
Circadian Robot Metamorphosis - Lunds universitet · Circadian Robot Metamorphosis ... dog can...
8
Circadian Robot Metamorphosis Anne C. van Rossum ⇤,⇤⇤ H. Jaap van den Herik ⇤⇤ ⇤ Almende B.V. Rotterdam, The Netherlands ⇤⇤ Tilburg University Tilburg center for Cognition and Communication The Netherlands Abstract Insects, fish, and amphibians undergo the in- triguing process of metamorphosis. Metamorphic processes are governed by gene regulatory net- works: complex networks of gene-protein inter- actions. Recent studies demonstrate the isolated use of artificial gene regulatory networks for (1) body development, (2) metamorphosis, or (3) epi- genetics. We will evolve a gene regulatory net- work by fitness uniform selection in such a way that a process of metamorphosis becomes cou- pled to a circadian rhythm of day and night. A fitness figure is established that defines the dif- ference in average body size as a distinguishing metric to enforce a metamorphic process. Our results show that a swarm of modular robots can be evolved that shows a pronounced difference in average body size during day versus night. Ad- ditional experiments show (1) robustness against virtual gene knockouts, (2) robustness against phase shifts, and (3) duty cycle adjustments of the circadian input. One of our conclusions is that light can provide epigenetic information for the process of metamorphosis. 1 Introduction Insects, fish, and amphibians undergo the intriguing pro- cess of metamorphosis, a process guided by information. This information is stored in the genome of an organism; however, additional information might just as well origi- nate from other sources. In some cases, the environment itself can contribute to the guidance of metamorphosis, in which case we speak of an epigenetic contribution. Modular robots are swarm robots with connectors (Kernbach et al., 2009). The connectors enable the in- dividual robots to aggregate to large robot organisms. In the FP7 project Replicator, cube-sized 10x10x10cm robots are developed with four connectors, one at each wind rose direction (see Fig. 1, left). The robot mod- ules can move on their own by two screw drives. In an assembled state, a robot organism in the form of a dog can perform locomotion by using robot modules as legs. The functionality which we will address in mod- ular robots is not locomotion, but open-ended develop- ment, more specifically, metamorphosis. Open-ended de- velopment deals with all body formation and reforma- tion processes after a robot organism has been formed (also homeostatic processes). Metamorphosis is a subset of open-ended development: it describes only transitions between different body forms which occur in this adult stage. Figure 1: Left: The first prototype of the Replica- tor/Symbrion robot. Right: A scenario of functional meta- morphosis. The dog-like organism is able to cross the bar on the floor. The snake is able to climb the obstacle. To reach the power outlet the organism has to morph from dog to snake. 1.1 Metamorphosis Metamorphosis of modular robots is intended to have a functional role. In Fig. 1 a scenario is sketched in which a dog-like robot morphs into a snake-like robot organism. The depicted scenario is very demanding on a cognitive level. In this case, metamorphosis is preceded by per- ception and classification of a power outlet. The process of metamorphosis is not separated from environmental input. Yet, the coupling with the environment has two distinct facets. First, metamorphosis is coupled to perceptual phenomena in the environment as is argued by (van Dijk et al., 2008) within embodied embedded cognition. In the usual case of sensory-motor learning, invariances in the perceptual input are registered in relation to the actuator commands that started to bring about those perceptual changes in the first place. In the 137 Johansson, B., !ahin, E. & Balkenius, C. (2010). Proceedings of the Tenth International Conference on Epigenetic Robotics: Modeling Cognitive Development in Robotic Systems. Lund University Cognitive Studies, 149.
Transcript of Circadian Robot Metamorphosis - Lunds universitet · Circadian Robot Metamorphosis ... dog can...

Circadian Robot Metamorphosis
Anne C. van Rossum⇤,⇤⇤ H. Jaap van den Herik⇤⇤
⇤Almende B.V.
Rotterdam, The Netherlands
⇤⇤Tilburg University
Tilburg center for Cognition and Communication
The Netherlands
Abstract
Insects, fish, and amphibians undergo the in-triguing process of metamorphosis. Metamorphicprocesses are governed by gene regulatory net-works: complex networks of gene-protein inter-actions. Recent studies demonstrate the isolateduse of artificial gene regulatory networks for (1)body development, (2) metamorphosis, or (3) epi-genetics. We will evolve a gene regulatory net-work by fitness uniform selection in such a waythat a process of metamorphosis becomes cou-pled to a circadian rhythm of day and night. Afitness figure is established that defines the dif-ference in average body size as a distinguishingmetric to enforce a metamorphic process. Ourresults show that a swarm of modular robots canbe evolved that shows a pronounced di�erence inaverage body size during day versus night. Ad-ditional experiments show (1) robustness againstvirtual gene knockouts, (2) robustness againstphase shifts, and (3) duty cycle adjustments ofthe circadian input. One of our conclusions isthat light can provide epigenetic information forthe process of metamorphosis.
1 Introduction
Insects, fish, and amphibians undergo the intriguing pro-cess of metamorphosis, a process guided by information.This information is stored in the genome of an organism;however, additional information might just as well origi-nate from other sources. In some cases, the environmentitself can contribute to the guidance of metamorphosis,in which case we speak of an epigenetic contribution.
