
Chapter -2shodhganga.inflibnet.ac.in/bitstream/10603/40866/12/12_chapter2.pdf · Chapter 2...
24
Chapter 2 Isolation and screening of xylanase producer 43 Chapter -2 Isolation, Screening and Identification of Potential Xylanolytic Fungal Culture for Xylanase Production Part of this chapter is published as: PRODUCTION OF XYLANASE UNDER SOLID STATE FERMENTATION BY Aspergillus tubingensis JP-1 AND ITS APPLICATION Bioprocess and Biosystems Engineering. DOI 10. 1007/s00449-011-0657-1
-
Upload
phungxuyen -
Category
Documents
-
view
219 -
download
0
Transcript of Chapter -2shodhganga.inflibnet.ac.in/bitstream/10603/40866/12/12_chapter2.pdf · Chapter 2...

Chapter 2 Isolation and screening of xylanase producer
43
Chapter -2
Isolation, Screening and Identification of Potential
Xylanolytic Fungal Culture for Xylanase Production
Part of this chapter is published as:
PRODUCTION OF XYLANASE UNDER SOLID STATE FERMENTATION
BY Aspergillus tubingensis JP-1 AND ITS APPLICATION
Bioprocess and Biosystems Engineering. DOI 10. 1007/s00449-011-0657-1

Chapter 2 Isolation and screening of xylanase producer
44
2.1 Introduction
In recent years, there has been an increasing interest in the effective utilization of
plant residues, wood crop residues for microbial degradation and transformation into
a specific product of industrial value. Agro-industrial residues such as sugarcane
bagasse, wheat straw, wheat bran, corn cobs, rice straw, rice bran, cassava waste are
considered to be the ideal substrates for enzyme production. Carbon sources used for
microbial production of both xylanase and cellulase are mostly insoluble in nature.
Hemicelluloses, the second most abundant polysaccharide next to cellulose consist of
β- 1, 4- linked pyranosyl backbone. The breakdown of hemicellulose requires the
action of various enzymes of which the endoxylanase is the most important.
Endoxylanase are responsible for the random hydrolysis of xylan, a major
hemicellulose component of agro-industrial residues, is advantageous for the
recovery of hexose and pentose sugars for use as raw materials in a wide number of
biotechnological processes (Beg et al., 2001, Kapoor et al., 2008, Saha, 2003).
The microbial endoxylanases (E.C. 3.2.1.8) have been widely isolated from a diverse
range of microorganisms, including fungi and bacteria, are known to produce
different types of xylanases (Sunna and Antranikian, 1997, Krisana et al., 2005, Lee
et al., 2009). The nature of these enzymes varies with different organisms, among
which filamentous fungi are especially interesting from an industrial point of view
due to extracellular release of xylanase, higher yield compared to bacteria and yeast
and production of several auxiliary enzymes that are necessary for debranching of
the substituted xylan. The use of cellulase free xylanase plays an important role in
animal feed, paper and pulp industry, bakery industry.
A success of any fermentation process depends both on isolation and screening of an
appropriate strain and on optimization of fermentation parameters for the production
of the enzymes (Haq et al., 2002, Khan et al., 2003). Screening of the large numbers
of micro-organisms requires efficient plate screening methods. A number of plate
screening assays for the detection of xylanases have been described in the literature
(Table 2.1).

Chapter 2 Isolation and screening of xylanase producer
45
Table 2.1 Plate screening assay for the detection of xylan degrading micro-
organisms.
Plate assay Substrate Reference
Screening of xylan degrading
microorganisms
Insoluble dye substrate Lee and Lee, 1997
Assay for endohydrolases Soluble dye labelled
polysaccharide
McCleary, 1988
Assay for endo-1,4- β-
xylanases, and endo- 1,4 β-
glucanases
Soluble chromogenic
substrate
Biely et al., 1985
Xylanase assay 2,2-bicinchoninate
(BCA)
Kenealy and Jeffries,
2003
Assay for Xylan degrading
microorganisms
Insoluble dye labeled
xylan with chromogenic
substrate
Ten et al., 2004
Assay for polysaccharide and
protein degrading organisms
Mixture of insoluble
dye labeled collagen
Ten et al., 2005
Taxonomic identification of the xylanase producing fungi is seen as an important
component of the research, since this would reveal related species that may produce
alternative varieties of xylanases. Another benefit of identification will be increased
understanding of physiological aspects and medium components required for optimal
production of xylanase from the host organism. Moreover, phylogenetic analysis is
an essential component of demonstrating novelty of the production organism for
patenting purposes, and to determine any prior isolation of a similar xylanase from a
similar host. Molecular characterization may currently be seen as the preferred
method of species identification, and several genera have undergone reclassification
based specifically on DNA sequence analysis rather than their structural or breeding
similarities. DNA sequence analysis methods are objective, reproducible and rapid
means of identification, and therefore they have been widely used (Liu et al., 2007,
Li et al., 2007, Bakri et al., 2010).The analysis of the ribosomal region by PCR
amplification using universal primers targeted to conserved regions within the

