
Comparative Vertebrate Anatomy Lab Book
181
0 A LABORATORY MANUAL FOR COMPARATIVE VERTEBRATE ANATOMY By Frank Logiudice Department of Biology University of Central Florida © 2008 Frank Logiudice
Transcript of Comparative Vertebrate Anatomy Lab Book

0
A LABORATORY MANUAL
FOR
COMPARATIVE VERTEBRATE ANATOMY
By Frank Logiudice
Department of Biology
University of Central Florida
© 2008 Frank Logiudice

1
This book is dedicated to Orville M. Berringer, a fellow faculty member and a good friend. I
learned many of the dissection techniques utilized in this manual while teaching “Human
Anatomy” with him. Budd Berringer was a dedicated mentor to both students and fellow
faculty. He was a strong advocate for the University of Central Florida’s prehealth professional
students. Its was due to his efforts that the University of Central Florida Prehealth Professional
Office was established. His hard work and dedication helped many UCF students to enter into
careers in medicine, dentistry, and veterinary medicine and his instruction gave them the tools to
succeed in professional school. I have had the honor of teaching with many spectacular faculty
but Budd will always stand out in my mind as the one who had a profound influence in how I
approach the teaching of anatomy.

2
TABLE OF CONTENTS
Introduction page 3
Chapter 1: Skeletal Structure page 8
Part A: Introduction page 8
Part B: The Axial Skeleton page 14
Topic B1: The Vertebrae page 14
Topic B2: The Ribs and Sternum page 23
Topic B3: The Skull page 23
Part C: The Appendicular Skeleton page 49
Topic C1: The Pectoral Girdle and Appendage page 49
Topic C2: The Pelvic Girdle and Appendage page 55
Chapter 2: Myology page 62
Part A: An Introduction page 62
Part B: Muscle Dissection Techniques page 65
Part C: The Muscles page 71
Topic C1: The Muscles of Squalus page 71
Topic C2: The Muscles of Necturus page 76
Topic C3: The Muscles of Felis page 83
Chapter 3: Visceral Organs and Angiology page 104
Part A: The Exposing the Visceral Organs and Angiology page 104
Topic A1: The Exposing the Visceral Organs and Angiology in Squalus page 104
Topic A2: The Exposing the Visceral Organs and Angiology in Necturus page 105
Topic A1: The Exposing the Visceral Organs and Angiology in Felis page 105
Part B: The Respiratory System page 106
Topic B1: The Respiratory System in Squalus page 106
Topic B2: The Respiratory System in Necturus page 108
Topic B3: The Respiratory System in Felis page 109
Part C: The Digestive System page 111
Topic C1: The Digestive System in Squalus page 112
Topic C2: The Digestive System in Necturus page 115
Topic C3: The Digestive System in Felis page 117
Part D: The Urogenital System page 122
Topic D1: The Urogenital System in Squalus page 122
Topic D2: The Urogenital System in Necturus page 126
Topic D3: The Urogenital System in Felis page 128
Part E: The Circulatory System page 135
Topic E1: The Circulatory System in Squalus page 135
Topic E2: The Circulatory System in Necturus page 150
Topic E3: The Circulatory System in Felis page 162

3
Introduction
Comparative vertebrate anatomy is a study of the vertebrate form from both an
anatomical point of view and from an evolutionary perspective. It is a study of vertebrate
morphology and of the evolutionary changes that have occurred in the vertebrate body over time.
These changes caused the formation of new species that were able to exploit an increasing range
of resources and ecological niches. Today vertebrates are an extremely successful subphylum of
animals.
One important aspect of studying comparative anatomy is dissection. Dissection will
allow us to have direct observation of anatomy and should give us an appreciation of
evolutionary developments.
We will be using primarily three species over the semester in our study of comparative
vertebrate anatomy. They are the spiny dogfish (Squalus sukleyi or Squalus acanthias), the
common mudpuppy (Necturus maculosus), and the common house cat (Felis catus). These three
species are very useful for observing a number of vertebrate anatomical features and
evolutionary developments. In particular they will allow us to see the various anatomical
modifications that occurred as vertebrates evolved for a terrestrial existence.
Kingdom: Animalia
Phylum: Chordata
Subphylum: Vertebrata
Class: Chondrichthyes
Order Squaliformes
Family: Squalidae
Genus: Squalus
Squalus acanthias and Squalus sukleyi are two very similar species commonly called
“spiny dogfish”. These are small sharks that are considered to be relatively abundant (although
like most shark species their numbers have dropped off markedly in recent years). S. acanthias
is found in the Northern Atlantic while S. sukleyi is located in the Northern Pacific. Both species
can form large schools that tend to segregate by size and gender. They are active predators and
feed on a variety of small fishes and cephalopods. Females tend to be larger than males with
adult males ranging from 60-90cm (24-35 inches) and adult females from 76-107 cm (30-42
inches) in length. Squalus are long living fish reaching an age comparable to that of humans.
Females reach maturity at 12 years and have a gestation period of two years. They give birth to
live young (about five to seven pups per litter).
Squalus is very useful in comparative vertebrate anatomy because they demonstrate what
may be considered to be basal vertebrate anatomy. These are the earliest living representatives

4
of the gnathostomata, the jawed vertebrates. Squalus and all sharks belong to the class
Chondrichthyes meaning that their skeleton is composed of cartilage. This is considered to be
the primitive vertebrate condition. Other vertebrates do retain some cartilage in the adult skeleton
but the bulk of the skeleton will be composed of bone. Also, the embryonic skeleton of
vertebrates will be composed of cartilage so Squalus is also useful from a developmental point of
view.
Kingdom: Animalia
Phylum: Chordata
Subphylum: Vertebrata
Class: Lissamphibia
Order Caudata
Family: Proteidae
Genus: Necturus
Necturus maculosus is a salamander species found in freshwater ponds, lakes, streams,
and rivers of eastern North America. They are nocturnal and purely aquatic. Necturus are
neotenic meaning that they are considered to be in a permanent larval state. They retain gills in
the adult state, which is reached in five or six years. This species can live to thirty years of age
or older. Necturus is a predator of small aquatic animals, in particular minnows and crayfish. N.
maculosus is the largest Necturus species and reaches a maximum length of 49 cm (19 inches).
Necturus maculosus is a nice example of the vertebrate transition between aquatic and
terrestrial existence. Although purely aquatic, these salamanders display tetrapod developments.
They have well-developed limbs with the standard tetrapod skeletal structures. However, like
fishes, it is the axial musculature that provides the main force for locomotion in these animals.
As a result, the axial musculature maintains a prominent segmental pattern as seen in fishes.

5
Kingdom: Animalia
Phylum: Chordata
Subphylum: Vertebrata
Class: Mammalia
Order Carnivora
Family: Felidae
Genus: Felis
Felis catus is a species that is well known to most students. The common housecat has
been a companion of humans for thousands of years. They are intelligent mammals with a social
structure. Felis is also an efficient predator of small animals. This may have been one reason
behind its domestication and has also led to a loss of many species of birds and small mammals
in areas in which cats have been introduced.
As a mammal it displays all of the features of a vertebrate that has evolved for terrestrial
existence. Like Necturus it has limbs but these have been modified over those of the mudpuppy
and most of the locomotory force comes from the appendages. The skeleton has an increase in
differentiation and muscle attachment points. The appendicular muscles are greater in size,
number, and complexity of action. Felis is also an endotherm and an amniote. As a result we
will be able to observe specializations that are the result of endothermy and structural features
found in amniotes that are not found in other vertebrates.
_____________________________________________________________________________
Before you enter into the lab you should take some time to become familiar with basic
anatomical terminology. This will make it much easier for you to assimilate the information
presented to you in your first, and subsequent, lab sessions. You also would do well to study the
lab material prior to each lab session. Due to security concerns, equipment costs, and other
logistical concerns you most likely will not have access to the laboratory any time other than
your assigned laboratory section. You will want to maximize that lab experience by working
with the actual anatomical specimens and not waste it reading your book. You can read your
book anywhere but you will only have a short period of time each week to interact with the lab
specimens. No matter how well depicted, no illustration equals working with the real thing.
To start you off this chapter will introduce you to some of the terminology of anatomy.
You will find that a familiarity with these terms will make your first task of learning osteological
features to be much easier.

6
1. Planes and Sections of the Body
To aid in the complex study of anatomy, it is often beneficial to divide into sections
through the use of planes (although this can also be done with surgical techniques as well).
(1) Sagittal Plane - a sagittal plane divides the body into right and left portions. If the
plane runs directly along the midline of the body it is termed a midsagittal plane. A
midsagittal plane will divide the body into equal right and left portions. If the plane
is off the midline it is said to be a parasagittal plane. A parasagittal plane will divide
the body into unequal right and left portions, the level of disparity depends on where
the plane is located.
(2) Frontal Plane (also termed Coronal) - a frontal plane divides the body into an anterior
and a posterior portion.
(3) Transverse Plane (also termed Horizontal or Cross Sectional) - a transverse plane
divides the body into upper (superior) a lower (inferior) portions.
2. Directional Terminology
!!!!!!!!!!! Directional terminology will also be invaluable to you in your studies.! You will notice
directional terms being used quite a bit in the naming of anatomical structures beginning with
osteological features.! One study method that may speed up your learning of these terms is to
consider them in opposite pairs (ex: superior vs. inferior).
!!!!!!!!!!! In directional terminology we are referring to the structure’s position in a body in the
anatomical position and relative to another structure.
!!!!!!!!!!! (1) Superior - to be above another structure. (Ex; the head is superior to the neck.)
!!!!!!!!!!! (2) Inferior - to be below another structure. (Ex; the neck is inferior to the head.)
(3) Anterior - to be in front of another structure. (Ex; the incisors are anterior to
the canines.)
!!!!!!!!!!! (4) Posterior - to be behind another structure. (Ex; the canines are posterior to the !!!!!!!!!!!
incisors.)
(5) Medial - to be along the midline axis of the body.! (Ex; the nose is medial relative to
the eyes.)
(6) Lateral - to be away from the midline of the body. (Ex; the eyes are lateral relative to
the nose.)
(7) Proximal - to be closer to the main axis of the body.! (Ex; the shoulder is proximal
relative to the wrist.)
(8) Distal - to be at a distance from, to be further from, the main axis of the body.! (Ex;
the wrist is distal relative to the shoulder.)
(9) Intermediate - to be between two structures, either between distal and proximal
structures or between lateral and medial structures.! The term “middle” can be used
for intermediate but middle and medial are not equivalent terms.! (Ex; your finger has
three bones called “phalanx” bones.! The one closest to your knuckle is the proximal
phalanx.! The one that your fingernail is on is the distal phalanx.! The one between is
intermediate, or middle, phalanx.)
(10) Deep - to be further from the surface of the body than is another structure, to be
beneath another structure.! (ex; the ribs are deep to the pectoralis major.)
(11) Superficial - to be closer to the surface of the body than is another structure.! (Ex;
the pectoralis major is superficial to the ribs.)

7
!!!!!!!!!!! (12) Cranial/Cephalad - to mean specifically “towards the head”.
!!!!!!!!!!! (13) Caudal - to mean specifically “towards the tail”.
Squalus Skeleton Demonstrating The Differences Between the Axial and Appendicular Skeleton
Axial:
Chondrocranium
Splanchnocranium
Ribs
Vertebral
Column
Pectoral
Girdle &
Fins
Appendicular:
Pelvic
Girdle &
Fins

8
Chapter 1: Skeletal Structure
Part A: An Introduction
The skeleton is the basic framework of the vertebrate body. It is the scaffolding on which
the muscles attach and which serves to protect the visceral organs. During the development, the
skeleton sets the pathway by which blood vessels and nerves grow. It is typically composed of
bone and cartilage.
The skeleton can be subdivided into the axial skeleton and the appendicular skeleton.
The axial skeleton is composed of the skeletal structures of the main axis of the body. These are
the skull, vertebral column, and rib cage. The appendicular skeleton is composed of the skeletal
elements of the limbs and their associated girdles.
Axial: Vertebral Column Ribs Splanchnocranium Neurocranium
Appendicular: Pelvic Girdle & Limbs Pectoral Girdle & Limbs
Necturus Skeleton Demonstrating The Differences Between the Axial and Appendicular Skeleton

9
Cat Skeleton Demonstrating The Differences Between the Axial and Appendicular Skeleton
In extant agnathans and chondrichthyes the skeleton is composed exclusively of cartilage
(with the exception of the teeth in chondrichthyans). There are four classes of cartilage: hyaline
cartilage, fibrocartilage, elastic cartilage, and calcified cartilage. The skeleton of Squalus is
composed primarily of hyaline cartilage. The jaws are composed of calcified cartilage (also
known as mineralized cartilage). Calcified cartilage is cartilage that has become more rigid due
to an invasion of calcium salts into the cartilage matrix. This adds strength to the tissue, which is
beneficial in shark jaws.
The cartilage organ is composed primarily of cartilage tissue. However it is a true organ
because it is composed of more than one type of tissue. A membrane termed the perichondrium
surrounds it. The perichondrium is a connective tissue having two layers. The outer layer is a
tough fibrous connective tissue that allows the organ to withstand mechanical forces (much like
an athletic bandage strengthening an ankle). The inner layer a single layer of multipotent cells
that can produce new cartilage cells and repair the cartilage organ.
In other vertebrates the embryonic skeleton is initially composed of cartilage. During
development the cartilage is replace by bone to varying degrees. In Neopterygians there is a
significant amount of cartilage in the adult skeleton while teleosts have a highly ossified
skeleton. The adult skeletons of Necturus and Felis are composed mostly of bone but some
cartilage remains.
Osteology is the study of bone and the skeleton. Bone organs are true organs being
composed of several tissues working together to perform a task. Bone organs typically contain

10
two classes of bone tissue: compact bone and spongy bone. Compact bone has a denser
histological arrangement making it stronger than spongy bone. As a result, compact bone will
form the outer layer of a bone organ while spongy bone forms the inner aspect of the bone organ.
Sometimes they are referred to as cortical bone and medullary bone respectively.
Compact Bone Tissue
Spongy Bone Tissue
There are four classes of bone organs based on shape: long bones, short bones, flat bones,
and irregular bones. Long bones are longer than they are wide. As we shall see, long bones will
have a distinctive internal architecture that distinguishes them from the other three classes. Long
bones include the femur and humerus. Short bones are simply that, short. They include many of
the carpal and tarsal bones (ex; pisiform, first cuneiform). There is a subclass of short bone
called the sesamoid bones. Sesamoid bones are short bones that do not attach to the skeleton
directly. Instead they are enveloped in tendon and ligament and attach indirectly to the
skeleton. Sesamoid bones are named for their sesame seed like shape. An example of a
sesamoid bone is the patella. Flat bones are thin, flattened bones. They include many of the
bones of the skull (ex; parietal) and the ribs. Certain flat bones have evolved an elongated
appearance (ex; ribs, clavicle) and may be confused with long bones initially. However their
internal structure is clearly not that of a long bone. Irregular bones are oddly shaped bones that
do not fit into the other three classes. They include the vertebrae.

11
The Four Classes of Bone Organs Based On Shape
Long bones have a distinctive structure. The rounded ends of a long bone, commonly
called “heads”, are termed epiphysis. The epiphysis that is closer to the main axis of the body is
called the proximal epiphysis. The epiphysis that is further from the main axis of the body is
called the distal epiphysis. Between the two epiphyses is the shaft of the bone anatomically
called the diaphysis. Internally both the epiphyses and the diaphysis have an outer layer of
cortical bone composed of compact bone tissue. In the epiphyses, internal to the cortical bone,
we see an extensive proliferation of spongy bone tissue forming the medullary bone. Within the
intertrabecular spaces red marrow is found. Red marrow is a hemopoietic tissue meaning that it
produces blood cells. In the diaphysis, internal to the cortical bone, we see very little spongy
bone. Instead there is a large open space called the medullary cavity. The medullary cavity is
filled with yellow marrow primarily. Yellow marrow is a densely packed adipose tissue.
In growing bone there exists a plate of hyaline cartilage between the epiphysis and the
diaphysis called the epiphyseal plate or epiphyseal growth plate . This is the region from which
the bone lengthens until an individual hits maturity. When growth stops the cartilaginous plate
ossifies and remains as a line of compact bone tissues situated between the diaphysis and the
epiphysis. This structure is termed the epiphyseal line.

12
Sagittal Section of a Long Bone
Covering the external surface of the bone organ is a connective tissue sheath called the
periosteum. The periosteum has two layers that are similar to those of the perichondrium. The
outer layer is a tough fibrous connective tissue. Extensions of this fibrous layer form ligaments
and play a role in forming tendons. Internally the periosteum has a single layer of multipotent
cells that can produce new bone cells and repair and remodel the bone organ. The periosteum is
attached to the bone by fibers called perforating fibers. Lining the internal surfaces of the bone
organ is the endosteum. The endosteum is similar to the inner layer of the periosteum. It is a
single layer of multipotent cells that can produce new bone cells and repair the bone organ.
The other three classes of bone organs have a very similar internal architecture to one
another. They have an outer layer of cortical bone (compact bone tissue) surrounding an inner
layer of medullary bone (spongy bone). The spongy bone houses red marrow. The other three
classes of bone organ lack: epiphyses, diaphysis, and a medullary cavity. Like long bones they
are covered by periosteum externally and endosteum internally.

13
Chapter 1. Table 1 Basic Terminology For Osteology
The following list covers many of the terms that you will come across as you study
osteology.! An understanding of these terms prior to entering the lab should make your study
time more efficient and effective.
(1) Foramen – a hole (ex; foramen magnum).
!!!!!!!!!!! (2) Fissure - a long narrow, opening in a bone (ex; superior orbital fissure).
(3) Meatus - meaning “canal” this is a canal-like opening in a bone (ex; external acoustic
meatus).! Often the term “canal” can be used interchangeably with “meatus”.
!!!!!!!!!!! (4) Fossa - a depression on the surface of a bone (ex; anterior cranial fossa).
!!!!!!!!!!! (5) Fovea - a pit in the surface of a bone (ex; fovea capitis).
(6) Sulcus - a groove on the surface of a bone (ex; lacrimal sulcus).! Often the term
“groove” can be used interchangeably with “sulcus”.
!!!!!!!!!!! (7) Tubercle - a rounded elevation on a bone (ex; pubic tubercle).
(8) Tuberosity -a roughened elevation on the surface of a bone (ex; ischial tuberosity).! In
certain cases the terms “tubercle” and “tuberosity” may be used interchangeably.
(9) Line - a line on the surface of a bone, a shallow linear elevation on the surface of a
bone (ex; soleal line).
(10) Crest - a linear elevation on the surface of a bone (ex; intertrochanteric crest).! A
crest will be a higher linear elevation than is a line.
(11) Process - an extension of a bone (ex; zygomatic process of the temporal bone)! If a
process attaches to another bone it will be named for the bone to which it articulates
(ex; temporal process of the zygomatic bone)..
(12) Condyle - a flared extension on a long bone that allows for better articulation (ex;
medial condyle of femur). Condyles allow for direct bone to bone attachment.! They
greatly increase! the surface area for an articulation adding to the strength and
stability of a joint.
(13) Condyloid - a condyle-like structure found on a bone that is not a long bone
(ex; condyloid process of the mandible).
!!!!!!!!!!! (14) Epicondyle - an elevation situated on a condyle (ex; medial epicondyle of the !!!!!!!!!!!
femur).! They allow for indirect bone to bone attachment (by serving as the !!!!!!!!!!
!!!!!!!!!!! anchoring point for ligaments) and for muscle attachment.
(15) Coronoid - a wing-like extension of a bone (ex; coronoid process of the ulna).
(16) Coracoid - meaning “crow’s beak” this is an extension of a bone that resembles the
bill of a crow (ex; coracoid process of the scapula).
!!!!!!!!!!! (17) Malleolus - a flattened projection (ex; medial malleolus of tibia).
!!!!!!!!!!! (18) Ramus - a bridge-like structure (ex; superior pubic ramus).
!!!!!!!!!!! (19) Ala - a wing shaped surface of a bone (ex; ala of sacrum).
(20) Facet - means an articulating surface (ex; superior articulating facet of vertebra).!
The terms “facet” and “articulating surface” can be used interchangeably.
(21) Demifacet - half of an articulating surface (ex; demifacet on body of a
thoracic vertebra).
(22) Cornu - meaning “horn”, it is a horn-like extension off of a bone (ex; sacral cornu)
!

14
!
Chapter 1: Skeletal Structure
Part B: The Axial Skeleton
Topic B1: The Vertebrae
In vertebrates vertebrae can be composed of either cartilage or, more commonly, bone.
They are successively arranged to form the vertebral column. The vertebral column surrounds
and protects the spinal cord, serves as a point of attachment for ligaments, tendons, and muscles,
and helps to bear the weight of the body.
Mammalian Vertebrae Demonstrating Some Basic Features
The modern vertebra typically consists of a centrum, a neural arch (aka; vertebral arch),
and one or more process (aka; apophyses) that project from either the centrum or the arch. The
centrum occupies the position occupied by the notochord during development. Due to their
method of development the vertebral centra are located intersegmentally, between two
myomeres. They develop along the notochord that had defined the long axis of the developing
embryo. The degree to which the notochord is maintained throughout an organism’s life varies.
The notochord may begin regression with the development of the vertebrae. The neural, or
vertebral, arch is located dorsal to the centrum. The vertebral arch serves to protect the spinal
cord. Successive arches, and their connecting ligaments, form the neural/vertebral canal. The
base of the arch is termed the pedicle and the roof and sides of the arch are termed the lamina.
Along with the vertebral arch some vertebrates will have vertebrae possessing a hemal arch.
When present the hemal arches are located in the tail, ventral to the centra and will house the
caudal artery. In amniotes they are called chevron bones.
The number and types of apophyses vary between vertebrate groups. Chondrichthyes can
be considered to have basal vertebrae since theirs have only one pair of apophysis. Mammals on

15
the other hand have numerous, well developed apophyses. The diapophyses (or transverse
processes) are the most common vertebral process. They articulate with ribs and serve for muscle
and ligament attachment (as do all of the apophyses to some degree). Another paired lateral
apophysis off of the centrum is the parapophyses. Parapophyses occur in vertebrates that have
biccipital ribs such as Necturus. (“Biccipital” means “two –headed”). The two heads are the
capitulum and tubercle. The parapophysis will attach to the capitulum of the rib. The tuberculum
of the rib will attach to the diapophysis of the vertebrae. Zygapophyses (or articulating
processes) allow for articulation between successive vertebrae. The typical vertebra will have
two pairs of zygapophyses. Located on the cranial aspect of the vertebra are the
prezygapophyses (a.k.a.; cranial articulating processes, superior articulating processes). These
are paired structures that allow one vertebra to attach to the vertebra cranial to it.
Postzygapophyses (a.k.a.; caudal articulating processes, inferior articulating processes) are
paired structures on the caudal aspect of the vertebrae that allow one vertebra to attach to the
vertebra caudal to it.
Squalus displays the typical chondrichthyan vertebra. The vertebrae are composed of
cartilage and consist of a centrum and vertebral arch. The chondrichthyan vertebrae are
considered to be the model for the primative vertebrate vertebrae. Although agnathans are more
primative and the vertebrae of extant species are considered to be incomplete, their placement in
the evolution of the vertebral column is speculative. The current agnathans vertebrae may
represent an early stage of vertebral development. However it is equally likely that the vertebrae
of extant species represents a derived characteristic; that the vertebrae have been reduced in
complexity over time.
Diapophysis
Dorsal Plate of Neural Arch
Dorsal Intercalary Plate of Neural Arch
Centrum
Ventral Plate Ventral Intercalary Plate
Trunk Vertebrae Caudal Vertebrae
Squalus Vertebrae
As is true for all fishes the vertebrae are divided into only two regional groups: trunk and
caudal. They are easily distinguishable. The trunk vertebrae possess poorly defined transverse
processes. In Squalus the trunk vertebrae will articulate with ribs. The caudal vertebrae have

16
hemal arches and do not articulate with ribs. The caudal vertebrae also display dispondyly.
Dispondyly means that there are two centra and two sets of neural and hemal arches in each tail
metamere. As a result the number of caudal centra is double that of caudal myomeres and caudal
spinal nerve pairs.
In sharks the notochord is retained in the adult. It is found throughout the length of the
vertebral column, where it is restricted to the centra. Shark centra are amphicelous being concave
at both ends. The vertebral arch consists of paired dorsal plates that will join to form the
vertebral arch. Between the arches are paired dorsal intercalary plates. The vertebral canal is
composed of the dorsal plates and the interdigitating dorsal intercalary plates. The dorsal
intercalary plates will each have a pair of perforations called the intervertebral foramina. These
openings allow for the passage of the spinal nerves. The caudal vertebrae will also have hemal
arches consisting of paired ventral plates. As occurred with the vertebral canal, between the
hemal arches are paired ventral intercalary plates. The ventral plates and the ventral intercalary
plates will form the hemal canal for the caudal artery.
Like Squalus the vertebrae of Necturus are amphicelous. Necturus, however, has a
vertebral column that shows a greater degree of regional differentiation than was seen in
Squalus. Along with trunk and caudal vertebrae there are one cervical vertebra and one sacral
vertebra. The cervical vertebra is a trunk vertebra that is modified to articulate with the occipital
condyles of the skull. This is the first step in the evolution of a neck, a tetrapod feature. The
cervical vertebra does not articulate with ribs. The sacral vertebra is a trunk vertebra that, like
the other trunk vertebrae, articulates with a pair of ribs. This rib pair is unique, however. These
are the sacral ribs. Sacral ribs articulate with the ilium of the pelvis.
Caudal Vertebrae Sacral Vertebra Trunk Vertebrae Cervical Vertebra
Necturus Demonstrating Regional Vertebral Variation
The trunk vertebrae of Necturus each have a pair of prezygapophyses and
postzygapophyses, a spinous process, a centrum, and a vertebral arch that will enclose the
vertebral canal. They will also have two pairs of processes to articulate with the biccipital ribs of
urodeles. They have a pair of diapophyses to articulate with the tuberculum of the rib and a pair
of parapophyses to articulate with the capitulum of the rib. This pattern is also true for the sacral
vertebra since the sacral ribs are also biccipital.
The caudal vertebrae show some variation. The first five resemble smaller trunk
vertebrae complete with parapophyses and diapophyses. After the fifth caudal vertebrae the
vertebrae more closely resemble the caudal vertebrae typical of fish. They will possess a hemal

17
arch to enclose the hemal canal and a vertebral arch. The diapophyses and parapophyses are lost
and the prezygapophyses and postzygapophyses are reduced.
Dorsal View: Prezygapophysis Neural Spine Postzygapophysis Parapophysis Diapophysis
Ventral View: Centrum Transverse Process
Necturus Trunk Vertebrae
Amniotes have a vertebral column showing a much greater degree of regional
specialization than that of other vertebrates. This can easily be seen in Felis. To deal with life on
land the vertebral column became specialized into five subdivisions. Cervical vertebrae are
located in the neck and are designed to increase head movement. Thoracic vertebrae are located
in the upper to mid-back. They are associated with long ribs designed to protect the viscera.
These vertebrae also provide some support for the pectoral girdle. Lumbar vertebrae are the
vertebrae of the lower back. Sacral vertebrae serve to support the pelvic girdle. Caudal vertebrae,
as we have seen previously, are the vertebrae of the tail.
All five regional classes of vertebrae have been modified over evolutionary time to better
perform different duties. As a result, they will have morphological features that should allow
you to distinguish between them. Cervical vertebrae can be differentiated from the other
regional classes by the presences of transverse foramen, holes in the diapophyses/transverse
processes. The transverse foramina allow for passage of the vertebral artery. Cervical vertebrae
tend to show the greatest degree of variation between one another. In amniotes the number of
cervical vertebrae is greater than what we saw in Necturus giving a longer, more flexible neck.
Felis, and most mammals, will have seven cervical vertebrae. They are often referred to by the

18
Representatives of the Five Regional Types of Vertebrae in Felis
Typical Feline Cervical Vertebra, Cranial View
letter “C” and the number of their placement. For example, the first cervical vertebra can be
called “C1”. The first and second cervical vertebrae are modified to increase movement of the
head on the neck. C1 (a.k.a.; atlas) is a ring-shaped bone lacking most of the centrum. The
cranial articulating processes are well developed to articulate with the skull. In Felis they
articulate with the two occipital condyles of the skull. (there is only one occipital condyle in
reptilian and avian skulls.) C2 (or axis) is a modified vertebra having a cranially projecting
process called the dens or odontoid process. The dens is actually the centrum of C1
that detached from C1 and fused to C2 during development. The dens allows for the head to
pivot on the neck

19
Felis C1, The Atlas, Ventral View
Felis C2, The Axis, Lateral View

20
The thoracic vertebrae are the most numerous regional type of vertebra at thirteen and
show some variation. They can be identified by a facet on transverse process for the tubercle of
a rib, and facets/demifacets on the body for the head of the rib. The presence of facets on the
transverse processes and facets or demifacets on the centra is due to the fact that all thoracic
vertebrae articulate with the ribs. The last two thoracic vertebrae and the first lumbar resemble
each other quite a bit. Look for the presence (or absence) of facets on their bodies to distinguish
them. Most times the head of one rib will overlap between two adjacent bodies. When this
occurs each body has half an articulating surface, a demifacet, of the half a rib head that it is
articulating with. Facets can be distinguished from demifacets by appearance (facets look like
full moons and demifacets resemble half moons) and relative location (demifacets are near the
edge of the body). Facets are found on the bodies of T1, T12, and T13. Demifacets are found on
the bodies of T1 through T11. (T1 attaches to ribs R1 and R2.)
Typical Feline Thoracic Vertebrae, Cranial View
There are seven lumbar vertebrae in Felis. Lumbar vertebrae are larger than are thoracic
vertebrae. They have larger centra and processes. This as a structural adaptation since they bear a

21
Felis Thorax Demonstrating a Demifacet
great deal of weight and still must have articulations that allow for a good range of motion in the
lower back. In addition they have mammillary and accessory processes/bodies. The
mammillary body is a rounded elevation on the cranial articulating process. The accessory
process is a more linear elevation that is near the caudal articulating process. The combination
of the accessory and caudal articulating processes allows for better articulation with the cranial
articulating process on the next vertebra. The more caudally located lumbar vertebrae have long,
cranially pointed transverse processes that help to anchor the erector spinae muscles
Typical Feline Lumbar Vertebra, Lateral View

