
Components of the macrolide binding site on the ribosome
10
Journal of Antimicrobial Chemotherapy (1985) 16, Suppl. A, 53-62 Components of the macrolide binding site on the ribosome F. Tejedor and Juan P. G. Ballesta Centro de Biologia Molecular, CSIC and UAM, Canto Blanco, Madrid-34, Spain Carbomycin A, niddamycin and tylosin, macrolide antibiotics that inhibit ribosomal activity, have a-fi unsaturated ketone groups in their structure that make them photo- reactive. These drugs can also take part in thermic reactions, probably through an addition mechanism to the double bond. Given of the photoactivity and thermic reactivity of their molecules, affinity labeling of the macrolide binding site on the ribosome has been performed using radioactive derivatives of these drugs. After either irradiating or incubating samples containing antibiotics and ribosomal particles, radioactivity appears covalent associated to the proteins. Ribosomal protein L27 is the major labeled component, indicating that this polypeptide, which seems to be part of the peptidyl transferase centre of the ribosome, also plays an important role on the macrolide binding site. Antibiotics have been extensively used as tools in molecular biology studies dealing with the structure and function of their cellular targets (Biicher & Sies, 1969). In the case of the ribosome, the identification of the components involved in the drug-binding sites has provided information on the structure of the functional domains of the particles affected by the antibiotics. For instance, the localization of the puromycin and chloramphenicol binding sites in the model of the large ribosomal subunit indicates the region of the particle where the peptidyl transferase centre is located (see Wittmann, 1983 for a recent review). Affinity labelling has been used for localization of the cellular components involved in the binding site of many cellular effectors (Chowdhry & Westheimer, 1979) and is a technique that gives clear-cut results, provided that appropriate controls are used. Macrolides have been considered as peptidyl transferase inhibitors since they affect the binding of its substrates. They also compete for binding to theribosomeswith typical peptide bond formation inhibitors such as chloramphenicol and lincomycin (Vazquez, 1979). However, it is also true that some macrolides are bad inhibitors of some peptide formation assays (Vazquez, 1975) and we have to recognize that many questions about their actual mode of action are still far from being answered. In fact, in Menninger's conclusions as to the mode of action of macrolides, he does not consider peptidyl transferase inhibition as the main effect of the activity of these drugs (Menninger, 1985). Clearly, the identification of the macrolide binding site on the ribosome can help us to understand its mode of action. Since some macrolides have, in their structure, potentially photoreactive chemical groups such as a-/? unsaturated ketones and can be made radioactive by reducing the aldehyde group at position 18, we have attempted the affinity labelling of the binding site of carbomycin A, niddamycin and tylosin. As shown in Figure 1 these compounds have the appropriate reactive groups. Reduction of the three antibiotics was carried out with either tritiated or non-radioactive borohydride, and the reduced drugs were purified by thin layer chromatography and 53 0305-7453/85/16A053+10 $02.00/0 © The British Society for Antimicrobial Chemotherapy Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491 by guest on 29 March 2018
-
Upload
duongthien -
Category
Documents
-
view
222 -
download
1
Transcript of Components of the macrolide binding site on the ribosome

Journal of Antimicrobial Chemotherapy (1985) 16, Suppl. A, 53-62
Components of the macrolide binding site on the ribosome
F. Tejedor and Juan P. G. Ballesta
Centro de Biologia Molecular, CSIC and UAM, Canto Blanco, Madrid-34, Spain
Carbomycin A, niddamycin and tylosin, macrolide antibiotics that inhibit ribosomalactivity, have a-fi unsaturated ketone groups in their structure that make them photo-reactive. These drugs can also take part in thermic reactions, probably through anaddition mechanism to the double bond. Given of the photoactivity and thermicreactivity of their molecules, affinity labeling of the macrolide binding site on theribosome has been performed using radioactive derivatives of these drugs. After eitherirradiating or incubating samples containing antibiotics and ribosomal particles,radioactivity appears covalent associated to the proteins. Ribosomal protein L27 isthe major labeled component, indicating that this polypeptide, which seems to be partof the peptidyl transferase centre of the ribosome, also plays an important role onthe macrolide binding site.
