
Cyanogenic Glycosides: Synthesis, Physiology, and ...
33
Cyanogenic Glycosides: Synthesis, Physiology, and Phenotypic Plasticity Roslyn M. Gleadow 1 and Birger Lindberg Møller 2, 3 1 School of Biological Sciences, Monash University, 3800 Victoria, Australia; email: [email protected] 2 Plant Biochemistry Laboratory, Department of Plant and Environmental Sciences; Center for Synthetic Biology “bioSYNergy”; and VILLUM Research Center for Plant Plasticity, University of Copenhagen, DK-1871 Copenhagen, Denmark 3 Carlsberg Laboratory, DK-1799 Copenhagen, Denmark Annu. Rev. Plant Biol. 2014. 65:155–85 First published online as a Review in Advance on February 24, 2014 The Annual Review of Plant Biology is online at plant.annualreviews.org This article’s doi: 10.1146/annurev-arplant-050213-040027 Copyright c 2014 by Annual Reviews. All rights reserved Keywords cyanogenesis, hydrogen cyanide, cytochrome P450, plant defense, pathogen interactions, herbivore interactions Abstract Cyanogenic glycosides (CNglcs) are bioactive plant products derived from amino acids. Structurally, these specialized plant compounds are character- ized as α-hydroxynitriles (cyanohydrins) that are stabilized by glucosylation. In recent years, improved tools within analytical chemistry have greatly in- creased the number of known CNglcs by enabling the discovery of less abundant CNglcs formed by additional hydroxylation, glycosylation, and acylation reactions. Cyanogenesis—the release of toxic hydrogen cyanide from endogenous CNglcs—is an effective defense against generalist herbi- vores but less effective against fungal pathogens. In the course of evolution, CNglcs have acquired additional roles to improve plant plasticity, i.e., estab- lishment, robustness, and viability in response to environmental challenges. CNglc concentration is usually higher in young plants, when nitrogen is in ready supply, or when growth is constrained by nonoptimal growth condi- tions. Efforts are under way to engineer CNglcs into some crops as a pest control measure, whereas in other crops efforts are directed toward their re- moval to improve food safety. Given that many food crops are cyanogenic, it is important to understand the molecular mechanisms regulating cyanogen- esis so that the impact of future environmental challenges can be anticipated. 155 Annu. Rev. Plant Biol. 2014.65:155-185. Downloaded from www.annualreviews.org Access provided by Monash University on 07/23/20. For personal use only.
Transcript of Cyanogenic Glycosides: Synthesis, Physiology, and ...

PP65CH06-Gleadow ARI 8 April 2014 21:52
Cyanogenic Glycosides:Synthesis, Physiology,and Phenotypic PlasticityRoslyn M. Gleadow1 and Birger Lindberg Møller2,3
1School of Biological Sciences, Monash University, 3800 Victoria, Australia;email: [email protected] Biochemistry Laboratory, Department of Plant and Environmental Sciences;Center for Synthetic Biology “bioSYNergy”; and VILLUM Research Center for PlantPlasticity, University of Copenhagen, DK-1871 Copenhagen, Denmark3Carlsberg Laboratory, DK-1799 Copenhagen, Denmark
Annu. Rev. Plant Biol. 2014. 65:155–85
First published online as a Review in Advance onFebruary 24, 2014
The Annual Review of Plant Biology is online atplant.annualreviews.org
This article’s doi:10.1146/annurev-arplant-050213-040027
Copyright c© 2014 by Annual Reviews.All rights reserved
Keywords
cyanogenesis, hydrogen cyanide, cytochrome P450, plant defense,pathogen interactions, herbivore interactions
Abstract
Cyanogenic glycosides (CNglcs) are bioactive plant products derived fromamino acids. Structurally, these specialized plant compounds are character-ized as α-hydroxynitriles (cyanohydrins) that are stabilized by glucosylation.In recent years, improved tools within analytical chemistry have greatly in-creased the number of known CNglcs by enabling the discovery of lessabundant CNglcs formed by additional hydroxylation, glycosylation, andacylation reactions. Cyanogenesis—the release of toxic hydrogen cyanidefrom endogenous CNglcs—is an effective defense against generalist herbi-vores but less effective against fungal pathogens. In the course of evolution,CNglcs have acquired additional roles to improve plant plasticity, i.e., estab-lishment, robustness, and viability in response to environmental challenges.CNglc concentration is usually higher in young plants, when nitrogen is inready supply, or when growth is constrained by nonoptimal growth condi-tions. Efforts are under way to engineer CNglcs into some crops as a pestcontrol measure, whereas in other crops efforts are directed toward their re-moval to improve food safety. Given that many food crops are cyanogenic, itis important to understand the molecular mechanisms regulating cyanogen-esis so that the impact of future environmental challenges can be anticipated.
155
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
CNglc: cyanogenicglycoside
Cyanogenesis: therelease of HCN fromendogenous naturalproducts containing astabilizedα-hydroxynitrilefunctional group
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156STRUCTURE AND BIOSYNTHESIS OF CYANOGENIC GLYCOSIDES . . . . . . 158
Structural Diversity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158Biosynthesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159Turnover . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
CYANOGENESIS AND THE CYANIDE BOMB . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162HERITABILITY, POLYMORPHISM, AND QUANTITATIVE VARIATION . . . . 163
Presence/Absence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 163Quantitative Inheritance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165Where and When: Age Versus Developmental Stage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
HERBIVORES, PATHOGENS, AND WOUNDING RESPONSES. . . . . . . . . . . . . . . 166Herbivory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 166Pathogen Resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 167Wounding and Induction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168
ENVIRONMENTAL PLASTICITY AND OPTIMAL GROWTH . . . . . . . . . . . . . . . . 169Effects of Nitrogen Supply . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169Atmospheric CO2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 170Nonoptimal or Stressful Growth Conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
RESOURCE ALLOCATION: PURPOSES, COSTS, AND REGULATION. . . . . . . 172Energy Costs and Trade-Offs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 172Nitrogen as the Currency for Calculating Costs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 174
INTRODUCTION
Humans have a complicated relationship with hydrogen cyanide (HCN): It is toxic, and yet plantsthat generate HCN are an important part of the human diet. All plants produce tiny amounts ofHCN as an additional product in the biosynthesis of ethylene, but some plant species can releaselarge amounts from endogenously stored cyanogenic glycosides (CNglcs). CNglcs are bioactivenatural products (also called specialized plant products or secondary metabolites) derived fromamino acids with oximes and cyanohydrins (α-hydroxynitriles) as key intermediates. Glucosylationof the labile cyanohydrin results in the formation of a CNglc (133). CNglcs are stable compounds,but when the β-glycosidic linkage is hydrolyzed through the action of a β-glycosidase, the la-bile cyanohydrin that is formed dissociates to release HCN in a process known as cyanogenesis(137) (Figure 1). Acute or chronic exposure to HCN can lead to intoxication, mild to severeillness, and in extreme cases even death in humans (120) and animals (51) because HCN inhibitsthe activity of metalloenzymes, principally cytochrome c oxidase, the final enzyme in the respi-ratory electron transport chain (115). Hydrolysis of CNglcs results in the concomitant releaseof carbonyl compounds that may further augment the toxic and repellant effects of HCN (156).The oxime intermediates in the biosynthesis of CNglcs may also be toxic, especially toward fungi(99, 132).
CNglcs and the specific catabolic enzymes required to catalyze their hydrolysis are stored inseparate compartments within cells or in different tissues, preventing accidental autotoxicity (137).The two components are brought into contact upon tissue disruption, as caused by chewing her-bivores or when cell integrity is destroyed by physical processes, such as by freezing or maceration
156 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
HCN
α-Hydroxynitrile
Glucose
β-Glucosidase
Aldehyde/ketone
OximeAmino acid
α-Hydroxynitrile lyase (or spontaneous)Cyanogenic
monoglucoside
α-Hydroxynitrile(cyanohydrin)
UDP-glucose
UDP-glucosyltransferase
P450aa P450oxSugar
OC
N
R2
R1
Figure 1Biosynthesis and bioactivation of cyanogenic glycosides (CNglcs). CNglcs are synthesized from specific amino acids in a series ofreactions catalyzed by two multifunctional, membrane-bound cytochrome P450s (P450aa and P450ox) and a soluble UDP-glucosyl-transferase, with an oxime and an α-hydroxynitrile (cyanohydrin) as key intermediates. Cyanogenesis occurs when the β-glucosidiclinkage is hydrolyzed by a specific β-glycosidase to form an unstable α-hydroxynitrile (cyanohydrin) that dissociates into hydrogencyanide (HCN) and a ketone either spontaneously at high pH or catalyzed by an α-hydroxynitrilase. Abbreviations: aa, amino acid; ox,oxime.
Cyanide potential(HCNp): themaximum amount ofHCN that could bereleased from a giventissue; it provides analternative measure ofthe total CNglcconcentration
Cyanogenic capacity(HCNc): thecatabolic rate of HCNrelease from CNglcs;it is dependent on thetotal CNglc contentand the activity of thecatabolic enzymesβ-glycosidase andα-hydroxynitrilase
of the plant material as part of food processing (70). The cyanide potential (HCNp) is definedas the total amount of HCN that could be released by complete conversion of all cyanogeniccompounds in a given tissue to HCN (39); the cyanogenic capacity (HCNc) is the catabolic rateof HCN release from CNglcs (15).
CNglcs are effective deterrents to generalist herbivores (17, 70, 190, 204), and this is mostlikely the main evolutionary driver in their occurrence across the plant kingdom (3, 9, 124). Insome plant species, CNglcs have been implicated in the control of germination (52, 162) andbud burst via cyanohydrin and HCN formation (20). CNglcs may also serve as transport formsof carbon and nitrogen (179), and endogenous turnover processes may release the nitrogen fromCNglcs in the form of ammonia (90, 140, 157). More recently, it has been proposed that CNglcsmay also function in modulating oxidative stress (98, 133, 140).
CNglcs are unusually widespread in the plant kingdom. They are found in the oldest of ter-restrial plants (the ferns) and in gymnosperms and angiosperms (205). More than 3,000 plantspecies from all major vascular plant taxa, belonging to over 130 families of flowering plants, arecyanogenic; these represent 11% of the total number of plant species tested, with a clear over-representation of cultivated plants. This proportion is, therefore, somewhat higher than the 5%found in screens of natural population systems (65, 128). Throughout history, humans appear tohave inadvertently selected cyanogenic plants for cultivation, perhaps because of their increasedresistance to herbivores or because the need to process the plant material before consumptionmakes them less attractive to trespassers (93, 124).
The focus of this review is on the formation, turnover, and degradation of CNglcs and how theircontribution to phenotypic plasticity may optimize adaptation to environmental challenges. Thesynthesis and regulation of bioactive natural products are influenced by changes in the CO2 contentof the atmosphere (62, 64) and by changes in the climate (31, 194). It is important to be awareof and understand the consequences of such changes so that appropriate measures can be takento either reduce risks or capitalize on benefits because, for example, increased levels of CNglcsmay alter the host specificity of insects or pests. This is highly relevant for CNglcs, consideringthe option to bioengineer CNglc production into naturally noncyanogenic plant species in orderto improve their resistance to herbivores and pests as an alternative to the use of pesticides andfungicides (56, 190). Attempts are also being made to use classical mutagenesis-based breedingtechniques, or RNA-interference or antisense technology, to reduce the toxicity of edible tissuesof crop plants that accumulate large amounts of these compounds in order to make food crops saferfor consumption (25, 95). In nature, some plant species, such as white clover (Trifolium repens), are
www.annualreviews.org • Cyanogenic Glycosides 157
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
clearly polymorphic with respect to their ability to produce and degrade CNglcs. CNglcs are oneof many types of modulators of plant plasticity, and under most environmental conditions theyare not necessarily essential components. Given the known roles of CNglcs and their integrationinto plant primary metabolism, such manipulations may result in unexpected effects over thecourse of plant ontogeny (33). New understandings of the diverse roles of CNglcs shed light onthe physiological mechanisms underlying their contribution to improved phenotypic plasticity inaddition to their classical role as a constitutive defense system. At the end of the review, we listkey knowledge gaps and directions for future research on CNglcs.
STRUCTURE AND BIOSYNTHESIS OF CYANOGENIC GLYCOSIDES
Structural Diversity
All of the CNglcs so far identified are derived from aromatic and aliphatic amino acids (tyro-sine, phenylalanine, valine, isoleucine, and leucine) and a few nonproteinogenic amino acids: the(2S,1′R) and (2S,1′S) epimers of 2-(2′-cyclopentenyl)-glycine as well as (2S,1′S,2′S)-2-(2′-hydroxy-3′-cyclopentenyl)-glycine (36, 39, 133, 177). The most common CNglcs are the monoglucosidesdhurrin [(S)-4-hydroxymandelonitrile-β-D-glucopyranoside], prunasin [(R)-mandelonitrile-β-D-glucopyranoside], linamarin (2-hydroxyisobutyronitrile-β-D-glucopyranoside), and lotaustralin[(R)-2-hydroxy-2-methyl-butyronitrile-β-D-glucopyranoside] (Figure 2), which are derived fromtyrosine, phenylalanine, valine, and isoleucine, respectively. Cyanohydrins derived from tyrosine,phenylalanine, and 2-(2′-cyclopentenyl)-glycine (and notably not valine) contain a chiral center,giving rise to pairs of epimeric CNglcs like dhurrin and taxiphyllin, prunasin and sambunigrin,and deidaclin and tetraphyllin A, respectively (149).
The core structures of CNglcs may be further modified by single or multiple hydroxylations.These decorations are particularly common among the isoleucine- and leucine-derived CNglcs,such as those found in barley (143) and the genetic model plant Lotus japonicus (53, 187). Hydrox-ylation may also result in cleavage of the aromatic ring system, as observed in the biosynthesis ofthe tyrosine-derived triglochinin (142). It has not been resolved whether the hydroxylated CNglcswith a cyclopentanoid core structure are derived from hydroxylated parent amino acids or whetherhydroxylation proceeds as a late step in their synthesis (36).
The number of CNglcs found in nature is further expanded by the structural diversity ofthe sugar moiety. In the CNglcs currently known, the cyanohydrin is always stabilized by a β-glucosidic linkage to D-glucose. In cyanogenic diglycosides, the second sugar moiety may also bea D-glucose residue, with the most common examples being amygdalin (prunasin-6′-glucoside)and linustatin (linamarin-6′-glucoside), in both of which the second glucose residue is attachedby a β-1,6 linkage. However, the second glucose residue may also be bound in β-1.2, β-1.3,and β-1.4 linkages, as in eucalyptosin A, C, and B, respectively (139). Further structural diversityarises by incorporation of, e.g., arabinose, xylose, and apiose residues as additional sugar residues,as observed in vicianin, xeranthin, lucumin, and oxyanthin (4, 129, 163, 175). Yet another level ofdiversity arises from additional combinations of acylation and glycosylation reactions, as observedin oxyanthin 5′′-benzoate (163). The presence of these derivatives is typically restricted to specificstages in plant ontogeny or to specific tissues (129, 131). Isolation and structural characterizationare thus challenging and dependent on advanced liquid chromatography–mass spectrometry andNMR instrumentation, so only within the past few decades has the widespread occurrence of thesecomplex derivatives of the classical CNglcs been fully realized. Rather than being considered arather small class of specialized molecules, CNglcs are now thought of as a rapidly expanding anddiverse group of compounds.
