
Proceedings Vol 2 - APWSS - Asian-Pacific Weed Science Society

Pacific Science, vol. 68, no. 1 July 16, 2013 (Early view)
Climate change and weed impacts on small
island ecosystems: Lantana camara L. (Magnoliopsida: Verbenaceae) distribution in
Fiji
By: Subhashni Taylor* and Lalit Kumar Abstract Oceanic island ecosystems are particularly vulnerable to invasion by exotic species and this vulnerability may be intensified by the effects of climate change, particularly if favourable climate conditions allow exotic invaders to spread to new areas. Effective management of such species requires knowledge of their potential distribution under current and future climate. This research examines the value of species distribution modelling in invasive species management in small island ecosystems using the specific case of Lantana camara L. invasion in Fiji. A niche model of L. camara was developed using CLIMEX species distribution modelling (SDM) software. Several sources of data were used to develop the model including phenological observations and geographic distribution records. The model was used to estimate its potential distribution under historical climate. The CSIRO-Mk3.0 Global Climate Model (GCM) was used to explore the impacts of climate change on its potential future distribution. The model was run with the A1B and A2 scenarios for 2030 and 2070. Large parts of both the major islands of Fiji, Viti Levu and Vanua Levu, were shown to have high climatic suitability for L. camara. However, under future climate scenarios, the climatic suitability for L. camara in Fiji was projected to contract. The results illustrate that SDMs can play a useful role in the formulation of cost-effective invasive species management strategies and the resulting species distribution maps have broad applicability throughout the many islands of the South Pacific region. *Corresponding Author E-mail: [email protected]

2
Introduction
The Pacific Islands region has been designated as one of the 25 biodiversity "hot spots" for
conservation priority in the world, and the flora of the Pacific Islands has been estimated to
comprise over 3300 endemic plant species that are unique to the region (Myers et al. 2000). The
combined threats of climate change and invasive species puts them at risk of extinction (Meyer
2000, Myers et al. 2000). Invasive species have many negative impacts on the native flora and
communities of islands and the added impacts of climate change can further exacerbate such
threats, particularly if favourable climate conditions allow invasives to expand their distribution.
Therefore, it is important to protect and maintain this rich but fragile natural heritage from the
impacts of invasive species.
A review of invasive species in the Pacific region (Sherley et al. 2000) has outlined a
number of issues that are barriers to effective management of such species in the region. Some
problems that are highlighted in the report include: (1) shortage and inaccessibility of scientific
information on basic biology, including distribution, for risk assessment and effective
management; (2) inadequate quarantine and risk assessment facilities; (3) lack of awareness of
the impacts of invasive species on biodiversity; and (4) threats to biodiversity are not yet well
understood.
Biosecurity agencies require information on the potential distribution and relative
abundance of invasive species under current and future climate scenarios if they are to carry out
risk assessments and formulate effective long term management strategies (Kriticos et al. 2011a).
Species distribution models (SDMs), bioclimatic models, and ecological niche models (ENMs)
are valuable tools that can be used in such instances (Guisan and Zimmerman 2000). A species’
preferred climate is inferred from its distribution data, commonly termed the ‘environmental
envelope approach’ (Barry and Elith 2006). The major assumption behind such models is that

3
climate is the primary factor defining the potential range of plants and other poikilotherms
(Woodward 1987). A range of software has been developed which can be used to model species’
current and future distributions (Peterson et al. 2011). CLIMEX (Hearne Scientific Software
2007, Sutherst et al. 2007) is one such software that has been frequently used in invasive species
distribution modelling (Poutsma et al. 2008, Sutherst and Bourne 2009, Chejara et al. 2010,
Kriticos et al. 2011a). A range of information types, such as experimental observations of a
species’ growth response to temperature and soil moisture, current distribution and seasonal
phenology data, can be used within CLIMEX to model potential distributions of species. In a
comparison of CLIMEX with two correlative modelling methods (MaxEnt and Boosted
Regression Trees), Webber et al. (2011) found that CLIMEX was better placed to project a
species’ distribution in a novel climate such as a new continent, or under a future climate
scenario. Furthermore, a review of the various climate-based packages intended for potential
species distribution assessment found that ‘CLIMEX was the most suitable climate modelling
package for undertaking Weed Risk Assessments because it can support model-fitting to a global
plant distribution, includes a climate change scenario mechanism, and provides an insight into
the plant’s ecological response to climate’ (Kriticos and Randall 2001).
In this paper, we examine the utility of bioclimatic models in invasive species
management in small island ecosystems. Limited research has been carried out in this respect,
particularly in the context of the islands of the South Pacific (however, see Christenhusz and
Toivonen 2008, Rödder and Lötters 2010). We ask the following question: Can species
distribution modelling be used to deal with some of the issues that are seen as barriers to
effective management of invasive species in the South Pacific, specifically the issues related to
lack of information on invasive species distribution?

