
Cooperative Biotin Binding by Streptavidin
6
THE JOURNAL OF BIOLOGICAL CHEMLSTRY a 1990 by The Amwican Society for Biochemktry and Molecular Biology, Inc. Cooperative Biotin Binding by Streptavidin Vol. 265, No. 6, Iswe of February 25, pp. 3369-3373,199O Printed in U.S. A. ELECTROPHORETIC BEHAVIOR AND SUBUNIT ASSOCIATION OF STREPTAVIDIN IN THE PRESENCE OF 6 M UREA* (Received for publication, October 4, 1989) Takeshi Sano$ and Charles R. Cantor+8 From the Department of Genetics and Development, College of Physicians and Surgeons, Columbia University, New York, New York 10032 We describe the cooperativity in the biotin binding of streptavidin. We have developed an electrophoretic method which can separate streptavidin molecules with bound biotin from those without biotin. In 6 M urea, the electrophoretic mobility of streptavidin in polyacrylamide gels becomes significantly faster upon biotin binding. When streptavidin was titrated with biotin, only two major bands were observed on the gel, consisting of streptavidin molecules without bound bio- tin and those saturated with biotin. The change in mobility is due partly to the negative charge of the bound biotin, but it must reflect conformational changes of the protein molecule associated with biotin binding. Gel filtration chromat.ography showed that the streptavidin molecule dissociates into two subunit dimers in the presence of 6 M urea. These results sug- gest that the biotin binding by the streptavidin subunit dimer is cooperative and that some communication must exist between the two subunits. The streptavidin-biotin and avidin-biotin systems provide att.ractive methods for detection, characterization, and isola- tion of various biological materials (for review see Green, 1975; Bayer et ul., 1979; Bayer and Wilchek, 198@ Shamsud- din and Harris, 1983; Wilchek and Bayer, 1984,1988; Fuccillo, 1985; Buckland, 1986; Uhlen, 1989). The remarkably high and specific biotin-binding affinity of streptavidin and avidin (Kd - lo-l5 M) and the potential ahility of biotin to be incorpo- rated into various biological substances facilitate a variety of applications. The streptavidin molecule consists of four iden- tical subunits without any carbohydrate moiety and binds four D-biotin molecules (Chaiet et ul., 1964; Chaiet and Wolf, 1964; Green, 1975). In contrast, avidin is a glycoprotein with one carbohydrate chain/subunit, and the subunit composition of a molecule is heterogeneous because of differences in the carbohydrate moiety (Fraenkel-Conrat et ul., 1952; Huang and DeLange, 1971; DeLange and Huang, 1971; Brush and White, 1982; Gatti et ul., 1984; Gope et ul., 1987). There is great interest in the unusual biotin-binding prop- erty of streptavidin and avidin, since such a high and specific * This work was supported by Grant CA39782 from the National Cancer Institute, National Institutes of Health. The costs of publi- cation of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “uduertise- men? in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ Present address: Dept. of Molecular and Cell Biology, University of California, Berkeley, CA 94720. § To whom correspondence should be addressed. Present address: Human Genome Ctr., Lawrence Berkeley Laboratory, One Cyclotron Rd., MS I-213, Berkeley, CA 94720. Tel.: 415-486-6800. affinity is not generally observable in other biological systems. Earlier reports have shown that the formation of the strept- avidin-biotin or avidin-biotin complex does not involve any covalent bonds (Green, 1975). The biotin binding of the four subunits of avidin is considered to be random (Green, 1964, 1966, 1975), but the biotin-binding affinity of the isolated subunit is lower than that of the intact tetramer (Green and Toms, 1973). In spite of numerous studies, the nature of the biotin binding of streptavidin and avidin is not fully under- stood. Recently, two groups (Hendrickson et ul., 1989; Weber et ul., 1989) independently determined the three-dimensional structure of the streptavidin molecule by x-ray cryst,allogra- phy. These studies revealed that many hydrogen bonds and van der Waals interactions are involved in the formation of the streptavidin-biotin complex. These intermolecular inter- actions produce the high and specific binding affinity. Con- formational changes in streptavidin, associated with biotin binding, also apparently contribute to the stabilization of the streptavidin-biotin complex. In the present work, we have developed an electrophoretic method for detecting biotin binding to streptavidin. This electrophoresis system, containing 6 M urea, can separate streptavidin molecules w&h bound biotin from those without biotin. Based on the elect,rophoretic behavior of st,reptavidin in this system and on the subunit association of streptavidin in the presence of 6 M urea, we demonstrate that there is cooperativity in biotin-binding of streptavidin. This implies the existence of some kind of communication between the subunits. EXPERIMENTAL PROCEDURES Mute&k-Streptavidin and BSA’ (fraction V) were obtained from Boehringer Mannheim and used without further purification. Urea (ultrapure grade) was from Schwartz/Mann Biotech or International Biotechnologies. Amplify, a scintillator for fluorography, and D-[car- borzyl-T] biotin were from Amersham Corp. Sephacryl S-300 HR was from Pharmacia LKB Biotechnology Inc. M, standard proteins for gel filtration chromatography were from Boehringer Mannheim or