
Chlamydomonas reinhardtii のののの葉緑体葉緑体 …...
23
Chlamydomonas reinhardtii Chlamydomonas reinhardtii Chlamydomonas reinhardtii Chlamydomonas reinhardtii の葉緑体 葉緑体 葉緑体 葉緑体での での での でのコラーゲンタンパク コラーゲンタンパク コラーゲンタンパク コラーゲンタンパク発現系 発現系 発現系 発現系の構築 構築 構築 構築 Construction of the collagen protein expression system in Construction of the collagen protein expression system in Construction of the collagen protein expression system in Construction of the collagen protein expression system in Chlamydomonas Chlamydomonas Chlamydomonas Chlamydomonas reinhardtii reinhardtii reinhardtii reinhardtii chloroplast chloroplast chloroplast chloroplast 物質・環境システム工学コース 1075018 大濱 真
Transcript of Chlamydomonas reinhardtii のののの葉緑体葉緑体 …...

Chlamydomonas reinhardtiiChlamydomonas reinhardtiiChlamydomonas reinhardtiiChlamydomonas reinhardtiiのののの葉緑体葉緑体葉緑体葉緑体でのでのでのでのコラーゲンタンパクコラーゲンタンパクコラーゲンタンパクコラーゲンタンパク発現系発現系発現系発現系のののの構築構築構築構築
Construction of the collagen protein expression system in Construction of the collagen protein expression system in Construction of the collagen protein expression system in Construction of the collagen protein expression system in Chlamydomonas Chlamydomonas Chlamydomonas Chlamydomonas
reinhardtiireinhardtiireinhardtiireinhardtii chloroplast chloroplast chloroplast chloroplast
物質・環境システム工学コース 1075018 大濱 真

2
目次目次目次目次
1.要旨 3 頁
2.序章 4 頁
3.実験材料と手法 5-10頁
4.結果と考察 10-21頁
5.参考文献 22-23頁

3
要旨要旨要旨要旨
植物細胞を有用タンパク質発現の場として使用する利点として、①培地のコストが安い、②ヒトに感染
するウィルスが無く安全性の確保が容易、などが上げられる。本研究では、陸上植物よりも増殖速度が速
く培養が容易な藻類をタンパク質発現の場として利用する為の系の構築を目標とする。一般的には、外来
遺伝子は核ゲノムに導入されるが、より大量のタンパク発現を実現するため、葉緑体への外来遺伝子の導
入を行った。葉緑体を用いることの優位性として、①ゲノムのコピー数が多い、②RNAi などの外来遺伝子
の発現を阻害する機構がない、などがある。
単細胞緑藻クラミドモナス(Chlamydomonas reinhardtii)は、古くから遺伝子研究に用いられ、これまで
に、核ゲノム、葉緑体、ミトコンドリアなどの全塩基配列が決定しているモデル生物である。また、藻類
の中で唯一外来遺伝子の導入が可能な藻である。本研究では、藻類における機能性タンパク質発現のモデ
ルケースとして、クラミドモナスの葉緑体において、ヒト由来コラーゲンα2type1タンパクの発現を試み
た。コラーゲン Type1 は細胞内で、皮膚、骨、腱の主成分として存在している。コラーゲンタンパクは分
子量約130万のペプチド鎖3本で構成される。生体内でコラーゲンは、3本鎖のうち2本はα1、残り
1本をα2という異なるペプチド鎖で構成されており、この3本鎖が 3 重螺旋の構造をとり存在すること
が知られている。しかし、これまでの研究で、α2鎖だけでも3重螺旋構造をとることは可能であり、コ
ラーゲンとしての機能を持つことが確認されている。発現用コンストラクトには強力な転写活性が期待で
きる内在遺伝子 rbcL の rbcL3’UTR部位を利用した。また、遺伝子導入確認のマーカーとしてスペクチノ
マイシン耐性賦与遺伝子(aadA)を付加した。作成したコンストラクトをボンバートメント法でクラミドモ
ナスに導入し、相同組み換えによって葉緑体に遺伝子が導入されたと思われる株を、スペクチノマイシン
耐性をマーカーとして選抜した。得られた株に対して、コンストラクト導入部位の確認、コラーゲンα
2type1用ポリクローナル抗体(Takara bio.委託生産)を用いた抗原抗体反応によるコラーゲンタンパクの
発現の解析を行った。その結果、aadA とコラーゲン遺伝子の存在は確認できたが、目的としていた葉緑体
の rbcLへ導入されていないことが解った。また、タンパク質の解析では、ポリクローナル抗体に反応する
タンパク質の存在は確認できたが、その分子量は目的産物の分子量よりも小さいものだった。発現されて
いると思われるタンパク質について、より確実な発現を検証するために、コラーゲンタンパクの単離を試
みたが、コラーゲンタンパクのみを回収することできなかった。形質転換体のタンパク質発現の不安定さ、
薬剤への耐性などから、今回得られた形質転換体では、コンストラクトが葉緑体ではなく偶然核に導入さ
れ、1度は核内のプロモーターを利用してコラーゲンタンパクが発現されたが、その後 PTGS などの外来遺
伝子阻害機構の影響を受けて発現が停止したと考えられる。