4
We answer this question by developing a model of the climate responses of the highly
noxious weed, Lantana camara L. based on its native and invasive distribution using the
CLIMEX software package. We then use this model to examine its potential current distribution
and future distribution under climate change on the island of Fiji in the South Pacific.
Materials and Methods
Study Area
The Fijian Islands archipelago is located in the Pacific Ocean between longitudes 174º E and
178º W and latitudes 12º to 22º S. It comprises approximately 330 islands of which one third are
inhabited. The archipelago covers about 1.3 million km2 of the South Pacific Ocean with the
total land area being 18,333 km2. Viti Levu and Vanua Levu are the two main islands, making up
15,985 km2 of the land area (Figure 1). The total population is just over 800,000 with most
people living on the two main islands (Fiji Islands Bureau of Statistics 2011).
Figure 1 near here
Lantana camara L.
Lantana camara L. (hereafter called Lantana) has been ranked among one of the world’s 100
worst invasive alien species (Lowe et al. 2000). Its native range is Central and northern South
America and the Caribbean (Day et al. 2003). However, it is now a major weed in many tropical
and subtropical countries with its global distribution encompassing approximately 60 countries
or island groups between 35º N and 35º S (Day et al. 2003). Lantana was introduced to the
Pacific islands after European contact and since then has rapidly naturalized and become a
noxious weed in this region (Figure 1). It has been reported on many islands such as Hawaii,
Cook Islands, Fiji, Samoa, Vanuatu, New Caledonia and Solomon Islands (Thaman 1974,
Waterhouse 1997, Day et al. 2003, Global Invasive Species Database 2012) and is considered

5
among the most significant invasive taxa found on the majority of the islands in the region
(Meyer 2000). Lantana forms dense stands that exclude native seedlings by inhibiting their
germination, seedling growth and survival through competition (Sharma et al. 2005).
Furthermore, a single plant can produce up to 12 000 seeds per year which are dispersed over
long distances by birds and mammals (Stock et al. 2009). The ability to spread through layering
also makes it a successful invader (Day et al. 2003, Stock et al. 2009). These characteristics,
combined with a favourable climate, an abundance of open habitats and a relative absence of
natural competition, have allowed Lantana to become naturalized in many of the Pacific island
groups (Thaman 1974). It can potentially cause a reduction in biodiversity and increased
susceptibility to fire intrusion (Berry et al. 2011). Other potential impacts include decline in soil
fertility and an alteration of ecosystem processes (Day et al. 2003). Lantana has been classified
as a “dominant invader” and one of the most important threats to biodiversity in many Pacific
island nations due to its widespread occurrence and its ability to form dense stands thereby
severely impacting native biota (Meyer 2000).
Distribution of Lantana in Fiji
The distribution of Lantana in Fiji was ascertained from a variety of sources, including Smith
(1991), Day and Neser (2000), A. Macanawai (Principal Research Officer, Plant Protection
Section, Department of Agriculture, Koronivia Research Station, Nausori, Fiji, personal
communication) and A. Tye (Former Invasive Species Adviser, Secretariat of the Pacific
Regional Environment Programme). In Fiji, Lantana has been reported along road sides in most
provinces of Viti Levu and in some coconut plantations on Vanua Levu and Taveuni (A.
Macanawai, personal communication). It has also been reported as a garden escapee and being
widely distributed in Fiji in cultivated areas, particularly on the drier sides of the large islands
(A. Tye, personal communication) as well as in the cooler and wetter eastern and mountainous

6
regions (Day and Neser 2000). On Viti Levu, Lantana was reported in Lautoka, Nadarivatu,
Yaqara, Navua, Colo-i-Suva and Lami while in Vanua Levu it was found in Labasa and
Savusavu as well as on Taveuni Island (Smith 1991). Smith’s (1991) observations were made
over 20 years ago and therefore will need to be interpreted with caution although the personal
communication data is more recent.
CLIMEX Software
A model of the potential distribution of Lantana under current and future climate scenarios was
developed using CLIMEX software (Hearne Scientific Software 2007, Sutherst et al. 2007). An
eco-physiological growth model forms the basis of this software where it is assumed that a
population experiences a favourable season with positive growth and an unfavourable season
with negative population growth. Parameters that describe a species’ response to climate can be
inferred from its geographic range and phenological observations (Sutherst et al. 2007). The user
can then apply these parameters to novel climates to project the potential range in new regions or
climate scenarios (Kriticos et al. 2011a). An annual growth index (GIA) describes the potential
for population growth during favourable climate conditions. The GIA is determined from the
temperature index (TI) and moisture index (MI) which represent the species’ temperature and
moisture requirements for growth. The probability that the population can survive unfavourable
conditions is described by four stress indices (cold, wet, hot and dry) and up to four interaction
stresses (hot–dry, hot–wet, cold–dry and cold–wet). The growth and stress indices are calculated
weekly and combined into an overall annual index of climatic suitability, the Ecoclimatic index
(EI) which is theoretically scaled from 0 to 100. An EI value of zero indicates that the species
will not be able to survive at that location, 1–10 indicate marginal habitats, 10–20 can support
substantial populations while EI values >20 are highly favourable (Sutherst et al. 2007). For a
detailed description of parameters, see Sutherst and Maywald (1985). In this study, parameters

7
describing Lantana’s climatic preferences were fitted using the methodology described in
Sutherst and Maywald (1985), Chejara et al (2010) and Kriticos et al (2011a).
Meteorological Data and Climate Change Scenarios
The modelling component of this study employed the CliMond 10´ gridded climate data
(Kriticos et al. 2011b). Five variables, average minimum monthly temperature (Tmin), average
maximum monthly temperature (Tmax), average monthly precipitation (Ptotal) and relative
humidity at 09:00 h (RH09:00) and 15:00 h (RH15:00), were used to represent historical climate
(averaging period 1950–2000). The same five variables were used to depict potential future
climate in 2030 and 2070, based on the CSIRO-Mk3.0 Global Climate Model (GCM) (Gordon et
al. 2002) with the A1B and A2 SRES scenarios (IPCC 2000). The A1B scenario describes a
world with a balanced use of fossil and non-fossil resources while A2 describes a varied world
with high population growth but slow economic development and technological change. The B
family of scenarios were not assessed here because of the findings of Rahmstorf et al (2007) that
recent global temperature increases were much higher than the hottest IPCC scenarios. The
CSIRO-Mk3.0 GCM was selected for this study because it performed well compared to other
GCMs at representing basic aspects of the observed climate at a regional scale (Hennessy and
Colman 2007, Kriticos et al. 2011b).
Fitting CLIMEX Parameters
Information on the global distribution of Lantana was downloaded from the Global Biodiversity
Information Facility (GBIF). The GBIF is a database of natural history collections around the
world for various species which is available for download. A total of 4126 records were
downloaded of which only 1740 records were used in parameter fitting because many records did
not have geolocations or were repetitions and were subsequently discarded. Further data on
Lantana’s exotic distribution in South Africa (SAPIA 2006) and Asia (Biswas 1934, Jafri 1974,

