
CIENCIAS BIOLOGICAS Y - 148.206.53.84148.206.53.84/tesiuami/UAM LOTE 5/UAM20176.pdf · decir que...
72
MATRICULA; 92233479 ‘$1 /LICENCIATURA: HDROBIOLOGIA &’ UNIDAD: IZTAF’ALAPA /DIVISION: CIENCIAS BIOLOGICAS Y DE LA SALUD I .ail TELEFONO: 7277979 PIN 69045 19 ,I_ 4 TRIMESTRELECTIVO: 99-1 i /TITULO DEL TRABAJO: COMPORTAMIENTO SOCIAL DE LA TONINA (Tursrops truncutus) EN CONDICIONES DE CAUTIVERIO LUGAR DONDE SE REALIZO EL SERVICIO: UNAM UAM-I FECHA DE INICIO: FEBRERO 24 DE 1998 /FECHA DE TERMiNACION: ENERO DE 1999 i 1 CLAVE DE REGISTRO: H 003 98 & 4 - -LtcqL(- 9 BIOL. ROSA MARTHA ORTEGA L. &Af A?LO”r.7 . M. EN C. CARLOS ALVAREZ SILVA
Transcript of CIENCIAS BIOLOGICAS Y - 148.206.53.84148.206.53.84/tesiuami/UAM LOTE 5/UAM20176.pdf · decir que...

MATRICULA; 92233479
‘$1 /LICENCIATURA: HDROBIOLOGIA
&’ UNIDAD: IZTAF’ALAPA
/DIVISION: CIENCIAS BIOLOGICAS Y DE LA SALUD I
.ail TELEFONO: 7277979 PIN 69045 19 , I _
4 TRIMESTRELECTIVO: 99-1 i
/TITULO DEL TRABAJO: COMPORTAMIENTO SOCIAL DE LA TONINA (Tursrops truncutus) EN CONDICIONES DE CAUTIVERIO
LUGAR DONDE SE REALIZO EL SERVICIO: UNAM UAM-I
FECHA DE INICIO: FEBRERO 24 DE 1998
/FECHA DE TERMiNACION: ENERO DE 1999 i 1
CLAVE DE REGISTRO: H 003 98 &
4- -LtcqL(- 9 BIOL. ROSA MARTHA ORTEGA L.
&Af A?LO”r.7 . M. EN C. CARLOS ALVAREZ SILVA

A c m w n t i d ~
UNIVERSIDAD AUTOPIOMA METROPOLITANA DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD SECRETAR~A ACADEMICA
A QUIEN CORRESPONDA:
Por medio de la presente se hace constar que el:
del Departamento de HlDROBlOLOG~A de la División de Ciencias Biológicas y de la Salud, asesoró el siguiente Servicio Social:
TITULO "COMPORTAMIENTO SOCIAL DE LA TONINA (Tursiops
ALUMNA RODRíGUEZ PALACIO M6NiCA CRISTINA MATRiCULA 92233479 LICENCIATURA HlDROBlOLOGíA PERIODO
Se extiende la presente para los fines que al interesado convengan, en la Ciudad de México, Distrito Federal a seis de septiembre de mil novecientos noventa y nueve.
M. en C. CARLOS ALVAREZ SILVA
truncatus) EN CONDICIONES DE CAUTIVERIO"
FEBRERO 24, 1998 a FEBRERO 16, 1999
k, "CASA ABIERTA AL TIE A T E N T A M E N TE,
J I SECRETARIO ACADÉMICO
UNIDAD IZTAPAIAPA AV. M ~ C ~ O B C L L ~ y ia PU~SIM. coi. wc~n6na. D.F. os340 T~J. (5) 723 83 51, Fax (5)612 80 83 e-mail: cdcb.yxanum.uam.mx.

ICE
'4n~dcntes ........................................................ 1
Objetivo Genemi .................................................. 23
Introáwión ....................................................... 6
Objetivos Particulares .......................................... 23
Hipótesis de Trabajo ............. ........I.. ................... 23
-lid ................................................................ 24
Etograma 28 ; 1
Metodo10gh U t i l d y Actividades Realizadas 26
..............................................................
34 Objetivos y Metas Alcanzadas
Resultados ...................... : ..................................... 35
Análisis de Resultados ......................................... 50
Resultadcs y Discusiones .: ................................... 53
Conclusiones Finales y Recomendaciones .......... 63
Literatura Citada .................................................. 66
.

ANTECEDENTES
Los comportamientos son patrones en el tiempo, por lo que cualquier investigución del comportamiento se ocum de secuencias que a difenisea de las caracttres corporaies, no son siempre visibles. Estos, se manifiestan la mayoría de las veces, en movimientos musculares, aunque algunas veces también en actividades glandulares o aiteraciones pigmentarias (cambio de color).
Cuando un etólogo observa a un anúnai en una actividad debemllnads, se le piantea la pregunta de porqué este animal se comporta de ral f m a y no de otra Cupado oímos cantar un pájaro, nos podemos preguntar por qué lo hace en didad, tratamos primen, de ver en que forma contribuye este canto a la coaservaci6a de la especie, que ventaja selectiva ofrece ai animal, y con estas preguntas, el etólogo se acerca a la etología Cuando por fin se encuentra la función del comportamiento, se pede entonces. comenzar a investigar su desarrollo. en el desarrollo de organismos jóvenes se observa un proceso de diferenciación e integración de las formas de comportamiento (Eibesfel4 1974).
Un comportamiento siempre tiene una causa, esta se puede hallar tanto en esamulos sensoriales externos como en los mecanismos esamulocams del sístcma nervioso antral. las hormonas o los estimulos seasorialcs intcmos. LM causas fisioi6gicns del comportamiento son tema de estudio para el etólogo, tendiendo as1 un puente hacia la fisiología, cuyos métodos utiliza a menudo.
El estudio comparado del comportamiento, apareció como disciplina que investigaba la, áreas de las ciencias naturales, desarrollándose a partir de la mlogía, especialmente gracias a las investigaciones de K. Loreaz y N. Tinbergen, y se bas6 en el descubrbnisnto de las adaptaciones filogenéticas en el comportamiento. Sin embargo el cbnooimiento de estas determinanta relativamente independientes de la experiencia iadindual es más anti@.
En 1716, Pemau (citado por Eibesfeldt, 1974) sabía que los animales poseea habilidades innatas distintas de las adquiriáas, jmtrones de comportamiento que no deben apemiex por la imitación de un modelo o por medio de otros tip de adiestramiento. Descnbi6 el comportamiento de distintas aves y determinó qué especies debfan aprender el canto de sus padres y cuáles conocían el canto típico de la especie a través de su memoria genética, al llegar a la madurez sexual.
Por otro lado Spiilding, en el mismo año, demostró la maduración de las formas de comportamiento innatas colocando golondrinas en jauias tan estreches que no les permitían mover las alas, a pesar de esto, estas aves volaban pcrfcciammtc desde la primera vez que les dio la oportunidad pam hacerlo (Eibesfeldt, 1974).
.
1

P I
?.,.
Al finalizar el siglo XIX, el concepto de la evolución por selección natural había alcanzado gran aceptación. La etología enfocaba entonces los procesos y el progreso de la evoluci6n cerebral. Los primeros y más destacados investigadores en este campo fueron Darwin y Spencer (1959 y 1855 respectivamente; citado por Klopfer, 1976), quienes explícitamente reconocieron, ambos, una continuidad en los estados mentales y las capacidades conduciistas, desde los mamíferos inferiores hasta el hombre. Ambos se interesan>n, asimismo en las leyes que gobiernan la evolución mental. En particular, los principios danvinianos de antítesis y hábitos asociados han demostrado su validez hasta el día de hoy.
La dirección indicada por Darwin fue seguida por Romanes (1884, citado por Klopfer, 1976), quien estableció el estudio en toda forma del comportamiento compativo, y preparó el temno para el primero de los modernas estudiosos del comportamiento animai: Lloyd Morgan. Las advertencias de hforgan para que no se hicieran suposiciones superííuas desempeñaron un -1 importante en el desano110 de los procedimientos objetivos y funcionales en el estudio del comportamiento.
Las aportaciones de James también fueron vitales para los conductistas que trataban de liberarse de algunas de las engañosas suposiciones del lamarckianismo. Jamcs en 1890, presentó una defdción completamente mecanicista del instinto. Descnbia los instintos como correlacionados con los órganos. De la misma manera que un animai tiene ciutos órganos posee también la capacidad innata de uiilizwlos, y esta capncidad se basarla en una organización neural detemhada.
Lloyd Morgan (1894, 1900; citado por Klopfer, 1976) se expresa de forma semejantc al decir que ladestnictura del sistema nervioso central en quc se basan los instintos es un resultado del desarro110 filogenético (Eibesfddt, 1974 y Klopfer, 1976). Finalmente Loeb (1890-1918; citadoporKlopfer, 1976)ysuescuelamecaaicistaencontran>nlametodologla de las ciencias fisicas en las raíces especulativas de le etología (Kiopfcr, 1976).
Sin embargo, se considera que los verdaderos precursores de la Etobgía h n Charles Darwin, Ch. whitman, O. Heinroth y W. Craig (1872, 1899 y 1919, 1911 y 1918 respectivamente; citados por Eibesfcldt, 1974); siendo Darwin el primero en introducir el
trabajo sobre los movimientos de expresión en el hombre y los animaia. Heinrodi d 6 la sistemática de especies afines de ánades, y Whitman la de las palomas. En su búsqueda de canicteres útiles para la sistematica se encontraron con los comportamientos innatos constantes que caracterizan, igual que las estructuras morfolbgices a dearmiaadss categorías sistemáticas, y en cuyas similitudes pudieron leer el parenkm más o qmos cercano entre las unidades taxonómicas. A esto Heimoth lo Ilam6 "arteige Triebhandungen" (impulsos propios de la especie)
punto de vista filogenético comparado en la investigaación del compoaamiciao ,-m
Craig fue el primero en diferenciar el acto consumatono "estereoti 'pado" del comportamiento apetitivo inicial variable, con el que el animai busca m a situación determinada apropiada de estímulo liberador.
L
I

i
rc'
' I r
r l
,. , ,,. ,_<I__I '
J. Uexküll(l921; citado por Eibesfeldt, 1974) tuvo gran influencia en el desarrollo de la etología, investigó por medio de experimentos las interrelaciones entre los organismos y su medio ambiente, demostrando que un animal percibe con sus órganos sensoriales svlo una parte limitada del medio que le rodea, algunas de estas propiedades del medio percibidas le sirven de sea; según él, Sólo tienen señales aquellos objetos que tienen importancia para la vida del animal, y con ello se convierten en portadores de seilal para el sujeto (Eibesfeldt, 1974).
La aparición de un objeto portador de señal en el campo de percepción de un sujeto tiene siempre un efecto que confíere un significado funcional al portador de la señal. El significado funcional extingue siempre la seilalización. La señal puede extinguirse. objetivamente, cuando se trata de alimento que es c o m i d o , o bien extinguido sub~etivamenie cuando se pmenca saciedad, con lo que cierre el filtro del 6rgano sensod. Tan pronto como el significado f u n c i d del objeta extuigue su sellal. se c h el ciclo funcional que parte del objeto y, pasando por el sujeto, vuelve de nwo al objeto (Uexkoil, 1937; citado por Eibesfeldt, 1974).
Lorenz (1935, 1937; citado por Eibesfeldt, 1!474), fue el primero en reco~lcet Ir importancia de estos descubrimientos, apoyado por una serie de observaciones propias como base para la inducción, propuso una síntesis, que constituye el fuadamento del estudio comparado del comportamiento. Al principio, el núcleo de su9 ObSenaEiones fueron los patrones innatos de comportamiento, reconoció la espontaneidad que se bpua en la base de los movimientos instintivos, particuiaridad fisiológica que había sido ohrldada por los reflexólogos cláisicos. investigó que estímulos clave desencadenaben un determinado comportamiento antes de cuaiquia experiencia y estudió la filogenia y la ontogenia de los patrones innatos de comportamiento. En la conjunción del iiistinto y el adicsmmw enamtK5uaalultvjpformitde iateprrrcióndekiRImtey E6la4Uíra y arb fenhem áe impronta <BescubR6 UMI dispaaíoión innata al apmdízaáe dc espacial
p i i r a k iniportancut . s u b r s y 6 U M I y o t r a v e z f a t r a s c Q d e m a d a n i t o s ~ cienciies que mtaa del llemhe.
. .
Tmbmgen (I937; citado por Kbpkz. fW6) estudi& d mmhiui~ &&tis==@- cuandohacen nx&r sus huevos, y ambos expiicaron en haaños stgnkntes mticbosa tas conceptm basicas de Is Etoi~gís. Lorem y Thbgen, son am&e&os losvwda&ms fundadonsdel es$udiQ comparado del ComporClfluai tosigitieadohtrn&iwt&Wliiiilíg, Craig, Heinroth y Huxiey, observan>n a los animales en su medio aritural o en condiciona seminaturales de cautividad. Su primer objetivo fue el compilar un archivo completo o "etograma" del comportamiento de las especies estudiadas, y iuego descubrir la función de cada pauta de conducta. Un primer blanco de su atención fue la exhibición, puta deslurollada para servir a una función principalmente comunicativa (Klopfer, 1976).
En un principio, la etología recaía en la investigación del instllito, aunque sin limitarse a ello. pudiendo defullr instinto como un mecanismo nervios0 jerárquicamente organizado
3

susceptible a ciertos impulsos de origen interno y externo que lo estimulan, desencadenen y dirigen, y responden a tales impuisos con movimientos coordenados que contribuyen a manten& el individuo y la especie (Tinbergen, 1970).
La Etología es pues, una Ciencia Natural, nacida de la Biologíu, íii la que ha tomado el método descriptivo comparado y el método analítico fisiológico (morfología y fisiología del comportamiento). Su posición dentro de la teoría del conocimiento es el realismo crítico, orientada según el neodarwini ' 'smo y prticip en un fructífero intercambio de ideas con otras escuelas del estudio del comportamiento orientadas se& las Ciencias Naturales, especialmente con el Conductismo y con algunas escuelas rusas (Eibesfeldt, 1974).
En general, todas las investigaciones, deben empezar por una descripción y ordenación de los fenómenos a estudiar, por lo tanto, la base de todo estudio etológico es el &grama, catiilogo exacto de todas las formas de comportamiento propias del animal. Pam su realización, se escogen unidades de comportamiento que no sean demasiado p#lue&s y por lo tanto poco significativas para ser difermciadas, ni demasiado . B r a n d e s P p U e s resultarían demasiado variables (por ejemplo: escarbar, erguir la cola y cabeza, roer, eta).
El etograma es en sí, el punta de partida de la investigación, Tinbeqen (1951; citado por Eibesfeldt, 1974) dejó en claro que debería acentuarse espccialmcntc la importancia de un inventario completo de las pautas de conducta de una especie, la razón de tal énfasis es que
<Fie influyen sobre la pauta de conducta en cuestión.
Ya en 1906 Jennings (citado por Eibesfeldt, 1974) habló de la necesidad de empezar el estudio del comportam¡ento de una especie auimai con un inventario de lap de comportamiento la que llamó sistema de acción.
La descripción de una forma de comportamiento debe incluir cuaiquier detalle del proceso. Pero una descripción Asica de este tipo no es nunca completa en la pdctka, ya que el observador omite lo que no le parece importante, por ello es necesario hacer uso ds Camaras de video las cuales son el principal instrumento del etólogo, ya que aquí se fijan las formas de comportamiento y se co- p una comparación fimira, adcnuis al analizarla a cámara lenta o dpida, nos proporciona ¡nfomaciones no BoEcsI(I1cJ a la observación directa (Eibesfeldt, 1974).
Casi siempre, se describen las formas de comportamiento se& su finicón, en estos asos se desplaza la atención desde la coordinación de movimientos que lleva a la meta basta aquella. Transportar o construir un nido, son conceptos funcionales, pcío sin cmbugo presuponen una in5erpretación por parte del ObBerVador. y ahí radica el peligro de esb
otra causa, en la función del comportamiento. Desde el punto de vista biológico, la funci6n se mide por la supervivencia Sin embargo, la supervivencia frecuentemente se e~eguni de maneras bastante indirectas.
una investigación prematura puede dejar de tom en cuenta factom
procedimiento (Hmde, 1966). Y es que las escuelas de etología se han interesado ,PorWU
4

Para poder registrar la duración, frecuencia y posición relativa entre ellos de una secuencia de patrones de movimiento, sin por eso dejar de observar al animal, se suele emplear un registro múltiple (Eibesfeldt, 1974).
La mayoría de observaciones se realizan en aninales cautivos, lo que tiene ciertas desventajas, el animal no puede o explorar libremente, lo que provoca perturbaciones, sobre todo en los mamíferos que nomalmente t i e m una gran actividad, en estos casos, a menudo se pueden observar movimientos del animal siempre por el mismo camino, oscilaciones del cuerpo y otros movimientos estereotipados, los cuales pueden tener causas muy diversas (Klopfer, 1976).
Un armadillo en el zoológico de Amsterdam cesaba en sus movimientos estereotipados cuando el suelo de su jaula, desnudo hasta entonces, se recubría con una capa de 20 cm de tierra, de fonna que el animal pudiera enterrarse para dormir. Los movimientos estereotipados aparecían de nuevo cuando se limpiaba el suelo de la jaula. Heidigcr (1 942; citado por Eibesfeldt, 1974) estudió una serie de pemirbaciones debidas a la cautividad A menudo los mamíferos superiores carecen de posibilidades de desarrollar sus actividades, por lo que en distintos mlógicos se ha introducido una terapia por el trabajo. La cría suele ser difícil, pues los animales no se aparean o no educan a sus hijos. Las observaciones de campo, ayudan a evitar estas perturbaciones.
Koening (1951; citado por Eibesfeldt, 1974) observó, por ejemplo, que los paros barbudos mantenidos en cautividad llegaban a criar, pero expulsaban a los polluelos del Ndo poco después de haber salido del cascarón; la causa se encontraba en la alimentación que era muy abundante y entonces, los padres saciaban pronto a los polluelos y estos ya no abrían el pico, lo cual no ocurre en la naturaleza, puesto que los padres deben buscar alimento mh tiempo.
En condiciones naturales el polluelo que no abre el pico esta enfermo o muerto, y los padres lo echan del nido, los paros barbudos en cautividad se comportaban ante los polluelos que no abren el pico como si estuvieran muertos, para eviiar esta patwbación, bastó con disminuir el alimento que se les suministraba.
Otras muchas perturbaciones en animales cautivos tienen causas semejantea. Si el investigador conoce a los animales, las podrá comprender y evitar pronto, es falso que el animal en cautiverio se comporte de manera diferente y anormal, y por ello las observaciones realizadas tengan menos valor, como se afirma. Muchos trabajos excelentes demuestran lo contrario.
Algunos detalles de comportamiento sólo pod& ser estudkdos a través de UII largo contacto con la especie animal que se trata. Pueden minimizerse las pcmirbaciones debidas ai cautiverio, dejando al animal relativamente libre en su ambiente natural. Las observaciones de laboratorio y de campo se complementao entre si, y es inútil discutir acerca de las ventajas y desventajas de cada uno de los metodoÉi (Tinbergen, 1970 y Eibesfeldt, 1974).
.

