
Construction an Improved D-Arabinose Pathway in ...jb.asm.org/content/165/3/704.full.pdf ·...
6
Vol. 165, No. 3 JOURNAL OF BACTERIOLOGY, Mar. 1986, p. 704-709 0021-9193/86/030704-06$02.00/0 Copyright © 1986, American Society for Microbiology Construction of an Improved D-Arabinose Pathway in Escherichia coli K-12 JOANNE M. BARTKUSt AND ROBERT P. MORTLOCK* Department of Microbiology, New York State College of Agriculture and Life Sciences, Cornell University, Ithaca, New York 14853 Received 26 August 1985/Accepted 27 November 1985 A ribitol catabolic, pathway was transduced into Escherchia coli K-12 in an effort to determine whether the ribitol pathway would confer an advantage to D-arabinose-positive mutants growing on D-arabinose as the sole carbon source. Competition studies in chemostats showed that ribitol-positive strains, with a selection coefficient of 9%/h, have a significant competitive advantage over ribitol-negative strains. Ribitol-positive strains grown in batch culture also exhibited a shorter lag period than did ribitol-negative strains when transferred from glucose to D-arabinose. Repeated transfer of a ribitol-positive strain of E. coli K-12 on D-arabinose yielded a strain with further improved growth on D-arabinose. This "evolved" strain was found to constitutively synthesize L-fucose permease, isomerase, and kinase but had lost the ability to grow on L-fucose, apparently owing to the loss of a functional aldolase. This constitutive mutation is not linked to the fucose gene cluster and may be similar to an unlinked constitutive mutation described by Chen et al. (J. Bacteriol. 159:725-729, 1984). Although the pentose D-arabinose is not usually metabo- lized by enteric bacteria, mutants which can utilize D- arabinose as a sole source of carbon and energy can be readily isolated. Two types of regulatory mutations have been found which enable cells to grow on D-arabinose. In the first type the enzymes of the L-fucose pathway are constitu- tively synthesized, while in the second type the fucose enzymes are induced by an intermediate of D-arabinose metabolism. All of the D-arabinose-positive isolates of Kleb- siella pneumoniae PRL-R3 and about half of the D- arabinose-positive isolates of Klebsiella aerogenes W-70 exhibit the constitutive phenotype. The remaining half of the D-arabrnose-positive isolates of K. aerogenes exhibit the inducible phenotype, but only the inducible phenotype has been found in D-arabinose-positive isolates of Escherichia coli K-12 (15, 17, 23). D-Arabinose is transported into D-arabinose-positive mu- tants via L-fucose permease and isomerized to D-ribulose by L-fucose isomerase (EC 5.3.1.3). The fate of the D-ribulose is determined by whether or not the cells contain a ribitol catabolic pathway. D-Ribulose is an intermediate and in- ducer of the ribitol pathway, and those cells which are induced for the enzymes of the ribitol pathway use D- ribulokinase to phosphorylate D-ribulose in the C-5 position, forming D-ribulose-5-phosphate. Further metabolism pro- ceeds through the pentose phosphate pathway (Fig. 1). Both K. pneumoniae and K. aerogenes have been shown to metabolize D-arabinose in this fashion (17, 23). In cells not containing a ribitol pathway, as is the case in E. coli K-12, the D-ribulose formed by the isomerization of D-arabinose is phosphorylated in the C-1 position by L-fuculokinase (EC 2.7.1.52), and the resulting D-ribulose-1-phosphate is cleaved by the action of L-fuculose-l-phosphate aldolase (EC 4.1.2.17) to yield dihydroxyacetone phosphate and glycoaldehyde (15) (Fig. 1). Although E. coli B, like E. coli K-12, does not contain a * Corresponding author. t Present address: Department of Biochemistry, Molecular and Cell Biology, Cornell University, Ithaca, NY 14853. ribitol catabolic pathway (18), D-arabinose-utllizing strains of E. coli B have been found which phosphorylate D-ribulose in the C-5 position by the action of a D-ribulokinase (EC 2.7.1.47) (3, 16). The origin of this D-ribulokinase activity has not been determined. In addition, E. coli B is unable to grow on L-fucose owing to the lack of a functional L- fuculose-1-phosphate aldolase, and utilization of D- arabinose must occur via the pentose phosphate pathway as is seen in Klebsiella species (3, 11). Because the D-ribulose formed by isomerization of D- arabinose is a natural intermediate of the ribitol pathway, we predict that the presence of a ribitol catabolic pathway would confer an advantage to E. coli K-12 when grown on D-arabinose, and we also predict that there would be a bias toward evolution of a D-arabinose pathway which utilized the D-ribulokinase of the ribitol pathway rather than the L-fuculokinase of the fucose pathway. To test these predic- tions, ribitol catabolic genes were transduced into a D- arabinose-positive isolate of E. coli K-12. We were able to demonstrate the advantage of the ribitol-positive strain over the ribitol-negative strain in a chemostat with D-arabinose as the carbon source. We then selected for further improve- ment of growth of the ribitol-positive E. coli K-12 strain on D-arabinose and examined the resultant strain to determine the nature of the "evolved" pathway. Our observations led us to make predictions on further improvements that could be made on this evolved pathway. MATERIALS AND METHODS Bacterial strains and culture conditions. The bacterial strains used are listed in Table 1. Bacteriophage P1 was prepared by heat induction of SG404 (19). JMFTC-1 was isolated by random TnJO mutagenesis followed by selection for an insertion near the fucose gene cluster (2, 8). Strain JM1112 was isolated by plating JM1000, an E. coli K-12 strain which was made a proline auxotroph and is resistant to rifampin, on mineral salts agar containing 0.4% D-arabinose as the sole carbon source. Strain JM2112 was constructed by transduction of the ribitol-D-arabitol genes of E. coli K20 into JM1112. Strain JM2110 is a D-arabinose-positive isolate of E. 704 on July 1, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of Construction an Improved D-Arabinose Pathway in ...jb.asm.org/content/165/3/704.full.pdf ·...