22
Cats possess three sacral vertebrae. These three vertebrae fuse together to form the
sacrum. Although S1 is a large bone, S2 and S3 show a reduction in size. This is due to the fact
that the sacrum articulates with the innominate bones. The pelvis takes over the bulk of the
weight-bearing role. Before identifying the features of the sacrum, you should take the time to
examine S1 and to identify the basic vertebral features.
Felis Sacrum
To deal with its role in the pelvic girdle the sacrum has a number of specialized features.
The easiest to observe is the lateral mass. The lateral mass is actually the fused transverse
processes of the three sacral vertebrae. They are designed to improve the articulation and
weight-bearing role of the iliosacral joint. The descriptive term for the cranial surface of the
lateral mass is ala since it resembles the wing of a bird. The roughened portion of the lateral
mass, where it joins with a correspondingly shaped surface on the ilium (to form the iliosacral
joint), is termed the articular surface. The articular surface loosely resembles a human ear and so
can be referred to as “auricular surface”. (Auricle, or pinna, is the anatomical term for what is
commonly called the “ear”.) The sacral crests are linear elevations found on the dorsal aspect of
the sacrum. One is the median sacral crest. It is a singular elevation formed from the sacral
spinous processes. There are also a pair of lateral sacral crests. These are linear elevations on
the lateral mass. A third set of sacral crest are the paired articular/auricular sacral crests. This is
another pair of linear elevations of the lateral mass located lateral to the lateral sacral crest and
located immediately dorsal to the articular surface. These sacral crests serve as anchoring points
of the erector spinae muscles.
The sacrum is shaped much like an inverted arrowhead. The point of the arrowhead is
termed the apex of the sacrum. There is an opening near the apex of the sacrum called the sacral
hiatus. The sacral hiatus allows for passage of the spinal nerves. This purpose is also served by
the sacral foramina. Sacral foramina are openings between adjacent sacral vertebrae by which the

23
sacral spinal nerves exit the vertebral column. Sacral foramina are the intervertebral foramina of
the sacrum. Flanking the sacral hiatus are horn-like projections called sacral cornua. The sacral
cornua point caudally so as to attach to the first caudal vertebrae by ligaments.
The caudal vertebrae of cats are variable in number. Those located closest to the sacrum
resemble smaller versions of the lumbar vertebrae and show the typical apophyses. The more
distal caudal vertebrae have a very reduced structure. They are centra having reduced
zygapophyses (the role being taken over by ligaments) and a dorsally incomplete vertebral arch.
The caudal vertebrae of Felis will have hemal arches. These inverted “V-shaped” osseous
structures tend to detach from the vertebrae. Due to their appearance they are termed chevron
bones.
Topic B2: The Ribs and Sternum
Ribs are flat bones that articulate with the vertebrae and extend into the body wall. They
are formed intersegmentally, as are vertebral centra. True ribs arise by endochondral
ossification. Some reptiles possess abdominal ribs (a.k.a.; gastralia). These ribs do not arise
endochondrally and so are not considered to be true ribs. Some fishes have two sets of ribs,
dorsal and ventral ribs, associated with each trunk vertebrae. Dorsal ribs project laterally into the
body wall and into the skeletogenous septum. The skeletogenous septum is a collagenous
structure that divides the epaxial and hypaxial muscle groups. Ventral ribs project ventrally as do
mammalian ribs. The occurrence of these ribs varies among fishes. Most fishes have only
ventral ribs. Sharks and a few other fishes have only dorsal ribs. Although the ventral ribs are
positioned where tetrapod ribs are located it is believed that the dorsal ribs of fishes actually gave
rise to the tetrapod rib.
In Squalus the ribs are dorsal ribs. They are short cartilaginous structures that project
into the skeletogenous septum. They articulate with the transverse processes of the trunk
vertebrae.
Squalus Ribs Necturus Ribs

24
In Necturus the ribs are also short structures that project into the skeletogenous septum.
The ribs of urodeles are biccipital meaning “two headed”. The two heads both are on the
vertebral extremity of the rib and are the capitulum and tuberculum. The capitulum articulates
with the parapophysis of the trunk vertebrae. The tuberculum articulates with the diapophysis of
the trunk vertebrae.
In Felis the ribs typically attach at two locations: the thoracic vertebrae and the sternum.
Although not all of the ribs attach to the sternum, most do and many of those will attach by a
cartilage called costal cartilage. Mammal ribs have reduced the Y-shape of the biccipital ribs
found in their reptilian ancestors. The capitulum has become a shorter, more rounded structure
termed the head. The tuberculum has been reduced to a small, round elevation on the neck of the
rib called the tubercle. Both are on the vertebral extremity of the rib. The rib extends ventrally
towards the sternum. As it does it bends closer to the midline. This curve is called the angle.
The sternal extremity attaches to the sternum either by its own costal cartilage (vertebrosternal),
by its costal cartilage attaching to the costal cartilage of a rib cranial to it (vertebrochondral) or
not at all (vertebral).
Vertebral Extremity:
Angle Tubercle Neck Head
Sternal Extremity
Felis Rib Felis Thorax Showing the Sternum
The sternum is found only in tetrapods. Its presence, size, and anatomical features
correlate to the extent that the front limbs are used in locomotion. In most mammals the sternum
is composed of bony plates called sternebrae. Felis has seven sternebrae. The first is called the
manubrium. The last sternebra is termed the xiphisternum. Attached to the xiphisternum, is the
xiphoid. The xiphoid is one of the last skeletal features to ossify. (In humans it ossifies at about
the age of 45.)

25
Topic B3: The Skull
The skull is a complex structure. In comparative vertebrate anatomy the term ”skull” can
be divided into the cranial skeleton and the visceral skeleton. The cranial skeleton is the skull
proper. It can be thought of as the skull minus the jaw. There are two components to the cranial
skeleton: the neurocranium and the dermatocranium. Joints called sutures typically join the
bones of the skull.
The neurocranium is the primary brain case. The neurocranium is also referred to as the
chondrocranium, or endocranium. The neurocranium protects the brain and certain special senses
such as vision and olfaction. It is composed of cartilage in primitive vertebrates such as Squalus.
Over evolutionary time it has become increasingly ossified. The neurocranium begins as
cartilage and is subsequently partially or completely replaced by bone in all vertebrates
excepting the chondrichthyes. For this reason the neurocranium of chondrichthyes is referred to
as the chondrocranium. As a result, the bony component of the neurocranium is composed of
bones that have formed by endochondral ossification
Primitive vertebrates had bones in the integument that were part of a protective covering
termed dermal armor. The integumentary bones of the head migrated subdermally to form the
dermatocranium. Since these bones were once integumentary structures that developed by
intramembranous ossification, the bones of the dermatocranium today will also arise by
intramembranous ossification.
The visceral skeleton is also known as the splanchnocranium. As was the case with the
neurocranium, it is composed of cartilage in primitive vertebrates. It also became ossified as
membrane bones were added from the integument surrounding the skull. The splanchnocranium
is composed of four skeletal structures. One is the embryonic upper jaw cartilage called the
palatopterygoquadrate and its replacement bones. A second is the embryonic lower jaw cartilage
called the Meckel’s cartilage and its replacement bones. A third is the skeleton of the
Squalus Chondrocranium (white tagged) and Splanchnocranium (green tagged)

26
hyoid. A fourth is the skeleton of the branchial arches, the gill arches and their derivatives in
tetrapods.

27
The skull of Squalus consists of the chondrocranium and the splanchnocranium. Both the
chondrocranium and splanchnocranium of chondrichthyes are composed entirely of cartilage in
the adult. The only exception is the teeth. In sharks and rays the neurocranium demonstrates
complete fusion of all its components into a singular cartilaginous structure, the chondrocranium.
Some other fishes have a primarily cartilaginous neurocranium in the adult (ex; sturgeons).
However, they lack a complete roof, or tectum, of cartilage to protect the brain. This role is
handled by the dermatocranium. Squalus, as is true for all sharks and rays, lacks a
dermatocranium.
The walls of the chondrocranium are fully developed in the adult and include a posterior
occipital wall that is lacking in agnathans. The occipital wall of the chondrocranium will have an
opening for the spinal cord called the foramen magnum. On each side of the foramen magnum
will be the occipital condyles that will articulate with the first vertebra. The chondrocranium will
house and protect the brain and some of the special senses. It will have a pair of orbits in which
the eyes are located. The orbit will not fuse to the chondrocranium to allow for movement of the
eye. The orbit is bracketed by the anterior orbital process, supraorbital crest, and postorbital
processes dorsally and the antorbital shelf and infraorbital shelf ventrally running cranial to
caudal in both cases. It also contains a small mushroom-shaped structure called the optic
pedicle. The capsules of those special senses that do not require movement to function will fuse
Olfactory Capsule (damaged) Deep Ophthalmic Foramen
Olfactory Foramen Trochlear Foramen Trigeminofacial Foramen
Optic Foramen Occulomotor Foramen Hyomandibular Foramen
Basal Plate showing Notochord
Squalus – Chondrocranium, Lateral View Showing Foramina

28
to the chondrocranium. The otic capsules house the internal ear and are fully fused to the
posteriolateral walls. The internal ear allows for the sensation of hearing, static equilibrium, and
dynamic equilibrium. The olfactory capsules are fully fused to the anteriolateral walls and will
contain the olfactory bulbs.
The floor of the chondrocranium is composed of the basal plate and the ethmoid plate.
The basal plate will fuse to the otic capsules during development. It will contain the notochord.
The notochord is visible as a ridge extending cranially from the foramen magnum on the ventral
aspect of the chondrocranium. An opening will remain in the chondrocranium where the basal
and ethmoid plates fused during development. This opening is the hypophyseal fenestra. This
opening remains to accommodate the carotid arteries and the pituitary gland (aka; hypophysis).
The portion of the opening visible on the ventral surface of the chondrocranium is the carotid
canal. The hypophyseal fenestra has developed a saddle shaped structure called the sella turcica
to house the pituitary gland. The ethmoid plate component of the floor of the chondrocranium is
fused to the olfactory capsules. The ethmoid plate will extend anterior to the olfactory capsules
as a blade-like extension called the rostrum. The rostrum will support the snout of the shark and
is very thin walled. The rostrum is the final portion of the chondrocranium to chondrify,
especially at the junction between the rostrum and the rest of the chondrocranium.
Olfactory Capsule Rostral Fenestra Precerebral Cavity Epiphyseal Foramen
Endolymphatic Foramen Endolymphatic Fossa Perilymphatic Foramen
Squalus-Dorsal Chondrocranium Showing Foramina

29
Otic Capsule Optic Pedicle Supraotic Crest Postorbital Process Supraorbital Crest
Hyomandibular Foramen Trigeminofacial Foramen Optic Foramen
Squalus-Chondrocranium Showing Optic Pedicle
The chondrocranium will have a number of openings to allow for passage of blood
vessels and nerves. These include the olfactory foramina on the anterior aspect of the olfactory
capsules. There are a number of foramina associated with the orbit. They include the superficial
ophthalmic foramina, deep ophthalmic foramina, occulomotor foramina, optic canals, trochlear
foramina, and trigeminofacial foramina. Dorsal and medial to the orbits will be the epiphyseal
foramen. On the posteriodorsal aspect of the chondrocranium is a depression called the
endolymphatic fossa that will house endolymphatic foramina and a second depression called the
perilymphatic fossa that will house the perilymphatic foramina. The otic capsule will have the
hyomandibular foramina (to anchor the articulation of the hyomandibula with the
chondrocranium). A pair of extensions will be found on the caudal aspect of the otic capsule
called the postotic processes. These processes will each have an opening covered by a thin plate
of cartilage termed the postotic fenestra. Between the postotic process and foramen magnum will
be the glossopharyngeal foramen and the vagus canal respectively.

30
Centrum of First Vertebra Vagus Foramen Postotic Process
Occipital Condyle Foramen Magnum Glossopharyngeal Foramen Postotic Fenestra
Squalus – Chondrocranium, Posterior View (upside down)
The visceral skeleton, or splanchnocranium, derives from the pharyngeal arches. In
fishes the splanchnocranium forms the jaws and gill arches. In Squalus the splanchnocranium
consists of the cartilages of each pharyngeal arch and the median basihyal and basibranchial
cartilages of the pharyngeal floor. The skeleton of each arch follows very closely to a basic
pattern. All but the first and last pharyngeal arches support gills.
The first pharyngeal arch is called the mandibular arch and forms the jaws. The
mandibular arch is a pharyngeal arch that has been modified for feeding. It consists of two
cartilages forming the upper and lower jaw respectively. The palatopterygoquadrate cartilage
forms the upper jaw. It has two pairs of processes. One pair of processes enters into the orbits
(one process per orbit) and so they are termed the orbital processes. There is also a pair of
quadrate processes, each projecting laterally. The quadrate cartilages, along with the
hyomandibula, are responsible for the suspension of the jaws from the chondrocranium. The
lower jaw is composed of Meckel’s cartilage. It will articulate with the palatopterygoquadrate
and the hyoid on each side. A pair of slender labial cartilages flanks the jaws. They have no
known function.
The second pharyngeal arch is called the hyoid arch. It is also a modified pharyngeal arch
although its role in feeding is more limited. The hyoid arch consists of 5 cartilages: 1

31
Occipital Condyle Postotic Process Postorbital Process Antorbital Process
Basal Plate Postorbital Shelf Antorbital Shelf
Supraotic Crest Carotid Foramen Supraorbital Crest Rostral Fenestra Rostral Carina
Superficial Ophthalmic Foramina
Squalus-Chondrocranium, Ventral View
singular and 2 paired groups. Dorsally are the right and left hyomandibular cartilages /
hyomandibulae. These are short cartilages that articulate with the posterior aspect of the
chondrocranium. Immediately below the hyomandibular cartilages are the right and left
ceratohyal cartilages. The ceratohyal cartilages bear gills and extend under the chondrocranium.
The ventral aspects of the ceratohyal cartilages attach with the singular basihyal cartilage located
midventrally. Meckel’s cartilage and the palatopterygoquadrate cartilage articulate with each
other at the angle of the mouth and will also articulate with the hyomandibular cartilages of the
hyoid arch. Ligaments support these articulations. The hyomandibulae are bound by ligaments to
the otic capsules and suspends the jaws and the entire branchial skeleton from the
chondrocranium. This is an example of a hyostolic jaw suspension.
Pharyngeal arches 3 through 7 have the same basic structure. All but #7 bear gills. The
dorsal most portion of these pharyngeal arches are the pharyngobranchial cartilages. The
pharyngobranchial cartilages attach ventrally to the epibranchial cartilages. The epibranchial
cartilages attach ventrally to the ceratobranchial cartilages. Both the epibranchial and

32
ceratobranchial cartilages typically will bear gill rakers and gill rays. The ceratobranchial
cartilages attach ventrally to the hypobranchial cartilages. The paired hypobranchial cartilages
attach to the singular, midventrally located basibranchial cartilage. There are only two
basibranchial cartilages. The third pharyngeal arch attaches to the first basibranchial while
arches 4 through 7 share a common, larger basibranchial cartilage.
First Visceral Arch: Palatopterygoquadrate + Meckel’s Cartilage
First Epibranchial Last Pharyngeobranchial
Second Visceral Arch: Basihyal + Ceratohyal + Hyomandibula Gill Rays
Squalus- Splanchnocranium, Lateral View

33
First Visceral Arch: Meckel’s Cartilage + Palatopterygoquadrate
Second Visceral Arch: Ceratohyal + Basihyal
First Ceratobranchial First Hypobranchial Basibranchial
Squalus-Visceral Splanchnocranium

34
Otic Capsule Quadrate Process Orbital Process Antorbital Process Antorbital Shelf
Palatopterygoquadrate Meckel’s Cartilage Labial Cartilage (encased in flesh)
Squalus Chondrocranium and Splanchnocranium Showing Jaw Articulation
In Necturus the skull consists of the splanchnocranium and neurocranium. Unlike
Squalus, the neurocranium of Necturus is mostly ossified and consists of both endochondral and
membrane bones. The endochondral bones derive from components of the cartilaginous
chondrocranium during development. The membrane bones are homologous to the dermal
armor of more primitive vertebrates and so are collectively called the dermatocranium. As a
result the neurocranium of Necturus is also referred to as the “neurocranial-dermatocranial
complex”.
The neurocranium of the developing Necturus is initially entirely cartilaginous. Some of
these cartilages are maintained in the adult (such as the antorbital cartilages) but many are
replaced by bone through endochondral ossification. As is the case with Squalus, the floor of the
cartilaginous neurocranium includes the ethmoid plate. This plate will partly ossify to give the
ethmoid bone. Posteriolateral to the ethmoid plate will be the right and left quadrate cartilages.
The quadrate bones will replace these paired cartilages. The quadrate bones will articulate with
the right and left palatopterygoid bones (one of the dermal bones that will ensheath the
palatopterygoquadrate cartilage). Posterior to the quadrate cartilages in the developing skull will
be the otic capsules. They will include the prootic and opisthotic cartilages, which will be
replaced by the prootic and opisthotic bones respectively. Between them will be some remnant

35
cartilage in the adult. This cartilage has a perforation called the fenestra ovalis. The fenestra
ovalis will be covered by a membrane (the internal tympanic membrane) on which sits the
columnella/stapes. The columnella is the first auditory ossicle to appear in vertebrates. It will
have a tiny, spike-like projection called the stylus and will conduct sounds waves into the otic
capsule. (Note: the quadrate bone will become one of the mammalian ossicles, the incus.)
Caudal and medial to the otic capsules will be the foramen magnum and the cartilages that
surround it. These cartilages will be the supraoccipital arch, basioccipital arch, and exoccipitals.
Through endochondral ossification the supraoccipital, basioccipital, and exoccipital bones will
replace these cartilages respectively. The exoccipital bones will possess the occipital condyles to
articulate with the cervical vertebra.
Exoccipital Parietal Squamosal Quadrate Palatopterygoid Frontal Premaxilla
Otic Capsule
Necturus – Skull, Dorsal View
The dermatocranium will include bones that will ensheath the palatopterygoquadrate
cartilage to form the new upper jaw. These bones include the premaxillae, vomers (which also
form a portion of the primary palate), and palatopterygoids (which articulate with the quadrate
bones). All of these bones bear teeth. The roof of the skull is formed by the frontal and parietal
bones. The parietal bone will form the roof of the otic capsule. Lateral to the parietal bone will

36
be the squamosal. The squamosal will articulate with the quadrate anteriorly. The floor of the
skull will be ensheathed by the dermatocranial bone called the parasphenoid. The parasphenoid,
along with the vomers, will form the primary palate. The parasphenoid and parietals will obscure
the basioccipital and supraoccipital bones respectively.
Epibranchials Squamosal Quadrate Angular Palatopterygoid Dentary Premaxilla
Opisthotic Parietal Frontal
Necturus – Skull, Dorsal View 2

37
Premaxilla Vomer Frontal Palatopterygoid Parietal Squamosal
Dentary Splenial Meckel’s Cartilage Angular Quadrate
Necturus – Skull, Lateral View
Exoccipital Foramen Magnum
Necturus – Skull, Posterior View

38
Premaxilla Ethmoid Vomer Parasphenoid Prootic Fenestra Ovalis Opisthotic
Palatopterygoid Quadrate Squamosal Stylus Columnella Exoccipital
Necturus– Skull, Ventral View
The splanchnocranium of Necturus includes the jaws, hyoid, and gill arches. The hyoid
and gill arches remain as cartilage throughout the life of the mudpuppy. The cartilages of the
embryonic jaws will become ensheathed in bone derived from dermal armor. As we have
already seen, the palatopterygoquadrate becomes encased in the premaxillae, vomers, and
palatopterygoid (some times referred to as the “pterygoid”). The dentary, splenial, and angular
bones will cover most of the Meckel’s cartilage of the lower jaw. Some Meckel’s cartilage will
remain near the jaw joint above the angular. The jaw joint is between the angular and quadrate
bones. Both the dentary and splenial will bear teeth. The dentary is equivalent to the
mammalian mandible. The mandibular arch is the first visceral arch.
The hyoid arch will be composed of two pairs of cartilages. The basal cartilage pair is
the hypohyals that will attach to the first basibranchial cartilage of the gill arches. Attached to
the hypohyals will be the ceratohyals. The hyoid comprises the second visceral arch.

39
Angular Meckel’s Cartilage Splenial Dentary Mental Symphysis
Angular Dentary
Necturus Mandible
(Top-Left, Internal View Bottom-Right, External View)
Necturus possesses three pairs of gills (visceral arches 3, 4, and 5). Their skeletal
structures are cartilaginous throughout life. Unlike the gills of Squalus, the gills of Necturus are
external. These structures can be moved by muscles to better direct them into water currents so
as to pick up more oxygen. The first gill arch pair is the best developed. The second and third
are anchored on to the first. The skeleton of the gills in Necturus has two basibranchial
cartilages. One is larger and anterior to the other. This first basibranchial cartilage will attach to
the hyoid and the first pair of ceratobranchial cartilages. The second basibranchial will be partly
ossified and attaches only to the first basibranchial. There are two pairs of ceratobranchial
cartilages. The first pair is large and well developed. They will attach to the first basibranchial
cartilage inferiorly and the first pair of epibranchial cartilages superiorly. The second pair of
ceratobranchial is very reduced and attach to the first ceratobranchials. These cartilages support
the second and third pairs of epibranchials. There are three pairs of epibranchial cartilages. The
first pair attached to the first ceratobranchial cartilages while the second and third pair attach to
the second ceratobranchials. The first epibranchial is the largest of the three and the third is the
smallest.

40
Dentary Splenial Angular Ceratohyal Basibranchial 2 Ceratobranchial1 & 2
Hypohyal Basibranchial 1 Epibranchial 1, 2, & 3
Note; Viscera Arch 1 = Jaws/Mandibular Arch
Visceral Arch 2 = Hyoid/Hyoid Arch
Visceral Arch 3 = Ceratobranchial 1 + Epibranchial 1
Visceral Arch 4 = Ceratobranchial 2 + Epibranchial 2
Visceral Arch 5 = Ceratobranchial 2 + Epibranchial 3
Necturus - Splanchnocranium
The skull of Felis is more complex that that of either Necturus or Squalus. Mammalian
skulls have evolved features that will differentiate them from the skulls of other vertebrates.
There are four developments distinguishing mammalian skulls. One is the development of the
dentary as the only bone of the lower jaw. A second is an altered site of articulation for the
lower jaw. This ties in with the third development, the presence of three pairs of auditory
ossicles. A fourth development is alterations of the secondary palate (Note: Squalus and
Necturus have only a primary palate). The secondary palate separates the nasal and oral cavities.
The neurocranium is incomplete dorsally resulting in the presence of fontanels in the skull of
a newly born mammal. These fontanels allow for increased growth of the skull and the ability of

41
Some Features of the Skull in Felis
the skull to pass through the birth canal. Endochondral ossification of the neurocranium will
result in a number of bones. The neurocranium contributes the basioccipital, basisphenoid, and
presphenoid bones to form the floor of the cranium. It will also contribute a portion of the lateral
walls of the cranium with the alisphenoid and orbitosphenoid bones. Each alisphenoid will
possess a wing shaped structure that projects posteriorly called the pterygoid process. This
process will have a hook-like projection called the pterygoid hammulus. The pterygoid
processes and hammuli help to anchor the soft palate. The supraoccipital and exoccipital will
form the posterior wall. In addition the embryonic ethmoid plate will partially ossify to give rise
to the ethmoid bone. The ethmoid bone and cartilages will house the olfactory bulbs and support
the olfactory mucosa. Ossification areas in the otic capsules will form the petrosal (a.k.a.;
periotic) bones that house the structures of the middle ear. As was the case with Squalus, the
hypophysis rests in the sella turcica. In mammals the sella turcica is a component of the
basisphenoid.
Due to its evolutionary origins the dermatocranium will contribute bones through
intramembranous ossification. In mammals it will contribute paired and unpaired bones. Paired
dermatocranial bones include: premaxillae, maxillae, parietals, malars (a.k.a.; jugals), nasals,
lacrimals, and squamosal bones. Unpaired bones include the frontal and the interparietal. The
temporal complex consists of both endochondral and membrane bones. The squamous bone is of
dermatocranial origin and contributes to the lateral walls of the skull. The tympanic bulla is a
large swelling near the external auditory meatus. It is made up of two components: tympanic
and endotympanic. The tympanic bone surrounds the tympanic membrane and is derived from
the angular bone of the dermatocranium in nonmammals. Near the tympanic bulla is another
portion of the temporal complex called the mastoid. The mastoid is also of dermatocranial origin
and will have a projection called the mastoid process. The temporal complex includes the
endochondrally derives petrous portion. The squamosal portion is the new site of jaw

42
Premaxilla Ethmoid Turbinates Malar Orbit Frontal Temporal Fossa Squamosal Parietal Sagittal Crest
Maxilla Perpendicular Plate of Ethmoid Lacrimal Interparietal Supraoccipital Lambdoidal Crest
Felis Skull Dorsal View
Felis Skull Ventral View

43
articulation. The remaining bones of the quadrate-articular joint migrate into the ear to become
the auditory ossicles (the articular becomes the malleus and the quadrate becomes the incus).
Felis Skull Ventral View

44
Felis Skull Lateral View
Premaxilla Maxilla Lacrimal Canal Planum Frontal Parietal Squamosal Sagittal Crest
Lacrimal Postorbital Processes Supraoccipital
Canine Premolars Malar Zygomatic Process of Malar External Auditory Canal Tympanic Bulla
Zygomatic Process of Squamosal Mastoid
Felis Skull Lateral View

45
Felis Skull, Sagittal Sections
Mammalian skulls have openings within certain bones called cranial sinuses. In Felis the
sinuses are found in the frontal, ethmoid, maxilla, and sphenoid bones. Mammals will have a
primary and secondary palate. Primary palates are found in all vertebrates. The primary
mammalian palate includes an unpaired vomer that makes up part of the nasal septum. The nasal
septum also includes a portion of the ethmoid, mesethmoid, and cartilage. The secondary palate

46
is found only in amniotes. It is a complete palate in mammals and includes the palatine, the
maxillae, and the pterygoid processes of the sphenoids.
In humans the number of bones in the skull has been reduced when compared to that of
the cat. The basioccipital and exoccipitals form the occipital bone. The basisphenoid,
presphenoid, alisphenoids, and orbitosphenoids all join to form the sphenoid bone. The
squamous, mastoid, and petrous form the temporal bone.
Felis Skull, Sagittal Sections
All mammal skulls are synapsid. Synapsid skulls will have an opening on each side
called the temporal fossa. This serves as an anchoring point for the muscles that close the jaw.
Under the temporal fossa will be a bridge-like structure called the zygomatic arch. The
zygomatic arch is formed by the zygomatic process of the malar joining with the zygomatic
process of the squamosal.
The skull of Felis has numerous openings. The openings of the skull typically allow for
the passage of nerves and/or blood vessels. (For example, the hypoglossal canal allows for the
passage of cranial nerve 12, the hypoglossal nerve.) A number of these openings are located in
or near the orbit and include: the lacrimal canal, orbital fissure, infraorbital foramen, optic
canal/foramen, foramen rotundum, and foramen ovale. Others are found on the ventral aspect of
the skull such as the carotid canal/foramen, jugular foramen, hypoglossal canal/foramen, anterior
palatine foramen, posterior palatine foramen, and stylomastoid foramen. The external acoustic
canal is located at the tympanic bulla. The foramen magnum is located on the posterior aspect of
the skull. A few foramina are internal structures such as the internal acoustic canal and olfactory
foramina of the cribriform plate.
An interesting internal aspect of the feline skull is the tentorium. This bony partition
serves to separate the cerebrum from the cerebellum. You will also notice that these bones

47
making up the inner aspect of the cranium will have the impression of cerebral blood vessels and
of the cortex.
Felis Skull Showing a Close Up of Some Foramina
The splanchnocranium of Felis includes the jaws, the hyoid, and some of the
laryngeal cartilages. The upper jaw is incorporated into the premaxillae and maxillae. The
lower jaw has become one bone (actually two that join to form one bone during development),
the dentary or mandible. This represents a reduction in the number of bones making up the
lower jaw when compared to Necturus or to a reptile. Over evolutionary time the size of the
muscles of mastication became larger. This required more room to anchor on to the lower jaw, in
particular the dentary component of the lower jaw. As a result, the dentary became larger and
the other bones of the reptilian lower jaw were reduced in size. Some bones were lost and others
developed new roles. The articular and quadrate bones of the reptilian jaw joint are two of the
three auditory ossicles in mammals. In mammals the lower jaw articulates at the temporal bone
by means of the mandibular condyle. The mandible has a masseteric fossa and coronoid process
on each side to allow for muscle attachment.