f-
II .,.,_,
P ta.,.,
I- 1 '.Y
INTRODUCCION
Los cetaceos conforman un grupo de mamíferos muy pticuiar, que se diferencia de los demás mamíferos acuática porque estan adaptados para reaiizar absolutamente todas sus funciones dentro del agua, medio en )cl que pasan toda su vida, En este grupo se incluyen gran número de especies, todas con estriLctunis anatómicas y estrategias fisiológicas particulares.
Su cuerpo tiene forma de torpedo, lo cuál les da ventaja para moverse más fácilmente en el agua, esta forma casi no tiene partes salientes que opongnn resistencia al movimiento y, además, la reducción de la superñcie del cuerpo que se encuentra en contacto con el agua, junto con otras vasias adaptaciones les ayuda a conservar y mantmer su calor corporal. Esto es muy importante ya que en el ague, el calor se pierde 25 vccesmásrápidoque en el aire. Sin adaptaciones para contramstat este fenómeno, y otros m8s, la vida de los mamíferos acuátim no seria posible.
Los cetaceos son m a m i f a que provienen de anmtros que originain~cnte vivieron en las partes secas de los continentes y no en el mar. Poco a poco y con el paso de muchos millones de años, estos animales han evolucionado hasia las formas que ahora conommm.
orden taxonómico de los ceteceoS ha sido dividido en dos subbrdencs: h Misticetos o ballenas barbadas y los Odontocctos o &áceos con dicnies, que es el grupo al cual pertmccen los delfines (PérezXortéq et ul., 1994).
Para estudiarlos más facilmente y baddose en su origen y afinidades f i l o g d t i ~ el
Hace cincuenta miilones de años, aparecieron en el planeta, los antcawm de los c&ícm~, la mayoría de las -es presentes eran de tallas palucees y mediaow, ilcgmdo a alcanzar una longitud máxima de 21 m. Sus cucrjm eran muy largos, tenían extrcmida&a posteriores muy reducidas y el hoCic0 muy alargado. Estas criaauas habitaron aguas somera9 de ambientes costeios o de mar abierto. Sus restos fbiles han sido enconidos CII depósitos del Eoceno Medio y Tardío.
Aunque en la actualidad sólo viven cetaceos pertenecientes a los subórdeaes Mist id y Odontoccti, sus más antiguos ancestros se catalogan en otro suborden que en la actualidad no tiene representantes vivos: el suborden Arcbxccti, a cuyos Fepnsentsntrr se les ilama Ungulados, sin embargo, deescubrimientos recientes. como por ejemplo el hailazgo de fósiles cctáceos muy primitivos en depósitos de la región paquistani del Himplayq propiciaron una reorganización que acomoda los cetáceos y a los artiodtkiiloa (b6vidos, &vidos, etc.) como descendientes de un anccstm común (Em, 1987).
A r q w algunos Arqlleocetos primitivos fueron inicialmaite clslificoda, como
. Los odontocetos nctuaies presentan UIUL ashetría en el cráneo. El desmollo de esta cualidad les facilita realizar la ecolocación, función con la que pWaen Onent~e y detedar tanto a presas como a posibles deprededom. El arreglo asirnétrico de ciatM cstrucairas, les facilita el poder dhigir sonidos de alta frecuencia (mas de 150 kcps) que muitan ideales par "ver con el o í & ya que pueden recibir el eco de los objetos en forma similar a
6

como se da el fenómeno de ewuóicación en los murciélagos (Aguayo et al., 1983) y bjas frecuencias, alrededor de 15 kcps, los cuales son mas adecusdos para la comunicación.
Estos sonidos se dan en forma de clicks, producidos por movimientos de aire reciclado, sacos y válvulas de los pms nasales, este aire es enfocado a través de una estnictura de tejido adiposo conocido como “melón” que actúa como un cristalino acústico, entonces lo proyectan al medio ambiente y estos al rebtar en un obstaculo vuelven ai oído del animal después de ser ampiificados por una masa de grasa de la mandíbula inferior. entonces el cerebro analiza el tiempo que ha tardado el sonido en volver y de ello deduce con exactitud la naturaieza, dimensiones y distancia de los objetos que le rcdean (Evaas. 1987 y Yañez Me- 1990).
Una superñcie plana en la parte posterior de cada premaxilsr en la mayoria de los fbiies y de los Odontocetos existentes, indica el sitio de un saco premaxih y pumibiemente cuando los primeros Odontocetos del oligoceno iniciaron la ecolocalización. Sin cmbqo> los cráneos de estos ancestros de los delfines eran simétricus, por lo que sc ha sugerido que la asimetría de los tejidos blandos que los recubrfan precedió a la de las paries beas (Mead, 1975). Ya que el cráneo de los Arqwocetos ad como el de los Odontoccm pnmitivos era simétrico, una diferencia importante que ayuda a distinguir entre los fchiles de estos dos grupos es la tendencia a la telescopkacibn o alargamiento de la parte fiontsl de la estnictura ósea, así como el desplazamiento del orificio m d , CanCterIatica que aim no se presentaba en los Arquecwtos o, si acaso, era incipiente.
Durante el Oligoceno. apancieron algunos cctdceos qua por JUS . s e CoaJideranf~asinttnntdiasentrsIrwArqusocetosylos~acaialcsELwsennaalss ya estaban m8s adaptados ai ambiente acuático con la aparición de cataaerfstiuc cano im siguientes: M cuerpo mss hi drOdin8mic0, la pérdida de la mayor putc del peio de su cuerpo, cambio de posición de su9 orificios respiratorios junto con el desarro110 de pliegues que les permitían cerrarlos completamente; @dida de un cutiio funcid, reducci6n de las extremidades y de la cintura pélvica y desarrollo de lóbulos caudales en posici6a horizontal y una aleta dorsal, apéndice que después se perdió en algunos grupos.
Al iguai que los Arqueocetos, los Odontocetos primitivos eran heterodontos, es decir que tenían dientes de difemtes formas y funciones. Sin embargo. hacia fims del Oligocena esta caTacteristicayahabia cambiado en el grupo de los OQntocetos pues para entonces tenían numerosos dientes dispuestos en una larga fila en cada una de ISS maxila y huesos dentanos. cada diente, a i i a y aui UM solSra& era de Ismiwaa fanna de hasdcmis, w decir, que tos OdontoCetoJ deesaepaca etaahomodwtos al I & d pueIcsáeftínes8ct\ieks (Ptrez €e et et, 1994).
Et proceso de IaesrohcuCión da OdoatPoatos, del que sesdkbu@t%m aspector conocidas, tuvo lugei; a lo 1- de varias millonas de allos y dio o&e e sasantay tm espacies * que w asnqieú en miata y siete l?émlv% diez f&&l?s y - superf8milias.
.
.

La suprfamilia Platanistoidea comprende a los delfines de río, de los que no existen representantes en México. La superfamilia Delfinoidea incluye a todas las marsopas y delfines además del narval y la belugas. La superfamilia Ziphioidea está formada por las ballenas picudas; y la superfamilia Physeteroidea, cuyos representantes son los cachalotes (Barnes, 1984).
La familia Delphinidae es un grupo de origen reciente, ya que su registro fósil se extiende hasta hace aproximadamente 11 millones de años, y esta es la familia más divema dentro de los cetáceos. Son de hábitos pelágicos, aunque algunos gustan de aguas profundas. En cuanto a su alimentación ciertas especies son estmófagas, alimcntendosc exclusivamente de peces o de calamares, oíros son genedias, presentado UM alimentación más variada en la que incluyen peces, moluscos y cnist8ceos. Se cne que las especies más primitivas de esta familia eran eviuigas ya que su rostro em de longitud y anchura de tamafb mediano (Barnes, 1984).
Al iguai que otros marnifetos deprcdaúores, los delñnes presentan un cerebra cksanallado, y en algunas especies la relación entre el volumen del cerebm y el tamaft0 del cuerpo es alta.
Existen algunas polémicas en cuanto a la inteligencia de los dolfim, ea wmúu que so hable de que son muy inteligentes, sin embargo todas las especies tienen hs mismas capacidades.
El desarrollo de diferenteJ estructurss morfol6gicas y sw funciones, a veces y de manem parcial, dependen del medio en ei que habiten y de la manera en que sc hayan adagtado pararesponderaeste.
Las pruebas para medir la inteligencia son muy discutibles y SUB resuitmios no sw aceptados por muchos especialistas. Si no hay una maaem efectiw de medir la inteligencia en los humanos, mucho menos existe un método pua haccrio UXI nnimaies. Simplemente podemos decir que las deífíws están adaptacb a su ambkntc y apmvecb~ los recursos de que disponen de manera eficiente.
Cons ided las diEorentes especies da delfines padewiai dsoir qua h iim& time una amplia distribución, q s ~ i b t e ai aguas tropicaies, -opic&s y tentplad# La didbución de lap especies caaterss ag mejor comicida dcbi$o 8 la h#i&d tauto económica como funciami de estudiarlas en la cerca& a la casta
En agues mexicanas, encontramosoercode 20 espcciesdifaente4do deitlnes, sin inchiir a otros Odontocetos como los caohalotes, baileiias picudap y matsopas @%ma COrtbL et ai., 1%).
8

CLASEICACiON TAXONOMICA
P . i
RENO ANIMAL PHYLUM CHORDATA
SUBPHYLUM VERTEBRATA
SUBCLASE EUTHERIA ORDEN CETACEA
CLASE-
SUBORDEN ODONTOCETI FAMiLL4DELP”IDM GENERO Tursrops
ESPECIE Tursiop mmcatus
El nombre del género fue dado por Grevais en el año de 1855, en la revista Histoire Naiurelle des mamiferes: 2,323 (citado por YaeU Medina, 1990).
La especie fue dada por Montagu en el año de 1821 (citado por Y& Medina, 1990). Def’mw muicofup, el género ha sido cambiado, citado en la revista Mem. Wemcrian Nat. His, Soc. 3,75. Esta especie tambih puede ser encontrada con el nombre de Turaiop hasro (Grasse citado por Yañez Medina, 199C).
La tonina o Taasiopr tnmcaEw (Montagu, 1821; citado por Yaflez Medina, 1990). que es el que se estudiará en el pnsente trabajo, es también conocido como del& nsriz de botella, delfin n e w o tursión El patrón de color de esta especie es muy variado, generalmeaa es gris oscuro en el dorso, desvaneciéndose de manera gradual hacia la6 cosfpQs, hasta u11 tono más claro en el vientre. Son muy conocidos en todo el mundo, por w ~ ~ i ó n en los acuarios, películas y progamas de televisión.
Tiene la aleta dorsal ubicada en posición caitral, es alta, esbelta y falcado. La cabeza es robusta con hocico corto, grueso y bien difennciado del melón. El cuerpo es robusto en forma de torpedo con una quiüa moderada en el pedímcuio de la aleta caudal. Las atetss pectorales son de tamafb moderadoy ahusado en la punta La longitud del cuerpo varía entre 3 y 4 metros, por lo que los animales de esta especie se cmmtran entre los dslfines de mayor tamaño. Los machos generabate son mhs grandes que las hembras. El peso promedio oscila entre 150 y 200 kilogramos. La gestación tiene une duraciúa aproximadamente de un año y la lactancia dura entre 12 y 18 meses. Las crías al nacer midenalrededorde1y1.3metros(Y~ezMedm,l990yPQczCortez etaL.1994)
La alimentación de las toninas es muy variada, se dice que estos animales son oporhmistar ya que tienen la capacidad de ap~~vechar una amplia variedad de alimentos dependiendo de lo que haya a disposición. En su dieta se incluyen cefalópodos, peces Pequeeos y
Y cnistsceos. A menudo se les observa detrsis de barcos pesqunos como serdinaos camaroneros de los cuales obtienen los desechos arrojados al mar. También se Ies llega a observar frecuentemente en tomo a otras actividades pesqueras.
.
9

P
F'
Gallo (1989) observó grupos de toninas alimentándose de bonitos (Euthynnw Irneahlp), en la Punta Noreste de la isla de Santa Catalina, en el Golfo de California, México. Estas observaciones se hicieron a bordo de una pangs de fibra de vidrio de 7 metros de eslora, además. se usa~~nunos binoculares de 8 x 40 y un cuentabdtos. Abajo de la panga se podía ver pasar a los bonitos pcrseguidos por las toninas y se escuchaban siibidos producidos por estas úitimas, también se encontmon lobos marinos (Zaloph culifornrunus) en la ~ I I I I de alimentación de las tonutas.
Un caso interesante que se niaciona con la aiimentación de las toninas, es el reportado por Gallo en 1986, en donde un ejempkr hembrp captudo en Cioded dd Carmen, Campeche, f a W a las 26 horas deehabet sidvcppgñsQ, pese a ios esfucrnm rreüzaáos para aywhio. La muerte tue debida a le Íngcstidndc bagn (Bagre marhs),ya que u~llt espina dorsal del bagn, etravesó SU e9tówsgq llegando su pinba hirsta d ocas- UD absceso *. Tanlbib st han reportado asos de mtnrtt del ddfht comrinDe l~rmcu&fph#S por inelusi6n deesplta de msntanaye. que se mes incidmtal
En cuarrto al C O l I q m m m m . depgrcgici6ndeindividaosesvarinbk. E n l a s p o b I a c ~ coatetps es ram Cllumtrar manadas de m4s de so ej*nptms, mieirtns que en alta mpI
cuando e1 animel está cn busca de aiimnrta (Galbet d., 1989).
pucden ilegar a congrejpe manadas de aigunos ciento# de individuoa. En ocprfoa#i pueden encenrmrse acornpenadPo de otrru especias famiruido maMdu mixtas, muy freouentemente se asocian con calderones o bailenas piloto Globice&lu inucror~ktbi y con baiiena de aleta iWuenoperu phyJotiu hecho que ha sido obsenrado ampliamente (Gallo, 1984). También se ha visto asociado con lobw marinos Z mllfonrimnu. en la laguna de Ojo de Liebre y en la isla Santa Cataiba, en el Golfo de California. México (Oaiio, 1989).
La distribución de esfa especie es por lo gmcfai coster~, auaqire lambih se pnseinui en regiones peihgicas. Se encuentran principalmente en zonas impides y templadas del mundo.
Estos delfines no se asocian con el atún y, por 10 tanto, prácticamente no se ven involucrados en la pesquería de esta especie. Los registros de tonhas CCrcadlLI por atuneros son en lances sobre manadas mixtas y, en algunas ocasiones, en lams sobn objetos flotantes.
Estos animales tienen una gran versatilidad ecológica, 10 cual los hace que apWech.una gran variedad de hábitats.En la desembocadura del Río Colorado, ni la parte mis m i e b del Mar de Cortés, se han visto a atas animales alimentándose casi comptatamonte fizera del agua de manera similar a la seealadp e ilustrada por NOms y Ni&in (1992; citado por Delgado ei ai., 1994). Al nadar k i a La htia, en playas de pendiente suave, awndm a bs pews entre su cuerpoy la orilla dd río. Cuando los p ~ a n w cncucatnil atrepdoe entra el delñn y ladena, &tan parn scrpgilmeatc captursdos eil el hocico dd tunMh D~pu&+ el

