
Characterization - pdfs.semanticscholar.org · with a Polytron homogenizer (Brinkmann Instru-ments,...
11
Biochem. J. (1982) 206, 267-277 267 Printed in Great Britain Characterization of the enzymic capacity for cysteine desulphhydration in liver and kidney of the rat Martha H. STIPANUK and Pamela Witte BECK Division ofNutritional Sciences, Cornell University, Ithaca, NY 14853, U.S.A. (Received 9 February 1982/Accepted 26 April 1982) The contribution of cystathionine y-lyase, cystathionine,f-synthase and cysteine amino- transferase coupled to 3-mercaptopyruvate sulphurtransferase to cysteine desulphhydra- tion in rat liver and kidney was assessed with four different assay systems. Cyst- athionine y-lyase and cystathionine J-synthase were active when homogenates were incubated with 280 mM-L-cysteine and 3 mM-pyridoxal 5'-phosphate at pH 7.8. Cysteine aminotransferase in combination with 3-mercaptopyruvate sulphurtransferase catalysed essentially all of the H2S production from cysteine at pH 9.7 with 160mM-L-cysteine, 2mM-pyridoxal 5'-phosphate, 3mM-2-oxoglutarate and 3mM-dithiothreitol. At more- physiological concentrations of cysteine (2 mM) cystathionine y-lyase and cystathionine ,6-synthase both appeared to be active in cysteine desulphhydration, whereas the amino- transferase pathway did not. The effect of inhibition of cystathionine y-lyase by a suicide inactivator, propargylglycine, in the intact rat was also investigated; there was no significant effect of propargylglycine administration on the urinary excretion of total 35S, 35SO42- or [35S]taurine formed from labelled dietary cysteine. The desulphhydration of cyst(e)ine may be cata- lysed by several enzymes present in mammalian tissues. These include cysteine aminotransferase (EC 2.6.1.3) in conjunction with 3-mercapto- pyruvate sulphurtransferase (EC 2.8.1.2), cyst- athionine y-lyase (EC 4.4.1.1) and cystathionine fi- synthase (EC 4.2.1.22). The reactions catalysed by these enzymes are summarized in Scheme 1. The reactions may be important not only in cysteine catabolism but also as a source of metabolically active reduced sulphur. Other known pathways of cysteine degradation in animal tissues lead to release of sulphur in the 4+ or 6+ oxidation state. This highly oxidized sulphur is irreversibly lost to the pool of metabolically active reduced sulphur because of the absence of sulphate- and sulphite-reducing systems in animal tissues (Greenberg, 1975). Cysteine participates in transamination reactions with various amino acceptors, and it is likely that several non-specific aminotransferases catalyse its transamination to 3-mercaptopyruvate (Meister et al., 1954; Ip et al., 1977; Ubuka et al., 1977a,b, 1978). Aspartate aminotransferase catalyses the transamina- *tion of cysteine with 2-oxoglutarate in vitro and may also be active in vivo (Taniguchi & Kimura, 1974). The keto acid of cysteine is decomposed to inorganic sulphur and pyruvate by both desulphura- tion and'trans-sulphuration reactions (Meister et al., Vol. 206 1954; Kun & Fanshier, 1958, 1959a,b). These reac- tions are catalysed by the same enzyme, 3-mercapto- pyruvate sulphurtransferase, which apparently has an absolute specificity for 3-mercaptopyruvate as the sulphur donor (Fiedler & Wood, 1956). The sulphurtransferase can transfer the sulphur from 3-mercaptopyruvate to sulphite to form thio- sulphate or to other sulphur acceptors such as thiol compounds, sulphinates and cyanide (Meister et al., 1954; Kun & Fanshier, 1958, 1959a,b). When no sulphur-accepting substrate is present the atomic sulphur of the enzyme-sulphur complex is apparently released and may be polymerized to form elemental sulphur, reduced to sulphide or oxidized to sulphate (Meister et al., 1954; Kun & Fanshier, 1959a). Free H2S may be formed by reduction of the atomic sulphur or released from products such as thiosulphate and persulphides in subsequent reactions, as summarized in reactions IA-1C of Scheme 1. H2S may be released from thiosulphate, as illustrated in the inset in Scheme 1, in a reaction catalysed by thiosulphate sulphurtrans- ferase (EC 2.8.1.1) or a glutathione-dependent thio- sulphate reductase (Koj et al., 1967; Koj, 1968; Uhteg & Westley, 1979). Cystathionine -lyase catalyses the desulphhydra- tion of certain disulphides, including cystine (Cavallini et al., 1962a,b; Szczepkowski & Wood, 0306-3283/82/080267-1 1$01.50/1 X 1982 The Biochemical Society
Transcript of Characterization - pdfs.semanticscholar.org · with a Polytron homogenizer (Brinkmann Instru-ments,...

Biochem. J. (1982) 206, 267-277 267Printed in Great Britain
Characterization of the enzymic capacity for cysteine desulphhydration inliver and kidney of the rat
Martha H. STIPANUK and Pamela Witte BECKDivision ofNutritional Sciences, Cornell University, Ithaca, NY 14853, U.S.A.
(Received 9 February 1982/Accepted 26 April 1982)
The contribution of cystathionine y-lyase, cystathionine,f-synthase and cysteine amino-transferase coupled to 3-mercaptopyruvate sulphurtransferase to cysteine desulphhydra-tion in rat liver and kidney was assessed with four different assay systems. Cyst-athionine y-lyase and cystathionine J-synthase were active when homogenates wereincubated with 280mM-L-cysteine and 3 mM-pyridoxal 5'-phosphate at pH 7.8. Cysteineaminotransferase in combination with 3-mercaptopyruvate sulphurtransferase catalysedessentially all of the H2S production from cysteine at pH 9.7 with 160mM-L-cysteine,2mM-pyridoxal 5'-phosphate, 3mM-2-oxoglutarate and 3mM-dithiothreitol. At more-physiological concentrations of cysteine (2mM) cystathionine y-lyase and cystathionine,6-synthase both appeared to be active in cysteine desulphhydration, whereas the amino-transferase pathway did not. The effect of inhibition of cystathionine y-lyase by a suicideinactivator, propargylglycine, in the intact rat was also investigated; there was nosignificant effect of propargylglycine administration on the urinary excretion of total35S, 35SO42- or [35S]taurine formed from labelled dietary cysteine.
