
Current Status of Aedes aegypti Insecticide...
8
Research Article Current Status of Aedes aegypti Insecticide Resistance Development from Banjarmasin, Kalimantan, Indonesia P. H. Hamid , 1 V. I. Ninditya, 1 J. Prastowo, 1 A. Haryanto, 1 A. Taubert, 2 and C. Hermosilla 2 1 Veterinary Medicine, Gadjah Mada University, Jl. Fauna No. 2, Karangmalang, Yogyakarta, Indonesia 2 Institute of Parasitology, Biomedical Research Centre Seltersberg, Justus Liebig University Giessen, Schubertstr. 81, Giessen, Germany Correspondence should be addressed to P. H. Hamid; penny [email protected] Received 24 June 2018; Revised 2 November 2018; Accepted 5 December 2018; Published 20 December 2018 Academic Editor: Gabriele Gentile Copyright © 2018 P. H. Hamid et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Aedes aegypti represents the principal vector of many arthropod-borne diseases in tropical areas worldwide. Since mosquito control strategies are mainly based on use of insecticides, resistance development can be expected to occur in frequently exposed Ae. aegypti populations. Surveillance on resistance development as well as testing of insecticide susceptibility is therefore mandatory and needs further attention by national/international public health authorities. In accordance, we here conducted a study on Ae. aegypti resistance development towards several oſten used insecticides, i.e., malathion, deltamethrin, permethrin, -cyhalothrin, bendiocarb, and cyfluthrin, in the periurban area of Banjarmasin city, Kalimantan, Indonesia. Our results clearly showed resistance development of Ae. aegypti populations against tested insecticides. Mortalities of Ae. aegypti were less than 90% with the highest resistance observed against 0.75% permethrin. Collected mosquitoes from Banjarmasin also presented high level of resistance development to 0.1% bendiocarb. Molecular analysis of voltage-gated sodium channel (Vgsc) gene showed significant association of V1016G gene point mutation in resistance Ae. aegypti phenotypes against 0.75% permethrin. However, F1534C gene point mutation did not correlate to Ae. aegypti insecticide resistance to 0.75% permethrin. Irrespective of periurban areas in Kalimantan considered as less densed island of Indonesia, Ae. aegypti-derived resistance to different routinely applied insecticides occurred. Our findings evidence that Ae. aegypti insecticide resistance is most likely spreading into less populated areas and thus needs further surveillance in order to delay Ae. aegypti resistance development. 1. Background Ae. aegypti is the key factor in spreading and transmission of various infectious diseases such as Dengue Fever (DF), Chikungunya, Yellow Fever (YF), and most recently also Zika [1, 2]. Among these viral diseases, DF was classified as most important mosquito-borne disease in the world by WHO due to its wide global burden and associated consequences of economic losses in public health systems [3]. Indonesia is included as “red area” on current WHO risk map for DF together with almost all nations across equator line with tropical/subtropical climates [4]. Due to its large population, Indonesia is the most affected country in South East Asia. Indonesia is an island chain located in equator which is bridging two continents, namely, Asia and Australia. Our investigated island in this study, Kalimantan or also known as Borneo, has more rural areas than urban/ periurban areas surrounded by wet-rain forest, palm oil plantations, and coal mining companies. Escalating progress of man- made environment influence has resulted in increasing human movements rapidly through modern transportation networks from different islands to less unpopulated islands within Indonesia. History on how Ae. aegypti mosquitoes were introduced and inhabited the island of Kalimantan is still unclear. A feasible scenario of spread by sailors and their ships in colonialism era might have played relevant role in successful colonization of mosquitoes, similar to what happened in the past in Philippines and Hawaii [5]. Since insect control mainly relies on chemical compound usage all over Indonesian archipelago, we evaluated here resistance status of Ae. aegypti populations to frequently insecticides which may reflect changes of ecological bases/habitats in Hindawi BioMed Research International Volume 2018, Article ID 1735358, 7 pages https://doi.org/10.1155/2018/1735358
Transcript of Current Status of Aedes aegypti Insecticide...

Research ArticleCurrent Status of Aedes aegypti Insecticide ResistanceDevelopment from Banjarmasin Kalimantan Indonesia
P H Hamid 1 V I Ninditya1 J Prastowo1 A Haryanto1
A Taubert2 and C Hermosilla 2
1Veterinary Medicine Gadjah Mada University Jl Fauna No 2 Karangmalang Yogyakarta Indonesia2Institute of Parasitology Biomedical Research Centre Seltersberg Justus Liebig University Giessen Schubertstr 81 Giessen Germany
Correspondence should be addressed to P H Hamid penny hamidugmacid
Received 24 June 2018 Revised 2 November 2018 Accepted 5 December 2018 Published 20 December 2018
Academic Editor Gabriele Gentile
Copyright copy 2018 P H Hamid et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Aedes aegypti represents the principal vector of many arthropod-borne diseases in tropical areas worldwide Sincemosquito controlstrategies are mainly based on use of insecticides resistance development can be expected to occur in frequently exposed Aeaegypti populations Surveillance on resistance development as well as testing of insecticide susceptibility is therefore mandatoryand needs further attention by nationalinternational public health authorities In accordance we here conducted a study on Aeaegypti resistance development towards several often used insecticides ie malathion deltamethrin permethrin 120582-cyhalothrinbendiocarb and cyfluthrin in the periurban area of Banjarmasin city Kalimantan Indonesia Our results clearly showed resistancedevelopment of Ae aegypti populations against tested insecticides Mortalities of Ae aegypti were less than 90 with the highestresistance observed against 075 permethrin Collected mosquitoes from Banjarmasin also presented high level of resistancedevelopment to 01 bendiocarbMolecular analysis of voltage-gated sodium channel (Vgsc) gene showed significant association ofV1016G gene point mutation in resistanceAe aegypti phenotypes against 075 permethrin However F1534C gene pointmutationdid not correlate toAe aegypti insecticide resistance to 075 permethrin Irrespective of periurban areas inKalimantan consideredas less densed island of Indonesia Ae aegypti-derived resistance to different routinely applied insecticides occurred Our findingsevidence thatAe aegypti insecticide resistance is most likely spreading into less populated areas and thus needs further surveillancein order to delay Ae aegypti resistance development
1 Background
Ae aegypti is the key factor in spreading and transmissionof various infectious diseases such as Dengue Fever (DF)Chikungunya Yellow Fever (YF) andmost recently also Zika[1 2] Among these viral diseases DF was classified as mostimportant mosquito-borne disease in the world by WHOdue to its wide global burden and associated consequencesof economic losses in public health systems [3] Indonesiais included as ldquored areardquo on current WHO risk map for DFtogether with almost all nations across equator line withtropicalsubtropical climates [4] Due to its large populationIndonesia is the most affected country in South East Asia
Indonesia is an island chain located in equator whichis bridging two continents namely Asia and Australia Ourinvestigated island in this study Kalimantan or also known
as Borneo has more rural areas than urban periurbanareas surrounded by wet-rain forest palm oil plantationsand coal mining companies Escalating progress of man-made environment influence has resulted in increasinghuman movements rapidly through modern transportationnetworks from different islands to less unpopulated islandswithin Indonesia History on how Ae aegypti mosquitoeswere introduced and inhabited the island of Kalimantan isstill unclear A feasible scenario of spread by sailors andtheir ships in colonialism era might have played relevantrole in successful colonization of mosquitoes similar to whathappened in the past in Philippines and Hawaii [5] Sinceinsect control mainly relies on chemical compound usageall over Indonesian archipelago we evaluated here resistancestatus of Ae aegypti populations to frequently insecticideswhich may reflect changes of ecological baseshabitats in
HindawiBioMed Research InternationalVolume 2018 Article ID 1735358 7 pageshttpsdoiorg10115520181735358
2 BioMed Research International
Kalimantan Current problems faced by Kalimantan aremainly massive deforestation due to expansion of housingareas extension of new palm oil plantations coal mining-and logging-activities [6] Ecological changes in vegetationcomposition of tropical rain forests creation of new planta-tionwatering systems and dam systems for logging economicactivities are nowadays considered to have serious impact onepidemiology and spreading of various human vector-bornediseases Clear example is evidenced by recent ecosystemdestruction in Africa where use of dams and culture ofrice in paddy-fields produced large expanses of water whichare suitable breeding spots for mosquitoes and water snailsboth relevant vectors of human diseases such as malaria andschistosomiasis in sub-Saharan Africa [7] Deforestation isalso evidently enhancing landscape becoming warmer andtherefore much more hospitable for mosquito colonization[8] Studies on malaria mosquito vectors evidently showedthat deforestation and resulting impact on microclimaticenvironments greatly influenced life cycles of mosquitoesby reducing vector generation times increasing reproduc-tive rates enhancing larvalndashpupa survival rates and finallyenhancing vector populations [7 8]
Due to its flexible feeding as well as breeding habitatsAe aegypti has rapidly evolved in last decades as extremelyadapted species to human environments Ae aegyptirsquos synan-thropic behavior is relatively closer to humans when com-pared to other Aedes species eg Ae albopictus and thusleading to significantly higher spread of Ae aegypti-transmitted diseases [9] Reducing Ae aegypti populationsand minimizing interactions with humans are the mostreasonable way to control transmitted diseases because com-plete eradication of suitable vectors andor viruses is ratherunrealistic control approaches [10] Furthermore appropriatevaccines and therapies for most human diseases caused byarboviruses are still under development
