
Contractility, Metabolism and Pharmacological Reactions...
13
Contractility, Metabolism and Pharmacological Reactions of Isolated Gas-Perfused Cat Hearts By Lloyd P. Gobel, Ivan Bihler, and Peter E. Dresel ABSTRACT Cat hearts were perfused through the aorta with substrate-free Krebs solution for 5 min, then with 5% CO 2 -95% O 3 saturated with water vapor at 37.5°C. Heart rate decreased rapidly but contractility was maintained at pre-gas levels for 3 to 4 hours, decreasing to 25% at 10 hours. Driving the preparations at 168 beats/min did not appreciably change the time course of change in contractility. The hearts developed more tension at all resting tensions and were able to per- form more work than hearts perfused with substrate-free Krebs solution, but their work capacity was less than that of hearts perfused with glucose-Krebs solution. They reacted normally to sympathomimetic amines and blocking agents. They were very sensitive to increases in perfusion pressure, developing completely re- versible arrhythmias at pressures 80 to 150$ above the control (60 mm Hg). Most hearts developed contractile altemans which usually was not accompanied by electrical alternation. Glycogen, lactate, pyruvate content decreased rapidly, the first to a residual level and the others to undetectable levels. ADDITIONAL KEY WORDS glycogen loss heart mechanical alternation prolonged survival rate Starling relationship pressure arrhythmias • As early as 1902, Magnus (1) maintained a beating cat heart for 1 hour while perfus- ing it with gaseous oxygen. Burns et al. (2) repeated this experiment and described the relationship between the perfusing pressure and the flow of gas through the coronary ar- teries. Sabiston et al. (3) and Talbert et al. (4) described a technique for gas perfusion with which electrical, but not mechanical, activity could be maintained for periods ap- proaching 8 hours. The liquid media used to perfuse the iso- From the Department of Pharmacology and Thera- peutics, University of Manitoba, Faculty of Medicine, Winnipeg, Manitoba, Canada. This work was aided by grants from the Medical Research Council of Canada, the American Heart Association and the Muscular Dystrophy Association of Canada. A preliminary report was presented to the Fed. Am. Soc. Expd. Biol. in April 1965. (Federation Proc. 24: 365, 1965.) Dr. Gabel's work was supported by a U. S. Public Health Service Training Grant (T.I. CM 1054) from the Division of General Medical Sciences. His present address is Wamer-Lambert Research Institute, Morris Plains, New Jersey. Dr. Bihler is a Medical Research Associate of the Medical Research Council of Canada. Accepted for publication September 16, 1966. lated heart perform several functions: (A) maintenance of a constant temperature and ionic environment, (B) supply of oxygen, (C) supply of energy-yielding substrates, and (D) removal of metabolic end products. Per- fusion with a gaseous medium may maintain a constant temperature and a supply of oxy- gen, but is obviously incapable of supplying or removing metabolites other than CO 2 . It is well documented (5-7) that the iso- lated heart fails rapidly when perfused with a substrate-free liquid medium. This paper describes a procedure for gas perfusion of the isolated cat heart which allows contractility to remain stable for periods exceeding 3 hours, and it compares gas- and liquid-perfused hearts with respect to contractility, perfor- mance of work and response to pharmacol- ogical agents. Methods PERFUSION SYSTEM We were unable to maintain contractility using the procedures of Burns et al. (2) or of Sabis- ton et al. (3) primarily because of difficulties in regulating the temperature of the gaseous per- fusate. An apparatus was constructed which per- mitted both close temperature control of the CircuUtion Rtinrcb. Vol. XIX. Novembtr 1966 891 by guest on July 5, 2018 http://circres.ahajournals.org/ Downloaded from
Transcript of Contractility, Metabolism and Pharmacological Reactions...

Contractility, Metabolism and PharmacologicalReactions of Isolated Gas-Perfused Cat Hearts
By Lloyd P. Gobel, Ivan Bihler, and Peter E. Dresel
ABSTRACTCat hearts were perfused through the aorta with substrate-free Krebs solution
for 5 min, then with 5% CO2-95% O3 saturated with water vapor at 37.5°C.Heart rate decreased rapidly but contractility was maintained at pre-gas levelsfor 3 to 4 hours, decreasing to 25% at 10 hours. Driving the preparations at 168beats/min did not appreciably change the time course of change in contractility.The hearts developed more tension at all resting tensions and were able to per-form more work than hearts perfused with substrate-free Krebs solution, but theirwork capacity was less than that of hearts perfused with glucose-Krebs solution.They reacted normally to sympathomimetic amines and blocking agents. Theywere very sensitive to increases in perfusion pressure, developing completely re-versible arrhythmias at pressures 80 to 150$ above the control (60 mm Hg).Most hearts developed contractile altemans which usually was not accompaniedby electrical alternation. Glycogen, lactate, pyruvate content decreased rapidly,the first to a residual level and the others to undetectable levels.
ADDITIONAL KEY WORDSglycogen loss heartmechanical alternation
prolonged survivalrate Starling relationship
pressure arrhythmias
• As early as 1902, Magnus (1) maintaineda beating cat heart for 1 hour while perfus-ing it with gaseous oxygen. Burns et al. (2)repeated this experiment and described therelationship between the perfusing pressureand the flow of gas through the coronary ar-teries. Sabiston et al. (3) and Talbert et al.(4) described a technique for gas perfusionwith which electrical, but not mechanical,activity could be maintained for periods ap-proaching 8 hours.
The liquid media used to perfuse the iso-
From the Department of Pharmacology and Thera-peutics, University of Manitoba, Faculty of Medicine,Winnipeg, Manitoba, Canada.