8
Thakur 1992, Chen and Gilbert 1994, Press et al. 2000) were used for fitting stress parameters.
Both native and exotic distribution data were used for model parameterization to ensure that the
complete range of environmental conditions in which lantana may occur was covered. Growth
parameters were fitted using seasonal phenology data from the southern states of Brazil (Winder,
1980, 1982). Although Winder’s study was restricted to Lantana tiliaefolia and L. glutinosa,
these data were used in parameter fitting based on the ecological similarity of these two species
to the weedy taxa of Lantana. Each parameter was iteratively adjusted until a reasonable match
was seen between the potential and known distribution of Lantana in these areas. The intention
was to get maximum EI values near known vigorous populations and to minimize EI values
outside the recorded distribution of Lantana. The parameters were checked to ensure that they
were biologically reasonable (Table 1). For a detailed explanation of the parameter-fitting
procedure and model validation, refer to Taylor et al. (2012) and Taylor and Kumar (2012). The
fitted parameters were then used to project lantana’s potential distribution in Fiji under historical
climate (averaging period 1950–2000) and climate change scenarios.
Results
Historical Climate
Figure 2 shows the projected distribution in the Pacific region for the islands that could be shown
at this scale. These were Vanuatu, New Caledonia and Solomon Islands. The presence data
shown in Figure 1 accords well with the projected distribution on these three islands in Figure 2.
The observed distribution of Lantana in Fiji is shown in Figure 3 while the potential distribution
under historical climate is shown in Figure 4. The current distribution shows a good match with
the modelled distribution. Most of Vanua Levu,Viti Levu and Taveuni are shown to be highly
suitable or suitable for Lantana.

9
Future Climate
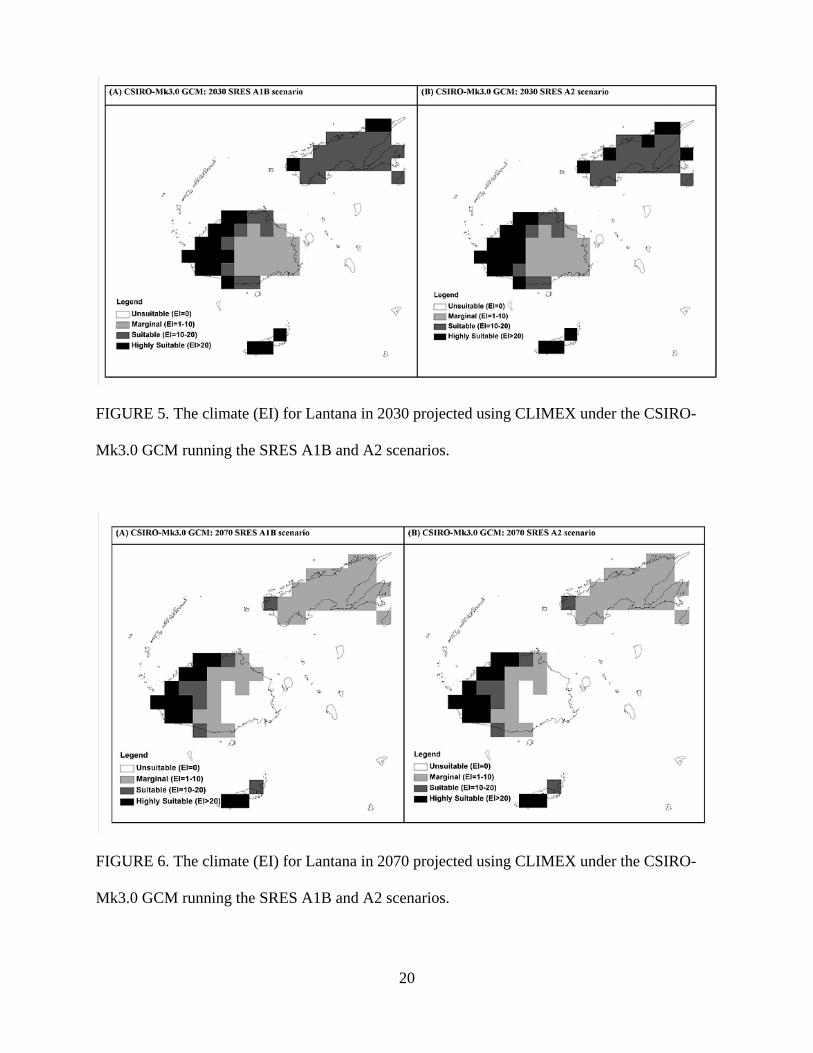
Potential distribution under future climate scenarios using the CSIRO-Mk3.0 GCM are shown in
Figures 5 and 6. A contraction in suitable climate areas was seen for this climate change model,
particularly throughout the island of Vanua Levu but also on the eastern half of Viti Levu
(Figures 5 and 6) with this trend even clearer in the 2070 scenario. The two emission scenarios
showed very similar results for both time periods.
The changes in wet stress between current climate and 2070 was also mapped (Figure 7)
as this was the only stress that appeared to be impacted by climate change. Increasing levels of
wet stress is seen on the eastern side of Viti Levu and throughout Vanua Levu.
Table 1 near here
Figures 2, 3, 4, 5, 6 and 7 near here
Discussion
The utility of species distribution modelling was examined as a tool for the purposes of
informing the formulation of long term management strategies for invasive species on small
island ecosystems. This was done using the specific case of Lantana invasion in Fiji. The results
show that under current climate, large parts of Fiji are conducive to Lantana growth and spread
(Figure 4) and this accords well with the observed distribution (Figure 3). The likelihood of
further spread of this highly noxious weed from infested to uninfested areas is possible given the
rapid increase in travel and transportation. Furthermore, habitat disturbance caused by activities
such as logging, land clearing for agriculture and introduction of large mammals such as cattle
and sheep provide further suitable habitats for Lantana invasion (Thaman 1974, Day et al. 2003).
In particular, containment efforts around the three southern provinces on Viti Levu where