Sigma. Other reagents were analytical grade. Preparation of Recombinunt Streptavidin-Recombinant streptav- idin was expressed in Escherichia coli and purified according to the methods described previously (San0 and Cantor, 1990). We used a lysogen BL2l(DE3) transformed with pLysE and pTSA-2, which carry the T7 lysozyme gene and the streptavidin gene, respectively. Electrophoresis-Polyacrylamide gel electrophoresis in the pres- ence of 6 M urea (6 M urea-PAGE) basically followed the method of Perrie and Perry (1966), using an 8.0% acrylamide gel with a stacking gel of 3.5% acrylamide (SieGankowski aid Dreizin, 1978). The gei consisted of a 37.5 to 1 weight ratio of acrvlamide to N.w-methvl- enebisacrylamide in 6 M uria, 25 mM Tris,-80 rnM glyciie (pH 8.;). The running buffer was composed of 25 mM Tris, 80 mM glycine (pH ’ The abbreviations used are: BSA, bovine serum albumin; PAGE, polyacrylamide gel electrophoresis; SDS, sodium dodecyl sulfate. 3369 by guest on January 12, 2019 http://www.jbc.org/ Downloaded from
Transcript of Cooperative Biotin Binding by Streptavidin

THE JOURNAL OF BIOLOGICAL CHEMLSTRY a 1990 by The Amwican Society for Biochemktry and Molecular Biology, Inc.
Cooperative Biotin Binding by Streptavidin
Vol. 265, No. 6, Iswe of February 25, pp. 3369-3373,199O Printed in U.S. A.
ELECTROPHORETIC BEHAVIOR AND SUBUNIT ASSOCIATION OF STREPTAVIDIN IN THE PRESENCE OF 6 M UREA*
(Received for publication, October 4, 1989)
Takeshi Sano$ and Charles R. Cantor+8 From the Department of Genetics and Development, College of Physicians and Surgeons, Columbia University, New York, New York 10032
We describe the cooperativity in the biotin binding of streptavidin. We have developed an electrophoretic method which can separate streptavidin molecules with bound biotin from those without biotin. In 6 M urea, the electrophoretic mobility of streptavidin in polyacrylamide gels becomes significantly faster upon biotin binding. When streptavidin was titrated with biotin, only two major bands were observed on the gel, consisting of streptavidin molecules without bound bio- tin and those saturated with biotin. The change in mobility is due partly to the negative charge of the bound biotin, but it must reflect conformational changes of the protein molecule associated with biotin binding. Gel filtration chromat.ography showed that the streptavidin molecule dissociates into two subunit dimers in the presence of 6 M urea. These results sug- gest that the biotin binding by the streptavidin subunit dimer is cooperative and that some communication must exist between the two subunits.
The streptavidin-biotin and avidin-biotin systems provide att.ractive methods for detection, characterization, and isola- tion of various biological materials (for review see Green, 1975; Bayer et ul., 1979; Bayer and Wilchek, 198@ Shamsud- din and Harris, 1983; Wilchek and Bayer, 1984,1988; Fuccillo, 1985; Buckland, 1986; Uhlen, 1989). The remarkably high and specific biotin-binding affinity of streptavidin and avidin (Kd - lo-l5 M) and the potential ahility of biotin to be incorpo- rated into various biological substances facilitate a variety of applications. The streptavidin molecule consists of four iden- tical subunits without any carbohydrate moiety and binds four D-biotin molecules (Chaiet et ul., 1964; Chaiet and Wolf, 1964; Green, 1975). In contrast, avidin is a glycoprotein with one carbohydrate chain/subunit, and the subunit composition of a molecule is heterogeneous because of differences in the carbohydrate moiety (Fraenkel-Conrat et ul., 1952; Huang and DeLange, 1971; DeLange and Huang, 1971; Brush and White, 1982; Gatti et ul., 1984; Gope et ul., 1987).
There is great interest in the unusual biotin-binding prop- erty of streptavidin and avidin, since such a high and specific
* This work was supported by Grant CA39782 from the National Cancer Institute, National Institutes of Health. The costs of publi- cation of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “uduertise- men? in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Dept. of Molecular and Cell Biology, University of California, Berkeley, CA 94720.
§ To whom correspondence should be addressed. Present address: Human Genome Ctr., Lawrence Berkeley Laboratory, One Cyclotron Rd., MS I-213, Berkeley, CA 94720. Tel.: 415-486-6800.
affinity is not generally observable in other biological systems. Earlier reports have shown that the formation of the strept- avidin-biotin or avidin-biotin complex does not involve any covalent bonds (Green, 1975). The biotin binding of the four subunits of avidin is considered to be random (Green, 1964, 1966, 1975), but the biotin-binding affinity of the isolated subunit is lower than that of the intact tetramer (Green and Toms, 1973). In spite of numerous studies, the nature of the biotin binding of streptavidin and avidin is not fully under- stood. Recently, two groups (Hendrickson et ul., 1989; Weber et ul., 1989) independently determined the three-dimensional structure of the streptavidin molecule by x-ray cryst,allogra- phy. These studies revealed that many hydrogen bonds and van der Waals interactions are involved in the formation of the streptavidin-biotin complex. These intermolecular inter- actions produce the high and specific binding affinity. Con- formational changes in streptavidin, associated with biotin binding, also apparently contribute to the stabilization of the streptavidin-biotin complex.