48
Felis Mandible
Felis Mandible
The hyoid apparatus of Felis is a delicate bony structure that is often lost from articulated
skeletal specimens. It is composed of 11 bones, 5 paired and 1 unpaired. There is a central,
singular basihyal. It is located immediately superior to, and superficial to, the larynx. Extending
posteriorly from the basihyal are the paired thyrohyals. The thyrohyal bones will be on either
side of the thyroid cartilage of the larynx. The remaining hyoid components extend cranially
from the basihyal. The first pair to come off of the basihyal is the ceratohyals. The thyrohyals
are attached to the ceratohyals and are near the angle of the mandible. The stylohyals arise from
the thyrohyals and extend posteriorly to lie near the mastoid bones. In humans the stylohyal will
break off of the hyoid apparatus and attach to the temporal bone to form the styloid process. The

49
tympanohyals arise from the stylohyals and extend posteriorly. They are located near the
tympanic bulla.
Thyrohyal Basihyal Ceratohyal
Thyrohyal Ceratohyal Epihyal Stylohyal
Felis Hyoid Apparatus, posterior view (damaged)
Chapter 1: Skeletal Structure

50
Part C: The Appendicular Skeleton
Topic C1: The Pectoral Girdle and Appendage
Both the pectoral limb and girdle are entirely cartilaginous in sharks. The pectoral limb
of Squalus is the pectoral fin. The pectoral girdle consists of the coracoid bar (composed of a
two coracoid cartilages), the paired scapulae, and the paired suprascapulae. The scapula has the
point of articulation between the pectoral fin and girdle called the glenoid. The proximal
cartilages of the pectoral fin are the basalia. There are three basalia per fin. Anteriormost is the
propterygium, then the middle basalia is the mesopterygium, and the posterior element is the
metapterygium. Two rows of radial cartilages attach to the basal cartilages. Anchored into the
distal row of radialia are the numerous ceratotrichia.
The tetrapod pectoral limb and girdle evolved from the pectoral fin and girdle. The
Ceratotrichia Radial Cartilages Metapterygium Suprascapular Cartilage
Mesopterygium Propterygium Glenoid Surface Scapula Coracoid Bar
Squalus Pectoral Girdle
basalia were rearranged with one bone being proximal and the other two becoming paired and
more distal. The proximal basalia became the humerus. The paired distal basalia became the
radius and ulna. The radialia became the carpal bones. Although the evolution of the pectoral
limb is well documented by the fossil record, the origin of the digits is still under some debate.
The basic pectoral girdle developed in Devonian Period fishes and has been modified by
subsequent vertebrates. These primitive fishes had pectoral girdles composed of 7 paired

51
elements. Three pairs are endochondral bones of the endoskeleton. These were the suprascapula,
scapula, and coracoid. Four pairs are membrane bones derived from the ancestral dermal armor.
These were the postemporal, supracleithrum, cleithrum, and clavicle. Since chondrichthyes are
cartilaginous, the pectoral girdle of Squalus consists only of the endochondral component. Over
time the number of components to the tetrapod pectoral girdle were reduced. The suprascapular
bones of the endochondral component were lost in bony fishes and most tetrapods. In tetrapods
the intramembranous component has mostly been lost leaving only the clavicle. A new
membrane bone developed in amphibians, the interclavicle. The interclavicle has been retained
in most vertebrates including many reptiles, birds, and monotreme mammals. The scapula is
maintained in all limbed tetrapods since it has the glenoid, the point of articulation between the
humerus and pectoral girdle. Both the clavicle and coracoid brace the pectoral girdle against the
ribcage. As a result, over evolutionary time, one has been lost and the other has been maintained.
Modern amphibians and reptiles possess a coracoid but lack a clavicle. In birds the clavicles have
been maintained and the coracoids have been lost. In mammals the clavicles have become the
dominant bracing bone. The coracoids have been reduced to small projections of the scapula.
The pectoral girdle of Necturus is composed of both bone and hyaline cartilage. There is
a pair of cartilaginous coracoids. These coracoids are ventrally oriented. Extending cranially
from the coraocoids are the long, slender procoracoids. The dorsal component of the pectoral
girdle is the paired scapula and suprascapula. The scapula will have both a bony and
cartilaginous component. The glenoid is on the cartilage component. The suprascapula is a thin
piece of cartilage and is attached to the bony component of the scapula.

52
Necturus Pectoral Girdle and Limb (both are from the right side of the body)
The pectoral limb includes a humerus, radius, ulna, carpal bones, metacarpal bones, and
the phalanges. The humerus is the bone of the “arm” in Necturus. The radius and ulna are
paired bones of the “forearm”. They do not show as many distinguishing features in amphibians
as they do in mammals and may be hard to tell apart. There are differences. The proximal
epiphysis of the ulna is the larger epiphysis to articulate with the humerus while the distal
epiphysis of the radius is the larger to articulate with the carpals. The carpal bones are small
short bones arranged into two rows to support the metacarpus. There are four metacarpal bones
to support the four digits. The digits, or phalanges, are composed of three phalanx bones each:
proximal, middle, and distal.
Felis Clavicle
The pectoral girdle of Felis consists of the scapula and a very reduced clavicle. The size
of the clavicle in mammals corresponds to the degree that it is required for locomotion. In bats
and moles they are large while in cetaceans they are lost. In cats the clavicle’s role has been
replaced by the pectoral musculature. The pectoral musculature forms a sling that supports the
pectoral limb. As a result, the clavicle of Felis is fairly small and easily lost. Many students will
not even notice it when they are dissecting the cat. The scapula is a flat bone possessing a
number of projections for anchoring muscle including the coracoid process, acromion, and spine.
It has three depressed surfaces. Dorsally there are the supraspinous fossa and infraspinous fossa.
Ventrally there is the subscapular fossa.

53
Felis Scapula
The arm has one bone (each), the humerus. The humerus is a long bone and has the
features typical to that class of bone organ. The proximal epiphysis is rounded to allow for
rotational movement and is called the “head”. The humerus has a number of points for muscle
attachment including the greater tubercle, lesser tubercle, deltoid ridge, pectoral ridge/crest,
epicondyles, and supracondylar crests. Between the greater and lesser tubercle is the
intertubercular/biccipital groove. One of the two tendons of the biceps brachii runs through this
groove. The distal epiphysis is expanded to allow for greater articulation giving the lateral and
medial condyles. The condylar surfaces are shaped to better articulate with the radius and ulna.
The surface for the ulna is the trochlea; a spool-shaped surface that allows for the hinge-like
movement of the humerus-ulna. The surface for the radius is the capitulum; a rounded surface
that allows for the rotational movements of the radius. The radius-capitulum articulation actually
represents a ball-and-socket joint.

54
Felis Humerus
The forearm is composed of two bones, the radius and ulna that are parallel to each other.
Running the length of their diaphyses, on their opposing surfaces are the interosseous crests.
The interosseous crest anchors the interosseous ligament which helps to maintain the proper
alignment between these bones. The ulna will articulate with the humerus at the trochlea. It will
have a notch that the trochlea fits into called the semilunar, ulnar, or trochlear notch. The
trochlear notch is found between two processes, the coronoid process and the olecranon. The
olecranon is an anchoring point for the triceps brachii. Distally the ulna narrows and will have a
spike-like process called the styloid process. The radius is wider distally to support the first row
of carpal bones. It will also have a styloid process on its distal epiphysis. The proximal epiphysis
will articulate with both the humerus (at the capitulum) and the ulna (at the radial notch).
Styloid Process Shaft Coronoid Process Trochlear Notch Olecranon

55
Radial Notch (on coronoid process) Interosseous Crest Distal Epiphysis
Felis Ulna
(Top Ulna-Right, lateral view Bottom Ulna-Right, Medial View)
L. Radius, Ventral Radial Tuberosity Interosseous Crest Distal Epiphysis Styloid Process
R. Radius, Dorsal Fovea for Capitulum Diaphysis Distal Epiphysis Styloid Process
Felis Radius
The carpus is composed of 7 bones arranged into two rows. The proximal row is
composed of the scapholunar on the radial side, the triquetrum on the ulnar side, and the
pisiform. The scapholunar is actually two fused carpal bones, the scaphoid and lunate as is seen
in humans. The distal row is made up of the trapezium (on the radial side), trapezoid, capitate,
and hamate (on the ulnar side). The metacarpus is composed of five bones numbered first
through fifth from the dewclaw side. Typically there are five digits. All but the first are
composed of three bones: proximal phalanx, middle phalanx, and distal phalanx that has the
claw. The dewclaw has only two phalanx bones, the proximal and distal, and as a result a special
name is given to this digit: pollex.

56
Felis Right Forefoot, Dorsal View
Topic C2: The Pelvic Girdle and Appendage
The pelvic fin and girdle of Squalus are composed of cartilage. While the structure of the
pelvic fin is similar to that of the pectoral fin, the girdles have some notable differences. The
pelvic girdle in fishes is poorly developed and is not as closely associated with the vertebral
column as is the pectoral. The pelvic girdle of Squalus consists of an ischiopubic bar. The
ischiopubic bar will have small, lateral projections called iliac tubercles/processes and the
articulating surface of the pelvis, the acetabulum. The pelvic fin has only two basalia, the
propterygium and the metapterygium. The propterygium is small and can be misidentified as a
radial cartilage. The metapterygium is large and will support a single row of radial cartilages
Acetabular Surface Ischiopubic Bar Iliac Process Propterygium

57
Ceratotrichia Radial Cartilages Clasper Cartilage Metapterygium
Squalus Male Pelvic Girdle
Acetabular Surface Ischiopubic Bar Iliac Process Propterygium
Ceratotrichia Radial Cartilages Metapterygium
Squalus Female Pelvic Girdle
which in turn support the ceratotrichia. The metapterygium will support the clasper cartilages in
male chondrichthyes. The claspers are the intromittent organs in these fishes.
Sacrum Sacral Rib Ilium

58
Pubis Puboischiadic Plate Ischium
Necturus Pelvis
The pelvic girdle of Necturus consists of both bone and cartilage. Tetrapod pelvic girdles
will begin as cartilage but tend to ossify during development. The greater strength of bone
allows the pelvis to withstand the pressure that it will be exposed to during life. However, since
Necturus is aquatic, the pelvis of this species is not exposed to the amount of mechanical stress
typical for a tetrapod. There are three paired components to a tetrapod pelvic girdle: the ilium,
ischium, and pubis. The ilium will articulate with the sacrum. Collectively these three pairs of
skeletal structures will have the joint cavity, the acetabulum. The combination of the pelvic
girdle and sacrum forms the structure known as the pelvis. In Necturus the ilium is a slender
bone that articulates with the sacrum by means of a sacral rib. The pubis is cranial and the
ischium is caudal relative to one another. The pubis is composed of cartilage and will be pointed
in Necturus. The ischia are composed of both cartilage and bone. The pubis and ischial
cartilages will grow together during development to form the puboischiadic plate. The
puboischiadic plate will have a pair of openings called the obturator foramen for passage of the
obturator nerve.
The thigh of Necturus has the femur, which articulates at the acetabulum of the pelvic
girdle. The tibia and fibula are the paired bones of the leg. As is the case with the radius and
ulna these paired bones each possess an interosseous margin to anchor the interosseous ligament.
The two bones are similar in size and appearance. The fibula is the laterally oriented bone. The
tarsal bones are small short bones that support four metatarsals. The four metatarsals support
four phalanges composed of three phalanx bones each

59
Necturus Pelvic Girdle and Limb
Felis Os Coxa
The pelvic girdle of Felis is composed of the paired hip bones and the sacrum. The hip
bone is called the os coxa or innominate and is made up of three fused bones: the ilium, ischium,
and pubis. The two os coxae articulate with the sacrum to form the pelvis. The os coax has a

60
large opening called the obturator foramen that decreases the weight of the pelvis and allows for
passage of the obturator nerve. The ilium is the dorsal bone of the os coxa. The ischium and
pubis are ventral relative to the ilium. The pubis is cranial and the ischium is caudal.
Felis Femur
The thigh is composed of one long bone, the femur. The femur, like the humerus, as a
rounded portion to the proximal epiphysis termed a “head”. As is typical of the appendicular
skeleton, the femur has many points of muscular attachment including: the greater trochanter,
lesser trochanter, intertrochanteric crest, gluteal tuberosity, linea aspera, lateral and medial
epicondyles, and popliteal surface. The distal epiphysis is expanded, giving rise to the lateral and
medial condyles, to allow for greater articulation with the leg. Anteriorly the distal epiphysis has
a roughened point for attachment of the patellar tendon called the patellar surface. This allows
for attachment of a sesamoid bone called the patella. The patella reinforces the knee joint.
The leg is composed of two bones, the tibia and fibula that are parallel to each other. Running
the length of their diaphyses, on their opposing surfaces are the interosseous margins. The
interosseous margin anchors the interosseous ligament that helps to maintain the proper
alignment between these bones. The tibia is the weight bearing bone of the leg and will articulate
with the femur. To facilitate this the tibia will have well developed condyles. Near the condyles
will be a roughened elevation, the tibial tuberosity, which anchors the patellar tendon. The distal
epiphysis of the tibia has a flattened process called the medial malleolus. In mammals the fibula
is much more slender than the tibia. The proximal epiphysis will have a styloid process and the
distal epiphysis will have the lateral malleolus.

61
apex
Felis Patella
Felis Tibia
Styloid Process L. Fibula Medial View Interosseous Margin Distal Epiphysis
Lateral Malleolus Peroneal Groove L. Fibula Lateral View Head of Fibula Styloid Process
Felis Fibula

62
The hind foot of Felis is composed of the tarsals, metatarsals, and phalanx bones. There
are seven tarsal bones arranged into three rows. The proximal row contains the talus and the
calcaneus. The talus articulates with the tibia and the calcaneus articulates with the fibula and
talus. The middle row is composed of one short bone, the navicular. The distal row is composed
of four bones. There are three cuneiforms and they articulate with the navicular. They are
named the medial, intermediate, and lateral cuneiforms. Each cuneiform supports one metatarsal.
Lateral to the lateral cuneiform is the cuboid. The cuboid supports the fourth metatarsal. Felis
has four metatarsals bones in the pes. They are numbered one through four counting medial to
lateral. Each metatarsal supports one phalange that is composed of three phalanx bones.
Distal Phalanx Middle Phalanx Proximal Phalanx Metatarsal Cuboid Calcaneus
Lateral Cuneiform Intermediate Cuneiform Medial Cuneiform Navicular Talus
Felis Hind Foot Dorsal View

63
Chapter 2: Myology
Part A: An Introduction
Myology is a study of muscles; muscle cells, muscle tissue, and muscle organs. In this
chapter we will be focusing on skeletal muscle organs. Skeletal muscle organs are composed
primarily of skeletal muscle tissue but also include blood vessels, nerves, and connective tissue
sheaths. There are three connective tissue sheaths associated with skeletal muscle organs. A
muscle cell will be elongated. This allows the muscle to contract, to shorten its length causing it
to pull on a bone and produce a movement. As a result, a muscle cells is called a myofiber or
fiber. Each myofiber in a skeletal muscle organ will be ensheathed by a thin connective tissue
membrane called the endomysium. Groups of endomysium encased myofibers form an
organizational unit called a fascicle. The fascicle will be covered in another connective tissue
sheath called the perimysium. Groups of fascicles will form a skeletal muscle organ. A tough
connective tissue sheath called the epimysium will cover the organ. The epimysium is rich in
collagen bundles. During development it will extend from the muscle to meet with extensions
from the periosteum of a bone to form a tendon. The epimysium is sometimes referred to as
“deep fascia”.
In this chapter you will be required to learn a number of skeletal muscle organs, their
attachments, and their actions. Skeletal muscles are termed “skeletal muscles” because they
typically attach to the skeleton. This attachment will be either direct (by the epimysium fusing to
the periosteum) or indirect (by means of a tendon or aponeurosis). The two attachments are
termed origin and insertion. The origin is the attachment that does not move when the muscle is
contracted. The insertion is the attachment that does move when the muscle contracts. When
learning skeletal muscles you may find it to be more efficient to identify the muscle, follow it to
its origin and insertion, and then think of the movement that it’s contraction will cause. This
should allow you to learn this material faster and to retain it longer. Also. it also helps to match
up these muscles to your own body and to actually exercise the muscle a few times to feel out the
origin, insertion, and action.
It is useful to review how skeletal muscles are named prior to attempting to learn them.
Many of the muscles that we will study follow these rules and, once you understand these rules,
their names should be relatively easy for you to learn. Sometimes more than one of these criteria
will be used to name the muscle:
(1) Location - some names describe the location of the muscle.(ex; the intercostal
muscles are located between the ribs)
(2) Shape - some names describe the shape of the muscle (ex; deltoid, trapezius).
(3) Size - some names describe the size of the muscle relative to other muscles (ex;
major, minor, maximus, minimus, longus, brevis).
(4) Direction - some names describe the direction in which the muscle’s fibers
run. Muscle “fibers” refers to muscle fascicles, not muscle cells. Fascicle patterns are
visible. Different arrangements of fascicles are classified and reflected in muscle
shapes and functions:
(a) Parallel - the long axis of the fascicles run parallel to the long axis of
the muscle (ex; biceps brachii). Although the fibers are running parallel to
the long axis of the muscle, the muscle may or may not be parallel to the midline
of the body. This results in different types of parallel fiber patterns:

64
[1] Rectus (meaning “straight”) - the fibers are running parallel to the body
midline (ex; rectus abdominis)
[2] Oblique - the fibers run obliquely to the body midline (ex; external
abdominal oblique).
[3] Transverse - the fibers are running at a right angle/perpendicular to the
body midline (ex; transversus abdominis).
(b) Pennate - the fascicles are short and attach obliquely to a tendon which runs the
length of the muscle. There are three types:
[1] Unipennate - the fascicles only attach to one side of the tendon (ex;
extensor digitorum longus).
[2] Bipennate - the fascicles attach to two sides of the tendon (ex; soleus ).
[3] Multipennate - the fascicles attach to more than two sides of the tendon (ex;
deltoid).
(c) Convergent - the origin of the muscle is broad and the muscle narrows as it
approaches the insertion (ex; pectoralis major).
(d) Circular/Sphincter - the fascicles are arranged in concentric rings (ex;
orbicularis oculi).
: (5) Location of Origin and/or Insertion – in this case the muscle is named for the
bone or bones that it is attached to. When both the origin and the insertion are present
in the name of a muscle, the origin name usually comes first (ex.; brachioradialis,
sternomastoid).
(6) Action - the muscles are named for the action which they produce (ex; the
supinator supinates the forearm). Many times the region of the body
that the muscle is located in will help you to remember it’s action. For example the
muscles of the dorsal aspect of the limbs are generally extensors and those of the
ventral aspect of the limbs are typically flexors.
(a) Types of Movements
[1] Extensors - increase the angle between two bones
[2] Flexors – decrease the angle between two bones.
[3] Abductors – move a limb away from the midline of the body
[4] Adductors – move a limb towards the midline of the body
[5] Protractors – thrust a part of the body forward (ex; mandible,
or tongue)
[6] Retractors – pull a part of the body back
[7] Levators – raise a structure of the body
[8] Depressors – lower a structure of the body
[9] Rotators – rotate a body part on its axis
[10] Supinators – rotate the palm/sole upwards
[11] Pronators – rotate the palm/sole downwards
[12] Tensors – increase the tension on a structure
[13] Sphincters – decrease the size of an opening.
[14] Dilators – increase the size of an opening.
You should also remember that the muscles were first named in humans. When muscles
were identified in other species that had the same basic origin and insertion they were given the

65
same name as they have in humans. However, humans being bipedal have some unique
myological features. For example: only in hominids is the gluteus maximus the largest gluteal
muscle to power our unusual mode of locomotion. More typically the gluteus medius is larger
than the gluteus maximus. However, due to the rules of muscle nomenclature, the smaller
gluteus maximus is still called the “maximus”.
Skeletal muscle organs can be divided up into axial muscles and appendicular muscles.
Axial muscles are the skeletal muscles of the trunk and tail. They run from the tail to beneath the
pharynx. In all vertebrates axial muscles will display metamerism, a primitive, but retained,
characteristic. The metameric axial muscles combined with flexible metameric vertebral column
allows for locomotion in fishes, some aquatic tetrapods, and limbless terrestrial tetrapods. Axial
muscles are metameric because of their origin from segmental mesodermal somites. This
metameric pattern is especially visible in fishes where the axial muscles are arranged into
segments called myomeres separated by tendinous myosepta (a.k.a.; myocommata).
Appendicular muscles are the muscles of the limbs and their associated girdles. Since most
fishes use axial muscles for locomotion, appendicular muscles in fish are of a limited function.
As a result they are low in number and variety, are uncomplicated, and have little mass. The
appendicular muscles of tetrapods have become increasingly numerous and complex as a result
of the needs of terrestrial life. Correspondingly the role of axial muscles has been reduced in
tetrapods
Skeletal muscle organs can either work together (synergistic) or against one another
(antagonistic) to perform a task. In an antagonistic muscle interaction two (or more) muscles
produce opposite movements on a structure. One muscle will be termed the “agonist” and the
other will be termed the “antagonist”. The agonist or prime mover produces a particular
movement (ex; the biceps brachii causes flexion of the forearm). The antagonist opposes or
reverses the movement of the agonist (ex; the triceps brachii causes extension of the forearm).
When the agonist is acting the antagonist is either at rest or only slightly contracted so as to
control the extent of motion produced by the agonist. The antagonist can also be the agonist for
another motion and in that case it is the previous agonist that acts as the antagonist. For example:
if instead of forearm flexion, we are discussing forearm extension the triceps brachii would be
the agonist and the biceps brachii would be the antagonist.
Most movements will also involve one or more muscles that assist the prime mover.
These muscles are termed synergists. Synergists work either by adding force to a movement or
by reducing undesirable extra movements (ex; the quadriceps group is composed of the vastus
lateralis, vastus medialis, vastus intermedius, and rectus femoris. All four muscles work together
to extend the leg.
In this lab we will be studying two types of skeletal muscle organs: somatic and
branchiomeric muscles. Somatic muscles are those muscle organs that control the “soma”, the
body. All somatic muscle organs are composed of skeletal muscle tissue. Typically they are
innervated by branches of spinal nerves (the exception is a few of the tongue muscles) and are
voluntary (however reflex contractions will occur). Somatic muscles are derivatives from
myotomes of mesodermal somites. As a result somatic muscles are sometimes also referred to as
myotomal or somitic muscles. Branchiomeric muscle organs (a.k.a.; pharyngeal arch muscle
organs) are also composed of skeletal muscle tissue. Branchiomeric somatic muscle organs are
the muscles of the pharyngeal arches and their ontogenetic and phylogenetic derivatives. Like
somatic muscle organs, branchiomeric somatic muscle organs are derived from myotomes.

66
However, they are derived from the anteriormost somites and the unsegmented paraxial
mesoderm in the head. Paraxial mesoderm shows incomplete segmentation. Individual
subdivisions are termed somitomeres. Somitomeres differ from somites in that they are not fully
segmented and lack some of the components found in somites. As a result, these muscle organs
are also referred to as somitomeric muscles. They are innervated by cranial nerves instead of
spinal nerves. Their innervation also differs from that of the typical skeletal muscle tissue in that
there is both a voluntary and an involuntary aspect of control. The voluntary control that you
have over swallowing is due to a branchiomeric muscle in your esophagus. The involuntary
aspect of this muscle includes the “gag” reflex.
Chapter 2: Myology
Part B: Muscle Dissection Techniques
Muscle dissection takes patience and good technique to provide the student with a
specimen that clearly demonstrates the structures to be examined. For the effective study of
muscle organs it is essential that you have a good anatomical specimen.
You will be dissecting both superficial and deep muscles. Superficial muscles are located
close to the integument. They will be exposed by carefully removing the skin and fascia. Fascia
is a general term for the hypodermis and epimysium covering a muscle organ. The hypodermis
is referred to as “superficial fascia” and the epimysium is referred to as “deep fascia”. The
degree of difficulty involved in removing the fascia will be based on a number of factors such as
the age of the animal (the deep fascia is tougher in older animals) and the amount of fat on the
animal. We want to remove most of the epimysium to make it easier to see the “fiber pattern” of
a muscle organ. The fiber pattern is actually the pattern of the fascicles in a muscle organ.
When they are exposed they can often tell a student how the muscle will act. To a lesser extent
the fascicle pattern will also make it easier to see the attachments of the muscle to the skeleton.
The best strategy is to do one side of the animal at a time. That way if an error is made you will
be aware of a potential problem when you perform the same dissection on the other side. The
entire animal should be dissected superficially first, on both sides. Once you have completed the
superficial dissection evaluate your performance. The side with the greatest number of errors
should be your deep side.
In the deep dissection you will be exposing the muscles that lay beneath the superficial
muscles. This is done by “cutting and reflecting” the superficial muscle. If the muscle’s origin
and insertion are intact, you cut the belly of the muscle width-wise and then fold the two cut
halves back to the origin and insertion (called “reflecting”). Because of this approach the impact
of many of the mistakes that you made during the superficial muscle dissection will be reduced.
The technique to expose the muscles in a superficial dissection is called the “pressing
technique”. Utilizing the scalpel and micro-tooth tissue forceps you will press the skin away
from the hypodermis and the fascia away from the muscle. Of the three animals that we are
studying Felis is the most convenient to practice the pressing technique on. Mammals have a
well-defined hypodermis which will give you a margin of error as you become comfortable with
this technique. Squalus has the collagenous component of the dermis anchoring directly on to
the epimysia and the skin is difficult to get through due to its numerous denticles. Necturus has
fairly thin skin that also anchors on to the epimysia of its muscles in many places. Necturus is

67
also a good deal smaller than the other two specimens which will magnify the effects of any
slight mistake.
The first thing that you need to familiarize yourself with is how to hold the scalpel. You
should be working with a metal scalpel handle. Plastic handles do not afford the stability for
precise dissections. The scalpel handle has roughened portion behind the blade. Rest the scalpel
handle against your middle finger at the roughened portion. Your thumb should be placed on the
opposite side, also at the roughened portion, and slightly behind the location of your middle
finger. This way the handle is gripped between your middle finger and thumb. Your index finger
should be placed on the edge of the handle between your middle finger and thumb. The index
finger will act as a guide for the blade while dissecting. This gives you a more precise grip
resulting in a higher degree of control when working with the scalpel. The blade will be
controlled by your fingertips. The movement of your scalpel hand is more like the painting of a
portrait than the slicing of meat.
When removing the integument you will first be removing the dermis from the
hypodermis. The blade should be positioned so that the cutting edge is at a 45o angle from the
hypodermis. The edge will be pointed towards the dermis in the direction of movement of the
scalpel. This will seem odd to you at first. You would expect to have the blade facing the
hypodermis. However, if you were to hold the blade that way and made a mistake, you would
slice into the hypodermis which we are trying to preserve for practice purposes. If you are
holding the scalpel correctly and make a mistake you are more likely to slice into the skin which
you are removing and are not concerned about. What allows this technique to work is tension.
While you are moving the scalpel you will be pulling on the skin with the micro-tooth tissue
forceps. This will stretch and strain collagen bundles that anchor the skin to the hypodermis
making it easier for the blade to slice through them and remove the skin. You will be able to see
these collagen bundles with the unaided eye as you work to remove layers from above the
muscles. The key is to maintain tension by moving the forceps hand along with the scalpel hand.
(It is easy to forget to keep the forceps hand moving and, once the tension is reduced, you will
most likely slice into the abovelying skin.) You will actually feel more fatigue with your forceps
hand than with your scalpel hand by the end of the lab period. You should work the scalpel along
a lane. Once you come to the end of the lane it is time to start a new one. However, you do not
want to pick up the blade from the animal and move it a new location; that increases the chances
of making a mistake. Instead keep the scalpel where it is and rotate it 90o. Now the blade is
facing in the opposite direction and you can make a second lane, overlapping with the first.
Repeat this again and again until you have removed the skin.
When working on the fascia you should remove one layer at a time using the pressing
technique. This is tedious but necessary to avoid damaging the muscles that you wish to study.
Remove the hypodermis first but be careful. The integument of Felis contains cutaneous skeletal
muscle fascicles that make up the muscle of the integument, the cutaneous maximus. The
cutaneous maximus looks a bit like the muscles that you are trying to save and will fuse on to
some of those muscles in certain locations. You should first remove the portion of the
hypodermis superficial to these muscle fibers so that you can observe them and safely remove
them from your specimen. Another difficulty is that there are a number of superficial blood
vessels and structures that you will wish to preserve (ex; radial artery, vein, and nerve and the
brachioradialis of the forearm). These will often be partially concealed in adipose of the
hypodermis. The fatter the animal the more likely it is that you will lose structures by

68
injudiciously removing the adipose. For working in fatty areas it is better to use another
technique, the “separating technique”.
In the separating technique you utilize your fine-point dissection scissors instead on your
scalpel. Place the closed scissors into the fat and gently open the blades. This will cause the fat
to fragment without damaging other structures. Close the blades before you move the scissors to
another location and repeat. The fat will break up and expose the structures that you are trying to
save. As with the scalpel you want to hold the fine-point dissection scissors in a grip that will
allow you to be as precise as possible. There are two grips that work well, although the first one
is a little better than is the second. The first grip is to place your thumb and ring finger into the
rings of the scissors handle. Rest the weight of the scissors on the middle finger and place the tip
of your index finger against the screw holding the two blades together. This will decrease the
shaking of the blades and allow for more precision as you work the scissors. The degree of
shaking may seem minimal but you will be trying to observe some small structures that can be
easily damaged. The second grip is to place your thumb and middle finger into the rings of the
scissors handle. The index finger can either be placed against the screw holding the blades
together or you can rest the scissors on the index finger. This grip is not quite as precise as the
first one but is more stable than the grip that you are used to using.
Once you have removed the hypodermis you will move on to the deep fascia, the
epimysium. This part of the dissection will require patience. It can take a bit of time to remove
this layer but it is worth it. Again you will be using the pressing technique and, since you are
very close to the muscle that you wish to preserve, you will need to be very careful with your
technique.
Demonstration of the Proper Grip for Holding the Fine-point Dissection Scissors

69
Demonstration of the Proper Grip for Holding the Scalpel
Making the Initial Incision
Note: The scalpel is held at a 45o angle to the underlying muscle, pointed in the direction of the
cut and facing away from the muscle that you wish to save.

70
Expanding the Incision
The tension on the collagen fibers from the fine-tooth tissue forceps makes it easier to slice through them.
As you “hit” the edges of the incision you will expand the incision.
Reversing Direction
Once you have completed one “lane” using the pressing technique you reverse direction to start a new
lane. Rotate the blade 90o. Now you are ready to work in the opposite direction.

71
The Pressing Technique
Notice how the collagen bundles are straining due to the forceps allowing the scalpel to more
easily cleave through them.