Chapter 2 Isolation and screening of xylanase producer
46
r RNA complex, followed by DNA sequencing of the internal transcribed spacer
(ITS) region has been widely used to identify a broad range of the fungi to their
species level (Henry et al., 2000, Chen et al., 2001, Pryce et al., 2003, Liu et al.,
2008).
Considering the industrial importance of the xylanases in animal feed, paper and pulp
industry, food industry, production of xylose and the production of fuel and chemical
feed stocks, an extensive isolation and screening programme was initiated to select
the potential xylanase producing microorganisms from soil and identify and
characterize the higher xylanase producing isolate.
Objectives
• To isolate potential xylanase producing fungi from the natural environment
and identification of higher xylanase producing strain.
• Screening for the production of higher xylanolytic enzymes by isolated
culture under submerged fermentation and under solid state fermentation
using various lignocellulosic wastes.
2.2 Materials and Methods
2.2.1 Chemicals
Birch wood xylan was procured from Sigma (Sigma St. Louis, MO, USA), whereas,
Potato Dextrose agar, Malt extract were procured from Hi-Media Labs, Mumbai,
India and all other chemicals used were of analytical grade.
Wheat straw, wheat bran, rice bran, sorghums straw and sugarcane bagasse were
collected locally and used as lignocellulosic substrates.

Chapter 2 Isolation and screening of xylanase producer
47
2.2.2 Isolation and Screening of Xylanase Producing Culture
2.2.2.1 Primary Screening
Samples of soil and decomposing organic matter were collected from agricultural
land, forests and gardens of Anand, Amreli and Vadodara districts of Gujarat, India,
for the isolation of xylanolytic microorganisms. The dilution plate method was
employed for the isolation of fungal strains (Johnson and Curl, 1972). One gram of
soil was suspended in 100 ml sterile distilled water. The suspension was serially
diluted to 10-7. 100 µl aliquot of the appropriately diluted samples from 10-2 to 10-7
were plated on Malt Extract Agar plate containing (g L-1) malt extract 20, birch wood
xylan 5.0, agar-agar 20 and streptomycin sulphate 0.025 to eliminate bacterial
contamination. The plates were incubated at 30°C for 4-5 days. The colonies
surrounded by zone of hydrolysis on the ME xylan agar plate were selected as
xylanase producing organism. The potential isolates were further confirmed using
Defined xylan agar (DX) media containing (g L-1): Birch wood xylan 10, KH2PO4
1.0, (NH4)2SO4 2.0, NaCl 1.0, agar-agar 20, pH 6.0) (Yang et al., 1995). The
inoculated plates were incubated for 2-4 days at 30°C. The colonies which produced
distinct zone of hydrolysis were selected for further screening in a liquid medium.
The potential isolates were sub-cultured and maintained at 4oC on potato dextrose
(PDA) agar slant and used for further study.
2.2.2.2 Quantitative Screening
Based on the preceding screening performance, five xylanolytic fungal strains were
quantitatively screened for xylanase production in Defined xylan (DX) broth ((g L-1):
Birch wood xylan 10, KH2PO4 1.0, (NH4)2SO4 2.0, NaCl 1.0, pH 6.0). Erlenmeyer
flasks (250 ml) containing 100 ml DX broth were inoculated with spore suspension
of 2 x 106 spores ml-1 of different isolated cultures and incubated on a rotatory shaker
at 120 rpm for 12 days at 30°C. The enzyme from each flask was filtered through
muslin cloth and the filtrate obtained was centrifuged at 7000 x g for 15 min. at 4°C.
The clear supernatant thus obtained was used as a source of xylanase.