Modular robots are swarm robots with connectors(Kernbach et al., 2009). The connectors enable the in-dividual robots to aggregate to large robot organisms.In the FP7 project Replicator, cube-sized 10x10x10cmrobots are developed with four connectors, one at eachwind rose direction (see Fig. 1, left). The robot mod-ules can move on their own by two screw drives. Inan assembled state, a robot organism in the form of adog can perform locomotion by using robot modules aslegs. The functionality which we will address in mod-
ular robots is not locomotion, but open-ended develop-ment, more specifically, metamorphosis. Open-ended de-velopment deals with all body formation and reforma-tion processes after a robot organism has been formed(also homeostatic processes). Metamorphosis is a subsetof open-ended development: it describes only transitionsbetween di�erent body forms which occur in this adultstage.
Figure 1: Left: The first prototype of the Replica-tor/Symbrion robot. Right: A scenario of functional meta-morphosis. The dog-like organism is able to cross the baron the floor. The snake is able to climb the obstacle. Toreach the power outlet the organism has to morph from dogto snake.
1.1 Metamorphosis
Metamorphosis of modular robots is intended to have afunctional role. In Fig. 1 a scenario is sketched in whicha dog-like robot morphs into a snake-like robot organism.The depicted scenario is very demanding on a cognitivelevel. In this case, metamorphosis is preceded by per-ception and classification of a power outlet. The processof metamorphosis is not separated from environmentalinput. Yet, the coupling with the environment has twodistinct facets.
First, metamorphosis is coupled to perceptualphenomena in the environment as is argued by(van Dijk et al., 2008) within embodied embeddedcognition. In the usual case of sensory-motor learning,invariances in the perceptual input are registered inrelation to the actuator commands that started to bringabout those perceptual changes in the first place. In the
137
Johansson, B., !ahin, E. & Balkenius, C. (2010). Proceedings of the Tenth International Conference on Epigenetic Robotics: Modeling Cognitive Development in Robotic Systems. Lund University Cognitive Studies, 149.

case of metamorphosis, morphing actions are coupledto sensing actions. Second, metamorphosis is coupledto a specific time scale. Periodic phenomena in theenvironment can be reflected faithfully within the robotorganism. For instance, entrainment1 to a circadianrhythm (Sauze and Neal, 2008) is used for sailing robotsto predict the amount of solar energy later in the day.
1.2 Epigenetics
The environment can be seen as in competition or collab-oration with the genome in providing information aboutthe metamorphic process. One of the founders of sys-tems biology (Waddington, 1942) coined the term epige-netics to denote specific additional interactions with theenvironment in a developmental process.
The circadian rhythm, i.e, the rhythm of day andnight, is a type of epigenetic input that proves to beof extreme importance in the kingdom of animals aswell as in the kingdom of plants. Amongst others, circa-dian rhythms have been implied in growth vigor in plants(Ni et al., 2008). Besides that, disruption of the circa-dian cycle might even be a factor in breast and prostatecancer (Zhu et al., 2006).
1.3 Gene regulatory networks
A gene regulatory network (GRN ) is an interacting net-work of genes and proteins. Genes express proteins,that in their turn, activate or silence genes (see sub-section 3.1). GRNs have been used in modular roboticsfor the co-evolution of robot bodies and robot brains(Bongard, 2002). Open-ended development – in whichthere is no static end result of the gene regulatory net-work, but in which it continues to operate into adult-hood — has been used to implement homeostatic abil-ities (Quick et al., 2003). It is not meant, here and by(Quick et al., 2003), to denote a monotonic increasinglevel of complexity due to a developmental process.
GRNs have also been used to implement a circa-dian cycle. An evolved clock has been implemented(Knabe et al., 2006), but this clock has not been usedfunctionally for, for instance, locomotion or behavioraladaptation. The approach is promising to implementmetamorphosis, because the authors showed (1) robust-ness with respect to shifts in phase, (2) limited shifts infrequency, and (3) noise conditions. Observe that circa-dian light is a straightforward epigenetic input function.Future work will need to include cognition to act uponthe perception of a complex object like a power outlet.2
1Entrainment: an endogenous rhythm that entrains to envi-ronmental stimuli. For the case of metamorphosis we only needcoupling with an exogenous rhythm.
2The output of perceptual modules will be supplied to the generegulatory network. From the viewpoint of the GRN — and froma research perspective — this input is not di�erent from directcircadian light. Further research is need to address coupling with
Our problem statement reads: How to evolve a circadianrhythm for robot metamorphosis using a gene regulatorynetwork?
1.4 Overview
Section 2 describes the background of epigenetic robotics.Section 3 describes the model: the gene regulatory net-work; the genetic encoding, and the coupling with theenvironment. Section 4 describes the performed experi-ments and results. Section 5 contains our conclusion.
2 Background
Let us consider epigenetic robotics in the light of previ-ous research directions (subsection 2.1) and in the wayinformation can be fed to an epigenetic system (subsec-tion 2.2).