4
序章序章序章序章
現在、植物を有用タンパク質生産の場として利用するための基礎研究が増加している。植物は、光合成
により二酸化炭素と水から糖類を生産し、アミノ酸を自給できるため、動物細胞でタンパク質を生産させ
る場合に比べて、コストを抑えることが可能である。実際に、ヒト抗体(mAb)を各種の細胞で生産させた
場合のコストは、動物細胞では\15,000/g であるのに対して、植物細胞では\5/g と非常に安価であるとい
う試算報告がある(Dove A. 2002)。また、植物では動物やヒトに感染するようなウィルスが存在しないの
で生産されたタンパク質の安全性が高く、医療などの分野でも広く利用できると考えられる。植物細胞で
のタンパク生産の中で、葉緑体を用いた外来遺伝子が注目されている。葉緑体はシアノバクテリアの共生
により進化したものだと言われており、遺伝子の発現系や制御系は原核生物に近いと考えられる。よって
真核生物の核でみられるPTSG などの外来遺伝子発現抑制機構が無い、あるいは未発達であると考えられ、
外来遺伝子の安定した発現が期待できる。また、原核生物の遺伝子工学的知識を応用し、タンパク質の生
産をさせることも可能であると考えられる。
本研究では、陸上生物よりも圧倒的に速い増殖速度を持ち培養も比較的簡単な、藻類を有用タンパク質
の生産の場として用いる。中でも、古くから研究が行われている単細胞緑藻 Chlamydomonas reinhardtii
(以下 C. reinhardtii)の葉緑体を有用タンパク質の生産場所として選択した。C. reinhardtiiはモデル
生物として研究されており、核、葉緑体、ミトコンドリアへの外来遺伝子導入法が確立されている。また、
核、葉緑体、ミトコンドリアなどの全塩基配列が決定しており、多大なリソースの蓄積もある。C.
reinhardtiiの葉緑体の特徴として、①片親遺伝でmt+株の遺伝子のみが次世代に受け継がれていく、②ゲ
ノムのコピー数が多い(100コピー)、③細胞1個体に葉緑体は1つで、細胞体積の大半を占める、④遺伝
子組み換えが相同組み換えによって起こり、目的部位への遺伝子導入が可能、などがあげられる。
C. reinhardtiiで実際に発現が確認されている外来遺伝子として、スペクチノマイシン/ストレプトマイ
シン耐性賦与遺伝子 aadA(Goldshcmidt-Clermont M.1991)、βガラクトシダーゼ(GUS)タンパク質をコー
ドする uidA、蛍光タンパク質をコードする GFP(Franklin S.et al.2002)がある。本研究では、藻類にお
ける機能性タンパク質発現のモデルケースとして C. reinhardtiiの葉緑体を用いて、バイオマテリアルや
化粧品などとしての利用が期待できる有用タンパクであるコラーゲンタンパクα2type1の発現を試みた。
コラーゲンタンパクは、多種多様で 1(type1)~19(type19)まで分類されるが、中でももっとも一般
的なtype1を発現させるタンパクとして選択した。Type1は皮膚、骨、腱などの主成分で、細胞内マトリッ
クスでは線維状で存在している。コラーゲンは長さが約 300 ナノメートル、直径が 1.5 ナノメートルの棒
状の分子であり、分子量約130万の3本の鎖で構成される。3本鎖のうち2本はα1、残り1本をα2とい
う、異なるペプチド鎖がそれぞれ左巻きの螺旋を巻きながら、右巻きの 3 重螺旋構造を作る。コラーゲン
タンパクの主な部分はグリシン-x-y の繰り返し配列で構成され、x にはプロリンなど、yにはヒドロキシ
プロリンなどがからなる。また、コラーゲンタンパクの繰り返し配列の前後にはそれぞれpre、proのペプ
チドがあり、preは細胞外輸送、proはコラーゲン分子の螺旋化に関係しており、細胞外マトリックスに出
た後切断され、凝集して線維状になる。(藤本 大三郎.1998、1999)これまでに、タバコやイーストで発
現が試みられ、タバコでは全タンパク1kg中0.1g程度(Ruggiero F.2000)、イーストでは全タンパク1kg
中 3~4g程度(Olsen DR.2001)のコラーゲンタンパクが得られたという報告がある。

5
実験材料実験材料実験材料実験材料とととと手法手法手法手法
C. reinhardtiiC. reinhardtiiC. reinhardtiiC. reinhardtii株株株株およびおよびおよびおよび培養条件培養条件培養条件培養条件
本研究に用いた、C. reinhardtiiは野生株であるCC-125mt+と細胞壁欠損株であるCW-15(Chlamydomonas
Genetic Center [c/o Dr. Elizabeth Harris,Department of Botany, Duke University, Durham, NC 27706,
USA]譲渡)である。培養は、TAP 液体培地(Elizabeth H Harris.1989)および TAP 寒天培地(1%寒天)を
用い25℃、3000LUX 下で明暗周期23.5 時間(明)0.5 時間(暗)の条件で行った。液体培地の場合は同条
件下で、125rpmで振盪培養を行った。
形質転換用形質転換用形質転換用形質転換用コンストラクトコンストラクトコンストラクトコンストラクトのののの作製作製作製作製
COL1A2LCOL1A2LCOL1A2LCOL1A2L
本研究で使用するのはヒトコラーゲンα2type1の cDNA(5411bp)を鋳型とし、前部telo配列を含む繰
り返し部分約3.3kb(図1a)をXbaⅠサイト付加するプライマー(COL1A2-primer1)とNotⅠサイト付加す
るプライマー(COL1A2-primer2)を用いてPCR反応(94℃1分→[94℃30秒→45℃30秒→72℃2分]×5サ
イクル→[94℃30秒→62℃30秒→72℃2分]×25サイクル→72℃2分→4℃∞)で増幅し、できた産物を0.8%
アガロースゲル電気泳動にて確認した。アガロースゲルから目的の産物を、Ultra-free DA(Millipore)
にを用いて、エタノール沈殿で濃縮した。濃縮したDNAをpT7Blue T-vector(Novagen)のEcoRVサイトに
Ligation Kit Ver2 SolutionⅠ(Takara bio inc.)を用いてライゲーションし、Competent high DH5α(Toyobo)
を用いてクローニングを行った後、シークケンスによって塩基配列の確認をした。ここで得られたプラス
ミドをCOL1A2Lと名付けた。
COL1A2SCOL1A2SCOL1A2SCOL1A2S
前部telo配列を含む繰り返し部分の1/3程度の短い配列約1.1kb(図1a)を持つコンストラクトも作製
した。ヒトコラーゲンα2TypeⅠのcDNAを鋳型として、XbaⅠサイト付加するプライマー(COL1A2-primer1)
と NotⅠサイト付加するプライマー(COL1A2-primer3)を用いてPCR 反応(94℃1分→[94℃30 秒→48℃30
秒→72℃2分]×5サイクル→[94℃30秒→62℃30秒→72℃2分]×25サイクル→72℃2分→4℃∞)を行い、
できた産物を0.8%アガロース電気泳動にて確認した。その産物を、Ultra-free DAにより目的のバンドを
抽出後、エタノール沈殿で濃縮した。濃縮したDNAをpT7Blue T-vectorのEcoRVサイトにLigation Kit Ver2
SolutionⅠを用いてライゲーションし、Competent high DH5α を用いてクローニングを行った後、シーク
ケンスによって塩基配列の確認をした。ここで得られたプラスミドをCOL1A2Sと名付けた。
選択選択選択選択マーカーマーカーマーカーマーカー
遺伝子導入の確認をするための選択マーカーとして、Goldschmit-Clermont M.より供与された、スペク
チノマイシン耐性賦与遺伝子を持つ aadA(oglycoside adenine transferase gene)カセットpUC-atpX-AAD
の一部または全域を利用した。(Goldschmit-Clermont M.1991)
コンストラクトコンストラクトコンストラクトコンストラクト atpAatpAatpAatpA----COL1A2LCOL1A2LCOL1A2LCOL1A2L、、、、atpAatpAatpAatpA----COL1A2SCOL1A2SCOL1A2SCOL1A2S
atpA遺伝子のプロモーターを利用した形質転換用コンストラクトは、pCV2ベクター(西村 俊祐.2004)
を貰い受けて改良を行った。(図1b)まず、pCV2ベクターを制限酵素XbaⅠとNotⅠで切断した。切断した
DNA断片を0.8%アガロースゲルで電気泳動を行い、目的のバンドを切り出したゲルをUltra-free DAを使
用して5000gで 10分間遠心して抽出後、エタノール沈殿で濃縮した(pCV2-reと命名)。プラスミドCOL1A2L