Chapter 2 Isolation and screening of xylanase producer
48
2.2.2.3 Identification of the Selected Isolate
Identification of the best isolate was based on structural morphology and nucleotide
sequence analysis. PCR amplification using universal primers targeted to conserved
regions within the r RNA complex, followed by DNA sequencing of the internal
transcribed spacer (ITS) regions (partial 18S r RNA ITS 1, 5.8S r RNA ITS 2, and
partial 28S r RNA gene sequencing). Molecular identification of the potential isolate
was performed by Bangalore GeNei (Bengaluru, India).
2.2.3 Xylanase Production under SSF
Erlenmeyer flasks (250 ml) containing 5 g of each different lignocellulosic substrates
(wheat straw, wheat bran, rice bran, sugarcane bagasse, sorghum straw) were
autoclaved at 121°C for 30 min. and mixed with 25 ml of Mandels and Sternburg’s
(MS) medium (Mandels and Sternburg’s, 1976). MS medium comprising of (g L-1):
Peptone, 1.0, (NH4)2SO4, 1.4, KH2PO4, 2.0, Urea, 0.3, CaCl2, 0.3, MgSO4.7H2O, 0.3
and trace elements (mg L-l): FeSO4.7H2O, 5.0, MnSO4.H2O, 1.6, ZnSO4.7H2O, 1.4,
CoCl2, 2.0, Tween 80, 0.1% (v/v), pH 6.0.). The flasks were inoculated with 2 × 106
spores ml-1 of spore suspension prepared from a week old PDA slants of the culture
grown at 30°C. Sterile distilled water added with 0.1% (v/v) Tween 80 was used to
prepare fungal spore suspension. The inoculated flasks were incubated at 30°C under
static conditions for 10 days. The enzyme from each flask was harvested using 10 ml
of 50mM sodium citrate buffer (pH 5.0). The contents were squeezed through muslin
cloth and the enzyme extract obtained was centrifuged at 7000 x g at 4oC for 15 to 20
min. The clear supernatant thus obtained was used as the enzyme sample for assay.
2.2.4 Enzyme Assays
2.2.4.1 Xylanase (E.C. 3.2.1.8)
Xylanase activity was measured using 1% birchwood xylan (4-O methyl
glucuronoxylan) solution as a substrate (Bailey et al., 1992). The reaction mixture
contained 0.9 ml of 1% (w/v) birchwood xylan (prepared in 50 mM sodium citrate
buffer, pH 5.0) and 0.1 ml of suitably diluted enzyme extract, incubated at 50°C for
10 min. The release of reducing sugar was measured as xylose equivalents using the

Chapter 2 Isolation and screening of xylanase producer
49
dinitrosalicylic acid method (Miller, 1959). One unit of xylanase is defined as the
amount of enzyme that liberates 1μmole of reducing sugar as xylose equivalent ml-1
min-1 under assay condition.
2.2.4.1.2 β- Xylosidase (E.C. 3. 2. 1.37)
β- Xylosidase activity was measured using p-nitro phenyl-β-D-xylopyranoside
(pNβxp) as a substrate (Judith and Nei, 2002). The reaction mixture contained 1 ml
of 1 mM pNβxp (prepared in 50 mM sodium citrate buffer) and 1 ml of suitably
diluted enzyme extract. The reaction mixture was incubated at 50°C for 10 min and
the reaction was terminated by adding 2 ml of 1 M Na2CO3. One unit of β-xylosidase
is defined as the amount of enzyme required to liberate 1μmole p-nitro phenol ml-1
min-1 under assay condition.
2.2.4.1.3 Filter Paper Cellulase Activity (FPA)
Filter paper activity was measured according to IUPAC recommendations employing
filter paper as substrate (Ghose, 1987). The release of reducing sugars in 60 min at
50oC at pH 5.0 (50 mM citrate buffer) was measured as glucose equivalent using
dinitrosalicylic acid method (Miller, 1959). One unit of cellulase (FPA) is defined as
the amount of enzyme that liberates 1 μmole of reducing sugar as glucose equivalent
ml-1 min-1 under assay condition.
2.2.4.1.4 Endogluconase (1, 4-β-D-glucan-4-gluconohydrolase (E.C. 3.2.1.4))
Endogluconase (CMCase) activity was measured using carboxy methyl cellulose
(CMC) as a substrate (Mandels et al., 1974). The assay system contained 0.5 ml of
appropriately diluted enzyme with 0.5 ml of 2% (w/v) CMC in 50 mM citrate buffer,
pH 5.0. The reaction mixture was incubated at 50oC for 30 min. and the release of
reducing sugar was measured as glucose equivalents using the dinitrosalicylic acid
method (Miller, 1959). One unit of endogluconase is defined as the amount of
enzyme that liberates 1μmole of reducing sugar as glucose equivalent ml-1 min-1
under assay condition.