2.1 Evo-devo, eco-devo, epigenetics
Epigenetics originally (see also subsection 1.2) denotedenvironmental influence during development. We do notaim here for experimental predictions in developmentalpsychology. Our (old-fashioned) interpretation of epige-netics is best explained visually, see Fig. 2.
genesG0
genesG1
genesG2
GRN{P,G0}
GRN{P,G1}
GRN{P,G2}
body{M,D0}
swarmS2
brain{F0}
selection�
environm.EIenv
IbrainIbody
Figure 2: Overview of three types of influence on the generegulatory network: Ibody, Ibrain, and Ienv.
Epigenetic robotics can be seen as a natural sequel totwo contemporary research directions. First, within evo-devo3 research a so-called “indirect encoding” method isused. It defines an intermediate level of transcription
several types of environmental information, but that falls outsidethe scope of this paper.
3Evo-devo stands for “evolutionary development”.
138

between the genotype and the phenotype in the form ofproteins. It can be seen as data decompression.4 Theevo-devo (Dusheck, 2002) approach is depicted in Fig. 2by the intermediate GRN block between the level of thegenotype (the genes) and the level of the phenotype (thebody and the brain). Second, the eco-devo5 researchdirection is concerned with the influence of the environ-ment on the developmental process. This is depictedin Fig. 2 by the inclusion of the input from the blockenvironment. Third, and topic of this paper, epigeneticrobotics extends the eco-devo approach to processes afterdevelopment. Genome as well as environment continueto influence the modular robot during its entire lifetime.
Let us consider Fig. 2 in more detail. The candidatesolutions for the evolutionary algorithm are the gene reg-ulatory networks. The same GRN operates on all robotmodules in a swarm. So, the term swarm Si is used to re-fer to a collection of individual robot modules plus robotorganisms that all have the same genome Gi. A swarmSi is defined as the set {{M,Di}, Fi} with {M,Di} asthe body and Fi as the brain. Di is the connection graphbetween the modules M . The evolutionary process oper-ates on a population of swarms. It does not just operateon a population of individuals.
2.2 Body, brain, and environment
Figure 2 illustrates that three kinds of informationalsources impinge on the gene regulatory network {P,Gi}.(The trivial information contained by the genes is left outin this enumeration.) The three types of sources are asfollows
1. The body {M,Di}. For instance, local injurysignals can bring about post-embryonic regenera-tion in hydra, planarians, fish, and amphibians(Alvarado and Tsonis, 2006). In metamorphosis thebody influences through Ibody the regulatory networkto bring about a new body shape. For instance, whena snake-like body form has been obtained, this factalone might induce a subsequent body change to adog-like body form (cf. Fig 1).
2. The brain Fi. It is the cognitive control mech-anism of the process of development or metamor-phosis. Of course the body does also influencethe brain, but in this case the reverse process ismeant. For instance, the disappearance of a maleindividual can induce a sexual transformation in fish(Bass and Groberb, 2000). The disappearance of themale is first processed by the female brain. A sec-ond example concerns the metamorphosis of reef-building corals. The larvae sense the environment
4The number of neurons (in the order of 1010) succeeds thenumber of genes (in the order of 105) (Gierer, 1988) so a develop-mental process is needed to get every neuron in place.
5Eco-devo abbreviates “ecological development”.
and their neurons interpret external clues by emittingneuropeptides, namely the family of GLWamides6
(Iwao et al., 2002).
3. The environment E. Epigenetics is concerned withthe direct influence Ienv of the environment on thegene regulatory network. Lacking a brain, plantsare natural candidates for epigenetics. For instance,phytochromes, a family of photoreceptors on plants,target five identified genes in Arabidopsis thaliana7.The transcript levels of those genes increase when theplant is under a dense canopy and promote shade-avoidance (Jiao et al., 2007).
3 Model
A gene regulatory network will be evolved that exhibits ametamorphic rhythm coupled to a circadian input. Thismodel consists of four components:
• Gene regulatory network (subsection 3.1). Genes andproteins interact with each other. Those interactionsneed to be described formally.
• Genetic encoding (subsection 3.2). A precise repre-sentation of a series of genes in an artificial genomeand the transcription method.
• Evolutionary algorithm (subsection 3.3). Detailsabout the character of natural selection and geneticoperators (cross-over and mutation).
• Coupling (subsection 3.4). A specification of the cou-pling of the GRN with GRNs on connected robotsand the environment.
3.1 Regulatory network
A metamorphic process can be described by a regula-tory network. A regulatory network is a tuple {P,G}.It consists of a set of regulatory entities (g0, . . . , gL) ⇥G, and a set of regulated elements, artificial proteins,(p0, . . . , pM ) ⇥ P with pi ⇥ R�0. The proteins are up-dated synchronously according to Eq. 1. We will use theshorthand pi to indicate p[t]i and pi for p[t+�t]i (withvarying �t).
U(⇧gij) : pi = ⇥(pi � ⌅+⇧
j⇥Ni
⇤ij) (1)
⇤ij =
�⌅⇤
⌅⇥
⇤ij if �ij < pj < ⇥ij
�⇤ij if ⇥ij < pj < �ij
0 else
(2)
6One of the first discovered neuropeptides involved in the pro-cess of metamorphosis was isolated from sea anemone and has thevery appropriate name Metamorphosin-A (Iwao et al., 2002).
7A 20-25 cm tall flowering plant with one of the smallest knownplant genomes, popular as model organism.