6
および COL1A2S を制限酵素 XbaⅠと NotⅠで切断した後、0.8%アガロースゲルで電気泳動を行い、目的の
バンドを切り出したゲルを Ultra-free DA により抽出後、エタノール沈殿で濃縮した。得られた DNA 断片
をpCV2-reベクターのXbaⅠサイトとNotⅠサイトにLigation Kit Ver2 SolutionⅠで挿入した。得られた
コンストラクトをatpA-COL1A2L、atpA-COL1A2S(図1c)と名付けた
rbcLrbcLrbcLrbcL----COL1A2SCOL1A2SCOL1A2SCOL1A2S
rbcL遺伝子を用いた形質転換用コンストラクトは、atpA-COL1A2Sを元にして作製した。rbcL遺伝子の終
止コドンから下流を中心に、相同組み換えを目的とした遺伝子コード領域をそれぞれ、rbcL 相同領域上流
60bpを付加する合成オリゴマー(rbcLT/F)、rbcL相同領域下流60bpを付加する合成オリゴマー(rbcLT/R)
を用いて、テールPCR(94℃1分→[94℃30秒→40℃30秒→72℃1分]×5サイクル→94℃30秒、Takara bio.
Pyrobest DNAポリメラーゼ使用)でaadA-COL1A2Sの前後に相同領域を付加した。その後、PCR Purification
Kit(Qiagen)を使用して精製し、回収されたPCR産物を鋳型にして付加したテール部を含むaadA-COL1A2S
領域を増幅するプライマーセット(rbcLT/P/F2、rbcLT/P/R2)を用いてPCR(94℃1分→[94℃30秒→46℃
30 秒→72℃2 分]×30 サイクル→72℃2 分)を行い、配列を含む全域を増幅した。PCR Purification Kit
で PCR 産物を回収後、回収した産物を鋳型にして更にテール部を付加するための合成オリゴマーセット
(rbcL 相同領域上流 70p:rbcLT/F2、rbcL 相同領域下流 68bp:rbcLT/R2)でテール PCR(94℃1 分→[94℃
30 秒→46℃30 秒→72℃2 分]×5 サイクル→94℃30 秒、Pyrobest DNA ポリメラーゼ使用)で貼り付けた。
PCR産物をPCR Purification Kitで回収し、テールを含む配列を増幅するプライマーセット(rbcLT/P/F3、
rbcLT/P/R3)を用い、PCR(94℃1分→[94℃30秒→50℃30秒→72℃2分]×30サイクル→72℃2分、Pyrobest
DNAポリメラーゼ使用、反応系100μlで5本分)を行った。PCR Purification Kitで回収後、エタノール
沈殿を行い濃縮した。得られたコンストラクトをrbcL-COL1A2S(図1d)と名付けた。

7
atpA5’ aadA CTB
pT7-Blue
XbaⅠ NotⅠ
rbcL3’
b.b.b.b.
a. a. a. a.
pre c-pro c-telo Collagen α2 typeⅠ n-telo n-pro
COL1A2S
COL1A2L
COL1A2-primer1 COL1A2-primer2 COL1A2-primer3
c. c. c. c.
atpA5’ aadA
COL1A2L
pT7-Blue
XbaⅠ NotⅠ
rbcL3’ COL1A2S
rbcLT/R2
d. d. d. d.
atpA5’ aadA COL1A2S
pT7-Blue
XbaⅠ NotⅠ
rbcL3’
rbcLT/F rbcLT/R
aadA COL1A2S
rbcLT/F2
rbcL相同組み換え領域
rbcLT/P/F3 rbcLT/P/R3
aadA COL1A2S
図1. 形質転換用コンストラクト
a.Collagen α2typeⅠ、b.pCV2ベクター、c. atpA-COL1A2L、atpA-COL1A2S、d.rbcL-COL1A2S