Chapter 2 Isolation and screening of xylanase producer
50
2.2.4.1.5 Protease The protease activity was measured using casein as the substrate (Gessesse et al.,
2003). The reaction mixture contained 1 ml of casein solution (0.5% w/v, prepared in
50 mM sodium citrate buffer) and 1 ml of suitably diluted enzyme extract. The
mixture was incubated at 50oC for 20 min and the reaction was terminated by adding
2 ml of 10% trichloro-acetic acid. The tyrosine released was measured by Lowry’s
method. One unit of protease activity is defined as the amount of enzyme that
liberates 1 μmole of tyrosine ml-1 min-1 under assay condition.
2.2.4.2 Protein Estimation
Soluble protein content was determined according to Lowry’s method (Lowry et al.,
1951) using bovine serum albumin as the standard.
2.3 Results and Discussion
Isolation and Screening of Xylanase Producing Microorganisms
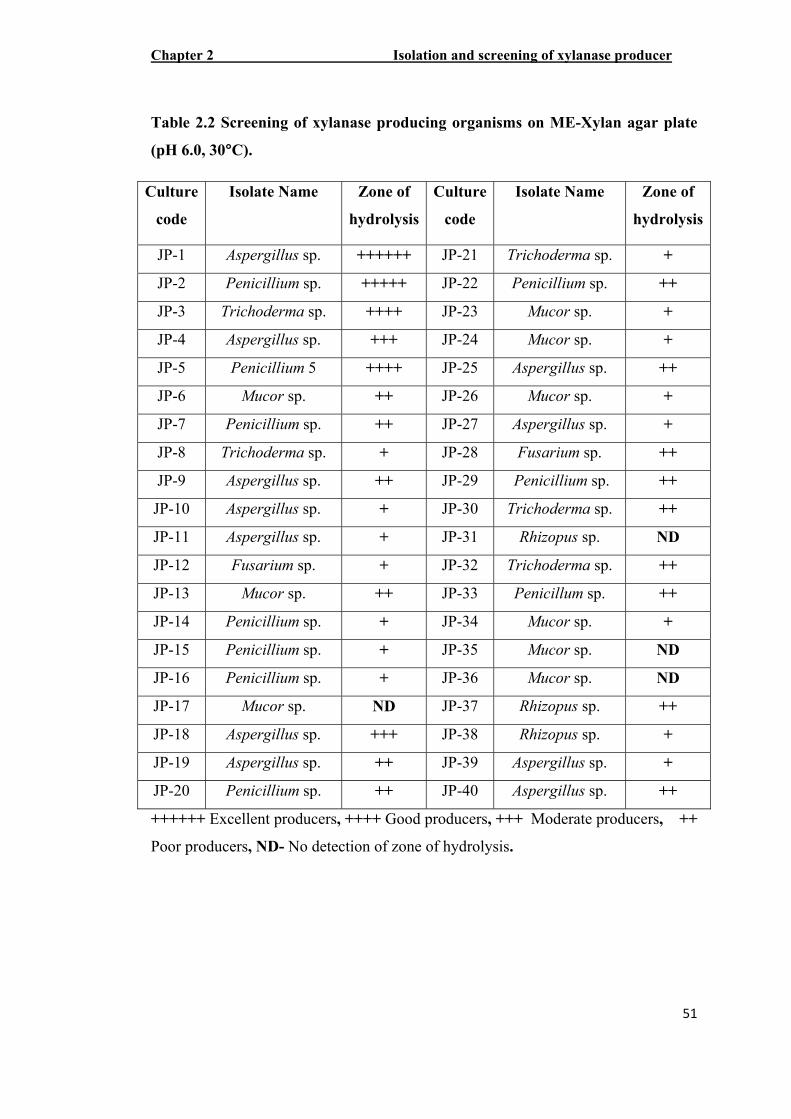
Based on the initial screening programme, a total of 40 isolates were capable of
exhibiting xylanolytic activities on ME xylan agar plate with varying zone of
clearance (Table 2.2). Among the 40 isolates, 5 isolates were selected for further
confirmation using the Defined xylan agar (DX) media (Plate. 2.1 (a-e)). The isolate
JP-1, demonstrated reproducible zones of hydrolysis of 30 to 40 mm diameter on the
DX agar plate (Table 2.3). Secondary screening (quantitative) for the xylanase
production was carried out by growing the isolates in the defined xylan broth under
shaking condition. It was observed that JP-1 produced higher xylanase (185.34 U ml-
1) on 8th day of cultivation. Compared to JP-1, lower xylanase production was
obtained with JP-2 (59.3 U ml-1), JP-3 (48.7 U ml-1), JP-4 (51.3 U ml-1), and JP-5
(43.5 U ml-1) (Table 2.4). The isolate JP-1 was found to be potent xylanase
producing microorganism and thus selected for our further studies.
Chapter 2 Isolation and screening of xylanase producer
51
Table 2.2 Screening of xylanase producing organisms on ME-Xylan agar plate
(pH 6.0, 30°C).
Culture
code
Isolate Name Zone of
hydrolysis
Culture
code
Isolate Name Zone of
hydrolysis
JP-1 Aspergillus sp. ++++++ JP-21 Trichoderma sp. +
JP-2 Penicillium sp. +++++ JP-22 Penicillium sp. ++
JP-3 Trichoderma sp. ++++ JP-23 Mucor sp. +
JP-4 Aspergillus sp. +++ JP-24 Mucor sp. +
JP-5 Penicillium 5 ++++ JP-25 Aspergillus sp. ++
JP-6 Mucor sp. ++ JP-26 Mucor sp. +
JP-7 Penicillium sp. ++ JP-27 Aspergillus sp. +
JP-8 Trichoderma sp. + JP-28 Fusarium sp. ++
JP-9 Aspergillus sp. ++ JP-29 Penicillium sp. ++
JP-10 Aspergillus sp. + JP-30 Trichoderma sp. ++
JP-11 Aspergillus sp. + JP-31 Rhizopus sp. ND
JP-12 Fusarium sp. + JP-32 Trichoderma sp. ++
JP-13 Mucor sp. ++ JP-33 Penicillum sp. ++
JP-14 Penicillium sp. + JP-34 Mucor sp. +
JP-15 Penicillium sp. + JP-35 Mucor sp. ND
JP-16 Penicillium sp. + JP-36 Mucor sp. ND
JP-17 Mucor sp. ND JP-37 Rhizopus sp. ++
JP-18 Aspergillus sp. +++ JP-38 Rhizopus sp. +
JP-19 Aspergillus sp. ++ JP-39 Aspergillus sp. +
JP-20 Penicillium sp. ++ JP-40 Aspergillus sp. ++
++++++ Excellent producers, ++++ Good producers, +++ Moderate producers, ++
Poor producers, ND- No detection of zone of hydrolysis.