Antibiotics have been extensively used as tools in molecular biology studies dealing withthe structure and function of their cellular targets (Biicher & Sies, 1969). In the case ofthe ribosome, the identification of the components involved in the drug-binding sites hasprovided information on the structure of the functional domains of the particles affectedby the antibiotics. For instance, the localization of the puromycin and chloramphenicolbinding sites in the model of the large ribosomal subunit indicates the region of theparticle where the peptidyl transferase centre is located (see Wittmann, 1983 for a recentreview). Affinity labelling has been used for localization of the cellular componentsinvolved in the binding site of many cellular effectors (Chowdhry & Westheimer, 1979)and is a technique that gives clear-cut results, provided that appropriate controls are used.
Macrolides have been considered as peptidyl transferase inhibitors since they affectthe binding of its substrates. They also compete for binding to the ribosomes with typicalpeptide bond formation inhibitors such as chloramphenicol and lincomycin (Vazquez,1979). However, it is also true that some macrolides are bad inhibitors of some peptideformation assays (Vazquez, 1975) and we have to recognize that many questions abouttheir actual mode of action are still far from being answered. In fact, in Menninger'sconclusions as to the mode of action of macrolides, he does not consider peptidyltransferase inhibition as the main effect of the activity of these drugs (Menninger, 1985).Clearly, the identification of the macrolide binding site on the ribosome can help us tounderstand its mode of action.
Since some macrolides have, in their structure, potentially photoreactive chemicalgroups such as a-/? unsaturated ketones and can be made radioactive by reducing thealdehyde group at position 18, we have attempted the affinity labelling of the bindingsite of carbomycin A, niddamycin and tylosin. As shown in Figure 1 these compoundshave the appropriate reactive groups.
Reduction of the three antibiotics was carried out with either tritiated or non-radioactiveborohydride, and the reduced drugs were purified by thin layer chromatography and
530305-7453/85/16A053+10 $02.00/0 © The British Society for Antimicrobial Chemotherapy
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018
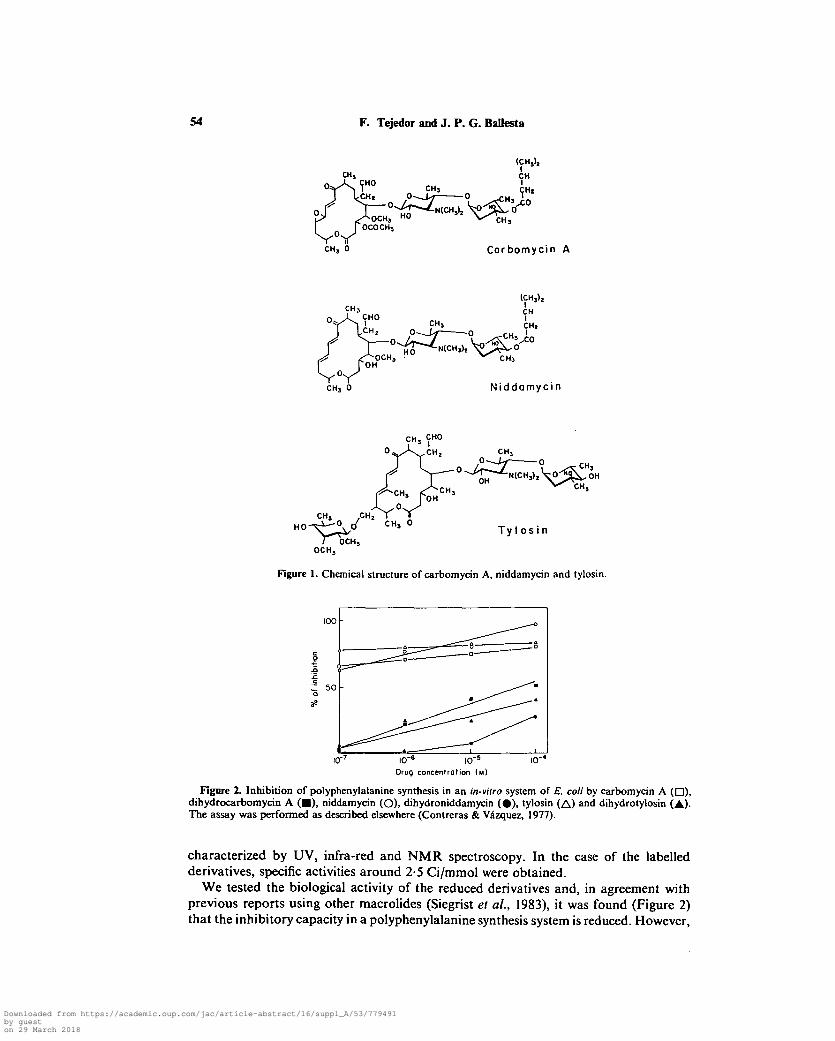
54 F. Tejedor and J. P. G. Ballesta
CHj 0 Carbomycin A
o ^ S ?H0
CHj 0
O H
(CHj)2
CH
Niddamycin
CHO
CHj .CHJ
TyIos in
OCH,
Figure 1. Chemical structure of carbomycin A, niddamycin and tylosin.