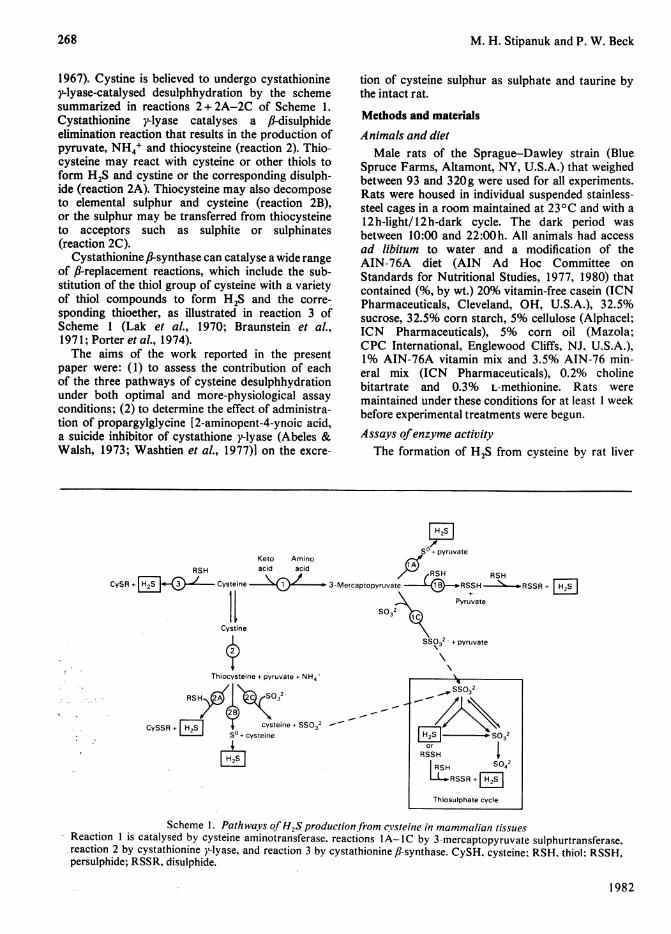
The desulphhydration of cyst(e)ine may be cata-lysed by several enzymes present in mammaliantissues. These include cysteine aminotransferase(EC 2.6.1.3) in conjunction with 3-mercapto-pyruvate sulphurtransferase (EC 2.8.1.2), cyst-athionine y-lyase (EC 4.4.1.1) and cystathionine fi-synthase (EC 4.2.1.22). The reactions catalysedby these enzymes are summarized in Scheme 1.The reactions may be important not only in cysteinecatabolism but also as a source of metabolicallyactive reduced sulphur. Other known pathways ofcysteine degradation in animal tissues lead to releaseof sulphur in the 4+ or 6+ oxidation state. Thishighly oxidized sulphur is irreversibly lost to thepool of metabolically active reduced sulphur becauseof the absence of sulphate- and sulphite-reducingsystems in animal tissues (Greenberg, 1975).
Cysteine participates in transamination reactionswith various amino acceptors, and it is likely thatseveral non-specific aminotransferases catalyse itstransamination to 3-mercaptopyruvate (Meister et al.,1954; Ip et al., 1977; Ubuka et al., 1977a,b, 1978).Aspartate aminotransferase catalyses the transamina-*tion of cysteine with 2-oxoglutarate in vitro andmay also be active in vivo (Taniguchi & Kimura,1974). The keto acid of cysteine is decomposed toinorganic sulphur and pyruvate by both desulphura-tion and'trans-sulphuration reactions (Meister et al.,
Vol. 206
1954; Kun & Fanshier, 1958, 1959a,b). These reac-tions are catalysed by the same enzyme, 3-mercapto-pyruvate sulphurtransferase, which apparently hasan absolute specificity for 3-mercaptopyruvate asthe sulphur donor (Fiedler & Wood, 1956). Thesulphurtransferase can transfer the sulphur from3-mercaptopyruvate to sulphite to form thio-sulphate or to other sulphur acceptors such asthiol compounds, sulphinates and cyanide (Meisteret al., 1954; Kun & Fanshier, 1958, 1959a,b).When no sulphur-accepting substrate is presentthe atomic sulphur of the enzyme-sulphur complexis apparently released and may be polymerizedto form elemental sulphur, reduced to sulphideor oxidized to sulphate (Meister et al., 1954; Kun &Fanshier, 1959a). Free H2S may be formed byreduction of the atomic sulphur or released fromproducts such as thiosulphate and persulphides insubsequent reactions, as summarized in reactionsIA-1C of Scheme 1. H2S may be released fromthiosulphate, as illustrated in the inset in Scheme 1,in a reaction catalysed by thiosulphate sulphurtrans-ferase (EC 2.8.1.1) or a glutathione-dependent thio-sulphate reductase (Koj et al., 1967; Koj, 1968;Uhteg & Westley, 1979).
Cystathionine -lyase catalyses the desulphhydra-tion of certain disulphides, including cystine(Cavallini et al., 1962a,b; Szczepkowski & Wood,
0306-3283/82/080267-1 1$01.50/1 X 1982 The Biochemical Society
M. H. Stipanuk and P. W. Beck
1967). Cystine is believed to undergo cystathioniney-lyase-catalysed desulphhydration by the schemesummarized in reactions 2+ 2A-2C of Scheme 1.Cystathionine y-lyase catalyses a ,B-disulphideelimination reaction that results in the production ofpyruvate, NH4+ and thiocysteine (reaction 2). Thio-cysteine may react with cysteine or other thiols toform H2S and cystine or the corresponding disulph-ide (reaction 2A). Thiocysteine may also decomposeto elemental sulphur and cysteine (reaction 2B),or the sulphur may be transferred from thiocysteineto acceptors such as sulphite or sulphinates(reaction 2C).
Cystathionine fl-synthase can catalyse a wide rangeof f-replacement reactions, which include the sub-stitution of the thiol group of cysteine with a varietyof thiol compounds to form H2S and the corre-sponding thioether, as illustrated in reaction 3 ofScheme 1 (Lak et al., 1970; Braunstein et al.,1971; Porter et al., 1974).The aims of the work reported in the present
paper were: (1) to assess the contribution of eachof the three pathways of cysteine desulphhydrationunder both optimal and more-physiological assayconditions; (2) to determine the effect of administra-tion of propargylglycine [2-aminopent-4-ynoic acid,a suicide inhibitor of cystathione y-lyase (Abeles &Walsh, 1973; Washtien et al., 1977)] on the excre-
tion of cysteine sulphur as sulphate and taurine bythe intact rat.
Methods and materialsAnimals and diet
Male rats of the Sprague-Dawley strain (BlueSpruce Farms, Altamont, NY, U.S.A.) that weighedbetween 93 and 320g were used for all experiments.Rats were housed in individual suspended stainless-steel cages in a room maintained at 230C and with a12 h-light/ 12 h-dark cycle. The dark period wasbetween 10:00 and 22:00h. All animals had accessad libitum to water and a modification of theAIN-76A diet (AIN Ad Hoc Committee onStandards for Nutritional Studies, 1977, 1980) thatcontained (%, by wt.) 20% vitamin-free casein (ICNPharmaceuticals, Cleveland, OH, U.S.A.), 32.5%sucrose, 32.5% corn starch, 5% cellulose (Alphacel;ICN Pharmaceuticals), 5% corn oil (Mazola;CPC International, Englewood Cliffs, NJ, U.S.A.),1% AIN-76A vitamin mix and 3.5% AIN-76 min-eral mix (ICN Pharmaceuticals), 0.2% cholinebitartrate and 0.3% L-methionine. Rats weremaintained under these conditions for at least 1 weekbefore experimental treatments were begun.Assays ofenzyme activity
The formation of H2S from cysteine by rat liver
S + pyruvateKeto Amino
RSH acid acid 1jjRSH RSHCySR + H Cysteine 3-Mercaptopyruvate RSSH --RSSR + H2
Pyruvate
Cystine p{$) ~~~~~~~~~~SSX03'+ pyruvate
Thiocysteine + pyruvate + NH4-
RSH 2 32
CySSR + cysteine 4SSO32 -
So°+ cysteine
H 2
Scheme 1. PathwaVs ofH2S production from cs'steine in mammalian tissues-Reaction 1 is catalysed by cysteine aminotransferase, reactions lA-IC by 3-mercaptopyruvate sulphurtransferase,reaction 2 by cystathionine y-lyase, and reaction 3 by cystathionine JJ-synthase. CySH. cysteine: RSH. thiol: RSSH,persulphide;,RSSR, disulphide.
