
Control y Modulación de la selectividad de diferentes ... · Tesis Doctoral. José Miguel Palomo...
167
Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química orgánica en medios macro orgánica en medios macro orgánica en medios macro orgánica en medios macro-acuosos acuosos acuosos acuosos Universidad Autónoma de Madrid Facultad de Ciencias Departament Universidad Autónoma de Madrid Facultad de Ciencias Departament Universidad Autónoma de Madrid Facultad de Ciencias Departament Universidad Autónoma de Madrid Facultad de Ciencias Departament o de Química Orgánica o de Química Orgánica o de Química Orgánica o de Química Orgánica Jose Jose Jose Jose Miguel Palomo Carmona Miguel Palomo Carmona Miguel Palomo Carmona Miguel Palomo Carmona Tesis Doctoral Tesis Doctoral Tesis Doctoral Tesis Doctoral
Transcript of Control y Modulación de la selectividad de diferentes ... · Tesis Doctoral. José Miguel Palomo...
Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes Control y Modulación de la selectividad de diferentes derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química derivados de lipasas en reacciones de química
orgánica en medios macroorgánica en medios macroorgánica en medios macroorgánica en medios macro----acuosos acuosos acuosos acuosos
Uni
vers
idad
Aut
ónom
a de
Mad
rid F
acul
tad
de
Cie
ncia
s D
epar
tam
ent
Uni
vers
idad
Aut
ónom
a de
Mad
rid F
acul
tad
de
Cie
ncia
s D
epar
tam
ent
Uni
vers
idad
Aut
ónom
a de
Mad
rid F
acul
tad
de
Cie
ncia
s D
epar
tam
ent
Uni
vers
idad
Aut
ónom
a de
Mad
rid F
acul
tad
de
Cie
ncia
s D
epar
tam
ent o
de
Quí
mic
a O
rgán
ica
o de
Quí
mic
a O
rgán
ica
o de
Quí
mic
a O
rgán
ica
o de
Quí
mic
a O
rgán
ica
JoseJoseJoseJose Miguel Palomo CarmonaMiguel Palomo CarmonaMiguel Palomo CarmonaMiguel Palomo Carmona
Tesis DoctoralTesis DoctoralTesis DoctoralTesis Doctoral

JoséJoséJoséJosé Miguel Palomo Carmona Miguel Palomo Carmona Miguel Palomo Carmona Miguel Palomo Carmona
Control y Control y Control y Control y mmmmodulación de la selectividad de odulación de la selectividad de odulación de la selectividad de odulación de la selectividad de derivados inmovilizados de distintas derivados inmovilizados de distintas derivados inmovilizados de distintas derivados inmovilizados de distintas lipasaslipasaslipasaslipasas,,,, en en en en reacciones de química orgánicareacciones de química orgánicareacciones de química orgánicareacciones de química orgánica desarrolladas desarrolladas desarrolladas desarrolladas
en en en en sistemas sistemas sistemas sistemas macromacromacromacro----acuososacuososacuososacuosos
Memoria presentada para optar al grado de Doctor Memoria presentada para optar al grado de Doctor Memoria presentada para optar al grado de Doctor Memoria presentada para optar al grado de Doctor por por por por la Universidad Autónoma de Madridla Universidad Autónoma de Madridla Universidad Autónoma de Madridla Universidad Autónoma de Madrid
DirectorDirectorDirectorDirectoreseseses:::: José José José José Manuel Guisán Seijas Manuel Guisán Seijas Manuel Guisán Seijas Manuel Guisán Seijas Roberto Fernández Lafuente Roberto Fernández Lafuente Roberto Fernández Lafuente Roberto Fernández Lafuente Instituto de Catálisis Instituto de Catálisis Instituto de Catálisis Instituto de Catálisis yyyy Universidad Universidad Universidad Universidad AutónomaAutónomaAutónomaAutónoma de Madrid de Madrid de Madrid de Madrid Petroleoquímica, CSICPetroleoquímica, CSICPetroleoquímica, CSICPetroleoquímica, CSIC Facultad de ciencias. Facultad de ciencias. Facultad de ciencias. Facultad de ciencias. Madrid Madrid Madrid Madrid Departamento de Química Orgánica Departamento de Química Orgánica Departamento de Química Orgánica Departamento de Química Orgánica
Madrid 2003Madrid 2003Madrid 2003Madrid 2003

La realización de esta tesis, ha sido posible, en parte, gracias a una Beca de formación de personal
investigador concedida por la Comunidad de Madrid.
En primer lugar, me gustaría agradecer de manera muy especial la oportunidad, la confianza y el apoyo
que el Dr. José Manuel Guisán Seijas ha depositado en mí durante estos años.
También, un especial agradecimiento quiero brindar al Dr. Roberto Fernández Lafuente por su
inestimable ayuda en todo momento tanto a nivel de trabajo experimental como en el aprendizaje
obtenido a lo largo de estos años que contemplan mi carrera científica.
A los Drs. Vicente Gotor, Lucía Ramírez y Maria Luisa Rúa por la ayuda prestada para la realización de
esta tesis.
A los Drs. Javier Soria Ruiz y Sagrario Mendioroz Echevarría, directores sucesivos del Instituto de Catálisis,
por las facilidades prestadas.
Al Dr. José Luis García Ruano, tutor de esta Tesis doctoral, por su disponibilidad y ayuda en todo
momento.
Un cariñoso agradecimiento a Mari Carmen Ceinos, por su buena disposición, paciencia y apoyo
durante la elaboración de este manuscrito.
Además, de manera especial un entrañable recuerdo a mis compañeros del laboratorio con los que he
compartido estos años y con los que he forjado una gran amistad, tanto al inicio de esta tesis, Gloria,
Olguita, “choni” Mateo, Rosa o Manolillo, como los que han ido apareciendo a lo largo de estos años, el
Beni, la Betanzos, la Wilson (Piquetin), Rodrigo, Claudita, Aurelio; además de las nuevas adquisiciones del
laboratorio Tamarismo, Fernando, Miguelito, Georgette y Noelia.
Un recuerdo muy especial para Gloria Muñoz (Glorita), gracias a su inestimable ayuda en la realización
de esta tesis, en su paso por nuestro laboratorio, y por la amistad ofrecida durante muchos años.
También un saludo especial a las dos brujas, la Berlus y la Vale, por su buen humor y paciencia durante
las largas horas de trabajo.
Una mención a todos aquellos que han pasado por aquí durante todo este tiempo Víctor Balcao, Rubens,
Paulo, Celia, Marcelo, Angelica, Eduardo, Andrés Illanes, Carmen Luisa, etc…
A mis colegas Javi, Mario, Alex, Pepe, Andi y Martin por su amistad y por los grandes momentos que
hemos vivido juntos.
A Ángel (Perrunis), por la gran amistad forjada durante mi estancia en Holanda y las sugerencias
aportadas en la escritura de los manuscritos relacionados con esta tesis.
Por ultimo y muy sinceramente a mi familia: mis padres (Fernando y Fuensanta), a mi hermano Fernando,
a mis abuelas, tíos, tías, primos y primas; y especialmente a Angeelika (mi tikikene), los que
verdaderamente me conocen y han estado a mi lado apoyándome tanto en los buenos como en los
malos momentos tanto en mi carrera científica como en mi vida privada.

Índice Índice Índice Índice iiii
Índice Introducción GeneralIntroducción GeneralIntroducción GeneralIntroducción General
A. Lipasas 1
B. Absorción de las lipasas sobre estructuras hidrofóbicas
3
C. Estabilización de lipasas 4
D. Modulación de las propiedades catalíticas de las lipasas. 4
Objetivos GeneralesObjetivos GeneralesObjetivos GeneralesObjetivos Generales 7
Capítulo 1. InteracciónCapítulo 1. InteracciónCapítulo 1. InteracciónCapítulo 1. Interacción de lipasas con estructuras hidrofóbicas de lipasas con estructuras hidrofóbicas de lipasas con estructuras hidrofóbicas de lipasas con estructuras hidrofóbicas
-Introducción 9
1. Interacción lipasa-lipasa 9
2. Interacción de lipasas con proteínas hidrofóbicas 10
3. Características físico-químicas de las distintas lipasas 11
-Objetivos 13
-Resultados y Discusión 14
1. Tendencia general de las lipasas a autoasociarse formando estructuras bimoleculares: modificación de las propiedades funcionales.
14
1.1 Purificación de lipasas. 14
1.2 Análisis de la autoasociación de lipasas. 15
1.3 Efecto de la concentración de enzima sobre la actividad catalítica de lipasas. 17
1.4 Efecto de la concentración de la lipasa en el perfil temperatura/actividad. 19
1.5 Efecto de la concentración de enzima sobre la estabilidad térmica. 20
1.6 Enantioselectividad de lipasas a diferente concentración enzimática. 20
2. Adsorción de lipasas sobre proteínas hidrofóbicas: Hidrofobinas. 23
2.1 Inmovilización de hidrofobinas. 23
2.2 Inmovilización de lipasas sobre el soporte glioxil-agarosa-hidrofobina. 23
2.3 Efecto de la inmovilización sobre las propiedades catalíticas de las lipasas. 25
-Conclusiones 27

Índice ii
-Parte Experimental 29
1. Materiales 29
2.Ensayos de determinación de actividad enzimática. 29
2.1 Hidrólisis de p-nitrofenilésteres. 29
2.2 Hidrólisis del butirato de etilo. 30
3. Determinación de la concentración de proteína. 30
4. Purificación de lipasas. 30
5. Electroforesis en geles de poliacrilamida-SDS-PAGE. 31
6. Experimento de gel-filtración de lipasas purificadas. 31
7. Inmovilización de hidrofobina sobre glioxil-agarosa. 32
8. Inmovilización de lipasas sobre el soporte glioxil-agarosa-hidrofobina. 32
9. Inmovilización de HLL a diferentes concentraciones sobre soporte PEI-Sepabeads. 32
10. Reacciones de hidrólisis. 33
11. Ensayo de actividad-temperatura de las distintas preparaciones de lipasas. 33
12. Inactivación térmica. 34
CapCapCapCapítítítítulo 2. Estabilidad y actividad de diferentes derivados inmovilizados de lipasa: ulo 2. Estabilidad y actividad de diferentes derivados inmovilizados de lipasa: ulo 2. Estabilidad y actividad de diferentes derivados inmovilizados de lipasa: ulo 2. Estabilidad y actividad de diferentes derivados inmovilizados de lipasa: ventaja de estabilizar la conformación abierta. ventaja de estabilizar la conformación abierta. ventaja de estabilizar la conformación abierta. ventaja de estabilizar la conformación abierta.
-Introducción 35
1. Métodos de inmovilización de las distintas conformaciones de lipasas. 35
1.1 Adsorción interfacial sobre soportes hidrófobicos: inmovilización de la conformación abierta de lipasas.
35
1.2 Inmovilización de lipasas por unión covalente limitada. 36
1.2.1 Inmovilización de lipasa sobre soportes agarosa-bromocianógeno (BrCN). 36
1.2.2 Inmovilización de lipasas sobre soportes glutaraldehido-agarosa. 36
1.3 Inmovilización covalente multipuntual de lipasas sobre soporte glioxil-agarosa. 37
2. Lipasas de termófilos. 39
-Objetivos 41
----Resultados y Discusión 42

Índice Índice Índice Índice iiiiiiiiiiii
1. purificación de lipasas. 42
2. Inmovilización de las distintas lipasas: efecto sobre la actividad enzimática. 43
2.1 Inmovilización de CAL-B. 43
2.2 Inmovilización de CRL. 44
2.3 Inmovilización de MML. 45
2.4 Inmovilización de BTL2. 45
2.5 Inmovilización de TTL. 46
2.6 Inmovilización de TAL. 46
3. Efecto de la inmovilización en la estabilidad de los distintos derivados inmovilizados de diferentes lipasas
47
3.1 Estabilidad de CAL-B. 47
3.2 Estabilidad de CRL. 48
3.3 Estabilidad de MML. 50
3.4 Estabilidad de BTL2. 51
3.5 estabilidad de TAL. 52
4. Efecto de la inmovilización sobre la temperatura óptima de lipasas 53
5. Diferente comportamiento de lipasas termófilas tras la adsorción sobre octadecil-Sepabeads.
54
5.1 Efecto de la inmovilización en la lipasa de T. thermophilus (TTL) 55
5.2 Efecto de la inmovilización en la lipasa de T. aquaticus (TAL) 56
-Conclusiones 57
-Parte Experimental 58
1. Materiales. 58
2. Purificación de lipasas. 58
3. Electroforesis en gel poliacrilamida-SDS-PAGE. 59
4. Electroforesis en condiciones no desnaturalizantes (nativa). 59
5. Inmovilización de lipasas sobre distintos soportes. 59
5.1 Preparación de derivado inmovilizado por adsorción interfacial de las distintas lipasas.
59

Índice iv
5.2 Preparación de los derivados covalentes de lipasas. 60
5.2.1 Preparación del derivado agarosa-bromocianógeno (BrCN). 60
5.2.2 Inmovilización de lipasas sobre soportes glutaraldehido. 60
5.2.3 Inmovilización de lipasas sobre soporte glioxil-agarosa. 60
6. Ensayos de determinación de actividad. 61
6.1 Hidrólisis de Butirato de butilo. 61
7. Estabilidad de los diferentes derivados inmovilizados de lipasas. 61
8. Perfil de Temperatura-actividad enzimática de las diferentes preparaciones de lipasas. 61
Capítulo 3. Modulación de la enantioselectividad de diferentes lipasas mediante Capítulo 3. Modulación de la enantioselectividad de diferentes lipasas mediante Capítulo 3. Modulación de la enantioselectividad de diferentes lipasas mediante Capítulo 3. Modulación de la enantioselectividad de diferentes lipasas mediante ingeniería conformacional: resolución de derivados del ácido (±)ingeniería conformacional: resolución de derivados del ácido (±)ingeniería conformacional: resolución de derivados del ácido (±)ingeniería conformacional: resolución de derivados del ácido (±)----2222----hidroxifenilachidroxifenilachidroxifenilachidroxifenilacéééético.tico.tico.tico.
-Introducción 62
1. Ingeniería Conformacional de lipasas. 62
2. Inmovilización de lipasas con distintas orientaciones. 63
2.1 Adsorción interfacial sobre soportes hidrofóbicos. 63
2.2. Inmovilización covalente a través de pocas uniones. 63
2.3 Inmovilización mediante enlace covalente multipuntual. 63
2.4 Adsorción iónica de lipasas. 64
2.5 Inmovilización sobre soportes heterofuncionales: Adsorción física + unión covalente 64
3. Derivados del ácido 2-hidroxi-fenilacético (ácido mandélico) 66
-Objetivos 69
-Resultados y Discusión 70
1. Hidrólisis enantioselectiva de ésteres derivados del ácido mandélico catalizada por los derivados inmovilizados de la lipasa de Candida antarctica B (CAL-B).
71
1.1 Especificidad de los distintos derivados inmovilizados frente a distintos sustratos. 71
1.1.1 Efecto del pH sobre la actividad de los derivados inmovilizados frente a los compuestos quirales 1111 y 7777.
72
1.2 Enantioselectividad de los distintos derivados inmovilizados de CAL-B catalizando la hidrólisis de mandelato de metilo [(±)-1111].
73
1.2.1 Efecto de la temperatura sobre la enantioselectividad de los derivados de CAL-B. 74

Índice Índice Índice Índice vvvv
1.2.2 Efecto de la presencia de co-disolvente en el medio de reacción sobre el valor de E de los distintos derivados de CAL-B.
75
1.3 Hidrólisis enantioselectiva del ácido (±)-2-O-butiril-2-fenilacético [(±)-7777] catalizada por derivados inmovilizados de CAL-B.
76
2. Hidrólisis enantioselectiva de ésteres derivados del ácido mandélico catalizada por los derivados inmovilizados de la lipasa de Candida rugosa (CRL).
77
2.1. Especificidad de los distintos inmovilizados de CRL en la hidrólisis de varios sustratos 77
2.1.1. Efecto del pH sobre la actividad específica de los distintos derivados inmovilizados de CRL
78
2.2 Enantioselectividad de los derivados inmovilizados de CRL en la hidrólisis de (±)-ésteres derivados del ácido mandélico.
79
2.2.1. Influencia del pH sobre la enantioselectividad de los derivados de CRL 79
3. Hidrólisis enantioselectiva del ácido (±)-2-O-butiril-2-fenilacético [(±)-7777] catalizada por derivados inmovilizados de la lipasa de Mucor miehei (MML).
80
3.1 Actividad de los diferentes derivados inmovilizados de MML en la hidrólisis de (±)-7: 7: 7: 7: efecto de las condiciones experimentales.
80
3.2 Enantioselectividad de los diferentes derivados inmovilizados de MML en la hidrólisis de (±)-7.7.7.7.
81
3.2.1 Efecto de la temperatura sobre el valor de E de los distintos derivados de MML. 82
3.2.2 Efecto del pH del medio sobre la enantioselectividad de los derivados de MML 82
4. Hidrólisis enantioselectiva del ácido (±)-2-O-butiril-2-fenilacético [(±)-7777] catalizada por los distintos derivados inmovilizados de la lipasa de Bacillus thermocatenulatus (BTL2).
83
4.1 Efecto de las condiciones experimentales sobre la actividad de los derivados de BTL2 en la hidrólisis de (±)-7.7.7.7.
83
4.1.1 Efecto del pH sobre la actividad de los derivados de BTL2. 83
4.1.2 Efecto de la temperatura sobre la actividad de los derivados de BTL2. 84
4.2 Enantioselectividad de los diferentes derivados inmovilizados de BTL2 en distintas condiciones experimentales.
84
4.2.1 Influencia del pH sobre la enantioselectividad de los derivados. 85
4.2.2 efecto de la temperatura sobre el valor de E. 86
5. Hidrólisis enantioselectiva del ácido (±)-2-O-butiril-2-fenilacético [(±)-7777] catalizada por los distintos derivados inmovilizados de la lipasa de Thermus thermophilus (TTL).
87
5.1 Actividad específica de los distintos derivados inmovilizados de TTL. 87
5.2 Enantioselectividad de los distintos derivados inmovilizados de TTL catalizando la hidrólisis de (±)-7 7 7 7 a 25ºC.
88
6. Hidrólisis enantioselectiva del compuesto [(±)-7777] catalizada por derivados inmovilizados de la lipasa de Thermus aquaticus (TAL).
88

Índice vi
6.1 Actividad específica de los distintos derivados inmovilizados de TAL a 25ºC. 88
6.2 Enantioselectividad de los distintos derivados inmovilizados de TTL catalizando la hidrólisis de (±)-7 7 7 7 a 25ºC.
89
-Conclusiones 90
-Parte experimental 91
1. Materiales 91
2. Inmovilización de los distintos lipasas sobre los distintos soportes 91
3. Hidrólisis enzimática de los diferentes sustratos 92
4. Determinación del exceso enantiomérico y la Enantioselectividad 93
5. Síntesis del ácido (±)-2-O-butiril-2-fenilacético [(±)-7777] 94
Capitulo 4. Hidrólisis enantioselectiva en medios acuosos de nuevos compuestos Capitulo 4. Hidrólisis enantioselectiva en medios acuosos de nuevos compuestos Capitulo 4. Hidrólisis enantioselectiva en medios acuosos de nuevos compuestos Capitulo 4. Hidrólisis enantioselectiva en medios acuosos de nuevos compuestos precursores de la (S)precursores de la (S)precursores de la (S)precursores de la (S)----(+)(+)(+)(+)----Zopiclona Zopiclona Zopiclona Zopiclona catalizada por la lipasa de catalizada por la lipasa de catalizada por la lipasa de catalizada por la lipasa de Candida antarcticaCandida antarcticaCandida antarcticaCandida antarctica B B B B
-Introducción 95
-Objetivos 97
-Resultados y discusión 98
1. Resolución enantioselectiva del carbonato de vinilo [(±)-10101010] utilizando derivados inmovilizados de CAL-B.
99
2. Estudio de las mejores condiciones de reacción. 100
3. Hidrólisis enantioselectiva de [(±)-10101010] empleando una concentración de 10 mM. 102
4. Reversivilidad de la adsorción de CAL-B sobre octadecil-Sepabeads. 103
5. Resolución enzimática de nuevos precursores de la Zopiclona. 104
-Conclusiones 108
-Parte experimental 109
1. Materiales. 109
2. Síntesis de compuestos. 109
3. Hidrólisis enzimática de carbonatos precursores de la Zopiclona. 112
4. Determinación del exceso enantiomérico y la Enantioselectividad. 113
Capítulo 5. RCapítulo 5. RCapítulo 5. RCapítulo 5. Resolución enantioselectiva de precursores de fármacos catalizada por esolución enantioselectiva de precursores de fármacos catalizada por esolución enantioselectiva de precursores de fármacos catalizada por esolución enantioselectiva de precursores de fármacos catalizada por lipasas inmovilizadas .Síntesis de (lipasas inmovilizadas .Síntesis de (lipasas inmovilizadas .Síntesis de (lipasas inmovilizadas .Síntesis de (----))))----Paroxetina y (S)Paroxetina y (S)Paroxetina y (S)Paroxetina y (S)----EnalaprilEnalaprilEnalaprilEnalapril
-Introducción 115

Índice Índice Índice Índice viiviiviivii
1. Derivados de 4-arilpiperidinas. Síntesis de la (-)-Paroxetina. 115
2. Ácido 2-hidroxi-4-fenilbutanoico, precursor en la síntesis del (S)-Enalapril. 119
-Objetivos 121
-Resultados y Discusión 122
1. Resolución enantioselectiva de (±)-trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxilato de etilo [(±)]-19, 19, 19, 19, intermedio en la síntesis de la (-)-Paroxetina, catalizada por derivados inmovilizados de CAL-B.
122
1.1 Actividad específica de diferentes lipasas de origen microbiano frente a (±)-trans-19.19.19.19. 122
1.2 Actividad específica de los distintos derivados inmovilizados de CAL-B. 123
1.3 Búsqueda de las mejores condiciones de reacción. 124
1.4 Resolución enantioselectiva de (±)-trans 19 19 19 19 catalizada por los derivados inmovilizados de CAL-B.
126
1.5 Resolución a escala semi-preparativa de (±)-trans 19 19 19 19.... 127
2. Resolución enantioselectiva de (±)-2-hidroxi-4-fenilbutirato de etilo [(±)-2222], , , , precursor en la síntesis del (S)-Enalapril, catalizada por los derivados inmovilizados de la lipasa de Bacillus themocatenulatus (BTL2).
129
2.1 Actividad específica de los derivados de BTL2 en la hidrólisis de (±)-2222 129
2.2 Hidrólisis enantioselectiva de (±)-2 2 2 2 catalizada por los distintos derivados inmovilizados de BTL2.
130
2.3 Efecto de la concentración del disolvente sobre el valor de E. 131
2.4 Reversibilidad de la adsorción de BTL2 sobre octadecil-Sepabeads. 133
-Conclusiones 134
-Parte experimental 135
1. Compuestos. 135
2. Preparación de los derivados inmovilizados de lipasas. 135
3. Hidrólisis enzimática [(±)-trans-19191919] y (±)-2222.... 135
4. Determinación del exceso enantiomérico y la enantioselectividad. 136
Discusión general y coDiscusión general y coDiscusión general y coDiscusión general y conclusionesnclusionesnclusionesnclusiones 137
BibliografíaBibliografíaBibliografíaBibliografía 144

Introducción General
1
A. LipasasA. LipasasA. LipasasA. Lipasas
Las lipasas son enzimas con un gran interés en química orgánica ya que presentan
una amplia especificidad por sustratos muy diferentes, lo cual contrasta, en algunos casos,
con una elevada regio y enantioselectividad1-21. Estas propiedades han convertido a las lipasas
en las enzimas con mayor uso en biotransformaciones, por ejemplo en resolución de mezclas
racémicas7-13,17-18.
Sin embargo, a la hora de utilizar la lipasa como catalizador de una reacción de
química orgánica, es preciso considerar algunas peculiaridades de su mecanismo de acción.
De este modo, mediante técnicas cristalográficas, se han obtenido dos conformaciones
diferentes de lipasas.
Figura 1Figura 1Figura 1Figura 1. Distintas conformaciones de lipasas.
En medios acuosos homogéneos, la lipasa cristaliza con el centro activo totalmente
aislado del medio de reacción por una cadena polipeptídica, llamada tapadera (flat o lid)
(Figura 1a), haciendo inaccesible la entrada de los sustratos (conformación cerrada). Esta
cadena polipeptídica presenta en su cara interna una serie de residuos hidrofóbicos que
interaccionan con las zonas hidrofóbicas que rodean al sitio activo de las lipasas.
Sin embargo, cuando la enzima se cristaliza en presencia de sustratos de gran
tamaño, se obtiene una conformación totalmente distinta, lo que se denomina lipasa abierta,
en la cual la tapadera se ha desplazado, interaccionando por medio de puentes salinos,
puentes de hidrogeno etc. con otra zona de la superficie de la lipasa, dejando libre el centro
activo de la enzima (Figura 1b).
C ompletamenteC ompletamenteC ompletamenteC ompletamenteinacces ibleinacces ibleinacces ibleinacces ible
a los sus tratosa los sus tratosa los sus tratosa los sus tratos
Abierta en presenc iaAbierta en presenc iaAbierta en presenc iaAbierta en presenc iade sus tratos óde sus tratos óde sus tratos óde sus tratos ó
es truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicas
C ompletamenteC ompletamenteC ompletamenteC ompletamenteinacces ibleinacces ibleinacces ibleinacces ible
a los sus tratosa los sus tratosa los sus tratosa los sus tratos
Abierta en presenc iaAbierta en presenc iaAbierta en presenc iaAbierta en presenc iade sus tratos óde sus tratos óde sus tratos óde sus tratos ó
es truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicas
C ompletamenteC ompletamenteC ompletamenteC ompletamenteinacces ibleinacces ibleinacces ibleinacces ible
a los sus tratosa los sus tratosa los sus tratosa los sus tratos
C ompletamenteC ompletamenteC ompletamenteC ompletamenteinacces ibleinacces ibleinacces ibleinacces ible
a los sus tratosa los sus tratosa los sus tratosa los sus tratos
Abierta en presenc iaAbierta en presenc iaAbierta en presenc iaAbierta en presenc iade sus tratos óde sus tratos óde sus tratos óde sus tratos ó
es truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicas
Abierta en presenc iaAbierta en presenc iaAbierta en presenc iaAbierta en presenc iade sus tratos óde sus tratos óde sus tratos óde sus tratos ó
es truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicases truc turas hidrofóbicas
L idL idL idL id
L idL idL idL id
aaaa bbbb

Introducción General
2
Estos datos han permitido establecer que en el mecanismo catalítico de las lipasas
ocurre lo que se denomina activación interfacial de lipasas21-32. Según esta propuesta, en
sistemas homogéneos las lipasas se encuentra en una conformación cerrada, con el centro
activo bloqueado, las cuales en presencia de interfases hidrofóbicas se adsorben a la misma
desplazando la “tapadera” y originando la conformación abierta. Este mecanismo de acción
permite a las lipasas actuar en las interfases (Esquema 1). Evidentemente, dado que el
sustrato natural de las lipasas son las grasas y aceites, esta actividad en la interfase es un
requerimiento indispensable para la función biológica de las lipasas.
Esquema 1Esquema 1Esquema 1Esquema 1. Activación interfacial de lipasas con interfases hidrofóbicas.
Sin embargo, el hecho es que las lipasas en medio acuoso presentan actividad
catalítica, en muchos casos comparable a la de esterasas no lipásicas, lo que nos sugiere la
hipótesis de partida de la presente tesis doctoral: las lipasas en sistemas acuosos
homogéneos se encuentran en un cierto equilibrio entre la conformación cerrada y una/s
conformación/s abierta/s que permiten que las lipasas sean catalíticamente activas en
ausencia de interfases (Esquema 2). La posible existencia de ese equilibrio abre nuevas
posibilidades en el diseño y control de las biotransformaciones catalizadas por lipasas.
Esquema Esquema Esquema Esquema 2222. Equilibrio entre las distintas conformaciones de lipasas en medios acuosos
homogéneos
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
Interfase hidrofóbicaInterfase hidrofóbicaInterfase hidrofóbicaInterfase hidrofóbica
lidlidlidlidCentro activoCentro activoCentro activoCentro activo
Lipasa activadaLipasa activadaLipasa activadaLipasa activadainterfacialmenteinterfacialmenteinterfacialmenteinterfacialmente
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
Interfase hidrofóbicaInterfase hidrofóbicaInterfase hidrofóbicaInterfase hidrofóbica
lidlidlidlidCentro activoCentro activoCentro activoCentro activo
Lipasa activadaLipasa activadaLipasa activadaLipasa activadainterfacialmenteinterfacialmenteinterfacialmenteinterfacialmente
Lipasa abiertaLipasa abiertaLipasa abiertaLipasa abiertay activay activay activay activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
Lipasa abiertaLipasa abiertaLipasa abiertaLipasa abiertay activay activay activay activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbicazonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicazonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica

Introducción General
3
B. B. B. B. AAAAbsorción de las lipasas sobre estructuras hidrofbsorción de las lipasas sobre estructuras hidrofbsorción de las lipasas sobre estructuras hidrofbsorción de las lipasas sobre estructuras hidrofóóóóbicasbicasbicasbicas
Como ya hemos dicho anteriormente, la conformación abierta de las lipasas muestra
al medio un gran bolsillo hidrofóbico, constituido por: i) la cara interna del lid y ii) toda la zona
hidrofóbica que rodea al centro activo, que muestra una gran afinidad por la superficie
hidrofóbica del sustrato natural, las gotas de aceite y grasa emulsionadas o no.
Sin embargo, este gran bolsillo hidrofóbico hace que la conformación abierta de las
lipasas tenga una gran afinidad por cualquier tipo de superficie hidrofóbica. De hecho, se ha
descrito que las lipasas se adsorben a multitud de estructuras hidrofóbicas: gotas de grasa33,
soportes hidrofóbicos34-40, burbujas de aire41, otras proteínas42, etc (Esquema 3).
Esquema Esquema Esquema Esquema 3333. Adsorción de lipasas sobre distintas estructuras hidrofóbicas.
Esto puede generar una serie de problemas a la hora de estudiar las propiedades de
las lipasas, que podrían ser diferentes en su conformación abierta y cerrada.
En esta tesis doctoral nos centraremos en el análisis de dos posibles interacciones y
sus posibles implicaciones en cuanto al estudio de las lipasas (Esquema 3):
- Interacciones lipasa-lipasa. Si nuestra hipótesis es correcta y las lipasas en medios
homogéneos están en equilibrio entre la conformación abierta y la cerrada, sería posible que
dos formas abiertas interaccionasen entre si, implicando los grandes bolsillos hidrofóbicos
expuestos en la conformación de la lipasa abierta.
Lipasa abierta y activaLipasa abierta y activaLipasa abierta y activaLipasa abierta y activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
soportesoportesoportesoporte
Gota de grasaGota de grasaGota de grasaGota de grasa
Proteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbica
Lipasa abierta y activaLipasa abierta y activaLipasa abierta y activaLipasa abierta y activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbicazonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicazonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
soportesoportesoportesoportesoportesoportesoportesoporte
Gota de grasaGota de grasaGota de grasaGota de grasa
Proteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbicaProteína hidrofóbica

Introducción General
4
- Interacción de las conformaciones abiertas de la lipasa con proteínas hidrofóbicas
contaminantes de la preparación comercial.
C. C. C. C. Estabilización de lipasasEstabilización de lipasasEstabilización de lipasasEstabilización de lipasas
Disponer de derivados enzimáticos suficientemente estables en las condiciones de
operación es uno de los cuellos de botella en el diseño de biotransformaciones. En este caso,
en el que tenemos dos estructuras de la proteína, es interesante disponer de datos que
muestren cual de las dos conformaciones es la más estable, la conformación que tiene fijada
la estructura abierta ó la que se encuentra en equilibrio, mayoritariamente cerrada (Esquema
4).
Esquema Esquema Esquema Esquema 4444. Inmovilización de las distintas conformaciones de lipasas
Para ello podemos comparar diferentes derivados de lipasas, utilizando derivados
obtenidos por activación interfacial sobre soportes hidrofóbicos con otro tipo de derivados.
D. D. D. D. Modulación de las propiedades catalíticas de las lipasas.Modulación de las propiedades catalíticas de las lipasas.Modulación de las propiedades catalíticas de las lipasas.Modulación de las propiedades catalíticas de las lipasas.
Teniendo en cuenta los grandes cambios conformacionales que sufren las lipasas y el
gran número de fuerzas que se implican en el mantenimiento de la estructura de las
diferentes conformaciones, el centro activo de la lipasa parece ser fácilmente alterable, con lo
que se podría conseguir modular las propiedades de las lipasas con relativa facilidad.
En esta tesis doctoral este estudio de la modulación de las propiedades de las lipasas
ha sido el núcleo central. Para ello se han utilizado dos herramientas distintas:

Introducción General
5
Ingeniería del medio.Ingeniería del medio.Ingeniería del medio.Ingeniería del medio.
En el esquema 5 puede verse la complejidad que implica el gran cambio
conformacional que sufren las lipasas. Centrándonos en la conformación abierta, la estructura
de esta estará definida por la interacción de un gran bolsillo hidrofóbico bien con una gota de
substrato o bien con el medio de reacción, mientras que por otro lado la tapadera debe de
acomodarse mediante un cierto número de interacciones electroestáticas, puentes de
hidrogeno, etc.
Parece evidente que cualquier cambio en las condiciones del medio puede alterar de
forma significativa todo el balance de interacciones: los disolventes favoreciendo el bolsillo
hidrofóbico y fortaleciendo las interacciones electrostáticas, el pH alterando el tipo y cantidad
de interacciones entre el lid y el resto de la superficie de la proteína, la fuerza iónica haciendo
más desfavorable la presencia del bolsillo hidrofóbico y debilitando las interacciones del lid
con el resto de la proteína, etc. De esta forma, las condiciones experimentales deberían de
afectar de forma dramática no solo al equilibrio entre la conformación abierta y cerrada de la
lipasa, sino también a la forma exacta del centro activo y por lo tanto, a sus propiedades
catalíticas.
Esquema Esquema Esquema Esquema 5555. Efecto de las condiciones experimentales en el mecanismo de apertura y
cierre de la lipasa.
Interacciones por puentes de hidrógenoInteracciones por puentes de hidrógenoInteracciones por puentes de hidrógenoInteracciones por puentes de hidrógenoy puentes salinosy puentes salinosy puentes salinosy puentes salinos
Bolsillo hidrofóbicoBolsillo hidrofóbicoBolsillo hidrofóbicoBolsillo hidrofóbico
Interacciones hidrofóbicasInteracciones hidrofóbicasInteracciones hidrofóbicasInteracciones hidrofóbicas
pH, copH, copH, copH, co----disolventes, disolventes, disolventes, disolventes, tªtªtªtª
CambioCambioCambioCambioExperimentalesExperimentalesExperimentalesExperimentales
Interacciones electrostáticasInteracciones electrostáticasInteracciones electrostáticasInteracciones electrostáticas

Introducción General
6
Ingeniería del derivado Ingeniería del derivado Ingeniería del derivado Ingeniería del derivado
Nuestro grupo de investigación ha desarrollado diferentes metodologías que
permiten inmovilizar proteínas de forma controlada y dirigida a través de diferentes zonas de
su superficie (aquellas con mayor densidad de cargas positivas o negativas, las zonas mas
hidrofóbicas, con mayor densidad de histidinas, de lisinas, etc). Además, es posible,
controlando el grado de activación del soporte y las condiciones de inmovilización, controlar el
grado de unión enzima soporte y por lo tanto la rigidez de la zona inmovilizada. Finalmente,
mediante el uso de polímeros policatiónicos es posible generar un ambiente altamente
hidrofílico alrededor de la lipasa, lo cual puede dar lugar a una alteración en sus propiedades.
Teniendo en cuenta los grandes cambios conformacionales que implican la apertura
y cierre de las lipasas, el hecho de preparar derivados inmovilizados de lipasas, implicando
zonas más o menos alejadas del centro activo, confiriendo más o menos rigidez o alterando
el microambiente, podría también alterar en gran medida sus propiedades catalíticas e
incluso su interacción con el medio de reacción (Esquema 6). De esta forma, la hipótesis sería
que diferentes derivados de una misma lipasa podrían tener unas propiedades catalíticas
(especificidad, selectividad, etc) muy diferentes.
Esquema 6Esquema 6Esquema 6Esquema 6....Diferentes metodologías de inmovilización.

Objetivos Generales
7
El objetivo principal de esta Tesis Doctoral es la Ingeniería de las reacciones de
hidrólisis selectivas catalizadas por derivados inmovilizados de lipasas en sistemas macro-
acuosos. Intentamos optimizar las características funcionales de diferentes lipasas
inmovilizadas (actividad, selectividad y estabilidad) para poder tener muchas posibilidades de
resolver muchísimos procesos de hidrólisis regio- y estéreo-selectivas.
En este caso las lipasas inmovilizadas tienen que actuar (inmovilizadas en el interior
de la estructura porosa de los soportes) en un medio completamente libre de interfases
hidrofóbicas. Para estudiar este problema establecemos una primera hipótesis de partida: en
ausencia de interfases las lipasas se encuentran preferentemente en su forma cerrada e
inactiva pero en equilibrio con una pequeña fracción de moléculas abiertas y activas. A partir
de esta hipótesis y de las subsiguientes hipótesis también planteadas en la introducción nos
planteamos los siguientes sub-objetivos:
1.- Estudio del comportamiento de las lipasas solubles en medios completamente acuosos y
de su posible interacción con diferentes estructuras hidrofóbicas. De este modo pretendemos
optimizar el manejo de las lipasas solubles y el diseño de nuevos mejores protocolos de
inmovilización.
2.- Inmovilización y estabilización de lipasas mesófilas y termófilas para obtener derivados
muy estables en condiciones industriales de reacción (pe. temperaturas elevadas y presencia
de co-disolventes orgánicos para mejorar lo solubilidad de sustratos moderadamente
hidrofóbicos).
3.- Control y modificación de la selectividad de lipasas inmovilizadas mediante:
a.- el uso de lipasas de diferentes fuentes
b.- la preparación de diferentes tipos de derivados: derivados de la forma abierta,
derivados de la forma mayoritariamente cerrada, derivados unidos por unión covalente uni- o

Objetivos Generales
8
multi-puntual por diferentes regiones de la estructura de las lipasas, derivados rodeados de
diferentes micro-ambientes, etc.
c.- la utilización de diferentes condiciones experimentales: diferentes pHs, diferentes
temperaturas, diferentes concentraciones de co-disolventes orgánicos, etc. para modificar el
mecanismo de apertura de las lipasas.
Este último sub-objetivo (núcleo central de esta Tesis) se desarrollo utilizando inicialmente dos
sustratos ésteres quirales modelo:
- éster metilico del ácido R,S mandélico donde el carbono quiral se encontraba en la
parte donador de acilo del sustrato éster.
- R,S ácido 2-O-butiril-2-fenilacético donde el carbono quiral se encuentra en la parte
nucleófilo del sustrato éster.
Posteriormente se utilizaron diferentes esteres quirales de interés en química farmacéutica.

Capítulo 1
9
INTRODUCCIÓN INTRODUCCIÓN INTRODUCCIÓN INTRODUCCIÓN
Debido al complejo mecanismo catalítico de las lipasas, estas se encuentran en un
cierto equilibrio entre dos conformaciones, mayoritariamente desplazado hacia la
conformación cerrada en medio acuoso. Sin embargo, las lipasas en presencia de una
interfase hidrofóbica sufren lo que se conoce como la activación interfacial21-32, dando lugar a
la fijación de la conformación abierta sobre dicha interfase.
Esta activación interfacial puede ser promovida por diferentes estructuras
hidrofóbicas, como pueden ser la propia lipasa o bien proteínas hidrofóbicas (Esquema 7),
además de por gotas de grasa33, soportes hidrofóbicos34-40, gotas de aire41 o lipopolisacáridos42.
Esquema Esquema Esquema Esquema 7.7.7.7. Mecanismo de adsorción de lipasas sobre estructuras hidrofóbicas
1. Interacción Lipasa1. Interacción Lipasa1. Interacción Lipasa1. Interacción Lipasa----LipasaLipasaLipasaLipasa.
Teniendo en cuenta la gran superficie hidrofóbica que se genera en una lipasa tras la
apertura del lid (la molécula de lipasa como conformación abierta), exponiéndose las áreas
hidrofóbicas de alrededor del sitio catalítico además de la parte hidrofóbica del lid, sería
posible pensar que toda esta superficie actuase como una interfase para otra molécula de
lipasa próxima a ella en su conformación cerrada, produciéndose la activación interfacial de
esta segunda, generándose así una estructura dimérica a través de dos moléculas de lipasa
(Esquema 8).
L ipasa abierta y ac tivaL ipasa abierta y ac tivaL ipasa abierta y ac tivaL ipasa abierta y ac tiva
L ipasa cerradaL ipasa cerradaL ipasa cerradaL ipasa cerradae inac tivae inac tivae inac tivae inac tiva
Ac tivación Ac tivación Ac tivación Ac tivación Interfac ialInterfac ialInterfac ialInterfac ial
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofí licahidrofí licahidrofí licahidrofí lica
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbicazonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofí licahidrofí licahidrofí licahidrofí licazonazonazonazona hidrofí licahidrofí licahidrofí licahidrofí lica
soportesoportesoportesoporte
soportesoportesoportesoporte
Gota de grasaGota de grasaGota de grasaGota de grasa
hidrofobinahidrofobinahidrofobinahidrofobina

Interacción de lipasas con estructuras hidrofóbicas
10
Esquema 8Esquema 8Esquema 8Esquema 8.... Mecanismo propuesto para la agregación de lipasas.
De hecho, estructuras diméricas de varias lipasas han sido identificadas mediante
estudios de rayos X, mostrándose que estas pueden cristalizar como dímeros en sus
conformaciones abiertas43-45. Además, varios autores han presentado en la literatura la
existencia de formas agregadas inespecíficas de algunas lipasas (estructuras oligoméricas),
aunque esto era debido a las drásticas condiciones experimentales empleadas (p.e muy alta
fuerza iónica) 46-54.
De este modo es posible pensar que las lipasas pueden presentar una tendencia a
auto asociarse dando estructuras con propiedades diferentes que pueden promover ciertos
problemas en su caracterización bioquímica, ya que este hipotético dímero puede presentar
diferente estabilidad (el hipotético “dímero” podría estar formado por dos estructuras abiertas
de lipasas), actividad o enantioselectividad (presentando un centro activo de lipasa accesible,
aunque parcialmente bloqueado por la presencia de otro) (Esquema 8).
2. Interacción de lipasas con proteínas hidrofóbicas.2. Interacción de lipasas con proteínas hidrofóbicas.2. Interacción de lipasas con proteínas hidrofóbicas.2. Interacción de lipasas con proteínas hidrofóbicas.
Debido a que las preparaciones comerciales de lipasas suelen presentar impurezas,
además de las interacciones lipasa-lipasa estudiadas anteriormente, también pueden
producirse interacciones con proteínas del extracto, como por ejemplo proteínas hidrofóbicas.
Esto puede generar una serie de problemas a la hora de caracterizar las lipasas comerciales
ya que las propiedades catalíticas dependen del grado de pureza, por ejemplo debido a la
mayor o menor concentración de proteína hidrofóbica.
++++++++++++
LipasaLipasaLipasaLipasa cerradacerradacerradacerrada
LipasaLipasaLipasaLipasa abiertaabiertaabiertaabierta
++++++++++++
LipasaLipasaLipasaLipasa cerradacerradacerradacerrada
LipasaLipasaLipasaLipasa abiertaabiertaabiertaabierta

Capítulo 1
11
Sin embargo, si somos capaces de controlar y fortalecer esta interacción entre lipasa
y proteína hidrofóbica, podríamos transformar este aparente problema en una herramienta.
A modo de ejemplo, se escogió la proteína más hidrofóbica conocida, la hidrofobina,
en interacción con las lipasas.
Las hidrofobinas son proteínas presentes únicamente en hongos, en las paredes de
las hifas55, involucradas en la formación de estructuras aéreas confiriendo hidrofobicidad a la
superficie de los hongos en contacto con el aire56. Estas funciones, son posibles gracias a su
particular habilidad de auto asociarse con interfases hidrofílica-hidrofóbica formando
películas muy finas altamente amfipáticas (5-12 nm)57-58. Estos ensamblajes son insolubles en
SDS caliente, lo cual permite la extracción de la hidrofobina de las paredes de la célula.
El basidiomiceto Pleurotus ostreatus contiene al menos cinco genes diferentes de
hidrofobinas, los cuales han sido ailslados, clonados, secuenciados y mapeados59-61. Las
expresiones de cuatro de ellos (vmh1, vmh2, vmh3 y POH2) han sido encontradas en
crecimiento vegetativo mientras la expresión del otro gen (fbh1) se encontró durante la
formación específica del cuerpo frutal59. Las hidrofobinas específicas del crecimiento vegetativo
han sido purificadas de las paredes de la célula, aunque también se han aislado del líquido
del medio de cultivo. Examinando la secreción de hidrofobinas a lo largo del crecimiento del
hongo, se ha observado que las distintas hidrofobinas podrían tener diferentes roles a lo largo
del ciclo vital de los hongos.
Su carácter hidrofóbico y sus propiedades de auto asociación hacen muy difícil el
manejo de las hidrofobinas en forma libre, debido a que rápidamente agregan. Sin embargo,
la auto asociación de hidrofobina a superficies hidrofóbicas o hidrofílicas ha sido empleado
como una interesante propiedad para aplicaciones médicas y técnicas, junto con el uso de las
películas de hidrofobina para inmovilizar diversas proteínas o enzimas62-63.
3333. . . . CaracterísticasCaracterísticasCaracterísticasCaracterísticas físicofísicofísicofísico----químicasquímicasquímicasquímicas de las distintas lipasas. de las distintas lipasas. de las distintas lipasas. de las distintas lipasas.
En esta tesis se emplearan diferentes lipasas procedentes de levaduras u hongos las
cuales se encuentran bien caracterizadas.
La lipasa de Candida antarctica B (CAL-B) es una enzima con un pequeño bolsillo
hidrofóbico entorno al centro activo y aunque se considera carente de lid, Uppenberg y col 44
determinaron la existencia de una pequeña cadena polipeptídica en alfa-hélice, la cual puede

Interacción de lipasas con estructuras hidrofóbicas
12
ser considerado como un potencial lid. Presenta un peso molecular entorno a 33 kDa y un
punto isoeléctrico (pI) de 6. Esta enzima ha sido utilizada como catalizador en múltiples
aplicaciones en química fina64-65 .
La lipasa de Mucor miehei presenta un peso molecular de 29 kD 27; por su parte la
lipasa de Humicola lanuginosa (HLL) pesa entorno a los 30 kD 65 y la lipasa de Pseudomonas
fluorescens (PFL) presenta un peso molecular de 33 kDa. La lipasa de Candida rugosa (CRL)
presenta dos diferentes isoenzimas, una de ellas con un peso de 63 kD y la otra de 60 kD 66.
No se han descrito isoenzimas o isoformas para el resto de lipasas estudiadas. Además todas
estas enzimas se han empleado ampliamente en resolución de compuestos quirales 9,20.

Capítulo 1
13
OOOOBJETIVOSBJETIVOSBJETIVOSBJETIVOS
En este capítulo nos centraremos, teniendo en cuenta los grandes cambios
conformacionales implicados en el mecanismo de catálisis de estas enzimas, en el estudio de
la interacción de lipasas sobre diferentes estructuras hidrofobicas (otra molécula de lipasa,
proteínas hidrofóbicas) y el efecto que esto puede tener sobre las propiedades funcionales de
las moléculas de lipasas.