Insecticides utilization is considered as the most efficienttools in vector control programs Insecticide applicationscan thereby vary from aerosol-space spraying coils lotionsclothes or curtains embedded with certain active insecticidecompounds and mass fogging to usage of larvicides in breed-ing waters Consequences of national policy breakdown byusage of massive insecticide-based controls with the sameactive compounds might result in insecticide resistancedevelopment As such Ae aegypti resistance development tocommonly used insecticides has been reported fromdifferentcountries worldwide such as Colombia [11] Brazil [12] GrandCayman [13]Thailand [14] India [15] Malaysia [16] Mexico[17] and China [18] Consistently to these findings we previ-ously reported on urban Ae aegypti resistance developmentin the cities Denpasar [19] and Jakarta [20] and evidenc-ing increased resistance to commonly used insecticides inIndonesia
Mechanism of arthropod insecticide resistance develop-ment is linked to several pathways which are basically involv-ing metabolic mechanisms and target sites [21] Main targetsite for pyrethroidchlorogenic resistance is represented bymutations in voltage-gated sodium channel (Vgsc) genes ofAe aegypti [21] Detailed Vgsc profiling ofAedes has identifiedat least seven pointmutations of insecticide knockdown (kdr)
which can clearly lead to reduced sensitivity of arthropodsodium voltage channels to exposed insecticides [22] Iden-tification of Ae aegypti specific Vgsc genes showing S989P-I1011MV- V1016GI- F1269C- and F1534C-point mutationsis related to this type of resistance [13 23ndash26] At leasttwo of these mutations ie V1016GI and F1534C wereresponsible for pyrethroid resistance as examples in Asia[14 23 27] and Latin America [24 28] Synergistic resistancemay also occur by additional point mutations in differentlocations ofVgsc genes ofAedes thereby increasing thousandsof times resistance capacities mainly towards permethrin-derived insecticides [29]
Kalimantan is well-known endemic area for not onlyrelevant arthropod-borne viral infections ie DF and YFbut also for important arthropod-borne protozoa such asPlasmodium mainly transmitted by endemic Anophelinemosquito species Chemical compounds of pesticides andinsecticides are widely utilized in Kalimantan for severalpurposes Such compounds are not only used for controllingmalaria andDF vectors but also for spraying hectares of plan-tations which is targeting agricultural pest insects Incidenceof resistance development may even occur simultaneouslywhen control programs raised against insects transmittingdiseases to humans coincide with control programs againstplant pests Consequently resistance surveillance has to beperformed periodically even in less populated (periurban) oragricultural areas to design strategies for avoiding or mini-mizing double insecticides exposure of local insect vectorsAlthough the city centre of Banjarmasin is also containinga relatively dense human population several urban partsalso show less density Our sampling points were performedin periurban areas of Banjarmasin corresponding to lesspopulated boroughs and further directly bordering forestedareasThese sampling sites were in contrast with our previoussampling sites in Denpasar and Jakarta [20 21] which allwere allocated close to the city centre [19 20] In thisstudy we explored resistance development of Ae aegypti inBanjarmasin Kalimantan to several types of commonly usedinsecticides Our results provide first important informationof current resistance status of Ae aegypti populations in thisinvestigated area
2 Methods
21 Ethics Approval and Consent to Participate No ethi-cal clearance was requested as these experiments includedexclusively Aedes mosquitoes Moreover no mosquito bloodfeedingwas here used and only F0 specimenswere used for allexperiment settings The allocation of ovitraps on residentialhousing areas represented any potential risk of infectionneither for humans nor domestic animals and therefore noethical consent was here issued
22 Mosquito Samples All mosquitoes in this experimentwere collected from periurban areas of the city Banjarmasin(3∘19101584052010158401015840S 114∘36101584029310158401015840E) in South Kalimantan IndonesiaThese collection sites were designed to cover periurbanareas of the northern (3∘16101584023910158401015840S 114∘35101584016810158401015840E) south-ern (3∘21101584046010158401015840S 114∘34101584055410158401015840E) and eastern (3∘18101584009910158401015840S
BioMed Research International 3
114∘36101584042610158401015840E) parts of the city All mosquito samples werecollected in July 2017 Ovitraps were placed after obtainingverbal permission from locals as performed in our experi-ments previously [19 20] Briefly ovitraps were constructedfrom glass with a black staining outside A filter paperwas placed in the mouth of ovitrap glass and thereafterfilled up to 34 with tap water Filled tap water and filterpapers were regularly replaced every week during collectiondates Egg-containing filter papers were carefully collectedand dried at room temperature (RT) stored and thereaftertransported in plastic containers to the Gadjah Mada Univer-sity in Yogyakarta Indonesia Mosquito eggs were allowedto hatch in the insectary of the Gadjah Mada UniversityFreshly hatched Ae aegypti-larvae were maintained in plasticcontainerswith tapwater daily fedwith chicken liver (wet anddried) until reaching adult stage All adult mosquitoes rearedfrom these collected eggs were thereafter fed with a 10 sugarsolution absorbed into cotton balls within specially designedentomological cages of the insectary Emerged mosquitoes(F0) up to three days old were exclusively used for allinsecticide-related experiments
23 Applied Insecticide Susceptibility Tests (ITS) Insecti-cide susceptibility tests (IST) were conducted according tothe international approved WHO protocols for Anophe-line mosquitoes diagnostic doses [30] Briefly the kits andinsecticide-impregnated filter papers were prepared andsupplied by the Vector Control Research Unit Univer-sity Sains Malaysia as officially WHO collaborating centrewithin Southeast Asia Impregnated filter papers tested werecontaining 5 malathion 005 deltamethrin 075 per-methrin 005 120582-cyhalothrin 01 bendiocarb and 015cyfluthrin In each IST assay a minimum of 120-150alive adult mosquitoes from three areas of Banjarmasinwere divided into 6 tubes each containing at least 20-25mosquitoes Four tubes (4 replicates) served as replicates for1 insecticide exposure and two tubes were additionally usedas controls The mosquito mortality was calculated by per-centage according to the following formula total numberof dead mosquitoestotal sample size X 100 as reportedelsewhere [30] Abbottrsquos formula was not used in this studysince control mortalities were always less than 5 Thetests were again performed in triplicate A total number ofmosquitoes here testedwere 402 specimens for 5malathion404 for 005 deltamethrin 420 for 075 permethrin 402for 005 120582-cyhalothrin 420 for 01 bendiocarb and 402for 015 cyfluthrin Somemosquito survivors as well as deadmosquitoes from these IST bioassays were kept at minus20∘Cfor further molecular resistance development analysis Resis-tance status of mosquito populations was defined accordingto the WHO recommendation corresponding to a thresholdof lt 90mortality [30]
24 DNA Isolation Isolation of DNA was performed usingPureLink Genomic DNA Isolation Kit (Invitrogen) accord-ing to manufacturer instruction Additionally we addedcombination with occasional vortexing using glass beads toease lysis of mosquitoes in the early step of DNA isolation
Dead and surviving mosquitoes obtained from IST bioassayswere extracted individually
25 V1016G and F1534C Genotyping Genotyping of themutants V1016G and F1534C was performed according toprevious report in literature [14 31] for allele-specific PCRassays For detection of V1016G the PCR consisted of 1120583l of10 pmol forward primer 51015840ACCGACAAATTGTTTCCC31015840 05120583l of 10 pmol of each reverse primer 51015840GCGGGCAGG-GCGGCGGGGGCGGGGCCAGCAAGGCTAAGAAAAG-GTTAACTC31015840 and 51015840GCGGGCAGCAAGGCTAAGAAAA-GGTTAATTA31015840 125 120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume PCR reactions were performed as follows 94∘C at2min 35 cycles of 30 s at 94∘C 30 s at 55∘C 30 s at 72∘C anda final elongation step for 2min at 72∘C PCR amplificationproducts were then loaded onto a 3 agarose gelThe F1534Cdetection PCR consisted of 1120583l of 10 pmol forward primer51015840GCGGGCTCTACTTTGTGTTCTTCATCATATT31015840 05 120583lof 10 pmol of the forward primer 51015840GCGGGCAGGGCG-GCGGGGGCGGGGCCTCTACTTTGTGTTCTTCATCAT-GTG31015840 and 1 120583l of reverse primer 51015840TCTGCTCGTTGA-AGTTGTCGAT31015840 125120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume Reactions were performed as follows 94∘C at 2min35 cycles of 30 s at 94∘C 30 s at 60∘C 30 s at 72∘C and afinal elongation step for 2min at 72∘C PCR amplificationproducts were then again loaded onto a 3 agarose gel
26 Statistical Analysis Statistical analysis and graphicalpresentation of obtained data were processed by using thecommercial available software GraphPad Prism 702 Inorder to determine mutations associated with resistancephenotype and odds ratio estimation using recessive modelwith 95 confidence and Fisherrsquos exact test were hereperformed respectively Chi square test was performed toevaluate Hardy-Weinberg Equilibrium (HWE) Differenceswere regarded as significant at the level of P lt 005
3 Results
31 Adult Ae aegypti Resistance Development to Tested Insec-ticides All adult mosquitoes (n = 2450) reared in laboratoryfrom different locations of Banjarmasin were tested againstvarious insecticides impregnated papers according to WHOprotocols [30] Based on IST results all Ae aegypti popula-tions showed various degree of resistance to insecticides withmortalities less than 90 except for malathion 5 (Figure 1)Resistance profiles to permethrin 075 and to bendiocarb01 were less than 50 (Figure 1) The highest resistancedevelopment observed was against permethrin when com-pared to any other tested insecticides except bendiocarb 01(005deltamethrin t =5059Plt 005 005 120582-cyhalothrint = 7159 P lt 001 5 malathion t = 1363 P lt 0001 01bendiocarb t = 0831 P ge 05 015 cyfluthrin t = 742 P lt001)
32 Mosquito Genotyping of Voltage-Gated Sodium Channel(Vgsc) Gene PCR genotyping showed distribution of V1016G
4 BioMed Research International
Table 1 Association of V1016G and F1534C with resistance to 075 permethrin
type ofmutation phenotypes genotypes OR
(95 CI)p value ofFisherrsquos testVV and VG GG
V1016G resistant 109 43susceptible 71 12 233 002 (S)
FF and FC CC OR(95 CI)
p value offisherrsquos test
F1534Cresistant 109 3