158 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
OHOHO
OH
O
OH
CNH
a Prunasin
d Linamarin
g Epiheterodendrin
j Proteacin k Triglochinin l Dhurrin
m Xeranthin
h Tetraphyllin B i Oxyanthin 5''-benzoate
e Linustatin f Lotaustralin
b Sambunigrin c Amygdalin
OHO
HO
OH
O
OH
CNH
OHOHO
O
O
OH
CNHO
HOHO
OH
OH
OHOHO
OH
O
OH
CN
OHO
HO
OH
O
OH
CN
OHO
HO
O
O
OH
CNOHO
HO
OH
OH
OHO
HO
OH
O
OH
CN OHO
HO
OH
O
OH
OH
CN
OHOHO
O
O
OH
CNH
OHOHO
OH
O
OH
CNH
OO OH
OH
HO
HO
OHO
HO
OH
OH
NC
OHO
O
OH
OHO
HO
OH
O
OH
CNH
OH
HO
OH OH
O
O
OHOHO
O
O
OH
CNHOO
HOOH
OHO
OH OH
O
O
OHO
HOHO
HO
O
OH
Figure 2The structural diversity of cyanogenic glycosides (CNglcs). Prunasin (a) and sambunigrin (b) are epimers derived from phenylalanine.Amygdalin (c) is a prunasin-derived diglucoside. Linamarin (d ) and linustatin (e) are mono- and diglucosides (respectively) derived fromvaline. Lotaustralin ( f ) is derived from isoleucine and typically co-occurs with linamarin. Epiheterodendrin ( g) is derived from leucine.Tetraphyllin B (h) is derived from the nonproteinaceous amino acid 2-cyclopentenyl glycine following an additional hydroxylation.Oxyanthin 5′′-benzoate (i ) is an example of a more complex CNglc derived from phenylalanine and containing glucose and apioseresidues. Proteacin ( j ), triglochinin (k), and dhurrin (l ) are all derived from tyrosine. Xeranthin (m) is one of the most structurallycomplex CNglcs identified so far, being an acylated tetraglycoside derived from phenylalanine following an additional 3-hydroxylation.The sugar residues involved are glucose, xylose, and apiose, the latter of which is acylated by a cinnamic acid residue.
Biosynthesis
The biosynthetic pathway for CNglcs was first elucidated in Sorghum bicolor (122, 134, 182).In sorghum, the CNglc dhurrin is synthesized from L-tyrosine in a series of steps catalyzedby two multifunctional cytochrome P450 enzymes (CYP79A1 and CYP71E1) (8, 99, 100, 109,183) and a family 1 UDP-glucosyltransferase (UGT85B1) (94) in combination with the P450redox partner NADPH-dependent cytochrome P450 oxidoreductase (POR) (79) (Figure 3). The
www.annualreviews.org • Cyanogenic Glycosides 159
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
L-Ty
rosi
ne
CY
P7
9A
1
(E)-
p-H
ydro
xyp
he
nyl
-
ace
tald
eh
yde
oxi
me
CY
P7
1E
1
p-H
ydro
xym
an
de
lon
itri
le
HO
O OH
OH
N
HO
OH N
HO
O
N
HO
Glu
cose
UD
P-g
luco
se
UG
T8
5B
1
Glu
cose
Dh
urr
ina
se
p-H
ydro
xym
an
de
lon
itri
le
OH N
HO
Dh
urr
in
α-H
ydro
xyn
itri
lely
ase
p-H
ydro
xyb
en
zald
eh
ydeO
HO
HC
N +
H2S
Cys
tein
e
β-C
yan
oa
lan
ine
syn
tha
se β-C
yan
oa
lan
ine
NH
2N
H2
NH
2
O
OH
N
2H
2O
Nit
rila
se
L-A
spa
rag
ine
O
H2N
O
OH p-
Hyd
roxy
ph
en
yla
ceto
nit
rileN
HO
NH
3
Nit
rila
se h
ete
rom
er
NIT
4p-
Hyd
roxy
ph
en
yla
ceti
c a
cid
O
OH
HO
L-A
spa
rtic
aci
d
O
HO
O
OH
+o
rN
H3
NA
DP
+
Alt
ern
ate
turn
ov
er
pa
thw
ay
Glu
cose
NA
DP
H+
O2
NA
DP
H+
O2
NA
DP
+
Glu
tath
ion
eS-
tra
nsf
era
se?
+
Glu
tath
ion
e-
S-ly
ase
Glu
tath
ion
e Glu
tath
ion
e
con
jug
ate
Bio
syn
the
sis
Ca
tab
oli
sm
De
tox
ifica
tio
n
H2N
Figu
re3
Sche
me
illus
trat
ing
the
synt
hesi
san
dtu
rnov
erof
dhur
rin
inSo
rghu
mbi
colo
r.T
hest
eps
inth
epu
tativ
eal
tern
ativ
etu
rnov
erpa
thw
ayre
sulti
ngin
the
conv
ersi
onof
dhur
rin
into
4-hy
drox
yphe
nyla
ceto
nitr
ileha
veno
tbee
nde
fined
(90,
157)
,but
itis
know
nth
atSb
NIT
4A/B
2,a
nitr
ilase
hete
rodi
mer
,cat
alyz
esth
eco
nver
sion
of4-
hydr
oxyp
heny
lace
toni
trile
to4-
hydr
oxyp
heny
lace
ticac
idan
dam
mon
ia(9
0).p
-Hyd
roxy
phen
ylac
etic
acid
gluc
osid
eha
sbee
nsh
own
toac
cum
ulat
ein
etio
late
dso
rghu
mse
edlin
gs(7
8,90
).
160 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
first step is the conversion of tyrosine to (E)-p-hydroxyphenylacetaldoxime (183). This step, cat-alyzed by CYP79A1, is rate limiting in the biosynthesis of dhurrin in young sorghum seedlings(32). The (E)-p-hydroxyphenylacetaldoxime is then converted to p-hydroxymandelonitrile in aCYP71E1-catalyzed reaction (8). Glucosylation of p-hydroxymandelonitrile catalyzed by the sol-uble UGT85B1 enzyme generates dhurrin (94). p-Hydroxymandelonitrile is labile at neutraland alkaline conditions, and if it is not rapidly glucosylated, it dissociates into HCN and p-hydroxybenzaldehyde.
The enzymes catalyzing dhurrin formation in sorghum (CYP79A1, CYP71E1, UGT85B1,and POR) are thought to form a metabolon to promote rapid channeling of the toxic and labileintermediates into dhurrin formation and to prevent undesired metabolic crosstalk (91, 97, 113,144). CYP79A1, CYP71E1, and POR are anchored to the endoplasmic reticulum via a transmem-brane domain positioned near their N termini (8, 109), whereas UGT85B1 is a soluble enzyme(94). Metabolon assembly may be mediated by direct protein–protein interactions or by specificlipids (91, 190). It remains to be understood how the glycosides are transported from their initialproduction site in the cytosol not only to their storage site, typically in the vacuole (169), but also tothe specific tissues such as the epidermal cell layer and epicuticular wax of the barley leaf (88, 143).
The types of intermediates identified in the CNglc pathway in sorghum have also been foundin other species, such as flax (Linum usitatissimum) (44), white clover (T. repens) (87), cassava(Manihot esculenta) (6), L. japonicus (53), almond (Prunus amygdalus) (167), and Triglochin maritima(142). Thus, the key intermediates involved in biosynthesis of CNglcs appear to be the sameacross the plant kingdom. In addition to sorghum, the genes encoding the entire pathway forCNglc synthesis have now been identified in cassava (6, 96, 105) and L. japonicus (188). Genesencoding CYP79s and UGTs involved in CNglc synthesis have also been identified in whiteclover (150) and almond (57), respectively.
The aliphatic CNglcs linamarin and lotaustralin normally co-occur in plants, but the ratiosbetween the two CNglcs differ between species. The cassava genome contains two paralogs ofeach of the three biosynthetic genes involved in linamarin and lotaustralin formation. Preliminaryanalysis shows identical regulation and coexpression of the different genes and their paralogs (96,105). In L. japonicus, the paralogous genes CYP79D3 and CYP79D4 show differential expression,with CYP79D3 preferentially expressed in areal parts of the plant and CYP79D4 expressed exclu-sively in the roots (53). The CYP79s are considered the signature enzyme of the CNglc pathway,and all tested current members of this P450 family catalyze conversion of amino acids to the cor-responding oximes. Both CYP79 and the second P450 in the pathway, catalyzing the conversionof oxime to cyanohydrin, belong to the CYP71 clan of P450s (7). The latter P450 and the UGTthat catalyzes the final step in CNglc synthesis have a much broader substrate specificity comparedwith that of CYP79 (80, 94, 100).
Turnover
In plants containing large amounts of CNglcs, the nitrogen and sugar content of the CNglcs mayrepresent significant percentages of the total organic matter present (140). For example, in newlyformed shoots of Eucalyptus cladocalyx, as much as 25% of leaf nitrogen can be tied up in prunasin(64); in tips of young etiolated sorghum seedlings, dhurrin constitutes 30% of the dry matter(78); and in the epidermal cell layer of barley leaves, glucose bound in CNglcs constitutes 90% ofthe total soluble carbohydrate content (160). Under such circumstances, but also in general, theability of plants to reuse the carbon and nitrogen deposited in CNglcs (e.g., to balance resourcedemands in primary metabolism) offsets some of the energy and resource costs associated withtheir synthesis and storage (140).
www.annualreviews.org • Cyanogenic Glycosides 161
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
The classical catabolic pathway proceeds after tissue disruption and involves hydrolysis by aβ-glycosidase and dissociation of the cyanohydrin formed by the action of an α-hydroxynitrilase(39, 86) (Figures 1 and 3). This pathway is typically referred to as the bioactivation pathway orcyanogenesis. Part of the HCN may be recaptured and reincorporated into primary metabolismin a reaction catalyzed by β-cyanoalanine synthase that involves stoichiometric consumption ofcysteine (26, 158). Subsequent action of nitrilase 4 (NIT4) family enzymes results in asparagine andaspartate formation (90). More recently, an alternative pathway independent of prior tissue damagewas discovered and studied in sorghum (90). p-Hydroxyphenylacetonitrile is an intermediate inthis turnover pathway and is converted into p-hydroxyphenylacetic acid by the action of thenitrilase heteromer NIT4A/NIT4B2s with a concomitant release of ammonia (90) (Figure 3).The turnover rates achieved by this mechanism may be quite high, as demonstrated in youngsorghum plants, where it may reach 0.8% of the total CNglc present per hour (1). Rapid turnoverof dhurrin is also observed upon extended growth of sorghum plants (32).
CYANOGENESIS AND THE CYANIDE BOMB
The process resulting in the release of HCN from CNglcs by the action of a β-glycosidaseis referred to as cyanogenesis, and sometimes also as the cyanide bomb (137). Hydrolysis ofglucosinolates by the action of myrosinases represents a similar binary system, in this case resultingin the release of thiocyanates and related compounds, generally referred to as the mustard oil bomb(205). In both these binary systems, compartmentalization is essential to prevent autotoxicity andto ensure that the “bomb” is detonated as a targeted response, e.g., to herbivore attack (137).The most common type of compartmentation is at the level of the organelle. CNglcs, at least inthe leaves, are typically confined to the vacuole (e.g., 40, 98, 169). Within the stems and petiolesof cassava, linamarin is confined to vesicle-like structures within the latex (48). The location ofthe β-glycosidases involved in hydrolysis of CNglcs is more variable. Depending on the species,they may be present in the apoplastic space, bound to the cell wall, in the cytoplasm, in smallvesicles, or in the chloroplast (48, 58, 101, 111, 165, 191). Less is known about the location of theα-hydroxynitrilases, but the evidence available suggests that they are cytoplasmic in both sorghum(111) and Hevea brasiliensis (84).
Separation of CNglcs and their degradative enzymes may also occur at the tissue level. Insorghum, for example, dhurrin is found almost exclusively in the epidermal cell layer of the leafblade, whereas dhurrinase and α-hydroxynitrilase are located predominantly in the mesophyllcells (111, 169, 191). A similar epidermal localization of CNglcs has also been observed in limabeans (Phaseolus lunatus) (58). In barley (Hordeum vulgare), the CNglcs and β- and γ-hydroxynitrileglucosides are likewise localized in the epidermal cells of the leaf blade, whereas β-glucosidaseactivity related to their hydrolysis is restricted to the endosperm of the germinating seed (143).Within fruits, some plants concentrate the CNglcs in the seeds [e.g., H. brasiliensis (179), Prunusserotina (186), and Canthium schimperianum (174)], whereas in others, the cyanogenic component isthe fruit itself [e.g., Passiflora edulis (34)]. Some species have only trace amounts of CNglcs in the dryseeds [e.g., sorghum (120) and clover (87, 195)], whereas others have completely acyanogenic seeds[e.g., E. cladocalyx (69)]. Prunasin and amygdalin are located in the parenchyma of the cotyledons inthe seeds of rosaceous stone fruits, whereas the β-glucosidase and α-hydroxynitrilase are locatedin the procambium (159, 165, 186).
The β-glycosidases responsible for hydrolysis of CNglcs have been isolated and their genesidentified in several species. Sorghum contains two β-glucosidases, the dhurrinases Dhr1 andDhr2, which hydrolyze dhurrin with high specificity and organ-specific expression (35). Dhr1accumulates in the mesocotyl and the root tip, whereas Dhr2 accumulates in the leaves (35, 191),
162 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
although the functional significance of this difference is unknown. White clover contains a singlelinamarase encoded by the Li locus (152). L. japonicus contains the two paralogous β-glucosidasesBGD2 and BGD4. BGD2 is able to hydrolyze all the nitrile glucosides present in L. japonicus,including linamarin and lotaustralin. In contrast, BGD4 specifically hydrolyzes the isoleucine-derived β- and γ-hydroxynitrile glucosides designated as rhodiocyanoside D and A, respectively(136, 188).
The β-glucosidases discussed above have cyanogenic monoglucosides as substrates. Bitter al-monds contain large amounts of the cyanogenic diglucoside amygdalin and smaller amounts ofthe monoglucoside prunasin (167). Amygdalin is hydrolyzed by amygdalin hydrolase, resulting inthe formation of the monoglucoside prunasin, which may then be cleaved by a prunasin hydrolase.Amygdalin hydrolase and prunasin hydrolase constitute small enzyme families (110). No evidencefor formation of the diglucoside gentiobiose was observed upon the action of amygdalin hydro-lase (167). In contrast, β-glycosidase-catalyzed cleavage of the cyanogenic diglycoside vicianinresults in formation of the diglycoside vicianose, composed of arabinose and glucose (4). Thus,depending on the plant species and CNglc, β-glycosidase-catalyzed hydrolysis of CNglcs harbor-ing more than a single sugar moiety may proceed sequentially as well as by direct initial releaseof the aglycone. Cyanogenic diglycosides are frequently referred to as storage or transport forms.Formation of the diglycosides may prevent their hydrolysis by β-glycosidases, as documented inH. brasiliensis, where the linamarase hydrolyzing the monoglucoside linamarin was shown to notcleave the co-occurring diglucoside linustatin (76).
HERITABILITY, POLYMORPHISM, AND QUANTITATIVE VARIATION
Presence/Absence
Cyanogenesis is a stable, heritable trait. In some species, all individuals are cyanogenic, whereasothers are highly polymorphic, with phenotypically acyanogenic individuals (e.g., 42, 54, 67, 87,130, 170, 202). Relatively few species are monomorphic (e.g., 95, 121, 125, 128). Cyanogenesisin white clover is a classic example of Mendelian inheritance with two independently segregatingloci—one indicative of the ability to produce CNglcs (Ac) and the other indicative of the presenceof the specific β-glucosidase able to degrade the CNglcs (Li ) (43, 81, 87, 151, 152). Plants mustbe homozygous recessive at one of the loci to be acyanogenic. Both phenotypes persist in thepopulation as a consequence of selection for both cyanogenic and acyanogenic phenotypes underdifferent environmental and herbivore pressures, giving rise to a balanced polymorphism (43, 87,104). Plants with the Acaclili genotype are not naturally cyanogenic, although they are still able tosynthesize CNglcs. Plant material containing CNglcs but without enzymes able to degrade themmay pose a toxicity risk because the CNglcs may be broken down to release HCN in some animals,such as in cattle rumens (51). For example, Turnera ulmifolia may contain the specific β-glucosidaseeven when the capacity to synthesize CNglcs is absent (170), whereas Acacia species that are knownto synthesize CNglcs always lack the requisite β-glucosidase for natural cyanogenesis (41).