This work was aided by grants from the MedicalResearch Council of Canada, the American HeartAssociation and the Muscular Dystrophy Associationof Canada.
A preliminary report was presented to the Fed. Am.Soc. Expd. Biol. in April 1965. (Federation Proc.24: 365, 1965.)
Dr. Gabel's work was supported by a U. S. PublicHealth Service Training Grant (T.I. CM 1054) fromthe Division of General Medical Sciences. His presentaddress is Wamer-Lambert Research Institute, MorrisPlains, New Jersey.
Dr. Bihler is a Medical Research Associate of theMedical Research Council of Canada.
Accepted for publication September 16, 1966.
lated heart perform several functions:(A) maintenance of a constant temperatureand ionic environment, (B) supply of oxygen,(C) supply of energy-yielding substrates, and(D) removal of metabolic end products. Per-fusion with a gaseous medium may maintaina constant temperature and a supply of oxy-gen, but is obviously incapable of supplyingor removing metabolites other than CO2.
It is well documented (5-7) that the iso-lated heart fails rapidly when perfused witha substrate-free liquid medium. This paperdescribes a procedure for gas perfusion ofthe isolated cat heart which allows contractilityto remain stable for periods exceeding 3 hours,and it compares gas- and liquid-perfusedhearts with respect to contractility, perfor-mance of work and response to pharmacol-ogical agents.
MethodsPERFUSION SYSTEM
We were unable to maintain contractility usingthe procedures of Burns et al. (2) or of Sabis-ton et al. (3) primarily because of difficulties inregulating the temperature of the gaseous per-fusate. An apparatus was constructed which per-mitted both close temperature control of the
CircuUtion Rtinrcb. Vol. XIX. Novembtr 1966 891
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

892 GABEL, BIHLER, DRESEL
0LA3S WOOL
HUMIDIFIER I
no v
TEMPERATURE CONTROL
TELETHERMOMETER
«v :
— RESISTANCE WIRE
8 3
; FAN
f ,T T %
L 1m
TRAP
— :
j
1
3<
, t •i ••
• -
TRANSDUCER PR RWtRVCTRS
FIGURE 1
Apparatus for gas and liquid perfusion. For explanation see text.
humidified gas mixture at various flow rates andrapid switching from liquid to gas perfusion (Fig.1). The heart was perfused by the Langendorfftechnique from a cannula with four side armsclose to its tip. Two thermistor probes (YSI Mod-els 402 and 403) extended directly into the per-fusate flowing through side arm A. One was usedto monitor temperature, the other functioned inconjunction with a temperature regulator (YSIModel 63) which controlled the flow of currentthrough a heating coil placed around the glasstube leading to the cannula. This coil servedonly as a final temperature adjustment (see be-low) and maintained the temperature of bothliquid and gaseous perfusates at 37.5 ± 0.4°C.During gas perfusion, side arm B was connectedby a stopcock (S-3) and a trap to a gas pressureregulator (PR) constructed by immersing a glasstube into a column of water to a depdi corres-ponding to 60 mm Hg. The trap collected liquidperfusate contained in the system at the time ofchange from liquid to gas perfusion. Side arm Cwas connected to a Statham P 23D pressuretransducer (PT). A thin plastic tube used forinjection of drugs passed through side arm D andterminated in the cannula close to its point ofinsertion into the aorta.
Liquid perfusates of different composition werestored in reservoirs placed level with the heart,pressurized to 60 mm Hg with 95% O2 and 5%
CO2. The liquid passed through a water-jacketedspiral condenser which served as a heat exchang-er, and then through stopcocks S-l and S-2 be-fore reaching the cannula. Stopcock S-l selectedeither reservoir; stopcock S-2 selected either gasor liquid perfusate.
The 95% O2 and 5% CO2 mixture for perfusionwas preheated in a water-jacketed condenserand was then humidified in a water-jacketed gasscrubbing bottle containing distilled water. Itthen passed through glass wool which removedwater droplets and was led to stopcock S-2.
The heart was enclosed in a water-jacketedLucite box. High humidity was maintained inthe chamber by a fan which circulated air pastseveral vertical strips of moist filter paper. A fun-nel under the heart passed through the bottomof the box for recovery of liquid perfusate. Thewater circulating through all jacketed componentswas supplied from a constant-temperature bathat40.0±0.5°C.
In those experiments in which liquid perfusatewas used exclusively, the perfusion system waschanged to allow recirculation of the perfusate.Our apparatus was similar to that of Bleehenand Fisher (8) except that the perfusate wasrecirculated by a pump instead of an air lift andthe pressure was supplied by the equilibratinggas instead of gravity. The total volume of liquidin our system was 200 to 250 ml.
Circulation Resurcb, Vol. XIX, Novtmbtr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

THE GAS-PERFUSED CAT HEART 893
PREPARATION OF THE HEART
Kittens of either sex weighing 0.4 to 1.2 kgwere fed a diet of meat, fish and eggs ad libitumfor at least 2 days before use. The animals werekilled by a blow on the head and the hearts re-moved and placed in a beaker of cold Krebs-Hen-seleit-bicarbonate solution. Anticoagulants werenot used because of their possible effects ontissue metabolism (9).
The hearts were cleaned of extraneous tissueand a small slit was made in the walls of eachventricle. The aorta was then cannulated and theheart connected to the perfusion apparatus. A5-min period of liquid perfusion preceded gasperfusion in all experiments. Ten seconds of gasperfusion was used to clear the vasculature be-fore weighing hearts which had last been per-fused with liquid. The liquid perfusate was sub-strate-free Krebs-Henseleit solution gassed with95* O2 and 5% CO.,; it contained NaCl, 118.0IDM; KC1, 4.7 mM; KH2PO4) 1.2 mM; CaCl2 •2H.O, 2.5 mM; MgSO4, 2.4 mM and NaHCO,,,26.6 mM. Glucose (2.0 g/liter) was added onlyfor the experiments in which work capacity wasdetermined.