1- u .__,. J ,-c
deifin se desliza hacia aguas mk profundas dándole oportunidad de alimentarse a otro miembro de la manada
Los tursiones aprovechan los cambios de la marea, que son muy extremos en el Golfo de California, para entrar y salir al río. También han sido vistos a grandes distancies rio arriba en algunos atimtes de la Laguna de TénniwS en el estado de Campeche y dentro de otros cuerpos de agua dulce y lagunas costeras, en diferentes partes del Golfo de México ( P k Cortes et al., 1989).
En México se distribuyen prá&camente en toda la franja costera de ambos litorales siendo abunáantes en algunas regiones. üebido a su comportamento acrobático y a su adaptación al cautiverio, la tonina del Golfo de México es muy demand& para su captura con fines recreativos en espectaculos de acuarios. Algunos estudios de abundancia y distriiución en esta región han sido desarrollados con relación a ata actividad para poder llevar a cabo una adecuada administración y manejo del recurso.
Se ha visto que aunque ilegan a observarse cries en difernites épocas del afb, existen pica en los que se presenta una mayor abundaucia de ellas (Delgado, 1991). También se hii observado que aunque la especie se encuentra distribuida a io largo de todo el litad, las concentraciones son mayores durante algunos meses, consecumtmmtc en otras temporadas se da un mayor praa0 de dispersión.
Ademas de estudios como los mencionados se e n c m en proyacto o en poctso algimas otrasi investigaciones que pretenden proporcionar infonnación sobrs la ccología de esta especie. En la Bahía de la Paz se han hecho estudios sobre la manen en que les toninas utilizan el hábitat (Acevedo, 1989). En el norte del Gdfo dt califamu . seban desarrollado estudios, que están enfocados a detamuier ' a d e m a s d e l u s o d d ~ l a manera en que lo comparten con otras especia de cctáccos (Sdber, 1990 y Ptnz Coíttz et al., 1994).
Por otxo lado es imprtante d a r que existe confusión en cuanto a la taxmmia de las toninas. A diferencia de io que se ha manejado, p.obablemente se tratri de más de ma especie. Al medir y comparar cr$rneosy OtrOBrestaB óseos coltctaQe en las ployas ea posible realizar análisis estadísticos que pemiian detcmunar ' si existen difkmciaa suficientes entre las poblaciones, como para poder decir que se trata de capecica disiintm.
Para estudios como el mencionado, es importante contar con colecciones c i d c m debidamente registradas y abiertas a los investigadores iatertsadw y ad pwmittr el intercambio de informacÍ6n (Pérez CorttZ et al., 1994).
Los delfines, poseen señales sociales complejas y un rico repertono de =mento; de ahi que puedan interactuar con otros miembros de su grupo de ma~cflu muy elaboradSS. Entre los ejemplos pueden citarse las &des de agresi6n que son ídilcs para esttblecxr y mantener jerarquías de dominancia, las seeales de galanteo, los sonidos o rnovimieatoS de
11

advertencia cuando se acerca un atacante en potencia y muchas otras M e s que contribuyen ai funcionamiento del gnipo y de cada uno de sus miembros.
El esfuerzo destinado a acumular datos sobre el comportamiento y los sistemas sociales de los delfines resulta dificultado por el hecho de que la mayor parte de su comunicaci6n se realiza bajo el agua. Es muy dificil acercarse a un grupo de delfines en un bote y permanecer con ellos el tiempo suficiente para empezar a comprender su sistema sociai. Todo lo que se ve es un grupo de aletas dorsales cuando los animaies suben a respirar a la superficie, y después se les pierde de vista cuando se desplazan bajo el agua incluso cuando el agua es clara y esta en calma, la proximidad del bote puede molestarlos, y es diñcil separar lo que es natural en su comportamiento de lo que no es y umstituye simplemente una reacción frente a la presencia del bote.
Debido a esto y a otros problemas, la mayor parte de las primeras observaciones se hicieron con delfines en cautiverio, y aunque la cautividad es una situación anormal, los investigadores podían estudiarlos durante períodos prolongados de tiempo. El trabgjo más conocido fue el de Tavolga y Essapian, de Marineland, Florida, quien observó un grupo de 12 toninas en una gran piscina, durante cinco años, descubriendo relaciones de dominancia y jemqukación. Y en generai, los cetaceos más estudiacEos son los T. mamtw, donde muchos de los aspectos del conocimiento que se tienen sobre su fisiología y comportamiento, se han adquirido durante los últimos 20 años, especialmente esaidiando a los animales en cautiverio de los delfinarios de todo el mundo (Wbing, 1979).
Tornillin, A.. G. (1957 citado por Yablokov ef al., 1974), Caidwell et al., (1972); Aguayo (1983) y algunos autores observaron que estos organismos forman grupos de aproximadamente 15 a 20 organismos, y también pequeños grupos de 2 a 3. Generalmeate viven en manadas, pero se ha observado una diferenciación estacionai de acuerdo ai scxq durante casi todo el año los machos adultos forman ~nrpos independienta, y sólo se mezclan con el grupo de hembras y crías en el período de apareamiento. La base de la manada son estas úitimas con sus crías de diferentes edades, este primer agrupamiento permanece por muchos años.
En la playa de Bahía Tiburón en el occidente de Australia viven deiíincs nariz de botella que nadan a poca profundidad e interactiuin con los turistas. en este lugar viven CQCB de 200 animales, y son estudiados por los antropólogos y primatblogos de Harvani Viven en un rango determinado en una comunidad flexible que en temporadas sc reúnen en gnipos por tallas, y condicí6n social. operando en un grupo relativamente rígido y cerrado. Las hembras cooperan con las crías aún no siendo sus madres, actúan como niñeras o tuum, como guías protectom, y para reunir a la manada en caso de peligro. Cuando los maehos alcanzan la madurez sexual, dejan la familia y forman grupos separados. NormaimentC los machos jóvenes forman p p o s a m e donde predomina una conducta h o m d , ya que no tienen bien definida su identidad sexual (Booth, 1988).
Se ha reportado, una jerarquiuición dentro de las manadas, que a veces resulta muy compleja, usualmente el macho de mayor tamaño ocupa una posición dominaate, este
12

animal (alfa) manifiesta su superioridad moviendo la cola, asumiendo poses que asocian con clics y sonidos característicos; él es el que primero se alimenta, y en periodo de apareamiento se presentan luchas entre el macho alfa y los demás delfines, el macho alfa ataca a los delfines jóvenes que están cortejando a las hembras; el resto del tiempo nada aparte del grupo y tiene poco contacto con las hembras y crías. (Tavolg y Essapian, 1957; Caldwefl et al., 1972; Cousteau, 1975; Booth, 1988; Peters, 1980)
En algunas manadas no se ha observado una jerarquía bien definida, Ekl'kovich en 1959 (citado por Yañez Medina, 1990) pudo ver que dos hembras jugaban el pap1 de guías y protectoras, pero no manifestaban rasgos típicos de dominancia, ni gozaban de ningún privilegio. Sin embargo, Sverdrup en 1970 (citado por Yakz Medina, 1990), ha encontrado hembras dominantes en algunas agrupaciones, que son las primems en twa~ el alimento, inspeccionar nuevos objetos y proteger le manada
En caso de peligro, Caldwll et d., (1972) ha observado que algunos machos j6venes (exploradores) examinan la situación y tratan de encontrar una salida para la maneda, los machos alfa no exploran, esto lo hacen especimenes de menor jerarquía, que son menos aptos para la reproduwión, protegiendo así la estructura social y el potencial reproductivo del grupo. En el caso de no hnber exploradores, ninguno de los miembros adultos asumen el papel, por io que se cree, que estos grupos son más susceptibles de ser aeapados (Peters, 1980 y Yañw Mcdina, 1990).
tas diferencias en el ComportamientD de T. t m m , se puuien atribuir ai hecho de que las relaciones sociales en las poblaciones de la misma especie. pueden varior y presentiir especificidad definida en grupos diferentes, esta habilidad de la estructm socid es un signo de alto nivel en las relaciones sociales. Esta infonnaci611, o la myorip da eii4 fue tomada de agrupciones en cautiverio y se debe tomar en cuenta que en este estado las comunidades son artificiaimente sobrepobladas, por lo que hay un incremento de dominancia.
Yablokov (1974) cree que en la vida salvaje se manifiesta menos, ya que esta balanceada y constituida por miembros del mismo rango. Un orden rígido de dominancia y subordinación se crea inevitablemente, bajo condiciones artificiales de sobrepoblación, por lo que no se pueden extrapolar las reiaciones sociales formadíis en cosnunidades experimentales, a la organización del grupo en la vida libre (Y- Medina, 1990).
.
13

P
r L. I f" I F'. 1
COMPORTAMIENTO SEXUAL:
El ritual de cortejo de los delfines, está constituido por varias etapas:
1) interacción amistosa: Las observaciones indican que el macho selecciona una hembra y la corteja por algunos días, lo cual puede durar semanas, sin embargo durante este período puede mostrar interés en otras y esto puede interrumpirse por algún tiempo.
2) Persecución: En esta etapa, la hembra tiende a aiejarse del macho, saltando fuera del agua, 61 la persigue y se reanuda el juego amomso, fases de cortejo y cópula según Yablokov et al., 1974.
3) Entrecruzamiento al nadar: Aquí, el macho se coloca bajo el vientre de la hembra, tocáucbia coa su &ctn dorsai, en esta fast se plesentan eFcceiones que dpran pocos SegMdOs (Tavolgp y Essspíon. 1957; Yabiokov et ul., 1974).
4) Poses: El mrschodurante e¡ Cortjo nada alrededor de la haah edogtamb Varias poses, como si despicgma las diferentes pertes de su cuerpo. tolrt. ulll posición vcrticcit sobre el agua y In dpea r í t a n i ~ con la ooli (Tavolga y Esapan, 195% YaMokov ef ai., 1974).
5) Caricias eo el Brea geniep1: i.as caricias ~ 8 8 el hocico y IW afeopa, st hicea f r e c ~ c o n l a p a r t e a n t e r i o r & l vienbe, ~delapSretsspectoratesy&a~gaiítaf (Tavolga y Essapias k957; Yawdtov et ai.,l9?4>
6 ) M o r d e d u t a s : N a d a a d o i a d o a a h r d o c o a L r r e ~ ~ ~ e o i r k E b a t r P ~ e s ~ pectorples dc elh& hace Iomismo coa hi alea d o d y albciiiatiwuaai tcoohoc;seidDZIiO dentro de la boca, tal juego puede durar de entre 10 y 15 minutos (Tavolga y Essapian, 1957; Yabiokov et n1.,1974).
7) Cópiax En la cójndu, eT macho wive u h hembra de if& y por oigunoa sqtmh p e n n a n e c e e n c a n t r r c t a c o n s u v i c a t r e . ~ & ~ e s t e n e a p o s i c i ó n ~ y m K B a lentanmte(Ya& Medins im).
otros atdoresmásrecientes, Bpn reddo Iasetapps en&otrespeaia~ pBses,caxiciesoee la nariz y mordisqueos suaves (Bentley Ward, 1996; comunicación penonal).
Caidwell et al., (1972) grabó sonidos emitidos por el macho durante el cortejo, loo d e s son semejantes a un ladrido o gruíiido. El mismo autor Seflaló lo siguiente "Este sonidcl fue escuchado por primera vez, hacia los tres meses de edad, cuando el bebé tratd de copilar con su madre". La homosexualidad y la masturbación son comunes en deiñnes cautivos (Tavolga y Essapian, 19S7; Caidweil et al., 1972), pero no sc ha considemdo d. Se observó que un macho puede pmcunvse a otro macho como c o m m d. Yablokov et al., (1974) indica que un delffn común (O. delphis) inicia su dvidad sexual, más a menudo con un macho T. truncutza, que m hembras de su misma especie. Caldwii,
14

menciona que cuando un nuevo miembro es introducido ai estanque, el macho ai% lleva a cabo el acto sexuai cm él para demostnu su dominancia
Todas las observaciones sobre el comportamiento sexual se han realuado bejo coadicioncs alteradas (en cautiverio), por lo que las desviaciones de las formas heterosexuales, constituyen una forma de compensación para gastar las energías en un estanque (Yablokov et al., 1974).
Durante la preñeq las hembras tienden a acompafbrse de otras hembras, aiejándose del resto de la población del estanque, con sólo breves contactos con los otros miembros de la comunidad. A través de toda la gestaci6n las actividades de la hembra decmxn y toma hábitos mhs lentos (Tavolga y Essupian, 1957).
El aumento de la región abdominal se nota aproximadamente a los cuatro meses pero hay una constante pérdida y ganancia de peso; casi en el sexto mes, se incrementa el tameflo de las giánduias mamarias, hacia los nueve meses se observa por primera vez una flexión del cuerpo, la cual ocurre cuando el animal está en posición horizoatt& lleveadose a cabo de la siguiente manera: Las aletas caudales y pedúnculo se mueven hacia la ngión veatral, llegando a formar un angUio de aproximadamente 135 grados con el cuerpo, a menudo la cabeza también se inclina y la hembra forma un arco, esta posición puede ser maatmi& por algunos segundos; cuándo el animal se relaja, la cola se eleva casi hasb los mismos grados que ocupan en la posici6n anterior, posteriormente vueiven a mdaz nomakmc.
Al onceavo mes aumenta la huencia de las flexioncs por <ua; ai inicio del doaavo mes las ejecuta constantemente por una hora o más, seguido por un pertodo sin y asi sucesivamente, esto continúa hasta que el nacimiento tiene lugar, poco anta de esto se observa a la hembra flexionarsc por varias horas ai día
Terminando la gestación, pmenta o m actividades, baja varias veccs al fondo del estanque (en el que se acaricia), para rasc~vse vi-* el área ventrai. BcrriíonaEmente seña observado qua íwertpargmes fibras anba de kabcFha también se baja hacia la pila de roc89 en ek pB0 Bictndrrpr ap€kaa&sa extremo pasterim y mrqnedo su gorcióa veatrd SObR: cIk, &&I&U cm frirrp ConstderaM e. Qtnzesatoopetivi<EacleJesEitgn~ c.mI*irririeiaa*v* Y E=Plaft, 1w.
variap semanas an(es del parto se presei#tm caiabíae ea; hn aii8iies
minutos, e inuledía- dcspiés de nspñar Ihnm&m&, p6Ta npe(b B &&kl nuevamente. Cuando más cerca está e1 nacimiento, la inusuai respiración es más ñwucníe y coIlsecuentemente ella pasa más tiempo en la superficie y sólo baja para rascame con los cepillos. Durante este tiempo se ha observado que abre la boca ampiiiimente y saca la lengua, tocando el labio maxilar, el significado de esta acci6n cs dtscomm 'do (Tavolga y Essapian, 1957).
cantínuamcntc seemcucriaamla~k,nrprsndD*air -ssaádirp~

En este período aumentan las defecaciones y respiraciones, pude ser debido a la presión del útero crecido sobre los órganos internos. La alimentación es normal durante toda la gestación; en las Últimas semanas, la abemira genital SI& una serie de dilataciones. En el doceavo mes, se pueden ver los movimientos del feto; y se O~SWM unas combas en el abdomen materno, que cambian constantemente de posición y que no se deben a los múscuios de la madre. Cuando el feto ejecuta algún movimiento violento, la madre se detiene y espera a que cese, para después continuar nadando.
Y desde la primera semana del octavo mes aparece una comba en el abdomen materno, dirigido hacia el lado derecho, persistiendo por varios días; poster¡ormente se obscrva una pmtusión cerca de las aletas pectoraies, permaneciendo así hasta en final de la gestación.
11 'i ..
F"' '..,."
COMPORTAMENTO DURANTE Y DESPUES DEL PARTO:
Un poco antes del parto, la madre nada lentamente, generaimente BcOmpBBBda por otra hembra, el nacimiento se inicia con la salida de las aletas caudaies del beb6, las cuaiw se encuentran en un plano horizontal, poco después salen 3i4 parte3 de este, cusnáo la madre se flexiona, el bebé sale un poco más, pero cuando se reiaja, mtmcede; esta situación puede pmnaneier por varios minutos, posteriormente sale todo el pedúaculo y al poco ti- sc observa a la cría subiendo hacia la superficie para tomar su primera respiración; iuego de esto se le ha observado nadando inciertamente entre las aletas d o d a de la maQe y la "tía" (niñera o madrina). En el bebé se pueden observar unas baadas sin pigmatos. El parto puede durar de 50 a 60 minutos .
El comportamiento del resto de la población del estanque, es una actitud n o d y sólo cuando alguno de ellos se acerca a la madfe, esta nada rápidamente, huyendo de 4. Todo el parto se lleva a cabo mientras la moQe nada lentamente, casi al ñnal de éste, la conducta de la comunidad cambia y se nOea una gran txcitaOión demostrada por silbidoa y otros niidos.
Después del nacimiento la piel del abdomen de la madre tiene una apariencia de papcl crepé, probablemente resultado de las fuertes contracciones de sus músculos Cuatro hwaJ después del nacimiento, se ve a la madrc hacer un rápido movimiento y la piacenta cs expulsada, sin dejar rastro de sangre. Cinco horas después del parto, el infante h t n su hocico sobre uno de los lados matemos, haciendo esto durante algunes horas, sin reacción de la madre, aproximadamente tres horas después de esto, ella se ladea y el infante busco el pezón para alimentarse por primera v a (Tavolga y Essepiau, 1957; Petcrs, 1980 y Y*& Medina, 1990)
Los delfines recién nacidos, están bien sdaptados a la vida indepmdiente, ven, oyen, nadan e intercambian información sónica con la madre llegando a distinguirla de otros delfints. En las primeras semanas el recién nacido no se aleja de su madre. y pnstnta formas

/"' t,
r L''- I
f- I , .,
r"' I
espedalesdesucomporEarmento . ,que le faeiütan sobmivir: nada atrss y a un lado de la aletadorsal de lamadre y latía, protegiéndose por ambos lados.
Cuando el atoaato sigueasu madre, mlnc8 scooloca atrás de e lk por muyrápido que este naáe, él siempre va a su lado, pera esto no t h e que peder muchas rmrgk, ya que las leyes hidrodinamicas io ayudan (Caldwelí et ui., 1972).
La madre impide cualquier intento del bebé para unirse con otms delñlics aduitos, si 61 la
medio minuto (T'avoiga y Essapiau, 1957); tal vigilancia por pate de su madre, se justifica, pues se han dado casos en que machos adultos atacan a los recih nacidos, dendotes morded- graves (Mann, 1973).
Si se pierde el bebé, este busca a su madre, nadando en círculos a altas velocidades y silbando continuamente, cerca de la superficie, en el área donde estuvo por última vez c011 ella. Si surge al@ problema en el estanque, todm los miembros de ia manada se agrupan alrededor de la m&e y del recién nacido.
Belbvich. Knishinslrayli y Gurdch (citado por bldwell et uf., 1972), oboewan>n las relaciones entre una hembra adulta y un del& de aproximadamente un año de edad, qw nadaba persistentanente alrededor de ella después de ser capturado. En los primuus m a w de cautiverio, los do4 fueron casi inseparables, 61 absndonaba a SU msQt di6 por W periodos de tiempo (1415 minutos); para jugarcoh otros j&enes,
La hembra a menudo jugaba con su delfín y esto coasida en se@- uno ni om y darse ligeros moediscos ea las aletas y acariciarse IIMI ai otro en el rostro; pbm Wqmimoattr, ao colocó al infante en otro estanque, después de esto se escwhwon sefuies de alarma eatrs los dos y se negaron a comer duraute el timp que estuvieron saparados; el joven delfh two que ser devuelto a su madre, pues sc temi6 por su vida DcspuGs da ciao0 mews us observ6 al del& en compañia de adultos más a menuáo, nueve meses m8s Eardt 61 tenfa poco contacto con su madre. En caso de peligro, aunque sean aduitos de 4 y 6 a eilos buscan la protección materna, lo cual indica que el Vinculo es muy fuerte, tambih se sabe que madre y cría SeparadSS por varios años, se reconocen cuando sc reúnen cuando un delfm permanece mucho tiempo con ella, es muy benéfico, ya que aprende e imita las experiencias de los adultos.
Hay numerosas observaciones de la vida libre que indican un fuQte sentido pater~I en estos organismos, se ha visto que cuando la madre está vieja y enferma, las crias(ya organismos aduitos), se turnan para cuidada y ayudaria a mantenerse en superfGcie sosteniéndola con sus hocicos (Caldwell et al., 1972; Peters, 1980).
dewbedece lo castíea, Ilwándolo al fondo del estanque, mananihidolo alli por cerca de