10
Lantana currently is not known to occur (Figure 3) and where climatic suitability is shown to be
marginal (Figure 4) may be effective in preventing spread to these uninvaded regions.
An important aspect of invasive species management is to educate the citizens and to
raise public awareness of the impacts of such species, not only on agriculture but also on native
biodiversity. The maps of potential Lantana distribution produced in this study can be a useful
tool in public awareness campaigns. Furthermore, maps of potential distribution resulting from
such modeling exercises may also be valuable for other aggressive invaders that have been
identified in many South Pacific countries such as African tulip tree (Spathodea campanulata),
guava (Psidium guajava), giant sensitive plant (Mimosa diplotricha), mile-a-minute (Mikania
micrantha) and elephant grass (Pennisetum purpureum) amongst others (Sherley et al. 2000).
They can be employed to enlist the help of local communities in the management of existing
infestations and the prevention of further invasion, particularly through transfer from invaded
areas to weed free areas. Potential distribution maps can also inform coordination and
collaboration efforts on management so that areas of high conservation status can be targeted for
weed control.
The overall impact of climate change upon the distribution of Lantana in Fiji will be a
decrease in its range (Figures 5 and 6). The climate change modelling indicates that all of Vanua
Levu and western Viti Levu will remain climatically suitable for Lantana invasion until 2030.
However, climatic suitability decreases in 2070 (Figure 6) with only the west coast of Viti Levu
and the island of Taveuni remaining at risk of Lantana invasion. The major reason for this
decrease in suitable climate is due to increasing wet stress (Figure 7) caused by increased soil
moisture levels. Wet stress accumulates if the soil moisture level exceeds the wet stress threshold
(SMWS). This threshold value is set at greater than or equal to the limiting high soil moisture
value (SM3). Although Lantana is often found in tropical areas receiving 3000mm of rainfall per

11
year, it can successfully survive such high levels of rainfall only if the soil is well drained (Day
et al. 2003) since it has a tendency to rot in boggy or hydromorphic soils (Hamilton 1919,
Thaman 1974). The modelling results show that climate change has little impact on the
distribution of the other stresses i. e. cold, heat and dry stresses. Furthermore, the long term
reduction in climatic suitability in 2070 is a useful finding because it identifies areas that would
benefit from a concerted effort of weed control measures in the short term.
The results from this study have broader implications for plant invasions in the South
Pacific region, given the increasing concern about the threats posed by plant invaders to the
survival of the remaining native plants and habitats in island ecosystems (Mueller-Dombois and
Fosberg 1998, Denslow and Johnson 2006, Denslow et al. 2009). The collection of data on the
distribution of aggressive invaders under current and future climates is vital to the formulation of
prevention, education and management plans. Furthermore,valuable information on present and
future invasion risks that is gained from maps such as Figures 2 to 7 may encourage land
managers to consider the future implications of present-day management decisions. This is
particularly pertinent given that many invaders (for example, Lantana) were originally
introduced into new areas as garden ornamentals or pasture species which then turned out to be
major threats to native and agricultural systems.
Distribution maps such as the ones presented in this study can offer opportunities for the
formulation of cost-effective alternatives such as: (1) targeted elimination of outlying
populations of noxious weeds based on perceived future threats; (2) categorization of areas for
appropriate types of weed management as the distribution maps help to identify areas that need
to be prioritized for eradication and areas where containment would be more cost-effective; (3)
development of early-warning and response systems for potential invaders; (4) development of
strategic control initiatives, such as containment lines, designed to stop the proliferation of a

12
weed into uninfested areas; (5) formulation of long term plans to monitor possible incursions into
areas containing environmental assets of high value; and (6) making informed decisions about
the provision of resources for weed control, particularly in areas where climatic suitability is
expected to decrease in the future. Such modeling exercises are especially valuable in identifying
regions currently at risk and in the future from weed species that may not be present, or recently
arrived with limited distributions such as ‘sleeper’ weeds which may be currently present as
benign populations. Distribution maps for such species can be used to undertake a program of
surveillance and local eradication of possible sleeper populations.
Model Limitations and Future Research
CLIMEX is based on the response of a species to climate. Non-climatic factors such as dispersal
potential and biotic interactions are not explicitly included in the modelling process. Although
niche modelling is underpinned by Hutchinson’s (Hutchinson 1957) fundamental and realized
niche concepts, there are conflicting views on what the models actually represent (Araújo and
Guisan 2006). Some researchers consider that niche models provide an estimate to the species’
fundamental niche (Soberon and Peterson 2005) while others think that the models present a
‘‘spatial representation of the realized niche’’ (Guisan and Zimmerman 2000, Pearson and
Dawson 2003). Therefore, in assessing the risk of invasion, it is important to remember that there
may be discrepancies between the realized and fundamental niche of the invasive species being
modelled. Other factors such as competition and lack of dispersal opportunities may lead to an
underestimation of the threat posed by the invader in some areas (Rödder and Lötters 2010). The
issue of realized and fundamental niches is particularly relevant in the case of invaders where
biological control agents have been released although this is of concern mainly if the control
agents have established successfully and provided some measure of control. In the case of
Lantana, 36 species of biological control agents have been released in 32 countries (Julien and