MEASUREMENTS
Isometric contractility was measured with aGrass FT-03 force-displacement transducer at-tached to a string sewn to the ventricular apex;resting tension was defined as the diastolic ten-sion exerted on the apex. Isotonic contractilityand external work were measured electronicallywith an apparatus in which work was estimatedfrom the lifting of imposed weights attached tothe apex of the ventricle (10).
Heart rates were determined from the record-ings of contractile force. Some hearts were drivenelectrically through small clip electrodes placedon the right atrial appendage and ventricularapex. Suprathreshold rectangular pulses (4 to8 v) of 5 msec duration were supplied by aGrass SD-5 stimulator. Electrograms were ob-tained using electrodes placed in the samepositions as the stimulating electrodes. Record-ings were made with a Grass polygraph.
Analytical Mcthodi
Only the ventricles, cut below the fat padsurrounding the circumflex artery, were used foranalysis. They were blotted with tissue paperand then dropped into liquid nitrogen. The froz-en tissue was placed in steel test tubes previ-ously cooled with liquid nitrogen and powderedby percussion. The powder was mixed by stirringand stored at — 20°C. Aliquots of tissue powderwere weighed rapidly and analyzed as follows:
Water content. This was calculated from theweight loss after drying to constant weight at80°C in vacuo. This value was used to calculateCircmUiio* Restrcb, Vol. XIX, Novemttr 1966
the dry weight of aliquots used for other analyses.Glycogen and total carbohydrate. Samples
were hydrolyzed in 501 NaOH and glycogenprecipitated with cold 80* methanol. Othersamples were homogenized in 5% trichloraceticacid to extract total tissue carbohydrate. Theglycogen precipitate and the trichloroacetic acidextract were analyzed spectrophotometrically bythe sulfuric acid method of Kemp and Kits VonHejningen (11). Results are expressed as glucoseequivalents.
Lactate and pyruvate. Cold 656 perchloric acidextracts were analyzed by enzymatic methods(12, 13). The reagents were obtained fromSigma Chemical Co., St. Louis, Mo.
Results are expressed as micromoles per gramdry weight. Data for each group of hearts areexpressed as means ± their standard errors. Thesignificance of differences between groups wasestimated by Student's t test.
DRUGS USED
Stock solutions of epinephrine bitartrate, nor-epinephrine bitartrate, isoproterenol hydrochlo-ride, pronethalol hydrochloride and cocaine hydro-chloride containing 10 mg/ml of the bases weremade in acidified 0.9% NaCl, and further dilu-tions made with the same vehicle on the day ofeach experiment; 0.05 to 0.1 ml of the appropriatedilution was made to 1 ml with Krebs-Hense-leit solution immediately before administration.In later experiments it was found that more re-producible results were obtained if the drugswere injected in a volume of 5 ml.
ResultsCONTRACTILITY AND HEART RATE
Isometric contractile force and heart rateat a constant resting tension of 10 g weremeasured in two groups of hearts. The firstwas perfused for 2 hours with recirculatingsubstrate-free Krebs-Henseleit solution (6hearts). The second group (4 hearts) wasperfused with gas for 10 hours after 5 minof perfusion with liquid. Contractile forceand rate at the end of 5 min of liquid per-fusion were taken as the initial (control) val-ues in both groups.
Figures 2A and 2B show that the changes incontractility and rate resulting from long peri-ods of perfusion were strikingly different un-der the two conditions. Figure 2A showsthat the rate of liquid-perfused hearts de-clined throughout the 2-hour period of theexperiments, reaching 73 ± 6* of the initial
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

894 GABEL, BIHLER, DRESEL
value of 163 ± 8 beats/min at the end of theexperiments. The rates of gas-perfused heartsfell rapidly during the first hour to a value of43 ± 3* of initial but declined only slowlyduring the remaining 9 hours. The contractileforce of liquid-perfused hearts declined con-tinuously throughout the experiment (Fig.2B). It was 50* of the initial value after 80min and 10* of initial after 120 min. The con-tractility of gas-perfused hearts increased sig-nificantly during the first 20 min and remainedabove the initial level for 100 min. No statis-tically significant fall below the initial level
was observed before the fourth hour of gasperfusion. The contractile force continued todecline slowly, reaching a value of 50* of ini-tial at 7 hours and of 25* of initial after 10hours, at which time the experiments wereterminated. A third group (4 hearts) was per-fused with gas for 7 hours while being drivenat a rate (168 beats/min) close to the initialrate of liquid-perfused hearts. The contractilityof these hearts was only slightly less than thatof the other gas-perfused group. The increasein contractile force above the initial value,of 75 min duration, was not statistically sig-
HEART RATE
3 4 3 6HOURS OF PERFU90N
10
PERCENTOF
CONTROL
123'
IOO'
73 '
SO-
25 '
CONTRACTILE FORCE
OAS
1M/MIN
3 4 3 6HOURS OF PERFU9ON
10
FIGURE 2
Top, time course of change in heart rate during perfusion with gas and with substrate-free Krebssolution. Bottom, time course of isometric contractile force; same hearts as above. Additionalgroup labeled 168 beats/min was driven electrically at that rate. Bars show SE; 4 hearts/group.
CircmUiion Research, Vol. XIX, Novunbtr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

THE GAS-PERFUSED CAT HEART 895
nificant; 50* of control was reached after 5hours and 25% of control after 7 hours.