72
Chapter 2: Myology
Part C: The Muscles
Topic C1: The Muscles of Squalus
In fishes axial muscles are the predominant muscles of locomotion. The muscles of the
fins play a minor role in locomotion, typically steering or stabilizing the thrust produced by the
axial muscles. As a result the appendicular muscles of fishes in general, and of Squalus in
particular, will be of limited size, diversity, and function.
The first step in the muscle dissection of Squalus is to remove the skin using the pressing
technique. The collagen of the dermis anchors on to the collagen of the epimysium in Squalus.
As a result you will be able to see the muscles as soon as you remove the skin. Many times the
muscle will begin to fray as you are pressing away the skin. You should increase tension with
the forceps hand to decrease the amount of fraying. Some areas will be very difficult to remove
the skin from without damaging underlying structures. The skin around the gills is a good
example of that. You will need to remove the skin from the base of the chondrocranium to the
base of the first dorsal fin dorsally and from the Meckel’s cartilage to the point opposite the first
dorsal fin ventrally. You will also need to remove the skin from the base of the pectoral fins,
both dorsally and ventrally. There will typically be one layer of fascia left to remove after the
animal has been skinned. This fascia will be tightly attached to the muscle and will require
patience to remove.
The muscles of the body wall and tail in fishes such as Squalus consist of myomeres
separated by myosepta into which the longitudinally arranged muscle cells attach. These muscles
are well developed being the muscles for locomotion. The myomeres have a “zigzag” pattern
immediately beneath the skin. The segmental arrangement of the axial muscles is easy to
observe in Squalus. There is one myomere per vertebra and one spinal nerve per myomere. As is
true for all fishes other than agnathans, the myomeres of Squalus are divided into dorsal and
ventral masses. The dorsal and ventral muscle masses are the epaxial and hypaxial muscles
respectively. A sheet of tendinous material called the horizontal skeletogenous septum separates
the epaxial and hypaxial muscles. The horizontal skeletogenous septum stretches between the
vertebral column and the skin along the entire trunk and tail. It will also anchor on to the dorsal
ribs of Squalus. The muscles of the pectoral and pelvic girdles interrupt the metameric
arrangement of myomeres. These girdles interrupt only the hypaxial muscle segmentation. The
epaxial muscles are dorsal to the girdles and as a result will not be affected. The epaxial muscles
will extend cranially on to the skull. Those epaxial muscles located dorsal to the gills are termed
epibranchial muscles. Those hypaxial muscles located ventral to the gills are termed
hypobranchial muscles. The innervation of the epaxial and hypaxial muscles is also metameric.
Epaxial muscles are innervated by the dorsal rami of spinal nerves. Hypaxial muscles are
innervated by the ventral rami of spinal nerves. This pattern of innervation is seen tetrapods as
well.

73
Epibranchial Muscles Horizontal Septum Myomere Myoseptum Epaxial Muscles
Branchiomeric Muscles Hypaxial Muscles
(disrupted segmental pattern due to pectoral girdle)
Squalus Muscles Showing the Relationship of Epaxial and Hypaxial Muscles
The hypobranchial muscles can be seen on the ventral surface of Squalus. They are
located between the pectoral girdle and Meckel’s cartilage. The hypobranchial muscles will be
closely associated with the branchiomeric muscles. To observe them you will need to cut and
reflect the intermandibularis. This can be done removing the right or left muscle at the
midventral raphe and then folding it to the Meckel’s cartilage .The hypobranchial muscles are:
common coracoarcurals (2), coracomandibular (1), coracohyoids (2), and the coracobranchials
(10).

74
Squalus Muscles, Ventral View:
Deep Hypaxial, Hypobranchial, Branchiomeric, and Ventral Appendicular Muscles
(*Note the difference in the fiber pattern between the interhyoid and intermandibular)
The Branchiomeric Muscles
Muscle Origin Insertion Action
First Ventral Constrictor (Intermandibular) Mid-ventral Raphe Meckel's cartilage Raise the floor of the mouth
Second Ventral Constrictor (Interhyoid) Hyomandibula Ceratohyal Compresses oropharynx
Third through Sixth Ventral Constrictors Vertical Raphe Vertical Raphe Compresses oropharynx
Coracobranchials Coracoid Bar Ventral Gill Cartilages Expands oropharynx
First Dorsal Constrictor - Spiracular Otic Capsule Palatopterygoquadrate Aids to elevate palatoquadrate
- Mandibular Adductor Palatopterygoquadrate Mandibular Arch Closes mouth
Second Dorsal Constrictor Otic Capsule Hyomandibula Compresses oropharynx
Third through Sixth Dorsal Constrictors Vertical Raphe Vertical Raphe Compresses oropharynx
First Levator (Palatopterygoquadrate
Levator) Otic Capsule Palatopterygoquadrate Elevates palatopterygoquadrate
Second Levator (Hyoid Levator) Epihyoideus Epihyoideus Elevates hyoid
Third through Sixth Levators (Cucullaris) Fascia of Epaxial Muscle Scapula Elevates scapula
Coracomandibular Coracohyoid Common Coracoarcural
Intermandibular
Interhyoid*
Coracomandibular Coracohyoid Common Coracoarcural
Intermandibular
Interhyoid*

75
The branchiomeric muscles derive from splanchnic mesoderm (unlike the hypobranchial
muscles that derived from somatic mesoderm). They control the movements of the
splanchnocranium, the jaws and gill arches. They surround the oropharynx and can be observed
ventrally, laterally, and dorsally. The branchiomeric muscles are the dorsal constrictors, the
ventral constrictors, and the levators (all numbered first through sixth). The first dorsal
constrictor is actually two muscles: the spiracular and the mandibular adductor. The mandibular
adductor increased in size and points of attachment during tetrapod evolution and became three
masticatory muscles of mammals: the masseter, temporalis, and pterygoideus. The first ventral
constrictor is also known as the intermandibularis. The intermandibularis gave rise to the
mammalian mylohyoid and possibly the digastric muscles. Immediately deep to it is the second
ventral constrictor, the interhyoideus. It can be observed by cutting and reflecting the
intermandibularis. The interhyoideus can be distinguished from the intermandibularis by its
fiber pattern. The intermandibularis has an oblique pattern while the interhyoid has a transverse
fiber pattern. The first and second levators elevate the palatopterygoquadrate to open the mouth.
The third through sixth levators are also known as the cucullaris. The cucullaris is homologous
to the trapezius of amniotes.
Squalus Muscles, Ventral View:
Superficial Hypaxial, Hypobranchial, Branchiomeric, and Ventral Appendicular Muscles
The Hypobranchial Muscles
Muscle Origin Insertion Action
Common Coracoarcural Coracoid Bar Mandibular Adductor Aids to open mouth
Coracomandibular Coracoid Bar Meckel's Cartilage Opens mouth
Coracohyoid Coracoid Bar Basihyal of Hyoid Braces jaws for feeding
Coracobranchials Coracoid Bar Ventral Gill Cartilages Expands oropharynx

76
The appendicular muscles are reduced to levators (extensors) and depressors (flexors) of
the fins. The two observed in this lab are the pectoral levator and the pectoral depressor. During
vertebrate development the long plane of the limb is defined by its skeletal components. This
establishes a dorsal plane and a ventral plane. The mesoderm of the dorsal plane differentiates
into extensor muscles and the mesoderm of the ventral plane differentiates into flexor muscles.
You can observe this pattern in the fins of Squalus and limbs of Necturus and Felis.
Squalus Muscles Dorsal View: Superficial Branchiomeric Muscles
The Appendicular Muscles
Muscle Origin Insertion Action
Pectoral Levator Scapula Pterygiophores Elevate/extend pectoral fin
Pectoral Depressor Coracoid Bar Pterygiophores Depress/flex pectoral fin

77
Topic C2: The Muscles of Necturus
In Necturus we have a nice example of the transition between aquatic and terrestrial
existence and its correlation in the musculature. Necturus has retained the primitive metameric
pattern of epaxial and hypaxial muscles seen in fishes. This allows these aquatic urodeles to
swim like fish, utilizing the axial musculature and lateral undulations of the vertebral column.
Amniotes gradually lost much of the metamerism of the axial muscles. The disappearance of the
epaxial myotomes led to the development of long, strap-like or pennate muscle bundles in
amniotes. These changes in muscles correlated with changes that increased the flexibility of the
vertebral column so as to improve the movement of the trunk for life on land. (Think about how
a whale swims versus a shark.) Changes also occurred in the lateral body wall. Broad muscular
sheets arranged in layers gradually replaced the hypaxial muscles. The muscle fascicles of each
sheet are arranged in a different direction than those of neighboring muscle sheets. This fiber
pattern is possible since these muscles are no longer used for locomotion. That role has been
taken over by the appendicular muscles. Instead the muscles of the lateral wall serve to support
the viscera in a “muscular sling”. Intermediary stages in this transition can be observed in extant
species such as Necturus which shows epaxial and hypaxial myomeres. However, the hypaxial
myomeres are arranged into three sheets: external abdominal oblique, internal abdominal
oblique, and transversus abdominis. We see the same three muscle sheets in amniotes such as
Felis.
The appendicular muscles of Necturus are much better developed than are those of fishes
but not as well developed as are those of amniotes. One example of this is the latissimus dorsi.
In urodeles, such as Necturus, it is a thin, triangular muscle originating from the fascia of epaxial
muscle near the shoulder. In amniotes, such as Felis, it is a broad sheet, covering a much larger
area, originating from the transverse processes of cervical, thoracic, and lumbar vertebrae. In
Necturus the origin of the latissimus is so weak that you may remove the muscle when skinning
your specimen. The pattern of tetrapod muscle evolution is an increase in the size, number, and
strength of attachments over time. This can be observed by comparing Felis to Necturus.
The skin of Necturus is thin and care must be taken with its removal. As previously
mentioned, without attention it is easy to remove some muscles including all of the dorsal
pectoral musculature and branchial levators. There is very little hypodermis in amphibians. In
many locations the dermis will be immediately superficial to the muscle. It is best to begin at the
tail. Using the pressing technique you can start by removing the skin from the tail and working
your way cranially. Observe the numerous large glands on the underside of the skin in the tail.
With good technique the skin can be safely removed. When you get to the ventral head you
should change direction, however. You should resume pressing the skin away at the dentary and
work your way caudally. This is to avoid the gular fold, a fold of skin at the posterior aspect of
the lower jaw. It will have some fibers of the interhyoid attached to it. If you were to attempt to
remove it going caudal to cranial you could easily remove the interhyoid and intermandibular
muscles along with the skin. You do not need to change direction while skinning the dorsal
head.
The axial muscles of Necturus display a metameric pattern and demonstrate myomeres
and myosepta. They can be divided into hypaxial and epaxial muscles by the horizontal
skeletogenous septum. The epaxial muscles of the trunk and dorsal gill region are called the
dorsalis trunci. The hypaxial muscles of the trunk are arranged into three broad sheets: external
abdominal oblique, internal abdominal oblique, and transversus abdominis; running superficial to

78
deep. The external abdominal oblique has its fibers running at a 45o angle to the midline of the
body. The internal abdominal oblique also has its fibers running at a 45o angle to the midline of
the body but these fibers are perpendicular to those of the external abdominal oblique. The
fibers of the transversus abdominis are perpendicular to the midline of the body as its name
indicates. The right and left muscle sheets do not meet medially. Between them is the rectus
abdominis. These paired muscles meet at the linea alba of the midline and extend from the pubis
to the pectoral region. The rectus abdominis will show regular segmentation with its myomeres
being separated by myosepta, a pattern that we see even in ourselves. The portion of the
hypaxial muscles in the ventral gill region is the rectus cervicis.
The muscles of the head and gill region include hypobranchial, epibranchial, and
branchiomeric muscles. The epibranchial muscles are the cervical portion of the dorsalis trunci.
The hypobranchial muscles include the rectus cervicis and geniohyoid. The rectus cervicis will
have a segmental pattern. However, that metameric pattern is disrupted by cranial elements of
the pectoral girdle. Posterior slips of the rectus cervicis will attach to the procoracoids to give
rise to the omoarcual muscle. With the development of the neck the hypobranchial muscles
became more strap-like as can be seen with the geniohyoid and branchiohyoid. The
branchiomeric muscles have begun to take on other roles although the branchial levators do serve
to move the gills. The mandibular adductor of Squalus has expanded to become the anterior
mandibular adductor, external mandibular adductor, and mandibular depressor of Necturus.
These muscles are homologous to the temporalis, masseter, and (possibly) digastric of mammals
Necturus, Some Axial MusclesThe Axial Muscles of the Trunk
Muscle Origin Insertion Action
Dorsalis Trunci Myosepta & Vertebrae Myosepta & Vertebra Laterally flexes trunk
External Abdominal
Oblique
Myosepta & Horizontal
Septum
Myosepta & Horizontal
Septum Laterally flexes trunk
Internal Abdominal
Oblique
Myosepta & Horizontal
Septum
Myosepta & Horizontal
Septum Laterally flexes trunk
Transversus Abdominis Horizontal Septum Linea Alba Laterally flexes trunk
Rectus Abdominis Puboischiadic Plate Coracoid Ventrally flexes trunk

79
Necturus Dorsal Muscles of the Head and Trunk
The Muscles of the Head and Pharyngeal Region
Muscle Origin Insertion Action
Anterior Mandibular
Levator Parietal & Frontal Dentary Raise lower jaw
External Mandibular
Levator Parietal & Frontal Dentary Raise lower jaw
Mandibular Depressor Parietal & Squamosal Angular Depress lower jaw
Dorsalis Trunci Myosepta & Vertebrae Parietal & Opisthotic Laterally turns head
Branchial Levators Fascia of Epibranchials Epihyals Raise gills
Intermandibular Dentary & Angular Median raphe Tenses floor of oropharynx
Interhyoid Fascia of Branchiohyoid Median raphe Tenses floor of oropharynx
Geniohyoid Dentary Second basibranchial Moves hyoid cranially
Branchiohyoid First gill arch Ceratohyoid Moves hyoid cranially
Rectus Cervicis Coracoid Ceratohyoid Moves hyoid cranially

80
Necturus Ventral Muscles of the Head and Pharyngeal Region
respectively. As in Squalus the intermandibularis and interhyoid are superficial muscles of the
ventral head. They are distinguished by their fiber patterns and can be separated by lightly using
the separation technique with the tips of your scissors. (To observe the geniohyoid and
branchiohyoid you must cut and reflect the intermandibular and interhyoid.) The
intermandibularis is homologous to the mylohyoid and possibly the anterior portion of the
digastric in mammals.
The appendicular muscles of tetrapods are much more complex and diverse than are
those of fishes. They can be divided up into the muscles of the pectoral girdle and anterior limb
and the muscles of the pelvic girdle and posterior limb. The appendicular muscles are typically
innervated by the ventral rami of the spinal nerves and are under somatic motor control. During
development somatic mesoderm will grow into the limb bud and differentiate into a dorsal mass
and a ventral mass. . The dorsal mass will develop into extensor muscles primarily while the
ventral mass will develop into flexor muscles primarily. This pattern can be seen in fishes and
tetrapods. In some amniotes (such as humans) this pattern is harder to see since the limb has
become rotated but it does still exist. These muscles are easily observed after skinning Necturus.
Only judicious application of the separating technique with the tips of your scissors will be
needed to better define the borders of most muscles.
The dorsal muscles of the pectoral girdle tend to be thin muscles that originate from
fascia of neighboring epaxial muscles. It is this loose attachment that makes them easy to
remove or damage during skinning. The latissimus dorsi and cucullaris (which is homologous to
the trapezius) are much smaller, thinner, and more weakly attached than is seen in amniotes. The
cucullaris is of a different embryonic origin than are other muscles being a branchiomeric muscle

81
instead of a somatic muscle. The dorsal muscles of the pectoral girdle include the latissimus
dorsi, cucullaris, pectoriscapularis, and the dorsalis scapulae. The dorsalis scapulae is most
likely homologous to the deltoid. The muscles of the ventral pectoral girdle are the pectoralis,
supracoracoideus, and procoracohumeralis. The pectoralis and supracoracoideus have close, but
slightly different insertions, allowing for different ranges in the movements produced by them.
The arm of Necturus has three muscles. One is the triceps brachii, which has three points of
origin (or heads) and serves as an antagonist to the other two muscles. The other two are the
humeroantebrachialis and the coracobrachialis. These muscles are synergists to one another,
producing flexion of the forearm. They can be distinguished from each other by the fact that the
coracobrachialis is the more medial of the two muscles. The humeroantebrachialis, although it
has only one point of origination, is homologous to the biceps brachii.
Procoracohumeralis Cucullaris Humeroantebrachialis
Omoarcual Triceps Brachii External Abdominal Oblique
Pectoriscapularis Dorsalis Scapulae Latissimus Dorsi
Necturus Myology of the Pectoral Girdle and Forelimb, Dorsal View

82
Geniohyoid Branchiohyoid Rectus Cervicis Procoracohumeralis Humeroantebrachialis
Omoarcual Coracohumeralis
Intermandibularis Interhyoid Supracoracoideus Pectoralis
Necturus Myology: Branchial Muscles, Forelimb, and Pectoral Girdle
Muscles of the Forelimb and Pectoral Girdle
Muscle Origin Insertion Action
Cucullaris Fascia on dorsalis trunci, cervical portion Scapula Move scapula craniodorsally
Pectoriscapularis Fascia on dorsalis trunci, cervical portion Scapula Move scapula cranially
Dorsalis scapulae Suprascapular cartilage Humerus Abduct humerus
Latissimus dorsi Fascia of dorsalis trunci, thoracic portion Humerus Move humerus caudally
Procoracohumeralis Procoracoid cartilage Humerus Move humerus cranially
Supracoracoideus Linea alba on coracoid Humerus Adduct humerus
Pectoralis Linea alba on coracoid Humerus Adduct humerus
Triceps Brachii Coracoid, Scapula, & Humerus Ulna Extends forearm
Humeroantebrachialis Humerus Radius Flexes forearm
Coracobrachialis Coracoid Humerus Flexes forearm
While the components of the pectoral girdle are loosely attached to one another and are
capable of independent movement, such is not the case with the pelvic girdle. The elements of
the pelvic girdle are tightly joined together and are also joined to the vertebral column
(especially in amniotes). So unlike pectoral girdle muscles which can originate on the axial

83
skeleton and inserting on the girdle, pelvic girdle muscles originate on the pelvis and insert into
the limb. The locomotory muscles of the posterior limb extend from bones/cartilages of the
pelvic girdle (ilium, ischium, and pubis) to the femur and/or the fibula/tibia, from the femur to
the tibia/fibula, and from the tibia/fibula to the bones of the pes. In modern amphibians and
reptiles the muscles are typically named for their origin and insertion. Such is the case with most
of the pelvic and hind limb muscles of Necturus. These muscles tend to have broad insertions.
This causes them to lack the more precise movements that we see in amniotes. For example, the
ischioflexorius inserts over a broad area of the tibia and the plantar fascia of the pes. There are
five pairs of ventrally oriented muscles: puboischiofemoralis externus, puboischiofemoralis
internus, pubotibialis, puboischiotibialis, and ischioflexorius. Due to the shape of the
puboischiadic plate the origin of the puboischiofemoralis externus comes to a notable point.
There are three pairs of dorsally oriented muscles: iliotibialis, ilioextensorius, and iliofibularis.
Puboischiofemoralis Externus Puboischiotibialis
Pubotibialis Puboischiofemoralis Internus
Necturus Muscles of the Pelvic Girdle and Hind Limb, Ventral View

84
External Abdominal Oblique Iliotibialis Ilioextensorius Iliofibularis
Necturus Muscles of the Pelvic Girdle and Hind Limb, Dorsal View
Muscles of the Pelvis and Hind Limb
Muscle Origin Insertion Action
Puboischiofemoralis externus Puboischiadic plate Femur Adducts femur
Puboischiofemoralis internus Puboischiadic plate Femur Moves femur anteriorly
Pubotibialis Pubis Tibia Adducts femur & extends leg
Puboischiotibialis Puboischiadic plate Tibia Adducts femur & flexes leg
Ischioflexorius Ischium Tibia & Pes Flexes leg & foot
Iliotibialis Ilium Tibia Extends leg
Ilioextensorius Ilium Tibia Extends leg
Iliofibularis Ilium Fibula Extends leg
Topic C3: The Muscles of Felis
In amniotes such as Felis the appendicular muscles display little of the metameric pattern
that we saw with Squalus and Necturus. The epaxial muscles of amniotes are strap-like and run
the length of the vertebral column, extending from the base of the skull into the tail to varying

85
extents. These muscles function to extend the vertebral column and to laterally flex the torso and
are collectively known as the erector spinae muscles. Epaxial muscles are divided into four
groups: intervertebrals, longissimus, spinales, and iliocostales. The intervertebral muscles are the
deepest epaxials and are the only ones to retain the metameric condition. They help to maintain
posture. The multifidus spinae will be the only one visible in this dissection. The longissimus is
the longest epaxial muscle. It is located dorsal and lateral to the transverse processes of the
vertebrae. The longissimus is the dominant extensor of the mammalian vertebral column
although it does not usually play a role in mammalian locomotion. This muscle typically has
three bundles that insert in varying locations. The bundles are named for these points of
insertion: longissimus capitis inserts on the head, longissimus cervicis inserts on the neck, and
longissimus dorsi inserts on the trunk. In the lumbar region the longissimus dorsi has three
distinct bundles with the medial bundle extending into the tail to form the extensor caudae
lateralis. You can observe these three bundles by cutting and reflecting a portion of the multi-
layered lumbodorsal aponeurosis. The spinales are also located dorsal to the transverse processes
and medial to the longissimus. There are a number of spinales muscles including spinalis capitis,
spinalis cervicis, and spinalis dorsi. The iliocostales are a thin sheet of muscles originating on the
ilium and inserting on the ribs. The spinalis dorsi, longissimus dorsi, and iliocostalis can be
observed by cutting and reflecting the spinotrapezius and latissimus dorsi. This will expose a
portion of aponeurosis in the thoracic region that can be carefully removed to show the spinalis
dorsi and longissimus dorsi. A third muscle will be visible ventral to the longissimus dorsi, the
serratus dorsalis. The serratus dorsalis displays a segmental pattern and serves to help stabilize
the vertebral column. It has two portions: serratus dorsalis cranialis and serratus dorsalis
caudalis. They can be differentiated by the fiber pattern. Gently peel the serratus dorsalis from
its attachment to the longissimus to observe the iliocostalis. You should be able to observe
tendons of the iliocostalis attaching to the ribs. The epaxial muscle of the dorsal neck is the
splenius and can be observed by cutting and reflecting the clavotrapezius.
Felis Erector Spinae Muscles

86
Felis Erector Spinae MusclesErector Spinae Muscles of Felis
Muscle Origin Insertion Action
Serratus Dorsalis, Cranialis R1-R9 Mid-dorsal Raphe Pulls ribs cranially
Caudalis R10-R13 Lumbar Vertebrae Pulls ribs caudally
Longissimus dorsi Sacral and Caudal Vertebrae Thoracic & Cervical Vertebrae, Occipital Extends vertebral column
Spinalis Dorsi T10-T13 Thoracic & Cervical Vertebrae Extends vertebral column
Iliocostalis Ilium Ribs Laterally flexes spine
Splenius Mid-dorsal fascia line Occipital Raises head dorsally
Multifidus Dorsi Vertebral transverse processes Spinous process of vertebra cranial to origin Extends vertebral column
Hypaxial muscles can be broken up into four groups: subvertebrals, oblique sheets,
transverse sheets, and the rectus abdominis. Subvertebral muscles are longitudinally arranged
muscles located deep to the transverse processes in the roof of the coelom from atlas to pelvis.
They are fairly powerful flexors of the spine and serve as antagonists to the epaxial muscles. Due
to their location they will not be a part of this dissection. Oblique and transverse muscles are
located in the lateral body wall and as a result are sometimes referred to as “parietals”. Felis will
have the same oblique and transverse muscle sheets that we saw in Necturus: external abdominal
oblique, internal abdominal oblique, and transversus abdominis (running superficial to deep).
These are very thin muscles and you must be careful when exposing them. You will be able to
differentiate these three muscles by their fiber patterns. Mammals also have a diaphragm that is
the main muscle for respiration. The diaphragm is an oblique muscle. The cremaster muscle is an
outgrowth of the internal abdominal oblique muscle and is found in male mammals. It is a
portion of the spermatic cord and serves to retract the testes. The rectus muscles are limited to
the anterior abdominal wall. Along with the oblique and transverse muscles, the rectus muscles
form a muscular sling to support the viscera. There are two rectus muscles. One is the rectus

87
abdominis which is found in all tetrapods. It extends the length of the abdomen, on either side of
the linea alba, from pubis to sternum and assists in flexing the trunk. The second is the
pyramidalis. It is an outgrowth of the rectus abdominis and is found only in mammals. It serves
to support the pouch in marsupials and is vestigial in placental mammals.
Felis Abdominal Muscles
Felis Abdominal Muscles
Muscle Origin Insertion Action
External Abdominal Oblique Lumbodorsal Aponeurosis & Ribs Linea Alba and Pubis Twists trunk
Internal Abdominal Oblique Lumbodorsal Aponeurosis Linea Alba Twists trunk
Transversus Abdominis R10-R13 Linea Alba Twists trunk
Rectus Abdominis Sternum & Ribs/Pubis Pubis/ Sternum & Ribs Flexes trunk
Branchiomeric muscles are found in all amniotes including Felis. In amniotes the
branchiomeric muscles still operate the jaws but the muscles that were once of the gill arches
now serve other purposes. In all gnathostomes the muscles of the first arch, the mandibular arch,
are chiefly jaw muscles. In tetrapods the adductor mandibulae has expanded and developed into
up into three muscles: masseter, temporalis, and pterygoideus. These three muscles have spread
over the skull and provide the bulk off the force for jaw movements. The intermandibularis of
Squalus and Necturus has become the mylohyoid in tetrapods. A portion of the intermandibularis

88
may have also given rise to the anterior belly of the digastric muscle. In tetrapods hyoid arch
muscles continue to serve some of the same functions in tetrapods as they did in fishes but they
will also have some new roles. In urodeles and many reptiles the muscles of the second arch give
rise to the depressor mandibulae which serves to open the mouth. In mammals the depressor
mandibulae becomes the posterior belly of the digastric muscle. In amphibians and lower reptiles
a thin sheet of muscle called the sphincter colli is superficial to the branchiohyoideus. In higher
reptiles the sphincter colli extended upwards on to the skull and became the platysma. The
platysma increased in size in mammals and gave rise to the muscles of facial expression. These
are really modified cutaneous muscles and care must be taken when removing the skin so as to
avoid damaging these muscles. Mammals also have a stylohyoid muscle which serve to attach
the hyoid to the styloid process of the skull. The branchiomeric muscles of arches 3 through 7
have been pretty much lost in tetrapods. The remaining muscles from the third arch are:
stylopharyngealis (for swallowing) and stylohyoid (posterior belly) in some mammals. In
mammals the muscles remaining from the fourth arch are the intrinsic muscles of the mammalian
larynx: cricothyroid, cricoarytenoid, and thyroarytenoid. The trapezius, a branchiomeric muscle
of the amniote pectoral girdles, is actually a derivative of the cucullaris.
Felis Superficial Neck and Branchiomeric Muscles

89
Sternothyroid Thyrohyoid Hyoglossus Genioglossus Geniohyoid (Digastric & Mylohyoid cut)
Sternomastoid Clavotrapezius Stylohyoid Mylohyoid Digastric
Sternohyoid Masseter
Felis Tongue and Neck Muscles
In the head and neck of Felis we will also find hypobranchial and tongue muscles.
Hypobranchial and tongue muscles are anterior extensions of the hypaxial muscles. The
hypobranchial muscles of amniotes include: sternomastoid, cleidomastoid, sternohyoid,
sternothyroid, thyrohyoid, and geniohyoid. The sternothyroid and thyrohyoid may be observed
by cutting and reflecting the sternohyoid. The muscles of the tongue develop from mesenchyme
that has migrated from anterior hypobranchial muscle blastemas. Tongue muscles can be either
extrinsic or intrinsic. Extrinsic tongue muscles originate outside of the tongue (on the hyoid or
chin for example) and insert in the tongue. They include the genioglossus, hyoglossus, and
styloglossus. These muscles may be observed but cutting and reflecting the mylohyoid. Intrinsic
tongue muscles originate and insert in the tongue. They are restricted to mammals, birds, and
some retiles. Since hypobranchial and tongue muscles originate from the anteriormost somites
of the trunk they are innervated by cervical spinal nerves in general, although CN XII will play a
role in some cases.