susceptible 91 2 125 1 (NS)OR= odds ratioCI= confidence intervalS= significantNS= nonsignificant
Cyflothrin 015
Bendiocarb 01
-cy
halothrin 005
Permeth
rin 075
Deltameth
rin 005
Malathion 5
0
20
40
60
80
100
m
orta
litie
s
Figure 1 Resistance profiles to different insecticides tested inAe aegypti from Banjarmasin Indonesia 5 malathion (n= 402)005deltamethrin (n= 404) 075 permethrin (n= 420) 005 120582-cyhalothrin (n= 402) 01 bendiocarb (n= 420) and 015cyfluthrin (n= 402)The bars are percentage of mortality after expo-sure to insecticides with error bars represents standard deviation
and F1534C in Vgsc gene domains II and III In resistantmosquitoes the wild-type VV homozygotes were 014 VGheterozygotes were 057 and GG homozygotes were 028 Insusceptible group VV frequencies were 014 VG were 071and GG were 014 Distribution frequencies of total VV VGandGG genotypes in Banjarmasin were 0145 0621 and 023respectively Total V1016 frequency was 0455 and 1016G was0545 respectively (Figure 2(a)) HWE calculation of V1016and 1016G showed P lt 005 The descriptive distributionpattern of GG genotype in resistant and susceptible mosquitopopulations is provided in Table 1 Odd ratio (OR) estimationusing recessive model of total GG genotype in V1016Gmutations showed significant association (Fisherrsquos exact testP = 002 OR = 233 95 CI = 116-459) with permethrin-resistance phenotypes (see Table 1)
F1534C distributions of F to C point mutations in Vgscgene domain III were provided in Figure 2(b) In resistantphenotype homozygotes frequencies of CC were 003 andin heterozygotes FC frequencies were 062 respectively
Susceptible mosquito samples showed CC frequencies of 002and of 048 for FC frequencies (Figure 2(b)) Distributionfrequencies of total FF FC and CC genotypes in Banjarmasinwere 042 056 and 002 Total F1534 frequency was 0698and 1534C was 0302 (Figure 2(b)) HWE calculation ofF1534 and 1534C showed P lt 005 The CC genotype wasrarely detected in Ae aegypti populations originating fromBanjarmasin Odd ratio (OR) of F1534C mutation showedno significant association (Fisherrsquos exact test P = 1 OR =125 95 CI = 025-717) with phenotypes tested againstpermethrin (Table 1) Heterozygotes were widely distributedin Ae aegypti population compared to homozygotes of bothwild-type and mutant alleles
4 Discussion
In contrast with our previous reports from other Indonesiancities [19 20] Ae aegypti population in Banjarmasin was stillsusceptible to 5 malathion since mortality rate achievedmore than 90 Surprisingly resistance to bendiocarb wasas high as for permethrin The resistance to permethrin isamong the highest corroborating previous reports conductedin Indonesia [19 20 32 33] The genotype frequency ofV1016G in Banjarmasin was significantly deviated fromHWE Mosquitoes carrying heterozygote genotype of VGwere higher when compared to previous research conductedin Indonesia The genotype frequency of F1534C in Ban-jarmasin was also significantly departed from HWE Inaccordance with other reports the CC genotype is foundrarely in the mosquito population in Banjarmasin [19 2032 33] Considering that knockdown resistance is generallysupposed to be a recessive trait [34] we used recessive modelto evaluate the association of resistant phenotype with eachsingle point mutation Molecular analysis of voltage-gatedsodium channel (Vgsc) gene showed significant associationof V1016G gene point mutation in resistance A aegyptiphenotype against 075 permethrin However F1534C genepoint mutation did not correlate to Ae aegypti insecticideresistance development to 075 permethrin in this experi-ment
Our results contribute with additional pivotal data ofglobal development of resistance to insecticides occurringwidely worldwide even in islands or geographic areas which
BioMed Research International 5
phenotypes no of samples
genotypes
VV VG GG
n freq n freq n freq
resistant 152 22 014 87 057 43 028
susceptible 83 12 014 59 071 12 014VV VG GG
00
02
04
06
08
genotype
frequ
ency
(a)
phenotypes no of samples
genotypes
FF FC CC
n freq n freq n freq
resistant 112 40 036 69 062 3 003
susceptible 93 46 049 45 048 2 002
FF FC CC00
02
04
06
genotype
frequ
ency
(b)
Figure 2 Distribution of V1016G and F1534C of Ae aegypti in Banjarmasin (a) VV VG GG and FF FC CC genotypes frequency
were considered less exposed to overuse of insecticidesHowever we speculate on possible insecticide cross-useagainst insects affecting crops and animals As such nationaland international oil palm companies in Kalimantan haveused massively agrochemicals in past decades to boost oilproduction during critical palm culture periods exposed tovarious insects such as Oryctes spp Tirathaba spp Thoseaspp and species of order LepidopteraThis phenomenon hasalso been reported in other geographic areas [35]where heavyuse of insecticides occurred without selective managementand thereby unnecessarily exposing synanthropic insectssuch as Ae aegyptimosquitoes
Despite possibilities of insecticide cross-use betweencrops and animalhuman-related insects mosquito incursionfrom different areas is now becoming in focus Severalinvestigations evidently showed that Ae aegypti can be foundin airport entry points andor seaports [36] Additionallywind-blown dispersal of various mosquito species has beendemonstrated in the past Since massive movements of goodtrades are increasing worldwide especially by the use ofonline systems these might also contribute in mosquitointroduction in previously nonendemic areas and should beconsidered as risk factors by public health authorities Nowa-days everything can be bought via Internet and afterwardsdelivered with ease by international parcel services fromlarge modern cities to far less populated and remote areasThis phenomenon needs further attention since changes intrade mechanism through revolution industry 40 can not
be avoided Additionally Ae aegypti eggs are known to behighly resistant as dried stages and as such to be foundin containers of ships crossing long oceanic distances andstill surviving for more than a year This survival strategyof Ae aegypti allows this species to be transported withease using global maritime terrestrial and air-borne traveland trade networks Mosquito larvae can hatch easily onceeggs become rehydrated in new place and to successfullycomplete their life cycle All above-mentioned anthropogenicfactors seem significantly to contribute in spread of infectiousagents worldwide crossing political borders and boundariesThis study in Banjarmasin (KalimantanBorneo Island) alsoshowed that distances and geographic barriers eg straitsandor seas can not impede resistance development as pre-viously demonstrated in our studies in the cities of Denpasar(Bali island) [19] and Jakarta (Java Island) [20] Anotherplausible scenario of resistance spread was the recolonizationof certain species which occurred for instance in a regiondeclared as being lsquomosquito-free geographic areasnationsrsquoin Brazil and probably originating from areas in whichmosquito eradication was never completely achieved [37]We stressed the point that control programs have to benationinternational teamworks and furthermore to be coor-dinated simultaneously between regions across boundariessince insecticide resistance phenotypesgenotypes in a regionmay be introduced from other islands
Putting all together insecticide resistance-related inves-tigations are needed for better understanding molecular
6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

2 BioMed Research International
Kalimantan Current problems faced by Kalimantan aremainly massive deforestation due to expansion of housingareas extension of new palm oil plantations coal mining-and logging-activities [6] Ecological changes in vegetationcomposition of tropical rain forests creation of new planta-tionwatering systems and dam systems for logging economicactivities are nowadays considered to have serious impact onepidemiology and spreading of various human vector-bornediseases Clear example is evidenced by recent ecosystemdestruction in Africa where use of dams and culture ofrice in paddy-fields produced large expanses of water whichare suitable breeding spots for mosquitoes and water snailsboth relevant vectors of human diseases such as malaria andschistosomiasis in sub-Saharan Africa [7] Deforestation isalso evidently enhancing landscape becoming warmer andtherefore much more hospitable for mosquito colonization[8] Studies on malaria mosquito vectors evidently showedthat deforestation and resulting impact on microclimaticenvironments greatly influenced life cycles of mosquitoesby reducing vector generation times increasing reproduc-tive rates enhancing larvalndashpupa survival rates and finallyenhancing vector populations [7 8]
Due to its flexible feeding as well as breeding habitatsAe aegypti has rapidly evolved in last decades as extremelyadapted species to human environments Ae aegyptirsquos synan-thropic behavior is relatively closer to humans when com-pared to other Aedes species eg Ae albopictus and thusleading to significantly higher spread of Ae aegypti-transmitted diseases [9] Reducing Ae aegypti populationsand minimizing interactions with humans are the mostreasonable way to control transmitted diseases because com-plete eradication of suitable vectors andor viruses is ratherunrealistic control approaches [10] Furthermore appropriatevaccines and therapies for most human diseases caused byarboviruses are still under development
Insecticides utilization is considered as the most efficienttools in vector control programs Insecticide applicationscan thereby vary from aerosol-space spraying coils lotionsclothes or curtains embedded with certain active insecticidecompounds and mass fogging to usage of larvicides in breed-ing waters Consequences of national policy breakdown byusage of massive insecticide-based controls with the sameactive compounds might result in insecticide resistancedevelopment As such Ae aegypti resistance development tocommonly used insecticides has been reported fromdifferentcountries worldwide such as Colombia [11] Brazil [12] GrandCayman [13]Thailand [14] India [15] Malaysia [16] Mexico[17] and China [18] Consistently to these findings we previ-ously reported on urban Ae aegypti resistance developmentin the cities Denpasar [19] and Jakarta [20] and evidenc-ing increased resistance to commonly used insecticides inIndonesia
Mechanism of arthropod insecticide resistance develop-ment is linked to several pathways which are basically involv-ing metabolic mechanisms and target sites [21] Main targetsite for pyrethroidchlorogenic resistance is represented bymutations in voltage-gated sodium channel (Vgsc) genes ofAe aegypti [21] Detailed Vgsc profiling ofAedes has identifiedat least seven pointmutations of insecticide knockdown (kdr)