100 -
icr* io~=
Drug concentration ( M )
10'
Figure 2. Inhibition of polyphenylalanine synthesis in an in-vitro system of E. coli by carbomycin A ( • ) ,dihydrocarbomycin A ( • ) , niddamycin (O), dihydroniddamycin ( • ) , tylosin (A) and dihydrotylosin (A)-The assay was performed as described elsewhere (Contreras & Vazquez, 1977).
characterized by UV, infra-red and NMR spectroscopy. In the case of the labelledderivatives, specific activities around 2-5 Ci/mmol were obtained.
We tested the biological activity of the reduced derivatives and, in agreement withprevious reports using other macrolides (Siegrist el al., 1983), it was found (Figure 2)that the inhibitory capacity in a polyphenylalanine synthesis system is reduced. However,
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018
Macrolide binding site 55
0 0
5 0
0 0
5 0
;—•>. (o)
• \
\ -
* V (b)
- \
•
IO"e I0"7 I0"6 I0'5 IO"e I0"7 I0"6 I0"5
Drug concentrotion ( M )
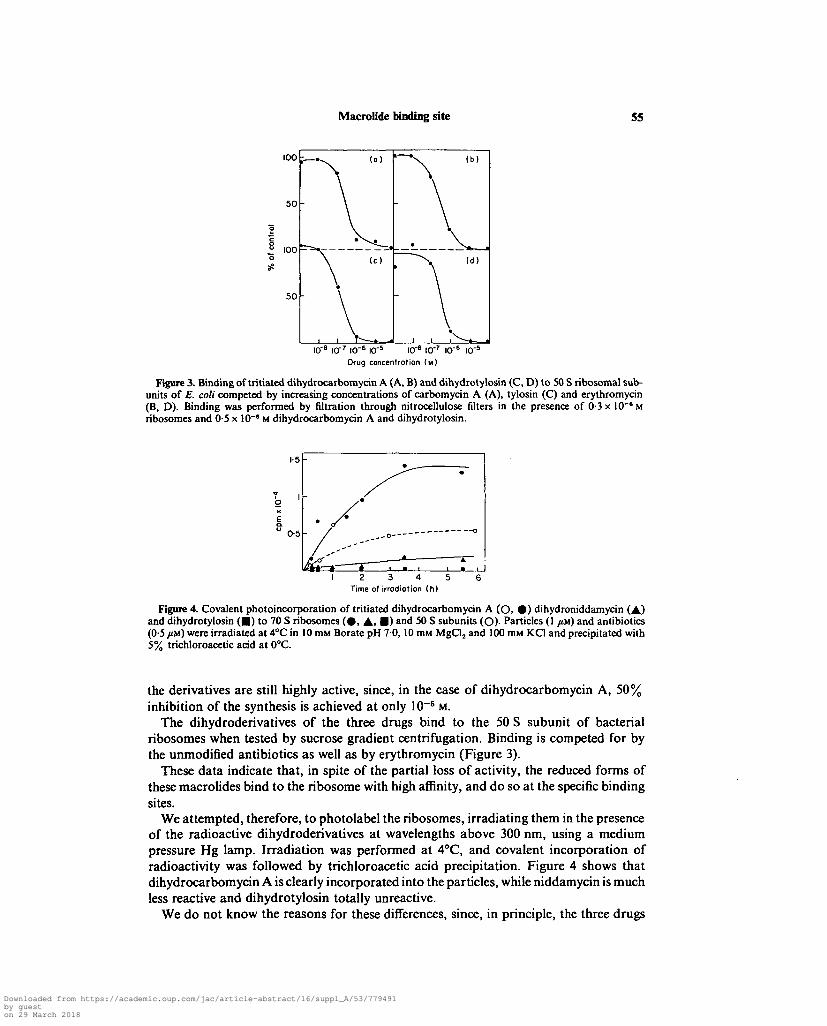
Figure 3. Binding of tritiated dihydrocarbomycin A (A, B) and dihydrotylosin (C, D) to 50 S ribosomal sub-units of E. coli competed by increasing concentrations of carbomycin A (A), tylosin (C) and erythromycin(B, D). Binding was performed by nitration through nitrocellulose filters in the presence of 0-3 x 10~a Mribosomes and 0-5 x 10"' M dihydrocarbomycin A and dihydrotylosin.