Interacción de lipasas con estructuras hidrofóbicas
14
RESULTADOS Y DISCUSIONRESULTADOS Y DISCUSIONRESULTADOS Y DISCUSIONRESULTADOS Y DISCUSION
1.1.1.1. Tendencia general de las lipasTendencia general de las lipasTendencia general de las lipasTendencia general de las lipasas a as a as a as a auto asociarseauto asociarseauto asociarseauto asociarse formando estructuras formando estructuras formando estructuras formando estructuras
bimoleculares: modificación de las propiedades bimoleculares: modificación de las propiedades bimoleculares: modificación de las propiedades bimoleculares: modificación de las propiedades funcionalesfuncionalesfuncionalesfuncionales....
1.1 Purificación de lipasas 1.1 Purificación de lipasas 1.1 Purificación de lipasas 1.1 Purificación de lipasas
Para estudiar la existencia de estructuras macromoleculares de las lipasas y su influencia
en las propiedades funcionales de la misma, es importante trabajar con lipasas puras. Por
tanto el primer paso, consistió en la purificación de las distintas lipasas seleccionadas para
este estudio (CAL-B, MML, CRL, HLL, PFL). Esta purificación se realizó a través de la adsorción
interfacial de las distintas lipasas sobre geles octil-agarosa a baja fuerza iónica, obteniéndose
una inmovilización cuantitativa y rápida como esta descrito en la literatura34, confirmándose
por electroforesis en condiciones desnaturalizantes la adsorción únicamente de la lipasa
sobre el soporte (Figura 2). La obtención de las lipasas puras en solución se describe en la
parte experimental al final de este capítulo.
Figura 2Figura 2Figura 2Figura 2. Electroforesis SDS. Electroforesis SDS. Electroforesis SDS. Electroforesis SDS----PAGE de las diferentes preparaciones de lipasas. PAGE de las diferentes preparaciones de lipasas. PAGE de las diferentes preparaciones de lipasas. PAGE de las diferentes preparaciones de lipasas. a)a)a)a) Línea 1.-Marcadores de bajo peso molecular. Línea 2.- Preparación comercial de PFL. Línea 3.- PFL adsorbida sobre octil-agarosa. Línea 4.- preparación comercial de HLL. Línea 5.- HLL adsorbida sobre octil-agarosa. Línea 6.- Preparación comercial de CAL-B, Línea.- 7. CAL-B adsorbida sobre octil-agarosa. b)b)b)b) Línea 1.-Marcadores de bajo peso molecular. Línea 2.- Preparación comercial de MML. Línea 3.- MML adsorbida sobre octil-agarosa. Línea 4.- Preparación comercial de CRL. Línea 5.- CRL adsorbida sobre octil-agarosa.
9494949466666666
45454545
30303030
20,120,120,120,1
1111 2222 3333 4444 55551111 2222 3333 4444 5555 6666 7777
PmPmPmPm ((((KDaKDaKDaKDa)))) aaaa bbbb9494949466666666
45454545
30303030
20,120,120,120,1
1111 2222 3333 4444 55551111 2222 3333 4444 55551111 2222 3333 4444 5555 6666 77771111 2222 3333 4444 5555 6666 777711111111 22222222 33333333 44444444 55555555 66666666 77777777
PmPmPmPm ((((KDaKDaKDaKDa)))) aaaa bbbb
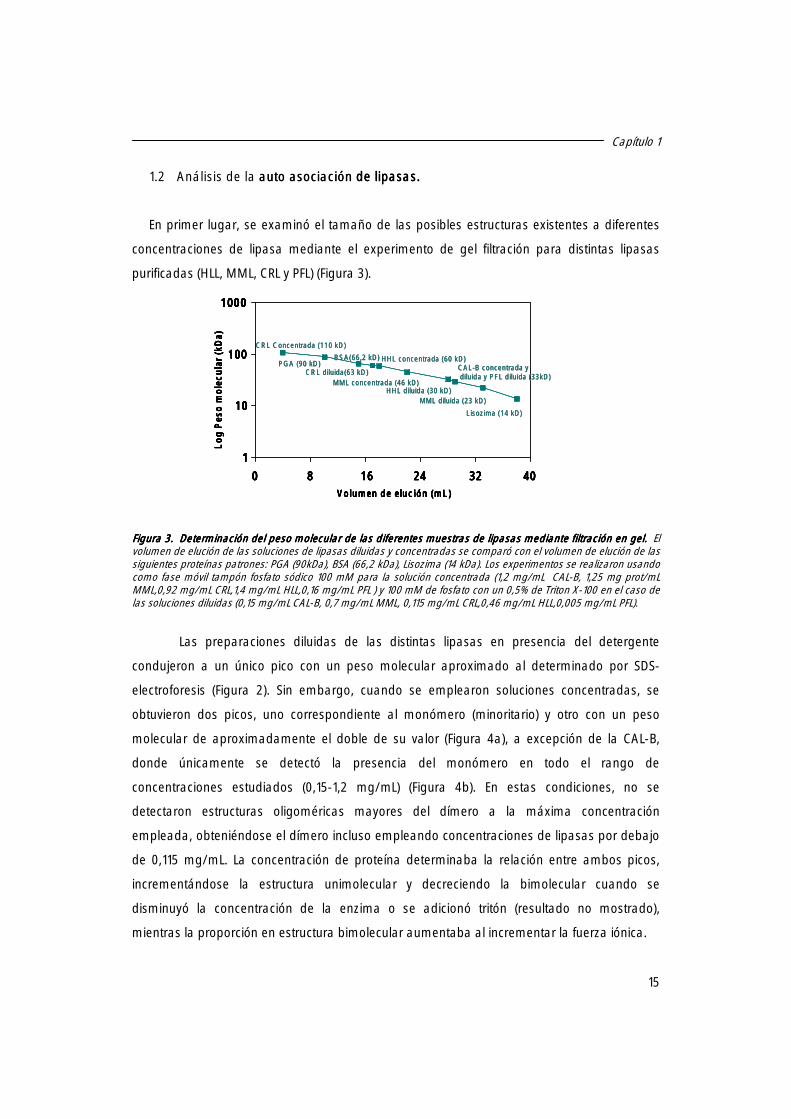
Capítulo 1
15
1.2 Análisis de la auto asociaciónauto asociación de lipasas. de lipasas. de lipasas.
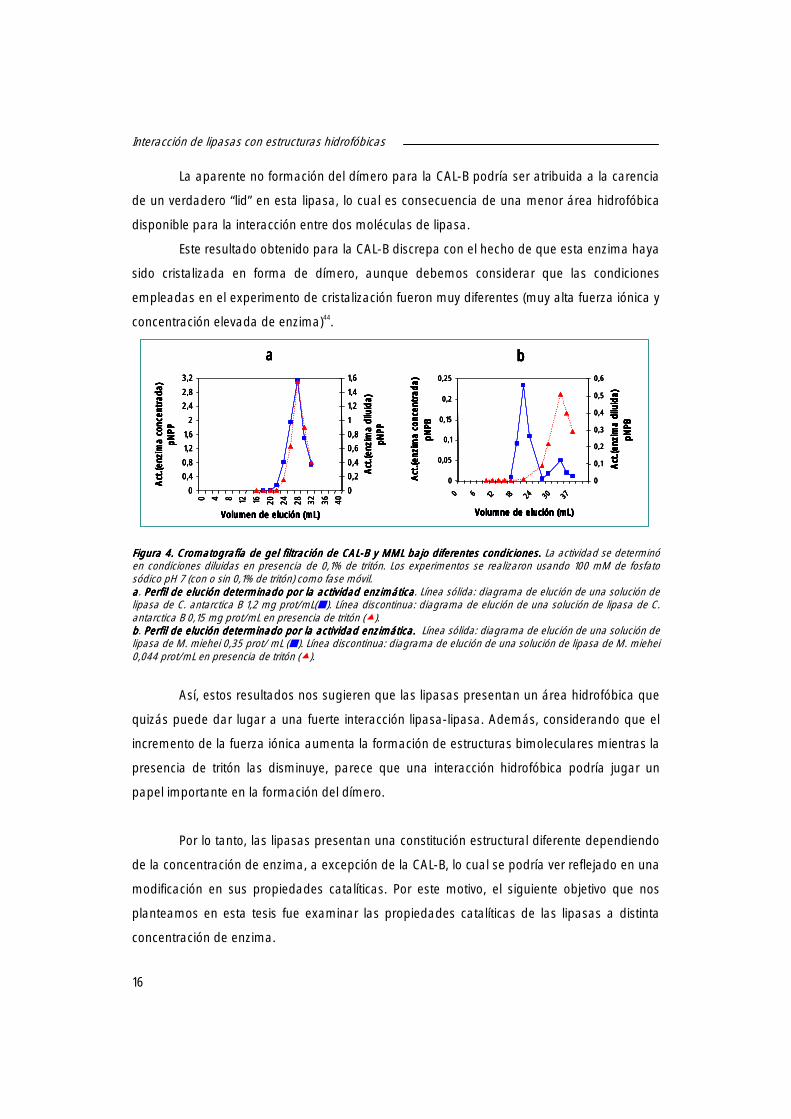
En primer lugar, se examinó el tamaño de las posibles estructuras existentes a diferentes
concentraciones de lipasa mediante el experimento de gel filtración para distintas lipasas
purificadas (HLL, MML, CRL y PFL) (Figura 3).
Figura 3Figura 3Figura 3Figura 3. . . . Determinación del peso molecular de las diferentes muestras de lipasas mediante filtración en gel. Determinación del peso molecular de las diferentes muestras de lipasas mediante filtración en gel. Determinación del peso molecular de las diferentes muestras de lipasas mediante filtración en gel. Determinación del peso molecular de las diferentes muestras de lipasas mediante filtración en gel. El volumen de elución de las soluciones de lipasas diluidas y concentradas se comparó con el volumen de elución de las siguientes proteínas patrones: PGA (90kDa), BSA (66,2 kDa), Lisozima (14 kDa). Los experimentos se realizaron usando como fase móvil tampón fosfato sódico 100 mM para la solución concentrada (1,2 mg/mL CAL-B, 1,25 mg prot/mL MML,0,92 mg/mL CRL,1,4 mg/mL HLL,0,16 mg/mL PFL ) y 100 mM de fosfato con un 0,5% de Triton X-100 en el caso de las soluciones diluidas (0,15 mg/mL CAL-B, 0,7 mg/mL MML, 0,115 mg/mL CRL,0,46 mg/mL HLL,0,005 mg/mL PFL).
Las preparaciones diluidas de las distintas lipasas en presencia del detergente
condujeron a un único pico con un peso molecular aproximado al determinado por SDS-
electroforesis (Figura 2). Sin embargo, cuando se emplearon soluciones concentradas, se
obtuvieron dos picos, uno correspondiente al monómero (minoritario) y otro con un peso
molecular de aproximadamente el doble de su valor (Figura 4a), a excepción de la CAL-B,
donde únicamente se detectó la presencia del monómero en todo el rango de
concentraciones estudiados (0,15-1,2 mg/mL) (Figura 4b). En estas condiciones, no se
detectaron estructuras oligoméricas mayores del dímero a la máxima concentración
empleada, obteniéndose el dímero incluso empleando concentraciones de lipasas por debajo
de 0,115 mg/mL. La concentración de proteína determinaba la relación entre ambos picos,
incrementándose la estructura unimolecular y decreciendo la bimolecular cuando se
disminuyó la concentración de la enzima o se adicionó tritón (resultado no mostrado),
mientras la proporción en estructura bimolecular aumentaba al incrementar la fuerza iónica.
1111
1 01 01 01 0
1 001 001 001 00
1 0001 0001 0001 000
0000 8888 1 61 61 61 6 24242424 32323232 40404040
CALC AL --B concentrada yB concentrada ydiluida y P F L diluida (33kD) diluida y P F L diluida (33kD)
L isozimaL isozima (1 4 ( 1 4 kDkD))
P GA (90 P GA (90 kDkD ))HHL concentrada (60 HHL concentrada (60 kDkD))
MML concentrada (46 MML concentrada (46 kDkD))HHL diluida (30 HHL diluida (30 kDkD))
CR L diluida(63 C R L diluida(63 kDkD ))
C R L C oncentrada (1 1 0 C R L C oncentrada (1 1 0 kDkD))
MML diluida (23 MML diluida (23 kDkD))
BS A(66,2 B S A(66,2 kDkD))
Volumen de Volumen de Volumen de Volumen de eluc ieluc ieluc ieluc ióóóónnnn (((( mLmLmLmL ))))
Lo
g P
eso
mol
ecu
lar
(L
og
Pes
o m
olec
ula
r (
Lo
g P
eso
mol
ecu
lar
(L
og
Pes
o m
olec
ula
r ( k
Da
kDa
kDa
kDa )) ))
1111
1 01 01 01 0
1 001 001 001 00
1 0001 0001 0001 000
0000 8888 1 61 61 61 6 24242424 32323232 40404040
1111
1 01 01 01 0
1 001 001 001 00
1 0001 0001 0001 000
0000 8888 1 61 61 61 6 24242424 32323232 40404040
CALC AL --B concentrada yB concentrada ydiluida y P F L diluida (33kD) diluida y P F L diluida (33kD)
L isozimaL isozima (1 4 ( 1 4 kDkD))
P GA (90 P GA (90 kDkD ))HHL concentrada (60 HHL concentrada (60 kDkD))
MML concentrada (46 MML concentrada (46 kDkD))HHL diluida (30 HHL diluida (30 kDkD))
CR L diluida(63 C R L diluida(63 kDkD ))
C R L C oncentrada (1 1 0 C R L C oncentrada (1 1 0 kDkD))
MML diluida (23 MML diluida (23 kDkD))
BS A(66,2 B S A(66,2 kDkD))
Volumen de Volumen de Volumen de Volumen de eluc ieluc ieluc ieluc ióóóónnnn (((( mLmLmLmL ))))
Lo
g P
eso
mol
ecu
lar
(L
og
Pes
o m
olec
ula
r (
Lo
g P
eso
mol
ecu
lar
(L
og
Pes
o m
olec
ula
r ( k
Da
kDa
kDa
kDa )) ))
Interacción de lipasas con estructuras hidrofóbicas
16
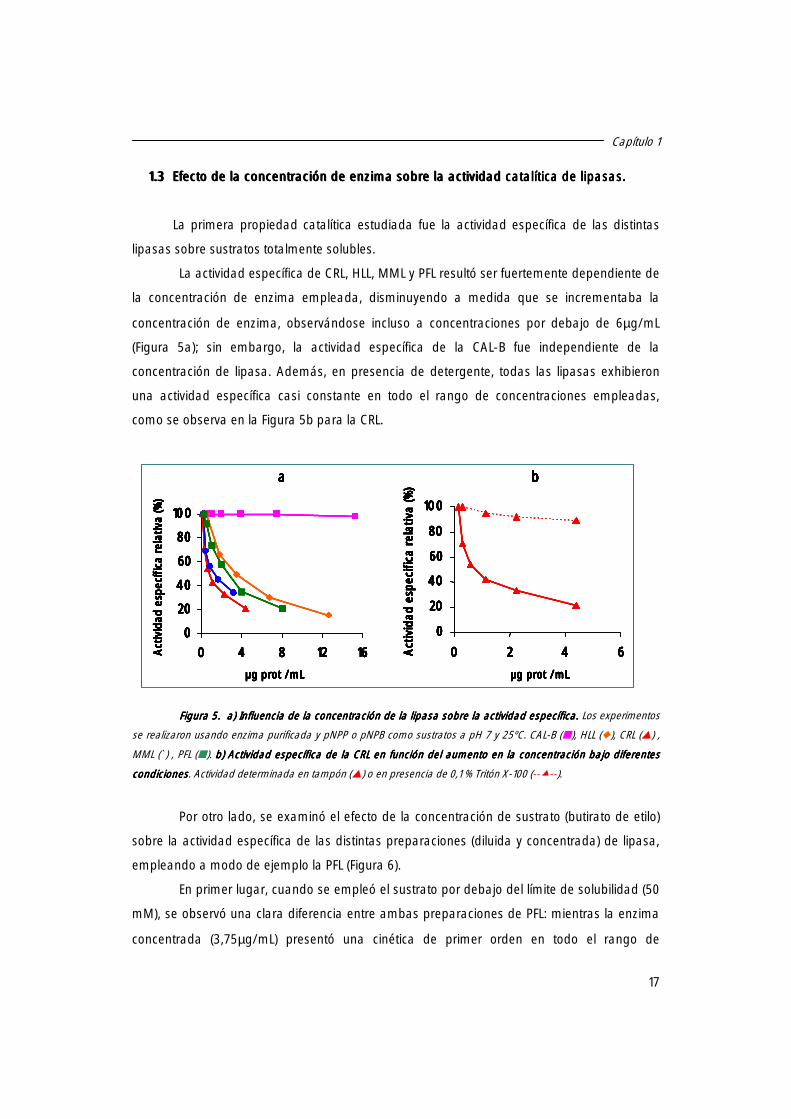
La aparente no formación del dímero para la CAL-B podría ser atribuida a la carencia
de un verdadero “lid” en esta lipasa, lo cual es consecuencia de una menor área hidrofóbica
disponible para la interacción entre dos moléculas de lipasa.
Este resultado obtenido para la CAL-B discrepa con el hecho de que esta enzima haya
sido cristalizada en forma de dímero, aunque debemos considerar que las condiciones
empleadas en el experimento de cristalización fueron muy diferentes (muy alta fuerza iónica y
concentración elevada de enzima)44.
Figura Figura Figura Figura 4444. Cromatografía de gel filtración de CAL. Cromatografía de gel filtración de CAL. Cromatografía de gel filtración de CAL. Cromatografía de gel filtración de CAL----B y MML bajo diferentes condiciones. B y MML bajo diferentes condiciones. B y MML bajo diferentes condiciones. B y MML bajo diferentes condiciones. La actividad se determinó en condiciones diluidas en presencia de 0,1% de tritón. Los experimentos se realizaron usando 100 mM de fosfato sódico pH 7 (con o sin 0,1% de tritón) como fase móvil. aaaa. Perfil de elución determinado por la actividad enzimáticaPerfil de elución determinado por la actividad enzimáticaPerfil de elución determinado por la actividad enzimáticaPerfil de elución determinado por la actividad enzimática. Línea sólida: diagrama de elución de una solución de lipasa de C. antarctica B 1,2 mg prot/mL(�). Línea discontinua: diagrama de elución de una solución de lipasa de C. antarctica B 0,15 mg prot/mL en presencia de tritón (�). bbbb. Perfil de elución determinado por la actividad enzimática. Perfil de elución determinado por la actividad enzimática. Perfil de elución determinado por la actividad enzimática. Perfil de elución determinado por la actividad enzimática. Línea sólida: diagrama de elución de una solución de lipasa de M. miehei 0,35 prot/ mL (�). Línea discontinua: diagrama de elución de una solución de lipasa de M. miehei 0,044 prot/mL en presencia de tritón (�).
Así, estos resultados nos sugieren que las lipasas presentan un área hidrofóbica que
quizás puede dar lugar a una fuerte interacción lipasa-lipasa. Además, considerando que el
incremento de la fuerza iónica aumenta la formación de estructuras bimoleculares mientras la
presencia de tritón las disminuye, parece que una interacción hidrofóbica podría jugar un
papel importante en la formación del dímero.
Por lo tanto, las lipasas presentan una constitución estructural diferente dependiendo
de la concentración de enzima, a excepción de la CAL-B, lo cual se podría ver reflejado en una
modificación en sus propiedades catalíticas. Por este motivo, el siguiente objetivo que nos
planteamos en esta tesis fue examinar las propiedades catalíticas de las lipasas a distinta
concentración de enzima.
bbbb
0
0,05
0,1
0,15
0,2
0,25
0 6 12 18 24 30 37
VolumneVolumneVolumneVolumne de de de de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPB
pNPB
pNPB
pNPB
0
0,1
0,2
0,3
0,4
0,5
0,6
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
pNPB
pNPB
pNPB
pNPB
aaaa
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
0 4 8 12 16 20 24 28 32 36 40
Volumen de Volumen de Volumen de Volumen de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPP
pNPP
pNPP
pNPP
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
pN
PPpN
PPpN
PPpN
PP
bbbb
0
0,05
0,1
0,15
0,2
0,25
0 6 12 18 24 30 37
VolumneVolumneVolumneVolumne de de de de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPB
pNPB
pNPB
pNPB
0
0,1
0,2
0,3
0,4
0,5
0,6
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
pNPB
pNPB
pNPB
pNPB
bbbb
0
0,05
0,1
0,15
0,2
0,25
0 6 12 18 24 30 37
VolumneVolumneVolumneVolumne de de de de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPB
pNPB
pNPB
pNPB
0
0,1
0,2
0,3
0,4
0,5
0,6
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
pNPB
pNPB
pNPB
pNPB
0
0,05
0,1
0,15
0,2
0,25
0 6 12 18 24 30 37
VolumneVolumneVolumneVolumne de de de de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPB
pNPB
pNPB
pNPB
0
0,1
0,2
0,3
0,4
0,5
0,6
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
pNPB
pNPB
pNPB
pNPB
aaaa
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
0 4 8 12 16 20 24 28 32 36 40
Volumen de Volumen de Volumen de Volumen de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPP
pNPP
pNPP
pNPP
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
pN
PPpN
PPpN
PPpN
PP
aaaa
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
0 4 8 12 16 20 24 28 32 36 40
Volumen de Volumen de Volumen de Volumen de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPP
pNPP
pNPP
pNPP
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
pN
PPpN
PPpN
PPpN
PP
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
0 4 8 12 16 20 24 28 32 36 40
Volumen de Volumen de Volumen de Volumen de elucielucielucielucióóóónnnn ((((mLmLmLmL))))
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
Act
.(enz
ima
conc
entr
ada)
A
ct.(e
nzim
a co
ncen
trad
a)
pNPP
pNPP
pNPP
pNPP
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
A
ct.(e
nzim
a di
luid
a)
Act
.(enz
ima
dilu
ida)
pN
PPpN
PPpN
PPpN
PP
Capítulo 1
17
1.31.31.31.3 Efecto de la concentración de enzima sobre la actividad Efecto de la concentración de enzima sobre la actividad Efecto de la concentración de enzima sobre la actividad Efecto de la concentración de enzima sobre la actividad catalítica de lipasascatalítica de lipasascatalítica de lipasascatalítica de lipasas....
La primera propiedad catalítica estudiada fue la actividad específica de las distintas
lipasas sobre sustratos totalmente solubles.
La actividad específica de CRL, HLL, MML y PFL resultó ser fuertemente dependiente de
la concentración de enzima empleada, disminuyendo a medida que se incrementaba la
concentración de enzima, observándose incluso a concentraciones por debajo de 6µg/mL
(Figura 5a); sin embargo, la actividad específica de la CAL-B fue independiente de la
concentración de lipasa. Además, en presencia de detergente, todas las lipasas exhibieron
una actividad específica casi constante en todo el rango de concentraciones empleadas,
como se observa en la Figura 5b para la CRL.
Figura Figura Figura Figura 5555. a) Influencia de la concentración de la lipasa sobre . a) Influencia de la concentración de la lipasa sobre . a) Influencia de la concentración de la lipasa sobre . a) Influencia de la concentración de la lipasa sobre la actividad especla actividad especla actividad especla actividad especíííífica. fica. fica. fica. Los experimentos
se realizaron usando enzima purificada y pNPP o pNPB como sustratos a pH 7 y 25ºC. CAL-B (�), HLL (�), CRL (�) ,
MML (` ) , PFL (�). b)b)b)b) Actividad especActividad especActividad especActividad especíííífica de la CRL en función del aumento en la concentración bajo fica de la CRL en función del aumento en la concentración bajo fica de la CRL en función del aumento en la concentración bajo fica de la CRL en función del aumento en la concentración bajo diferentes diferentes diferentes diferentes
condicionescondicionescondicionescondiciones. Actividad determinada en tampón (�) o en presencia de 0,1 % Tritón X-100 (--�--).
Por otro lado, se examinó el efecto de la concentración de sustrato (butirato de etilo)
sobre la actividad específica de las distintas preparaciones (diluida y concentrada) de lipasa,
empleando a modo de ejemplo la PFL (Figura 6).
En primer lugar, cuando se empleó el sustrato por debajo del límite de solubilidad (50
mM), se observó una clara diferencia entre ambas preparaciones de PFL: mientras la enzima
concentrada (3,75µg/mL) presentó una cinética de primer orden en todo el rango de
µg µg µg µg protprotprotprot /mL/mL/mL/mL
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 4444 8888 12121212 16161616Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fica
rel
ativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 4444 8888 12121212 16161616Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fica
rel
ativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
Act
ivid
ad e
spec
ifica
rel
ativ
a (%
) A
ctiv
idad
esp
ecifi
ca r
elat
iva
(%)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 4444 8888 12121212 16161616Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fica
rel
ativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
)
µg µg µg µg protprotprotprot /mL/mL/mL/mL
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 4444 8888 12121212 16161616Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fica
rel
ativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
)
0000
20202020
40404040
60606060
80808080
100100100100
0000 4444 8888 12121212 16161616Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fica
rel
ativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
) fic
a re
lativ
a (%
)
Interacción de lipasas con estructuras hidrofóbicas
18
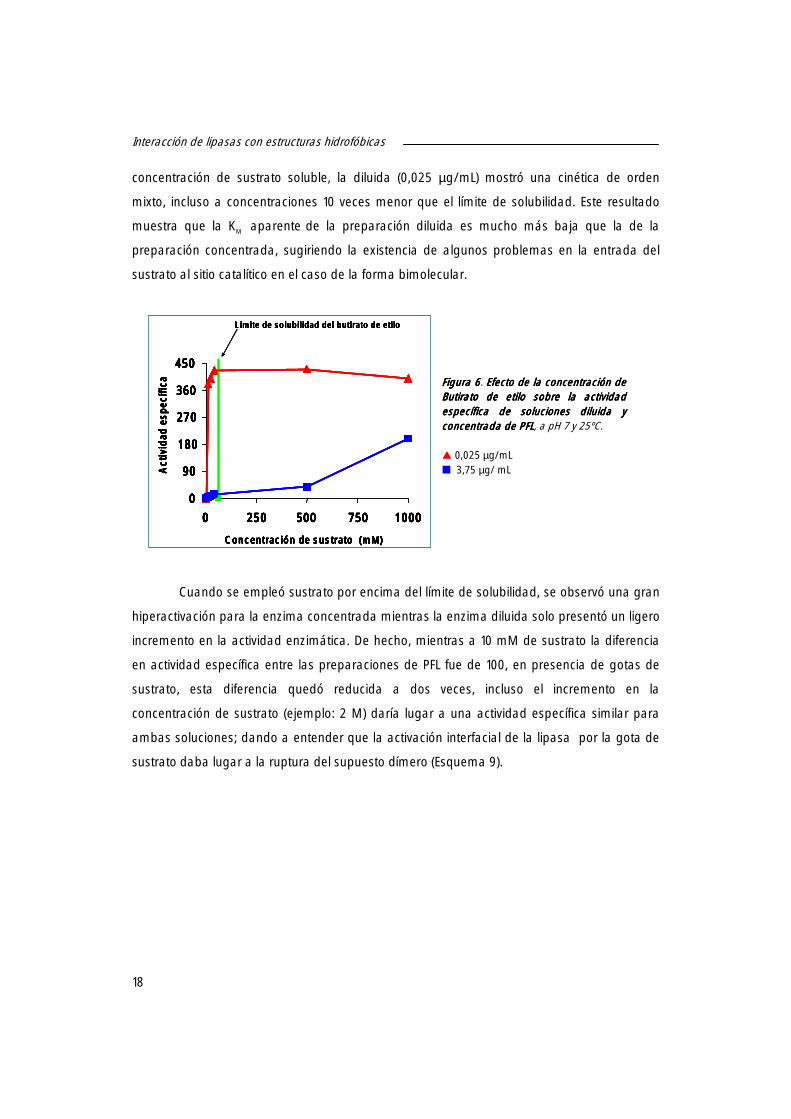
concentración de sustrato soluble, la diluida (0,025 µg/mL) mostró una cinética de orden
mixto, incluso a concentraciones 10 veces menor que el límite de solubilidad. Este resultado
muestra que la KM aparente de la preparación diluida es mucho más baja que la de la
preparación concentrada, sugiriendo la existencia de algunos problemas en la entrada del
sustrato al sitio catalítico en el caso de la forma bimolecular.
Figura Figura Figura Figura 6666. EEEEfecto de la confecto de la confecto de la confecto de la concentración de centración de centración de centración de Butirato de etiloButirato de etiloButirato de etiloButirato de etilo sobre la actividad sobre la actividad sobre la actividad sobre la actividad específica de soluciones diluida y específica de soluciones diluida y específica de soluciones diluida y específica de soluciones diluida y concentrada de PFLconcentrada de PFLconcentrada de PFLconcentrada de PFL, a pH 7 y 25ºC. � 0,025 µg/mL � 3,75 µg/ mL
Cuando se empleó sustrato por encima del límite de solubilidad, se observó una gran
hiperactivación para la enzima concentrada mientras la enzima diluida solo presentó un ligero
incremento en la actividad enzimática. De hecho, mientras a 10 mM de sustrato la diferencia
en actividad específica entre las preparaciones de PFL fue de 100, en presencia de gotas de
sustrato, esta diferencia quedó reducida a dos veces, incluso el incremento en la
concentración de sustrato (ejemplo: 2 M) daría lugar a una actividad específica similar para
ambas soluciones; dando a entender que la activación interfacial de la lipasa por la gota de
sustrato daba lugar a la ruptura del supuesto dímero (Esquema 9).
L imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti lo
0000
90909090
1 801 801 801 80
270270270270
360360360360
450450450450
0000 250250250250 500500500500 750750750750 1 0001 0001 0001 000
C onc entrac iC onc entrac iC onc entrac iC onc entrac ióóóón de s us trato (n de s us trato (n de s us trato (n de s us trato ( mMmMmMmM))))
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fic
afic
afic
afic
a
L imi te de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti lo
0000
90909090
1 801 801 801 80
270270270270
360360360360
450450450450
0000 250250250250 500500500500 750750750750 1 0001 0001 0001 000
C onc entrac iC onc entrac iC onc entrac iC onc entrac ióóóón de s us trato (n de s us trato (n de s us trato (n de s us trato ( mMmMmMmM))))
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fic
afic
afic
afic
a
L imi te de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti loL imite de solubil idad del butirato de eti lo
0000
90909090
1 801 801 801 80
270270270270
360360360360
450450450450
0000 250250250250 500500500500 750750750750 1 0001 0001 0001 000
C onc entrac iC onc entrac iC onc entrac iC onc entrac ióóóón de s us trato (n de s us trato (n de s us trato (n de s us trato ( mMmMmMmM))))
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fic
afic
afic
afic
a
0000
90909090
1 801 801 801 80
270270270270
360360360360
450450450450
0000 250250250250 500500500500 750750750750 1 0001 0001 0001 000
C onc entrac iC onc entrac iC onc entrac iC onc entrac ióóóón de s us trato (n de s us trato (n de s us trato (n de s us trato ( mMmMmMmM))))
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
Act
ivid
ad e
spec
íí íí fic
afic
afic
afic
a
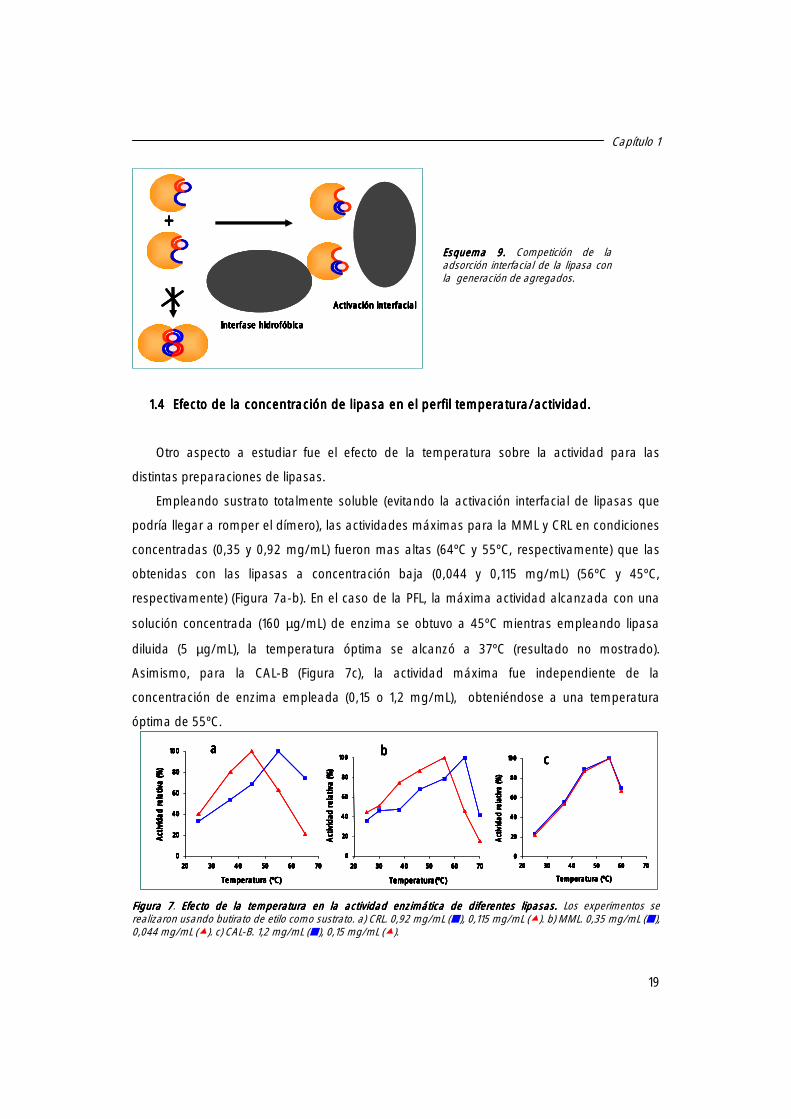
Capítulo 1
19
Esquema Esquema Esquema Esquema 9999.... Competición de la adsorción interfacial de la lipasa con la generación de agregados.
1.41.41.41.4 Efecto de la concentración de liEfecto de la concentración de liEfecto de la concentración de liEfecto de la concentración de lipasa pasa pasa pasa enenenen el perfil temperatura/actividad. el perfil temperatura/actividad. el perfil temperatura/actividad. el perfil temperatura/actividad.
Otro aspecto a estudiar fue el efecto de la temperatura sobre la actividad para las
distintas preparaciones de lipasas.
Empleando sustrato totalmente soluble (evitando la activación interfacial de lipasas que
podría llegar a romper el dímero), las actividades máximas para la MML y CRL en condiciones
concentradas (0,35 y 0,92 mg/mL) fueron mas altas (64ºC y 55ºC, respectivamente) que las
obtenidas con las lipasas a concentración baja (0,044 y 0,115 mg/mL) (56ºC y 45ºC,
respectivamente) (Figura 7a-b). En el caso de la PFL, la máxima actividad alcanzada con una
solución concentrada (160 µg/mL) de enzima se obtuvo a 45ºC mientras empleando lipasa
diluida (5 µg/mL), la temperatura óptima se alcanzó a 37ºC (resultado no mostrado).
Asimismo, para la CAL-B (Figura 7c), la actividad máxima fue independiente de la
concentración de enzima empleada (0,15 o 1,2 mg/mL), obteniéndose a una temperatura
óptima de 55ºC.
Figura Figura Figura Figura 7777. Efecto de la temperatura en la actividad enziEfecto de la temperatura en la actividad enziEfecto de la temperatura en la actividad enziEfecto de la temperatura en la actividad enzimática de diferentes lipasas.mática de diferentes lipasas.mática de diferentes lipasas.mática de diferentes lipasas. Los experimentos se realizaron usando butirato de etilo como sustrato. a) CRL. 0,92 mg/mL (�), 0,115 mg/mL (�). b) MML. 0,35 mg/mL (�), 0,044 mg/mL (�). c) CAL-B. 1,2 mg/mL (�), 0,15 mg/mL (�).
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
++++
InterfaseInterfaseInterfaseInterfase hidrofóbicahidrofóbicahidrofóbicahidrofóbica
ActivaciónActivaciónActivaciónActivación interfacialinterfacialinterfacialinterfacial
++++
InterfaseInterfaseInterfaseInterfase hidrofóbicahidrofóbicahidrofóbicahidrofóbica
ActivaciónActivaciónActivaciónActivación interfacialinterfacialinterfacialinterfacial
InterfaseInterfaseInterfaseInterfase hidrofóbicahidrofóbicahidrofóbicahidrofóbica
ActivaciónActivaciónActivaciónActivación interfacialinterfacialinterfacialinterfacial

Interacción de lipasas con estructuras hidrofóbicas
20
1.51.51.51.5 Efecto de la concentración de Efecto de la concentración de Efecto de la concentración de Efecto de la concentración de eeeenzima nzima nzima sobre la estabilidad térmica sobre la estabilidad térmica sobre la estabilidad térmica
En este caso se examinó la estabilidad de las lipasas a diferentes concentraciones a
50ºC (Figura 8). Se observó como cuando la concentración de la lipasa era mayor, aumentaba
su estabilidad en todos los casos, excepto para la CAL-B, donde la estabilidad fue
independiente de la concentración de enzima. En algunos casos, la diferencia de estabilidad
entre la preparación concentrada y diluida fue muy significativa; por ejemplo, la actividad de la
preparación diluida (0,09 mg/mL) de HLL (principalmente en forma monomérica) disminuyó
hasta el 40% en 20 horas, mientras la concentrada (0,74 mg/mL) (enriquecida en estructura
dimérica) mantuvo más del 80% de su actividad inicial después de 30 horas de incubación a
dicha temperatura. Además, al añadir un 0,6% de tritón (v/v) sobre la preparación
concentrada de HLL se determinó una estabilidad similar a la obtenida con la preparación
diluida, lo que refuerza la idea de que la interacción lipasa-lipasa en el dímero es hidrofóbica
y la presencia del tritón produce la ruptura de ese dímero.
Figura Figura Figura Figura 8888. Influencia de la concentración lipásica en la estabilidad de la enzima. Las inactivaciones se realizaron Influencia de la concentración lipásica en la estabilidad de la enzima. Las inactivaciones se realizaron Influencia de la concentración lipásica en la estabilidad de la enzima. Las inactivaciones se realizaron Influencia de la concentración lipásica en la estabilidad de la enzima. Las inactivaciones se realizaron a 50ºC y pH 7.a 50ºC y pH 7.a 50ºC y pH 7.a 50ºC y pH 7. a. H. lanuginosa. 0,74 mg/mL (�), 0,09 mg/mL(�) b. M.miehei. 0,35 mg/mL (�), 0,044 mg/mL(�) . c. C.antarctica B. 1,2 mg/mL (�), 0,15 mg/mL (�).
1.61.61.61.6 Enatioselectividad de lipasas a diferente concentración enzimática. Enatioselectividad de lipasas a diferente concentración enzimática. Enatioselectividad de lipasas a diferente concentración enzimática. Enatioselectividad de lipasas a diferente concentración enzimática.
La resolución de mezclas racémicas es una de las aplicaciones más interesantes de
las lipasas.
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030
Tiempo (horas) Tiempo (horas) Tiempo (horas) Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030
Tiempo (horas) Tiempo (horas) Tiempo (horas) Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030
Tiempo (horas) Tiempo (horas) Tiempo (horas) Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030
Tiempo (horas) Tiempo (horas) Tiempo (horas) Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030
Tiempo (horas) Tiempo (horas) Tiempo (horas) Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
0000
20202020
40404040
60606060
80808080
100100100100
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
Act
ivid
ad r
esid
ual %
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Capítulo 1
21
De este modo se analizó, en primer lugar, el efecto de la concentración de enzima
sobre la enantioselectividad de la PFL en la hidrólisis de 2-hidroxi-4-fenilbutanoato de etilo
totalmente soluble.
La preparación diluida presentó un moderado valor de enantioselectividad (E=7),
mientras la preparación concentrada resultó ser altamente enantioselectiva (E=100), al igual
que sucedía cuando se empleaba PFL adsorbida interfacialmente sobre un soporte octil-
agarosa67a.También esta descrito en la literatura que la enantioselectividad de la PFL fue mayor
cuando se encontraba adsorbida sobre gotas de sustrato (por encima del limite de
solubilidad, 6 mM en agua) que frente a sustrato soluble 67b.
Por tanto, teniendo en cuenta que el valor de E era similar para la enzima
concentrada o adsorbida sobre un soporte hidrofóbico (octil-agarosa), podemos concluir que
la estructura bimolecular realmente se forma como hemos descrito en el esquema 8.
1.6.1 Efecto de la concentración de enzima sobre la enantioselectividad de HLL.
Además, teniendo en cuenta los resultados obtenidos anteriormente, se examinó la
relevancia que podría tener este fenómeno de agregación incluso tras la inmovilización. Para
ello, se inmovilizaron soluciones de HLL con distinta concentración (0,09 y 0,74 mg lipasa/mL)
sobre un soporte Sepabeads recubierto con un polímero PEI (fuerte intercambiador aniónico
diseñado para inmovilizar proteínas por adsorción iónica)68, analizándose la
enantioselectividad en la resolución de varios esteres quirales [(±)-mandelato de metilo [(±)-1111] y
2-hidroxi-4-fenilbutanoato de etilo [(±)-2222]) mediante reacción de hidrólisis a 25ºC. La
inmovilización de la lipasa en el interior de los poros del soporte aseguró la interacción de la
enzima únicamente con sustrato soluble, evitándose así la posible activación interfacial de la
lipasa sobre gotas de sustrato, lo cual podría romper nuestro supuesto dímero cuando se
empleó una concentración de enzima de 0,74 mg/mL.
Así, se encontraron que los resultados obtenidos al utilizar el derivado inmovilizado de
HLL diluida o concentrada fueron muy diferentes (Tabla 1).En la hidrólisis del sustrato (±)-1111, la
lipasa inmovilizada bajo condiciones diluidas no presentó enantiopreferencia hacia ningún
enantiómero mientras bajo condiciones concentradas dio un exceso enantiomérico moderado
(15%). Sin embargo en la resolución del compuesto (±)-2222, la enzima inmovilizada bajo
condiciones diluidas presentó un exceso enantiomérico de solo un 4% mientras cuando se

Interacción de lipasas con estructuras hidrofóbicas
22
utilizó el derivado PEI de la enzima inmovilizada en condiciones concentradas se consiguió
obtener un exceso enantiomérico del 57% en las mismas condiciones de reacción.
Tabla 1. Efecto de la concentración de enzima durante la inmovilización sobre la enantioselectividad Tabla 1. Efecto de la concentración de enzima durante la inmovilización sobre la enantioselectividad Tabla 1. Efecto de la concentración de enzima durante la inmovilización sobre la enantioselectividad Tabla 1. Efecto de la concentración de enzima durante la inmovilización sobre la enantioselectividad
de HLL en la hidrólisis de diferentes ésteres racémicosde HLL en la hidrólisis de diferentes ésteres racémicosde HLL en la hidrólisis de diferentes ésteres racémicosde HLL en la hidrólisis de diferentes ésteres racémicos. aee= exceso enantiomérico al 50% de conversión, bE=enantioselectividad.
Por tanto, este resultado muestra como las propiedades de los derivados
inmovilizados pueden ser distintas cambiando las condiciones de la inmovilización (alterando
la relación monómero/dímero) (Esquema 10).
Esquema Esquema Esquema Esquema 10101010. Posible inmovilización/congelación de las diferentes estructuras pseudo-cuaternarias de la enzima mediante la adsorción iónica sobre soportes recubiertos con PEI.
Derivado Inmovilizado
Concentración de enzima en La inmovilización (mg/mL)
(1)(1)(1)(1) (2)(2)(2)(2)
Enantiómero Preferido
eea Eb Enantiómero Preferido
eea Eb
PEI-HLL 0,09 R 0 1 R 4 1,077
PEI-HLL 0,74 R 16 1,4 R 57,5 4
OH
COOMe
OH
COOMe
OH
COO -
+-( )-1 (R) -3H2O
+ (1)
OH
COOEt
OH
COOEt
OH
COO -
+-( )-2 (R) -4 (S) -4H2O
+ (2)
OH
COOMe
OH
COOMe
OH
COO -
+-( )-1 (R) -3H2O
+ (1)
OH
COOMe
OH
COOMe
OH
COO -OH
COO -
+-( )-1+-( )-1 (R) -3(R) -3H2OH2O
+ (1)
OH
COOEt
OH
COOEt
OH
COO -
+-( )-2 (R) -4 (S) -4H2O
+ (2)OH
COOEt
OH
COOEt
OH
COO -OH
COO -
+-( )-2+-( )-2 (R) -4(R) -4 (S) -4(S) -4H2OH2O
+ (2)
++++
SoporteSoporteSoporteSoporte PEIPEIPEIPEI
++++
SoporteSoporteSoporteSoporte PEIPEIPEIPEI
++++
SoporteSoporteSoporteSoporte PEIPEIPEIPEI

Capítulo 1
23
2222 Adsorción de lipasas sobre proteínas hidrofóbicAdsorción de lipasas sobre proteínas hidrofóbicAdsorción de lipasas sobre proteínas hidrofóbicAdsorción de lipasas sobre proteínas hidrofóbicas: Hidrofobinas.as: Hidrofobinas.as: Hidrofobinas.as: Hidrofobinas.
2.12.12.12.1 Inmovilización de hidrofobinasInmovilización de hidrofobinasInmovilización de hidrofobinasInmovilización de hidrofobinas
Las hidrofobinas son proteínas que presentan una alta razón de aminoácidos
hidrofóbicos respecto a los hidrofílicos56. Tienden a agregarse fácilmente, incluso a
concentraciones bajas dando agregados insolubles en agua. Además, el manejo de las
hidrofobinas purificadas en solución presenta grandes dificultades y requiere condiciones
estrictas y disolventes desnaturalizantes. Así, la mayoría de los protocolos de solubilización de
estas hidrofobinas no son compatibles con los protocolos de inmovilización, y el uso de co-
disolventes no fue suficiente para conseguir solubilizarlas, incluso empleando altas
concentraciones (75%).Sin embargo, las hidrofobinas fueron totalmente solubles en soluciones
que presentaban un alto porcentaje de detergente (tritón X-100). De este modo, se decidió
emplear una mezcla de tritón (40%) y dioxano (50%) manteniéndose la proteína dispersa
facilitando su inmovilización.
De este modo, la hidrofobina se inmovilizó sobre glioxil-agarosa mediante unión
covalente de la proteína al soporte69. La inmovilización de la hidrofobina procedió de forma
más lenta que para otras proteínas aunque prácticamente toda la hidrofobina ofrecida (0,5
mg/mL) se inmovilizó sobre el soporte en 24 horas.
2.22.22.22.2 InmInmInmInmovilización de lipasas sobre el soporte glioxilovilización de lipasas sobre el soporte glioxilovilización de lipasas sobre el soporte glioxilovilización de lipasas sobre el soporte glioxil----agarosaagarosaagarosaagarosa----hidrofobina.hidrofobina.hidrofobina.hidrofobina.
Cuando las lipasas se ofrecieron a glioxil-agarosa-hidrofobina a pH 7, la
inmovilización fue más lenta que frente a octil-agarosa34, alcanzándose hasta un 50% de
enzima inmovilizada entre 1 y 2 horas, observándose el efecto de la hiperactivación de la
enzima, aunque ese aumento de actividad fue menor frente a la glioxil-agarosa-hidrofobina
que cuando se empleó el octil-agarosa34 (Figura 9).
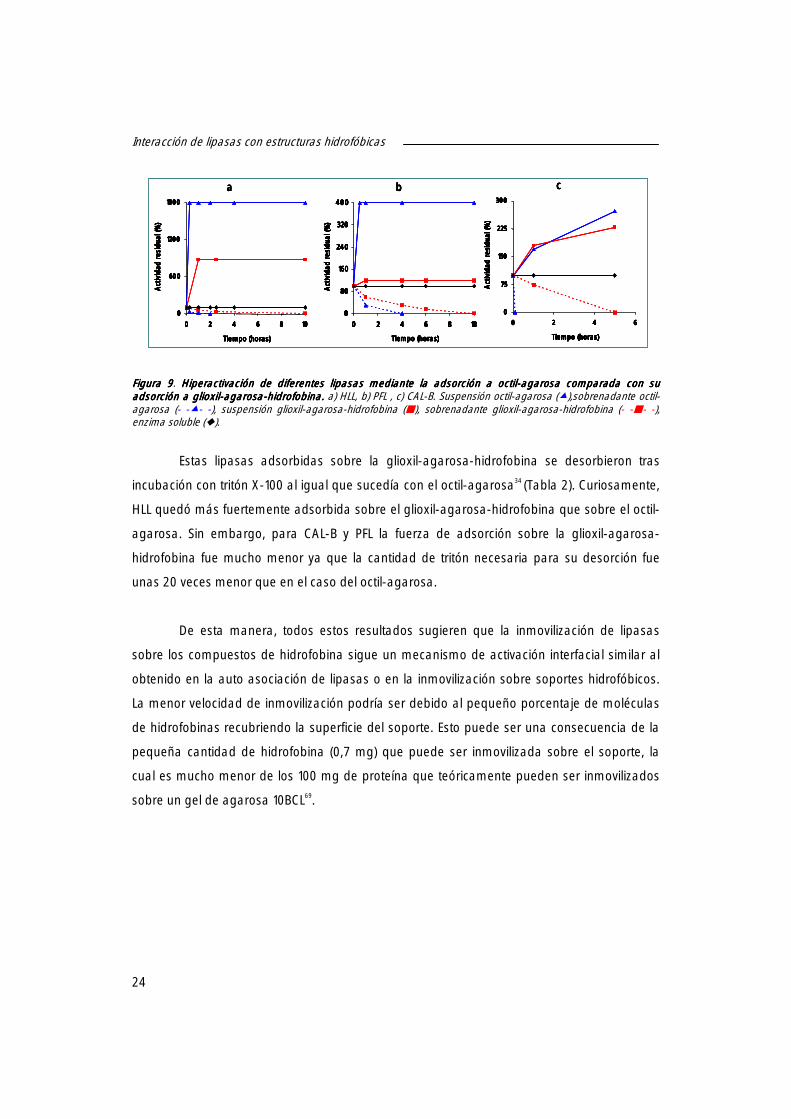
Interacción de lipasas con estructuras hidrofóbicas
24
Figura Figura Figura Figura 9999. Hiperactivación de difHiperactivación de difHiperactivación de difHiperactivación de diferentes lipasas mediante la adsorción a octilerentes lipasas mediante la adsorción a octilerentes lipasas mediante la adsorción a octilerentes lipasas mediante la adsorción a octil----agarosa comparada con su agarosa comparada con su agarosa comparada con su agarosa comparada con su adsorción a glioxiladsorción a glioxiladsorción a glioxiladsorción a glioxil----agarosaagarosaagarosaagarosa----hidrofobina.hidrofobina.hidrofobina.hidrofobina. a) HLL, b) PFL , c) CAL-B. Suspensión octil-agarosa (�),sobrenadante octil-agarosa (- -�- -), suspensión glioxil-agarosa-hidrofobina (�), sobrenadante glioxil-agarosa-hidrofobina (- -�- -), enzima soluble (�).
Estas lipasas adsorbidas sobre la glioxil-agarosa-hidrofobina se desorbieron tras
incubación con tritón X-100 al igual que sucedía con el octil-agarosa34 (Tabla 2). Curiosamente,
HLL quedó más fuertemente adsorbida sobre el glioxil-agarosa-hidrofobina que sobre el octil-
agarosa. Sin embargo, para CAL-B y PFL la fuerza de adsorción sobre la glioxil-agarosa-
hidrofobina fue mucho menor ya que la cantidad de tritón necesaria para su desorción fue
unas 20 veces menor que en el caso del octil-agarosa.
De esta manera, todos estos resultados sugieren que la inmovilización de lipasas
sobre los compuestos de hidrofobina sigue un mecanismo de activación interfacial similar al
obtenido en la auto asociación de lipasas o en la inmovilización sobre soportes hidrofóbicos.
La menor velocidad de inmovilización podría ser debido al pequeño porcentaje de moléculas
de hidrofobinas recubriendo la superficie del soporte. Esto puede ser una consecuencia de la
pequeña cantidad de hidrofobina (0,7 mg) que puede ser inmovilizada sobre el soporte, la
cual es mucho menor de los 100 mg de proteína que teóricamente pueden ser inmovilizados
sobre un gel de agarosa 10BCL69.
aaaa
0000
600600600600
1200120012001200
1800180018001800
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
80808080
160160160160
240240240240
320320320320
400400400400
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
75757575
150150150150
225225225225
300300300300
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
600600600600
1200120012001200
1800180018001800
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
600600600600
1200120012001200
1800180018001800
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
600600600600
1200120012001200
1800180018001800
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
80808080
160160160160
240240240240
320320320320
400400400400
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
80808080
160160160160
240240240240
320320320320
400400400400
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
80808080
160160160160
240240240240
320320320320
400400400400
0000 2222 4444 6666 8888 10101010
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
75757575
150150150150
225225225225
300300300300
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
75757575
150150150150
225225225225
300300300300
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
75757575
150150150150
225225225225
300300300300
0000 2222 4444 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Capítulo 1
25
Tabla 2. Fuerza de adsorción de lipasas sobre soportes Tabla 2. Fuerza de adsorción de lipasas sobre soportes Tabla 2. Fuerza de adsorción de lipasas sobre soportes Tabla 2. Fuerza de adsorción de lipasas sobre soportes hidrofóbicos (octilhidrofóbicos (octilhidrofóbicos (octilhidrofóbicos (octil----agarosa y glagarosa y glagarosa y glagarosa y glioxilioxilioxilioxil----agarosaagarosaagarosaagarosa----hidrofobina). hidrofobina). hidrofobina). hidrofobina). Los números expresan la cantidad de detergente necesario para desorber 100% de la lipasa del soporte. El análisis se llevo acabo midiendo la actividad enzimática de sobrenadantes y suspensiones en la hidrólisis de pNPP.
2.32.32.32.3 EfectoEfectoEfectoEfecto de la inmovilización sobre las propiedades catalíticas de las lipasas. de la inmovilización sobre las propiedades catalíticas de las lipasas. de la inmovilización sobre las propiedades catalíticas de las lipasas. de la inmovilización sobre las propiedades catalíticas de las lipasas.
En cuanto a la estabilidad, el derivado inmovilizado glioxil-agarosa-hidrofobina de CAL-B
presentó una estabilidad muy superior a la enzima soluble y algo mayor a la obtenida para el
derivado octil-agarosa-CAL-B (a 50ºC y pH 7) reteniendo hasta un 85% de la actividad inicial
después de 70 horas de incubación en estas condiciones (Figura 10).
Figura 10Figura 10Figura 10Figura 10. Estabilidad de . Estabilidad de . Estabilidad de . Estabilidad de diferentes diferentes diferentes diferentes derivados de CALderivados de CALderivados de CALderivados de CAL----B a B a B a B a 50ºC y pH 7.50ºC y pH 7.50ºC y pH 7.50ºC y pH 7. � glioxil-agarosa-hidrofobina � octil-agarosa � glioxil -agarosa � soluble
En cuanto a la enantioselectividad (Tabla 3), cuando se estudió la reacción de hidrólisis
del mandelato de metilo [(±)-1111] catalizada por derivados octil-agarosa y glioxil-agarosa-
hidrofobina de CAL-B se observó como estos presentaron valores de exceso enantiomérico al
50% de conversión muy similares [ee=65% para octil (E=9), 69% para glioxil-hidrofobina (E=11)],
mientras la enzima soluble mostró un valor muy bajo de enantioselectividad (E=3). Cuando se
Lipasa Soporte Tritón X-100(%)
CAL-B Octil-agarosa 1 Glioxil-agarosa-hidrofobina 0,05
PFL Octil-agarosa 1 Glioxil-agarosa-hidrofobina 0,05
HLL Octil-agarosa 0,5
Glioxil-agarosa-hidrofobina 1
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
80808080
90909090
100100100100
0000 20202020 40404040 60606060 80808080
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
% A
ctiv
idad
resi
dua
% A
ctiv
idad
resi
dua
% A
ctiv
idad
resi
dua
% A
ctiv
idad
resi
dua
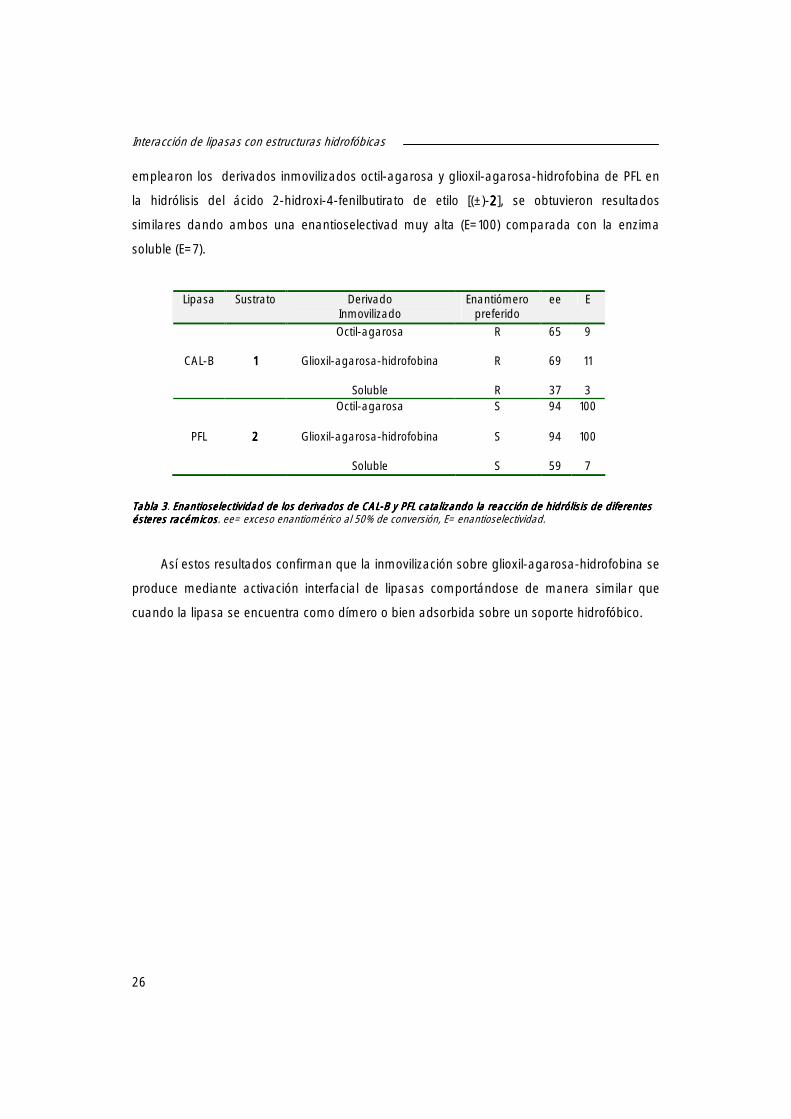
Interacción de lipasas con estructuras hidrofóbicas
26
emplearon los derivados inmovilizados octil-agarosa y glioxil-agarosa-hidrofobina de PFL en
la hidrólisis del ácido 2-hidroxi-4-fenilbutirato de etilo [(±)-2222], se obtuvieron resultados
similares dando ambos una enantioselectivad muy alta (E=100) comparada con la enzima
soluble (E=7).
Lipasa Sustrato Derivado Inmovilizado
Enantiómero preferido
ee E
Octil-agarosa
R 65 9
CAL-B 1111 Glioxil-agarosa-hidrofobina
R 69 11
Soluble R 37 3 Octil-agarosa
S 94 100
PFL 2222 Glioxil-agarosa-hidrofobina
S 94 100
Soluble S 59 7
Tabla 3Tabla 3Tabla 3Tabla 3. Enantioselectividad de los derivados de CALEnantioselectividad de los derivados de CALEnantioselectividad de los derivados de CALEnantioselectividad de los derivados de CAL----B y PFL catalizando la reacción de hidrólisis B y PFL catalizando la reacción de hidrólisis B y PFL catalizando la reacción de hidrólisis B y PFL catalizando la reacción de hidrólisis de diferentes de diferentes de diferentes de diferentes ésteres racémicosésteres racémicosésteres racémicosésteres racémicos. ee= exceso enantiomérico al 50% de conversión, E= enantioselectividad.
Así estos resultados confirman que la inmovilización sobre glioxil-agarosa-hidrofobina se
produce mediante activación interfacial de lipasas comportándose de manera similar que
cuando la lipasa se encuentra como dímero o bien adsorbida sobre un soporte hidrofóbico.

Capítulo 1
27
CONCLUSIOCONCLUSIOCONCLUSIOCONCLUSIONESNESNESNES
Los resultados obtenidos muestran como incluso a moderadas concentraciones, la
mayoría de lipasas en solución exhiben una fuerte tendencia a auto asociarse dando una
mezcla de estructuras diméricas y monoméricas que puede complicar la caracterización de
las propiedades intrínsecas de la molécula de la lipasa soluble.
De hecho, esta tendencia provocó importantes cambios en las propiedades de la
mayoría de lipasas: parece que las propiedades de la enzima dependen de esta estructura
pseudo-cuaternaria de la lipasa.
Algunas evidencias como los cambios en las propiedades catalíticas, el hecho de que
únicamente se detectan estructuras bimoleculares como agregados en las condiciones de alta
concentración de lipasa, la generalidad de esta tendencia en las lipasas con lid, la aparente
naturaleza hidrofóbica de la interacción y la competición entre la auto asociación y la
activación interfacial, sugieren que el mecanismo mostrado en el Esquema 8 es el mejor para
explicar esta auto asociación entre lipasas constituyendo agregados bimoleculares.
Por lo tanto, las dramáticas alteraciones estructura-función de la lipasa soluble con la
concentración de la enzima debería ser considerado en cualquier estudio que implique el
empleo de lipasas (incluso en preparaciones inmovilizadas), siendo posible la elección de
lipasa inmovilizada como dímero o bien como monómero según el protocolo de manipulación
de esta (Esquema 10).
Teniendo en cuenta que esa interacción es de naturaleza hidrofóbica y la presencia
de tritón da lugar a la ruptura del dímero, es completamente necesaria la presencia de este
detergente para asegurar la estructura monomérica de la lipasa.
Además la interacción de lipasas sobre proteínas hidrofóbicas (hidrofobinas),lo cual a
priori se plantea como un problema, se ha transformado en una herramienta ya que la
interacción de lipasas con superficies hidrofóbicas provoca una alteración en sus propiedades
lo cual dependerá del grado de pureza y de la concentración de ésta. La inmovilización de la
hidrofobina sobre un soporte covalente permite adsorber las lipasas selectivamente a través
de un mecanismo similar al observado (activación interfacial) cuando se empleó un soporte
hidrofóbico más convencional (octil-agarosa) o tras la formación del dímero para la mayoría
de lipasas. En este caso, las moléculas hidrofóbicas de hidrofobina deberían actuar como
gotas hidrofóbicas provocando la activación de la lipasa. Así se observó una hiperactivación y

Interacción de lipasas con estructuras hidrofóbicas
28
las propiedades de estabilidad y enantioselectividad de las lipasas en este soporte fueron
similares a las presentadas cuando se inmovilizaron sobre los soportes hidrofóbicos.
Por lo tanto, los resultados encontrados sugieren como la interacción de lipasas sobre
distintas estructuras hidrofóbicas transcurre a través de un mecanismo de activación
interfacial, fijándose la conformación abierta de la enzima a dichas estructuras.

Capítulo 1
29
PARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTAL 1.1.1.1. MaterialesMaterialesMaterialesMateriales
Las lipasas de Candida antarctica B (Novozym 525 L) (CAL-B), Mucor miehei (MML)
(Novozym 388) y Humicola lanuginosa (Novozym 871) (HLL) fueron suministradas por la
compañía Novo-Nordisk (Dinamarca). La Lipasa de Candida rugosa (CRL) (Type VII) fue de
Sigma Chemical Co. La lipasa de Pseudomonas fluorescens, PS”Amano” lot. LPSA21250457
(PFL) fue obtenida de Amano enzyme Inc. (NAGOYA, Japon). Las hidrofobinas (purificadas de
micela vegetativa de basidiomiceta P. ostreatus) fueron donadas por Lucía Ramírez
(departamento de producción agraria, Universidad Publica de Navarra, Pamplona).
El gel de agarosa 10% entrecruzado (BCL) fue donado por la compañía Hispanagar SA
(Burgos). Octil-agarosa 4BCL, DEAE-agarosa y los marcadores de peso molecular fueron de
Pharmacia Biotech (Uppsala). Octadecil-Sepabeads fue amablemente donado por Resindion
srl (Milán, Italia). Además polietilenimina (PEI) (60 kDa), Tritón X-100, butirato de etilo, p-
nitrofenil propionato (pNPP), p-nitrofenil butirato (pNPB) y (±)-mandelato de metilo [(±)-1111] se
obtuvieron de sigma. La mezcla racémica de 2-hidroxi-4-fenilbutanoato de etilo [(±)-2222] fue
donada por Vita Invest (S.A). Los soportes PEI-Sepabeads y glioxil-agarosa se prepararon
como esta descrito en la literatura 68-69.
2. Ensayos de determinación de actividad enzimática. 2. Ensayos de determinación de actividad enzimática. 2. Ensayos de determinación de actividad enzimática. 2. Ensayos de determinación de actividad enzimática.
2.1 Hidrólisis de p-nitrofenilesteres
El ensayo se llevó a cabo midiendo el aumento en el valor de absorbancia a 348 nm
producido por la formación de p-nitrofenol en la hidrólisis de 0,4 mM de pNPP o pNPB en 25
mM de tampón fosfato sódico a pH 7 y 25ºC. Para iniciar la reacción, 0,05 mL de solución
lipásica o suspensión se añadió a 2,5 mL de sustrato. Una unidad internacional de actividad
de pNPP o pNPB se define como la cantidad de enzima necesaria para hidrolizar 1 µmol de
pNPP o pNPB por minuto (IU) bajo las condiciones descritas anteriormente.

Interacción de lipasas con estructuras hidrofóbicas
30
2.2 Hidrólisis del butirato de etilo.
Este ensayo se realizó empleando un pHstato Mettler Toledo D50, midiendo el ácido
butírico producido por la hidrólisis enzimática del butirato de etilo a diferentes concentraciones
en tampón fosfato sódico 50 mM a pH 7 y 25ºC. El hidróxido sódico empleado como agente
de titulación fue de una concentración de 50 mM.
3. Determinación de la concentración de proteína.3. Determinación de la concentración de proteína.3. Determinación de la concentración de proteína.3. Determinación de la concentración de proteína.
La concentración proteica de la preparación comercial de las diferentes lipasas se
determinó utilizando el método de Bradford70. La curva de calibración se obtuvo con
seroalbúmina bovina (BSA) para determinar las concentraciones de proteína en un rango
comprendido entre 0,1 y 1,5 mg/mL. Así CAL-B presentó 12 mg proteína/mL, HLL contenía 7,2
mg/mL, MML 3,5 mg/mL, CRL presentó 9,2 mg/mL (en una concentración de 50 mg de
polvo/mL) y PFL 0,155 mg/mL (a una concentración de 25 mg/mL de polvo)
4444. Purificación de lipasas.. Purificación de lipasas.. Purificación de lipasas.. Purificación de lipasas.
La metodología general empleada para purificar lipasas de otras posibles proteínas
presentes en el extracto (por ejemplo esterasas) fue la cromatografía por adsorción interfacial
sobre geles octil-agarosa34 a muy baja fuerza iónica (5 mM) y pH 7 promoviendo así la
inmovilización sobre el gel únicamente de la lipasa de interés. Periódicamente, muestras de
sobrenadante y suspensión fueron recogidas y analizadas por ensayo de actividad pNPP o
pNPB. Siguiendo este protocolo se obtuvo una inmovilización cuantitativa de la actividad de la
lipasa.
Las lipasas adsorbidas se lavaron con tritón X-100 (1% para CAL-B, 0,5% para CRL,
0,6% para MML, 2% para HLL y 2% para PFL) en tampón fosfato sódico 5 mM y pH 7 siendo así
desorbidas. La mezcla enzima/detergente se diluyó adsorbiéndose a DEAE-agarosa a pH 8.0,
y fue lavada con agua destilada para eliminar todo el detergente. Después de esto, la enzima
adsorbida se extrajo del soporte empleando 200 mM de fosfato sódico a pH 8, dializado
frente a una solución de fosfato sódico 5 mM pH 7 y concentrado utilizando la ultra filtración
con tubos Centricon™ (Millipore).

Capítulo 1
31
5. Electrofores5. Electrofores5. Electrofores5. Electroforesis en geles de poliacrilamidais en geles de poliacrilamidais en geles de poliacrilamidais en geles de poliacrilamida----SDSSDSSDS PAGEPAGEPAGE
El análisis de la enzima adsorbida en el derivado fue confirmado por electroforesis
SDS-PAGE mostrando una única banda con peso molecular correspondiente a la lipasa nativa.
La técnica utilizada fue la descrita por Laemmli71, utilizando geles de poliacrilamida en
placa (100×75×1 mm) a una concentración del 12%.
Las muestras fueron tratadas durante 5-10 minutos a 100ºC en presencia de tampón
de ruptura (Tris-HCl 62,5 mM pH 6,8 – SDS 2% -βmercaptoetanol 5% - glicerol 10% y azul de
bromofenol al 0,005%).
La electroforesis se realizó a temperatura ambiente y 150 mV de corriente constante
utilizando como electrolito Tris-HCl 25 mM-glicina 192 mM y SDS 1%.
La visualización de las proteínas se realizó mediante tinción con azul brillante de Coomassie R-
25072.
Se utilizaron marcadores de bajo peso molecular: fosforilasa b (94 KD), BSA (66,2 KD),
ovoalbúmina (45 KD), anhidrasa carbónica (30 KD) e inhibidor de tripsina (20,1 KD).
6666. Experimento de gel. Experimento de gel. Experimento de gel. Experimento de gel----filtración de lipasas purififiltración de lipasas purififiltración de lipasas purififiltración de lipasas purificadas.cadas.cadas.cadas.
Los análisis de gel-filtración se realizaron en una columna de dimensiones 10 mm x
510 mm empaquetada con gel agarosa 10 BCL con un volumen total de 40 mL. El eluyente
empleado fue tampón fosfato sódico 100 mM (ocasionalmente con 0,1% tritón X-100) a pH 7 y
los experimentos se realizaron a 25ºC y a un flujo de 0,25 mL/min. La columna se equilibró
previamente pasando 400 mL del correspondiente tampón. En todos los casos se cargo 0,2
mL de enzima a diferentes concentraciones. Las muestras se fraccionaron en alícuotas de 1 mL
analizándose posteriormente su actividad mediante el ensayo con pNPP o pNPB empleando
muy baja concentración de enzima y 0,1% de tritón para evitar así la agregación de la enzima
durante la determinación de la actividad. La masa molecular de las preparaciones de las
distintas lipasas se estimó mediante la calibración empleando proteínas de peso molecular
conocido de referencia [PGA (90 kD), BSA (66,2 KD) y Lisozima (14 kD)].

Interacción de lipasas con estructuras hidrofóbicas
32
7777. Inmovilización . Inmovilización . Inmovilización . Inmovilización de hidrofobinas sobre glioxilde hidrofobinas sobre glioxilde hidrofobinas sobre glioxilde hidrofobinas sobre glioxil----agarosaagarosaagarosaagarosa....
Para inmovilizar las moléculas de hidrofobina sobre la superficie del soporte de manera
dispersa, una pequeña cantidad de hidrofobinas (teóricamente la máxima carga de este
soporte es superior a 100 mg de proteína por gramos de soporte y se añadió solo 0,7 mg de
hidrofobinas) se incubó con el gel glioxil-agarosa69. La proteína (0,1 mg/mL) se disolvió en una
mezcla de 50% dioxano/ 40% tritón X-100/10% tampón bicarbonato sódico 200 mM a pH=10,5
y la inmovilización se mantuvo durante 16 horas. Después de este tiempo, la mezcla fue
reducida mediante la adición de un volumen igual de 2 mg/mL de borohidruro sódico en
tampón bicarbonato sódico 100 mM (pH=10,5) durante 30 minutos y posteriormente lavada
con 100 volúmenes de agua destilada. El glioxil-agarosa reducido (usado como referencia en
la inmovilización de lipasas) se trató exactamente bajo las mismas condiciones.
8888. . . . Inmovilización de lipasas sobre el soporte glioxilInmovilización de lipasas sobre el soporte glioxilInmovilización de lipasas sobre el soporte glioxilInmovilización de lipasas sobre el soporte glioxil----agarosaagarosaagarosaagarosa----hidrofobina hidrofobina hidrofobina hidrofobina
La lipasa purificada se diluyó 10 veces empleando agua destilada (para diluir el Tritón
X-100) ofreciéndose 1 mL de soporte glioxil-agarosa-hidrofobina (concentración 0,1 mg/mL). El
seguimiento de la inmovilización se realizó mediante el análisis a distintos tiempos de
muestras de suspensión y sobrenadante. La inmovilización se realizo a baja fuerza iónica.
Además, una suspensión referencia se preparó usando glioxil-agarosa reducido para
asegurar que la adsorción de la lipasa sobre el glioxil-agarosa-hidrofobina fue promovida por
la presencia de la hidrofobina.
9999. Inmovili. Inmovili. Inmovili. Inmovilización de HLL a diferentes concentraciones sobre soporte PEIzación de HLL a diferentes concentraciones sobre soporte PEIzación de HLL a diferentes concentraciones sobre soporte PEIzación de HLL a diferentes concentraciones sobre soporte PEI----Sepabeads.Sepabeads.Sepabeads.Sepabeads.
La inmovilización fue realizada empleando un soporte PEI-Sepabeads68 (con peso
molecular de PEI de 60 kDa). En el caso de la enzima concentrada, una solución 15 mL de
tampón fosfato sódico (5 mM) conteniendo 0,74 mg/mL de lipasa se ofreció a 3 gramos de
soporte. Para la enzima diluida, se añadieron 120 mL conteniendo 0,0925 mg/mL de lipasa
con 0,6% de tritón (v/v) sobre tres gramos de soporte. La carga enzimática en ambos casos
fue de 3,7 mg de lipasa/mL de soporte. Las inmovilizaciones se realizaron a pH 7 y 25ºC con
un rendimiento de inmovilización cercano al 100% en ambos casos.

Capítulo 1
33
10101010. Reacciones de hidrólisis.. Reacciones de hidrólisis.. Reacciones de hidrólisis.. Reacciones de hidrólisis.
Soluciones de [(±)-1111] (10 mM, 10 mL) o [(±)-2222] (2mM, 20 mL) se prepararon en tampón
fosfato sódico 25 mM a 25ºC y pH=7 o 7,5 respectivamente. A estas, se añadió 1 mL del
derivado enzimático en cada caso analizándose la conversión mediante HPLC en fase reversa.
En todos los casos el pH se mantuvo constante mediante un pHstato.
Se empleó una columna Kromasil C18 (25x0,4 cm) suministrada por Análisis Vinicos.
Los ensayos se llevaron a cabo por triplicado. La fase móvil fue una mezcla isocrática de
acetonitrilo: 10 mM de tampón fosfato amónico en agua bi-ionizada (30:70) (para [(±)-1111]),
(40:60 para [(±)-2222]) a pH 3,00 y con un flujo de 1,5 mL/min. La absorbancia empleada fue de
254 nm (detector UV Spectra Physic SP 8450). Los tiempos de retención de los compuestos
fueron: ácido mandélico (3333) (2,9 min), [(±)-1111] (8 min), ácido 4-fenil-2-hidroxibutanoico (4)4)4)4) (3,5
min), [(±)-2222] (8,3 min).
Igualmente se analizó el exceso enantiomérico del ester sin hidrolizar de ambos sustratos
mediante análisis en HPLC quiral. La columna empleada fue una CHIRACEL OD-R, la fase móvil
fue una mezcla isocrática de acetonitrilo y agua 30:70 para [(±)-1] 1] 1] 1] y 40:60 para [(±)-2222], flujo 0,5
mL/min analizado a una adsorbancia de 254 nm. La enantioselectividad se expresó en
ambos casos como el valor de E calculado a partir del exceso enantiomérico de sustrato (ee) y
del grado de conversión (c) (al 50% de hidrólisis) de acuerdo a la ecuación previamente
descrita por Chen y col 73.
E= ln [(1-c)(1-ee)] / ln [1-c)(1+ee)]
11111111. Ensayo de actividad. Ensayo de actividad. Ensayo de actividad. Ensayo de actividad----temperatura de las distintas preparaciones de lipasas.temperatura de las distintas preparaciones de lipasas.temperatura de las distintas preparaciones de lipasas.temperatura de las distintas preparaciones de lipasas.
El efecto de la temperatura sobre la actividad enzimática fue examinado en la
hidrólisis del butirato de etilo (50 mM) a pH 7 y 25 mM de fosfato sódico empleando un
pHstato. El tampón se preincubó a la temperatura deseada antes de adicionar el sustrato y la
enzima.

Interacción de lipasas con estructuras hidrofóbicas
34
11112222. Inactivación térmica.. Inactivación térmica.. Inactivación térmica.. Inactivación térmica.
Para estudiar la estabilidad de las diferentes lipasas, las diferentes preparaciones se
incubaron a 50ºC y pH 7. Se fueron tomando diferentes alícuotas analizándolas con el ensayo
de pNPP o pNPB.

Capítulo 2
35
INTRODUCCIÓNINTRODUCCIÓNINTRODUCCIÓNINTRODUCCIÓN
Una vez más debemos referirnos al complejo mecanismo que rige la catálisis en las
lipasas. Como vimos anteriormente, estas enzimas en medios acuosos presentan poca
actividad ya que se encuentran en un cierto equilibrio entre dos conformaciones, una abierta,
y activa, y otra cerrada, considerada inactiva. En medios acuosos esta estructura abierta es
más desfavorable que la estructura cerrada de la lipasa 74-75, con lo que el equilibrio debe estar
desplazado hacia esta segunda estructura.
La inmovilización de la lipasa mediante unión covalente multipuntual69 podría dar
lugar a una mayor estabilización de la enzima que las correspondientes enzimas en solución
ó que las moléculas inmovilizadas a través de un único enlace. Asimismo, la adsorción
interfacial sobre superficies hidrofóbicas da lugar a una fijación de la conformación abierta de
la lipasa 43-45,76-78, lo cual también podría verse reflejado en una mayor actividad y estabilidad.
Por tanto, mediante estas metodologías puede ser posible conseguir la estabilización
de las distintas conformaciones que existen en las lipasas.
1111. . . . Métodos de Métodos de Métodos de Métodos de inmovilización inmovilización inmovilización inmovilización de las distintasde las distintasde las distintasde las distintas conformaciones de lipasas. conformaciones de lipasas. conformaciones de lipasas. conformaciones de lipasas.
1.11.11.11.1 Adsorción interfacial sobre soportes hidrofóbicosAdsorción interfacial sobre soportes hidrofóbicosAdsorción interfacial sobre soportes hidrofóbicosAdsorción interfacial sobre soportes hidrofóbicos: inmovilización de la : inmovilización de la : inmovilización de la : inmovilización de la
conformación abierta de conformación abierta de conformación abierta de conformación abierta de lipasas.lipasas.lipasas.lipasas.
La tendencia de las lipasas a absorberse a superficies hidrofóbicas (activación
interfacial) se ha empleado para desarrollar un protocolo de inmovilización específico para
lipasas, que consiste en la adsorción interfacial de las lipasas sobre soportes hidrofóbicos [por
ejemplo octil-agarosa34, octadecil-Sepabeads79 e incluso sobre hidrofobinas inmovilizadas
sobre glioxil-agarosa78 (capítulo 1)] a baja fuerza iónica. En estas condiciones, las lipasas
"confunden" la superficie del soporte con la superficie hidrofóbica de las gotas de su sustrato
natural, de forma que se adsorben muy fuertemente a estos soportes, a través del “lid” y del
gran bolsillo hidrofóbico que rodea al centro activo, fijándose así la conformación abierta.
Así, este tipo de inmovilización permite en un único paso la purificación,
inmovilización e hiperactivación interfacial de las lipasas (Esquema 11).

Estabilidad de derivados inmovilizados de lipasas
36
1.21.21.21.2 InInInInmovilización movilización movilización movilización de de de de lipasas lipasas lipasas lipasas por unión cpor unión cpor unión covalente ovalente ovalente limitada.limitada.limitada.limitada.
1.2.1 Inmovilización de lipasas sobre soporte agarosa-bromocianógeno (BrCN)
La inmovilización de enzimas sobre un soporte agarosa activado con grupos bromuro
de cianógeno tiene lugar a través de la unión covalente entre la enzima y el soporte (Esquema
12). Esta inmovilización se realiza a pH 7, donde la interacción de la enzima sobre el soporte
posiblemente procede por el grupo amino más reactivo de la proteína (el amino terminal) de
modo que casi no existe rigidificación de la enzima.
Esquema Esquema Esquema Esquema 12121212. . . . Inmovilización de enzimas sobre agarosa bromocianógeno
1.2.2 Inmovilización de lipasas sobre soporte glutaraldehído-agarosa
La utilización de un soporte agarosa recubierto en su superficie de grupos amino80 y
posteriormente activado superficialmente con grupos glutaraldehído81 permite la inmovilización
de las enzimas a través de la zona con mayor carga negativa cuando se realiza a pH 7 y a
Esquema Esquema Esquema Esquema 11111111.... Adsorción interfacial de lipasas sobre superficie hidrofóbica
Lipasa abiertaLipasa abiertaLipasa abiertaLipasa abiertay activay activay activay activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
Activación Activación Activación Activación InterfacialInterfacialInterfacialInterfacial
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicasoportesoportesoportesoporte
Lipasa abiertaLipasa abiertaLipasa abiertaLipasa abiertay activay activay activay activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
Activación Activación Activación Activación InterfacialInterfacialInterfacialInterfacial
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicasoportesoportesoportesoporte
Lipasa abiertaLipasa abiertaLipasa abiertaLipasa abiertay activay activay activay activa
Lipasa cerradaLipasa cerradaLipasa cerradaLipasa cerradae inactivae inactivae inactivae inactiva
Activación Activación Activación Activación InterfacialInterfacialInterfacialInterfacial
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicasoportesoportesoportesoporte
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlica
zonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbicazonazonazonazona hidrofóbicahidrofóbicahidrofóbicahidrofóbica
zonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicazonazonazonazona hidrofÍlicahidrofÍlicahidrofÍlicahidrofÍlicasoportesoportesoportesoportesoportesoportesoportesoporte
• Inmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminal• Unión covalente Unión covalente Unión covalente Unión covalente unipuntualunipuntualunipuntualunipuntual
OOOO
OOOONNNN
• Inmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminal• Unión covalente Unión covalente Unión covalente Unión covalente unipuntualunipuntualunipuntualunipuntual
OOOO
OOOONNNN
• Inmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminalInmovilización a través del amino terminal• Unión covalente Unión covalente Unión covalente Unión covalente unipuntualunipuntualunipuntualunipuntual
OOOO
OOOONNNN
OOOO
OOOONNNN

Capítulo 2
37
baja fuerza iónica (Ej. 25 mM). En primer lugar se produce una adsorción de la enzima por la
zona más rica en cargas negativas con posterior unión covalente a través de algún/os grupos
aminos presentes en la proteína (Esquema 13).
Esquema Esquema Esquema Esquema 13131313. . . . Inmovilización de enzimas sobre soporte glutaraldehído-agarosa
1.31.31.31.3 IIIInmovilización covalente multnmovilización covalente multnmovilización covalente multnmovilización covalente multipuntualipuntualipuntualipuntual de lipasas sobre soporte glioxil agarosa. de lipasas sobre soporte glioxil agarosa. de lipasas sobre soporte glioxil agarosa. de lipasas sobre soporte glioxil agarosa.
La inmovilización irreversible de enzimas por formación de enlaces covalentes con el
soporte82, supone grandes ventajas a la hora de utilizar catalizadores estables. Si además el
proceso permite la formación de numerosos enlaces entre cada molécula de enzima y el
soporte con el que se une (multi-interacción), las posibles distorsiones de la enzima se verán
limitadas.
Así, la inmovilización por enlace covalente multipuntual de enzimas puede
conseguirse a través del empleo de geles de agarosa activados de forma que en su superficie
presenten una monocapa de aldehídos sencillos (gel glioxil) moderadamente alejados de la
pared del soporte y totalmente expuestos al medio, de forma que los grupos amino de la
enzima pueden reaccionar con ellos69 (Esquema 14).
Las principales características del sistema elegido son:
1- Buena congruencia geométrica entre el soporte y la enzima.
2- La unión unipuntual aldehído-amino (bases de Schiff) es muy débil y reversible, de forma
que si una unión introduce una importante distorsión en la estructura tridimensional de la
proteína, este enlace pueda romperse.
3- Reducción final de las reversibles bases de Schiff a enlaces amino secundarios muy
estables utilizando borohidruro, agente adecuado para difundir por el complicado entramado
• Inmovilización a través del área de grupos amino más reactivos• Impedimentos estéricos por multiinteraciones moleculares • Modificaciones químicas significativas
HHHH2222NNNN
• Inmovilización a través del área de grupos amino más reactivos• Impedimentos estéricos por multiinteraciones moleculares • Modificaciones químicas significativas
HHHH2222NNNNHHHH2222NNNNHHHH2222NNNN

Estabilidad de derivados inmovilizados de lipasas
38
macromolecular formado por la enzima unida multipuntualmente al soporte y que además es
capaz de reducir los grupos activos remanentes en el soporte.
4- La primera interacción de la enzima con el soporte, se produce a través de al menos dos
puntos por lo que en este tipo de interacción la enzima se debe auto-orientar hacia el soporte
por su zona/s más rica/s en grupos amino reactivos (lisinas).
Las variables que se han descrito pueden controlar el grado de multiinteracción entre
la enzima y el soporte son:
a) La temperatura de inmovilización: El uso de temperaturas moderadamente altas podría
promover una mayor flexibilidad de la estructura de la proteína, lo que favorecería su
reactividad con los grupos del soporte.
b) El tiempo de inmovilización: De forma que consigamos un correcto alineamiento de las
moléculas de enzima y el soporte.
c) El pH del medio: Importante ya que marca la reactividad de los grupos de la superficie de
la proteína (Ej. Lisinas reactivas, tienen un pK de en torno a 10 y son poco reactivas a pH
neutro).
Etc.
Esta estrategia de estabilización-inmovilización de enzimas ha sido aplicada con éxito
a numerosas enzimas, como α-quimotripsina, tripsina, carbopeptidasa A, termolisina, β-
galactosidasa, lipasas, D-aminoácido oxidasas, etc83-90.
La ventaja fundamental de este sistema radica en que tras la reducción de los
derivados, la superficie del soporte es sumamente hidrofílica e inerte. Sin embargo, este
sistema no cumple todas las características del sistema ideal:
1- La necesidad de inmovilizar las enzimas a un pH elevado (pH 10), en el que las enzimas
son poco estables.
2- El uso de borohidruro en la reducción del enlace tipo base de Schiff, puede representar un
problema cuando se trata de cantidades muy grandes de derivado (Ej. preparación a nivel
industrial).