8
形質転換形質転換形質転換形質転換
葉緑体の形質転換には PDS-1000/He システム(Bio-Rad)(パーティクルガン法)(Goldschmit-Clermont
M.1991)を使用した。DNA コンストラクト 5μgを(atpA-COL1A2L、atpA-COL1A2S、rbcL-COL1A2S)をスペ
ルミジン(16nM)とCaCl2(1M)で600nm金粒子に付着させる。対数増殖期(2~5×106個/ml)のCC-125mt
+(プレート1枚あたり5ml)をろ過装置(Mllipore 47mm ガラス製フィルターホルダー)を用いてHybondTM-N
+ナイロンメンブレンフィルター(Amaersham Biosciences)上に細胞が一層になるように吸引ろ過した。
CW-15は吸引ろ過によりメンブレン上で破裂して死滅する可能性が高いために、TAP寒天培地(スペクチノ
マイシン100μg/ml:spc100mg/ml)に一層になるように撒いて乾燥させた。PDS-1000/Heのチャンバー内を
25mmHg に減圧し、1000psi の He ガス圧で金粒子を CC-125mt+、CW-15 に撃ち込だ。その後、TA 寒天培地
(spc100μg/ml)で 2 週間程培養し、出てきたコロニーを TAP 寒天培地(spc100μg/ml)で単細胞分離し
た。
導入遺伝子導入遺伝子導入遺伝子導入遺伝子のののの確認確認確認確認
形質転換後、単離培養したコロニーをTAP液体培地(spc100μg/ml)で対数増殖期のまで培養し、DNeasy
Plant Kit(Qiagen)によってDNAを回収し、PCR(94℃2分→[94℃30秒→51℃30秒→72℃2分]×40サイ
クル→72℃2分)(aadA領域を増幅するプライマーセットaadA/F、aadA/R、COL1A2S領域を増幅するプライ
マーセット:COL1A2S-primer1、COL1A2S/R)によって外来遺伝子が目的場所へ導入されたかを確認した。ま
た、導入位置の確認はPCR(94℃2分→[94℃30秒→53℃30秒→72℃90秒]×40サイクル→72℃2分)(導
入されていれば2.5kb付近にバンドが出るプラーマーセット:rbcLC/F、rbcLC/R)により確認を行った。(図
2)
タンパクタンパクタンパクタンパク質質質質のののの抽出抽出抽出抽出
PCR により、コンストラクトの導入が確認された細胞をTAP液体培地(spc100μg/ml)3日~7日振盪培
養で対数増殖期まで増殖させ、微量遠心機(15000rpm、2分)で細胞をペレット状にした。ペレットに2×
SDS サンプルバッファー(0.25M Tris-HCLl[ph6.8]、SDS、超純水、グリセロール、βメルカプトエタノー
ル)を加え、ガラスビーズを入れて約2分ボルテックスして細胞壁を破壊した。その後、5分間ボイルして
サンプルとした。(大野、西村 1997)
SDSSDSSDSSDS----PAGEPAGEPAGEPAGE によるによるによるによるタンパクタンパクタンパクタンパク質質質質のののの分離分離分離分離ととととウェスタンブロットウェスタンブロットウェスタンブロットウェスタンブロット
18%ランニングゲル(0.75M Tris-HCl、10%SDS、超純水、Long Ranger gelsolution:BMARockland,ME USA、
TEMED、10%APS)、の上部に 4%スタッキングゲル(0.25M Tris-HCl、10%SDS、超純水、Long Ranger
gelsolution:BMARockland,ME USA、TEMED、10%APS)を作製し、タンパク質の分離を行った。電気泳動の条
件は、20mAで 1時間(スタッキングゲルの通過)、ランニングゲルに到達後は40mAで 1時間電気泳動を行
った。その後、電気泳動後のゲルからランニングゲルを切り出し、プロテイントランスファーバッファー
(0.25mM Tris、192mM グリシン、20% メタノール)の中で 20 分間振盪する。ランニングゲルと同じ大き
さに切った HybondTM-P:PVDF メンブレンをメタノールに 10 秒間浸し、超純水で 5 分間振盪し、プロテイン
トランスファーバッファーで 10 分間振盪して平衡化した後、定電流(ゲル面積 1cm2あたり 2mA)で 90 分
間ブロッティングを行った。(大野、西村 1997)

9
ドットブロットドットブロットドットブロットドットブロット
HybondTM-P:PVDF メンブレンを切り出し(1サンプルあたり1cm2必要)、メタノールに10秒程浸し、超純
水で 5 分間振盪して洗浄した後、PBS バッファー(75mM Na2HPo4、25mM NaH2Po4、100mM NaCl)に浸しドッ
トブロットに用いた。調整したメンブレンを PBS バッファーに浸した 3MM ペーパー(Watman)の上に乗せ
た状態で、2×SDSサンプルバッファーで処理したサンプルを1μlスポットした後、乾いた3MMペーパー上
で乾燥させた。
イムノブロティングイムノブロティングイムノブロティングイムノブロティング
ウェスタンブロッティングまたはドットブロットを行った PVDF メンブレンをブロッキングバッファー
(PBSバッファー、Tween20 01.%v/v、スキムミルク5%v/v:雪印)(2ml/cm2)に浸し、4℃で一晩または室温
で 1 時間振盪して抗体の非特異的な結合に対するブロッキングを行った。その後、メンブレンを PBS-T バ
ッファー(PBSバッファー、Tween20 01.%v/v)(2ml/cm2)で2回洗い、イムノブロティングに使用した。
ブロッキング後のメンブレンをコラーゲンα2type1に特異的に反応する1次抗体MS-527(Takara bio inc.
委託生産)を1/5000含む PBS-Tバッファー(2ml/cm2)に浸し、室温で1時間振盪した。PBS-Tバッファー
を交換しながら2回洗浄し、PBS-Tバッファー(4ml/cm2)室温で15分間振盪した。その後、PBS-Tバッフ
ァーを交換しながら3回洗浄(室温で5分間振盪)した。洗浄の終わったメンブレンに2次抗体Mouse IgG,
HRP linked whole antibody from sheep(Amaersham Biosciences)を1/25000倍含むPBS-Tバッファーで
希釈した液(2ml/cm2)に浸し、1時間振盪する。PBS-Tバッファーを交換しながら2回洗浄し、PBS-Tバッ
ファー(4ml/cm2)室温で15分間振盪した。その後、PBS-Tバッファーを交換しながら3回洗浄(室温で5
分間振盪)した。HRP 基質(ECL Plus Western Blotting Detection System :Amaersham Biosciences)を
メンブレンに塗布し、室温で 5 分間置き、Image Reader LAS-1000 ver1.0(Fujifilm)を用いて化学蛍光
を検出した。
コラーゲンコラーゲンコラーゲンコラーゲン抗体抗体抗体抗体のののの検証検証検証検証
HybondTM-P:PVDF メンブレンにウシコラーゲン(Bovine Collagen TypeⅠ:BD Biosciences)、ヒトコラー
ゲン(Human Collagen TypeⅠ:BD Biosciences)、熱処理(5 分間ボイル)したウシコラーゲン、ヒトコラ
ーゲン、アミノ酸合成したαⅠTypeⅠ抗原、CC-125mt+から抽出したタンパクとαⅠTypeⅠ抗原を混合した
ものをブロットした。上記と同じ条件でブロッキング後、1次抗体ヒトコラーゲン抗体(Rabbit Antibody to
Human Collagen TypeⅠ:BIODESIGN)1/5000、2次抗体Rabbit IgG, HRP linked whole antibody from donkey
(Amaersham Biosciences)1/25000で上記のイムノブロッティングと同様に、化学蛍光の検出を行った。
コラーゲンタンパクコラーゲンタンパクコラーゲンタンパクコラーゲンタンパクのののの抽出抽出抽出抽出
コンストラクトの導入が確認された形質転換体を対数増殖期まで培養し、5000rpm で 10 分遠心を行い、
ペレット化した。ペレットに0.6ml Cold Homogenigation buffer(50mM Tris[ph7.4]、150mM NaCl、2.5mMEDTA)
に溶かした。1M DTTと 1M PMSFをそれぞれ2μl加え、2ml サンプルカップにガラスビーズを0.1g程度加
えて約2分ボルテックスで破砕した。さらに、0.6mlの Cold Extraction Buffer(1.0M Acetic acid、0.4M
NaCl)を加え、4℃で12時間以上振盪した。15000rpmで 20分遠心し、酸不溶性タンパクを沈殿させたもの