0-5 -
3 4Time of irrodiotion (h)
Figure 4. Covalent photoincorporation of tritiated dihydrocarbomycin A (O, 0 ) dihydroniddamycin (A)and dihydrotylosin ( • ) to 70 S ribosomes ( 0 , A, • ) and 50 S subunits (O)- Particles (1 /M) and antibiotics(0-5 im) were irradiated at 4°C in 10 nut Borate pH 70, 10 miu MgCl2 and 100 mM KC1 and precipitated with5% trichloroacetic acid at 0°C.
the derivatives are still highly active, since, in the case of dihydrocarbomycin A, 50%inhibition of the synthesis is achieved at only 10~8 M.
The dihydroderivatives of the three drugs bind to the 50 S subunit of bacterialribosomes when tested by sucrose gradient centrifugation. Binding is competed for bythe unmodified antibiotics as well as by erythromycin (Figure 3).
These data indicate that, in spite of the partial loss of activity, the reduced forms ofthese macrolides bind to the ribosome with high affinity, and do so at the specific bindingsites.
We attempted, therefore, to photolabel the ribosomes, irradiating them in the presenceof the radioactive dihydroderivatives at wavelengths above 300 nm, using a mediumpressure Hg lamp. Irradiation was performed at 4°C, and covalent incorporation ofradioactivity was followed by trichloroacetic acid precipitation. Figure 4 shows thatdihydrocarbomycin A is clearly incorporated into the particles, while niddamycin is muchless reactive and dihydrotylosin totally unreactive.
We do not know the reasons for these differences, since, in principle, the three drugs
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

56 F. Tejedor and J. P. G. Ballesta
Table I. Photocovalent binding of dihydrocarbomycin A as a functionof the drug concentration
Drugconcentration
(M)
1 x 10"'1-8x10-'
5 x 10-'1 x 10-"
1-8 xlO-6
5 x 10-"
Boundradioactivity(cpmx 10"3)
01010-261-421 981-75
Ribosomes were irradiated in the presence of the indicated concentrations of antibiotic for 2 h at 4°C andthen precipitated with 5% trichloroacetic acid at 0°C and filtered through glass fibre filters.
2
'•5 1
-i <
!10 20 30 40
2
3
•5
21
1
0-5
(b)
-
-
11 \x
A.I
|
1 .4
i"
2
3
•S
2
1
0-5
(c)
-
-
-A/ \J "
• - 1
f\i •i •
i '°-I •* \i •,-
J '•10 20 30 40 10 20 30 40
Fraction number
Figure S. Covalent binding of tritiated dihydrocarbomycin A to ribosomes tested by sucrose gradient cen-trifugation of irradiated particles, (a) Ribosomes and antibiotics were irradiated at 4°C; (b) samples wereincubated at 30°C in the dark; (c) samples were kept at 4°C without irradiation. After centrifugation, thesamples were fractionated and after checking their A2I0 (O), precipitated with trichloroacetic acid as inFigure 4.
have similar photoreactive groups. The photoincorporation is not related to the totalbinding to the particles since dihydrotylosin binds very efficiently to the ribosomes whentested by sucrose gradient centrifugation (not shown). We tend to think that thestructural characteristics of the molecules of the antibiotic are responsible for thesedifferences in reactivity. In the case of dihydrotylosin, the presence of one methyl groupat position C-12 and one sugar residue at position C-14, both near the photoreactivegroup, might interfere with the photoincorporation reaction.