Although the genetics of cyanogenesis has been studied in clover for more than 50 years, theactual basis of the mutation was only recently established: The nonfunctional ac allele is the result ofa deletion in CYP79D15, the gene encoding the first P450 in the pathway (151). Lack of linamaraseis linked to partial or complete loss of the Li gene from the white clover genome (152). Thereis no evidence of linkage between Ac and Li in clover at the phenotypic (87) or molecular level(151, 152), despite the obvious advantages of inheriting both together. A recently described ethylmethanesulfonate–generated mutation in sorghum, tcd1, that renders CYP79A1 nonfunctional isalso recessive and follows a Mendelian pattern of inheritance (25), also without any indication of
www.annualreviews.org • Cyanogenic Glycosides 163
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
CYP79
CYP71E and CYP736
UGT85CYP79D2
CYP71E7
CYP71E UGT85K5
CYP79A1 UGT85B1 CYP71E1
CYP71
CYP79D3 CYP79D4
CYP736p
UGT85K3
Sorg
hum
bic
olor
Lo
tus j
apon
icus
M
anih
ot e
scul
enta
Chromosome 3, contig CM0241 (region 140–560 kb)
Chromosome 1 (region 1,040–1,170 kb)
Scaffold 08265 (region 850–950 kb)
UGT85K4
CYP736A2
CYP736p CYP736p
10 kb
10 kb
20 kb
Rho
Figure 4Schematic representation of the clustering of cyanogenic glucoside biosynthetic genes in the genomes of Lotus japonicus (chromosome3), Sorghum bicolor (chromosome 1), and Manihot esculenta (chromosome number not yet established because the genome is still in draftform). Functional genes are shown with arrows indicating their orientation. Confirmed genes in cyanogenic glucoside biosynthesis arelabeled above each bar, with CYP79 genes in pink, CYP71E and CYP736 genes in green, and UGT85 genes in blue. Adapted fromReference 187.
linkage. The last step in cyanogenesis is the dissociation of the cyanohydrin with concomitantrelease of HCN. This process is catalyzed by an α-hydroxynitrilase but occurs spontaneously atalkaline pH. Little is known about the α-hydroxynitrilases catalyzing the dissociation process (24).
In L. japonicus, sorghum, and cassava, the three genes coding for the specific enzymes thatcatalyze the formation of CNglcs are clustered on a single chromosome (187, 189) (Figure 4).Clustering of nonhomologous genes encoding enzymes for synthesis of plant defense compoundsis a recently discovered phenomenon. It was first reported in maize (Zea mays) for the synthesisof the benzoxazinoid defense compound DIMBOA (2,4-dihydroxy-1,4-benzoxazin-3-one) (61).Some terpenoid pathways are also organized in gene clusters. This applies to synthesis of avenacinin oat (Avena strigosa) (138, 161), synthesis of momilactone and multifunctional phytocassanes-oryzalides/oryzadiones in rice (Oryza sativa) (196), and synthesis of steroidal glycoalkaloids inpotato (Solanum tuberosum) and tomato (Solanum lycopersicum) (89). The genes encoding synthesisof the opiate alkaloid noscapine in the opium poppy (Papaver somniferum) are also clustered (201).
In CNglc biosynthesis, the individual enzymes catalyzing a specific step are encoded by genesthat are related between species but not necessarily by orthologous genes. Likewise, the organiza-tion of the three known gene clusters encoding CNglc synthesis in the three species investigatedis very different (187). Together, these point to independent evolution of CNglc biosynthesis inseveral higher-plant lineages by repeated recruitment and neofunctionalization of members fromsimilar multigene families (187, 189). Some insects are able to synthesize cyanogenic glucosides denovo (202, 203), as observed in the Burnet moth (Zygaena filipendulae). The insect genes CYP405A2,CYP332A3, and UGT33A1, which encode the entire biosynthetic pathway, have been recruited
164 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
from the same two multigene families as in plants. The low sequence identity between the plantand insect genes clearly demonstrates that the insect pathway has evolved by convergent evolutionand not following horizontal gene transfer or by divergent evolution, as might have been assumedconsidering that the P450s involved are multifunctional and catalyze highly unusual reactions (72,92). Thus, plants and insect have independently found a way to package a cyanide time bomb tofend off herbivores, predators, and pests. Interestingly, the genes coding for the cyanogenic β-glucosidases are not clustered with the biosynthetic genes (187, 189), consistent with the observedindependence between β-glucosidase activity and HCNp in nonmodel species (67, 172).
Quantitative Inheritance
CNglc concentration (HCNp) shows high quantitative variability even within genetically identicalplants of single clones (31, 193). Broad-sense heritability, calculated by comparing the cyanogenicpotential of half sibs, has been found to be approximately 0.8 in several species (e.g., 19, 72, 173).Given that the inheritance of the trait is well known, this is likely to be due to other factors, suchas differences in physiology or in uptake and allocation of resources. The activity of cyanogenicβ-glycosidases varies quantitatively as well and appears to be a determining factor in the rateof HCN release from CNglcs (HCNc) (11). A strong positive correlation between HCNp andβ-glucosidase activity has been reported in Eucalyptus polyanthemos (73) but was not detected innatural populations of E. cladocalyx (68).
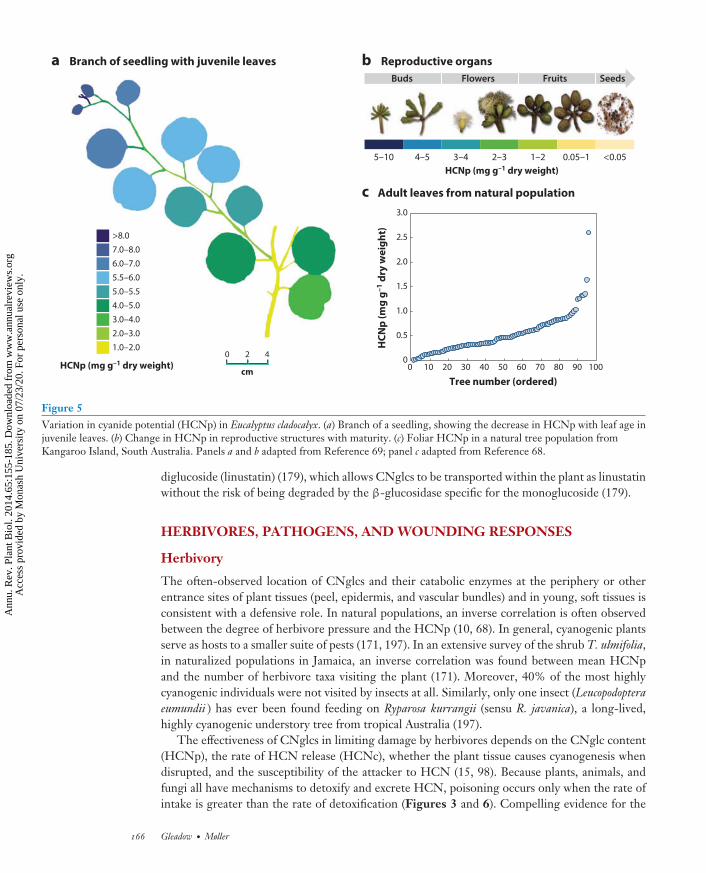
Where and When: Age Versus Developmental Stage
One of the difficulties in comparing the cyanogenic status of individual plants is that theHCNp varies ontogenetically, phenologically, and chronologically. HCNp is generally highest inseedlings and decreases with plant age (45, 69, 114, 193, 199). For example, in E. cladocalyx, in theseries Sejunctae, seedlings have a high HCNp (74) (Figure 5). Notable exceptions to the patterndescribed above are the cyanogenic Eucalyptus species from the series Maidenaria. They are essen-tially acyanogenic as seedlings (<10 ppm HCN), becoming cyanogenic only after 6–12 months(74). Initiation of prunasin synthesis in Maidenaria appears to be linked to a change from juvenile,dorsiventral leaf anatomy to adult isobilateral leaves. A similar pattern occurs in lima beans, whereonly secondary leaves are cyanogenic (17). Interestingly, coppice leaves in E. cladocalyx, which bearjuvenile leaf morphology but sprout from epicormic buds on mature trees, have an HCNp similarto that of leaves of seedlings (74). Newly formed tissues are also nearly always more cyanogenicthan older tissues (42), as in E. cladocalyx, where HCNp is as high in newly formed shoots andyoung reproductive organs of adult plants as it is in seedlings (69) (Figure 5). This may corre-late with the transcript levels of the CYP79 genes involved, as in sorghum, where the CYP79A1transcript levels are higher in young seedlings (32), and in L. japonicus, where expression of thetwo CYP genes governing the synthesis of lotaustralin and linamarin (CYP79D3 and CYP736A2)is highest in the apical leaves (53, 188).
CNglcs may accumulate in all parts of a plant [e.g., as in cassava (125)], only in the abovegroundparts [e.g., as in Eucalyptus (69) and white clover (87, 185)], or only in vegetative tissues [e.g., asin sorghum (120)]. In plants that synthesize more than a single CNglc, the ratio between thedifferent types of CNglcs may differ in above- and belowground parts. This pattern may varywith reproductive stage as well. Some T. ulmifolia populations, for example, lose their cyanogeniccapacity around flowering, whereas others do not (173). Vegetative and reproductive tissues ofplants may also contain different types of CNglcs, and the content in roots may differ from thatin shoots. Most Prunus species contain prunasin in the leaves but store the related diglucoside,amygdalin, in the seeds (165, 186). H. brasiliensis also contains a monoglucoside (linamarin) and a
www.annualreviews.org • Cyanogenic Glycosides 165
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.
PP65CH06-Gleadow ARI 8 April 2014 21:52
Tree number (ordered)
0 10 20 30 40 50 60 70 80 90 1000
0.5
1.0
1.5
2.0
2.5
3.0
HCNp (mg g–1 dry weight)
HC
Np
(m
g g
–1 d
ry w
eig
ht)
5–10 4–5 3–4 2–3 1–2 0.05–1 <0.05
0 2 4
HCNp (mg g–1 dry weight)
1.0–2.0
2.0–3.0
3.0–4.0
4.0–5.0
5.0–5.5
5.5–6.0
6.0–7.0
7.0–8.0
>8.0
cm
a Branch of seedling with juvenile leaves b Reproductive organs
c Adult leaves from natural population
Buds Flowers Fruits Seeds
Figure 5Variation in cyanide potential (HCNp) in Eucalyptus cladocalyx. (a) Branch of a seedling, showing the decrease in HCNp with leaf age injuvenile leaves. (b) Change in HCNp in reproductive structures with maturity. (c) Foliar HCNp in a natural tree population fromKangaroo Island, South Australia. Panels a and b adapted from Reference 69; panel c adapted from Reference 68.
diglucoside (linustatin) (179), which allows CNglcs to be transported within the plant as linustatinwithout the risk of being degraded by the β-glucosidase specific for the monoglucoside (179).
HERBIVORES, PATHOGENS, AND WOUNDING RESPONSES
Herbivory
The often-observed location of CNglcs and their catabolic enzymes at the periphery or otherentrance sites of plant tissues (peel, epidermis, and vascular bundles) and in young, soft tissues isconsistent with a defensive role. In natural populations, an inverse correlation is often observedbetween the degree of herbivore pressure and the HCNp (10, 68). In general, cyanogenic plantsserve as hosts to a smaller suite of pests (171, 197). In an extensive survey of the shrub T. ulmifolia,in naturalized populations in Jamaica, an inverse correlation was found between mean HCNpand the number of herbivore taxa visiting the plant (171). Moreover, 40% of the most highlycyanogenic individuals were not visited by insects at all. Similarly, only one insect (Leucopodopteraeumundii ) has ever been found feeding on Ryparosa kurrangii (sensu R. javanica), a long-lived,highly cyanogenic understory tree from tropical Australia (197).
The effectiveness of CNglcs in limiting damage by herbivores depends on the CNglc content(HCNp), the rate of HCN release (HCNc), whether the plant tissue causes cyanogenesis whendisrupted, and the susceptibility of the attacker to HCN (15, 98). Because plants, animals, andfungi all have mechanisms to detoxify and excrete HCN, poisoning occurs only when the rate ofintake is greater than the rate of detoxification (Figures 3 and 6). Compelling evidence for the
166 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
Thiosulfate ThiocyanateSulfiteHCN +Rhodanese
+
Figure 6Method of detoxifying hydrogen cyanide (HCN) commonly found in mammals. HCN is converted to theless toxic thiocyanate and partly excreted in the urine. Both this and the β-cyanoalanine pathway thatdominates in plants (Figure 3) rely on the availability of the sulfur-containing amino acid cysteine.
effectiveness of CNglcs in herbivore defense comes from transgenic studies. Tattersall et al. (190)transferred the entire pathway for synthesis of the CNglc dhurrin from sorghum into Arabidopsisthaliana and then fed the transgenic plants to the flea beetle (Phyllotreta nemorum), a commonherbivore of crucifers. The larvae ate 80% less tissue of the Arabidopsis leaves transformed tocontain dhurrin than they ate of the leaves of control plants.
From the plant’s point of view, avoidance of herbivore attack is an optimal strategy because iteliminates tissue loss. HCN and concomitantly released aliphatic or aromatic carbonyl compoundshave distinct odors that can act as feeding deterrents (11, 155). Interestingly, only ∼80% of humanscan detect the odor of HCN (77). The marzipan odor of almonds, although sometimes attributedto HCN, arises from benzaldehyde, which is manufactured as “almond essence.” Taste is alsoimportant. Several studies have detected a correlation between bitterness and HCNp (108, 116),as recognized in the common names of highly cyanogenic varieties of Prunus, such as bitter andsweet cherries (P. emarginata and P. avium, respectively) and almonds (P. amygdalus syn. P. dulcius).Traditional farmers use the bitterness of cassava as an indicator of the HCNp of cassava (124),although this could be due to the strong association with the occurrence of a bitter apiosyl glucosideand not just the cyanogenic glucosides (108).
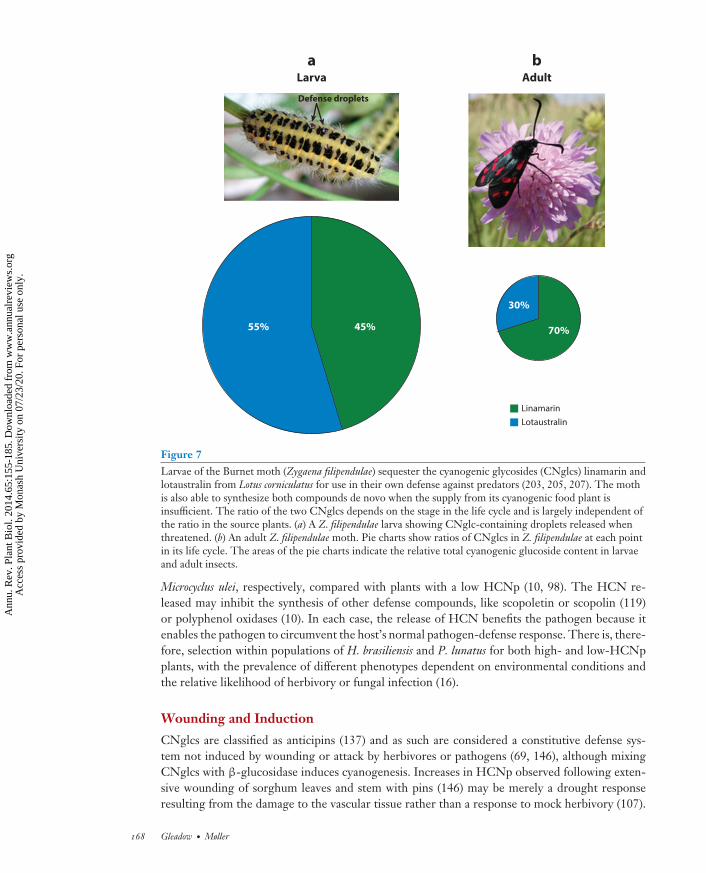
Several specialist herbivores not only tolerate CNglcs but also actually sequester them for usein their own arsenal of defense compounds against predators (145, 155, 172, 203). For example,larvae of Euptoieta hegesia (Lepidoptera) that sequester CNglcs from their host (T. ulmifolia L.)are more distasteful to their Anolis predators (172). Larvae of the Burnet moth (Z. filipendulae;Lepidoptera) are able to sequester the CNglcs linamarin and lotaustralin from their cyanogenichost plants, typically bird’s-foot trefoil (202–207). If the larvae cannot sequester sufficient amountsof the CNglcs, then replenishment is achieved by de novo synthesis in the larvae (see Figure 7for details). The pathway for CNglc synthesis in Z. filipendulae has been elucidated using genomicmethods. A sequence comparison of the insect and plant genes involved clearly shows that theinsect pathway has evolved completely independent of any known plant and represents a fascinatingexample of convergent evolution. Whether the insect genes involved in CNglc synthesis areclustered on the insect genome, as has been found in cyanogenic plants (see Figure 4), has notbeen analyzed. Tortoise beetles (Paropsis atomaria; Coleoptera) are also cyanogenic, excreting aliquid that contains HCN when threatened (135). They also appear to be capable of de novosynthesis when feeding on Eucalyptus hosts that are noncyanogenic (135).
