
Chromatographic forms of terminal deoxynucleotidyl transferase in normal lymphoid cells and in...
6
LeukemiaResearch V ol. 6, N o. 6, pp. 775 -780, 1982. 0145-2126/82/060775-06503.00/0 Printedin Great Britain. Pergamon PressLtd. CHROMATOGRAPHIC FORMS OF TERMINAL DEOXYNUCLEOTIDYL TRANSFERASE IN NORMAL LYMPHOID CELLS AND IN LEUKEMIA CELLS AT PRESENTATION AND RELAPSE* RICHARD BELL, ANNE LILLQUIST, HERBERT ABEI.SON 1 and RONALD MCCAFFREY Section of Medical Oncology, The Evans Department of Clinical Research, Boston University Medical Center, Boston, MA 02118, U.S.A. and t Sidney Farber Cancer Institute, Boston, MA 02115, U.S.A. (Received 17 May 1982. Accepted 29 June 1982) Abslraet--Normal thymocyte and bone marrow terminal deoxynucleotidyl transferase (TdT) have distinguishing characteristics by phosphocellulose chromatography in Tris buffer: marrow TdT elutes as a single peak at 0.3 M salt, whereas thymocyte TdT separates into two forms, one at 0.3 M salt and one at 0.4 M salt. Since the majority of TdT-positive acute leukemias are anatomi- cally bone marrow-derived, one would have predicted the presence of a bone marrow TdT- phosphocellulose chromatographic pattern in such patients. However, in 376 consecutive, un- treated TdT-positive acute lymphoblastic leukemias (ALL) studied by us we have invariably en- countered the two-peak thymocyte-type phosphocellulose pattern. The TdT patterns in the thymic-dependent, TdT-positive lym,~homa of AKR mice, and the TdT-positive bone marrow- derived, thymic-independent Abelson virus leukemia of Balb/C mice duplicate the situation in human ALL: a thymocyte pattern is seen in both the marrow-derived and thymus-derived diseases. This chromatographic difference between leukemia-associated and normal marrow-associated TdT in both murine and human leukemia suggested that phosphoceUulose-TdT patterns might be useful for monitoring residual marrow tumour cell burden in TdT-positive leukemia. This has not turned out to be the case: in eight patients studied in early relapse the blast cell TdT pattern was the single-peak 0.3 M species. Therefore, leukemic cell TdT cannot reliably be distinguished from normal marrow cell TdT. The chromatographic behaviour of TdT may be regulated by phos- phorylation-dephosphorylation, the 0.3 M salt peak can be converted to the 0.4 M salt species by treatment with protein kinase and ATP, and the 0.4 M species can be converted to the 0.3 M form by exposure to alkaline phosphatase. Thus, apparently anatomic compartment-specific forms of TdT may only reflect differing cellular metabolic activity. Key words." Lymphoblastic leukemia, CML blast crisis, murine leukemia, terminal deoxynucleo- tidyl transferase, relapse. INTRODUCTION TERMINAL deoxynucleotidyl transferase (TdT) is a unique DNA polymerase which is ex- pressed in the blast cells of almost all human cases of acute lymphoblastic leukemia [7, 8, 10, 11, 12]. Since TdT is normally expressed only in thymocytes and a population of primi- tive bone marrow lymphoid cells, TdT-positive leukemia cell populations have been thought to be developmentally related to one or both of these TdT-positive normal cohorts [4, 6, 9, 11]. Normal thymocyte and bone marrow TdT have distinguishing characteristics by phosphocellulose chromatography. Human bone marrow TdT elutes from phosphocellu- * Supported by Grants CA18662, A-105877, FR-128, CAL-19514, CA15187 and GM21747 from the U.S. National Institutes of Health. Ronald McCaffrey and Herbert Abelson were the recipients of Research Career Development Awards CA00099 and CA00075 from the U.S. National Institute of Health. Correspondence to: Ronald P. McCaffrey, Section of Medical Oncology, University Hospital, 75 E. Newton Street, Boston, MA 02118, U.S.A. 775
-
Upload
richard-bell -
Category
Documents
-
view
212 -
download
0
Transcript of Chromatographic forms of terminal deoxynucleotidyl transferase in normal lymphoid cells and in...

Leukemia Research V ol. 6, N o. 6, pp. 775 -780, 1982. 0145-2126/82/060775-06503.00/0 Printed in Great Britain. Pergamon Press Ltd.