10
に3.4M NaClを 1.3ml加え、4℃で12時間以上放置した。その後、35000gで 1時間遠心し、コラーゲンを
沈殿させ、上清を除き、10μlの0.1M 酢酸に溶かして回収した。(Edward J. 1982)
結果結果結果結果とととと考察考察考察考察
形質転換体形質転換体形質転換体形質転換体のののの確認確認確認確認
3種類のコンストラクト(atpA-COL1A2L、atpA-COL1A2S、rbcL-COL1A2S)をそれぞれパーティクルガン法
によりCC-125mt+とCW-15に 10枚ずつ計60プレートに導入した。その結果、rbcL-COL1A2S コンストラク
トが導入されたと思われる CC-125mt+を 1 株得ることができた(COL1A2S-mt と命名)。atpA-COL1A2L、
atpA-COL1A2S コンストラクトについては撃ち込みに用いた DNA 濃度が薄く、うまく形質転換されなかった
と考えられる。得られた形質転換体からDNAを回収し、PCRによって導入された遺伝子の確認を行った。導
入された遺伝子については、aadA前部からCOL1AS後部(1.9kb)、COL1A2S前後部(1.1kb)、aadA前後部(800bp)
がそれぞれ撃ち込みに使用したコンストラクトと同じ位置にバンドが確認できることから、目的のバンド
が導入されていることが確認できた。(図 3)導入位置の確認には rbcL/C/F と rbcL/C/R を用い、PCR を行
ったが、特異的な増幅(2.5kb 付近)は確認できず、目的である葉緑体への導入は確認できなかった。(図
4)
図 2. 葉緑体への導入とコンストラクト導入の確認
葉緑体内に上記の図のように導入される予定である。rbcL3’UTR側の相同組み換えにより導入される。
atpA-COL1A2L、atpA-COL1A2S は atpA5’と相同組み換えを起こし、atpA を破壊する形で導入される。矢印はコンス
トラクト導入確認のためのプライマー位置を表す。
rbcL-COL1A2S
rbcL
aadA COL1A2S
atpA
atpA5’ aadA COL1A2L
atpA5 aadA COL1A2S
atpA-COL1A2L
atpA-COL1A2S
aadA/F
aadA/R
aadA/F
aadA/R
Primer1 COL1A2S/R
rbcLC/FrbcLC/R
Primer2
Primer3
Primer1
Primer1
葉緑体DNA

11
図 3. PCRによる導入遺伝の確認
プライマーを用いて導入されたコンストラクトの確認。
1. aadA 遺伝子の前部から COL1A2S 遺伝子の後部(約 1.9kb)を特異的に増幅するプライマーセット
aadA/F+COL1A2S/R。
2. COL1A2s遺伝子の前後部(約1.1kb)を特異的に増幅するプライマーセットprimer1+COL1A2S/R。
3. aadA遺伝子の前後部(約800b)を特異的に増幅するプライマーセットaadA/F+aadA/R。
aadA COL1A2S
aadA/F
aadA/R
Primer1
COL1A2S/R
COL1A2S-mt
rbcL-COL1A2Sコンストラクト
マーカー
1
2
3
1
2
3

12
コラーゲンコラーゲンコラーゲンコラーゲン抗体抗体抗体抗体のののの特異性特異性特異性特異性のののの検証検証検証検証
本研究では、タンパク質の検出のために細胞を熱処理する。そのため、熱処理をしたコラーゲンとも反
応する抗体が必要である。そこで、商品として販売されているヒトコラーゲン抗体(Rabbit Antibody to
Human Collagen TypeⅠ:Biodesign)について、熱変性コラーゲンと反応が可能か検証を行った。その結果、
熱処理をしていないウシコラーゲン(Bovine Collagen TypeⅠ:BD Biosciences)、ヒトコラーゲン(Human
Collagen TypeⅠ:BD Biosciences)との特異的反応は確認できるが、熱処理(5分間ボイル)を行ったヒト
コラーゲンでは反応が弱くなり、熱変性をしたウシコラーゲンについては、反応が確認できなかった。(図
5)今回検証したヒトコラーゲン抗体はタンパク質のアミノ酸配列認識だけではなく、立体構造も認識して
いること考えられ、今後の実験への使用が困難であることが解った。
図4. 導入遺伝子の位置確認
プライマーを用いて導入されたコンストラクトの確認を行った。コントロールとしてCC-125+を使用。
rbcLC/F と rbcLC/R で PCR を行うと遺伝子が正しく導入されていた場合は 2.5kb 付近にバンドが確認で
きる。しかし、上記の写真からは特異的なバンドは確認できない。
rbcL aadA COL1A2S rbcL3’UTR
rbcLC/F
rbcLC/R
マーカー
CC-125mt+
COL1A2S-mt
2.5kb