Pathogen Resistance
HCN can inhibit hyphal growth of some fungi (153) and has been reported to protect against fun-gal attack, e.g., against the rice blast fungus (181). Likewise, release of the oxime intermediate infungus-infected plants as a result of a concomitant reactive oxygen burst may promote resistance(132, 133). However, in general, CNglcs are not noted for being particularly effective againstfungal pathogens; fungi may be able to convert HCN into formamide through the action of theenzyme cyanide hydratase, providing a ready source of reduced nitrogen (21), or may be able tocarry out cyanide-resistant respiration [e.g., Stemphylium loti (153)]. P. lunatus and H. brasiliensisplants with high HCNp are more sensitive to the pathogens Colletotrichum lindemuthianum and
www.annualreviews.org • Cyanogenic Glycosides 167
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.
PP65CH06-Gleadow ARI 8 April 2014 21:52
aLarva
bAdult
55% 45% 70%
30%
Linamarin
Lotaustralin
Defense droplets
Figure 7Larvae of the Burnet moth (Zygaena filipendulae) sequester the cyanogenic glycosides (CNglcs) linamarin andlotaustralin from Lotus corniculatus for use in their own defense against predators (203, 205, 207). The mothis also able to synthesize both compounds de novo when the supply from its cyanogenic food plant isinsufficient. The ratio of the two CNglcs depends on the stage in the life cycle and is largely independent ofthe ratio in the source plants. (a) A Z. filipendulae larva showing CNglc-containing droplets released whenthreatened. (b) An adult Z. filipendulae moth. Pie charts show ratios of CNglcs in Z. filipendulae at each pointin its life cycle. The areas of the pie charts indicate the relative total cyanogenic glucoside content in larvaeand adult insects.
Microcyclus ulei, respectively, compared with plants with a low HCNp (10, 98). The HCN re-leased may inhibit the synthesis of other defense compounds, like scopoletin or scopolin (119)or polyphenol oxidases (10). In each case, the release of HCN benefits the pathogen because itenables the pathogen to circumvent the host’s normal pathogen-defense response. There is, there-fore, selection within populations of H. brasiliensis and P. lunatus for both high- and low-HCNpplants, with the prevalence of different phenotypes dependent on environmental conditions andthe relative likelihood of herbivory or fungal infection (16).
Wounding and Induction
CNglcs are classified as anticipins (137) and as such are considered a constitutive defense sys-tem not induced by wounding or attack by herbivores or pathogens (69, 146), although mixingCNglcs with β-glucosidase induces cyanogenesis. Increases in HCNp observed following exten-sive wounding of sorghum leaves and stem with pins (146) may be merely a drought responseresulting from the damage to the vascular tissue rather than a response to mock herbivory (107).
168 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
Wounding induced a surge in HCN release in lima beans and Hevea by increasing β-glucosidaseand α-hydroxynitrilase activity rather than increasing HCNp (11, 98). Herbivores may elicit a dif-ferent response to mechanical damage mediated through chemical signals or enzyme activities intheir saliva, as has been found in Nicotiana attenuata (46). Many plants increase methyl jasmonateand salicylic acid levels in response to herbivory and pathogen attack (49, 168). In sorghum,microarray studies have revealed extensive transcriptional changes in response to wounding orexposure to methyl jasmonate and salicylic acid (47, 164); these changes include induction ofdhurrinase and α-hydroxynitrilase transcripts and temporary induction of CYP71E1, one of thegenes governing dhurrin biosynthesis (208), similar to the enzyme activity change in lima beansand Hevea described above. Rather than being a constitutively expressed defense system, CNglcbiosynthesis and cyanogenesis obviously respond to environmental and developmental changes.
ENVIRONMENTAL PLASTICITY AND OPTIMAL GROWTH
Effects of Nitrogen Supply
In general, plants supplied with high levels of nitrogenous fertilizers (ammonia or nitrate) have anincreased content of CNglcs (Figure 8, Table 1). Highly fertilized fields of forage sorghum, forexample, can sometimes become toxic to livestock (HCNp > 600 ppm) (200). At some stages ofplant ontogeny, like the seedling stage, the HCNp is likely transcriptionally regulated (32), whereasat other stages regulation may be controlled at multiple levels. A link between nitrogen supply andCNglc deployment has also been observed in legumes, where the rate of colonization by nitrogen-fixing rhizobia has been associated with higher concentrations of linamarin and lotaustralin anddecreased herbivory in both clover (106) and lima beans (14). Not all plants respond to nitrogenin this way. In a study by Busk & Møller (32) (see above), dhurrin concentration did not increasein very young seedlings grown at high levels of potassium nitrate. Moreover, transcript levels
Nitrogen
Drought
CO2
Young tissue(growth)
Shade
Retardation of growth
Acceleration of maturation
Downregulation of RuBisCO
Pro
mo
tio
n o
f n
ew
gro
wth
Imp
rove
d w
ate
r u
se e
ffici
en
cy?
Extremetemperatures
SugarO
CN
R2
R1
CNglc
Figure 8Scheme illustrating environmental plasticity in cyanogenic glycosides (CNglcs). CNglcs are typically higherin plants where growth is limited by resources other than nitrogen. Conditions associated with an increase inCNglcs are shown as arrows, those associated with a decrease in CNglcs are shown as lines with circles onthe end, and those where the effect is uncertain are indicated with a dashed line. Interplays between thedifferent parameters, as they are thought to affect CNglcs, are also indicated (Table 1).
www.annualreviews.org • Cyanogenic Glycosides 169
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.
PP65CH06-Gleadow ARI 8 April 2014 21:52
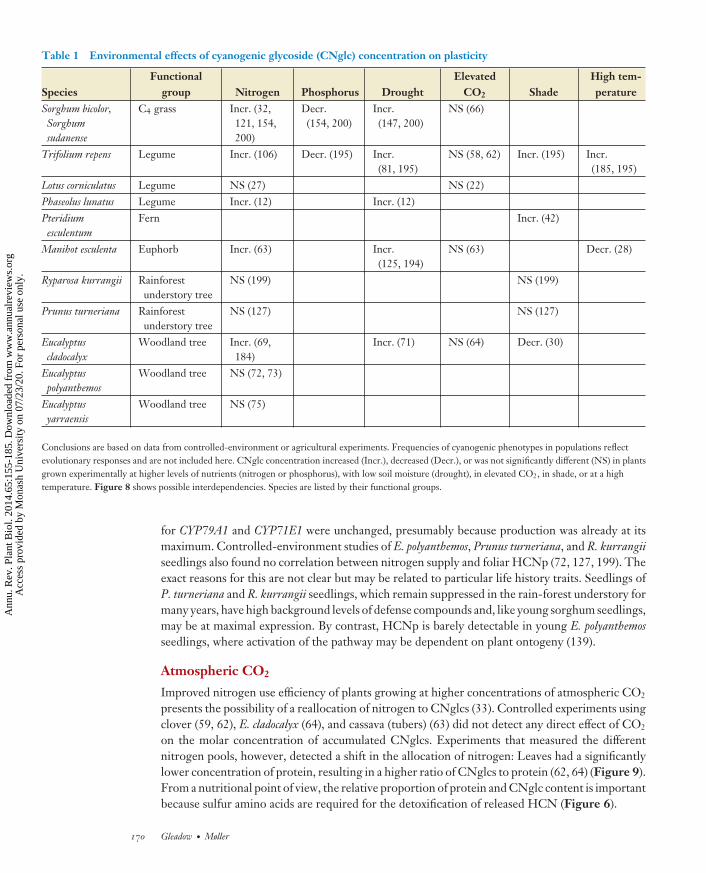
Table 1 Environmental effects of cyanogenic glycoside (CNglc) concentration on plasticity
SpeciesFunctional
group Nitrogen Phosphorus DroughtElevated
CO2 ShadeHigh tem-perature
Sorghum bicolor,Sorghumsudanense
C4 grass Incr. (32,121, 154,200)
Decr.(154, 200)
Incr.(147, 200)
NS (66)
Trifolium repens Legume Incr. (106) Decr. (195) Incr.(81, 195)
NS (58, 62) Incr. (195) Incr.(185, 195)
Lotus corniculatus Legume NS (27) NS (22)Phaseolus lunatus Legume Incr. (12) Incr. (12)Pteridiumesculentum
Fern Incr. (42)
Manihot esculenta Euphorb Incr. (63) Incr.(125, 194)
NS (63) Decr. (28)
Ryparosa kurrangii Rainforestunderstory tree
NS (199) NS (199)
Prunus turneriana Rainforestunderstory tree
NS (127) NS (127)
Eucalyptuscladocalyx
Woodland tree Incr. (69,184)
Incr. (71) NS (64) Decr. (30)
Eucalyptuspolyanthemos
Woodland tree NS (72, 73)
Eucalyptusyarraensis
Woodland tree NS (75)
Conclusions are based on data from controlled-environment or agricultural experiments. Frequencies of cyanogenic phenotypes in populations reflectevolutionary responses and are not included here. CNglc concentration increased (Incr.), decreased (Decr.), or was not significantly different (NS) in plantsgrown experimentally at higher levels of nutrients (nitrogen or phosphorus), with low soil moisture (drought), in elevated CO2, in shade, or at a hightemperature. Figure 8 shows possible interdependencies. Species are listed by their functional groups.
for CYP79A1 and CYP71E1 were unchanged, presumably because production was already at itsmaximum. Controlled-environment studies of E. polyanthemos, Prunus turneriana, and R. kurrangiiseedlings also found no correlation between nitrogen supply and foliar HCNp (72, 127, 199). Theexact reasons for this are not clear but may be related to particular life history traits. Seedlings ofP. turneriana and R. kurrangii seedlings, which remain suppressed in the rain-forest understory formany years, have high background levels of defense compounds and, like young sorghum seedlings,may be at maximal expression. By contrast, HCNp is barely detectable in young E. polyanthemosseedlings, where activation of the pathway may be dependent on plant ontogeny (139).
Atmospheric CO2
Improved nitrogen use efficiency of plants growing at higher concentrations of atmospheric CO2
presents the possibility of a reallocation of nitrogen to CNglcs (33). Controlled experiments usingclover (59, 62), E. cladocalyx (64), and cassava (tubers) (63) did not detect any direct effect of CO2
on the molar concentration of accumulated CNglcs. Experiments that measured the differentnitrogen pools, however, detected a shift in the allocation of nitrogen: Leaves had a significantlylower concentration of protein, resulting in a higher ratio of CNglcs to protein (62, 64) (Figure 9).From a nutritional point of view, the relative proportion of protein and CNglc content is importantbecause sulfur amino acids are required for the detoxification of released HCN (Figure 6).
170 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
Ambient CO2
(380 ppm)
a b
Twice ambient CO2
(780 ppm)
Prunasin
RuBisCO
Other protein
Other nitrogen sinks(nucleic acids, etc.)
Figure 9Allocation of nitrogen to prunasin [measured as cyanide potential (HCNp)], RuBisCO, total other protein,and other nitrogen sinks (such as nucleic acids and thylakoid membranes) in the leaves of Eucalyptus cladocalyxseedlings grown at either (a) ambient or (b) approximately twice the ambient concentrations of atmosphericCO2. The area of each pie is proportional to the total nitrogen concentration of the leaves in each treatment(64).
Nonoptimal or Stressful Growth Conditions
CNglc concentrations are generally higher when growth is limited by environmental factors suchas light, temperature, or drought (Figure 8, Table 1). Three explanations are often presentedto account for this: (a) CNglcs are concentrated in a smaller amount of plant tissue (178), (b) theplants are phenologically younger owing to delayed growth (126), or (c) there is active upregulationat the transcriptional level (32, 47, 164, 208). Active upregulation could be either an adaptation toprotect existing plant tissue from herbivores or a mechanism for moderating oxidative stress (133,178). Transcriptomic analysis of sorghum seedlings experiencing osmotic stress found more than1,000 genes with altered expression, including many involved in defense-related pathways (29,47). Abscisic acid, an important signal molecule in drought responses and regulation of stomatalaperture, also covaries with HCNp (5, 18). Evidence for a direct effect of stress on CNglc synthesiscomes from the study of cell cultures. Osmotically stressed suspension cultures of Eschscholtziacalifornica showed higher production rates of dhurrin and triglochinin (85), which suggests thatthe synthesis of these compounds was induced by the osmotic stress.
The magnitude of the increase in HCNp in response to low soil moisture depends on theseverity and duration of the stress, the ontogenic stage, and the availability of other resources (71,147, 194). In E. cladocalyx, the HCNp of extant leaves increased by 70% after a prolonged droughtwhen the nitrogen supply was high but only by 30% when the nitrogen supply was limited (70).Similar responses in cyanogenic crop plants may have important ramifications for human health(see sidebar, Cyanide and Cassava). In cassava, drought-stressed tubers may become more toxicbecause of a direct increase in concentration (194) and relocation of linamarin from leaves totubers (103). This increased HCNp in drought-stressed cassava is not permanent and decreasesafter plants are rewatered (194). A similar situation exists in forage sorghum, where a severe orextended drought period may result in toxic levels of dhurrin and necessitate long-term silage toenable use of the sorghum plants as fodder (25, 147).
CNglcs may also provide additional plasticity to plant development by controlling seed ger-mination in response to rainfall events. Eremophila maculata grows in arid regions, and the fruitwalls of its seeds contain germination inhibitors in the form of water-soluble glycosides, including
www.annualreviews.org • Cyanogenic Glycosides 171
Erratum
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
CYANIDE AND CASSAVA
A single dose of cyanide of ∼1–3 mg per kilogram of body weight is lethal for most vertebrates (148). Cassavahas a high HCNp, and it is the only staple food that is potentially lethal if not adequately processed (124). TheHCNp of cassava flour bought at markets in Mozambique in a year suffering from drought was greater than200 ppm (50), well above the World Health Organization’s recommended maximum level of 10 ppm. Under suchspecial circumstances, where water required for proper processing is limited and the population is restricted toa monotonous cassava diet high in cyanohydrins or cyanide, ingestion of the toxic constituents may cause acutecyanide poisoning or development of konzo, a neurological disease causing permanent paralysis of the lower limbs(124). The concentration of the CNglcs linamarin and lotaustralin increases in cassava tubers when the plant suffersfrom severe drought (124). Elite cassava lines that yield well in different soil types and when subjected to differentenvironmental challenges concomitant with having a low HCNp are in high demand (95).
the CNglc eucalyptosin A. Heavy rainfall causes the CNglc to leach out of the fruit wall andthereby trigger germination under optimal growth conditions (162). In rosaceous plants (e.g.,almonds), cold acclimation and bud break and flower development may be controlled by cyanohy-drin or HCN release resulting from endogenous hydrolysis of the CNglcs present (20). The samemechanisms may explain why spraying with cyanamide (which causes HCN liberation) can induceand synchronize bud break and flowering in some rosaceous plants.
Only a few studies have considered the effect of temperature on HCNp. In T. repens, HCNpis typically higher in plants growing at nonoptimal temperatures, e.g., below 15◦C or above 25◦C(185, 195).
Foliar HCNp has been found experimentally to be higher, lower, or unaffected in plants grownin the shade (e.g., in clover, E. cladocalyx, and P. turneriana, respectively) (30, 81, 127). The differentresponses appear to reflect the environments to which the species are adapted. In E. cladocalyx,which is adapted to open woodlands, nitrogen was reallocated from prunasin production to thephotosynthetic apparatus in shade-grown plants, likely to maximize light interception and increaseCO2 fixation (30). In contrast, P. turneriana seedlings, adapted to long periods under a rain-forestcanopy, reallocated CNglcs away from growing leaf tips to the existing leaves, presumably toprotect existing productive leaves from herbivory (127).