Covalent binding also takes place at the specific binding sites for macrolides in theribosome since they can be saturated at low antibiotic to ribosome ratios, as shown inTable I, presenting the binding of dihydrocarbomycin A to 70 S ribosomes as a functionof the drug concentration. Moreover, the covalent binding is also competed for by theunmodified drug (not shown).
The distribution of the covalently-bound radioactivity among the ribosomal sub-particles was followed by sucrose gradient centrifugation of the labelled ribosomes. Itwas found that dihydrocarbomycin A is covalently bound to both ribosomal subunits(Figure 5 a). The distribution is clearly seen in panel A, where ribosomes were irradiatedat 4°C and spun down. In panel B, ribosomes were incubated at 30°C, without irradiationbeing applied to the gradients. In panel C, ribosomes were neither irradiated nor incu-
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

Macrolide binding site 57
_ 10 -
10 20 30 40 10 20 30 40Fraction number
10 20 30 40
Figure 6. Covalent binding of tritiated dihydroniddamycin to ribosomes. The experiment was performed asdescribed in Figure 5.
Table II. Quantification of protein labelling by dihydrocarbomycin A and dihydroniddamycin
Protein
L27L28L2L6/11L8L30L32/33S12S9S20/21S18S19
of total radioactivity incorporated in ribosomes labelled by
Dihydrocarbomycin A:
50 SPhotoreaction
70Ssubunits ribosomes
65-811-311-52-82-93-5000000
48-6105-410117-65-3
24-95-2000
Dihydroniddamycin
Photoreaction70S
ribosomes
29-617-71514-704.90
10-48-73-800
Thermic reaction70S
ribosomes
21022-9
7 12-4000
14-510-413-84-63-8
Bidimensional gel electrophoresis of proteins was performed by the methods of Kaltschmidt & Wittmann(1983) and Subramanian (1974) in the cases of dihydrocarbomycin and dihydroniddamycin respectively. Inthe second technique, although the sensitivity is higher, the resolution and identification of the proteins is notso well established as in the first system, and in some cases the radioactive spot coincides with more than oneprotein. In these cases, we have listed the protein that has also been identified as radioactive in the Kaltschmidt& Wittmann system.
bated. As expected, covalent incorporation is dependent on the previous irradiationof the samples.
Next, the same experiment was performed using dihydroniddamycin with surprisingresults. In this case (Figure 5b), although there is also an irradiation-dependentincorporation of antibiotic as shown in panel A, the incubation of the sample at 30°Cwithout previous irradiation also induces a high level of covalent incorporation ofradioactivity into the particles (panel B), even noticeable at 4°C (panel C).
Under these conditions, dihydrotylosin is not incorporated into the ribosomes eitherby irradiation or by incubation. It is, therefore, clear from these experiments that at leastdihydroniddamycin is able to take part in a thermic reaction with some ribosomalcomponents without previous irradiation.
We have tried to characterize the thermic as well as the photochemical reactions ofthese macrolides. The thermic reaction seems to involve the formation of an adduct
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

58 F. Tejedor and J. P. G. Ballesta
Id
SI 2
B
CM
Figure 7. Two-dimensional gel electrophoresis of ribosomal proteins from 70 S particles irradiated in thepresence of tritiated dihydrocarbomycin A. (A) Stained gel; (B) autoradiography. Electrophoresis was carriedout according to the method of Kaltschmidt & Wittmann (1970).
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

Macrolide binding site 59
between the a-fi unsaturated ketone and one half-cystine residue of the ribosomalproteins. This must be similar to what has been shown by Le Goffic and his colleagues(Siegrist et ai, 1983) for the reaction of rosaramycin and /S-mercaptoethanol.