Vol. 165, No. 3JOURNAL OF BACTERIOLOGY, Mar. 1986, p. 704-7090021-9193/86/030704-06$02.00/0Copyright © 1986, American Society for Microbiology
Construction of an Improved D-Arabinose Pathway inEscherichia coli K-12
JOANNE M. BARTKUSt AND ROBERT P. MORTLOCK*Department ofMicrobiology, New York State College ofAgriculture and Life Sciences, Cornell University,
Ithaca, New York 14853
Received 26 August 1985/Accepted 27 November 1985
A ribitol catabolic, pathway was transduced into Escherchia coli K-12 in an effort to determine whether theribitol pathway would confer an advantage to D-arabinose-positive mutants growing on D-arabinose as the solecarbon source. Competition studies in chemostats showed that ribitol-positive strains, with a selectioncoefficient of 9%/h, have a significant competitive advantage over ribitol-negative strains. Ribitol-positivestrains grown in batch culture also exhibited a shorter lag period than did ribitol-negative strains whentransferred from glucose to D-arabinose. Repeated transfer of a ribitol-positive strain of E. coli K-12 onD-arabinose yielded a strain with further improved growth on D-arabinose. This "evolved" strain was foundto constitutively synthesize L-fucose permease, isomerase, and kinase but had lost the ability to grow onL-fucose, apparently owing to the loss of a functional aldolase. This constitutive mutation is not linked to thefucose gene cluster and may be similar to an unlinked constitutive mutation described by Chen et al. (J.Bacteriol. 159:725-729, 1984).
Although the pentose D-arabinose is not usually metabo-lized by enteric bacteria, mutants which can utilize D-arabinose as a sole source of carbon and energy can bereadily isolated. Two types of regulatory mutations havebeen found which enable cells to grow on D-arabinose. In thefirst type the enzymes of the L-fucose pathway are constitu-tively synthesized, while in the second type the fucoseenzymes are induced by an intermediate of D-arabinosemetabolism. All of the D-arabinose-positive isolates of Kleb-siella pneumoniae PRL-R3 and about half of the D-arabinose-positive isolates of Klebsiella aerogenes W-70exhibit the constitutive phenotype. The remaining half of theD-arabrnose-positive isolates of K. aerogenes exhibit theinducible phenotype, but only the inducible phenotype hasbeen found in D-arabinose-positive isolates of Escherichiacoli K-12 (15, 17, 23).
D-Arabinose is transported into D-arabinose-positive mu-tants via L-fucose permease and isomerized to D-ribulose byL-fucose isomerase (EC 5.3.1.3). The fate of the D-ribulose isdetermined by whether or not the cells contain a ribitolcatabolic pathway. D-Ribulose is an intermediate and in-ducer of the ribitol pathway, and those cells which areinduced for the enzymes of the ribitol pathway use D-ribulokinase to phosphorylate D-ribulose in the C-5 position,forming D-ribulose-5-phosphate. Further metabolism pro-ceeds through the pentose phosphate pathway (Fig. 1). BothK. pneumoniae and K. aerogenes have been shown tometabolize D-arabinose in this fashion (17, 23). In cells notcontaining a ribitol pathway, as is the case in E. coli K-12,the D-ribulose formed by the isomerization of D-arabinose isphosphorylated in the C-1 position by L-fuculokinase (EC2.7.1.52), and the resulting D-ribulose-1-phosphate iscleaved by the action of L-fuculose-l-phosphate aldolase(EC 4.1.2.17) to yield dihydroxyacetone phosphate andglycoaldehyde (15) (Fig. 1).Although E. coli B, like E. coli K-12, does not contain a
* Corresponding author.t Present address: Department of Biochemistry, Molecular and
Cell Biology, Cornell University, Ithaca, NY 14853.
ribitol catabolic pathway (18), D-arabinose-utllizing strainsof E. coli B have been found which phosphorylate D-ribulosein the C-5 position by the action of a D-ribulokinase (EC2.7.1.47) (3, 16). The origin of this D-ribulokinase activityhas not been determined. In addition, E. coli B is unable togrow on L-fucose owing to the lack of a functional L-fuculose-1-phosphate aldolase, and utilization of D-arabinose must occur via the pentose phosphate pathway asis seen in Klebsiella species (3, 11).Because the D-ribulose formed by isomerization of D-
arabinose is a natural intermediate of the ribitol pathway, wepredict that the presence of a ribitol catabolic pathwaywould confer an advantage to E. coli K-12 when grown onD-arabinose, and we also predict that there would be a biastoward evolution of a D-arabinose pathway which utilizedthe D-ribulokinase of the ribitol pathway rather than theL-fuculokinase of the fucose pathway. To test these predic-tions, ribitol catabolic genes were transduced into a D-arabinose-positive isolate of E. coli K-12. We were able todemonstrate the advantage of the ribitol-positive strain overthe ribitol-negative strain in a chemostat with D-arabinose asthe carbon source. We then selected for further improve-ment of growth of the ribitol-positive E. coli K-12 strain onD-arabinose and examined the resultant strain to determinethe nature of the "evolved" pathway. Our observations ledus to make predictions on further improvements that couldbe made on this evolved pathway.