RESOURCE ALLOCATION: PURPOSES, COSTS, AND REGULATION
Energy Costs and Trade-Offs
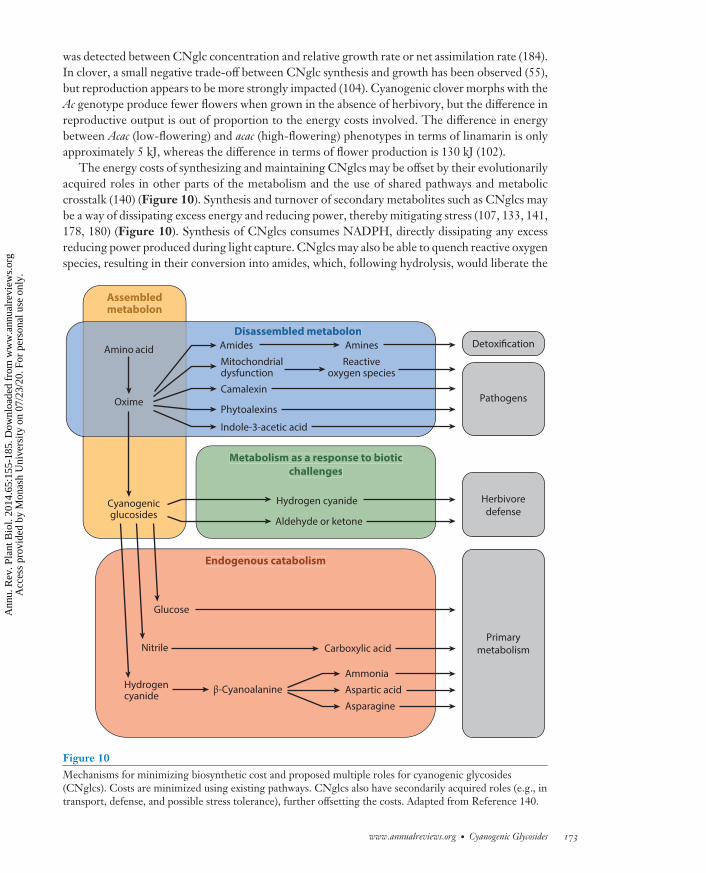
Resource allocation trade-off theories assume that there must be a cost associated with synthesizingand maintaining defensive compounds (38, 83). For CNglcs, this has been hard to demonstrate,leading some to speculate that the costs are too small to measure (107). The average energy costof dhurrin synthesis has been calculated as 2.1 g glucose (g dhurrin)−1 (60). This is low in termsof the overall energy budget, although additional costs would have to be assigned to maintenance,turnover, and transport as well as the synthesis of the β-glucosidase and α-hydroxynitrilase re-quired for rapid HCN release. The bottom line is that the cost of deploying a CNglc-based defensemust be less than the benefit gained, e.g., through reduced grazing (81) or increased metabolicplasticity. If there are costs in CNglc deployment, then it should be possible to measure a sacrificein terms of plant growth or reproductive output. Highly cyanogenic E. polyanthemos seedlingswere found to grow more slowly than seedlings with low HCNp (72). However, in a similar studyusing E. cladocalyx seedlings in which ontogenetic effects were kept to a minimum, no trade-off
172 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.
PP65CH06-Gleadow ARI 8 April 2014 21:52
was detected between CNglc concentration and relative growth rate or net assimilation rate (184).In clover, a small negative trade-off between CNglc synthesis and growth has been observed (55),but reproduction appears to be more strongly impacted (104). Cyanogenic clover morphs with theAc genotype produce fewer flowers when grown in the absence of herbivory, but the difference inreproductive output is out of proportion to the energy costs involved. The difference in energybetween Acac (low-flowering) and acac (high-flowering) phenotypes in terms of linamarin is onlyapproximately 5 kJ, whereas the difference in terms of flower production is 130 kJ (102).
The energy costs of synthesizing and maintaining CNglcs may be offset by their evolutionarilyacquired roles in other parts of the metabolism and the use of shared pathways and metaboliccrosstalk (140) (Figure 10). Synthesis and turnover of secondary metabolites such as CNglcs maybe a way of dissipating excess energy and reducing power, thereby mitigating stress (107, 133, 141,178, 180) (Figure 10). Synthesis of CNglcs consumes NADPH, directly dissipating any excessreducing power produced during light capture. CNglcs may also be able to quench reactive oxygenspecies, resulting in their conversion into amides, which, following hydrolysis, would liberate the
Detoxification
Pathogens
Herbivore
defense
Primary
metabolism
Amino acid
Cyanogenicglucosides
Amides
Reactiveoxygen species
Mitochondrialdysfunction
Phytoalexins
Indole-3-acetic acid
Hydrogen cyanide
Aldehyde or ketone
Glucose
Nitrile
Hydrogencyanide
β-Cyanoalanine
Carboxylic acid
Ammonia
Aspartic acid
Asparagine
Oxime
Camalexin
Amines
Assembledmetabolon
Disassembled metabolon
Metabolism as a response to biotic
challenges
Endogenous catabolism
Figure 10Mechanisms for minimizing biosynthetic cost and proposed multiple roles for cyanogenic glycosides(CNglcs). Costs are minimized using existing pathways. CNglcs also have secondarily acquired roles (e.g., intransport, defense, and possible stress tolerance), further offsetting the costs. Adapted from Reference 140.
www.annualreviews.org • Cyanogenic Glycosides 173
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
nitrogen in the form of easily accessible ammonium (180). The proposed alternative turnoverpathway (157) (Figure 3) also results in amide formation. CNglcs may also react chemically withH2O2, opening up the possibility that the pathway helps the plant to reduce damage caused byexcess reactive oxygen species (90, 133, 180).
CNglcs are only one of many defenses at a plant’s disposal. Defense strategies are likely tovary with different selective pressures (magnitude and type) and with developmental stage (2, 17).Attempts to understand the complete defense syndrome are an increasingly important area forresearch (2). Trade-offs between strategies are likely, e.g., between CNglcs and tannins (27, 74) orvolatile organic compounds (13). The higher HCNp in younger plants and plant parts is consistentwith the optimal allocation theory of plant defense (123), but as leaves expand, there may simplybe a trade-off with leaf toughness and other forms of chemical defense (198). New techniquesunder development—such as in situ polymerase chain reaction (95, 166), Raman spectroscopy(192), Fourier transform infrared spectroscopy (82), and bioimaging using mass spectrometry(117)—will likely help pinpoint localization and answer these questions.
Nitrogen as the Currency for Calculating Costs
Given that nitrogen is the limiting plant nutrient in most terrestrial systems, it may be moremeaningful to calculate costs in terms of nitrogen. An analysis of E. cladocalyx trees sampledacross a discontinuous natural population indicated that, on average, for every nitrogen moleculeeffectively incorporated into the leaves as prunasin, additional nitrogen was added on top of thisinvestment, possibly to increase the capacity to generate additional energy by photosynthesis (68,184). The roles of CNglcs in nitrogen storage and in defense are not necessarily mutually exclusive.The reduced rate of photosynthesis in stressed plants makes it economically attractive for plantsto store nitrogen in a reduced form, ready to be remobilized when conditions improve, rather thanas photosynthetic proteins or nitrate (147). In H. brasiliensis, the carbon and nitrogen resources inCNglcs can, for example, be mobilized and used for growth and latex production (112).
Some species appear to have become obligate cyanotypes that are dependent on CNglcs fornormal development at specific stages of ontogenesis. Cassava plantlets engineered using RNA-interference technology to have a highly reduced CNglc content exhibit poor growth or lethality atthe time point where the wild-type plant would start to produce CNglcs and have a higher require-ment for nitrogen supply (95). In a similar fashion, acyanogenic tcd1 seedlings of sorghum, in whichthe CYP79A1 protein harbors a single amino acid change that inactivates the enzyme, exhibit aslower initial growth rate than their wild-type siblings (25). In several species, studies have docu-mented the ability of cyanogenic plants to remobilize the nitrogen bound to CNglcs into ammoniaand its subsequent use in primary metabolism in the synthesis in amino acids without the releaseof gaseous HCN (37, 118, 125, 176) (see Figure 3). The ability of plants to remobilize the sugarand nitrogen stored in CNglcs, combined with the ability to transport CNglcs to specific partsof the plants guided by the generation of differently glycosylated transport forms, illustrates howturnover of CNglcs may play an important role in balancing primary metabolism, especially duringmajor changes in plant ontogeny, and thereby improve plant plasticity (e.g., 133, 159, 176, 178).
SUMMARY POINTS
1. Cyanogenic glycosides (CNglcs) are glycosides of cyanohydrins that release toxichydrogen cyanide (HCN) and ketones when hydrolyzed by β-glycosidases and α-hydroxynitrilases in a process referred to as cyanogenesis.
174 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
2. The biosynthesis of CNglcs and the process of cyanogenesis are now well understood inseveral species, and the relevant genes have been identified.
3. CNglcs are a structurally much more diverse class of bioactive products than originallythought, encompassing additional and different sugars than glucose that are linked bymultiple linkage types and further decorated by acylation reactions.
4. Cyanogenesis is an effective defense against generalist herbivores but is not particularlyeffective against fungal pathogens. Many fungi efficiently convert HCN into ammoniaand carbon dioxide. Some insect specialists have evolved mechanisms to sequester or denovo synthesize CNglcs and use them as their own defense against predators and as asource of reduced carbon and nitrogen.
5. Cyanogenesis follows classical Mendelian inheritance patterns, but its quantitative ex-pression at a particular point in space and time is highly plastic.
6. The genes encoding the enzymes involved in CNglc biosynthesis are clustered to promotestable inheritance of the defense pathway in dynamic ecosystems.
7. CNglc concentration is generally higher in young plants, where growth is constrained,or where nitrogen is in ample supply.
8. Factors affecting CNglc concentration can be explained in terms of a resource-basedtrade-off between plant growth and defense. The difficulty in calculating such costs mayarise because the production costs are actually low and because CNglcs have secondarilyacquired important roles in nitrogen transport and storage and offer improved toleranceto oxidative stress, offsetting the direct costs of production.
9. CNglcs are defense compounds that through evolution have gained additional functionsin plant primary metabolism manifested in the course of plant ontogeny. Many plantspecies are polymorphic with respect to cyanogenesis, demonstrating that CNglcs arenot essential plant constituents but rather constituents that under specific environmentalconditions and ontogenetic stages improve phenotypic plasticity by increasing the abil-ity of plants to adapt to environmental challenges. Given the high proportion of foodcrops that are cyanogenic, it is important to be able to predict the response to changingenvironments in order to maintain and improve food safety.
FUTURE ISSUES
1. Most of our knowledge of CNglcs has been derived from studying a limited number ofplant species. Recurrent evolution of the biosynthetic pathway has recently been demon-strated. In a similar manner, the routes for endogenous turnover may be highly speciesdependent. Likewise, the functional roles that CNglcs may play in a particular plantspecies to improve plant plasticity toward a particular environmental challenge and inthe course of plant ontogeny may vary substantially. This also applies to the ability ofCNglcs to ward off herbivores and pests and thus improve plant defense. Even at themost basic level, we need to take into account whether knowledge of the roles of CNglcsin one plant species can be generalized or whether the observed effects are unique.
www.annualreviews.org • Cyanogenic Glycosides 175
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
2. Many basic questions remain about the interaction of CNglcs with primary metabolismand their ability to facilitate plant plasticity in a variety of settings, including in responseto environmental challenges. New roles for CNglcs are being proposed and need furtherscrutiny. Knowledge of the molecular mechanisms—e.g., with respect to increased stresstolerance—may be used to improve other crop plants through classical or molecularbreeding. Given that so many food crops are cyanogenic, understanding the effects ofclimate change on CNglc accumulation and turnover is important for human food safety.Likewise, climate change–induced alterations in CNglc content may severely affect thefeeding behavior of herbivores and pests, changes that could be either beneficial orharmful (or neither) with respect to humans and the global environment.
3. Experimental data in the literature are frequently confounded by developmental and sizedifferences. There is a need for additional experimentation that focuses on ontogenet-ically relevant comparisons and takes into account the differences in abiotic and bioticchallenges that plants are exposed to in the course of their life cycles (126). Experimentsin which the biological variation among plants is minimal may provide helpful guidelineson how to design large-scale field experiments (126).
4. The molecular regulation of CNglc production and turnover is poorly understood. Com-parisons of the transcriptomes of acyanogenic and cyanogenic morphs, coupled with themetabolomics of the different nitrogen pools, would allow investigation of the physio-logical changes that accompany phenotypic plasticity.
5. The consequences for plant growth and agroecology of bioengineering cyanogenesisinto and out of crop plants—as an alternative to the use of pesticides and to reduce theirtoxicity, respectively—need to be carefully evaluated. Cassava is propagated by cuttings,so the negative effects of removing CNglcs at the seedling stage may not have any practicaleffects in cassava cultivation, but careful investigations to identify negative effects in thecourse of plant ontogeny need to be strengthened and encouraged.
6. Several studies have indicated that there may be a trade-off between cyanogenesis andother types of defense. The emergence of new model species with mutations in differentparts of the biosynthetic and catabolic pathways as well as mutant collections in cropplants presents unique opportunities to discover how the potentially conflicting demandsof responding to abiotic stress, herbivory, and attack by pathogens can be reconciled.
7. Use of crops for green manure as a substitute for chemical fertilizers and pesticides isan important approach for more sustainable agricultural practices. Green manure fromcyanogenic crops like white clover and sorghum is rich in nitrogen and releases HCNthat may be utilized for pest control. The risk of leaching HCN into the environmentfollowing application to sandy and loamy soils needs to be investigated to avoid unin-tended threats to the quality of groundwater and surface waters (23). This aspect needsconsideration in assessments of the risk associated with use of crops as green manure toreplace chemical fertilizers and pesticides as well as in genetic engineering approachesto design crops with improved pest resistance.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
176 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
ACKNOWLEDGMENTS
B.L.M. acknowledges financial support from the Villum Foundation to the VILLUM ResearchCenter for Plant Plasticity, from the Novo Nordisk Foundation to the Plant Pathway DiscoverySection at the Center for Biosustainability, and from the University of Copenhagen UCPH Excel-lence Programme for Interdisciplinary Research to Center for Synthetic Biology “bioSYNergy.”We gratefully acknowledge the support of the Australian Research Council (DP130101049 toB.L.M. and R.M.G.). We also thank Samantha Fromhold and Mohammed Saddik Motawia forhelp with figure preparation.