In the case of the photoreaction, the reaction mechanism is still poorly understood.It is clear that SH-containing groups are involved. However, other chemical residues ofproteins must take part in the reaction as well since, as will be shown later, proteinswithout cysteine in their amino acid sequence are also labelled.
We labelled ribosomes with dihydrocarbomycin A and dihydroniddamycin, bothunder conditions for photoincorporation (4°Q and also for thermic incorporation (30°Cin the dark) in the case of dihydroniddamycin.
The RNA and the proteins were separated, and in all cases more than 90% of theradioactivity was associated with the proteins. Separation of proteins was achieved bytwo-dimensional gel electrophoresis and the radioactive spots detected by fluorography.Figure 6 shows the results obtained when the proteins of 70 S ribosomes labelled withdihydrocarbomycin A are separated by the Kaltschmidt & Wittmann (1970) two-dimensional electrophoresis technique. The main radioactive spot in the autoradiogramcorresponds to protein L27 which, due to the incorporation of more than one residue,gives at least two radioactive spots which move to a north-west position of theunmodified protein. Other labelled proteins are SI2, L2, L32/33, S9, L30. Similar resultswere obtained with dihydroniddamycin.
We have quantified these results by densitometry of the spots and the data arepresented in Table II. It is clear that in all the photoincorporation experiments, eitherwith dihydrocarbomycin A or dihydroniddamycin, protein L27 is the main labelledproduct. In 50 S subunits, it represents 65% of the total incorporated radioactivity.Proteins L28 and L2 are also considerably labelled and to a lesser extent proteins L6/L11,L30 and L32/33. In the small subunit, protein S12 is considerably labelled bydihydrocarbomycin A which also labels S9. Dihydroniddamycin labels S12 and S9 toa quite similar extent.
The thermic incorporation of dihydroniddamycin, on the other hand, presentsqualitative as well as quantitative differences with respect to photoincorporation. Ingeneral, the result looks less specific, labelling more proteins in the 30 S subunit like SI8,S19 and S20/21.
The first question that arises after the analysis of these results is the meaning of the30 S subunit labelling. Since it has been clearly established by different functional studies(Vazquez, 1979) that the binding site for macrolides is on the large subunit, the questionas to whether the labelling found in the small subunit has functional significanceimmediately arises.
At the moment, it is difficult to answer this question. Proteins SI2 and S9, labelledin the small subunit in all cases, are close neighbours of proteins L27 and L2 and thereforeit is not surprising that they are radioactively labelled. This does not mean, however,that they are part of the binding site, although their labelling suggest that this site is atthe interface between both subunits. This is the case for other antibiotics such aspuromycin (Nicholson et al., 1982), chloramphenicol (Pongs & Messer, 1976), tobramycin(Tangy, Capman & Le Goffic, 1983), pleuromutilin (Hogenauer et ai, 1981), andpactamycin (Tejedor, Amils & Ballesta, 1985). On the other hand, the fact that mutationsin the small subunit affect sensitivity of bacteria to macrolides, as found by severalauthors (Saltzman & Apirion, 1976; Pardo & Rosset, 1977), is compatible with a directinvolvement of this subunit in the binding process, and, if so, proteins S12 and S9 mighthave a role in the macrolides' interaction.
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

60 F. Tejedor and J. P. G. BaUesta
Figure 8. Model of a possible binding site for macrolides after Eilat et al. (1974). M represents the macrolidemolecule. The lines between proteins indicate close proximity as detected by cross-linking experiments (Coverel al., 1981).
Figure 9. Model of the 50 S ribosomal subunit including some protein markers as detected by the groupsof Lake & Stonier (Wittmann, 1983) as well as the position of the binding site for elongation factor EFG andantibiotics such as puromycin (Pu), chloramphenicol (Cm) and thiostrepton (Ts). Exit, indicates the place wherethe polypeptide growing chain has been detected emerging from synthesizing ribosomes (Bernabeu & Lake,1982). The possible position of the macrolide binding site around protein L27 has been marked as Ma.