MATERIALS AND METHODS
Bacterial strains and culture conditions. The bacterialstrains used are listed in Table 1. Bacteriophage P1 wasprepared by heat induction of SG404 (19). JMFTC-1 wasisolated by random TnJO mutagenesis followed by selectionfor an insertion near the fucose gene cluster (2, 8). StrainJM1112 was isolated by plating JM1000, an E. coli K-12 strainwhich was made a proline auxotroph and is resistant torifampin, on mineral salts agar containing 0.4% D-arabinoseas the sole carbon source. Strain JM2112 was constructed bytransduction of the ribitol-D-arabitol genes ofE. coli K20 intoJM1112. Strain JM2110 is a D-arabinose-positive isolate ofE.
704
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
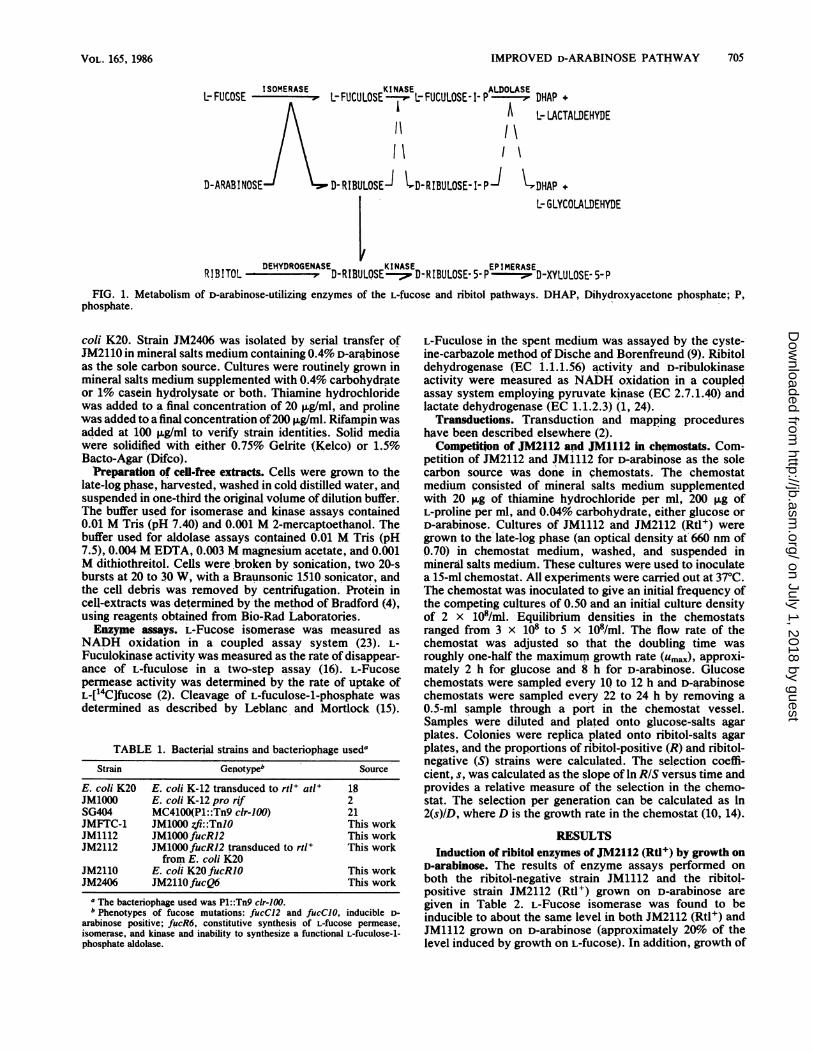
IMPROVED D-ARABINOSE PATHWAY 705
ISOMERASE KINASE ALDOLASEL-FUCOSE - ~ L-FUCULOSE--" L-FUCULOSE-1-P----7DHAP +
L-LACTALDEHYDE~~~~~~~~~~~~I\11\D-ARABINOSE D-RIBULOSEJ tD-RIBULOSE-I-Pj V,DHAP +
KINASE EPIMERASEL- GLYCOLALDEHYDE
RIBITOL - D D-RIBULOSE--- D-RIBULOSE-5-P-------s'D-XYLULOSE- 5-PFIG. 1. Metabolism of D-arabinose-utilizing enzymes of the L-fucose and ribitol pathways. DHAP, Dihydroxyacetone phosphate; P,
phosphate.