Capítulo 2
39
Esquema Esquema Esquema Esquema 14141414. . . . Inmovilización covalente multipuntual de enzimas sobre soportes glioxil-agarosa
2. Lipasas de Termófilos.2. Lipasas de Termófilos.2. Lipasas de Termófilos.2. Lipasas de Termófilos.
Las lipasas presentan poca estabilidad en condiciones de alta temperatura, pH, etc.,..
(MML, CRL….). Sin embargo, existen lipasas provenientes de organismos termófilos, las cuales
son estables a temperaturas superiores a los 50ºC.
Debido a su extremada estabilidad a temperaturas elevadas (termofília) y en
disolventes orgánicos, las lipasas de termófilos han llegado a ser de especial interés para
aplicaciones biotecnológicas 91-94.
Así, las lipasas empleadas en la industria de detergentes necesitan ser estables en
presencia de surfactantes y pH altos, además la mayoría de los procesos industriales operan
a temperaturas por encima de 45ºC, por lo que la lipasa debe ser activa y estable a estas
temperaturas.
Actualmente la mayoría de lipasas termófilas que han sido purificadas o
caracterizadas provienen de Bacillus 95-96, como las lipasas de B. thermocatenulatus 97-99, B.
thermoleovorans 100, B.subtilis101 o B. stearothermophilus102, no obstante también existen lipasas
provenientes de Thermus como T. thermophilus, T.aquaticus, o T. flavus aunque su
caracterización y utilización no se encuentran descritas en la literatura.
El termófilo Bacillus thermocatenulatus (BTL) produce dos lipasas, llamadas BTL1 y BTL2 97-98. El gen para la lipasa BTL2 fue clonado y expresado en Escherichia coli. La lipasa de BTL2
es una proteína de 43 kDa que muestra alta estabilidad a 50°C, y pH alcalino (9,0–11,0) y en
Derivado enzimáticoDerivado enzimáticoDerivado enzimáticoDerivado enzimáticomultipuntualmultipuntualmultipuntualmultipuntual
IncubaciónIncubaciónIncubaciónIncubaciónpH 10pH 10pH 10pH 10
Reacción de grupos amino Reacción de grupos amino Reacción de grupos amino Reacción de grupos amino cercanoscercanoscercanoscercanos
ReducciónReducciónReducciónReducción
pH 10pH 10pH 10pH 10
NHNHNHNH2222
NHNHNHNH2222
NHNHNHNH2222+
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCC NNNN
CCCCHHHH
OOOO
CCCCHHHH
OOOOCCCC NNNN
CCCC NNNNCCCC NNNNCCCC NNNNCCCC NNNN
Derivado enzimáticoDerivado enzimáticoDerivado enzimáticoDerivado enzimáticomultipuntualmultipuntualmultipuntualmultipuntual
IncubaciónIncubaciónIncubaciónIncubaciónpH 10pH 10pH 10pH 10
Reacción de grupos amino Reacción de grupos amino Reacción de grupos amino Reacción de grupos amino cercanoscercanoscercanoscercanos
ReducciónReducciónReducciónReducción
pH 10pH 10pH 10pH 10
NHNHNHNH2222
NHNHNHNH2222
NHNHNHNH2222+
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCCHHHH
OOOO
CCCC NNNN
CCCCHHHH
OOOO
CCCCHHHH
OOOOCCCC NNNN
CCCC NNNNCCCC NNNNCCCC NNNNCCCC NNNN

Estabilidad de derivados inmovilizados de lipasas
40
disolventes orgánicos (2-propanol, acetona, metanol). Estas propiedades hacen de la BTL2
una lipasa interesante para aplicaciones biotecnologicas99.
Sin embargo, podría ser posible mejorar la estabilidad de estas enzimas de termófilos
mediante el empleo de las técnicas de inmovilización comentadas anteriormente. Además,
podríamos pensar en el empleo de lipasas de termófilos, una vez inmovilizadas, a
temperaturas donde normalmente estas presentan bajas actividades (por ejemplo: 25ºC), lo
cual nos permitiría mayores posibilidades de utilización, por ejemplo en procesos de química
fina.

Capítulo 2
41
OBJETIVOSOBJETIVOSOBJETIVOSOBJETIVOS
Considerando que las lipasas se emplearán industrialmente en forma inmovilizada, es
interesante caracterizar las distintas lipasas usando sistemas en fase sólida. Ya se vio en el
capítulo anterior como es importante conocer la estructura de la lipasa que se emplea.
Así, el objetivo que nos planteamos en este capítulo es el estudio de la actividad y
estabilidad (frente a distintas condiciones experimentales) de diferentes derivados de distintas
lipasas (CAL-B, CRL, MML, BTL2, TTL, TAL), examinándose el efecto de la inmovilización sobre
las propiedades de la lipasa en dos casos:
- La inmovilización mediante unión covalente multipuntual.
- La inmovilización por adsorción interfacial.

Estabilidad de derivados inmovilizados de lipasas
42
RESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓN
1.1.1.1. Purificación de lipasas Purificación de lipasas Purificación de lipasas Purificación de lipasas
Las diferentes lipasas fueron purificadas empleando el gel octil-agarosa34. En el
capítulo 1 se presentó la purificación para las lipasas de C. antarctica B (CAL-B), M. miehei
(MML), H. lanuginosa (HLL), C. rugosa (CRL) y Ps. fluorescens (PFL).
En el caso de las lipasas de Thermus thermophilus (TTL) y Thermus aquaticus (TAL) el
seguimiento de la inmovilización se realizó mediante el ensayo de hidrólisis de butirato de
butilo a 45ºC para TTL y a 65ºC para TAL, observándose una rápida y total inmovilización,
confirmándose la adsorción únicamente de la lipasa por electroforesis en condiciones
desnaturalizantes tras tinción de plata, como se observa en la Figura 11 para la TAL.
Figura 11Figura 11Figura 11Figura 11.... Gel de electroforesis SDSGel de electroforesis SDSGel de electroforesis SDSGel de electroforesis SDS----PAGE de PAGE de PAGE de PAGE de la adsorción sobre octil de TAL.la adsorción sobre octil de TAL.la adsorción sobre octil de TAL.la adsorción sobre octil de TAL. a) Línea 1.- Marcadores de bajo peso molecular. Línea 2.- Preparación comercial de TAL. Línea 3.- TAL adsorbida sobre derivado octil.
En cuanto a la purificación de la lipasa de Bacillus thermocatenulatus (BTL2), el
extracto se ofreció a octil-agarosa, donde se adsorbieron otras proteínas presente en el
extracto crudo además de la lipasa de interés. Mediante un gradiente de desorción con tritón,
empleando un 0,2%, fue posible obtener la lipasa pura como se puede ver en la electroforesis
de la Figura 12a, ya que ésta se desorbía, mientras las demás proteínas se mantenían
adsorbidas sobre el gel octil.
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
1111
30303030
45454545
6666666694949494
14.414.414.414.4
20.120.120.120.1
2222 3333
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
1111
30303030
45454545
6666666694949494
14.414.414.414.4
20.120.120.120.1
2222 3333
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
11111111
3030303030303030
4545454545454545
66666666666666669494949494949494
14.414.414.414.414.414.414.414.4
20.120.120.120.120.120.120.120.1
22222222 33333333

Capítulo 2
43
Además para comprobar que nuestra enzima era la única lipasa del extracto se
realizó una electroforesis nativa con tinción específica de lipasas empleando α-naftilacetato
como sustrato (Figura 12b). Se obtuvo una única banda frente a este sustrato de manera que
se puede asumir que prácticamente toda la actividad observada se debe a nuestra lipasa.
Figura 1Figura 1Figura 1Figura 12222.... Geles de electroforesis de BTL2. Geles de electroforesis de BTL2. Geles de electroforesis de BTL2. Geles de electroforesis de BTL2.
a)a)a)a) SDSSDSSDSSDS----PAGEPAGEPAGEPAGE. Línea 1.- Marcadores de bajo peso molecular. Línea 2.- extracto crudo de BTL2. Línea 3.- BTL2 adsorbida sobre derivado octil. Línea 4.- gel octil después de la desorción con 0,2% de tritón. Línea 5. BTL2 desorbida del gel octil-agarosa. La concentración de proteína fue de 2,5 mg/mL y en el caso de la enzima desorbida el sobrenadante se concentró con centricón hasta 10 veces.
b)b)b)b) Electroforesis nativaElectroforesis nativaElectroforesis nativaElectroforesis nativa. Línea 1.- Extracto crudo de BTL2. Línea 2.- Lipasa desorbida.
2.2.2.2. IIIInmovilización nmovilización nmovilización nmovilización de las distintas lipasas: efecto sobre la actividad enzimática.de las distintas lipasas: efecto sobre la actividad enzimática.de las distintas lipasas: efecto sobre la actividad enzimática.de las distintas lipasas: efecto sobre la actividad enzimática.
2.12.12.12.1 Inmovilización de Inmovilización de Inmovilización de Inmovilización de CALCALCAL
En primer lugar, se prepararon derivados inmovilizados de la lipasa de C antarctica B
(Tabla 4), la cual se diferencia de las demás lipasas en la ausencia de un verdadero lid. La
inmovilización de esta enzima sobre una resina octadecil-Sepabeads (soporte hidrofóbico)
transcurrió de forma rápida (1 hora), observándose una hiperactivación de la enzima frente a
butirato de etilo. Sin embargo, frente a los demás soportes, la inmovilización de la enzima fue
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
30303030
45454545
6666666694949494
20.120.120.120.1
1111 55552222 3333 4444
aaaa bbbb
1111 2222
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
30303030
45454545
6666666694949494
20.120.120.120.1
1111 55552222 3333 4444
aaaa bbbb
1111 2222
PmPmPmPm (kDa)(kDa)(kDa)(kDa)
30303030
45454545
6666666694949494
20.120.120.120.1
1111 55552222 3333 4444
aaaa
30303030
45454545
6666666694949494
20.120.120.120.1
1111 55552222 3333 4444
3030303030303030
4545454545454545
66666666666666669494949494949494
20.120.120.120.120.120.120.120.1
11111111 5555555522222222 33333333 44444444
aaaa bbbb
1111 2222
bbbb
1111 222211111111 22222222
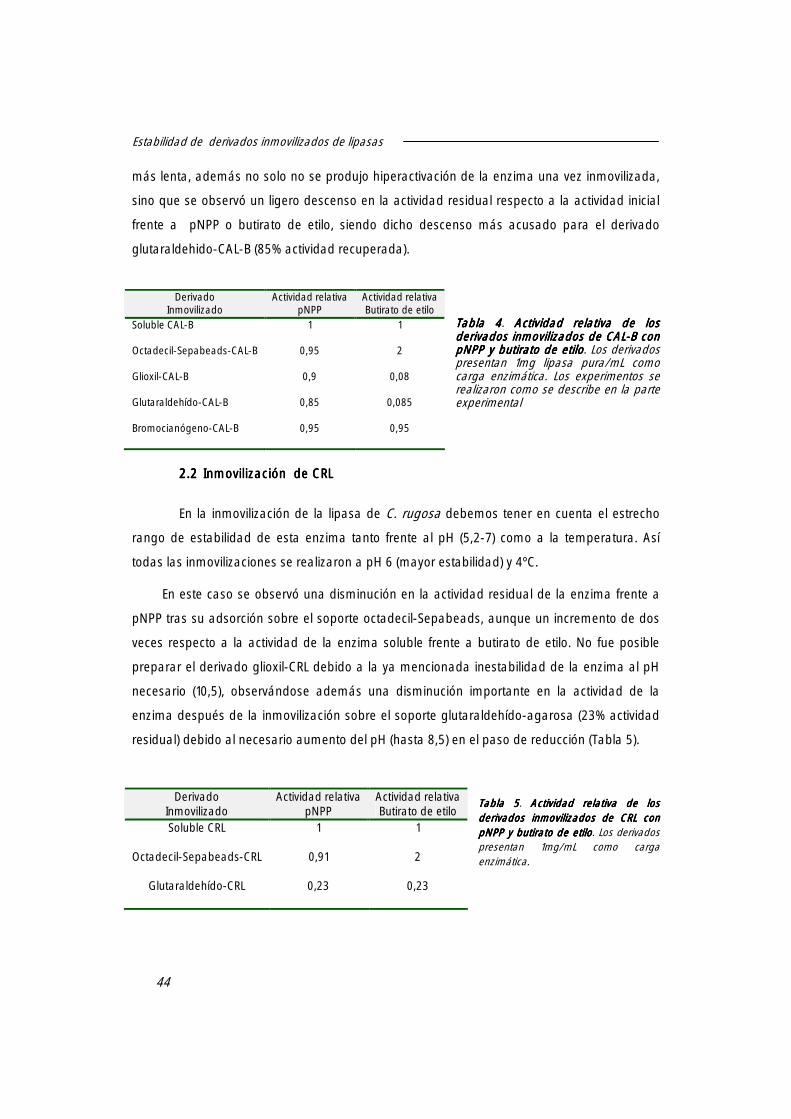
Estabilidad de derivados inmovilizados de lipasas
44
más lenta, además no solo no se produjo hiperactivación de la enzima una vez inmovilizada,
sino que se observó un ligero descenso en la actividad residual respecto a la actividad inicial
frente a pNPP o butirato de etilo, siendo dicho descenso más acusado para el derivado
glutaraldehido-CAL-B (85% actividad recuperada).
Tabla 4Tabla 4Tabla 4Tabla 4. Actividad reActividad reActividad reActividad relativa de los lativa de los lativa de los lativa de los derivados inmovilizados de CALderivados inmovilizados de CALderivados inmovilizados de CALderivados inmovilizados de CAL----B con B con B con B con pNPP y butirato de etilopNPP y butirato de etilopNPP y butirato de etilopNPP y butirato de etilo. Los derivados presentan 1mg lipasa pura/mL como carga enzimática. Los experimentos se realizaron como se describe en la parte experimental
2.22.22.22.2 Inmovilización Inmovilización Inmovilización de CRL de CRL de CRL
En la inmovilización de la lipasa de C. rugosa debemos tener en cuenta el estrecho
rango de estabilidad de esta enzima tanto frente al pH (5,2-7) como a la temperatura. Así
todas las inmovilizaciones se realizaron a pH 6 (mayor estabilidad) y 4ºC.
En este caso se observó una disminución en la actividad residual de la enzima frente a
pNPP tras su adsorción sobre el soporte octadecil-Sepabeads, aunque un incremento de dos
veces respecto a la actividad de la enzima soluble frente a butirato de etilo. No fue posible
preparar el derivado glioxil-CRL debido a la ya mencionada inestabilidad de la enzima al pH
necesario (10,5), observándose además una disminución importante en la actividad de la
enzima después de la inmovilización sobre el soporte glutaraldehído-agarosa (23% actividad
residual) debido al necesario aumento del pH (hasta 8,5) en el paso de reducción (Tabla 5).
Tabla 5Tabla 5Tabla 5Tabla 5. Actividad relativa de los Actividad relativa de los Actividad relativa de los Actividad relativa de los derivados inmovilizadosderivados inmovilizadosderivados inmovilizadosderivados inmovilizados de CRL con de CRL con de CRL con de CRL con pNPP y butirato de etilopNPP y butirato de etilopNPP y butirato de etilopNPP y butirato de etilo. Los derivados presentan 1mg/mL como carga enzimática.
Derivado Inmovilizado
Actividad relativa pNPP
Actividad relativa Butirato de etilo
Soluble CAL-B
1 1
Octadecil-Sepabeads-CAL-B
0,95 2
Glioxil-CAL-B 0,9 0,08
Glutaraldehído-CAL-B
0,85 0,085
Bromocianógeno-CAL-B
0,95 0,95
Derivado Inmovilizado
Actividad relativa pNPP
Actividad relativa Butirato de etilo
Soluble CRL
1 1
Octadecil-Sepabeads-CRL
0,91 2
Glutaraldehído-CRL
0,23 0,23

Capítulo 2
45
2.32.32.32.3 InmovilizaciónInmovilizaciónInmovilizaciónInmovilización de MML de MML de MML de MML
Otra de las enzimas estudiadas fue la lipasa de M. miehei. La inmovilización de la
MML sobre octadecil-Sepabeads fue muy rápida, como ya se había visto para las demás
lipasas estudiadas, dando lugar a una hiperactivación de la enzima de dos veces respecto a
la actividad pNPB y hasta de cinco veces frente a butirato de etilo; mientras que, cuando la
enzima fue inmovilizada covalentemente sobre glioxil-agarosa o glutaraldehido-agarosa se
observó una disminución en la actividad final de un 30% y un 50%, respectivamente frente a
ambos sustratos (Tabla 6).
Tabla 6Tabla 6Tabla 6Tabla 6. Actividad relativa de los Actividad relativa de los Actividad relativa de los Actividad relativa de los derivados inmovilizados de MML con derivados inmovilizados de MML con derivados inmovilizados de MML con derivados inmovilizados de MML con pNPB pNPB pNPB pNPB y butirato de etiloy butirato de etiloy butirato de etiloy butirato de etilo. Los derivados presentan 1mg/mL como carga enzimatica.
2.42.42.42.4 InmovilizaciónInmovilizaciónInmovilizaciónInmovilización de BTL2 de BTL2 de BTL2 de BTL2
La inmovilización de la BTL2 sobre octadecil-Sepabeads fue también muy rápida
mostrando una cierta hiperactivación de la enzima frente a pNPP y un aumento de hasta tres
veces en la actividad frente a butirato de etilo respecto a la actividad enzimática inicial.
Cuando la lipasa se inmovilizó sobre glioxil-agarosa por unión covalente multipuntual se
produjo una ligera disminución en la actividad de un 25% (Tabla 7)
Tabla 7Tabla 7Tabla 7Tabla 7. Actividad relativa de los . Actividad relativa de los . Actividad relativa de los . Actividad relativa de los derivados inmovilizados de BTL2 con derivados inmovilizados de BTL2 con derivados inmovilizados de BTL2 con derivados inmovilizados de BTL2 con pNPP pNPP pNPP pNPP y butirato de etiloy butirato de etiloy butirato de etiloy butirato de etilo. Los derivados presentan 1 mg/mL como carga enzimática.
Derivado Inmovilizado
Actividad relativa pNPP
Actividad relativa Butirato de etilo
Soluble MML
1 1
Octadecil-Sepabeads-MML
2 5
Glioxil-MML
0,7 0,73
Glutaraldehído-MML
0,5 0,6
Derivado Inmovilizado
Actividad relativa pNPP
Actividad relativa Butirato de etilo
Soluble BTL2
1 1
Octadecil-Sepabeads-BTL2
1,2 3,4
Glioxil-BTL2
0,75 0,76

Estabilidad de derivados inmovilizados de lipasas
46
2.52.52.52.5 Inmovilización Inmovilización Inmovilización Inmovilización de TTL de TTL de TTL de TTL
La inmovilización de TTL sobre octadecil-Sepabeads por adsorción interfacial fue, una vez
más, muy rápida, destacándose la hiperactivación de la enzima 4 veces respecto a la soluble
frente a butirato de butilo. Sin embargo la inmovilización sobre glioxil-agarosa dio lugar a un
ligero descenso en la actividad de la enzima de un 20% (Tabla 8).
Tabla 8Tabla 8Tabla 8Tabla 8. Actividad relativa de los derivados . Actividad relativa de los derivados . Actividad relativa de los derivados . Actividad relativa de los derivados inmovilizados de TTL con Butirato de butiloinmovilizados de TTL con Butirato de butiloinmovilizados de TTL con Butirato de butiloinmovilizados de TTL con Butirato de butilo. Los derivados presentan 1 mg/mL como carga enzimática. Los experimentos se realizaron como se describe en la parte experimental.
2.62.62.62.6 InmovilizaciónInmovilizaciónInmovilizaciónInmovilización de TAL de TAL de TAL de TAL Para la TAL, la inmovilización sobre octadecil-Sepabeads no produjo un incremento
en la actividad de la enzima respecto a la inicial catalizando la hidrólisis del butirato de butilo,
incluso se observó un leve descenso de ésta. Asimismo, la inmovilización sobre glioxil-
agarosa produjo un descenso en la actividad de la enzima inmovilizada de un 15% respecto a
la actividad inicial (Tabla 9).
Tabla 9Tabla 9Tabla 9Tabla 9. Actividad relativa de los derivados . Actividad relativa de los derivados . Actividad relativa de los derivados . Actividad relativa de los derivados inmovilizados de TAL con butirato de butiloinmovilizados de TAL con butirato de butiloinmovilizados de TAL con butirato de butiloinmovilizados de TAL con butirato de butilo. Los derivados presentan 1 mg/mL como carga enzimática. Los experimentos se realizaron como se describe en la parte experimental.
Por tanto, a la vista de los resultados obtenidos, podemos decir que la hiperactivación de la
enzima tras su inmovilización sobre el soporte octadecil-Sepabeads fue mucho más
significativa empleando esteres del ácido butírico como sustrato que pNPP, quizás debido a
que ésta primera es una molécula de menor tamaño y más hidrofóbica lo cual la hace más
accesible al centro activo de la enzima. Para los demás derivados inmovilizados no se produjo
variación significativa en la actividad enzimática al cambiar de sustratos.
Derivado Inmovilizado
Actividad relativa pNPP
Soluble TTL
1
Octadecil-Sepabeads-TTL
4
Glioxil-TTL
0,8
Derivado Inmovilizado
Actividad relativa pNPP
Soluble TAL
1
Octadecil-Sepabeads-TAL
0,95
Glioxil-TAL
0,85
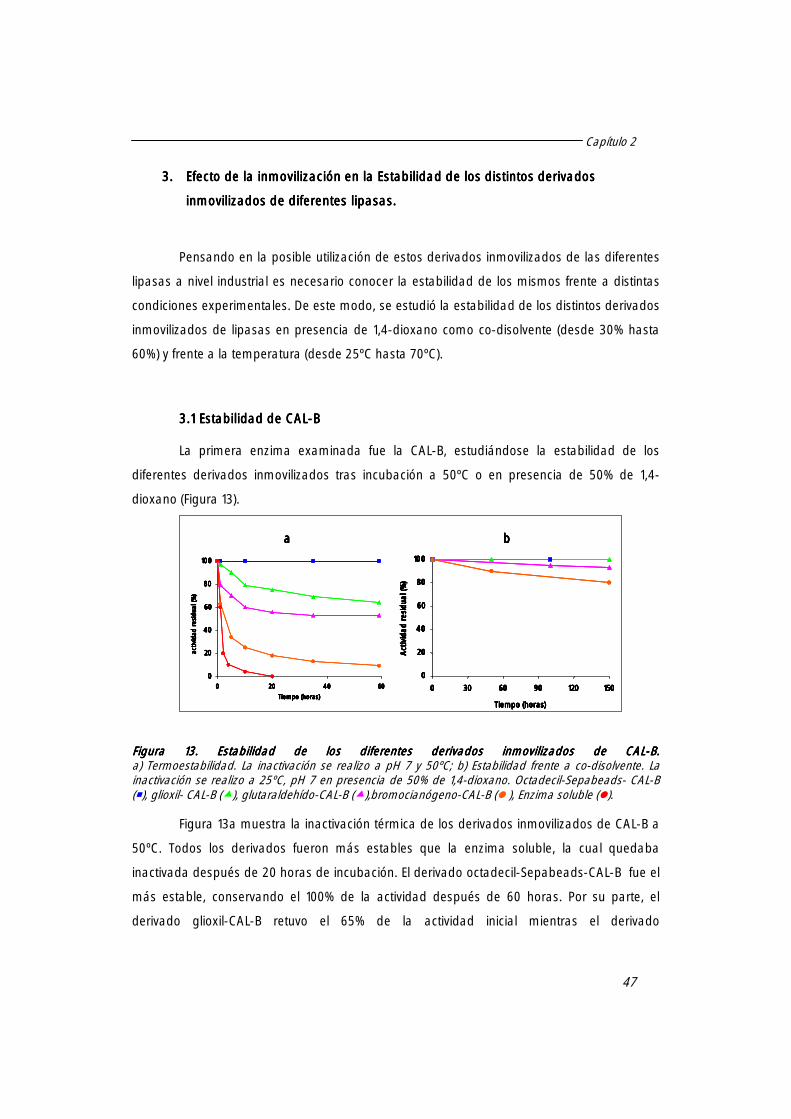
Capítulo 2
47
3.3.3.3. Efecto de la inmovilización en la Efecto de la inmovilización en la Efecto de la inmovilización en la Efecto de la inmovilización en la EEEEstabilidad de los distintos derivados stabilidad de los distintos derivados stabilidad de los distintos derivados stabilidad de los distintos derivados
inmovilizados de inmovilizados de inmovilizados de inmovilizados de diferentes diferentes diferentes diferentes lipasas.lipasas.lipasas.lipasas.
Pensando en la posible utilización de estos derivados inmovilizados de las diferentes
lipasas a nivel industrial es necesario conocer la estabilidad de los mismos frente a distintas
condiciones experimentales. De este modo, se estudió la estabilidad de los distintos derivados
inmovilizados de lipasas en presencia de 1,4-dioxano como co-disolvente (desde 30% hasta
60%) y frente a la temperatura (desde 25ºC hasta 70ºC).
3.1 Estabilidad de CAL3.1 Estabilidad de CAL3.1 Estabilidad de CAL3.1 Estabilidad de CAL----BBBB
La primera enzima examinada fue la CAL-B, estudiándose la estabilidad de los
diferentes derivados inmovilizados tras incubación a 50ºC o en presencia de 50% de 1,4-
dioxano (Figura 13).
Figura 1Figura 1Figura 1Figura 13333. Estabilidad de los diferentes derivados inmovilizados de CAL. Estabilidad de los diferentes derivados inmovilizados de CAL. Estabilidad de los diferentes derivados inmovilizados de CAL. Estabilidad de los diferentes derivados inmovilizados de CAL----B.B.B.B. a) Termoestabilidad. La inactivación se realizo a pH 7 y 50ºC; b) Estabilidad frente a co-disolvente. La inactivación se realizo a 25ºC, pH 7 en presencia de 50% de 1,4-dioxano. Octadecil-Sepabeads- CAL-B (�), glioxil- CAL-B (�), glutaraldehído-CAL-B (�),bromocianógeno-CAL-B (� ), Enzima soluble (�).
Figura 13a muestra la inactivación térmica de los derivados inmovilizados de CAL-B a
50ºC. Todos los derivados fueron más estables que la enzima soluble, la cual quedaba
inactivada después de 20 horas de incubación. El derivado octadecil-Sepabeads-CAL-B fue el
más estable, conservando el 100% de la actividad después de 60 horas. Por su parte, el
derivado glioxil-CAL-B retuvo el 65% de la actividad inicial mientras el derivado
aaaa bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
activ
idad
res
idua
l (%
)ac
tivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Estabilidad de derivados inmovilizados de lipasas
48
bromocianógeno-CAL-B solo conservó un 15% de la actividad después de 60 horas, sugiriendo
esto que se han introducido algunas uniones covalentes multipuntuales entre el enzima y el
soporte glioxil-agarosa, aumentando la rigidez del enzima y por tanto su estabilidad.
El derivado octadecil-Sepabeads-CAL-B fue de nuevo el más estable en presencia de
50% de co-disolvente (Figura 13b), junto con el derivado glioxil-CAL-B, manteniendo ambos el
100% de actividad después de 200 horas de incubación, mientras el derivado BrCN-CAL-B solo
conservó un 70% de la actividad inicial.
La solubilización de fármacos normalmente requiere el empleo de altas temperaturas
junto con la presencia de una determinada cantidad de co-disolvente en el medio. Por ello, se
estudió la estabilidad de los diferentes derivados de CAL-B a 37°C y con un 40% de co-
disolvente (Tabla 10). En estas condiciones, los derivados octadecil-Sepabeads y glioxil CAL-B
mantuvieron toda la actividad tras 170 horas de incubación, mientras los derivados
glutaraldehído-CAL-B y BrCN-CAL-B solo retuvieron un 57% y un 40% de la actividad inicial,
respectivamente.
Tabla 10. Estabilidad de los derivados Tabla 10. Estabilidad de los derivados Tabla 10. Estabilidad de los derivados Tabla 10. Estabilidad de los derivados inmovilizados de CALinmovilizados de CALinmovilizados de CALinmovilizados de CAL----B a 37ºC y en presencia de B a 37ºC y en presencia de B a 37ºC y en presencia de B a 37ºC y en presencia de 40% de 1,440% de 1,440% de 1,440% de 1,4----dioxano. dioxano. dioxano. dioxano. Los números están referidos a la actividad residual después de 170 h de incubación.
3.2 Estabilidad de CRL3.2 Estabilidad de CRL3.2 Estabilidad de CRL3.2 Estabilidad de CRL
En el caso de CRL, en cuanto a la termoestabilidad, el derivado octadecil-Sepabeads-
CRL mantuvo totalmente su actividad después de 100 horas a 45ºC, mientras el derivado
covalente glutaraldehído-CRL conservó un 60% de la actividad inicial; sin embargo, la lipasa
soluble era totalmente inactiva en dos horas (Figura 14a).
Derivado Inmovilizado
Actividad residual (%)
Octadecil-Sepabeads-CAL-B
100
Glioxil-CAL-B
100
Glutaraldehído-CAL-B
57
BrCN-CAL-B 40
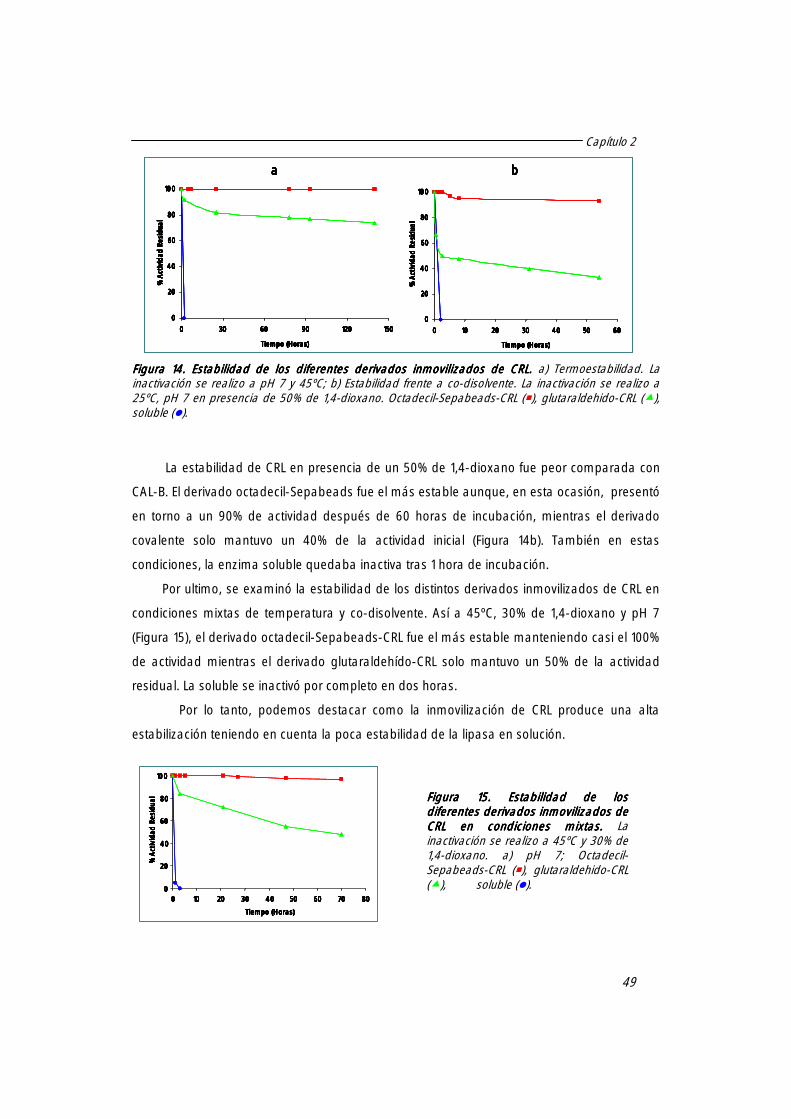
Capítulo 2
49
Figura 1Figura 1Figura 1Figura 14444. Estabilidad de los diferentes derivados inmovilizados de CRL. . Estabilidad de los diferentes derivados inmovilizados de CRL. . Estabilidad de los diferentes derivados inmovilizados de CRL. . Estabilidad de los diferentes derivados inmovilizados de CRL. a) Termoestabilidad. La inactivación se realizo a pH 7 y 45ºC; b) Estabilidad frente a co-disolvente. La inactivación se realizo a 25ºC, pH 7 en presencia de 50% de 1,4-dioxano. Octadecil-Sepabeads-CRL (�), glutaraldehido-CRL (�), soluble (�).
La estabilidad de CRL en presencia de un 50% de 1,4-dioxano fue peor comparada con
CAL-B. El derivado octadecil-Sepabeads fue el más estable aunque, en esta ocasión, presentó
en torno a un 90% de actividad después de 60 horas de incubación, mientras el derivado
covalente solo mantuvo un 40% de la actividad inicial (Figura 14b). También en estas
condiciones, la enzima soluble quedaba inactiva tras 1 hora de incubación.
Por ultimo, se examinó la estabilidad de los distintos derivados inmovilizados de CRL en
condiciones mixtas de temperatura y co-disolvente. Así a 45ºC, 30% de 1,4-dioxano y pH 7
(Figura 15), el derivado octadecil-Sepabeads-CRL fue el más estable manteniendo casi el 100%
de actividad mientras el derivado glutaraldehído-CRL solo mantuvo un 50% de la actividad
residual. La soluble se inactivó por completo en dos horas.
Por lo tanto, podemos destacar como la inmovilización de CRL produce una alta
estabilización teniendo en cuenta la poca estabilidad de la lipasa en solución.
Figura 15Figura 15Figura 15Figura 15. Estabilidad de los . Estabilidad de los . Estabilidad de los . Estabilidad de los diferentes derivados inmovilizadodiferentes derivados inmovilizadodiferentes derivados inmovilizadodiferentes derivados inmovilizados de s de s de s de CRL en condiciones mixtas. CRL en condiciones mixtas. CRL en condiciones mixtas. CRL en condiciones mixtas. La inactivación se realizo a 45ºC y 30% de 1,4-dioxano. a) pH 7; Octadecil-Sepabeads-CRL (�), glutaraldehido-CRL (�), soluble (�).
bbbbaaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
bbbbaaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
bbbbaaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070 80808080
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070 80808080
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070 80808080
Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)Tiempo (Horas)
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
% A
ctiv
idad
Res
idua
l%
Act
ivid
ad R
esid
ual
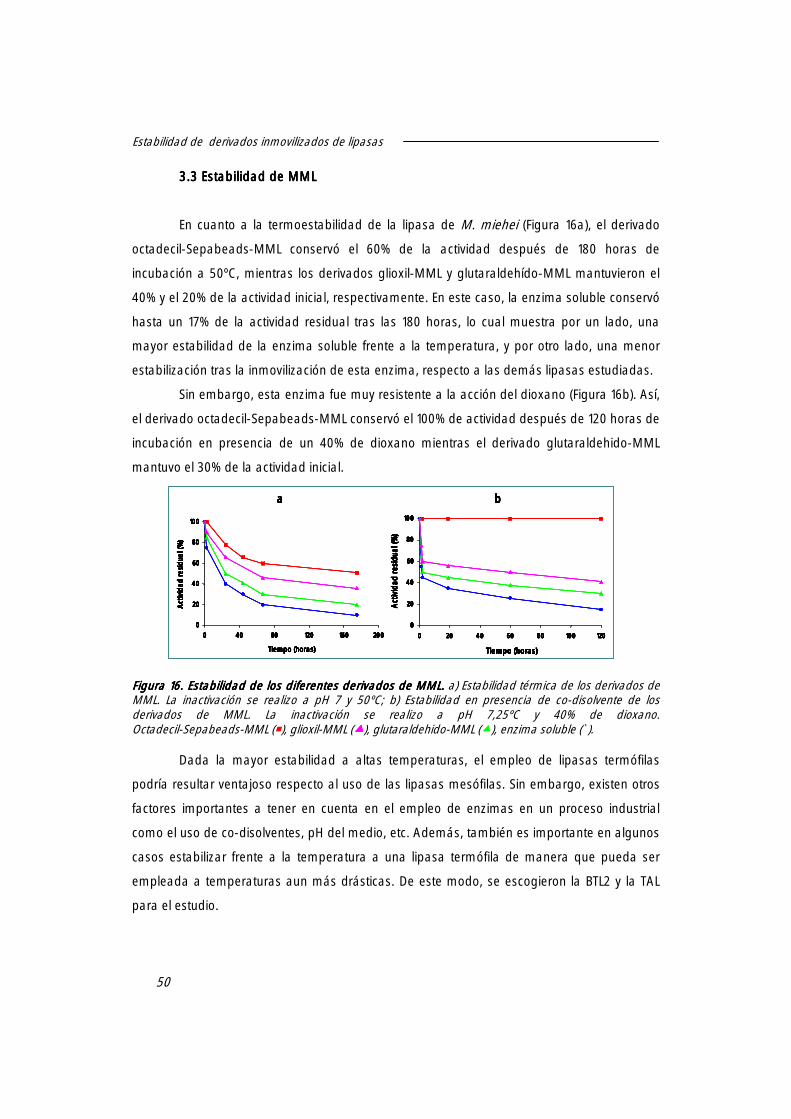
Estabilidad de derivados inmovilizados de lipasas
50
3.3 Estabilidad de MML3.3 Estabilidad de MML3.3 Estabilidad de MML3.3 Estabilidad de MML
En cuanto a la termoestabilidad de la lipasa de M. miehei (Figura 16a), el derivado
octadecil-Sepabeads-MML conservó el 60% de la actividad después de 180 horas de
incubación a 50ºC, mientras los derivados glioxil-MML y glutaraldehído-MML mantuvieron el
40% y el 20% de la actividad inicial, respectivamente. En este caso, la enzima soluble conservó
hasta un 17% de la actividad residual tras las 180 horas, lo cual muestra por un lado, una
mayor estabilidad de la enzima soluble frente a la temperatura, y por otro lado, una menor
estabilización tras la inmovilización de esta enzima, respecto a las demás lipasas estudiadas.
Sin embargo, esta enzima fue muy resistente a la acción del dioxano (Figura 16b). Así,
el derivado octadecil-Sepabeads-MML conservó el 100% de actividad después de 120 horas de
incubación en presencia de un 40% de dioxano mientras el derivado glutaraldehido-MML
mantuvo el 30% de la actividad inicial.
Figura 1Figura 1Figura 1Figura 16666.... Estabilidad de los diferentes derivados de MML. Estabilidad de los diferentes derivados de MML. Estabilidad de los diferentes derivados de MML. Estabilidad de los diferentes derivados de MML. a) Estabilidad térmica de los derivados de MML. La inactivación se realizo a pH 7 y 50ºC; b) Estabilidad en presencia de co-disolvente de los derivados de MML. La inactivación se realizo a pH 7,25ºC y 40% de dioxano. Octadecil-Sepabeads-MML (�), glioxil-MML (����), glutaraldehido-MML (�), enzima soluble (` ).
Dada la mayor estabilidad a altas temperaturas, el empleo de lipasas termófilas
podría resultar ventajoso respecto al uso de las lipasas mesófilas. Sin embargo, existen otros
factores importantes a tener en cuenta en el empleo de enzimas en un proceso industrial
como el uso de co-disolventes, pH del medio, etc. Además, también es importante en algunos
casos estabilizar frente a la temperatura a una lipasa termófila de manera que pueda ser
empleada a temperaturas aun más drásticas. De este modo, se escogieron la BTL2 y la TAL
para el estudio.
aaaa bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 40404040 80808080 120120120120 160160160160 200200200200
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060 80808080 100100100100 120120120120
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 40404040 80808080 120120120120 160160160160 200200200200
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060 80808080 100100100100 120120120120
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 40404040 80808080 120120120120 160160160160 200200200200
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 40404040 80808080 120120120120 160160160160 200200200200
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060 80808080 100100100100 120120120120
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 20202020 40404040 60606060 80808080 100100100100 120120120120
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Capítulo 2
51
3.4 Estabilidad de BTL23.4 Estabilidad de BTL23.4 Estabilidad de BTL23.4 Estabilidad de BTL2
Para la BTL2, el estudio de la estabilidad de los distintos derivados inmovilizados
reflejó las diferencias más significativas entre ellos tras incubación a 65ºC o en presencia de
30% de 1,4-dioxano (Figura 17).
Previamente, se estudió la estabilidad de los mismos a 50ºC observándose que todos
los derivados inmovilizados mantenían el 100% de actividad residual tras 400 horas, mientras
la enzima soluble conservaba el 80% de actividad en estas condiciones.
La Figura 17a muestra la inactivación térmica de los derivados de BTL2 a 65ºC. En este
caso, tras 50 horas de incubación, el derivado octadecil-Sepabeads-BTL2 fue el más estable
manteniendo el 100% de actividad mientras el derivado inmovilizado glioxil-BTL2 conservaba
un 80%. En estas mismas condiciones, la enzima soluble era totalmente inactiva transcurridas
50 horas.
Figura 1Figura 1Figura 1Figura 17777. Estabilidad de los diferentes derivados inmovilizados de BTL2. . Estabilidad de los diferentes derivados inmovilizados de BTL2. . Estabilidad de los diferentes derivados inmovilizados de BTL2. . Estabilidad de los diferentes derivados inmovilizados de BTL2. a) Estabilidad térmica: la inactivación se realizo a pH 7 y 65ºC; b) Estabilidad en presencia de codisolvente: la inactivación se realizo a pH 7,25ºC y 30% de dioxano. Octadecil-Sepabeads-BTL2 (�), glioxil-BTL2 (����), enzima soluble (` ).
Respecto a la estabilidad en presencia de co-disolvente en el medio (25°C y 30% de
dioxano), el derivado octadecil-Sepabeads-BTL2 conservó, una vez mas, el 100% de actividad
después de 20 horas de incubación (Figura 17b), mientras el derivado glioxil-BTL2 mantuvo un
90% de la actividad. En este caso, la enzima soluble conservó hasta un 40% de su actividad
inicial. Hay que añadir que el derivado octadecil-Sepabeads siguió siendo totalmente estable
tras incubaciones hasta 50% de dioxano en un periodo de incubación de 72 horas.
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 5555 10101010 15151515 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 5555 10101010 15151515 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 5555 10101010 15151515 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 5555 10101010 15151515 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 5555 10101010 15151515 20202020
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
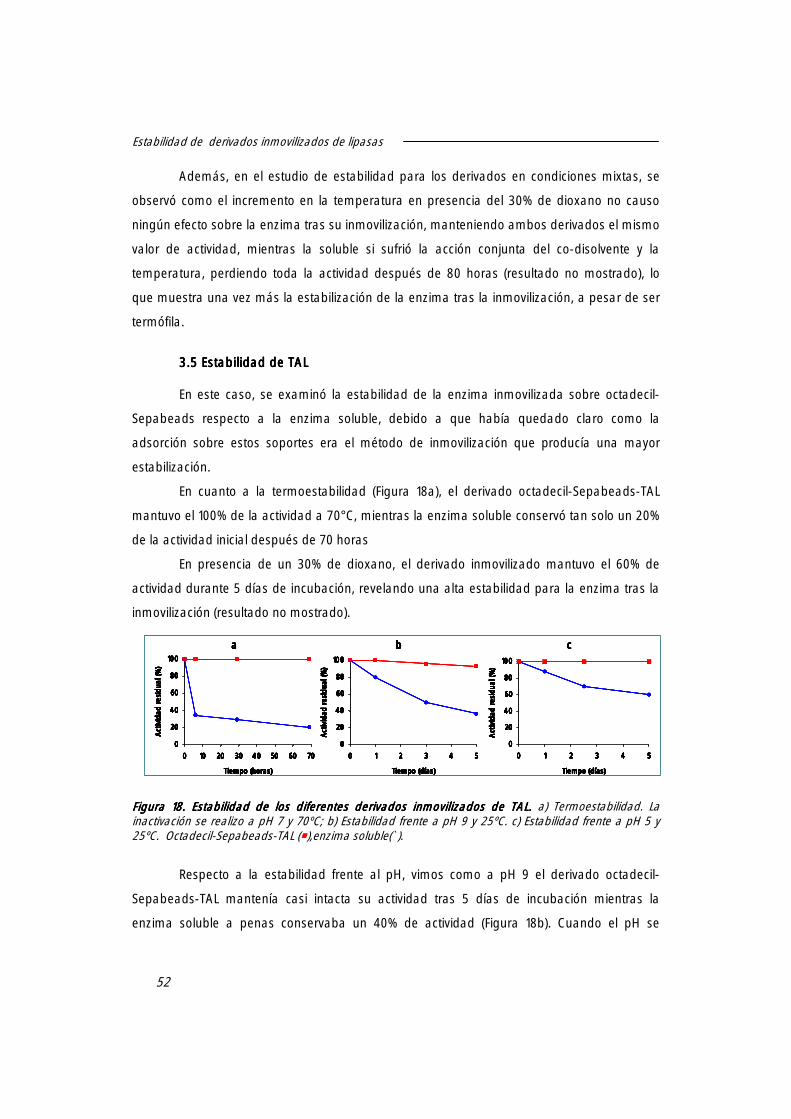
Estabilidad de derivados inmovilizados de lipasas
52
Además, en el estudio de estabilidad para los derivados en condiciones mixtas, se
observó como el incremento en la temperatura en presencia del 30% de dioxano no causo
ningún efecto sobre la enzima tras su inmovilización, manteniendo ambos derivados el mismo
valor de actividad, mientras la soluble si sufrió la acción conjunta del co-disolvente y la
temperatura, perdiendo toda la actividad después de 80 horas (resultado no mostrado), lo
que muestra una vez más la estabilización de la enzima tras la inmovilización, a pesar de ser
termófila.
3.5 3.5 3.5 3.5 Estabilidad de TALEstabilidad de TALEstabilidad de TALEstabilidad de TAL
En este caso, se examinó la estabilidad de la enzima inmovilizada sobre octadecil-
Sepabeads respecto a la enzima soluble, debido a que había quedado claro como la
adsorción sobre estos soportes era el método de inmovilización que producía una mayor
estabilización.
En cuanto a la termoestabilidad (Figura 18a), el derivado octadecil-Sepabeads-TAL
mantuvo el 100% de la actividad a 70°C, mientras la enzima soluble conservó tan solo un 20%
de la actividad inicial después de 70 horas
En presencia de un 30% de dioxano, el derivado inmovilizado mantuvo el 60% de
actividad durante 5 días de incubación, revelando una alta estabilidad para la enzima tras la
inmovilización (resultado no mostrado).
Figura 1Figura 1Figura 1Figura 18888. Estabilidad de los diferentes derivados inmovilizados de TAL. . Estabilidad de los diferentes derivados inmovilizados de TAL. . Estabilidad de los diferentes derivados inmovilizados de TAL. . Estabilidad de los diferentes derivados inmovilizados de TAL. a) Termoestabilidad. La inactivación se realizo a pH 7 y 70ºC; b) Estabilidad frente a pH 9 y 25ºC. c) Estabilidad frente a pH 5 y 25ºC. Octadecil-Sepabeads-TAL (�),enzima soluble(` ).
Respecto a la estabilidad frente al pH, vimos como a pH 9 el derivado octadecil-
Sepabeads-TAL mantenía casi intacta su actividad tras 5 días de incubación mientras la
enzima soluble a penas conservaba un 40% de actividad (Figura 18b). Cuando el pH se
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 10101010 20202020 30303030 40404040 50505050 60606060 70707070
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 1111 2222 3333 4444 5555
Tiempo (dTiempo (dTiempo (dTiempo (díííías)as)as)as)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Capítulo 2
53
disminuyó hasta 5, de nuevo el derivado inmovilizado fue totalmente activo tras cinco días,
mientras en este caso, la soluble mantuvo hasta un 60% de actividad residual (Figura 18c).
Todos estos resultados muestran como el derivado interfacialmente adsorbido, octadecil-
Sepabeads, fue siempre el derivado inmovilizado más estable, manteniendo el 100% de
actividad en la mayoría de las lipasas estudiadas, tanto en inactivación térmica como en
presencia de disolvente, además en condiciones mixtas de co-disolvente y temperatura,
estabilizándose incluso lipasas termófilas las cuales ya era estables a condiciones bastante
drásticas. Por tanto, parece que la conformación abierta de la lipasa es más estable incluso
que la lipasa inmovilizada mediante unión covalente multipuntual.
4.4.4.4. Efecto de la inmovilización sobre la temperatura Efecto de la inmovilización sobre la temperatura Efecto de la inmovilización sobre la temperatura Efecto de la inmovilización sobre la temperatura óóóóptima de lipasasptima de lipasasptima de lipasasptima de lipasas
La estabilización de la forma abierta de la lipasa tras la adsorción interfacial sobre la
resina octadecil-Sepabeads ha quedado patente tras los estudios realizados anteriormente.
Teniendo en cuenta este hecho, quisimos ver que efecto podía tener esta estabilización en la
temperatura óptima de las distintas lipasas, comparándose la curva actividad-temperatura de
la enzima soluble en condiciones en las que se encuentra totalmente libre sin existir ningún
tipo de autoasociación lipásica (mayoritariamente en su forma cerrada) con el derivado
octadecil-Sepabeads (estructura abierta). Esto se realizó en condiciones donde el sustrato era
totalmente soluble (evitando así la adsorción interfacial de la lipasa soluble sobre gotas de
sustrato). En el caso de la lipasa de C. rugosa, la enzima libre presentó un máximo de
actividad a 45ºC mientras la temperatura óptima del derivado octadecil-Sepabeads fue de
55ºC (Figura 19). En todos los demás casos también se observó un aumento del óptimo de
temperatura en unos diez grados, de 55ºC a 65ºC para la lipasa de C. antarctica B, de 56ºC a
64ºC para la MML (resultados no mostrados).
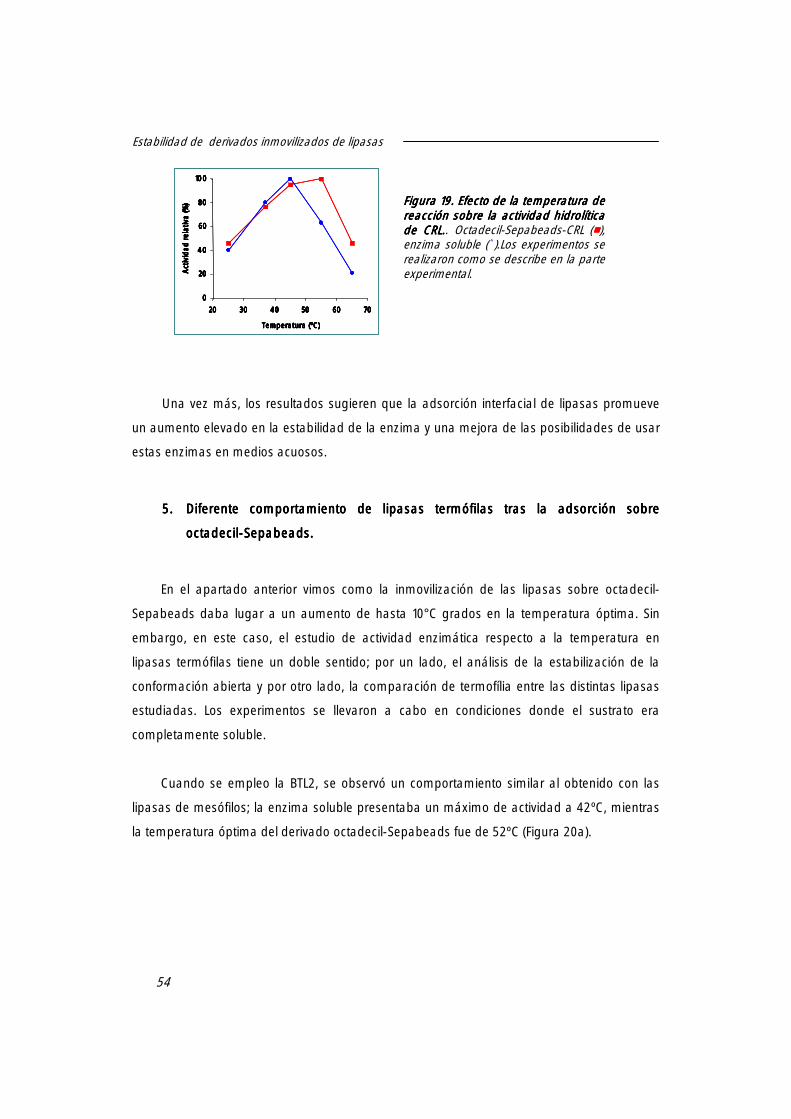
Estabilidad de derivados inmovilizados de lipasas
54
Figura Figura Figura Figura 19191919. Efecto de la temperatura de . Efecto de la temperatura de . Efecto de la temperatura de . Efecto de la temperatura de reacción sobre la areacción sobre la areacción sobre la areacción sobre la actividad hidrolítica ctividad hidrolítica ctividad hidrolítica ctividad hidrolítica de CRL.de CRL.de CRL.de CRL.. Octadecil-Sepabeads-CRL (����), enzima soluble (` ).Los experimentos se realizaron como se describe en la parte experimental.
Una vez más, los resultados sugieren que la adsorción interfacial de lipasas promueve
un aumento elevado en la estabilidad de la enzima y una mejora de las posibilidades de usar
estas enzimas en medios acuosos.
5.5.5.5. Diferente comportamiento de lipasaDiferente comportamiento de lipasaDiferente comportamiento de lipasaDiferente comportamiento de lipasas s s s termófilastermófilastermófilastermófilas tras la tras la tras la tras la adsorciónadsorciónadsorciónadsorción sobre sobre sobre sobre
octadeciloctadeciloctadeciloctadecil----Sepabeads. Sepabeads. Sepabeads. Sepabeads.
En el apartado anterior vimos como la inmovilización de las lipasas sobre octadecil-
Sepabeads daba lugar a un aumento de hasta 10°C grados en la temperatura óptima. Sin
embargo, en este caso, el estudio de actividad enzimática respecto a la temperatura en
lipasas termófilas tiene un doble sentido; por un lado, el análisis de la estabilización de la
conformación abierta y por otro lado, la comparación de termofília entre las distintas lipasas
estudiadas. Los experimentos se llevaron a cabo en condiciones donde el sustrato era
completamente soluble.
Cuando se empleo la BTL2, se observó un comportamiento similar al obtenido con las
lipasas de mesófilos; la enzima soluble presentaba un máximo de actividad a 42ºC, mientras
la temperatura óptima del derivado octadecil-Sepabeads fue de 52ºC (Figura 20a).
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
Act
ivid
ad r
elat
iva
(%)
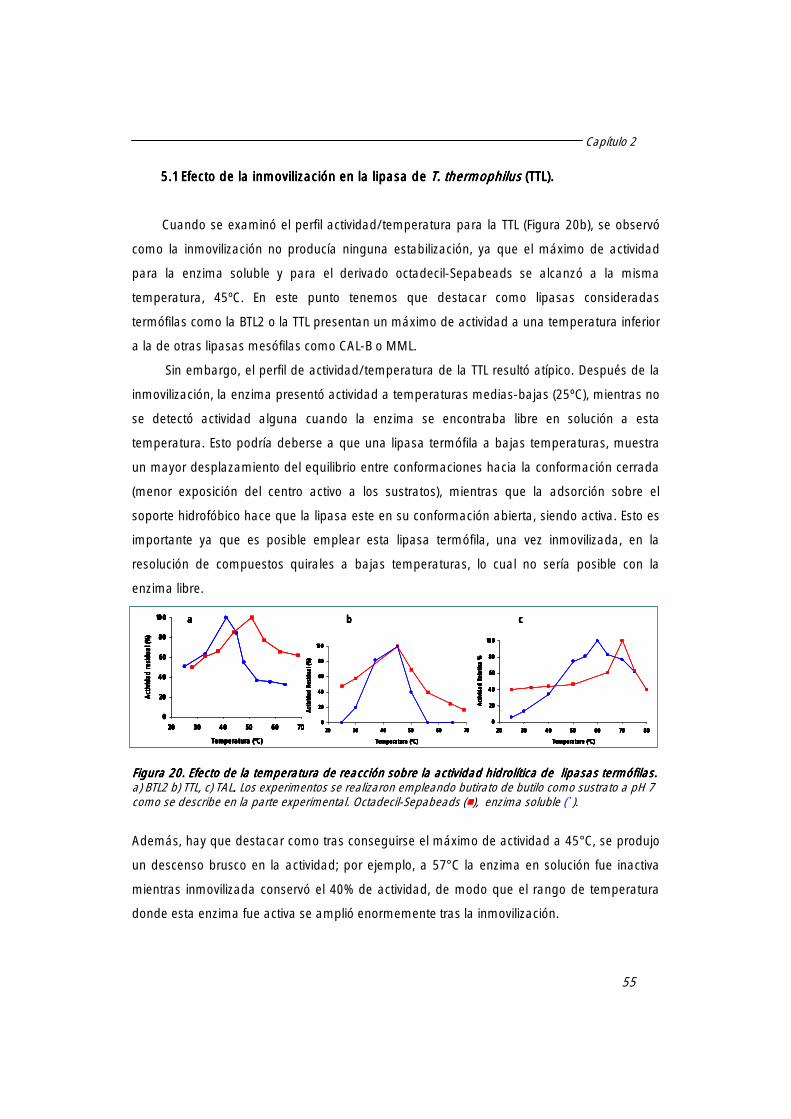
Capítulo 2
55
5555.1 Efecto de la inmovilización.1 Efecto de la inmovilización.1 Efecto de la inmovilización.1 Efecto de la inmovilización en la lipasa de en la lipasa de en la lipasa de en la lipasa de T. thermophilusT. thermophilusT. thermophilusT. thermophilus (TTL). (TTL). (TTL). (TTL).
Cuando se examinó el perfil actividad/temperatura para la TTL (Figura 20b), se observó
como la inmovilización no producía ninguna estabilización, ya que el máximo de actividad
para la enzima soluble y para el derivado octadecil-Sepabeads se alcanzó a la misma
temperatura, 45ºC. En este punto tenemos que destacar como lipasas consideradas
termófilas como la BTL2 o la TTL presentan un máximo de actividad a una temperatura inferior
a la de otras lipasas mesófilas como CAL-B o MML.
Sin embargo, el perfil de actividad/temperatura de la TTL resultó atípico. Después de la
inmovilización, la enzima presentó actividad a temperaturas medias-bajas (25ºC), mientras no
se detectó actividad alguna cuando la enzima se encontraba libre en solución a esta
temperatura. Esto podría deberse a que una lipasa termófila a bajas temperaturas, muestra
un mayor desplazamiento del equilibrio entre conformaciones hacia la conformación cerrada
(menor exposición del centro activo a los sustratos), mientras que la adsorción sobre el
soporte hidrofóbico hace que la lipasa este en su conformación abierta, siendo activa. Esto es
importante ya que es posible emplear esta lipasa termófila, una vez inmovilizada, en la
resolución de compuestos quirales a bajas temperaturas, lo cual no sería posible con la
enzima libre.
Figura 2Figura 2Figura 2Figura 20000.... Efecto de la temperatura de reacción sobre la actividad hidrolítica de lipasas termófilas. Efecto de la temperatura de reacción sobre la actividad hidrolítica de lipasas termófilas. Efecto de la temperatura de reacción sobre la actividad hidrolítica de lipasas termófilas. Efecto de la temperatura de reacción sobre la actividad hidrolítica de lipasas termófilas. a) BTL2 b) TTL, c) TAL. . . . Los experimentos se realizaron empleando butirato de butilo como sustrato a pH 7 como se describe en la parte experimental. Octadecil-Sepabeads (����), enzima soluble (` ).
Además, hay que destacar como tras conseguirse el máximo de actividad a 45°C, se produjo
un descenso brusco en la actividad; por ejemplo, a 57°C la enzima en solución fue inactiva
mientras inmovilizada conservó el 40% de actividad, de modo que el rango de temperatura
donde esta enzima fue activa se amplió enormemente tras la inmovilización.
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070 80808080
TemperatureTemperatureTemperatureTemperature ((((ººººCCCC))))
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070 80808080
TemperatureTemperatureTemperatureTemperature ((((ººººCCCC))))
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
aaaa
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
cccc
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070 80808080
TemperatureTemperatureTemperatureTemperature ((((ººººCCCC))))
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070 80808080
TemperatureTemperatureTemperatureTemperature ((((ººººCCCC))))
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
Act
ivid
ad R
elat
iva
%A
ctiv
idad
Rel
ativ
a %
bbbb
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
20202020 30303030 40404040 50505050 60606060 70707070
Temperatura (Temperatura (Temperatura (Temperatura (ººººCCCC))))
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)
Act
ivid
ad R
esid
ual (
%)

Estabilidad de derivados inmovilizados de lipasas
56
5.2 Efecto de la 5.2 Efecto de la 5.2 Efecto de la 5.2 Efecto de la inmovinmovinmovinmovilizaciónilizaciónilizaciónilización sobre sobre sobre sobre la lipasa de la lipasa de la lipasa de la lipasa de T. aquaticusT. aquaticusT. aquaticusT. aquaticus ( ( ( (TALTALTALTAL))))
El estudio de estabilidad de esta enzima mostró como una vez inmovilizada sobre
octadecil-Sepabeads se consiguió una alta estabilización en comparación con la enzima en
solución.
Asimismo, cuando se examinó el perfil de actividad/temperatura de TAL (Figura 20c),
se observó que tras la inmovilización, la enzima alcanzaba un máximo de actividad a los 70°C
mientras que cuando la enzima se encontraba en solución este máximo era alcanzado a los
60°C. Este hecho nos muestra, por un lado, la mayor estabilidad de la conformación abierta
de la lipasa, y por otro, que la TAL fue la lipasa de mayor termofília.
No obstante, un comportamiento interesante fue que tras la inmovilización, el perfil de
actividad/temperatura de la enzima cambió respecto a la enzima soluble. A 25°C la enzima
inmovilizada presentaba casi un 40% más de actividad que la lipasa soluble la cual era casi
inactiva, aunque en este caso la actividad de la enzima se mantuvo intacta en el derivado
hasta los 50°C, a partir de donde empezó a incrementarse. Por lo tanto, esta enzima tras su
inmovilización presenta una actividad casi constante desde los 25°C a los 80°C, siendo más
alta entre los 65 y 75°C.