Chapter 2 Isolation and screening of xylanase producer
52
Table 2.3 Screening of potential xylanase producing organisms on defined
Xylan (DX) agar plate (pH 6.0, 30°C).
Culture code Isolate Name Zone diameter (mm)
JP-1 Aspergillus sp. 40
JP-2 Penicillium sp. 30
JP-3 Trichoderma sp. 29
JP-4 Aspergillus sp. 25
JP-5 Penicillium sp. 28
Plate 2.1 (a) Plate 2.1 (b)
Plate 2.1 (c) Plate 2.1 (d)

Chapter 2 Isolation and screening of xylanase producer
53
Plate 2.1 (e) Plate 2.1 (f)
Plate 2.1 Plate assay showing zone of hydrolysis by a xylanase
producing isolates (a) Aspergillus sp. JP-1 grown on ME xylan agar
and (b) JP-1 (c) JP-2, (d) JP-3, (e) JP-4, (f) JP-5 , grown on DX agar
medium.
Table 2.4 Xylanase production by potent isolates using Defined xylan broth at
30°C under shaking condition for 12 days.
Aspergillus sp. JP-1
Penicillium sp. JP-2
Trichoderma sp. JP-3
Aspergillus sp. JP-4
Penicillium sp. JP-5 Days Xylanase
activity (U ml-1)
Xylanase activity (U ml-1)
Xylanase activity (U ml-1)
Xylanase activity (U ml-1)
Xylanase activity (U ml-1)
1 11.42 5.35 4.7 4.6 3.5 2 31.87 17.5 15.8 18.6 10.7 3 50.11 28.36 24.42 22.9 19.3 4 84.29 40.9 33.87 30.6 24.3 5 111.8 58.9 42.9 38.9 33.8 6 126.35 62.3 45.2 43.14 38.4 7 173.19 65 44.2 49.5 40.9 8 185.34 59.3 48.7 51.3 43.5 9 173.19 51.8 45.7 44.8 45.9 10 138.47 46.2 45.01 41.2 40.4 11 125.5 39.35 28.15 31.6 29.6 12 101.8 22.72 23.81 20.12 24.2