coli K20. Strain JM2406 was isolated by serial transfer ofJM2110 in mineral salts medium containing 0.4% D-arabinoseas the sole carbon source. Cultures were routinely grown inmineral salts medium supplemented with 0.4% carbohydrateor 1% casein hydrolysate or both. Thiamine hydrochloridewas added to a final concentration of 20 ,ug/ml, and prolinewas added to a final concentration of200 pug/ml. Rifampin wasadded at 100 ,ug/mnl to verify strain identities. Solid mediawere solidified with either 0.75% Gelrite (Kelco) or 1.5%Bacto-Agar (Difco).Preparation of cell-free extracts Cells were grown to the
late-log phase, harvested, washed in cold distilled water, andsuspended in one-third the original volume of dilution buffer.The buffer used for isomerase and kinase assays contained0.01 M Tris (pH 7.40) and 0.001 M 2-mercaptoethanol. Thebuffer used for aldolase assays contained 0.01 M Tris (pH7.5), 0.004 M EDTA, 0.003 M magnesium acetate, and 0.001M dithiothreitol. Cells were broken by sonication, two 20-sbursts at 20 to 30 W, with a Braunsonic 1510 sonicator, andthe cell debris was remoyed by centrifugation. Protein incell-extracts was determined by'the method of Bradford (4),using reagents obtained fom Bio-Rad Laboratories.Enzyme assays. L-Fucose isomerase was measured as
NADH oxidation in a coupled assay system (23). L-Fuculokinase activity was measured as the rate of disappear-ance of L-fuculose in a two-step assay (16). L-Fucosepermease activity was determined by the rate of uptake ofL-["4C]fucose (2). Cleavage of L-fuculose-1-phosphate wasdetermined as described by Leblanc and Mortlock (15).
TABLE 1. Bacterial strains and bacteriophage useda
Strain Gepiotypeb Source
E. coli K20 E. coli K-12 transduced to rtl+ atl+ 18JM1000 E. coli K-12 pro rif 2SG404 MC4100(P1::Tn9 cl-400) 21JMFTC-1 JM1000 zfi::TnlO This workJM1112 JMlOOOfucR12 This workJM2112 JMlOOOfucRl2 transduced to rtl+ This work
from E. coli K20JM2110 E. coli K20 fucR10 This workJM2406 JM2110fucQ6 This work
a The bacteriophage used was P1::Tn9 clr-100.b Phenotypes of fucose mutations: fucCi2 and fucCiO, inducible D-
arabinose positive; fucR6, constitutive synthesis of L-fucose permease,isomerase, and kinase and inability to synthesize a functional L-fuculose-l-phosphate aldolase.
L-Fuculose in the spent medium was assayed by the cyste-ine-carbazole method of Dische and Borenfreund (9). Ribitoldehydrogenase (EC 1.1.1.56) activity and 'D-ribulokinaseactivity were measured as NADH oxidation in a coupledassay system employing pyruvate kinase (EC 2.7.1.40) andlactate dehydrogenase (EC 1.1.2.3) (1, 24).
Transductions. Transduction and mapping procedureshave been described elsewhere (2).
Competition of JM2112 and JM1112 in ch,emostats. Com-petition of JM2112 and JM1112 for D-arabinose as the solecarbon source was done in chemostats. The chemostatmedium consisted of mineral salts medium supplementedwith 20 pg of thiamine hydrochloride per ml, 200 ,ug ofL-proline per ml, and 0.04% carbohydrate, either glucose orD-arabinose. Cultures of JM1112 and JM2112 (Rtl+) weregrown to the late-log phase (an optical density at' 660 nm of0.70) in chemostat medium, washed, and suspended inmineral salts medium. These cultures were used to inoculatea 15-ml chemostat. All experiments were carried out at 370C.The chemnostat was inoculated to give an initial frequency ofthe competing cultures of 0.50 and an initial culture densityof 2 x 108/ml. Equilibrium densities in the chemostatsranged from 3 x 108 to 5 x 108/ml. The flow rate of thechemostat was adjusted so that the doubling time wasroughly one-half the maximum growth rate (umr,), approxi-mately 2 h for glucose and 8 h for D-arabinose. Glucosechemostats were sampled every 10 to 12 h and D-arabinosechemostats were sampled every 22 to 24 h by removing a0.5-ml sample through a port in the chemostat vessel.Samples were diluted and plated onto glucose-salts agarplates. Colonies were replica plated onto ribitol-salts agarplates, and the proportions of ribitol-positive (R) and ribitol-negative (S) strains were calculated. The selection coeffi-cient, s, was calculated as the slope of ln RIS versus time andprovides a relative measure of the selection in the chemo-stat. The selection per generation can be calculated as ln2(s)/D, where D is the growth rate in the chemostat (10, 14).
RESULTSInduction of ribitol enzymes of JM2112 (Rtl+) by growth on
D-arabinose. The results of enzyme assays performed onboth the ribitol-negative strain JM1112 and the ribitol-positive strain JM2112 (Rtl+) grown on D-arabinose aregiven in Table 2. L-Fucose isomerase was found to beinducible to about the same level in both JM2112 (Rtl+) andJM1112 grown on D-arabinose (approximately 20% of thelevel in'duced by growth on L-fucose). In addition, growth of
VOL. 165, 1986
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

706 BARTKUS AND MORTLOCK
JM2112 (Rtl+) on D-arabinose resulted in the induction ofribitol dehydrogenase and D-ribulokinase to near wild-typeinduced levels. These data indicate that the D-ribuloseformed by the isomerization of D-arabinose was inducing theribitol catabolic enzymes and that at least some of thisD-ribulose was metabolized through the kinase of the ribitolpathway and phosphorylated in the C-5 position. That therewas no difference in the levels of L-fucose isomerase in thesestrains indicates that enough D-ribulose was beingphosphorylated in the C-1 position to keep the fucoseenzymes induced in JM2112 (Rtl+).