13
COL1A2COL1A2COL1A2COL1A2 用用用用ポリクローナルポリクローナルポリクローナルポリクローナル抗体抗体抗体抗体のののの使用使用使用使用のののの検討検討検討検討
標品ヒトコラーゲン用抗体が使用できないため、Collagen α2 Type1 遺伝子のtelo 部を含む13アミノ
酸(DRGPRGERGPPGC)に対するポリクローナル抗体作製を Takara bio inc. に委託した。作製した COL1A2
用ポリクローナル抗体について ECL Plus Western Blotting Detection System (Amaersham Biosciences)
を用いて、抗血清の特異性を検証した。その結果、COL1A2S 抗原および、COL1A2S-mt から抽出したサンプ
ルに対して特異的に反応することが確認できた。また、COL1A2S-mtにも反応が確認できることから、COL1A2S
遺伝子が発現していることも解った。(図6)
図5. ヒトコラーゲン抗体の特異性の検証
ヒトコラーゲン、ウシコラーゲン、熱処理ヒトコラーゲン、熱処理ウシコラーゲンを直接PDVFメンブレ
ンにスポットした後、ヒトコラーゲン抗体を用いたイムノブロッティングを行い、特異性を検証した。
ブロットしたサンプルは、1.ヒトコラーゲン、2.ウシコラーゲン、3.熱処理ヒトコラーゲン、4.熱処理
ウシコラーゲン、5.合成α1Type1抗原、6.CC-125mt++合成α1type1抗原
1111 2222 3333
4444 5555 6666
14
発現発現発現発現タンパクタンパクタンパクタンパクのののの確認確認確認確認
COL1A2S-mt 株が、増殖速度が非常に遅く、死に至りやすいという不安定な特徴を持つため、安定してコ
ラーゲンタンパクが発現されているのかを確認した。COL1A2S-mt を継代培養し、様々な時間毎にタンパク
抽出および COL1A2 用ポリクローナル抗体と ECL Plus Western Blotting Detection System (Amaersham
Biosciences)を用いたコラーゲンタンパクの検出を試みた。コントロールには C. reinhardtiiの野生株で
あるCC-125mt+からCOL1A2S-mtと同じ手法でタンパクを抽出したものを使用した。単一株より抽出したタ
ンパクにも関わらず、タンパク抽出を行った時期によって、コラーゲンの検出度が異なり、タンパクの発
現が非常に不安定であることが解った。(図7)
1111 2222 3333
4444 5555 6666
7777 8888
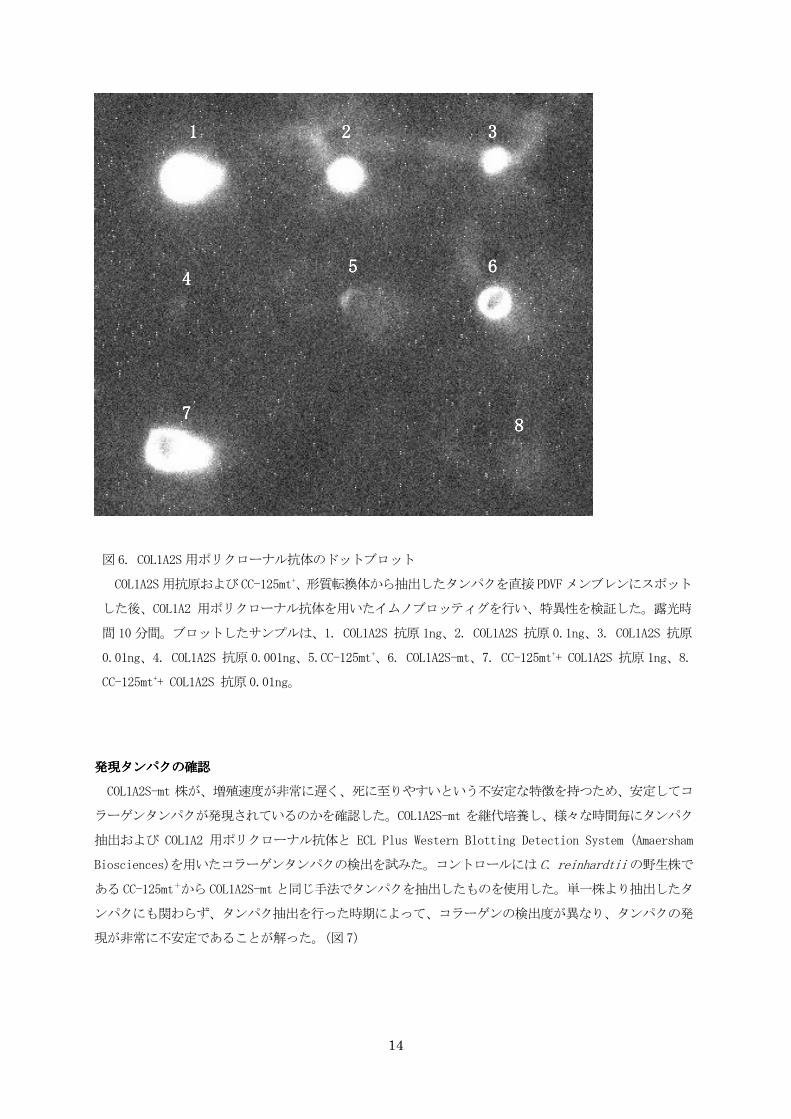
図6. COL1A2S用ポリクローナル抗体のドットブロット
COL1A2S用抗原およびCC-125mt+、形質転換体から抽出したタンパクを直接PDVFメンブレンにスポット
した後、COL1A2 用ポリクローナル抗体を用いたイムノブロッティグを行い、特異性を検証した。露光時
間 10 分間。ブロットしたサンプルは、1. COL1A2S 抗原 1ng、2. COL1A2S 抗原 0.1ng、3. COL1A2S 抗原
0.01ng、4. COL1A2S 抗原 0.001ng、5.CC-125mt+、6. COL1A2S-mt、7. CC-125mt++ COL1A2S 抗原 1ng、8.
CC-125mt++ COL1A2S 抗原0.01ng。