C H R O M A T O G R A P H I C F O R M S O F T E R M I N A L D E O X Y N U C L E O T I D Y L T R A N S F E R A S E IN N O R M A L L Y M P H O I D C E L L S A N D IN L E U K E M I A C E L L S A T
P R E S E N T A T I O N A N D R E L A P S E *
RICHARD BELL, ANNE LILLQUIST, HERBERT ABEI.SON 1 and RONALD MCCAFFREY Section of Medical Oncology, The Evans Department of Clinical Research,
Boston University Medical Center, Boston, MA 02118, U.S.A. and t Sidney Farber Cancer Institute, Boston, MA 02115, U.S.A.
(Received 17 May 1982. Accepted 29 June 1982)
Abslraet--Normal thymocyte and bone marrow terminal deoxynucleotidyl transferase (TdT) have distinguishing characteristics by phosphocellulose chromatography in Tris buffer: marrow TdT elutes as a single peak at 0.3 M salt, whereas thymocyte TdT separates into two forms, one at 0.3 M salt and one at 0.4 M salt. Since the majority of TdT-positive acute leukemias are anatomi- cally bone marrow-derived, one would have predicted the presence of a bone marrow TdT- phosphocellulose chromatographic pattern in such patients. However, in 376 consecutive, un- treated TdT-positive acute lymphoblastic leukemias (ALL) studied by us we have invariably en- countered the two-peak thymocyte-type phosphocellulose pattern. The TdT patterns in the thymic-dependent, TdT-positive lym,~homa of AKR mice, and the TdT-positive bone marrow- derived, thymic-independent Abelson virus leukemia of Balb/C mice duplicate the situation in human ALL: a thymocyte pattern is seen in both the marrow-derived and thymus-derived diseases. This chromatographic difference between leukemia-associated and normal marrow-associated TdT in both murine and human leukemia suggested that phosphoceUulose-TdT patterns might be useful for monitoring residual marrow tumour cell burden in TdT-positive leukemia. This has not turned out to be the case: in eight patients studied in early relapse the blast cell TdT pattern was the single-peak 0.3 M species. Therefore, leukemic cell TdT cannot reliably be distinguished from normal marrow cell TdT. The chromatographic behaviour of TdT may be regulated by phos- phorylation-dephosphorylation, the 0.3 M salt peak can be converted to the 0.4 M salt species by treatment with protein kinase and ATP, and the 0.4 M species can be converted to the 0.3 M form by exposure to alkaline phosphatase. Thus, apparently anatomic compartment-specific forms of TdT may only reflect differing cellular metabolic activity.
Key words." Lymphoblastic leukemia, CML blast crisis, murine leukemia, terminal deoxynucleo- tidyl transferase, relapse.
INTRODUCTION
TERMINAL deoxynucleotidyl transferase (TdT) is a unique DNA polymerase which is ex- pressed in the blast cells of almost all human cases of acute lymphoblastic leukemia [7, 8, 10, 11, 12]. Since TdT is normally expressed only in thymocytes and a population of primi- tive bone marrow lymphoid cells, TdT-positive leukemia cell populations have been thought to be developmentally related to one or both of these TdT-positive normal cohorts [4, 6, 9, 11]. Normal thymocyte and bone marrow TdT have distinguishing characteristics by phosphocellulose chromatography. Human bone marrow TdT elutes from phosphocellu-
* Supported by Grants CA18662, A-105877, FR-128, CAL-19514, CA15187 and GM21747 from the U.S. National Institutes of Health. Ronald McCaffrey and Herbert Abelson were the recipients of Research Career Development Awards CA00099 and CA00075 from the U.S. National Institute of Health.
Correspondence to: Ronald P. McCaffrey, Section of Medical Oncology, University Hospital, 75 E. Newton Street, Boston, MA 02118, U.S.A.
775

776 RICHARD BELL et al.
lose as a single peak at 0.3 M salt. In contrast, human thymocyte TdT separates into two chromatographic forms on phosphocellulose: one form elutes at 0.3 M salt (similar to bone marrow TdT); the second elutes at 0.4 M salt [9, I0]. As the majority of human TdT- positive leukemias do not have T-cell markers and are thought to be anatomically bone marrow-derived [6], one would have predicted the occurrence of a bone marrow type TdT-phosphocellulose chromatographic pattern in the blast cells of the majority of TdY- positive human leukemias. However, this is not the case: in a large series untreated con- secutive cases of TdT-positive acute lymphoblastic leukemia (ALL), the majority of which were of the non-T, non-B type, we universally encountered the thymocyte-type TdT- phosphocellulose chromatographic pattern. In order to explore this phenomenon in more detail, we studied the blast cell TdT chromatographic characteristics of two types of TdT- positive murine lymphoblastic leukemia: the thymic-derived disease of AKR mice [13] and the bone marrow-derived, thymic-independent Abelson virus-induced disease of Balb/C mice [1].