139

The regulatory update rule U for each entity ⌥gij de-scribes the ingoing edges from the vertices pj in neigh-bourhood Ni to vertex pi. To calculate the new quantityof pi a non-linearity ⇥ij is used. The edge ⌥gij is not ascalar, but a vector [�ij ,⇥ij , ⇤ij ], with �ij ,⇥ij , ⇤ij � R⇥0.The protein pj can either up-regulate or down-regulatepi. If �ij < ⇥ij the protein pi is up-regulated, if �ij > ⇥ij
it is down-regulated. The rate of regulation is defined by⇤ij . If the regulating entity does not fall in between thelimits defined by �ij and ⇥ij nothing happens. Thatis, the protein pi is only decayed with a rate defined by⇧. The proteins are capped by a sigmoid function, �(x)which enforces the protein quantity to be between ⌅min
and ⌅max.
3.2 Genetic encoding
Each update rule as defined in Eq. 1 and Eq. 2 is rep-resented by a gene gl. The genome G is a series ofcodons [c0, . . . , cN�1] with N the length of the genome.Each gene gl is transcribed by analog genetic encod-ing (Durr et al., 2006). A start codon is defined bymk : ci mod 10 = 0. The array of ci is iteratedtill an instance of mk is found. Then gl is transcribedby using the values directly subsequent to m. Heregl = [⌃(pi), ⌃(pj), ⌃(⇤ij), ⌃(�ij), ⌃(⇥ij)], with ⌃ a func-tion that normalizes the codon value to the number ofaccepted types of the corresponding field. For instance,the function ⌃(pi) normalizes to the number of proteintypes.
This type of encoding allows for multiple phenom-ena. (1) The number of genes depends on the num-ber of start codons, not on the (codon) length N of thegenome. There is a variable set of genes. (2) The knock-out of a gene gl can reveal an underlying start codonthat was previously transcribed as one of the fields of gl.This allows for an encoding in which the knockout of agene might reveal a gene that was previously not tran-scribed because it was overlapped before the knockout.Such genes are called phase-0 same-strand overlappinggenes (Sabath et al., 2008)8. (3) A new gene does notnecessarily destroy an old gene. A mutation in a non-transcribed or non-coding part of the genome can giverise to a start codon. This new gene can be tried out inparallel to all existing genes.
8There might actually be selective pressure against phase-0overlap (Kingsford et al., 2007). It is not argued here that theintroduction of start codons will necessarily lead to overlappinggenes. Only the possibility for overlap is o�ered. Moreover, phase-0 overlap is relatively rare for biological genes compared to phase-1and phase-2 overlap because phase-0 overlap tends to be longer.In our case, artificial genes have a fixed size: overlap is not deter-mined by a stop codon. Phase-0 overlap does not necessarily needto be rare using fixed-sized genes.
3.3 Evolutionary algorithm
The evolutionary selection method applied is fitness uni-form optimization (Hutter et al., 2006). Contrary topure elitist schemes, this evolutionary algorithm rewardssparsely populated fitness levels above densely popu-lated levels. Set fmin the minimum fitness level, fmax
the maximum fitness level in the population of swarmsS = {S0, . . . , SH�1}. Here Si is swarm i with its owngene regulatory network {P,G}. H is the number ofswarms. In the fitness optimization selection scheme arandom value fs is drawn from the interval |fmin, fmax|.Then the swarm Si � S with fitness nearest to fs iscopied to the new generation. The deletion scheme ran-domly removes a swarm from the old generation. Theselection plus deletion operation is iterated H times tillthe new generation obtains the same size H as the oldgeneration. Fitness uniform optimization seems not toreward for higher fitness values; however, this is decep-tive. High fitness levels are probably sparsely populatedand will be selected for indirectly. The method makes acontinuous e⇤ort to maintain diversity in the population.Within each fitness level there is genetic drift. The twogenetic operators cross-over and mutation are defined.A cross-over takes place with probability Pc = 0.7. Mu-tation probability is at a level of Pm = 0.1, which meansthat on average NPm random codons are mutated. Thebiggest drawback of uniform selection is that the numberof fitness levels (or bins) need to be limited compared tothe number of swarms. If the lowest set of fitness lev-els are all dropped in the same bin, the evolutionaryalgorithm has a hard time to bootstrap. A square rootfunction is used over the fitness values, which results insmaller bins at lower levels (see section 4).