Effect of ribitol pathway on growth of JM2112 (Rtl+) onD-arabinose in batch culture. The growth characteristics ofJM1112 and JM2112 (Rtl') with D-arabinose as the solecarbon source were determined. For measurement ofgrowthrates, cells were grown in 250-ml sidearm flasks containing20 ml of mineral salts medium supplemented with D-arabinose. Concentrations of D-arabinose used to measuregrowth rates ranged from 0.04 to 0.4%. To determine induc-tion lags, the inoculum was grown in mineral salts mediumcontaining glucose, washed, and suspended in D-arabinose-salts medium. We defined induction lag as the length of timeelapsed before growth of the glucose-grown inoculum can bedetected in D-arabinose medium. Because D-ribulose is anintermediate of the ribitol pathway and is the natural sub-strate of the kinase of that pathway, we reasoned that growthof JM2112 (Rtl+) on D-arabinose should be more efficientthan growth of JM1112 on D-arabinose. Although we coulddetect no improvement in the growth rate of JM2112 (Rtl+)at any concentration of D-arabinose tested, each strainhaving a doubling time of 4.2 h, we did observe an approx-imately fivefold decrease in the length of time requiredbefore growth of unadapted cells could begin on 0.4%D-arabinose, 10 to 15 h for JM1112 versus 2 to 4 h forJM2112. The presence of the ribitol pathway in JM2112(Rtl') eliminates the need to metabolize D-arabinose throughthe aldolase of the fucose pathway, and we believe that thiswas responsible for the short induction lag observed in thisstrain. Since L-fuculose-1-phosphate aldolase has beenshown to be regulated separately from the early fucoseenzymes (12), we believe the length of the induction lag(approximately 12 h) seen for JM1112 may have been due topoor induction of the aldolase in D-arabinose-grown cellsrelative to the induction of the early enzymes, although wehave no direct evidence to support this idea.
Competitive advantage of JM2112 (Rtl+) over JM1112 onD-arabinose. Figure 2 shows the results of competition of
TABLE 2. Comparison of enzymes induced by growth of JM1112and JM2112 on D-arabinose
Strain Growth Activitybsubstratea RDH DRK LFI
JM1112 (Rtl-) CH <0.001 <0.005 <0.005CHF <0.001 <0.005 0.72D-ARA <0.001 <0.005 0.20
JM2112 (Rtl+) CH 0.10 <0.005 0.02CHF 0.10 <0.005 1.08D-ARA 2.56 0.38 0.20
a CH, Casein hydrolysate; CHF, casein hydrolysate plus L-fuicose; D-ARA,D-arabinose.
b Specific activities expressed as micromoles ofNADH oxidized per minuteper milligram of protein. RDH, Ribitol dehydrogenase; DRK, D-ribulokinase;LFI, L-fUCose isomerase.
5
1
10 20 30 40 50 60 70
Time in Hours
FIG. 2.-/Competition of JM1112 and JM2112 in chemostats. Theselection coefficient, s, is the slope of In RIS versus time, where R isthe proportion of ribitol-positive cells and S is the proportion ofribitol-negative cells. For D-arabinose, s = 0.094, and for D-gIucose,s = 0.006. Symbols: 0, D-arabinose as the limiting carbon source;U, D-glucose as the limiting carbon source.
JM1112 and JM2112 (Rtl+) in chemostats containing 0.04%D-arabinose or 0.04% D-glucose as the limiting carbonsource. Under the conditions employed, the limit of resolu-tion is s = 0.005 or 0.5% selection per h (14). A positive slopeindicates selection in favor of JM2112 (Rtl+), the ribitol-positive strain. The selection coefficient of 0.094 ± 0.002(dilution rate, D = 0.10) calculated for the D-arabinosechemostat reveals JM2112 (Rtl+) to be at a distinct advan-tage over JM1112, while there was no significant selection ineither direction in the glucose chemostat (s = 0.006 + 0.003;D = 0.31). In experiments with appropriately marked batchcultures, we could find no evidence for genetic exchangebetween JM2112 (Rtl+) and JM1112. Since JM1112 andJM2112 (Rtl+) are isogenic except for the ribitol catabolicgenes, we concluded that the selective advantage of JM2112(Rtl+) on D-arabinose was due to metabolism of D-ribulosethrough the ribitol pathway as opposed to metabolismthrough the fucose pathway.
Isolation of a strain showing improved growth on D-arabinose. After approximately 15 serial transfers of theribitol-positive strain JM2110 in D-arabinose-mineral saltsmedium (300 generations), a strain was isolated which ex-hibited improved growth on D-arabinose. This strain, desig-nated JM2406, was found to have a growth rate on D-arabinose which was 56% faster than the growth rate ofJM2110 on D-arabinose, with a doubling time of 3 h forJM2406 versus 4.6 h for JM2110. In addition, the 2- to 4-h lagobserved when JM2110 (Rtl+) was transferred from glucoseto D-arabinose was shortened in JM2406 to less than 0.5 h.This improved growth ofJM2406 on D-arabinose was accom-panied by the loss of the ability of this strain to grow onL-fucose; and addition of L-fucose to an exponentially grow-
J. BACTERIOL.
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IMPROVED D-ARABINOSE PATHWAY 707
/ I
2 4 6 8 10
Time in Hours
FIG. 3. Fucose toxicity and excretion of L-fuculose in strain
JM2406. L-Fucose was added at the arrow to one of two parallel
cultures growing on casein hydrolysate, and the culture supernatant
was assayed for the presence of L-fuculose. Symbols: *, JM2406
growing on casein hydrolysate; *, JM2406 growing on casein
hydrolysate, L-fucose added at arrow; 0, L-fuculose secreted into
the medium by the culture growing in the presence of L-fucose
(micrograms of L-fuculose per 50 Ill of culture supernatant). OD66,Optical density at 660 nm.
ing culture of JM2406 resulted in the cessation of growth and
in excretion of L-fuculose into the medium (Fig. 3). This
fucose toxicity and excretion of fuculose suggested a strain
containing a nonfunctional L-fuculose-1-phosphate aldolase.