COMPORTAMENTO INTRA E iNTERESPECiFiC0:
Comúnmente estos organismos se prestan ayuda en caso de enfemedad o debilidad. En general la asistencia puede ser de dos tipos: una en que los miembros de la manada permanecen cerca del animal lastimado y oúa, la más común, en la que varios miembros de la manada sostienen y acarrean al animal lastimado, lo que previene su hundimiento.
Los delfines nariz de botella, colocan sus cabezas bajo las aletas peaorales del lastimado, manteniéndolo en la superficie hasta que se halla repuesto y esto puede tardar mucho tiempo. Algunas veces se ha observado que los asistentes son reemplazados por oms miembros de la manada .
Algunos investigadores han encontrado que sólo los jóvenes de ambos sexos son ayudados por las hembras adultas, comúnmente los machos adultos no recibea ayuda La ayuda se da mas fkilmente a miembros de la misma manada que a los de otra. La asistencia puede ser dada por miembros de ambos sexos, pero se cree que las hembras, la mayoria de las veccs son volmiarias ; este particular comportamiento debe estar relacionado con el instinto matemal.
Esta ayuda no está restringida a individuos de la misma especie, sino que también puede ser dada a especímenes de géneros diferentes, especialmente cuando ellos haupanmiecido juntos por largo tiempo. Se han visto ballenas piloto (Giobrcephuia melaem) i!ewado hasta la superficie a individuos de varias especies de delfines, tomándolos COD su boos por el pedúnculo caudal.
La acción de empujar hacia arriba, no sólo se efectúa entre miembros de dif- especies, sino también con recién nacidos mwrtos, y en general erlquier objeto inanimado. Esta útil e importante adaptación de los ccEbceos, a la vida acuática, se debe a que cuando el animal es lastimado o enferma, pierde la respiración antes que nada y estc instinto a ayudar en estos organismos, a menudo supera el instinto de propia conseneción.
Se han reportado cams de delfines T. truncatus que empujan náufragas hasta bpinym Para comprobar esto, Caldwell en 1972, realhi una serie de experimnrtoS, enconirando que lo hacen para sacarlo de su territorio y no para ayudarlo, aunque dios pueden SQ
entrenados para defendex y ayudar a humanos en el agua. El mismo autoriwlicaque d comportamiento cooperativo es una acción refleja y que un delfín puede preveair a otro de ser lastimado (Yañez Medina,lW). . COMPORTAMIENTO DE TElUüTORIALIDAü:
Tursiops mmcatus, muestra una tendencia a mantener un área muy restringida dentro de su hábitat. Algunos investigadores que los han estudiado en vida libre, dicen que un gnipo
18

r'
siempre se encuentra en la bahía, mientras que otro se encuentra a io largo de las playas, ambas defienden su microtemtwio y rara vez se alejan de él.
Estos microtemtoriw también &sten en los estanques; si el espacio lo permite, cada delffn tiene su lugar favorito, donde es localizado en sus lapsos de descanso, cuando hay actividad como alimentación o juegos, tiende a entrar a otros microterritonos, pero concluido esto vuelve a su lugar, el cual defiende de cualquier intruso, haciendo sonidos con las mandíbulas, con gestos de la boca abierta, o sólo viéndolo (Caldweil eral., 1972).
Los lugares preferidos, parecen ser los cercanos a la superficie o donde halla un chorro de agua, ya que gozan teniendo una coniente de esta sobre eiios.
Se han hecho observaciones bajo el agua, demostrando que las manadas salvajes tienden a mantener tales microterritorios dentro de una columna de agua (una íiucción de agua que se extiende desde la superficie hasta el fondo), es más deseable para ellos estar en la cima de la columna, pues así no tienen que nadar tanto pira poder respirar, pero como es esperado, los animales dominantes ocupan estas posiciones y los que están en les columnas inferiores, tendrán serios problemas si cruzan por su tenitono, esto es obvio en un estanque, ya que el espacio es limitado, pero en la vida libre la cantidad de superficie del agua no es una factor limitante, el porqué Iw delfines persisten en vivir en difmtes capes no ha sido entendido.
Según otros autores, estos animaies no realizan migraciones, sino que sc mueven en un gran territorio en la búsqueda de alimento, por lo que la territorialidad eu vid. Libn w esta muy demarcada (Cousteau, 1975,1990, Peters, 1980)
COMPORTAMIENTO ALIMENTICIO
Dentro del grupo de los delfuies nariz de botella no se obsmaa luchas por el aiimento. Yablokov (1974) menciona que los primeros en alimentame en ua eskqw son los jóvenes, los adultos no tratan de tomar el aiimento antes que elloa, sólo se otweiva competencia en forma de juego, cuando tomaban los peces de la mano del entrenador y esta consisüa en sumirse unos a otros, colocando su vientre sobre la cabeza del otro, sin presentarse una verdadera lucha.
En cuanto a las crias nacidas en cautiverio, la mayoria de las veces mucren antes de cumplir un año de edad, debido a que se les alimenta con peces mwtae, y estos los contaminan de virus, pnrásitos y bacterias para las cuaies estos no tienea antiwerpm.
ia alimentación en cautiverio, esta supeditada a l habajo, ei comportamiento es condicionado o lo que se conoce como condicionamiento opemntc, pw lo que no hay búsqueda ni otros patrones típicos de vida libre (Cousteau, 1975 y Yaik Medina, 1990).
19

L - ''.. m ,
!
IC''
" ' 1 P
r"' I r"" I
COMPORTAMIENTO DE JUEGO:
La curiosidad y los juegos, son los rasgos principales de estos animales y la mayor parte de su tiempo lo usan aparte de comer y descansar, en jugar. Los recién nacidos empiezan a jugar u las dos semanas de edad, el bebé trata de unirse. a los juegos de los adultos y es muy interesante el comportamiento imitativo a esta edad.
Los jóvenes prefieren jugar con los miembros de su misma edad Normalmente el más activo e inquisitivo inicia el juego y los demás io siguen; ellos pucden morderse, pegarse en la cola, empujarse contra la pared o sumergirse unos a otros, pero sin llegar a la lucha Verdadera
Cuando uno es alimentado o mimado, los otros comienzan a empujarlo y a desarrollar una lucha por alcanzar tal privilegio. Mientras se alimentan, uno le quita el pez ai otro, lanzándolo lejos, después lo vuelven a atrapar y así conünúan.
Debido a que la piel de los delfines es muy sensible, les gusta ser acariciados, por lo que en los tanques se colocan cepiiios en el fondo. El contacto corporal eatn ellos es muy importante por lo que acostumbirui nadar muy juntos, muchas veccs tocandose unos a otros (Carifío Preciado, 1994 y Yañez Mcdina, 1990).
Otra forma de juego es elevarse e; forma vertical fuera del agua, y golpearia rítmicamente con la cola. Algunos juguetes preferidos son: peces que habitan en el estaaque, tomigiu y pelícanos. A los peces los atram los lanzan por el aire y los &jan escspar para que sc escondan, luego los buscan y los vuelven a atrapar, pero sin hacerles dafb, a i m delíhu los empujan para que naden m8s rápido. A las tortugas las colocan su lomo y las ~carnan por todo el estanque. A los pelícanos los salpican y los molestan.
Tursrops truncufw presenta una gran Vanedad de juegos su movilidad y actitud juguetona combinados con su alta capacidad para imitar y aprender, les han dado la reputackh de actores de primera en los circos, haciendo uso de cualquier objeto. Son especialisras en pasarse un aro del rostro a las aletas pectorales, y de estas a la dorsai o lanzárselos unos a otros, jugar con pelotas y muchas otras cosas. En Estados Unidos y en Japón los han entrenado para jugarbasquetbol. llevan y traen figuras que les son easdhdsscn láminas, k quitan un cigarro de la boca al entrenador, etc. Se ha visto que otras especia de del& no pueden aprender tan exacto, aún pesando largo tiempo en contacto con T. mmcah~ (Yablokov et al., 1974). . LOS juegos, son una expresión del deseo de comunicarse en alguna forma con organismis de la misma o diferente especie. Hay la creencia de que en estos animales los juegos son esencialmente educativos; mientras juegan se ponen en contacto con todos los elementos del medio, y ello los prepara para un comportamiento apropiado cuando son mayores (Yablokov et al., 1974).
P- S I ~
20

La mayor parte del día pasan nadando y jugando activamente. Caldwell (1972) en sus observaciones ha encOntIpLd0 que los delfines más viejos tienden a descansar gran parte del tiempo, dejando su fosa nasal fuera del agua, así que no tienen que elevarse para respirar, los jóvenes por otro lado, se les ha visto por más de 30 horas en continua actividad McBride y Hebb (1948; citado por Moukhmetor, 1590) indican que ellos son aparentemente animales diurnos, pero que después de alimentarse toman una siesta por breves períodos de tiempo.
Se pensaba que los delfines dormían flotando en posición horizontal y sobre su lado ventral, maiitemendo su fosa itasril fwra del agua y cou lor ojos semicerrados. Sa encontraron ejemplares que descansaban con los ojos abiertos y se pensó, que dguws duermen así, generalmentt con el menor ruido se despertaban y volvkui a su actividad (Caldweii et al., 1972). Pero estudios recienten ban dem& que los delfincs ti- un suef$o lenta unihemht&ico, en el cuál el eatpdo del encétih registm cmdm lentas en un hemisfeno mientras que el otro hemisfen6 cppta un8 deancn#iuac ' ibn, #I decir, que mienass un hemisferio duemie, el Otro se ment#m . aspielto. cupndoun~s f e r i a duerme el otro sc mantiene aciivo cm un nivel elevado del tono muscular y dc la actividad refleja del enctfalo, indispcnsablcs para ia natizacíón n o d e inofiiisiva de los actos respiratorios (Moukhmetor, 1990).
COMPORTAMIENTO E INTERACCION CON LOS "0s:
Las condiciones de estar cautivos, los afecta notablemente, ya que se n@hm puranes de conducta de c~trcq los patrones observados en vida libn no son i@cs a loa que presaitlui en cautiverio, presentan patrom fijos estenotipicos, rígidos y rspstidos an sus
de la movimientos espaciales, aunque relacionados muy d4biimente toll la alimentnción, entre otras wnductas (GygaX; 1993).
En vida libre e incluso en cautiverio- se ha visto que los delfines prefieren mas la compiidfo
~ e ~ i o s ; l a Q r r s o s m 6 t r c l r o m a u b s J a e F ~ 0 6 ~ ~ . , , " ~ ~ f ~ R , C O Q
de !w NfWl c Ir& les ohdaos, se krai Feslc4$do npig (FaS40 da ClFri: Q6mapPndwraDI OIP
b)timo w tb T. trpemrna. rmao GrtA#e piww, w e cbm admk CiriFlgSrsdrr aGm*atos~wlas~dctaffaento~~Ismftrkcatidrr&Nsan Zek& (Ceustaaq l!?7$ caldwett el d., tsn y IW4],
Orra Eossti6nque ba m@mdb m d m bíphtmb ( ~ i rtparlu( rEoeBaicainiu laa atritwyea al ínttrato @aumm% - Y a ~ k m ~ * ~ * 4 % b h 6 f W ~ ~
rñcloBD 8 bba8frU y W q U C b?N * lSC&C4% robrc St beib. @Ciip-
buques como si quisieran cargarlos (Cousicau, 1975); otros lo relacionan coll el instiat0 maternal; y algunos a que es el ruido del motor lo que los atrae. pues no se ha visto que sigan veleros (Rodríguez, 1974), pero no se ha emunhdo una rcspcsia collcrcta y satisfactoria.
21

6'- " * ., ,
r ../. _.
r
En algunas regiones del mundo los delfines son muy apreciados. Como en Nouamgh, poblado de Mauritania, donde ellos llevan grandes banm de peces (mjol), pnra que los pescadores los puedan atrapar. Las causa por lo cual hacm esto son desconocidas (Cousteau, 1975). Actualmente mmcnfus está siendo entrenado para servir como mensajero entre la superficie y alguna estación submarina, además de ser utilizados como ayudantes y protectores de los buzos, ya que su capacidad para aprender los ha capacitado para recibir órdenes y llevarlas a cabo sin cometer ningún errw (Caldwell et al., 1972; Cousteau, 1975). Desafortudamente gracias a esta habilidad se les ha entrenado para transportar material bélico en algunos países @stados Unidos, Rusia y China) (Mcintyre. 1974).
En México, desde 1991, a niños de edad preescolar, menores con trastomos n e m l ó ~ m o autistas se les ha brindado la oporiunidad de inttrachiar con los delfines entrados especialmente para este trabajo. Una vez establecida una relación familiar, el infante nada con estos animales durante 12 minutos, dos veces por semana. Estas sesiones a pesar de encontrarse en fase experimental, han demostrado que los niños adquieren una paz espiritual y psicol6gica
Los chicos normaies aumentan su sensibilidad y se les estimula su curiosidad y avidez de aprendizaje. Los autistas y menores con trastomos neurológicos muestran mejorías mecánicas, gracias a las micmdas de uitrasonido emitidas por los dclfíncs hacia las partes irnovilizadas, que estimulan el sistema nervioso ccntral.
Los delfines son animales muy sensibles, confiados, iniuitivos y dóciles Conservar d e b s en cautiverio para que el público se distraiga y los científicos investiguen puede ser un nuevo comienzo para la investigación y evaluación de anomaiías de comportamiento en cetáceos cautivos, los estudios de estas anomalías pueden ayudar a mejorer sus condiciones en cautiverio para el beneficio tanto de ellos como de nosotroB mismos.
I I
t .
22

OBJETIVO GENERAL
observación y adisis del comportamiento social de los delfmes Tursiops truncatus en condiciones de Cautiverio.
OBJETIVOS PARTICULARES
- Comparar la conducta de los delfines Twsiops truncum en los diferentes acuuioa
- Registrar la conducta en los diferentes acuarios dependiendo de la presencia de machos y hembras.
- Observar y establecer la conducta de deüines al cambiar la composición de los individuos.
HIPOTESIS DE TRABAJO
El comportamiento social de los delfines estará en función del sexo, edad y número de componentes del grupo, así como de las condiciones del estampe, espcio, la aümmtaCión y tensiones debidas al público yio entremdores .
23

MATERIAL
Se trnbaj6 en dos d e l f i o s , uno en la ciudad de Cuernavaca, Morelos y el otro en la ciudad de México, Distrito Federal.
El delfinario del parque recreativo Jungla Mggica, situado en Cuernavaca, Morelos. Tiene una capacidad de 3 millones y medio litros de agua, con una salinidad que oscila entre 55 y 65 partes por mil, (de hecho cuando la salinidad bajó por el período de lluvias le echaron al estanque 28 toneladas de sai) y se encuentra a temperatura ambiente.
Dos delfines hembras, Cometa y Beta, con un tiempo en cautiverio de 28 y 19 años respectivamente. Cometa mide 3 metros y 10 centímetros, llegando a pesar de 300 a 350 kilogramos. Beta mide dos metros y medio con un peso aproximado de 270 kilogramos.
La alimentación es a base de peces (jurel, sierra, cocinero) , consumiendo hasta 6 kilos diarios de peces cada uno, la aiimentaci6n se les da por raciones que dependen del trabajo. Los medicamentos y las vitaminas se les incorporan junto con el pescado (Cevoun Vit A, B, C, Tevagram BIZ, Multivitaminas, Melox, Digenor Phis, y muy poais antibióticos). A Beta le están suministrando Melox porque tiene úlcera.
Durante la investigaci611, Cometa se enfermó del ojo izquierdo paca io cual se le administr6 terramicina, esto fue en Julio 15 de 19%. Mas tarde enfam6 del est6mago con una íüerte tos por io que estuvo un tiempo delicada de salud, tiempo en el que suspendieron las funciones al público para no forzarla a trabajar. Los medicamentos que se le suministraron fueron 4 Prodolinas diarias durante tres días, 10 Melox y 4 Cataplam diarios durante 3 días, todo esto dentro del pacado. Se recuperó en unos quince días aproximadamente bajo el estricto cuidado del entrenador, el señor Ricardo Roberto Ruiz Aranda (Red. Ruii) d cual lleva 4 años trabajando con ellas, quién aprendió el trabajo gracias al witrcnador anterior. La segunda entrenadora es la señorita Verónica Miyauchi Castillo.
A partir del 27 de agosto llegaron al delfinario 2 deüines cubanos los aialsr timen aproximadamente 10 años de edad y año y medio de cautiverio, son hembra y macho, y fueron destinados a las áreas 1 y 2, separándolos del resto del delfinario por medio da UM
malla Los delfines permaneerán en Jungla Mágica hasta el mes de enero de 1997 donde serán trasladados al Perú. Estos delfines traen microchips en la base de la aleta dorsai para evitar robos y confusiones.
El otro delfinario donde se hicieron las observaciones es el ubicado en d parque de diversiones Atlantis, en la ciudad de México D.F. El delfinario tiene una capacidad de un millón y medio de litros de agua, lo mantienen a una salinidad x y a tempaatura ambiente.
En el parque se encuentran dos delfines adultos Tnt6n e isis (macho y hembra respectivamente), Tritón tiene 16 años de edad aproximadamente y 7 años de cautiwrio, isis tiene aproximadamente 8 años de edad y 3 de cautiverio, y otros ddfhca denominados para los fines de este estudio como hembra 1, d o l , macho 2 y macho 3 tienen aproximadamente 1 año y medio de estar cautivos.
24