which can clearly lead to reduced sensitivity of arthropodsodium voltage channels to exposed insecticides [22] Iden-tification of Ae aegypti specific Vgsc genes showing S989P-I1011MV- V1016GI- F1269C- and F1534C-point mutationsis related to this type of resistance [13 23ndash26] At leasttwo of these mutations ie V1016GI and F1534C wereresponsible for pyrethroid resistance as examples in Asia[14 23 27] and Latin America [24 28] Synergistic resistancemay also occur by additional point mutations in differentlocations ofVgsc genes ofAedes thereby increasing thousandsof times resistance capacities mainly towards permethrin-derived insecticides [29]
Kalimantan is well-known endemic area for not onlyrelevant arthropod-borne viral infections ie DF and YFbut also for important arthropod-borne protozoa such asPlasmodium mainly transmitted by endemic Anophelinemosquito species Chemical compounds of pesticides andinsecticides are widely utilized in Kalimantan for severalpurposes Such compounds are not only used for controllingmalaria andDF vectors but also for spraying hectares of plan-tations which is targeting agricultural pest insects Incidenceof resistance development may even occur simultaneouslywhen control programs raised against insects transmittingdiseases to humans coincide with control programs againstplant pests Consequently resistance surveillance has to beperformed periodically even in less populated (periurban) oragricultural areas to design strategies for avoiding or mini-mizing double insecticides exposure of local insect vectorsAlthough the city centre of Banjarmasin is also containinga relatively dense human population several urban partsalso show less density Our sampling points were performedin periurban areas of Banjarmasin corresponding to lesspopulated boroughs and further directly bordering forestedareasThese sampling sites were in contrast with our previoussampling sites in Denpasar and Jakarta [20 21] which allwere allocated close to the city centre [19 20] In thisstudy we explored resistance development of Ae aegypti inBanjarmasin Kalimantan to several types of commonly usedinsecticides Our results provide first important informationof current resistance status of Ae aegypti populations in thisinvestigated area
2 Methods
21 Ethics Approval and Consent to Participate No ethi-cal clearance was requested as these experiments includedexclusively Aedes mosquitoes Moreover no mosquito bloodfeedingwas here used and only F0 specimenswere used for allexperiment settings The allocation of ovitraps on residentialhousing areas represented any potential risk of infectionneither for humans nor domestic animals and therefore noethical consent was here issued
22 Mosquito Samples All mosquitoes in this experimentwere collected from periurban areas of the city Banjarmasin(3∘19101584052010158401015840S 114∘36101584029310158401015840E) in South Kalimantan IndonesiaThese collection sites were designed to cover periurbanareas of the northern (3∘16101584023910158401015840S 114∘35101584016810158401015840E) south-ern (3∘21101584046010158401015840S 114∘34101584055410158401015840E) and eastern (3∘18101584009910158401015840S
BioMed Research International 3
114∘36101584042610158401015840E) parts of the city All mosquito samples werecollected in July 2017 Ovitraps were placed after obtainingverbal permission from locals as performed in our experi-ments previously [19 20] Briefly ovitraps were constructedfrom glass with a black staining outside A filter paperwas placed in the mouth of ovitrap glass and thereafterfilled up to 34 with tap water Filled tap water and filterpapers were regularly replaced every week during collectiondates Egg-containing filter papers were carefully collectedand dried at room temperature (RT) stored and thereaftertransported in plastic containers to the Gadjah Mada Univer-sity in Yogyakarta Indonesia Mosquito eggs were allowedto hatch in the insectary of the Gadjah Mada UniversityFreshly hatched Ae aegypti-larvae were maintained in plasticcontainerswith tapwater daily fedwith chicken liver (wet anddried) until reaching adult stage All adult mosquitoes rearedfrom these collected eggs were thereafter fed with a 10 sugarsolution absorbed into cotton balls within specially designedentomological cages of the insectary Emerged mosquitoes(F0) up to three days old were exclusively used for allinsecticide-related experiments
23 Applied Insecticide Susceptibility Tests (ITS) Insecti-cide susceptibility tests (IST) were conducted according tothe international approved WHO protocols for Anophe-line mosquitoes diagnostic doses [30] Briefly the kits andinsecticide-impregnated filter papers were prepared andsupplied by the Vector Control Research Unit Univer-sity Sains Malaysia as officially WHO collaborating centrewithin Southeast Asia Impregnated filter papers tested werecontaining 5 malathion 005 deltamethrin 075 per-methrin 005 120582-cyhalothrin 01 bendiocarb and 015cyfluthrin In each IST assay a minimum of 120-150alive adult mosquitoes from three areas of Banjarmasinwere divided into 6 tubes each containing at least 20-25mosquitoes Four tubes (4 replicates) served as replicates for1 insecticide exposure and two tubes were additionally usedas controls The mosquito mortality was calculated by per-centage according to the following formula total numberof dead mosquitoestotal sample size X 100 as reportedelsewhere [30] Abbottrsquos formula was not used in this studysince control mortalities were always less than 5 Thetests were again performed in triplicate A total number ofmosquitoes here testedwere 402 specimens for 5malathion404 for 005 deltamethrin 420 for 075 permethrin 402for 005 120582-cyhalothrin 420 for 01 bendiocarb and 402for 015 cyfluthrin Somemosquito survivors as well as deadmosquitoes from these IST bioassays were kept at minus20∘Cfor further molecular resistance development analysis Resis-tance status of mosquito populations was defined accordingto the WHO recommendation corresponding to a thresholdof lt 90mortality [30]
24 DNA Isolation Isolation of DNA was performed usingPureLink Genomic DNA Isolation Kit (Invitrogen) accord-ing to manufacturer instruction Additionally we addedcombination with occasional vortexing using glass beads toease lysis of mosquitoes in the early step of DNA isolation
Dead and surviving mosquitoes obtained from IST bioassayswere extracted individually
25 V1016G and F1534C Genotyping Genotyping of themutants V1016G and F1534C was performed according toprevious report in literature [14 31] for allele-specific PCRassays For detection of V1016G the PCR consisted of 1120583l of10 pmol forward primer 51015840ACCGACAAATTGTTTCCC31015840 05120583l of 10 pmol of each reverse primer 51015840GCGGGCAGG-GCGGCGGGGGCGGGGCCAGCAAGGCTAAGAAAAG-GTTAACTC31015840 and 51015840GCGGGCAGCAAGGCTAAGAAAA-GGTTAATTA31015840 125 120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume PCR reactions were performed as follows 94∘C at2min 35 cycles of 30 s at 94∘C 30 s at 55∘C 30 s at 72∘C anda final elongation step for 2min at 72∘C PCR amplificationproducts were then loaded onto a 3 agarose gelThe F1534Cdetection PCR consisted of 1120583l of 10 pmol forward primer51015840GCGGGCTCTACTTTGTGTTCTTCATCATATT31015840 05 120583lof 10 pmol of the forward primer 51015840GCGGGCAGGGCG-GCGGGGGCGGGGCCTCTACTTTGTGTTCTTCATCAT-GTG31015840 and 1 120583l of reverse primer 51015840TCTGCTCGTTGA-AGTTGTCGAT31015840 125120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume Reactions were performed as follows 94∘C at 2min35 cycles of 30 s at 94∘C 30 s at 60∘C 30 s at 72∘C and afinal elongation step for 2min at 72∘C PCR amplificationproducts were then again loaded onto a 3 agarose gel
26 Statistical Analysis Statistical analysis and graphicalpresentation of obtained data were processed by using thecommercial available software GraphPad Prism 702 Inorder to determine mutations associated with resistancephenotype and odds ratio estimation using recessive modelwith 95 confidence and Fisherrsquos exact test were hereperformed respectively Chi square test was performed toevaluate Hardy-Weinberg Equilibrium (HWE) Differenceswere regarded as significant at the level of P lt 005
3 Results
31 Adult Ae aegypti Resistance Development to Tested Insec-ticides All adult mosquitoes (n = 2450) reared in laboratoryfrom different locations of Banjarmasin were tested againstvarious insecticides impregnated papers according to WHOprotocols [30] Based on IST results all Ae aegypti popula-tions showed various degree of resistance to insecticides withmortalities less than 90 except for malathion 5 (Figure 1)Resistance profiles to permethrin 075 and to bendiocarb01 were less than 50 (Figure 1) The highest resistancedevelopment observed was against permethrin when com-pared to any other tested insecticides except bendiocarb 01(005deltamethrin t =5059Plt 005 005 120582-cyhalothrint = 7159 P lt 001 5 malathion t = 1363 P lt 0001 01bendiocarb t = 0831 P ge 05 015 cyfluthrin t = 742 P lt001)
32 Mosquito Genotyping of Voltage-Gated Sodium Channel(Vgsc) Gene PCR genotyping showed distribution of V1016G
4 BioMed Research International
Table 1 Association of V1016G and F1534C with resistance to 075 permethrin
type ofmutation phenotypes genotypes OR
(95 CI)p value ofFisherrsquos testVV and VG GG
V1016G resistant 109 43susceptible 71 12 233 002 (S)
FF and FC CC OR(95 CI)
p value offisherrsquos test
F1534Cresistant 109 3
susceptible 91 2 125 1 (NS)OR= odds ratioCI= confidence intervalS= significantNS= nonsignificant
Cyflothrin 015
Bendiocarb 01
-cy
halothrin 005
Permeth
rin 075
Deltameth
rin 005
Malathion 5
0
20
40
60
80
100
m
orta
litie
s
Figure 1 Resistance profiles to different insecticides tested inAe aegypti from Banjarmasin Indonesia 5 malathion (n= 402)005deltamethrin (n= 404) 075 permethrin (n= 420) 005 120582-cyhalothrin (n= 402) 01 bendiocarb (n= 420) and 015cyfluthrin (n= 402)The bars are percentage of mortality after expo-sure to insecticides with error bars represents standard deviation
and F1534C in Vgsc gene domains II and III In resistantmosquitoes the wild-type VV homozygotes were 014 VGheterozygotes were 057 and GG homozygotes were 028 Insusceptible group VV frequencies were 014 VG were 071and GG were 014 Distribution frequencies of total VV VGandGG genotypes in Banjarmasin were 0145 0621 and 023respectively Total V1016 frequency was 0455 and 1016G was0545 respectively (Figure 2(a)) HWE calculation of V1016and 1016G showed P lt 005 The descriptive distributionpattern of GG genotype in resistant and susceptible mosquitopopulations is provided in Table 1 Odd ratio (OR) estimationusing recessive model of total GG genotype in V1016Gmutations showed significant association (Fisherrsquos exact testP = 002 OR = 233 95 CI = 116-459) with permethrin-resistance phenotypes (see Table 1)