In agreement with the observations of Cat-tell and Gold (14) and Luisada and Weiss(15), we observed that hearts maintained onliquid tended to shorten gradually throughoutthe period of perfusion and it was necessaryto adjust the force-displacement transducer inorder to maintain the 10 g resting tension.This did not occur in gas-perfused hearts.When gas perfusion was started after 2 hoursof liquid perfusion, diastolic tension decreasedto the control within 3 min but contractileforce increased only 20*.
Contractions of alternate strength (contrac-tile alternans) were observed in 90* of morethan 50 hearts perfused with gas. This usuallybegan during the first 15 min. The differencebetween alternate contractions, very small ini-tially, increased during the next 45 min andthen remained constant for the remainder ofthe experiment. Contractility during alternansis expressed as the arithmetic mean of thealternating beats. The curve relating contrac-tility to time for any 1 heart shows a slightdip of less than 5 min duration immediatelyafter onset of contractile alternans, followed bya continuing increase in average contractility.
This is not seen in the composite curve (Fig.2B) because of variations in the time at whichalternation began.
Electrograms were obtained from 37 heartswith contractile alternans. No electrical alter-nans was seen in 27 of these (73*). Theweaker beat in each of these hearts beganonly after relaxation from the stronger beatwas complete (Fig. 3B). The weaker beat ineach of the 10 hearts which did show electricalalternans began before relaxation was com-plete (Fig. 3A).
Many experimentally induced arrhythmiasare affected by changes in perfusion pres-sure (16, 17). We observed that increasingthe perfusion pressure from 60 to more than100 mm Hg caused multifocal ventriculararrhythmias which continued until the pres-sure was lowered (Fig. 4). In 6 gas-perfusedhearts the mean pressure threshold for theinitiation of the arrhythmia was 124 ± 9 mmHg, whereas it was 187 ± 12 mm Hg in 6liquid-perfused hearts. The contractility andelectrical activity of the latter group did notreturn to control conditions when the pres-sure was lowered. Gas-perfused hearts didnot show these irreversible changes. The pres-sure threshold for arrhythmia production could
A
^ ^ W \ f ^ ^
30-
10-0
B
" !A/V\/WWWWWWW\AAA100 -
I SEC
FIGURE 3
Contractile alternans during gas perfusion. The upper tracing is an electrogram; lower tracingshows isometric contractile force. In A, electrical alternans accompanies mechanical alternans.In B, recording from another heart shows mechanical alternans only.
CmuUlicm Rumrcb, Vol. XIX, Noptnktr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

896 GABEL, BIHLER, DRESEL
PERFUSION PRESSURE!!mm Hg
FIGURE 4
50-
43'
40
35
I - 30'
o
ra is
10
KREBS
Response of a gas-perfused heart to increased perfusion pressure. Electrogram recorded withelectrodes on right atrial appendage and on ventricular apex. The arrows indicate thresholdpressure (120 mm Hg) for induction of arrhythmia and for return to sinus rhythm on decreasingthe perfusion pressure.
min, were perfused with liquid for a 10-minperiod. The effect of graded increases in rest-ing tension on isometric developed tensionswas determined. Gas perfusion was then be-gun and the procedure repeated as soon ascontractile force became stable (5 to 15 min).Figure 5 shows the relationship of restingtension to developed tension, both expressedin terms of heart weight. This method of ex-pressing the tensions was used because themore commonly used units (grams per squaremillimeter) were obviously impractical inwhole hearts. The tension developed at anygiven resting tension was greater during gasperfusion than during liquid perfusion. Im-posed resting tensions were not allowed to ex-ceed 35 g/g heart because preliminary experi-ments showed that excessive tensions causeirreversible changes in contractility.
We have compared the external work doneby hearts perfused with gas, with substrate-free Krebs solution and with Krebs to which2 g/liter glucose had been added. Four hearts,driven at 180 beats/min, were perfused ini-tially with Krebs plus glucose, and succes-sively heavier weights suspended from theapex. Work and power output were recordedelectronically. The time required to estimatework with the series of 5 weights was lessthan 10 min. The hearts were then per-fused with substrate-free Krebs solution andthe procedure repeated. A second determin-ation during Krebs plus glucose perfusion wasdone in order to ensure that the preparationhad not been altered by the substrate-free
CircuUlicm Research, Vol XIX, Novtmber 1966
6 10 13
RESTING TENSION
20 23 30 35
g / g h«ort wt.
FIGURE 5
Relationship between resting tension and isometricdeveloped tension in hearts perfused first withKrebs solution and then with gas. Heart rate was 180beats/min. Regression lines calculated by method ofleast squares.
be determined repeatedly in the same heartand was found to remain remarkably con-stant. The atria did not appear to be affectedby the increases in perfusion pressure.