1982
_--W SS032 O
ES-~~~~S,2oor
RSSH i|RH SO42
LRSSR+EH1
Thiosulphate cycle
268
4

Cysteine desulphhydration in rat tissues
and kidney homogenates was measured with fourdifferent assay systems, referred to below as cysteinedesulphhydration assays I, II, III and IV. Alldeterminations were made in triplicate. The incuba-tion mixtures for assays I and II were based onpreliminary investigations of varied substrate andcofactor concentrations and pH to establish therequirements for maximal H2S formation by bothrat liver and kidney homogenates. A linear depen-dence of activity on tissue concentration and incuba-tion time was observed with both assay systems.Assay II incubation mixture included a keto acid(2-oxoglutarate), which is required for the amino-transferase pathway of H2S production, and dithio-threitol, which was added to minimize oxidation ofcysteine to cystine. 2-Oxoglutarate and dithiothreitolwere omitted from assay I incubation mixture, whichcontained 280 mM-L-cysteine (free base), 3 mm-pyridoxal 5'-phosphate, 100 mM-Tris/HCl buffer,pH 7.8, and 0.3 ml of a 4% (w/v) liver homogenateor 10% (w/v) kidney homogenate in 50mM-potas-sium phosphate buffer, pH 6.8, in a final volume of1.0ml; the pH of the complete incubation mixturewas adjusted to 7.8. The incubation mixture forassay II contained 160mM-L-cysteine, 2mM-pyridoxal 5'-phosphate, 3 mM-dithiothreitol, 3mM-2-oxoglutarate, 100mM-Tris/HCl buffer, pH 9.7, and0.3ml of a 1% (w/v) liver or kidney homogenatein 50mM-potassium phosphate buffer, pH6.8, in afinal volume of 1.0 ml; the pH of the completeincubation mixture was adjusted to 9.7. Whenhomogenates of brain, heart and gastrocnemiusmuscle were used for these two assays, 40% (w/v)homogenates were used for assay I and 0.5%, 1%and 4% (w/v) homogenates of heart, skeletal muscleand brain respectively were used for assay II.The incubation conditions for cysteine desulph-
hydration assays III and IV were fixed at 2mM-cysteine and pH 7.4 to be closer to physiologicalconditions. Concentrations of pyridoxal 5'-phos-phate, 2-oxoglutarate and dithiothreitol were thosethat yielded maximal activity in rat liver and kidneyhomogenates. A linear dependence of activity ontissue concentration and incubation time was ob-served. Assay III incubation mixture contained,in a final volume of 1.0 ml, 2mM-cysteine, 0.05mM-pyridoxal 5'-phosphate, 100mM-potassium phos-phate buffer, pH7.4, and 0.3ml of 40% (w/v) liveror 30% (w/v) kidney homogenate in 50mM-potassium phosphate buffer, pH6.8; the pH of thecomplete incubation mixture was adjusted to 7.4.The incubation mixture for assay IV was the sameas that for assay III except that 0.5 mM-2-oxo-glutarate and 0.05 mM-dithiothreitol were included.
All incubations were performed in 25 ml Erlen-meyer flasks fitted with septum stoppers andplastic centre wells (Kontes, Vineland, NJ, U.S.A.).All components of the incubation mixtures and the
Vol. 206
tissue homogenates were added to the flasks whilethe flasks were in an ice bath. Centre wells werefilled with 0.5ml of a 15:1 (v/v) mixture of 1%(w/v) zinc acetate and 12% (w/v) NaOH and afolded 2cm x 2.5cm rectangle of Whatman no. 1filter paper for trapping evolved H2S (Fogo &Popowsky, 1949). Each flask was flushed with N2for 20 s and then sealed.The reactions were initiated by transferring the
flasks to a 370C shaking water bath at 20s intervals.After 15min at 370C, reactions were stopped byinjection of 0.5 ml of 50% (w/v) trichloroacetic acidthrough the diaphragm of each septum stopper..Flasks were left in the shaking water bath at 370Cfor 1h after the reactions were stopped to allowcomplete trapping of H2S. Blank incubations wereperformed for all samples in a similar manner exceptthat the tissue preparations were first heated at1000C for 2min.H2S was measured as described by Siegel (1965).
The centre wells and contents were transferred totest tubes, and 3.5 ml of water was added to eachtube. To each tube, 0.4 ml of 20mM-NN-dimethyl-p-phenylenediamine sulphate in 7.2M-HCl was added.This was followed immediately by addition of 0.4mlof 30mM-FeCl3 in 1.2M-HCl. After 10min, theabsorbance of the resulting solution at 670nm wasdetermined with a Gilford Stasar III spectrophoto-meter (Gilford Instrument Laboratories, Oberlin,OH, U.S.A.). The 6670 for sulphide was 3.38 x107M-1 cm-I; standard sulphide solutions were cali-brated by reaction of the sulphide with iodine andtitration of the excess iodine with standard thio-sulphate (Roy & Trudinger, 1970).
Cystathionine y-lyase, cysteine: 2-oxoglutarateaminotransferase and cystathionine 6-synthase ac-tivities were determined as previously described(Stipanuk & Benevenga, 1977; Stipanuk, 1979). Theproducts and reactions monitored were the forma-tion of 2-oxobutyrate by cleavage of cystathionine,the formation of glutamate by transamination ofcysteine and 2-oxoglutarate and the synthesis ofcystathionine from serine and homocysteine.The protein concentration of tissue preparations
was determined by the method of Lowry et al.,(1951). Bovine serum albumin was used as thestandard.
All rats were killed by decapitation between09:00 and 12:00h. The liver, both kidneys, heart,brain and the right gastrocnemius muscle wereimmediately removed, rinsed with ice-cold 0.9%NaCl, placed on ice and then homogenized in4 vol. of 50mM-potassium phosphate buffer, pH 6.8,with a Polytron homogenizer (Brinkmann Instru-ments, Westbury, NY, U.S.A.) The 20% (w/v)homogenates were subsequently diluted with addi-tional homogenization buffer to yield the appropriatetissue concentration.
269

M. H. Stipanuk and P. W. Beck
DL-Propargylglycine (Sigma Chemical Co., St.Louis, MO, U.S.A.) was added to some tissuehomogenates, which were then incubated at 370Cfor 5min before measurement of enzyme activity;the amount of propargylglycine added to each tissuepreparation was calculated to yield 2 mM-prop-argylglycine when the tissue preparation was dilutedin the complete incubation mixture. S-Adenosyl-L-methionine (chloride salt; Sigma) was added totissue homogenates to yield a concentration of 2mmin 20% (w/v) tissue homogenates; these homo-genates were incubated at 370C for 15min beforemeasurements of enzyme activity were made. Con-trol determinations were made on untreated tissuepreparations, which were from the same rats andhad been incubated in the same manner as thetreated tissue preparations.A group of six rats was given 30,umol of DL-
propargylglycine per lOOg body wt. by intraperi-toneal injection of a 60mM solution of DL-propargyl-glycine in 0.9% NaCl 3 h before the rats werekilled. A second group of five rats was given236,umol of L-ethionine (Sigma) per lOOg body wt.by intraperitoneal injection of a solution of 154mM-L-ethionine in 0.9% NaCl 2.5h before the rats werekilled. Two other groups of rats with similar bodyweights were injected with the correspondingvolumes of saline and used as control groups. Ratswere randomly assigned to treatment and controlgroups. Injections were made at 09:00h.