3.4 Neighbor coupling
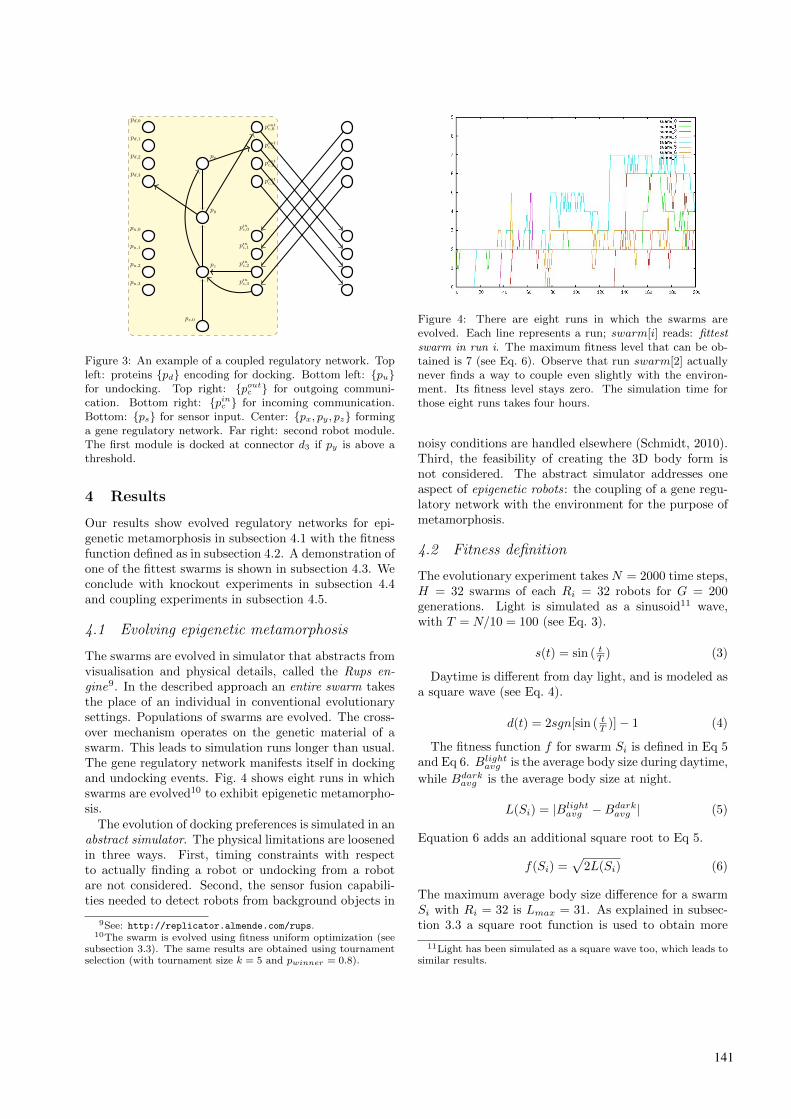
A set of the total number of proteins is used to couplethe regulatory network with the environment as well aswith its neighbors (Fig. 3).
There are five sets of proteins: {pd}, {pu}, {ps}, {pinc },and {poutc }, each of them a proper subset of {pi}. Thesets {pd} and {pu} have each four elements, correspond-ing to the number of connectors on a robot module.The set ps has one element, corresponding to a sen-sor input. The sets {pinc } and {poutc } are proteins usedfor communication. Communication is asymmetricallycross-coupled: protein poutc,k � {poutc } on module mA in-
crements pinc,l � {pinc } on module mB , with k = l thelocal index in each set. In our experiments we set:d = [0, . . . , 3], u = [4, . . . , 7], s = [17], cin = [10, . . . , 13],and cout = [14, . . . , 17].
140
px
py
pz
poutc,2
pd,2
poutc,3
poutc,1
poutc,0
pd,3
pd,1
pd,0
pinc,0pu,0
ps,0
pinc,1
pinc,2
pinc,3
pu,1
pu,2
pu,3
Figure 3: An example of a coupled regulatory network. Topleft: proteins {pd} encoding for docking. Bottom left: {pu}for undocking. Top right: {poutc } for outgoing communi-cation. Bottom right: {pinc } for incoming communication.Bottom: {ps} for sensor input. Center: {px, py, pz} forminga gene regulatory network. Far right: second robot module.The first module is docked at connector d3 if py is above athreshold.
4 Results
Our results show evolved regulatory networks for epi-genetic metamorphosis in subsection 4.1 with the fitnessfunction defined as in subsection 4.2. A demonstration ofone of the fittest swarms is shown in subsection 4.3. Weconclude with knockout experiments in subsection 4.4and coupling experiments in subsection 4.5.