15
ウェスタンブロットウェスタンブロットウェスタンブロットウェスタンブロットによるによるによるによるタンパクタンパクタンパクタンパクのののの解析解析解析解析
ドットブロットにより特異的タンパクの発現が確認できた株について、SDS-PAGE でタンパクを分離し、
イムノブロットを行うことで目的のコラーゲンが発現されているか確認した。COL1A2S遺伝子が正しく翻訳
され、タンパク質が発現した場合、33kDa 付近に特異的なバンドが確認できるはずだが、COL1A2S-mt を用
いたサンプルでは、目的産物より分子量の小さいバンドしか確認できなかった。(図8)
図7. ドットブロット
タンパク質を直接PDVFメンブレンにスポットした後、COL1A2用ポリクローナル抗体を用いたイムノ
ブロッティングを行い、コラーゲンの発現を検証した。露光時間は10分間。ブロットしたサンプルは、
1. CC-125mt+、2. COL1A2S-mt _0620、3. COL1A2S-mt _0711、4. COL1A2S-mt _0724、5. COL1A2S-mt _0725、
6. COL1A2S-mt _0728、7. COL1A2S-mt _0819、8. COL1A2S-mt _0917、9. COL1A2S-mt _0918。2~9のサ
ンプル名横の数字はタンパクを回収した日付。0724株と0819株について特異的な反応が見られた。
1111 2222 3333
4444 5555 6666
7777 8888 9999

16
コラーゲンタンパクコラーゲンタンパクコラーゲンタンパクコラーゲンタンパクのののの単離単離単離単離
コラーゲンタンパクを塩析出(Edward J. 1982)により単離を試みた。通常、コラーゲンタンパクは酸可
溶性であることが知られている。COL1A2S-mtから抽出したサンプルを酸可溶性分画と酸不溶性分画に分け、
酸可溶性分画に NaCl を加えてタンパクを析出させた後、0.1M 酢酸に溶かして使用する。回収したサンプ
ルをPDVF メンブレンにスポットした後、COL1A2 用ポリクローナル抗体を用いたイムノブロッティング
を行い、回収コラーゲンの有無を検証した。塩析には 10 月 15 日に回収したサンプルを使用した。また、
ポジティブコントロールには熱処理サンプル r-COL1A2S_0724(先のドットブロットで確認)を使用した。
結果、塩析出タンパクおよび上清の両サンプルともにコラーゲンタンパクが検出されなかった。(図 9)原
因として、発現したコラーゲンが本来の長さの 1/3 しかないため、本来の物性が得られず、酸不溶性分画
に混入した可能性がある為、酸不溶性分画中タンパクについても、コラーゲンタンパクの検出を試みた。
1111 2222 3333 4444 5555 6666 7777
図8. ウェスタンブロッティング
COL1A2S-mtより抽出したサンプルをSDS-PAGEを用いて分離し、イムノブロッティングにより解析し
た。露光時間は10分間。ロードしたサンプルは、1. CC-125mt+、2. COL1A2S-mt _0620、3. COL1A2S-mt
_0724、4. COL1A2S-mt _0819、5. COL1A2S-mt _0711、6. COL1A2S-mt _0917、7. マーカー。2~6のサ
ンプル名横の数字はタンパクを回収した日付。
33kDa

17
酸不溶性分画酸不溶性分画酸不溶性分画酸不溶性分画のののの確認確認確認確認
塩析出コラーゲンがどの分画にあるかを確認するために、酸不溶性分画も回収してイムノブロッティン
グを行った。酸不溶性分画はそのままスポットし、ポジティブコントロールとして熱処理サンプル
COL1A2S-mt _0724(先のドットブロットで確認)を使用した。結果、酸不溶性分画にも酸可溶性分画にも
コラーゲンタンパクは確認できなかった。(図 10)5.が反応しているのはブロットしたタンパクが多すぎた
ためのノイズであった。
1111 2222 3333
4444 5555 6666
7777 8888 9999
図9. 塩析出コラーゲンのドットブロット
形質転換体に対し塩析を行い、析出タンパクとその上清を直接 PDVF メンブレンにスポットした後、
COL1A2用ポリクローナル抗体を用いたイムノブロッティングを行い、コラーゲンタンパクの検出を試み
た。露光時間10分間。サンプルは、1. 上清廃液(酢酸)、2. 上清廃液濃縮(酢酸)、3. 上清廃液(PBS)、
4. 上清廃液濃縮(PBS)、5. 塩析出サンプル(酢酸)、6. 塩析出サンプル(PBS)、7. CC-125mt+、8.
COL1A2S-mt _0724、9. COL1A2S-mt _1015。8、9.のサンプル名横の数字はタンパクを回収した日付、1
~6(サンプル溶液名)。

18
同様の実験をCOL1A2S-mtではなく、CC-125mt+にヒトコラーゲンタンパクを混合したサンプルで行うこと
で塩析方法の検討を行った。しかし、はすべての分画で特異的な反応は見られなかった。(データ不掲載)
このことから、今回使用した塩析法に不備があると考えられる。しかし、図9でr-COL1A2S_1015株の反応
が確認できていないことなどを考慮すると、COL1A2S-mt のコラーゲンタンパクの発現が止まっている可能
性が高いため、COL1A2S-mt でタンパクの発現が止まっている可能性を検証するため発現タンパクの再確認
を行った。使用したサンプルは継代培養を行っていた細胞から新しく調整したサンプルと、以前にドット
ブロットにより発現が確認できたCOL1A2S-mt _0724株(別途寒天培地[spc100μg/ml]にて保存)を培養し
て使用した。また、ポジティブコントロールとして COL1A2S 抗原を使用した。結果、すべての株で特異的
な反応は見られず、以前に発現が確認されたCOL1A2S-mt _0724株を培養した株でも確認できなかったため、
今日COL1A2S-mtでは、コラーゲンタンパクの発現が止まっていると考えられる。(図11)
1111 2222 3333
4444 5555 6666
図10. 塩析出サンプルの再ドットブロット
塩析出サンプルと酸不溶性分画サンプルを直接 PDVF メンブレンにスポットした後、COL1A2 用ポリク
ローナル抗体を用いたイムノブロッティングを行い、塩析出を検証した。露光時間10分間。ブロットし
たサンプルは、1.酸不溶性分画、2.塩析出サンプル(酢酸)、3. 上清廃液(酢酸)、4. 上清廃液濃縮(酢
酸)、5. CC-125mt+、6. COL1A2S-mt _0724。2~4(サンプル溶液名)