In the present work, we have developed an electrophoretic method for detecting biotin binding to streptavidin. This electrophoresis system, containing 6 M urea, can separate streptavidin molecules w&h bound biotin from those without biotin. Based on the elect,rophoretic behavior of st,reptavidin in this system and on the subunit association of streptavidin in the presence of 6 M urea, we demonstrate that there is cooperativity in biotin-binding of streptavidin. This implies the existence of some kind of communication between the subunits.
EXPERIMENTAL PROCEDURES
Mute&k-Streptavidin and BSA’ (fraction V) were obtained from Boehringer Mannheim and used without further purification. Urea (ultrapure grade) was from Schwartz/Mann Biotech or International Biotechnologies. Amplify, a scintillator for fluorography, and D-[car-
borzyl-T] biotin were from Amersham Corp. Sephacryl S-300 HR was from Pharmacia LKB Biotechnology Inc. M, standard proteins for gel filtration chromatography were from Boehringer Mannheim or Sigma. Other reagents were analytical grade.
Preparation of Recombinunt Streptavidin-Recombinant streptav- idin was expressed in Escherichia coli and purified according to the methods described previously (San0 and Cantor, 1990). We used a lysogen BL2l(DE3) transformed with pLysE and pTSA-2, which carry the T7 lysozyme gene and the streptavidin gene, respectively.
Electrophoresis-Polyacrylamide gel electrophoresis in the pres- ence of 6 M urea (6 M urea-PAGE) basically followed the method of Perrie and Perry (1966), using an 8.0% acrylamide gel with a stacking gel of 3.5% acrylamide (SieGankowski aid Dreizin, 1978). The gei consisted of a 37.5 to 1 weight ratio of acrvlamide to N.w-methvl- enebisacrylamide in 6 M uria, 25 mM Tris,-80 rnM glyciie (pH 8.;). The running buffer was composed of 25 mM Tris, 80 mM glycine (pH
’ The abbreviations used are: BSA, bovine serum albumin; PAGE, polyacrylamide gel electrophoresis; SDS, sodium dodecyl sulfate.
3369
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

3370 Cooperative Biotin Binding by Streptavidin
8.6) without urea. The protein sample was adjusted to a urea concen- tration of 6 M by diluting with sample buffer consisting of 10 M urea, 25 mM Tris, 80 mM glycine (pH 8.6). Alternatively, solid urea was added to a protein solution to a final concentration of 6 M. Dialysis of the protein sample against 6 M urea, 25 mM Tris, 80 mM glycine (pH 8.6) also worked well; however, the dilution procedure is easier and reduces the time needed.
PAGE analysis under nondenaturing conditions (nondenaturing PAGE) was performed according to the method of Davis (1964) using a 7.5% acrylamide gel with a stacking gel of 3% acrylamide. PAGE analysis in the presence of SDS (SDS-PAGE) was conducted in the discontinuous buffer system of Laemmli (1970). Proteins were stained with 0.15% Coomassie Brilliant Blue R-250 (Sigma) dissolved in 45% methanol, 10% acetic acid.
For fluorography, each denaturing gel was washed twice with 5 volumes of 20% ethanol, 10% acetic acid for 10 min with gentle shaking to remove urea from the gel. For nondenaturing gels this step was omitted. The gel was then immersed in 3 volumes of 50% Amplify, 20% ethanol for 30 min with gentle shaking and dried in UUCUO. The dried gel was exposed to Kodak X-Omat AR film at -70 ‘C.
Gel Fihation Chromatography-Gel filtration chromatography was carried out at room temperature (-20 ‘YJ using a Sephacryl S- 300 HR column (1.6 X 95 cm) equilibrated with 6 M urea, 25 mM Tris, 80 mM glycine (pH 8.6). Streptavidin (200 pg, 15-nmol subunits) in 400 ~1 of 6 M urea, 25 mM Tris, 80 mM glycine (pH 8.6) was applied onto a column and eluted with the same buffer at a flow rate of 35 ml h-l. The &I, was estimated by calibration using Mr standard proteins (BSA, 67,000; ovalbumin, 43,000; chymotrypsinogen A, 25,000; cytochrome c, 12,500).
Determinahon oj Biohn-binding Ability-Biotin-binding ability of streptavidin was determined by the bentonite adsorption method (Korenman and O’Malley, 1967, 1970) using D-[curbonyl-‘4C]biotin (52 mCi mmol-‘).