Effect ofpropargylglycine on the 24 h urinary excre-tion ofcysteine sulphur as sulphate and taurine
Twenty-four rats that weighed between 122 and162g were randomly distributed among six treat-ment groups. Two of these groups were fed adlibitum for 2 days on an L-amino acid diet that wasthe same as the 20%-casein diet except that thecasein was replaced by 16.94% of an L-amino acidmixture, the amounts of sucrose and corn starchwere each increased to 34.18% and the diet con-tained 6 mmol of DL-propargylglycine per kg of diet.The L-amino acid mixture was a modification of thatof Rogers & Harper (1965) and contained (% ofdiet, by wt.) 1.12% arginine, 0.30% histidine, 0.80%isoleucine, 1.08% leucine, 1.76% lysine hydro-chloride, 0.40% methionine, 0.60% cysteine, 1.14%phenylalanine, 0.17% tryptophan, 0.80% valine,0.34% alanine, 0.34% aspartic acid, 3.41% glutamicacid, 2.28% glycine, 0.34% proline, 0.34% serine,0.34% tyrosine, 0.60% asparagine and 0.79%sodium acetate. The amino acid diet was usedbecause a diet in which the cysteine content could bevaried and in which cysteine was present in free formwas needed for the tracer study on day 3. The otherfour groups of rats were fed on a control diet thatcontained no propargylglycine; the rats in two ofthese groups were fed ad libitum and those in the
remaining two groups were pair-fed to the rats givenpropargylglycine. Food was placed in the cages justbefore the beginning of the dark cycle each day.Food intake and body weight of the rats weremeasured daily.
Between 09:00 and 10:OOh on the third day, 5gof the L-amino acid diet mixed with 4ml of distilledwater and labelled with 2,uCi of L-l 35S]cysteine(Amersham Corp., Arlington Heights, IL, U.S.A.)was administered to each rat by gastric intubation.The diet given to each rat was the same as that eatenon the previous 2 days except that the L-cysteine(free base) content of the diet was 0.2% or 2.6%rather than 0.6%. The preparation and administra-tion of the liquid diets and the determination of theexact amount of L-[ 5S]cysteine administered toeach rat were performed as previously described(Stipanuk, 1979).
Rats were placed in individual metabolic cagesimmediately after they received the labelled diets,and urine was collected for 24h in vials that con-tained 5ml of 2% (w/v) phenol. Rats had accessto water but not to food during this 24 h period. Ratswere killed at the end of the 24 h period. The bladdercontents of each rat were added to the urine collec-tions, and the 'unabsorbed' radioactivity in thegastrointestinal tracts of the rats was determinedas previously described (Stipanuk & Benevenga,1977; Stipanuk, 1979). The amounts of total 3ISradioactivity, [35Sltaurine and 35SO42- in the 24hurine collections were determined by methods pre-viously described (Stipanuk, 1979). Results arereported as percentages of the absorbed (admin-istered minus unabsorbed) dose of L- 35S]cysteine.To ascertain that cystathionine y-lyase activity
was inhibited in tissues of the rats that were givenpropargylglycine, 18 additional rats (104-129gbody wt.) were randomly assigned to the same sixtreatment or control groups and fed on the sameL-amino acid diets for 2 days. On the third day at08:00h, these rats were given the same L-amino aciddiets by gastric intubation except that the liquiddiets were not labelled with L-[35S]cysteine. At 8hafter intubation of the liquid diets, the rats werekilled by decapitation and liver and kidneys wereremoved. The cystathionine y-lyase activity and therate of H2S production from cysteine (desulphhydra-tion assay system I) were determined for liver andkidney.
StatisticsData were analysed by using either Student's t
test or paired t test [Minitab-80.1 (PennsylvaniaState University, University Park, PA, U.S.A.)employed with the Cornell University ComputerSystem]. Levels of significance have been reportedwhen P <0.10.
1982
270

Cysteine desulphhydration in rat tissues
Results
Tissue distribution ofcapacityfor H2S productionThe distribution of the capacity for H2S produc-
tion in rat tissues was determined with assay systemsI and II, which yielded maximal rates of H2S forma-tion in the absence (I) and in the presence (II) of2-oxoglutarate and dithiothreitol. As shown in Table1, H2S production in assay system II was muchhigher than that in system I in all tissues. H2S pro-duction in assay system I was greatest in liver andkidney, which corresponds to the distribution ofcystathionine y-lyase and cystathionine fJ-synthaseactivities in rat tissues (Tallan et al., 1958; Muddet al., 1965; Sturman et al., 1970; Finkelstein et aL,1971). H2S production in assay system II wasgreatest in heart, liver and kidney; this activity dis-tribution is similar to that observed for cysteine:2-oxoglutarate aminotransferase (Ishimoto, 1979;S.-M. Kuo, T. C. Lea & M. H. Stipanuk, unpublishedwork), as well as aspartate aminotransferase(Herzfeld & Greengard, 1971), which can catalysethe transamination of cysteine (Taniguchi & Kimura,1974). The pH value and substrate and cofactorconcentrations that yielded maximal rates of H2Sproduction from cysteine in rat liver with assay IIwere almost the same as those found to give maximaland similar rates of glutamate production in thetransamination of cysteine with 2-oxoglutarate(S.-M. Kuo, T. C. Lea & M. H, Stipanuk, un-published work). Thus assay system II appears tomeasure primarily, if not entirely, the combinedactivity of cysteine aminotransferase and 3-mer-captopyruvate sulphurtransferase.
Influence of pyridoxal S'-phosphqte and 2-oxo-glutarate on H2S productionAs shown in Table 2 for assay system I, which
yielded maximal activity for rat liver and kidneyhomogenates in the absence of 2-oxoglutarate, theomission of pyridoxal 5'-phosphate resulted in a
Table 1. Distribution of enzymic capacity for cysteinedesulphhydration in rat tissues
Experimental details are given in the text. Values aremeans + S.D. for four or five rats whose body weightsranged from 190 to 269 g.
Cysteine desulphhydration(umol of H2S/min per g)
TissueLiverKidneyHeartBrainSkeletal muscle
Assay I0.645 + 0.0820.250 + 0.0720.037 + 0.0190.018 + 0.0100.010+0.004
Assay II3.78 + 0.522.26 +0.154.11 +0.141.42 + 0.091.67 + 0.10
78% and a 45%o decrease in H2S production in liverand kidney respectively. On the other hand, omissionof pyridoxal 5'-phosphate from assay system II,which yielded a maximal rate of H2S production forrat liver and kidney homogenates in the presenceof 2-oxoglutarate, had no effect on H2S productionby either liver or kidney. Omission of pyridoxal5'-phosphate from the more-physiological assaysystems, III and IV, resulted in a 22% and a 43%decrease for liver and a 35% and a 29% decrease forkidney in H2S production in assay systems III andIV respectively. Thus exogenous pyridoxal 5'-phos-phate appeared to be essential for maximal activityin assay systems I, III and IV but not in assaysystem II. The similar requirement for pyridoxal$'-phosphate in assay systems I, III and IV suggeststhat the same enzyme is involved in catalysis ofH2S production in the three systems and that adifferent enzyme is involved in assay system II.
Addition of 2-oxoglutarate increased H2S produc-tion in assay system I by 256% and 369% in liverand kidney respectively, whereas omission of 2-oxoglutarate from assay system II resulted in a 92%and a 96% decrease in activity in liver and kidneyrespectively. Addition of 2-oxoglutarate to assaysystem III had no significant effect on H2S produc-tion by either liver or kidney; omission of 2-oxo-:glutarate from assay system IV similarly had noeffect on H2S production by liver but decreasedH2S production by kidney by 21%. Theseobservations suggest either that cysteine trans-amination did not play a large role in cysteinedesulphhydration under the more-physiologicalincubation conditions used in assay systems IIIand IV or that endogenous keto acids in the tissuepreparations were adequate for cysteine trans-amination in these assay systems. The near-absolutedependency of activity in assay system II on theaddition of 2-oxoglutarate supports the previousconclusion that this assay system primarily is ameasure of the combined activities of cysteineaminotransferase and mercaptopyruvate sulphur-transferase.The influence of dithiothreitol on the rate of
cysteine desulphhydration in each of the four assaysystems was also investigated. Because cystathioniney-lyase uses cystine rather than cysteine as itssubstrate, we added dithiothreitol to minimizeformation of cystine, and hence the desulphhydra-tion of cystine by cystathionine y-lyase. Althoughaddition of 3 mM-dithiothreitol to assay system IIresulted in a small and consistent increase in HISproduction, no statistically significant effect ofdithiothreitol was observed in any of the assaysystems. It seems likely that dithiothreitol has non-specific effects that may enhance the production ofH2S by some pathways and decrease the productionof H2S by other pathways.