90
Felis Tongue and Neck Muscles
Felis Tongue and Neck Muscles
Muscle Origin Insertion Action
Sternomastoid Manubrium Temporal Laterally flex neck
Sternohyoid Manubrium Hyoid Pulls hyoid posteriorly
Digastric Basioccipital Mandible Depresses mandible
Mylohyoid Mandible Median Raphe Raises floor of mouth
Stylohyoid Stylohyal Hyoid Elevates hyoid
Masseter Zygomatic Arch Mandible Elevates mandible
Temporalis Temporal Fossa Mandible Elevates mandible
Sternothyroid Manubrium Thyroid cartilage Pulls larynx caudally
Thyrohyoid Thyroid cartilage Hyoid Raises larynx
Cricothyroid Cricoid cartilage Thyroid cartilage Puts tension of the vocal cords
Cleidomastoid Clavicle Temporal Laterally flex neck
Geniohyoid Mandible Hyoid Pulls mandible cranially
Hyoglossus Hyoid Tongue Retracts tongue
Styloglossus Stylohyal Tongue Retracts tongue
Genioglossus Mandible Tongue Protracts base of tongue

91
As has been mentioned previously, the appendicular muscles of tetrapods are more
numerous and complex than are those of fishes. In Felis the appendicular muscles are larger and
more diverse than those of Necturus. One example is the latissimus dorsi. It is a primary
appendicular muscle that inserts on the humerus. It has increased in size, strength, and the size of
its attachments over evolutionary time. In Necturus the latissimus dorsi is a thin triangular
muscle arising from fascia superficial to the epaxial myomeres of the shoulder region. In
mammals the origin covers a much greater area of the dorsal body and is directly on the spinous
processes of the vertebrae (most thoracic, all lumbar, some sacral, and even some caudal
vertebrae). The size of the latissimus dorsi and area of insertion on the humerus has also
increased.
Deep to the latissimus dorsi in Felis are three extrinsic muscle groups that insert on the scapula.
These three groups are the two levators of the scapula, the rhomboideus muscles, and the serratus
ventralis. The levators of the scapula in Felis are the levator scapulae ventralis and the levator
scapulae dorsalis. The rhomboideus group in Felis is composed of three pairs of muscles that
move the scapula and differ in their point of origin. Originating on the occipital bone is the
rhomboideus capitis (a.k.a.; occipitoscapularis). Originating on the cervical vertebrae is the
rhomboideus minors. The broadest origin is that of the rhomboideus major which originates on
some of the cervical and thoracic vertebrae. The serratus ventralis originates from some of the
ribs and inserts on the scapula. It can be seen in the axillary region immediately caudal to the
levator scapulae dorsalis (a.k.a.; levator scapulae) and ventral to the serratus dorsalis. The
trapezius of tetrapods is derived from the cucullaris of fish and is of branchiomeric origin. The
trapezius, like the latissimus dorsi, has increased in size over evolutionary time. In mammals it
has three portions: clavotrapezius, acromiotrapezius, and spinotrapezius. These three muscles
differ in their point of insertion to the pectoral girdle. Due to its evolutionary origin the trapezius
receives innervation from branchiomeric nerves, which is unique in appendicular muscles.
Felis Superficial Back and Dorsal Arm Muscles

92
Felis Deep Upper Back Muscles
Felis Upper Back Muscles
Muscle Origin Insertion Action
Clavotrapezius Lambdoidal Crest Clavicle Dorsally raise scapula
Acromiotrapezius Cervical Vertebrae Scapula Dorsally raise scapula
Spinotrapezius Thoracic Vertebrae Scapular muscle fascia Dorsally raise scapula
Clavobrachialis Clavicle Ulna Flexes forearm
Acromiodeltoid Scapula Humerus Flex and rotate humerus
Spinodeltoid Scapula Humerus Flex and rotate humerus
Levator Scapulae Ventralis C1 and Occipital Scapula Raises scapula cranially
Latissimus Dorsi Thoracic & Lumbar Vertebrae Humerus Pulls arms caudodorsally
Rhomboideus Major Cervical &Thoracic Vertebrae Scapula Pulls scapula dorsally
Rhomboideus Minor Cervical Vertebrae Scapula Pulls scapula dorsally
Rhomboideus Capitis Occipital Scapula Pulls scapula cranially
Supraspinatus Scapula Humerus Extends humerus
Infraspinatus Scapula Humerus Laterally rotates humerus
Teres Major Scapula Humerus Medially rotates humerus
Teres Minor Scapula Humerus Laterally rotates humerus
The ventral muscles of the pectoral girdle are collectively referred to as the “pectorals”.
These muscles have undergone expansive changes over evolutionary time due to an increased
need for strength. Primitively it originated on the coracoid cartilages (or bones) and a midventral

93
raphe on the neck as we saw in Necturus. In mammals it originates on the bones of the sternum
and some ribs. Over time the pectorals developed from one thin muscle into a varying number of
superficial and deep muscles. In Felis the pectoralis has developed into four muscles pairs. They
are the: pectoralis major, pectoralis minor, xiphihumeralis, and pectoantebrachialis. Deep to the
superficial pectoral muscles are small, strap-like muscles that help to move the ribs. By cutting
and reflecting the pectoral muscles you may observe: transversus costarum, scalenus (anterior,
medius, and posterior), and the external and the internal intercostals.
In Felis there are five postaxial muscles that originate on the scapula and insert on the
humerus. One easily observed muscle is the deltoid. Although the deltoid is a single muscle in
humans, in Felis and most other mammals it is actually three muscles: clavodeltoid (a.k.a.;
clavobrachialis), acromiodeltoid, and spinodeltoid. The deltoid is believed to be homologous to
the dorsalis scapulae that you saw in Necturus. Another muscle is the teres major. The teres
major most likely developed from a slip of the latissimus dorsi. An antagonist to the teres major
is the teres minor, both causing rotation of the humerus. The subscapularis is another of these
muscles and originates from the subscapular fossa. In Felis there are also three ventral muscles
from the scapula that serve to flex the arm. They are the: supraspinatus, infraspinatus, and
coracobrachialis. The supraspinatus and infraspinatus originate from the dorsal scapula (the
supraspinous and infraspinous fossae respectively). This, however, is a modified condition and
these muscles developed from the ventral extrinsic muscle of reptiles called the
supracoracoideus. So they are truly ventral muscles.
Felis Superficial Chest Muscles and Ventral Superficial Arm Muscles

94
Rectus Abdominis Scalenus Medius Transversus Costarum Scalenus Anterior Scalenus Posterior
Serratus Ventralis Levator Scapulae Subscapularis Teres Major
Felis Deep Chest Muscles
Felis Chest Muscles
Muscle Origin Insertion Action
Pectoantebrachialis Manubrium Fascia of forearm Adducts forelimb
Pectoralis Major Sternebrae Humerus Adducts forelimb
Pectoralis Minor Sternebrae Humerus Adducts forelimb
Xiphihumeralis Xiphoid process Humerus Adducts forelimb
Serratus Ventralis R1-R10 Scapula Pulls scapula ventrally
Levator Scapula C3-C7 Scapula Raises scapula cranially
Scalenus Ribs Cervical Vertebrae Flexes neck
External Intercostal Border of rib Border of adjacent rib Expands ribcage
Internal Intercostal Border of rib Border of adjacent rib Compresses ribcage
The muscles of the arm will typically extend (dorsal muscles) or flex (ventral muscles)
the forearm. In Felis the triceps brachii is a well developed muscle. It has three heads. The long
head originates from the infraglenoid tubercle of the scapula and inserts on the olecranon. The
two other heads (the lateral and medial heads will originate on the humerus and also insert on the
olecranon. Mammalian ventral muscles that insert and flex the forearm include: biceps brachii,
brachialis, anconeus, and epitrochlearis. The biceps brachii is homologous to the

95
humeroantebrachialis that you observed in Necturus. When removing the fascia from these
muscles work from the shoulder towards the elbow. Do not go below the elbow until the fascia is
removed from all of the arm muscles. The tendons of the arm muscles will blend in with a tough
connective tissue sheath superficial to the forearm muscles called the retinaculum. It is difficult
to separate the tendons (which you want to save) from the retinaculum until you can see the arm
muscles clearly.
Felis Superficial Dorsal Arm Muscles
Felis Arm Muscles
Muscle Origin Insertion Action
Triceps Brachii Scapula & Humerus Ulna Extends forearm
Anconeus Humerus Ulna Extends forearm
Epitrochlearis Latissimus dorsi, ventral border Ulna Extends forearm
Brachialis Humerus Ulna Flexes forearm
Biceps Brachii Humerus Radius Flexes forearm

96
Felis Deep Dorsal Arm Muscles
Felis Deep Ventral Arm Muscles

97
The muscles of the forearm in Felis can seem confusing but are easy to learn if you
remember two things. The dorsal muscles of the forearm are the supinators and extensors of the
wrist, manus, and digits while the ventral muscles of the forearm are the pronators and flexors of
the wrist, manus, and digits. The other thing to remember is which side of the forearm the ulna
is on and which side the radius is on. On the dorsal forearm the superficial muscles running
from radial side to ulnar side are: brachioradialis, extensor carpi radialis longus, extensor carpi
radialis brevis, extensor digitorum communis, extensor digitorum lateralis, and extensor carpi
ulnaris. On the ventral forearm the superficial muscles running from the ulnar side to the radial
side are: flexor carpi ulnaris, palmaris longus, flexor carpi radialis, and pronator teres. One
dorsal, superficial muscle that you must be careful with is the brachioradialis. This is a very thin
muscle in Felis and can easily be removed with the skin. Be very careful when skinning the
forearm to avoid this.
Felis Superficial Dorsal Forearm Muscles

98
Felis Superficial Ventral Forearm Muscles
Felis Forearm Muscles
Muscle Origin Insertion Action
Brachioradialis Humerus Radius Supinates forepaw
Extensor Carpi Radialis Longus Humerus Carpals Extends wrist/paw
Extensor Carpi Radialis Brevis Humerus Carpals Extends wrist/paw
Extensor Digitorum Communis Humerus Metacarpals Extends digits
Extensor Digitorum Lateralis Humerus Lateral Metacarpals Extends lateral digits
Extensor Carpi Ulnaris Humerus Carpals Extends wrist/paw
Flexor Carpi Ulnaris Humerus & Ulna Carpals Flexes wrist
Palmaris Longus Humerus Palmar Fascia Flexes palm & digits
Flexor Carpi Radialis Humerus Carpals Flexes wrist
Pronator Teres Humerus Radius Pronates forepaw

99
Felis Superficial Buttocks and Lateral Thigh Muscles
Felis Buttocks and Thigh Muscles
Muscle Origin Insertion Action
Gluteus Medius Sacral & Caudal Vertebrae Femur Abducts thigh
Gluteus Maximus Sacral & Caudal Vertebrae Femur Abducts thigh
Caudofemoralis Caudal Vertebrae Patella Abducts thigh & Extends leg
Tensor Fascia Latae Ilium IT Band Tighten IT band
Tenuissimus 2nd Caudal Vertebra Patella Abducts thigh
Biceps Femoris Ischium Patella & Tibia Flexes leg
Semitendinosus Ischium Tibia Flexes leg
Sartorius Ilium Tibia Adducts & Rotates thigh
Gracilis Ischium & Pubic Symphysis Tibia Adducts thigh
Semimembranosus Ischium Femur Extends thigh
Adductor Magnus Ischium & Pubis Femur Adducts thigh
Adductor Longus Pubis Femur Adducts thigh
Pectineus Pubis Femur Adducts & Extends thigh
Iliopsoas Ilium & Lumbar Vertebrae Femur Rotates and Flexes thigh
Rectus Femoris Ilium Patella Extends leg
Vastus Medialis Femur Patella Extends leg
Vastus Lateralis Femur Patella Extends leg
Vastus Intermedius Femur Patella Extends leg

100
Felis Superficial Ventral Thigh Muscles
The muscles of the buttocks will have their origins partially covered by the lumbodorsal
aponeurosis. As a result, care must be taken when removing the fascia from them. When the
origins begin to fray you have removed as much fascia from that area as is reasonably possible.
The superficial buttocks muscles will be the caudofemoralis, gluteus medius, and gluteus
maximus. The gluteus medius is actually the larger gluteal muscle while the gluteus maximus is
a thin flap of muscle in Felis. The tendon of the caudofemoralis will run beneath the biceps
femoris to insert on the patella. Immediately lateral to the gluteal muscles is the tensor fascia
latae. This muscle is attached to a broad tendon called the lateral fascia or iliotibial tract or IT
band. Take care not to damage this tendon until it is time to go deep. The superficial lateral
thigh muscles are the semitendinosus and biceps femoris. The biceps femoris is a large muscle
having prominent fascicles and will be close to the integument. Cutting and reflecting the biceps
femoris will make it easier to see the semitendinosus and a ventral thigh muscle, the
semimembranosus. While reflecting the biceps femoris, you can observe two slender structures
beneath the muscle, the sciatic nerve and the tenuissimus muscle. The tenuissimus is a slender
muscle that resembles a fascicle that broke free from the biceps femoris and will be smaller than
the sciatic nerve. The ventral superficial thigh muscles are the sartorius, gracilis, and
semimembranosus. In Felis the sartorius is a broad, flap-like muscle covering much of the
medial thigh and is visible on the lateral surface of the thigh as well. This is different from the
sartorius in humans which is a thin, strap-like muscle running from the ilium, across the thigh, to
the medial epicondyle. By cutting and reflecting the sartorius and the iliotibial tract you can
observe the quadriceps. Three of the four muscles are immediately apparent. These are the
vastus lateralis (beneath the iliotibial tract), rectus femoris, and vastus medialis (both are below

101
the sartorius). Cutting and reflecting the cigar-shaped rectus femoris will allow you to observe
the fourth muscle, the vastus intermedius. You will also cut and reflect the gracilis in your deep
dissection. This will expose the adductor muscles and make it easier to observe the
semimembranosus and semitendinosus. In Felis there are three adductor muscles; the adductor
magnus (a.k.a.; adductor femoris), adductor longus, and pectineus. After removing some fat and
fascia from the ventral junction of the hip and thigh you will be able to observe a small muscle
anterior to the pectineus. This muscle is the iliopsoas and it runs roughly perpendicular to the
neighboring inner thigh muscles. The iliopsoas is actually two muscle, iliacus and psoas major
that have fused together. When removing fascia from the thigh muscles care must be taken to
preserve their tendons of insertion, as was the case with the arm muscles. Again work from the
girdle out. Do not work below the knee until the thigh muscles are fully exposed to avoid
damaging these tendons.
A prominent feature of the leg is the calf muscle. The "calf" is actually two muscles that
share the calcaneal tendon. The superficial muscle is the gastrocnemius. Immediately deep to it is
the soleus. The soleus can also be distinguished from the gastrocnemius by its bipennate fiber
pattern. The soleus is more easily observed on the medial aspect of the leg. On the lateral aspect
you will see the flexor hallucis longus. The soleus is actually sandwiched between the
gastrocnemius and flexor hallucis longus on the lateral aspect but the soleus is hard to see on this
side. On the posterior-medial aspect of the leg you can observe the tibialis posterior and flexor
digitorum longus running between the soleus and tibia. Anterior to the tibia will be two muscles,
the tibialis anterior and the extensor digitorum longus. On the lateral aspect of the leg are three
slender muscles called the peroneals (a.k.a.; fibulares). The larger, more anterior peroneal
muscle is the peroneus longus. Posterior to it is the smaller peroneus tertius. Both muscles look
similar having the muscle below the knee and tapering into a slender tendon. Between the two is
the third peroneal muscle, the peroneus brevis.

102
Vastus Lateralis Vastus Medialis Rectus Femoris
Vastus Intermedius Rectus Femoris (half reflected)
Felis Quadriceps, Two Views

103
Felis Lateral Shin Muscles
Felis Leg Muscles
Muscle Origin Insertion Action
Gastrocnemius Femur Calcaneus Extends hindpaw
Soleus Tibia Calcaneus Extends hindpaw
Flexor Digitorum Longus Fibula & Tibia Phalanges Flexes digits
Flexor Hallucis Longus Fibula & Tibia Phalanges Flexes digits
Tibialis Posterior Fibula & Tibia Lateral Cuneiform Extends hindpaw
Tibialis Anterior Fibula & Tibia First Metatarsal Flexes hindpaw
Extensor Digitorum Longus Femur Phalanges Extends hindpaw
Peroneus Longus Fibula Metatarsals Flexes hindpaw
Peroneus Brevis Fibula 5th Metatarsal Extends hindpaw
Peroneus Tertius Fibula Metatarsals Extends & Abducts digits
Gastrocnemius Soleus Flexor Hallucis Longus Tibialis Posterior Peroneus Brevis
Tibialis Anterior Extensor Digitorum Longus
Peroneus Peroneus
Longus Tertius

104
Felis Anterior Leg Muscles
Felis Medial Leg Muscles

105
Part 3: Visceral Organs and Angiology
Part A: Exposing the Visceral Organs and Angiology
Part A1: Exposing the Visceral Organs and Blood Vessels in Squalus
Angiology is the study of blood vessels. Blood vessels are true organs, being composed
of tissues working together to perform a function. The wall of the typical blood vessel is
composed of three “tunics”: the tunica intima, the tunica media, and the tunica adventitia. The
nature of these tunics varies with the class and type of blood vessel. In your dissection you will
be able to observe two of the three classes of blood vessels, arteries and veins. Colored latex has
been injected into the arteries and veins to make them easier to work with and to study. Pink
latex signifies oxygenated blood while blue latex designates deoxygenated blood flow.
Typically oxygenated blood flows through arteries and deoxygenated blood flows through veins.
However, the situation is reversed in the mammalian pulmonary circuit.
To observe the visceral organs and blood vessels it is necessary to open the
pleuroperitoneal cavity. The coelom in most vertebrates is divided into a pericardial cavity and a
pleuroperitoneal cavity. This will be the case in both Squalus and Necturus. The pericardial
cavity contains the heart and the pleuroperitoneal cavity contains the other visceral organs. In
mammals such as Felis the coelom is divided differently. The diaphragm divides the thoracic
cavity from the abdominopelvic cavity. The thoracic cavity can be divided into the pericardial
cavity, two pleural cavities (that contain the lungs), and the mediastinum. The abdominopelvic
cavity can be divided into the abdominal and pelvic cavities. This division does not have a clear
boundary such as the diaphragm. Typically it is demarcated by the rim of the pelvis.
Your Squalus will already have a ventral incision made when the animal’s visceral blood
vessels were injected with latex. You may use this incision as a starting point. Using your fine
point scissors follow the incision as close to midventral as possible. Extend it cranially to the
coracoid bar and caudally to the cloaca. While extending the incision keep your scissors as far
from the pleuroperitoneal cavity as possible so as to avoid damaging the contents. Your next
step is to make two transverse incisions, one below the coracoid bar and one at the ischiopubic
bar. While folding back the hypaxial musculature you will observe that the inner aspect is
“shiny”. This is due to the parietal peritoneum, a serous membrane. You will also observe that
the visceral organs will also be shiny. This is due to the visceral peritoneum. Both the parietal
and visceral peritoneum are one continuous serous membrane. You will also notice folds of the
peritoneum running between the visceral organs and running within these folds will be visceral
blood vessels. To expose these vessels simply use your scissors and the separating technique to
break the vessels free of these peritoneal folds. The blood vessels in Squalus are very fragile and
care must be taken not to damage the vessel wall. In most cases exposing the blood vessel will be
good enough.
To observe the heart you will need to open the pericardial cavity. Take your fine point
scissors and insert them shallowly between the right and left common coracoarcurals. Separate
these two muscles and you will see a white membrane immediately deep to them. This is the
parietal pericardium. The parietal pericardium has a fibrous outer layer and an inner layer that is
a serous membrane. Carefully open the parietal pericardium to observe the heart.
One other area that will need to be opened for your study of vasculature will be the
oropharynx. Using the bone clippers cut through the mandibular arch, the hyoid arch, and the
gill arches at about the point where the ceratobranchial and epibranchial cartilages meet on one

106
side. Then using your scissors cut across the ventral oropharynx at the coracoid bar being
careful not to damage the heart. This will expose the roof of the oropharynx. You should be able
to see some of the blood vessels beneath the oral mucosa. To expose these blood vessels peel
back the oral mucosa.
Part A2: Exposing the Visceral Organs and Blood Vessels in Necturus
The technique required to expose the visceral organs and blood vessels in Necturus is
similar to that used for Squalus. To open the pleuroperitoneal cavity you should follow the
incision that was already made to inject the visceral vasculature. Extend this incision to the
pectoral and pelvic girdles. Then make two transverse incisions, one below the pectoral girdle
and one above the pelvic girdle. As with Squalus, be careful to pull up on the scissors so as to
avoid damaging the organs. You will be able to observe the parietal and visceral peritoneum as
you open your specimen. You will be able to observe most of the blood vessels by gently
separating them from the peritoneal folds that they are encased in. Unlike Squalus however, you
will need to follow the vessels on to the limbs. This can be done by following the vessels to their
exit point through the body wall and then separating the muscles concealing them.
To observe the heart you will need to open the pericardial cavity. Take your fine point
scissors and insert them shallowly between the right and left portions of the rectus cervicis along
the median raphe. Separate these two muscle portions and you will see a white membrane
immediately deep to them. This is the parietal pericardium. The parietal pericardium has a
fibrous outer layer and an inner layer that is a serous membrane. Carefully open the parietal
pericardium to observe the heart.
One other area that will need to be opened for your study of vasculature will be the
oropharynx. Using your fine point scissors cut through the jaw joint and the lateral wall of the
oropharynx. Then using your scissors cut across the ventral oropharynx at the coracoid plates
being careful not to damage the heart. This will expose the roof of the oropharynx. You should
be able to see some of the blood vessels beneath the oral mucosa. To expose these blood vessels
peel back the oral mucosa.
Part A3: Exposing the Visceral Organs and Blood Vessels in Felis
The coelom of Felis will need to be approached differently than it was in either Necturus
or Squalus. The abdominopelvic cavity can be opened by using your fine point scissors to make
a midventral incision through the linea alba from the diaphragm to the pubis. Keep the scissors
pulled up to avoid damaging the internal organs. Make a pair of transverse incisions, one
immediately below the diaphragm/last ribs and one at the iliac crest. You will be able to observe
the parietal and visceral peritoneum. You should be able to see how the visceral peritoneum is
reflected from the surface of the urinary bladder onto the rectus abdominis to form the parietal
peritoneum. Unlike Squalus and Necturus, Felis will require more work than simply separating
the vasculature from the peritoneal folds to expose blood vessels. There will also be adipose
tissue. Using the tips of your fine point scissors you will be able to break the fat away using the
separating technique. This step can be tedious and a little frustrating. However, after a while
you should be able to see some results. Often the best approach is to open your scissors parallel
to the blood vessels that you are cleaning. Keep in mind that these vessels should be a milky
pink, milky yellow (hepatic portal system), or a milky blue in color. If you have cleaned the
vessel so that its color is a bright pink, yellow, or blue you have actually removed the blood

107
vessel wall and have exposed the latex. This latex will dry out and break on you. You should try
to leave the vessel walls intact.
To open the thoracic cavity you will need to first remove the pectoral musculature. The
rib cage of Felis narrows cranially and it is easy to mistakenly cut through the axillary blood
vessels and brachial plexus. To avoid this use your pressing technique to slice the pectoral
muscles away from their origins along the sternum. You should be able to observe an aperture
where the blood vessels and nerves are exiting the thorax. Place the tip of your bone clipper into
that aperture. Clip through the ribs to about R10. While clipping through the ribs place the
fingers of your non-cutting hand into the thoracic cavity and push on the lungs. This will reduce
the chances that you will accidentally damage these delicate organs. At R10, take your scissors
and cut through the intercostals muscles, along the curve of the ribs, to the sternum. Use your
bone clippers to cut through the sternum and then take your scissors and follow the curve of the
ribs, through the intercostals muscles to the point opposite of where you were clipping through
the ribs. Take the bone clippers and clip through the ribs to the aperture on the opposite side of
where you had started. Now the rib cage can be opened and closed much like the hood of a car.
The anchoring point is the sternohyoid and sternothyroid muscles. Use the pressing technique to
carefully press these muscles away from the neck. This will begin to expose the organs and
blood vessels of the neck.
As was the case with Necturus, you will need to follow the blood vessels into the limbs.
This can be done by following the vessels to their exit point through the body wall and then
separating the muscles concealing them.
Part 3: Visceral Organs and Angiology
Part B: The Respiratory System
The respiratory system will differ markedly between your three anatomical specimens.
All of the organs to be studied in this lab will be part of the conducting portion of the respiratory
system. These are the organs and structures that are involved in ventilation, the exchange of
gases between the water/atmosphere and the gills/lungs. These gases will be exchanged across
the respiratory membrane in external respiration, the exchange of gases between the gills/lungs
and the blood stream. The respiratory membrane will be part of the respiratory portion of the
respiratory system. It will be well vascularized and have a great deal of surface area to increase
the efficiency of this exchange of gases.
Part B1: The Respiratory System in Squalus
Squalus, as is the case with most elasmobranchs, is pentanchid, having 5 pairs of gills.
Along with the gills elasmobranchs also have spiracles. The spiracles can be observed on the
dorsal aspect of the head, posterior to the eyes. Spiracles are modified gills that serve as one-way
intake valves. They are used in rays and skates resting on the sediment to bring in water. The
spiracle evolved into the mammalian eustachian tube. The eustachian tube connects the middle
ear and the pharynx and serves to equalize air pressure in the head.
The gill slits of elasmobranchs are said to be “naked” since, unlike most fishes, they are
not covered by an operculum. Each gill slit opens into a gill chamber. In the first four chambers
the anterior and posterior walls have a demibranch, a well-vascularized gill surface. The fifth

108
chamber has a demibranch only on the anterior wall (i.e.; the demibranch is missing from the
posterior wall). The demibranch on the anterior wall is called the pretrematic demibranch. The
demibranch on the posterior wall is called the posttrematic demibranch. Between the
posttrematic demibranch of one gill slit and the pretrematic demibranch of the next gill slit is a
partition called the interbranchial septum. The interbranchial septum is supported by long,
tapering, and sometimes branching cartilaginous gill rays that radiate from the gill cartilages.
Each demibranch’s functional surface consists of large numbers of lamellae of gill mucosa. This
gill mucosa will be well vascularized to maximize the exchange of gasses between gill capillaries
Posttrematic Demibranch Pretrematic Demibranch Gill Slit
Holobranch Interbranchial Septum Demibranch Spiracle
Squalus Dorsal Oropharynx Displaying Gill Structure
and the water. The folds increase surface area for external respiration. A holobranch is composed
of the two demibranchs of a single gill arch (the posttrematic demibranch of the previous gill
chamber and the pretrematic demibranch of the next gill chamber), the associated interbranchial
septum, cartilages, vasculature, nerves, branchiomeric muscles, and associated connective
tissues. The typical pentanchid shark has four holobranchs. Anterior to the first holobranch is the
pretrematic demibranch of the first gill chamber called the first demibranch. Since the fifth
chamber lacks a posttrematic demibranch there is no second demibranch. Elasmobranch gill
arches also have gill rakers. Gill rakers are stubby, projecting structures radiating from the
pharyngeal border to protect the gills from mechanical trauma. Respiratory water enters by the
mouth and spiracles. It exits by way of the gills. Most sharks use ram ventilation but some
species (and the all species of rays and skates) are able to pump water into the pharyngeal
chambers.

109
Part B2: The Respiratory System in Necturus
Necturus possesses both gills and lungs. These urodeles are neotenic, showing juvenile
characteristics in the adult. The gills are typically a juvenile condition that will be replaced by
lungs in the mature urodele. Although Necturus develops lungs the gills are retained and fully
functional in the adult animal. The lungs are not used for respiration but instead serve as
hydrostatic organs. The pulmonary artery and vein will be seen prominently on the outer surface
of the lung. The lungs of Necturus are elongated, simple sacs.
The gills are the primary source of respiration. The gills are external and easily observed.
They are feathery, well vascularized structures that can be moved to increase the mount of
oxygen crossing the respiratory surface. The exchange os gases between the water and the
respiratory membrane of the gills will be supplemented by respiration across the integument and
the pharyngoesophageal mucosa.
External Gills Lung
Necturus Gills and Lungs
After opening the oropharynx, observe the roof of the oropharynx. You will be able to
observe the internal nares which penetrate the primary palate. This development first appeared in
rhidipstian fishes and was a modification that allows the nares to be used for ventilation.
Necturus - Internal Nares Penetrating the Primary Palate

110
Part B3: The Respiratory System in Felis
In Felis the respiratory system begins with the external nares and nasal cavity. The nasal
cavity is connected to the pharynx by the internal nares. The pharynx begins at the internal nares
and extends down the neck to the level of the glottis. The pharynx communicates with the nose,
mouth, trachea, and esophagus so it serves as a common passageway for air and food. Its walls
are composed of skeletal muscle lined with a mucus membrane. The nature of the mucus
membrane varies regionally. There are three divisions to the pharynx: nasopharynx, oropharynx,
and laryngeopharynx. The nasopharynx is the superior most portion of the pharynx. It lays
posterior to the nasal cavity and extends to the level of the soft palate. It has four openings in its
walls: the two internal nares and the two eustachian tubes. The nasopharynx will exchange air
with the eustachian tubes to stabilize air pressure within the head. The oropharynx is the middle
portion of the pharynx. It extends from the soft palate to the level of the hyoid bone. The
oropharynx has only one opening, the fauces, from the mouth. The laryngeopharynx is a short,
inferior most segment running from the level of the hyoid to the larynx and esophagus.
The larynx is a short passageway between the glottis and trachea. Its walls are supported
by cartilage to withstand air pressure. Most nonmammalian tetrapods have only two pairs of
laryngeal cartilages; the cricoids and arytenoids. Mammals, however, have a greater variety to
the structure of the larynx since they use the greatest range of vocalizations. Most mammals have
the arytenoids and cricoid cartilages and also a third pair, the thyroids. The thyroid and cricoid
cartilages are easily observed and were exposed during your dissection of the neck muscles.
Felis Respiratory Organs Image 1

111
The trachea is a singular tube that carries air from the larynx to the lungs. Due to the
vacuum pressure of air during inhalation, cartilaginous plates reinforce the trachea. These plates
are C-shaped, being open dorsally/posteriorly so as to prevent the inhibition of swallowing. The
posterior wall of the trachea instead is composed of a smooth muscle, the trachealis muscle. You
should cut a window into the trachea to observe its internal structure. This is done by making a
three-sided cut. This will cause the cut section to remain attached at one side. In most tetrapods
the trachea ends when it branches into the right and left primary bronchi which will enter into the
right and left lung respectively. This branching point is called the “carina”. The bronchi are
branching structures that serve to conduct air into the various subsegments of the lungs. The
bronchi are structurally similar to the trachea. In mammals the bronchi branch into primary,
secondary, and tertiary bronchi. The tertiary bronchi branch into even smaller branches called
bronchioles. By using the separating technique you should be able to observe the primary and
secondary bronchi.
To observe the trachea and lungs you will need to remove the thymus. The thymus is a
glandular organ of the immune system. It can easily be teased away from the trachea and large
thoracic blood vessels. Interestingly the thymus can be used to determine the age of your animal.
When a mammal reaches puberty the thymus is at its greatest in terms of size and functionality.
The thymus will then begin to degenerate, a process called age-related involution. The
functional tissue will be progressively replaced by adipose tissue. Observe the thymus in your
anatomical specimen before removing it. How much of the thymus is healthy, lobulated tissue
and how much is fat?
Felis Respiratory Organs Image 2

112
The lungs are paired, sac like structures that serve as the site of external respiration.
Internally the lungs have a respiratory membrane that will be thin and well vascularized to
increase the efficiency of external respiration. Mammalian lungs are paired structures located in
the thoracic cavity. A muscular diaphragm separates the thoracic and abdominopelvic cavities.
The diaphragm is the primary muscle driving mammalian inspiration. The phrenic nerve controls
the diaphragm. You should be able to observe this nerve running along the inferior vena cava. It
is a branch of the brachial plexus. The diaphragm will be assisted to some degree by the
intercostals. Within the thoracic cavity each lung resides in its own chamber called the pleural
cavity. The two pleural cavities are separated by the mediastinum and pericardial cavity. The
mammalian lungs are highly compartmentalized being broken up into asymmetrical lobes,
lobules, segments, and bronchioles. Each subunit receives its own branches of the respiratory
tree; either bronchi or bronchioles. The smallest bronchioles terminate in alveoli; sac like
structures that serve as the site of external respiration in mammals. Felis has seven lobes to the
lungs, three on the left lung and four on the right lung. The left lung lobes are the: left anterior
lobe, left middle lobe, and left posterior lobe. The right lung lobes are the: right anterior lobe,
right middle lobe, right posterior lobe and (right) mediastinal lobe.
Part 3: Visceral Organs and Angiology
Part C: The Digestive System
The digestive system serves to process food brought into the body (ingestion) through
mechanical and chemical forces. Nutrients are released from the food and absorbed into the
body while indigestible food substances are eliminated from the body. The organs of the
digestive system can be divided into two groups: the gastrointestinal tract/ tube and the accessory
organs.
The gastrointestinal tract is also known a, the G. I. tube, the digestive tube, and as the
alimentary canal. Its nature varies throughout the vertebrates but will typically include the
mouth, esophagus, stomach, small intestine, and large intestine. The mouth, or oropharynx, is
not considered to be a portion of the digestive tube proper. It lacks the basic histology found in
the other organs of the digestive tube. So, although the oral cavity is the entrance into the
digestive tube, it is not structurally similar to the other organs of the digestive tube. The other
organs of the digestive tube have the same four tissue layers or tunics. The tunics will vary to
some degree between the different organs but overall will be fairly consistent. These tunics are:
tunica mucosa, tunica submucosa, tunica muscularis, and tunica serosa. The organs of the
digestive tube proper are very closely related to one another developmentally and evolutionarily
and this is reflected in the similarity of their construction and histology.
The accessory digestive organs are located outside of the digestive tube proper. They
include the: teeth, tongue, extrinsic salivary glands, pancreas, liver, and gall bladder. The
relationship between the accessory digestive organs and the digestive tube is easiest to see with
the accessory glands. Accessory glands, such as the pancreas, exist outside of the tube and
release their products into the digestive tube by way of ducts.