De~erminahon of Protein Concentration-Protein concentrations of commercial streptavidin and BSA were determined using the extinction coefficients Z?g& 3.4 (Green and Melamed. 1966) and ---. I!&‘& 0.63 (Webster, 1970), respeitively. Protein conc&tration of the recombinant streptavidin was determined by the micro-biuret method (Itzhaki and Gill, 1964).
RESULTS
Characterization of Protein Preparations-The streptavidin preparation from Boehringer Mannheim showed a single sharp band at iVr 13,500 on SDS-PAGE analysis (data not shown). This iL!fr was smaller than that of the mature strept- avidin subunit (16,500) obtained from the deduced amino acid sequence of the streptavidin gene of Streptomyces avidinii (Argaraiia et al., 1986), suggesting that this preparation had been truncated at both termini to a minimum size, so-called core streptavidin (Ptihler et al., 1987; Hendrickson et al., 1989; Bayer et al., 1989). This preparation bound 3.9 molecules of biotin/molecule (subunit tetramer, M* 54,000), which is the almost full biotin-binding ability (4 molecules of biotin/mol- ecule). BSA also showed a single band at Mr 67,000 on SDS- PAGE analysis and was used as the negative control in the following experiments.
The recombinant streptavidin expressed in E. coli has an intact C terminus, and the structure of N terminus is almost the same as core streptavidin with additional seven amino acids (Sane and Cantor, 1990). This preparation showed a single band at Mr 15,900 on SDS-PAGE analysis and bound 3.8-4.0 molecules of biotin/molecule.
6 M Urea-PAGE Analysis-We employed 6 M urea-PAGE for detecting the biotin-binding of streptavidin. The migration of a protein molecule in 6 M urea-PAGE is affected by various factors, including Mr, net charge, and conformation of the protein molecule. In 6 M urea, protein molecules are usually unfolded and display a smaller mobility than in the native state.
The addition of biotin led to a marked increase in the mobility of streptavidin on 6 M urea-PAGE (Fig. 1A). There was no effect of biotin on the mobility of BSA. The fluorogram
Aab Bab -- -m BlotIn - + - + -+ -+
FIG. 1. Polyacrylamide gel electrophoresis of streptavidin in the presence of 6 M urea. Electrophoresis was carried out in the presence of 6 M urea basically according to Perrie and Perry (1966) using a 7.5% acrylamide gel. A, stained protein; B, fluorogram. Strept- avidin (4 pg, 0.30-nmol subunit) with or without biotin (0.18 pg, 0.74 nmol) was applied in each lane after the addition of urea to 6 M (kznes a). For fluorography, D-[curboxyl-‘4C]biotin (0.18 pg, 0.73 nmol) was used. BSA (5 fig, 75 pmol) with or without biotin was run as a negative control (kznes b). Note that the amount of unbound biotin was reduced extensively because the gel was washed before fluorography.
FIG. 2. Titration of streptavidin with biotin monitored by polyacrylamide gel electrophoresis in the presence of 6 M urea. A, stained protein; B, fluorogram. Conditions are exactly the same as described in Fig. 1 except for the molar ratio of biotin to streptavidin subunit, indicated above each lane.
of the gel (Fig. 1B) shows that streptavidin has bound biotin under the condition of the PAGE analysis, suggesting that the migration changes resulted from biotin binding. The fluorogram also indicates that there is little or no dissociation of bound biotin molecules from streptavidin in 6 M urea. It is thus clear that 6 M urea-PAGE can be used for the detection of biotin binding by streptavidin.
Some minor protein bands were observable together with the major one. These bands also migrated faster in the pres- ence of biotin than in the absence, similar to the major protein band. In addition, the protein in these bands also bound biotin. It is quite likely that most of these minor bands consisted of aggregated streptavidin molecules, which are usually present in most streptavidin preparations (Ptihler et al., 1987; Bayer et al., 1989).