Vol. 206
271

M. H. Stipanuk and P. W. Beck
Table 2. Effect ofaddition or omission ofassay components on the production ofH2S by rat liver and kidney homogenates, Experimental details are given in the text. Values for control activities are means ± S.D. for 11 rats. Values expressed
as percentages of control are means + S.D. for three to seven rats. The level of significance was determined with thepaired t test.
Control activity(,umol of H2S/min per g)
Cysteine-desulphhydration ,assay system Liver
I: 280mM-L-cysteine, 3 mM- 0.403 ± 0.154pyridoxal 5'-phosphate, pH 7.8
II: 160mM-i,cysteine, 2 mM- 2.31 ± 0.57pyridoxal 5'-phosphate,
3 mM-2-oxo&lutarate, 3 mM-dithiothreitol, pH 9.7
III: 2mM-L-cysteine, 0.05 mM- 0.017 + 0.003pyridoxal 5'-phosphate, pH 7.4
IV: 2mM-L-cysteine, 0.05 mM- 0.019 ± 0.006pyridoxal 5'-phosphate,0.5 mM-2-oxoglutarate,0.05 'mM-dithiothreitol,
pH7.4
Kidney0.192 +0.065
1.96 + 0.44
Addition or omission-Pyridoxal 5'-phosphate
+2-Oxoglutarate (3mM)
-Pyridoxal 5'-phosphate-2-Oxoglutarate
0.013 + 0.001 -Pyridoxal 5'-phosphate
+2-Oxoglutarate (0.5 mM)0.018 + 0.003 -Pyridoxal 5'-phosphate
-2-Oxoglutarate
H2S production(% of control)
Liver Kidney22+7 55+7
P <0.003 P <0.008356+21 469+72P<0.0001 P<0.002100+8 96+ 118+4 4+4
P< 0.0001 P < 0.0001
78± 15 64+6P<0.007 P<0.000196+29 137+3757+9 71 +9
P<0.0001 P<0.000198+ 15 79+8
P<0.04
Effects ofethionine and S-adenosylmethionine on theactivities ofenzymes involved in H2S productionWe used two approaches in an effort to enhance
cystathionine fl-synthase activity selectively.Ethionine (246ymnIol/100g body wt.) or saline wasadministered to intact rats, which were killed 2.5 hlater for determination of enzyme activities in liverand kidney of these rats. The second approach wasto incubate homogenates of liver and kidney fromuntreated rats with S-adenosylmethionine before thedetermination of enzyme activities.
Administration of ethionine to the intact rat didnot markedly indrease hepatic cystathionine ,B-synthase activity, as had been reported by Koracevicand co-workerg (Koracevic, 1975; Koracevic et al.,1979) and Finkelstein et al. (1975). Only a 33%increase (P<0.07) in the activity of hepatic cysta-thionine f?-synthase was observed. A 16% increase(P<0.07) in the rate of cysteine desulphhydrationby rat liver in assay system I was also observed. Noincrease in either activity was observed for kidney.The increases in both cystathionine ,8-synthase acti-vity and H2S production from cysteine in assaysystem I that were observed for liver, although small,are consistent with involvement of cystathionine,i-synthase in the catalysis of cysteine desulphhydra-tion by rat liver.Compared with the treatment of intact rats with
ethionine, incubation of liver and kidney homo-genates with S-adenosylmethionine had similar butsomewhat greater effects. As shown in Table 3, the
activity of cystathionine fl-synthase was increasedin liver by 44% and in kidney by 27%, but theseincreases were not statistically significant. The 44%increase in hepatic cystathionine ,B-synthase activityis smaller than the 166% increase reported byFinkelstein et al. (1975) but similar to the responsethat we observed to treatment of intact rats withethionine. Pretreatment of homogenates with S-adenosylmethionine increased H2S production fromcysteine as measured in assay system I by 39% inliver and 56% in kidney, but it had no effect on H2Sproduction as measured in assay system II, whichprimarily measures cysteine aminotransferase plusmercaptopyruvate sulphurtransferase activities. S-Adenosylmethionine was also effective in increasingH2S production from cysteine in both of the more-physiological assay systems; increases of 47% and42% with assay system III and of 75% and 20%with assay system IV were seen for liver and kidneyrespectively. No effect of S-adenosylmethionine oncystathionine y-lyase or cysteine aminotransferaseactivity was observed.
Effect of propargylglycine on the activities ofenzymes involved in H2S production
Abeles and co-workers (Abeles & Walsh, 1973;Washtien et al., 1977) have reported the irreversibleinactivation of cystathionine y-lyase by low concen-trations (60gM) of propargylglycine. Uren et al.(1978) observed cystathioninaemia and cystathio-ninuria in mice treated for 4 days with daily injec-
1982
272

Cysteine desulphhydration in rat tissues
tions of 12,umol of DL-propargylglycine per lOOgbody wt. DL-Propargylglycine was used in thepresent study as a specific covalent inhibitor ofcysteine desulphhydration catalysed by cystathioniney-lyase. Data in Table 4 show that incubation oftissue preparations with propargylglycine inhibitedcystathionine y-lyase activity by 93% in both liverand kidney; propargylglycine did not significantlyaffect cysteine: 2-oxoglutarate aminotransferaseactivity in liver or kidney or cystathionine ,-synthaseactivity in liver. Cystathionine ,B-synthase activity inkidney homogenates was slightly (13%) lower intreated than in control preparations.
Incubation of liver and kidney homogenates withpropargylglycine before the assay decreased H2Sproduction with assay system I by 87% and 74%in liver and kidney respectively, but had no signi-
ficant effect with assay system II. In the more-physiological assays of cysteine desulphhydration,pretreatment with propargylglycine decreased H2Sproduction by liver and kidney homogenates by68% and 30% respectively with assay system IIIand by 51% and 32% respectively with assay systemIV.
These results indicate that cystathionine y-lyasecatalysed little or none of the observed desulph-hydration of cysteine in assay system II, whereasit was responsible for most of the desulphhydrationof cysteine in assay system I. The residual activityin assay system I in the presence of propargylglycinemay have been due to H2S production catalysedby other enzymes (cystathionine f,-synthase orcysteine aminotransferase plus mercaptopyruvatesulphurtransferase) or to a small amount of cyst-
Table 3. Effect of incubation of rat liver and kidney homogenates with S-adenosyl-L-methionine on the activities ofenzymes that catalyse the production ofH2Sfrom cysteine
Experimental details are given in the text. Values are means ± S.D. for four rats. The level of significance was deter-mined by the paired t test.