LITERATURE CITED
1. Adewusi SRA. 1990. Turnover of dhurrin in green sorghum seedlings. Plant Physiol. 94:1219–242. Agrawal AA. 2011. New synthesis—trade-offs in chemical ecology. J. Chem. Ecol. 37:230–313. Agrawal AA, Hastings AP, Johnson MTJ, Maron JL, Salminen J-P. 2012. Insect herbivores drive real-
time ecological and evolutionary change in plant populations. Science 338:113–164. Ahn YO, Saino H, Mizutani M, Shimizu B, Sakata K. 2007. Vicianin hydrolase is a novel cyanogenic
β-glycosidase specific to β-vicianoside (6-O-α-L-arabinopyranosyl-β-D-glucopyranoside) in seeds ofVicia angustifolia. Plant Cell Physiol. 48:938–39
5. Alves AAC, Setter TL. 2004. Abscisic acid accumulation and osmotic adjustment in cassava under waterdeficit. Environ. Exp. Bot. 51:259–71
6. Andersen MD, Busk PK, Svendsen I, Møller BL. 2000. Cytochromes P450 from cassava (Manihot es-culenta Crantz) catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin andlotaustralin: cloning, functional expression in Pichia pastoris, and substrate specificity of the isolatedrecombinant enzymes. J. Biol. Chem. 275:1966–75
7. Bak S, Beisson F, Bishop G, Hamberger B, Hofer R, et al. 2011. Cytochromes P450. Arabidopsis Book9:e0144
8. Bak S, Kahn RA, Nielsen HN, Møller BL, Halkier BA. 1998. Cloning of three A-type cytochromes P450,CYP71E1, CYP98, and CYP99 from Sorghum bicolor (L.) Moench by a PCR approach and identificationby expression in Escherichia coli of CYP71E1 as a multifunctional cytochrome P450 in the biosynthesisof the cyanogenic glucoside dhurrin. Plant Mol. Biol. 36:393–405
9. Bak S, Paquette S, Morant M, Morant A, Saito S, et al. 2006. Cyanogenic glycosides: a case study forevolution and application of cytochromes P450. Phytochem. Rev. 5:309–29
10. Ballhorn DJ. 2011. Constraints of simultaneous resistance to a fungal pathogen and an insect herbivorein lima bean (Phaseolus lunatus L.). J. Chem. Ecol. 37:141–44
11. Ballhorn DJ, Heil M, Lieberei R. 2006. Phenotypic plasticity of cyanogenesis in lima bean Phaseoluslunatus—activity and activation of β-glucosidase. J. Chem. Ecol. 32:261–75
12. Ballhorn DJ, Kautz S, Jensen M, Schmitt I, Heil M, Hegeman AD. 2011. Genetic and environmentalinteractions determine plant defences against herbivores. J. Ecol. 99:313–26
13. Ballhorn DJ, Kautz S, Lion U, Heil M. 2008. Trade-offs between direct and indirect defences of limabean (Phaseolus lunatus). J. Ecol. 96:971–80
14. Ballhorn DJ, Kautz S, Schadler M. 2013. Induced plant defense via volatile production is dependent onrhizobial symbiosis. Oecologia 172:833–46
15. Ballhorn DJ, Lieberei R, Ganzhorn JU. 2005. Plant cyanogenesis of Phaseolus lunatus and its relevancefor herbivore-plant interaction: the importance of quantitative data. J. Chem. Ecol. 31:1445–73 16. Shows that the
trade-off between theproduction of CNglcsand volatile organiccompounds results fromthe balance betweenrelative rates ofherbivory and pathogenattack.
16. Ballhorn DJ, Pietrowski A, Lieberei R. 2010. Direct trade-off between cyanogenesis and resis-tance to a fungal pathogen in lima bean (Phaseolus lunatus L.). J. Chem. Ecol. 98:226–36
17. Ballhorn DJ, Schiwy S, Jensen M, Heil M. 2008. Quantitative variability of direct chemical defense inprimary and secondary leaves of lima bean (Phaseolus lunatus) and consequences for a natural herbivore.J. Chem. Ecol. 34:1298–301
18. Bari R, Jones JDG. 2009. Role of plant hormones in plant defence responses. Plant Mol. Biol. 69:473–8819. Barnett RD, Caviness CE. 1968. Inheritance of hydrocyanic acid production in two sorghum × sudan-
grass crosses. Crop Sci. 8:89–91
www.annualreviews.org • Cyanogenic Glycosides 177
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
20. Barros PM, Goncalves N, Saibo NJM, Oliveira MM. 2012. Cold acclimation and floral development inalmond bud break: insights into the regulatory pathways. J. Exp. Bot. 63:4585–96
21. Basile LJ, Willson RC, Sewell BT. 2008. Genome mining of cyanide-degrading nitrilases from filamen-tous fungi. Appl. Microbiol. Biotechnol. 80:427–35
22. Bazin A, Goverde M, Erhardt A, Shykoff JA. 2002. Influence of atmospheric carbon dioxide enrichmenton induced response and growth compensation after herbivore damage in Lotus corniculatus. Ecol. Entomol.27:271–78
23. Bjarnholt N, Laegdsmand M, Hansen HC, Jacobsen OH, Møller BL. 2008. Leaching of cyanogenicglucosides and cyanide from white clover green manure. Chemosphere 72:897–904
24. Bjarnholt N, Møller BL. 2008. Hydroxynitrile glucosides. Phytochemistry 69:1947–6125. Uses ethylmethanesulfonate–generated sorghummutants with alteredcyanogenic status toprovide a new cropmodel for study.
25. Blomstedt CK, Gleadow RM, O’Donnell N, Naur P, Jensen K, et al. 2012. A combined biochem-ical screen and TILLING approach identifies mutations in Sorghum bicolor L. Moench resultingin acyanogenic forage production. Plant Biotechnol. J. 10:54–66
26. Blumenthal SG, Hendrickson HR, Abrol YP, Conn EE. 1968. Cyanide metabolism in higher plants. III.The biosynthesis of β-cyanoalanine. J. Biol. Chem. 243:5302–7
27. Briggs MA. 1990. Chemical defense production in Lotus corniculatus L. I. The effects of nitrogen sourceon growth, reproduction and defense. Oecologia 83:27–31
28. Brown A. 2011. Effect of temperature and drought on the growth and nutritive value of cassava. Hon. Thesis,Monash Univ., Victoria, Aust.
29. Buchanan CD, Lim S, Salzman RA, Kagiampakis I, Morishige DT, et al. 2010. Sorghum bicolor’s tran-scriptome response to dehydration, high salinity and ABA. Plant Mol. Biol. 58:699–720
30. Burns AE, Gleadow RM, Woodrow IE. 2002. Light alters the allocation of nitrogen to cyanogenicglycosides in Eucalyptus cladocalyx. Oecologia 133:288–94
31. Burns AE, Gleadow RM, Zacarias A, Cuambe CE, Miller RE, Cavagnaro TR. 2012. Variations in thechemical composition of cassava (Manihot esculenta Crantz) leaves and roots as affected by genotypic andenvironmental variation. J. Agric. Food Chem. 60:4946–56
32. Busk PK, Møller BL. 2002. Dhurrin synthesis in sorghum is regulated at the transcriptional level andinduced by nitrogen fertilization in older plants. Plant Physiol. 129:1222–31
33. Cavagnaro TR, Gleadow RM, Miller RE. 2011. Plant nutrient acquisition and utilisation in a high carbondioxide world. Funct. Plant Biol. 38:87–96
34. Chassagne D, Crouzet JC, Bayonove CL, Baumes RL. 1996. Identification and quantification of passionfruit cyanogenic glycosides. J. Agric. Food Chem. 44:3817–20
35. Cicek M, Esen A. 1998. Structure and expression of a dhurrinase (β-glucosidase) from Sorghum. PlantPhysiol. 116:1469–78
36. Clausen V, Frydenvang K, Koopmann R, Jørgensen LB, Abbiw DK, et al. 2002. Plant analysis bybutterflies: occurrence of cyclopentenylglycines in Passifloraceae, Flacourtiaceae, and Turneraceae anddiscovery of the novel nonproteinogenic amino acid 2-(3′-cyclopentenyl)glycine in Rinorea. J. Nat. Prod.65:542–47
37. Clegg DO, Conn EE, Janzen DH. 1979. Developmental fate of the cyanogenic glycoside linamarin inCosta Rican wild lima bean seeds. Nature 278:343–44
38. Coley PD, Bryant JP, Chapin FS III. 1985. Resource availability and plant antiherbivore defense. Science230:895–99
39. Conn EE. 1980. Cyanogenic compounds. Annu. Rev. Plant Physiol. 31:433–5140. Conn EE. 1991. The metabolism of a natural product: lessons learned from cyanogenic glycosides. Planta
Med. 57:1–941. Conn EE, Seigler DS, Maslin BR, Dunn J. 1989. Cyanogenesis in Acacia subgenus Aculezferum. Phyto-
chemistry 28:817–2042. Cooper-Driver G, Finch S, Swain T. 1977. Seasonal variation in secondary plant compounds in relation
to the palatability of Pteridium aqulinum. Biochem. Syst. Ecol. 5:177–8343. Corkhill L. 1942. Cyanogenesis in white clover (Trifolium repens L.) V. The inheritance of cyanogenesis.
N.Z. J. Sci. Technol. B 23:178–9344. Cutler AJ, Conn EE. 1981. The biosynthesis of cyanogenic glucosides in Linum usitatissimum (linen flax)
in vitro. Arch. Biochem. Biophys. 212:468–74
178 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
45. Dahler JM, McConchie C, Turnbull CGN. 1995. Quantification of cyanogenic glycosides in seedlingsof three Macadamia (Proteaceae) species. Aust. J. Bot. 43:619–28
46. Dinh ST, Baldwin IT, Galis I. 2013. The HERBIVORE ELICITOR-REGULATED1 gene enhancesabscisic acid levels and defenses against herbivores in Nicotiana attenuata plants. Plant Physiol. 162:2106–24
47. Dugas DV, Monaco MK, Olsen A, Klein RR, Kumari S, et al. 2011. Functional annotation of thetranscriptome of Sorghum bicolor in response to osmotic stress and abscisic acid. BMC Genomics 12:514
48. Elias M, Nambisan B, Sudhakaran PR. 1997. Characterization of linamarase of latex and its localizationin petioles in cassava. Arch. Biochem. Biophys. 341:222–28
49. Erb M, Meldau S, Howe GA. 2012. Role of phytohormones in insect-specific plant reactions. TrendsPlant Sci. 17:250–59
50. Ernesto M, Cardoso AP, Nicala D, Mirione E, Massaza F, et al. 2002. Persistent konzo and cyanogentoxicity from cassava in northern Mozambique. Acta Trop. 22:357–62
51. Finnie JW, Windsor PA, Kessell AE. 2011. Neurological diseases of ruminant livestock in Australia. II.Toxic disorders and nutritional deficiencies. Aust. Vet. J. 89:247–53
52. Flamatti GR, Waters MT, Scaffidi A, Merritt DJ, Ghisalberti EL, et al. 2013. Karrikin and cyanohydrinsmoke signals provide clues to new endogenous plant signaling compounds. Mol. Plant 6:29–37
53. Forslund K, Morant M, Jørgensen B, Olsen CE, Asamizu E, et al. 2004. Biosynthesis of the nitrileglucosides rhodiocyanoside A and D and the cyanogenic glucosides lotaustralin and linamarin in Lotusjaponicus. Plant Physiol. 135:71–84
54. Foulds W. 1982. Polymorphism for cyanogenesis in Lotus australis Andr. populations at GreenoughFront Flats, Western Australia. Aust. J. Bot. 30:211–17
55. Foulds W, Grime JP. 1972. The response of cyanogenc and acyanogenic phenotypes of Trifolium repensto soil moisture supply. Heredity 28:181–87
56. Franks TK, Powell KS, Choimes S, Marsh E, Iocco P, et al. 2006. Consequences of transferring threesorghum genes for secondary metabolite (cyanogenic glucoside) biosynthesis to grapevine hairy roots.Transgenic Res. 15:181–95
57. Franks TK, Yadollahi A, Wirthensohn MG. 2008. A seed coat cyanohydrin glucosyltransferase is asso-ciated with bitterness in almond (Prunus dulcis) kernels. Funct. Plant Biol. 35:236–46
58. Frehner M, Conn EE. 1987. The linamarin β-glucosidase in Costa Rica wild bean (Phaseolus lunatus L.)is apoplastic. Plant Physiol. 84:1296–300
59. Frehner M, Luscher A, Hebeisen T, Zanetti S, Schubiger F, Scalet M. 1997. Effects of elevated partialpressure of carbon dioxide and season of the year on forage quality and cyanide concentration of Trifoliumrepens L. from a FACE experiment. Acta Oecologica 18:297–304
60. Gershenzon J. 1989. The cost of plant chemical defense against herbivory: a biochemical perspective. InInsect–Plant Interactions, ed. EA Bernays, pp. 105–73. Bacon Raton, FL: CRC
61. Gierl A, Frey M. 2001. Evolution of benzoxazinone biosynthesis and indole production in maize. Planta213:493–98
62. Gleadow RM, Edwards E, Evans JR. 2009. Changes in nutritional value of cyanogenic Trifolium repensat elevated CO2. J. Chem. Ecol. 35:476–47
63. Gleadow RM, Evans JR, McCaffery S, Cavagnaro TR. 2009. Growth and nutritive value of cassava(Manihot esculenta Cranz.) are reduced when grown in elevated CO2. Plant Biol. 11:76–82
64. Gleadow RM, Foley WJ, Woodrow IE. 1998. Enhanced CO2 alters the relationship between photosyn-thesis and defence in cyanogenic Eucalyptus cladocalyx F. Muell. Plant Cell Environ. 21:12–22
65. Gleadow RM, Haburjak J, Dunn JE, Conn ME, Conn EE. 2008. Frequency and distribution ofcyanogenic glycosides in Eucalyptus L’Herit. Phytochemistry 69:1870–74
66. Gleadow RM, Møller BL, Cavagnaro T, Miller R, Stuart P, et al. 2013. Impacts of climate change onallocation of N to cyanogenic glycosides. Pharm. Biol. 50:623 (Abstr.)
67. Gleadow RM, Veechies AC, Woodrow IE. 2003. Cyanogenic Eucalyptus nobilis is polymorphic for bothprunasin and specific β-glucosidases. Phytochemistry 63:699–704
68. Gleadow RM, Woodrow IE. 2000. Polymorphism in cyanogenic glycoside content and cyanogenicβ-glucosidase activity in natural populations of Eucalyptus cladocalyx. Aust. J. Plant Physiol. 27:693–99
www.annualreviews.org • Cyanogenic Glycosides 179
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
70. Shows that theeffectiveness of CNglcsas a general defensedepends on HCNp, dietquality, and mode offeeding.
69. Gleadow RM, Woodrow IE. 2000. Temporal and spatial variation in cyanogenic glycosides in Eucalyptuscladocalyx. Tree Physiol. 20:591–98
70. Gleadow RM, Woodrow IE. 2002. Constraints on effectiveness of cyanogenic glycosides inherbivore defense. J. Chem. Ecol. 28:1301–13
71. Gleadow RM, Woodrow IE. 2002. Defense chemistry of cyanogenic Eucalyptus cladocalyx seedlings isaffected by water supply. Tree Physiol. 22:939–45
72. Goodger JQD, Ades PK, Woodrow IE. 2004. Cyanogenesis in Eucalyptus polyanthemos seedlings: heri-tability, ontogeny and effect of soil nitrogen. Tree Physiol. 24:681–88
73. Goodger JQD, Capon RJ, Woodrow IE. 2002. Cyanogenic polymorphism in Eucalyptus polyanthemosSchauer subsp. vestita L. Johnson and K. Hill (Myrtaceae). Biochem. Syst. Ecol. 30:617–30
74. Goodger JQD, Gleadow RM, Woodrow IE. 2006. Growth cost and ontogenetic expression patterns ofdefence in cyanogenic Eucalyptus spp. Trees 20:757–65
75. Goodger JQD, Woodrow IE. 2002. Cyanogenic polymorphism as an indicator of genetic diversity inthe rare species Eucalyptus yarraensis (Myrtaceae). Funct. Plant Biol. 29:1445–52
76. Gruhnert C, Biehl B, Selmar D. 1994. Compartmentation of cyanogenic glucosides and their degradingenzymes. Planta 195:36–42
77. Gwilt JR. 1960. Odour of potassium cyanide. Lancet 276:119778. Halkier BA, Møller BL. 1989. Biosynthesis of the cyanogenic glucoside dhurrin in seedlings of Sorghum
bicolor (L.) Moench and partial purification of the enzyme system involved. Plant Physiol. 90:1552–5979. Halkier BA, Møller BL. 1991. Involvement of cytochrome P-450 in the biosynthesis of dhurrin in
Sorghum bicolor (L.) Moench. Plant Physiol. 96:10–1780. Hansen KS, Kristensen C, Tattersall DB, Jones PR, Olsen CE, et al. 2003. The in vitro substrate
regiospecificity of recombinant UGT85B1, the cyanohydrin glucosyltransferase from Sorghum bicolor.Phytochemistry 64:143–51
81. Hayden KJ, Parker IM. 2002. Plasticity in cyanogenesis of Trifolium repens L.: inducibility, fitness costsand variable expression. Evol. Ecol. Res. 4:155–68
82. Heraud P, Caine S, Sanson G, Gleadow RM, Wood BR, McNaughton D. 2007. Focal plane arrayinfrared imaging: a new way to analyse leaf tissue. New Phytol. 173:216–25
83. Herms DA, Mattson WJ. 1992. The dilemma of plants: to grow or defend. Q. Rev. Biol. 67:283–33584. Hickel A, Hasslacher M, Griengl H. 1996. Hydroxynitrile lyases: functions and properties. Physiol. Plant.