As to the proteins labelled in the 50 S subunit, it must be noted that most of themhave been related by other techniques to the peptidyl transferase centre; these areproteins L2, L27, L6/11 and L32/33. This is in agreement with the interaction of thesemacrolides with that active domain of the ribosome. More concretely, proteins L2, L27and L32/33 have been associated with the P site of the peptidyl transferase (Saltzman& Apirion, 1976) which suggests that macrolides interact mainly with that part of theactive centre.
Some data on the inhibitory activity of macrolides indicate that a certain length ofthe polypeptide chain in the peptidyl-tRNA, around three to six peptide bonds, isrequired for inhibition of the peptidyl transferase activity (Vazquez, 1975). These results,together with the fact, tested in in-vitro experiments (Contreras & Vazquez, 1977), thatmacrolides do not bind to polysomes, can be interpreted as suggesting that theseantibiotics inhibit protein synthesis after the formation of the first few peptide bonds,by binding to the polypeptide chain exit channel. It is interesting to note that Cantor
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

Macrolide binding site 61
and co-workers (Eilat et al., 1974), have proposed a model for the P site and the growingpolypeptide exit groove based on affinity labelling results, that fits in nicely with ourresults. Figure 7 presents a model of a possible macrolide interaction, based on Cantor'sresults in which the drug binding at protein L27 would block the exit channel, hinderingthe growing polypeptide after the formation of a few peptide bonds. This model is alsocompatible with the alternative mode of action that Menninger (1985) presents in detailin this supplement. The steric hindrance that the macrolide molecule imposes on thegrowing polypeptide can destabilize the peptidyl-tRNA during translocation from theA site to the P site, inducing the release of the tRNA molecule, as stated in Menninger'smodel.
In any case, the region where the macrolides bind to the 50 S subunit must be aroundthe position of protein L27, which, as shown in Figure 9, has been localized to one sideof the central protuberance of the large subunit, side by side with the chloramphenicol-binding site, which also interacts with the puromycin site. If we assume that chloram-phenicol and puromycin are inhibitors of the A site of peptidyl transferase, whilemacrolides seem to be P site inhibitors, these two parts of the active centre mustcorrespond to the regions where the binding site of the respective antibiotics in the modelare situated.
Le Goffic and co-workers, using the macrolide rosaramycin under different labellingconditions, have found a pattern of labelled proteins that differs from our results(Siegrist, Moreau & Le Goffic, 1984). They find labelling in proteins LI, L5, L6 and SI.This discrepancy may be due to the different labelling conditions and/or to the differentdrug used. In any case, we have to keep in mind that affinity labelling results are con-ditioned by the specificity of the reaction that takes place and it is possible that one com-ponent of the binding site not labelled by one probe, is labelled by a different one withdistinct reactivity. Therefore, different results obtained with varying derivatives or similardrugs are not necessarily incompatible, but can be complementary.
Bearing in mind these limitations of the technique, we are at present performing affinitylabelling experiments with other derivatives of macrolides, which have groups withdifferent reactivity in other positions of the molecule. We are trying to map other possiblecomponents of their binding site that would provide additional data to confirm oreventually discard the model proposed above, and that might also help to resolve thediscrepancies between the results of Le Goffic and ourselves.
Acknowledgements
This work was supported by institutional grants to the Centro de Biologia Molecularfrom Fondo de Investigaciones Sanitarias (Instituto Nacional de Prevision) and by per-sonal grants to J. P. G. Ballesta from Comision Asesora de Investigacion Cientifica yTecnica and from Fundacion Eugenio Rodriguez Pascual.
References
Bernabeu, C. & Lake, J. A. (1982). Nascent polypeptide chains emerge from the secretory domainof the large ribosomal subunit: Immunomaping of the nacent chain. Proceedings of theNational Academy of Sciences, U.S.A. 79, 3111-5.
Bucher, Th. & Sies, H. (1969). Inhibitors: Tools in Cell Research. Springer-Verlag, Berlin,Heidelberg, New York.
Chowdhry, V. & Westheimer, F. H. (1979). Photoaffinity labeling of biological systems. AnnualReview of Biochemistry 48, 293-325.