MATERIALS AND METHODS
Reagents. Tritiated deoxyguanosine triphosphate ([3H]dGTP), sp. act. 10 Ci/mmol, was purchased from New England Nuclear, Boston, Mass.; oligodeoxyadenylic acid, chain length 12-18 residues (oligo dA12q8 ), from Collaborative Research, Waltham, Mass.; phosphocellulose, p-11, from Whatman, Kent, U.K.
Cells and tissues. AKR, Balb/C, C57BI/6J and DBA mice were obtained from Jackson Laboratories, Bar Harbor, Maine. Normal marrow and thymus were obtained from 8-week-old animals. Spontaneous AKR lymphoblastic disease was studied in retired male breeders aged 6-8 months and Abelson virus leukemia was induced in Balb/C mice [1]. Normal and leukemic cells and tissues were harvested from diseased animals immediately after killing and stored as previously described [2, 3].
Cells from normal human bone marrow and normal human thymus were obtained and studied as previously described [10]. Blast cells from a total of 452 consecutive patients with TdT-positive acute lymphoblastic leukemia were studied; of these, 376 were studied at the time of initial presentation and 76 were studied at or during relapse. The majority of patients were children under age 15; the age range was 2-70 years. Surface marker data are available to us on 190 of the 376 cases studied at first presentation. Of these 79070, or 150, had non-T, non-B phenotype; six had B-cell markers; the remainder had T-cell markers. Surface marker data were not obtained on the relapsed cases. Blast cells from 130 cases of blastic chronic myelogenous leukemia were also studied. Marker data, beyond TdT status, were not obtained on these patients.
TdTextraction and chromatography. All operations were carried out at 0-4°C. In a typical experiment a cell pellet (1-10 x i0 s cells) was suspended in 1 ml TEM buffer (0.05 M Tris-HC1, pH 7.7, 1 mM EDTA and I mM 2-mercaptoethanol) containing 10 mM phenylmethylsulphonylfluoride and 5°70 ethanol, and subjected to three cycles of rapid freezing (dry ice/acetone bath) and thawing (37°C water bath). Triton X-100 was added to a final concentration of 0.5% and after 15 min, KC1 was added to a final concentration of 1.5 M. This homogen- ate was stirred for 2 h, dialysed overnight against two 100-volume changes of TEM buffer containing 50 mM KCI. The dialysate was centrifuged at 10,1300 g for 10 min, followed by another 100,000 g centrifugation. The final supernatant was used for phosphocellulose chromatography. Phosphocellulose (Whatman P-I 1) was pre- pared by the method of Burgess [5], equilibrated with 50 mM KC1 in TEMG-BSA (TEM plus 20°7o glycerol and I mg/ml bovine serum albumin) and 0.3 x 12 cm columns were poured. Supernatant was applied to the column at a flow rate of 0.2 ml/min, which was maintained. After a 25-ml wash with 50 mM KC1 in TEMG-BSA, the column was eluted with a 60-ml gradient of 0.05-1 M KCI in TEMG-BSA buffer. Fractions of 1 ml were col- lected and assayed for TdT.
TdTassays. All assays were performed at 37°C for 30 rain in 0. l-ml total volumes using 30 ~. of the phospho- cellulose column fractions. Reaction mixtures contained 0.05 M Tris-HCl (pH 8.3), 2 mM dithiothreitol, 0.6 mM manganese chloride, 1 /ag of a polymer of deoxyadenylic acid of chain length 12-18 residues (oligo dAi2_lS) and 8.5/aM tritium-labelled deoxyguanosine triphosphate ([3H]dGTP) at asp. act. of 1200 counts/min per pmol. Reactions were terminated and processed as previously described [I0].