13
Griffiths 1998) but it remains a serious problem in some parts of the world including Australia
and parts of the South Pacific (Day and Neser 2000). Consequently, the release of biocontrol
agents does not appear to have influenced the distribution and/or abundance of Lantana in these
areas. However, climate change will have implications for any new biocontrol agents that may be
released in the future since their distributions are also likely to change with climate change
(Kriticos et al. 2009). Hence, establishment of ongoing monitoring of current and future
biological control programs not only for Lantana but other invaders as well will be crucial so that
changes may be discovered in a timely manner and appropriate action taken.
A further cause of uncertainty in model projections is related to uncertainty in future
global greenhouse gas emission patterns (Kriticos et al. 2006). As such, the models based on
future climate scenarios provide an indication of the direction and magnitude of change that may
be expected in the future.
Although there have been rapid advances in the development of gridded datasets for
global climate normals (Hijmans et al. 2005, Mitchell and Jones 2005), these grids may be
inappropriate for small study regions (Mitchell and Jones 2005). Many of the small island states
of the South Pacific have small land areas with a limited number of meteorological stations.
Thus, there is a need for fine resolution climatic datasets developed specifically for these
countries as the currently available data are unlikely to reflect the climatic variation that can
occur over small distances on these mountainous islands. However, in the absence of such
datasets, this study has utilized the best available data to carry out the climate change modelling.
Furthermore, there is very limited spatial data available on endangered species and communities.
Such datasets would enhance the utility of species distribution models by identifying areas where
invaders and endangered species co-occur.

14
Acknowledgements
The National Climate Change Adaptation Research Facility (NCCARF) kindly provided a grant
that allowed the principal author to purchase the CLIMEX software as well as attend a training
workshop for CLIMEX. This work benefitted from a travel grant by the National Climate
Change Adaptation Research Facility (NCCARF) to S.T.
15
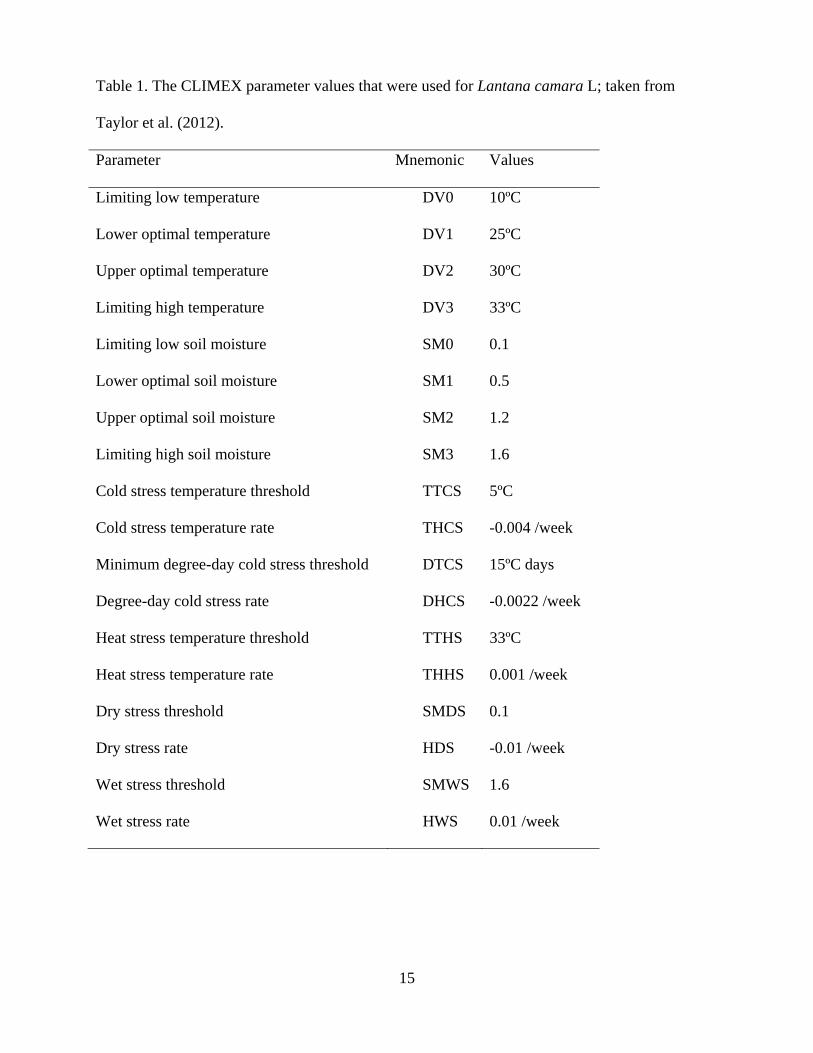
Table 1. The CLIMEX parameter values that were used for Lantana camara L; taken from
Taylor et al. (2012).
Parameter Mnemonic Values
Limiting low temperature DV0 10ºC
Lower optimal temperature DV1 25ºC
Upper optimal temperature DV2 30ºC
Limiting high temperature DV3 33ºC
Limiting low soil moisture SM0 0.1
Lower optimal soil moisture SM1 0.5
Upper optimal soil moisture SM2 1.2
Limiting high soil moisture SM3 1.6
Cold stress temperature threshold TTCS 5ºC
Cold stress temperature rate THCS -0.004 /week
Minimum degree-day cold stress threshold DTCS 15ºC days
Degree-day cold stress rate DHCS -0.0022 /week
Heat stress temperature threshold TTHS 33ºC
Heat stress temperature rate THHS 0.001 /week
Dry stress threshold SMDS 0.1
Dry stress rate HDS -0.01 /week
Wet stress threshold SMWS 1.6
Wet stress rate HWS 0.01 /week