F1534C distributions of F to C point mutations in Vgscgene domain III were provided in Figure 2(b) In resistantphenotype homozygotes frequencies of CC were 003 andin heterozygotes FC frequencies were 062 respectively
Susceptible mosquito samples showed CC frequencies of 002and of 048 for FC frequencies (Figure 2(b)) Distributionfrequencies of total FF FC and CC genotypes in Banjarmasinwere 042 056 and 002 Total F1534 frequency was 0698and 1534C was 0302 (Figure 2(b)) HWE calculation ofF1534 and 1534C showed P lt 005 The CC genotype wasrarely detected in Ae aegypti populations originating fromBanjarmasin Odd ratio (OR) of F1534C mutation showedno significant association (Fisherrsquos exact test P = 1 OR =125 95 CI = 025-717) with phenotypes tested againstpermethrin (Table 1) Heterozygotes were widely distributedin Ae aegypti population compared to homozygotes of bothwild-type and mutant alleles
4 Discussion
In contrast with our previous reports from other Indonesiancities [19 20] Ae aegypti population in Banjarmasin was stillsusceptible to 5 malathion since mortality rate achievedmore than 90 Surprisingly resistance to bendiocarb wasas high as for permethrin The resistance to permethrin isamong the highest corroborating previous reports conductedin Indonesia [19 20 32 33] The genotype frequency ofV1016G in Banjarmasin was significantly deviated fromHWE Mosquitoes carrying heterozygote genotype of VGwere higher when compared to previous research conductedin Indonesia The genotype frequency of F1534C in Ban-jarmasin was also significantly departed from HWE Inaccordance with other reports the CC genotype is foundrarely in the mosquito population in Banjarmasin [19 2032 33] Considering that knockdown resistance is generallysupposed to be a recessive trait [34] we used recessive modelto evaluate the association of resistant phenotype with eachsingle point mutation Molecular analysis of voltage-gatedsodium channel (Vgsc) gene showed significant associationof V1016G gene point mutation in resistance A aegyptiphenotype against 075 permethrin However F1534C genepoint mutation did not correlate to Ae aegypti insecticideresistance development to 075 permethrin in this experi-ment
Our results contribute with additional pivotal data ofglobal development of resistance to insecticides occurringwidely worldwide even in islands or geographic areas which
BioMed Research International 5
phenotypes no of samples
genotypes
VV VG GG
n freq n freq n freq
resistant 152 22 014 87 057 43 028
susceptible 83 12 014 59 071 12 014VV VG GG
00
02
04
06
08
genotype
frequ
ency
(a)
phenotypes no of samples
genotypes
FF FC CC
n freq n freq n freq
resistant 112 40 036 69 062 3 003
susceptible 93 46 049 45 048 2 002
FF FC CC00
02
04
06
genotype
frequ
ency
(b)
Figure 2 Distribution of V1016G and F1534C of Ae aegypti in Banjarmasin (a) VV VG GG and FF FC CC genotypes frequency
were considered less exposed to overuse of insecticidesHowever we speculate on possible insecticide cross-useagainst insects affecting crops and animals As such nationaland international oil palm companies in Kalimantan haveused massively agrochemicals in past decades to boost oilproduction during critical palm culture periods exposed tovarious insects such as Oryctes spp Tirathaba spp Thoseaspp and species of order LepidopteraThis phenomenon hasalso been reported in other geographic areas [35]where heavyuse of insecticides occurred without selective managementand thereby unnecessarily exposing synanthropic insectssuch as Ae aegyptimosquitoes
Despite possibilities of insecticide cross-use betweencrops and animalhuman-related insects mosquito incursionfrom different areas is now becoming in focus Severalinvestigations evidently showed that Ae aegypti can be foundin airport entry points andor seaports [36] Additionallywind-blown dispersal of various mosquito species has beendemonstrated in the past Since massive movements of goodtrades are increasing worldwide especially by the use ofonline systems these might also contribute in mosquitointroduction in previously nonendemic areas and should beconsidered as risk factors by public health authorities Nowa-days everything can be bought via Internet and afterwardsdelivered with ease by international parcel services fromlarge modern cities to far less populated and remote areasThis phenomenon needs further attention since changes intrade mechanism through revolution industry 40 can not
be avoided Additionally Ae aegypti eggs are known to behighly resistant as dried stages and as such to be foundin containers of ships crossing long oceanic distances andstill surviving for more than a year This survival strategyof Ae aegypti allows this species to be transported withease using global maritime terrestrial and air-borne traveland trade networks Mosquito larvae can hatch easily onceeggs become rehydrated in new place and to successfullycomplete their life cycle All above-mentioned anthropogenicfactors seem significantly to contribute in spread of infectiousagents worldwide crossing political borders and boundariesThis study in Banjarmasin (KalimantanBorneo Island) alsoshowed that distances and geographic barriers eg straitsandor seas can not impede resistance development as pre-viously demonstrated in our studies in the cities of Denpasar(Bali island) [19] and Jakarta (Java Island) [20] Anotherplausible scenario of resistance spread was the recolonizationof certain species which occurred for instance in a regiondeclared as being lsquomosquito-free geographic areasnationsrsquoin Brazil and probably originating from areas in whichmosquito eradication was never completely achieved [37]We stressed the point that control programs have to benationinternational teamworks and furthermore to be coor-dinated simultaneously between regions across boundariessince insecticide resistance phenotypesgenotypes in a regionmay be introduced from other islands
Putting all together insecticide resistance-related inves-tigations are needed for better understanding molecular
6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

BioMed Research International 3
114∘36101584042610158401015840E) parts of the city All mosquito samples werecollected in July 2017 Ovitraps were placed after obtainingverbal permission from locals as performed in our experi-ments previously [19 20] Briefly ovitraps were constructedfrom glass with a black staining outside A filter paperwas placed in the mouth of ovitrap glass and thereafterfilled up to 34 with tap water Filled tap water and filterpapers were regularly replaced every week during collectiondates Egg-containing filter papers were carefully collectedand dried at room temperature (RT) stored and thereaftertransported in plastic containers to the Gadjah Mada Univer-sity in Yogyakarta Indonesia Mosquito eggs were allowedto hatch in the insectary of the Gadjah Mada UniversityFreshly hatched Ae aegypti-larvae were maintained in plasticcontainerswith tapwater daily fedwith chicken liver (wet anddried) until reaching adult stage All adult mosquitoes rearedfrom these collected eggs were thereafter fed with a 10 sugarsolution absorbed into cotton balls within specially designedentomological cages of the insectary Emerged mosquitoes(F0) up to three days old were exclusively used for allinsecticide-related experiments
23 Applied Insecticide Susceptibility Tests (ITS) Insecti-cide susceptibility tests (IST) were conducted according tothe international approved WHO protocols for Anophe-line mosquitoes diagnostic doses [30] Briefly the kits andinsecticide-impregnated filter papers were prepared andsupplied by the Vector Control Research Unit Univer-sity Sains Malaysia as officially WHO collaborating centrewithin Southeast Asia Impregnated filter papers tested werecontaining 5 malathion 005 deltamethrin 075 per-methrin 005 120582-cyhalothrin 01 bendiocarb and 015cyfluthrin In each IST assay a minimum of 120-150alive adult mosquitoes from three areas of Banjarmasinwere divided into 6 tubes each containing at least 20-25mosquitoes Four tubes (4 replicates) served as replicates for1 insecticide exposure and two tubes were additionally usedas controls The mosquito mortality was calculated by per-centage according to the following formula total numberof dead mosquitoestotal sample size X 100 as reportedelsewhere [30] Abbottrsquos formula was not used in this studysince control mortalities were always less than 5 Thetests were again performed in triplicate A total number ofmosquitoes here testedwere 402 specimens for 5malathion404 for 005 deltamethrin 420 for 075 permethrin 402for 005 120582-cyhalothrin 420 for 01 bendiocarb and 402for 015 cyfluthrin Somemosquito survivors as well as deadmosquitoes from these IST bioassays were kept at minus20∘Cfor further molecular resistance development analysis Resis-tance status of mosquito populations was defined accordingto the WHO recommendation corresponding to a thresholdof lt 90mortality [30]
24 DNA Isolation Isolation of DNA was performed usingPureLink Genomic DNA Isolation Kit (Invitrogen) accord-ing to manufacturer instruction Additionally we addedcombination with occasional vortexing using glass beads toease lysis of mosquitoes in the early step of DNA isolation
Dead and surviving mosquitoes obtained from IST bioassayswere extracted individually
25 V1016G and F1534C Genotyping Genotyping of themutants V1016G and F1534C was performed according toprevious report in literature [14 31] for allele-specific PCRassays For detection of V1016G the PCR consisted of 1120583l of10 pmol forward primer 51015840ACCGACAAATTGTTTCCC31015840 05120583l of 10 pmol of each reverse primer 51015840GCGGGCAGG-GCGGCGGGGGCGGGGCCAGCAAGGCTAAGAAAAG-GTTAACTC31015840 and 51015840GCGGGCAGCAAGGCTAAGAAAA-GGTTAATTA31015840 125 120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume PCR reactions were performed as follows 94∘C at2min 35 cycles of 30 s at 94∘C 30 s at 55∘C 30 s at 72∘C anda final elongation step for 2min at 72∘C PCR amplificationproducts were then loaded onto a 3 agarose gelThe F1534Cdetection PCR consisted of 1120583l of 10 pmol forward primer51015840GCGGGCTCTACTTTGTGTTCTTCATCATATT31015840 05 120583lof 10 pmol of the forward primer 51015840GCGGGCAGGGCG-GCGGGGGCGGGGCCTCTACTTTGTGTTCTTCATCAT-GTG31015840 and 1 120583l of reverse primer 51015840TCTGCTCGTTGA-AGTTGTCGAT31015840 125120583l of Dream Taq Green PCR MasterMix (Thermo Fisher Scientific) in a 25120583l total reactionvolume Reactions were performed as follows 94∘C at 2min35 cycles of 30 s at 94∘C 30 s at 60∘C 30 s at 72∘C and afinal elongation step for 2min at 72∘C PCR amplificationproducts were then again loaded onto a 3 agarose gel