4.1 Evolving epigenetic metamorphosis
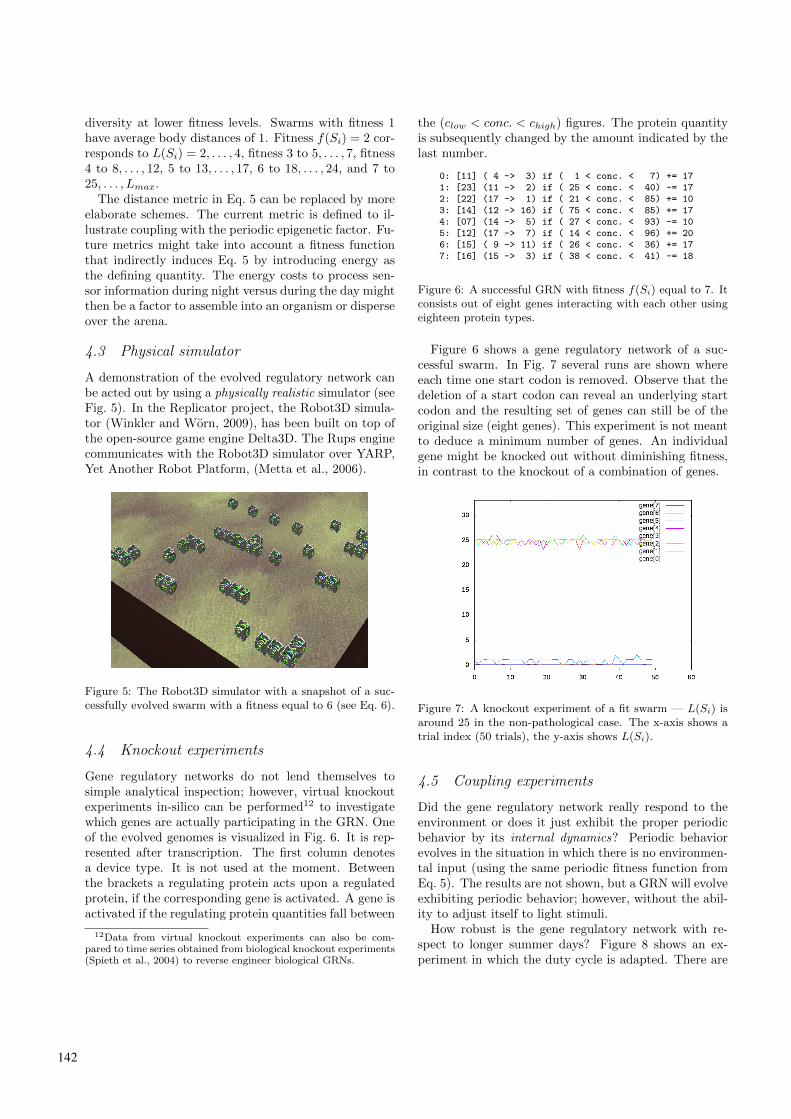
The swarms are evolved in simulator that abstracts fromvisualisation and physical details, called the Rups en-gine9. In the described approach an entire swarm takesthe place of an individual in conventional evolutionarysettings. Populations of swarms are evolved. The cross-over mechanism operates on the genetic material of aswarm. This leads to simulation runs longer than usual.The gene regulatory network manifests itself in dockingand undocking events. Fig. 4 shows eight runs in whichswarms are evolved10 to exhibit epigenetic metamorpho-sis.
The evolution of docking preferences is simulated in anabstract simulator. The physical limitations are loosenedin three ways. First, timing constraints with respectto actually finding a robot or undocking from a robotare not considered. Second, the sensor fusion capabili-ties needed to detect robots from background objects in
9See: http://replicator.almende.com/rups.10The swarm is evolved using fitness uniform optimization (see
subsection 3.3). The same results are obtained using tournamentselection (with tournament size k = 5 and pwinner = 0.8).
Figure 4: There are eight runs in which the swarms areevolved. Each line represents a run; swarm[i] reads: fittestswarm in run i. The maximum fitness level that can be ob-tained is 7 (see Eq. 6). Observe that run swarm[2] actuallynever finds a way to couple even slightly with the environ-ment. Its fitness level stays zero. The simulation time forthose eight runs takes four hours.
noisy conditions are handled elsewhere (Schmidt, 2010).Third, the feasibility of creating the 3D body form isnot considered. The abstract simulator addresses oneaspect of epigenetic robots : the coupling of a gene regu-latory network with the environment for the purpose ofmetamorphosis.
4.2 Fitness definition
The evolutionary experiment takes N = 2000 time steps,H = 32 swarms of each Ri = 32 robots for G = 200generations. Light is simulated as a sinusoid11 wave,with T = N/10 = 100 (see Eq. 3).
s(t) = sin ( tT ) (3)
Daytime is di�erent from day light, and is modeled asa square wave (see Eq. 4).
d(t) = 2sgn[sin ( tT )]� 1 (4)
The fitness function f for swarm Si is defined in Eq 5and Eq 6. Blight
avg is the average body size during daytime,
while Bdarkavg is the average body size at night.
L(Si) = |Blightavg �Bdark
avg | (5)
Equation 6 adds an additional square root to Eq 5.
f(Si) =�2L(Si) (6)
The maximum average body size di�erence for a swarmSi with Ri = 32 is Lmax = 31. As explained in subsec-tion 3.3 a square root function is used to obtain more
11Light has been simulated as a square wave too, which leads tosimilar results.
141
diversity at lower fitness levels. Swarms with fitness 1have average body distances of 1. Fitness f(Si) = 2 cor-responds to L(Si) = 2, . . . , 4, fitness 3 to 5, . . . , 7, fitness4 to 8, . . . , 12, 5 to 13, . . . , 17, 6 to 18, . . . , 24, and 7 to25, . . . , Lmax.
The distance metric in Eq. 5 can be replaced by moreelaborate schemes. The current metric is defined to il-lustrate coupling with the periodic epigenetic factor. Fu-ture metrics might take into account a fitness functionthat indirectly induces Eq. 5 by introducing energy asthe defining quantity. The energy costs to process sen-sor information during night versus during the day mightthen be a factor to assemble into an organism or disperseover the arena.
4.3 Physical simulator
A demonstration of the evolved regulatory network canbe acted out by using a physically realistic simulator (seeFig. 5). In the Replicator project, the Robot3D simula-tor (Winkler and Worn, 2009), has been built on top ofthe open-source game engine Delta3D. The Rups enginecommunicates with the Robot3D simulator over YARP,Yet Another Robot Platform, (Metta et al., 2006).
Figure 5: The Robot3D simulator with a snapshot of a suc-cessfully evolved swarm with a fitness equal to 6 (see Eq. 6).