Comparison of L-fucose and ribitol enzymes in JM2l110 and
JM2406. Levels of induced and uninduced L-fucose and
ribitol enzymes in JM2110 and JM2406 are shown in Table 3.The ribitol enzymes of both JM2110 and JM2406 wereinduced by growth on D-arabinose, indicating that at leastpart of the D-ribulose formed as a result of the isomerizationof D-arabinose by L-fucose isomerase is being metabolizedvia the pentose phosphate pathway. As expected, JM2406lacked a functional L-fuculose-1-phosphate aldolase, whichapparently accounts for the inability of this strain to grow onL-fucose. Surprisingly, JM2406 was also found to synthesizeconstitutively the permease, isomerase, and kinase of theL-fucose pathway, possibly explaining the absence of a lagwhen JM2406 is transferred to D-arabinose from glucose.Although the levels of the constitutive fucose enzymes inJM2406 are low, these enzymes can be induced to higherlevels in the presence of L-fucose.The constitutive mutation in JM2406 is unlinked to the
L-fucose genes. Bacteriophage P1 grown on JMFTC-1 wasused in two-factor crosses. The transposon insertionzpfi::TnJO was used as the selected marker, and tetracycline-resistant transductants were scored for acquisition of awild-type fucose phenotype. Both the aldolase-negative andthe constitutive mutation were found to be less than 1%cotransducible with TnJO. All transductants which regainedthe L-fuculose-l-phosphate aldolase lost the constitutivemutation as well. The marker zfi::TnJO is 40% cotransduc-ible with the fucose gene cluster (2), and a mutation in thealdolase structural gene should map within that region (22).Our data indicate that the nonfunctional aldolase is not dueto a mutation in the structural gene for the aldolase. Itappears that the nonfunctional, or more likely noninducible,aldolase and the constitutive synthesis of the L-fucosepermease, isomerase, and kinase are due to a single muta-tion, possibly in a regulatory element not yet identified aspart of the L-fucose regulon. We designated this new fucoseallele fucQ, and the mutation in this allele resulting in theconstitutive synthesis of the permease, isomerase, andkinase of the fucose pathway was designated fucQ6.
DISCUSSIONAlthough we could not detect a growth rate advantage of
JM2112 (Rtl+), a ribitol-positive E. coli K-12 strain, in batchcultures growth on D-arabinose, the chemostat provides amuch more sensitive means to measure small differences ingrowth rates. In D-arabinose chemostats, there was a signif-icant selection (9.4%/h) for the ribitol-positive strain over theribitol-negative strain. This, coupled with the decreased lagexhibited by the ribitol-positive strain transferred from glu-
TABLE 3. Induction of L-fucose and ribitol enzymes in JM2110 and JM2406
Strain Growth Activity of fucose enzymesb Activity of ribitol enzymescStralnsubstrate" LFPd LFI' LFKf LFAd RDHd DRKd
JM2110 (fucRl0) CH 0.02 0.02 0.05 0.06 0.10 NDCHF 12.80 1.08 0.77 0.20 0.10 NDD-ARA NDi 0.15 0.20 ND 3.48 ND
JM2406 (fucQ6) CH 1.11 0.26 0.28 <0.005 0.10 <0.005CHF ND 1.17 0.63 <0.005 0.10 <0.005D-ARA ND 0.26 0.25 ND 2.41 0.38
a See Table 2, footnote a.b LFP, L-Fucose permease; LFI, L-fucose isomerase; LFK, L-fuculokinase; LFA, L-fuculose-l-phosphate aldolase.c RDH, Ribitol dehydrogenase; DRK, D-ribulokinase.d Expressed as nanomoles of L-[14C]fucose transported per minute per milligram (dry weight).e Expressed as micromoles of NADH oxidized per minute per milligram of protein.f Expressed as micromoles of L-fuculose-l-phosphate formed per minute per milligram of protein.8 ND, Not determined.
VOL. 165, 1986
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

708 BARTKUS AND MORTLOCK
cose to D-arabinose, indicates to us that the presence of aribitol pathway does indeed confer a significant advantage toa cell confronted with D-arabinose as a carbon source.Repeated transfer of a ribitol-positive transductant of E.