26 Statistical Analysis Statistical analysis and graphicalpresentation of obtained data were processed by using thecommercial available software GraphPad Prism 702 Inorder to determine mutations associated with resistancephenotype and odds ratio estimation using recessive modelwith 95 confidence and Fisherrsquos exact test were hereperformed respectively Chi square test was performed toevaluate Hardy-Weinberg Equilibrium (HWE) Differenceswere regarded as significant at the level of P lt 005
3 Results
31 Adult Ae aegypti Resistance Development to Tested Insec-ticides All adult mosquitoes (n = 2450) reared in laboratoryfrom different locations of Banjarmasin were tested againstvarious insecticides impregnated papers according to WHOprotocols [30] Based on IST results all Ae aegypti popula-tions showed various degree of resistance to insecticides withmortalities less than 90 except for malathion 5 (Figure 1)Resistance profiles to permethrin 075 and to bendiocarb01 were less than 50 (Figure 1) The highest resistancedevelopment observed was against permethrin when com-pared to any other tested insecticides except bendiocarb 01(005deltamethrin t =5059Plt 005 005 120582-cyhalothrint = 7159 P lt 001 5 malathion t = 1363 P lt 0001 01bendiocarb t = 0831 P ge 05 015 cyfluthrin t = 742 P lt001)
32 Mosquito Genotyping of Voltage-Gated Sodium Channel(Vgsc) Gene PCR genotyping showed distribution of V1016G
4 BioMed Research International
Table 1 Association of V1016G and F1534C with resistance to 075 permethrin
type ofmutation phenotypes genotypes OR
(95 CI)p value ofFisherrsquos testVV and VG GG
V1016G resistant 109 43susceptible 71 12 233 002 (S)
FF and FC CC OR(95 CI)
p value offisherrsquos test
F1534Cresistant 109 3
susceptible 91 2 125 1 (NS)OR= odds ratioCI= confidence intervalS= significantNS= nonsignificant
Cyflothrin 015
Bendiocarb 01
-cy
halothrin 005
Permeth
rin 075
Deltameth
rin 005
Malathion 5
0
20
40
60
80
100
m
orta
litie
s
Figure 1 Resistance profiles to different insecticides tested inAe aegypti from Banjarmasin Indonesia 5 malathion (n= 402)005deltamethrin (n= 404) 075 permethrin (n= 420) 005 120582-cyhalothrin (n= 402) 01 bendiocarb (n= 420) and 015cyfluthrin (n= 402)The bars are percentage of mortality after expo-sure to insecticides with error bars represents standard deviation
and F1534C in Vgsc gene domains II and III In resistantmosquitoes the wild-type VV homozygotes were 014 VGheterozygotes were 057 and GG homozygotes were 028 Insusceptible group VV frequencies were 014 VG were 071and GG were 014 Distribution frequencies of total VV VGandGG genotypes in Banjarmasin were 0145 0621 and 023respectively Total V1016 frequency was 0455 and 1016G was0545 respectively (Figure 2(a)) HWE calculation of V1016and 1016G showed P lt 005 The descriptive distributionpattern of GG genotype in resistant and susceptible mosquitopopulations is provided in Table 1 Odd ratio (OR) estimationusing recessive model of total GG genotype in V1016Gmutations showed significant association (Fisherrsquos exact testP = 002 OR = 233 95 CI = 116-459) with permethrin-resistance phenotypes (see Table 1)
F1534C distributions of F to C point mutations in Vgscgene domain III were provided in Figure 2(b) In resistantphenotype homozygotes frequencies of CC were 003 andin heterozygotes FC frequencies were 062 respectively
Susceptible mosquito samples showed CC frequencies of 002and of 048 for FC frequencies (Figure 2(b)) Distributionfrequencies of total FF FC and CC genotypes in Banjarmasinwere 042 056 and 002 Total F1534 frequency was 0698and 1534C was 0302 (Figure 2(b)) HWE calculation ofF1534 and 1534C showed P lt 005 The CC genotype wasrarely detected in Ae aegypti populations originating fromBanjarmasin Odd ratio (OR) of F1534C mutation showedno significant association (Fisherrsquos exact test P = 1 OR =125 95 CI = 025-717) with phenotypes tested againstpermethrin (Table 1) Heterozygotes were widely distributedin Ae aegypti population compared to homozygotes of bothwild-type and mutant alleles
4 Discussion
In contrast with our previous reports from other Indonesiancities [19 20] Ae aegypti population in Banjarmasin was stillsusceptible to 5 malathion since mortality rate achievedmore than 90 Surprisingly resistance to bendiocarb wasas high as for permethrin The resistance to permethrin isamong the highest corroborating previous reports conductedin Indonesia [19 20 32 33] The genotype frequency ofV1016G in Banjarmasin was significantly deviated fromHWE Mosquitoes carrying heterozygote genotype of VGwere higher when compared to previous research conductedin Indonesia The genotype frequency of F1534C in Ban-jarmasin was also significantly departed from HWE Inaccordance with other reports the CC genotype is foundrarely in the mosquito population in Banjarmasin [19 2032 33] Considering that knockdown resistance is generallysupposed to be a recessive trait [34] we used recessive modelto evaluate the association of resistant phenotype with eachsingle point mutation Molecular analysis of voltage-gatedsodium channel (Vgsc) gene showed significant associationof V1016G gene point mutation in resistance A aegyptiphenotype against 075 permethrin However F1534C genepoint mutation did not correlate to Ae aegypti insecticideresistance development to 075 permethrin in this experi-ment
Our results contribute with additional pivotal data ofglobal development of resistance to insecticides occurringwidely worldwide even in islands or geographic areas which
BioMed Research International 5
phenotypes no of samples
genotypes
VV VG GG
n freq n freq n freq
resistant 152 22 014 87 057 43 028
susceptible 83 12 014 59 071 12 014VV VG GG
00
02
04
06
08
genotype
frequ
ency
(a)
phenotypes no of samples
genotypes
FF FC CC
n freq n freq n freq
resistant 112 40 036 69 062 3 003
susceptible 93 46 049 45 048 2 002
FF FC CC00
02
04
06
genotype
frequ
ency
(b)
Figure 2 Distribution of V1016G and F1534C of Ae aegypti in Banjarmasin (a) VV VG GG and FF FC CC genotypes frequency
were considered less exposed to overuse of insecticidesHowever we speculate on possible insecticide cross-useagainst insects affecting crops and animals As such nationaland international oil palm companies in Kalimantan haveused massively agrochemicals in past decades to boost oilproduction during critical palm culture periods exposed tovarious insects such as Oryctes spp Tirathaba spp Thoseaspp and species of order LepidopteraThis phenomenon hasalso been reported in other geographic areas [35]where heavyuse of insecticides occurred without selective managementand thereby unnecessarily exposing synanthropic insectssuch as Ae aegyptimosquitoes
Despite possibilities of insecticide cross-use betweencrops and animalhuman-related insects mosquito incursionfrom different areas is now becoming in focus Severalinvestigations evidently showed that Ae aegypti can be foundin airport entry points andor seaports [36] Additionallywind-blown dispersal of various mosquito species has beendemonstrated in the past Since massive movements of goodtrades are increasing worldwide especially by the use ofonline systems these might also contribute in mosquitointroduction in previously nonendemic areas and should beconsidered as risk factors by public health authorities Nowa-days everything can be bought via Internet and afterwardsdelivered with ease by international parcel services fromlarge modern cities to far less populated and remote areasThis phenomenon needs further attention since changes intrade mechanism through revolution industry 40 can not
be avoided Additionally Ae aegypti eggs are known to behighly resistant as dried stages and as such to be foundin containers of ships crossing long oceanic distances andstill surviving for more than a year This survival strategyof Ae aegypti allows this species to be transported withease using global maritime terrestrial and air-borne traveland trade networks Mosquito larvae can hatch easily onceeggs become rehydrated in new place and to successfullycomplete their life cycle All above-mentioned anthropogenicfactors seem significantly to contribute in spread of infectiousagents worldwide crossing political borders and boundariesThis study in Banjarmasin (KalimantanBorneo Island) alsoshowed that distances and geographic barriers eg straitsandor seas can not impede resistance development as pre-viously demonstrated in our studies in the cities of Denpasar(Bali island) [19] and Jakarta (Java Island) [20] Anotherplausible scenario of resistance spread was the recolonizationof certain species which occurred for instance in a regiondeclared as being lsquomosquito-free geographic areasnationsrsquoin Brazil and probably originating from areas in whichmosquito eradication was never completely achieved [37]We stressed the point that control programs have to benationinternational teamworks and furthermore to be coor-dinated simultaneously between regions across boundariessince insecticide resistance phenotypesgenotypes in a regionmay be introduced from other islands
Putting all together insecticide resistance-related inves-tigations are needed for better understanding molecular
6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

4 BioMed Research International
Table 1 Association of V1016G and F1534C with resistance to 075 permethrin
type ofmutation phenotypes genotypes OR
(95 CI)p value ofFisherrsquos testVV and VG GG
V1016G resistant 109 43susceptible 71 12 233 002 (S)
FF and FC CC OR(95 CI)
p value offisherrsquos test
F1534Cresistant 109 3
susceptible 91 2 125 1 (NS)OR= odds ratioCI= confidence intervalS= significantNS= nonsignificant
Cyflothrin 015
Bendiocarb 01
-cy
halothrin 005
Permeth
rin 075
Deltameth
rin 005
Malathion 5
0
20
40
60
80
100
m
orta
litie
s
Figure 1 Resistance profiles to different insecticides tested inAe aegypti from Banjarmasin Indonesia 5 malathion (n= 402)005deltamethrin (n= 404) 075 permethrin (n= 420) 005 120582-cyhalothrin (n= 402) 01 bendiocarb (n= 420) and 015cyfluthrin (n= 402)The bars are percentage of mortality after expo-sure to insecticides with error bars represents standard deviation
and F1534C in Vgsc gene domains II and III In resistantmosquitoes the wild-type VV homozygotes were 014 VGheterozygotes were 057 and GG homozygotes were 028 Insusceptible group VV frequencies were 014 VG were 071and GG were 014 Distribution frequencies of total VV VGandGG genotypes in Banjarmasin were 0145 0621 and 023respectively Total V1016 frequency was 0455 and 1016G was0545 respectively (Figure 2(a)) HWE calculation of V1016and 1016G showed P lt 005 The descriptive distributionpattern of GG genotype in resistant and susceptible mosquitopopulations is provided in Table 1 Odd ratio (OR) estimationusing recessive model of total GG genotype in V1016Gmutations showed significant association (Fisherrsquos exact testP = 002 OR = 233 95 CI = 116-459) with permethrin-resistance phenotypes (see Table 1)