Titration of Streptavidin and Biotin-A titration of strept- avidin with biotin was carried out and monitored by 6 M urea- PAGE. Two major stained protein bands were observed on the gel, corresponding to the mobility of free streptavidin and streptavidin saturated with biotin (Fig. 2.4). At molar ratios
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from
Cooperative Biotin Binding by Streptavidin
of biotin to streptavidin subunit between 0 and 1, some faint bands were also observed at positions between the two major 0.06
VO
3371
Chymobypsinow A BSA Odbumin Cylochmmec
bands. Some of these are aggregated molecules; others may Ill 11
represent streptavidin partially saturated with biotin. A fluo- 0.05
rogram of the gel (Fig. 2I3) shows that only the fast moving E 1
major band and the fastest moving of the minor bands bound i i
0.04
biotin. The migration changes were saturated at a molar ratio of biotin to streptavidin subunit of 1, indicating that the migration changes are solely the result of biotin binding. The failure to see large amounts of protein between the two major stained bands indicates either that the binding of biotin by streptavidin in 6 M urea displays considerable cooperativity or that the streptavidin molecule dissociates into monomeric subunit in 6 M urea. If the former is true, the cooperativity in O.OOL biotin binding must be the result of subunit interactions, 0 SO 100 150 zoo
remaining in 6 M urea. Elution Volume (ml) Nondenaturing PAGE Analysis-We next examined the FIG. 4. Gel filtration chromatography of streptavidin in the
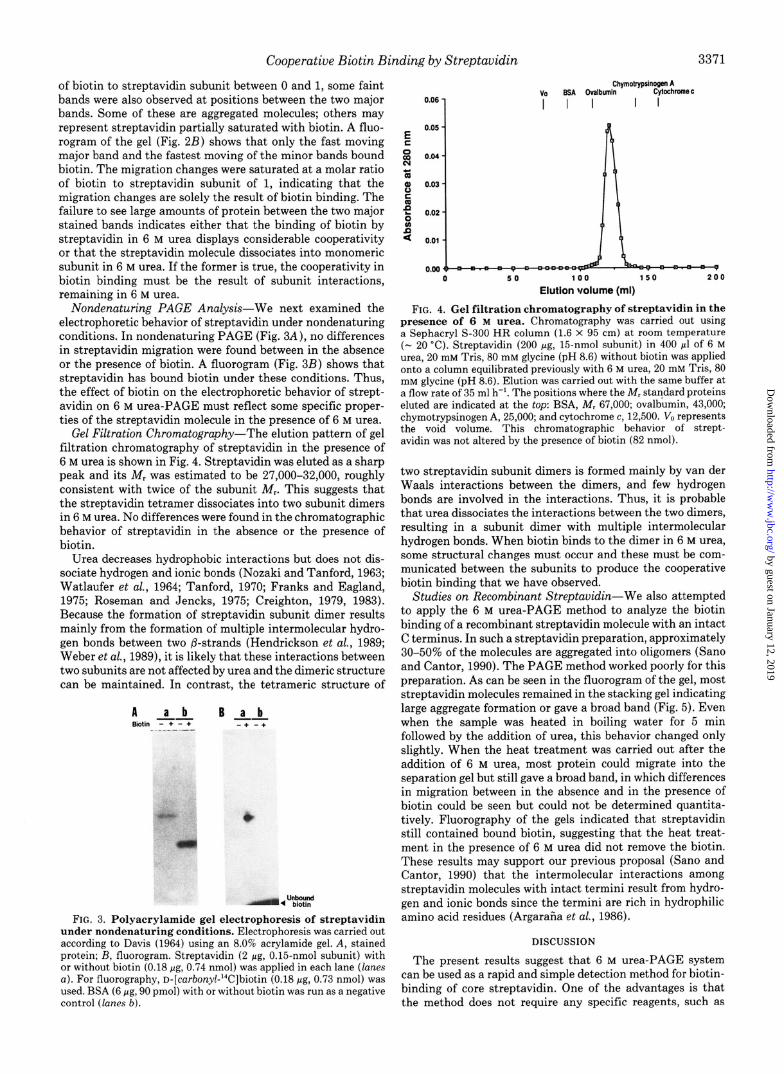
electrophoretic behavior of streptavidin under nondenaturing presence of 6 M urea. Chromatography was carried out using conditions. In nondenaturing PAGE (Fig. 3A), no differences a Sephacryl S-300 HR column (1.6 x 95 cm) at room temperature in streptavidin migration were found between in the absence (- 20 ‘C). Streptavidin (200 pg, 15nmol subunit) in 400 ~1 of 6 M or the presence of biotin. A fluorogram (Fig. 3B) shows that urea, 20 mM Tris, SO mM glycine (pH 8.6) without biotin was applied
streptavidin has bound biotin under these conditions. Thus, onto a column equilibrated previously with 6 M urea, 20 mM Tris, 80
the effect of biotin on the electrophoretic behavior of strept- mM glycine (pH 8.6). Elution was carried out with the same buffer at a flow rate of 35 ml h-‘. The positions where the Mr standard proteins
avidin on 6 M urea-PAGE must reflect some specific proper- eluted are indicated at the top: BSA, Mr 67,000; ovalbumin, 43,000; ties of the streptavidin molecule in the presence of 6 M urea. chymotrypsinogen A, 25,000; and cytochrome c, 12,500. V,) represents
Gel Filtration Chromatography-The elution pattern of gel the void volume. This chromatographic behavior of strept- filtration chromatography of streptavidin in the presence of avidin was not altered by the presence of biotin (82 nmol).