98:891–9885. Hosel W, Berlin J, Hanzlik TN, Conn EE. 1987. In-vitro biosynthesis of 1-(4′-hydroxyphenol)-2-
nitroethane and production of cyanogenic compounds in osmotically stressed cell suspension cultures ofEschscholtzia californica Cham. Planta 166:176–81
86. Hughes J, Decarvalho JPC, Hughes MA. 1994. Purification, characterization, and cloning of α-hydroxynitrile lyase from cassava (Manihot esculenta Crantz). Arch. Biochem. Biophys. 311:496–502
87. Hughes MA. 1991. The cyanogenic polymorphism in Trifolium repens L. (white clover). Heredity 66:105–15
88. Ibenthal WD, Pourmohseni H, Grosskopf S, Oldenburg H, Shafieiazad S. 1993. New approaches towardsbiochemical mechanisms of resistance susceptibility of Gramineae to powdery mildew (Erysiphe graminis).Angew. Bot. 67:97–106
89. Itkin M, Heinig U, Tzfadia O, Bhide AJ, Shinde B, et al. 2013. Biosynthesis of antinutritional alkaloidsin solanaceous crops is mediated by clustered genes. Science 341:175–79
90. Jenrich R, Trompetter I, Bak SS, Olsen CE, Møller BL, Piotrowski M. 2007. Evolution of heteromicnitrilase complexes in Poaceae with new functions in nitrile metabolism. Proc. Natl. Acad. Sci. USA104:18848–53
91. Jensen K, Osmani SA, Hamann T, Naur P, Møller B. 2011. Homology modeling of the three membraneproteins of the dhurrin metabolon: catalytic sites, membrane surface association and protein-proteininteractions. Phytochemistry 72:2113–23
92. Jensen NB, Zagrobelny M, Hjernø K, Olsen CE, Houghton-Larsen J, et al. 2011. Convergent evolutionin biosynthesis of cyanogenic defence compounds in plants and insects. Nat. Commun. 2:273
93. Jones DA. 1998. Why are so many food plants cyanogenic? Phytochemistry 47:155–62
180 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
94. Jones PR, Møller BL, Hoj PB. 1999. The UDP-glucose:p-hydroxymandelonitrile-O-glucosyltransferasethat catalyzes the last step in synthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor: isolation,cloning, heterologous expression, and substrate specificity. J. Biol. Chem. 274:35483–91
95. Jørgensen K, Bak S, Busk PK, Sørenson C, Olsen CE, et al. 2005. Cassava plants with a depletedcyanogenic glucoside content in leaves and tubers. Distribution of cyanogenic glucosides, their site ofsynthesis and transport, and blockage of the biosynthesis by RNA interference technology. Plant Physiol.139:363–74
96. Jørgensen K, Morant AV, Morant M, Jensen NB, Olsen CE, et al. 2011. Biosynthesis of the cyanogenicglucosides linamarin and lotaustralin in cassava: isolation, biochemical characterization, and expressionpattern of CYP71E7, the oxime-metabolizing cytochrome P450 enzyme. Plant Physiol. 155:282–92
97. Jørgensen K, Rasmussen AV, Morant M, Nielsen AH, Bjarnholt N, et al. 2005. Metabolon formationand metabolic channeling in the biosynthesis of plant natural products. Curr. Opin. Plant Biol. 8:280–91
98. Shows that somefungi increase HCNc byinducing catabolicenzyme activity tosuppress naturalphytoalexin responses.
98. Kadow D, Voß K, Selmar D, Lieberei R. 2012. The cyanogenic syndrome in rubber tree Heveabrasiliensis: tissue-damage-dependent activation of linamarase and hydroxynitrile lyase acceler-ates hydrogen cyanide release. Ann. Bot. 109:1253–62
99. Kahn RA, Bak S, Svendsen I, Halkier BA, Møller BL. 1997. Isolation and reconstitution of cytochromeP450ox and in vitro reconstitution of the entire biosynthetic pathway of the cyanogenic glucoside dhurrinfrom sorghum. Plant Physiol. 115:1661–70
100. Kahn RA, Fahrendorf T, Halkier BA, Møller BL. 1999. Substrate specificity of the cytochrome P450enzymes CYP79A1 and CYP71E1 involved in the biosynthesis of the cyanogenic glucoside dhurrin inSorghum bicolor (L.) Moench. Arch. Biochem. Biophys. 363:9–18
101. Kakes P. 1985. Linamerase and other β-glucosidases are present in the cell walls of Trifolium repens L.leaves. Planta 166:156–60
102. Kakes P. 1989. An analysis of the costs and benefits of the cyanogenic system in Trifolium repens L. Theor.Appl. Genet. 77:111–18
103. Kakes P. 1994. The function of cyanogenesis in cassava. Acta Hortic. 375:79–85104. Kakes P. 1997. Difference between the male and female components of fitness associated with the gene
Ac in Trifolium repens. Acta Bot. Neerl. 46:219–23105. Kannangara R, Motawia MS, Hansen NKK, Paquette SM, Olsen CE, et al. 2011. Characterization and
expression profile of two UDP-glucosyltransferases, UGT85K4 and UGT85K5, catalyzing the last stepin cyanogenic glucoside biosynthesis in cassava. Plant J. 68:287–301
106. Kempel A, Brandl R, Schadler M. 2009. Symbiotic soil microorganisms as players in aboveground plant-herbivore interactions: the role of rhizobia. Oikos 118:634–40
107. Kerchev PL, Fenton B, Foyer CH, Hancock RD. 2012. Plant responses to insect herbivory: interactionsbetween photosynthesis, reactive oxygen species and hormone signalling pathways. Plant Cell Environ.35:441–53
108. King NLR, Bradbury JH. 1995. Bitterness of cassava: identification of a new apiosyl glucoside and othercompounds that affect its bitter taste. J. Sci. Food Agric. 68:223–30
109. Koch BM, Sibbesen O, Halkier BA, Svendsen I, Møller BL. 1995. The primary sequence of cy-tochrome P450tyr, the multifunctional N-hydroxylase catalyzing the conversion of L-tyrosine top-hydroxyphenylacetaldehyde oxime in the biosynthesis of the cyanogenic glucoside dhurrin in Sorghumbicolor (L.) Moench. Arch. Biochem. Biophys. 323:177–86
110. Koepke T, Schaeffer S, Harper A, Dicenta F, Edwards M, et al. 2013. Comparative genomics analysisin Prunoideae to identify biologically relevant polymorphisms. Plant Biotechnol. J. 11:883–93
111. Kojima M, Poulton JE, Thayer SS, Conn EE. 1979. Tissue distributions of dhurrin and of enzymesinvolved in its metabolism in leaves of Sorghum bicolor. Plant Physiol. 63:1022–28
112. Kongsawadworakul P, Viboonjun U, Romruensukharom P, Chantuma P, Ruderman S, Chrestin H.2009. The leaf, inner bark and latex cyanide potential of Hevea brasiliensis: evidence for involvement ofcyanogenic glucosides in rubber yield. Phytochemistry 70:730–39
113. Kristensen C, Morant M, Olsen CE, Ekstrom CT, Galbraith DW, et al. 2005. Metabolic engineeringof dhurrin in transgenic Arabidopsis plants with marginal inadvertent effects on the metabolome andtranscriptome. Proc. Natl. Acad. Sci. USA 102:1779–84
www.annualreviews.org • Cyanogenic Glycosides 181
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
114. Lamont BB. 1993. Injury-induced cyanogenesis in vegetative and reproductive parts of two Grevilleaspecies and their F1 hybrid. Ann. Bot. 71:537–42
115. Leavesley HB, Li L, Prabhakaran K, Borowitz JL, Ison GE. 2008. Interaction of cyanide and nitric oxidewith cytochrome c oxidase: implications for acute cyanide toxicity. Toxicol. Sci. 101:101–11
116. Lee J, Zhang G, Wood E, Castillo CR, Mitchell AE. 2013. Quantification of amygdalin in nonbitter,semibitter, and bitter almonds (Prunus dulcis) by UHPLC-(ESI)QqQ MS/MS. J. Agric. Food Chem.61:7754–59
117. Li B, Knudsen C, Hansen NK, Jørgensen K, Kannangara R, et al. 2013. Visualizing metabolite distribu-tion and enzymatic conversion in plant tissues by desorption electrospray ionization mass spectrometryimaging. Plant J. 74:1059–71
118. Lieberei R, Selmar D, Biehl B. 1985. Metabolisation of cyanogenic glucosides in Hevea brasiliensis. PlantSyst. Evol. 150:49–63
119. Lieberei R. 2007. South American leaf blight of the rubber tree (Hevea spp.): new steps in plant domes-tication using physiological features and molecular markers. Ann. Bot. 100:1125–42
120. Loyd RC, Gray E. 1970. Amount and distribution of hydrocyanic acid potential during the life cycle ofplants of three sorghum cultivars. Agron. J. 62:394–97
121. McBee GG, Miller FR. 1980. Hydrocyanid acid potential in several sorghum breeding lines as affectedby nitrogen fertilization and variable harvests. Crop Sci. 20:232–35
122. McFarlane IJ, Lees EM, Conn EE. 1975. The in vitro biosynthesis of dhurrin, the cyanogenic glycosideof Sorghum bicolor. J. Biol. Chem. 250:4708–13
123. McKey D. 1974. Adaptive patterns in alkaloid physiology. Am. Nat. 108:305–20124. McKey D, Cavagnaro TR, Cliff J, Gleadow RM. 2010. Chemical ecology in coupled human and natural
systems: people, manioc, multitrophic interactions and global change. Chemoecology 20:109–33125. McMahon JM, White WLB, Sayre RT. 1995. Cyanogenesis in cassava (Manihot esculenta Crantz).
J. Exp. Bot. 46:731–41126. Miller RE, Gleadow RM, Cavagnaro TR. 2014. Age versus stage: Does ontogeny modify the effect of
phosphorus and arbuscular mycorrhizas on above- and below-ground defence in forage sorghum? PlantCell Environ. 37:929–42
127. Miller RE, Gleadow RM, Woodrow IE. 2004. Cyanogenesis in tropical Prunus turneriana: characterisa-tion, variation and response to low light. Funct. Plant Biol. 31:491–503
128. Miller RE, Jensen R, Woodrow IE. 2006. Frequency of cyanogenesis in tropical rainforests of Far NorthQueensland, Australia. Ann. Bot. 97:1017–44
129. Miller RE, McConville MJ, Woodrow IE. 2006. Cyanogenic glycosides from the rare Australian endemicrainforest tree Clerodendrum grayi (Lamiaceae). Phytochemistry 67:43–51
130. Miller RE, Simon J, Woodrow IE. 2006. Cyanogenesis in the Australian tropical rainforest endemicBrombya platynema (Rutaceae): chemical characterisation and polymorphism. Funct. Plant Biol. 33:477–86
131. Miller RE, Tuck KL. 2013. Reports on the distribution of aromatic cyanogenic glycosides in Australiantropical rainforest tree species of the Lauraceae and Sapindaceae. Phytochemistry 92:146–52
132. Møller BL. 2010. Dynamic metabolons. Science 330:1328–29133. Møller BL. 2010. Functional diversifications of cyanogenic glucosides. Curr. Opin. Plant Biol. 13:337–46134. Møller BL, Conn EE. 1979. The biosynthesis of cyanogenic glucosides in higher plants: N-
hydroxytyrosine as an intermediate in the biosynthesis of dhurrin by Sorghum bicolor (L.) Moench.J. Biol. Chem. 254:8575–83
135. Moore BP. 1967. Hydrogen cyanide in the defensive secretions of larval Paropsini (Coleoptera:Chrysomelidae). Aust. J. Entomol. 6:36–38
136. Morant AV, Bjarnholt N, Kragh ME, Kjaergaard CH, Jorgensen K, et al. 2008. The β-glucosidasesresponsible for bioactivation of hydroxynitrile glucosides in Lotus japonicus. Plant Physiol. 147:1072–91
137. Morant AV, Jørgensen K, Jørgensen C, Paquette SM, Sanchez-Perez R, et al. 2008. β-glucosidases asdetonators of plant chemical defense. Phytochemistry 69:1795–813
138. Mugford ST, Louveau T, Melton R, Qi X, Bakht S, et al. 2013. Modularity of plant metabolic geneclusters: a trio of linked genes that are collectively required for acylation of triterpenes in oat. Plant Cell25:1078–92
182 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
139. Neilson EH, Goodger JQD, Motawia MS, Bjarnholt N, Frisch T, et al. 2011. Phenylalanine derivedcyanogenic diglucosides from Eucalyptus camphora and their abundances in relation to ontogeny andtissue type. Phytochemistry 72:2325–34
140. Shows that plantsreduce defense coststhrough the use ofshared biosyntheticpathways andintegration into theprimary metabolism.
140. Neilson EH, Goodger JQD, Woodrow IE, Møller BL. 2013. Plant chemical defence: at whatcost? Trends Plant Sci. 18:250–58
141. Nielsen AZ, Ziersen B, Jensen K, Lassen LM, Olsen CE, et al. 2013. Redirecting photosynthetic reducingpower toward bioactive natural product synthesis. ACS Synth. Biol. 2:308–15
142. Nielsen JS, Møller BL. 1999. Biosynthesis of cyanogenic glucosides in Triglochin maritima and theinvolvement of cytochrome P450 enzymes. Arch. Biochem. Biophys. 368:121–30
143. Nielsen KA, Olsen CE, Pontoppidan K, Møller BL. 2002. Leucine-derived cyanoglucosides in barley.Plant Physiol. 129:1066–75
144. Nielsen KA, Tattersall DB, Jones PR, Møller BL. 2008. Metabolon formation in dhurrin biosynthesis.Phytochemistry 69:88–98
145. Nishida R. 2002. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol.47:57–92
146. O’Donnell NH. 2012. Regulation of cyanogenesis in forage sorghum. PhD Thesis, Monash Univ., Victoria,Aust.
147. O’Donnell NH, Møller BL, Neale AD, Hamill JD, Blomstedt CK, Gleadow RM. 2013. Effects ofPEG-induced osmotic stress on growth and dhurrin levels of forage sorghum. Plant Physiol. Biochem.73:83–92
148. Oke OL. 1969. The role of hydrocyanic acid in nutrition. World Rev. Nutr. Diet. 11:170–98149. Olafsdottir ES, Jorgensen LB, Jaroszewski JW. 1992. Substrate specificity in the biosynthesis of cy-
clopentanoid cyanohydrin glucosides. Phytochemistry 31:4129–34150. Olsen KM, Hsu S-C, Small LL. 2008. Evidence on the molecular basis of the Ac/ac adaptive cyanogenesis
polymorphism in white clover (Trifolium repens L.). Genetics 179:517–26
151. Provides amolecular analysis of Acand Li alleles in cloverthat shows recentevolution but noevidence for balancingselection.