Les dan de comer sierra o arenque o Cocinero o smell, dependiendo las temporadas. UM cantidad de 6 kilos por individuo, en el transcurso del día, y los cambian frecuentemente de estanques, no permaneciendo la misma población con que inicié mis observaciones hasta el final.
Su entrenadora es la señorita Angeles de Alba y la segunda entrenadora es la señorita Christei Pellegrín.
El veterinario que las atiende a todos es el Doctor José Luis Sol6nano.
' I ?
25

METODOLOGIA UTILIZADA Y
ACiWJDADES REALIZADAS
Para obtener una mejor y más clara información en la toma de datos, se ha decidido utilizar dos técnicas de descripción del comportamiento.
-Em-: en ésta tomamos la apariencia, postura y movimientos del sujeto. Por ejemplo: la rata aprieta el interruptor, o el ave desliza la punta del pico por sus plumas.
-Consecuencia: son los efectos del comportamiento del sujeto sobre el medio, sobre los individuos o sobre sí mismo. Por ejemplo: la rata enciende la lw,o el ave se acicala.
Existen a su vez, cuatro tipos de medidas para ellas, que son:
-Latencia: minutos, horas que transcurren desde la aparición de un suceso y el fin -Frecuencia: número de veces que aparece la pauta de comportamiento en la unidad de tiempo. -Duración: longitud de tiempo en el que se prolonga la pauta de comportamiento. -Intensidad: altura de un salto, intensidad sonora (Altman, 1974 y Martln y Bateson, 1991).
Para nuestro estudio, se di primero que todo un muestre0 i d libitum, en el período comprendido de marzo 4 al 6 de mayo de 1996, una vez por semana con un periodo de observación promedio de 3 horas por vez, en el delfinario del parque de diversiones Jungla Mágica en Cuemavaca, Morelos. Este mueatre0 nos sirvió de basc para determinar bien el método e instrumento de registro más apropiado para el caso, así como para mejorar nuestro etograma inicial de la especie.
En nuestro inshumento de registro (Tabla No 1), en el cuadro donde se indica el tipo de conducta, se registraron éstas de manera estructural y cuando lo llegó a ameritar, en el cuadro donde van las observaciones se colocaron las consecuencias de dicho comportamiento, el muestre0 fue de barrido, con intervalos de tiempo de cada 5 minutos.
Las conductas, que se colocaron en el cuadro forman parte del etogrsma definido para esta especie en condiciones de cautiverio, cabe anotar que no todos los miembros de una especie se comportan del modo típico de la especie, por lo que en nuestro etogmma añadimos patrones particulares de conducta observados en los individuos Beta y Cometa, que son los delfines ubicados en Jungla Mágica y este se extendió para los deffines del parque de diversiones Atlantis ubicado en México D.F., asimismo a lo largo de los períodos de observación y la toma de datos realizados en Atlantis, se fue enriqueciendo el etograma con algunas comportamientos típicos de los delfines presentes en este lugar y se notó la ausencia de otros presentes en los delfines Beta y Cometa.
Así mismo, se midió por h e n c i a y duración los comportamientos, i n d i d o e1 Brea en el que se realizaron y todo se registró en formatos como el siguiente, para luego análizar los resultados.
26

Las observaciones sistemáticas se realizaron en Jungla Mágica de mayo 13 a Septiembre 30, una vez por semana en un tiempo fijo de tres horas. Y en Atlantis se realizaron de Agosto 5 Septiembre 30, en un tiempo fijo igual al antenor. Por lo cuál en Jungla Mágica se realizaron mucho más observaciones que en el otro delfinario.
Antes de realizarse las observaciones mencionadas se destinó un tiempo prudente para la identificación y reconocimiento de los individuos, esto se basó en marcas características en las aletas dorsales manchas de diferentes tonalidades en su cuerpo, así como en el tamaiio corporal de los delfines estudiados.
El área del delfinario de Jungla Mágica se dividió en 6 subárcas, las cuales están marcadas en el mapa anexo (Figura No 1). A partir de una división hecha por los directivos del parque, en el área del delfinario por medio de una red con el fin de separar a Beta y Cometa de un pareja de delfines (macho y hembra) traídos de Cuba, se dividió nuevamente el área ahora asignada a ellas quedando 2 subáreas para los delfines cubanos y 6 para Beta y Cometa, como lo indica el mapa anexo (Figura No 2).
En el parque de diversiones Atlantis, se dividió el área correspondiente al delfinario en 5 subáreas que indicamos en el mapa anexo (Figura No 3)
Las áreas de interacción con los entrenadores fieron la 2 y la 4 para el delfinario de At-,, y la 1 y actualmente la 4 para el delfingo de Jungla Mágica
El tiempo en que no se vieron, Beta y los delfines cubanos en el caso de Jungla Mágica e Isis en el caso de Atlantis, se debió a que estuvieron fuera del banido de obsavación. Para d casa de los primeros, estaban en el área 1 lugar que por una pared divkoria, no es posibisver desde el escenario o el iugar de observación.
.

ETOGRAMA:
COMPORTAMIENTO DE CORTEJO
1) Persecusiones: En esta etapa la hembra tiende a alejarse de macho, saltando fuera del agua, él la persigue el comportamiento es similar al de persecusión descrito para el comportamiento social de los delfines, pero va acompañado de otras fases del comportamiento de cortejo.
2) Entrecruzamiento al nadar: Aquí, el macho nada vigorosamente cerca de la hembra y pasa por encima de ella se cuerpo, la hembra voltea regularmente antes de cada d i o y nada alrededor del macho y este hacen lo mismo, mejorando el vigor en cada contacto con ella o asediando otra vez.
3) Posts: El macho durante el cortejo nada nirededor de la hembra adoptando varias poses como si desplegara las diferentes partes de su cuerpo, generalmente toma un posición vertical sobre el agua y la golpea rítmicamente con la cola, en la posición vertical tambiCn se aprecian ereccioncs de corta duración.
4) Cariciaa con la nariz: Las caricias con el hocico. se hacen tkcuentements en la parte anterior del vientre, de las alaa~pectoralles y área genitaz esto ebrealizadotantn por el macho como por la hembra y se iieg6 a deectar inclusive entre machos.
5) Mordisqueos: Nadando lado a lado con la hembra el macho sujeta con la boca las aletar pectorales de ella, hace lo mismo con la aleta dorsal y ake rdmente c o b tl dorso dentro de la boca; tal juego puede durar entre 10 y 15 minutos. Algunas veces la hembm muerde la región genital del macho logrando una erección por parte del macho.
6) Erecciones: El macho exhibe erecciones durante las numerosaa fases de la actividad sexual.
7) Cópula: En la cópula el macho vuelve a la hembra de lado y por algunos segundos permanece en contacto con su vientre. Ambos deüincs están en poaición horizoasal y nadan lentamente. Se observan penetraciones parciales, ercccionas sin 10- paieoor y penetración total en la cual no se ve el pene Sólo hasta que se sepana Nota: Se considera juego amor090 al conjunto de las conductas descritas del No 1 al 5.
COMPORTAMIENTO S O W
Penecusiones: Se persiguen nadando, generalmente a altas velocidades, luego sa topan y muy juntas se mordisquean el cusrpo y se golpean con las aletas en posición opuesta una de la otra (cabeza de una con la cola de la otra). Se persiguen luego rápidamente y sa acorralan en un área menor (si existe), sumergiéndose con violencia, después de esto ya no nadan simultáneamente y a veces toman aire sincronizados.
24
28
- - - -_ . -

Nado recto o demho: movimiento continuo hacia adelante con el lado dorsal fuera del agua en un tiempo en el que no muestra ningún otro comportamiento.
Nado lateral: Movimiento continuo hacia adelante con el lado lateral por fuera del agua.
Giros: Cambios de dirección al nadar de más de 1500 al cruzar a un Brea mayor en longitud y profundidad.
Saltos: Levantar como mínimo la cabeza fuera del agua en un movimiento continuo hacia adelante, la cabeza parece que esta rewtrando al agua. O bien son saltos altos en los cuáies queda afuera todo su cuerpo.
Nado sincronhdo: Los delfines nadan uno junto al otro en la misma dirección, repitiendo los mismos movimientos. Estan perfectamente bien coordinados.
Nado en círculos: Nadar en círculos en ruta fija y turnándose a Iugares predecibles. Parm que no tuviera ningún propósito el hacia donde se dirigen
Quedarse quieto: El animal permanece un rato quieto como recostado en el fondo del estanque.
Mirar a un punto fijo: Mirar con la cabeza fuera del agua hacia el cuarto donde esta el entrenador, al pasillo por donde van a entrar las entrsnadorss, a las cubaas de comida, al observador, o a oiras personas que se acercan al acuario.
Juegos: Estos pueden ser con un pez, sin tragarío, por largo tiempo, con monedas del tondo del estanque, hojas o cualquier otro objeto, e involucrar en el juego a las personas que se acerquen, llevándoles el objeto, para que se lo tYai lejos y ellos &o nuwamentq y así sucesivamente sin cansarse de elio. Un juego pnsente en Tritón (el macho que peraaanció todo el tiempo en Atlantis) es el echar agua en la c(v& cuando se acercan pasonas al estanque a verlos.
Vertical: Suben o bajan en el agua en posición vertical.
Boca arriba: Nado con el Vientre expuesto golpeando el agua con una o las dos aletas pectorales.
Salir: Sacar el cuerpo fiera del agua sobre el escenario y hacer caravana.
Sobar: En los bordes salientes del acuario a manera de d o n e s , soban o res&¡egan.su cuerpo por varias partes como vientre, lomo, cabeza, cara y aletas.
Flotar: Flotan de lado, dejando expuestos el ojo y parte de la cabeza en la supañcie del agua.
I
29

.
30

,_. J
I I f 19
.. a
a 8 a"
31
.
I

32

rn
R II II I R II II
n TABLA No 1
pCHA: OBSERVADOR HORA DE COMIENZO HORA FIN& BETA B >GRUPO: COMETA: C
SESION DE OBSERVACION
I I I 33

OBJETIVOS Y METAS ALCANZADAS
-Se comparó la conducta de los delfines en los diferentes d o s .
-Se registraron las variaciones de conducta en los individuos, dependiendo de la presencia de machos o hembras
-Se elaboró un etograma definido para esta especie en condiciones de cautiverio.
-Se elaboraron gráficas rqresentativas del tiempo dedicado por los orgaaismos a cada UM de las conductas, respecto ai tiempo total de obsavaci6n.
-Se determinaron las áreas representativas para los diferentes comportamientos.
-Se interpretaron los datos y se sacaron conclusiones ai respecto.
-Se comprobb el territorialism marcado en esta especie s610 presente en condiciones de cautivaio
-Se elaboró un instrumento adecuado para medii este tipo de conducta en de&@ que se encuentran en condiciones de cautiverio.
-Se registraron los cambios en la conducta social de los indicviduos, ai cambiarles in comp6siciÓn de 10s mismos.
-Se comprobó que el comportamiento social de los delfines en condiciones de cautiverio está en función del sexo, edad y número de mmpomntes del grupo, así como de h a condiciones del estanque, espacio, alimentación y tensiones debidas tanto ai público como a los ennenadores.
34

RESULTADOS
.
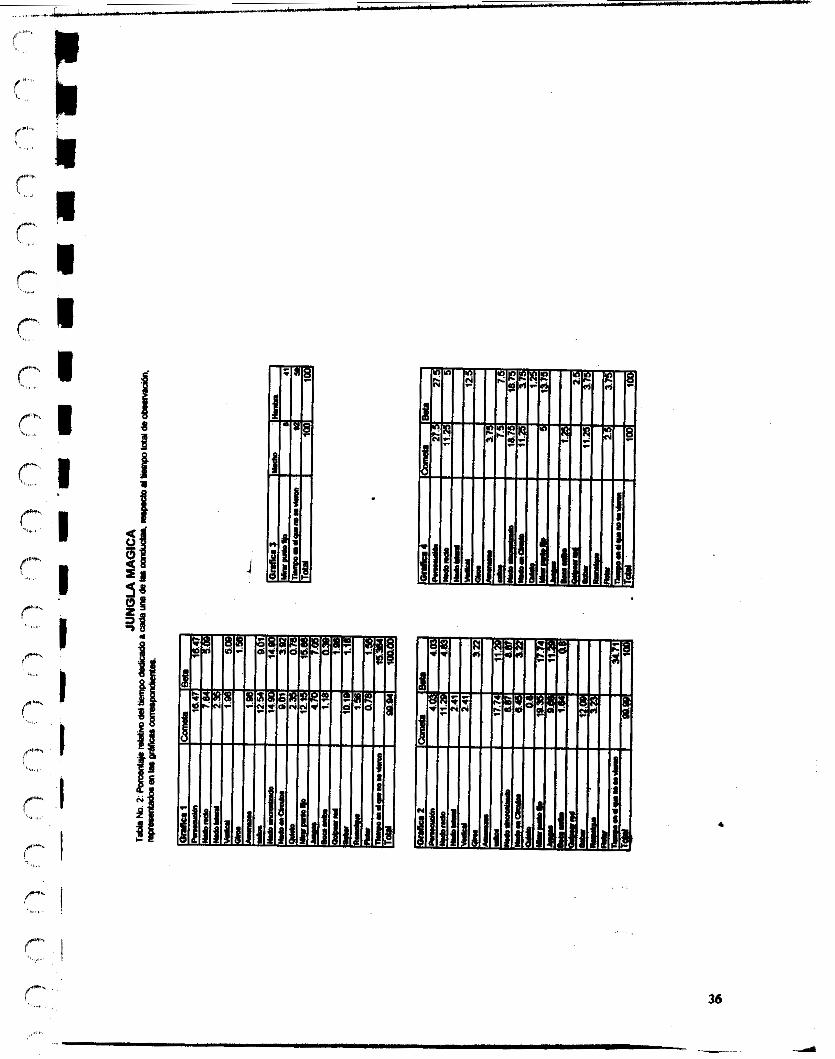
I I I I I I 1 I I I
I
.

/""
',--, I r̂' I
" I p"
P"" - 1 c I r l ír- I
F".
I i ,
/-. ./,-
8 d
8 ni
8 i
8 <o
8 m
8 8 8 8 8 5 5 f c e
U
o U
8 - o c
n
n

' I
f" ' I
F - 1
I" "
F J a 8
i
38
I
1

1-
t

. -
Y i r
t
c 8 e! ! (3
! B
I
nb
N
.
P
.- I

t- * '-' . r-
' $ t . ' . ~
. . . . . , '
+"
. .
4f
.
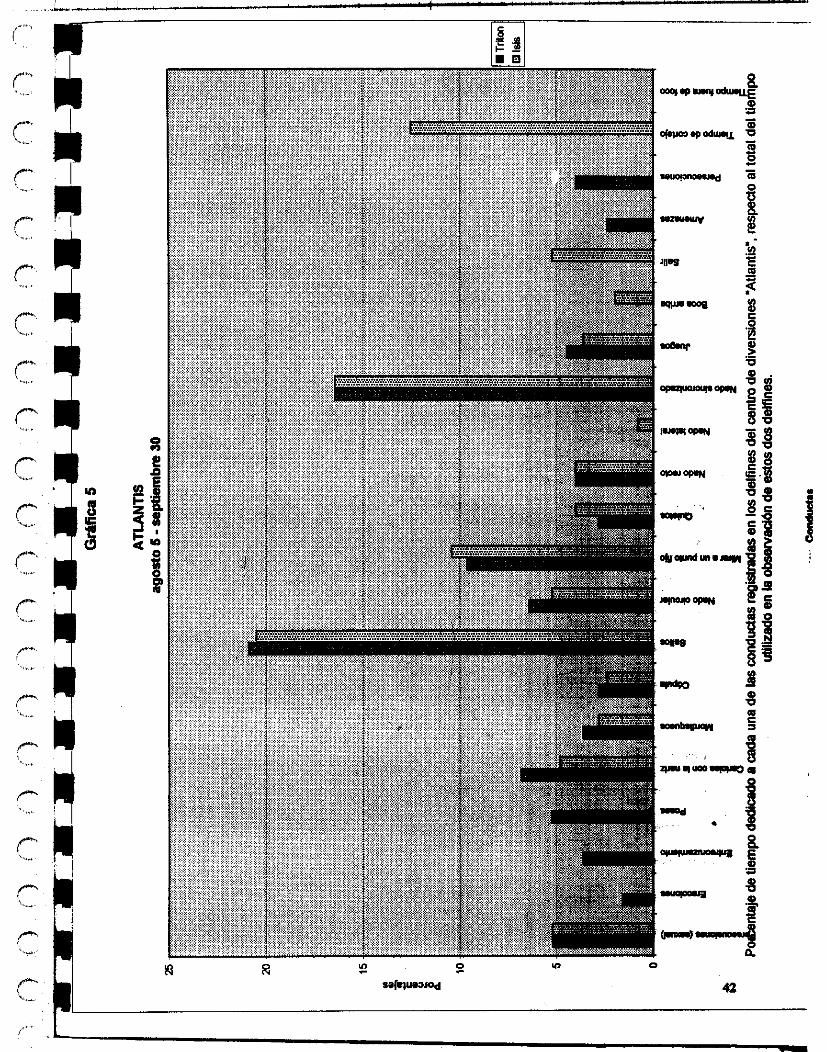
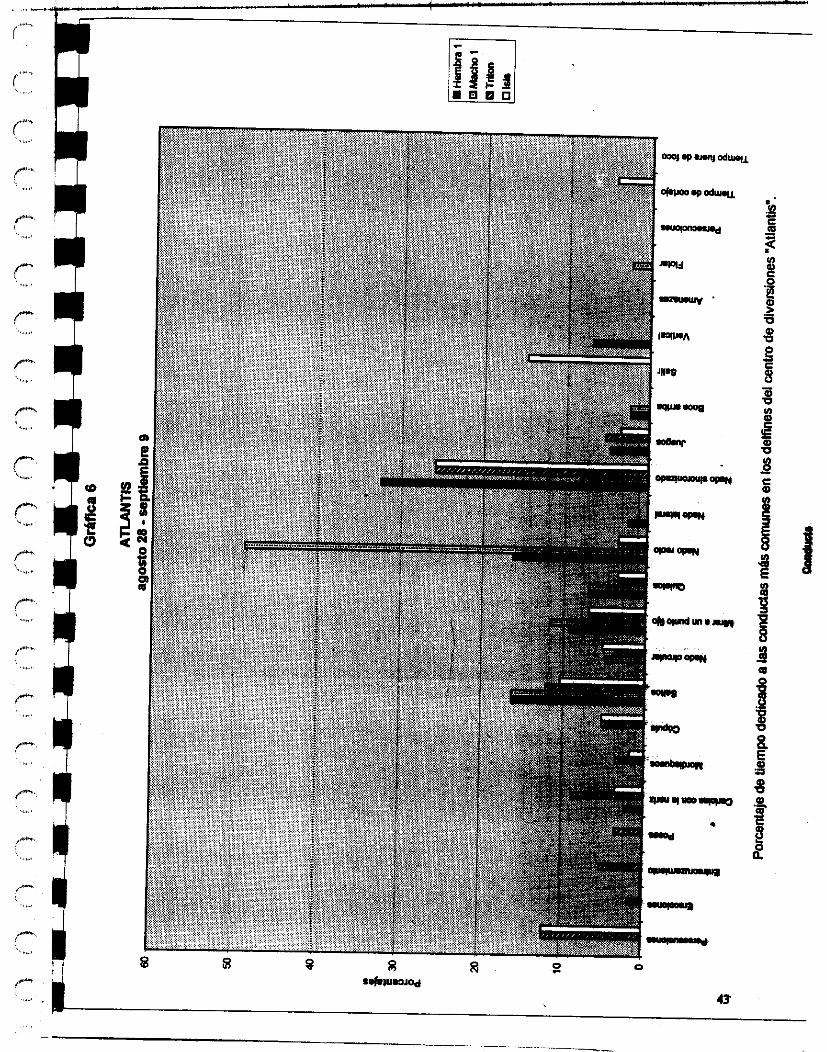
c Q