F1534C distributions of F to C point mutations in Vgscgene domain III were provided in Figure 2(b) In resistantphenotype homozygotes frequencies of CC were 003 andin heterozygotes FC frequencies were 062 respectively
Susceptible mosquito samples showed CC frequencies of 002and of 048 for FC frequencies (Figure 2(b)) Distributionfrequencies of total FF FC and CC genotypes in Banjarmasinwere 042 056 and 002 Total F1534 frequency was 0698and 1534C was 0302 (Figure 2(b)) HWE calculation ofF1534 and 1534C showed P lt 005 The CC genotype wasrarely detected in Ae aegypti populations originating fromBanjarmasin Odd ratio (OR) of F1534C mutation showedno significant association (Fisherrsquos exact test P = 1 OR =125 95 CI = 025-717) with phenotypes tested againstpermethrin (Table 1) Heterozygotes were widely distributedin Ae aegypti population compared to homozygotes of bothwild-type and mutant alleles
4 Discussion
In contrast with our previous reports from other Indonesiancities [19 20] Ae aegypti population in Banjarmasin was stillsusceptible to 5 malathion since mortality rate achievedmore than 90 Surprisingly resistance to bendiocarb wasas high as for permethrin The resistance to permethrin isamong the highest corroborating previous reports conductedin Indonesia [19 20 32 33] The genotype frequency ofV1016G in Banjarmasin was significantly deviated fromHWE Mosquitoes carrying heterozygote genotype of VGwere higher when compared to previous research conductedin Indonesia The genotype frequency of F1534C in Ban-jarmasin was also significantly departed from HWE Inaccordance with other reports the CC genotype is foundrarely in the mosquito population in Banjarmasin [19 2032 33] Considering that knockdown resistance is generallysupposed to be a recessive trait [34] we used recessive modelto evaluate the association of resistant phenotype with eachsingle point mutation Molecular analysis of voltage-gatedsodium channel (Vgsc) gene showed significant associationof V1016G gene point mutation in resistance A aegyptiphenotype against 075 permethrin However F1534C genepoint mutation did not correlate to Ae aegypti insecticideresistance development to 075 permethrin in this experi-ment
Our results contribute with additional pivotal data ofglobal development of resistance to insecticides occurringwidely worldwide even in islands or geographic areas which
BioMed Research International 5
phenotypes no of samples
genotypes
VV VG GG
n freq n freq n freq
resistant 152 22 014 87 057 43 028
susceptible 83 12 014 59 071 12 014VV VG GG
00
02
04
06
08
genotype
frequ
ency
(a)
phenotypes no of samples
genotypes
FF FC CC
n freq n freq n freq
resistant 112 40 036 69 062 3 003
susceptible 93 46 049 45 048 2 002
FF FC CC00
02
04
06
genotype
frequ
ency
(b)
Figure 2 Distribution of V1016G and F1534C of Ae aegypti in Banjarmasin (a) VV VG GG and FF FC CC genotypes frequency
were considered less exposed to overuse of insecticidesHowever we speculate on possible insecticide cross-useagainst insects affecting crops and animals As such nationaland international oil palm companies in Kalimantan haveused massively agrochemicals in past decades to boost oilproduction during critical palm culture periods exposed tovarious insects such as Oryctes spp Tirathaba spp Thoseaspp and species of order LepidopteraThis phenomenon hasalso been reported in other geographic areas [35]where heavyuse of insecticides occurred without selective managementand thereby unnecessarily exposing synanthropic insectssuch as Ae aegyptimosquitoes
Despite possibilities of insecticide cross-use betweencrops and animalhuman-related insects mosquito incursionfrom different areas is now becoming in focus Severalinvestigations evidently showed that Ae aegypti can be foundin airport entry points andor seaports [36] Additionallywind-blown dispersal of various mosquito species has beendemonstrated in the past Since massive movements of goodtrades are increasing worldwide especially by the use ofonline systems these might also contribute in mosquitointroduction in previously nonendemic areas and should beconsidered as risk factors by public health authorities Nowa-days everything can be bought via Internet and afterwardsdelivered with ease by international parcel services fromlarge modern cities to far less populated and remote areasThis phenomenon needs further attention since changes intrade mechanism through revolution industry 40 can not
be avoided Additionally Ae aegypti eggs are known to behighly resistant as dried stages and as such to be foundin containers of ships crossing long oceanic distances andstill surviving for more than a year This survival strategyof Ae aegypti allows this species to be transported withease using global maritime terrestrial and air-borne traveland trade networks Mosquito larvae can hatch easily onceeggs become rehydrated in new place and to successfullycomplete their life cycle All above-mentioned anthropogenicfactors seem significantly to contribute in spread of infectiousagents worldwide crossing political borders and boundariesThis study in Banjarmasin (KalimantanBorneo Island) alsoshowed that distances and geographic barriers eg straitsandor seas can not impede resistance development as pre-viously demonstrated in our studies in the cities of Denpasar(Bali island) [19] and Jakarta (Java Island) [20] Anotherplausible scenario of resistance spread was the recolonizationof certain species which occurred for instance in a regiondeclared as being lsquomosquito-free geographic areasnationsrsquoin Brazil and probably originating from areas in whichmosquito eradication was never completely achieved [37]We stressed the point that control programs have to benationinternational teamworks and furthermore to be coor-dinated simultaneously between regions across boundariessince insecticide resistance phenotypesgenotypes in a regionmay be introduced from other islands
Putting all together insecticide resistance-related inves-tigations are needed for better understanding molecular
6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom
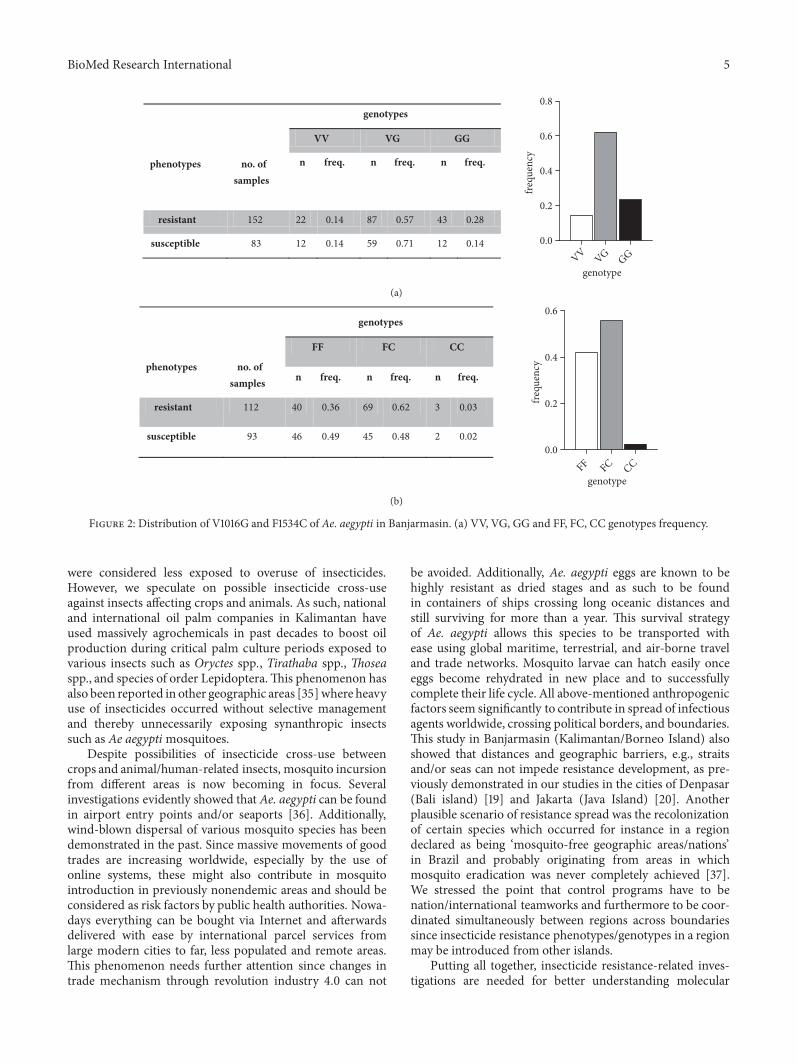
BioMed Research International 5
phenotypes no of samples
genotypes
VV VG GG
n freq n freq n freq
resistant 152 22 014 87 057 43 028
susceptible 83 12 014 59 071 12 014VV VG GG
00
02
04
06
08
genotype
frequ
ency
(a)
phenotypes no of samples
genotypes
FF FC CC
n freq n freq n freq
resistant 112 40 036 69 062 3 003
susceptible 93 46 049 45 048 2 002
FF FC CC00
02
04
06
genotype
frequ
ency
(b)
Figure 2 Distribution of V1016G and F1534C of Ae aegypti in Banjarmasin (a) VV VG GG and FF FC CC genotypes frequency
were considered less exposed to overuse of insecticidesHowever we speculate on possible insecticide cross-useagainst insects affecting crops and animals As such nationaland international oil palm companies in Kalimantan haveused massively agrochemicals in past decades to boost oilproduction during critical palm culture periods exposed tovarious insects such as Oryctes spp Tirathaba spp Thoseaspp and species of order LepidopteraThis phenomenon hasalso been reported in other geographic areas [35]where heavyuse of insecticides occurred without selective managementand thereby unnecessarily exposing synanthropic insectssuch as Ae aegyptimosquitoes
Despite possibilities of insecticide cross-use betweencrops and animalhuman-related insects mosquito incursionfrom different areas is now becoming in focus Severalinvestigations evidently showed that Ae aegypti can be foundin airport entry points andor seaports [36] Additionallywind-blown dispersal of various mosquito species has beendemonstrated in the past Since massive movements of goodtrades are increasing worldwide especially by the use ofonline systems these might also contribute in mosquitointroduction in previously nonendemic areas and should beconsidered as risk factors by public health authorities Nowa-days everything can be bought via Internet and afterwardsdelivered with ease by international parcel services fromlarge modern cities to far less populated and remote areasThis phenomenon needs further attention since changes intrade mechanism through revolution industry 40 can not
be avoided Additionally Ae aegypti eggs are known to behighly resistant as dried stages and as such to be foundin containers of ships crossing long oceanic distances andstill surviving for more than a year This survival strategyof Ae aegypti allows this species to be transported withease using global maritime terrestrial and air-borne traveland trade networks Mosquito larvae can hatch easily onceeggs become rehydrated in new place and to successfullycomplete their life cycle All above-mentioned anthropogenicfactors seem significantly to contribute in spread of infectiousagents worldwide crossing political borders and boundariesThis study in Banjarmasin (KalimantanBorneo Island) alsoshowed that distances and geographic barriers eg straitsandor seas can not impede resistance development as pre-viously demonstrated in our studies in the cities of Denpasar(Bali island) [19] and Jakarta (Java Island) [20] Anotherplausible scenario of resistance spread was the recolonizationof certain species which occurred for instance in a regiondeclared as being lsquomosquito-free geographic areasnationsrsquoin Brazil and probably originating from areas in whichmosquito eradication was never completely achieved [37]We stressed the point that control programs have to benationinternational teamworks and furthermore to be coor-dinated simultaneously between regions across boundariessince insecticide resistance phenotypesgenotypes in a regionmay be introduced from other islands
Putting all together insecticide resistance-related inves-tigations are needed for better understanding molecular
6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

6 BioMed Research International
based resistance mechanism and to assess resistance statusin exposed mosquito populations and helping to reduceoveruse of certain insecticides in national control programsPublic health local agencies may also consider combinationof biological and nonbiological strategies in controlling thesevectors Additionally impact of climate and geographic fac-tors (eg altitude temperature annual precipitation rela-tive humidity and biogeographic region) on mosquito-hostinteractions and spread of thesemosquitoes into nonendemicareas of Kalimantan is relevant topics to be considered infuture investigations in one of the most biodiversed islandsof the planet
5 Conclusions