STARLING'S RELATIONSHIP
Five hearts, driven electrically at 180 beats/
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

THE GAS-PERFUSED CAT HEART 897
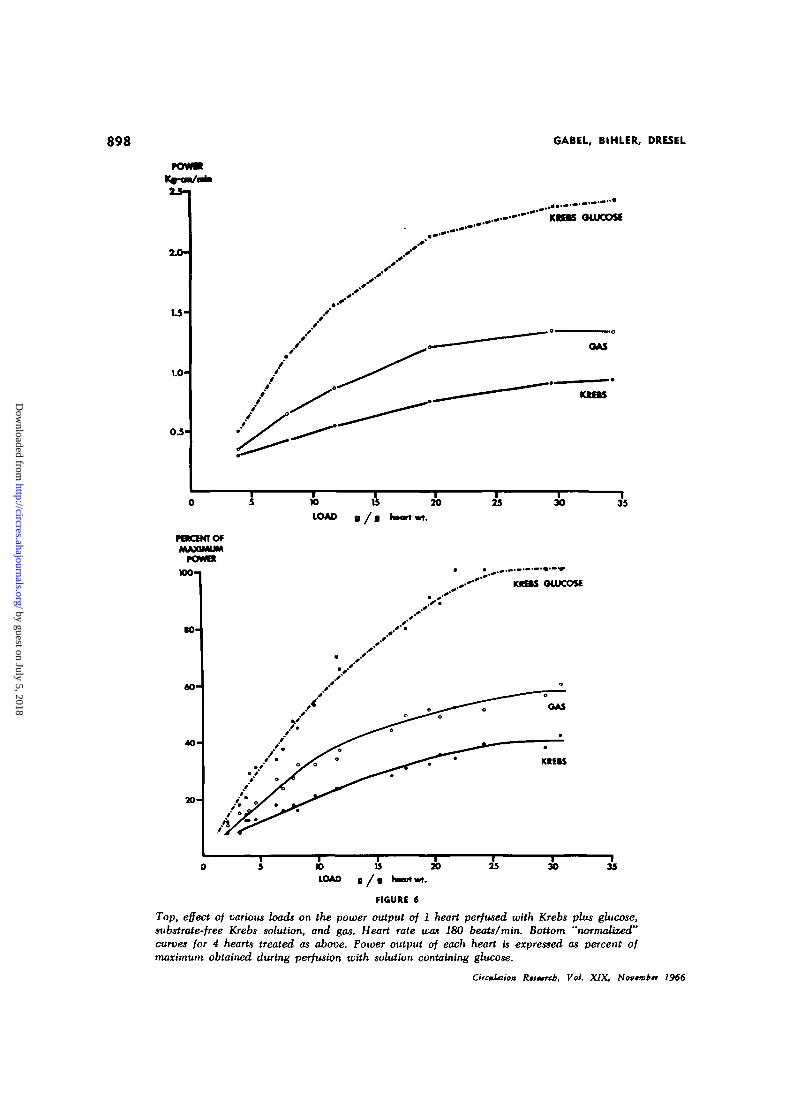
medium. The procedure was then repeatedduring gas perfusion. Figure 6A shows acharacteristic series of curves obtained in 1heart. Except at the smallest and the largestloads used, power output during gas perfusionwas approximately 1.5 times that during sub-strate-free liquid perfusion, whereas poweroutput in the presence of glucose was morethan 3 times that in its absence. The resultsfrom the entire group of preparations, whichdiffered from each other in weight, have beencombined by expressing the power output interms of the maximum power obtained duringperfusion with Rrebs plus glucose. Figure 6Bshows these "normalized" curves to have thesame quantitative relationship to each other.
METABOLIC STUDIES
We have studied the tissue content of vari-ous metabolites during gas and substrate-freeliquid perfusion. A 10-g resting tension wasapplied to the hearts during these studies andcontractility was recorded to ensure that thepreparations were similar to those used inthe physiological studies. The experimentswere terminated by quickly cutting off theventricles during continued gas perfusion, blot-ting them and freezing in liquid nitrogen.The control group of hearts was treated inthe same manner within 30 sec of removalfrom the animals. The results are summarizedin Table 1.
Table 1 shows that the water content ofthe control group was higher than that of
hearts perfused for a brief period, presumablybecause their vasculature had not been clearedof fluid. The increase in tissue water due toprolonged perfusion with liquid is well known.Tissue water did not decrease during pro-longed gas perfusion, which indicates thatadequate humidification of the perfusate andof the environment was achieved in our sys-tem.
Tissue glycogen content decreased rapidlyduring the initial 5 min of liquid perfusionand did not change greatly when liquid per-fusion was continued for 2 hours. Gas per-fusion for 5 min after 5 min of liquid per-fusion resulted in a further significant decreasein glycogen content to a level which remainedstable for the next 10 hours. Nonglycogencarbohydrate levels changed in a mannersimilar to that of glycogen. The short periodof gas perfusion produced a similar rapidfurther fall in the levels of lactate and pyru-vate which, however, continued so that theirlevels in individual hearts were no longermeasurable.
PHARMACOLOGICAL STUDIES
We determined the response of oxygen-perfused hearts to a number of drugs whichwere administered in 1 or 5 ml Krebs solution.It was essential that the solution be admin-istered into the perfusion cannula rapidlyenough to pass through the coronary vesselsas a single bolus. Slow injection causedthe formation of multiple gas-liquid inter-
TABLE 1
Effect of Perfusions on Water and Carbohydrate Contents
Perfusion
Control(0 time)
Krebs, 5 minGas, 5 minGas, 3 hoursGas, 10 hoursKrebs, 2 hours
Watercontent
%
80.9 ± 1.5
76.6 ± 0.4f76.1 ± 0.376.3 ± 0.476.8 ± 0.584.3 ± 1.9t
Glycogen/tmole«»/f
72.4 ± 3.8
23.8 ± 2.0f13.8 ± O.lt13.4 ± 0.513.6 ± 0.119.6 ± 0.7
Other carbo-hydrates
/imole»»/t
47.9 ± 4.6
19.3 ± 1.9f15.6 ± 1.59.2 ± 0.9f
10.0 ± 0.617.4 ± 1.8t
Lactatefimola/g
95.7 ± 7.6
33.2±4.1t10.6 ± 2.5t0.1*
—9.2 ± 1.9f
Pyruvate>imoles/g
0.16 ± 0.02
0.08 ± O.Olf<0.03<0.03<0.03<0.03
Means ± SE, four hearts per group; results calculated per gram dry weight of tissue.*As glucose equivalents.tSignificantly different (P < 0.05) from preceding group and control.^Pooled sample.