6 M urea is shown in Fig. 4. Streptavidin was eluted as a sharp peak and its Mr was estimated to be 27,000-32,000, roughly two streptavidin subunit dimers is formed mainly by van der consistent with twice of the subunit Mr. This suggests that Waals interactions between the dimers, and few hydrogen the streptavidin tetramer dissociates into two subunit dimers bonds are involved in the interactions. Thus, it is probable in 6 M urea. No differences were found in the chromatographic that urea dissociates the interactions between the two dimers, behavior of streptavidin in the absence or the presence of resulting in a subunit dimer with multiple intermolecular biotin. hydrogen bonds. When biotin binds to the dimer in 6 M urea,
Urea decreases hydrophobic interactions but does not dis- some structural changes must occur and these must be com- sociate hydrogen and ionic bonds (Nozaki and Tanford, 1963; municated between the subunits to produce the cooperative Watlaufer et al., 1964; Tanford, 1970; Franks and Eagland, biotin binding that we have observed. 1975; Roseman and Jencks, 1975; Creighton, 1979, 1983). Studies on Recombinant Streptavidin-We also attempted Because the formation of streptavidin subunit dimer results to apply the 6 M urea-PAGE method to analyze the biotin mainly from the formation of multiple intermolecular hydro- binding of a recombinant streptavidin molecule with an intact gen bonds between two @-strands (Hendrickson et al., 1989; C terminus. In such a streptavidin preparation, approximately Weber et al., 1989), it is likely that these interactions between 30-50% of the molecules are aggregated into oligomers (Sano two subunits are not affected by urea and the dimeric structure and Cantor, 1990). The PAGE method worked poorly for this can be maintained. In contrast, the tetrameric structure of preparation. As can be seen in the fluorogram of the gel, most
streptavidin molecules remained in the stacking gel indicating A a b large aggregate formation or gave a broad band (Fig. 5). Even -- B ~4 sii,,” - + - + -+ -+ when the sample was heated in boiling water for 5 min . ----
followed by the addition of urea, this behavior changed only slightly. When the heat treatment was carried out after the addition of 6 M urea, most protein could migrate into the separation gel but still gave a broad band, in which differences in migration between in the absence and in the presence of biotin could be seen but could not be determined quantita- tively. Fluorography of the gels indicated that streptavidin still contained bound biotin, suggesting that the heat treat- ment in the presence of 6 M urea did not remove the biotin. These results may support our previous proposal (Sane and Cantor, 1990) that the intermolecular interactions among
-4%Z streptavidin molecules with intact termini result from hydro- gen and ionic bonds since the termini are rich in hydrophilic
FIG. 3. Polyacrylamide gel electrophoresis of streptavidin amino acid residues (Argaraiia et al., 1986). under nondenaturing conditions. Electrophoresis was carried out according to Davis (1964) using an 8.0% acrylamide gel. A, stained DISCUSSION protein; B, fluorogram. Streptavidin (2 Kg, 0.15.nmol subunit) with or without biotin (0.18 pg, 0.74 nmol) was applied in each lane (lanes The present results suggest that 6 M urea-PAGE system a). For fluorography, n-[carbonyl-“C]biotin (0.18 gg, 0.73 nmol) was can be used as a rapid and simple detection method for biotin- used. BSA (6 pg, 90 pmol) with or without biotin was run as a negative binding of core streptavidin. One of the advantages is that control (/arm b). the method does not require any specific reagents, such as
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

3372 Cooperative Biotin Binding by Streptavidin
A B C ---
FIG. 5. Polyacrylamide gel electrophoresis of recombinant streptavidin in the presence of 6 M urea. Electrophoresis was carried out as described in Fig. 1. Only the fluorogram of the gel was shown. Recombinant streptaiidin (4 Lg, 0.25-nmol subunit) &th or without D-~ca~~ony~-14Clbiotin (0.18 ILL?. 0.73 nmol) was annlied in each lane after the addition oi urea?0 6 M. The following heat treatments were made on the protein sample before the application to the gel: A, no heat treatment; L3, the protein sample was heated in boiling water for 5 min before the addition of urea; C, the protein sample was heated in boiling water for 5 min in the presence of 6 M urea.
biotin molecules labeled with radioisotopes or conjugated to some enzymes. The sensitivity depends on the staining method for protein. Thus Coomassie Brilliant Blue R-250 staining, which is used most frequently for proteins in poly- acrylamide gel, detects as little as 1 pg of streptavidin, al- though streptavidin has a lower reactivity with Coomassie Brilliant Blue than most of other proteins (Sharma and Tihon, 1988). The use of the silver-staining method would increase this sensitivity.
Here we discuss the factors affecting the electrophoretic behavior of streptavidin in the presence of 6 M urea. Because biotin is an acidic molecule (p1 - 3.5), biotin binding of streptavidin causes a shift in net charge, which would give faster migration on PAGE. However, if this factor were dom- inant, a similar result should be also obtained on nondena- turing PAGE, contrary to what we observed (Fig. 3). Thus, some additional factors must be involved in changes in elec- trophoretic behavior in 6 M urea.