Capítulo 2
57
CONCLUSIONESCONCLUSIONESCONCLUSIONESCONCLUSIONES
Los diferentes métodos de inmovilización han determinado un comportamiento
similar en la estabilidad de lipasas de distinto origen, encontrándose que la conformación
abierta de las lipasas tras la absorción hidrofóbica sobre resina octadecil-Sepabeads fue, en
todos los casos, más estable incluso que la lipasa inmovilizada por enlace covalente
multipuntual.
Cuando se emplearon enzimas termófilas, donde ya existe una estabilidad frente a
temperatura elevada, se obtuvo una estabilización tras la adsorción de la conformación
abierta de la lipasa sobre soportes hidrofóbicos. Además, la inmovilización sobre octadecil-
Sepabeads de TTL y TAL, permitió el empleo de estas termófilas en un amplio rango de
temperaturas; por ejemplo a 25ºC, condiciones donde la enzima libre era muy poca o
totalmente inactiva.
Además, en el perfil actividad-temperatura se observó como, en los casos de CRL,
MML, CAL-B, BTL2, TAL, la temperatura óptima de la enzima aumentó en torno a 10ºC respecto
a la soluble tras la adsorción sobre el soporte octadecil-Sepabeads, mientras para la TTL, la
temperatura óptima de la enzima soluble y del derivado fue la misma.

Estabilidad de derivados inmovilizados de lipasas
58
PARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTAL
1.1.1.1. MaterialesMaterialesMaterialesMateriales
La lipasa de Bacillus thermocatenulatus (BTL2) fue cedida por Maria luisa Rua (Universidad
de Vigo).Las lipasas de Thermus thermophilus (62296) (TTL) y Thermus aquaticus (TAL) (62293)
fueron obtenidas de Fluka. Los geles octil-Sepharose 4BCL y bromuro de cianógeno (BrCN-
activado Sepharose 4BCL) fueron obtenido de Pharmacia Biotech (Uppsala, Suecia).
Los geles de agarosa 6 BCL y 10 BCL fueron donados por la compañía Hispanagar SA
(Burgos). Octadecil-Sepabeads fue donado por Resindion srl (Milán, Italia). Además el
glutaraldehido al 25%, tritón X-100, butirato de etilo, butirato de butilo, p-nitrofenil propionato
(pNPP), p-nitrofenil butirato (pNPB), α-naftilacetato y Fast Blue RR se obtuvieron de sigma. Los
soportes glioxil-agarosa69 y glutaraldehído-agarosa80-81 se prepararon como esta descrito en la
literatura.
2.2.2.2. Purificación de lipasas. Purificación de lipasas. Purificación de lipasas. Purificación de lipasas.
La purificación de BTL2, TTL y TAL se realizó como fue descrito en el capítulo 1 para
CAL-B, CRL, HHL, MML y PFL. La concentración proteica de TTL fue 0,95 mg/mL (10 mg
polvo/mL); TAL presentó 0,5 mg/mL (10 mg polvo/mL) y el extracto de BTL2 contenía 15 mg/mL
determinada utilizando el Método de Bradford 70.
La cinética de inmovilización se llevo a cabo espectrofotométricamente a través de la
hidrólisis de pNPP (como se describe en el capítulo 1) para BTL2 a 348 nm, pH 7 y 25ºC o
mediante pHstato con butirato de butilo para TTL a 45ºC y para TAL a 65ºC midiendo actividad
de sobrenadante y suspensión de manera periódica. Siguiendo este protocolo se obtuvo una
inmovilización cuantitativa de la actividad de las distintas lipasas, con rendimientos finales de
inmovilización en torno al 100%. Después de la inmovilización el derivado enzimático fue
lavado abundantemente con agua destilada y se procedió a la desorción de la lipasa a pH 7 y
25ºC mediante la adición de Tritón X-100 necesaria (0,2% para BTL2, 1% para TTL y 1% para
TAL).

Capítulo 2
59
3.3.3.3. Electroforesis en gel poliaElectroforesis en gel poliaElectroforesis en gel poliaElectroforesis en gel poliacrilamidacrilamidacrilamidacrilamida----SDSSDSSDSSDS----PAGEPAGEPAGEPAGE
La electroforesis se realizó como se describe en el capítulo 1.La visualización de las
proteínas se realizó mediante tinción con plata.
4.4.4.4. Electroforesis en condiciones no desnaturalizantesElectroforesis en condiciones no desnaturalizantesElectroforesis en condiciones no desnaturalizantesElectroforesis en condiciones no desnaturalizantes (nativ (nativ (nativ (nativoooo))))
La preparación de la electroforesis nativa se realizó de la misma manera que la SDS-
PAGE descrita en el capítulo 1, en ausencia de SDS y βmercaptoetanol, además no se hirvieron
las muestras.
La determinación de actividad en el extracto de BTL2 se llevo a cabo mediante
electroforesis nativa con tinción específica de lipasas. Para ello se realizó la electroforesis en
condiciones nativas con 1% de tritón en el electrolito, cargando 10µl de extracto crudo con 2,5
% de tritón y de lipasa previamente desorbida con tritón con una concentración final, en
ambos casos, de 2 mg/mL. Después de esto, el gel se incubó en una solución de tampón
fosfato sódico 0,1 M pH=6,2 conteniendo 0,02% de α-naftilacetato (disuelto en acetona) y Fast
Blue RR (0,05%)103. La actividad se detectó en 15 minutos.
5.5.5.5. InmovilizacInmovilizacInmovilizacInmovilización de lipasas sobre distintos soportesión de lipasas sobre distintos soportesión de lipasas sobre distintos soportesión de lipasas sobre distintos soportes
Se prepararon derivados inmovilizados con concentración de lipasa en torno a 1 mg
de lipasa pura/mL, evitando así problemas de difusión. En todos los casos se obtuvo más de
un 95% de enzima inmovilizada. Tras la purificación se mantuvo el tritón en la solución
enzimática evitándose así la formación de estructuras bimoleculares de lipasa76-77. El
seguimiento de la inmovilización se llevo a cabo por hidrólisis de pNPP (CAL-B, CRL, HLL, BTL2),
pNPB (MML) a 25ºC o butirato de butilo (TTL a 45ºC, TAL a 65ºC) y la fuerza iónica empleada
fue de 5 mM (tampón fosfato sódico pH 7, bicarbonato pH 9-11), a excepción de CRL donde se
empleó 25 mM.
5555.1 .1 .1 .1 Preparación de derivado inmovilizado por adsorción interfacial de las Preparación de derivado inmovilizado por adsorción interfacial de las Preparación de derivado inmovilizado por adsorción interfacial de las Preparación de derivado inmovilizado por adsorción interfacial de las
distintasdistintasdistintasdistintas lipasas. lipasas. lipasas. lipasas.
La lipasa purificada se diluyó unas 10 veces empleando agua destilada (para diluir el
Tritón X-100) y se añadió la resina octadecil-Sepabeads. El seguimiento de la inmovilización se
realizó mediante el análisis de muestras de suspensión y sobrenadante a distintos tiempos. La

Estabilidad de derivados inmovilizados de lipasas
60
inmovilización se realizó a baja fuerza iónica, pH 7 y 25ºC. En algunos casos, preparaciones
de enzima no purificadas daban resultados similares, de forma que estos soportes también
pueden ser utilizados para inmovilización selectiva de lipasas al igual que el octil-agarosa34,79.
5555.2 Preparación de .2 Preparación de .2 Preparación de .2 Preparación de los los los los derivadoderivadoderivadoderivadossss covalente covalente covalente covalentessss de lipasas. de lipasas. de lipasas. de lipasas.
5.2.1 Preparación del derivado bromocianógeno-agarosa (BrCN).
A 9 mL de una solución de lipasa purificada en tampón fosfato sódico 5 mM y pH 7 se
añadió 3 gramos de soporte BrCN. Después de 3 horas de inmovilización el derivado se
incubó con etanolamina 0,5 M a pH 7 durante 20 minutos para acabar de destruir grupos
libres. El rendimiento de inmovilización fue superior al 95%.
5.2.2 Inmovilización de lipasas sobre soportes glutaraldehído.
El soporte glutaraldehído-agarosa se ofreció a la enzima purificada. Después de 5
horas de inmovilización, como punto final de la reacción entre el enzima y el soporte, se
añadió un volumen de 100 mM de bicarbonato sódico conteniendo 2 mg/mL de borohidruro
sódico a pH 8,5 durante 30 minutos, tras lo cual el derivado se lavó con abundante agua
destilada.
5.2.3 Inmovilización de lipasas sobre soporte glioxil-agarosa.
Este protocolo de inmovilización fue seleccionado para obtener preparaciones de
lipasas inmovilizadas mediante enlaces covalentes multipuntuales, donde era posible esperar
cierta rigidificación de la estructura de la enzima 69.
Glioxil-agarosa se añadió a las diferentes soluciones enzimáticas, y el pH se aumentó
hasta 10,5. Únicamente en el caso de la CRL no fue posible la inmovilización debido a la
inactivación de esta a este pH. Después de 5 horas se obtuvo una inmovilización completa de
la enzima, manteniendo la inmovilización durante 20 horas, asegurándonos así una unión
covalente multipuntual entre enzima y soporte. Estas multi-interacciones entre la enzima y el
soporte se finalizaron al añadir 1 mg de borohidruro sódico por mL de suspensión durante 30
minutos lavando posteriormente el derivado inmovilizado con abundante agua.

Capítulo 2
61
6.6.6.6. Ensayos de determinación de actividadEnsayos de determinación de actividadEnsayos de determinación de actividadEnsayos de determinación de actividad
El ensayo de hidrólisis enzimática con los sustratos pNPP, pNPB y butirato de etilo ya fue
descrito en el capítulo anterior.
6.1 Hidrólisis de Butirato de butilo.6.1 Hidrólisis de Butirato de butilo.6.1 Hidrólisis de Butirato de butilo.6.1 Hidrólisis de Butirato de butilo.
Este ensayo se realizó empleando un pHstato Mettler Toledo D50, midiendo el ácido
butírico producido por la hidrólisis enzimática del butirato de butilo 30 mM en tampón fosfato
sódico 50 mM a pH 7 y 45ºC (para TTL) y a 65ºC (para TAL). Hidróxido sódico 10 mM fue
empleado como agente titulante.
7.7.7.7. Estabilidad de los diferentes derivados inmovilizados de lipasas.Estabilidad de los diferentes derivados inmovilizados de lipasas.Estabilidad de los diferentes derivados inmovilizados de lipasas.Estabilidad de los diferentes derivados inmovilizados de lipasas.
Los diferentes derivados inmovilizados se incubaron en las diferentes condiciones
descritas (pH, tª, presencia de co-disolventes) y periódicamente se fueron tomando muestras
de estas reacciones de inactivación, ensayándose la actividad de estas mediante los métodos
anteriormente descritos.
8.8.8.8. Perfil de TemperaturaPerfil de TemperaturaPerfil de TemperaturaPerfil de Temperatura----actividad enzimática de las diferactividad enzimática de las diferactividad enzimática de las diferactividad enzimática de las diferentes prepaentes prepaentes prepaentes preparaciones de raciones de raciones de raciones de
lipasas.lipasas.lipasas.lipasas.
El efecto de la temperatura sobre la actividad enzimática de las preparaciones de las distintas
lipasas fue comprobado en la hidrólisis del butirato de etilo (50 mM) o butirato de butilo (30
mM) en tampón fosfato sódico 50 mM a pH 7. El tampón fue preincubado para alcanzar la
temperatura deseada antes de añadir el sustrato y la enzima.

Capitulo 3
62
IIIINTRODUCCIÓNNTRODUCCIÓNNTRODUCCIÓNNTRODUCCIÓN
1.1.1.1. Ingeniería Conformacional de lipasas.Ingeniería Conformacional de lipasas.Ingeniería Conformacional de lipasas.Ingeniería Conformacional de lipasas.
El complejo mecanismo catalítico presente en las lipasas hace posible pensar que la
alteración de este mecanismo de apertura y cierre o de la forma exacta de la conformación
abierta de la lipasa, podría dar lugar a una modificación en las propiedades catalíticas de la
misma. De esta forma, la posibilidad de controlar y modular este equilibrio se puede convertir
en una poderosa herramienta de mejora de las propiedades de las lipasas como enzimas de
interés industrial.
Por lo tanto, la hipótesis que planteamos se basa en que esta alteración del equilibrio
podría llevarse a cabo mediante el empleo de diferentes técnicas de inmovilización en las
cuales se involucren distintas áreas de la enzima, en donde sea posible controlar: i) la
orientación de la lipasa sobre el soporte, ii) la rigidez de las lipasas inmovilizadas, y iii) los
micro-ambientes que rodean las moléculas de lipasas inmovilizadas. Todo esto podría
provocar una modificación en el mecanismo de apertura y cierre de la lipasa, dando lugar a
una alteración en la forma exacta de la conformación abierta (activa) final, viéndose esto
reflejado en una modificación de sus propiedades de actividad y especificidad (enantio o
regioselectividad) (Esquema 15).
Esquema 1Esquema 1Esquema 1Esquema 15555. . . . Ingeniería conformacional de lipasas
IngenieríaIngenieríaIngenieríaIngeniería ConformacionalConformacionalConformacionalConformacional
CondiciónCondiciónCondiciónCondición 2222
CondiciónCondiciónCondiciónCondición 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
Forma cerrada
Forma abierta
Forma Forma Forma Forma cerradacerradacerradacerrada
LidLidLidLid
Centro Centro Centro Centro activoactivoactivoactivo
IngenieríaIngenieríaIngenieríaIngeniería ConformacionalConformacionalConformacionalConformacional
CondiciónCondiciónCondiciónCondición 2222
CondiciónCondiciónCondiciónCondición 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
Forma cerrada
Forma abierta
Forma Forma Forma Forma cerradacerradacerradacerrada
LidLidLidLid
Centro Centro Centro Centro activoactivoactivoactivo
IngenieríaIngenieríaIngenieríaIngeniería ConformacionalConformacionalConformacionalConformacional
CondiciónCondiciónCondiciónCondición 2222
CondiciónCondiciónCondiciónCondición 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 1111
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
DerivadoDerivadoDerivadoDerivado inmovilizadoinmovilizadoinmovilizadoinmovilizado 2222
Forma cerrada
Forma abierta
Forma Forma Forma Forma cerradacerradacerradacerrada
LidLidLidLid
Centro Centro Centro Centro activoactivoactivoactivo
Forma Forma Forma Forma cerradacerradacerradacerrada
LidLidLidLid
Centro Centro Centro Centro activoactivoactivoactivo

Ingeniería conformacional de lipasas
63
Además, un cambio en las condiciones de reacción podría llegar a tener un gran
efecto en las propiedades catalíticas de las lipasas, debido quizás a la modificación que se
produciría sobre las interacciones globales de las conformaciones presentes en el equilibrio
(abierta y cerrada), alterándose, de nuevo, la forma exacta de la conformación abierta de la
enzima (Esquema 10).
Esta modulación de las propiedades catalíticas de las lipasas podría denominarse
“Ingeniería Conformacional”, lo cual ha sido empleado para modular el comportamiento de
diferentes enzimas que sufren grandes cambios conformacionales durante la catálisis (por
ejemplo, penicilina G acilasa)104-105.
2. 2. 2. 2. Inmovilización de lipasas Inmovilización de lipasas Inmovilización de lipasas Inmovilización de lipasas conconconcon distintas orientaciones. distintas orientaciones. distintas orientaciones. distintas orientaciones.
2.1 2.1 2.1 2.1 Adsorción interfacial sobre soportes hidrofóbicos.Adsorción interfacial sobre soportes hidrofóbicos.Adsorción interfacial sobre soportes hidrofóbicos.Adsorción interfacial sobre soportes hidrofóbicos.
Como ya se comentó en el capítulo anterior, la interacción de lipasas sobre soporte
hidrofóbicos se produce a través del sitio activo, fijándose la conformación abierta.
2.2 Inmovilización covalente a través de pocas uniones.2.2 Inmovilización covalente a través de pocas uniones.2.2 Inmovilización covalente a través de pocas uniones.2.2 Inmovilización covalente a través de pocas uniones.
La inmovilización de lipasas sobre soportes glutaraldehído-agarosa y bromocianógeno-
agarosa, a pH 7, ocurre a través de algunas uniones covalentes, sin embargo la orientación
de la enzima en cada caso es distinta.
Mientras en el derivado agarosa-bromocianógeno, la enzima queda unida al soporte
principalmente por el amino terminal; en el derivado glutaraldehido, la naturaleza del soporte
provoca, en primer lugar, una adsorción de la enzima, sobre grupos amino del soporte,
orientándose por la zona más rica en cargas negativas y posteriormente, unión covalente a
través de algunos grupos aminos de la enzima cercanos al soporte.
2.3 Inmovilización mediante enlace covalente multip2.3 Inmovilización mediante enlace covalente multip2.3 Inmovilización mediante enlace covalente multip2.3 Inmovilización mediante enlace covalente multipuntual.untual.untual.untual.
En este caso, como ya se comentó en el capítulo anterior de esta tesis, la unión entre
la enzima y el soporte se produce a través de las zonas más ricas en lisinas de la proteína,
siendo una interacción muy intensa a través de un elevado número de enlaces.

Capitulo 3
64
2.4 2.4 2.4 2.4 Adsorción iónica de lipasas.Adsorción iónica de lipasas.Adsorción iónica de lipasas.Adsorción iónica de lipasas.
Se han utilizado unos nuevos soportes intercambiadores aniónicos, obtenidos por
modificación de grupos reactivos del soporte (p.e., aldehído, epóxido), mediante la reacción
de estos con grupos aminos no ionizados (primarios, secundarios y terciarios) de un polímero
policatiónico, polietilenimina68,106-107. El empleo de estos polímeros poli-catiónicos establece que
la fuerza de unión sea mucho mayor que utilizando soportes aniónicos convencionales (DEAE),
ya que el número de grupos cargados del soporte se ve altamente incrementado (unos 1000
µmol/mL de gel empaquetado). Además, estos grupos proceden de un polímero flexible
permitiendo una fuerte adsorción muy poco distorsionante: el polímero se adapta a la
proteína en lugar de forzar a la proteína a adaptarse al polímero.
Las lipasas quedaran inmovilizadas sobre este tipo de soporte a través de la zona con
mayor densidad de cargas negativas sobre la superficie de la proteína; por tanto, las
condiciones de pH deben ser las adecuadas, permitiendo a la enzima quedar sobre el
polímero, generándose un ambiente muy hidrofílico alrededor de un importante porcentaje de
la superficie externa de la proteína (Esquema 16).
Esquema 1Esquema 1Esquema 1Esquema 16666.... Adsorción iónica de lipasas sobre soportes PEI
2.52.52.52.5 Inmovilización sobre soportes heterofuncionales: Inmovilización sobre soportes heterofuncionales: Inmovilización sobre soportes heterofuncionales: Inmovilización sobre soportes heterofuncionales: Adsorción fAdsorción fAdsorción fAdsorción fíííísica + unión covalentesica + unión covalentesica + unión covalentesica + unión covalente....
Los soportes Eupergit/Sepabeads comerciales son soportes epoxiacrílicos, con una
mayor o menor hidrofobicidad. Estos presentan poros con grandes superficies recubiertas de
grupos epóxido; los cuales, en principio, son capaces de reaccionar con varios grupos de la
proteína dependiendo de las condiciones, dando directamente uniones muy estables: grupos
amino (amino secundario), tioles (tio-éteres), hidroxilo (éteres). Sin embargo, los grupos
• Una superficie flexible conteniendo una alta concentraciónde grupos amino
• Fuerte adsorción iónica• Orientación: área cercana a la zona con mayor carga neta negativa
• Una superficie flexible conteniendo una alta concentraciónde grupos amino
• Fuerte adsorción iónica• Orientación: área cercana a la zona con mayor carga neta negativa
• Una superficie flexible conteniendo una alta concentraciónde grupos amino
• Fuerte adsorción iónica• Orientación: área cercana a la zona con mayor carga neta negativa

Ingeniería conformacional de lipasas
65
epóxidos, en condiciones suaves (pH 7, baja fuerza iónica) presentan una reactividad muy
baja con las enzimas solubles.
De esta manera, se ha propuesto que el proceso de inmovilización de enzimas a
soportes con grupos epóxido se produce siguiendo un mecanismo en dos etapas 108-109
(Esquema 17):
1º.- la adsorción física de la enzima sobre el soporte, lo cual permite acercamiento de
la enzima al soporte (tradicionalmente, una adsorción hidrofóbica),
2º.-la unión covalente entre la enzima adsorbida y los grupos epóxido del soporte.
Esquema 1Esquema 1Esquema 1Esquema 17777. . . . Adsorción física más inmovilización covalente sobre soportes epóxido
No obstante, esta adsorción física de las proteínas sobre el soporte no tiene por que
ser exclusivamente hidrofóbica; así, la modificación de un pequeño porcentaje de grupos
epóxido del soporte con distintos compuestos (iminodiacético (IDA), etilendiamina (EDA),
quelato de cobre (IDA-Cu) puede provocar diferente tipo de adsorción de las moléculas de
proteína sobre el soporte (Esquema 18). Este tipo de soportes heterofuncionales adsorberán
las proteínas a través de diferentes zonas dependiendo del compuesto introducido en el
soporte, lo cual marcará la orientación final de la enzima inmovilizada:
-Soporte epóxido-etilendiamina: la proteína se absorberá sobre este soporte a través
de las zonas de su superficie más ricas en cargas negativas.
-Soporte epóxido-iminodiacético: la adsorción de la proteína se producirá a través de
las zonas de la superficie con mayor número de cargas positivas.
-Soporte epóxido-iminodiacético-Cu: la adsorción de la proteína se realizará a través
de la zona con un mayor número de residuos de histidinas.
Inmovilización covalente a través de grupos (NH2,SH,OH) cercanos al bolsillo hidrofóbico
1M 1M 1M 1M fosfatofosfatofosfatofosfato pH 7pH 7pH 7pH 7
AdsorciAdsorciAdsorciAdsorcióóóónnnn hidrofhidrofhidrofhidrofóóóóbicabicabicabica
SoporteSoporteSoporteSoporte hidrofóbicohidrofóbicohidrofóbicohidrofóbico
InmovilizaciInmovilizaciInmovilizaciInmovilizacióóóónnnn covalentecovalentecovalentecovalenteintramolecularintramolecularintramolecularintramolecular
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
1111
2222
OOOO
HHHH2222NNNN
BolsillosBolsillosBolsillosBolsillos hidrofhidrofhidrofhidrofóóóóbicosbicosbicosbicos En En En En la la la la superficiesuperficiesuperficiesuperficie de la de la de la de la proteínaproteínaproteínaproteína
Inmovilización covalente a través de grupos (NH2,SH,OH) cercanos al bolsillo hidrofóbico
1M 1M 1M 1M fosfatofosfatofosfatofosfato pH 7pH 7pH 7pH 7
AdsorciAdsorciAdsorciAdsorcióóóónnnn hidrofhidrofhidrofhidrofóóóóbicabicabicabica
SoporteSoporteSoporteSoporte hidrofóbicohidrofóbicohidrofóbicohidrofóbico
InmovilizaciInmovilizaciInmovilizaciInmovilizacióóóónnnn covalentecovalentecovalentecovalenteintramolecularintramolecularintramolecularintramolecular
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
1111
2222
OOOO
HHHH2222NNNN
BolsillosBolsillosBolsillosBolsillos hidrofhidrofhidrofhidrofóóóóbicosbicosbicosbicos En En En En la la la la superficiesuperficiesuperficiesuperficie de la de la de la de la proteínaproteínaproteínaproteína
Inmovilización covalente a través de grupos (NH2,SH,OH) cercanos al bolsillo hidrofóbico
1M 1M 1M 1M fosfatofosfatofosfatofosfato pH 7pH 7pH 7pH 7
AdsorciAdsorciAdsorciAdsorcióóóónnnn hidrofhidrofhidrofhidrofóóóóbicabicabicabica
SoporteSoporteSoporteSoporte hidrofóbicohidrofóbicohidrofóbicohidrofóbico
InmovilizaciInmovilizaciInmovilizaciInmovilizacióóóónnnn covalentecovalentecovalentecovalenteintramolecularintramolecularintramolecularintramolecular
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
1111
2222
OOOO
HHHH2222NNNN
BolsillosBolsillosBolsillosBolsillos hidrofhidrofhidrofhidrofóóóóbicosbicosbicosbicos En En En En la la la la superficiesuperficiesuperficiesuperficie de la de la de la de la proteínaproteínaproteínaproteína
1M 1M 1M 1M fosfatofosfatofosfatofosfato pH 7pH 7pH 7pH 7
AdsorciAdsorciAdsorciAdsorcióóóónnnn hidrofhidrofhidrofhidrofóóóóbicabicabicabica
SoporteSoporteSoporteSoporte hidrofóbicohidrofóbicohidrofóbicohidrofóbico
InmovilizaciInmovilizaciInmovilizaciInmovilizacióóóónnnn covalentecovalentecovalentecovalenteintramolecularintramolecularintramolecularintramolecular
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
1111
2222
OOOOOOOO
HHHH2222NNNNHHHH2222NNNNHHHH2222NNNNHHHH2222NNNN
BolsillosBolsillosBolsillosBolsillos hidrofhidrofhidrofhidrofóóóóbicosbicosbicosbicos En En En En la la la la superficiesuperficiesuperficiesuperficie de la de la de la de la proteínaproteínaproteínaproteínaBolsillosBolsillosBolsillosBolsillos hidrofhidrofhidrofhidrofóóóóbicosbicosbicosbicos En En En En la la la la superficiesuperficiesuperficiesuperficie de la de la de la de la proteínaproteínaproteínaproteína

Capitulo 3
66
Tras la adsorción de la enzima, esta se une covalentemente al soporte por reacción de grupos
amino, tiol o hidroxilo de la enzima.
3333. Derivados del ácido . Derivados del ácido . Derivados del ácido . Derivados del ácido 2222----hidroxihidroxihidroxihidroxi----fenilacético (ácido mandélico)fenilacético (ácido mandélico)fenilacético (ácido mandélico)fenilacético (ácido mandélico)
Los isómeros puros del ácido mandélico (3333) y sus correspondientes ésteres son muy
utilizados en síntesis orgánica.
Los ésteres del isómero R son empleados en la síntesis enzimática del antibiótico
cefamandol 110 y el ácido ópticamente puro puede ser utilizado para la resolución de
racematos111. Además, algunos derivados O-acilados del ácido mandélico se han empleado
como desplazantes en fase estacionaria quiral 112.Por su parte, el ester metílico del ácido
mandélico (1111) es empleado como intermediario en la síntesis de la Pemolina (Cylert®) (2-
Amino-5-fenil-4-(5H) oxazolona), estimulante del sistema nervioso central. (Esquema 19).
Esquema 19Esquema 19Esquema 19Esquema 19
Hasta el momento, la resolución óptica del ácido mandélico se ha llevado a cabo
mediante métodos diastereoméricos empleando como agentes, alcaloides o aminas
Esquema 1Esquema 1Esquema 1Esquema 18888. . . . Adsorción e inmovilización de lipasas con diferentes orientaciones
NH
O NH2
O
OH
O
OMe
1111
NH
O NH2
O
OH
O
OMe
1111
NH
O NH2
O
OH
O
OMe
1111
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
NHNHNHNH3333++++
NHNHNHNH 3333++++
NHNHNHNH 3333++++
NHNHNHNH 3333++++
E DAE DAE DAE DA----S epabeadsS epabeadsS epabeadsS epabeads
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
NHNHNHNH3333++++
NHNHNHNH 3333++++
NHNHNHNH 3333++++
NHNHNHNH 3333++++
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
NHNHNHNH3333++++NHNHNHNH3333++++
NHNHNHNH 3333++++NHNHNHNH 3333++++
NHNHNHNH 3333++++NHNHNHNH 3333++++
NHNHNHNH 3333++++NHNHNHNH 3333++++
E DAE DAE DAE DA----S epabeadsS epabeadsS epabeadsS epabeads
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----
NHNHNHNH ++++COOCOOCOOCOO ----
COOCOOCOOCOO ----
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----
NHNHNHNH ++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH ++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH ++++COOCOOCOOCOO ----
COOCOOCOOCOO ----
IDAIDAIDAIDA----S epabeadsS epabeadsS epabeadsS epabeads
F orma F orma F orma F orma cerradacerradacerradacerrada L idL idL idL id
Centro Centro Centro Centro ac tivoac tivoac tivoac tivo
F orma F orma F orma F orma cerradacerradacerradacerrada L idL idL idL id
Centro Centro Centro Centro ac tivoac tivoac tivoac tivo
E upergi tE upergi tE upergi tE upergi t ----CuCuCuCu
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
HisHisHisHis
HisHisHisHisNHNHNHNH++++
C OOC OOC OOC OO ----
COOCOOCOOCOO ----C uC uC uC u +2+2+2+2
NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----CuCuCuCu +2+2+2+2
E upergi tE upergi tE upergi tE upergi t ----CuCuCuCu
OOOO OOOO
OOOO OOOO
OOOOOOOO
OOOO OOOO
HisHisHisHis
HisHisHisHis
HisHisHisHis
HisHisHisHisNHNHNHNH++++
C OOC OOC OOC OO ----
COOCOOCOOCOO ----C uC uC uC u +2+2+2+2
NHNHNHNH++++C OOC OOC OOC OO ----
COOCOOCOOCOO ----NHNHNHNH++++C OOC OOC OOC OO ----
COOCOOCOOCOO ----NHNHNHNH++++C OOC OOC OOC OO ----
COOCOOCOOCOO ----C uC uC uC u +2+2+2+2
NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----CuCuCuCu +2+2+2+2
NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----NHNHNHNH++++COOCOOCOOCOO ----
COOCOOCOOCOO ----CuCuCuCu +2+2+2+2
His His His His His His His His
NHNHNHNH 3333++++
NHNHNHNH3333++++
---------------- ----
His His His His His His His His
NHNHNHNH 3333++++
NHNHNHNH3333++++
His His His His His His His His
NHNHNHNH 3333++++
NHNHNHNH3333++++
His His His His His His His His
NHNHNHNH 3333++++
NHNHNHNH3333++++
---------------- -------------------- ----
---------------- -------------------- ----
NHNHNHNH3333++++
NHNHNHNH 3333++++
NHNHNHNH3333++++
NHNHNHNH 3333++++
NHNHNHNH3333++++
NHNHNHNH 3333++++
<

Ingeniería conformacional de lipasas
67
ópticamente activas (por ejemplo L-fenilalanina o L-metionina), aunque presentan el
inconveniente de un coste elevado, lo cual los hace inviables para la producción del ácido a
escala industrial.
Además, los enantiómeros del ácido mandélico presentan gran interés, ya que son
precursores en la síntesis de varios fármacos.
La Oxibutinina (Esquema 20), receptor antagonista muscarínico, es un fármaco de
gran interés en la industria farmacéutica el cual está indicado para el tratamiento de la
incontinencia urinaria. Sin embargo, efectos secundarios no deseados (sequedad de
mucosas, visión borrosa, nauseas, desvelo y palpitaciones cardiovasculares) limitan su utilidad
clínica.
Teniendo en cuenta que el isómero S presenta toda la actividad farmacológica, debía
buscarse una estrategia de síntesis para la obtención del fármaco enantioméricamente puro.
Durante el desarrollo de la síntesis asimétrica del (S)-ácido (Esquema 20), precursor quiral del
S-oxibutinina, químicos de Sepracor desarrollaron un nuevo y práctico proceso empezando
por el ácido S-mandélico (S-3333) y ciclohexanona113. Para la economía del proceso sintético, es
necesario optimizar la etapa anterior donde se lleva a cabo la resolución enantioselectiva del
ácido mandélico de forma enzimática 114.
Esquema Esquema Esquema Esquema 20202020. . . . Ruta retrosintética de la S-oxibutinina
Otro ejemplo es el (+)-Goniodiol, [(1´R,2´S,5R)-5-(1´,2´-dihidroxi-2-feniletill)-pent-2-eno-5-
lactona)] (Esquema 21), una estiril-lactona que exhibe una potente y selectiva citotoxicidad
frente al carcinoma de pulmón humano115. Debido a sus características estructurales y a su
potente actividad biológica, varios grupos se plantearon la síntesis de esta estiril-lactona a
partir del isómero R del ácido mandélico116-118; empleando el 2,3-O-isopropiliden-D-
gliceraldehído117 como bloque estructural, o basándose en reacciones asimétricas de
formación estereoselectiva de enlace carbono-carbono con complejos de cromo tricarbonil(η6-
OH
O
H OHOH
O
OH
O
OHO
N
S -ácidoS -Oxibutinina S -3333
Capitulo 3
68
areno)119 o bien mediante reacciones de dihidroxilación y epoxidación asimétrica de
Sharpless118-120 .
Esquema Esquema Esquema Esquema 21212121. . . . Ruta retrosintética de (+)-Goniodiol
O
OH
OH
O OTBS
CO2Et
OH
O
OH
R -3333(+)-Goniodiol

Ingeniería conformacional de lipasas
69
OBJETIVOS OBJETIVOS OBJETIVOS OBJETIVOS
En el presente capítulo nos centraremos en el estudio de las propiedades de actividad
y enantioselectividad de los distintos derivados inmovilizados de diferentes lipasas (CAL-B,
CRL, MML, BTL2, TTL, TAL) en donde se pretende ver si la hipótesis propuesta en la
introducción es correcta (la enzima presenta muy distintas propiedades dependiendo de
la metodología de inmovilización empleada).
Además se estudiará el efecto que ligeros cambios en las condiciones de reacción
(pH,Tª,..) pueden producir sobre las propiedades de los distintos derivados inmovilizados.
Para ello se empleará la resolución enzimática de varios compuestos quirales,
derivados del ácido mandélico, como sustratos modelos mediante reacción de hidrólisis
en medios acuosos.

Capitulo 3
70
RESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓNRESULTADOS Y DISCUSIÓN
Mediante el uso de distintos sustratos como modelos se intentó estudiar las
propuestas de la “Ingeniería Conformacional” en la modulación de las propiedades catalíticas
de distintas lipasas. Los sustratos empleados fueron el butirato de etilo (5555) ,éster alifático de
cadena corta, y dos compuestos quirales: mandelato de metilo [(±)-1111], con centro asimétrico
en el donador de acilo; y el ácido 2-O-butiril-2-fenilacético [(±)-7777], donde el centro quiral se
encuentra en el resto acilo, sustratos de peor reconocimiento para las lipasas.
La reacción empleada fue la hidrólisis en medios acuosos catalizada por los
derivados inmovilizados de las distintas lipasas (Esquema 22).
Esquema Esquema Esquema Esquema 22222222. Reacción de hidrólisis de los distintos sustratos empleando diferentes derivados inmovilizados de CAL-B.
áááácido 2cido 2cido 2cido 2----OOOO----butirilbutirilbutirilbutiril----2222---- fenilaceticofenilaceticofenilaceticofenilacetico
Butirato de etiloButirato de etiloButirato de etiloButirato de etilo
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
áááácidocidocidocido butbutbutbut ííííricoricoricorico
(5)(5)(5)(5)
((((±))))----1111
((((±))))----7777
SSSS----1111
Mandelato de Mandelato de Mandelato de Mandelato de metilometilometilometilo
(6)(6)(6)(6)
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
****
O
O
O
OH
OH
COOH
OH
COOH
O
O
OHO
O
O
OHO
++++
++++
O
OEt
O
OH ++++ EtOH
SSSS---- áááácido cido cido cido mandelicomandelicomandelicomandelico (2)(2)(2)(2)
COOMe
OH
OH
COOH COOMe
OH
OH
COOH COOMe
OH
++++
++++
RRRR----7777
****
áááácido 2cido 2cido 2cido 2---- OOOO----butirilbut irilbut irilbut iril---- 2222---- fenilaceticofenilaceticofenilaceticofenilacetico
Butirato de et iloButirato de et iloButirato de et iloButirato de et ilo
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
áááácidocidocidocido butbutbutbut ííííricoricoricorico
(5)(5)(5)(5)
((((±))))---- 1111
((((±))))---- 7777
SSSS---- 1111
Mandelato de Mandelato de Mandelato de Mandelato de metilometilometilometilo
(6)(6)(6)(6)
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
****
O
O
O
OH
OH
COOH
OH
COOH
O
O
OHO
O
O
OHO
++++
++++
O
OEt
O
OH ++++ EtOH
SSSS---- áááácido cido cido cido mandelicomandelicomandelicomandelico (2)(2)(2)(2)
COOMe
OH
OH
COOH COOMe
OH
OH
COOH COOMe
OH
++++
++++
RRRR----7777
****
áááácido 2cido 2cido 2cido 2---- OOOO----butirilbut irilbut irilbut iril---- 2222---- fenilaceticofenilaceticofenilaceticofenilacetico
Butirato de et iloButirato de et iloButirato de et iloButirato de et ilo
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
áááácidocidocidocido butbutbutbut ííííricoricoricorico
(5)(5)(5)(5)
((((±))))---- 1111
((((±))))---- 7777
SSSS---- 1111
Mandelato de Mandelato de Mandelato de Mandelato de metilometilometilometilo
(6)(6)(6)(6)
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
HHHH2222O/lipasasO/lipasasO/lipasasO/lipasas
****
O
O
O
OH
OH
COOH
OH
COOH
O
O
OHO
O
O
OHO
++++
++++
O
OEt
O
OH ++++ EtOH
SSSS---- áááácido cido cido cido mandelicomandelicomandelicomandelico (2)(2)(2)(2)
COOMe
OH
OH
COOH COOMe
OH
OH
COOH COOMe
OH
++++
++++
RRRR----7777
****

Ingeniería conformacional de lipasas
71
1.1.1.1. Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de éééésteres steres steres steres derivados del áderivados del áderivados del áderivados del ácido mandélico cido mandélico cido mandélico cido mandélico
catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de la lipasa de la lipasa de la lipasa de la lipasa de Candida Candida Candida Candida
antarctica Bantarctica Bantarctica Bantarctica B ( ( ( (CALCALCALCAL----BBBB))))....
1.1 Especificidad de los distintos derivados inmovilizados frente a distintos sustratos.1.1 Especificidad de los distintos derivados inmovilizados frente a distintos sustratos.1.1 Especificidad de los distintos derivados inmovilizados frente a distintos sustratos.1.1 Especificidad de los distintos derivados inmovilizados frente a distintos sustratos.
La primera propiedad catalítica analizada para los distintos derivados inmovilizados
de CAL-B fue la actividad específica en la reacción de hidrólisis de los distintos sustratos (Tabla
11).
En primer lugar, la reacción de hidrólisis se realizó a pH 7, con el objetivo de estudiar
dos efectos sobre la actividad catalítica de esta lipasa: i) la inmovilización sobre distintos
soportes y ii) la naturaleza del sustrato.
Empleando el butirato de etilo (5555), el derivado octadecil-Sepabeads-CAL-B presentó la
actividad específica más alta respecto a la de los demás derivados inmovilizados de CAL-B,
destacando la diferencia de actividad con el derivado Eupergit-Cu-CAL-B, el cual fue ochenta y
una veces menos activo. La actividad para los demás derivados respecto a octadecil-
Sepabeads-CAL-B fue entre tres y seis veces inferior.
Cuando se empleó el isómero R del mandelato de metilo (1111), se observó una
disminución significativa en el valor de actividad para todos los derivados inmovilizados.
Además, en este caso, el derivado PEI-CAL-B fue el más activo de todos los derivados de CAL-
B, con una actividad entre cinco y ocho veces superior a la de los derivados octadecil-
Sepabeads, glioxil o glutaraldehído de CAL-B. Una vez más, el derivado Eupergit-Cu-CAL-B
junto con el derivado bromocianógeno-CAL-B (BrCN) presentaron las actividades más bajas.
En la hidrólisis del ácido 2-O-butiril-2-fenilacético [(±)-7777], los derivados inmovilizados
de CAL-B presentaron actividades extremadamente más bajas respecto a las obtenidas frente
a los sustratos anteriores, quizás debido al mayor tamaño del resto acilo en el compuesto 7777.
En este caso, la mayor actividad específica para la CAL-B se encontró tras su inmovilización
sobre el soporte glutaraldehíido-agarosa, el cual fue 3 veces más activo que el derivado
glioxil-CAL-B (también derivado covalente). Sin embargo, las diferencias de actividades
respecto a los demás derivados de CAL-B fueron de más de 18 y 30 veces.
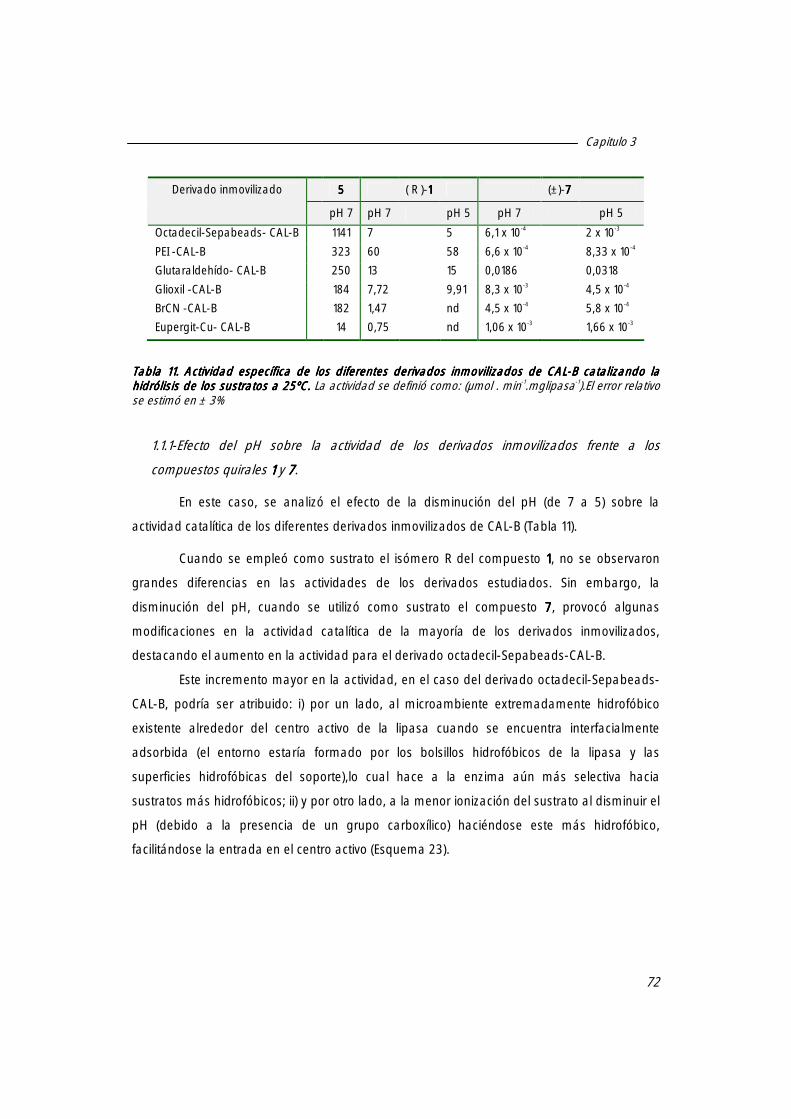
Capitulo 3
72
Tabla 11.Tabla 11.Tabla 11.Tabla 11. Actividad especActividad especActividad especActividad especíííífica de los diferentes derivados inmovilizados de CALfica de los diferentes derivados inmovilizados de CALfica de los diferentes derivados inmovilizados de CALfica de los diferentes derivados inmovilizados de CAL----B catalizando la B catalizando la B catalizando la B catalizando la hidrólisis de los sustratos a 25ºC.hidrólisis de los sustratos a 25ºC.hidrólisis de los sustratos a 25ºC.hidrólisis de los sustratos a 25ºC. La actividad se definió como: (µmol . min-1.mglipasa-1).El error relativo se estimó en ± 3%
1.1.1-Efecto del pH sobre la actividad de los derivados inmovilizados frente a los
compuestos quirales 1111 y 7777.
En este caso, se analizó el efecto de la disminución del pH (de 7 a 5) sobre la
actividad catalítica de los diferentes derivados inmovilizados de CAL-B (Tabla 11).
Cuando se empleó como sustrato el isómero R del compuesto 1111, no se observaron
grandes diferencias en las actividades de los derivados estudiados. Sin embargo, la
disminución del pH, cuando se utilizó como sustrato el compuesto 7777, provocó algunas
modificaciones en la actividad catalítica de la mayoría de los derivados inmovilizados,
destacando el aumento en la actividad para el derivado octadecil-Sepabeads-CAL-B.
Este incremento mayor en la actividad, en el caso del derivado octadecil-Sepabeads-
CAL-B, podría ser atribuido: i) por un lado, al microambiente extremadamente hidrofóbico
existente alrededor del centro activo de la lipasa cuando se encuentra interfacialmente
adsorbida (el entorno estaría formado por los bolsillos hidrofóbicos de la lipasa y las
superficies hidrofóbicas del soporte),lo cual hace a la enzima aún más selectiva hacia
sustratos más hidrofóbicos; ii) y por otro lado, a la menor ionización del sustrato al disminuir el
pH (debido a la presencia de un grupo carboxílico) haciéndose este más hidrofóbico,
facilitándose la entrada en el centro activo (Esquema 23).
Derivado inmovilizado 5555 ( R )-1111 (±)-7777
pH 7 pH 7 pH 5 pH 7 pH 5
Octadecil-Sepabeads- CAL-B 1141 7 5 6,1 x 10-4 2 x 10-3
PEI -CAL-B 323 60 58 6,6 x 10-4 8,33 x 10-4
Glutaraldehído- CAL-B 250 13 15 0,0186 0,0318
Glioxil -CAL-B 184 7,72 9,91 8,3 x 10-3 4,5 x 10-4
BrCN -CAL-B 182 1,47 nd 4,5 x 10-4 5,8 x 10-4
Eupergit-Cu- CAL-B 14 0,75 nd 1,06 x 10-3 1,66 x 10-3

Ingeniería conformacional de lipasas
73
Esquema Esquema Esquema Esquema 23232323. . . . Efecto del pH en la actividad de la enzima adsorbida sobre soporte octadecil-Sepabeads catalizando la hidrólisis del ácido 2-O-butiril-2-fenilacético
Por tanto, vemos como distintos derivados inmovilizados de la misma lipasa
presentan distinta actividad frente a un mismo sustrato; y, un mismo derivado presenta
diferente actividad frente a distintos sustratos. Además, el efecto del pH sobre la actividad
catalítica también es diferente dependiendo del derivado inmovilizado utilizado.
1.2. Enantioselectividad de los distintos derivados inmovilizados de CAL1.2. Enantioselectividad de los distintos derivados inmovilizados de CAL1.2. Enantioselectividad de los distintos derivados inmovilizados de CAL----B catalizando la B catalizando la B catalizando la B catalizando la
hidrólisis de mandelato de metilo [hidrólisis de mandelato de metilo [hidrólisis de mandelato de metilo [hidrólisis de mandelato de metilo [(±)-1].1].1].1].
La propiedad catalítica más interesante de las lipasas es la enantioselectividad frente a
sustratos quirales. En este caso, analizamos el efecto de la inmovilización sobre la
enantioselectividad de la lipasa de C. antarctica B en la hidrólisis de (±)-1111.
En las condiciones de pH 7 y 25ºC (Tabla 12), todos los derivados inmovilizados
presentaron una preferencia estereoquímica similar, hidrolizando el isómero R. El derivado PEI-
CAL-B fue el más enantioselectivo (E=67), mientras el valor de E más bajo se obtuvo con el
derivado BrCN-CAL-B (E=7,4). Las enantioselectividades encontradas para los demás
derivados inmovilizados fueron muy similares (E=12-20).
O
O
COOH
O
O
COO-
CadenaCadenaCadenaCadena octadeciloctadeciloctadeciloctadecil (hidrof(hidrof(hidrof(hidrofóóóóbica)bica)bica)bica)
ZonaZonaZonaZona hidrofhidrofhidrofhidrofóóóóbicabicabicabica
Mucho Mucho Mucho Mucho mmmmáááásssslentolentolentolento
1111
2222
VVVV2222 >>V>>V>>V>>V1111
O
O
COOH
O
O
COO-
CadenaCadenaCadenaCadena octadeciloctadeciloctadeciloctadecil (hidrof(hidrof(hidrof(hidrofóóóóbica)bica)bica)bica)
ZonaZonaZonaZona hidrofhidrofhidrofhidrofóóóóbicabicabicabica
Mucho Mucho Mucho Mucho mmmmáááásssslentolentolentolento
1111
2222
VVVV2222 >>V>>V>>V>>V1111
O
O
COOH
O
O
COO-
CadenaCadenaCadenaCadena octadeciloctadeciloctadeciloctadecil (hidrof(hidrof(hidrof(hidrofóóóóbica)bica)bica)bica)
ZonaZonaZonaZona hidrofhidrofhidrofhidrofóóóóbicabicabicabica
Mucho Mucho Mucho Mucho mmmmáááásssslentolentolentolento
1111
2222
VVVV2222 >>V>>V>>V>>V1111
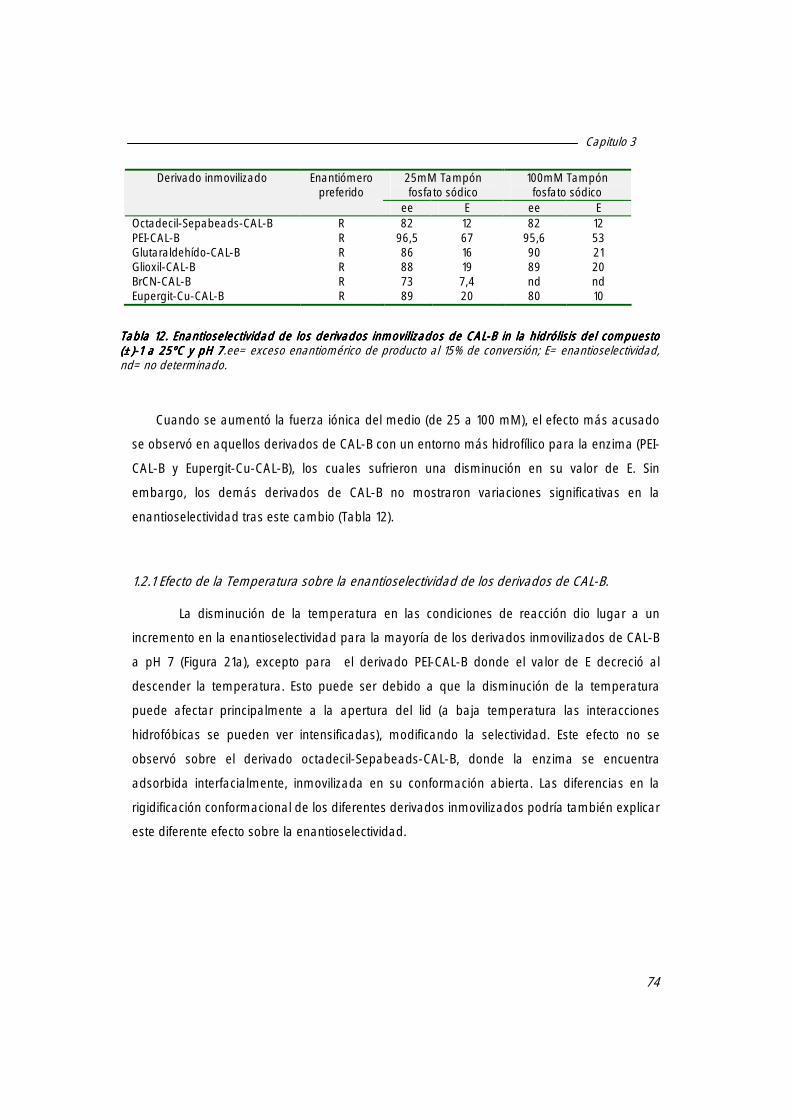
Capitulo 3
74
Derivado inmovilizado Enantiómero preferido
25mM Tampón fosfato sódico
100mM Tampón fosfato sódico
ee E ee E Octadecil-Sepabeads-CAL-B R 82 12 82 12 PEI-CAL-B R 96,5 67 95,6 53 Glutaraldehído-CAL-B R 86 16 90 21 Glioxil-CAL-B R 88 19 89 20 BrCN-CAL-B R 73 7,4 nd nd Eupergit-Cu-CAL-B R 89 20 80 10
Tabla 12.Tabla 12.Tabla 12.Tabla 12. Enantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CAL----B in la hidrólisis del compuestB in la hidrólisis del compuestB in la hidrólisis del compuestB in la hidrólisis del compuesto o o o (±)(±)(±)(±)----1 1 1 1 a 25ºC y pHa 25ºC y pHa 25ºC y pHa 25ºC y pH 7777.ee= exceso enantiomérico de producto al 15% de conversión; E= enantioselectividad, nd= no determinado.
Cuando se aumentó la fuerza iónica del medio (de 25 a 100 mM), el efecto más acusado
se observó en aquellos derivados de CAL-B con un entorno más hidrofílico para la enzima (PEI-
CAL-B y Eupergit-Cu-CAL-B), los cuales sufrieron una disminución en su valor de E. Sin
embargo, los demás derivados de CAL-B no mostraron variaciones significativas en la
enantioselectividad tras este cambio (Tabla 12).
1.2.1 Efecto de la Temperatura sobre la enantioselectividad de los derivados de CAL-B.
La disminución de la temperatura en las condiciones de reacción dio lugar a un
incremento en la enantioselectividad para la mayoría de los derivados inmovilizados de CAL-B
a pH 7 (Figura 21a), excepto para el derivado PEI-CAL-B donde el valor de E decreció al
descender la temperatura. Esto puede ser debido a que la disminución de la temperatura
puede afectar principalmente a la apertura del lid (a baja temperatura las interacciones
hidrofóbicas se pueden ver intensificadas), modificando la selectividad. Este efecto no se
observó sobre el derivado octadecil-Sepabeads-CAL-B, donde la enzima se encuentra
adsorbida interfacialmente, inmovilizada en su conformación abierta. Las diferencias en la
rigidificación conformacional de los diferentes derivados inmovilizados podría también explicar
este diferente efecto sobre la enantioselectividad.

Ingeniería conformacional de lipasas
75
FiguraFiguraFiguraFigura 22221111. Efecto de la temperatura sobre la enantioselectividEfecto de la temperatura sobre la enantioselectividEfecto de la temperatura sobre la enantioselectividEfecto de la temperatura sobre la enantioselectividad de los diferentes derivados de ad de los diferentes derivados de ad de los diferentes derivados de ad de los diferentes derivados de CALCALCALCAL----B en la hidrólisis del sustrato (±)B en la hidrólisis del sustrato (±)B en la hidrólisis del sustrato (±)B en la hidrólisis del sustrato (±)----1111; (a)(a)(a)(a) Enantioselectividad (E) a pH 7; (b)(b)(b)(b) Enantioselectividad (E) a pH 5. Glutaraldehído-CAL-B (` ),glioxil-CAL-B (�),octadecil-CAL-B (�),PEI-CAL-B (�). Los experimentos se realizaron a 25 mM de tampón fosfato sódico y 10 mM de sustrato a pH 7.
Cuando se alteró el pH del medio, de pH 7 a pH 5 (Figura 21b), la tendencia en los
valores de enantioselectividad para los diferentes derivados inmovilizados fue la misma que a
pH 7. Sin embargo, en el caso del derivado PEI-CAL-B, la disminución de enantioselectividad a
4ºC fue más acusada a este pH.
1.2.2 Efecto de la presencia de co-disolvente en el medio de reacción sobre el valor de E de los
distintos derivados de CAL-B.
La presencia de cierta cantidad de co-disolvente en el medio de reacción fue otro
factor que influyó sobre la enantioselectividad de los diferentes derivados.
La Figura 22 muestra las enantioselectividades de algunos derivados inmovilizados de
CAL-B frente a distintas concentraciones de 1,4-dioxano como co-disolvente (pH 7 y 25ºC).
Podemos ver como el derivado octadecil-Sepabeads-CAL-B no sufrió ningún cambio en el
valor de enantioselectividad a los distintos porcentajes de dioxano, mientras otros derivados
experimentaron decrecimientos bruscos, o drásticos aumentos, en el valor de E a una
determinada concentración de co-disolvente, como sucedió para los derivados PEI-CAL-B (E de
67 a 25 con 20% de dioxano) o glutaraldehído-CAL-B (E de 19 a 60 con 15% de dioxano).
aaaa
000010101010202020203030303040404040505050506060606070707070
0000 20202020 40404040 60606060
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
bbbb
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020 30303030
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
aaaa
000010101010202020203030303040404040505050506060606070707070
0000 20202020 40404040 60606060
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
bbbb
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020 30303030
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
aaaa
000010101010202020203030303040404040505050506060606070707070
0000 20202020 40404040 60606060
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
aaaa
000010101010202020203030303040404040505050506060606070707070
0000 20202020 40404040 60606060
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
000010101010202020203030303040404040505050506060606070707070
0000 20202020 40404040 60606060
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
bbbb
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020 30303030
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
bbbb
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020 30303030
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020 30303030
Temperatura(Temperatura(Temperatura(Temperatura(ººººCCCC))))
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
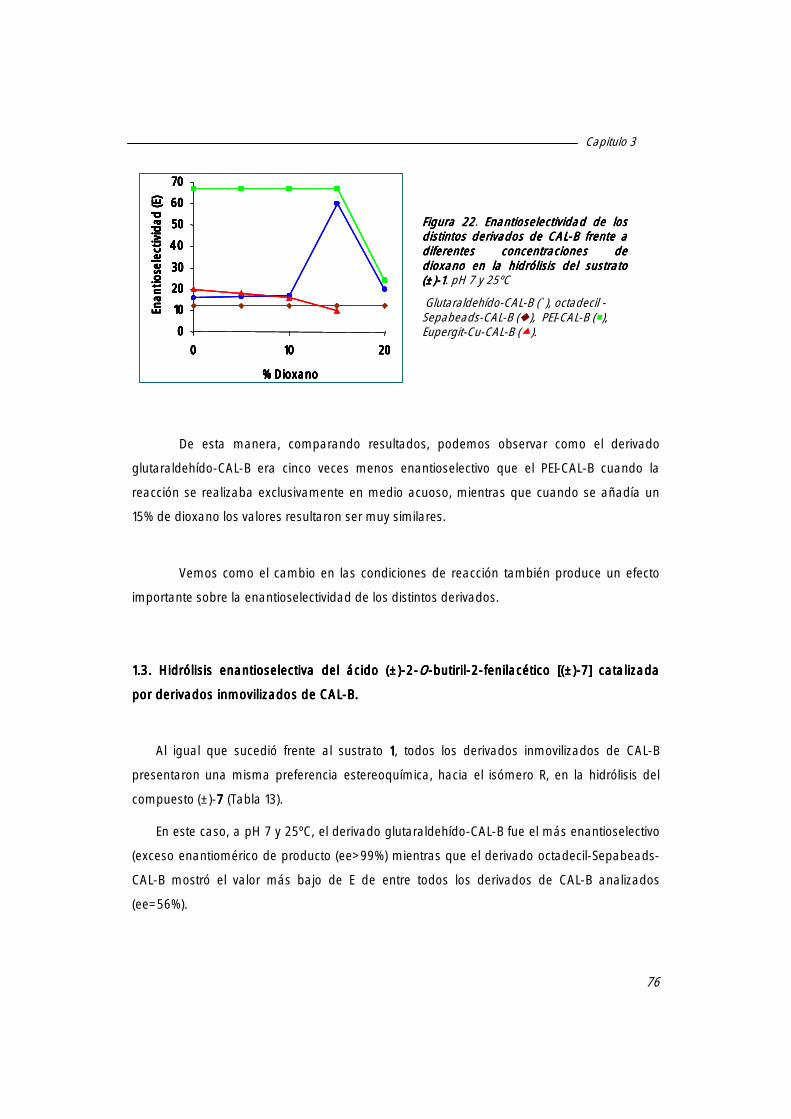
Capitulo 3
76
Figura 2Figura 2Figura 2Figura 22222. Enantioselectividad de los Enantioselectividad de los Enantioselectividad de los Enantioselectividad de los distintos derivados de CALdistintos derivados de CALdistintos derivados de CALdistintos derivados de CAL----B frente a B frente a B frente a B frente a diferentes concentraciones de diferentes concentraciones de diferentes concentraciones de diferentes concentraciones de dioxano en la hidrólisis del sustrato dioxano en la hidrólisis del sustrato dioxano en la hidrólisis del sustrato dioxano en la hidrólisis del sustrato (±)(±)(±)(±)----1111. pH 7 y 25ºC
Glutaraldehído-CAL-B (` ), octadecil -Sepabeads-CAL-B (�), PEI-CAL-B (�), Eupergit-Cu-CAL-B (�).
De esta manera, comparando resultados, podemos observar como el derivado
glutaraldehído-CAL-B era cinco veces menos enantioselectivo que el PEI-CAL-B cuando la
reacción se realizaba exclusivamente en medio acuoso, mientras que cuando se añadía un
15% de dioxano los valores resultaron ser muy similares.
Vemos como el cambio en las condiciones de reacción también produce un efecto
importante sobre la enantioselectividad de los distintos derivados.
1.3. Hidrólisis enantioselectiva del ácido (±)1.3. Hidrólisis enantioselectiva del ácido (±)1.3. Hidrólisis enantioselectiva del ácido (±)1.3. Hidrólisis enantioselectiva del ácido (±)----2222----OOOO----butirilbutirilbutirilbutiril----2222----fenilacfenilacfenilacfenilacéééético tico tico tico [[[[(±)(±)(±)(±)----7] catalizada 7] catalizada 7] catalizada 7] catalizada
por derivados inmovilizados de CALpor derivados inmovilizados de CALpor derivados inmovilizados de CALpor derivados inmovilizados de CAL----B.B.B.B.
Al igual que sucedió frente al sustrato 1111, todos los derivados inmovilizados de CAL-B
presentaron una misma preferencia estereoquímica, hacia el isómero R, en la hidrólisis del
compuesto (±)-7 7 7 7 (Tabla 13).
En este caso, a pH 7 y 25ºC, el derivado glutaraldehído-CAL-B fue el más enantioselectivo
(exceso enantiomérico de producto (ee>99%) mientras que el derivado octadecil-Sepabeads-
CAL-B mostró el valor más bajo de E de entre todos los derivados de CAL-B analizados
(ee=56%).
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020
% % % % DioxanoDioxanoDioxanoDioxano
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020
% % % % DioxanoDioxanoDioxanoDioxano
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
0000
10101010
20202020
30303030
40404040
50505050
60606060
70707070
0000 10101010 20202020
% % % % DioxanoDioxanoDioxanoDioxano
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
Enan
tiose
lect
ivid
ad (E
)En
antio
sele
ctiv
idad
(E)
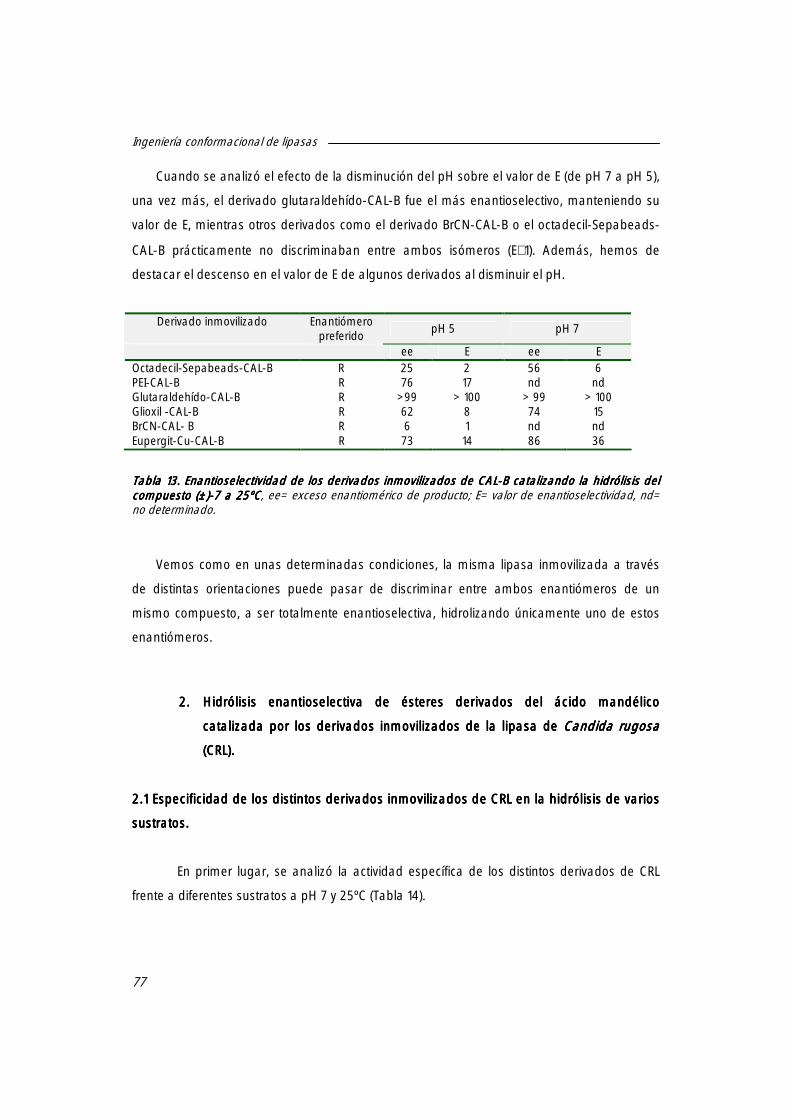
Ingeniería conformacional de lipasas
77
Cuando se analizó el efecto de la disminución del pH sobre el valor de E (de pH 7 a pH 5),
una vez más, el derivado glutaraldehído-CAL-B fue el más enantioselectivo, manteniendo su
valor de E, mientras otros derivados como el derivado BrCN-CAL-B o el octadecil-Sepabeads-
CAL-B prácticamente no discriminaban entre ambos isómeros (E≅ 1). Además, hemos de
destacar el descenso en el valor de E de algunos derivados al disminuir el pH.
Tabla 13.Tabla 13.Tabla 13.Tabla 13. Enantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CALEnantioselectividad de los derivados inmovilizados de CAL----B catalizando la hidrólisis del B catalizando la hidrólisis del B catalizando la hidrólisis del B catalizando la hidrólisis del compuesto (±)compuesto (±)compuesto (±)compuesto (±)----7 a 25ºC7 a 25ºC7 a 25ºC7 a 25ºC, ee= exceso enantiomérico de producto; E= valor de enantioselectividad, nd= no determinado.
Vemos como en unas determinadas condiciones, la misma lipasa inmovilizada a través
de distintas orientaciones puede pasar de discriminar entre ambos enantiómeros de un
mismo compuesto, a ser totalmente enantioselectiva, hidrolizando únicamente uno de estos
enantiómeros.
2.2.2.2. Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de Hidrólisis enantioselectiva de ésteresésteresésteresésteres derivados del ácido mandélico derivados del ácido mandélico derivados del ácido mandélico derivados del ácido mandélico
catalizada por catalizada por catalizada por catalizada por los derivados inmovilizados de la lipasa de los derivados inmovilizados de la lipasa de los derivados inmovilizados de la lipasa de los derivados inmovilizados de la lipasa de Candida rugosaCandida rugosaCandida rugosaCandida rugosa
(CRL)(CRL)(CRL)(CRL)....
2.1 Especificidad de los distintos d2.1 Especificidad de los distintos d2.1 Especificidad de los distintos d2.1 Especificidad de los distintos derivados inmovilizados de CRL en la hidrólisis de varios erivados inmovilizados de CRL en la hidrólisis de varios erivados inmovilizados de CRL en la hidrólisis de varios erivados inmovilizados de CRL en la hidrólisis de varios
sustratos.sustratos.sustratos.sustratos.
En primer lugar, se analizó la actividad específica de los distintos derivados de CRL
frente a diferentes sustratos a pH 7 y 25ºC (Tabla 14).
Derivado inmovilizado Enantiómero preferido
pH 5 pH 7
ee E ee E Octadecil-Sepabeads-CAL-B R 25 2 56 6 PEI-CAL-B R 76 17 nd nd Glutaraldehído-CAL-B R >99 > 100 > 99 > 100 Glioxil -CAL-B R 62 8 74 15 BrCN-CAL- B R 6 1 nd nd Eupergit-Cu-CAL-B R 73 14 86 36
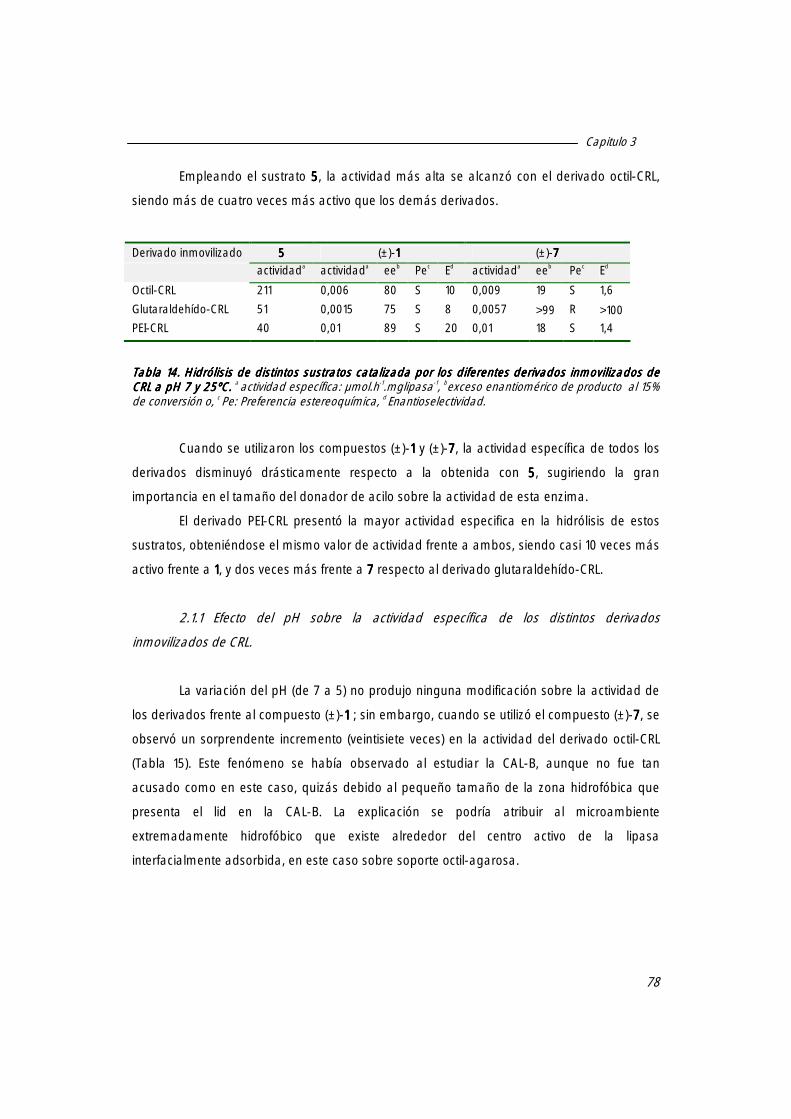
Capitulo 3
78
Empleando el sustrato 5555, la actividad más alta se alcanzó con el derivado octil-CRL,
siendo más de cuatro veces más activo que los demás derivados.
Derivado inmovilizado 5555 (±)-1111 (±)-7777 actividada actividada eeb Pec Ed actividada eeb Pec Ed
Octil-CRL 211 0,006 80 S 10 0,009 19 S 1,6
Glutaraldehído-CRL 51 0,0015 75 S 8 0,0057 >99 R >100 PEI-CRL 40 0,01 89 S 20 0,01 18 S 1,4
TablaTablaTablaTabla 14. 14. 14. 14. Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de CRL a pH 7 y 25ºC.CRL a pH 7 y 25ºC.CRL a pH 7 y 25ºC.CRL a pH 7 y 25ºC. a actividad específica: µmol.h-1.mglipasa-1, bexceso enantiomérico de producto al 15% de conversión o, c Pe: Preferencia estereoquímica, d Enantioselectividad.
Cuando se utilizaron los compuestos (±)-1111 y (±)-7777, la actividad específica de todos los
derivados disminuyó drásticamente respecto a la obtenida con 5555, sugiriendo la gran
importancia en el tamaño del donador de acilo sobre la actividad de esta enzima.
El derivado PEI-CRL presentó la mayor actividad especifica en la hidrólisis de estos
sustratos, obteniéndose el mismo valor de actividad frente a ambos, siendo casi 10 veces más
activo frente a 1111, y dos veces más frente a 7777 respecto al derivado glutaraldehído-CRL.
2.1.1 Efecto del pH sobre la actividad específica de los distintos derivados
inmovilizados de CRL.
La variación del pH (de 7 a 5) no produjo ninguna modificación sobre la actividad de
los derivados frente al compuesto (±)-1111 ; sin embargo, cuando se utilizó el compuesto (±)-7777, se
observó un sorprendente incremento (veintisiete veces) en la actividad del derivado octil-CRL
(Tabla 15). Este fenómeno se había observado al estudiar la CAL-B, aunque no fue tan
acusado como en este caso, quizás debido al pequeño tamaño de la zona hidrofóbica que
presenta el lid en la CAL-B. La explicación se podría atribuir al microambiente
extremadamente hidrofóbico que existe alrededor del centro activo de la lipasa
interfacialmente adsorbida, en este caso sobre soporte octil-agarosa.