CircuUtio» Rtstrcb, Vol. XIX, Novtmktr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from
898 GABEL, BIHLER, DRESEL
PBtCBtTOfMJUQMUM
rovmKM-I
• 0 -
60-
40
KIBS OLUCOSf
KftftS
LOAD B / •
20 30 35
KKBS OWCOSE
KIllS
0 5 1 0 1 5 2 0 2 5 3 0 3 5LOAD g / a hnH wt.
FIGURE 6
Top, effect of various loads on the power output of 1 heart perfused with Krebs plus glucose,substrate-free Krebs solution, and gas. Heart rate was 180 beats/min. Bottom "normalized?'curves for 4 hearts treated as above. Power output of each heart is expressed as percent ofmaximum obtained during perfusion with solution containing glucose.
CircmUiUm Kutrcb, Vol. XIX. Novtmbtr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

THE GAS-PERFUSED CAT HEART 899
faces which would not pass through the heartat the usual perfusion pressure.
Injections of Krebs solution resulted in atransient (less than 20 sec) increase in con-tractility of approximately 10*. There was nochange in heart rate. Epinephxine and nor-epinephrine (0.1 to 0.5 fig) increased contrac-tility and rate by 50 to 150*. Dose-responsecurves were determined for isoproterenol in4 hearts. This amine was 10 times more po-tent than epinephxine, a dose of 0.05 \ig,producing 50% of maximal increase in contrac-tility. The durations of action of these sympa-thomimetics were not increased over thoseproduced by similar injections into liquid-perfused hearts. This is probably due to therapid decomposition of the amines at the highoxygen tensions, although uptake into tissuestorage sites cannot be discounted.
The stimulus voltage of driven preparationswas increased to 10 X threshold in order to de-termine the response of the gas-perfused heartto release of endogenous catecholamines (18).Figure 7A shows the positive inotropic re-sponse to this procedure. The magnitude andduration of the response were potentiated
by the prior administration of 1 yxg of co-caine (Fig. 7B). Pronethalol (1 to 10 /ng),administered after washing out the cocaineby brief perfusion with Krebs solution blockedthis response (Fig. 7C). Figure 7D showsthat prior treatment of an animal with reser-pine makes the gas-perfused heart unrespon-sive to increases in driving voltage.
Discussion
The question arises whether the capillarycirculation is perfused by the gas mixture orwhether the heart remains viable because oxy-gen passing through the open left ventricularcavity diffuses into the tissue. We citethe following evidence against the latter pos-sibility :
(A) Although we can observe the escapeof gas from the slit in the free wall of theleft ventricle, the resistance of the systemprovides an argument against this being theonly route of egress of gas. In a number ofexperiments the aortic valve opened sud-denly, apparently spontaneously; this re-sulted in a precipitous fall in perfusion pres-sure and a decrease of 80 to 100* in the force
50 V 5V
A
50V
B
C D
5OV 5V 50 V 5V
FIGURE 7
Effect of drugs on the contractile response of gas-perfused hearts to catecholamines releasedby supramaximal electrical stimulation. Heart rate was 180 beats/min. A = Control; B = aftercocaine (1/ig); C = cocaine washed out, treated with pronethalol (lfLg); D — as in A, but heartobtained from an animal treated with reserpine 24 hours previously.
Circulation Rtsurch, Vol. XIX, Novtmitr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

900 GABEL, BIHLER, DRESEL
of contraction of the heart. These preparationscould often be restored by gentle manipula-tion of the left ventricle. Contractility returnedto normal together with the return of per-fusion pressure. We have also found thatslitting of the ventricle is a convenient, ratherthan an essential, aspect of the preparation.
(B) Cardiogreen dye or diluted india ink(5 ml) were injected through the aortic can-nula into several hearts after 2 to 6 hoursof gas perfusion. This volume of solutionpasses through the hearts in 20 to 30 sec.Dissection showed that most or all of themuscle mass had been stained. The exact dis-tribution of dye was somewhat variable,however, small portions of the free walls ofthe ventricles sometimes showing only lightpink coloration.
(C) Although our preparation has main-tained contractility for periods longer thanthose reported by other workers, it is notunique. Dog hearts have been kept alive invitro and in situ either by orthograde or byretrograde perfusion of the coronary vascu-lature with gaseous oxygen. Camishion et al.(19) have shown that tissue oxygen tensions,measured with embedded oxygen electrodesduring retrograde perfusion, exceed those ob-served during perfusion with blood. Talbertet al. (4) showed that gas flows of 700 to1,500 ml/min at pressures of 20 to 50 mmHg are required during retrograde perfusion.Sabiston et al. (3) measured flows of 300 to1,000 ml/min at 80 to 120 mm Hg duringorthograde perfusion. These flows were ob-served under conditions where gas escape wasimpossible. We conclude that gas will flowthrough the coronary vasculature at reason-able pressures if perfusion with a salinemedium precedes gas perfusion and the for-mation of bubbles is studiously avoided. How-ever, none of the studies, including our own,presents definitive evidence that gas flow oc-curs through capillary beds.