16
FIGURE 1. Island nations of the South Pacific Ocean where Lantana is present (P) and is one of
the top ten weeds (marked with an asterisk). Presence and importance data is adapted from
Thaman (1974), Holm et al. (1991) and Waterhouse (1997) (Inset shows the major islands in
Fiji).

17
FIGURE 2. The climate (EI) for Lantana in Vanuatu, New Caledonia and Solomon Islands based
on CLIMEX under historical climate (averaging period 1950-2000).

18
FIGURE 3. The current observed distribution of Lantana in Fiji.

19
FIGURE 4. The climate (EI) for Lantana in Fiji based on CLIMEX under historical climate
(averaging period 1950-2000).
20
FIGURE 5. The climate (EI) for Lantana in 2030 projected using CLIMEX under the CSIRO-
Mk3.0 GCM running the SRES A1B and A2 scenarios.
FIGURE 6. The climate (EI) for Lantana in 2070 projected using CLIMEX under the CSIRO-
Mk3.0 GCM running the SRES A1B and A2 scenarios.

21
FIGURE 7. The changes in wet stress from historical climate to 2070 projected using the SRES
A1B and A2 scenarios.
Literature cited
Araújo, M. B. and A. Guisan. 2006. Five (or so) challenges for species distribution modelling. J.
Biogeogr. 33:1677–1688.
Barry, S. and J. Elith. 2006. Error and uncertainty in habitat models. J. Appl. Ecol. 43:413–423.
Berry, Z. C., K. Wevill and T. J. Curran. 2011. The invasive weed Lantana camara increases fire
risk in dry rainforest by altering fuel beds. Weed Res. 51:525–533.
Biswas, K. 1934. Some foreign weeds and their distribution in India and Burma. Indian Forester.
60:861–865.
Brooks, T. M., R. A. Mittermeier, C. G. Mittermeier, G. A. B. Da Fonseca, A. B. Rylands, W. R.
Konstant, P. Flick, J. Pilgrim, S. Oldfield, G. Magin, and C. Hilton-Taylor. 2002. Habitat
Loss and Extinction in the Hotspots of Biodiversity. Conserv. Biol. 16:909–923.
Chejara, V. K., D. J. Kriticos, P. Kristiansen, B. M. Sindel, R. D. B. Whalley, and C. Nadolny.
2010. The current and future potential geographical distribution of Hyparrhenia hirta.
Weed Res. 50:174–184.

22
Chen, S. and M. G. Gilbert. 1994. Flora of China. Available: http://www.efloras. org. [Accessed
15 September 2010].
Christenhusz, M. and T. Toivonen. 2008. Giants invading the tropics: the oriental vessel fern,
Angiopteris evecta (Marattiaceae). Biol. Invasions. 10:1215–1228.
Danin, A. 2000. The inclusion of adventive plants in the second edition of Flora Palaestina.
Willdenowia. 30:305–314.
Day, M. D. and S. Neser. 2000. Factors Influencing the Biological Control of Lantana camara in
Australia and South Africa. Pages 897-908 in N. R. Spencer, ed. Proceedings of the X
International Symposium on Biological Control of Weeds. Montana State University,
Montana.
Day, M. D., C. J. Wiley, J. Playford, and M. P. Zalucki. 2003. Lantana: Current management
status and future prospects. ACIAR Monograph. Canberra: Australian Centre for
International Agricultural Research.
Denslow, J. S. and M. T. Johnson. 2006. Biological Control of Tropical Weeds: Research
Opportunities in Plant–Herbivore Interactions. Biotropica. 38:139–142.
Denslow, J. S., J. C. Space, and P. A. Thomas. 2009. Invasive Exotic Plants in the Tropical
Pacific Islands: Patterns of Diversity. Biotropica. 41:162–170.
Fiji Islands Bureau Of Statistics. 2011. Fiji Facts and Figures [Online]. Suva: Fiji Islands Bureau
of Statistics. Available: http://www.statsfiji.gov.fj/Releases/FFF2010.pdf [Accessed 16
July 2012].
Garibaldi, A., P. Pensa, A. Minuto, and M. L. Gullino. 2008. First report of Sclerotinia
sclerotiorum on Lantana camara in Italy. Plant Dis. 92:1369.
Global Biodiversity Information Facility. 2010. GBIF Data Portal. Available:
http://www.gbif.org/. [Accessed 15 September 2010].