Ingeniería conformacional de lipasas
79
Tabla 15. Tabla 15. Tabla 15. Tabla 15. Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de Hidrólisis de distintos sustratos catalizada por los diferentes derivados inmovilizados de CRL a pH 5 y 25ºC.CRL a pH 5 y 25ºC.CRL a pH 5 y 25ºC.CRL a pH 5 y 25ºC. a actividad específica: µmol.h-1.mglipasa-1 b exceso enantiomérico, cPreferencia estereoquímica, d Enantioselectividad.
2.2 Enantioselectividad de los derivados inmovilizados de CRL en la hidrólisis de 2.2 Enantioselectividad de los derivados inmovilizados de CRL en la hidrólisis de 2.2 Enantioselectividad de los derivados inmovilizados de CRL en la hidrólisis de 2.2 Enantioselectividad de los derivados inmovilizados de CRL en la hidrólisis de (±)(±)(±)(±)----
ésteresésteresésteresésteres derivados del ácido mandélico. derivados del ácido mandélico. derivados del ácido mandélico. derivados del ácido mandélico.
El estudio de enantioselectividad para esta lipasa en la reacción de hidrólisis del
compuesto (±)-1111 a pH 7 mostró una misma especificidad para todos los derivados de CRL,
hacia la hidrólisis del isómero S (Tabla 14).
El derivado PEI-CRL mostró el valor de enantioselectividad más alto (E=20) mientras los
demás derivados inmovilizados presentaron resultados similares (E=8-10).
Cuando se utilizó el compuesto (±)-7777, el derivado glutaraldehído-CRL fue el más
enantioselectivo (E>100) hidrolizando el isómero R (inversión en la preferencia enantiomérica),
mientras los demás derivados mostraron muy bajos valores de E (E=1,4), hidrolizando
preferentemente el isómero S (Tabla 14).
2.2.1 Influencia del pH sobre la enantioselectividad de los derivados de CRL
La alteración en los valores de enantioselectividad de los distintos derivados de CRL
tras la disminución del pH (de 7 a 5) quedó reflejada en la hidrólisis de ambos sustratos (Tabla
15).
De esta manera, en la hidrólisis de (±)-1111, el aumento más significativo, tras el cambio
en el pH, se produjo sobre el derivado PEI-CRL, el cual presentó un valor de E superior a 100.
Cuando se empleó el compuesto (±)-7777, se observó un sorprendente incremento en la
enantioselectividad del derivado octil-CRL, pasando de una E=1,6 a pH 7 (casi no existe
Derivado inmovilizado (±)-1111 (±)-7777 actividada eeb Pec Ed actividada eeb Pec Ed
Octil-CRL 0,005 94 S 40 0,243 96 S 85
Glutaraldehído-CRL 0,0017 80 S 10 0,0081 >99 R >100 PEI-CRL 0,01 >99 S >100 0,0098 12 S 1,2

Capitulo 3
80
discriminación entre enantiómeros por parte de la enzima) hasta una E=85 (reconoce casi
únicamente el isómero S) a pH 5, mientras los demás derivados conservaron su valor de E.
Por tanto, dependiendo del derivado y de las condiciones empleadas, es posible
obtener enantioméricamente puros: ambos isómeros del compuesto 2222 y ambos isómeros del
compuesto 7777, a partir del compuesto (±)-7777.
3.3.3.3. Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido (±)(±)(±)(±)----2222----OOOO----butirilbutirilbutirilbutiril----2222----fenilacetico fenilacetico fenilacetico fenilacetico [[[[(±)(±)(±)(±)----7] 7] 7] 7]
catalizada catalizada catalizada catalizada por derivados inmovilizados de la lipasa de por derivados inmovilizados de la lipasa de por derivados inmovilizados de la lipasa de por derivados inmovilizados de la lipasa de Mucor mieheiMucor mieheiMucor mieheiMucor miehei (MML).(MML).(MML).(MML).
3.1 Activid3.1 Activid3.1 Activid3.1 Actividad de los diferentes derivados inmovilizados de MML en la hidrólisis de ad de los diferentes derivados inmovilizados de MML en la hidrólisis de ad de los diferentes derivados inmovilizados de MML en la hidrólisis de ad de los diferentes derivados inmovilizados de MML en la hidrólisis de (±)(±)(±)(±)----7: 7: 7: 7:
Efecto de las condiciones experimentales.Efecto de las condiciones experimentales.Efecto de las condiciones experimentales.Efecto de las condiciones experimentales.
Hasta ahora, se ha visto la influencia de las condiciones experimentales sobre la
actividad específica de distintos derivados de dos lipasas (CAL-B y CRL). En este caso, se
estudió este efecto sobre un nuevo tipo de derivados inmovilizados utilizando la lipasa de
Mucor miehei, donde la enzima se encuentra inmovilizada sobre soportes Sepabeads-
heterofuncionales (Tabla 16).
En este caso, el derivado IDA-Sepabeads-MML mostró el valor más alto de actividad
en la hidrólisis de (±)-7 7 7 7 a pH 7 y 25ºC, mientras el derivado IDA-Cu2+-Sepabeads-MML fue el
menos activo (27 veces menos actividad que el derivado IDA-Sepabeads-MML). Las
actividades obtenidas para los demás derivados fueron entre 7 y 14 veces más bajas respecto
a la actividad del derivado IDA-Sepabeads-MML, para los derivados EDA-Sepabeads-MML y
Octadecil-Sepabeads-MML, respectivamente. Esto demuestra, una vez más, el efecto de la
metodología de inmovilización sobre la actividad de la lipasa.
3.1.1 Efecto de la temperatura sobre la actividad específica de los distintos derivados
inmovilizados de MML.
La disminución en la temperatura (de 25ºC a 4ºC) provocó un decrecimiento en la
actividad para todos los derivados de MML. Este descenso de actividad fue más acusado en

Ingeniería conformacional de lipasas
81
los derivados IDA-Sepabeads-MML y IDA-Cu2+-Sepabeads-MML, disminuyéndose su valor 8 y
13 veces, respectivamente; sin embargo, para el derivado octadecil-Sepabeads-MML apenas
se observó variación en la actividad.
Este efecto sobre la actividad catalítica podría ser explicado si pensamos como el
descenso de la temperatura puede afectar principalmente a la apertura del lid (a baja
temperatura las interacciones hidrofóbicas se pueden ver intensificadas), además teniendo en
cuenta la distinta rigidificación conformacional que existiría en cada derivado inmovilizado.
Derivado inmovilizado AE 25ºC pH 7 AE 25ºC pH 7 AE 25ºC pH 7 Octadecil-Sepabeads-MML 0,04 0,027 0,07
IDA- Cu +2- Sepabeads-MML 0,02 0,0015 0,0057
IDA-Sepabeads-MML 0,55 0,067 0,0326
EDA-Sepabeads-MML 0,096 0,02 0,076
Tabla 16. Tabla 16. Tabla 16. Tabla 16. Actividad específica (AE) de los distintos derivados de MML catalizando la hidrólisis de Actividad específica (AE) de los distintos derivados de MML catalizando la hidrólisis de Actividad específica (AE) de los distintos derivados de MML catalizando la hidrólisis de Actividad específica (AE) de los distintos derivados de MML catalizando la hidrólisis de (±)(±)(±)(±)----7777. . . . La actividad se definió como µmol min-1mglipasa-1. El error relativo se estimó en ±4%
3.1.2. Influencia del pH sobre la actividad específica de los derivados de MML.
Por otro lado, cuando se disminuyó el pH de 7 a 5, el derivado octadecil-Sepabeads-
MML experimentó un incremento en la actividad específica de casi dos veces mientras los
demás derivados sufrían un descenso en la actividad, destacando la disminución tan acusada
del derivado IDA-Sepabeads-MML (17 veces menos activo).
Este efecto en la actividad del derivado octadecil-Sepabeads-MML, como había sido
observado para los correspondientes derivados de CAL-B y CRL, podía estar relacionado a la
mayor hidrofobicidad del sustrato a este pH.
3.23.23.23.2 Enantioselectividad de los diferentes derivados inmovilizados de MMLEnantioselectividad de los diferentes derivados inmovilizados de MMLEnantioselectividad de los diferentes derivados inmovilizados de MMLEnantioselectividad de los diferentes derivados inmovilizados de MML en la hidrólisis en la hidrólisis en la hidrólisis en la hidrólisis
de de de de (±)-7. 7. 7. 7.
En primer lugar, se analizó el efecto causado por las distintas metodologías de
inmovilización en el valor de enantioselectividad para los derivados de MML a pH 7 y 25ºC
(Tabla 17).
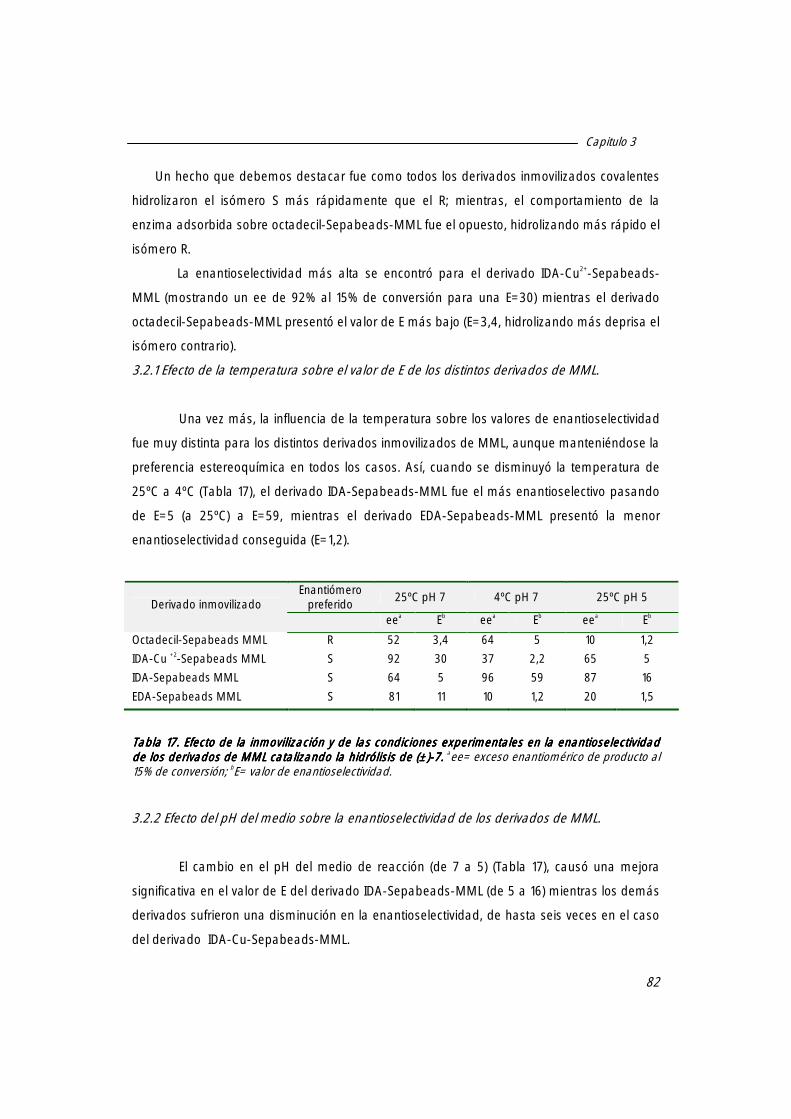
Capitulo 3
82
Un hecho que debemos destacar fue como todos los derivados inmovilizados covalentes
hidrolizaron el isómero S más rápidamente que el R; mientras, el comportamiento de la
enzima adsorbida sobre octadecil-Sepabeads-MML fue el opuesto, hidrolizando más rápido el
isómero R.
La enantioselectividad más alta se encontró para el derivado IDA-Cu2+-Sepabeads-
MML (mostrando un ee de 92% al 15% de conversión para una E=30) mientras el derivado
octadecil-Sepabeads-MML presentó el valor de E más bajo (E=3,4, hidrolizando más deprisa el
isómero contrario).
3.2.1 Efecto de la temperatura sobre el valor de E de los distintos derivados de MML.
Una vez más, la influencia de la temperatura sobre los valores de enantioselectividad
fue muy distinta para los distintos derivados inmovilizados de MML, aunque manteniéndose la
preferencia estereoquímica en todos los casos. Así, cuando se disminuyó la temperatura de
25ºC a 4ºC (Tabla 17), el derivado IDA-Sepabeads-MML fue el más enantioselectivo pasando
de E=5 (a 25ºC) a E=59, mientras el derivado EDA-Sepabeads-MML presentó la menor
enantioselectividad conseguida (E=1,2).
Derivado inmovilizado Enantiómero
preferido 25ºC pH 7 4ºC pH 7 25ºC pH 5
eea Eb eea Eb eea Eb
Octadecil-Sepabeads MML R 52 3,4 64 5 10 1,2
IDA-Cu +2-Sepabeads MML S 92 30 37 2,2 65 5
IDA-Sepabeads MML S 64 5 96 59 87 16
EDA-Sepabeads MML S 81 11 10 1,2 20 1,5
Tabla 17. Tabla 17. Tabla 17. Tabla 17. Efecto de la inmovilización y de las condiciones experimentales en la enantioselectividad Efecto de la inmovilización y de las condiciones experimentales en la enantioselectividad Efecto de la inmovilización y de las condiciones experimentales en la enantioselectividad Efecto de la inmovilización y de las condiciones experimentales en la enantioselectividad de los derivados de MML catalizando la hidrólisis de de los derivados de MML catalizando la hidrólisis de de los derivados de MML catalizando la hidrólisis de de los derivados de MML catalizando la hidrólisis de (±)(±)(±)(±)----7777.... aee= exceso enantiomérico de producto al 15% de conversión; bE= valor de enantioselectividad.
3.2.2 Efecto del pH del medio sobre la enantioselectividad de los derivados de MML.
El cambio en el pH del medio de reacción (de 7 a 5) (Tabla 17), causó una mejora
significativa en el valor de E del derivado IDA-Sepabeads-MML (de 5 a 16) mientras los demás
derivados sufrieron una disminución en la enantioselectividad, de hasta seis veces en el caso
del derivado IDA-Cu-Sepabeads-MML.
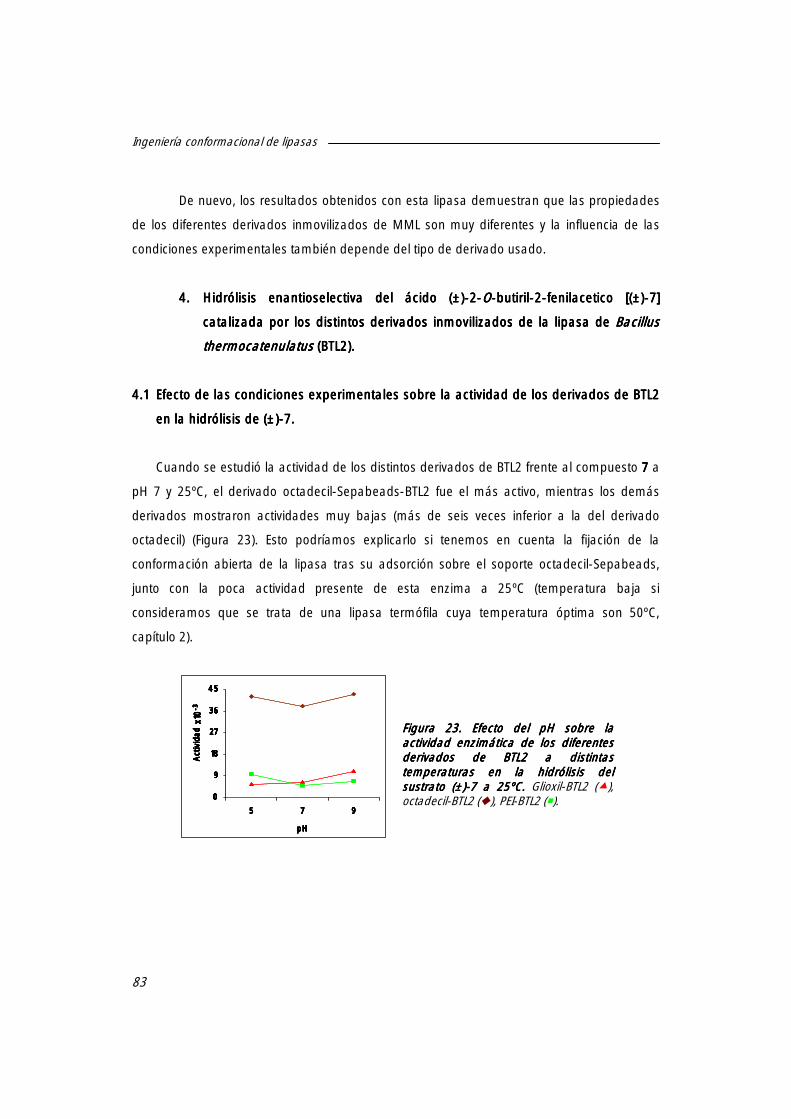
Ingeniería conformacional de lipasas
83
De nuevo, los resultados obtenidos con esta lipasa demuestran que las propiedades
de los diferentes derivados inmovilizados de MML son muy diferentes y la influencia de las
condiciones experimentales también depende del tipo de derivado usado.
4.4.4.4. Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido (±)(±)(±)(±)----2222----OOOO----butirilbutirilbutirilbutiril----2222----fenilacetico fenilacetico fenilacetico fenilacetico [[[[(±)(±)(±)(±)----7] 7] 7] 7]
catalizada por catalizada por catalizada por catalizada por los distintos los distintos los distintos los distintos derivados inmovilizados de derivados inmovilizados de derivados inmovilizados de derivados inmovilizados de la lipasa de la lipasa de la lipasa de la lipasa de Bacillus Bacillus Bacillus Bacillus
thermocatenulatusthermocatenulatusthermocatenulatusthermocatenulatus ( ( ( (BTL2BTL2BTL2BTL2))))....
4.14.14.14.1 Efecto de las condiciones experimentales sobre la actividad de los derivados de BTL2 Efecto de las condiciones experimentales sobre la actividad de los derivados de BTL2 Efecto de las condiciones experimentales sobre la actividad de los derivados de BTL2 Efecto de las condiciones experimentales sobre la actividad de los derivados de BTL2
en la hidrólisis de en la hidrólisis de en la hidrólisis de en la hidrólisis de (±)(±)(±)(±)----7.7.7.7.
Cuando se estudió la actividad de los distintos derivados de BTL2 frente al compuesto 7777 a
pH 7 y 25ºC, el derivado octadecil-Sepabeads-BTL2 fue el más activo, mientras los demás
derivados mostraron actividades muy bajas (más de seis veces inferior a la del derivado
octadecil) (Figura 23). Esto podríamos explicarlo si tenemos en cuenta la fijación de la
conformación abierta de la lipasa tras su adsorción sobre el soporte octadecil-Sepabeads,
junto con la poca actividad presente de esta enzima a 25ºC (temperatura baja si
consideramos que se trata de una lipasa termófila cuya temperatura óptima son 50ºC,
capítulo 2).
FiguraFiguraFiguraFigura 22223333. . . . Efecto del pH sobre la Efecto del pH sobre la Efecto del pH sobre la Efecto del pH sobre la actividad enzimática de los actividad enzimática de los actividad enzimática de los actividad enzimática de los diferentes diferentes diferentes diferentes derivados de BTL2 a distintas derivados de BTL2 a distintas derivados de BTL2 a distintas derivados de BTL2 a distintas temperaturas en la hidrólisis del temperaturas en la hidrólisis del temperaturas en la hidrólisis del temperaturas en la hidrólisis del sustrato (±)sustrato (±)sustrato (±)sustrato (±)----7777 a 25ºC. a 25ºC. a 25ºC. a 25ºC. Glioxil-BTL2 (�), octadecil-BTL2 (�), PEI-BTL2 (�).
x10
x10
x10
x10
-- -- 33 33
0000
9999
18181818
27272727
36363636
4 54 54 54 5
5555 7777 9999
pHpHpHpH
Act
ivid
adA
ctiv
idad
Act
ivid
adA
ctiv
idad
x10
x10
x10
x10
-- -- 33 33
0000
9999
18181818
27272727
36363636
4 54 54 54 5
5555 7777 9999
pHpHpHpH
Act
ivid
adA
ctiv
idad
Act
ivid
adA
ctiv
idad
x10
x10
x10
x10
-- -- 33 33
0000
9999
18181818
27272727
36363636
4 54 54 54 5
5555 7777 9999
pHpHpHpH
Act
ivid
adA
ctiv
idad
Act
ivid
adA
ctiv
idad
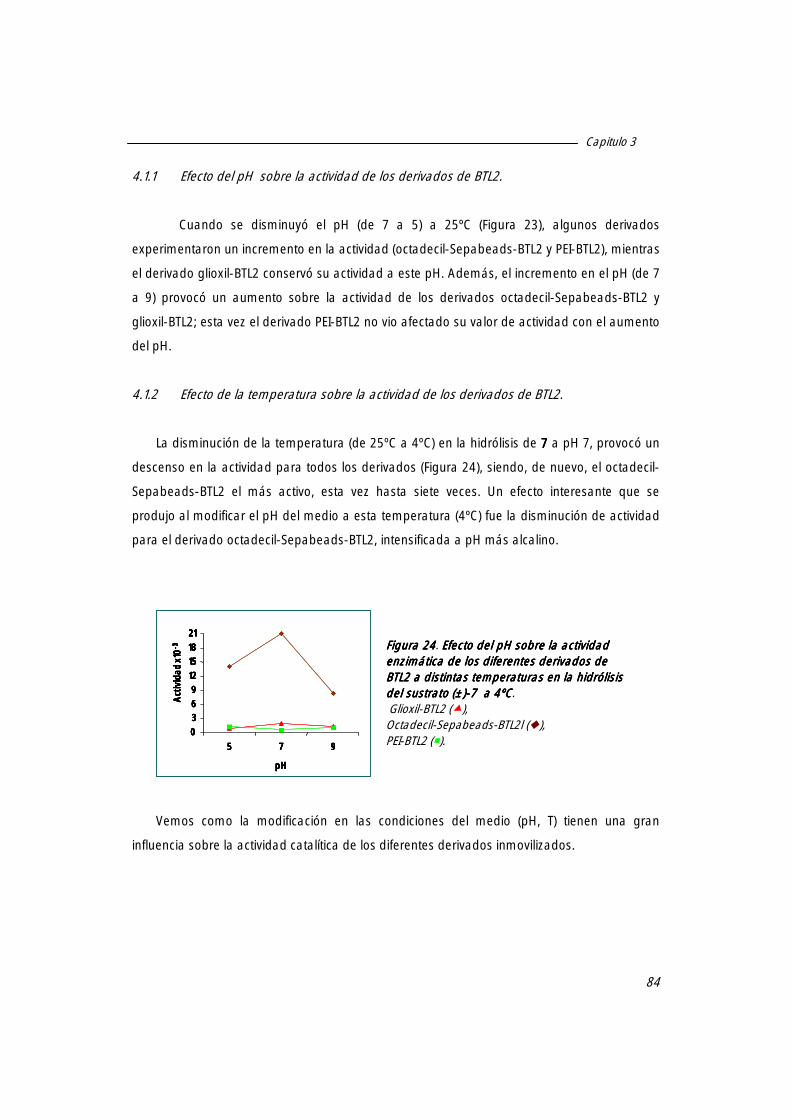
Capitulo 3
84
4.1.1 Efecto del pH sobre la actividad de los derivados de BTL2.
Cuando se disminuyó el pH (de 7 a 5) a 25ºC (Figura 23), algunos derivados
experimentaron un incremento en la actividad (octadecil-Sepabeads-BTL2 y PEI-BTL2), mientras
el derivado glioxil-BTL2 conservó su actividad a este pH. Además, el incremento en el pH (de 7
a 9) provocó un aumento sobre la actividad de los derivados octadecil-Sepabeads-BTL2 y
glioxil-BTL2; esta vez el derivado PEI-BTL2 no vio afectado su valor de actividad con el aumento
del pH.
4.1.2 Efecto de la temperatura sobre la actividad de los derivados de BTL2.
La disminución de la temperatura (de 25ºC a 4ºC) en la hidrólisis de 7777 a pH 7, provocó un
descenso en la actividad para todos los derivados (Figura 24), siendo, de nuevo, el octadecil-
Sepabeads-BTL2 el más activo, esta vez hasta siete veces. Un efecto interesante que se
produjo al modificar el pH del medio a esta temperatura (4ºC) fue la disminución de actividad
para el derivado octadecil-Sepabeads-BTL2, intensificada a pH más alcalino.
Vemos como la modificación en las condiciones del medio (pH, T) tienen una gran
influencia sobre la actividad catalítica de los diferentes derivados inmovilizados.
Figura 2Figura 2Figura 2Figura 24444. Efecto del pH sobre la actividad Efecto del pH sobre la actividad Efecto del pH sobre la actividad Efecto del pH sobre la actividad enzimática de los diferentes derivados de enzimática de los diferentes derivados de enzimática de los diferentes derivados de enzimática de los diferentes derivados de BTL2 a distintas temperaturas en la hidrólisis BTL2 a distintas temperaturas en la hidrólisis BTL2 a distintas temperaturas en la hidrólisis BTL2 a distintas temperaturas en la hidrólisis del sustrato (±)del sustrato (±)del sustrato (±)del sustrato (±)----7777 a 4ºa 4ºa 4ºa 4ºCCCC. Glioxil-BTL2 (�), Octadecil-Sepabeads-BTL2l (�), PEI-BTL2 (�).
x10
x10
x10
x10
-- -- 33 33
0000333366669999
12121212151515151818181821212121
5555 7777 9999
pHpHpHpH
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad x
10x10
x10
x10
-- -- 33 33
0000333366669999
12121212151515151818181821212121
5555 7777 9999
pHpHpHpH
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad x
10x10
x10
x10
-- -- 33 33
0000333366669999
12121212151515151818181821212121
5555 7777 9999
pHpHpHpH
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad
Act
ivid
ad
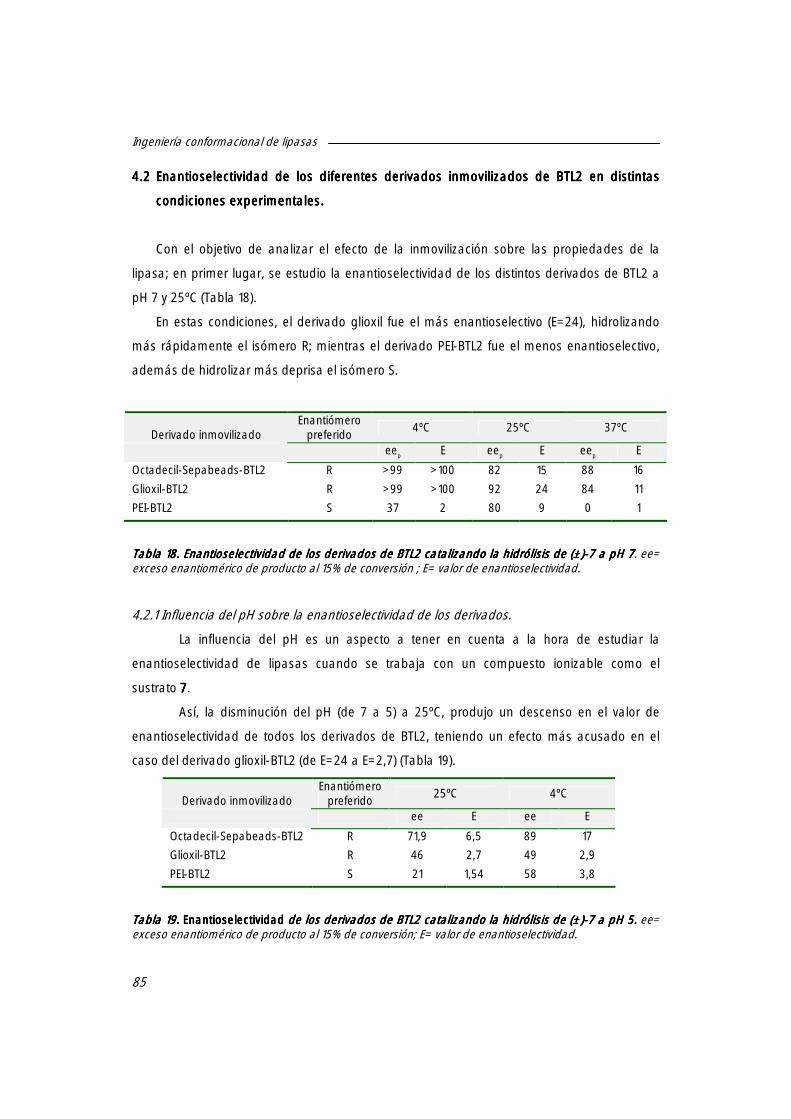
Ingeniería conformacional de lipasas
85
4.24.24.24.2 Enantioselectividad de los diferentes derivados inmovilizados de BTL2 en distintas Enantioselectividad de los diferentes derivados inmovilizados de BTL2 en distintas Enantioselectividad de los diferentes derivados inmovilizados de BTL2 en distintas Enantioselectividad de los diferentes derivados inmovilizados de BTL2 en distintas
condiciones experimentales.condiciones experimentales.condiciones experimentales.condiciones experimentales.
Con el objetivo de analizar el efecto de la inmovilización sobre las propiedades de la
lipasa; en primer lugar, se estudio la enantioselectividad de los distintos derivados de BTL2 a
pH 7 y 25ºC (Tabla 18).
En estas condiciones, el derivado glioxil fue el más enantioselectivo (E=24), hidrolizando
más rápidamente el isómero R; mientras el derivado PEI-BTL2 fue el menos enantioselectivo,
además de hidrolizar más deprisa el isómero S.
Derivado inmovilizado Enantiómero
preferido 4ºC 25ºC 37ºC
eep E eep E eep E
Octadecil-Sepabeads-BTL2 R >99 >100 82 15 88 16
Glioxil-BTL2 R >99 >100 92 24 84 11
PEI-BTL2 S 37 2 80 9 0 1
Tabla 18. EnantioselectividadTabla 18. EnantioselectividadTabla 18. EnantioselectividadTabla 18. Enantioselectividad de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de (±)(±)(±)(±)----7 a pH 77 a pH 77 a pH 77 a pH 7. ee= exceso enantiomérico de producto al 15% de conversión ; E= valor de enantioselectividad.
4.2.1 Influencia del pH sobre la enantioselectividad de los derivados.
La influencia del pH es un aspecto a tener en cuenta a la hora de estudiar la
enantioselectividad de lipasas cuando se trabaja con un compuesto ionizable como el
sustrato 7777.
Así, la disminución del pH (de 7 a 5) a 25ºC, produjo un descenso en el valor de
enantioselectividad de todos los derivados de BTL2, teniendo un efecto más acusado en el
caso del derivado glioxil-BTL2 (de E=24 a E=2,7) (Tabla 19).
Derivado inmovilizado Enantiómero
preferido 25ºC 4ºC
ee E ee E
Octadecil-Sepabeads-BTL2 R 71,9 6,5 89 17
Glioxil-BTL2 R 46 2,7 49 2,9
PEI-BTL2 S 21 1,54 58 3,8
Tabla 19. Tabla 19. Tabla 19. Tabla 19. EnantioselectividadEnantioselectividadEnantioselectividadEnantioselectividad de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de de los derivados de BTL2 catalizando la hidrólisis de (±)(±)(±)(±)----7 a pH 57 a pH 57 a pH 57 a pH 5.... ee= exceso enantiomérico de producto al 15% de conversión; E= valor de enantioselectividad.
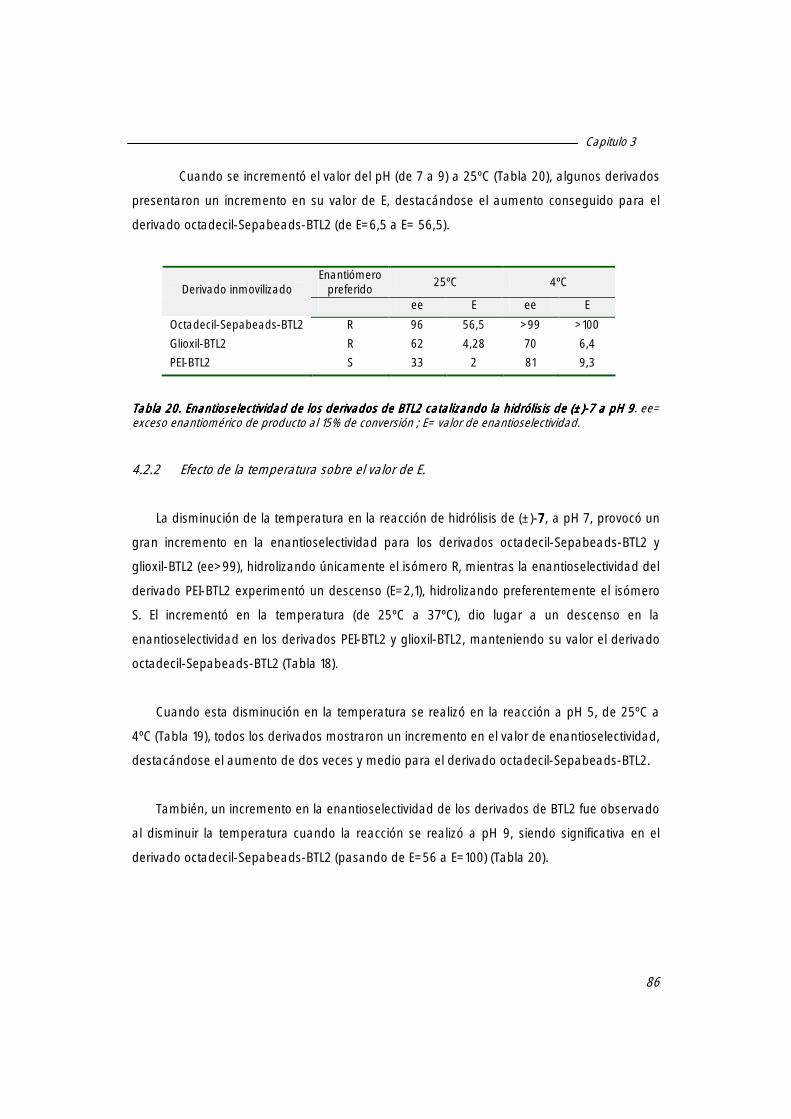
Capitulo 3
86
Cuando se incrementó el valor del pH (de 7 a 9) a 25ºC (Tabla 20), algunos derivados
presentaron un incremento en su valor de E, destacándose el aumento conseguido para el
derivado octadecil-Sepabeads-BTL2 (de E=6,5 a E= 56,5).
Derivado inmovilizado Enantiómero
preferido 25ºC 4ºC
ee E ee E
Octadecil-Sepabeads-BTL2 R 96 56,5 >99 >100
Glioxil-BTL2 R 62 4,28 70 6,4
PEI-BTL2 S 33 2 81 9,3
Tabla 20. Tabla 20. Tabla 20. Tabla 20. Enantioselectividad de los derivados de BTL2 catalizando la hidrólisis de Enantioselectividad de los derivados de BTL2 catalizando la hidrólisis de Enantioselectividad de los derivados de BTL2 catalizando la hidrólisis de Enantioselectividad de los derivados de BTL2 catalizando la hidrólisis de (±)(±)(±)(±)----7 a pH 97 a pH 97 a pH 97 a pH 9. ee= exceso enantiomérico de producto al 15% de conversión ; E= valor de enantioselectividad.
4.2.2 Efecto de la temperatura sobre el valor de E.
La disminución de la temperatura en la reacción de hidrólisis de (±)-7777, a pH 7, provocó un
gran incremento en la enantioselectividad para los derivados octadecil-Sepabeads-BTL2 y
glioxil-BTL2 (ee>99), hidrolizando únicamente el isómero R, mientras la enantioselectividad del
derivado PEI-BTL2 experimentó un descenso (E=2,1), hidrolizando preferentemente el isómero
S. El incrementó en la temperatura (de 25ºC a 37ºC), dio lugar a un descenso en la
enantioselectividad en los derivados PEI-BTL2 y glioxil-BTL2, manteniendo su valor el derivado
octadecil-Sepabeads-BTL2 (Tabla 18).
Cuando esta disminución en la temperatura se realizó en la reacción a pH 5, de 25ºC a
4ºC (Tabla 19), todos los derivados mostraron un incremento en el valor de enantioselectividad,
destacándose el aumento de dos veces y medio para el derivado octadecil-Sepabeads-BTL2.
También, un incremento en la enantioselectividad de los derivados de BTL2 fue observado
al disminuir la temperatura cuando la reacción se realizó a pH 9, siendo significativa en el
derivado octadecil-Sepabeads-BTL2 (pasando de E=56 a E=100) (Tabla 20).

Ingeniería conformacional de lipasas
87
Vemos una vez más, como se obtienen diferencias significativas en los valores de E para la
BTL2 al emplear diferentes protocolos de inmovilización y a su vez al modificar las condiciones
del medio.
5555 Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido Hidrólisis enantioselectiva del ácido (±)(±)(±)(±)----2222----OOOO----butirilbutirilbutirilbutiril----2222----fenilacfenilacfenilacfenilacéééético tico tico tico [[[[(±)(±)(±)(±)----7] catlizada7] catalizada7] catalizada7] catalizada
por derivados inmovilizados de por derivados inmovilizados de por derivados inmovilizados de por derivados inmovilizados de la la la la lipasalipasalipasalipasa de de de de Thermus thermophilusThermus thermophilusThermus thermophilusThermus thermophilus (TTL) (TTL) (TTL) (TTL)....
5.1. Actividad espec5.1. Actividad espec5.1. Actividad espec5.1. Actividad especíííífica de los distintos derivados inmovilizados de fica de los distintos derivados inmovilizados de fica de los distintos derivados inmovilizados de fica de los distintos derivados inmovilizados de TTL.TTL.TTL.TTL.
El estudio de la actividad catalítica de los derivados de TTL a 25ºC hacia distintos
sustratos se hace aun más interesante teniendo en cuenta la inactividad de la enzima soluble
en estas condiciones.
Frente al sustrato 5555, el derivado octadecil-Sepabeads-TTL fue el más activo, casi ocho
veces más que el derivado glioxil-TTL. Cuando se empleó el compuesto 7777, la actividad de los
derivados disminuyó frente a la obtenida con 5 5 5 5 siendo el derivado glioxil-TTL el más activo,
ahora casi dos veces más que el octadecil-Sepabeads-TTL (Tabla 21).
La disminución en el valor del pH del medio (de 7 a 5) en la hidrólisis de 7777 (Tabla 19),
condujo a un descenso en el valor de actividad de ambos derivados, aunque el derivado
glioxil-TTL continuó siendo el más activo frente a este compuesto.
Tabla 21Tabla 21Tabla 21Tabla 21. Resolución hidrolítica de (±)Resolución hidrolítica de (±)Resolución hidrolítica de (±)Resolución hidrolítica de (±)----7 catalizada por derivados inmovilizados de TTL a 25ºC.7 catalizada por derivados inmovilizados de TTL a 25ºC.7 catalizada por derivados inmovilizados de TTL a 25ºC.7 catalizada por derivados inmovilizados de TTL a 25ºC.aaaa. Actividad específica (AE): µmol.h-1.mglipasa-1 bexceso enantiomérico de producto. cPreferencia estereoquímica d Enantioselectividad.
Derivado inmovilizado 5555 (±)-7777
pH 7 pH 5 AEa AEa eeb Pec Ed AEa eeb Pec Ed
Octadecil-Sepabeads-TTL 2,5 0,015 80 R 9,2 6,55x10-3 21 R 1,53
Glioxil-TTL 0,32 0,033 63 R 4 0,0186 18 R 1,44

Capitulo 3
88
5.2. Enantioselectividad de los distintos derivados inmovilizados de 5.2. Enantioselectividad de los distintos derivados inmovilizados de 5.2. Enantioselectividad de los distintos derivados inmovilizados de 5.2. Enantioselectividad de los distintos derivados inmovilizados de TTLTTLTTLTTL catalizando la catalizando la catalizando la catalizando la
hidrólisis de hidrólisis de hidrólisis de hidrólisis de (±)-7 a 25ºC.7 a 25ºC.7 a 25ºC.7 a 25ºC.
En cuanto a la enantioselectividad de esta enzima frente al compuesto (±)-7 7 7 7 a pH 7
(Tabla 21), cabe destacar como todos los derivados hidrolizaron más deprisa el isómero R.
Además, el derivado octadecil-Sepabeads-TTL fue más enantioselectivo que el derivado
glioxil-TTL (E=9,2 frente a E=4).La disminución en el pH (de 7 a 5) tuvo un efecto negativo sobre
ambos derivados, decreciéndose la enantioselectividad hasta un valor de en torno a 1,5.
6666 Hidrólisis enantioselectiva del compuestoHidrólisis enantioselectiva del compuestoHidrólisis enantioselectiva del compuestoHidrólisis enantioselectiva del compuesto [[[[(±)(±)(±)(±)----7] catalizada por derivados 7] catalizada por derivados 7] catalizada por derivados 7] catalizada por derivados
inmovilizados de la inmovilizados de la inmovilizados de la inmovilizados de la lipasa de lipasa de lipasa de lipasa de Thermus aquaticusThermus aquaticusThermus aquaticusThermus aquaticus ( ( ( (TAL).TAL).TAL).TAL).
6.1. Actividad espec6.1. Actividad espec6.1. Actividad espec6.1. Actividad especíííífica de los distintos derivados inmovifica de los distintos derivados inmovifica de los distintos derivados inmovifica de los distintos derivados inmovilizados delizados delizados delizados de TAL TAL TAL TAL a 25ºC. a 25ºC. a 25ºC. a 25ºC.
Cuando se examinó el comportamiento de esta enzima, se observó como era la más
termoresistente de las tres lipasas de termófilos estudiadas, además de no presentar
demasiada actividad a temperaturas de 25ºC.
Por esto, quizás el comportamiento de esta lipasa frente a distintos sustratos podía
ser interesante. Como primera propiedad se analizó la actividad específica de dos derivados a
pH 7 frente a varios sustratos (Tabla 22).
Empleando el sustrato 5555, el derivado octadecil-Sepabeads-TAL fue el más activo con
una diferencia de cuarenta y seis veces respecto a la actividad del derivado glioxil-TAL. Sin
embargo, cuando se empleo el compuesto 7777 al mismo pH (pH 7), las actividades de ambos
derivados de TAL fueron similares, destacándose el descenso en la actividad del derivado
octadecil-TAL, mas de 400 veces menor que la actividad obtenida con el compuesto 5555.
El cambio de pH en la reacción con el compuesto 7777 (de pH 7 a 5) determinó un
descenso en la actividad para de ambos derivados de TAL, presentando de nuevo actividades
muy parecidas.
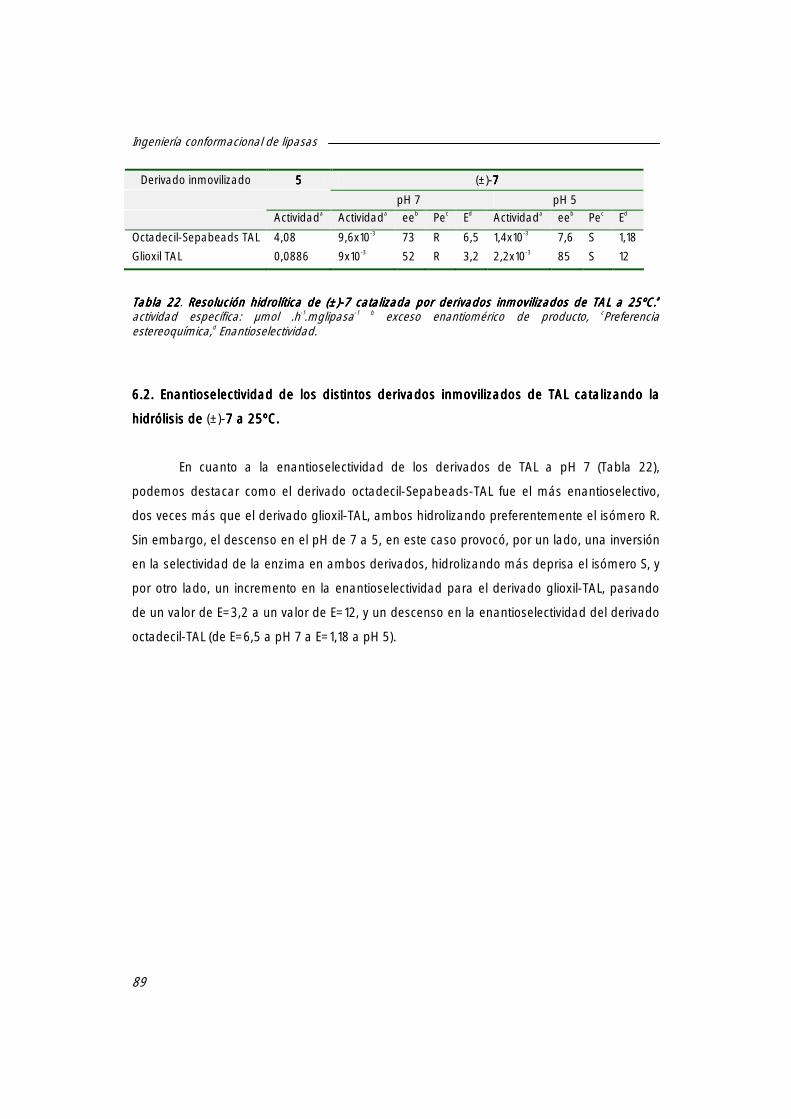
Ingeniería conformacional de lipasas
89
Derivado inmovilizado 5555 (±)-7777
pH 7 pH 5 Actividada Actividada eeb Pec Ed Actividada eeb Pec Ed
Octadecil-Sepabeads TAL 4,08 9,6x10-3 73 R 6,5 1,4x10-3 7,6 S 1,18
Glioxil TAL 0,0886 9x10-3 52 R 3,2 2,2x10-3 85 S 12
Tabla 22Tabla 22Tabla 22Tabla 22. Resolución hidrolítica de (±)Resolución hidrolítica de (±)Resolución hidrolítica de (±)Resolución hidrolítica de (±)----7 catalizada por derivados inmovilizados de TAL a 25ºC.7 catalizada por derivados inmovilizados de TAL a 25ºC.7 catalizada por derivados inmovilizados de TAL a 25ºC.7 catalizada por derivados inmovilizados de TAL a 25ºC.aaaa
actividad específica: µmol .h-1.mglipasa-1 b exceso enantiomérico de producto, cPreferencia estereoquímica,d Enantioselectividad.
6.2. Enantioselectividad de los distintos derivados inmovilizados de TAL catalizando la 6.2. Enantioselectividad de los distintos derivados inmovilizados de TAL catalizando la 6.2. Enantioselectividad de los distintos derivados inmovilizados de TAL catalizando la 6.2. Enantioselectividad de los distintos derivados inmovilizados de TAL catalizando la
hidrólisis de hidrólisis de hidrólisis de hidrólisis de (±)-7 a 25ºC.7 a 25ºC.7 a 25ºC.7 a 25ºC.
En cuanto a la enantioselectividad de los derivados de TAL a pH 7 (Tabla 22),
podemos destacar como el derivado octadecil-Sepabeads-TAL fue el más enantioselectivo,
dos veces más que el derivado glioxil-TAL, ambos hidrolizando preferentemente el isómero R.
Sin embargo, el descenso en el pH de 7 a 5, en este caso provocó, por un lado, una inversión
en la selectividad de la enzima en ambos derivados, hidrolizando más deprisa el isómero S, y
por otro lado, un incremento en la enantioselectividad para el derivado glioxil-TAL, pasando
de un valor de E=3,2 a un valor de E=12, y un descenso en la enantioselectividad del derivado
octadecil-TAL (de E=6,5 a pH 7 a E=1,18 a pH 5).

Capitulo 3
90
CONCLUSIONESCONCLUSIONESCONCLUSIONESCONCLUSIONES
Todos los resultados obtenidos sugieren que las propiedades de una misma lipasa
pueden ser fuertemente moduladas mediante lo que denominamos “ingeniería
conformacional”: inmovilización dirigida (alterando la rigidez, la orientación y el
microambiente) e ingeniería del medio.
Así, se observó como para unas condiciones de reacción determinadas, la misma
lipasa inmovilizada sobre distintos soportes (por ejemplo, presentando diferente orientación y
microambiente) exhiben muy diferentes propiedades catalíticas:
-Diferente actividad enzimática (incluso en un factor de 80 veces, por ejemplo, entre
los derivados octadecil-Sepabeads-CAL-B y Eupergit-Cu-CAL-B en la hidrólisis del compuesto 5555
-Diferente enantioselectividad, variando de E≅ 1 para un determinado derivado
inmovilizado a más de 100 para otro, como por ejemplo entre los derivados octil-CRL y
glutaraldehido-CRL en la hidrólisis de (±)-7777 a pH 7, además de observarse una inversión en la
selectividad de la enzima.
Al mismo tiempo, pequeños cambios en las condiciones de reacción, tuvieron efectos
muy significativos en las propiedades de los distintos derivados inmovilizados de cada lipasa;
como por ejemplo, en la hidrólisis del compuesto 7 7 7 7, el derivado octadecil-Sepabeads-BTL2,
pasó de tener una E=15 (a 25ºC) a más de 100 (a 4ºC), o el glioxil-TAL de tener E=4 (a pH 7)
hacia el isómero R a E=12 (a pH 5),y además, hacia el isómero contrario.
Todos estos resultados parecen soportar la hipótesis inicial expuesta en el esquema
15 de la introducción. El hecho de que estemos empleando lipasa purificada en todos los
casos, puede implicar que las diferencias entre los diferentes derivados inmovilizados, sólo
pueden ser explicadas por el distinto comportamiento de la enzima inmovilizada sobre los
diferentes soportes.
Por tanto vemos como esta metodología de la ingeniería conformacional es general
para las lipasas.

Ingeniería conformacional de lipasas
91
PARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTAL
1.1.1.1. MaterialesMaterialesMaterialesMateriales
Los geles de agarosa 6% y 10% entrecruzada (BCL) fueron donados por la compañía
Hispanagar SA (Burgos). Octadecil-Sepabeads y Sepabeads FP-EP (epóxido) fueron donados
por Resindion srl (Milán, Italia). La resina Eupergit-C fue donada por Röhm GMBH (Degussa-
Hüls Gruppe) (Darmstadt, Alemania). Además la polietilenimina (PEI) con una masa molecular
de 25 kDa, glutaraldehído al 25%, ácido iminodiacético (IDA), etilendiamina (EDA) y tritón X-100
se obtuvieron de sigma.
Los soportes PEI-agarosa68,106-107, glioxil-agarosa69,82-90, glutaraldehído-agarosa80-81,IDA-
Sepabeads, EDA-Sepabeads, quelatos de cobre Sepabeads (IDA-Cu2+) o quelatos de cobre
sobre Eupergit C (Eupergit-Cu)108-109 se prepararon como esta descrito en la literatura.
2222. Inmovilización de las distin. Inmovilización de las distin. Inmovilización de las distin. Inmovilización de las distintas lipasas sobre los distintos soportestas lipasas sobre los distintos soportestas lipasas sobre los distintos soportestas lipasas sobre los distintos soportes
Se prepararon derivados inmovilizados con concentración de lipasa purificada de 1
mg de lipasa/mL, evitando así problemas de difusión. En todos los casos se obtuvo más de un
95% de enzima inmovilizada. La metodología empleada en la purificación y en el seguimiento
de inmovilización para cada lipasa se realizó como se describe en el capítulo 2.
La preparación de los derivados inmovilizados octadecil-Sepabeads, glioxil,
bromocianógeno y glutaraldehído fue descrita en el capítulo 2.
2.1 Inmovilización de lipasas por adsorción iónica
La inmovilización fue realizada empleando soportes PEI-agarosa. 120 mL de una solución
de lipasa pura en tampón bicarbonato 5 mM pH 9 se ofreció a 3 g de soporte y se siguió la
inmovilización por medición de alícuotas de sobrenadante y suspensión. En el caso de CRL a
inmovilización se realizó a pH 7. Después de 3 horas el derivado inmovilizado se filtró y se lavó
con abundante agua destilada.

Capitulo 3
92
2.2 Inmovilización de lipasas sobre soportes Sepabeads heretofuncionales y Eupergit-Cu
7g de soporte (EDA-Sepabeads, IDA-Sepabeads, IDA-Cu2+-Sepabeads y Eupergit-Cu)
se suspendió sobre 25 mL de solución de lipasas purificada en tampón fosfato sódico 25 mM
a pH 7 y 25ºC. Periódicamente se analizó el estado de la inmovilización covalente mediante el
seguimiento de la actividad enzimática de sobrenadante y suspensión frente a pNPP. La
inmovilización fue realizada en condiciones donde en primer lugar se producía una absorción
física de las moléculas de enzima sobre la superficie del soporte dando lugar así a la posterior
unión covalente. Después de la inmovilización, los derivados se incubaron en presencia de
glicina 3 M a pH 8,5 durante 24 horas para bloquear los grupos epóxidos remanentes y
promover la hidrofilización de la superficie del soporte. Finalmente los derivados se lavaron
abundantemente con agua destilada.
En la Tabla 23 se representa la actividad pNPP recuperada para los distintos derivados
de las diferentes lipasas tras su inmovilización en los diferentes soportes.
Lipasa Derivado inmovilizado Actividad relativa pNPP PEI 0,8 CAL-B BrCN 0,95 Eupergit-Cu 1 CRL PEI 0,62 PEI 0,78 EDA-Sepabeads 0,44 MML IDA-Sepabeads 0,3 IDA-Cu+2-Sepabeads 0,3 BTL2 PEI 0,9
Tabla 23Tabla 23Tabla 23Tabla 23. Activi. Activi. Activi. Actividad relativa (dad relativa (dad relativa (dad relativa (ttttanto por uno) de los derivados inmovilizados de lipasa con pNPPanto por uno) de los derivados inmovilizados de lipasa con pNPPanto por uno) de los derivados inmovilizados de lipasa con pNPPanto por uno) de los derivados inmovilizados de lipasa con pNPP. Los derivados presentan 1 mg lipasa pura/mL como carga enzimática. La lipasa soluble tiene el valor de uno en cada caso. 3333. Hidrólisis enzimática de los diferentes sustratos.. Hidrólisis enzimática de los diferentes sustratos.. Hidrólisis enzimática de los diferentes sustratos.. Hidrólisis enzimática de los diferentes sustratos.
La actividad enzimática de los diferentes derivados inmovilizados de las distintas
lipasas se analizó en la reacción de hidrólisis en medio acuoso de varios sustratos. El butirato
de etilo (5555) se disolvió a 25ºC en una solución de 100 mL de tampón fosfato sódico 25 mM a
pH 7 hasta una concentración de 10 mM y se añadió 0,1 gramos de derivado inmovilizado. El

Ingeniería conformacional de lipasas
93
mandelato de metilo [(±),(-),(+)-1111] se disolvió en una solución de tampón fosfato sódico 25 mM
a distintas condiciones (pH, Tª, co-disolvente) hasta una concentración de 10 mM y se añadió
0,2 gramos de catalizador. En el caso del ácido 2-O-butiril-2-fenilacético [(±)-7777], este se
disolvió en diferentes condiciones en una solución de 6 mL de tampón fosfato sódico 10 mM
hasta una concentración de 0,5 mM y se añadió 0,5 g de derivado inmovilizado. El pH de la
reacción se mantuvo constante en cada caso mediante la adición automática de NaOH en un
pHstato Mettler Toledo D50.
3.1 Análisis en HPLC.
El grado de hidrólisis se analizó por HPLC en fase reversa (Spectra Physic SP 100)
acoplado a un detector UV (Spectra Physic SP 8450) y utilizando como columna una Kromasil
C18 (25 x 0,4 cm) suministrada por Análisis Vinicos (España). Cada ensayo se realizó por
triplicado. La fase móvil fue una mezcla isocrática de acetonitrilo y tampón fosfato amónico 10
mM en proporción 30:70 para el sustrato 1111 y 35:65 para los compuestos 5555 y 7777, a pH 2,95 a un
flujo de 1,5 mL/min. La absorbancia empleada en el análisis fue 254 nm (para 1 1 1 1 y 7 7 7 7) o 225 nm
(para 5555). Los tiempos de retención fueron los siguientes: ácido mandélico (2222) [2,9 min (a 30:70),
3,1 min (a 35:65)], mandelato de metilo (1111) (8 min), ácido butírico (6666) (2,95 min), butirato de etilo
(5555) (16 min), ácido 2-O-butiril-2-fenilacético (7777) (18 min).
La actividad específica (AE) se definió como µmol de sustrato hidrolizado por minuto (o
por hora, donde la actividad fue realmente baja) y por miligramo de lipasa pura, y fue
calculada mediante la siguiente ecuación:
[S].c.V [S]=concentración de sustrato (mM) AE= c= conversión (tanto por uno) m.t V= volumen de reacción m= mg de lipasa t= tiempo de reacción (min o horas)
La actividad enzimática se define de la misma forma, donde m=gramos de catalizador. 4444. Determinación del exceso enantiomérico y la Enantioselectividad. Determinación del exceso enantiomérico y la Enantioselectividad. Determinación del exceso enantiomérico y la Enantioselectividad. Determinación del exceso enantiomérico y la Enantioselectividad
A diferentes grados de conversión (entre 10-15% conversión) se analizó el exceso
enantiomérico (ee) del ácido formado a través de HPLC en fase quiral reversa. La columna

Capitulo 3
94
empleada fue una Chiracel OD-R mientras la fase móvil fue una mezcla isocrática de
acetonitrilo y NaClO4/HClO4 0,5 M en proporción 5:95 a pH 2,3. El análisis se realizó a un
flujo de 0,5 mL/min y la absorbancia empleada fue de 225 nm. El isómero S del ácido
mandélico (2222) aparece a un tiempo de retención de 39 minutos y el isómero R a 42
minutos.
La Enantioselectividad se expresó como el valor E calculado a partir del exceso
enantiomérico de producto (ee) y del grado de conversión (c) (en tanto por uno), de
acuerdo a la ecuación73:
ln [1-c(1+ee)] E= ln [1-c(1-ee)]
5555. Síntesis del ácido 2. Síntesis del ácido 2. Síntesis del ácido 2. Síntesis del ácido 2----OOOO----butirilbutirilbutirilbutiril----2222----fenilacéticofenilacéticofenilacéticofenilacético [(±)-7777]
Los espectros de 1H- y 13C-RMN se obtuvieron empleando un espectrometro Bruker AC-
300 (1H-250 MHz y 13C-75,5 MHz) con TMS (tetremetilsilano) como patrón interno
A 3,042 gramos de ácido mandélico [(±)-2222] (20 mmoles) en 200 mL de eter dietílico se
añadió 2,88 mL de NEt3 (20 mmoles). 2,131 mL (20 mmoles) de cloruro de butirilo en 100 mL de
éter se adicionó lentamente sobre la disolución anterior en agitación continua. La mezcla se
mantuvo a 25ºC durante 4 horas, obteniendo un rendimiento de síntesis del 50% (el
seguimiento de la síntesis se llevo a cabo por HPLC, condiciones anteriormente descritas).
Después de esto se extrajo con agua y eter dietílico. La fase orgánica fue secada sobre sulfato
sódico, filtrado y evaporado el disolvente empleando un rotavapor a vacío. Se obtuvo un
aceite amarillento (1,769 g). Pm: 222,1, 1H-RMN (CDCl3): δ(ppm): 0,95 (m, 3H), 1,7 (septete, 2H),
2,4 (m,2H) , 5,9 (s,1H, CH), 7,4 (m, 4H), 13C NMR (CDCl3): 173(C=O), 170 (C=O), 131,8 (C), 127,8
(CH), 127,3 (CH), 126,1 (CH), 76,018 (CH), 34,24 (CH2), 16,76 (CH2) ,12,065 (CH3).