Urea has a structure similar to part of the biotin molecule, the ureido group. The residues of streptavidin responsible for binding the ureido group of biotin are aromatic and polar amino acids (Hendrickson et ul., 1989; Weber et ul., 1989), and several water molecules may occupy these sites in the absence of biotin. Upon the addition of urea, these water molecules are probably replaced by urea molecules (Green, 1963, 1975). Because the Kd of streptavidin-urea complex must be much higher (for the avidin-urea complex, Kd - 3.6 x lo-’ M; Green, 1963, 1975) than that of the streptavidin- biotin complex (-lo-” M), streptavidin does not release bound biotin molecules even if 6 M urea is added. Even though urea is neutral, its p1 is slightly higher than that of streptav- idin (p1 - 5.5-6). Thus, urea binding of streptavidin may result in only a slight charge change.
Crystallographic studies (Weber et al., 1989) show that the binding of biotin induces conformational changes in the strept- avidin subunit. Two surface loops of the streptavidin subunit, which are disordered in the absence of biotin, are ordered to
bury the biotin molecule. These transitions should not occur when urea binds to streptavidin, because they are induced by the carboxyl portion of biotin molecule. These results of the loop ordering and possibly other factors such as ionic bond formation are to flatten the streptavidin subunit slightly and make it more compact. It is understandable that this could result in faster streptavidin migration in a gel matrix. Thus, one might be tempted to argue that a similar conformational change induced in streptavidin subunit by biotin binding may be responsible for the faster mobility on 6 M urea-PAGE. However, the gel filtration chromatographic behavior of strept- avidin was not altered by biotin binding. This might indicate that overall size and shape changes play an insignificant role in the biotin-induced shift in PAGE mobility. One dominant factor must be net charge changes caused by binding of urea or biotin.
The quaternary structure of the streptavidin molecule is also changed by biotin binding (Weber et ul., 1989). Because the migration of streptavidin on nondenaturing PAGE, where streptavidin molecule is tetrameric, was not altered by biotin binding (Fig. 3), it is likely that these changes are hidden within the tetrameric structure and do not affect its size or shape in a major way. In urea the streptavidin molecule presumably has a somewhat unfolded structure and the biotin- induced conformational changes or charge changes may be larger than in the native state, because more amino acid residues are exposed at the surface of the molecule.
We conclude that changes in the net charge of the strept- avidin subunit dimer caused by the transition from bound urea and bound biotin is the principle cause of the increase in electrophoretic mobility of streptavidin in 6 M urea. Con- formational changes in streptavidin induced by biotin binding may also contribute to the migration changes, especially if these lead to the loss of protons. The dissociation of streptav- idin into two subunit dimers and the unfolded structure, which are induced by urea, may amplify these changes.
Green (1964, 1966, 1975) suggested from fluorescence quenching measurements that the biotin binding of avidin is not cooperative. On the other hand, Weber et ul. (1989) suggested the existence of some communication between the subunits. In a titration of streptavidin with biotin on 6 M urea-PAGE (Fig. 2), only two major bands were observed on the gel. If the biotin binding of the subunit dimer is sequential or random, a protein band corresponding to a subunit dimer with one biotin molecule should be the major species observed on the gel at molar ratios of biotin to streptavidin subunit near 0.5. Such a protein band is expected to have a mobility between the bands corresponding to free subunit dimers and biotin-saturated dimers. However, our results showed no such major band. These results reveal that the biotin binding of the streptavidin subunit dimer is cooperative and that some communication exists between the subunits in the dimer. Biotin binding of the subunit probably causes some confor- mational change in the other subunit within the dimer. This notion is consistent with the differences in the crystal struc- ture between free streptavidin and saturated streptavidin, suggesting the presence of subunit communication (Weber et cd., 1989). It is not clear, however, why the cooperativity in biotin binding seen in the isolated dimer in 6 M urea is lost in the tetramer.
In the native tetrameric molecule, a tryptophan residue located in a loop near the p-strand closest to the C terminus of the subunit is involved in the biotin binding of the other subunit (Hendrickson et al., 1989; Weber et al., 1989). This is apparently why the isolated subunit has a lower affinity for biotin. This interaction occurs through the junction between
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

Cooperative Biotin Binding by Streptavidin 3373
the two subunit dimers and is lost by the dissociation into two subunit dimers in 6 M urea. Thus, this interaction should not be involved in the cooperativity in biotin binding in the subunit dimer.
A&no&edgrne&s-We thank Drs. Arno Pahler and Wayne A. Hendrickson for many valuable suggestions and discussion and Dr. Mitsuo Ikebe for information on urea-PAGE analyses.