19
1111 2222 3333
4444 5555 6666
7777 8888 9999
図11. 形質転換体の再ドットブロット
タンパク質を直接PDVFメンブレンにスポットした後、COL1A2用ポリクローナル抗体を用いたイムノ
ブロッティングを行い、コラーゲンの発現を検証した。露光時間は10分間。ブロットしたサンプルは、
1. CC-125mt+、2. COL1A2S-mt _0724(古株)、3. COL1A2S-mt _1126(新株)、4. COL1A2S-mt _1121(新
株)、5. COL1A2S-mt _1215(新株)、6. COL1A2S-mt _1216(新株)、7. COL1A2S-mt _1217(新株)、8.
CC-125mt++COL1A2S抗原、9.COL1A2S抗原。3~7のサンプル名横の数字はタンパクを回収した日付。

20
導入遺伝子導入遺伝子導入遺伝子導入遺伝子のののの再確認再確認再確認再確認
タンパク発現の停止が確認された原因を検討すべく、導入遺伝子の再確認を行った。プライマーセット
は外来遺伝子導入を確認に用いた aadA/F、aadA/R、COL1A2S-primer1、COL1A2S/R を使用した。COL1A2S遺
伝子と aadA遺伝子は確認することができたが、aadA前部から COL1A2S後部までのプライマーセットで反応
が確認できなかった。(図12)aadA遺伝子、COL1A2S遺伝子はそれぞれ確認できており、欠損は起こってい
ないことが分かった。
aadA COL1A2S
aadA/F Primer1
aadA/R COL1A2S/R マーカー
マーカー
COL1A2S-mt _0724
COL1A2S-mt _1128
COL1A2S-mt _1216
COL1A2S-mt _1218
1111
2222
3333
1111
2222 3333
1111 2222 3333
1111
2222
3333
図12. PCRによる導入遺伝子の再確認
プライマーを用いて導入されたコンストラクトの確認。
1. aadA 遺伝子の前部から COL1A2S 遺伝子の後部(約 1.9kb)を特異的に増幅するプライマーセット
aadA/F+COL1A2S/R。
2. COL1A2s遺伝子の前後部(約1.1kb)を特異的に増幅するプライマーセットprimer1+COL1A2S/R。
3. aadA遺伝子の前後部(約800b)を特異的に増幅するプライマーセットaadA/F+aadA/R。

21
今回、atpAプロモーターと rbcLの3’UTRを利用したコンストラクト計3種類作製し、コラーゲンタン
パクの発現を試みた。atpA 用コンストラクトで形質転換体が得られなかったのは、形質転換の方法に不備
があったものと考えられるため、今後、導入する DNA 濃度を上げる等の検討をする必要がある。形質転換
体が得られた rbcL用コンストラクトで発現したコラーゲンタンパクについては、ポリクローナル抗体に特
異的に反応するタンパク質の存在は確認することができたが、予想していたものより分子量が小さく、ま
た、コラーゲンを抽出することもできなかった。導入したコンストラクトの欠損は生じていない為、発現
された思われるコラーゲンが実際の物より短いうえに、ヒドロキシル化などの修飾を受けていないためで
あること、抽出に使用したサンプルのタンパク発現が停止していたことが主な原因であると考えた。この
ことから、葉緑体用に作製したコンストラクトが葉緑体ではなく核に導入され、偶然プロモーターを拾っ
て発現した。そして核で発現してタンパク質を生産していたが、PTGS などの外来遺伝子阻害機構の影響を
受けてタンパク発現が止まってしまったと考えた。また、撃ち込んで間もないころに比べて培養速度が速
くなったため、TAP液体培地(スペクチノマイシン500μg/ml:spc100mg/ml)での培養を試してみたが、死
滅せずに培養できたことから aadA遺伝子に関しては、PTGSなどの影響を受けておらず、葉緑体で発現して
いると考えられる。結果として、撃ち込んで間もないころは aadA遺伝子、COLA2S遺伝子ともに偶然核に導
入されていたが、時間経過により aadA遺伝子は発現しているものの、COL1A2S遺伝子は外来遺伝子阻害機
構により発現が停止し、タンパク質の発現が止まったと考えられる。

22
参考文献参考文献参考文献参考文献
Dove A. (2002)
Uncorking the biomanufacturing bottleneck.
Nat Biotechnol.Aug;20(8):777-9
Edward J. Miller R. Kent Rhodes. (1982)
Preparation and characterization of the different types of collagen
Methods Enzymol vol82;39-40
Elizabeth H Harris. (1989)
The Chlamydomonas Sourcebook.
San Diego:Academic
Franklin S. Ngo B. Efuet E. Mayfield SP. (2002)
Development of a GFP reporter gene for Chlamydomonas reinhardtii chloroplast.
Plant J. 2002 Jun;30(6):733-44
Goldschmidt-Clermont M. (1991)
Transgenic expression of aminoglycoside adenine transferase in the chloroplast: a selectable marker
of site-directed transformation of chlamydomonas.
Nucleic Acids Research. Aug 11;19(15):4083-89
Olsen DR. Leigh SD. Chang R. McMullin H. Ong W. Tai E. Chisholm G. Birk DE. Berg RA. Hitzeman RA.
Toman PD.(2001)
Production of human type I collagen in yeast reveals unexpected new insights into the molecular
assembly of collagen trimers.
J Biol Chem. Jun 29;276(26):24038-43.
Ruggiero F. Exposito JY. Bournat P. Gruber V. Perret S. Comte J. Olagnier B. Garrone R. Theisen
M.(2000)
Triple helix assembly and processing of human collagen produced in transgenic tobacco plants.
FEBS Lett. Mar 3;469(1):132-6.
大野 茂男、西村 善文(1997)
細胞工学別冊 プロトコールシリーズ タンパク実験プロトコール ①機能解析編
秀潤社

23
西村 俊祐 (2004)
Chlamydomonas reinhardtiiの葉緑体での外来遺伝子発現系の構築:コレラ毒素Bサブユニットタンパク質
の発現
高知工科大学 大学院 修士論文
藤本大三郎(1998)
コラーゲンの秘密に迫る-食品・化粧品からバイオマテリアルまで-
裳華房
藤本大三郎(1999)
コラーゲン物語
東京化学同人


