coli K-12 on D-arabinose resulted in the isolation of strainJM2406 which showed improved growth on D-arabinose.Since the presence of a ribitol pathway was found to beadvantageous to a cell growing on D-arabinose, we predictedthat the evolution of a pathway for D-arabinose in a ribitol-positive E. coli strain would favor the utilization of the ribitolpathway, with the D-ribulose formed by isomerization ofD-arabinose being phosphorylated in the C-5 position. In-deed, the D-arabinose pathway which evolved in JM2406routes the D-ribulose through the ribitol catabolic pathwayresulting in improved growth on D-arabinose. This improvedgrowth of JM2406 with respect to its parent strain JM2110does not appear to be due to increased levels of the fucose orribitol enzymes, and, in fact, although growth of JM2406 hasbeen improved, this improvement was attained at the ex-pense of the ability to utilize L-fucose. The inability ofJM2406 to grow on L-fucose appears to be due to theinability to synthesize a functional L-fuculose-1-phosphatealdolase. It is possible that the combination of constitutivesynthesis of the fucose enzymes and the loss of the aldolaseactivity allows all of the D-ribulose formed by the isomeriza-tion of D-arabinose to be phosphorylated in the C-5 position,whereas in the parent strain, JM2110, a portion of theribulose must be phosphorylated in the C-1 position to keepthe fucose enzymes induced. It is conceivable that thiswould result in the improved growth of JM2406 on D-arabinose. This evolved pathway in JM2406 resembles theD-arabinose pathway of E. coli B, in which D-ribulose isphosphorylated in the C-5 position and L-fuculose-1-phosphate aldolase is not present (3, 11, 16). A survey of anumber of E. coli B strains has failed to produce a strainwhich is wild-type for the ability to utilize L-fucose (E.Elsinghorst, personnal communication), and it is not clearhow the D-arabinose pathway evolved in E. coli B.Although E. coli B appears to contain a true D-
ribulokinase (3), E. coli B cannot utilize ribitol (18). Sinceribitol dehydrogenase is not necessary for D-arabinose utili-zation, it is tempting to speculate that elimination of thisunnecessary activity would be a step in the evolution of theD-arabinose pathway of JM2406 and may already haveoccurred in E. coli B.Another predicted improvement of the D-arabinose path-
way of JM2406 would be alteration of the positive regulatorof the fucose enzymes to accept either D-arabinose orD-ribulose as an inducer. The apparent inducer of the fucoseenzymes in both K. aerogenes and E. coli is L-fuculose-1-phosphate, and the alternate inducer has been shown inKlebsiella species to be the D-ribulose-l-phosphate formedby the phosphorylation of D-ribulose by L-fuculokinase (2,23). Strain JM2406 appears to have taken a step in thisdirection by altering the regulation of the L-fucose permease,isomerase, and kinase so that they are constitutively synthe-sized, thereby making L-fuculokinase a dispensible enzymeactivity.Although constitutive mutants of the fucose enzymes are
readily isolated in Klebsiella species by selection on D-arabinose, such mutations have not been observed in ribitol-negative strains of E. coli K-12 grown on D-arabinose.Constitutive mutants of the fucose enzymes in E. coli K-12have been isolated by selection for improved growth onL-1,2-propanediol (12). Such selection results in the consti-tutive synthesis of L-fuculose-1-phosphate aldolase and in
the noninducibility of L-fucose permease, isomerase, andkinase (7, 12). Fucose-positive revertants of these aldolase-constitutive mutants constitutively synthesize L-fucosepermease, isomerase, and kinase as well (13). The mutationwhich allows the constitutive synthesis of these early fucoseenzymes has been termed a suppressor mutation because itis not linked to the fucose gene cluster (5, 6). We believe thatthe constitutive mutationfucQ6 in JM2406 is similar to thissuppressor mutation because it results in constitutive syn-thesis of the early fucose enzymes and is unlinked to thefucose gene cluster. We have not yet determined whetherthe apparent noninducibility of the aldolase is also a result ofthe fucQ6 mutation or is a result of a second mutation closeto fucQ6, although indirect evidence indicates that thesephenotypes are due to a single mutation. If the constitutivesynthesis of the L-fucose permease, isomerase, and kinaseand the noninducible aldolase are indeed due to a singlemutation, it may well be that this suppressor locus actuallycodes for an as yet unidentified regulator of the fucoseenzymes. We have isolated by a different method a strain,JM3406, which constitutively synthesizes L-fucose per-mease, isomerase, and kinase (2). The constitutive mutationin JM3406, fucR6, maps within the fucose gene cluster andhas no apparent effect on the ability of the cell to induceL-fuculose-1-phosphate aldolase (2). If constitutive synthesisof the fucose enzymes confers an advantage to cells growingon D-arabinose, then it would be expected that either thefucR6 or the fucQ6 mutation should arise in response toselection on D-arabinose. We have, however, been unable toisolate the fucR6 mutation by direct selection on D-arabinose. The reason for inability to isolate this mutation bydirect selection is unclear. It may be that thefucR6 mutationarises at low frequency with respect to the fucQ6 mutation,or it may be that losing the unnecessary aldolase activity, asoccurs in the fucQ6 mutation, conferred more of an advan-tage to the cells growing on D-arabinose than did constitutivesynthesis of the fucose enzymes alone. Additionally, if thefucQ6 regulatory mutation does result in constitutive syn-thesis and aldolase deficiency at the same time, and if it is themost common constitutive mutation to appear in E. coliK-12, then this type of mutation could only be isolated bydirect selection on D-arabinose in cells that contained theribitol pathway. Therefore, all previous D-arabinose-positivemutants of E. coli K-12 had to be of the inducible phenotype.Improved growth on a novel substrate resulting in loss of
the ability to utilize a natural substrate has been observedbefore. As mentioned previously, improved growth on L-1,2-propanediol results in loss of the ability to grow on L-fucose(12), and Scangos and Reiner (20) have reported that growthof ribitol-positive E. coli on high levels of xylitol results intoxicity of ribitol to these cells. The borrowing of existingenzymes for the utilization of a novel substrate may be acommon mechanism for the acquisition of a new metaboliccapability. It is clear, however, that under certain circum-stances the acquisition of the new capability can result inconcomittant loss of an existing function, a phenomenonwhich results in no net increase in the metabolic diversity ofthe cell. Although it would be rare in nature for a microor-ganism to encounter a single compound as the only carbonsource available, periodic events of this type or the exhaus-tion of a preferred carbon source could lead to the acquisi-tion of a new capability and loss of an old one.