REFERENCES
Argaraiia, C. E., Kuntz, I. D., Birkin, S., Axel, R., and Cantor, C. R. (1986) Nucleic Acids Res. 14, 1871-1882
Bayer, E. A., and Wilchek, M. (1980) Methods Biochem. AT&. 26, l- 46
Bayer, E. A., Skutelsky, E., and Wilchek, M. (1979) Methods Enzymol, 62,308-315
Bayer, E. A., Ben-Hur, H., Hiller, Y., and Wilchek, M. (1989) Biochem. J, 259,369-376
Brush, R. C., and White, H. B., III (1982) Biochemistry 21, 5334- 5341
Buckland, R. M. (1986) Nature 320, 557-558 Chaiet, L., and Wolf, F. J. (1964) Arch. Biochem. Biophys. 106, l-5 Chaiet, L., Miller, T. W., Tausing, F., and Wolf, F. J. (1964) Anti-
microb. Agents Chemother. 3, 28-32 Creighton, T. E, (1979) J. Mol. Biol. 129, 235-264 Creighton, T. E. (1983) Proteins-Structure and Mole&w- Principles,
pp. 149-152 and 291-295, W. H. Freeman, New York Davis, B. J. (1964) Ann. N. Y. Acad. Sci. 121,404-427 DeLange, R. J., and Huang, T.-S. (1971) J. Biol. Chem. 246, 698-
709 Fraenkel-Conrat, H., Snell, N. S., and Ducay, E. D. (1952) Arch.
Biochem. Biophys, 39, 97-107 Franks, F., and Eagland, D. (1975) Crit. Reu. Biochem. 3, 165-219 Fuccillo, D. A. (1985) EioTechniques 3, 494-501 Gatti, G., Bolognesi, M., Coda, A., Chiolerio, F., Filippini, E., and
Malcovati, M. (1984) J. MoZ. Biol. 178, 787-789 Gape, M. L., Kleinanen, R. A., Kristo, P. A., Conneely, 0. M., Beattie,
W. G., Zaruchi-Shulz, T., O’Malley, B. W., and Kulomaa, M. S. (1987) Nucleic Acids Res. 15, 3595-3606
Green, N. M. (1963) Biochem. J. 89,599-609 Green, N. M. (1964) Biochem. J. 90, 564-568 Green, N. M. (1966) Biochem. J. 101, 774-780 Green, N. M. (1975) Adv. Protein Chem. 29,85-133 Green, N. M., and Melamed, M. D. (1966) Biochem. J. 100,614-621 Green, N. M., and Toms, E. J. (1973) Biochem. J. 133,687-700 Hendrickson, W. A., Pahler, A., Smith, J. A., Satow, Y., Merritt, E.
A., and Phizackerley, R. P. (1989) Proc. Natl. Acad. Sci. U. S. A. 86,2190-2194
Huang, T.-S., and DeLange, R. J. (1971) J. Biol. Chem. 246, 686- 697
Itzhaki, R. F., and Gill, D. M. (1964) Anul. Biochem. 9,401-410 Korenman, S. G., and O’Malley, B. W. (1967) Biochim. Biophys. Acta
140,174-176 Korenman, S. G., and O’Malley, B. W. (1970) Methods Enzymol.
18A,427-430 Laemmli, U. K. (1970) Nuture 227,680-685 Nozaki, Y., and Tanford, C. (1963) J. Biol. Chem. 238. 4074-4081 Pahler, A.; Hendrickson, W. A., Kolks, M. A. G., Argaraiia, C. E.,
and Cantor, C. R. (1987) J. Biol. Chem. 262, 13933-13937 Perrie, W. T., and Perry, S. V. (1970) Biochem. J. 119,31-38 Roseman, M., and Jencks. W. P. (1975) J. Am. Chem. Sot. 97.631-
640 Sano, T., and Cantor, C. R. (1990) Proc. Natl. Acud. Sci. U. S. A. 87,
142-146 Shamsuddin, A. M., and Harris, C. C. (1983) Arch. Puthol. Lab. Med.
107,514-517 Sharma, H. K., and Tihon, C. (1988) Anul. Biochem. 170,135-139 Siemankowski, R. F., and Dreizen, P. (1978) J. Biol. Chem. 253,
8648-8658 Tanford, C. (1970) Adv. Protein Chem. 24, l-95 Uhlen, M. (1989) Nuture 340. 733-734 Watlaufer, D. B., Malik, S. K.; Stoller, L., and Coffin, R. L. (1964) J.
Am. Chem. Sot. 84,508-514 Weber, P. C., Ohlendorf, D. H., Wendoloski, J. J., and Salemme, F.
R. (1989) Science 243,85-88 Webster, G. C. (1970) Biochim. Biophys. Acta 207,371-373 Wilchek, M., and Bayer, E. A. (1984) Zmmunol. Today 5,39-43 Wilchek, M., and Bayer, E. A. (1988) Anul. Biochem. 171, l-32
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

T Sano and C R Cantorassociation of streptavidin in the presence of 6 M urea.
Cooperative biotin binding by streptavidin. Electrophoretic behavior and subunit
1990, 265:3369-3373.J. Biol. Chem.
http://www.jbc.org/content/265/6/3369Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/6/3369.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 12, 2019http://w
ww
.jbc.org/D
ownloaded from

















![Conducting Polymer-Based Composite Materials for ...example, George et al. modified the surface of PPy through biotin-streptavidin coupling [45]. Due to the ability of attaching any](https://static.fdocuments.net/doc/165x107/6144dc7934130627ed509edb/conducting-polymer-based-composite-materials-for-example-george-et-al-modiied.jpg)