. ._
L

U .
.
4s
E
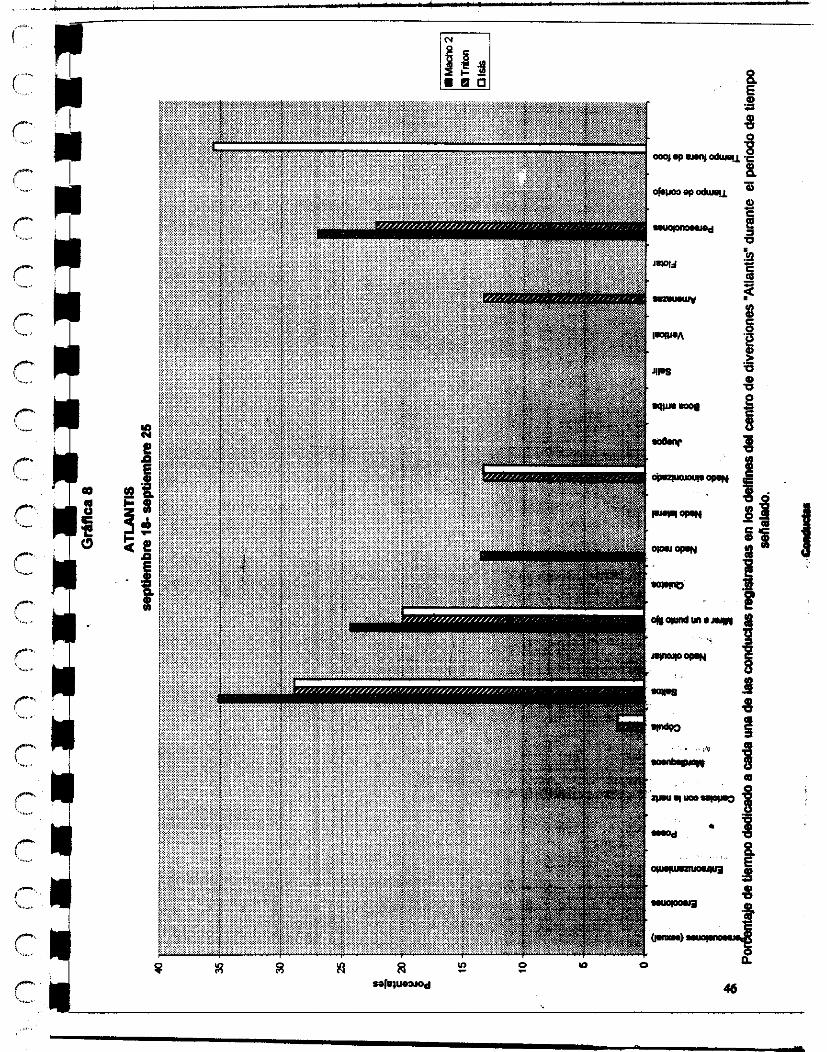
B E

8 E

TABLA No 5
JUNGLA MAGICA
AREAS REPRESENTATIVAS PARA LOS DIFERENTES COMPORTAM~ENTOS
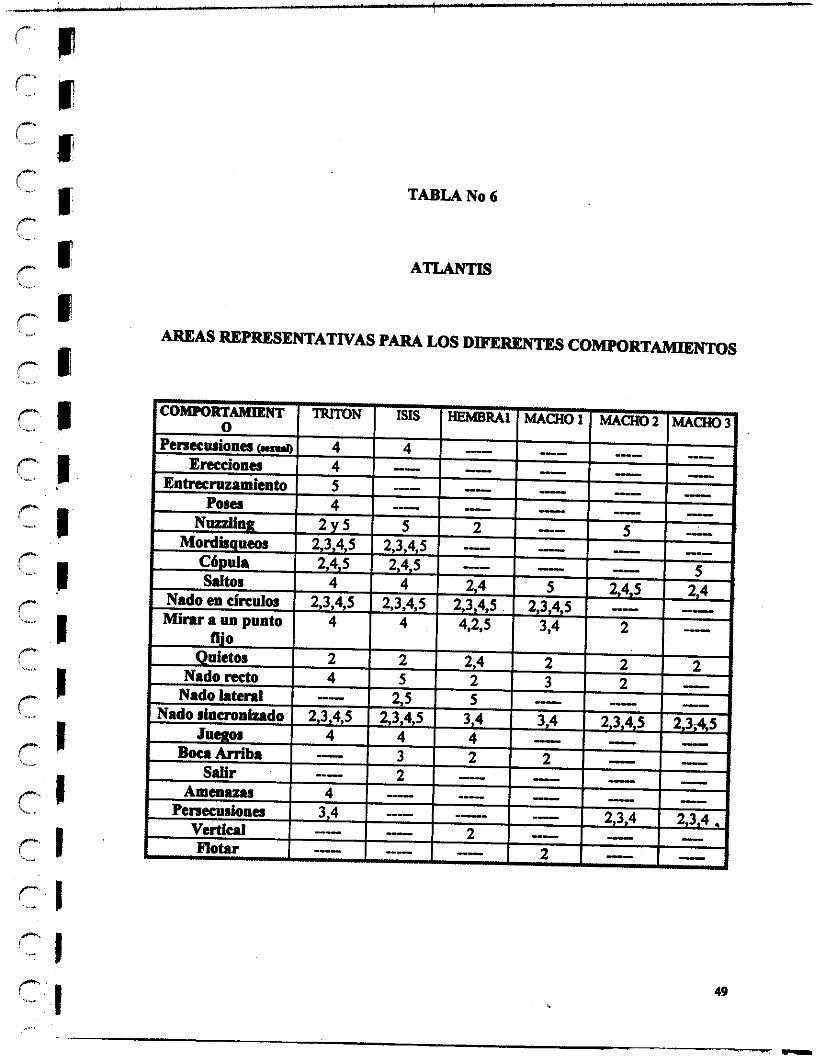
TABLA No 6
ATLANTIS
AREAS REPRESENTATIVAS PARA LOS DIFERENTES COMPORTAMIENTOS
49

.F'
P 1
" I I P
!- I- c
ANALISIS DE RESIJLTADOS
JüNGLA MAGICA
Observando la Gráfica No 1, así como los nsultados de las Tablas No 2 y No 5, que corresponden al tiempo total de observaciones realizadas a los delfines hembra Cometa y Beta y a la relacibn de los diferentes cornpottdentos con las Breas del delfinano, respectivamente, vemos que hay una clara superioridad en la utilización del territorio por parte de Cometa quien peameció más tiempo en toda d área mayor del delfinario y Beta sólo en el área 1, área que queda fuera de la vista del obJavrdor la mayor parte del tiempo, por lo que se le denominó a esta actividad, tiempo fuera de foco.
En esta misma gráfica (Gráfica No 1 y Tabla No 21 dondc se marc% todo el tiempo en general sobresalen los comportamientos de persecusión, srltoo, nado siucronhdo, mirar a un punto fijo, tiempo fuera de foco, siendo los otros aparentemente menm significativos.
Observamos también que las persecusionea aparentemente fueron muy aitas, pero al analizar el período en el cual estuvieron loa dos dcíñnas solos en el delibero sin la posterior separación-reducción de áreas (Gráfica No 2 y Tabla No 2) vemoa que estM ocuparon un bajo porcentaje de sus interacciones, siendo más significativo aquí otras conductas como snitor y mirar a un punto rijo.
En la Gráfica No 4 y Tabla No 2, se ve claramente el inaCmanto de las pvsccPrlona ya que Beta no tiene mas su espacio habitual e invade el de Comeia. Entoaces es en este perlodo de tiempo donde además surgen amenam por parte de la delfín alfa.
En cuanto al nado sincronkndo actividad también importante se observa que duraate todo el tiempo se mantuvo relativamente estable, aún después de la inchisión da la pareja de delfines cubanos.
En la M c a # 2 mirar a un punto íijo se mantiene al niv& máa elevado du las actividades realizadas por Beta y Cometa, cuando las condiciones del deiñndo eran estables, conducta que disminuye en la W c a 4, en donde la actividad mas rcprwcntativa de ese período de observaciones fue la de las persecusiones, como ya antes se dijo. La mismo ocurre con los saltos, nado recto, nado sincronizado, juegos y robar.
En la Gráfica No 4 y Tabla No 2 desaparecen algunas actividades hportautw como fue)oii nado Iiterd, giros, juego# y remolque. Y a su vez apancieron nuevos comportamrent ' osa saber, amenazas, golpear la red, flotar y sobar aparece en Beta, rewiitado da su nueva situación.

.,f.,
, / . , < I I
Con reiación a los delfines cubanos, no hubo intmicciones notablw e m estos y las hembras, al estar separados ambas parejas por una rcd (Gráfica No 2 y Tabla No 2), &Io se present6 el comportamiento de mirar a un punto fijo a trav6s de la red, en dirección a Beta y Cometa, la cual nunca saltaron.
Cabe anotar que las condiciones en Jungia Mágica fueron las más estables para los delfines, en cuanto a trato, alimentación y espacio disponible.
En la Gráñca No 5, que corresponde ai tiempo total de observaciones independientes realizadas a Isis y TritÓn, lo correspondiente ai comportamiento de cortejo no es notable por haber sido separado en sus diferentes fases. Dentro del cortejo lo mas notorio fueron las eariciaa con la nariz siendo efechiadar mas por trítón que por Isis, ai igual que el resto de actividades de este comportamiento donde el macho toma la iniciativa
Las actividades desarrolladas por más tiempo fueron las de srltos y nado rincroahdo, siguiéndole en importancia el mirar a un punto fijo, siendo las otras actividades menos relevantes para la pareja.
En la Gráñca No 6, donde se compara el comportamiento de Isis y Tntón ante la presencia del Macho 1 y la Hembra 1, vemos que el nado sincronlzado de Isis, Trit6n y Hembra 1, ocupa los valores más elevados, siguiéndole aaitos, y se mantiene el comportamienta de cortejo en la pareja inicial, aumentando las peiaceusionea sexuales y llegando a la e6pufr.
Para los cuatro individuos observados el comportamiento de mitos se mantuvo aproximadamente igual, siendo para estos igualmente importante el nado recto principalmente para el macho 1. Para este el nado shcronizado se dio muy bajo y las veces que se presentó fue con la Hembra 1 y no con Tnth e Isis.
Para macho 1 y hembra 1, las variaciones en el comportamiento no se dieron, por el contrario se ven menos enriqueciúas que para la pareja inicial de delfines Los comportamientos o bien no se presentaron o se presentaron a un bajo nivel.
En la Gráñca No 7, podemos ver muy decrwiente la variación de coductaa, aquí la Hembra 1, Tntón e Isis muestran un decremento considerable en su amplia gama de comportamiento, donde prevalecen los siltos y el mirar a un punto njo, Is siguen en importancia nado circular y quieto, para la Hembra 1 y para la pareja inicial, saltar. &o circular y lo más bajo fue mirar a un punto fijo y quictos.
La GrBfíca No 8 en cuanto a Isis y TritÓn, se observan con un alto valor da duración los saitos y el mirar a un punto fijo, y para Tntón se presentan como actividad nueva Iiis amenazas, las cuales lanza ai macho nuevo, llamado Macho 2. Aquí debido a'las interacciones aparece un nuevo ítem denominado tiempo f iera de foe0 en d cual sólo regie6 las conductas de los machos dentas que Isis permaneció fuers de la vista
Se nota una baja en las diferentes conductas y para el Macho 2, el cual pmmta mayor cantidad de saltos, siguiéndole en importancia las perseeusloner por Tntón (el delñn ai&), mirar a un punto fijo y el nado recto.

Por úitimo tenemos la W c a No 9, en la cual además de Isis y Tritón, hay dos machos presentes Macho 2 y Macho3, este úitimo fue introducido al estanque en plena sesión de obsmrac¡ón. Este día, el Último de obsewaciones, Tritón e Isis mostraron un alto valor en el nado aincronhdo, junto con los otro dos individuos, todos presentaron elevados el nivel de saltor y las persecusioner, de connotación no sexual. Se presentaron Firiciaa con la nariz hacia el ejemplar nuevo también de un alto valor y una cbpuia del delfin alfa con el Macho 3. El comportamiento que más bajo nivel presenta en esta grátia, es el de
P
'.
quietos para los 4 animales.
.
52

*^ .
F
F'
!e-. I'
, ' .I
RESULTADOS Y DISCUSIONES
Los mamífms sociales como son los delfines, poseen SeAalCs compiejas y un rico repertorio de comportamiento. Las señales de agresión que manifiestan, les son Wes para establecer y mantener jerarquías de dominancia, y contribuyen al funcionamiento del grupo y de cada uno de sus miembros. Esta vida en facilita la alimentación, comunicación, apmdizaje y respuesta adecuada a los ciclos ambientales.
Los delfines en vida libre, muestran patrones diurnos y estacionaleJ en su comportamiento. En un esnidio realuado en Australia y Argentina, se determinó que los delfines pnscntaron tres actividades comunes compuestas de comwicntos individuaice:
en las difemtes estaciones que se realizaron las observacioms. Asf como en los difenntes lugares donde se traba~6 (Brager, 1993).
En los párrafos siguientes se dan explicaciones posibles de las con<hictos de los delñnes en cautiverio, observados en este trabajo, c o ~ l a s wn la cooducts da dclfiner en libertad, así como con la de otras especies sociales.
Búsqueda de alimento, viajar y sacislizar, y estos comportsmicntos vanm en intensidad
Persecnrión y Amenazas:
Según Lo& (1971), la agresión dentro de una especie d i m una funci6a cmwmdm, y sirve por tanto para regular la distribución por un territorio, aunque esta no es la única, como lo SeAaló Darwin en 1842, quién vio que el combate rmtro rivales inachos, SC~CC~OM los mejores y más fuertes en favor de la Selección Natural, y además Sim para pmdigar mejores cuidados de lacria Y es que nunca se ha ObSaMdo que d objetivo de esta seael aniquilamiento de los congémes, aunque cuando las luchas o duelos sc desandlm en circunstancias excepcionales, no naturales ni pvistas por el d s m o de la evoiución como por ejemplo en cautiwrio, el comportamiento agresivo piede tener co- destructoras.
Las especies que no tienen inhibiciones para causar daño a los de su mUma espacie tienen gran capacidad de huida, por esto no hay presión selectiva que produzca inhibiciones para impedir que se mate a los congéneres.
En las condiciones antinaturales de la cautividad, en que el vencido no pieds huir rápidamente del vencedor, este siempre scab por matar a aquel lenta y cnielmente. Ea el capítulo dedicado a la moral y las armas del Anillo del Rey Salomón, Loreaz. 1103 muestfe como dos palomas aparentemente pacíficas que compertían la misme jada al atacarse entre sí y al no tener por donde huir termina una matando a la otra de una manera penosa
53

Esto se ha presentado en delfines en cautiverio donde a una amenaza o una seaal de agresión el otro delfín de menor jerarquía, no puede huir o alejarse y el animal amenazante arremete muchas veces contra él hasta llegar a lastimado seriamente como ocunió hace años en el Delfinario de Jungia Mggica, según reporta su entrenador, en el cuál la delfín alfa en este caso Cometa, y un macho introducido al estanque tuvieron que luchar por la jerarquía terminando con la muerte de este macho.
En Atlantis actualmente el delfin alfa es un macho: Tritón, el cuál mantiene su jerarquía amenmmdo, persiguiendo y copuiando con los individuos que frecwntemente son llevados al estanque. El individuo reportado para Atlantis como Macho 1, siempre se mantuvo a una distancia individual del resto del grupo y aquí cabe anotar que en vida libre normalmente los machos subadultos forman su grupo aparte, es un grupo denominado por muchos investigadores como de solteros, lo cual podría explicar esta exclusión apamite del grupo. Sin embargo esta exclusión desde mi punto particuiar de vista se debió al temtorialismo marcado por Trit6n en el área de todo el eshaque, así como a las dimensiones más pequeRas en compnración con el deliinario de Jungla Mágica, y al gran número de animales ahí presentes.
El grupo que se sincronizb, estaba compuesto por 2 laembras y el macho Tnth, ai Macho 1 io mantuvieron a p , y además este macho, fue lestimado por Tnt6n en su cabeza, amnneció con un hematoma el cuál two que curar el doctor y fuc SePeraQ dei grupo hacinandolo ai área 1 y separándolo de los demás por mediode una red, era un macho de tamaño más chico que Tritón de aproximadamente 4 años de edad, segiia me informó la entrenadora. Esto concuerda con la literatura donde nos reportan que el dela de mayor tamaño siempre a m d a los de menor y éi nunca es a m d a menaa que ambos animales compartan igud o similar talla corporal.
Según Noms y Dohi, de la Universidad de California en Santa Cruz, la función puede s a 1s de o r e a los miembros del grupo para enfireritarse a diferentes situaciones. Por ejemplo, las amenazas y Perscepsioncs por parte de los delñnes mayores, puede hacer que las hembras más pequeñas y los animales jóvenes se congnguea en el centro del grupo, donde se hallarán mejor ptegdos de depredadores potenciales como y tiburones (WIirsing, 1979).
En general una porción del espacio puede distribuirse de diferentes mamme, la decisión referente a que individuo poseerá dekmuda ' sección puede basarse en el momento de la llegada, como sucede en el caso de muchas aves en las que la prioridad territorial pmvoca
rango individual dentro de una jerarquía de dominio, como sucede en el caso en que la gallina doméstica divide los lugares disponibles de la perch del gallincn, &lop&, 1970).
los primeros regresos a los refugios de invierno. La otra aitmistnni . edePurdeide1
54

