S., Boselli, M. and Clemens, B.
E�ects of generalized interictal EEG discharges on sleep stability:
assessment by means of cyclic alternating pattern. Epilepsia, 1992,
33: 317±326.
Terzano, M. G., Parrino, L., Anelli, S. and Halasz, P. Modulation of
generalized spike-and-wave discharges during sleep by cyclic alter-
nating pattern. Epilepsia, 1989, 30: 772±781.
Terzano, M. G., Parrino, L., Boselli, M., Spaggiari, M. C. and Di
Giovanni, G. Polysomnographic analysis of arousal responses in
OSAS by means of the cyclic alternating pattern (CAP). J. Clin.
Neurophysiol., 1996, 13: 145±155.
Terzano, M. G., Parrino, L. and Spaggiari, M. C. The cyclic
alternating pattern sequences in the dynamic organization of sleep.
Electroenceph. Clin. Neurophysiol., 1988, 69: 437±447.
Terzano, M. G. and Parrino, L. Clinical application of cyclic
alternating pattern. Physiol. Behav., 1993, 54: 807±813.
Terzano, M. G. and Parrino, L. Evaluation of EEG cyclic alternating
pattern during sleep in insomniacs and controls under placebo and
acute treatment with zolpidem. Sleep, 1993, 15: 64±70.
Vaughn, B. V., Quint, S. R., Messenheimer, J. A. and Robertson, K.
R. Heart period variability in sleep. Electroenceph. Clin. Neuro-
physiol., 1995, 94: 155±162.
Zucconi, M., Oldani, A., Ferini-Strambi, L. and Smirne, S. Arosual
¯uctuations in non-rapid eye movement parasomnias: the role of
cyclic alternating pattern as a measure of arousal instability. J. Clin.
Neurophysiol., 1995, 12: 147±154.
18 R. Ferri et al.
Ó 2000 European Sleep Research Society, J. Sleep Res., 9, 13±18


