Regular surveillance on insecticide resistance development ofAe aegypti mosquitoes is mandatory to be performed andto cover all main islands of Indonesia to set better goals andallow proper evaluation of on-going mosquito control strate-gies It is recommended that insecticide resistance on otherdisease-transmitting mosquito genera such asAnopheles andCulex shall be performed in parallel within this investigatedarea
Data Availability
The data used to support the findings of this study areincluded within the article
Conflicts of Interest
The authors declare that they have no conflicts of interestP H Hamid V I Ninditya A Haryanto and J Prastowoperformed the experiments analyzed the data and wasinvolved in the project administration P H Hamid CHermosilla and A Taubert coordinated the work and wrotethe manuscript All authors read and approved the finalversion of the manuscript
Acknowledgments
We deeply thank voluntary citizens of Banjarmasin forallowing ovitrap placements in their yards and homes Exper-iments of this work were partially supported and financed byDIKTI- (Direktorat Jenderal Pendidikan Tinggi Indonesia-)INSINAS research grant to P H Hamid
References
[1] N E A Murray M B Quam and A Wilder-Smith ldquoEpidemi-ology of dengue past present and future prospectsrdquo Journal ofClinical Epidemiology vol 5 no 1 pp 299ndash309 2013
[2] G Benelli and H Mehlhorn ldquoDeclining malaria rising ofdengue and Zika virus insights for mosquito vector controlrdquoParasitology Research vol 115 no 5 pp 1747ndash1754 2016
[3] WHOGlobal Strategy for Dengue Prevention and Control 2012-2020 WHO Press Geneva Switzerland 2012
[4] WHODengue Countries or Areas at Risk WHOPress GenevaSwitzerland 2012
[5] J R Powell and W J Tabachnick ldquoHistory of domesticationand spread of Aedes aegyptindasha reviewrdquo Memorias do InstitutoOswaldo Cruz vol 108 pp 11ndash17 2013
[6] K M Carlson R Heilmayr H K Gibbs et al ldquoEffect of oilpalm sustainability certification on deforestation and fire inIndonesiardquo Proceedings of the National Acadamy of Sciences ofthe United States of America vol 115 no 1 pp 121ndash126 2018
[7] E J Kweka E E Kimaro and SMunga ldquoEffect of deforestationand land use changes on mosquito productivity and develop-ment in western kenya highlands Implication for malaria riskrdquoFrontiers in Public Health vol 4 p 238 2016
[8] Y A Afrane G Zhou BW Lawson A K Githeko andG YanldquoEffects of microclimatic changes caused by deforestation onthe survivorship and reproductive fitness of Anopheles gambiaein western Kenya highlandsrdquo The American Journal of TropicalMedicine and Hygiene vol 74 no 5 pp 772ndash778 2006
[9] Y Tsuda W Suwonkerd S Chawprom S Prajakwong and MTakagi ldquoDifferent spatial distribution of Aedes aegypti andAedes albopictus along an urban-rural gradient and the relatingenvironmental factors examined in three villages in northernThailandrdquo Journal of the American Mosquito Control Associa-tion vol 22 no 2 pp 222ndash228 2006
[10] A C Morrison E Zielinski-Gutierrez T W Scott and RRosenberg ldquoDefining challenges and proposing solutions forcontrol of the virus vector Aedes aegyptirdquo PLoS Medicine vol5 no 3 p e68 2008
[11] I Fonseca-Gonzalez M L Quinones A Lenhart and W GBrogdon ldquoInsecticide resistance status of Aedes aegypti (L)from Colombiardquo Pest Management Science vol 67 no 4 pp430ndash437 2011
[12] E P Lima M H S Paiva A P de Araujo et al ldquoInsecticideresistance in Aedes aegypti populations from Ceara BrazilrdquoParasites amp Vectors vol 4 no 1 p 5 2011
[13] A FHarris S Rajatileka andH Ranson ldquoPyrethroid resistanceinAedes aegypti fromGrand CaymanrdquoTheAmerican Journal ofTropical Medicine and Hygiene vol 83 no 2 pp 277ndash284 2010
[14] J Yanola P Somboon CWaltonWNachaiwieng P Somwangand L-A Prapanthadara ldquoHigh-throughput assays for detec-tion of the F1534C mutation in the voltage-gated sodiumchannel gene in permethrin-resistant Aedes aegypti and thedistribution of this mutation throughout Thailandrdquo TropicalMedicineamp International Health vol 16 no 4 pp 501ndash509 2011
[15] R B S Kushwah C L Dykes N Kapoor T Adak and OP Singh ldquoPyrethroid-Resistance and Presence of Two Knock-downResistance (kdr)Mutations F1534C andaNovelMutationT1520I in Indian Aedes aegyptirdquo PLOS Neglected TropicalDiseases vol 9 no 1 p e3332 2015
[16] I H Ishak Z Jaal H Ranson and C S Wondji ldquoContrastingpatterns of insecticide resistance and knockdown resistance(kdr) in the dengue vectors Aedes aegypti and Aedes albopictusfromMalaysiardquo Parasites amp Vectors vol 8 no 1 p 181 2015
[17] G P Garcıa A E Flores I Fernandez-Salas et al ldquoRecentrapid rise of a permethrin knock down resistance allele inAedesaegypti in Mexicordquo PLOS Neglected Tropical Diseases vol 3 no10 p e531 2009
[18] C-X Li P E Kaufman R-D Xue et al ldquoRelationship betweeninsecticide resistance and kdr mutations in the dengue vectorAedes aegypti in Southern Chinardquo Parasites amp Vectors vol 8 p325 2015
[19] P H Hamid J Prastowo A Widyasari A Taubert and CHermosilla ldquoKnockdown resistance (kdr) of the voltage-gated
BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

BioMed Research International 7
sodium channel gene of Aedes aegypti population in DenpasarBali Indonesiardquo Parasites amp Vectors vol 10 no 1 2017
[20] P H Hamid J Prastowo A Ghiffari A Taubert and C Her-mosilla ldquoAedes aegypti resistance development to commonlyused insecticides in Jakarta Indonesiardquo PLoS ONE vol 12 no12 2017
[21] J Hemingway N J Hawkes L McCarroll andH Ranson ldquoThemolecular basis of insecticide resistance in mosquitoesrdquo InsectBiochemistry and Molecular Biology vol 34 no 7 pp 653ndash6652004
[22] Y Du Y Nomura G Satar et al ldquoMolecular evidence fordual pyrethroid-receptor sites on a mosquito sodium channelrdquoProceedings of the National Acadamy of Sciences of the UnitedStates of America vol 110 no 29 pp 11785ndash11790 2013
[23] C Brengues N J Hawkes F Chandre et al ldquoPyrethroid andDDT cross-resistance in Aedes aegypti is correlated with novelmutations in the voltage-gated sodium channel generdquo Medicaland Veterinary Entomology vol 17 no 1 pp 87ndash94 2003
[24] K Saavedra-Rodriguez L Urdaneta-Marquez S Rajatileka etal ldquoA mutation in the voltage-gated sodium channel geneassociated with pyrethroid resistance in Latin American Aedesaegyptirdquo Insect Molecular Biology vol 16 no 6 pp 785ndash7982007
[25] H Kawada Y Higa O Komagata et al ldquoWidespread distribu-tion of a newly found point mutation in voltage-gated sodiumchannel in pyrethroid-resistant Aedes aegypti populations inVietnamrdquo PLOS Neglected Tropical Diseases vol 3 no 10 pe527 2009
[26] R Srisawat N Komalamisra C Apiwathnasorn et al ldquoField-collected permethrin-resistant Aedes aegypti from centralThailand contain point mutations in the domain IIS6 of thesodium channel gene (kdr)rdquo Southeast Asian Journal of TropicalMedicine and Public Health vol 43 no 6 pp 1380ndash1386 2012
[27] S Plernsub J Saingamsook J Yanola et al ldquoAdditive effect ofknockdown resistance mutations S989P V1016G and F1534Cin a heterozygous genotype conferring pyrethroid resistance inAedes aegypti in Thailandrdquo Parasites amp Vectors vol 9 no 1 p417 2016
[28] L C Alvarez G Ponce K Saavedra-Rodriguez B Lopez andA E Flores ldquoFrequency of V1016I and F1534C mutations inthe voltage-gated sodium channel gene in Aedes aegypti inVenezuelardquoPestManagement Science vol 71 no 6 pp 863ndash8692015
[29] K Hirata O Komagata K Itokawa A Yamamoto T Tomitaand S Kasai ldquoA Single Crossing-Over Event in Voltage-Sensitive Na+ Channel Genes May Cause Critical Failure ofDengue Mosquito Control by Insecticidesrdquo PLOS NeglectedTropical Diseases vol 8 no 8 2014
[30] WHO Test procedures for insecticide resistance monitoring inmalaria vectormosquitoesWorldHealthOrganizationGeneva2013
[31] S A Stenhouse S Plernsub J Yanola et al ldquoDetection of theV1016G mutation in the voltage-gated sodium channel gene ofAedes aegypti (Diptera Culicidae) by allele-specific PCR assayand its distribution and effect on deltamethrin resistance inThailandrdquo Parasites amp Vectors vol 6 no 1 p 253 2013
[32] J R Wuliandari S F Lee V L White W Tantowijoyo AA Hoffmann and N M Endersby-Harshman ldquoAssociationbetween three mutations F1565C V1023G and S996P in thevoltage-sensitive sodium channel gene and knockdown resis-tance in aedes aegypti from yogyakarta Indonesiardquo Insects vol6 no 3 pp 658ndash685 2015
[33] S Sayono A P N Hidayati S Fahri et al ldquoDistribution ofvoltage-gated sodium channel (NAV) alleles among the aedesaegypti populations in central Java province and its associationwith resistance to pyrethroid insecticidesrdquo PLoS ONE vol 11no 3 p e0150577 2016
[34] T G E Davies A O OrsquoReilly L M Field B A Wallaceand M S Williamson ldquoKnockdown resistance to DDT andpyrethroids From target-site mutations to molecular mod-ellingrdquo Pest Management Science vol 64 no 11 pp 1126ndash11302008
[35] H A A Khan W Akram K Shehzad and E A ShaalanldquoFirst report of field evolved resistance to agrochemicals indengue mosquito Aedes albopictus (Diptera Culicidae) fromPakistanrdquo Parasites amp Vectors vol 4 no 1 p 146 2011
[36] M-M Kuan and F-Y Chang ldquoAirport sentinel surveillanceand entry quarantine for dengue infections following a feverscreening program in Taiwanrdquo BMC Infectious Diseases vol 12p 182 2012
[37] P Kotsakiozi A Gloria-Soria A Caccone et al ldquoTracking thereturn ofAedes aegypti toBrazil themajor vector of the denguechikungunya and Zika virusesrdquo PLOS Neglected Tropical Dis-eases vol 11 no 7 p e0005653 2017
Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom

Hindawiwwwhindawicom
International Journal of
Volume 2018
Zoology
Hindawiwwwhindawicom Volume 2018
Anatomy Research International
PeptidesInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Journal of Parasitology Research
GenomicsInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawiwwwhindawicom
The Scientific World Journal
Volume 2018
Hindawiwwwhindawicom Volume 2018
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Neuroscience Journal
Hindawiwwwhindawicom Volume 2018
BioMed Research International
Cell BiologyInternational Journal of
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Biochemistry Research International
ArchaeaHindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Genetics Research International
Hindawiwwwhindawicom Volume 2018
Advances in
Virolog y Stem Cells International
Hindawiwwwhindawicom Volume 2018
Hindawiwwwhindawicom Volume 2018
Enzyme Research
Hindawiwwwhindawicom Volume 2018
International Journal of
MicrobiologyHindawiwwwhindawicom
Nucleic AcidsJournal of
Volume 2018
Submit your manuscripts atwwwhindawicom


