ACKNOWLEDGMENTThis material is based upon work supported by the National
Science Foundation under grant PCM-8402573.
J. BACTERIOL.
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IMPROVED D-ARABINOSE PATHWAY 709
LITERATURE CITED1. Anderson, R. L., and W. A. Wood. 1962. Purification and
properties of L-xylulokinase. J. Biol. Chem. 237:1027-1033.2. Bartkus, J. M., and R. P. Mortlock. 1986. Isolation of a mutation
resulting in constitutive synthesis of L-fucose catabolic en-zymes. J. Bacteriol. 165:710-714.
3. Boulter, J., B. Gielow, M. McFarland, and N. Lee. 1974.Metabolism of D-arabinose by Escherichia coli B/r. J. Bacteriol.117:920-923.
4. Bradford, M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
5. Chakrabartl, T., Y.-M. Chen, and E. C. C. Lin. 1984. Clusteringof genes for L-fucose dissimilation by Escherichia coli. J.Bacteriol. 157:984-986.
6. Chen, Y.-M., T. Chakrabartl, and E. C. C. Lin. 1984. Consti-tutive activation of L-fucose genes by an unlinked mutation inEscherichia coli. J. Bacteriol. 159:725-729.
7. Cocks, G. T., J. Aguilar, and E. C. C. Lin. 1974. Evolutiop ofL-1,2-propanediol catabolism in Escherichia coli by recruitmentof enzymes for L-fucose and L-lactate metabolism. J. Bacteriol.118:83-88.
8. Davis, R. W., D. Botstein, and J. R. Roth. 1980. Isolation ofTnlO insertions near particular genes, p. 21-26. In Advancedbacterial genetics. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
9. Dische, Z., and E. Borenfreund. 1951. A new spectrophotomet-ric method for the detection and determination of keto sugarsand trioses. J. Biol. Chem. 192:583-587.
10. Dykhuizen, D., and D. Hartl. 1981. Evolution of competitiveability in Escherichia coli. Evolution 35:581-594.
11. Green, M., and S. S. Cohen. 1956. Enzymatic conversion ofL-fucose to L-fuculose. J. Biol. Chem. 219:557-568.
12. Hacking, A. J., and E. C. C. Lin. 1976. Disruption of the fucosepathway as a consequence of genetic adaptation to propanediolas a carbon source in Escherichia coli. J. Bacteriol.126:1166-1172.
13. Hacking, A. J., and E. C. C. Lin. 1977. Regulatory changes in
the fucose system associated with the evolution of a catabolicpathway for propanediol in Escherichia coli. J. Bacteriol.130:832-838.
14. Hartl, D. L., and D. E. Dykhuizen. 1981. Potential for selectionamong nearly neutral allozymes of 6-phosphogluconate dehy-drogenase in Escherichia coli. Proc. NatI. Acad. Sci. USA78:63446348.
15. Leblanc, D. J., and R. P. Mortlock. 1971. Metabolism ofD-arabinose: a new pathway in Escherichia coli. J. Bacteriol.106:90-96.
16. Lim, R., and S. S. Cohen. 1966. D-Phosphoaribinoisomerase andD-ribulokinase in Escherichia coli. J. Biol. Chem. 241:4304-4315.
17. Oliver, E. J., and R. P. Mortlock. 1971. Growth of Aerobacteraerogenes on D-arabinose: origin of the enzyme activities. J.Bacteriol. 108:287-292.
18. Reiner, A. M. 1975. Genes for ribitol and D-arabitol catabolismin Escherichia coli: their loci in C strains and absence in K-12and B strains. J. Bacteriol. 123:530-536.
19. Rosner, J. L. 1972. Formation, induction and curing ofbacteriophage P1 lysogens. Virology 48:679-689.
20. Scangos, G. A., and A. M. Reiner. 1978. Acquisition of theability to utilize xylitol: disadvantages of a constitutive catabolicpathway in Escherichia coli. J. Bacteriol. 134:501-505.
21. Silhavy, T. J., M. L. Berman, and L. W. Enquist. 1984.Experiments with gene fusions. Cold Spring Harbor Labora-tory, Cold Spring Harbor, N.Y.
22. Skjold, A. C., and D. H. Ezekiel. 1982. Analysis of lambdainsertions in the fucose utilization region of Escherichia coliK-12: use of fuc and argA transducing bacteriophages to par-tially order the fucose utilization genes. J. Bacteriol.152:120-125.
23. St. Martin, E. J., and R. P. Mortlock. 1976. Natural and alteredinduction of the L-fucose catabolic enzymes in Klebsiella aero-genes. J. Bacteriol. 127:91-97.
24. Wood, W. A., M. J. McDonough, and L. B. Jacobs. 1961. Ribitoland D-arabitol utilization by Aerobacter aerogenes. J. Biol.Chem. 236:2190-2195.
VOL. 165, 1986
on July 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