23
Global Invasive Species Database. 2012. Lantana camara. Available:
http://www.issg.org/database/species/distribution.asp?si=56&fr=1&sts=&lang=EN
[Accessed 18 July 2012].
Gordon, H. B., L. D. Rotstayn, J. L. Mcgregor, M. R. Dix, E. A. Kowalczyk,S. P. O’Farrell, L. J.
Waterman, A. C. Hirst, S. G.Wilson, M. A. Collier, I. G. Watterson. and T. I. Elliott.
2002. The CSIRO Mk3 Climate System Model. CSIRO Atmospheric Research Technical
Paper No. 60. Aspendale: CSIRO Atmospheric Research.
Guisan, A. and N. E. Zimmerman. 2000. Predictive habitat distribution models in ecology. Ecol.
Model. 135:147–186.
Hamilton, A. A. 1919. An ecological study of the saltmarsh vegetation in the Port Jackson
district. Proc. Linn. Soc. NSW, 44:463–513.
Hearne Scientific Software. 2007. CLIMEX Software Version 3.0.2. Hearne Scientific Software
Pty Ltd, Melbourne.
Hennessy, K. J. and R. Colman. 2007. Global climate change projections. Pages 36-48 in K. B.
Pearce, P. N. Holper, M. Hopkins, W. J. Bouma, P. H. Whetton, K. J. Hennessy and S.
B. Power, eds. Climate change in Australia – Technical Report 2007. Melbourne:
CSIRO.
Hijmans, R. J., S. E. Cameron, J. L. Parra, P. G. Jones, and A. Jarvis. 2005. Very high resolution
interpolated climate surfaces for global land areas. Int. J. Climatol. 25:1965–1978.
Holm, L. G., D. L. Plucknett, J. V. Pancho, and J. P. Herberger. 1991. The World’s Worst
Weeds: Distribution and Biology. Krieger Publishing Company, Malabar.
Hulme, P. E. 2004. Invasions, islands and impacts: a Mediterranean perspective. Pages 337-361
in J. M. Fernandez-Palacios and C. Morici, eds. Island Ecology. Asociacion Espanola de
Ecologia Terrestre, Madrid.

24
Hulme, P. E. 2009. Trade, transport and trouble: managing invasive species pathways in an era
of globalization. J. Appl. Ecol. 46:10–18.
Hutchinson, G. E. 1957. Concluding remarks. Cold Spring Symposium on Quantitative Biology.
Yale University, New Haven, CT, USA.
IPCC. 2000. Special Report on Emissions Scenarios: A Special Report of Working Group III of
the Intergovernmental Panel on Climate Change. Cambridge, UK.
Jafri, S. 1974. Flora of Pakistan. Available: http://www.efloras.org. [Accessed 15 September
2010].
Julien, M. H. and M. W. Griffiths. 1998. Biological Control of Weeds: A World Catalogue of
Agents and their Target Weeds. CAB International, Wallingford.
Kriticos, D. J. and R. P. Randall. 2001. A comparison of systems to analyze potential weed
distributions. Pages 61-79 in R. H. Groves, F. D. Panetta and J. G. Virtue, eds. Weed
Risk Assessment. Collingwood: CSIRO Publishing.
Kriticos, D. J., N. S. Alexander, and S. Kolomeitz. 2006. Predicting the potential geographic
distribution of weeds in 2080. Pages 27-34 in C. Preston, J. H. Watts and N. D.
Crossman, eds.15th Australian Weeds Conference. Adelaide, Australia: Weed
Management Society of South Australia.
Kriticos, D. J., N. D. Crossman, N. Ota and J. K. Scott. 2009. Climate change and invasive plants
in South Australia. Canberra: CSIRO Climate Adaptation Flagship. 97 p.
Kriticos, D. J., M. S. Watt, K. J. B. Potter, L. K. Manning, N. S. Alexander, and N. Tallent-
Halsell. 2011a. Managing invasive weeds under climate change: considering the current
and potential future distribution of Buddleja davidii. Weed Res. 51:85–96.

25
Kriticos, D. J., B. L. Webber, A. Leriche, N. Ota, I. Macadam, J. Bathols, and J. K. Scott. 2011b.
CliMond: global high-resolution historical and future scenario climate surfaces for
bioclimatic modelling. Methods Ecol. Evol. doi: 10.1111/j.2041-210X.2011.00134.x
Lowe, S., M. Browne, S. Boudjelas, and M. De Poorter. 2000. 100 of the World’s Worst
Invasive Alien Species: A selection from the Global Invasive Species Database. World
Conservation Union (IUCN). Available: http://www. issg.org/booklet.pdf. [Accessed 21
September 2011].
Martín, J., P. Cardoso, M. Arechavaleta, P. Borges, B. Faria, C. Abreu, A. Aguiar, J Carvalho, A.
Costa, R. Cunha, F. Fernandes, R. Gabriel, R Jardim, C. Lobo, A. Martins, P Oliveira,P.
Rodrigues, L. Silva, D. Teixeira, I. Amorim, N. Homem, B. Martins, M. Martins, and E.
Mendonça. 2010. Using taxonomically unbiased criteria to prioritize resource allocation
for oceanic island species conservation. Biodiversity Conserv. 19:1659–1682.
Meyer, J.-Y. 2000. Preliminary review of the invasive plants in the Pacific Islands (SPREP
Member Countries). Pages 85-114 in G. Sherley ed. Invasive species in the Pacific: A
technical review and draft regional strategy. South Pacific Regional Environmental
Programme, Apia, Samoa.
Mitchell, T. D. and P. D. Jones. 2005. An improved method of constructing a database of
monthly climate observations and associated high-resolution grids. Int. J. Climatol.
5:693–712.
Mueller-Dombois, D. and F. R. Fosberg. 1998. Vegetation of the Tropical Pacific Islands.
Springer-Verlag, New York.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. Da Fonseca, and J. Kent. 2000.
Biodiversity hotspots for conservation priorities. Nature. 403:853–858.