RESULTS
TdT in normal tissues
Bone marrow. TdT activity solubilized from normal human and murine bone marrow

Chromatographic forms of TdT 777
m
o ' - 6
Q- 4
C~
, , , 2 I i-
n,, O 60
Z 40
~ 20 " 0
m
3 M A
B 0.30 M --0.38 M
, / \ 30 40 50
.2M c
3
1
D ::L ";" 3O 4O
F R A C T I O N N U M B E R
I 50
FIG, 1. Phosphocellulose chromatography of TdT derived from normal bone marrow and thymus. Panel A: human bone marrow; panel B: human thymus; panel C: pooled g-week-old DBA mouse
marrow; panel D: 8-week-old DBA mouse thymus.
elutes from phosphocellulose as a single peak. The human enzyme elutes at 0.3 M salt, while the murine enzyme elutes at 0.2 M salt (Figs. IA and C). This murine enzyme pattern is consistent across the mouse strains studied: C57B1/6J, AKR, Balb/C and DBA all show this single peak 0.2 M enzyme. Substrate-initiator preferences, testing all four possible oligodeoxynucleotides with each of the four possible deoxynucleoside triphos- phates [10], showed that the polymerization of dGMP onto an oligo dA primer was the most efficient initiator-substrate combination for both murine and human enzymes (data not shown).
Thymocytes. TdT activity from thymocytes resolves into two peaks on phosphocellu- lose. With human thymocytes a major peak (Peak II) is seen at 0.38 M salt, and a distinct, consistent 'shoulder' peak (Peak I) at 0.3 M salt. Murine thymocytes exhibit the same two-peak pattern, with activities seen maximally at 0.2 and 0.3 M salt (Figs. 1B and D). Upon rechromatography on phosphocellulose these peaks maintain their original elution position [10]. As with the bone marrow TdT, the polymerization of dGMP onto an oligo dA primer was the most efficient initiator-substrate combination for both Peak I and Peak II derived from human and murine thymus.
TdT in leukemia ceils
Human cells. All 452 cases of acute lymphoblastic leukemia included in this study were TdT-positive, as were 42 of the 130 cases of blastic chronic myelogenous leukemia. TdT solubilized from the 376 newly diagnosed cases of acute lymphoblastic leukemia in- variably exhibited the 'thymocyte' phosphocellulose chromatographic profile. The elution pattern shown in Fig. 2A is typical of such cases, and is obtained with blast cells from either the bone marrow or peripheral blood. Two peaks elute from the column: Peak I, at 0.3 M salt, and Peak II, at approx. 0.4 M salt. In a few instances, virtually all the activity was confined to the Peak II area. No examples of activity limited to the Peak I area were

778 RICHARD BELL etal.
? 0 ,e,-
X
tl,
Q ud I- ne 0 0.
o
Z a.
CD "0
:Z
A 0.3 M .39 M
100 j ~
5 0 - -
1
B .29 M .38 M
1 5 0 -
100
\ I
20 30 40 I
50
F R A C T I O N N U M B E R
FIG. 2. Phosphocellulose chromatography of TdT from human leukemic blast cells. Panel A: bone marrow blasts f rom a case of childhood c o m m o n ALL; panel B: circulating blasts f rom a case of TdT-positive CML blast crisis. The data shown are for a 42-year-old woman with lymphoid blast
crisis.
encountered in chromatograms from the 376 cases studied, including those with T-cell markers. This double-peak pattern does not appear to be an artefact of initial chromato- graphy. The isolated peaks maintain chromatographic identity on rechromatography [10]. Similarly, in the 42 TdT-positive cases of blastic chronic myelogenous leukemia a 'thymocyte' pattern has been invariably observed. A representative chromatogram for a patient from this group of 42 is shown in Fig. 2B.
The majority of the 76 cases of relapsed acute lymphoblastic leukemia also had this 'thymocyte' TdT-phosphocellulose pattern. However, a subgroup of eight of these 76 had a single-peak phosphocellulose pattern in the 0.3 M salt (bone marrow) area. The pattern for one such patient is shown in Fig. 3. These eight cases were all studied (using bone
Q LU I--
a,. O a. ? 60 n- o 0 " ~ x 40
~;t~ 20 O -r
31 M 4 M
2 0 3 0 4 0
FRACTION NUMBER
FIG. 3. Phosphocellulose chromatography of TdT from bone marrow blast cells f rom a case of ALL in early relapse. The data shown are for a 7-year-old girl with c o m m o n ALL.
Chromatographic forms of TdT 779
marrow blast cells) early on in relapse, which was diagnosed during routine monitoring of bone marrow status while maintenance chemotherapy was being administered. Seven of these eight had the 'common' phenotype on original presentation; the eighth case had no marker studies performed at presentation.