4.4 Knockout experiments
Gene regulatory networks do not lend themselves tosimple analytical inspection; however, virtual knockoutexperiments in-silico can be performed12 to investigatewhich genes are actually participating in the GRN. Oneof the evolved genomes is visualized in Fig. 6. It is rep-resented after transcription. The first column denotesa device type. It is not used at the moment. Betweenthe brackets a regulating protein acts upon a regulatedprotein, if the corresponding gene is activated. A gene isactivated if the regulating protein quantities fall between
12Data from virtual knockout experiments can also be com-pared to time series obtained from biological knockout experiments(Spieth et al., 2004) to reverse engineer biological GRNs.
the (clow < conc. < chigh) figures. The protein quantityis subsequently changed by the amount indicated by thelast number.
0: [11] ( 4 -> 3) if ( 1 < conc. < 7) += 171: [23] (11 -> 2) if ( 25 < conc. < 40) -= 172: [22] (17 -> 1) if ( 21 < conc. < 85) += 103: [14] (12 -> 16) if ( 75 < conc. < 85) += 174: [07] (14 -> 5) if ( 27 < conc. < 93) -= 105: [12] (17 -> 7) if ( 14 < conc. < 96) += 206: [15] ( 9 -> 11) if ( 26 < conc. < 36) += 177: [16] (15 -> 3) if ( 38 < conc. < 41) -= 18
Figure 6: A successful GRN with fitness f(Si) equal to 7. Itconsists out of eight genes interacting with each other usingeighteen protein types.
Figure 6 shows a gene regulatory network of a suc-cessful swarm. In Fig. 7 several runs are shown whereeach time one start codon is removed. Observe that thedeletion of a start codon can reveal an underlying startcodon and the resulting set of genes can still be of theoriginal size (eight genes). This experiment is not meantto deduce a minimum number of genes. An individualgene might be knocked out without diminishing fitness,in contrast to the knockout of a combination of genes.
Figure 7: A knockout experiment of a fit swarm — L(Si) isaround 25 in the non-pathological case. The x-axis shows atrial index (50 trials), the y-axis shows L(Si).
4.5 Coupling experiments
Did the gene regulatory network really respond to theenvironment or does it just exhibit the proper periodicbehavior by its internal dynamics? Periodic behaviorevolves in the situation in which there is no environmen-tal input (using the same periodic fitness function fromEq. 5). The results are not shown, but a GRN will evolveexhibiting periodic behavior; however, without the abil-ity to adjust itself to light stimuli.
How robust is the gene regulatory network with re-spect to longer summer days? Figure 8 shows an ex-periment in which the duty cycle is adapted. There are
142

R = 9 runs with a duty cycle ranging from 0.0, . . . , 1.0with steps of 1.0
R .
Figure 8: A duty cycle experiment from always light (dark)to always dark (left) and intermediate conditions. The y-axisshows L(Si). The duty cycle experiment uses a block waveinstead of a sine wave, which is why fitness stays low.
In Fig. 8 it can be seen that the GRN is not robustagainst duty cycle variations. The actual behavior ofan evolved swarm is revealing (see Fig. 9). In this ex-periment, the input deviates from the default day-nightrhythm by introducing “clouds”.
Figure 9: A swarm over N=4000 time steps. The x-axisshows time t, the y-axis shows the organism size. The sensorinput and day/night wave forms are qualitatively shown forconvenience.
Clouds in Fig. 9 correspond to the parts in the graphwhere the sensor input remains zero. Normally, theswarm assembles at night, and disassembles at daytime.However when it stays dark during the day (at t = 1200)it assembles and disassembles without sensory input foran additional cycle. If the next day it stays dark again,it does not disassemble anymore. This swarm does haveits own internal dynamics, but it needs to be regularlyreinforced by environmental input.
In Fig. 10 it can be seen that the GRN is considerablyrobust against phase shifts.
Figure 10: A phase experiment running from 0 phase to aphase equal to the period. The x-axis shows a trial index (50trials), the y-axis shows L(Si).
5 Conclusion
Light has tremendous regulating e�ects on cells in bio-logical systems, from plants, to bacteria, to humans. Themachinery of the cell, in the shape of a self-organizinggene regulatory network, responds to this influence dur-ing its entire lifetime. Light as an epigenetic phe-nomenon contributes together with the genome itself,to the development and metamorphosis of the organism.The introduction of the environment as an informationalsource for the development of organisms has been rarelyexplored.
We have implemented a model of a gene regula-tory network (GRN) on simulated modular robots. Aswarm is evolved that responds to environmental peri-odic stimuli by undergoing body transitions. The evo-lutionary algorithm is based on analog genetic encod-ing (Durr et al., 2006) and fitness uniform optimization(Hutter et al., 2006).
Several experiments tested the evolved gene regulatorynetwork for its coupling properties. One set of experi-ments tested the gene regulatory network against muta-tions. Knockout experiments, by erasing start codons,show that not all genes are essential. A second set ofexperiments was devised to test against variations in thecircadian input. Phase shifts do not seem to hamper theperformance too much. The duty cycle tests for di�erent“ratios” of day and night show degradation of GRN per-formance. On inspection of an evolved swarm it showsthat a cyclic rhythm needs to be regularly reinforced byenvironmental stimuli.