26
Pearson, R. G. and T. P. Dawson. 2003. Predicting the impacts of climate change on the
distribution of species: Are bioclimate envelope models useful? Global Ecol. Biogeogr.
12:361–371.
Peterson, A.T., J. Soberon, R. G. Pearson, R. P. Anderson, E. Martinez-Meyer, M. Nakamura
and M. B. Araujo. 2011. Ecological Niches and geographic distributions. Princeton
University Press, Princeton.
Poutsma, J., A. J. M. Loomans, B. Aukema, and T. Heijerman. 2008. Predicting the potential
geographical distribution of the harlequin ladybird, Harmonia axyridis, using the
CLIMEX model. BioControl. 53:103–125.
Press, J. R., K. K. Shrestha, and D. A. Sutton. 2000. Annotated Checklist of the Flowering Plants
of Nepal. Available: http://www.efloras.org. [Accessed 15 September 2010].
Pyšek, P. and D. M. Richardson. 2006. The biogeography of naturalization in alien plants. J.
Biogeogr. 33:2040–2050.
Rahmstorf, S., A. Cazenave, J. A. Church, J. E. Hansen, R. F. Keeling, and D. E. Parker. 2007.
Recent climate observations compared to projections. Science. 316:709.
Rödder, D. and S. Lötters. 2010. Potential Distribution of the Alien Invasive Brown Tree Snake,
Boiga irregularis (Reptilia: Colubridae). Pac. Sci. 64:11–22.
South African Plant Invaders Atlas. (sapia) 2006. Weeds and invasive plants quarter degree
square list per species: available reports. Available: http://www.agis.agric.za/wip/
[Accessed 21 September 2010].
Sanders, R. W. 1987. Taxonomic significance of chromosome observations in Caribbean species
of Lantana (Verbenaceae). Am. J. Bot. 74:914–920.
Sax, D. F., S. D. Gaines, and J. H. Brown. 2002. Species Invasions Exceed Extinctions on
Islands Worldwide: A Comparative Study of Plants and Birds. Am. Nat. 160:766–783.

27
Sharma, G. P., A. S. Raghubanshi, and J. S. Singh. 2005. Lantana invasion: An overview. Weed
Biol. Manag. 5:157–165.
Sherley, G., S. Timmins, and S. Lowe. 2000. Draft Invasive Species Strategy for the Pacific
Islands Region. Pages 1-6 in G. Sherley, ed. Invasive species in the Pacific: a technical
review and draft regional strategy. Apia, Samoa: South Pacific Regional Environment
Programme.
Smith, A. C. 1991. Flora Vitiensis Nova: A new flora of Fiji. National Tropical Botanical
Garden, Lawai, Kauai, Hawaii.
Soberon, J. and A. T Peterson. 2005. Interpretation of models of fundamental ecological niches
and species’ distributional areas. Biodiv. Inform. 2:1–10.
Steadman, D. W. 2006. Extinction and Biogeography of Tropical Pacific Birds. University of
Chicago Press, Chicago.
Stock, D., K. Johnson, A. Clark, and E. Van Oosterhout. 2009. Lantana Best Practice Manual
and Decision Support Tool. Yeerongpilly: The State of Queensland, Department of
Employment, Economic Development and Innovation.
Sutherst, R. W. and G. Maywald. 1985. A computerized system for matching climates in
ecology. Agric., Ecosyst. Environ. 13:281–299.
Sutherst, R. W., G. Maywald, and D. J. Kriticos. 2007. CLIMEX Version 3: User's Guide.
Melbourne: Hearne Scientific Software Pty Ltd.
Sutherst, R. W and A. S. Bourne. 2009. Modelling non-equilibrium distributions of invasive
species: a tale of two modelling paradigms. Biol. Invasions. 11:1231–1237.
Taylor, S., L. Kumar, N. Reid, and D. J. Kriticos. 2012. Climate change and the potential
distribution of an invasive shrub, Lantana camara L. PLoS ONE. 7: e35565.
doi:10.1371/journal.pone.0035565

28
Taylor, S. and L. Kumar. 2012. Sensitivity analysis of CLIMEX parameters in modelling
potential distribution of Lantana camara L. PLoS ONE. 7: e40969.
doi:10.1371/journal.pone.0040969
Thakur, M. L. 1992. Lantana weed (Lantana camara var. aculeate Linn) and its possible
management through natural insect pests in India. Indian Forester. 118:466–488.
Thaman, R. 1974. Lantana camara: Its introduction, dispersal and impact on islands of the
tropical Pacific ocean. Micronesica. 10:17–39.
Waterhouse, D. F. 1997. The major invertebrate pests and weeds of agriculture and plantation
forestry in the southern and western Pacific. Canberra: Australian Centre for International
Agricultural Research.
Webber, B. L., C. J. Yates, D. C. Le Maitre, J. K. Scott, D. J. Kriticos, N. Ota, A. Mcneill, J. Le
Roux, and G. Midgley. 2011. Modelling horses for novel climate courses: insights from
projecting potential distributions of native and alien Australian acacias with correlative
and mechanistic models. Div. Dist. 17:978–1000.
Winder, J. A. 1980. Factors affecting the growth of Lantana in Brazil. Ph.D. diss., University of
Reading, UK.
Winder, J. A. 1982. The effects of natural enemies on the growth of Lantana in Brazil. Bull.
Entomol. Res. 27:599–616.
Woodward, F. I. 1987. Climate and Plant Distribution. Cambridge University Press, Cambridge.


