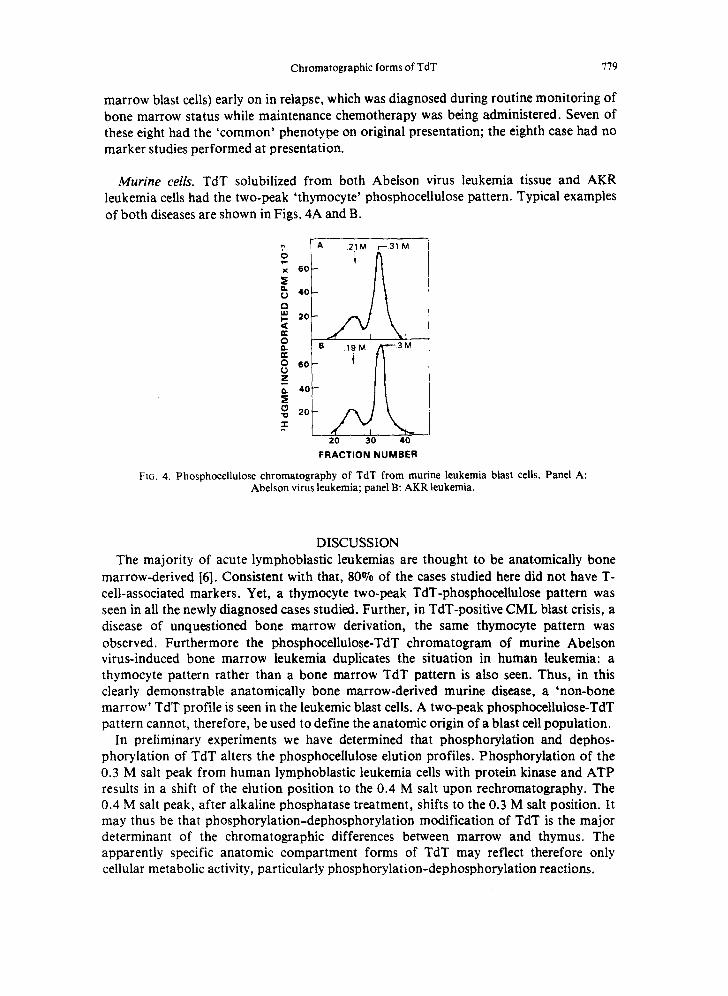
Murine cells. TdT solubilized from both Abelson virus leukemia tissue and AKR leukemia cells had the two-peak 'thymocyte' phosphocellulose pattern. Typical examples of both diseases are shown in Figs. 4A and B.
O m,- x : [
Q U,J I,- ,<
o er O
z m
r.9 '1=
A .21M r---,31M
4O
20
I
60 B . ~
40
20
20 30 40 FRACTION NUMBER
FIG. 4. Phosphocellulose chromatography of TdT from munne leukemia blast cells. Panel A: Abelson virus leukemia; panel B: AKR leukemia.
DISCUSSION The majority of acute lymphoblastic leukemias are thought to be anatomically bone
marrow-derived [6]. Consistent with that, 80% of the cases studied here did not have T- cell-associated markers. Yet, a thymocyte two-peak TdT-phosphocellulose pattern was seen in all the newly diagnosed cases studied. Further, in TdT-positive CML blast crisis, a disease of unquestioned bone marrow derivation, the same thymocyte pattern was observed. Furthermore the phosphocellulose-TdT chromatogram of murine Abelson virus-induced bone marrow leukemia duplicates the situation in human leukemia: a thymocyte pattern rather than a bone marrow TdT pattern is also seen. Thus, in this clearly demonstrable anatomically bone marrow-derived murine disease, a 'non-bone marrow' TdT profile is seen in the leukemic blast cells. A two-peak phosphocellulose-TdT pattern cannot, therefore, be used to define the anatomic origin of a blast cell population.
In preliminary experiments we have determined that phosphorylation and dephos- phorylation of TdT alters the phosphocellulose elution profiles. Phosphorylation of the 0.3 M salt peak from human lymphoblastic leukemia cells with protein kinase and ATP results in a shift of the elution position to the 0.4 M salt upon rechromatography. The 0.4 M salt peak, after alkaline phosphatase treatment, shifts to the 0.3 M salt position. It may thus he that phosphorylation-dephosphorylation modification of TdT is the major determinant of the chromatographic differences between marrow and thymus. The apparently specific anatomic compartment forms of TdT may reflect therefore only cellular metabolic activity, particularly phosphorylation-dephosphorylation reactions.