113
Part C1: The Digestive System in Squalus
In Squalus the mouth and oropharynx will contain a primary tongue and will be bordered
by numerous teeth arranged into multiple rows. Teeth are lost and replaced through out the
animal’s life, a characteristic termed polyphyodontic dentition. You will notice that the teeth are
all of the same basic shape. This is termed homodontic dentition and is typically a feature of
ectotherms. The primary tongue lacks muscle and so is incapable of independent movement.
It is a crescent-shaped elevation on the floor of the oropharynx that is shaped by the underlying
basihyal and ceratohyal cartilages.
The esophagus is a collapsible, muscular tube extending from the oropharynx to the
stomach. Its primary purpose in vertebrates is to conduct food to the stomach. At the point
where the esophagus meets the stomach there is a circular band of smooth muscle called the
gastroesophageal sphincter (a.k.a.; the cardiac or lower esophageal sphincter). The esophagus of
Squalus will have short, stubby evaginations of the tunica mucosa called esophageal papillae. It
is speculated that these evaginations may help to keep prey trapped in the digestive tract.
The stomach is a muscular outpocketing of the digestive tract that serves to store food.
The stomach can store food for an amazing length of time in sharks. There are documented
cases of captive sharks storing food in the stomach for up to a year. To allow for the storage of
food the stomach must be able to distend. The tunica mucosa will be arranged into folds called
rugae that allow the stomach to stretch. Rugae are found in all vertebrates that possess a
stomach. You should cut a window into the stomach. The stomach contents may be interesting
and an empty stomach will display the rugae. The stomach will have two sphincters associated
with it; the cardiac sphincter and the pyloric sphincter (located between the stomach and small
intestine). In Squalus the stomach is a J-shaped organ having a greater and a lesser curvature.
There are three portions to the elasmobranch stomach. The first portion is the cardia. It is
located where the esophagus meets the stomach. The short, middle portion is called the body.
The stomach will have a narrowing portion where the stomach meets the small intestine called
the pylorus. Peritoneal folds will extend off of the gastric tunica serosa. One extends between
the stomach and liver and is termed the lesser omentum. Another attaches the stomach to the
dorsal pleuroperitoneal wall and is called the mesogaster.
In Squalus, as is the case with most fishes from agnathans through teleosts, the intestine
is typically a straight tube from the stomach to cloaca. In chondrichthyes and certain other
nonteleost fishes the intestine has a spiral valve (a.k.a.; typhosole) that serves to increase surface
area to enhance the efficiency of digestion and nutrient absorption. This portion of the intestine is
called the spiral intestine. Some anatomists choose to consider the spiral intestine, and the short
segment before it, to be portions of the small intestine. The spiral intestine would be the ileum
and the short segment between it and the stomach would be the duodenum. You should a cut a
window into the ileum to observe the spiral valve. A peritoneal fold, called the mesentery
extends from the intestinal tunica serosa to anchor the organ to the dorsal pleuroperitoneal wall.
The portion of the intestine after the spiral valve is called the postvalvular intestine. Those
anatomists who subdivide the chondrichthyan intestine consider the postvalvular intestine to be
the colon of the large intestine. This segment will empty into the cloaca. In chondrichthyes
connected to the postvalvular segment of the intestine is the rectal gland. The rectal gland serves
for osmoregulation and does not play a role in digestion. A peritoneal fold arises from the tunica
serosa of both the rectal gland and the colon called the mesorectum. It serves to attach both
organs to the dorsal pleuroperitoneal wall.

114
Esophagus Stomach: Cardia Pylorus Body Ventral Lobe of Pancreas Spleen
Small Intestine: Duodenum Ileum/Spiral Intestine Spiral Valves
Squalus Digestive Organs 1
Colon Cloaca
Rectal Gland
Squalus Colon and Rectal Gland

115
The accessory digestive organs of Squalus include the liver, gall bladder, and pancreas.
The liver is a prominent gland found in the cranial two thirds of the pleuroperitoneal cavity. It
will have a great deal of oil inside of it. This oil allows the shark to use the liver as a hydrostatic
organ. The liver will produce bile for the break down of lipids. This accessory organ is well
vascularized. It receives nutrient rich blood from the stomach and intestine for processing. The
series of blood vessels that connect the liver to the stomach and intestine make up the hepatic
portal system. The liver of Squalus has a right, left, and median lobe. A peritoneal fold, the
falciform ligament, attaches the liver to the midventral body wall. Along the caudal apex of the
median lobe is the gall bladder. The gall bladder stores bile produced by the liver. The gall
bladder releases bile into the intestine near the pyloric junction by the cystic duct. An organ
located along the greater curvature of the stomach that may be mistaken for a lobe of the liver is
the spleen. Like the liver, the spleen is a well-vascularized organ. It serves as a portion of the
immune system and also serves as a reservoir for blood. The pancreas is a glandular organ that
produces a wide variety of digestive enzymes and buffering agents. In Squalus it has two
separate lobes. The ventral lobe is located on the ventral aspect of the duodenum near the
pyloric junction. The dorsal lobe is an elongated structure located dorsal to the stomach.
Running through the dorsal lobe of the pancreas is the lienomesenteric vein.
Pancreas: Ventral Lobe
Squalus Digestive Organs 2

116
Part C2: The Digestive System in Necturus
The digestive system of Necturus is similar to that of Squalus. The mouth and
oropharynx will contain a primary tongue and have a homodontic dentition, although the number
of tooth rows is much reduced. The oropharynx opens into an esophagus. This muscular tube
will conduct food from the oropharynx into the stomach. It is separated from the stomach by the
cardiac sphincter.
The stomach in Necturus is a J-shaped structure and will show a greater and a lesser
curvature. There are three portions to the stomach in Necturus: the cardia, the body, and the
pylorus. It is difficult to distinguish the cardiac region from the esophagus in Necturus. One
landmark is the attachment point of the gastric artery. That will be at the cardia. A peritoneal
fold from the gastric tunica serosa, the gastrosplenic ligament, will attach the spleen to the
stomach at the cardia. Ventrally another peritoneal fold called the hepatogastric ligament
attaches the liver to the stomach. A third peritoneal fold will attach the stomach to the dorsal
pleuroperitoneal wall called the mesogaster. The tunica mucosa will be arranged into rugae when
the stomach is empty. You should cut a window into the body of the stomach. Many times the
stomachs of these animals will contain crayfish and/or minnows. The stomach and small
intestine meet at the pyloric junction which will contain the pyloric sphincter.
The intestines of Necturus can be divided into the small and large intestines. Unlike
Squalus, the intestines of Necturus are not straight. They will show a slight degree of coiling.
This is particularly true for the small intestine. The small intestine in Necturus is divided into the
duodenum and ileum. The duodenum is the longer segment and will be responsible for the bulk
of nutrient break down and much of nutrient absorption. The ileum will complete the absorption
of nutrients. The small intestine is attached to the dorsal pleuroperitoneal wall by an
outpocketing of its tunica serosa called the mesentery. The mesentery also runs between the
coils of the small intestine and binds them together. The ileum terminates at the large intestine.
Necturus Digestive Organs 1

117
The large intestine of Necturus is composed almost exclusively of the rectum. The
rectum allows for primarily for the storage of feces prior to elimination. It will be attached to the
dorsal pleuroperitoneal wall by an outpocketing of its tunica serosa called the
mesorectum/mesocolon. The rectum will open into the cloaca.
Ileum Duodenum Stomach: Pylorus Body Cardia Liver (retracted) Esophagus
Oviduct Ovary Pancreas Spleen Lung
Necturus Digestive Organs 2
The accessory digestive organs of Necturus include the liver, gall bladder, and pancreas.
The liver is a prominent, grey, glandular organ dominating the ventral aspect of the
pleuroperitoneal cavity. It is lobulated but does not show lobes. As a result, it will appear as one
solid structure. The liver will produce bile. It is attached to the ventral abdominal wall by the
falciform ligament. The gall bladder can be found at the caudal extent of the liver, more on the
right side than midventral. It will store bile produced by the liver to be released into the
duodenum during digestion. The pancreas may be a little hard to distinguish initially. It is
roughly the same color as the duodenum and runs partially along it. You will notice it at the
pyloric junction and should be able to observe the pancreaticoduodenal artery entering it. The
pancreas it an active digestive gland. It will produce a variety of enzymes and buffering agents.

118
Part C3: The Digestive System in Felis
The digestive system of amniotes is more complex than is that of fishes or amphibians.
This is especially true for mammals, such as Felis, due to their endothermic nature. Food must be
processed much more rapidly, and efficiently, in endotherms than in ectotherms so as to be able
to maintain a constant internal temperature. Some of these modifications can be observed in and
around the oral cavity. One obvious feature is the cheeks. The cheeks allow and animal to keep
food in its mouth when chewing. You should be able to observe some of the salivary glands that,
unlike nonmammals, will produce enzymes to aid in digestion. You should be able to see the
parotid salivary gland on the posterior aspect of the masseter. The submandibular salivary
glands are found beneath the angle of the mandible. The tongue of Felis has both a primary and
secondary tongue component. The secondary tongue is glandular in nature and mobile. It will
also have keratinized papillae on the dorsal surface to rasp meat off of bone and to groom the fur.
Felis has a heterodontic dentition meaning that the teeth are of variable shapes. This allows for a
division of labor and is found in animals that chew their food. There are four types of teeth in
mammals. Incisors are chisel-shaped teeth designed to bite food. Canines are the fang teeth
designed for tearing meat. Premolars are intermediate shaped teeth that serve to both tear and
crush food. In Felis the premolar is called the carnassials and is modified to shear through
tendons. Molars are the posterior most teeth and serve to crush food. The nature and relative
numbers of these teeth types will vary based on a species mode of feeding. Another feature that
was not observed in either Squalus or Necturus is the presence of a secondary palate. This
divides the nasal cavity from the oral cavity.
Felis Upper Digestive Tract

119
Food (or to be more accurate, the bolus) will pass from the oral cavity into the pharynx
and then to the esophagus. The esophagus is a collapsible muscular tube that will transport the
food to the stomach. Unlike the other organs of the digestive tract it lacks a tunica serosa along
much of its length. Instead, the entire esophagus cranial to the diaphragm is covered by the
tunica adventitia. The tunica adventitia is a fibrous connective tissue that anchors the esophagus
to the tunica adventitia of the trachea, the tunica externa of the common carotid arteries, and the
epineurium of the vagus nerves. You will need to use the separating technique to extricate these
structures from each other. Also, due to its muscular nature and outer layer of connective tissue
the esophagus may initially mistaken for a neck muscle. You may wish to cut a window into the
esophagus. Notice how its inner aspect is folded to allow for distension during swallowing. It
will terminate immediately below the diaphragm at the cardiac sphincter at the stomach.
Felis Digestive Organs 1
The stomach of Felis is much like that of humans and is fairly simple for a mammal
(some rodents and ungulates have very complex stomachs). It is a J-shaped organ having a
greater and lesser curvature. Coming off of the greater curvature is a four-layered fold of the
tunica serosa called the greater omentum. The greater omentum hangs over the intestines. It will
contain ribbons of fat. Due to its location and appearance it is sometimes referred to as the “fatty
apron”. After opening the abdominopelvic cavity you will need to carefully remove this
peritoneal structure. Take care when removing it from the pancreas and spleen to avoid loosing

120
these organs. A second peritoneal fold arises from the lesser curvature. This is the lesser
omentum. The lesser omentum will attach the stomach to the caudate lobe of the liver. The
stomach of Felis has the four basic mammalian portions. The first portion is the cardia that
receives food from the esophagus. Near the cardia is the fundus, a dome-shaped portion for
storing food. The bulk of the stomach is the body where mechanical and chemical digestion
occurs. The terminal portion is the pylorus which meets the small intestine at the pyloric
junction. You should cut a window in the body of the stomach along the greater curvature. You
will be able to see the rugae if the stomach is empty and should even be able to differentiate
some of the tunics.
Felis Digestive Organs 2
The intestines of amniotes are distinctly divided into a small intestine and large intestine.
The small intestine is very long and highly coiled. It is named for its relatively smaller bore
diameter. This allows for more contact between the digested food and the tunica mucosa of the
intestine which increases the efficacy of nutrient absorption. There are other modifications to the
small intestine that have evolved to maximize nutrient absorption. One of these can be observed
by cutting a window into the intestine. The inner aspect will be covered with numerous finger-
like extensions of the tunica mucosa called villi. Villi dramatically increase surface area to
increase nutrient absorption. The small intestine of mammals, birds, and some reptiles will be
divided into three segments. The first segment is the duodenum. It begins at the pyloric junction
and will have one lobe of the pancreas running partially along its length. The middle segment is
the jejunum. The last segment is the ileum which meets the large intestine at the ileocolic (or
ileocecal) junction. You will notice numerous lymph nodes around the ileum. This is to help to

121
deal with pathogens that may have been ingested with food. Our appendix is a modification of
that. An extension of the intestinal tunica serosa called the mesentery will attach the small
intestine to the posterior abdominal wall and will attach its coils to one another.
The large intestine is shorter than the small intestine. It is named for its greater bore
diameter. Very little nutrient absorption occurs in the large intestine so contact between the
materials and the tunica mucosa is not as important. An extension of the colic tunica serosa,
called the mesocolon, will attach the large intestine to the posterior abdominal wall and will
attach its coils to one another. The large intestine has four principle portions: the cecum, the
colon, the rectum, and the anal canal. The cecum is a blind pocket located immediately below the
ileocolic junction. The ileocolic junction passes indigestible food into the first segment of the
colon, the ascending colon. The ascending colon will connect to the short transverse colon. The
transverse colon connects to the longest segment of the feline colon the descending colon. Most
mammals have only these three colic segments. Humans and apes have a fourth segment, the
sigmoid colon which empties into the rectum. In Felis the descending colon empties into the
rectum. It can be difficult to distinguish the descending colon from the rectum. One landmark is
the rim of the pelvis. The rectum begins below the rim of the pelvis. You will get a better view
of it and of the anal canal when you break the pelvis open for the urogenital system dissection.
The rectum serves for the storage of feces prior to elimination. The anal canal is a very short
segment that will most likely be constricted in your specimen. It serves to pass the feces out of
the body.
Felis Digestive System 3

122
The accessory digestive organs of Felis include the liver, gall bladder, and pancreas. The
liver is more restricted in its location than was the case for Squalus or Necturus but is more
lobulated. It can be initially divided into a right and left portion. Running from the fissure that
divides these two portions is a peritoneal fold called the falciform ligament. The falciform
ligament attaches the liver to the diaphragm. The left portion of the liver is divided into the left
medial, left lateral, and caudate lobe (located at the lesser curvature of the stomach. The right
medial lobe dominates the right portion of the liver. This lobe contains the gall bladder and is
sometimes mistaken for two lobes. The right lateral lobe is along the posterior abdominal wall
and is divided into a cranial and caudal portion. The liver produces bile and bile salts for the
emulsification of fats. The spleen, like the liver, is a well-vascularized organ. Both will be a
rich brown color and the spleen can be confused for a lobe of the liver. The spleen is located
along the greater curvature of the stomach. It is an immune organ that serves as a reservoir for
blood. The liver is well vascularized because it processes nutrient-rich blood. The hepatic portal
system is a series of veins that drain nutrient-rich blood from the capillary beds of the stomach,
small intestine, and large intestine and carry it into the liver. The liver will screen pathogens,
remove poisons, and store various nutrients received from this blood.
Liver Left Medial Lobe Fundus Body Pancreas: Gastric Lobe Duodenal Lobe Spleen
Gall Bladder Liver Right Median Lobe Duodenum Descending Colon Rectum
Felis Digestive System 4

123
The gall bladder is located in a depression, the cystic fossa, in the right medial lobe of the
liver. It will store bile produced by the liver. This bile will tend to give the gall bladder a
greenish hue. The gall bladder is drained by the cystic duct. The cystic duct meets with the right
and left hepatic ducts to form the common bile duct. The common bile duct will meet with the
main pancreatic duct to form the hepatopancreatic ampulla that brings bile and pancreatic juice
into the duodenum near the pyloric junction. You can observe the cystic, right hepatic duct, left
hepatic duct, and common bile duct by using your separating technique to remove the
surrounding peritoneum.
The pancreas is a lobulated glandular organ. In Felis it has two lobes. One lobe runs
along the greater curvature of the stomach and is sometimes referred to as the gastric lobe. The
other lobe runs a short distance along the duodenum and is sometimes called the duodenal lobe.
The two lobes are attached to each other (unlike the situation with Squalus). The pancreas
produces a variety of enzymes and buffering agents.
Part 3: Visceral Organs and Angiology
Part D: The Urogenital System
The urogenital system is actually two interrelated systems, the urinary and reproductive
systems. The kidneys are a nice example of ontogeny recapitulating phylogeny. The embryonic
kidney will go through a pronephric stage. The pronephric kidney will give rise to the
mesenephric kidney which is the adult kidney in Squalus and Necturus. In amniotes, such as
Felis, the metanephric kidney will replace the mesenephric kidney. All three types of kidneys
will work in close association with the blood vessels and will be well vascularized. The kidneys
will filter the blood and then modify this filtrate by removing substances from the blood (tubular
excretion) or from the filtrate (tubular resorption). Due to their development the kidneys will be
located against the musculature of the posterior abdominal wall or posterior pleuroperitoneal
wall. This will cause the kidneys to be dorsal to the parietal peritoneum, a situation termed
retroperitoneal. The ducts that drain the kidneys will be at least partially retroperitoneal as well.
Portions of the reproductive system will include the urinary system or have evolved from
urinary structures. For example, the mesenephric ducts of the mesenephric kidneys developed
into the vas deferens in amniotes. All three species will have two distinct genders. However, in
some cases the specimens may not have gone through puberty prior to being sacrificed. The
organs of these animals will be poorly developed. Typically your specimens will be sexually
mature adults but some of the Felis and Squalus specimens may not be. In some cases the uteri
of Felis and Squalus specimens will contain young. This range of developmental situations may
give you some valuable information depending on the range of specimens present in your lab.
Part D1: The Urogenital System in Squalus
In Squalus the urinary system consists of the paired mesenephric kidneys and
mesenephric ducts. The mesenephric kidneys run the length of the pleuroperitoneal cavity on the
dorsal wall. They flank the dorsal aorta and the paired posterior cardinal veins. In turn the renal
portal veins flank the mesenephros. The renal portal veins take blood rich in metabolic wastes
and deliver it into the afferent renal veins for processing by the mesenephros. The metabolites
will be removed from the blood stream and the blood will then enter into the posterior cardinal

124
veins by way of the efferent renal veins. The resultant urine will drain into the mesenephric
ducts. The mesenephric ducts will drain into the cloaca. These structures are very fragile and
easily damaged. Once you have removed the parietal peritoneum there is nothing left to do. The
clarity of your specimen will be based more on the quality of vascular injections than on any
efforts of yours.
Squalus male Urogenital System
In Squalus the male gonads, the testes, are located at the cranial aspect of the
pleuroperitoneal cavity near the posterior cardinal sinuses. They will be held in place by a
peritoneal fold called the mesorchium. The testes secrete sperm into the mesenephric duct. As a
result, the mesenephric duct varies between sexually mature and immature sharks. The
mesenephric duct will be a straight tube in an immature individual and a convoluted tubule in a

125
mature specimen. The caudal portion of the mesenephric duct will be expanded to give the
seminal vesicle. The seminal vesicles provide seminal fluid to the sperm. The caudal end of the
seminal vesicles will be further expanded to give the sperm sacs. Sperm will be stored in the
sperm sacs. During coitus the sperm will exit the sperm sacs and travel along a groove in the
clasper. The clasper will be inserted into the female cloaca to allow sperm to be introduced into
her urogenital system. You may notice a swollen structure immediately deep to the skin on the
ventral aspect of the clasper at its base. This is the siphon sac. The siphon sac draws in water to
aid in ejaculation during mating.
Squalus Male Urogenital System showing Testis
In female Squalus the gonads, the ovaries, are also located at the cranial aspect of the
pleuroperitoneal cavity near the posterior cardinal sinuses. They will be held in place by a
peritoneal fold called the mesovarium. The ovary will release ova into the oviduct (a.k.a; uterine
tube, fallopian tube). The oviduct begins at a funnel-shaped opening located immediately
posterior to the sinus venosus called the ostium. The oviducts develop from an embryonic tube
called the Muellerian Duct. So in females the mesenephric duct serves only as an excretory
structure. Along the oviduct will be the shell gland (a.k.a; nidamental gland). The shell gland is
located parallel to the caudal third of the ovary and will secrete a leathery shell around the
fertilized egg. The caudal portion of the oviducts will expand in a pregnant shark to become the
right and left uterus. Unlike mammals the chondrichthyan uteri do not unite. Both uteri open
separately into the cloaca. The uterus in a pregnant Squalus will contain numerous yolky eggs
(termed marcolethical) and several young. Young are borne ovoviparously after a gestation of
two years.

126
Squalus Male Urogenital System showing Seminal Vesicle
(Note: The Sperm Sacs are immediately caudal to the seminal vesicles)
Ovary (containing ova) Oviduct Kidney Uterus Cloaca
Squalus Female Urogenital System in an Ovulating Specimen

127
Part D2: The Urogenital System in Necturus
In Necturus the urinary system consists of the paired mesenephric kidneys and
mesenephric ducts. Unlike Squalus, however, the mesenephros do not run the length of the
pleuroperitoneal cavity. Instead they are caudally oriented. This condition leads too the kidneys
of certain jawed fishes and amphibians to be termed opisthonephros meaning “rear facing
kidney”. The kidneys will flank the dorsal aorta and post cava. The renal portal veins will flank
the kidneys. The kidneys taper cranially. Along the lateral margin will be the mesenephric ducts
which open into the cloaca. The cloaca will have an expansion termed the urinary bladder.
However, this structure is not a true bladder and is not homologous to the mammalian urinary
bladder. The kidneys will be easier to see in males since in females the ovaries obscure much of
the surrounding area, especially in ovulating specimens.
Necturus Male Urogenital System
In male Necturus the testes will be located immediately lateral to the middle aspect of the
mesenephros. They are actually embedded within the parietal peritoneum, a condition termed
“intraperitoneal” and held in place by the mesorchium. Numerous tiny white tubules run through
the mesorchium called the efferent ductules. The efferent ductules carry sperm into the kidney, in
particular into the mesenephric duct. As was the case with Squalus, the mesenephric ducts are
both excretory and reproductive structures in males. In a sexually mature Necturus the
mesenephric ducts will be convoluted along the cranial two thirds. Sperm will be carried along
the mesenephric ducts for release into the environment by way of the cloaca. Necturus practice

128
external fertilization. Males will have a swollen region external to the cloaca called the cloacal
gland. This gland produces secretions used in mating.
Necturus Female Urogenital System
(Note: the numerous large eggs are obscuring many structures)
In female Necturus the ovaries will also be located immediately lateral to the middle
aspect of the mesenephros. They are held in place by the mesovarium. When producing ova the
ovaries can become quite large and heavy with eggs. In this condition they will obscure many
lower pleuroperitoneal features. The mesenephric ducts in females are solely excretory n
function and will remain straight throughout life. The reproductive ducts, as is the case with
Squalus, are the oviducts that developed from Muellerian ducts. They will be large convoluted
tubules in a female producing eggs. The oviducts begin at the funnel-shaped ostium, one per
oviduct, located at the cranial aspect of the pleuroperitoneal cavity. The ostium has cilia. Ciliary
beating will draw the ovulated ova into the ostium and oviduct. Along the length of the oviducts
will be numerous glandular evaginations to produce the “jelly” typical of amphibian eggs. The
oviducts terminate at the cloaca. The terminal portion of the oviducts will be expanded and will
store eggs prior to copulation. These terminal expansions are termed uteri by some but are not
considered to be true uteri by most comparative anatomists. While the female is embraced by a

129
male (amplexus), the eggs will be released into the water. The male will be releasing his
spermatozoa simultaneously for external fertilization.
Part D3: The Urogenital System in Felis
In Felis the urinary system consists of the paired metanephric kidneys, the ureters, the
urinary bladder, and the urethra. The kidneys in amniotes are much more compact than are those
of other vertebrates. As is the case for all vertebrate kidneys, the metanephric kidney is
retroperitoneal. Three tissue layers that will also be retroperitoneal will surround it. The first
layer in the renal fascia. This is a layer of connective tissue that anchors the kidney to other
structures. The second layer is a layer of adipose tissue called the adipose capsule. The adipose
capsule serves to protect the kidneys from mechanical trauma. Depending on the amount of fat
in your specimen as a whole, there can be a good amount of adipose in the adipose capsule. Care
should be taken to remove this adipose since there are a number of blood vessels and structures
embedded within it. These structures include the ureters and the adrenal glands. The ureters will
be white, as is the adipose, and can easily be removed with the fat. The adrenal glands are small,
beige endocrine organs located cranial to the kidneys. Due to their small size they also are easily
lost. Use your fine tip scissors and separating technique to break the fat away. The deepest layer
is the renal capsule, a thin layer of fibrous connective tissue that covers the external surface of
the kidney.
Descending Colon Abdominal Aorta Posterior Vena Cava Renal Vein & Artery
Urinary Bladder Ureter Hilus Kidney
Felis Urinary System
When the kidney is unveiled you will notice that it is a bean-shaped organ with a
depression on its medial aspect. That depression is termed the hilus. The hilus is where the renal
artery enters the kidney and the renal vein and ureter exit the kidney. To observe the internal
structure of the kidney you will need to slice it open using your scalpel. Begin your incision on
the convex lateral surface and bring it through to the hilus. Do not cut into the hilus. If the
technique is performed properly the kidney will open and close like a book and the hilus will
serve as the binding of the book. Once you have opened the kidney you will be able to observe a

130
number of features. Gently peel away the renal capsule. The interior of the kidney will have
three differently colored zones. The outermost zone, which is typically a grayish hue, is the
renal cortex. The middle, brownish zone, is the renal medulla. You may notice that the border
between the cortex and medulla has a scalloped appearance. Each of these crescents is the base
of a renal pyramid. The renal pyramids are subunits of the renal medulla. The innermost, ivory
colored zone is the renal pelvis. The flap-like medullary structure projecting into the pelvis is
the renal papilla. The renal pelvis drains urine from the renal medulla. The renal pelvis
resembles the ureter since they are actually composed of the same tissues and developed from an
out branching of the mesenephric duct. The renal medulla and cortex are histologically and
developmentally distinct from the renal pelvis. During development the ureter and renal pelvis
invaded the nephrogenic mesoderm that produces the medulla and cortex of the metanephros.
You may also notice blood vessels throughout the kidney. The kidneys are extremely well
vascularized structures since they remove wastes from the blood stream.
Renal Papilla Renal Pelvis Base of a Renal Pyramid
Renal Capsule Renal Cortex Renal Medulla
Felis Internal Structure of the Kidney
The ureters are paired structures that carry urine from the kidney to the urinary pelvis.
The ureters are retroperitoneal along most of their length. However, the terminal portion
penetrates the parietal peritoneum to enter the pelvis and to drain into the urinary bladder. The
urinary bladders of placental and marsupial mammals are considered to be true bladders. Unlike
other “false bladders” they receive urine directly from the ureters and develop from the allantois,
an embryonic membrane. The ureters will enter the bladder from the caudal-dorsal border. This
allows the bladder more room for distension. The bladder is a muscular organ. If the urinary

131
bladder of your specimen is flaccid and stretched it will be empty. An empty bladder is safe to
cut open. Make an incision along the cranial border of the bladder. You should be able to see
that the internal structure is arranged into folds called rugae to allow for distension. You should
also be able to see the openings of three vessels, the two ureters and the one urethra, on the floor
of the urinary bladder. This feature is called the trigone of the bladder. If the bladder is a hard,
acorn-like structure it contains urine. It is advised that you do not make an incision into a
urinary bladder in this state.
Marsupial and placental mammals lack a cloaca. As a result, the true bladder can not
empty into a cloaca. Instead it empties into a tubular organ called the urethra. The urethra serves
to drain the bladder and to release waste into the external environment. In females the urethra is
a solely excretory organ while the urethra of males is both excretory and reproductive. As a
result the urethra will differ between the two genders. In females the urethra is a short organ. In
males the urethra will pass through the intromittent organ, the penis, and will necessarily be
longer. The male urethra will have three segments. The first segment to receive urine is situated
immediately below the urinary bladder. This segment will be surrounded by the prostate gland
and is called the prostatic urethra. Between the bladder proper and the prostatic urethra in Felis
is a narrow portion called the neck of the bladder. Although this is considered to be a portion of
the urinary bladder it is really a portion of the prostatic urethra. The prostatic urethra drains into
a short segment that penetrates the floor of the pelvis called the membranous urethra. The
membranous urethra drains into the longest segment called the penile urethra (a.k.a.; spongy
urethra) that passes through the length of the penis.
Felis Female Urogenital Organs

132
In mammals the structures of the female reproductive system will give rise to the
structures of the male reproductive system. For that reason we will begin with the female
reproductive system in Felis. The female reproductive system in mammals consists of a pair of
ovaries, a pair of oviducts, a singular uterus, and a singular vagina. The ovary is the female
gonad. It will be located in the lower abdominal cavity in Felis. It will be held in place by
peritoneal folds such as the suspensory ligament. On occasion you will be able to observe a
bulge on the surface of the ovary. This bulge is an ovum that was about to enter ovulation. The
oviduct is a tubular organ found on the lateral margin of the ovary. The oviduct serves to
transport the ovum/embryo to the uterus and also serves as the sight of fertilization. It has three
sections. The first section is the infundibulum. The infundibulum is a funnel-shaped opening and
will have extensions called fimbriae to increase surface area for capturing the ovulated ovum.
The middle and longest segment is the ampulla. The last segment is the isthmus, a narrow
portion that enters the uterus at the apex of the cornu.
Urethra Vagina Urinary Bladder Uterine Body Uterine Cornu Rectum
Felis Lower Female Urogenital System
Placental mammalian uteri come in two forms: simplex and bicornuate. The uterus of
Felis is bicornuate. It is composed of a right and left uterine horn (a.k.a.; uterine cornu) and the
uterine body. The uterine horns are held in place by peritoneal folds such as the broad ligament
and the round ligament. They will be located in the lateral aspect of the lower abdominal cavity.
Their size and width will be dependent on the reproductive state of your specimen. In a pregnant
animal the uterine horns can be quite large. The embryos and fetuses reside in the uterine horns.
They enter the uterine body only during parturition. The two uterine horns meet caudally at the
uterine body. To observe the uterine body, and other pelvic structures, you will need to open the

133
pelvis. Take your scalpel and make an incision through the muscles superficial to the pubic
symphysis. While doing this, exert pressure on the knees with your other hand. In a smaller
specimen this may be enough to open the pelvis. If it is not then take the bone clippers and cut
through the pubic symphysis. The pelvis will contain some adipose tissue that may be removed
through the separating technique. The body of the uterus will meet the vagina. Unless the
specimen is in the later stages of pregnancy the transition may be hard to recognize. The vagina
will be situated between the urethra (ventral) and rectum and anal canal (dorsal). It is a muscular,
tubular organ that will open into the urogenital vestibule.
Felis Male Urogenital System 1
The male reproductive system in mammals consists of a pair of testes, the scrotum, a
paired duct system, the penis, and a number of accessory sex glands. The testis is the male
gonad and will produce spermatozoa and male sex hormones. These organs are homologous to
the ovaries. To allow for spermatogenesis in mammals the testes must be kept at a temperature
that is slightly lower than is the internal core temperature of the animal. As a result, the testes
are held outside of the body in a cutaneous outpocketing called the scrotum. The amount of time
that the testes spend in the scrotum varies based on the reproductive strategy of the species. The
testes will reside in the scrotum only temporarily in mammals with set breeding seasons. In
species such as Felis and humans that are always reproductively active the testes reside in the
scrotum permanently. Using your scissors open one side of the scrotum, only one scrotal sac, to
expose one testis. The scrotum is homologous to the labia majora and labia minora of the female
vulva. The passageway between the scrotum and pelvic cavity is the inguinal canal. The testes
are held in place by collagen bundles.