I 1
1 6"'
i- II
" I i
I P
*" I ("
Cuando la organizsción espacial es una función de la posesión de territorios individuales, la distancia media entre los individuas permanecerá razonablemente constante. El desarrollo de la temtorialidad y de unas dimensiones 6ptimas del territorio serh pues el resultado de la Selección Natural. Y la forma en que podemos distribuir un espacio consiste :n permitir el uso mutuo de una área amplia por parte de un gnipo de individuos, estableciendo prioridades sociales dentro del grupo para determinar el uso de cierta parte del área en un tiempo determinado. Una banda de monos aulladores (Carpenter, 1934; citado por Klopfer, 1970) puede por ejemplo, distribuirse en un área considerable sin que ninguno de sus miembros retenga la posesión de determinada . partemásqueduranteun lapso relativamente corto. En determinado momento, sin embargo, no todas las porciones del área habitada son iguaimente accesibles a todos los individuos. Un joven subordinado tiene que contentarse con los sitios que de momento no son reclamados por sus superiores (Klopfer, 1970).
Comparando la organllación social del grupo de primates con la de los deifínes se pueden encontrar muchas similitudes, anaiizando lo propuesto por Carpenter y comparándolo con nuestros resultados, vemos que en el caso de Cometa y Bets, algunas veces el delñn alfa permitió la presencia en su territorio de Beta, y hubo momentos en que al ingresar Beta al temtorio de Cometa esta arremetió contra ella Esto queda claramenie mosirah, en cuanto a los nados sincronizados, que se mantuvieron todo el tiempo, en los ndoa en círcuhw, nado recto, vertical, quietos, juegos y en mirar 8 un punto fijo. de k3.s d e s ia mayoria de estas actividades fueron realizadas simultáneamente en el área mayor o área 3 y 4.
Cometa también ocupó el área 1 y 2 donde esiaba Beta, en este lugar nunca sopnsaicaron Penecusiones, ni otras d e s de agresión, otra prueba del territorialirno marcado, Cometa si podía estar en todas partes, más- no.
Giros:
Los giros como están descritos en el Eta- son para pasar de un área de menor tamaño a una de mayor e incluso al revés, por lo cuál &lo se presentaron en Beta, l a d pasaba al área mayor ocupada por el delfín alfa
SlltOS:
La conducta se puede alterar por situaciones de tensión, conflictos, fmtmi6n y alimento, si esto no varía es inflexible, se exceden en importancia a los beneíicioa O se exhiben como resultado de una condición patológica y puede uno afirmar que los estareosi Pos son anormalidades de adaptación La localización de la conducta de esterroti 'Po piede SQ
reflejo de los factores causales, como expectación por la llegada del alimeiao envokndo conflictos crónicos o f~stración. Y algunos esiendpos temunan ' rremplezandola conducta normal.
LI-. .

En relación con los siltos, estos en vida libre, se presentan más cuando los animaia estas separados y por ello puede tener una función de comunicación, sin embargo, en
frecu~nte y rápidamente en la expectación del alimento (Mason, 1991).
Debido a que estos animales acoBtumbran a vivir en grandes grupos y espacios abiertos, pueden reaccionar ai cautiverio con estos patrones de comportamiento estemtípico. Lo primen, que se observa es un repetido y rígido movimiento que no comspondc al observado en vida libre, y luego el animal muestra patrones de movimientos espaciales rígidos. Lomiz (1971) reportó para el león marino patwncs de movimientos especiales consistentes en nadar en chuios en diferentes nitas y tuni8ndoss a lugara predccibles.
Los patrones de comportamiento estereotipado se incrementan en fnniencia e intemidad en expectación del alimento, uno de estos detectados en el d o , es el de los mitos, los cuáles son caáa vez más altos, más sucesivos y en las áreas donde iidrrcpcaian ’ con los enímaáores.
condiciones de cautiverio los asociamw a una conducta estcreoti wYaque~-
Sin embargo, el comportamiento de los delfines no es iguai al de lobos marinos y sólo algunos patrones de comporiamiento pueden ser locaiizados en el espacia Esto podría ayudar a definir si ciertas rutas de nado co- a un estenotipo comprprdghrutas de animales en vida libre con las d a s de delfines en cautiverio.
Los siltoa observados para los d e f f i Macho 1 y Hembra 1, de Atlantis, eran sucesivos y de diferentes formas, como de lado, sólo 1acabezareentranQ al aguay mi0 Scne de trucos
desesperadamente por las mismas condiciones en que se u cambio de delfinario, nuevos animales con los que intaactuer y un espacio muy limitado.
Para el estudio llevado a cabo en Atlantis no se presentb un área aacterística de temtonalismo por alguno de los animales, las únicas áress que se pucden asociar a este y otros comportamientos son las áreas doude presentan intcracción ~ o l l loa entmisdorss. Continuando con el CBSO de Atlantis, en temporada de clases, debido a la ausencia de las entrenadoras quienes por tal motivo llegaban u11 poco más tarde a alhentarlos Is conducta estemotípica de los saitm fue evidente, en las dress antes mencionadas
En el estudio de Gygax, los delfines que mostreron mayor variedad de combtas fueroa los que tenían mayor tiempo cautivos, esto se conñmib en el presents trabajo. Es c v i b que las condiciones que rodean el cstado de cautiverio wn relevantes para el desturollo o no &e actividades sociales e interrelaciones entre los organismos. En Hswai en el Sea Life perk, existe un iaboratoíio que se dedica a estudiar a los delñnes, en este lugar no sc utüiza la comida como premio, por lo que han desandlado una serie de conductes muy paiticuieres debidas a su propia evolución. Enire mis excitados, producm uaa serie de burbujas um formas de donas en la superfície del agua. Pueden emitir un c h de burbujas pequeaaS a
seguramente aprewlidos en el tiempo de estar en cautiverio, y sc m m i b t h o tan
56

.-
' R
/*"'
través de los dientes las cuáles parecen ser signos o sonidos como mensajes vocales, y hay datos a cerca de esta actividad en vida libre.
Los delfines que ingresan al delfinario, pronto adquieren o aprenden esta llamada "cultura", ya que son hábiles para aprender e imitar, prueba de ello, es que según se reporta en el laboratorio que tiene una ventana hacia los delfines, los científicos hicieron burbujas de jabón en cierta ocasión, y a los pocos minutos uno de los delfines produjo una gande que subió lentamente al área del laboratorio en forma de dona (Marten et u&iW).
En este capo el del& no intenta agradar a los humanos para que a cambio le den de comer, simplemente se encarga de adaptarse a las condiciones del cautiveno interactuanQ y comunicandose con los demas individuos, esta actividad no se rrgistrú en el esiudio dentro del etograma previsto, pero se observó unas muy pocas veces en el delfinario de Jungla MAgiCa
Nado Sinemahado y Remolque:
Es bien sabido que los delfines apoyan a los individuos enfcmios o heridos ayUdendolos a subira mph. O bien permanecen cerca de él, para ayudarlo en ceso de que lo Ilegma necesitar, sosteniéndolo y acarreaadolo para pnrenir su himdimiento. Esta ayuda no se restringe a individuos de la misma especie, sino que también puedc ser dada a especimcnes de géneros diferentes, recih nacidos muertos y se han reportado ~8908 de delñnes quo remolcan a náufragos hacia la playa (Yañez Medina, iw).
En el delfinario de Jungia Mbgica se realizó la actividad de nado coll delfines en una sesión particular de esta actividad, el delfín alfa, Cometa, se dejo motmu por un niño de aproximadamente 8 años, y lo remold por lodo el área mayor del áeifínmio, como en otras ocasiones lo había hecho, lo raro de esta ocasión es que ya no quería soltar ai niño, y cuando este se soltaba para alejarse lo volvis a cargar rápkbente en su lomo. Esto se puede interpretar como ese apoyo brindado a individuos enfermoÉi a quienes ayudan a respirar sosteniéndolos o remolcándolos, en este caso, two que intcmap el entnnedor de forma abrupta para que dejara al niño.
Estos delfines no se dejan montar por todos los nijios, al igud que no se dejan acariciar por todo el mundo. A medida que pasa el tiempo Cometa y Beta, se han acostumbrado más a su nueva situación con un área menor, (cabe anotar que aunque se les rsdujo el -pot la introducción de los delfines cubanos, esta es aún mSúr grande que el dradel delfindo de Atlantis) y cada vez más realizaa actividades de nrido rincraalpdb, así cam0 Beti ahora presenta la conducta de robar, y la realizan casi simult$neamenfe.
El agua del delfínario de Jungia Mágica, se encontró verde en las Últimas tres observaciones debido a un filtro dañado, la presencia de nuevos indiMduos y por si fuera poco las lluvias. Los animales se etlcontraban ya más tranquilos, las Perseedones habían descendido y aumentaba el nado sineronizado, ¿a qué se debía esto? En estanques de

acuacuitura, muchos peces se tranquilizan cuando el agua esta verde pues así se protegen de los depndadons y se sienten más seguros. Hay reportes de delfines en aguas turbias cercanas a la costa donde conviven tranquilamente y lopran alimentarse más fácil pues ellos gracias a la ~ i ~ i ó n localizan pronto a sus presas y la turbidez no representa ningún obstáculo. Esto podría ser una explicación pero más podria explicarlo el hecho que ya hay creado un nuevo microterritorio para Beta que esta ubicado en el subárea 3' (Gráñca No 2 y Tabla No 5) como lo reporta el estar vertical, los juegaa y otras actividades que realiza en esta zona.
Nado en Círculos:
Ridgway (1990; citado por Gym 1993) reporta que los nados en círculos son comunes en deltines recién capturados, lo dehe como reacci6n de penico, pero este compor&uniento decrece según aumenta el tiempo en cautiverio. En el estudio reaiizado por Gym y en el presente estudio, este tipo de nado io asocio al &ea del deifínano, con una forma rectangular, redondeada en las esquinas, en Atiantis, donde para que los animales puedan nadar a altas velocidades sin chocar enire ellos debian presentar este tipo de nado.
Para Jungla Mbgica este tipo de nado se observó en un mayor porcentaje en Cometa en las áreas 3 y 4 y enBeta en las áreas 1 y 2 (Tabla No 5), aparte de ser su9 dnastenitoriales, esto también se puede asociar a la forma de estas áreas, las cuáies unidas (1-2 y 3,4; ver Figura No 1) rep-tan M rcctáuguio, &ea en el que pusden nsdar a altas veiocidades sin chocar contra parad, ya que en este caso no es entre e l k pres se aicoritraban sepaiadss. En la posterior separación por medio de una red realizaron este tipo de nado de manera similar a los delfines de Atlantis. a alta velocidad en sincmnización en las dress 3 y 4 (Figura No 2)
Quieto:
El permanecer quietos en el fondo del estampe como si se acostsran sobre el piso, tal vez indique cansancio por parte de los animaics, quienes no tienen una hora exacta de dormir, ya que en la literatura se reporta que ciertos 8 ~ p o s de delfines eri vida librc preñeren descansar en horas de la mailana, mientras otros eligen horas nochum& esto varía de acuerdo a los individuos. y a la estación, no ob4tantt, como no fue tan relevante en el estudio, no arroja mayor información
. Mirar a un Punto Fijo:
El delfin macho "cubano", ai igual que la hembra por estar separados por una red del resto del delfinario, 610 @an ser vistos al Bccrclllse estos al bna dos; en dicha bna lo íiaico que io& detectar en ellos fue la actividad de mirar a un pito fijo. Esto fue rtalipdo mayor tiempo por el macho quién inclusive no respondía a las seflalcs de alimentación- entrenamiento que daba su eatremior, y permanecía mirend0 hacia el h a donde se
58
~ L

r"
F J - 1 ( r l
("I
encontraban Cometa y Beta, coincidiendo algunas veces con ellas quienes a su vez io miraban a él.
Este individuo, se encontraba aparentemente en celo, porque al parecer tmth de cortejar a la hembra cubana y esta lo rechazb mordiéndolo, cosa que se verificó al día siguiente, presentabamarcas de dientes por el área de atrás de los ojos en su cabeza, tal vez alguna de las tres hembras estaba en celo, lo cuál produjo esta reaccibn en este macho. O bien es faita de aclimatación a su nuevo habitat lo que lo mantenía en esta conduc@ aimquc esto nunca ha sido reportado para justificar estados de tensión en deltina cautivos recién capturados y recitn trasladados.
En realidad esta fue una de las conductas mh altamente regisúadas para ambos delfinatios y para casi todos los animales estudiados, los delfines son animales muy inquietos y ai no tener otras cosas con las que interactuar en el delfinario, cosas ajenas a eilos, como juguetes yío cepillos, se distraen faciimente al ~cercdfseles personas diferentes, por lo que toman esta actitud, también se asocia a la hora de alimentsCión, yaque antes de la seperación de áreas en Atlantis, Beta pemianecfa mucho tiempo mirando hacia el lugar donde el entrenador esta y, donde üecuentemcnte estaban las cubetas de alimento.
Juegas y Boen Arriba:
Es muy di8 cil poder detedar a veces que tipo de comportamiento es aprendido o bicn que tipo de comportamiento es innato a la especie. Pata un investigador de campo como Raridy Wells, esto es mucho mh sencillo ya que este señor trabaja con animales cautivos al mismo tiempo que cbn animales en vida libre (Wolkomir et al., 1%). Sin embargo, por medio del muestre0 ad libitum, se pudo detectar parte de los comporlamientos aprmadoa por los animales como en et caso del denominado Boca arriba, el cuál fue iacorporido . alshowqut presentan los delfínes y después de haberlo aprendido io incorponvon a su rutina de comportamientos. Cabe anotar que antes de esto no lo nalizabaa.
En el comportamiento de jucgoa por ejemplo hay una mezcla de mtiniU apmdidas en cautiverio, y algunas pacas que qucdan de su comportamiento innata de la especie, o bien que fueron aprendidas durante su vida libre, ya que estos animales tiemn gran capacidad de imitación.
Por ejemplo, los delfines al sur de Carolina y Georgia en los EstaQs Unih de América acodan a los peces haciéndolos saltar en el lodo donde saicn a captinarlos wn la boca Este comportamiento es transmitido cui-te, ya que las crías imitan a SUY madres. Asimismo se ha observado que las crías juegan al gato y al ratón con los peces, MLipdolos ai aire, y nxapmhbllos nuevamente.
Este Último caso es registrado dentro de nuestro etograma, pero a difaenck de ellos. los peces con los que aquí juegan, están muertos. por lo demás es igual. (Ademe de jugar wn peces io hacen con cualquier otro objeto presente en el Delñnano). Wilson sugim que

imitan a los miembros del grupo que tienen mayor éxito al capturar peces y evitar ser depredsdos.
los otros tres miembros del gnipo. para que los individuos sean capeces de dcsempefkr Esto es ventajoso para cooperar al c a m el alimento, ya que conocen los movimientos de
varios papeles en el momento de reunir a un cardumen de peces (Waising, 1979).
Muchos de los juegos presentes en Tritón e Isis en Atlantis, son apnndidos, ya que de este forma juega la entrenadora con elios, entre estos incluimos el mojar escupiendo agua por la boca, el esconderse y miir, entre otros descritos previamente en el &grama
Beta y Cometa presentan más jpesor que conservan de su vida libre, tales como: Tirar un pez y cualquier otro objeto al aire, todos inanimados, a pesar de tener muchisimo más tiempo en cautiverio que los delñnes de Atlantis.
Golpear 1. Red:
Este comportamiento lo presentó Beta en las áreas 3 y 4 indistintamente, golpeando la la& hacia el área 2. Es notable como los delfines pudiendo dar saltor tan altas en vida libre, mí como en Cautiverio, no se atreven a saltar las redes. Esto csusab alta mortalidad en delfines quienes quedaban abapados en redes de cerco para el atún. Una -pera esto es que a pesar que su vista les haga ver que dai69 de la mi hay agua, el senti& de la ecolocación les marque el ñn del medio acuoso por la presencia de esa bamra ff sia y esto les impida saltar, porque al hacerlo i r l ~ en contra de su m e z a .
Sobar y Salir:
La piel de los delfines es completamente sensitiva y ellos íimmtcmentc sc acarician ll~o al otro con las aletas de sus flancos o i6rotaa otra vez uno con otro sus mrpoa Debido a esto, en algunos tanques colocau cepillos en el fondo (cana0 pncipdo, 1994 y Ya&z Medina, 1990).
Con relación al caso particular del comporiamiento denominado aobnr, es importan& notar que se presenta en Cometa con una duración mayor, y en Beta se comicom a manif*itu sólo cuando introducen a los delfines cubanos. donde uno de los ejempians es un macho joven.
Un comportamiento instintivo no ejecutado durante mucho tiempo como por ejemploel cortejo, hace bajar el valor liminal de los estímulos que lo desencadaian. El desceiw> del umbral de los estímulos desmcadcillsdores puede llegar incluso a am, en que el movimiento instintivo de que se trate sc dispare sin estúnulo extemo comprobable, está búsqueda que consiste en el caso de Cometa en sobar su cuerpo en un borde filoso a falta del contacto corporal y de una relación sexual (can& con la nariz, entncnizami 'ento al
.
60
I I