Activity (% of control)
Enzyme activity (product measured)Cysteine desulphhydration assay I (H2S)
Cysteine desulphhydration assay II (H2S)Cysteine desulphhydration assay III (H2S)
Cysteine desulphhydration assay IV (H2S)
Cystathionine y-lyase (2-oxobutyrate)Cystathionine 16-synthase (cystathionine)
Cysteine: 2-oxoglutarate aminotransferase (glutamate)
Liver139 ± 10P <0.005104 +4147 + 32P <0.06175 + 46P<0.05106 + 6144+29P<0.0197+2
Kidney156 + 26P < 0.02102 + 3142 + 26P<0.05120 + 11P<0.04101 +4127 + 40
109 +9
Table 4. Effect of incubation of rat liver and kidney preparations with DL-propargylglycine on the activities of enzYmesthat catalyse the production ofH2Sfrom cysteine
Experimental details are given in the text. Values are means + S.D. for four to six rats. The level of significance wasdetermined by the paired t test.
Enzyme activity (product measured)Cysteine desulphhydration assay I (H2S)
Cysteine desulphhydration assay II (H2S)Cysteine desulphhydration assay III (H2S)
Cysteine desulphhydration assay IV (H2S)
Cystathionine y-lyase (2-oxobutyrate)
Cystathioninef,-synthase (cystathionine)
Cysteine: 2-oxoglutarate aminotransferase (glutamate)
Activity (% of control)
Liver Kidney13+5 26+8
P<0.0001 P<0.000198+ 12 102+532+11 70+15
P<0.0001 P<0.00449+22 68+20
P<0.002 P<0.017+7 7+6
P<0.0001 P<0.000195+14 87+8
P<0.04103+6 106+4
Vol. 206
273

M. H. Stipanuk and P. W. Beck
athionine y-lyase activity that was not inhibitedby propargylglycine.The effect of an intraperitoneal injection of
DL-propargylglycine (30,umol per lOOg body wt.)3 h before the rats were killed is summarized in Table5. Results were similar to those shown in Table 4 forthe pretreatment of tissue preparations with prop-argylglycine. Cystathionine y-lyase activity wasnearly completely inhibited in both liver and kidney(95% and 96%), and cysteine desulphhydrationin assay system II was decreased by 67% in liverand 50% in kidney of rats that had been injectedwith propargylglycine.
Effect ofpropargylglycine on L-[ "Slcysteine metab-olism in the intact rat
Because propargylglycine is effective in inhibitingcystathionine y-lyase activity in the intact rat (Table5; Stipanuk, 1977; Uren et al., 1978), we investigatedthe effect of feeding with propargylglycine (6 mmol/kg of diet) for 3 days on the 24 h urinary excretionof sulphate and taurine from labelled dietary
cysteine. We hypothesized that we might see a de-crease in urinary SO42- excretion relative to taurineexcretion if cystathionine y-lyase-catalysed desulph-hydration of cysteine plays a major role in cysteinedegradation in the intact rat. The results arereported in Table 6. Two concentrations of cysteinewere included in the labelled diets that were intubatedon the third day, a typical dietary content, 0.2%,and an excess dose, 2.6%. Whereas the control ratsfed ad libitum gained weight, rats given propargyl-glycine consumed less diet than did the control ratsand lost weight. The food intake and body-weightchange of the pair-fed control groups were inter-mediate between those of the control groups fed adlibitum and the groups given propargylglycine. Be-cause rats were pair-fed for only 2 days and intakefor the first day was estimated from a preliminarystudy, there was some difference between the meantotal food intake of the rats given propargylglycineand that of their pair-fed controls.The excretion of total 3IS and "SO42- was similar
for rats treated with propargylglycine and both the
Table 5. Effect of injection of DL-propargylglycine on the activities ofenzymes that catalyse the production ofH2Sfromcysteine in rat liver and kidney
Experimental details are given in the text. Values are means + S.D. for five or six rats. Values followed by an asterisk(*) are significantly different from the control value at P < 0.0001 by Student's t text.
Activity (numol/min per g)
Enzyme activity (product measured)Cysteine desulphhydration assay I (H2S)Cysteine desulphhydration assay II (H2S)Cystathionine y-lyase (2-oxobutyrate)Cystathionine #-synthase (cystathionine)
LiverA- A
Control +Propargylglycine0.36 + 0.03 0.15 + 0.03*3.1 +0.3 3.0+0.3
0.80 + 0.26 0.04 + 0.02*0.10+0.02 0.12+0.04
Kidney
Control0.19 +0.032.0 +0.3
0.24 + 0.070.03 +0.01
+ Propargylglycine0.09 + 0.01*1.7 + 0.2
0.02 + 0.02*0.04 + 0.02
Table 6. Effect of the administration of DL-propargylglycine for 3 days on the 24 h urinary excretion of sulphate andtaurinefrom L-35Slcysteine by the rat
Experimental details are given in the text. Values are means + S.D. for four rats. Statistical significance was deter-mined by Student's t test; * indicates that the +propargylglycine value is different than that for the control group fedad libitum (P < 0.05), and t indicates that the +propargylglycine value is different than that for the pair-fed controlgroup (P < 0.05).
Dietary treatmentControl fed ad libitumPair-fed control+Propargylglycine
Control fed ad libitumPair-fed control+Propargylglycine
Intubatedlabelled diet
0.2% cysteine0.2% cysteine0.2% cysteine +
propargylglycine2.6% cysteine2.6% cysteine2.6% cysteine +
propargylglycine
Body-weightchange
(g/2 days)14 + 10+4
-8 ± l*t
17+2-5+4-7 + 1*
Foodintake
(g/2 days)28+416+210+3*t
31 +415+213+3*
Urinary excretion(% of absorbed dose)
Total 35S6.3 + 3.06.8 + 3.07.4 + 0.8
47 + 0.760 + 1050 + 24
35SO42-3.2 + 1.63.9 + 2.03.4 +0.8
29 + 1.646 + 1237+18
[35SiTaurine0.84 + 0.330.54 + 0.500.25 + 0.05*
9.3 + 1.95.6 + 1.27.1 +2.5
I "SiTaurine/35SO42-excretion
ratio0.30 + 0.170.12 + 0.050.08 + 0.03*
0.33 + 0.080.13 + 0.030.21 +0.09
1982
274
Cysteine desulphhydration in rat tissues
Table 7. Effect of the administration of DL-propargylglycine for 3 days on cysteine desulphhvdration and cvstathioniney-lyase activity in rat liver and kidney
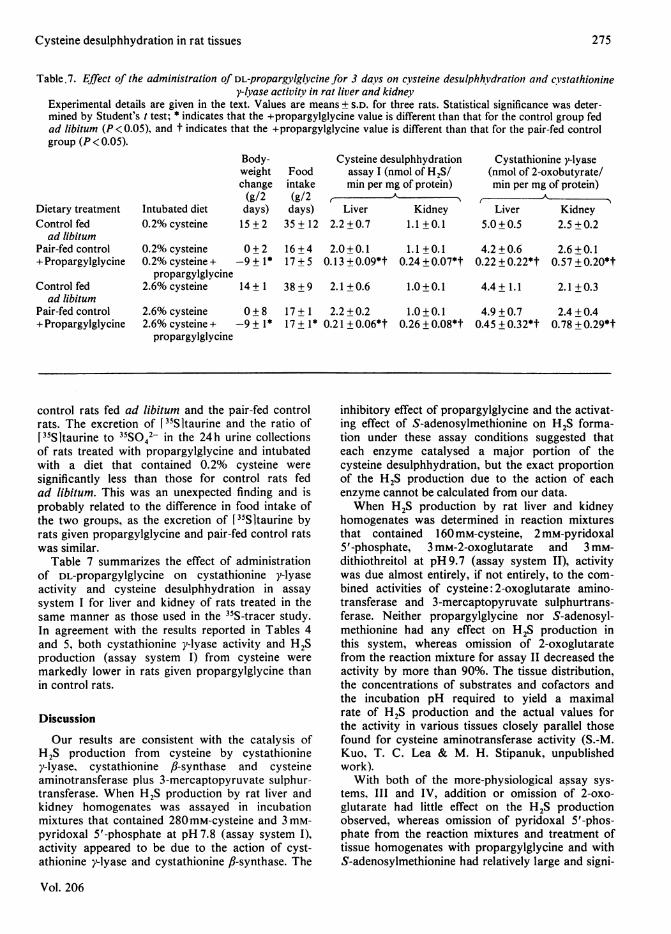
Experimental details are given in the text. Values are means ± S.D. for three rats. Statistical significance was deter-mined by Student's t test; * indicates that the +propargylglycine value is different than that for the control group fedad libitum (P < 0.05), and t indicates that the +propargylglycine value is different than that for the pair-fed controlgroup (P < 0.05).