Gas-perfused hearts beat more strongly,perform more work and fail much more slow-ly than do hearts perfused with substrate-freeKrebs solution. There are several possiblereasons for this. It is immediately apparent
that the oxygen content, but not necessarilythe Po-j of the perfusing medium, is greatly in-creased during gas perfusion. Lentini (20)reported that the optimal oxygen tension forisolated heart tissue is 700 mm Hg, a partialpressure exceeded in our liquid perfusate.Fisher and Williamson (21) and Opie et al.(22) showed that the oxygen content of liquidperfusates is sufficient to support metabolismin hypodynamic preparations. This may notbe true in hearts which are performing work.Blinks and Koch-Weser (23) concluded thatthe oxygen carrying capacity of liquid per-fusates is not sufficient to permit maximalworking capacity. Our experiments supporttheir contention by showing that gas per-fusion allowed a small increase in workcapacity. No definite conclusion can bereached until a method is found to measurethe oxygen consumption of hearts perfusedwith gas. However, maximal work productionduring oxygen perfusion was only 60$ of thatobtained when glucose was added to theliquid medium, showing clearly that the majordeterminant of work capacity is not oxygencontent. This confirms the experiments ofNeely et al. (24). The possibility that highoxygen may exert a direct pharmacologicaleffect on cardiac contractility cannot be elim-inated.
Another reason for better performance couldhave been a qualitative or quantitative changein the pattern of substrate utilization. Bothgas- and liquid-perfused hearts lost most oftheir carbohydrate stores within 15 min of iso-lation, indicating that there are no grosschanges in the rate of metabolism of thesestores. By analogy with the observations ofShipp et al. (25), it would appear probablethat gas-perfused hearts derive the energyfor contraction from the oxidation of lipids.We are now studying the lipid metabolism ofthese preparations.
The better performance of gas-perfusedhearts may be related to the unique natureof this preparation, in which the extracellularfluid does not equilibrate with a circulatingfluid but comes to at least partial equilibriumwith the intracellular space. It is possible
CirctUtiw Rutmrd, Vol. XIX, Novmttr 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

THE GAS-PERFUSED CAT HEART 901
that the accumulation in the extracellular fluidof nonvolatile substances lost from the intracel-lular space may help to maintain contractility.Adenosine and its phosphorylated deriva-tives, certain lipids, and metabolic cofactorssuch as nicotinamide have all been shownto be lost into the perfusing medium (6,26, 27). These substances are all known tohave, in higher concentrations, positive ino-tropic effects on the heart (28, 29, 30). Cate-cholamines also are lost in the perfusing me-dium (31), but our experiments showing thatinjected catecholamines act for only briefperiods and the observation that a beta-re-ceptor blocking agent has no negative inotrop-ic effect, appear to eliminate catecholaminesas essential for maintained contractility. Pre-liminary experiments have shown that a newsteady-state level of extracellular sodium andpotassium ions is achieved within 2 hoursof beginning gas perfusion. We have not asyet eliminated the possibility that the in-crease in contractility during this period isdue to the limitation on the net inward dif-fusion of sodium or outward diffusion ofpotassium ions. We believe that the observedability of the gas-perfused heart to maintaincontractility for long periods may result in aninteresting approach to the problem of cardiacfailure.
The gas-perfused heart may lend itself to avariety of other specialized studies of cardiacfunction. Drugs or other materials may beadministered with the assurance that theyand their metabolites are not being removedfrom the preparation by the perfusate. Quan-titative recovery of administered vehicle al-lows estimation of the material retained inthe tissue. We cannot be sure, however, thatgas-perfused tissue handles drugs in exactlythe same manner as do tissues bathed orperfused \vith liquid.
The absence of substrates in liquid perfu-sates lowers the threshold for induction ofarrhythmia in isolated hearts (32). The gas-perfused heart has a lower threshold than thesubstrate-free, liquid-perfused heart for thearrhythmia induced by increasing the per-fusion pressure. Pressure must exert its effect
CircmUtio* Riswcb, Vol. XIX, Novmitr 1966
via the coronary vasculature, since bothventricular cavities are open to the atmo-sphere. This supports the work of Mainwood(33) and of Panisset et al. (17). Our prepara-tion is of particular interest because pressure-induced changes in rhythm may be reproducedat will and occur at a constant threshold pres-sure. It may be of interest to study the effectsof vasoactive versus cardioactive drugs onthese arrhythmias.
The preparation may also be useful in thestudy of mechanical alternation which is notsecondary to arrhythmia or to the action ofpharmacological agents. It should be notedespecially that alternation occurred in ourpreparations during the phase of increasingtotal contractility and was maintainedthroughout the remainder of the experiment.
References1. MAGNUS, R.: Saugethierherzen bei Durchstro-
mung mit Gasen. Arch. Exptl. Path. Phar-malcol. 47: 200, 1902.
2. BURNS, B. D., ROBSON, J. G., AND SMITH, G. K.:Survival of mammalian tissues perfused withintravascular gas mixtures of oxygen and car-bon dioxide. Can. J. Biochem. Physiol. 36:499, 1958.
3. SABISTON, D. C., TALBEHT, J. L., RILEY, L. H.,JR., AND BLALOCK, A.: Maintenance of theheart by perfusion of the coronary circulationwith gaseous oxygen. Ann. Surg. 150: 361,1959.
4. TALBERT, J. L., RILEY, J. H., JR., SABISTON, D. C ,AND BLALOCK, A.: Retrograde perfusion ofthe coronary sinus with gaseous oxygen. Am.Surgeon 26: 189, 1960.
5. GARB, S., PENNA, M., AND SCHIABINE, A.: Speciesdifferences in the utilization of glucose forcontractile force by isolated perfused hearts.Am. J. Physiol. 180: 103, 1955.