Chapter 2 Isolation and screening of xylanase producer
54
Identification of Potential Xylanase Producer (JP-1)
Plate 2.2 Growth of the pure culture of JP-1 on Potato dextrose agar plate obtained after successive subculture.
Plate 2.3 Microscopic observation of the purified culture JP-1.
The initial identification of the isolate JP-1 was based on its morphological
characteristics. The colonial morphology of Aspergillus sp. JP-1 studied on PDA
(Potato dextrose agar) plate at 30°C shows that the initial white mycelia turns yellow
and finally to black upon maturation (Plate 2.2). The microscopic observation
showed that the isolate possessed distinct conidiophores terminated by a swollen
vesicle bearing flask shaped phialides. Spores show black coloration and were
produced in long chains from the ends of the phialides (Plate 2.3). The isolate JP-1
was microscopically identified to be a member of Aspergillus. The identification of
the isolate further confirmed using partial 18S r RNA, ITS1, 5.8S r RNA, ITS2 and
28S r RNA gene sequencing performed by Bangalore Genei, Bengaluru, India.
BLAST similarity search analysis based on ITS and 18S rRNA gene sequences,
revealed that the isolate belong to the genus Aspergillus. The closest phylogenetic
neighbour was found to be Aspergillus tubingensis EF 634380 (NRRL 35179) with
100% homology (Table 2.5). Through the alignment and cladistic analysis of
homologous nucleotide sequences of known fungi, phylogenetic relationship could
be inferred and the approximate phylogenetic position of the strain is shown in
Fig.2.1. The gene sequence has been deposited in the GenBank database of NCBI

Chapter 2 Isolation and screening of xylanase producer
55
under accession number EU 867248 (1147 bp) (Fig 2.2). The strain Aspergillus sp.
JP-1 was thus identified as Aspergillus tubingensis and named as Aspergillus
tubingensis JP-1.
EF634379 (Aspergillus niger isolate NRRL 35177)
EF634376 (Aspergillus niger isolate NRRL 35173)
JP1
EF634380 (Aspergillus tubingensis isolate NRRL
100
EF669601 (Aspergillus allahabadii isolate NRRL
U65304 (Aspergillus heteromorphus)
AF338641 (Aspergillus bombycis st. NRRL 29253)
AF104444 (Aspergillus bombycis st. NRRL 26010)
100
EF634403 (Aspergillus fumigatus isolate NRRL
AF459734 (Aspergillus carbonarius NRRL 4849)
AF459731 (Aspergillus carbonarius NRRL 346)
100
100
100
100
100
69
Figure 2.1 Phylogenetic dendogram of Aspergillus tubingensis JP-1 based on 18S rRNA, 5.5S rRNA, and partial 28S rRNA gene sequencing. Numbers following the names of the strains are accession numbers of published sequences. The bootstrapped unrooted tree was constructed by Neighbour-Joining method from the distance data generated by multiple alignments of nucleotide sequences.

Chapter 2 Isolation and screening of xylanase producer
56
AAGGATCATTACCGAGTGCGGGTCCTTTGGGCCCAACCTCCCATC
CGTGTCTATTATACCCTGTTGCTTCGGCGGGCCCGCCGCTTGTCGG
CCGCCGGGGGGGCGCCTTTGCCCCCCGGGCCCGTGCCCGCCGGA
GACCCCAACACGAACACTGTCTGAAAGCGTGCAGTCTGAGTTGAT
TGAATGCAATCAGTTAAAACTTTCAACAATGGATCTCTTGGTTCC
GGCATCGATGAAGAACGCAGCGAAATGCGATAACTAATGTGAAT
TGCAGAATTCAGTGAATCATCGAGTCTTTGAACGCACATTGCGCC
CCCTGGTATTCCGGGGGGCATGCCTGTCCGAGCGTCATTGCTGCC
CTCAAGCCCGGCTTGTGTGTTGGGTCGCCGTCCCCCTCTCCGGGG
GGACGGGCCCGAAAGGCAGCGGCGGCACCGCGTCCGATCCTCGA
GCGTATGGGGCTTTGTCACATGCTCTGTAGGATTGGCCGGCGCCT
GCCGACGTTTTCCAACCATTTTTTCCAGGTTGACCTCGGATCAGGT
AGGGATACCCGCTGAACTTAAGCATATCAATAAGCGGAGGAAAA
GAAACCAACCGGGATTGCCTCAGTAACGGCGAGTGAAGCGGCAA
GAGCTCAAATTTGAAAGCTGGCTCCTTCGGAGTCCGCATTGTAAT
TTGCAGAGGATGCTTTGGGTGCGGCCCCCGTCTAAGTGCCCTGGA
ACGGGCCGTCAGAGAGGGTGAGAATCCCGTCTTGGGCGGGGTGT
CCGTGCCCGTGTAAAGCTCCTTCGACGAGTCGAGTTGTTTGGGAA
TGCAGCTCTAAATGGGTGGTAAATTTCATCTAAAGCTAAATACTG
GCCGGAGACCGATAGCGCACAAGTAGAGTGATCGAAAGATGAAA
AGCACTTTGAAAAGAGAGTTAAACAGCACGTGAAATTGTTGAAA
GGGAAGCGCTTGCGACCAGACTCGCCCGCGGGGTTCAGCCGGCA
TTCGTGCCGGTGTACTTCCCCGTGGGCGGGCCAGCGTCGGTTTGG
GCGGCCGGTCAAAGGCCCCTGGAATGTAGTACCCTCCGGGGT
ACCTTATAGCCAGGGGTGCAATGCGGCCAGCCTGGACCGAGGAA
CGCGCTTCGGCACGGACGCTGGCATAATGGTCG
Figure 2.2 Aligned sequence data (1147 bp).