151. Olsen KM, Kooyers NJ, Small LL. 2013. Recurrent gene deletions and the evolution of adaptivecyanogenesis polymorphisms in white clover (Trifolium repens L.). Mol. Ecol. 22:724–38
152. Olsen KM, Sutherland BL, Small LL. 2007. Molecular evolution of the Li/li chemical defence polymor-phism in white clover (Trifolium repens L.). Mol. Ecol. 16:4180–93
153. Reviews thefunction of CNglcs as adefense againstpathogens.
153. Osbourn AE. 1996. Preformed antimicrobial compounds and plant defense against fungal attack.Plant Cell 8:1821–31
154. Patel CJ, Wright MJ. 1958. The effect of certain nutrients upon the hydrocyanic acid content of sudan-grass grown in nutrient solution. Agron. J. 50:645–47
155. Peterson SC. 1986. Breakdown products of cyanogenesis repellency and toxicity to predatory ants.Naturwissenschaften 73:627–28
156. Peterson SC, Johnson ND, LeGuyader JL. 1987. Defensive regurgitation of allelochemicals derivedfrom host cyanogenesis by eastern tent caterpillars. Ecology 68:1268–72
157. Piotrowski M. 2008. Primary or secondary? Versatile nitrilases in plant metabolism. Phytochemistry69:2655–67
158. Piotrowski M, Volmer J. 2006. Cyanide metabolism in higher plants: cyanoalanine hydratase is a NIT4homolog. Plant Mol. Biol. 61:111–22
159. Poulton JE, Li CP. 1994. Tissue level compartmentation of (R)-amygdalin and amygdalin hydrolaseprevents large-scale cyanogenesis in undamaged Prunus seeds. Plant Physiol. 104:29–35
160. Pourmohseni H, Ibenthal WD. 1991. Novel β-cyanoglucosides in the epidermal tissue of barley andtheir possible role in the barley-powdery mildew interaction. Angew. Bot. 65:341–50
161. Qi X, Bakht S, Leggett M, Maxwell C, Melton R, Osbourn A. 2004. A gene cluster for secondarymetabolism in oat: implications for the evolution of metabolic diversity in plants. Proc. Natl. Acad. Sci.USA 101:8233–38
162. Richmond GS, Ghisalberti EL. 1994. Seed dormancy and germination mechanisms in Eremophila(Myoporaceae). Aust. J. Bot. 42:705–15
163. Rockenbach J, Nahrstedt A, Wray V. 1992. Cyanogenic glycosides from Psydrax and Oxyanthus species.Phytochemistry 31:567–70
www.annualreviews.org • Cyanogenic Glycosides 183
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
164. Salzman RA, Brady JA, Finlayson SA, Buchanan CD, Summer EJ, et al. 2005. Transcriptional profilingof sorghum induced by methyl jasmonate, salicylic acid, and aminocyclopropane carboxylic acid revealscooperative regulation and novel gene responses. Plant Physiol. 138:352–68
165. Sanchez-Perez R, Belmonte FS, Borch J, Dicenta F, Møller BL, Jørgensen K. 2012. Prunasin hydrolasesduring fruit development in sweet and bitter almonds. Plant Physiol. 158:1916–32
166. Sanchez-Perez R, Jørgensen K, Motawia MS, Dicenta F, Møller BL. 2009. Tissue and cellular localizationof individual β-glycosidases using a substrate-specific sugar reducing assay. Plant J. 60:894–906
167. Sanchez-Perez R, Jørgensen K, Olsen CE, Dicenta F, Møller BL. 2008. Bitterness in almonds. PlantPhysiol. 146:1040–52
168. Santner A, Calderon-Villalobos LI, Estelle M. 2009. Plant hormones are versatile chemical regulatorsof plant growth. Nat. Chem. Biol. 5:301–7
169. Saunders JA, Conn EE. 1978. Presence of the cyanogenic glucoside dhurrin in isolated vacuoles fromsorghum. Plant Physiol. 61:154–57
170. Schappert PJ, Shore JS. 1994. Cyanogenesis in Turnera ulmifolia L. (Turneraceae). I. Phenotypic distri-bution and genetic variation for cyanogenesis on Jamaica. Heredity 74:392–404
171. Schappert PJ, Shore JS. 1999. Cyanogenesis, herbivory and plant defense in Turnera ulmifolia on Jamaica.Ecoscience 6:511–20
172. Schappert PJ, Shore JS. 1999. Effects of cyanogenesis polymorphism in Turnera ulmifolia on Euptoietahegesia and potential anolis predators. J. Chem. Ecol. 25:1455–79
173. Schappert PJ, Shore JS. 2000. Cyanogenesis in Turnera ulmifolia L. (Turneraceae): II. Developmentalexpression, heritability and cost of cyanogenesis. Evol. Ecol. Res. 2:337–52
174. Schwarz B, Wray V, Proksch P. 1996. A cyanogenic glycoside from Canthium schimperianum. Phytochem-istry 42:633–36
175. Schwind P, Wray V, Nahrstedt A. 1990. Structure elucidation of an acylated cyanogenic triglycoside,and further cyanogenic constituents from Xeranthemum cylindraceum. Phytochemistry 29:1903–11
176. Selmar D. 1993. Transport of cyanogenic glucosides: linustatin uptake by Hevea cotyledons. Planta191:191–99
177. Selmar D. 2010. Biosynthesis of cyanogenic glycosides, glucosinolates and non-protein amino acids.Annu. Plant Rev. 40:92–181
178. Proposes thatenhanced synthesis ofplant secondaryproducts under stressfulconditions provides amechanism fordissipating excessexcitation energy.
178. Selmar D, Kleinwachter M. 2013. Stress enhances the synthesis of secondary plant products:the impact of stress-related over-reduction on the accumulation of natural products. Plant CellPhysiol. 54:817–26
179. Selmar D, Lieberei R, Biehl B. 1988. Mobilization and utilization of cyanogenic glycosides: the linustatinpathway. Plant Physiol. 86:711–16
180. Sendker J, Nahrstedt A. 2009. Generation of primary amide glucosides from cyanogenic glucosides.Phytochemistry 70:388–93
181. Seo S, Mitsuhara I, Feng J, Iwai J, Hasegawa M, Ohashi Y. 2011. Cyanide, a coproduct of plant hormoneethylene biosynthesis, contributes to the resistance of rice to blast fungus. Plant Physiol. 155:502–14
182. Shimada M, Conn EE. 1977. The enzymatic conversion of p-hydroxyphenylacetaldoxime top-hydroxymandelonitrile. Arch. Biochem. Biophys. 180:199–207
183. Sibbesen O, Koch B, Halkier BA, Møller BL. 1995. Cytochrome P-450TYR is a multifunctional heme-thiolate enzyme catalyzing the conversion of L-tyrosine to p-hydroxyphenylacealdehyde oxime in thebiosynthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor (L.) Moench. J. Biol. Chem. 270:3506–11
184. Simon J, Gleadow RM, Woodrow IE. 2010. Allocation of nitrogen to chemical defence and plant func-tional traits is constrained by soil N. Tree Physiol. 30:1111–17
185. Stochmal A, Oleszek W. 1997. Changes of cyanogenic glucosides in white clover (Trifolium repens L.)during the growing season. J. Agric. Food Chem. 45:4333–36
186. Swain E, Li CP, Poulton JE. 1992. Tissue and subcellular localization of enzymes catabolizing(R)-amygdalin in mature Prunus serotina seeds. Plant Physiol. 100:291–300
187. Shows that genesencoding CNglcsynthesis in differentspecies are clustered,promoting stableinheritance offunctional pathways.
187. Takos AM, Knudsen C, Lai D, Kannangara R, Motawia MS, et al. 2011. Genomic clustering ofcyanogenic glucoside biosynthetic genes aids their identification in Lotus japonicus and suggeststhe repeated evolution of this chemical defence pathway. Plant J. 68:273–86
184 Gleadow · Møller
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP65CH06-Gleadow ARI 8 April 2014 21:52
188. Takos AM, Lai D, Mikkelsen L, Maher AH, Shelton D, et al. 2010. Genetic screening identifiescyanogenesis-deficient mutants of Lotus japonicus and reveals enzymatic specificity in hydroxynitrile glu-coside metabolism. Plant Cell 22:1605–19
189. Takos AM, Rook F. 2012. Why biosynthetic genes for chemical defense compounds cluster. Trends PlantSci. 17:383–88
190. Tattersall DB, Bak S, Jones PR, Olsen CE, Nielsen JK, et al. 2001. Resistance to an herbivore throughengineered cyanogenic glucoside synthesis. Science 293:1826–28
191. Thayer SS, Conn EE. 1981. Subcellular localisation of dhurrin β-glucosidase and hydroxynitrile lyasein the mesophyll cells of sorghum leaf blades. Plant Physiol. 67:617–22
192. Thygesen LG, Jørgensen K, Møller BL, Engelsen SB. 2004. Raman spectroscopic analysis of cyanogenicglucosides in plants: development of a flow injection surface-enhanced Raman scatter (FI-SERS) methodfor determination of cyanide. Appl. Spectrosc. 58:212–17
193. Till I. 1987. Variability of expression of cyanogenesis in white clover (Trifolium repens L.). Heredity59:265–71
194. Vandegeer R, Miller RE, Bain M, Gleadow RM, Cavagnaro TR. 2013. Drought adversely affects tuberdevelopment and nutritional quality of the staple crop cassava (Manihot esculenta Crantz). Funct. PlantBiol. 40:195–200
195. Vickery PJ, Wheeler JL, Mulcahy C. 1987. Factors affecting the hydrogen cyanide potential of whiteclover (Trifolium repens L.). Aust. J. Agric. Res. 38:1053–59
196. Wang Q, Hillwig ML, Peters RJ. 2011. CYP99A3: functional identification of a diterpene oxidase fromthe momilactone biosynthetic gene cluster in rice. Plant J. 65:87–95
197. Webber BL, Rentz DCF, Ueshima N, Woodrow IE. 2003. Leucopodoptera eumundii, a new genus andspecies of katydid from the tropical rainforests of North Queensland, Australia (Orthoptera: Tettigoni-idae: Phaneropterinae: Holochlorini). J. Orthoptera Res. 12:79–88
198. Webber BL, Woodrow IE. 2008. Intra-plant variation in cyanogenesis and the continuum of foliar plantdefense traits in the rainforest tree Ryparosa kurrangii (Achariaceae). Tree Physiol. 28:977–84
199. Webber BL, Woodrow IE. 2009. Chemical and physical plant defence across multiple ontogenetic stagesin a tropical rain forest understorey tree. J. Ecol. 97:761–71
200. Wheeler JL, Mulcahy C, Walcott JJ, Rapp GG. 1990. Factors affecting the hydrogen cyanide potentialof forage sorghum. Aust. J. Agric. Res. 41:1093–100
201. Winzer T, Gazda V, He Z, Kaminski F, Kern M, et al. 2012. A Papaver somniferum 10-gene cluster forsynthesis of the anticancer alkaloid noscapine. Science 336:1704–8
202. Zagrobelny M, Bak S, Møller BL. 2008. Cyanogenesis in plants and arthropods. Phytochemistry 69:1457–68
203. Zagrobelny M, Bak S, Olsen CE, Møller BL. 2007. Intimate roles for cyanogenic glucosides in the lifecycle of Zygaena filipendulae (Lepidoptera, Zygaenidae). Insect Biochem. Mol. Biol. 37:1189–97
204. Zagrobelny M, Bak S, Rasmussen AV, Jorgensen B, Naumann CM, Møller BL. 2004. Cyanogenicglucosides and plant–insect interactions. Phytochemistry 65:293–306
205. Zagrobelny M, Møller BL. 2011. Cyanogenic glucosides in the biological warfare between plants andinsects: the Burnet moth-birdsfoot trefoil model system. Phytochemistry 72:1585–92
206. Zagrobelny M, Motawia MS, Olsen CE, Bak S, Møller BL. 2013. Male-to-female transfer of 5-hydroxy-tryptophan glucoside during mating in Zygaena filipendulae (Lepidoptera). Insect Biochem. Mol. Biol.43:1037–44
207. Zagrobelny M, Olsen CE, Pentzold S, Furstenberg-Hagg J, Jørgensen K, et al. 2014. Sequestration, tis-sue distribution and developmental transmission of cyanogenic glucosides in a specialist insect herbivore.Insect Biochem. Mol. Biol. 44:44–53
208. Zhu-Salzman K, Salzman RA, Ahn J-E, Koiwa H. 2004. Transcriptional regulation of sorghum defensedeterminants against a phloem-feeding aphid. Plant Physiol. 134:420–31
www.annualreviews.org • Cyanogenic Glycosides 185
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP66-FrontMatter ARI 23 January 2015 17:42
Annual Review ofPlant Biology
Volume 66, 2015ContentsFrom the Concept of Totipotency to Biofortified Cereals
Ingo Potrykus � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
The Structure of Photosystem II and the Mechanism of WaterOxidation in PhotosynthesisJian-Ren Shen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �23
The Plastid Terminal Oxidase: Its Elusive Function Points to MultipleContributions to Plastid PhysiologyWojciech J. Nawrocki, Nicolas J. Tourasse, Antoine Taly, Fabrice Rappaport,
and Francis-Andre Wollman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �49
Protein Maturation and Proteolysis in Plant Plastids, Mitochondria,and PeroxisomesKlaas J. van Wijk � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �75
United in Diversity: Mechanosensitive Ion Channels in PlantsEric S. Hamilton, Angela M. Schlegel, and Elizabeth S. Haswell � � � � � � � � � � � � � � � � � � � � � � � � 113
The Evolution of Plant Secretory Structures and Emergence ofTerpenoid Chemical DiversityBernd Markus Lange � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 139
Strigolactones, a Novel Carotenoid-Derived Plant HormoneSalim Al-Babili and Harro J. Bouwmeester � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 161
Moving Toward a Comprehensive Map of Central Plant MetabolismRonan Sulpice and Peter C. McKeown � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 187
Engineering Plastid Genomes: Methods, Tools, and Applications inBasic Research and BiotechnologyRalph Bock � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 211
RNA-Directed DNA Methylation: The Evolution of a ComplexEpigenetic Pathway in Flowering PlantsMarjori A. Matzke, Tatsuo Kanno, and Antonius J.M. Matzke � � � � � � � � � � � � � � � � � � � � � � � � � 243
The Polycomb Group Protein Regulatory NetworkIva Mozgova and Lars Hennig � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 269
v
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.

PP66-FrontMatter ARI 23 January 2015 17:42
The Molecular Biology of Meiosis in PlantsRaphael Mercier, Christine Mezard, Eric Jenczewski, Nicolas Macaisne,
and Mathilde Grelon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 297
Genome Evolution in Maize: From Genomes Back to GenesJames C. Schnable � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 329
Oxygen Sensing and SignalingJoost T. van Dongen and Francesco Licausi � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 345
Diverse Stomatal Signaling and the Signal Integration MechanismYoshiyuki Murata, Izumi C. Mori, and Shintaro Munemasa � � � � � � � � � � � � � � � � � � � � � � � � � � � � 369
The Mechanism and Key Molecules Involved in Pollen Tube GuidanceTetsuya Higashiyama and Hidenori Takeuchi � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 393
Signaling to Actin Stochastic DynamicsJiejie Li, Laurent Blanchoin, and Christopher J. Staiger � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 415
Photoperiodic Flowering: Time Measurement Mechanisms in LeavesYoung Hun Song, Jae Sung Shim, Hannah A. Kinmonth-Schultz,
and Takato Imaizumi � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 441
Brachypodium distachyon and Setaria viridis: Model Genetic Systemsfor the GrassesThomas P. Brutnell, Jeffrey L. Bennetzen, and John P. Vogel � � � � � � � � � � � � � � � � � � � � � � � � � � � 465
Effector-Triggered Immunity: From Pathogen Perceptionto Robust DefenseHaitao Cui, Kenichi Tsuda, and Jane E. Parker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 487
Fungal Effectors and Plant SusceptibilityLibera Lo Presti, Daniel Lanver, Gabriel Schweizer, Shigeyuki Tanaka,
Liang Liang, Marie Tollot, Alga Zuccaro, Stefanie Reissmann,and Regine Kahmann � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 513
Responses of Temperate Forest Productivity to Insectand Pathogen DisturbancesCharles E. Flower and Miquel A. Gonzalez-Meler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 547
Plant Adaptation to Acid Soils: The Molecular Basis for CropAluminum ResistanceLeon V. Kochian, Miguel A. Pineros, Jiping Liu, and Jurandir V. Magalhaes � � � � � � � � � 571
Terrestrial Ecosystems in a Changing Environment: A DominantRole for WaterCarl J. Bernacchi and Andy VanLoocke � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 599
vi Contents
Ann
u. R
ev. P
lant
Bio
l. 20
14.6
5:15
5-18
5. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
Mon
ash
Uni
vers
ity o
n 07
/23/
20. F
or p
erso
nal u
se o
nly.






![Lecture 26 - Cyanophore Cyanogenic Glycosides [Compatibility Mode]](https://static.fdocuments.net/doc/165x107/54369a0f219acdd95f8b559d/lecture-26-cyanophore-cyanogenic-glycosides-compatibility-mode.jpg)