6. ROBB, J. S.: Maintenance of perfused mammalianhearts. Circulation Res. 1: 184, 1953.
7. ZACHARIAH, P.: Contractility and sugar perme-ability in the perfused rat heart. J. Physiol.(London) 158:59, 1961.
8. BLEEHEN, N. M., AND FISHER, R. B.: Action ofinsulin in the isolated rat heart J. Physiol.(London) 123:260, 1954.
9. MICHAJLJK, A., AND BRACDON, J. H.: Effect ofintravenous heparin on oxidation of fat. J.LipidRes. 1: 164, 1960.
10. GABEL, L. P.: Ph. D. Thesis, Department ofPharmacology and Therapeutics, Faculty ofMedicine, University of Manitoba, 1965.
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

902 GABEL, BIHLER, DRESEL
11. KEMP, A., AND KITS VON HEIJNINGEN, A. J. M.:
A colorimetric micro-method for the deter-mination of glycogen in tissues. Biochein. J.56: 646, 1954.
12. HORN, H. D., AND BHUNS, F. H.: Bestimmung
von L ( + ) Milchsaure mit Milchsauredehy-drogenase. Biochim. Biophys. Acta 21: 378,1956.
13. SECAL, S., BLAIR, A. E., AND WYNGAARDEN,
J. B.: An enzymatic spectrophotometric methodfor the determination of pyruvic acid in blood.J. Lab. Clin. Med. 48: 137, 1956.
14. CATTELL, MCK., AND GOLD, H.: Influence of dig-
italis glucosides on the force of contraction ofmammalian cardiac muscle. J. Pharmacol. Ex-pd. Therap. 62: 116, 1938.
15. LUISADA, A. A., AND WEISS, M. P.: Experiments
on the diastolic and resting lengths of cardiacstrips. Am. J. Physiol. 176: 123, 1954.
16. DAWES, G. S.: Experimental cardiac arrhythmiasand quinidine-like drugs. Pharmacol. Rev. 4:43, 1952.
17. PANISSET, J-C., CARLE, R., AND BEAULNES,
A.: Perfusion pressure in relation to the pro-duction and cessation of experimental cardiacarrhythmias. Can. J. Physiol. PharmacoL 42:21, 1964.
18. FuRCHCOTT, R. F. , DE GUBAREFF, T. , AND GROSS-
MAN, A.: Release of autonomic mediator incardiac tissue by suprathreshold stimuli.Science 129: 328, 1959.
19. CAMISHION, R. C , DAVTES, A. L., TOKUNAGA,
K., AND SOLTT, R. W.: Retrograde perfusionof the coronary arteries with gaseous oxygenduring cardiopulmonary bypass. Surgery 59:145, 1966.
20. LENTINI, E. A.: Myocardial developed tensionand oxygen supply. Am. J. Physiol. 207:341, 1964.
21. FISHER, R. B., AND WILLIAMSON, J. R.: Oxygen
uptake of the perfused rat heart. J. Physiol.(London) 158: 86, 1961.
22. OPIE, L. H., SHTPP, J. C , EVANS, J. R., AND
LEBOEUF, B.: Metabolism of glucose-U-C14 in
perfused rat heart. Am. J. Physiol. 203: 839,1962.
23. BLINKS, J. R., AND KOCH-WESER, J.: Physical
factors in the analysis of the actions of drugon myocardial contractility. Pharmacol. Rev.15: 531, 1963.
24. NEELY, J. R., LJEBERMEISTER, H., AND MORGAN,
H. E.: Regulation of carbohydrate metabolismin working muscle. 6th Intern. Cong. Biochem.1964, p. 661.
25. SHIPP, J. C , THOMAS, J. M., AND CREVASSE, L.:
Oxidation of carbon-14-labeled endogenouslipids by isolated perfused rat heart. Science143: 371, 1964.
26. KHAIRALLAH, P. A., AND MOMMAERTS, W. F.
H. M.: Nucleotide metabolism in cardiac ac-tivity. Circulation Res. 1: 8, 1953.
27. CLARK, A. J.: Action of ions and lipoids uponthe frog's heart. J. Physiol. (London) 47:66, 1913-1914.
28. CHEVILLARD, L., AND GUERIN, J.: Action des
derives adenyliques sur le coeur isole de mam-mifere. Compt. Rend. Soc. Biol. 149: 235,1955.
29. HAJDU, S., WEISS, H., AND TITUS, E.: Isolation
of a cardiac active principle from mammaliantissue. J. Pharmacol. Exptl. Therap. 120: 99,1957.
30. CALDER, R. M.: Effects of nicotinic acid onmyocardial systole, coronary flow, and ar-rhythmias of isolated heart. Proc. Soc. Exptl.Biol. Med. 65: 76, 1947.
31. Huxovifj, S., AND MUSCHOLL, E.: Die Nor-adrenalin-Abgabe aus dem Isolierten Kani-chenherzen bei sympathischer Nervenreizungund ihre Pharmakologische Beeinflussung.Arch. Expd. Padiol. Pharmakol. 244: 81,1962.
32. TETREAULT, L., AND BEAULNES, A.: On the
mode of action of glucose in die maintenanceof ventricular fibrillation. Can. J. Biochem.Physiol. 41: 1519, 1963.
33. MAINWOOD, G. W.: Mechanism of pacemakeracceleration due to increase in intra-arterialpressure in the isolated atrium. Can. J. Bio-chem. Physiol. 40: 805, 1962.
OrcuUium Rfjtrcb, Vol. XIX, Novmbv 1966
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

LLOYD P. GABEL, IVAN BIHLER and PETER E. DRESELHearts
Contractility, Metabolism and Pharmacological Reactions of Isolated Gas-Perfused Cat
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1966 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.19.5.8911966;19:891-902Circ Res.
http://circres.ahajournals.org/content/19/5/891World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 5, 2018http://circres.ahajournals.org/
Dow
nloaded from


