3-2
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018

62 F. Tejedor and J. P. G. BaUesta
Contreras, A. & Vazquez, D. (1977). Cooperative and antagonistic interactions of peptidyl-tRNAand antibiotics with bacterial ribosomes. European Journal of Biochemistry 74, 539-47.
Cover, J. A., Lambert, J. M., Norman, C. M. & Traut, R. R. (1981). Identification of proteins atthe subunit interface of E. coli ribosome by cross-linking with dimethyl 3.3'-dithiobis(propionimidate). Biochemistry 20, 2843-52.
Eilat, D., Pellegrini, H., Oen, Y., Lapidot, Y. & Cantor, C. R. (1974). A chemical mapping techniquefor exploring the location of proteins along the ribosome bound peptide chain. Journal ofMolecular Biology 88, 831-40.
Hogenauer, G., Egger, H., Ruf, C. & Stumper, B. (1981). Affinity labeling of Escherichia coli ribo-somes with a covalently bound derivative of the antibiotic pleuromutilin. Biochemistry 20,546-52.
Kaltschmidt, E. & Wittman, H. G. (1970). Ribosomal proteins, XII. Number of proteins in smalland large ribosomal subunits of Escherichia coli as determined by two-dimensional gelelectrophoresis. Proceedings of the National Academy of Sciences, U.S.A. 67, 1276-82.
Menninger, J. R. (1985). Functional consequences of binding macrolides to ribosomes. Journalof Antimicrobial Chemotherapy 16, Suppl. A, 23-34.
Nicholson, A. W., Hall, C. C, Strycharz, W. A. & Cooperman, B. S. (1982). Photoaffinity labelingof Escherichia coli ribosomes by an aryl azide analog of puromycin. On the identificationof two major covalently labeled ribosomal proteins and the mechanism of photoincorporation.Biochemistry 21, 3709-817.
Pardo, D. & Rosset, R. (1977). Properties of ribosomes form erythromycin resistant mutants ofEscherichia coli. Molecular and General Genetics 156, 267-71.
Pongs, O. & Messer, W. (1976). The chloramphenicol receptor site in Escherichia coli. In vivoaffinity labeling by monoiodamphenicol. Journal of Molecular Biology 101, 171-84.
Saltzman, L. & Apirion, D. (1976). Binding of erythromycin to the 50s ribosomal subunit is affectedby alterations in the 30s ribosomal subunit. Molecular and General Genetics 143, 301-6.
Siegrist, S., Velitchkovitch, S., Le Goffic, F. & Moreau, N. (1983). Mechanism of action of a 16-membered macrolide. Characteristics of dihydrorosamycin binding to Escherichia coli ribo-somes. Journal of Antibiotics 35, 866-74.
Siegrist, S., Moreau, N. & Le Goffic, F. (1984). Photolabeling of macrolide binding site on the70S E. coli ribosome. Comptes rendues, Academie des Sciences, Paris 299, 49-51.
Subramanian, A. R. (1974). Sensitive separation procedure for Escherichia coli ribosomal proteinsand the resolution of high-molecular weight components. European Journal of Biochemistry45, 541-6.
Tangy, F., Capmenu, M. L. & Le Goffic, F. (1983). Photoinduced labeling of Escherichia coliribosomes by a tobramycin analog. European Journal of Biochemistry 131, 581-7.
Tejedor, F., Amils, R. & Ballesta, J. P. G. (1985). Photoaffinity labeling of pactamycin bindingsite on eubacterial ribosomes. Biochemistry (in press).
Vazquez, D. (1975). In Antibiotics. III. Mechanisms of Action of Antimicrobial and AntitumorAgents (Concoran, J. W. & Hahn, F. E., Eds), pp. 459-79. Springer-Verlag, Berlin,Heidelberg, New York.
Vazquez, D. (1979). Inhibitors of Protein Synthesis. Springer-Verlag, Berlin, Heidelberg, New York.Wittman, H. G. (1983). Architecture of prokaryotic ribosomes. Annual Review of Biochemistry
52, 35-65.
Downloaded from https://academic.oup.com/jac/article-abstract/16/suppl_A/53/779491by gueston 29 March 2018


