134
Felis Male Urogenital System Focusing on the Duct System
Sperm will be released into a series of ducts. The testis itself has numerous convoluted
tubules called seminiferous tubules that serve as the site of gametogenesis. The testes are
drained by the epididymis. The epididymides are comma-shaped organs located on the lateral
margin of the testes. The epididymis has three portions: head, body, and tail. The head is the
widest portion. It is located on the superior aspect of the testis and contains the efferent ductules
that receive sperm from the testis. The epididymis tapers from the head into a body and a
narrowing tail. The body and tail contain the ductus epididymis. The tail of the epididymis
connects to the vas deferens (a.k.a.; ductus deferens). The vas deferens will carry sperm from
the scrotum into the pelvis and to the urethra. Along a portion of its length the vas deferens runs
with other organs in a composite structure called the spermatic cord. The spermatic cord
includes the vas deferens, testicular artery, testicular vein, testicular lymphatic, and testicular
nerve. All are covered by a connective tissue sheath and by the cremaster muscle (which is an
outgrowth of the rectus abdominis). The cremaster will allow the testes to be pulled closer to the
pelvis as required by ambient temperatures. You can observe the spermatic cord separating into
its composite structures once it enters the pelvis through the inguinal canal. The blood vessels
will travel cranially towards their branching points while the vas deferens will loop behind the
bladder. To observe the entrance of the vas deferens into the urethra, and other pelvic structures,
you will need to open the pelvis. The spermatic cords will most likely still be along the midline,
over the pubic symphysis and partially embedded in adipose. Use your fine tip scissors to
separate the two spermatic cords from one another and from the pubic symphysis. Next take your
scalpel and make an incision through the muscles superficial to the pubic symphysis. While

135
doing this, exert pressure on the knees with your other hand. In a smaller specimen this may be
enough to open the pelvis. If it is not then take the bone clippers and cut through the pubic
symphysis. The pelvis will contain some adipose tissue that may be removed through the
separating technique.
Neck of Bladder Prostate Urethra Penis: Crus Shaft Glans Prepuce (cut)
Urinary Bladder Spermatic Cord Epididymis: Head Body Tail Testis
Felis Male Urogenital System 2
The two vas deferens enter the urethra at the prostate. The prostate will appear as a slight
swelling surrounding a portion of the urethra. The size will correspond to whether or not your
specimen was of reproductive maturity when it was sacrificed. The prostate is an accessory male
sex gland that contributes to seminal fluid. It will also engorge with blood during the sexual
response to cause the urethra to be only a reproductive structure during mating. This is
necessary since the pH of urine will kill the spermatozoa. The prostate is homologous to the
uterus. The ejaculate (spermatozoa plus seminal fluid) will pass through the urethra to exit at the
urethral aperture at the distal extent of the glans penis. The penis is the intromittent organ of
mammals. It has three regions that can be observed through your dissection. The angle between
the membranous urethra and the base of the penis is termed the crus of the penis. Most of the
length of the penis is termed the shaft of the penis. It will terminate in a distal, expanded portion
called the glans penis. The glans penis is homologous to the clitoris of the female vulva. You
will notice that the dorsal surface of the glans penis will have numerous keratinized, spike-like
structures to aid in coitus. A fold of skin termed the prepuce will cover the glans penis. To
expose the penis you will need to cut through the prepuce. You can do this by pulling up on the
prepuce while inserting your scissors into the opening and using your separating technique until

136
the tip of the glans is exposed. Now you can safely use your scissors to cut through the dorsal
aspect of the prepuce. This will expose the length of the penis.
Part 3: Visceral Organs and Angiology
Part E: The Circulatory System
The circulatory system of vertebrates consists of blood, blood vessels, and a
muscular, pumping heart. Blood vessels are hollow organs that serve to carry blood throughout
the body. Their walls are composed of tissue layers called “tunics”. The innermost is the tunica
intima/tunica interna which is in direct contact with the blood. The second, middle, tunic is the
tunica media. The outermost tunic is the tunic externa/tunica adventitia. There are three classes
of blood vessels: arteries, veins, and capillaries. Arteries serve to carry blood from the heart to
the tissues of the body. Since they are carrying blood at its highest pressure, arteries have
proportionately the thickest walls with the tunica media being very thick. Capillaries are
microscopic blood vessels where materials are exchanged between the tissues and the blood. To
facilitate this, capillaries are very thin walled to maximize diffusion. They are microscopically
small and will not be a feature of your dissection. Veins serve to carry blood back to the heart
from the body’s tissues. Their walls are proportionately thinner than are those of arteries since
they carry blood at a lower pressure. Also, due to the lower pressure, veins will have numerous
valves projecting into the lumen to prevent the backflow of blood.
The heart is a muscular pump. It is located in the pericardial cavity and covered by a
serous membrane called the pericardium. The heart evolved from blood vessels. As a result, the
heart wall is composed of three tissue layers that are similar to the three tunics of large blood
vessels. The inner lining is termed the endocardium. The middle and thickest layer is the
myocardium. It is rich in cardiac muscle tissue to pump the blood. The epicardium is the outer
layer of the heart. The heart wall will receive its own blood supply through the coronary blood
vessels. Vertebrate hearts can be divided into single circuit and double circuit hearts. Single
circuit hearts are found in fishes. Blood passes form the heart to the gills, from the gills to the
rest of the body, and from the rest of the body back to the heart. Blood picks up oxygen from the
gills and delivers it to the rest of the body where the cells will use the oxygen for respiration.
The deoxygenated blood will then be returned to the heart. Double circuit hearts are found in
amniotes. There are two currents or circuits. One is the pulmonary circuit which carries
deoxygenated blood from the heart to the lungs to pick up oxygen. The oxygenated blood is then
carried from the lungs back to the heart. The other is the systemic circuit which carries
oxygenated blood from the heart to all of the tissues of the body. There oxygen is used for
respiration and carbon dioxide is dumped into the blood. The deoxygenated blood is returned to
the heart.
Part E1: The Circulatory System in Squalus
The heart of Squalus is typical for that of most fishes. It has four portions (in sequence of
receiving blood): sinus venosus, atrium, ventricle and conus arteriosus. Fish are said to have a
two-chambered heart, one atrium and one ventricle. The sinus venosus receives deoxygenated
blood from the body’s tissues. It is a thin walled vein having little muscle and being composed
mostly of fibrous connective tissue. The sinus venosus primarily serves to collect deoxygenated

137
Arteries of the Squalus Oropharynx
1. Internal Carotid Artery 2. Hyoidean Epibranchial Artery 3. Radix Aorta
4. Vertebral Artery 5-8 Efferent Branchial Arteries (I-IV) 9. Esophageal Artery
10. Dorsal Aorta

138
Squalus Arteries
1. Dorsal Aorta 2. Celiac 3. Gastrohepatic 4. Gastric
5. Hepatic 6. Pancreaticomesenteric 7. Intraintestinal 8. Duodenal
9. Pyloric 10. Anterior Intestinal 11. Gastrosplenic 12. Posterior Intestinal
13. Posterior Mesenteric 14. Iliac 15. Femoral 16. Caudal
17. Annular

139
Squalus Veins
1. Lateral Abdominal 2. Renal Portal 3. Posterior Cardinal 4. Dorsal Aorta
5. Hepatic Portal 6. Gastric 7. Leinomesenteric 8.Posterior Intestinal
9. Posterior Splenic 10. Pancreaticomesenteric 11. Pyloric 12. Intraintestinal
13. Anterior Splenic 14. Anterior Intestinal 15. Annular
Note: Systemic veins are in blue and veins of the hepatic portal system are in yellow.

140
Squalus Deep Vessels & Heart
1. Afferent Branchial a. 2. Ventral Aorta 3. Conus Arteriosus 4. Ventricle
5. Atrium 6. Sinus Venosus 7. Internal Jugular v. 8. External Jugular v.
9. Subclavian v. 10. Brachial v. 11. Lateral Abdominal v. 12. Posterior Cardinal Sinus
13. Genital Sinus 14. Dorsal Aorta 15. Posterior Cardinal v. 16. Renal Portal v.
17. Afferent renal v. 18. Efferent renal v. 19. Cloacal 20. Iliac
21. Femoral 22. Caudal a. 23. Caudal v. 24. Common Cardinal v.

141
blood from the tissues of the body and has little contractile action. (Blood is pulled into the sinus
venosus due to pressure caused by ventricular contractions.) Blood travels from the sinus
venosus, through the sinoatrial aperture, into the atrium. A pair of unidirectional valves guards
the sinoatrial aperture. These valves prevent the backflow of blood. Blood moves into the atrium
when it relaxes after emptying (systole). The atrium receives blood from the sinus venosus and
pushes it into the ventricle. This chamber is a thin walled muscular sack. Contractions of its
muscular component will push the blood through the atrioventicular aperture into the ventricle.
The atrioventicular aperture is also guarded by a pair of unidirectional valves which also prevent
the backflow of blood. The ventricle is a muscular, thick walled chamber that serves as the main
pumping chamber of the heart. It pumps blood into the conus arteriosus. The ventricle generates
the main force for fish circulation. The conus arteriosus receives blood from the ventricle and
conducts it to the gills by way of the ventral aorta. The conus arteriosus extends to the cranial
most extent of the pericardial chamber and joins with the ventral aorta. The wall of the conus
arteriosus is primarily composed of cardiac muscle and elastic connective tissue. The cardiac
muscle component of the conus arteriosus allows for steady blood pressure in the ventral aorta. It
also has a series of semilunar valves to prevent the backflow of blood into the ventricle. The
conus arteriosus is different between cartilaginous and bony fishes. Cartilaginous fishes, such as
Squalus, have a relatively longer conus arteriosus. Bony fishes, particularly teleosts, have a
shorter conus arteriosus. Since the conus arteriosus is shorter it has a muscular swelling at it s
base to compensate called the bulbus arteriosus.
Squalus Heart
The arteries serve to carry blood from the heart to the tissues of the body. Typically this
blood will be oxygenated but that is not always the case. In fishes, such as Squalus, arterial flow

142
begins with the ventral aorta receiving blood from the conus arteriosus and bringing it to the
gills. This blood is deoxygenated. Capillary beds in the gills will serve as the site of gas
exchange. The oxygenated blood will be recovered by the dorsal aorta and delivered to the
throughout the body. During development the ventral aorta extends craniad, under the pharynx,
and connects with the developing aortic arches. In the primitive gnathostome condition there
were six pairs of aortic arched connecting the ventral aorta and dorsal aorta. The first aortic
arches to develop are those of the mandibular arch. The other five arches develop shortly after.
However, before the sixth pair fully forms, the first pair will disappear leaving only branches
called the spiracular arteries. The second pair will sprout buds that will develop into the first
pretrematic arteries. Other buds will sprout off of the third through sixth pairs of aortic arches.
These buds will become the posttrematic arteries. The posttrematic arteries will give rise to
branches that will become the other pretrematic arteries. Soon during development arches 2
through 6 will develop occlusions where portions of the vessel will be lost. Segments located
ventral to the occlusions will give rise to the afferent branchial arteries while the segments
located dorsal to the occlusions will give rise to the efferent branchial arteries. At the same time
capillary beds will begin to form in the demibranchs. These capillary beds are special in that
they are drained by arterioles instead of venules (which is the typical condition). They are an
example of a rete mirabilia.
Esophageal A. Radix Aorta (poorly injected) Mock up of a Radix Aorta
Efferent Branchial Arteries Hyoidean Epibranchial A. Internal Carotid A.
Squalus Arteries of the Doral Oropharynx
In your dissection you should be able to observe the ventral aorta passing through the
floor of the oropharynx and branching into four pairs of afferent arteries going to the gills. These
structures tend to be poorly injected and will be hard to find. However, the roof of the

143
oropharynx will be fairly well injected and you should be able to observe its arteries. There will
be four pairs of efferent branchial arteries receiving oxygenated blood from the gills. They will
drain into the dorsal aorta that can be observed dorsal to the pleuroperitoneal cavity and at the
caudalmost extent of the oropharynx. The anterior extensions of the dorsal aorta are the paired
radix aortae (which are sometimes referred to as the “paired dorsal aortae”). They are small “L-
shaped” vessels located anterior to the first pair of efferent branchial arteries and immediately
lateral to the vertebrae. Immediately medial to these arteries are the difficult to find vertebral
arteries. Extending anteriorly from each first efferent artery will be a small artery that receives
blood directly from the first gill pair. This is the hyoidean epibranchial artery. The hyoidean
epibranchial artery will extend anteriorly along the roof of the oropharynx. Part way along its
length it will meet with the radix aorta. At its anterior extent it will branch into the stapedial
artery (which services the extrinsic ocular muscles) and the internal carotid artery. Coming off
of the second efferent branchial artery and extending caudally will be a wavy artery called the
esophageal artery. The subclavian arteries will arise from the dorsal aorta between the third and
fourth efferent branchials and will extend towards the pectoral fins. One of its branches will be
the brachial artery of the pectoral fin. Another will be the lateral artery that runs along the lateral
line and lateral abdominal vein. The vein is easy to find but the artery typically injects poorly.
Squalus Early Branches of the Dorsal Aorta

144
Celiac Trunk Gastrohepatic A. Gastric A. Posterior Intestinal A. Gastrosplenic A.
Hepatic A. Pancreaticomesenteric A. Anterior Intestinal A.
Squalus Visceral Arteries 1
The dorsal aorta will extend through the pleuroperitoneal cavity immediately ventral to the
vertebral column. It will give rise to the visceral arteries. The first branch will be the celiac
artery. This is a prominent artery that will divide into two branches: the gastrohepatic artery and
the pancreaticomesenteric artery. The gastrohepatic artery is a short vessel that will divide into
the gastric artery, to service the stomach, and the hepatic artery, to service the liver. The
pancreaticomesenteric artery gives rise to numerous conduits. One easily identified artery is the
anterior intestinal artery. The anterior intestinal will travel along the ventral wall of the intestine
and give rise to numerous annular arteries which run near the folds of the spiral valve. At the
point where the anterior intestinal arises three other vessels will also arise. One will enter into the
pyloric region of the stomach and is the pyloric artery. The second will be the duodenal artery
supplying a small portion of the intestine prior to the spiral valves. The third is the intraintestinal
artery which extends through the spiral valve. The second branch coming off of the dorsal aorta
is the posterior intestinal artery. It will enter the dorsal wall of the intestine. It will also give rise
to numerous annular arteries which will fuse to those that arose from the anterior intestinal
artery. Branching close to the posterior intestinal artery will be the gastrosplenic artery. The
gastrosplenic artery will enter into the spleen and traveling through the spleen will also send
branches into the stomach. In some specimens the posterior intestinal and gastrosplenic arteries

145
Pyloric A. Anterior Intestinal A. Annular Arteries Posterior Mesenteric A.
Gastrosplenic A. Posterior Intestinal A. Dorsal Aorta (poorly injected)
Squalus Visceral Arteries 2
Squalus Left Iliac Artery

146
arise from a common vessel that branches off of the dorsal aorta called the anterior mesenteric
artery. Typically the fourth major artery arising from the dorsal aorta will be the posterior
mesenteric which will service the rectal gland. Near the cloaca the dorsal aorta will branch into
the paired iliac arteries and caudal artery. The iliac arteries run into the pelvic fins and give rise
to the femoral arteries. The caudal artery runs through the tail and caudal fin.
Veins will drain blood from capillary beds. In a triple-injected specimen the veins will
have two colors. Yellow vessels designate veins of the hepatic portal system. The hepatic portal
system is a series of veins draining nutrient-rich blood from the digestive tract and carrying it to
the liver for processing. Blue vessels designate systemic veins. Systemic veins usually convey
blood that is high in carbon dioxide and other metabolic wastes to the heart and, subsequently,
the gills. The systemic vessels will also contain the renal portal system. This venous pathway
carries blood high in muscle waste products from the tail to the kidneys for cleansing. On
occasion an error with specimen processing may cause the colors to be reversed.
Hepatic Portal Vein Gastric V. Anterior Splenic V. Posterior Splenic V.
Pancreaticomesenteric V. Annular V. Anterior Intestinal V.
Squalus Hepatic Portal System 1
The largest vein of the hepatic portal system is the hepatic portal vein. It will run along
the common bile duct and the hepatic artery. It drains three veins: the gastric vein, the
pancreaticomesenteric vein, and the lienomesenteric vein. The gastric vein drains the capillary
beds of the stomach and will run along the gastric artery. The pancreaticomesenteric and

147
lienomesenteric veins are easy to confuse at first since they both run near each other and towards
the intestines. However, the pancreaticomesenteric vein runs with the pancreaticomesenteric
artery while the lienomesenteric vein runs through the dorsal lobe of the pancreas. The
pancreaticomesenteric vein has many of the same branches that you saw with the
pancreaticomesenteric artery. There is an anterior intestinal vein which is on the ventral aspect of
the intestine (parallel to the anterior intestinal artery) and will drain the ventral portions o the
annular veins. There is also a pyloric vein, draining the pyloric stomach, and an intraintestinal
vein, draining the spiral valves. The pancreaticomesenteric vein will also drain the anterior
splenic vein. The anterior splenic vein is a smaller vein running through the peritoneum along the
pyloric portion of the greater gastric curvature and draining the spleen. It is easy to lose when
removing the peritoneum. The lienomesenteric vein drains two fairly easily found veins: the
posterior intestinal vein and the posterior splenic vein. The posterior intestinal vein runs along
the dorsal aspect of the intestines (parallel to the posterior intestinal artery) and drains the
posterior portions of the annular veins. The posterior splenic vein will drain the caudal portion
of the spleen and typically runs parallel to the gastrosplenic artery.
Squalus Hepatic Portal System 2
The systematic veins anterior to the pleuroperitoneal cavity will not be injected and hard
to find at best. However, many systematic veins in the pleuroperitoneal cavity should be fairly
well injected and easier to work with. The renal portal system of the systemic vessels begins with
the caudal vein carrying blood from the tail into the pleuroperitoneal cavity. The caudal vein

148
feeds into the right and left renal portal veins. The renal portal veins run along, and immediately
lateral to, the mesenephric kidneys. Numerous small veins will branch off of the renal portal
vein and enter into the kidney. These are the afferent renal veins. The afferent renal veins
deliver the blood to the kidneys for the removal of metabolic wastes, excepting carbon dioxide.
(The tail is the main locomotory structure for swimming in sharks and the muscles will produce a
good deal of waste.) The blood will be processed by the kidneys and will then drain into a series
of small vessels called efferent renal veins. The afferent renal veins are on the lateral half of the
kidney and the efferent renal veins are on the medial half of the kidney. These vessels will drain
into the paired posterior cardinal veins. The posterior cardinal veins are located medial to the
mesenephros and immediately lateral to the dorsal aorta. These paired vessels will carry the
blood cranially to large venous sinuses. These are the right and left posterior cardinal sinuses. A
sinus is a thin walled but large vein and so the posterior cardinal sinuses are fragile. They will be
located behind the liver, near the gonads, in the cranial one fifth of the pleuroperitoneal cavity.
Immediately caudal to the posterior cardinal sinus, and draining into it, is the genital sinus. In
most specimens it will be difficult to differentiate the genital sinus from the posterior cardinal
sinus. The right and left posterior cardinal sinus will drain into the right and left common
cardinal veins respectively. The common cardinal veins will return blood to the heart by feeding
blood into the sinus venosus. Afferent Renal V.
Squalus Renal Portal System

149
Squalus Pleuroperitoneal Vessels
The femoral vein can be observed by separating the pelvic depressor muscles. It will
drain into the iliac vein which will run parallel to the iliac artery and drains into the lateral
abdominal vein. The lateral abdominal veins will receive blood from veins of the lateral wall
myomeres. In addition, the right and left lateral abdominal veins will both drain the cloacal
veins. The cloacal veins enter into the lateral abdominal vein at the same point as the iliac veins.
The lateral abdominal vein will run through the lateral body wall near to the lateral line and
lateral abdominal artery and will carry blood cranially. At the pectoral girdle it will drain the
subclavian vein. The subclavian vein carries blood from the brachial vein of the pectoral fin. At
its most cranial extent the lateral abdominal vein will drain into the common cardinal vein. The
common cardinal veins will return blood to the heart from all of the systematic vessels. It drains
the lateral abdominal, anterior cardinal, internal jugular veins and the posterior cardinal sinus.
Unfortunately it is usually difficult to observe this important vessel due to its location deep to the
coracoid bar and its fragility.

150
Squalus Showing the Right Posterior Cardinal Sinus
Part E2: The Circulatory System in Necturus
The heart of Necturus, like most amphibians, will display modifications due to aerial
respiration. These modifications allow oxygenated blood returning from the lungs or swim
bladder to be separated in the heart from deoxygenated blood returning from the other organs.
There are typically four modifications in the amphibian heart over that of gill breathing fishes.
One modification was the establishment of a complete or partial interatrial septum. This
establishes either the complete or partial separation of a right and left atrium. The interatrial
septum is complete in anurans and some urodeles. The septum is lacking in lungless urodeles. It
will be partial in Necturus although they do not use the lungs for respiration. In most species the
veins for the swimbladder/lung empty into the left atrium. So this blood will be oxygenated.
Deoxygenated blood enters the right atrium by way of the sinus venosus. Another modification
was the establishment of a partial interventricular septum or of ventricular trabeculae. Both
structures serve to separate oxygenated and deoxygenated blood. Necturus will possess a partial
interventricular septum. A third modification was the establishment of a spiral valve in the conus
arteriosus. The spiral valve also serves to separate oxygenated and deoxygenated blood.

151
It forces oxygenated blood into the aortic arches that go to the tissues. It also forces
deoxygenated blood into the aortic arches that go to the lungs or gills. Urodeles, such as
Necturus, lack the spiral valve. The fourth modification is the shortening of the ventral aorta to
point where it is virtually nonexistent. As a result blood goes directly from the conus arteriosus
to the appropriate vessel. Necturus, and urodeles in general, are the exception and have a
prominent ventral aorta.
Bulbous Arteriosus Left Atrium Parietal Pericardium
Right Atrium Conus Arteriosus Ventricle
Necturus Superficial Heart
The Necturus heart possesses two atria and one ventricle. It is sometimes referred to as a
“three chambered heart”. Atria are relatively thin walled structures that receive blood from veins
The right atrium receives deoxygenated blood from all of the tissues of the body by way of the
sinus venosus. The sinus venosus is a thin walled vessel located dorsal to the heart. A valve, the
sinoatrial valve, guards the opening between the sinus venosus and the right atrium. The left
atrium receives oxygenated blood from the gills by way of the pulmonary trunk. The two atria
are separated by a partial interatrial septum. Both atria will send blood into the ventricle.
However, the oxygenated and deoxygenated blood will be incompletely segregated by a partial
interventricular septum. The ventricular wall is more muscular, and thicker, than is that of the
atria. A pair of valves, the right and left atrioventricular valves (or AV valves), prevents the
backflow of blood between the ventricle and the two atria. The ventricle will send blood into the

152
Necturus Arteries & Heart
1. Bulbus Arteriiosus 2. Conus Arteriosus 3. R. Atrium 4. L. Atrium
5. Ventricle 6. External Carotid 7. Internal Carotid 8. Vertebral
9. Branchials 10. Radix Aorta 11. Subclavian 12. Pulmonary
13. Cutaneous 14. Axillary 15. Brachial 16. Segmentals
17. Dorsal Aorta 18. Gastric 19. Celiacomesenteric
20. Pancreaticoduodenal 21. Hepatic 22. Splenic 23. Mesenterics
24. Intestinals 25. Genitals 26. Renals 27. Iliac
28. Femoral 29. Caudal 30. Hypogastric

153
Necturus Veins
1. Common Cardinal 2. Subclavian 3. Brachial 4. Posterior Cardinal
5. Pulmonary 6. Post Cava 7. Genital 8. Renal
9. Renal Portal 10. Pelvic 11. Iliac 12. Femoral
13. caudal 14. Ventral Abdominal 15. Mesenteric 16. Intestinals
17. Hepatic Portal 18. Gastrosplenic 19. Splenic 20. Gastric
Note: Systemic veins are in blue and veins of the hepatic portal system are in yellow.