Dietary treatmentControl fed
ad libitumPair-fed control+ Propargylglycine
Control fedad libitum
Pair-fed control+ Propargylglycine
Intubated diet0.2% cysteine
0.2% cysteine0.2% cysteine +
propargylglycine2.6% cysteine
2.6% cysteine2.6% cysteine +
propargylglycine
Body-weightchange(g/2days)15+2
Foodintake(g/2days)35 + 12
Cysteine desulphhydrationassay I (nmol of H2S/min per mg of protein)
Liver Kidney2.2+0.7 1.1 +0.1
Cystathionine y-lyase(nmol of 2-oxobutyrate/min per mg of protein)
Liver Kidney5.0+0.5 2.5 +0.2
0+2 16+4 2.0+0.1 1.1 +0.1 4.2+0.6 2.6+0.1-9+1* 17+5 0.13+0.09*t 0.24+0.07*t 0.22+0.22*t 0.57+0.20*t
14+1 38±9 2.1+0.6 1.0+0.1 4.4+ 1.1 2.1 +0.3
0+8 17+ 1 2.2+0.2 1.0+0.1 4.9+0.7 2.4+0.4-9+1* 17+1* 0.21 +0.06*t 0.26+0.08*t 0.45+0.32*t 0.78+0.29*t
control rats fed ad libitum and the pair-fed controlrats. The excretion of [35Sltaurine and the ratio of[35Sltaurine to 3SO42- in the 24h urine collectionsof rats treated with propargylglycine and intubatedwith a diet that contained 0.2% cysteine weresignificantly less than those for control rats fedad libitum. This was an unexpected finding and isprobably related to the difference in food intake ofthe two groups, as the excretion of [35Sltaurine byrats given propargylglycine and pair-fed control ratswas similar.
Table 7 summarizes the effect of administrationof DL-propargylglycine on cystathionine y-lyaseactivity and cysteine desulphhydration in assaysystem I for liver and kidney of rats treated in thesame manner as those used in the 35S-tracer study.In agreement with the results reported in Tables 4and 5, both cystathionine y-lyase activity and H2Sproduction (assay system I) from cysteine weremarkedly lower in rats given propargylglycine thanin control rats.
Discussion
Our results are consistent with the catalysis ofH2S production from cysteine by cystathioniney-lyase, cystathionine f,-synthase and cysteineaminotransferase plus 3-mercaptopyruvate sulphur-transferase. When H2S production by rat liver andkidney homogenates was assayed in incubationmixtures that contained 280mM-cysteine and 3mM-pyridoxal 5'-phosphate at pH 7.8 (assay system I),activity appeared to be due to the action of cyst-athionine y-lyase and cystathionine J8-synthase. The
inhibitory effect of propargylglycine and the activat-ing effect of S-adenosylmethionine on H2S forma-tion under these assay conditions suggested thateach enzyme catalysed a major portion of thecysteine desulphhydration, but the exact proportionof the H2S production due to the action of eachenzyme cannot be calculated from our data.When H25 production by rat liver and kidney
homogenates was determined in reaction mixturesthat contained 160 mM-cysteine, 2 mM-pyridoxal5'-phosphate, 3 mM-2-oxoglutarate and 3 mM-dithiothreitol at pH 9.7 (assay system II), activitywas due almost entirely, if not entirely, to the com-bined activities of cysteine:2-oxoglutarate amino-transferase and 3-mercaptopyruvate sulphurtrans-ferase. Neither propargylglycine nor S-adenosyl-methionine had any effect on H2S production inthis system, whereas omission of 2-oxoglutaratefrom the reaction mixture for assay II decreased theactivity by more than 90%. The tissue distribution,the concentrations of substrates and cofactors andthe incubation pH required to yield a maximalrate of H2S production and the actual values forthe activity in various tissues closely parallel thosefound for cysteine aminotransferase activity (S.-M.Kuo, T. C. Lea & M. H. Stipanuk, unpublishedwork).
With both of the more-physiological assay sys-tems, III and IV, addition or omission of 2-oxo-glutarate had little effect on the H2S productionobserved, whereas omission of pyridoxal 5'-phos-phate from the reaction mixtures and treatment oftissue homogenates with propargylglycine and withS-adenosylmethionine had relatively large and signi-
Vol. 206
275

276 M. H. Stipanuk and P. W. Beck
ficant effects. Hence both cystathionine y-lyase andcystathionine fl-synthase appeared to be active inthe catalysis of cysteine desulphhydration in thesemore-physiological systems, whereas cysteineaminotransferase, which has a much higher pHoptimum (Ubuka et al., 1978), did not appear to beinvolved.A greater inhibition of H2S production by prop-
argylglycine in liver than in kidney was consistentlyobserved. This suggested that a greater proportionof the H2S production was catalysed by cysta-thionine y-lyase in liver than in kidney, but theresults of treatment of the tissue homogenates withS-adenosylmethionine did not suggest a greaterrelative role of cystathionine fl-synthase in thecatalysis of cysteine desulphhydration in kidneythan in liver. A possible explanation for thisobservation is that S-adenosylmethionine was notspecific for increasing cystathionine fi-synthase-catalysed cysteine desulphhydration and increasedcysteine desulphhydration by other pathways.
The physiological significance of cysteine desulph-hydration is difficult to assess. Inhibition of cyst-athionine y-lyase in the intact rat by propargyl-glycine had no significant effect on urinary excretionof 35S, "S0O42- and [35Sltaurine formed from labelleddietary cysteine. The desulphhydration of cysteinemay not be a quantitatively large route of cysteinecatabolism even though it may be physiologicallyimportant as a means of generating metabolicallyactive reduced sulphur, or flux through other path-ways leading to similar end products may increasewhen cystathionine y-lyase activity is inhibited.There are several possible routes of sulphate forma-tion from cysteine, which include both oxidation ofsulphide formed by the desulphhydration pathwaysconsidered here and oxidation of sulphite releasedby desulphination of the 3-sulphinylpyruvate formedby transamination or oxidative deamination ofcysteinesulphinate. The oxidation of cysteine tocysteinesulphinate followed by decarboxylation ofcysteinesulphinate to hypotaurine or conversionof cysteinesulphinate into 3-sulphinylpyruvate isthought by many investigators to be the major path-way of cysteine degradation in animal tissues(Wheldrake & Pasternak, 1967; Yamaguchi et al.,1973; Stipanuk, 1979).The desulphhydration of cysteine appears to be
catalysed by enzymes that are also physiologicallyactive with other substrates. Cystathionine ,6-synthase apparently has a much higher Km for L-cysteine (36mM) than for L-serine (2-8mM) or L-homocysteine (0.1-9 mM) (Nakagawa & Kimura,1968; Braunstein et al., 1969; Kraus et al., 1978).The Km of cysteine: 2-oxoglutarate aminotransferasefor L-cysteine is about 22 mm, whereas its Km foraspartate is about 0.5mm (Ubuka et al., 1978). Incontrast, the Km of rat liver cystathionine y-lyase
for L-Cystine has been estimated to be about 0.07mMcompared with 0.8-3.5 mm for L-Cystathionine and15-20mM for L-homoserine (Uren et al., 1978; Yaoet al., 1979). Thus cystine may be able to competefavourably with other substrates for cystathioniney-lyase even though intracellular cystine concen-trations are low (Greenberg, 1975).