Capítulo 4
95
INTRODUCCIONINTRODUCCIONINTRODUCCIONINTRODUCCION
La zopiclona (ácido 4-metil-1-piperazinacarboxílico-6-(5-cloro-2-piridinil)-6,7-dihidro-7-
oxo-5H-pirrolo[3,4-b]-pirazin-5-il ester) (8888) es una ciclopirrolona con propiedades hipnóticas de
acción no prolongada y algunos efectos laterales asociados. La molécula de zopiclona posee
un centro quiral aunque hasta el momento se comercializa en forma de mezcla racémica.
Estudios recientes121 han confirmado que el enantiómero dextrógiro (isómero S) es
aproximadamente dos veces más activo que el racémico, mientras que el isómero levógiro es
prácticamente inactivo. Además, según la patente EP 609210-B1, el isómero levógiro es el
causante de la mayor parte de los efectos secundarios asociados a la administración del
medicamento,lo cual hace más conveniente el uso del enantiómero S ópticamente puro.
Los enantiómeros de la zopiclona se pueden separar mediante cristalización
fraccionada de las sales diastereoisómeras formadas con un ácido ópticamente activo;como,
por ejemplo, el ácido (+)-O,O’-dibenzoiltartárico (patente EP 609210-B1) o los ácidos (+) o (–)-
málico122. También pueden ser separados mediante cromatografía sobre fase estacionaria
quiral 123.
Esquema Esquema Esquema Esquema 24242424
Por otro lado, en la obtención del fármaco opticamente puro se emplearon unos
carbonatos derivados (R1= fenilo opcionalmente sustituido) los cuales se obtuvieron a partir de
la reacción del alcohol 9 9 9 9 con un cloroformiato (Cl-CO-OR1)(Esquema 24).
Ademas se ha descrito la preparación de (S)-(+)-8888 a partir de un carbonato
ópticamente activo (con R1= vinilo), obtenido a su vez mediante resolución enzimática de una
NN
NO
N
O
O
N N
ClNN
N
O
O
N
H
Cl
O
OR1
NN
NO
N
OHH
Cl
(S)-(+)-8 carbonatos (±)-9
NN
NO
N
O
O
N N
ClNN
N
O
O
N
H
Cl
O
OR1
NN
NO
N
OHH
Cl
(S)-(+)-8 carbonatos (±)-9
NN
NO
N
O
O
N N
ClNN
N
O
O
N
H
Cl
O
OR1
NN
NO
N
OHH
Cl
(S)-(+)-8 carbonatos (±)-9
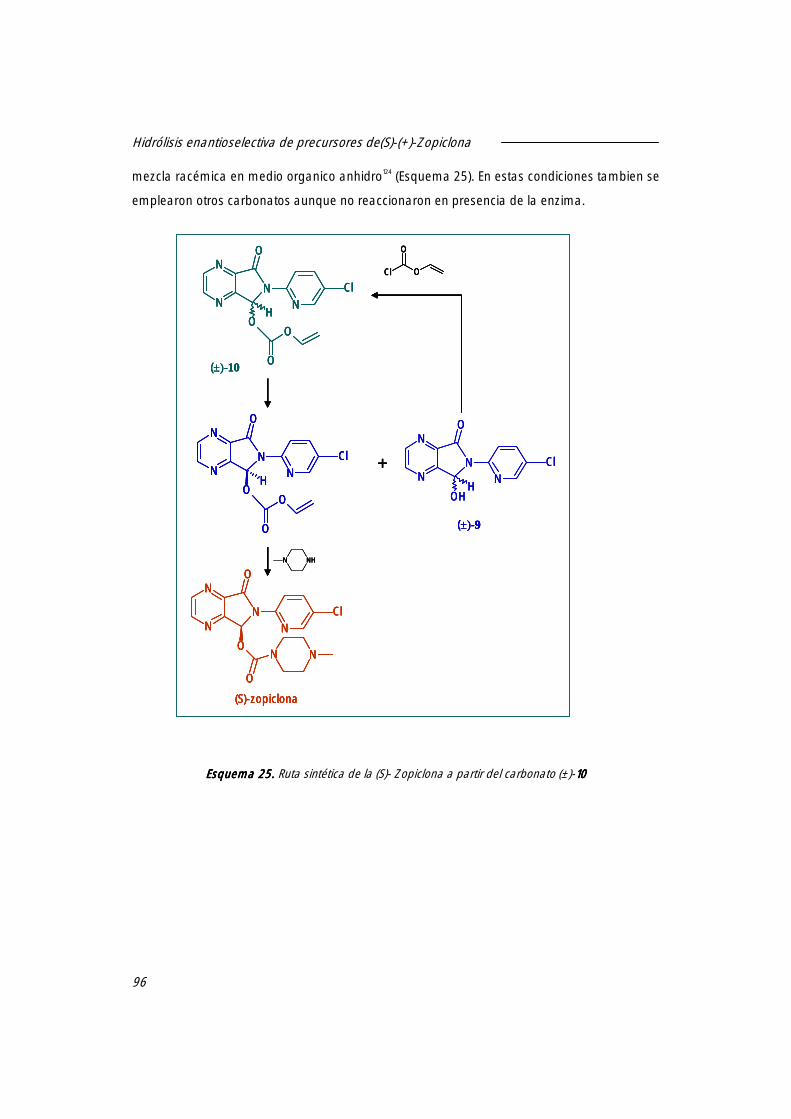
Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
96
mezcla racémica en medio organico anhidro124 (Esquema 25). En estas condiciones tambien se
emplearon otros carbonatos aunque no reaccionaron en presencia de la enzima.
Esquema Esquema Esquema Esquema 22225555. . . . Ruta sintética de la (S)- Zopiclona a partir del carbonato (±)-10101010
NN
NO
N
OHH
Cl
NN
N
O
O
N
H
Cl
O
O
NN
N
O
O
N
H
Cl
O
O
NN
NO
N
O
O
N N
Cl
+
N NH
Cl
O
O
(±)-9
(S)-zopiclona
(±)-10
NN
NO
N
OHH
Cl
NN
N
O
O
N
H
Cl
O
O
NN
N
O
O
N
H
Cl
O
O
NN
NO
N
O
O
N N
Cl
+
N NH
Cl
O
O
(±)-9
(S)-zopiclona
(±)-10
NN
NO
N
OHH
Cl
NN
N
O
O
N
H
Cl
O
O
NN
N
O
O
N
H
Cl
O
O
NN
NO
N
O
O
N N
Cl
+
N NH
Cl
O
O
(±)-9
(S)-zopiclona
(±)-10
NN
N
O
O
N
H
Cl
O
O
NN
N
O
O
N
H
Cl
O
O
NN
NO
N
O
O
N N
Cl
+
N NH
Cl
O
O
(±)-9
(S)-zopiclona
(±)-10

Capítulo 4
97
OBJETIVOSOBJETIVOSOBJETIVOSOBJETIVOS
En este capítulo se estudiará la aplicación de diferentes derivados inmovilizados de la
lipasa de C. antarctica B como biocatalizadores en la resolución enzimática de varios
compuestos mediante la reacción de hidrólisis en medios acuosos, en busca de:
i) el derivado inmovilizado óptimo, y
ii) nuevos intermedios ópticamente activos empleados como precursores en la
síntesis de la (S)-(+)-Zopiclona, enantiómero con las características biológicas
adecuadas.

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
98
RESULTADOS Y DISCUSIONRESULTADOS Y DISCUSIONRESULTADOS Y DISCUSIONRESULTADOS Y DISCUSION
En el capítulo anterior se demostró como la inmovilización dirigida de lipasas y la
ingeniería del medio tenía una gran influencia en las propiedades catalíticas de una misma
lipasa (actividad y enantioselectividad)125-128, empleándose esteres derivados del ácido
mandélico como sustratos modelos en la reacción de hidrólisis en medios acuosos; sin
embargo, en el presente capítulo emplearemos estos conocimientos en el estudio de un caso
concreto de aplicación industrial como la síntesis de la Zopiclona enantioméricamente pura.
Para ello se estudió la resolución enzimática mediante hidrólisis en medios acuosos de una
gran batería de precursores [(±)-10101010----16161616] (Esquema 26). Como catalizador se empleo la lipasa de
C. antarctica B inmovilizada sobre diferentes soportes. Hasta ahora la concentración de lipasa
pura empleada en nuestros derivados había sido de 1 mg/mL (capítulos 2 y 3), evitándose así
posibles problemas de difusión; sin embargo, la baja actividad presentada por los distintos
derivados de CAL-B frente a estos precursores nos permitió prepararlos con una concentración
de lipasa purificada de 12 mg/mL.
Esquema 2Esquema 2Esquema 2Esquema 26666.... Diferentes precursores de la zopiclona : (±)-5-sustituido-6-(5-cloropirid-2-il)-7-oxo-5,6-dihidropirrolo[3,4b] pirazina
NN
N
O
O
N
H
Cl
O
R
OClO
ClO
Cl
Cl ClO
R=
CH3
(±)-10101010
(±)-11111111
(±)-12121212
(±)-13131313
(±)-14141414
(±)-15151515
(±)-16161616
NN
N
O
O
N
H
Cl
O
R
OClO
ClO
Cl
Cl ClO
R=
CH3
(±)-10101010
(±)-11111111
(±)-12121212
(±)-13131313
(±)-14141414
(±)-15151515
(±)-16161616
NN
N
O
O
N
H
Cl
O
R
OClO
ClO
Cl
Cl ClO
R=
CH3
(±)-10101010
(±)-11111111
(±)-12121212
(±)-13131313
(±)-14141414
(±)-15151515
(±)-16161616

Capítulo 4
99
1.1.1.1. Resolución enantioselectiva del carbonato de vinilo [Resolución enantioselectiva del carbonato de vinilo [Resolución enantioselectiva del carbonato de vinilo [Resolución enantioselectiva del carbonato de vinilo [(±)-10101010] utilizando derivados ] utilizando derivados ] utilizando derivados ] utilizando derivados
inmovilizados de CALinmovilizados de CALinmovilizados de CALinmovilizados de CAL----B.B.B.B.
De todos los carbonatos empleados, el vinil carbonato (±)-10101010 ya había sido utilizado como
posible precursor de la (S)-Zopiclona; Gotor y col124 realizaron la resolución enzimática de este
compuesto en medio orgánico anhidro utilizando como nucleófilo una molécula de agua o de
alcohol.
Sin embargo, en esta tesis nos planteamos el estudio de la resolución enzimática de (±)-
10101010 en medios acuosos (Esquema 27) empleando concentraciones de co-disolvente más bajas
(restringidas a la solubilización del sustrato).
Esquema 2Esquema 2Esquema 2Esquema 27777. . . . Hidrólisis enantioselectiva de precursores de la zopiclona, catalizada por diferentes
derivados inmovilizados de CAL-B
En primer lugar se realizó un estudio de la actividad enzimática y la actividad específica de
una serie de derivados inmovilizados de CAL-B en la hidrólisis del sustrato (±)-10101010 a 25ºC bajo
una concentración dada empleando la menor cantidad de co-disolvente para solubilizarlo
(0,05 mM con 10% de 1,4-dioxano). Se definieron estas dos actividades debido a que solo
podemos determinar la actividad enzimática para el derivado comercial (ya que
desconocemos la cantidad de enzima por gramo de soporte de catalizador), de manera que
se pudiera hacer un estudio comparativo de los distintos derivados inmovilizados. En este
NN
N
O
O
N
H
Cl
O
R
NN
N
O
O
N
H
Cl
O
R
NN
NO
N
OHH
Cl
(±)-9
NN
NO
N
O
O
N N
Cl
+
(S)-(+)-8888 (Zopiclona)
CAL-B
H2O
(s)-(+)-10-16
+-( )-10-16
NN
N
O
O
N
H
Cl
O
R
NN
N
O
O
N
H
Cl
O
R
NN
NO
N
OHH
Cl
(±)-9
NN
NO
N
O
O
N N
Cl
+
(S)-(+)-8888 (Zopiclona)
CAL-B
H2O
(s)-(+)-10-16
+-( )-10-16
NN
N
O
O
N
H
Cl
O
R
NN
N
O
O
N
H
Cl
O
R
NN
NO
N
OHH
Cl
(±)-9
NN
NO
N
O
O
N N
Cl
+
(S)-(+)-8888 (Zopiclona)
CAL-B
H2O
(s)-(+)-10-16
+-( )-10-16+-( )+-( )-( )-10-16
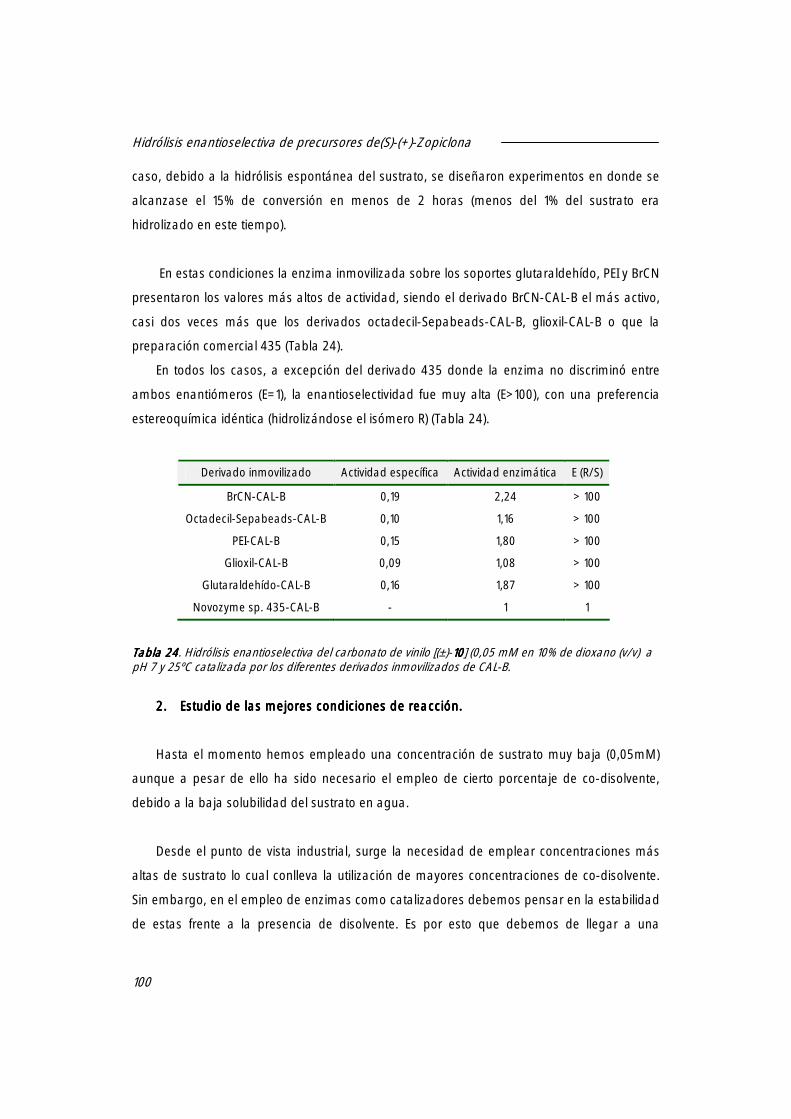
Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
100
caso, debido a la hidrólisis espontánea del sustrato, se diseñaron experimentos en donde se
alcanzase el 15% de conversión en menos de 2 horas (menos del 1% del sustrato era
hidrolizado en este tiempo).
En estas condiciones la enzima inmovilizada sobre los soportes glutaraldehído, PEI y BrCN
presentaron los valores más altos de actividad, siendo el derivado BrCN-CAL-B el más activo,
casi dos veces más que los derivados octadecil-Sepabeads-CAL-B, glioxil-CAL-B o que la
preparación comercial 435 (Tabla 24).
En todos los casos, a excepción del derivado 435 donde la enzima no discriminó entre
ambos enantiómeros (E=1), la enantioselectividad fue muy alta (E>100), con una preferencia
estereoquímica idéntica (hidrolizándose el isómero R) (Tabla 24).
Derivado inmovilizado Actividad específica Actividad enzimática E (R/S)
BrCN-CAL-B 0,19 2,24 > 100
Octadecil-Sepabeads-CAL-B 0,10 1,16 > 100
PEI-CAL-B 0,15 1,80 > 100
Glioxil-CAL-B 0,09 1,08 > 100
Glutaraldehído-CAL-B 0,16 1,87 > 100
Novozyme sp. 435-CAL-B - 1 1
Tabla 24Tabla 24Tabla 24Tabla 24. Hidrólisis enantioselectiva del carbonato de vinilo [(±)-10101010] (0,05 mM en 10% de dioxano (v/v) a pH 7 y 25ºC catalizada por los diferentes derivados inmovilizados de CAL-B.
2.2.2.2. Estudio de las mejores Estudio de las mejores Estudio de las mejores Estudio de las mejores condiciones de condiciones de condiciones de condiciones de reacción.reacción.reacción.reacción.
Hasta el momento hemos empleado una concentración de sustrato muy baja (0,05mM)
aunque a pesar de ello ha sido necesario el empleo de cierto porcentaje de co-disolvente,
debido a la baja solubilidad del sustrato en agua.
Desde el punto de vista industrial, surge la necesidad de emplear concentraciones más
altas de sustrato lo cual conlleva la utilización de mayores concentraciones de co-disolvente.
Sin embargo, en el empleo de enzimas como catalizadores debemos pensar en la estabilidad
de estas frente a la presencia de disolvente. Es por esto que debemos de llegar a una
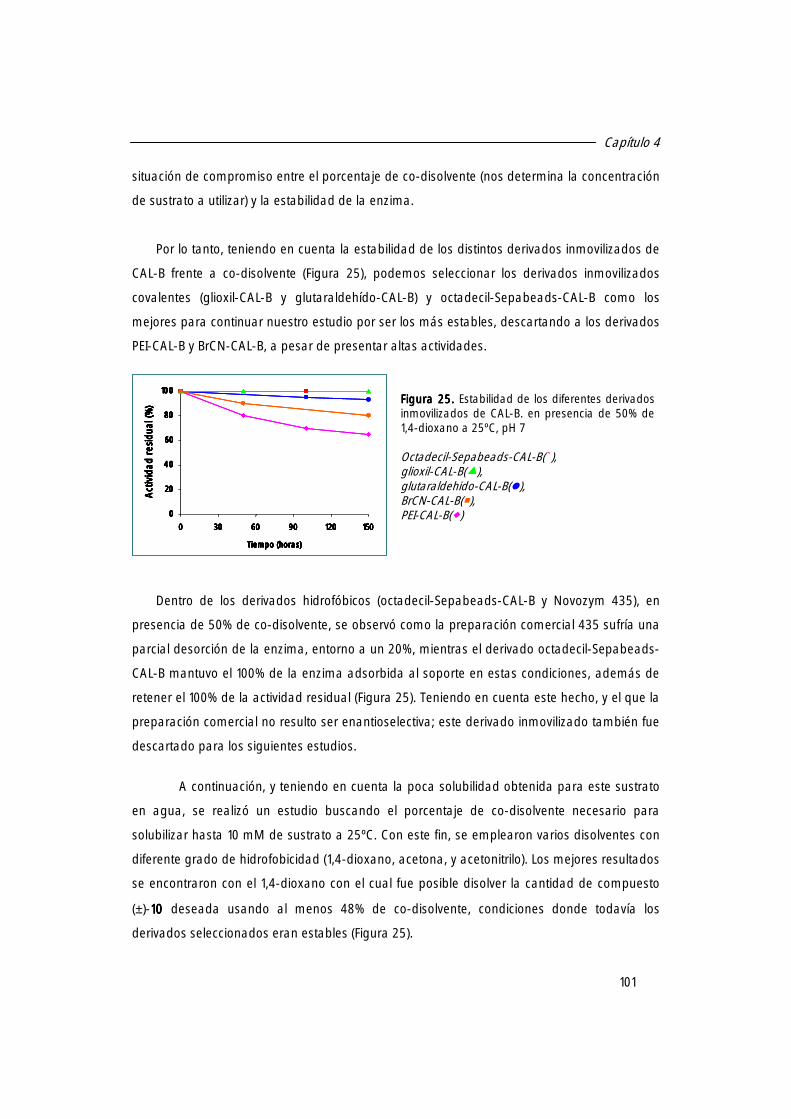
Capítulo 4
101
situación de compromiso entre el porcentaje de co-disolvente (nos determina la concentración
de sustrato a utilizar) y la estabilidad de la enzima.
Por lo tanto, teniendo en cuenta la estabilidad de los distintos derivados inmovilizados de
CAL-B frente a co-disolvente (Figura 25), podemos seleccionar los derivados inmovilizados
covalentes (glioxil-CAL-B y glutaraldehído-CAL-B) y octadecil-Sepabeads-CAL-B como los
mejores para continuar nuestro estudio por ser los más estables, descartando a los derivados
PEI-CAL-B y BrCN-CAL-B, a pesar de presentar altas actividades.
Figura 25Figura 25Figura 25Figura 25. . . . Estabilidad de los diferentes derivados inmovilizados de CAL-B. en presencia de 50% de 1,4-dioxano a 25ºC, pH 7 Octadecil-Sepabeads-CAL-B(` ), glioxil-CAL-B(����), glutaraldehido-CAL-B(�), BrCN-CAL-B(�), PEI-CAL-B(�)
Dentro de los derivados hidrofóbicos (octadecil-Sepabeads-CAL-B y Novozym 435), en
presencia de 50% de co-disolvente, se observó como la preparación comercial 435 sufría una
parcial desorción de la enzima, entorno a un 20%, mientras el derivado octadecil-Sepabeads-
CAL-B mantuvo el 100% de la enzima adsorbida al soporte en estas condiciones, además de
retener el 100% de la actividad residual (Figura 25). Teniendo en cuenta este hecho, y el que la
preparación comercial no resulto ser enantioselectiva; este derivado inmovilizado también fue
descartado para los siguientes estudios.
A continuación, y teniendo en cuenta la poca solubilidad obtenida para este sustrato
en agua, se realizó un estudio buscando el porcentaje de co-disolvente necesario para
solubilizar hasta 10 mM de sustrato a 25ºC. Con este fin, se emplearon varios disolventes con
diferente grado de hidrofobicidad (1,4-dioxano, acetona, y acetonitrilo). Los mejores resultados
se encontraron con el 1,4-dioxano con el cual fue posible disolver la cantidad de compuesto
(±)-10 10 10 10 deseada usando al menos 48% de co-disolvente, condiciones donde todavía los
derivados seleccionados eran estables (Figura 25).
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
0000
20202020
40404040
60606060
80808080
100100100100
0000 30303030 60606060 90909090 120120120120 150150150150
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)
Act
ivid
ad r
esid
ual (
%)

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
102
El aumento en la concentración de co-disolvente en el medio de reacción, junto al
incremento en la concentración de sustrato provocó un importante descenso en la hidrólisis
química del sustrato (menos del 0,5% durante el tiempo de reacción).
Para determinar el posible efecto inhibitorio sobre la actividad de los distintos
derivados inmovilizados al incrementarse la concentración de co-disolvente, se llevo a cabo la
reacción de hidrólisis del carbonato de vinilo a 0,05 mM pero empleando 50% de co-
disolvente (asegurando la total solubilización del sustrato) (Tabla 25), comparando la actividad
de los derivados con las obtenidas cuando se empleo 10% de co-disolvente (Tabla 24).
Se observó como el efecto fue distinto para cada derivado inmovilizado,
produciéndose una menor disminución en la actividad para el derivado glioxil-CAL-B de 120
veces al aumentar la concentración de dioxano.
Tabla 25. Tabla 25. Tabla 25. Tabla 25. ActiviActiviActiviActividad de los diferentes dad de los diferentes dad de los diferentes dad de los diferentes derivados inmoderivados inmoderivados inmoderivados inmovilizavilizavilizavilizados en la hidrólisis dos en la hidrólisis dos en la hidrólisis dos en la hidrólisis de (de (de (de (±±±±))))----10 10 10 10 a 0,05 mM, pH 7 y 25ºC con 50% de dioxano.
3. Hidrólisis 3. Hidrólisis 3. Hidrólisis 3. Hidrólisis enantioselectiva enantioselectiva enantioselectiva enantioselectiva de (de (de (de (±±±±))))----10 10 10 10 empleando una concentración de 10 mM.empleando una concentración de 10 mM.empleando una concentración de 10 mM.empleando una concentración de 10 mM.
Una vez seleccionados los mejores derivados de CAL-B, la concentración de sustrato,
así como el co-disolvente y su porcentaje en el medio de reacción, se llevo a cabo la reacción
de hidrólisis de (±)-10101010 en dichas condiciones (Tabla 26).
En cuanto a la actividad específica, los tres derivados inmovilizados presentaron
valores muy similares, siendo ligeramente más altos para los dos covalentes (glioxil-CAL-B y
glutaraldehído-CAL-B).
Sin embargo, referente a la enantioselectividad, podemos destacar como en general, todos
los derivados de CAL-B fueron altamente enantioselectivos (hidrolizando exclusivamente el
isómero R), aunque en un seguimiento más riguroso se determinó al derivado octadecil-
Sepabeads-CAL-B como el más enantioselectivo (ee>99%).
Derivado inmovilizado Actividad específica
(x 10 -3 )
Octadecil-Sepabeads-CAL-B 0,59
Glioxil- CAL-B 0,75
Glutaraldehído- CAL-B 0,70
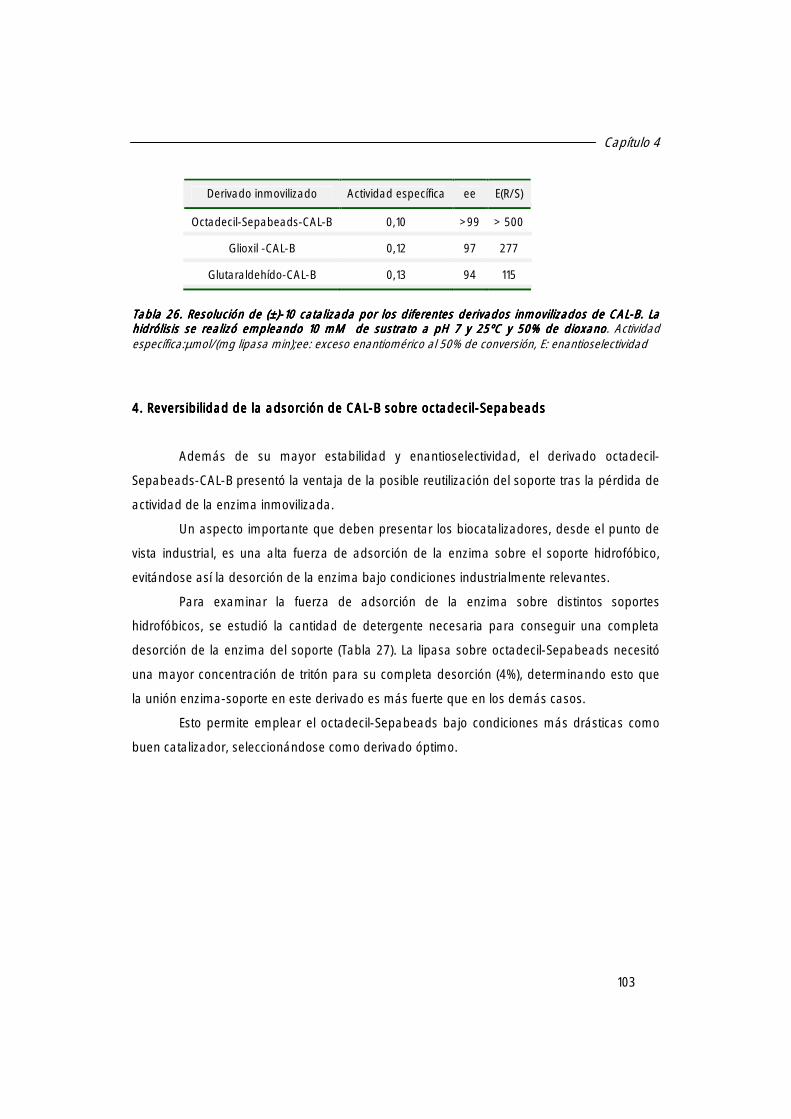
Capítulo 4
103
Tabla 26. Tabla 26. Tabla 26. Tabla 26. Resolución de (Resolución de (Resolución de (Resolución de (±±±±))))----10 catalizada por los diferentes derivados inmovilizados de CAL10 catalizada por los diferentes derivados inmovilizados de CAL10 catalizada por los diferentes derivados inmovilizados de CAL10 catalizada por los diferentes derivados inmovilizados de CAL----B. La B. La B. La B. La hidrólisis se realizó empleando 10 mM de sustrato a pH 7 y 2hidrólisis se realizó empleando 10 mM de sustrato a pH 7 y 2hidrólisis se realizó empleando 10 mM de sustrato a pH 7 y 2hidrólisis se realizó empleando 10 mM de sustrato a pH 7 y 25ºC y 50% de dioxano5ºC y 50% de dioxano5ºC y 50% de dioxano5ºC y 50% de dioxano. Actividad específica:µmol/(mg lipasa min);ee: exceso enantiomérico al 50% de conversión, E: enantioselectividad
4. 4. 4. 4. Reversibilidad de la adsorción de Reversibilidad de la adsorción de Reversibilidad de la adsorción de Reversibilidad de la adsorción de CALCALCALCAL----B sobre octadecilB sobre octadecilB sobre octadecilB sobre octadecil----Sepabeads Sepabeads Sepabeads Sepabeads
Además de su mayor estabilidad y enantioselectividad, el derivado octadecil-
Sepabeads-CAL-B presentó la ventaja de la posible reutilización del soporte tras la pérdida de
actividad de la enzima inmovilizada.
Un aspecto importante que deben presentar los biocatalizadores, desde el punto de
vista industrial, es una alta fuerza de adsorción de la enzima sobre el soporte hidrofóbico,
evitándose así la desorción de la enzima bajo condiciones industrialmente relevantes.
Para examinar la fuerza de adsorción de la enzima sobre distintos soportes
hidrofóbicos, se estudió la cantidad de detergente necesaria para conseguir una completa
desorción de la enzima del soporte (Tabla 27). La lipasa sobre octadecil-Sepabeads necesitó
una mayor concentración de tritón para su completa desorción (4%), determinando esto que
la unión enzima-soporte en este derivado es más fuerte que en los demás casos.
Esto permite emplear el octadecil-Sepabeads bajo condiciones más drásticas como
buen catalizador, seleccionándose como derivado óptimo.
Derivado inmovilizado Actividad específica ee E(R/S)
Octadecil-Sepabeads-CAL-B 0,10 >99 > 500
Glioxil -CAL-B 0,12 97 277
Glutaraldehído-CAL-B 0,13 94 115

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
104
Tabla 27. Tabla 27. Tabla 27. Tabla 27. Fuerza de unión de CALFuerza de unión de CALFuerza de unión de CALFuerza de unión de CAL----B sobre B sobre B sobre B sobre distintos soportes hidrofóbicos.distintos soportes hidrofóbicos.distintos soportes hidrofóbicos.distintos soportes hidrofóbicos. Los números expresan la cantidad de detergente necesario para desorber 100% de la lipasa del soporte. El análisis se llevo acabo midiendo la actividad enzimática de sobrenadantes y suspensiones en la hidrólisis de pNPP.
Por tanto, se llevo a cabo el curso de reacción de hidrólisis de (±)-10101010 utilizando como
catalizador este derivado inmovilizado (Figura 26). La relación volumen de reacción: cantidad
de derivado empleado fue de 4:1. La velocidad de reacción fue elevada, alcanzándose una
conversión de 45% en 55 minutos. Después de 48 horas, el valor de conversión no sobrepasó
el 50%, lo cual nos indicó la alta enantioselectividad mostrada por este derivado hacia el
sustrato (E>500)
Figura 2Figura 2Figura 2Figura 26666. Curso de reacción de hidrólisis Curso de reacción de hidrólisis Curso de reacción de hidrólisis Curso de reacción de hidrólisis enzimáticaenzimáticaenzimáticaenzimática de (±)-10101010 con el derivado octadecil-Sepabeads-CAL-B.
5.5.5.5. Resolución enzimática de Resolución enzimática de Resolución enzimática de Resolución enzimática de nuevos precursores de la Zopicnuevos precursores de la Zopicnuevos precursores de la Zopicnuevos precursores de la Zopiclonalonalonalona
En el apartado anterior se mostró como la resolución enzimática del compuesto (±)-10101010 fue
efectiva, especialmente con la lipasa inmovilizada sobre el soporte octadecil-Sepabeads.
Desafortunadamente, el alto coste del cloroformiato de vinilo, requerido para la síntesis del
compuesto (±)-10101010, es un inconveniente para el escalado del proceso. De esta manera, los
carbonatos (±)-14141414, , , , (±)-15151515 y (±)-11116666 (Esquema 26), que también son precursores de la Zopiclona,
fueron estudiados en condiciones similares, además de varios esteres derivados [(±)-11111111, , , , (±)-12121212
y [(±)-11113333] (Tabla 28). Debido a la alta estabilidad del derivado octadecil-Sepabeads-CAL-B, el
Lipasa Soporte Tritón X-100 (%)
Octil-agarosa 1
CAL-B Novozym 435 2
Octadecil-Sepabeads 4
0000
10101010
20202020
30303030
40404040
50505050
0000 1111 2222 3333 4444 5555 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Cove
rsi
Cove
rsi
Cove
rsi
Cove
rsi óó óó
nn nn(%
)(%
)(%
)(%
)
0000
10101010
20202020
30303030
40404040
50505050
0000 1111 2222 3333 4444 5555 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Cove
rsi
Cove
rsi
Cove
rsi
Cove
rsi óó óó
nn nn(%
)(%
)(%
)(%
)
0000
10101010
20202020
30303030
40404040
50505050
0000 1111 2222 3333 4444 5555 6666
Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Cove
rsi
Cove
rsi
Cove
rsi
Cove
rsi óó óó
nn nn(%
)(%
)(%
)(%
)
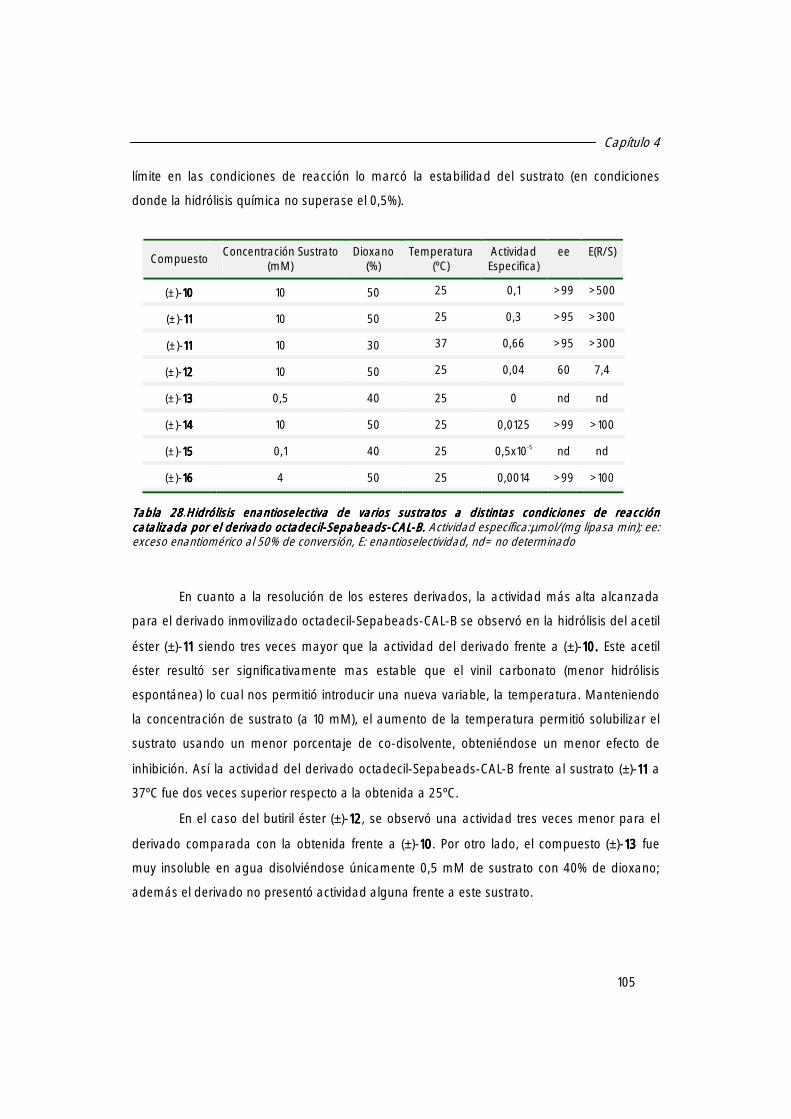
Capítulo 4
105
límite en las condiciones de reacción lo marcó la estabilidad del sustrato (en condiciones
donde la hidrólisis química no superase el 0,5%).
Tabla 28Tabla 28Tabla 28Tabla 28.Hidrólisis enantioselectiva de varios sustratos a distintas condiciones de reacción Hidrólisis enantioselectiva de varios sustratos a distintas condiciones de reacción Hidrólisis enantioselectiva de varios sustratos a distintas condiciones de reacción Hidrólisis enantioselectiva de varios sustratos a distintas condiciones de reacción catalizada por el derivado octadecatalizada por el derivado octadecatalizada por el derivado octadecatalizada por el derivado octadecilcilcilcil----SepabeadsSepabeadsSepabeadsSepabeads----CALCALCALCAL----B. B. B. B. Actividad específica:µmol/(mg lipasa min); ee: exceso enantiomérico al 50% de conversión, E: enantioselectividad, nd= no determinado
En cuanto a la resolución de los esteres derivados, la actividad más alta alcanzada
para el derivado inmovilizado octadecil-Sepabeads-CAL-B se observó en la hidrólisis del acetil
éster (±)-11 11 11 11 siendo tres veces mayor que la actividad del derivado frente a (±)-10. 10. 10. 10. Este acetil
éster resultó ser significativamente mas estable que el vinil carbonato (menor hidrólisis
espontánea) lo cual nos permitió introducir una nueva variable, la temperatura. Manteniendo
la concentración de sustrato (a 10 mM), el aumento de la temperatura permitió solubilizar el
sustrato usando un menor porcentaje de co-disolvente, obteniéndose un menor efecto de
inhibición. Así la actividad del derivado octadecil-Sepabeads-CAL-B frente al sustrato (±)-11 11 11 11 a
37ºC fue dos veces superior respecto a la obtenida a 25ºC.
En el caso del butiril éster (±)-11112222, se observó una actividad tres veces menor para el
derivado comparada con la obtenida frente a (±)-10101010. Por otro lado, el compuesto (±)-13 13 13 13 fue
muy insoluble en agua disolviéndose únicamente 0,5 mM de sustrato con 40% de dioxano;
además el derivado no presentó actividad alguna frente a este sustrato.
Compuesto Concentración Sustrato
(mM) Dioxano
(%) Temperatura
(ºC) Actividad Especifica)
ee E(R/S)
(±)-10101010 10 50 25 0,1 >99 >500
(±)-11111111 10 50 25 0,3 >95 >300
(±)-11111111 10 30 37 0,66 >95 >300
(±)-12121212 10 50 25 0,04 60 7,4
(±)-13131313 0,5 40 25 0 nd nd
(±)-14141414 10 50 25 0,0125 >99 >100
(±)-15151515 0,1 40 25 0,5x10-5 nd nd
(±)-16161616 4 50 25 0,0014 >99 >100

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
106
Referente a los nuevos carbonatos, el derivado octadecil-Sepabeads-CAL-B mostró
mucha menor actividad frente a todos ellos respecto de la obtenida frente al vinil carbonato
(±)-10. 10. 10. 10. De entre todos ellos, el clorometil carbonato (±)-14141414, con una solubilidad similar a la
mostrada por (±)-10101010, determinó la mejor actividad para el derivado inmovilizado octadecil-
Sepabeads-CAL-B, aunque casi 10 veces menor que frente a (±)-10101010. Por su parte el cloroetil
carbonato (±)-16161616 fue algo menos soluble y solo se pudo disolver 4 mM (a 25ºC) empleando la
concentración de dioxano máxima estudiada (50%), mientras el tricloroetil carbonato (±)-15151515 fue
muy insoluble (0,1 mM, 40% de dioxano) además de que el derivado presentó muy poca
actividad frente a este.
A partir de los resultados mostrados en la Tabla 28, se puede concluir que el tamaño
del grupo R del precursor tiene un efecto notable sobre la actividad del catalizador. Se pueden
obtener buenas velocidades de reacción cuando el grupo R es suficientemente pequeño,
como con los esteres derivados 11111111 y 12121212 o como con los carbonatos derivados 10101010 y 14141414.
Respecto a la enantioselectividad en estas reacciones, el derivado inmovilizado
presentó un bajo valor de E en la hidrólisis del compuesto (±)-12121212 mostrando un 60% de exceso
enantiomérico al 50% de conversión.
Por el contrario, usando los otros sustratos, se obtuvo un alto exceso enantiomérico
para el éster (±)-11111111 (ee>95% por 1H-RMN) y para los carbonatos (±)-11114444 y (±)-11116666 (ee>99% por
HPLC quiral).
Los mejores resultados para la resolución enzimática se obtuvieron con el éster (±)-11 11 11 11
(posee el grupo R más pequeño, un metilo).En la hidrólisis de este compuesto, empleando
como catalizador el octadecil-Sepabeads-CAL-B, el 50% de conversión se alcanzó en menos
de 45 minutos usando una relación gramos de catalizador: volumen de reacción 1:37. La
actividad específica y la enantioselectividad del derivado inmovilizado se mantuvieron intactas
durante al menos 10 ciclos de reacción (Figura 27).

Capítulo 4
107
Figura 2Figura 2Figura 2Figura 27777. Ciclos consecutivos de hidrólisis de (±)-11111111 catalizados por el derivado octadecil-Sepabeads- CAL-B. Cada ciclo de reacción duró 45 minutos en llegar al 50% de conversión.
Desafortunadamente, este compuesto no puede ser transformado en la Zopiclona
enantiopura debido a la racemización del centro estereogénico en sucesivos pasos124,129. Sin
embargo, los valores de actividad y enantioselectividad para la resolución del vinil carbonato
(±)-10101010 y del clorometil carbonato (±)-11114444 son suficientemente buenos como para ser utilizados
en la preparación de la (S)-(+)-Zopiclona.
Además, el compuesto (R)-9999,producto de la reacción de hidrólisis enzimática, se
racemiza espontáneamente en las condiciones de trabajo, con lo que, una vez purificado, se
puede emplear directamente para producir más carbonato racémico. De este modo se
aprovecha fácilmente toda la materia prima y se reduce la cantidad de material residual.
0000
10101010
20202020
30303030
40404040
50505050
Conv
ersi
on (%
)Co
nver
sion
(%)
Conv
ersi
on (%
)Co
nver
sion
(%)
Ciclo de reacción Ciclo de reacción Ciclo de reacción Ciclo de reacción
1º 10º0000
10101010
20202020
30303030
40404040
50505050
Conv
ersi
on (%
)Co
nver
sion
(%)
Conv
ersi
on (%
)Co
nver
sion
(%)
Ciclo de reacción Ciclo de reacción Ciclo de reacción Ciclo de reacción
1º 10º0000
10101010
20202020
30303030
40404040
50505050
Conv
ersi
on (%
)Co
nver
sion
(%)
Conv
ersi
on (%
)Co
nver
sion
(%)
Ciclo de reacción Ciclo de reacción Ciclo de reacción Ciclo de reacción
1º 10º

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
108
CONCLUSIONESCONCLUSIONESCONCLUSIONESCONCLUSIONES
El efecto de la inmovilización de la enzima sobre las propiedades catalíticas de
diferentes derivados inmovilizados de CAL-B fue estudiado en la resolución de varios
compuestos, precursores de la (S)-Zopiclona, en medios acuosos. Este trabajo ha demostrado
la fuerte influencia del tipo de derivado inmovilizado y de la naturaleza del sustrato.
El derivado inmovilizado octadecil-Sepabeads-CAL-B fue el más eficiente entre todos
los estudiados, presentando la más alta enantioselectividad (E>500), buena actividad y alta
estabilidad en las condiciones de reacción empleadas, además con la ventaja adicional de la
posible reutilización del soporte.
Empleando este catalizador ha sido posible resolver el vinil carbonato (±)-10101010 y el
clorometil carbonato (±)-11114444 con alta enantioselectividad (E>100), obteniéndose el enantiómero
(S)-(+) sin hidrolizar al 50% de conversión. Además, el acetil éster derivado (±)-11111111 fue el mejor
sustrato para esta enzima dando un exceso enantiomérico superior a 95% analizado por 1H-
RMN (E>300).

Capítulo 4
109
PARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTAL
1. Materiales 1. Materiales 1. Materiales 1. Materiales
La preparación comercial de CAL-B (Novozym 435) fue donada por Novo Nordisk (Dinamarca).
La purificación de la lipasa y preparación de los derivados inmovilizados se realizó como esta
descrito en los capítulos 2 y 3 de la presente tesis.
2222. Síntesis de compuestos. Síntesis de compuestos. Síntesis de compuestos. Síntesis de compuestos
Las rotaciones ópticas se midieron usando un polarímetro Pekin-Elmer 241 (unidades de
10-1 deg cm2 g-1). Los espectros de 1H- y 13C-RMN se obtuvieron empleando un espectrometro
Bruker AC-300 (1H-300 MHz y 13C-75,5 MHz) con TMS (tetrametilsilano) como patrón interno. El
espectro de masas se obtuvo en un Hewlett-Packard 1100 LC/MSD. En todos los casos la
síntesis se monitorizó por cromatografía en placa fina (TLC) usando una mezcla de
hexano:acetato de etilo 1:1 hasta la desaparición del producto de partida. Pasado el tiempo de
reacción, la mezcla de reacción se extrajo con agua y diclorometano. La fase organica se trató
con sulfato sódico para eliminar restos de agua, se filtró y se concentró a sequedad.
((((±±±±))))----6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----7777----oxooxooxooxo----5555----(viniloxicarboniloxi)(viniloxicarboniloxi)(viniloxicarboniloxi)(viniloxicarboniloxi)----5,65,65,65,6----dihidropirrolo[3,4b] pirazina dihidropirrolo[3,4b] pirazina dihidropirrolo[3,4b] pirazina dihidropirrolo[3,4b] pirazina
[(±)-10101010](Vinil carbonato). . . . A una suspensión de 6-(5-cloropirid-2-il)-5-hidroxi-7-oxo-5,6-
dihidropirrolo[3,4b]pirazina [(±)-9999] (1,0 g, 3,81 mmol) en diclorometano seco (8 mL) y piridina
seca (1,2 mL) se le adicionó, bajo atmósfera de N2, cloroformiato de vinilo (0,40 mL, 4,38 mmol,
1,15 eq) a 0ºC. Se agitó a temperatura ambiente durante 10 h. Rendimiento: 75%,polvo blanco,
P.e (ºC)=130-132,IR (KBr): 1770,1736 cm-1, 1H-RMN (CDCl3), δ(ppm): 8,91 (dd, 2H, 2 CH), 8,52 (d,
1H, CH, 3JHH= 8,98 Hz), 8,40 (d, 1H, CH, 4JHH= 2.58 Hz), 8,02 (s, 1H, CH), 7,82 (dd, 1H, CH, 4JHH=
2,58 Hz, 3JHH= 8,98 Hz), 7,20 (dd, 1H, CH, 3Jtrans=6,16 Hz, 2Jgem=13,84 Hz), 4,95 (dd, 1H, CH,
3Jcis=2,30 Hz, 2Jgem=13,84 Hz), 4,69 (dd, 1H, CH, 3Jcis=2,30 Hz, 3Jtrans=6,14 Hz). 13C-RMN (CDCl3):
162,3 (C=O), 153,9 (C=O), 151,0 (C), 148,3 (CH), 148,2 (CH), 147,2 (C), 146,5 (CH), 143,7 (C), 142,2
(CH), 138,1 (CH), 128,3 (C), 115,6 (CH), 98,8 (CH2), 80,3 (CH).EM (ESI+, m/z): 334(M+H)+,
356(M+Na)+

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
110
((((±±±±))))----6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----5555----((((OOOO----acetil)acetil)acetil)acetil)----7777----oxooxooxooxo----5,65,65,65,6----dihidropirrolo[3,4b]pirdihidropirrolo[3,4b]pirdihidropirrolo[3,4b]pirdihidropirrolo[3,4b]pirazina azina azina azina [(±)-
11111111](Acetil ester derivado). A una suspensión de (±)-9999 (600 mg, 2,29 mmol) en diclorometano
seco (10 mL) y piridina seca (0,74 mL) se le adicionó, bajo atmósfera de N2, cloruro de acetilo
(0,33 mL, 4,57 mmol, 2 eq) a 0ºC. Se agitó a temperatura ambiente durante 5 h. Rendimiento:
85%. P.e 170-173ºC; IR (KBr): 1735 cm-1. 1H RMN (CDCl3) δ 8,87 (dd, 2H, 2 CH), 8,51 (d, 1H, CH, 3JHH= 8,70 Hz), 8,37 (d, 1H, CH, 4JHH= 2,56 Hz), 8,13 (s, 1H, CH), 7,80 (dd, 1H, CH, 4JHH= 2,56 Hz,
3JHH= 8,70 Hz), 2,11 (s, 3H, CH3); 13C RMN (CDCl3) δ (ppm): 170,0 (C=O), 163,5 (C), 155,7 (C=O),
149,0 (CH), 148,6 (CH), 148,2 (C),147,4 (CH), 144,6 (C), 138,8 (CH), 129,0 (C), 116,7 (CH), 77,9 (CH),
21,4 (CH3). MS (ESI+) m/z (%): 327 [(M+Na)+, 100 %].
((((±±±±))))----6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----5555----((((OOOO----butirilbutirilbutirilbutiril))))----7777----oxooxooxooxo----5,65,65,65,6----dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina [(±)-
11112222](Butiril ester derivado). A una suspensión de (±)-9999 (600 mg, 2,29 mmol) en diclorometano
seco (10 mL) y piridina seca (0,74 mL) se le adicionó, bajo atmósfera de N2, cloruro de butirilo
(0,47 mL, 4,57 mmol, 2 eq) a 0ºC. Se agitó a temperatura ambiente durante 5 h. Rendimiento:
87%. P.e 139-142ºC; IR (KBr): 1724 cm-1 .1H RMN (CDCl3) δ 8,85 (dd, 2H, 2 CH), 8,49 (d, 1H, CH, 3JHH= 8,76 Hz), 8,33 (d, 1H, CH, 4JHH= 2,55 Hz), 8,13 (s, 1H, CH), 7,78 (dd, 1H, CH, 4JHH= 2,55 Hz,
3JHH= 8,76 Hz), 2,30 (t, 2H, CH2), 1,62 (q, 2H, CH2), 0,97 (t, 3H, CH3); 13C RMN (CDCl3) δ(ppm): 172,6
(C=O), 163,5 (C), 155,8 (C=O), 149,0 (CH), 148,5 (CH), 148,2 (C), 147,3 (CH), 144,6 (C), 138,8 (CH),
129,0 (C), 116,6 (CH), 77,7 (CH), 36,5 (CH2), 18,8 (CH2), 14,1 (CH3). MS (ESI+) m/z (%): 411 [(M+Na)+,
100 %], 355.
((((±±±±))))----6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----5555----((((OOOO----benzoil)benzoil)benzoil)benzoil)----7777----oxooxooxooxo----5,65,65,65,6----dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina dihidropirrolo[3,4b]pirazina [(±)-
11113333](Benzoill ester derivado).A una suspensión de (±)-9999 (500 mg, 1,90 mmol) en diclorometano
seco (10 mL) y piridina seca (0,62 mL) se le adicionó, bajo atmósfera de N2, cloruro de benzoilo
(0,44 mL, 3,81 mmol, 2 eq) a 0ºC. Se agitó a temperatura ambiente durante 4 h. Se trata con
sulfato sódico, se filtra y se concentra a sequedad. Rendimiento: 89%. P.e: 212-216ºC; IR (KBr):
1732 cm-1. 1H RMN (CDCl3) δ 8,86 (dd, 2H, 2 CH), 8,56 (d, 1H, CH, 3JHH= 8,72 Hz), 8,41 (s, 1H, CH),
8,28 (d, 1H, CH, 4JHH= 2,52 Hz), 7,96 (dd, 1H, CH, 4JHH= 2,52 Hz, 3JHH= 8,72 Hz), 7,79 (dd, 2H, 2
CH), 7,55 (d, 1H, CH), 7,42 (t, 2H, 2 CH); 13C RMN (CDCl3) δ (ppm): 166,1 (C=O), 163,6 (C), 155,9

Capítulo 4
111
(C=O), 149,1 (CH), 148,6 (CH), 148,1 (C), 147,5 (CH), 144,7 (C), 138,8 (CH), 134,4 (CH), 130,6 (CH),
129,3 (C), 129,1 (CH), 129,0 (C), 116,5 (CH). MS (ESI+) m/z (%): 389 [(M+Na)+, 60 %], 405 [(M+K)+,
100 %].
((((±±±±))))----5555----(clorometiloxicarboniloxi)(clorometiloxicarboniloxi)(clorometiloxicarboniloxi)(clorometiloxicarboniloxi)----6666----((((5555----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----7777----oxooxooxooxo5,65,65,65,6dihidropirrolodihidropirrolodihidropirrolodihidropirrolo
[3,4b] [3,4b] [3,4b] [3,4b] pirazina pirazina pirazina pirazina [(±)-11114444](clorometil carbonato).A una suspensión de (±)-9999 (1,0 g) en
diclorometano (10 mL) y piridina (1,2 mL) se le adicionó cloroformiato de clorometilo (1,0 ml) a 0
ºC. Se agitó a temperatura ambiente durante 17 h. Rendimiento: 86 %. P.e: 135-137 ºC, IR (cm-1):
1748, 1804, 1H-RMN (CDCl3), δ (ppm): 8,89 (dd, 2H, 2 CH), 8,50 (d, 1H, CH, 2JHH= 8,85 Hz), 8,37 (d,
1H, CH, 3JHH= 8,85 Hz), 7,97 (s, 1H, CH), 7,80 (dd, 1H, CH, 2JHH= 2,52 Hz, 3JHH= 8,88 Hz), 5,81 (dd,
2H, CH2), 13C-RMN (CDCl3), δ (ppm): 163,0 (C=O), 154,5 (C=O), 152,8 (C), 149,1 (CH), 148,0 (C),
147,4 (CH), 144,5 (C), 139,0 (CH), 129,2 (C), 116,3 (CH), 81,6 (CH), 73,2 (CH2), EM-ESI+: [M+Na]=
376,9, [M+H]= 355,0.
6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----7777----oxooxooxooxo----5555----(2,2,2(2,2,2(2,2,2(2,2,2----tricloroetiloxicarboniloxi)tricloroetiloxicarboniloxi)tricloroetiloxicarboniloxi)tricloroetiloxicarboniloxi)----5,65,65,65,6----dihidrodihidrodihidrodihidro
pirrolo [3,4b] pirrolo [3,4b] pirrolo [3,4b] pirrolo [3,4b] pirazinapirazinapirazinapirazina [(±)-11115555]( 2,2,2-tricloroetil carbonato). A una suspensión de (±)-9999 (1,0 g)
en diclorometano (30 mL) y piridina (1,2 mL) se le adicionó cloroformiato de 2,2,2-tricloroetilo
(0,8 mL) a 0 ºC. Se agitó a temperatura ambiente durante 5 h. Rendimiento: 98%. P.e: 201-203
ºC, IR (cm-1): 1788, 1745, 1H-RMN (CDCl3), δ (ppm): 8,91 (dd, 2H, 2 CH), 8,52 (d, 1H, CH, 2JHH= 8,77
Hz), 8,34 (d, 1H, CH,3JHH= 2,31 Hz), 8,03 (s, 1H, CH), 7,82 (dd, 1H, CH, 2JHH= 8,72 Hz, 3JHH= 2,56
Hz), 4,91 (m, 2H, CH2), 13C-RMN (CDCl3), δ (ppm): 162,4 (C=O), 153,9 (C=O) , 152,5 (C), 148,3 (CH),
147,3(C), 146,6 (CH), 143,8 (C), 138,3 (CH), 128,6 (C), 115,8 (CH), 93,8 (C), 80,8 (CH), 76,4 (CH2,
EM-ESI+: [M+Na]= 458,9.
5555----(2(2(2(2----cloroetiloxicarboniloxi)cloroetiloxicarboniloxi)cloroetiloxicarboniloxi)cloroetiloxicarboniloxi)----6666----(5(5(5(5----cloropiridcloropiridcloropiridcloropirid----2222----il)il)il)il)----7777----oxooxooxooxo----5,65,65,65,6----dihidropirrolodihidropirrolodihidropirrolodihidropirrolo
[3,4b]pirazina[3,4b]pirazina[3,4b]pirazina[3,4b]pirazina[(±)-11116666](2-cloroetil carbonato).A una suspensión de (±)-9999 (1,0 g) en diclorometano
(20 mL) y piridina (1,2 mL) se le adiciona cloroformiato de 2-cloroetilo (0,8 ml) a 0 ºC. Se agitó a
temperatura ambiente durante 7 h.Rendimiento: 98%. P.e: 177-178 ºC, IR (cm-1): 1766, 1741, 1H-
RMN (CDCl3), δ (ppm): 8,86 (dd, 2H, 2 CH), 8,52 (d, 1H, CH, 2JHH = 8,98 Hz), 8,39 (d, 1H, CH, 3JHH =
2,58 Hz), 7,99 (s, 1H, CH), 7,82 (dd, 1H, CH, 2JHH = 8,98 Hz, 3JHH = 2,58), 4,53 (m, 2H, CH2), 3,74 (t,

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
112
2H, CH2), 13C-RMN (CDCl3), δ (ppm): 162,8 (C=O), 154,6 (C=O), 153,6 (C), 148,6 (CH), 148,6 (CH),
147,8 (C), 147,1 (CH), 144,3 (C), 138,6 (CH), 128,9 (C), 116,3 (CH), 80,8 (CH), 68,5 (CH2), 41,3 (CH2),
EM-ESI+: [M+Na]= 391,0
3333. Hidrólisis enzimática d. Hidrólisis enzimática d. Hidrólisis enzimática d. Hidrólisis enzimática de carbonatos precursores de la e carbonatos precursores de la e carbonatos precursores de la e carbonatos precursores de la ZZZZopiclona.opiclona.opiclona.opiclona.
Las actividades enzimáticas de los diferentes derivados inmovilizados de CAL-B se
analizaron en la reacción de hidrólisis de los compuestos (±)-10101010----11116666 en diferentes condiciones.
En un primer experimento se disolvió (±)-10101010 en 10 mL de tampón fosfato sódico 10 mM
a pH 7, con un 10 % y un 50% de 1,4-dioxano, hasta una concentración de 0,05 mM,
añadiéndose a esta solución 300 mg de derivado inmovilizado en cada caso.
En segundo lugar se disolvió el sustrato (±)-10101010 hasta una concentración de 10 mM en
3,5 mL de tampón fosfato sódico 10 mM a pH 7 y 25ºC con un 50% de dioxano, añadiéndose
posteriormente 120 mg de derivado enzimático.
La actividad enzimática del derivado octadecil-Sepabeads-CAL-B se determinó en la
hidrólisis de (±)-11111111----16161616 en las siguientes condiciones.
Los esteres derivados (±)-11111111 y 12 12 12 12 se disolvieron hasta una concentración de 10 mM en
8 mL de tampón fosfato sódico 10 mM a pH 7 y 37ºC más un 30% de dioxano y pH 7 y 25ºC
con un 50% de dioxano, respectivamente, añadiéndose posteriormente 200 mg de derivado
inmovilizado en cada caso.
El benzoil ester derivado (±)-11113333 y el carbonato (±)-11115555 se disolvieron a 25ºC en 8 mL de
tampón fosfato sódico 10 mM a pH 7 con un 40% de dioxano hasta una concentración de
sustrato de 0,5 mM y 0,1 mM, respectivamente, añadiéndose a esta solución 100 mg de
derivado enzimático.
Los carbonatos (±)-11114444 y (±)-11116666 se disolvieron a 25ºC en 4 mL de tampón fosfato sódico
10 mM a pH 7 con un 50% de dioxano hasta una concentración de 10 mM y 4 mM
respectivamente, añadiéndose a esta solución 100 mg del derivado enzimático.
Durante la reacción, en todos los casos, el valor del pH se mantuvo constante
utilizando un pHstato (Mettler Toledo DL50 graphic).
El grado de hidrólisis de los diferentes sustratos se cuantificó por HPLC en fase reversa
(Spectra Physic SP 100) acoplado a un detector de UV (Spectra Physic SP 8450) sobre una

Capítulo 4
113
columna Kromasil C18 (25 x 0,4 cm) suministrada por Análisis Vinicos (España). Como fase
móvil se utilizó una mezcla isocrática de acetonitrilo (50%) y 10 mM de tampón fosfato
amónico (50%) ajustando el pH a 7. El flujo fue de 1,5 mL/min. La absorbancia de trabajo fue
de 270 nm. Las muestras se diluyeron hasta una concentración de 1 mM de sustrato. En estas
condiciones los tiempos de retención fueron: (±)-9999 (2,98 min), (±)-10101010 (11,3 min), (±)-11111111 (6,33 min),
(±)-11112222 (14,75 min), (±)-11113333 (18,5 min), (±)-11114444 (11 min), (±)-11115555 (27,5 min), (±)-11116666 (9,9 min).
La actividad específica se definió como µmol de sustrato hidrolizado por minuto por
mg de lipasa inmovilizada y la actividad enzimática como µmol de sustrato hidrolizado por
minuto por gramo de catalizador, calculada a partir de la ecuación descrita en el capítulo 3.
4444. Determinación del exceso enantiomérico y la Enantioselectividad.. Determinación del exceso enantiomérico y la Enantioselectividad.. Determinación del exceso enantiomérico y la Enantioselectividad.. Determinación del exceso enantiomérico y la Enantioselectividad.
Para determinar el exceso enantiomérico del ester sin hidrolizar de (±)-10101010 se recurrió al
análisis por HPLC quiral. La columna empleada fue una Chiracel OD; como fase móvil se utilizó
una mezcla de hexano:etanol:trietilamina (TEA) (40:60:0.1) con un flujo de 0,6 mL/min y 254
nm de absorbancia. Las muestras se diluyeron hasta una concentración de 0,5 mM de
sustrato en diclorometano recuperando la fase orgánica. Los tiempos de retención de los
isómeros R y S fueron 18,41 minutos y 19,85 minutos respectivamente.
El análisis de la pureza óptica de 11111111 se realizó mediante 1H-RMN utilizando como reactivo
de desplazamiento sales de europio(tris[3-(heptafluoropropilhidroximetilen)-4-canforato] de
europio) en CDCl3 tras una conversión del 50%. Para 7 gramos del sustrato se utilizan 4 mg de
la correspondiente sal de europio; el espectro 1H-RMN empleando como disolvente cloroformo
deuterado únicamente mostró la presencia del enantiómero S. Previamente había sido
comprobado el desdoblamiento del acetato racémico en tales proporciones. Por lo tanto la
pureza enantiomérica del acetato obtenido tras la hidrólisis fue mayor del 96%.
Para los cloro carbonatos (±)-11114444 y (±)-11116666 el exceso enantiomérico del ester sin hidrolizar se
analizo por HPLC quiral empleando una columna CHIRALPAK AS (4,6x250 mm). Para (±)-11114444 se
empleó una fase móvil de 60% hexano, 35% etanol y 5% isopropanol, a un flujo de 1 mL/min y
254 nm. Los tiempos de retención fueron: S-(+)-11114444 (10,34 min), R-(-)-11114444 (15,28 min). En el caso

Hidrólisis enantioselectiva de precursores de(S)-(+)-Zopiclona
114
del sustrato (±)-11116666 la fase móvil fue una mezcla de 60% hexano,40% isopropanol a un flujo de
0,8 mL/min y 254 nm. Los tiempos de retención fueron: (S)-(+)-11116666 (26 min),(R)-(-)-11116 6 6 6 (32,8).
La enantioselectividad se expresó como el valor E calculado a partir del exceso
enantiomérico del éster sin hidrolizar y el grado de conversión de acuerdo a la ecuación
previamente descrita por Chen y col73.