154
conus arteriosus. This artery will have numerous semilunar valves to prevent blood from flowing
backwards. The conus arteriosus in teleosts and amphibians are relatively shorter than was that
of Squalus. To compensate for this the conus arteriosus has a swollen, muscular structure called
the bulbous arteriosus which will help to propel the blood out of the pericardial cavity.
Efferent Common Trunk External Carotid A. Internal Carotid A. Radix Aorta Dorsal Aorta
Necturus Blood Vessels of the Head
The conus arteriosus will primarily send deoxygenated blood into the ventral aorta. The
ventral aorta is relatively long in urodeles such as Necturus for amphibians. It will carry blood
cranially and will branch into a right and left trunk that will immediately branch into an anterior
first afferent artery and a larger common (afferent) trunk. In turn the right and left common
trunks will branch into the second and third afferent branchial arteries on each side. The afferent
branchial arteries will bring oxygen-poor blood to the gills. The vasculature will ramify into
smaller arteries and capillary beds to allow for an exchange of gases. The oxygenated blood will
travel into anastomizing vessels that will coalesce into the three pairs of efferent branchial
arteries. The first efferent branchial artery will send off a branch that will service the floor of the
mouth called the external carotid artery. It will also send off a second, more medial, branch that
will service the roof of the mouth called the internal carotid artery. The second and third efferent
branchial arteries will join to form a second, efferent, common trunk. Where the second and
third efferent arteries join at the efferent common trunk another artery arises. This is the
pulmonary artery. The right and left pulmonary arteries will travel to the lungs and can be seen

155
on the dorsal pulmonary surface. The pulmonary arteries are the remains of the sixth aortic arch.
A pair of radix aortae will arise from the fusion of the efferent common trunk with the first
efferent artery on each side. These “L-shaped” paired arteries will fuse caudally, near the
pericardial cavity, to form the singular dorsal aorta. The dorsal aorta will carry blood the length
of the body and into the tail where it becomes the caudal artery.
Necturus Visceral Blood Vessels 1
The dorsal aorta runs immediately ventral to the vertebral column. It will branch into the
paired subclavian arteries in the pectoral region. The subclavian artery will exit the body wall
and become the axillary artery. The axillary artery will give rise to the brachial artery that runs
along the humerus. Pairs of arteries will arise segmentally from the dorsal aorta and run into the
lateral body wall. These are the parietal arteries. They will be located internal to the myosepta.
The first visceral branch to arise from the dorsal aorta is the gastric artery. This vessel will
service the stomach. The next visceral branch is the prominent celiacomesenteric artery. The
celiacomesenteric is a major artery of the digestive tract. It will give rise to four branches
(although branching may vary between individuals). The first branch will enter into the liver and
is the hepatic artery. The next branch, which arises almost at the same point as the hepatic, is the
splenic artery that services the spleen. The third branch is the pancreaticoduodenal artery and is a
continuation of the celiacomesenteric. As its name indicates, it enters into the pancreas and
passes through the pancreas to the duodenum. The pancreaticoduodenal will also service the
pylorus. The fourth branch, which occurs only in some specimens, will be the first mesenteric
artery. Several additional mesenteric arteries will arise separately from the dorsal aorta. Each
will give rise to a number of smaller arteries that will vascularize the duodenum and ileum called

156
the intestinal arteries. A series of paired arteries will originate from the dorsal aorta. Some will
service the testes or ovaries and are called generically the gonadal or genital arteries. More
specifically they can be named the testicular or ovarian arteries respectively. Another series of
paired arteries arising from the dorsal aorta in this region are the renal arteries. They are more
numerous than the genital arteries. These vessels will enter and vascularize the mesenephros.
Caudal to the last pair of renal arteries will arise the right and left iliac arteries. The iliac arteries
will exit the pelvis and enter into the thigh where they become the femoral arteries. A second
branch will arise from the iliac artery in the pelvis and will send branches into the cloaca and
bladder. This vessel is the hypogastric artery.
Necturus Visceral Blood Vessels 2

157
Necturus Visceral Blood Vessels 3
Necturus Visceral Blood Vessels 4

158
Necturus Pelvic Arteries
Veins will drain blood from capillary beds. As we saw in Squalus, in a triple-injected
Necturus specimen the veins will have two colors. Yellow vessels designate veins of the hepatic
portal system. Blue vessels designate systemic veins. Necturus will also have a renal portal
system component to the systemic division.
The hepatic portal system is a series of veins draining the capillary beds of the digestive
tract and delivering this nutrient-rich blood to the liver for processing. The hepatic portal vein is
located in a cleft along the dorsal aspect of the liver. It will enter the liver and break up into the
liver sinusoids. The hepatic portal vein drains the mesenteric, gastrosplenic, and the ventral
abdominal veins. The gastrosplenic vein drains the gastric vein and the splenic vein. It joins
with the mesenteric vein within the pancreas. The mesenteric vein is a singular vessel running
through the mesentery dorsal to the intestines. It will receive blood from a number of intestinal
veins that drain intestinal capillary beds. The ventral abdominal vein services both the hepatic
portal system and a portion of the systemic circulation called the renal portal system.
The renal portal system drains blood from the tail, pelvis, and hind limbs and delivers it
to the kidneys for the processing of metabolites. As in Squalus, the renal portal system of
Necturus includes a caudal vein that drains the tail. It will divide into the two renal portal veins.
The renal portal veins will run lateral to the mesenephros and give rise to numerous afferent
renal veins that will enter the kidneys. The right and left renal portal veins will continue to run

159
Intestinal Veins Mesenteric V. Pancreaticoduodenal V. Gastrosplenic V.
Necturus Hepatic Portal System

160
along the lateral surface of the kidney until at the cranial aspect of the mesenephros where they
will give rise to the right and left posterior cardinal veins. The renal portal veins will also
receive small branches from the body wall called parietal veins. In addition, the iliac veins drain
into the renal portal veins. The iliac vein will drain the hind limb via the femoral vein. At the
junction of the femoral and the iliac veins will arise the ventral abdominal vein. The ventral
abdominal vein connects into the hepatic portal system. This vessel is typically lost when the
abdominal wall is opened.
Renal Portal V. Genital Veins Hepatic V. Hepatic Portal V.
Efferent & Afferent Renal Veins Posterior Vena Cava Intestinal V.
Necturus Some Renal Portal Veins

161
Pulmonary V. Posterior Cardinal V. Dorsal Aorta Postcava Hepatic Portal V.
Parietal Arteries & Veins
Necturus Posterior Vena Cava and Some Other Veins
A prominent systemic vein of the posterior body is the posterior vena cava (a.k.a.;
postcava/post cava). It begins medial to the kidneys and runs through the pleuroperitoneal
cavity. At its starting point it will drain the efferent renal veins and the genital veins. As it
travels through the pleuroperitoneal cavity it enters into the liver. It travels cranially through the
liver, receiving blood from several hepatic veins along its length, and will eventually drain into
the hepatic sinus. The hepatic sinus drains into the common cardinal veins that will deliver

162
blood to the sinus venosus. The common cardinal veins ultimately drain most of the systemic
veins. The sinus venosus delivers blood to the right atrium. Once the posterior vena cava enters
the liver you will have a difficult time seeing its other connections. The posterior cardinal veins,
which arose from the renal portal veins, will travel along the dorsal wall of the pleuroperitoneal
cavity immediately flanking the dorsal aorta. These vessels receive blood along their lengths
from a number of parietal veins that drain the dorsal body wall cranial to the mesenephros. The
posterior cardinal veins will deliver blood to the common cardinal veins. Based on the quality of
injection, you may be able to observe the subclavian vein. It will also drain into the common
cardinal vein. The subclavian vein receives blood from the brachial vein by way of the axillary
vein. These three vessels will be running parallel to the similarly named arteries. Another vein
located in the pleuroperitoneal cavity is the pulmonary vein. The right and left pulmonary veins
drain the right and left lungs respectively. The two pulmonary veins will anastomize to form the
pulmonary trunk. The pulmonary trunk will enter into the left atrium. The systemic veins
anterior to the pleuroperitoneal cavity are rarely injected and so will be difficult to observe at
best.
Part E3: The Circulatory System in Felis
The heart of Felis is located in the pericardial cavity of the thoracic cavity. It will be
surrounded by the pericardium. As noted before, be careful when removing the pericardium so as
to avoid losing the vagus nerve (CN.X). The vagus nerve will help to modulate the heartbeat.
Mammals have a “four chambered heart”. The four chambers are two atria and two ventricles.
Unlike most vertebrates, adult mammals lack a sinus venosus. Instead the blood vessels that
ancestrally drained into the sinus venosus will drain directly into the right atrium. These vessels
are the superior vena cava and inferior vena cava. The ancestral location of the sinus venosus in
mammals is marked by the location of the sinoatrial node. Externally the atria in mammals have
outpocketings called auricles. The auricles will be a darker brown color, which makes them easy
to distinguish from the ventricles. Internally the right and left atria are completely separated by
an interatrial septum. The right atrium receives deoxygenated blood from the superior and
inferior vena cavae. The left atrium receives oxygenated blood from the lungs by way of the
pulmonary veins. Externally the right and left ventricles can be easily observed. They will be a
paler shade of brown than will be the auricles. The two ventricles are externally divided by a
groove on the ventral wall of the heart called the anterior interventricular sulcus. An anterior
interventricular artery and vein will be located in this sulcus and are part of the coronary blood
vessels. Internally the ventricles are separated by a complete interventricular septum to prevent
the mixing of oxygenated and deoxygenated blood. The right ventricle will send deoxygenated
blood into pulmonary circulation by way of the pulmonary trunk. The left ventricle will send
oxygenated blood into systemic circulation by way of the aorta, the largest artery of the body.
The walls of the four chambers will vary in thickness based on the amount of cardiac muscle
present. The amount of cardiac muscle in a chamber correlates with function. As a result
ventricles, which pump blood further than do atria, are more muscular than are atria.
Anastomizing muscular ridges on the luminal surface of the ventricle called trabeculae carnae
reinforce the ventricular walls. Amniote hearts have valves designed to prevent the backflow

163
Felis Arteries
1. R. Common Carotid 2. L. Common Carotid 3. Transverse Scapular 4. Thyrocervical
5. Internal Mammillary 6. L. Subclavian 7. Brachiocephalic 8. Vertebral
9. Costocervical 10. Ventral Thoracic 11. Long Thoracic 12. Thoracodorsal
13. R, Subclavian 14. Subscapular 15. Axillary 16. Brachial
17. Arch of Aorta 18. Pulmonary Trunk 19. Pulmonary Arteries 20. Thoracic Aorta
21. Abdominal Aorta 22. Celiac Trunk 23. Hepatic 24. (L.) Gastric
25. Splenic 26. Superior Mesenteric 27. Adrenolumbar 28. Renal
29. Posterior Pancreaticoduodenal 30. Mesenterics 31. Intestinals
32. R. Colic 33. Ileocolic 34. Middle Colic 35. L. Colic
36. Inferior Mesenteric 37. Genital 38. Adrenolumbar 39. External Iliac
40. Femoral 41. Popliteal 42. Saphenous 43. Internal Iliac
44. Caudal/Hypogastric/Median Sacral

164
Felis Veins
1. Anterior Facial 2. Posterior Facial 3. Transverse Jugular 4. Internal Jugular
5. Inferior Thyroid 6. External Jugular 7. Transverse Scapular 8. Brachiocephalic
9. Subclavian 10. Subscapular 11. Ventral Thoracic 12. Long Thoracic
13. Thoracodorsal 14. Brachial 15. Axillary 16. Costovertebral
18. Sternal 18. Azygos 19. Superior Vena Cava 20. Inferior Vena Cava
21. Pulmonary 22. Adrenolumbar 23. Renal 24. Genital
25. Ilioliumbar 26. Common Iliac 27. External Iliac 28. Internal Iliac
29. Femoral 30. Popliteal 31. Greater Saphenous 32. Caudal
33. Posterior Pancreaticoduodenal 34. Superior Mesenteric 35. Mesenterics
36. Intestinals 37. Ileocolic 38. Inferior Mesenteric 39. Hepatic Portal
Note: Systemic veins are in blue and veins of the hepatic portal system are in yellow.

165
of blood. Each valve consists of one or more flaps of fibrous connective tissue called cusps. The
atrioventicular (AV) valves prevent the backflow of blood from the ventricles into the atria. The
AV valves are anchored by connective tissue strands called cordae tendineae to bundles of
smooth muscle called papillary muscle. A second pair of valves, the semilunar valves prevent the
backflow of blood from the arteries (i.e.; aorta and pulmonary trunk) into the ventricles.
Left Ventricle Right Ventricle Right Auricle (of the R. Atrium)
Felis External Heart (L. Atrium is out of view)

166
Left Atrium
Felis Internal Heart
As is true for all amniotes, the circulatory system of Felis consists of a pulmonary circuit
and a systematic circuit. The pulmonary circuit begins with the right ventricle which sends
deoxygenated blood to the lungs by way of the pulmonary trunk. The pulmonary trunk will
divide into the right and left pulmonary arteries to service the right and left lung respectively.
External respiration will occur in the lungs and the oxygenated blood will return to the heart
through the pulmonary veins. The pulmonary veins will empty into the left atrium completing
the pulmonary circuit. The systemic circuit begins with the left ventricle. The left ventricle
sends this oxygen-rich blood out to the body via the aorta. The aorta will give rise to a number of
arteries that serve to carry the blood to all of the tissues of the body. Internal respiration will
occur in these tissues. The deoxygenated blood will be recovered from these tissues by a series
of veins. The veins will ultimately drain into the superior and inferior vena cavae. These two
large veins drain into the right atrium completing the systemic circuit.
The arteries can be observed throughout the body. We will begin with the thoracic cavity.
The pulmonary trunk may be observed exiting the heart and passing under the arch of the aorta.
The pulmonary trunk will divide into a right and left pulmonary artery. These two vessels will

167
Felis Upper Body Arteries1
further subdivide to send a vessel into each lobe of the lung. These pulmonary arteries can be
observed running between the heart and lungs. They will be blue instead of the typical red color
since they are carrying deoxygenated blood. The pulmonary arteries are the remnants of the sixth
aortic arch. The aorta of mammals is equivalent to the dorsal aorta of Squalus and Necturus. It
is a cane-shaped vessel beginning with an ascending aorta that extends a short way cranially. The
aorta then curves back and will run caudally throughout the thoracic and abdominopelvic
cavities. The curving portion is called the arch of the aorta and the caudally running portion is
generally called the descending aorta. Two major branches arise from the arch of the aorta. The
first is the brachiocephalic artery. The brachiocephalic artery will give rise to three primary
branches: right subclavian, right common carotid, and left common carotid. The two carotid
arteries will travel to the head and run medial to, and dorsolateral to, the trachea. Immediately
lateral to the common carotid arteries will be the right and left vagus nerves. A minor branch
will also arise from the brachiocephalic, the sternal artery. The sternal is a short vessel arising
half way along the ventral surface of the brachiocephalic. It will extend to the sternum. The
second branch coming off of the arch of the aorta is the left subclavian artery. (In humans three
major arteries branch off of the arch of the aorta: brachiocephalic, left common carotid, and left
subclavian.) Although they differ in length and point of origin, both the right and left

168
Brachial Thoracodorsal Long Thoracic Ventral Thoracic Common Carotids Thyrocervical Subscapular
Axillary Brachiocephalic Aorta Pulmonary Trunk Subclavian Vertebral Costocervical
Felis Upper Body Arteries 2
subclavian will run to the forelimb and will show a similar branching pattern. The subclavian
will extend cranially through the thorax and will curve to the arm. The curving point is under the
clavicle in humans explaining the name for this artery. Prior to the curving point the subclavians
will give rise to three vessels. The first branch is the vertebral artery that will travel dorsally
towards the vertebrae. The second branch arises close to the vertebral and is called the
costocervical artery. The costocervical will travel a short distance dorsally and then branch into
the costal and cervical arteries. The third branch occurs near the curving point. It is the internal
mammary artery. This vessel travels ventrally to exit the rib cage and service the mammaries. It
was cut when the rib cage was opened. At the curve of the subclavian artery another vessel will
arise, the thyrocervical artery. The thyrocervical artery will extend a short way into the neck. At
its terminus the thyrocervical will give rise to several small vessels. One will extend over the
shoulder and is the transverse scapular artery. At the point that the thyrocervical diverges from

169
the subclavian the subclavian enters the axilla and becomes the axillary artery. The axillary
artery will have three smaller arteries arise from it as it travels through the axilla. Typically all
three vessels will have been severed during your dissection and will appear as shortened nubs.
The first branching is the ventral thoracic artery (a.k.a.; anterior thoracic artery). This artery will
service the pectoralis major. The second artery to branch off of the axillary is the long thoracic
artery. The long thoracic artery will deliver blood to the pectoralis minor and xiphihumeralis.
The third artery to split from the axillary is the thoracodorsal artery. The thoracodorsal services
the latissimus dorsi and, due to its location, is the least likely of the three to be cut during your
dissection. At the terminus of the axillary artery one more artery will arise, the subscapular
artery. It enters into the subscapularis. The axillary at this point becomes the artery of the arm,
the brachial artery. The brachial artery will run along the humerus.
Common Carotids Transverse Scapular Thyrocervical Costocervical Vertebral Subclavian
Vagus N. Subscapular Brachial Axillary Thoracodorsal Long Thoracic Internal Mammary
Felis Upper Body Arteries 3
The descending aorta can be regionally divided into the thoracic aorta and the abdominal
aorta. The thoracic aorta will give rise to a number of small arteries that will run between the
ribs called intercostal arteries. The descending aorta becomes the abdominal aorta once it
penetrates the diaphragm. It will give rise to a number of visceral arteries and arteries of the
pelvis and hindlimb.
The first visceral branch to arise from the abdominal aorta is the celiac trunk. This is a
short, thick artery that will give rise to three other arteries. The first is the hepatic artery that will
enter the liver. The second is the left gastric artery that enters into the stomach along the greater
curvature. The third is the splenic artery which is actually the continuation of the celiac. It will
service the spleen. The second visceral branch is the superior mesenteric artery. This is large

170
Felis Cranial Abdominal Arteries
artery that vascularizes the pancreas, small intestine, and much of the large intestine. It will give
rise to a number of branches: posterior pancreaticoduodenal, mesenterics, ileocolic, right colic,
and middle colic arteries. The first branch of the superior mesenteric is the posterior
pancreaticoduodenal artery (a.k.a.; inferior pancreaticoduodenal) that will run through the
duodenal lobe of the pancreas and enter the duodenum near the pyloric junction. A number of
arteries will arise caudal to the branching point for the posterior pancreaticoduodenal. These are
the mesenteric arteries. The mesenteric arteries travel through the mesentery and will further
divide into smaller vessels servicing the walls of the small intestine called intestinal arteries.
Caudal to the diverging point of the mesenterics will arise the ileocolic artery, then the right colic
artery, and finally the middle colic artery. The ileocolic will enter into the colon at the ileocolic
junction. The right colic enters into the ascending colon near the ileocolic junction. The middle
colic will run along the wall of the transverse and descending colon until it anastomizes with the
left colic artery. Caudal to the branching point of the superior mesenteric will arise the paired
adrenolumbar arteries. These vessels will service the adrenal gland and a portion of the lumbar
musculature. Along the length of the abdominal aorta a series of smaller paired arteries will arise
and enter into the lumbar musculature called lumbar arteries. Immediately caudal to the
origination point of the adrenolumbar will arise a second prominent pair of arteries, the renal
arteries, which service the kidneys. Caudal to this point a smaller pair of vessels will branch off
of the abdominal aorta, the gonadal arteries. The gonadal arteries will arise at the same point in

171
Right Colic Ileocolic Middle Colic (cut) Superior Mesenteric Celiac Trunk Adrenolumbar
Felis Some Visceral Arteries 1
Felis Some Visceral Arteries 2

172
Felis Abdominopelvic Vessels 1
males and females. In females they are shorter vessels called ovarian arteries that will
vascularize the ovary and send branches along the uterine cornua. In males these vessels are
named testicular/spermatic arteries and will descend with the testes into the scrotum during
development. As a result, testicular arteries are longer and will be a portion of the spermatic
cord along part of their length. The next branch to originate from the abdominal aorta is the
singular inferior mesenteric artery. The inferior mesenteric will enter the caudal one third of the
descending colon and will send out a branch called the left colic. The left colic artery will travel
along the wall of the descending colon until it anastomizes with the middle colic artery. At the
crest of the ilium a pair of vessels will arise from the abdominal aorta termed the iliolumbar
arteries. The abdominal aorta terminates at the lesser pelvic brim giving rise to the paired
external iliac, paired internal iliac, and singular hypogastric arteries. The external iliac arteries
will travel along the greater pelvic brim and exit the pelvis through the body wall. At this point
they give rise to a major artery, the femoral, and several smaller vessels. The femoral will travel
through the thigh along the femur until it branches into two arteries near the knee. One branch,
the popliteal artery, dives deep and travels to the posterior knee. The other branch is the
saphenous artery and will be a continuation of the femoral traveling along the medial shin. The
internal iliac artery will dive deep into the pelvis and give rise to a number of smaller branches.
In humans the external and internal iliacs arise from a common vessel named the common iliac

173
artery. The hypogastric (a.k.a.; median sacral artery) will be located immediately ventral to the
sacrum and will travel into the tail as the caudal artery.
Felis Pelvic and Lower Abdominal Blood Vessels

174
Saphenous A.(cut) Popliteal A. Femoral A. Internal Iliac A. External Iliac A.
Greater Saphenous V. Popliteal V. Femoral V.
Felis Thigh Blood Vessels
Veins will drain blood from capillary beds. As we saw in Squalus and Necturus, in a
triple-injected Felis specimen the veins will have two colors. Yellow vessels designate veins of
the hepatic portal system. Blue vessels designate systemic veins. Felis, unlike Squalus and
Necturus, lack a renal portal system component to the systemic division.
The tissues of the body cranial to the heart will heart will be drained by a series of
systematic veins that will ultimately drain into the superior vena cava. The superior vena cava
drains into the right atrium. This vessels is the right precava of other vertebrates. The superior
vena cava is fed by: right and left brachiocephalic, sternal, right and left costovertebral, and
azygous veins. The azygous vein is dorsal to the superior vena cava on the right. It will run
caudally (giving it a “cane-shape”) and drains the intercostal veins. The azygous is found in
amniotes and is the remains of the right posterior cardinal vein of other vertebrates. The azygous
also receives blood from the hemizygous. The hemizygous is a smaller vessel than the azygous
and is found medially on the left side of the posterior thoracic wall. It will drain the left
intercostal veins. The hemizygous is the remains of the left posterior cardinal vein of
nonamniotes. The right and left costovertebral veins typically drain into the superior vena cava
(although they may feed into one or both brachiocephalic veins in some specimens). These veins
are located near the costocervical and vertebral arteries. The costovertebral vein will divide into

175
Felis Upper Body Veins
the costocervical and vertebral veins. Arising from the ventral surface of the superior vena at
approximately the halfway point of its length will be the sternal vein. The sternal vein will drain
the right and left internal mammary veins that will enter the ribcage from the surface of the
thorax. Depending on the amount of damage inflicted when you opened the ribcage, you may
see just the sternal or one or both of the two internal mammaries. The brachiocephalic veins are
a pair of large veins that drain the head and arms. They will receive blood from two primary
veins, the subclavian and jugular veins. The subclavian vein will vary in length but is typically
short (shorter than the subclavian artery that it runs with). It receives blood from the subscapular
and axillary veins. The subscapular vein drains smaller vessels of the subscapular region and can
be observed to enter the subscapularis. The axillary vein drains the ventral thoracic, long
thoracic, thoracodorsal, and brachial veins. The ventral thoracic, long thoracic, and
thoracodorsal veins will be similar to the similarly named arteries in terms of area serviced,
location, and having been cut during the dissection. The brachial vein will be located in the arm
and will run with the brachial artery. The jugular vein will branch into the external and internal
jugular veins. The internal jugular veins are rarely well injected. However, when they are, they
can be observed to run along the common carotids, in fact their tunica adventitiae will fuse
together. The external jugular vein is a prominent vessel and easy to observe. This vessel runs

176
Superior Vena Cava Sternal Costovertebral Brachiocephalic Subclavian External Jugular
Axillary Thoracodorsal Long Thoracic Ventral Thoracic
Felis Some Thoracic Veins
lateral to the internal jugular and will be much larger. Partway along its length it will
communicate with the transverse scapular vein. The transverse scapular will extend dorsally over
the shoulder and will drain the cephalic vein of the dorsolateral arm. At it’s most cranial extent
the external jugular will receive blood from three vessels: posterior facial, anterior facial, and
transverse jugular veins.
Another set of upper body blood vessels is the right and left pulmonary veins. These
vessels and their tibutaries are not a portion of the systemic circuit but instead are part of the
pulmonary circuit. They will transport oxygen-rich blood from the lungs to the left atrium
(which is the end of the pulmonary circuit).. As a result, these vessels will not be blue but will
instead be red.
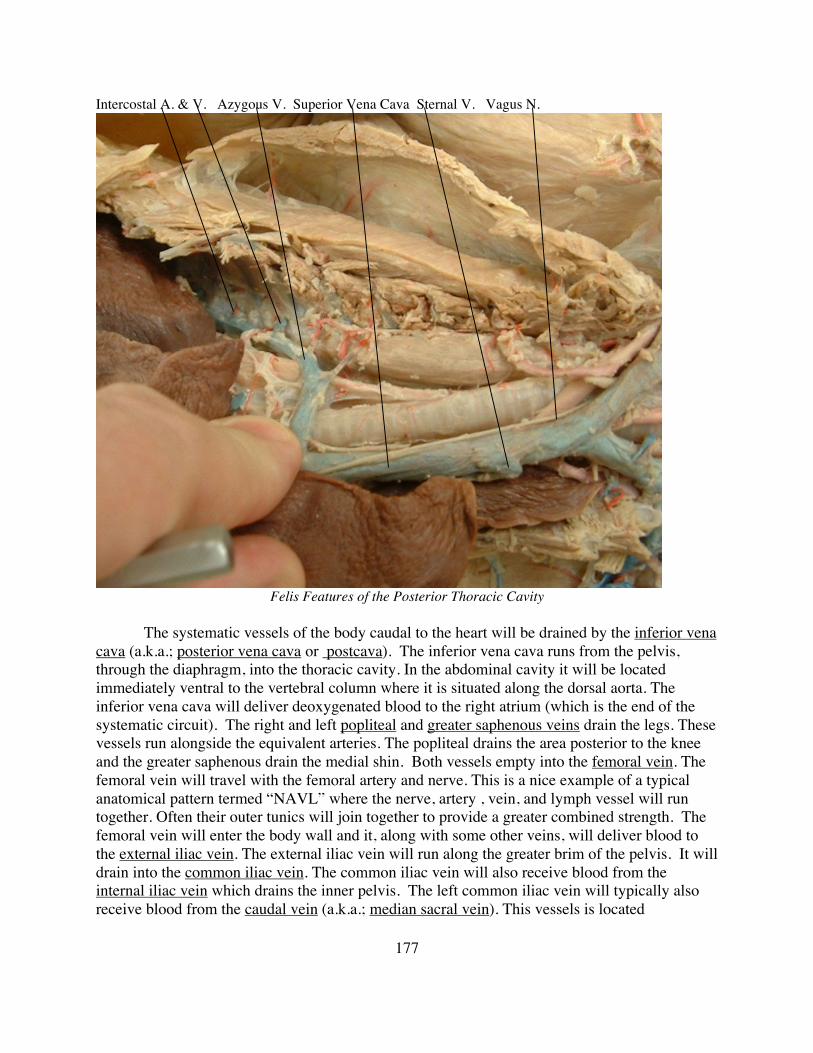
177
Intercostal A. & V. Azygous V. Superior Vena Cava Sternal V. Vagus N.
Felis Features of the Posterior Thoracic Cavity
The systematic vessels of the body caudal to the heart will be drained by the inferior vena
cava (a.k.a.; posterior vena cava or postcava). The inferior vena cava runs from the pelvis,
through the diaphragm, into the thoracic cavity. In the abdominal cavity it will be located
immediately ventral to the vertebral column where it is situated along the dorsal aorta. The
inferior vena cava will deliver deoxygenated blood to the right atrium (which is the end of the
systematic circuit). The right and left popliteal and greater saphenous veins drain the legs. These
vessels run alongside the equivalent arteries. The popliteal drains the area posterior to the knee
and the greater saphenous drain the medial shin. Both vessels empty into the femoral vein. The
femoral vein will travel with the femoral artery and nerve. This is a nice example of a typical
anatomical pattern termed “NAVL” where the nerve, artery , vein, and lymph vessel will run
together. Often their outer tunics will join together to provide a greater combined strength. The
femoral vein will enter the body wall and it, along with some other veins, will deliver blood to
the external iliac vein. The external iliac vein will run along the greater brim of the pelvis. It will
drain into the common iliac vein. The common iliac vein will also receive blood from the
internal iliac vein which drains the inner pelvis. The left common iliac vein will typically also
receive blood from the caudal vein (a.k.a.; median sacral vein). This vessels is located

178
Felis Pelvic and Lower Abdominal Blood Vessels
immediately ventral to the sacrum and will drain the tail. The right and left common iliacs will
join to form the caudal extent of the inferior vena cava. Normally this happens near the greater
pelvic brim but occasionally a specimen will appear to have the common iliacs joining near the
kidneys. In reality this is a case where the inferior vena cava has split and exists as a paired
structure until the two halves anastomize together near the renal veins. As the inferior vena cava
travels cranially through the abdominopelvic cavity it will receive blood from numerous small
lumbar veins. The first major vessels to deliver blood to the inferior vena cava are the iliolumbar
veins. These vessels, like the iliolumbar arteries, are located near the iliac crest. The next vein
to drain into the inferior vena cava is the right gonadal/genital vein. In females this would be the
ovarian and in males it would be the testicular/internal spermatic. The left gonadal vein delivers
blood into the left renal vein. This asymmetry is found in many species of placental mammal.
Notice the size of the ovarian veins when you study your specimen. The size can be correlated
with the female’s reproductive state at time of death. The next large veins to deliver blood into
the inferior vena cava are the right and left renal veins. Sometimes you will be able to observe
branches of the renal veins, called segmental veins, exiting at the hilus of the kidney. Cranial to
the renal veins will be the adrenolumbar veins. These vessels drain some of the lumbar
musculature and the adrenal glands. The adrenolumbar veins are the most cranial of the
abdominal systematic branches that you will be responsible for.

179
Felis Hepatic Portal Veins 1
The hepatic portal system is a series of veins draining the capillary beds of the digestive
tract and delivering this nutrient-rich blood to the liver for processing. The hepatic portal vein is
a large, but short, vessel that delivers blood into the liver. It can be seen on the posterior right
side of the stomach near the lesser curvature of the stomach. The hepatic portal vein receives
blood from the superior mesenteric vein. The superior mesenteric vein is a large, long vessel that
receives blood from the pancreas, small intestine, and large intestine. It will run near, but not
quite parallel to, the superior mesenteric artery. One small vessel that drains into the cranial
portion of this vein is the inferior or posterior pancreaticoduodenal vein. This vein can be
observed traveling from the duodenum through the dorsal lobe of the pancreas. The caudal
portion of the superior mesenteric will fan out into a number of veins that will empty into it.
These vessels include the numerous intestinal veins that drain capillary beds of the small
intestine. As they travel towards the superior mesenteric they will join together to form several
mesenteric veins. The mesenteric veins will deliver blood into the superior mesenteric. The
superior mesenteric will also receive blood from the ileocolic and inferior mesenteric veins. The
ileocolic will arise from the ileocolic junction. The inferior mesenteric will drain the remainder
of the large intestine and will receive blood from the middle colic and right colic veins. The

180
nutrient-rich blood will enter the sinusoids and will be processed by the liver. The resultant
blood, which will also be low in oxygen, will enter systemic circulation by draining into the
hepatic vein. The hepatic vein delivers the blood into the inferior vena cava immediately below
the diaphragm.
Superior Mesenteric V. Ileocolic A.& V. Right Colic A. Middle Colic A. Inferior Mesenteric. V. & A.
Felis Hepatic Portal Veins 1


