" ' I I
/ ( ,
I I
i""
cl '"' I
p"7 I
nadar) y el discernimiento inteligente de ma situación es a lo que Wallace Craig llamó comportamiento de apetencia (Klopfk, 1970).
Aunque bien pudiera atribuirse a una falta de contacto sexual que como ya se dijo es de gran importancia para los delfines, también podemos hablar simplemente de una conducta que siempre d e n e n estos animaies que es la de mantener contacto corporai con miembros de mi grupo o masada Una conducta que se me asemeja a esta es la de isis en el delfinano de A M s , &Hembra, d sa& al d o conducta que se pment6 Varias veces, hace lo que Cometa, por que en el delfinario de Atlantis el escenario es el único borde saliente sobre el d el animal puede realizar la conducta de sobar.
Flotar:
Este comportamiento lo presentó el Macho 1 en Atlantis y Cometa y Beta en Jungla Mapica, estas Últimas después de la separaci6n-reducci6n del área, siando muy bajo y poco significativo los datos arrojados.
Comportamiento de Cortejo: Perseewiwea, Entrcemzamiento, Erefeioaa, Poscs, Carieiir cm h Nariz, Mordirqueos y Cópula.
En cuanto al comportamiento de cortejo, los delfines parecen reaiizar gahtex y cópalas durante todo el año, no obsta& presentan un claro máximo de alumbramicmtos ai iú&( a veces dos como máximo). Esto indica que hay un cambio fisiol6gico en el mrcb o en la hembra, que hace que la concepión tenga lugar en mi período limitado. Este cambisse ha documentado en forma del aumento estacid del peso de los testículos en los machos& varias especies de delñues. Un apareamiento a lo lergo de todo el a80 implica además que el cortejo puede tener otra9 connotaoiones además de la sexuai (Wihiug, 1979).
Varios investigadores han sugerido que estas iiiteracciones pueden Saw asimismo para definir y refonar las jerarquías y loa lazos sociaieu. Este argumnito se apoya en la actividad homosexuat presente por lo menos en los delfínes cautivos. Lo cud se podría definir como comportamiento homosocial, la conducta sexual es pues, muy impmhnte para la vida de los delfines. (Booth, 1989 y Wisrsing, 1979)
En las observaciones miizadas a los delfines Tnt6n e isis, se detummó ' que el comportamiento de cortejo es largo y la copulaci6n es breve ya que la d ó n cs rápida y la introducción no dura más de un minuto. Aparte al llegar a la copuleoibn. no se manifitstan Persecwiona, ni entrecrnzamiento, sólo cuando están copulando se ve que se awmodan, con un nado más tranquilo.
.
61

F I h.",. I
Datar Anexor de Ejempiares no Tnbajrdos:
En Atlantis poseen varios estanques en los que constantemente están trasladando los animales. a los cuáles no tuve acceso.
En uno de estos estanques se tenia aislado un delfin macho, qwén emiti6 sonidos agudos muy fuertes e intensos, sonidos de baja y alta intensidad llegando a iastimar los oídos, provocando alteración en los individuos focales. ya que aumentaron su níunem de siiltor, pero no respondieron vdmente de la misma manera como podría pcnsarsc.
El día 4 de Septiembre, una de las presentadofas del show se a c e d a este estampe para acariciar al mismo delfin macho, y este cerró su maDdibula con la mano de la chica adentro, la chica asustada ha16 su mano y se causó una herida cwsiderablementc profintde en la mano izquierk con los dientes del animal.
.
62

F
CONCLUSIONES FINALES Y RECOMENDACIONES
En el pnsente trabajo, se cubrió el objaivo genemi que era la observación y d i s i s del comportamiento social de los delfines (Twmps htmcafus) en condicioss de cautiverio.
El trabajo en Jungla Magia fue m8s provechoso ya que se m t ó con apoyo por parte de los entrenadores y el persoaal del parque, ademas de que se realizanm observaciones por mucho más tiempo. Aparte de esto el delfinario es el más graude para México, y esto favorece las interrelaciones entre los individuos y se pdo determinar ciprammte las jerarquías represatadas en territorialismo, el cuái en defendido por medio de Peneeusioaesy amCIIiIZI-L
En Atlantis no es posible que los delfines marquen su territorio debido al menor espacio, la forma del estanque, y al constante cambio de los animales de un estanqrie a otro, pero sin embargo esto nos muestra la versatilidad de estos animales para adeptarsc a les dikmutcs situaciones, el discernimiento inteligente ante la nueva situación Ya que el macho alfa. Tritón, utilizó mecanismos diversos para demostrar su jerarquia con los individuos nuevos del grupo, tales como amenszas, Persceiux, iiaya6pil.conelMacho3 aiingresarestc ai estanque. M u c h de estas conductas prevalecen de las conductas moatrpdas por los orgsnismos de vida libre, sólo que dependiendo de las nuevBs situacioneJ que se les presenten$ asísonutillladas.
En el estudio realizado por Gygax en 1993, se enwntr6 que existia una débil cMdencia que los dos delfines moetraban patrones de comporiamiento estacotipdo. Hay que tomar en cuenta, que es muy importante para el bienestar de los animales, las condiciones del delfinario, así como las condiciones a m b i d e s y el trato que reciben 0.1n.q interaccioDa entre los entrena&= y el persod a cargo, así como del tipo de alimentación. Las condiciones del delfinario en el estudio de Gygax eran temperatun ambiente de 18O y temperatura del agua a O', siempxe tuvieron juguetes en el delfinario, cos8 que no se presenta en los dos delfinanos del presmte estudio.
Gygax (19!23), r e W su estudio en el periodo en el que el delñnario se ciarp al púbIico y determinó que los delfines sólo moshraron rutas rigidas y nado circular en las horas de la maflena, no mostrando ma área específica los comportamientos de dta, gim5 y respiraciones. El comportamiento social fue baJo en cuanto a diiración y frmmcia y la variedad de conductas disminuía a medida que aumentaban les sesiones de cntnnsmi 'ento.
El comportamiento de nado cirrrilir presente en el estudio de Gygax y en el estudio presente lo asocio a la forma del delfinario en Atlantis, con ma forma rcdmguh, redondeada en las esquinas, donde los animales para poda nadar a albu velocidad sin chow entre ellos debían presentar este tipo de nado. En la Jimgle Mágica lo obsavamos para Beta en las breas 1 y 2 y para Cometa en la 3 y 4 formando entre ellas un nceangulo (Figura No 1).
63

Por otro lado en el presente estudio, si se presentsron conductas estereotipias con relación al alimento, ya que cuando los delfines no habían sido alimentados mostraron un único y repetido comportamiento de ritw cercanos a la zona de intemlación con los entrenadores.
Las diferencias en ambos estudios, ya que en el de Gygax fueron muy bajos, se deben a los cambios de las condiciones externas que afectan a los delfines tales como temperatura, a menor temperatura como en el estudio de Gygax (P), baja el metabolismo y por lo tanto decrece el hambre o la ansiedad de ser alimentados, por otro lado el estudio de él fue realizado en un período en que el delfinario estaba cerrado al público, este estudio se real% en período de vacaciones principalmente y los animales estaban trabejando intensamente; y por úitimo es importante sellslar el tipo de dimentact 'ón brindado a los delfines ya que una dieta surtida y sí son alimentados a sus horas, los animales no tienen por que presentar este tipo de alteraciones en su conducta, esto puede expiicar las diferencias en los resultados.
Las condiciones en Jungla wca fueron las más estables para los delfines, ya que &lo los delfines fueron pertdmh cuando ingnsan>n los delfines cubam, y sin embargo el espacio que les queda disponible a Beta y Cometa es suficientemente grande, en este lugar cuando los delfines están erüermos no se les obliga a trabajar, y su entm&or sin ser muy efusivo con ellas, les daun bum trato, el cual es corrapondido pues respoadea ad.
En Atlantis el trato de las entrenadores a los d e l b s en muy meloso, o al menos ad lo fiie en mi presencia, pero se tiene un mal manejo de los animales, ya que los umstanm cambios de organismos en los estanques no es ma situación óptima pan ellos, siento que s610 reaccionan ante ya algo inminena como en el caso del Macho 1 que sólo cuándo present6 los hematomas en la cabeza, fue scparado de los demás animales. Y la respi*rta de los organismos ante los cambios de deharios se detecta en una bejaen la variedad de las conductas ( M i c a s No 7,8 y 9, y Tablas No 3 y 4).
Es evidente que la presencia de machos y hembras en el mismo delñnario c r a nuevas expectativas en los animales y nuevas htedaciones. se pnsentb en Atlpntis un comportamiento de cortqo bien defínido, llegando a la &pul., la &paL de 2 machoJ para demostrar jerarquía, formación de dos manadas, una mixta que era el macho aifi , y dos hembras y la otra compuesta por un s610 macho, esto como conducias htemmks PrrO fue aquí en donde fue mucho más alto el comportamiento estercotípico pnsente, que consistió en saltos en las áreas de interacción con los entrendom.
En Beta y Cometa se observó que ante la presencia de un nuevo macho en el estanque, aunque separado de ellas por una red, estas mostraron inta69 hacia él y vicevrrsa Ellas presentaron en p e d un amplio rango de conductas. las d e s se manhM 'emn relativamente estables al haber cambios en el delfineno, debido a que la perhirbación W)
fue grave.
.
64

Por lo tanto se pucde afirmar que el comportamiento social y la salud mentai de los de los delfines sí está en función del sexo, edad y número de los componentes del grupo, así como del estanque, la alimentación, las tensiones debidas ai púólico (a las jornadas de trabajos) y a los entrenadores.
Y espero sinceramente que los resuitadas obtenidos en el presente estudio ayuden a mejorar las condiciones en cautiverio de estos bellos e inteiigentes animeles.
.
65
.

LITERATURA CITADA
Acevedo, G., A, 1989. Uso del área por el Tursión (TurJlopr fruncobL) en la Ensenada de La Paz, durante el verano de 1987. Tesis de Licenciatura, Biólogo Marino, Universidad Auiónoma de Baja California Sur. 1 15 p.
Aguayo, L.A., J. Urban, y L. Rojas. 1993. Delfines en aguas mexicanas. Iohrmacióo Científica y Tecnológica. S(76): 4-12.
Altmann, J., 1974. Observational study of Behavior: Sampling Methods. Behavior. 49: 227-267.
Barnes, L.G., 1984. Whales, dolphins and Porpoises: Ongin and evolution of the Ceiacea in: Mammals, Notes for a short course orgaoiacd by P. D. Gmgerieh and C. E. Badglesy. T. W. Broadhead, editor. UniversiQ of Tennesse Dept of Geologycal sciences, Studies in Geology. Pp. 139-134.
Booth, W., 1988. The Social Life ofDolphins. Science. 240 (4857): 1273-1274.
Booth, W., 1989. Unraveling the Dolphin Soap Opera Oceanus. 32 (I): 76 - 79.
Bdger, S., B. Wiirsing, A. Acevedo, & T. Henningsm 1994. Association Patams of Bottlenose Dolphins (Twsiops truncatus) in Galveston Bay, Texas. Jooroai oí Mammaloey. 75(2): 431437.
truncarus) Mahe Mammah Science. 9(4): 434438. Bdger, S., 1993. D i d and Seasonal Behavior Patterns of Bottelnose Dolphuis (Tiasiopr
Caídwell, D.K., & M, C. Caldwell. 1972. The Word of the Botuenoicd DdpaUa. J. B. Lippincott Company, Philadelphia, New Yo&, U.S.A Pp: 13-157.
Cariño, P. L. F., 1994. Los Delfines de México. flotección Ecológica o Protección Comercial?. México Desconocido. Ed. Jilguero S.A. de C. V. U.S.A. 203: 62-68.
Cousteau, J. Y., 1975. Dolphios. Doubleday and Company. inc. New Yo&, U.S.A. 405 p.
Cousteau, J. Y., 1990. El Mundo de loa 0ee;mos. Editorial Folio. México. 455 p.
Delgado, E. A., 1991. Algunos Aspectos de la Ecologia de Poblaciones de las To& (TurriopS fruneoftrr M o n a 1821) en la Laguna de TemiinoS y Sonda de campechc. México. Tesis de Licenciatw, Biólogo. ENEP Iztacala, UNAM. 148 p.
Delgado, E. A,, J. G. Ortega O. y A. Sánchez. 1994. Varamientos de Mamífm Marinos Durante Primavera y Otoiío y su Relación con la Actividad Humana en el Norte del
66

P I
c
Golfo de California Anales del htituto de Ciencias del Mar y Urnnologfa U.N.A.M. 65 (2): 287-295.
Evans, P.G.H., 1987. The Natural History of Whales and Dolphina. Facts on File Publications. 343 p.
Eibesfeldt, E., 1974. Etologfi. Editorial Omega Barcelona Pp. 19-34.
Gallo, RJ.P., 1984. InteraCci6n de Calhnes (Globicephaia mcrorynch), con la Ballena de Aleta (Baiaemptera physah) y con tursiones (Tursrops t ~ l c a t w Gilli). Aisles del instituto de Biologúi U.N.A.M. Ser. Zooiogía. 55 (1): 331-333.
Gallo, R.J.P. y H. Hugentobler. 1986. Un caso de Muate de Tonina (TVSIO~S rnmcahu) por in@h de bagre (Eagre wrw). VeaeriUrh Mexkmn. 12: 213-214.
Gallo, R.J.P., 1989. El Bonito ( E u t h i m linear>a) (soombridae) como alimento por Toninas (Tursiga9 truncda Gilli) (Delphinidie) y por Loeo MMM, (z<dopihw cuirrfontrams) (Otanidae). Anales del in8tit8ta de Biolegc U.N.A.M. Ser Zoología, q 1 ) : 125-127.
Gallo, R.J.P. y F. T. Aguilar. 1989. Trauumtismo Vertebral en el DelAn Comb Delphima delphis (CktkeX Delphiaidae) por incIUsih de espino de lnanaRaya Lk?syTsis sp (Pisces: Dasyatidae). Ve te r i ~ r i i Mexicana. 2 0 277-279. I
I I I I I I I I I 67
Gygax, L., 1993. Short Comunnicahioa Spatial Movemnt Putenis and &:brviour of two Captive Bottlenose Dolphins (Tursiop mmcd): absence of stenotyptd behaviotu 01 lack of definition?. Applied Animal Behavhr Science. 38 337-344.
Hinde, R. A,, 19áó. Animal Behaviour. A synthab of Etbdogy and Comparativa Psy~hology. MC Graw-Hill Book Company. México. 857 p.
Klopfler, P. H., 1970. L. Conducta EcológleiL 19O Edicibn en Espaaoi. Editorial ürijalbo S.A. México. 506 p.
Klopfler, P. H., 1976. introdufción ai Comportnmitato Animal Un Sigio de Etdoglr. Breviarios del Fondo de Cultura Económica McxicO. 506 p.
Lorenz, K., 1971. Sobre la Agrerin el Pretendido Mal. Siglo XXI. México. 342 p.
Lorenz, K., 1971. El Anillo del Rey SPlomón. Siglo XXI. México. 342 p. . Mann, B.E., 1973. The Drama of the Oceans. Harry U Abrames Inc. Publuhcrs, New
York. U.S.A. Pp. 17-258.
Marten, K, K. Shariff, S. Psarakos & D. White. 1996. Ring Bubbles of Dolphins. Scientfi Ameriein. Pp. 65 - 69.

Martín, P. y P. Bateson. 1991. La Mediciin del Coiaporbmhta Alianza Editorial. Madnd 237 p.
Mason, G. J., 1991. Stereotypies: a critical review. Animal Behavior. Academic Press for the amciation for the study of Animal Behaviour and the Animal Lehavior Society. Cambridge. 41: 1015-1037.
Mead, J. G., 1975. Anatomy of the nassal pasages and facial complex in the DelpWdae (Mammalia: Cetacca). Smithsonian Contributions to Zoology. 207:l-72.
McIntyre, J., 1974. Mind in the Waters. Charles Schribnci'sons, New York, U.S.A. 250 p.
Moulthametor, L., 1990. El Sueño de los Marniferos Mannos. Mundo Cintíñca Ed La Recherche. 10 (100): 292-299.
Pé~zCortéz, h4. H., G. K. S i l k y M. W. Newcomer. 1989. Tursiones (Tursiw hun~ahrp) alimentándose de un banco de lodo en el deita dei itio Coiorado. xiv ReunMn internacional pan el Eihdio de loa Mami&- Marinos, La Paz, BCS. 101 p.
Pérez-Cortéz, H., O. Maravilla-Chávez y P. Ulloa-Ramirez. 1994. ikühts SOciaLirla de Pesca. México. Pp. 57 - 76.
Peters, R., 1980 Mammalian Communication a Beiuviornl Aarlysm of Meaning. BrocWCole publishing Company. Califomin. Pp. 185-212.
Rodríguez, F. F., 1974. The Lite of the Ocean. Orbie, London Pp. 236255.
Silber, G.K., 1990. bis~butionsl relations of CebacebnS in the Northern Gulf of CdilFomia, with special reference to the Vaquita, plikecwiu sima Pb. D. lbsis. 97 p.
Wolkomir, J. y R. Woikomir. 19%. Marine Explorer. Wild We Conmtiw. Wildlife Conservation Society. New York. 68 p. .
Wtbing, B., 1979. Delfines. Investigación y Cien& Scientific American. Prensa Científica S.A. 32: 82-91.
Wiirsing, B., M. WUrsing & F. Cipriano. 1989. Doíphias in Differmt Worlds. Oceanus. 32 (1): 71-75

I i
I r " '
1 r""
L
Yabiokov, A V., V. M. ñel'kovich & V. F. Borisov. 1974. Whrka and Dolphins. Joint Publication Research Service, Arlington, Virginia, U.S.A., Pp. 1-528.
Yañez-Medina, R, 1990. Algunos Aspectos sobre la Biología de b Delfines. Universidad Autónoma Metropolitaaa iztapahpa, Seminario de investigacibo 11. MCxico. 30 p.
.


