Although the physiological significance of cysteinedesulphhydration reactions in mammalian tissues isnot clear, our data indicate that cysteine desulph-hydration is catalysed by cystathionine y-lyase andcystathionine ,B-synthase and possibly by cysteineaminotransferase plus 3-mercaptopyruvate sulphur-transferase in liver and kidney incubation systemsthat contain 2 mM-L-cysteine and 0.05 mM-pyridoxal5'-phosphate at pH 7.4. These incubation conditionsapproach physiological values for substrate con-centrations and pH, and our results therefore suggestthat these enzymes are active in catalysis of cysteinedesulphhydration in the intact animal.
The authors gratefully acknowledge the excellent tech-nical assistance of Mrs. Teresa Lea and Mrs. LeahSacher. This work was supported by U.S. Public HealthService N.I.H. Grant AM-26959 and by a grant from theNutrition Foundation.
References
Abeles, R. H. & Walsh, C. T. (1973) J. Am. Chem. Soc.95, 6124-6125
AIN Ad Hoc Committee on Standards for NutritionalStudies (1977)J. Nutr. 107, 1340-1348
AIN Ad Hoc Committee on Standards for NutritionalStudies (1980) J. Nutr. 110, 1726
Braunstein, A. E., Goryachenkova, E. V. & Lac, N. D.(1969) Biochim. Biophys. Acta 171, 366-368
Braunstein, A. E., Goryachenkova, E. V., Tolosa, E. A.,Willhardt, I. H. & Yefremova, L. L. (1971) Biochim.Biophys. Acta 242, 247-260
Cavallini, D., Mondovi, B., DeMarco, C. & Scioscia-Santoro, A. (1962a) Arch. Biochem. Biophys. 96,456-457
Cavallini, D., Mondovi, B., DeMarco, C. & Scioscia-Santoro, A. (1962b) Enzymologia 24,253-266
Fiedler, H. & Wood, J. L. (1956) J. Biol. Chem. 222,387-397
Finkelstein, J. D., Kyle, W. E. & Harris, B. J. (1971)Arch. Biochem. Biophys. 146, 84-92
Finkelstein, J. D., Kyle, W. E., Martin, J. J. & Pick, A.-M.(1975) Biochem. Biophys. Res. Commun. 66, 81-87
Fogo, J. K. & Popowsky, M. (1949) Anal. Chem. 21,732-734
Greenberg, D. M. (1975) Metab. Pathways 3rd Ed. 7,505-528
Herzfeld, A. & Greengard, 0. (1971) Biochim. Biophys.Acta 237, 88-98
Ip, M. P. C., Thibert, R. J. & Schmidt, D. E., Jr. (1977)Can. J. Biochem. 55, 958-964
Ishimoto, Y. (1979) Physiol. Chem. Phys. 11, 189-19 1Koj, A. (1968) Acta Biochim. Pol. 15, 16 1-169
1982

Cysteine desulphhydration in rat tissues 277
Koj, A., Frendo, J. & Janik, Z. (1967) Biochem. J. 103,791-795
Koracevic, D. (1975) Experientia 31, 26-27Koracevic, D., Pejovic, M. & Djordjevic, V. (1979)
Arch. Int. Physiol. Biochim. 87, 105-108Kraus, J., Packman, S., Fowler, B. & Rosenberg, L. E.
(1978) J. Biol. Chem. 253, 6523-6528Kun, E. & Fanshier, D. W. (1958) Biochim. Biophys.
Acta 27, 659Kun, E. & Fanshier, D. W. (1959a) Biochim. Biophys.
Acta 32, 338-348Kun, E. & Fanshier, D. W. (1959b) Biochim. Biophys.
Acta 33, 26-28Lak, N. D., Gorgachenkova, E. V. & Braunstein, A. E.
(1970) Biokhimiya 35, 270-277Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall,
R. J. (195 1) J. Biol. Chem. 193, 265-275Meister, A., Fraser, P. E. & Tice, S. V. (1954) J. Biol.
Chem. 206, 561-575Mudd, S. H., Finkelstein, J. D., Irreverre, F. & Laster, L.
(1965)J. Biol. Chem. 240, 4382-4392Nakagawa, H. & Kimura, H. (1968) Biochem. Biophys.
Res. Commun. 32, 208-214Porter, P. N., Grishaver, M. S. & Jones, 0. W. (1974)
Biochim. Biophys. Acta 364, 129-139Rogers, Q. R. & Harper, A. E. (1965) J. Nutr. 87, 267-
273Roy, A. B. & Trudinger, P. A. (1970) The Biochemistry
of Inorganic Compounds of Sulfur, p. 69, CambridgeUniversity Press, London
Siegel, L. M. (1965) Anal. Biochem. 11, 126-132Stipanuk, M. H. (1977) Ph.D. Thesis, University of
Wisconsin, Madison; University Microfilms, AnnArbor
Stipanuk, M. H. (1979) J. Nutr. 109, 2126-2139Stipanuk. M. H. & Benevenga, N. J. (1977) J. Nutr.
107. 1455-1467Sturman, J. A., Rassin, D. K. & Gaull, G. E. (1970)
Int. J. Biochem. 1, 251-253Szczepkowski, T. W. & Wood, J. L. (1967) Biochim. Bio-
phys. Acta 139, 469-478Tallan, H. H., Moore, S. & Stein, W. H. (1958) J. Biol.
Chem. 230, 707-716Taniguchi, T. & Kimura, T. (1974) Biochim. Biophys.
Acta 64, 284-295Ubuka, T., Umemura, S., Ishimoto, Y. & Shimomura, M.
(1977a) Physiol. Chem. Phys. 9, 91-96Ubuka, T., Yuasa, S., Ishimoto, Y. & Shimomura, M.
(1977b) Physiol. Chem. Phys. 9, 241-246Ubuka, T., Umemura, S., Yuasa, S., Kinuta, M. &
Watanabe, K. (1978) Physiol. Chem. Phys. 10, 483-500
Uhteg, L. C. & Westley, J. (1979) Arch. Biochem. Bio-phys. 195, 211-222
Uren, J. R., Ragin, R. & Chaykovsky, M. (1978) Bio-chem. Pharmacol. 27, 2807-2814
Washtien, W., Cooper, A. J. L. & Abeles, R. H. (1977)Biochemistry 16, 460-463
Wheldrake, J. F. & Pasternak, C. A. (1967) Biochem. J.102, 45P-46P
Yamaguchi, K., Sakakibara, S., Asamizu, J. & Ueda, I.(1973) Biochim. Biophys. Acta 297,48-59
Yamaguchi, K., Shigehisa, S., Sakakibara, S., Hosokawa,Y. & Ueda, I. (1975) Biochem. Biophys. Acta 381,1-8
Yao, K., Kinuta, M. & Akagi, R. (1979) Physiol. Chem.Phys. 11, 257-260
Vol. 206


