780 RICHARD BELL etaL
Bone marrow-derived TdT-positive leukemic cells may thus contain the two-peak chromatographic forms of TdT on the basis of such phosphorylatiori-dephosphorylation reactions. In the eight 'early' relapse patients we studied, whose blast cells contained only the single-peak, normal marrow-type TdT, the TdT chromatogram would be explainable on this basis. Although we have not made direct observations on this, presumably these eight patients, if studied sequentially, over time, would evolve into a two-peak thymocyte- type TdT pattern. Such two-peak patterns were in fact seen in the 68 other relapsed patients, whose cells were obtained in established relapse. It is possible that the main- tenance chemotherapy which these eight patients were receiving may somehow be impli- cated in this altered phosphocellulose chromatography elution profile. Whatever the explanation for the observation of a single-peak profile in the eight patients, the finding of a 'non-leukemic' single-peak profile in these patients in early relapse compromises the potential use of qualitative bone marrow TdT-phosphocellulose elution profiles in the monitoring of the remission status of TdT-positive leukemias.
REFERENCES 1. ABELSON H. T. & RABSTEIN L. S. (1970) Virus-induced thymic independent disease in man. Cancer Res. 30,
2213. 2. BALTIMORE D., SILVERSTONE A. E., HARRISON Z. A. & MCCAFFREY R. (1977) Specialized DNA polymerases
in lymphoid cells. Cold Spring Harb. Syrup. quant. Biol. 41, 63. 3. BALTIMORE O., SILVERSTONE A. E., KUNG P. C., HARRISON T. A. & MCCAFEREY R. (1976) What cells con-
tain terminal deoxynucleotidyl transferase? In The Generation o f Antibody Diversity: A New Look. (CuNNINGHAM A. J., Ed.), p. 21. Academic Press, New York.
4. BOLLUM F. J. (1979) Terminal deoxynucleotidyl transferase as a hematopoietic cell marker. Blood54, 1203. 5. BURGESS R. R. (1969) A new method for the large scale purification of Escherichia coli deoxyribonucleic
acid-dependent ribonucleic acid polymerase. J. biol. Chem. 244, 6160. 6. GREAVES M. F. (1979) Cell surface characteristics of human leukemic cells. In Essays in Biochemistry
(CAMPBELL P. N. ,~ MARSHALL R. O., Eds.). Academic Press, New York. 7. KALWINSKY D. K., WEATHERRED W. H., DAHL G. V., BOWMAN W. P., MELVIN S. L., COLEMAN M. S. &
BOLLUM F. J. (1981) Clinical utility of initial terminal deoxynucleotidyl transferase determinations in child- hood acute leukemias. Cancer Res. 41, 2877.
8. KUNG P. C., LONC J. C., MCCAFFREV R. P., RATLIFF R. L., HARRISON T. A. & BALTIMORE D. (1978) Terminal deoxynucleotidyl transferase in the diagnosis of leukemia and malignant lymphoma. A m. J. Med. 64, 788.
9. MCCAFFREY R. P., HARRISON T. A., KUNG P. C., PARt(MAN R., SILVERSTONE A. E. & BALTIMORE, D. (1976) Terminal deoxynucleotidyl transferase in normal and neoplastic hematopoietic cells. In Modern Trends in Human Leukemia HNETH R., Ed.). Lehmanns Verlag, Munich, F.R.G.
10. MCCAFFREV R. P., HARRISON T. A., PARKMAn R. & BALTIMORE D. (1975) Terminal deoxynucleotidyl trans- ferase activity in human leukemic ceils and in normal human thymocytes. NewEngl. J. Med. 292, 775.
11. MCCAFFREV R. P., SMOLER D. F. & BALTIMORE D. (1973)Terminal deoxynucleotidyl transferase in a case of childhood acute lymphoblastic leukemia. Proc. natn. Acad. ScL U.S.A. 70, 521.
12. MARKS S. M., McCAFFREY R. P., ROSEI'rrHAL D. S. & MOLONEY W. C. (1978) Blastic transformation in chronic myelogenous leukemia: experience with 50 patients. Med. Ped. Oncol. 4, 159.
13. SIEGLER R. & RICH M. A. (1966) Influence of thymic mass on murine viral leukogenesis. Nature, Lond. 209, 313.







![Therapeutics A Novel Small-Molecule Inhibitor of Protein ...transferase–mediated dUTP nick end labeling (TUNEL)– positive cells], and abrogated the expression of several NF-κB–dependent](https://static.fdocuments.net/doc/165x107/6118340d0013286be8293532/therapeutics-a-novel-small-molecule-inhibitor-of-protein-transferaseamediated.jpg)