Chapter 2 Isolation and screening of xylanase producer
57
Table 2.5 Percentage homology based on nucleotide sequence of isolate JP-1 with other species of the Aspergillus genera.
SL. No.
ISOLATES PERCENTAGE HOMOLGY
1 2 3 4 5 6 7 8 9 10 11
1 JP1 * 100 99 99 92 96 92 92 92 97 97
2 EF634380 * 99 99 92 96 92 92 92 97 97
3 EF634376 * 100 92 96 92 92 92 97 97
4 EF634379 * 92 96 92 92 92 97 97
5 EF669601 * 92 91 91 93 92 92
6 U65304 * 93 93 92 96 96
7 AF338641 * 100 90 92 92
8 AF104444 * 90 92 92
9 EF634403 * 91 91
10 AF459734 * 100
11 AF459731 *

Chapter 2 Isolation and screening of xylanase producer
58
Screening of Xylanase Production under SSF
The technique for quantitative screening provided more valuable information than
qualitative analysis. Most studies on the production of xylanolytic enzymes by fungi
have been performed in liquid cultures, which do not reflect the natural living
conditions of these micro-organisms. Selection of an appropriate substrate is a key
factor in SSF which determines the success of the process. To minimize the
production cost, wheat straw was used for xylanase production under SSF system.
Five isolates (JP-1 to JP-5) were selected for xylanase production in solid state
fermentation. The crude enzyme extract obtained was subjected to enzyme profile
studies. Table 2.6 shows activity of xylanase and other related enzymes such as
Protease, Cellulase and Endoglucanase (CMCase). Higher xylanase production (1478
+ 11.03 U g-1) was obtained with isolate JP-1 after 8th day of cultivation (Table 2.6).
The amount of protease produced was low which may be due to the presence of
carbon sources indicating catabolic repression of its biosynthesis (Pandey et al.,
2000). Xylanase production in SSF was much higher than that in submerged
fermentation (SmF). Malarvizhi et al., (2003) reported 30 fold enhancement of
xylanase production under SSF than liquid culture with Ganoderma lucidum using
wheat bran as a substrate.

Chapter 2 Isolation and screening of xylanase producer
59
Table 2.6 Enzyme production by different isolates under solid state fermentation using wheat straw as a substrate at 30°C.
Isolate name
Xylanase
Activity
U g-1
Cellulase
Activity
(FPA) U g-1
Protease
Activity
U g-1
CMCase
Activity
U g-1
β- Xylosidase
Activity
U g-1
Protein
mg g-1
Aspergillus sp.JP-1 1478 + 11.3 0.669 + 0.17 0.608 + 0.13 0.117 + 0.08 4.3 + 0.2 11.03 + 0.12
Penicililum sp.JP-2 198.7 + 9.5 0.873 + 0.12 2.13 + 0.16 0.695 + 0.14 1.2 + 0.1 6.37 + 0.17
Trichoderma sp.JP-3 210.4 + 13.5 1.34 + 0.15 0.439 + 0.14 1.02 + 0.11 0.9 + 0.4 5.78 + 0.18
Aspergillus sp.JP-4 227.3 + 9.8 1.03 + 0.11 0.976 + 0.09 0.897 + 0.11 2.7 + 0.1 8.72 + 0.11
Penicillium sp.JP-5 289.4 + 13.2 0.562 + 0.07 0.389 + 0.1 0.582 + 0.13 1.6 + 0.09 5.86 + 0.19