Capítulo 5
115
INTRODUCCIONINTRODUCCIONINTRODUCCIONINTRODUCCION
1. Derivados de 41. Derivados de 41. Derivados de 41. Derivados de 4----arilpiperidinas. Síntesis de la (arilpiperidinas. Síntesis de la (arilpiperidinas. Síntesis de la (arilpiperidinas. Síntesis de la (----))))----ParoxetinaParoxetinaParoxetinaParoxetina
El grupo 4-arilpiperidina es un importante elemento estructural en un gran número de
compuestos biológicamente activos, posiblemente debido a su similitud con el farmacóforo
aril-alquilamina común para los neurotransmisores como la serotonina [5-hidroxi-triptamina
(5-HT)] o la dopamina (DA).
Los fármacos que modulan las acciones fisiológicas y fisiopatológicas del 5-HT son útiles
o potencialmente útiles en el tratamiento de una gran variedad de enfermedades humanas,
como la depresión, la ansiedad, el alcoholismo, el pánico crónico y trastornos alimenticios130.
Ejemplo de ellos son la Loperamida y el Trefentanil, que se emplean como analgésico, la
Anileridina como relajante muscular y la hormona del crecimiento MK0677 131(Esquema 28).
Esquema 2Esquema 2Esquema 2Esquema 28888
Otros de gran importancia son el Haloperidiol, la Meperidina y la Paroxetina (Paxil®,
Seroxat®) (Esquema 29)132.
La Paroxetina (11117777), (±)-4-(4´-fluorofenil)-3-[3,4-(metilendioxi)-fenoximetil] piperidina, es
un potente y selectivo inhibidor de la serotonina (5-HT) perteneciente a la familia de las
fenilpiperidinas cuyo enantiómero quiral (3S,4R) presenta la totalidad de la actividad
farmacológica, siendo responsable de los efectos antidepresivos. Debido a su importancia
biológica, varias estratégias de síntesis han sido desarrolladas para la preparación de este
Trefentanil Loperamida
AnileridinaMKD677
N
O
N
OF
N NN
O
N
N
OH
Cl
O
NHO
NH2
O
N
N
SO
O
NH2
N
OO
Trefentanil Loperamida
AnileridinaMKD677
N
O
N
OF
N NN
O
N
N
OH
Cl
O
NHO
NH2
O
N
N
SO
O
NH2
N
OO
Trefentanil Loperamida
AnileridinaMKD677
N
O
N
OF
N NN
O
N
N
OH
Cl
O
NHO
NH2
O
N
N
SO
O
NH2
N
OO
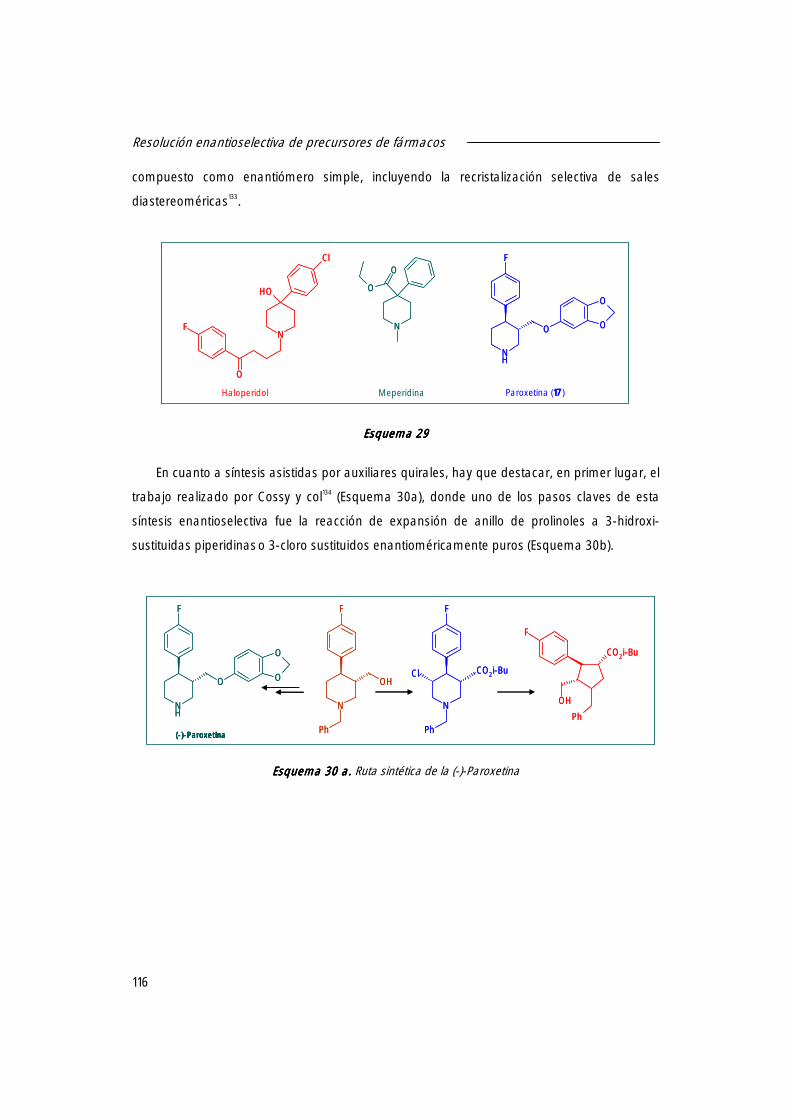
Resolución enantioselectiva de precursores de fármacos
116
compuesto como enantiómero simple, incluyendo la recristalización selectiva de sales
diastereoméricas133.
Esquema 2Esquema 2Esquema 2Esquema 29999
En cuanto a síntesis asistidas por auxiliares quirales, hay que destacar, en primer lugar, el
trabajo realizado por Cossy y col134 (Esquema 30a), donde uno de los pasos claves de esta
síntesis enantioselectiva fue la reacción de expansión de anillo de prolinoles a 3-hidroxi-
sustituidas piperidinas o 3-cloro sustituidos enantioméricamente puros (Esquema 30b).
Esquema Esquema Esquema Esquema 30303030 a a a a. . . . Ruta sintética de la (-)-Paroxetina
NH
F
O
O
O
F
OH
N
Ph
CO2i-Bu
F
N
Ph
Cl
F
CO2i-Bu
OH
Ph
((((---- ))))----ParoxetinaParoxetinaParoxetinaParoxetina
NH
F
O
O
O
F
OH
N
Ph
CO2i-Bu
F
N
Ph
Cl
F
CO2i-Bu
OH
Ph
((((---- ))))----ParoxetinaParoxetinaParoxetinaParoxetina
O
N
OH
F
Cl
Haloperidol
O
O
N
Meperidina
NH
F
O
O
O
Paroxetina (17171717)
O
N
OH
F
Cl
Haloperidol
O
O
N
Meperidina
NH
F
O
O
O
Paroxetina (17171717)Paroxetina (17171717)

Capítulo 5
117
Esquema Esquema Esquema Esquema 30303030 b b b b. Reacciones de expansión de anillo
En segundo lugar, Liu y col 133a propusieron una síntesis asistida por auxiliares quirales
diastereoselectivos partiendo de 2-piperidinonas quirales 4-sustituidas y trans-3,4-di-
sustituidas, obtenidas a partir de anhídridos glutáricos 3-sustituidos y (S)-metilbencilamina
(Esquema 31). Esta nueva metodología puede ser usada para la síntesis de 4-aril-2-
piperidinonas quirales, 2-piperidinonas y trans-3,4- piperidinas di-sustitudas. La metodología
desarrollada fue aplicada para la síntesis de Paroxetina homoquiral.
Esquema Esquema Esquema Esquema 31313131
Otras metodologías desarrolladas fueron la resolución biocatalítica133c y la
asimetrización de un intermedio diéster proquiral135.
Además, existen algunos procedimientos para la obtención de la Paroxetina a partir
del compuesto 1 1 1 19999, (±)-trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxilato de etilo, (Esquema
32)136, implicando una etapa de resolución. En la mayoría de los procesos descritos, como en
las patentes US 4007196 y EP 812827, se realiza la resolución utilizando métodos químicos que
emplean agentes de resolución costosos y condiciones contaminantes.
N
R*
R R*
OH
R
Cl
R*
RMsClEt3N
1) (CF3CO)2O2) 2Et3N3) NaOH
N
R*
R R*
OH
R
Cl
R*
RMsClEt3N
1) (CF3CO)2O2) 2Et3N3) NaOH
N
R*
R R*
OH
R
Cl
R*
RMsClEt3N
1) (CF3CO)2O2) 2Et3N3) NaOH
N
R1
R2
X
R3
N
R1
O
R3(X=O,H2)
R1
O OH
O
NHR*
O OO
R1
piperidina (X=H,H) y 2-piperidinona (X=O) trans-3,4-disustituidos
N
R1
R2
X
R3
N
R1
O
R3(X=O,H2)
R1
O OH
O
NHR*
O OO
R1
piperidina (X=H,H) y 2-piperidinona (X=O) trans-3,4-disustituidos
N
R1
R2
X
R3
N
R1
O
R3(X=O,H2)
R1
O OH
O
NHR*
O OO
R1
piperidina (X=H,H) y 2-piperidinona (X=O) trans-3,4-disustituidos
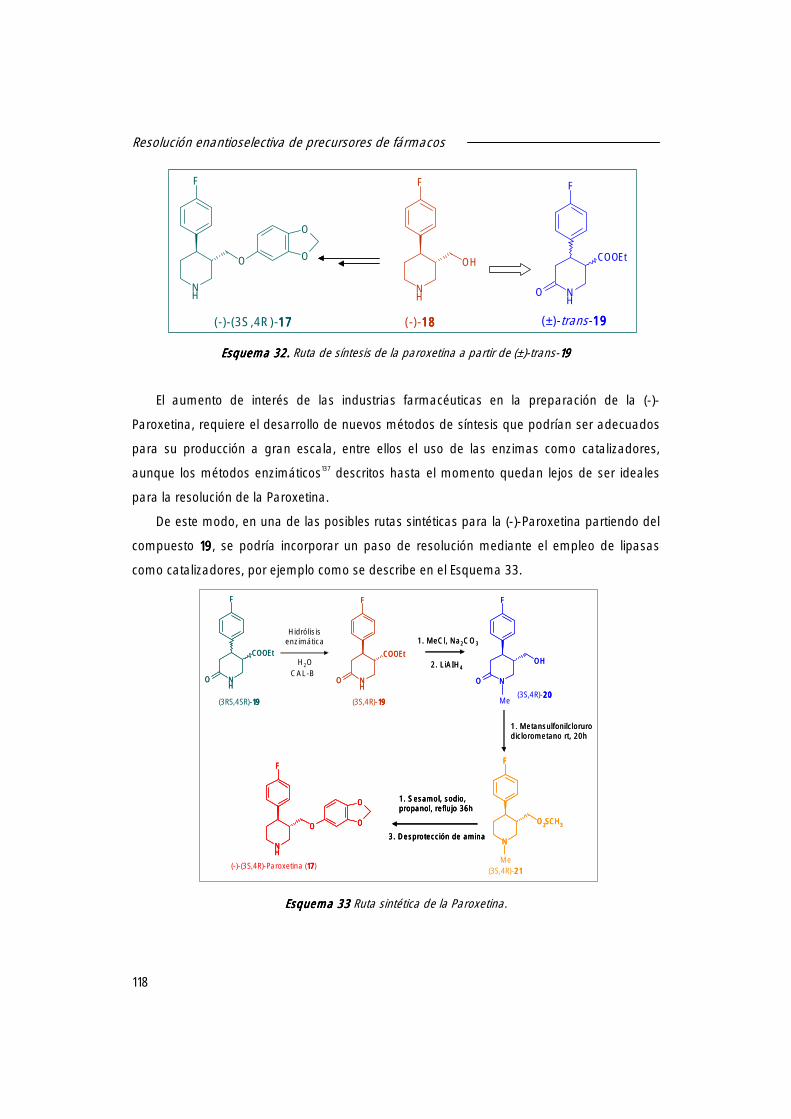
Resolución enantioselectiva de precursores de fármacos
118
Esquema Esquema Esquema Esquema 32323232.... Ruta de síntesis de la paroxetina a partir de (±)-trans-19191919
El aumento de interés de las industrias farmacéuticas en la preparación de la (-)-
Paroxetina, requiere el desarrollo de nuevos métodos de síntesis que podrían ser adecuados
para su producción a gran escala, entre ellos el uso de las enzimas como catalizadores,
aunque los métodos enzimáticos137 descritos hasta el momento quedan lejos de ser ideales
para la resolución de la Paroxetina.
De este modo, en una de las posibles rutas sintéticas para la (-)-Paroxetina partiendo del
compuesto 19191919, se podría incorporar un paso de resolución mediante el empleo de lipasas
como catalizadores, por ejemplo como se describe en el Esquema 33.
Esquema Esquema Esquema Esquema 33333333 Ruta sintética de la Paroxetina.
NH
F
O
O
O
NH
F
OH
NH
O
F
COOEt
(±)-trans-1 91 91 91 9(-)-(3S ,4R )-1 71 71 71 7 (-)-1 81 81 81 8
NH
O
F
COOEt
(3RS,4SR)-19191919
NH
O
F
COOEt
(3RS,4SR)-19191919
NH
O
F
COOEt
(3S,4R)-19191919
NH
O
F
COOEt
(3S,4R)-19191919
NO
F
OH
Me(3S,4R)-20202020
NO
F
OH
Me(3S,4R)-20202020
C AL -B
Hidrólisisenzimática
H2O
1 . MeC l, Na2C O3
2. L iAIH4
1 . MeC l, Na2C O3
2. L iAIH4
1 . Metansulfonilclorurodiclorometano rt, 20h1 . Metansulfonilclorurodiclorometano rt, 20h
N
F
O2SCH3
Me(3S,4R)-21212121(-)-(3S,4R)-Paroxetina (17171717)
NH
F
O
O
O
1 . S esamol, sodio,propanol, reflujo 36h
3. Desprotección de aminaN
F
O2SCH3
Me(3S,4R)-21212121
N
F
O2SCH3
Me(3S,4R)-21212121(-)-(3S,4R)-Paroxetina (17171717)
NH
F
O
O
O
(-)-(3S,4R)-Paroxetina (17171717)
NH
F
O
O
O
1 . S esamol, sodio,propanol, reflujo 36h
3. Desprotección de amina
1 . S esamol, sodio,propanol, reflujo 36h
3. Desprotección de amina

Capítulo 5
119
2. Ácido 22. Ácido 22. Ácido 22. Ácido 2----hidroxihidroxihidroxihidroxi----4444----fenilbutanoico, precursor en la síntesis del (S)fenilbutanoico, precursor en la síntesis del (S)fenilbutanoico, precursor en la síntesis del (S)fenilbutanoico, precursor en la síntesis del (S)----Enalapril.Enalapril.Enalapril.Enalapril.
Los inhibidores de la enzima que transforma la angiotensina (ACE) son una familia de
péptidos de importante relevancia en el control de la hipertensión y en el tratamiento de
deficiencias cardíacas. La mayoría de ellos poseen un elemento estructural común, un grupo
N-sustituido (S)-2-amino-4-fenilbutirato, como sucede en los agentes terapéuticos Enalapril,
Quinapril, Trandolapril y Moexipril (Esquema 34) 138-139.
Esquema Esquema Esquema Esquema 34343434.... Estructura química de algunos inhibidores ACE
En particular, el Enalapril, (S)-1-[N-[1-etoxicarbonil-3-fenilpropil]-L-alanil]-L-prolina (22222222) es
una prodroga; éster etílico de la Enalaprilat (S)-1-[N-[1-carboxi-3-fenilpropil]-L-alanil]-L-prolina).
Este tipo de fármacos derivados del ácido S-2-hidroxi-4-fenilbutanoico presentan un menor
poder de inhibición que los obtenidos a partir del isómero R-2-hidroxi-4-fenilbutanoico (R-4444),
por tanto será necesario desarrollar una técnica eficaz para la resolución de dicha mezcla
racémica (Esquema 35)
EEEEsquema 3squema 3squema 3squema 35555. Ruta sintética del (S)-Enalapril
NH
O
N COOH
COOEt
Enalapril (22222222)
N
NH
OCOOEt
COOH
Quinapril
NH
OCOOEt
H
H COOH
Trandolapril
N
NH
OCOOEt
COOHCH3O
CH3O
Moexipril
NH
O
N COOH
COOEt
Enalapril (22222222)
N
NH
OCOOEt
COOH
Quinapril
NH
OCOOEt
H
H COOH
Trandolapril
N
NH
OCOOEt
COOHCH3O
CH3O
Moexipril
NH
O
N COOH
COOEt
Enalapril (22222222)
N
NH
OCOOEt
COOH
Quinapril
NH
OCOOEt
H
H COOH
Trandolapril
N
NH
OCOOEt
COOHCH3O
CH3O
Moexipril
COOEt
OH OH
COOH NH
O
N COOH
COOEt
(S)-22222222(±)-2222 (R)-4444
COOEt
OH OH
COOH NH
O
N COOH
COOEt
(S)-22222222(±)-2222 (R)-4444

Resolución enantioselectiva de precursores de fármacos
120
En la literatura se han descrito varias metodologías de resolución quiral que conducen
a la obtención del ácido 2-hidroxi-4-fenilbutanoico o derivados mediante recristalización,
hidrólisis y reesterificación140. En otros casos se ha empleado la reducción estereoselectiva con
S-Alpine-borano del 2-oxo-4-fenilbutirato de etilo141.
Posteriormente fueron descritos métodos de resolución enzimática, los cuales
presentan una serie de ventajas respecto a los tradicionales métodos químicos. Así, Wong y
col142 resolvieron la mezcla racémica del 2-hidroxi-4-fenilbutanonitrilo empleando la lipasa de
Ps. Fluorescens (PFL), mientras otros autores llevaron a cabo la resolución del éster etílico del
ácido 2-hidroxi-4-fenilbutanoico [(±)-2222] empleando la misma lipasa143.Sin embargo, en todos
los casos se utilizó enzima soluble, lo cual desde un punto de vista industrial resulta menos
ventajoso frente al empleo de catalizadores inmovilizados. Así Baldano y col144 consiguieron
obtener el S-2222 empleando como catalizador la Penicilina G aminohidrolasa inmovilizada.

Capítulo 5
121
OBJETIVOSOBJETIVOSOBJETIVOSOBJETIVOS
El objetivo de este capítulo se centra en la aplicación de distintos derivados
inmovilizados de diferentes lipasas en la resolución quiral de sustratos precursores de
distintos fármacos mediante reacción de hidrólisis en medios acuosos. Por un lado, se
emplearán derivados inmovilizados de CAL-B en la hidrólisis del éster etílico del ácido
(3RS,4RS)-trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxílico, intermedio en la síntesis de la
(3S,4R)-(-)-Paroxetina. Por otro lado, se estudiará la hidrólisis enantioselectiva del 2-hidroxi-4-
fenilbutirato de etilo como intermedio en la síntesis de S-Enalapril catalizada por derivados
inmovilizados de la lipasa de B. thermocatenulatus (BTL2).

Resolución enantioselectiva de precursores de fármacos
122
RESULTADOSRESULTADOSRESULTADOSRESULTADOS Y DISCUSIÓN Y DISCUSIÓN Y DISCUSIÓN Y DISCUSIÓN
1. Resolución enantioselectiva de 1. Resolución enantioselectiva de 1. Resolución enantioselectiva de 1. Resolución enantioselectiva de (±)(±)(±)(±)----transtranstranstrans----4444----(4´(4´(4´(4´----flufluflufluorofenil)orofenil)orofenil)orofenil)----6666----oxopiperidinoxopiperidinoxopiperidinoxopiperidin----3333----carboxilato carboxilato carboxilato carboxilato
de etilo [(±)]de etilo [(±)]de etilo [(±)]de etilo [(±)]----19,19,19,19, intermedio en la síntesis de la (intermedio en la síntesis de la (intermedio en la síntesis de la (intermedio en la síntesis de la (----))))----Paroxetina, Paroxetina, Paroxetina, Paroxetina, catalizada por dcatalizada por dcatalizada por dcatalizada por derivados erivados erivados erivados
inmovilizados de CALinmovilizados de CALinmovilizados de CALinmovilizados de CAL----B.B.B.B.
Otro de los procesos de interés industrial estudiados en esta Tesis fue la resolución del
intermedio de la Paroxetina, (±)-trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxilato de etilo,
empleado en la ruta sintética del isómero enantiomericamente puro (3S,4R), mediante
hidrólisis enantioselectiva catalizada por lipasas. En este caso el objetivo principal consistió en
determinar las condiciones óptimas de esta resolución.
Esquema 3Esquema 3Esquema 3Esquema 36666. . . . Hidrólisis enantioselectiva de (±)-trans-19 19 19 19 catalizada por diferentes derivados inmovilizados de CAL-B.
1.11.11.11.1 Actividad específica de diferentes lipasas de origeActividad específica de diferentes lipasas de origeActividad específica de diferentes lipasas de origeActividad específica de diferentes lipasas de origen microbiano frente a (n microbiano frente a (n microbiano frente a (n microbiano frente a (±±±±))))----trans-19191919
En primer lugar, se realizó un chequeo de la actividad de distintas lipasas con el fin de
encontrar la lipasa con las mejores condiciones para llevar a cabo esta resolución. Para ello se
utilizaron varias lipasas, lipasa de Pseudomonas fluorescens (PFL), Humicola lanuginosa (HLL),
Candida rugosa (CRL) y Candida antarctica (fracción B) (CAL-B), inmovilizadas sobre gel octil-
agarosa, examinándose su actividad específica en la reacción de hidrólisis del sustrato (±)-
trans-19191919 (Esquema 36).
NH
O
F
COOEt
NH
O
F
COOH
NH
O
F
COOEt+
(3RS,4RS)-(±)-19191919 (3R,4S)-(+)-23232323 (3S,4R)-(-)-19191919
Derivados CAL-B
H2ONH
O
F
COOEt
NH
O
F
COOH
NH
O
F
COOEt+
(3RS,4RS)-(±)-19191919 (3R,4S)-(+)-23232323 (3S,4R)-(-)-19191919
Derivados CAL-B
H2ONH
O
F
COOEt
NH
O
F
COOH
NH
O
F
COOEt+
(3RS,4RS)-(±)-19191919 (3R,4S)-(+)-23232323 (3S,4R)-(-)-19191919
Derivados CAL-B
H2O

Capítulo 5
123
Las actividades específicas de las distintas lipasas inmovilizadas fueron muy bajas (Tabla
29), obteniéndose el valor más alto en actividad cuando se empleó la CAL-B (entre 9 y 30
veces más alta que con las demás lipasas), la cual fue escogida para realizar los siguientes
estudios.
Tabla 29Tabla 29Tabla 29Tabla 29. Actividad específicaActividad específicaActividad específicaActividad específica de los d de los d de los d de los derivados inmovilizados octilerivados inmovilizados octilerivados inmovilizados octilerivados inmovilizados octil----agarosa de las diferentes agarosa de las diferentes agarosa de las diferentes agarosa de las diferentes lipasas lipasas lipasas lipasas en la reacción de hidrólisis de 2 mM de (±)-trans 19 19 19 19 a pH 7 y 45ºC.
1.2 Actividad e1.2 Actividad e1.2 Actividad específicaspecíficaspecífica de los distintos de de los distintos de de los distintos de de los distintos derivados inmovilizados de CALrivados inmovilizados de CALrivados inmovilizados de CALrivados inmovilizados de CAL----B. B. B. B.
Una vez seleccionada la enzima, ésta fue inmovilizada sobre distintos soportes
(glioxil-agarosa, glutaradehído-agarosa, octadecil-Sepabeads), obteniéndose derivados
inmovilizados con una carga enzimática de 12 mg de lipasa/mL. El objetivo fue estudiar el
efecto de la inmovilización sobre la actividad del enzima, además de determinar unas
condiciones de reacción óptimas tanto para el sustrato como para los derivados
inmovilizados.
De este modo, en primer lugar, se analizó la actividad específica de los distintos
derivados inmovilizados de CAL-B en la hidrólisis de trans-11119999 en medios acuosos en
condiciones suaves (25ºC, 2 mM, pH 7). Los derivados glutaraldehído-CAL-B y glioxil-CAL-B
presentaron las actividades específicas más altas, 5 veces más que la actividad del derivado
octadecil-Sepabeads-CAL-B (Tabla 30).
Lipasa Actividad específica
Pseudomonas fluorescens (PFL) 0,0027
Candida antartica (CAL-B) 0,0257
Candida rugosa (CRL) 0,0009
Humicola lanuginosa (HLL) 0,0082

Resolución enantioselectiva de precursores de fármacos
124
Tabla 30Tabla 30Tabla 30Tabla 30. Actividad específica Actividad específica Actividad específica Actividad específica de los distintos derivados de los distintos derivados de los distintos derivados de los distintos derivados inmovilizados de CALinmovilizados de CALinmovilizados de CALinmovilizados de CAL----B B B B catalizando la hidrólisis de (±)-trans-19 19 19 19 a 2 mM a diferentes temperaturas. La concentración de lipasa en cada derivado inmovilizado fue de 12 mg/mL. Actividad especifica (µmol. mglip
-
1 .h-1).
1.2.1 Efecto de la temperatura sobre la actividad de CAL-B.
El incrementó en la temperatura, de 25ºC a 45ºC (Tabla 30), provocó un aumento en
el valor de la actividad para todos los derivados de CAL-B, siendo más importante en el
derivado octadecil-Sepabeads-CAL-B, donde la enzima aumentó su actividad 30 veces.
1.3 1.3 1.3 1.3 BúsquBúsquBúsquBúsquedaedaedaeda de las mejores condiciones de reacción. de las mejores condiciones de reacción. de las mejores condiciones de reacción. de las mejores condiciones de reacción.
Para llevar a cabo una reacción viable industrialmente, es necesario el empleo de
altas concentraciones de sustrato (ej. 50 mM). Debido a la baja solubilidad de este sustrato en
medio acuoso (limitado a 2 mM a 25ºC), además de emplear una mayor temperatura (45ºC),
fue necesario el empleo de co-disolventes para poder solubilizar concentraciones más
elevadas del mismo. Así, con el fin de optimizar la concentración de co-disolvente necesaria
en el medio de reacción para solubilizar hasta 100 mM del compuesto (±)-trans-19191919, se
emplearon varios disolventes miscibles en agua con diferente grado de hidrofobicidad (Figura
28). Los mejores resultados de solubilidad se encontraron para el acetonitrilo y el 1,4-dioxano.
Figura 2Figura 2Figura 2Figura 28888. Solubilidad de (±)Solubilidad de (±)Solubilidad de (±)Solubilidad de (±)----trans trans trans trans 19 en varios disolventes.19 en varios disolventes.19 en varios disolventes.19 en varios disolventes. Acetonitrilo (�), DMF (�), diglime (�), dioxano (` ), cellosolve (�).
Derivado inmovilizado Actividad
25ºC Actividad 45ºC
Glutaraldehído-CAL-B 0,005 0,057
Glioxil-CAL-B 0,0065 0,066
Octadecil-Sepabeads-CAL-B 0,001 0,030
20202020
40404040
60606060
80808080
100100100100
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
Sust
rato
sol
uble
(Su
stra
to s
olub
le (
Sust
rato
sol
uble
(Su
stra
to s
olub
le ( m
Mm
Mm
Mm
M)) ))
20202020
40404040
60606060
80808080
100100100100
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
Sust
rato
sol
uble
(Su
stra
to s
olub
le (
Sust
rato
sol
uble
(Su
stra
to s
olub
le ( m
Mm
Mm
Mm
M)) ))
20202020
40404040
60606060
80808080
100100100100
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
Sust
rato
sol
uble
(Su
stra
to s
olub
le (
Sust
rato
sol
uble
(Su
stra
to s
olub
le ( m
Mm
Mm
Mm
M)) ))
20202020
40404040
60606060
80808080
100100100100
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
20202020
40404040
60606060
80808080
100100100100
20202020
40404040
60606060
80808080
100100100100
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
000010101010 20202020 30303030
CodisolventeCodisolventeCodisolventeCodisolvente (%)(%)(%)(%)
Sust
rato
sol
uble
(Su
stra
to s
olub
le (
Sust
rato
sol
uble
(Su
stra
to s
olub
le ( m
Mm
Mm
Mm
M)) ))
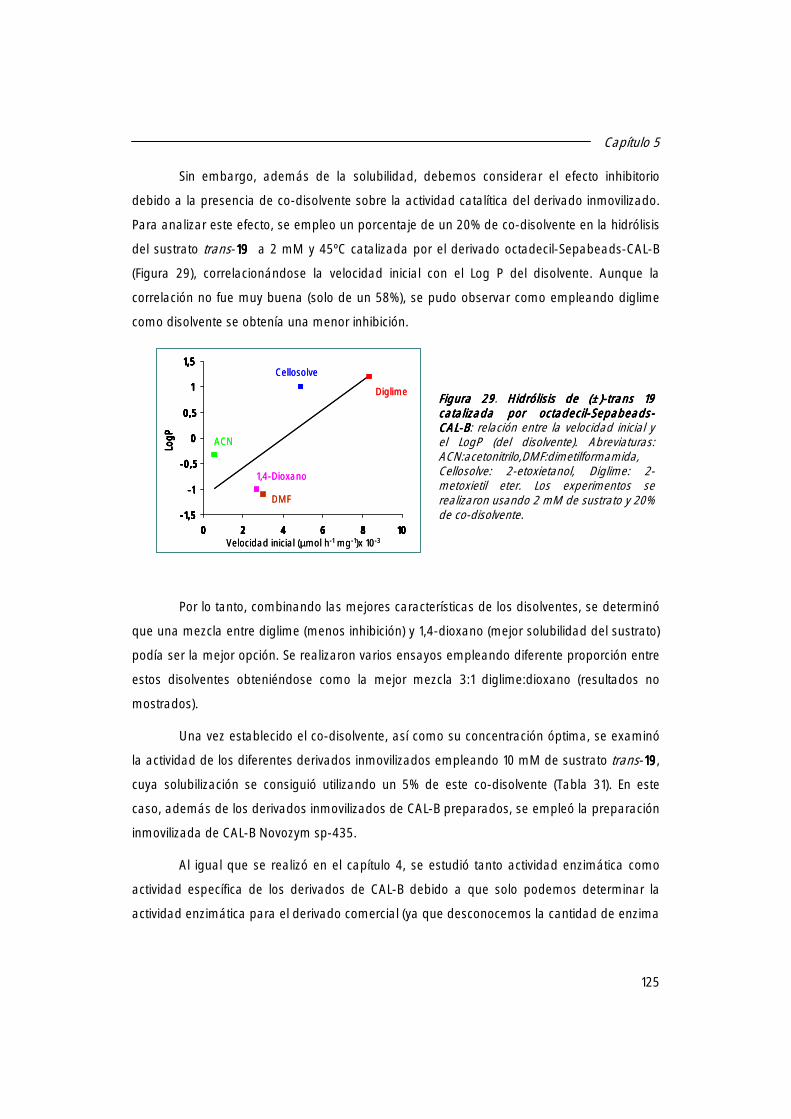
Capítulo 5
125
Sin embargo, además de la solubilidad, debemos considerar el efecto inhibitorio
debido a la presencia de co-disolvente sobre la actividad catalítica del derivado inmovilizado.
Para analizar este efecto, se empleo un porcentaje de un 20% de co-disolvente en la hidrólisis
del sustrato trans-19191919 a 2 mM y 45ºC catalizada por el derivado octadecil-Sepabeads-CAL-B
(Figura 29), correlacionándose la velocidad inicial con el Log P del disolvente. Aunque la
correlación no fue muy buena (solo de un 58%), se pudo observar como empleando diglime
como disolvente se obtenía una menor inhibición.
Figura Figura Figura Figura 29292929. Hidrólisis de (±)Hidrólisis de (±)Hidrólisis de (±)Hidrólisis de (±)----trans 19 trans 19 trans 19 trans 19 catalizada por octadecilcatalizada por octadecilcatalizada por octadecilcatalizada por octadecil----SepabeadsSepabeadsSepabeadsSepabeads----CALCALCALCAL----BBBB: relación entre la velocidad inicial y el LogP (del disolvente). Abreviaturas: ACN:acetonitrilo,DMF:dimetilformamida, Cellosolve: 2-etoxietanol, Diglime: 2-metoxietil eter. Los experimentos se realizaron usando 2 mM de sustrato y 20% de co-disolvente.
Por lo tanto, combinando las mejores características de los disolventes, se determinó
que una mezcla entre diglime (menos inhibición) y 1,4-dioxano (mejor solubilidad del sustrato)
podía ser la mejor opción. Se realizaron varios ensayos empleando diferente proporción entre
estos disolventes obteniéndose como la mejor mezcla 3:1 diglime:dioxano (resultados no
mostrados).
Una vez establecido el co-disolvente, así como su concentración óptima, se examinó
la actividad de los diferentes derivados inmovilizados empleando 10 mM de sustrato trans-19191919,
cuya solubilización se consiguió utilizando un 5% de este co-disolvente (Tabla 31). En este
caso, además de los derivados inmovilizados de CAL-B preparados, se empleó la preparación
inmovilizada de CAL-B Novozym sp-435.
Al igual que se realizó en el capítulo 4, se estudió tanto actividad enzimática como
actividad específica de los derivados de CAL-B debido a que solo podemos determinar la
actividad enzimática para el derivado comercial (ya que desconocemos la cantidad de enzima
ACN
Diglime
Cellosolve
1,4-Dioxano
DMF
----1,51,51,51,5
----1111
----0,50,50,50,5
0000
0,50,50,50,5
1111
1,51,51,51,5
0000 2222 4444 6666 8888 10101010
LogP
LogP
LogP
LogP
Velocidad inicial (µmol h-1 mg-1)x 10-3
ACN
Diglime
Cellosolve
1,4-Dioxano
DMF
----1,51,51,51,5
----1111
----0,50,50,50,5
0000
0,50,50,50,5
1111
1,51,51,51,5
----1,51,51,51,5
----1111
----0,50,50,50,5
0000
0,50,50,50,5
1111
1,51,51,51,5
0000 2222 4444 6666 8888 101010100000 2222 4444 6666 8888 10101010
LogP
LogP
LogP
LogP
Velocidad inicial (µmol h-1 mg-1)x 10-3
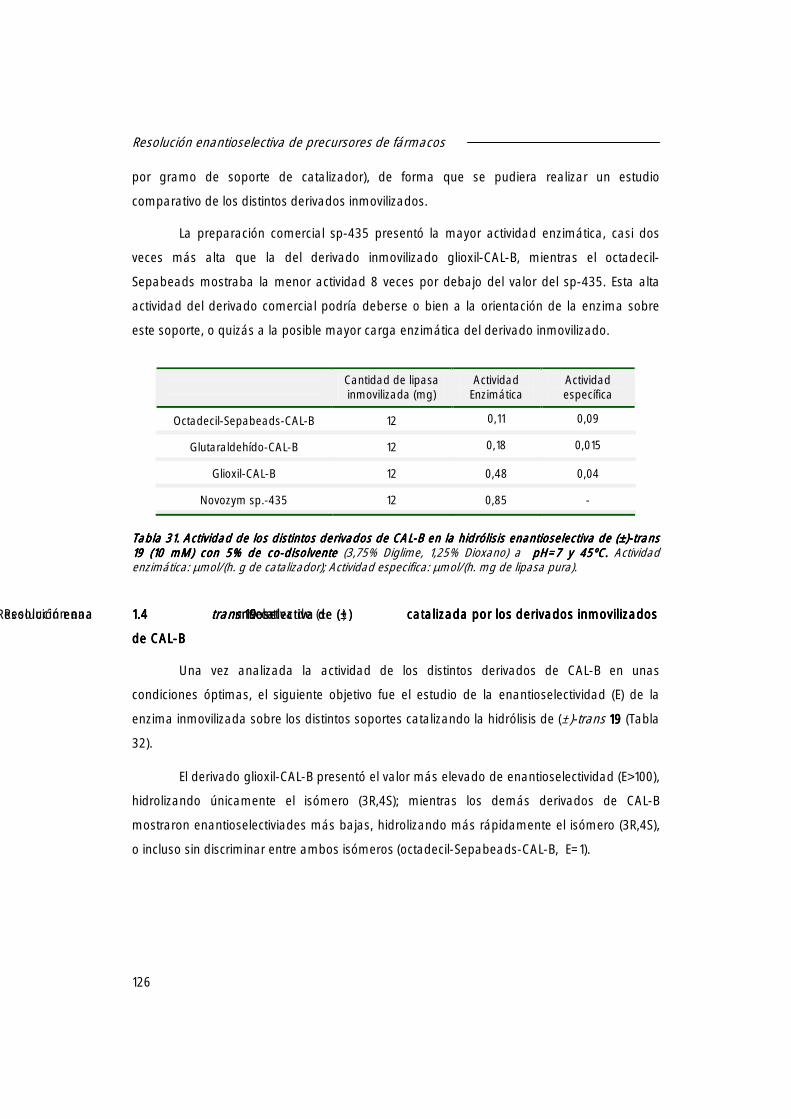
Resolución enantioselectiva de precursores de fármacos
126
por gramo de soporte de catalizador), de forma que se pudiera realizar un estudio
comparativo de los distintos derivados inmovilizados.
La preparación comercial sp-435 presentó la mayor actividad enzimática, casi dos
veces más alta que la del derivado inmovilizado glioxil-CAL-B, mientras el octadecil-
Sepabeads mostraba la menor actividad 8 veces por debajo del valor del sp-435. Esta alta
actividad del derivado comercial podría deberse o bien a la orientación de la enzima sobre
este soporte, o quizás a la posible mayor carga enzimática del derivado inmovilizado.
Tabla 31.Tabla 31.Tabla 31.Tabla 31. Actividad de los distintos derivados deActividad de los distintos derivados deActividad de los distintos derivados deActividad de los distintos derivados de CAL CAL CAL CAL----B en la hB en la hB en la hB en la hidrólisis enantioselectiva de (idrólisis enantioselectiva de (idrólisis enantioselectiva de (idrólisis enantioselectiva de (±±±±))))----trans trans trans trans 19 (10 mM) con 5% de co19 (10 mM) con 5% de co19 (10 mM) con 5% de co19 (10 mM) con 5% de co----disolventedisolventedisolventedisolvente (3,75% Diglime, 1,25% Dioxano) a pH=7 y 45ºC.pH=7 y 45ºC.pH=7 y 45ºC.pH=7 y 45ºC. Actividad enzimática: µmol/(h. g de catalizador); Actividad especifica: µmol/(h. mg de lipasa pura).
1.1.1.1.4444 enaResolución en aResolución ena nseletiva de (± )n tioselectiva de (±)ntioselectiva de (±)transtranstrans 1 1 1 19999 cat alizada por los derivados catalizada por los derivados catalizada por los derivados catalizada por los derivados inmovilizados inmovilizados inmovilizados inmovilizados
de CALde CALde CALde CAL----B B B B
Una vez analizada la actividad de los distintos derivados de CAL-B en unas
condiciones óptimas, el siguiente objetivo fue el estudio de la enantioselectividad (E) de la
enzima inmovilizada sobre los distintos soportes catalizando la hidrólisis de (±)-trans 19 19 19 19 (Tabla
32).
El derivado glioxil-CAL-B presentó el valor más elevado de enantioselectividad (E>100),
hidrolizando únicamente el isómero (3R,4S); mientras los demás derivados de CAL-B
mostraron enantioselectiviades más bajas, hidrolizando más rápidamente el isómero (3R,4S),
o incluso sin discriminar entre ambos isómeros (octadecil-Sepabeads-CAL-B, E=1).
Cantidad de lipasa inmovilizada (mg)
Actividad Enzimática
Actividad específica
Octadecil-Sepabeads-CAL-B 12 0,11 0,09
Glutaraldehído-CAL-B 12 0,18 0,015
Glioxil-CAL-B 12 0,48 0,04
Novozym sp.-435 12 0,85 -
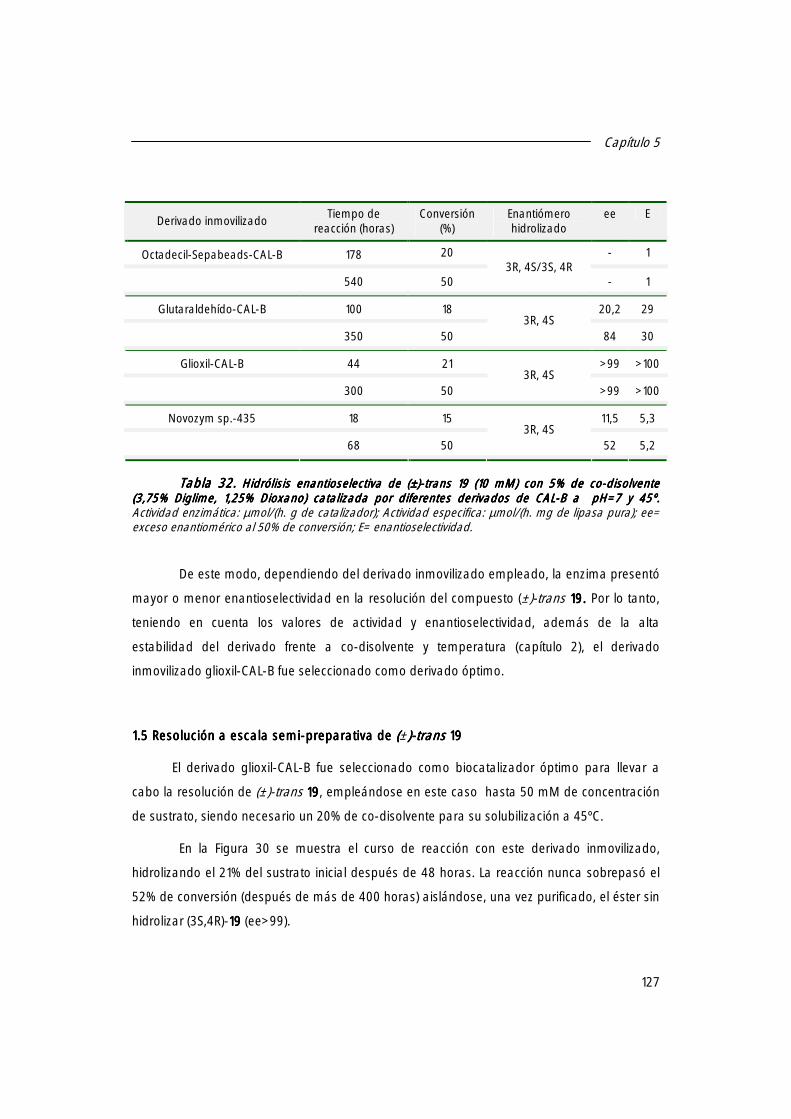
Capítulo 5
127
Tabla 3Tabla 3Tabla 3Tabla 32222.... Hidrólisis Hidrólisis Hidrólisis Hidrólisis enantioselectiva de (enantioselectiva de (enantioselectiva de (enantioselectiva de (±±±±))))----trans 19 (10 mM) con 5% de cotrans 19 (10 mM) con 5% de cotrans 19 (10 mM) con 5% de cotrans 19 (10 mM) con 5% de co----disolvente disolvente disolvente disolvente
(3(3(3(3,,,,75% Diglime, 175% Diglime, 175% Diglime, 175% Diglime, 1,,,,25% Dioxano) catalizada por diferentes derivados de CAL25% Dioxano) catalizada por diferentes derivados de CAL25% Dioxano) catalizada por diferentes derivados de CAL25% Dioxano) catalizada por diferentes derivados de CAL----B a pH=7 y 45º.B a pH=7 y 45º.B a pH=7 y 45º.B a pH=7 y 45º. Actividad enzimática: µmol/(h. g de catalizador); Actividad especifica: µmol/(h. mg de lipasa pura); ee= exceso enantiomérico al 50% de conversión; E= enantioselectividad.
De este modo, dependiendo del derivado inmovilizado empleado, la enzima presentó
mayor o menor enantioselectividad en la resolución del compuesto (±)-trans 1 1 1 19999. . . . Por lo tanto,
teniendo en cuenta los valores de actividad y enantioselectividad, además de la alta
estabilidad del derivado frente a co-disolvente y temperatura (capítulo 2), el derivado
inmovilizado glioxil-CAL-B fue seleccionado como derivado óptimo.
1.51.51.51.5 Resolución a escala semi Resolución a escala semi Resolución a escala semi Resolución a escala semi----preparativa de preparativa de preparativa de preparativa de ((((±))))----transtranstranstrans 19 19 19 19
El derivado glioxil-CAL-B fue seleccionado como biocatalizador óptimo para llevar a
cabo la resolución de (±)-trans 19 19 19 19, empleándose en este caso hasta 50 mM de concentración
de sustrato, siendo necesario un 20% de co-disolvente para su solubilización a 45ºC.
En la Figura 30 se muestra el curso de reacción con este derivado inmovilizado,
hidrolizando el 21% del sustrato inicial después de 48 horas. La reacción nunca sobrepasó el
52% de conversión (después de más de 400 horas) aislándose, una vez purificado, el éster sin
hidrolizar (3S,4R)-19191919 (ee>99).
Derivado inmovilizado Tiempo de reacción (horas)
Conversión (%)
Enantiómero hidrolizado
ee E
Octadecil-Sepabeads-CAL-B 178 20 - 1
540 50
3R, 4S/3S, 4R
- 1
Glutaraldehído-CAL-B 100 18 20,2 29
350 50 3R, 4S
84 30
Glioxil-CAL-B 44 21 >99 >100
300 50 3R, 4S
>99 >100
Novozym sp.-435 18 15 11,5 5,3
68 50 3R, 4S
52 5,2
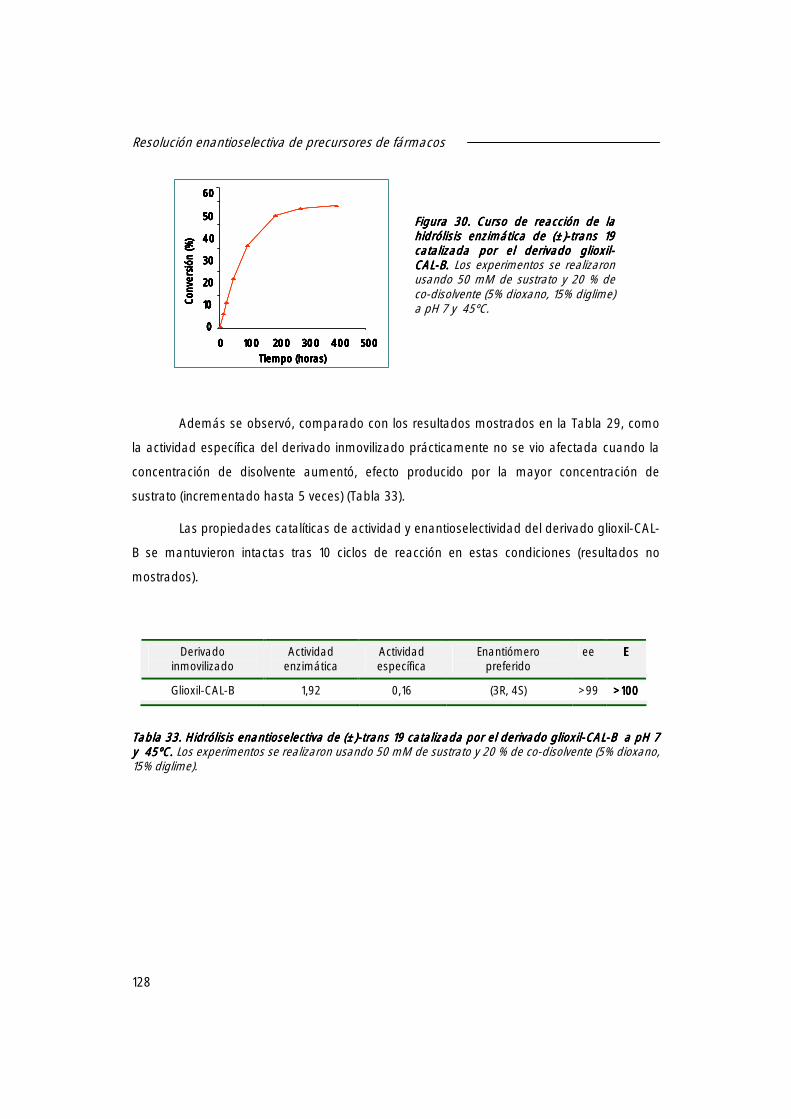
Resolución enantioselectiva de precursores de fármacos
128
Figura 3Figura 3Figura 3Figura 30000.... Curso de reacción de la Curso de reacción de la Curso de reacción de la Curso de reacción de la hidrólisis enzimática de (±)hidrólisis enzimática de (±)hidrólisis enzimática de (±)hidrólisis enzimática de (±)----trans 19 trans 19 trans 19 trans 19 catalizada por el derivado glioxilcatalizada por el derivado glioxilcatalizada por el derivado glioxilcatalizada por el derivado glioxil----CALCALCALCAL----B. B. B. B. Los experimentos se realizaron usando 50 mM de sustrato y 20 % de co-disolvente (5% dioxano, 15% diglime) a pH 7 y 45ºC.
Además se observó, comparado con los resultados mostrados en la Tabla 29, como
la actividad específica del derivado inmovilizado prácticamente no se vio afectada cuando la
concentración de disolvente aumentó, efecto producido por la mayor concentración de
sustrato (incrementado hasta 5 veces) (Tabla 33).
Las propiedades catalíticas de actividad y enantioselectividad del derivado glioxil-CAL-
B se mantuvieron intactas tras 10 ciclos de reacción en estas condiciones (resultados no
mostrados).
Tabla 33.Tabla 33.Tabla 33.Tabla 33. HiHiHiHidrólisis enantioselectiva de (±)drólisis enantioselectiva de (±)drólisis enantioselectiva de (±)drólisis enantioselectiva de (±)----trans 19 catalizada por el derivado glioxiltrans 19 catalizada por el derivado glioxiltrans 19 catalizada por el derivado glioxiltrans 19 catalizada por el derivado glioxil----CALCALCALCAL----B a pH 7 B a pH 7 B a pH 7 B a pH 7 y 45ºC. y 45ºC. y 45ºC. y 45ºC. Los experimentos se realizaron usando 50 mM de sustrato y 20 % de co-disolvente (5% dioxano, 15% diglime).
Derivado inmovilizado
Actividad enzimática
Actividad específica
Enantiómero preferido
ee EEEE
Glioxil-CAL-B 1,92 0,16 (3R, 4S) >99 >100>100>100>100
0000
10101010
20202020
30303030
40404040
50505050
60606060
0000 100100100100 200200200200 300300300300 400400400400 500500500500Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Conv
ersi
Conv
ersi
Conv
ersi
Conv
ersi
óó óó n (%
)n
(%)
n (%
)n
(%)
0000
10101010
20202020
30303030
40404040
50505050
60606060
0000 100100100100 200200200200 300300300300 400400400400 500500500500Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Conv
ersi
Conv
ersi
Conv
ersi
Conv
ersi
óó óó n (%
)n
(%)
n (%
)n
(%)
0000
10101010
20202020
30303030
40404040
50505050
60606060
0000
10101010
20202020
30303030
40404040
50505050
60606060
0000 100100100100 200200200200 300300300300 400400400400 5005005005000000 100100100100 200200200200 300300300300 400400400400 500500500500Tiempo (horas)Tiempo (horas)Tiempo (horas)Tiempo (horas)
Conv
ersi
Conv
ersi
Conv
ersi
Conv
ersi
óó óó n (%
)n
(%)
n (%
)n
(%)

Capítulo 5
129
2.2.2.2. Resolución enantioselectiva de Resolución enantioselectiva de Resolución enantioselectiva de Resolución enantioselectiva de ( ( ( (±±±±))))----2222----hidrhidrhidrhidroxioxioxioxi----4444----fenilbutirato de etilo fenilbutirato de etilo fenilbutirato de etilo fenilbutirato de etilo [(±)-2222], precursor en , precursor en , precursor en , precursor en
la síntesis del (S)la síntesis del (S)la síntesis del (S)la síntesis del (S)----Enalapril, Enalapril, Enalapril, Enalapril, catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de catalizada por los derivados inmovilizados de la lipasa de la lipasa de la lipasa de la lipasa de
Bacillus themocatenulatusBacillus themocatenulatusBacillus themocatenulatusBacillus themocatenulatus ( ( ( (BTL2BTL2BTL2BTL2))))....
Para finalizar, otra de las resoluciones estudiadas en esta tesis, a través de la reacción de
hidrólisis enzimática en medios acuosos, fue la realizada para (±)-2-hidroxi-4-fenilbutirato de
etilo, precursor del S-Enalapril (Esquema 37). Para ello se empleo la BTL2 inmovilizada sobre
distintos soportes.
Esquema 3Esquema 3Esquema 3Esquema 37777. Hidrólisis enantioselectiva de (±)-2 2 2 2 catalizada por derivados inmovilizados de
BTL2.
2.1 2.1 2.1 2.1 Actividad específica de los derivados de BTL2 en la hidrólisis de Actividad específica de los derivados de BTL2 en la hidrólisis de Actividad específica de los derivados de BTL2 en la hidrólisis de Actividad específica de los derivados de BTL2 en la hidrólisis de (±)-2 2 2 2
En primer lugar, se analizó la actividad específica de los distintos derivados
inmovilizados de BTL2 a pH 7,5 y 25ºC (Tabla 34).
El derivado inmovilizado glioxil-BTL2 presentó el valor más alto de actividad, siendo 26
veces mayor que la del derivado octadecil-Sepabeads-BTL2.
2.1.1 Efecto de la temperatura sobre la actividad.
Cuando la temperatura disminuyó a 4ºC (Tabla 34), la actividad del derivado
inmovilizado octadecil-Sepabeads-BTL2 descendió más de cinco veces respecto a la obtenida
a 25ºC. Sin embargo, la actividad específica de los otros derivados inmovilizados disminuyó a
la mitad de 25 a 4ºC. Además un incremento en la temperatura (de 25ºC hasta 37ºC) propició
un aumento en la actividad de hasta 3 veces para el derivado octadecil-Sepabeads-BTL2
(Tabla 34).
COOEt
OH
(±)-2222
OH
COOH
(R)-4444
+H2O
Derivados BTL2
COOEt
OH
(S)-2222
COOEt
OH
(±)-2222
COOEt
OH
(±)-2222
OH
COOH
(R)-4444
OH
COOH
(R)-4444
+H2O
Derivados BTL2
H2O
Derivados BTL2
COOEt
OH
(S)-2222
COOEt
OH
(S)-2222
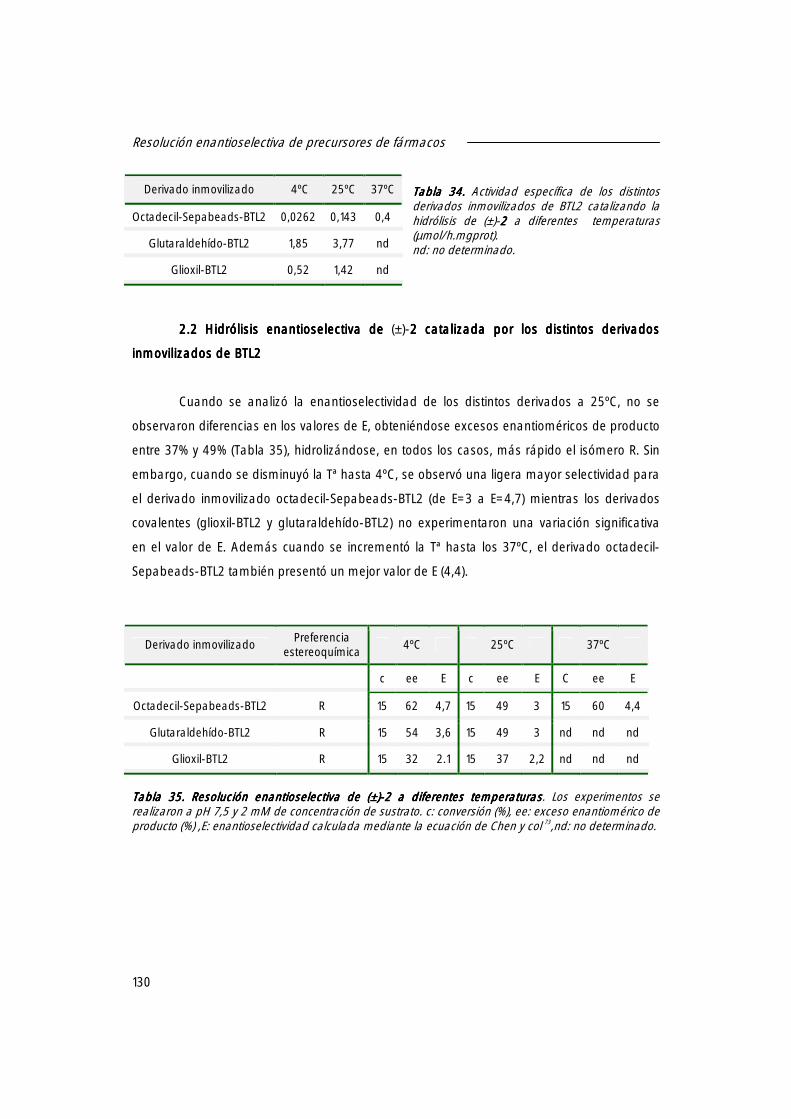
Resolución enantioselectiva de precursores de fármacos
130
Tabla 3Tabla 3Tabla 3Tabla 34444.... Actividad específica de los distintos derivados inmovilizados de BTL2 catalizando la hidrólisis de (±)-2222 a diferentes temperaturas (µmol/h.mgprot). nd: no determinado.
2.22.22.22.2 HiHiHiHidrólisis enantioselectiva de drólisis enantioselectiva de drólisis enantioselectiva de drólisis enantioselectiva de (±)-2 2 2 2 catalizada por los distintos derivados catalizada por los distintos derivados catalizada por los distintos derivados catalizada por los distintos derivados
inmovilizados deinmovilizados deinmovilizados deinmovilizados de BTL2 BTL2 BTL2 BTL2
Cuando se analizó la enantioselectividad de los distintos derivados a 25ºC, no se
observaron diferencias en los valores de E, obteniéndose excesos enantioméricos de producto
entre 37% y 49% (Tabla 35), hidrolizándose, en todos los casos, más rápido el isómero R. Sin
embargo, cuando se disminuyó la Tª hasta 4ºC, se observó una ligera mayor selectividad para
el derivado inmovilizado octadecil-Sepabeads-BTL2 (de E=3 a E=4,7) mientras los derivados
covalentes (glioxil-BTL2 y glutaraldehído-BTL2) no experimentaron una variación significativa
en el valor de E. Además cuando se incrementó la Tª hasta los 37ºC, el derivado octadecil-
Sepabeads-BTL2 también presentó un mejor valor de E (4,4).
Tabla 35Tabla 35Tabla 35Tabla 35.... Resolución enantioselectiva de (Resolución enantioselectiva de (Resolución enantioselectiva de (Resolución enantioselectiva de (±±±±))))----2222 a diferentes temperaturas a diferentes temperaturas a diferentes temperaturas a diferentes temperaturas. Los experimentos se realizaron a pH 7,5 y 2 mM de concentración de sustrato. c: conversión (%), ee: exceso enantiomérico de producto (%) ,E: enantioselectividad calculada mediante la ecuación de Chen y col 73,nd: no determinado.
Derivado inmovilizado 4ºC 25ºC 37ºC
Octadecil-Sepabeads-BTL2 0,0262 0,143 0,4
Glutaraldehído-BTL2 1,85 3,77 nd
Glioxil-BTL2 0,52 1,42 nd
Derivado inmovilizado Preferencia estereoquímica
4ºC 25ºC 37ºC
c ee E c ee E C ee E
Octadecil-Sepabeads-BTL2 R 15 62 4,7 15 49 3 15 60 4,4
Glutaraldehído-BTL2 R 15 54 3,6 15 49 3 nd nd nd
Glioxil-BTL2 R 15 32 2.1 15 37 2,2 nd nd nd

Capítulo 5
131
2.2.1 Efecto del disolvente sobre la enantioselectividad del derivado octadecil-Sepabeads-BTL2
catalizando la hidrólisis de (±)-2222....
Aunque el derivado octadecil-Sepabeads-BTL2 no fue el más activo catalizando la
hidrólisis de (±)-2222, si presentó un mayor valor de enantioselectividad. Así, centrándonos en
esta propiedad catalítica, intentaremos mejorar el valor de E de este derivado inmovilizado
mediante la modificación en las condiciones de reacción.
Para ello, en primer lugar se analizó el efecto de la presencia de co-disolvente sobre
el valor de E, añadiéndose un 10% de varios co-disolventes en el medio de reacción (Tabla 36).
De este modo, se observó un incremento en la enantioselectividad al añadir estos co-
disolventes, aunque el efecto más acusado tuvo lugar en presencia de un 10% de acetonitrilo
donde la lipasa inmovilizada presentó un valor de E=8 (con 0% de co-disolvente presentaba
una E=3).
Además, la actividad específica del derivado inmovilizado de BTL2 se incrementó
ligeramente de (0,14 a 0,2-0,23) en estas condiciones, respecto a la reacción sin presencia de
disolvente, aunque en los tres casos de manera similar.
Tabla 36. Tabla 36. Tabla 36. Tabla 36. Hidrólisis enantioselectiva de (±)-2222 catalizada por octadecil-Sepabeads-BTL2 a 2 mM, pH=7, 25ºC y 10% co-disolvente.
2.2.2.2.3333 Efecto de la concentración del disolvente sobre el va Efecto de la concentración del disolvente sobre el va Efecto de la concentración del disolvente sobre el va Efecto de la concentración del disolvente sobre el valor de E.lor de E.lor de E.lor de E.
El mejor valor de enantioselectividad hasta el momento se había obtenido en unas
determinadas condiciones (10% de acetonitrilo, 25ºC, pH 7,5 y usando el derivado inmovilizado
octadecil-Sepabeads-BTL2 como catalizador). Sin embargo, la concentración de co-disolvente
en el medio tiene una gran influencia sobre la selectividad de lipasas, como ya se observó en
el capítulo 3.
Disolvente Conversión (%) Actividad específica ee E (R/S)
Dioxano 16 0,20 73 7,3
Acetonitrilo 15 0,21 75 8
Diglime 15 0,23 72 6,8
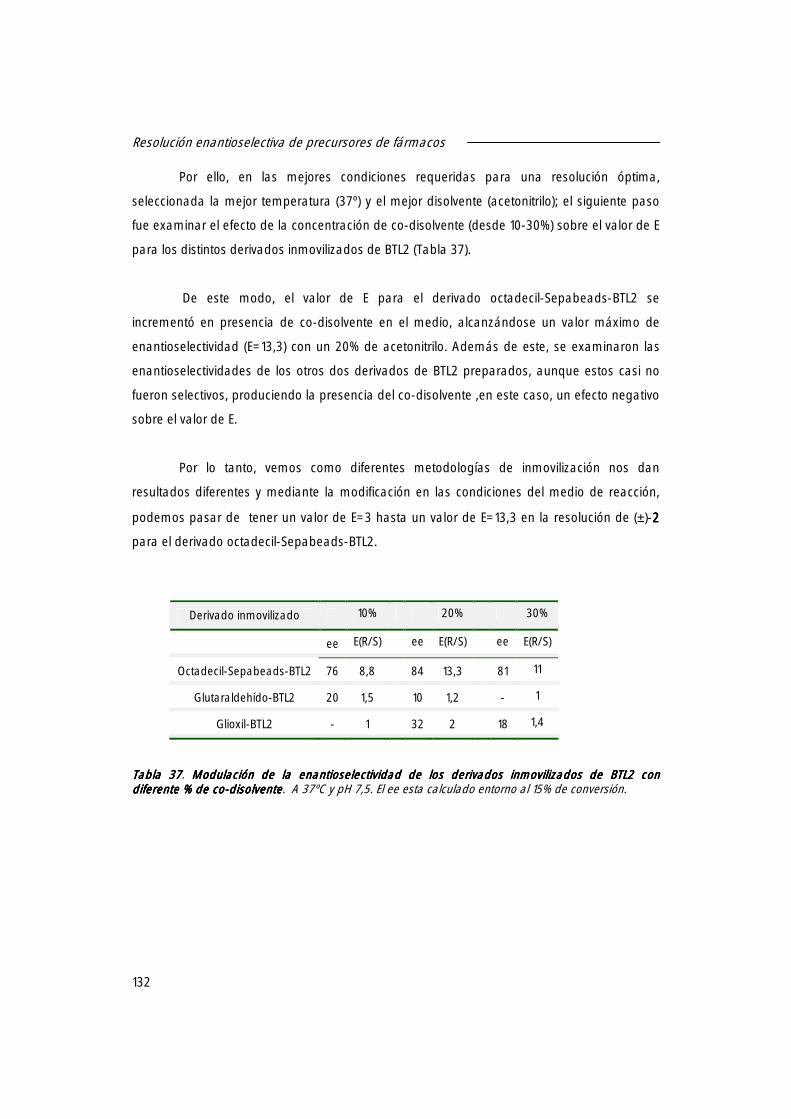
Resolución enantioselectiva de precursores de fármacos
132
Por ello, en las mejores condiciones requeridas para una resolución óptima,
seleccionada la mejor temperatura (37º) y el mejor disolvente (acetonitrilo); el siguiente paso
fue examinar el efecto de la concentración de co-disolvente (desde 10-30%) sobre el valor de E
para los distintos derivados inmovilizados de BTL2 (Tabla 37).
De este modo, el valor de E para el derivado octadecil-Sepabeads-BTL2 se
incrementó en presencia de co-disolvente en el medio, alcanzándose un valor máximo de
enantioselectividad (E=13,3) con un 20% de acetonitrilo. Además de este, se examinaron las
enantioselectividades de los otros dos derivados de BTL2 preparados, aunque estos casi no
fueron selectivos, produciendo la presencia del co-disolvente ,en este caso, un efecto negativo
sobre el valor de E.
Por lo tanto, vemos como diferentes metodologías de inmovilización nos dan
resultados diferentes y mediante la modificación en las condiciones del medio de reacción,
podemos pasar de tener un valor de E=3 hasta un valor de E=13,3 en la resolución de (±)-2222
para el derivado octadecil-Sepabeads-BTL2.
Tabla 37Tabla 37Tabla 37Tabla 37. Modulación de la enantioselectividad de los derivados inmovilizados de BTL2 con Modulación de la enantioselectividad de los derivados inmovilizados de BTL2 con Modulación de la enantioselectividad de los derivados inmovilizados de BTL2 con Modulación de la enantioselectividad de los derivados inmovilizados de BTL2 con diferente % de cdiferente % de cdiferente % de cdiferente % de coooo----disolventedisolventedisolventedisolvente. A 37ºC y pH 7,5. El ee esta calculado entorno al 15% de conversión.
Derivado inmovilizado 10% 20% 30%
ee E(R/S) ee E(R/S) ee E(R/S)
Octadecil-Sepabeads-BTL2 76 8,8 84 13,3 81 11
Glutaraldehído-BTL2 20 1,5 10 1,2 - 1
Glioxil-BTL2 - 1 32 2 18 1,4

Capítulo 5
133
2.4 Reversibilidad de la adsorción de BTL2 sobre octadecil2.4 Reversibilidad de la adsorción de BTL2 sobre octadecil2.4 Reversibilidad de la adsorción de BTL2 sobre octadecil2.4 Reversibilidad de la adsorción de BTL2 sobre octadecil----Sepabeads Sepabeads Sepabeads Sepabeads
Como ya se vio en el capítulo 4 para la CAL-B, además de la mejor
enantioselectividad, el derivado octadecil-Sepabeads-BTL2 presentó la ventaja de la posible
reutilización del soporte tras la pérdida de actividad de la enzima inmovilizada, aspecto
importante desde el punto de vista industrial.
Para examinar la fuerza de adsorción de la enzima sobre distintos soportes
hidrofóbicos, se estudió la cantidad de detergente necesaria para conseguir una completa
desorción de la enzima del soporte (Tabla 38). La lipasa sobre octadecil-Sepabeads necesitó
una mayor concentración de tritón para su completa desorción (1,5%), determinando esto una
unión más fuerte enzima-soporte en este derivado.
Esto permite emplear el octadecil-Sepabeads bajo condiciones más drásticas como
buen catalizador.
Tabla 38.Tabla 38.Tabla 38.Tabla 38. Fuerza de unión de BTL2 sobre soportes hidrofóbicos. Los números expresan la cantidad de detergente necesario para desorber 100% de la lipasa del soporte. El análisis se llevo acabo midiendo la actividad enzimática de sobrenadantes y suspensiones en la hidrólisis de pNPP.
Lipasa Soporte Tritón x-100 (%)
Octil-agarosa 0,2 BTL2
Octadecil-Sepabeads 1,5

Resolución enantioselectiva de precursores de fármacos
134
CONCLUSIONESCONCLUSIONESCONCLUSIONESCONCLUSIONES
Mediante la selección de la mejor lipasa (CAL-B) y el mejor derivado inmovilizado
(glioxil-CAL-B) fue posible llevar a cabo la resolución mediante reacción de hidrólisis de (±)-
trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxilato de etilo (trans-11119999).
Una alta enantioselectividad fue conseguida empleando el glioxil-CAL-B (E>100)
obteniéndose el enantiómero (3S,4R)-11119999 puro (éster sin reaccionar), el cual es el intermedio
con la configuración correcta para permitir la síntesis de (-)-11118888, precursor de la (-)-Paroxetina
(11117777).
Los resultados obtenidos con otros derivados inmovilizados de la misma enzima
mostraron enantioselectividades más bajas, por ejemplo glutaraldehído-CAL-B (E=30) e
incluso no fueron enantioselectivos, como el derivado octadecil-Sepabeads-CAL-B (E=1).
En la resolución del (±)-2-hidroxi-4-fenilbutirato de etilo [(±)-2222], se consiguió mejorar el
valor de enantioselectividad para el derivado octadecil-Sepabeads-BTL2 de E=3 a E=13
mediante la modificación en las condiciones de reacción del medio (ingeniería del medio).
Además, en unas mismas condiciones, el efecto de la inmovilización determinó pasar de una
E de 1,2 a 13, por ejemplo para el derivado glutaraldehído-BTL2 y octadecil-Sepabeads-BTL2,
respectivamente, a 37ºC, pH 7,5 y 20% de acetonitrilo.
Estos resultados refuerzan, aún más, la idea mostrada en el capítulo 3, sugiriendo
que las propiedades de una lipasa pueden ser moduladas mediante lo que hemos
denominado Ingeniería conformacional, siendo posible en algunos casos obtener un derivado
inmovilizado con alto valor de E frente al sustrato de interés.