Future studies are necessary to investigate the evo-lutionary trade-o�s between direct epigenetic influenceon the gene regulatory network and indirect influencevia an artificial brain. What are the circumstances inwhich cognitive and centralized preprocessing of envi-ronmental input is advantageous? Plants can exhibit avariety of phenotypic plasticity under influence of the
143

environment. What type of environments enforces us togo beyond robotic plants?
Acknowledgements
The authors would like to thank the anonymous refereesfor their constructive comments. The “REPLICATOR”project is funded by the European Commission withinthe work programme “Cognitive Systems, Interaction,Robotics” under grant agreement no. 216240.
References
Alvarado, A. and Tsonis, P. (2006). Bridging the re-generation gap: genetic insights from diverse animalmodels. Nature Reviews Genetics, 7(11):873–884.
Bass, A. and Groberb, M. (2000). Social and neuralmodulation of sexual plasticity in teleost fish. Brain,Behavior and Evolution, 57(5):293–300.
Bongard, J. (2002). Evolving modular genetic regula-tory networks. In Proceedings of the 2002 congresson evolutionary computation, pages 17–21. Citeseer.
Durr, P., Mattiussi, C., and Floreano, D. (2006). Neu-roevolution with analog genetic encoding. ParallelProblem Solving from Nature-PPSN iX, pages 671–680.
Dusheck, J. (2002). Evolutionary developmentalbiology: It’s the ecology, stupid! Nature,418(6898):578–579.
Gierer, A. (1988). Spatial organization and genetic in-formation in brain development. Biological Cyber-netics, 59(1):13–21.
Hutter, M., Legg, S., and IDSIA, S. (2006). Fitnessuniform optimization. IEEE Transactions on Evo-lutionary Computation, 10(5):568–589.
Iwao, K., Fujisawa, T., and Hatta, M. (2002). A cnidar-ian neuropeptide of the GLWamide family inducesmetamorphosis of reef-building corals in the genusacropora. Coral Reefs, 21(2):127–129.
Jiao, Y., Lau, O., and Deng, X. (2007). Light-regulatedtranscriptional networks in higher plants. NatureReviews Genetics, 8(3):217–230.
Kernbach, S., Hamann, H., Stradner, J., Thenius, R.,Schmickl, T., Crailsheim, K., van Rossum, A., Se-bag, M., Bredeche, N., Yao, Y., et al. (2009).On adaptive self-organization in artificial robot or-ganisms. In Proceedings of the 1st InternationalConference on Adaptive and Self-adaptive Systemsand Applications (ADAPTIVE 2009), pages 33–43,Athens/Glyfada, Greece. IEEE.
Kingsford, C., Delcher, A., and Salzberg, S. (2007). Aunified model explaining the o�sets of overlappingand near-overlapping prokaryotic genes. MolecularBiology and Evolution.
Knabe, J., Nehaniv, C., Schilstra, M., and Quick, T.(2006). Evolving biological clocks using genetic reg-ulatory networks. In Proceedings of the ArtificialLife X Conference, pages 15–21. Citeseer.
Metta, G., Fitzpatrick, P., and Natale, L. (2006). Yarp:Yet another robot platform. International Journalon Advanced Robotics Systems, 3(1):43–48.
Ni, Z., Kim, E., Ha, M., Lackey, E., Liu, J., Zhang,Y., Sun, Q., and Chen, Z. (2008). Altered circadianrhythms regulate growth vigour in hybrids and al-lopolyploids. Nature, 457(7227):327–331.
Quick, T., Nehaniv, C., Dautenhahn, K., and Roberts,G. (2003). Evolving embodied genetic regulatorynetwork-driven control systems. Advances in Arti-ficial Life, pages 266–277.
Sabath, N., Graur, D., and Landan, G. (2008). Same-strand overlapping genes in bacteria: compositionaldeterminants of phase bias. Biology Direct, 3(1):36.
Sauze, C. and Neal, M. (2008). A biologically inspiredapproach to long term autonomy and survival insailing robots. In proceedings of the InternationalRobotic Sailing Conference, May 23rd-24th, pages6–11.
Schmidt, T. (2010). Robotics: Environmental aware-ness through cognitive sensor fusion. Master’s the-sis, University of Groningen.
Spieth, C., Streichert, F., Speer, N., and Zell, A. (2004).Iteratively inferring gene regulatory networks withvirtual knockout experiments. Applications of Evo-lutionary Computing, pages 104–112.
van Dijk, J., Kerkhofs, R., van Rooij, I., and Hase-lager, P. (2008). Special section: Can there besuch a thing as embodied embedded cognitive neu-roscience? Theory & Psychology, 18(3):297.
Waddington, C. (1942). The epigenotype. Endeavour,1(1):18–20.
Winkler, L. and Worn, H. (2009). Symbricator3D –a distributed simulation environment for modularrobots. Intelligent Robotics and Applications, pages1266–1277.
Zhu, Y., Zheng, T., Stevens, R., Zhang, Y., and Boyle,P. (2006). Does “clock” matter in prostate can-cer? Cancer Epidemiology Biomarkers & Preven-tion, 15(1):3.
144


