Capítulo 5
135
PARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTALPARTE EXPERIMENTAL
1. 1. 1. 1. CompuestosCompuestosCompuestosCompuestos
Los compuestos (3RS,4RS)-trans-4-(4´-fluorofenil)-6-oxopiperidin-3-carboxilato de etilo [(±)-
trans-11119999] y (±)-2-hidroxi-4-fenilbutirato de etilo [(±)-2222] fueron donados por Vita Invest S.A.
(Barcelona, España).
2. 2. 2. 2. Preparación de los derivados inmovilizados de lipasas.Preparación de los derivados inmovilizados de lipasas.Preparación de los derivados inmovilizados de lipasas.Preparación de los derivados inmovilizados de lipasas.
La purificación y la preparación de los distintos derivados de CAL-B y de BTL2 se
realizaron como se describe en el capítulo 2. Los derivados octil-agarosa de las distintas
lipasas y los derivados de CAL-B presentaron una concentración de 12 mg de lipasa pura/mL,
mientras en el caso de la BTL2 se prepararon derivados con una concentración de 23 mg/mL
(máxima capacidad de carga de enzima en el soporte octadecil-Sepabeads).
3333. Hidrólisis enzimática de [(±). Hidrólisis enzimática de [(±). Hidrólisis enzimática de [(±). Hidrólisis enzimática de [(±)----transtranstranstrans----19191919] y (] y (] y (] y (±±±±))))----2222
El compuesto (±)-19 19 19 19 se disolvió en 20 mL de tampón fosfato sódico 25 mM pH 7 hasta
una concentración de 2 mM añadiendo a esto 1 gramo de derivado octil-agarosa de distintas
lipasas a 45ºC o 1 gramo de derivados CAL-B a distintas temperaturas. Asimismo, se disolvió
hasta 10 mM de sustrato conteniendo un 5% de co-disolvente (mezcla entre 1,4-dioxano y 2-
metoxietil éter (diglime) en proporción 1:3) a pH 7 y 45ºC, añadiéndose 1 gramo de derivado
enzimático.
Finalmente, la actividad del derivado glioxil-CAL-B se analizó mediante la adición de 1
g de derivado inmovilizado sobre una solución de 10 mL de sustrato (±)-19191919 con un 20% de co-
disolvente (dioxano/diglime 1:3) hasta una concentración de 50 mM a pH 7 y 45ºC.
La actividad de los derivados inmovilizados de BTL2 en la hidrólisis de (±)-2 2 2 2 se analizó
añadiendo 0,1 g de catalizador sobre una solución de 3 mL de tampón fosfato sódico 25 mM
a pH 7,5 con una concentración de 2 mM de sustrato a diferentes condiciones.

Resolución enantioselectiva de precursores de fármacos
136
En el transcurso de la reacción el valor del pH se mantuvo constante empleando un
pHstato Mettler Toledo DL50 graphic.
El grado de conversión se analizó por HPLC en fase-reversa (Spectra Physic SP 100)
sobre una columna Kromasil C18 (25x0,4 cm) suministrada por Análisis Vinicos. Los ensayos
se llevaron a cabo por triplicado. La fase móvil fue una mezcla isocrática de acetonitrilo: 10
mM de tampón fosfato amónico en agua bi-ionizada (30:70) (para el sustrato (±)-11119999), (40:60
para el sustrato (±)-2) 2) 2) 2) a pH 3,00 y con un flujo de 1 mL /min o 1,5 mL/min, respectivamente. La
absorbancia empleada fue de 270 nm o 254 nm, respectivamente (detector UV Spectra Physic
SP 8450). Los tiempos de retención de los compuestos fueron: ácido 22223333 (4,67 min), éster 11119999 (19
min), ácido 4444 (3,5 min), éster 2222 (8,3 min).
La actividad específica y actividad enzimática se definieron como µmol de sustrato por
hora y por mg de lipasa inmovilizada o por g de derivado inmovilizado, respectivamente;
calculándose como se describe en la parte experimental del capítulo 3.
4444. Determinación del exceso enantiomérico y la enantioselectividad.. Determinación del exceso enantiomérico y la enantioselectividad.. Determinación del exceso enantiomérico y la enantioselectividad.. Determinación del exceso enantiomérico y la enantioselectividad.
A diferentes grados de conversión, el exceso enantiomérico (ee) del éster 11119999 sin hidrolizar
se analizó por HPLC en fase quiral. La columna empleada fue una Chiral-AGP (100 x 4,0 mm),
la fase móvil fue una solución de tampón fosfato amónico 10 mM a pH 7 y los análisis se
realizaron a un flujo de 0,5 mL/min a una absorbancia de 210 nm. El tiempo de retención del
isómero (3R,4S) fue 14,73 minutos y 18,57 minutos para el isómero (3S,4R).
Igualmente se analizó el exceso enantiomérico del ácido 4444 formado mediante análisis en
HPLC quiral para una conversión del 15%. La columna empleada fue una CHIRACEL OD-R, la
fase móvil fue una mezcla isocrática de 20% de acetonitrilo y 80% de NaClO4/HClO4 0,5 M a
pH 2,3, flujo 0,5 mL/min analizado a una absorbancia de 254 nm.
La enantioselectividad se expresó en ambos casos como el valor de E calculado a partir
del exceso enantiomérico (ee) de sustrato (capítulo 1) ó de producto (capítulo 3) y del grado de
conversión (c) de acuerdo a la ecuación previamente descrita por Chen73.

Discusión general y conclusiones
137
Discusión General y Conclusiones Discusión General y Conclusiones Discusión General y Conclusiones Discusión General y Conclusiones
En la presente Tesis Doctoral se han evaluado y contrastado las muy interesantes
implicaciones biotecnológicas de un hipotético mecanismo de acción de lipasas en ausencia
de interfases hidrofóbicas; la actividad, selectividad y estabilidad de las lipasas en ausencia
de interfases hidrofóbicas esta determinada por un cierto equilibrio entre dos formas muy
diferentes de las lipasas (una forma cerrada e hidrofílica “versus” una forma abierta
conteniendo un gran bolsillo hidrofóbico expuesto al medio).
Esta hipótesis estaba inicialmente avalada por dos tipos de evidencias experimentales:
a.- A pesar de que la estructura cristalográfica de las lipasas en forma cerrada es
completamente inaccesible a sustratos pequeños, la mayoría de las lipasas bacterianas
(principalmente a concentraciones muy bajas y temperaturas moderadas o altas) muestran
una actividad esterásica relativamente alta hacia sustratos completamente solubles.
b.- experimentos previos reportados en la literatura y algunos experimentos obtenidos en
nuestro laboratorio muestran que algunas lipasas pueden presentar actividades y
selectividades muy diferentes dependiendo de los derivados inmovilizados utilizados y de las
condiciones de reacción.
Es decir, aun en ausencia de interfases hidrofóbicas, un cierto porcentaje de moléculas de
lipasas deben presentar su estructura abierta y menos estable y para ello es preciso que
complejos cambios conformacionales entre estructura cerrada y estructura abierta de las
lipasas estén involucrados en la catálisis de lipasas en ausencia de interfases hidrofóbicas.
A lo largo de esta Tesis Doctoral hemos ido encontrando muchísimas evidencias
experimentales que avalan esta hipótesis y que al mismo tiempo abren nuevas perspectivas
para una utilización más controlada y dirigida de las lipasas en biotransformaciones
industriales:

Discusión general y conclusiones
138
1.- A concentraciones elevadas y en ausencia de detergentes o interfases hidrofóbicas, las
lipasas tienden a auto-ensamblarse formando dímeros muy probablemente con los bolsillos
hidrofóbicos de su estructura abierta en íntimo contacto:
a.- En experimentos de filtración en gel las lipasas concentradas se eluyen a unos
volúmenes correspondientes al doble de su peso molecular y por el contrario las lipasas
diluidas y en presencia de detergentes se eluyen exactamente al volumen correspondiente a
su peso molecular.
b.- Experimentos de inactivación térmica nos muestran que las lipasas solubles
concentradas son mucho más estables que las lipasas solubles muy diluidas. De hecho las
estructuras abiertas reportadas en la literatura para las lipasas están mucho más
estructuradas que las estructuras cerradas (donde la cadena o lid que cubre el centro activo
tiene una estructura muy poco compacta).
c.- Muy interesantemente, los derivados inmovilizados (con las estructuras
enzimáticas completamente dispersas) de lipasas preparados a partir de disoluciones
concentradas de la enzima exhiben unas propiedades de actividad, selectividad y estabilidad
muy diferentes de las propiedades de los mismos derivados preparados a partir de enzima
muy diluida. Es decir, utilizando una u otra preparación de la misma enzima soluble podemos
modular las propiedades catalíticas de los derivados obtenidos.
2.- Las lipasas muestran una interesante tendencia a adsorberse sobre hidrofobinas
inmovilizadas. Las hidrofobinas son proteínas con una región superficial hidrofóbica muy
grande y una pequeña región superficial hidrofílica. Las hidrofobinas fueron inmovilizadas a
soportes hidrofílicos por la región más hidrofílica de la superficie de estas proteínas (pe., unión
covalente multipuntual entre hidrofobinas, por sus regiones más ricas en lisinas, y soportes
hidrofílicos glioxil-agarosa) quedando así expuestas al medio por sus regiones más
hidrofóbicas (una especie de gotas hidrofóbicas de tamaño y composición definida). Los
complejos formados entre las lipasas y las hidrofobinas inmovilizadas mostraron unas
propiedades similares a los complejos preparados entre lipasas y otras superficies

Discusión general y conclusiones
139
hidrofóbicas (pe., geles octil-agarosa) y parecen estar formados por la estructura abierta de
las lipasas con su bolsillo hidrofóbico en intimo contacto con la región hidrofóbica de las
hidrofobinas. De hecho, la enzima se adsorbe a las hidrofobinas inmovilizadas a muy baja
fuerza iónica donde las proteínas estándar no se adsorben a ningún soporte hidrofóbico, la
enzima se hiper-activa durante la adsorción y la enzima se desorbe con detergentes. La
posibilidad de utilizar hidrofobinas de diferente tamaño y con su superficie alterada (química o
genéticamente) podría abrirnos nuevos horizontes para la obtención de nuevos
biocatalizadores “híbridos” lipasa-hidrofobina con nuevas y mejores propiedades para las
biotransformaciones industriales.
3.- Las lipasas se adsorben muy fuertemente a soportes muy hidrofóbicos (Octadecil-
Sepabeads). La adsorción es muy similar a la previamente observada con otros geles
hidrofóbicos, pe., octil-agarosa. La adsorción ocurre vía adsorción interfacial de las lipasas
abiertas sobre las superficies hidrofóbicas. Ahora, al utilizar soportes más hidrofóbicos, las
propiedades de estos nuevos derivados son todavía mucho mejores con las observadas para
los derivados de lipasas adsorbidas a octil-agarosa, especialmente su estabilidad frente al
efecto del calor o de los co-disolventes orgánicos. De hecho estas formas abiertas de las
lipasas (formas más estructuradas) simplemente adsorbidas sobre superficies hidrofóbicas
son mucho mas estables que derivados de las formas cerradas (formas menos estructuradas)
estabilizados por inmovilización covalente multipuntual y ya mucho más estables que la
enzima soluble muy diluida o que derivados de la forma cerrada inmovilizados por uniones
covalentes menos intensas entre cada molécula de enzima y el soporte.
4.- Algunas lipasas de organismos termófilos también se podían inmovilizar por adsorción
interfacial sobre soportes muy hidrofóbicos y presentaban excelentes propiedades catalíticas:
estabilidad térmica todavía mucho mayor que las enzimas solubles, ya de por sí muy
estables, y una actividad a bajas temperaturas (pe, a 25 ºC ) cientos de veces mayor que la
bajísima actividad exhibida por las lipasas termófilas solubles a bajas temperaturas.
Todas estas evidencias sobre el mecanismo de acción de las lipasas en ausencia de
interfases hidrofóbicas nos llevaron seguidamente a plantarnos nuevas perspectivas para el
control y modulación de la selectividad de derivados inmovilizados de lipasas durante

Discusión general y conclusiones
140
biotransformaciones: la ingeniería conformacional de las reacciones catalizadas por lipasas
inmovilizadas.
La estructura de forma abierta de las lipasas esta determinada por dos interacciones físico-
químicas muy importantes:
a.- la interacción desfavorable de un gran bolsillo hidrofóbico con el medio acuoso al que esta
expuesto.
b.- la interacción favorable de la parte externa hidrofílica del “lid” que cerraba el centro activo
de la lipasa cerrada en intimo contacto con otro bolsillo hidrofílico “receptor” situado a la
vecindad del centro activo de la lipasas. En esta interacción intervienen interacciones
electrostáticas, puentes de hidrogeno, etc.
Así pues la estabilidad de la forma abierta de las lipasas esta muy influenciada por dos
interacciones de signo contrario: una interacción desfavorable del bolsillo hidrofóbico con el
medio y una interacción favorable de la región hidrofílica del “lid” y el bolsillo donde se aloja.
Si modificamos estas dos interacciones la forma exacta del centro activo de la estructura
abierta de las lipasas podrá sufrir ligeras modificaciones y por ello las propiedades catalíticas
podrían ser modificadas muy considerablemente. Por un lado intentamos la modificación de
estas interacciones como una consecuencia del proceso de inmovilización de las lipasas: a.-
induciendo mayor o menor rigidez diferentes regiones de su superficie que podrían estar más
o menos cercanas al sitio catalítico, b.- modificando el micro-ambiente que rodea cada
molécula de enzima inmovilizada (pe., introduciendo polímeros poli-catiónicos que podrían
desestabilizan la exposición del bolsillo hidrofóbico y al mismo tiempo interferir en las
interacciones entre el “lid” y su bolsillo receptor, etc.) c.- utilizando las formas abiertas de las
lipasas adsobidas interfacialmente a soportes hidrofóbicos, etc. . Por otro lado intentamos la
modificación de la estabilidad de la forma abierta de las lipasas trabajando a diferentes
temperaturas de reacción (la exposición del bolsillo hidrofóbico debe desestabilizarse a
medida que bajamos la temperatura) o trabajando a diferentes pHs (las interacciones
hidrofílicas entre el “lid” y el bolsillo receptor podrían alterarse también drásticamente). De este
modo, diferentes derivados de una misma lipasa podrían tener, y de hecho las tienen,

Discusión general y conclusiones
141
propiedades de actividad y selectividad muy diferentes y también un mismo derivado de una
misma lipasa podría comportarse, y de hecho lo hace, de un modo completamente diferente
a pH 5,0 y 4 ºC que a pH 7,0 y 25 ºC.
En estas circunstancias en esta Tesis hemos conseguido demostrar muy claramente que
diferentes derivados de una misma lipasa en diferentes condiciones experimentales podrían
exhibir muy diferentes actividades y selectividades y por ello utilizando una batería de
diferentes lipasas podríamos abordar con bastantes posibilidades de éxito la resolución de
mezclas racémicas de cualquier tipo de éster quiral por hidrólisis enantioselectiva: tanto
esteres modelo con carbonos quirales en la cadena del ácido carboxílico o en la molécula del
alcohol como esteres de relevancia químico-farmacéutica.
Finalmente, a modo de resumen de la siguiente Tesis Doctoral quisiéramos resaltar las
siguientes conclusiones:
1. Las lipasas tienden a formar estructuras diméricas incluso a concentraciones bajas, lo
cual fue determinado mediante experimentos de filtración en gel, donde las lipasas
concentradas se eluyen a unos volúmenes correspondientes al doble de su peso
molecular y por el contrario las lipasas diluidas y en presencia de detergentes se
eluyen exactamente al volumen correspondiente a su peso molecular.
2. Estas dos formas estructurales de lipasa (monómero y dímero) presentan
propiedades de estabilidad, actividad y enantioselectividad muy diferentes. De este
modo; por ejemplo, la actividad específica de CRL disminuyó al incrementarse la
concentración de enzima. La temperatura óptima de MML aumentó de 56ºC a 64ºC al
incrementarse la concentración de enzima.
3. La adsorción de lipasas sobre soportes recubiertos con polímeros catiónicos (PEI)
permite la inmovilización de la lipasa como dímero, mostrando una selectividad
distinta que cuando se encuentra como monómero. Por ejemplo, la lipasa de HLL en
condiciones diluida no fue casi enantioselectiva (E=1,07) mientras la lipasa
concentrada mostró un valor de E=4 frente a 2-hidroxi-4-fenilbutirato de etilo.

Discusión general y conclusiones
142
4. La adsorción de lipasas sobre hidrofobina, inmovilizada previamente de forma
covalente en soporte glioxil-agarosa, se produce mediante activación interfacial de la
misma manera que sucede sobre soportes hidrofóbicos, mostrando propiedades
catalíticas similares. Por ejemplo, la HLL sufrió una hiperactivación de hasta 9 veces
tras su inmovilización sobre hidrofobina. CAL-B inmovilizada sobre glioxil-hidrofobina
y sobre octil-agarosa presentó un valor de E=11 y E=9, respectivamente, en la
hidrólisis del mandelato de metilo.
5. La inmovilización de lipasa soporte octadecil-Sepabeads, donde se produce la fijación
de la conformación abierta de la lipasa, dio lugar a la mayor estabilización incluso
por encima de la obtenida tras la inmovilización mediante unión covalente
multipuntual. Por ejemplo, la CRL inmovilizada sobre soporte octadecil-Sepabeads
mantuvo el 100% de la actividad tras incubación a 45ºC durante 130 horas, mientras
la enzima unida de forma covalente multipuntual sobre glioxil-agarosa conservó el
70% de actividad, en las mismas condiciones y la enzima soluble perdió su actividad
después de 1 hora.
6. La inmovilización de lipasas procedentes de organismos termófilos sobre la resina
octadecil-Sepabeads dio lugar a un incremento notable en la estabilidad, además de
la posibilidad de utilizar estas enzima a temperaturas consideradas moderadas-
bajas (25ºC) para una termofila. Por ejemplo, TAL inmovilizada mantuvo 100% de
actividad a 70ºC después de 70 horas de incubación mientras la enzima libre solo
conservo un 20% en las mismas condiciones. TTL inmovilizada sobre octadecil-
Sepabeads presentó una actividad catalítica más de cien veces mayor que la de la
enzima libre a 25ºC.
7. Diferentes derivados inmovilizados de una misma lipasa presentan distinta actividad
frente a un mismo sustrato. Por ejemplo, la CAL-B inmovilizada sobre octadecil-
Sepabeads presentó 80 veces mayor actividad que cuando fue inmovilizada sobre
Eupergit-Cu en la hidrólisis del butirato de etilo.

Discusión general y conclusiones
143
8. Los distintos derivados de una misma lipasa presentan enantioselectividades muy
diferentes. Por ejemplo, la CRL inmovilizada sobre octil-agarosa no mostró
enantioselectividad (E≅ 1) mientras cuando se inmovilizó sobre soporte agarosa-
glutaraldehído presentó un valor de E mayor de 100 en la hidrólisis del ácido-2-O-
butiril-2-fenilacético a pH 7 y 25ºC. La CAL-B inmovilizada sobre agarosa-
bromocianógeno mostró un valor de E=7,4 mientras tras la inmovilización sobre
soporte agarosa-PEI presentó un valor de E=67 en la hidrólisis del mandelato de
metilo a pH 7 y 25ºC.
9. Pequeños cambios en las condiciones experimentales en las que se realiza la
reacción de hidrólisis (pH,Tª,co-disolvente, fuerza iónica,…) pueden también
ocasionar grandes alteraciones en la selectividad de un mismo derivado enzimático
de una misma lipasa. Por ejemplo, el derivado octadecil-Sepabeads-BTL2, donde se
observó un cambio en la enantioselectividad pasando de una E=15 (a 25ºC) a un valor
de E mayor de 100 (a 4ºC) en la hidrólisis del ácido-2-O-butiril-2-fenilacético.
10. Hemos conseguido resolver enantioselectivamente diferentes precursores,
intermedios claves en la síntesis de distintos fármacos, mediante el empleo de CAL-B
inmovilizada sobre distintos soportes como catalizador. Por ejemplo, la resolución del
precursor de la (-)-Paroxetina empleando el derivado glioxil-CAL-B (E>100). La
resolución de la (+)-Zopiclona empleando el derivado octadecil-Sepabeads-CAL-B
con un alto valor de enantioselectividad (E>100).

Bibliografía
144
1. Reetz, M. T.; Rüggeberg, C. J.; Dröge, M. J.; Quax, W. J. Immobilization of chiral
enzyme inhibitors on solid supports by amide-forming coupling and olefin metathesis.
Tetrahedron. 2002200220022002, 58, 8465-8473.
2. Koeller, K.M; Wong C-H. Enzymes for chemical synthesis. Nature. 2001200120012001,409,232-240.
3. Bornscheuer, U.T.; Kazlauskas, R. J. Hydrolases in Organic synthesis: Regio and
Stereoselective Biotransformations, Wiley-VCH, Weinheim (1999).
4. Nara, S. J.; Harjani, J. R.; Maikrao, M.M; Salunkhe, M.; Mane, A. T.; Wadgaonkar, P.P.
Lipase-catalysed polyester synthesis in 1-butyl-3-methylimidazolium
hexafluorophosphate ionic liquid. Tetrahedron Letters.2003200320032003, 44, 1371-1373.
5. Guieysse, D; Salagnad, C.; Monsan, P.; Remaud-Simeon, M. Lipase-catalyzed
enantioselective transesterification toward esters of 2-bromo-tolylacetic acids,
Tetrahedron:Asymmetry. 2003200320032003,14,317-323.
6. Murakami, M.; Kamaya, H.; Kaneko, C.; Sato, M. Synthesis of optically active 1,3-
dioxin-4-one derivatives having a hydroxymethyl group at the 2-position and their use
for regio-, diastereo-, and enantioselective synthesis of substituted cyclobutanols.
Tetrahedron: Asymmetry, 2003200320032003,14, 201-215.
7. Sobolev, A.; Franssen, M.C.R.; Poikans, J.; Duburs, G.;de Groot, A. Enantioselective
lipase-catalysed kinetic resolution of acyloxymethyl and ethoxycarbonylmethyl esters
of 1,4-dihydroisonicotinic acid derivatives. Tetrahedron: Asymmetry.2002200220022002,,,, 13, 2389-
2397.
8. Solymár, M.; Fülöp, F.; Kanerva, L.T. Candida antarctica lipase A–a powerful catalyst
for the resolution of heteroaromatic -amino esters. Tetrahedron: Asymmetry.2002200220022002.
13, 2383-2388.
9. a) Queiroz,N.; Nascimento, M.G. Pseudomonas sp. lipase immobilized in polymers
versus the use of free enzyme in the resolution of (R,S)-methyl mandelate. Tetrahedron
Letters.2002200220022002, 43, 5225-5227, b) Singh-Chimni, S.; Singh, S.; Kumar,S.; Mahajan,S.
Kinetic resolution of heteroaryl -hydroxy sulfides catalyzed by Humicola lanuginosa
lipase, Tetrahedron: Asymmetry. 2002200220022002,13, 511-517.
10. González-Sabín, J.; Gotor, V.; Rebolledo, F. CAL-B-catalyzed resolution of some
pharmacologically interesting –substituted isopropylamines. Tetrahedron:
Asymmetry. 2002200220022002,13, 1315-1320.

Bibliografía
145
11. Kamal,A.; Sandbhor,M.; Ramana, K.V. One-pot lipase-catalyzed synthesis of
enantiopure secondary alcohols from carbonyl compounds: a new protocol for lipase-
mediated resolution. Tetrahedron: Asymmetry. 2002200220022002, 13, 815-820.
12. García-Urdiales,E.; Rebolledo, F.; Gotor,V. Study of the enantioselectivity of the CAL-B-
catalysed transesterification of -substituted -propylmethanols and -substituted
benzyl alcohols. Tetrahedron: Asymmetry.2001200120012001, 12, 3047-3052.
13. Palomo, Jose M.; Fernández-Lorente, G.; Mateo, C.; Fuentes, M.; Guisan J.M.;
Fernández-Lafuente, R. Enzymatic production of (3S,4R)-(-)-4-(4'-fluorophenyl)-6-oxo-
piperidin-3-carboxylic acid using a commercial preparation from Candida antarctica
A: the role of a contaminant esterase. Tetrahedron: Asymmetry. 2002200220022002, 13, 2653-2659.
14. Sharma,R.; Chisti, Y.; Banerjee, U.C. Production,purification,characterization,and
applications of lipases. Biotechnology advances. 2001200120012001,19,627-662.
15. Rubio, C.; Latxague, L.; Déléris, G.; Coulon,D. Enzymatic hydrolysis of
(chloromethyldimethylsilyl)-2-propenyl acetate isomers: atypic specificity of Candida
antarctica lipase. J. Biotech.2001200120012001, 92, 61-66.
16. Bonrath,W.; Karge, R.; Netscher, T. Lipase-catalyzed transformations as key-steps in
the large-scale preparation of vitamins. J. Mol Catal B: Enzymatic.2002200220022002, 19-20, 67-72.
17. Conde, S.; Fierros, M.; Rodríguez-Franco, M.I.; Puig, C. Resolution of 1-(4-amino-3-
chloro-5-cyanophenyl)-2-bromo-1-ethanol by lipase mediated enantioselective
alcoholysis, hydrolysis and acylation. Tetrahedron: Asymmetry.1998199819981998, 9, 2229-2232. 18. Salazar,L.; Bermudez, J.L.; Ramírez, C.; Llama, E.F.; Sinisterra, J.V. Resolution of 3- -
naphthoxy-1,2-propanediol using Candida antarctica lipase. Tetrahedron: Asymmetry.
1999199919991999, 10, 3507-3514.
19. López-Serrano, P.; Jongejan, J. A.; van Rantwijk, F.;Sheldon, R. A. Enantioselective
acylation of -aminonitriles catalysed by Candida antarctica lipase. An unexpected
turnover-related racemisation. Tetrahedron: Asymmetry. 2001200120012001,12, 219-228.
20. Overbeeke, P.L.A.; Jongejan, J.A. Enantioselectivity of Candida rugosa lipase in the
hydrolysis of 2-chloropropionic acid methyl ester. Journal of Molecular Catalysis B:
Enzymatic.2003200320032003, 21, 89-91.
21. Sarda, L. ; Desnuelle, P. Action de la lipase pancréatique sur les esters en emulsión.
Biochim. Biophys. Acta. 1958195819581958,30, 513-521.

Bibliografía
146
22. Brzozowski, A.M.; Derewenda, U.; Derewenda, Z.S.; Dodson, G.G.; Lawson, D.M.;
Turkenburg, J.P.; Bjorkling, F.; Huge-Jensen, B.; Patkar, S.A.; Thim, L. A model for
interfacial activation in lipases from the structure of a fungal lipase-inhibitor complex.
Nature. 1991199119911991,351, 491-494.
23. Norin, M.; Haeffner, F.; Achour, A.; Norin, T.; Hult, K. Computer modeling of substrate
binding to lipases from Rhizomucor Miehei, Humicola Lanuginosa and Candida
rugosa. Protein Sci. 1994199419941994,3, 1493-1503.
24. Mingarro, Y.; Abad, C.;Braco, L. Interfacial activation based molecular bioimprinting of
lipolytic enzymes. Proc. Natl. Acad. Sci. U.S.A. 1995199519951995, 92, 3308-3312.
25. Brady, L.; Brzozowski, A.M.; Derewenda, Z.S.; Dodson, E.; Dodson, G.; Tolley, S.;
Turkenburg, J.P.; Christiansen, L.; Huge-Jensen, B.; Norskov, L.; Thim, L.; Menge, U. A
serine protease triad forms the catalytic center of a triacylglycerol lipase. Nature.
1990199019901990,343, 767-770.
26. Miled, N.; Beisson, F.; de Caro, J.; de Caro, A.; Arondel, V.; Verger, R. Interfacial
catalysis by lipases. J Mol Catal B: Enzymatic. 2001200120012001, 11, 165-171
27. Derewenda, Z.S.; Derewenda, U. The cristal and molecular structure of the
Rhizomucor miehei tryacylglyceride lipase at 1.9 A resolution. J. Mol. Biol. 1992199219921992, 227,
818-839.
28. Grochulski, P.; Li, Y.; Schrag, J.D.; Bouthillier, F.; Smith, P.; Harrinson, D.; Rubin, B.;
Cygler, M.. Insights into interfacial activation from an open structure of Candida
rugosa lipase. J. Biol. Chem. 1993199319931993,268, 12843-12847.
29. Ampon, K.; Basri, M.; Salleh, A.B.; Wan Yunus; W.M.Z.; Razak, C.N.A. Immobilization
by adsorption of hydrophobic lipase derivatives to porous polymer beads for use in
ester synthesis. Biocatalysis. 1994199419941994, 10, 341-351
30. Kerzel, P.; Mersmann, A. Adsorption behavior of lipase from Stafilococcus carnosa on
a hydrophobic adsorbent. J. Chromatogr. 1992199219921992,584, 109-113.
31. Norin, M.; Boutleje, J.; Holmberg, E.; Hult, K. Lipase immobilized by adsorption. Effect
of the support hydrophobocity on the reaction rate of ester synthesis in cyclohexane.
Appl. Microb. technol. 1988198819881988,28, 527-530.
32. Derewenda, U.; Brzozowski, A.M.; Lawson, D. M.; Derewenda, Z.S. Catalysis at the
interface:the anatomy of a conformational change in a triglyceride lipase.
Biochemistry. 1992199219921992,31, 1532-1541.

Bibliografía
147
33. Basri, M.; Ampon, K.; Yunus, W.M.Z.; Razak, C.N.A.; Salleh, A.B. Enzymatic synthesis of
fatty esters by hydrophobic lipase derivatives immobilized on organic polymer beads.
J. Am .Oil Chem. Soc. 1995199519951995,72, 407-411.
34. Bastida, A.; Sabuquillo, P.; Armisen, P.; Fernández-Lafuente, R.; Huguet, J.; Guisán,
J.M. “A single step purification, immobilization and hyperactivation of lipases via
interfacial adsorption on strongly hydrophobic supports” .Biotechnol. Bioeng. 1998199819981998,58,
486-493.
35. Fernández-lafuente, R.; Armisén, P.; Sabuquillo, P.; Fernández-Lorente, G.; Guisán,
JM. Immobilization of lipases by selective adsorption on hydrophobic supports.
Chemistry and Physics of Lipids. 1998199819981998,93, 185-197.
36. Al-Duri, B.; Robinson, E.; McNerlan, S.; Bailie, P. Hydrolysis of edible oils by lipases
immobilized on hydrophobic supports: effects of internal support structure. J.Am.Oil
Chem. Soc. 1995199519951995,72, 1351-1359.
37. Sabuquillo, P.; Reina, J.; Fernández-Lorente, G.; Guisán, J.M.; Fernández-Lafuente, R.
“Interfacial affinity chromatography of lipases: separation of different fractions by
selective adsorption on supports activated with hydrophobic groups” Biochem.
Biophys. Acta. 1998199819981998,1388, 337-348.
38. Sugiura, M.; Isobe, M. (b). Studies on the mechanism of the lipase reaction. II.
Inhibition of lipase activity by emulsion of organic solvents. Chem. Pharm.
Bull.1975197519751975,23, 1221-1225.
39. Sugiura, M.; Isobe, M. Studies on the mechanism of lipase reaction. III. Adsorption of
Chromobacterium lipase on hydrophobic glass beads. Chem. Phar. Bull.1976197619761976, 24,72-
78.
40. Reetz, M.T. Entrapment of biocatalysts in hydrophobic sol-gel materials for use in
organic chemistry. Adv. Matter. 1997199719971997,9, 943-954.
41. Sugiura, M.; Isobe, M. (a) .Comparative studies on the adsorption of lipase and
various proteins at the air-water interface. Biochi. Biophys. Acta. 1975197519751975,397,,,, 412-417.
42. Taipa, M. A; Liebeton, K.; Costa, J. V.; Cabral, J. M.; Jaeger, K.E. Lipase from
Chromobacterium viscosum: biochemical characterization indicating homology to the
lipase from Pseudomonas glumae .Biochim.Biophys.Acta. 1995199519951995,1256,396-402.
43. Ghosh,D.; Wawrzak, Z.; Pletnev, V.Z.; Li,N.; Kaiser, R.; Pangborn, W.; Jornvall, H.;
Erman, M.; Duax, W.L.. Structure.1995199519951995, 3,279-288.

Bibliografía
148
44. a)Uppenberg, J.; Hansen, M. T.; Patkar, S.; Jones, T. A. The sequence, crystal
structure determination and refinement of two crystal forms of lipase B from Candida
antarctica Structure. 1994199419941994, 2, 293-308. b)Uppenberg, J.;Ohrner, N.; Norin, M.; Hult, K.;
Kleywegt, G. J.; Patkar, S.; Waagen, V.; Anthonsen, T.; Jones, T. A. Crystallographic
and molecular-modeling studies of lipase B from Candida antarctica reveal a
stereospecificity pocket for secondary alcohols. Biochemistry. 1995199519951995,34,16838.
45. Pernas, M.A.; López, C.; Rúa, M.L.; Hermoso, J. Influence of the conformational
flexibility on the kinetics and dimerisation process of two Candida rugosa lipase
isoenzymes. FEBS. Lett. 2001200120012001,501, 87-91.
46. Rúa, M.L.; Schmidt-Dannert, C.;Wahl, S.; Sprauer, A.; Schmid, R.D.... Thermoalkalophilic
lipase of Bacillus thermocatenulatus; Large-scale production, purification and
properties: aggregation behaviour and its effect on activity, J. Biotechnol. 1997199719971997, 56, 89-
102.
47. Dünhaupt, A.; Lang, S.; Wagner, F. Pseudomonas cepacia lipase: studies on
aggregation, purification and on the cleavage of olive oil. Biotechnol. Lett.
1992199219921992,14,953-958.
48. Jaeger, K.E.; Adrian, F.J.; Meyer, H.E.;Hancock, R.E.W.; Winkler, U.K. Extracellular lipase
from Ps. Aeruginosa is an amphiphilic protein. Biochim. Biophys. Acta. 1992199219921992. 1120, 315-
321.
49. Lesuise, E.; Schanck, K.; Colson, C. Purification and preliminary characterization of the
extracellular lipase of Bacillus subtilis 168, an extremely basic pH-tolerant enzyme. Eur.
J. Biochem. 1993199319931993,216,155-160.
50. Schmidt-Dannert, C.; Sztajer,H.; ,Stöcklein,W; Mengen, U.; Schimd, R. D.
Screening,purification and properties of a thermophilic lipase from Bacillus
Thermocatenulatus . Biochim. Biophys. Acta. 1994199419941994. 1301,105-114.
51. Sugihara, A.; Tani, T.; Tominaga, Y. Purification and characterisation of a novel
thermostable lipase from Bacilus sp. J. Biochem . 1991199119911991. 109, 211-216.
52. Sugihara, A.; Ueshima, M.; Shimada, Y.; Tsunasawa, S.; Tominaga, Y. Purification and
characterization of a novel thermostable lipase from Pseudomonas cepacia.
J.Biochem . 1992199219921992. 112,598-603.
53. Liou, Y-C.; Marangoni, A.; Yada, R.Y. Aggregation behaviour of Candida rugosa lipase.
Food Res. Int. 1998199819981998. 31, 243-248.

Bibliografía
149
54. Stuer, W.; Jaeger, K-E.;Winkler, U.K. Purification of extracellular lipase from Ps.
aeruginosa. J. Bacteriol.,1986198619861986, 168, 1070-1074.
55. Wessels, J. G. H. Hydrophobins, unique fungal proteins, Mycologist. The international
Journal of General Mycology. 2000200020002000,14,153-159.
56. Wosten, H. A. Hydrophobins: multipurpose proteins. Annu. Rev. Microbiol. 2001200120012001,
55,625-46.
57. Wösten, H. A. B. ; Asgeirsdottir, S. A.; Krook, J. H.; Drenth, J. H. H.; Wessels, J. G. H.
The fungal hydrophobin Sc3p self-assembles at the surface of aerial hyphae as a
protein membrane constituting the hydrophobic rodlet layer. Eur. J. Cell Biol.
1994199419941994,63,122-129.
58. Wosten, H. A.; M. Richter, et al. Structural proteins involved in emergence of microbial
aerial hyphae.Fungal Genet Biol. 1999199919991999, 27,153-60.
59. Peñas, M. M.; Asgeirsdóttir, S. A.; Lasa, I.; Culiañez-Macià, F. A.; Pisabarro, A. G.;
Wessels, J. G. H.; Ramírez, L. Identification, characterization, and in situ detection of a
fruit-body-specific hydrophobin of Pleurotus ostreatus. Appl. Environ. Microbiol. 1998199819981998,
64,4028-4034.
60. Asgeirsdottir, S. A.; de Vries, O. M. "Identification of three differentially expressed
hydrophobins in Pleurotus ostreatus (oyster mushroom)." Microbiology.1998199819981998,144,
2961-9.
61. Peñas, M. M.; Rust, B.; Larraya, L.M.; Ramírez, L.; Pisabarro, A.G. Differentially
regulated vegetative mycelium specific hydrophobins of the edible basidiomycete
Pleurotus ostreatus." Appl. Environ. Microbiol. 2002200220022002, 68,3891-3898.
62. Scholtmeijer, K.; Wessels, J. G. "Fungal hydrophobins in medical and technical
applications." Appl Microbiol. Biotechnol.2001200120012001, 56,1-8
63. Gregory, G.; Martin, G.; Cannon, C.; McCormick, C.L. Biomacromolecules. 2000200020002000, 1, 49-
60
64. Anderson, E. M.; Larson, K. M.; Kirk*, O. One biocatalyst-many applications:The use of
Candida Antarctica B lipase in organic synthesis. Biocatal. Biotransform. 1998199819981998,16, 181-
204.
65. a) Martinelle, M.; Holmquist, M.; Hult, K. On the interfacial activation of Candida
antarctica lipase A and B as compared with Humicola lanuginosa lipase. Biochim.
Biophys. Acta. 1995199519951995,1258, 272-276,b) Lawson, D.M.; Brzozowski, A.M.; Rety, S.; Verma,

Bibliografía
150
C.; Dodson, G.G. Probing the nature of substrate binding in Humicola lanuginosa
lipase through X-ray crystallography and intuitive modelling. Protein Eng. 1994199419941994,7, 543-
550.
66. Lotti, M.; Tramontano, A.; Longhi, S.; Fuseti,F.; Brocca, S.; Pizzi, E.; Alberniga,
L.”Variability within the Candida rugosa lipase family “. Protein. Eng. 1994199419941994,7,531-537.
67. a) Fernandez-Lorente,G.; Terreni, M.; Mateo, C.; Bastida, A.; Fernández-Lafuente, R.;
Dalmases, P.; Huguet, J.; Guisan, J. M.. Modulation of lipase properties in macro-
aqueous systems by controlled enzyme immobilisation: Enantioselective hydrolysis of
a chiral ester by immobilised Pseudomonas lipase. Enzyme Microb. Technol. 2001200120012001,28,
389-396.b) Liese, A.; Kragl, U.; Kierkels, H.; Schulze, B. Membrane reactor
development for the kinetic resolution of ethyl 2-hydroxy-4-phenylbutyrate. Enzyme
Microb. Technol. 2002200220022002,30, 673-687.
68. Torres, R.; Mateo,C.; Fuentes, M.; Palomo, Jose M.; Ortiz,C.; Fernández-Lafuente, R.;
Guisan, J.M.; Tam, A.; Daminati, M. Reversible immobilization of invertase on Sepabeads-
polyethylenimine: stabilization of a multimeric enzyme. Biotechnol. Prog.2002200220022002, 18,1221-
1226.
69. Guisán, J.M. Aldehyde-agarose gels as activated supports for immobilization-
stabilization of enzymes. Enzyme. Microb. Technol. 1988198819881988,10, 375-382.
70. Bradford, M.M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem.
1976197619761976,72,248-254.
71. Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature. 1970197019701970, 277,680-685.
72. Swank, R.T.; Munkres, K.D. Molecular weight analysis of oligopeptides by
electrophoresis in poliacrylamide gel with sodium dodecyl sulfate. Anal. Biochem.
1971,1971,1971,1971, 39, 462-477.
73. Chen,C.S.; Fujimoto Y.; Girdaukas,G.; Sih, C.J. Quantitative analyses of biochemical
kinetic resolutions of enantiomers. J.Am.Chem.Soc. 1982198219821982,104,7294-7299.
74. Mingarro, Y.; Abad, C.;Braco, L. Interfacial activation based molecular bioimprinting of
lipolytic enzymes. Proc. Natl. Acad. Sci. U.S.A. 1995199519951995, 92, 3308-3312.

Bibliografía
151
75. Malcata, F.X.; Reyes, H.R.; García, H.S.; Hill, C.G. Jr.; Amundson, C.H. Kinetics and
mechanisms of reactions catalysed by immobilized lipases. Enzyme Microb. Technol.
1992199219921992,14, 426-446.
76. Palomo, Jose M.; Fuentes,M.; Fernández-Lorente, G.; Mateo,C.; Guisan, J.M.; Fernández-
Lafuente, R. General trend of Lipase to auto-assemble giving bi-molecular aggregates
greatly modifies the enzyme functionality. Biomacromolecules.2003200320032003,4,1-6.
77. Fernández-Lorente, G.; Palomo, Jose M.; Fuentes,M.; Mateo,C.; Guisan, J.M.; Fernández-
Lafuente, R.Self-assembly of Pseudomonas fluorescens lipase into bimolecular
aggregates dramatically affects functional properties Biotechnol. Bieng. 2003200320032003,82,232-
237
78. Palomo, Jose M....; Peña, M.; Fernández-Lorente, G.;Mateo, C.; Pisabarro, A.G.;
Fernández-Lafuente, R.; Ramirez. L.; Guisan, J.M. Solid Phase Handling of
Hydrophobins: A New Tool to Study Lipases. Biomacromolecules.2003200320032003,4,204-210.
79. Palomo, José M.; Muñoz, G.; Fernández-Lorente, G.; Mateo, C.; Fernández-
Lafuente,R.; Guisán, J.M. Interfacial adsorption of lipases on very hydrophobic support
(Octadecyl Sepabeads): Immobilization, hyperactivation and stabilization of the open
form of lipases. J. Mol. Cat B: Enzymatic. 2002200220022002,19-20,279-286.
80. Fernandez-Lafuente,R.; Rosell,C.M.; Rodriguez, V.; Santana,C.; Soler,G.; Bastida,A.;
Guisan, J.M. Preparation of activated supports containing low pK amino groups. A
new tool for protein immobilization via carboxyl coupling method. Enzyme Microb.
Technol. 1993199319931993, 15, 546-550.
81. Fernández-Lafuente, R.; Rodríguez, V.; Guisán, J.M. The coimmobilization of -amino
acid oxidase and catalase enables the quantitative transformation of -amino acids
( -phenylalanine) into -keto acids (phenylpyruvic acid). Enzyme Microb. Technol.
1998199819981998,23,28-33.
82. Leckband, D.; Langer, R.An approach for the stable immobilization of proteins.
Biotechnol Bioeng. 1991991991990000,37, 227-237.
83. Guisán, J.M.; Blanco, R.M. Stabilization of Trypsin by multiple-point attachment to
aldehyde-agarose gels. Ann. NY Acad. Sci. 1987198719871987,501, 67-72.

Bibliografía
152
84. Otero, C.; Ballesteros, J.M.; Guisán, J.M. Immobilization/stabilization of lipase from
Candida rugosa. Appl. Biochem. Biotechnol. 1991199119911991, 19, 163-175.
85. Blanco, R.M.; Guisán, J.M.(a). “Stabilization of enzymes by multipoint covalent
attachment to agarose-aldehyde gels. Borohydride reduction of tripsin-agarose
derivatives”. Enzyme Microb. Technol. 1989198919891989,11, 360-366.
86. Blanco, R.M.; Calvete, J.J.; Guisán, J.M. (b). “Immobilization-stabilization of enzymes;
variables that control the intensity of the trypsin (amine)-agarose (aldehído) multipoint
attachment”. Enzyme Microb. Technol. 1989198919891989,11, 353-359.
87. Blanco, R.M.; Guisán, J.M. “Protecting effect of competitive inhibitors during very
intense insolubilized enzyme- activated support multipoint attachments: trypsin
(amine)- agarosa (aldehyde) system”, Enzyme Microb. Technol. 1988198819881988,10, 227-232.
88. Pedroche, J.; Yust, M.M.; Girón-Calle, J.; Vioque, J.; Alaiz, M.; Mateo, C.; Guisán, J.M.;
Millán, F. Stabilization–immobilization of carboxypeptidase A to aldehyde–agarose
gels: A practical example in the hydrolysis of casein. Enzyme Microb. Technol. 2002200220022002,
31, 711-718.
89. Fernández-Lafuente, R., Rosell, C.M., Guisán, J.M., Caanan-Haden, L. and Rodes, L.
“Facile synthesis of artificial enzyme nano- environments via solid- phase chemistry of
immobilized derivatives dramatic stabilization of penicillin acylase versus organic
solvents”. Enzyme Microb. Technol. 1999199919991999, 24, 96-10
90. Alvaro, G.; Fernández-Lafuente, R.; Blanco, R.M.; Guisán, J.M. “Immobilization-
stabilization of penicillin acylase from E.coli”, Appl. Biochem. Biotech. 1990199019901990, 26, 186-
195.
91. Herbert, R.A.A perspective on the biotechnological potential of extremophiles. TIBTECH.
1992199219921992,10,395–402.
92. Jaeger, K.E.; Ransac, S.; Dijkstra, B.W.; Colson, C.; van Heuvel, M.; Misset, O. Bacterial
lipases. FEMS Microbiol. Rev. 1994199419941994,15, 29-63.
93. Niehaus,F.; Bertoldo, C.; Kähler, M.; Antranikian, G. Extremophiles as a source of novel
enzymes for industrial application. Appl Microbiol Biotechnol.1999199919991999,51, 711–729.
94. Pennisi, E. In industry, extremophiles begin to make their mark. Science.1997199719971997,276,
705–706.

Bibliografía
153
95. Sharma, R.; Soni,S.K.; Vohra,R.M.; Gupta, L.K.;Gupta, J.K. Purification and
characterisation of a thermostable alkaline lipase from a new thermophilic Bacillus sp.
RSJ-1 R. Process Biochem. 2002200220022002,37,1075-1084.
96. Kim, H.K.; Sung, M.H.; Kim, H.M.; Oh, T.K. Occurrence of thermostable lipase in
thermophilic Bacillus sp. strain 398. Biosci. Biotech. Biochem. 1994199419941994, 58, 961–962.
97. Schmidt-Dannert, C.; Sztajer, H.; Stöcklein, W.; Menge, U.; Schmid, R.D. Screening,
purification and properties of a thermophilic lipase from Bacillus thermocatenulatus.
Biochim. Biophys. Acta. 1994 1994 1994 1994 ,1214, 43–53.
98. Schmidt-Dannert, C.; Rúa, M.L.; Atomi, H.; Schmid, R.D. Thermoalkalophilic lipase of
Bacillus thermocatenulatus. 1. Molecular cloning, nucleotide sequence, purification
and some properties. Biochim. Biophys. Acta.1996199619961996,1301, 105–114.
99. Rúa, M.L.; Schmidt-Dannert, C.; Wahl, S.; Sprauer,A.; Schmid, R.D. Thermoalkalophilic
lipase of Bacillus thermocatenulatus: Large-scale production, purification and
properties: aggregation behaviour and its effect on activity. J.Biotechnol. 1997199719971997, 56, 89-
102.
100. Lee, D.W.; Koh, Y.S.; Kim, K.J.; Kim, B.Y.; Choi, H.J.; Kim, D.S.; Suhartono, M.T.; Pyun,
Y.R. Isolation and characterization of a thermophilic lipase from Bacillus
thermoleovorans ID-1. FEMS Microbiol. Lett. 1999199919991999,,,,179,393–400.
101. Dartois, V.; Baulard, A.; Schank, K.; Colson, C. Cloning, nucleotide sequence and
expression of a lipase gene from Bacillus subtilis. Biochim. Biophys. Acta.
1995199519951995,263,3194-3201.
102. Kim, H.K.; Park, S.Y.; Lee, J.K.; Oh, T.K. Gene cloning and characterization of
thermostable lipase from Bacillus stearothermophilus L1. Biosci. Biotechnol Biochem.
1998199819981998,62,66-71.
103. Backeljau, T. Estimating of genic similarity within and between Arion hortensis s.l. and
A. intermedius by means of isolectric focused esterase patterns in hepatopancreas
homogenates (Mollusca, Pulmonata: Arionidae). Zeitschrift für Zoologische Systematik
und Evolutionsforschung.1985198519851985,23, 38–49.
104. Terreni, M.; Pagani, G.; Ubiali, D.; Fernández-Lafuente, R.; Mateo, C., Guisán, J. M.
“Modulation of Penicillin Acylase Properties via Immobilization Techniques: One-pot
Chemoenzymatic Synthesis of Cephamandole from Cephalosporin C” Bioorg Medic.
Chem. Let. 2001200120012001,18,2429-2432.

Bibliografía
154
105. Rocchietti, S.; San Vicente Urrutia, A.; Pregnolato, M.; Tagliani, A.; Guisàn, J.M.;
Fernàndez-Lafuente,R.; Terreni, M. "Influence of the enzyme derivative preparation
and reaction conditions on the enantioselectivity of Penicillin G Acylase" Enzyme
Microb. Technol. 2002200220022002,31,88-93.
106. Pessela, B.C.C.; Fernández-Lafuente,R.; Fuentes,M.; Vián, A.; García, J.L.; Carrascosa,
A.V.; Mateo, C.; Guisán, J.M. Reversible immobilization of a thermophilic -
galactosidase via ionic adsorption on PEI-coated Sepabeads. Enzyme Microb.
Technol. 2002002002003333, 32, 369-374.
107. Mateo,C.; Abian, O; Fernández-Lafuente, R.; Guisán, J.M. “Reversible enzyme
immobilization via a very strong and non-distorting adsorption on supports-
polyethyleneimine composites”. Biotechnol. Bioeng. 2000200020002000,68, 98-105.
108. Mateo, C.; Fernández-Lorente, G.; Abian, O.; Fernández-Lafuente, R.; Guisán, J.M.
"Multifunctional epoxy-supports: a new tool to improve the covalent immobilization of
proteins. The promotion of physical adsorptions of proteins on the supports before
their covalent linkage". Biomocromolecules. 2000200020002000,1,739-745.
109. Mateo, C.; Fernández-Lorente, G.; Cortes, E.; Garcia, J.L.; Fernández-Lafuente, R.;
Guisan, J.M. "One step purification, covalent immobilization and additional
stabilization of poly-His tagged proteins using novel heterofunctional chelate-epoxy
supports" Biotechnol. Bioeng. 2001200120012001, 76, 269-277.
110. Savidge, T.A. Biotechnology of Industrial Antibiotics. (Vandame, E.J., Ed) Marcel Decker
New York 177-224.1984.
111. Wilen, S.H. Topics in Stereochemistry N.L.Allinger, E.L. Eliel eds vol. 6. 1971.
112. Quintero, G.; Vo, M.; Farkas ,G.; Vigh, G. Series of homologous displacers for
preparative chiral displacement chromatographic separations on Cyclobond-II
columns.J.Chromatogr. A.1995199519951995,693,1-5.
113. Grover P.T.; Bhongle, N.N.; Wald, S.A; Senanayake, C.H. Chiral mandelic acid template
provides a highly practical solution for (S)-oxybutynin synthesis. J. Org Chem. 2000200020002000,
65, 6283-6287.
114. Kazuhiko, T.; Kazuo, O.; Kazunori, T.; Takao, T.; Youichi, S. Dedoublement et
determination de la configuration absolute des enantiomeres de Lácide (thienyl-3)-2
cyclohexyl-2 Acetique. Application a la synthese désters Anticholinergiques du
Quinuclidinol-3”. Chem. Pharm. Bull. 1992199219921992,40, 1415-1423.

Bibliografía
155
115. Fang,X.P.;Anderson,J. E. ;Chang,C.J.;McLaughlin,J.L.;Fanwick,P.E.J.Nad.Prod.1991199119911991,54,1
034-1043.
116. Tsubuki,M.;Kanai,K.; Honda, T. Goniodiol and 9-deoxygoniopypyrone :syntheses and
absolute configurations. J.Chem.Soc.Chem Común.1992199219921992,1640-1641,
117. Mukai, C.; Hirai, S.; Hanaoka, M. J. Org. Chem.1997199719971997,62,6619-6626,
118. Yang, Z.C.; Zhou, W.S. Heterocycles.1997199719971997,45,367-383.
119. Tsubuki, M.; Kanai, K.; Honda, T. Concise syntheses of novel styryllactones, (+)-
goniofufurone,(+)-goniopypyrone,(+)-goniotriol,(+)-8-acetylgoniotriol and (+)-
althollactone. Synlett.1993199319931993,653-655.
120. Xu, D.; Sharpless, K.B. Tetrahedron lett. 1994199419941994,35,4685-4688.
121. Goa, K L; Heel, R. C.; Zopiclone. A review of its pharmacodynamic and
pharmacokinetic properties and therapeutic efficacy as an hypnotic Drugs. 1986198619861986,32,
48-65.
122. Blaschke, G; Hempel, G; Müller, W E. Preparative and analytical separation of the
zopiclone enantiomers and determination of their affinity to the benzodiazepine
receptor binding site. Chirality. 1993199319931993,5, 419-421.
123. Parissi-Poulou, M.; Piperaki, , , , S. Enantiomeric separation of zopiclone, its metabolites
and products of degradation on a -cyclodextrin bonded phase. J. Chromatography
A. 1996199619961996, 729, 19-28.
124. Gotor, V.; Limeres, F.; García, R.; Bayod, M.; Brieva, R. Enzymatic resolution of (±)-6-(5-
chloropyridin-2-yl)-7-vinyloxy-carbonyloxy-6,7-dihydro[5H]pyrrolo[3,4-b]pyrazin-5-one.
Synthesis of (+)-zopiclone. Tetrahedron: Asymmetry. 1997199719971997, 8, 995-997.
125. Fernández-Lorente, G.; Fernández-Lafuente, R.; Palomo , J.M.; Mateo, C.; Bastida, A.;
Coca, J.; Haramboure, T.; Hernández-Justiz, O.; Terreni, M.; Guisán, J. M. ”Biocatalyst
Engineering exerts a dramatic effect on Selectivity of Hydrolysis catalyzed by
Immobilized Lipases in aqueous medium. J. Mol. Cat B: Enzymatic. 2001200120012001, 11, 649-656.
126. Palomo, Jose M.; Fernandez-Lorente, G.; Mateo,C.; Ortiz, C.; Fernandez-Lafuente,
R.,Guisan, J.M. Modulation of the enantioselectivity of lipases via controlled
immobilization and medium engineering: Hydrolytic resolution of Mandelic acid esters.
Enzyme Microb Technol. 2002200220022002,31,775-783.
127. Palomo, Jose M.; Fernandez-Lorente, G.; Mateo, C.; Fuentes, M.; Fernandez-Lafuente R.;
Guisan, J.M. Modulation of the enantioselectivity of Candida antarctica B lipase via

Bibliografía
156
conformational engineering: kinetic resolution of (±)-α-hydroxy-phenylacetic acid
derivatives. Tetrahedron: Asymmetry. 2002200220022002,13,1337-1345.
128. Palomo, Jose M. ; Muñoz, G. ; Fernandez-Lorente, G. ; Mateo, C. ; Fuentes, M.; Guisan,
J.M. ; Fernandez-Lafuente, R. Modulation of Mucor miehei lipase properties via
directed immobilization on different hetero-functional epoxy resins. Hydrolytic
resolution of (RS)-2-butyroyl-2-phenylacetic acid J. Mol. Cat B: Enzymatic.2003200320032003, 21,201-
210.
129. Solares, L.F.; Díaz,M.; Brieva,R.; Sánchez,V.M.; Bayod, M.; Gotor,V. Enzymatic
resolution of new carbonate intermediates for the synthesis of (S)-(+)-zopiclone.
Tetrahedron: Asymmetry. 2002200220022002, 13, 2577-2582.
130. Robertson, D. W.; Krushinski, J. H.; Fuller, R. W.; Leander, J. D. Absolute configurations
and pharmacological activities of the optical isomers of fluoxetine, a selective
serotonin-uptake inhibitor. J. Med. Chem. 1988198819881988,31,1412-1417.
131. Harriman, G.C.B.; Shao, J.; Luly, J.R. Synthesis of 4-substituted 4-aryl
piperidines.Tetrahedron Letters....2000200020002000, 41,8853-8856.
132. Lomenzo, S. A.; Izenwasser, S.; Gerdes, R. M.; Katz, J.L.; Kopajtic, T.; Trudell, M.
L.Synthesis, dopamine and serotonin transporter binding affinities of novel analogues
of meperidine. Bioorg. Med. Chem. Lett. 1999199919991999, 9, 3273–3276.
133. a) Liu, L. T.; Hong, P.-C.; Huang, H.-L.; Chen, S.-F.; Wang, C.-L.L.;Wen, Y.-S.
Tetrahedron:Asymmetry. 2001200120012001, 12, 419–426; b) Christensen, J.A.; Squires, R.F. 4-
Phenylpiperidine compounds. 1977. US 4007196.,c) Curzons, A.D.; Powell, L.W.; Keay,
A.M. Process for stereospecific hydrolysis of Piperidinedione derivatives. 1993.
WO9322284.
134. Cossy, J.; Mirguet, O.; Gomez Pardo, D.; Demurs, J-R. Ring expansion: formal total
synthesis of (-)-Paroxetine. Tetrahedron Letters. 2001200120012001,42, 5705-5707.
135. Yu, M.S.; Lantos,I.; Peng,Z.; Yu,J.; Cacchio,T. Asymmetric synthesis of (-)-paroxetine
using PLE hydrolysis. Tetrahedron Letters.2002002002000000,41,5647-5651
136. Kiyoshi, S.; Nobushige, I.; Tadashi, K.; Masami, I.; Shigeya, Y.; Taro, I.; Teiji, Y.;
Yoshihiro, K.; Yayoi, T. Piperidine derivatives as intermediates for the preparation of
Paroxetine and process for their preparation. 1997.EP0812827
137. a) Gao, Y.; Zeep, C.M. and Heefner, D.L. Method of preparing optically pure precursors
of Paroxetine. 1993. WO9403426; b) de Gonzalo, G.; Brieva, R.; Sánchez, V.M.; Bayod,

Bibliografía
157
M.; Gotor, V. Enzymatic resolution of trans-4-(4´-fluorophenyl)-3-
hydroxymethylpiperidines,key intermediates in the synthesis of (-)-Paroxetine.
J.Org.chem. 2001200120012001,66, 8947-8953.
138. a)Wyvratt, M. J.; Patchett, A. A. Med. Res. Rev. 1985, 5, 483-531. (b) Pandurangi, R. S.;
Katti, K. V.; Stillwell, L.; Barnes, C. L. J. Am. Chem. Soc. 1998, 120, 11364-11373. For
recent examples, see: (c) Sawayama, T.; Tsukamoto, M.; Sasagawa, T.; Nishimura, K.;
Deguchi, T.; Takeyama, K.; Hosoki, K. Chem. Pharm. Bull. 1990, 38, 110-115. (d)
Blackburn, C.; Pingali, A.; Kehoe, T.; Herman, L. W.; Wang, H., Kates, S. A. Bioorg.
Med. Chem. Lett. 1997, 7, 823-826.
139. Natoff, I. L.; Redshaw, S. Drugs of the Future 1987198719871987, 12, 475-483.
140. Flynn, G.; Beight, W. An efficient synthesis of ethyl (R)-2-Hydroxy-4-Phenylbutyrate: a
usuful intermediate in the synthesis of converting enzyme inhibitors. Tetrahedron
Letters. 1988198819881988,29, 423-426.
141. Yanagisawa, H.; Ishihara, S.; Ando, A.; Kanazaki, T.; Miyamoto, S.; Koike, H.; Lijima, Y.;
Oizumi, K.; Matsushita, Y.; Hata, T. Angiotensin-converting enzyme inhibitors:
Perhydro-1,4-Thizaepin-5-One derivatives. J. Med. Chem. 1987198719871987,30, 1984-1991.
142. Chan, S.; Kevin, K.; Lin, C.; Wong, C. Lipase-catalyzed irreversible transesterification
using enol esters: Resolution of cyanohydrins and synthesis of ethyl (R)-2-hydroxy-4-
phenyl butyrate and (S)-Propanolol. Tetrahedron Letters. 1989198919891989, 30, 1917-1920.
143. Regenye, R.; Partridge, F.; Coffen, D. Kinetic resolution of 2-substitued esters catalysed
by a lipase Ex. Pseudomonas fluorescens. J. Org. Chem. 1990199019901990,55, 812-815.
144. Baldano, E.; D`Arrigo, P.; Pedrocchi-Fantoni, G.; Rosell, C.M.; Servi, S.; Taglini, A.;
Terreni, M. PenG Acylase catalyzed resolution of Phenylacetate esters of secondary
alcohols. Tetrahedron: Asymmetry. 1993199319931993,4, 1031-1034.


