Chapter 2 Isolation and screening of xylanase producer
60
Xylanase Production under SSF using Aspergillus tubingensis JP-1
Carbon source is the most important factor during the growth and metabolic process
of micro-organisms. The choice of an appropriate carbon source and the cost of
substrate play a crucial role in the economics of xylanase production. Various agro-
waste residue viz., wheat straw, wheat bran, rice bran, sorghum straw and sugarcane
bagasse were studied using MS medium as the moistening agents in the proportion of
1:5. Among the lignocellulosic materials tested as carbon source, wheat straw was
found to be far more effective for xylanase production. Maximum xylanase and β-
xylosidase production obtained was (1478 ± 11.31 U g-1) and (4.3± 0.2 U g-1) on 8th
day of cultivation using wheat straw as a substrate. Further increase in the
fermentation time, resulted in reduction of xylanase activity. In comparison with
wheat straw, when sorghum straw, rice bran, wheat bran and sugarcane bagasse were
used as substrates, 30, 66, 76, and 98% decrease in xylanase yield was observed,
respectively (Fig. 2.3). Cellulase and protease production were negligible with all the
other substrates used. Maximum cellulase activity (0.693+ 0.01 U g-1) and protease
activity (0.713 + 0.11 U g-1) were obtained with sorghum straw on 8th day of
cultivation (Table 2.7). Significant difference in xylanase production was observed,
when wheat straw was used as a carbon source. This may be attributed to its
hemicelluloses nature, favorable degradability and the presence of some nutrients
(Gomes et al., 1993). The results obtained with wheat straw as a choice of substrates
are in agreement with Bakri et al., (2003) and Yang et al., (2006) who reported the
use of wheat straw for xylanase production using Penicillium canescens and
Paecilomyces themophila, respectively.

Chapter 2 Isolation and screening of xylanase producer
61
Figure 2.3 Effect of different lignocellulosic substrates on xylanase production
by Aspergillus tubingensis JP-1.
Table 2.7 Xylanase production under Solid State Fermentation using different
lignocellulosic substrates at 30°C by Aspergillus tubingensis JP-1.
Substrates
Xylanase
Activity
(U g-1)
Protease
Activity
(U g-1)
Cellulase
Activity
(FPA) (U g-1)
Protein
(mg g-1)
Wheat Bran 364.11 + 12.71 0.557 + 0.09 0.024 + 0.005 13.16 + 0.11
Rice Bran 508.65 + 16.48 1.12 + 0.11 0.037 + 0.005 6.89 + 0.09
Sugarcane bagasse 17.15 + 2.87 0.678 + 0.11 0.064 + 0.002 3.11 + 0.11
Wheat straw 1478.5 + 11.31 0.608 + 0.13 0.669 + 0.007 10.99 + 0.13
Sorghum straw 1038.5 + 5.36 0.713 + 0.11 0.693 + 0.01 11.03 + 0.12

Chapter 2 Isolation and screening of xylanase producer
62
2.4 Conclusion
Large numbers of samples from various niches are necessary to be screened in order
to obtain potential strains of importance. Soil Samples collected from various regions
of Gujarat were screened for isolation of xylanase producers. Among the 40 different
cultures isolated, 5 cultures were capable of exhibiting higher xylanolytic activities.
Quantitative estimation for xylanase production was performed to find an efficient
xylanase producer from the isolated cultures. The isolate JP-1 was found to be the
most efficient producer of xylanase with negligible cellulase and protease. The
isolate JP-1 was later identified as Aspergillus tubingensis JP-1 (GenBank Accession
No. EU 867248) by partial 18S r RNA, ITS1, 5.8S r RNA, ITS2 and partial 28S r
RNA gene sequencing. Maximum production of xylanase by Aspergillus tubingensis
JP-1 was obtained with wheat straw on 8th day of cultivation (1478 + 11.31 U g-1 of
dry substrate). It is also important to determine the economically suitable optimized
medium and suitable culture conditions in which the isolate can produce high amount
of xylanase. The ability to produce high titers of xylanase with poor cellulase and
protease makes this strain a promising entrant for industrial applications.

Chapter 2 Isolation and screening of xylanase producer
63
2.5 References
Bailey, M.J., Biely, P., and Poutanen, K. (1992). Inter laboratory testing of methods for assay of xylanase activity. J. Biotechnol. 23: 257-270. Bakri, Y., Jacqes, P., and Thonort, P. (2003). Xylanase production by Penicillium canescence 10-10c in solid state fermentation. Appl. Biochem. Biotechnol. 105-108: 737-747. Bakri, Y., Masson, M., and Thonart, P. (2010). Isolation and identification of two new fungal strains for xylanase production. Appl. Biochem. Biotechnol. 162: 1626-1634. Beg, Q.K., Kapoor, M., Mahajan, L., and Hoondal, G.S. (2001). Microbial xylanases and their industrial applications: a review. App. Microbiol. Biotechnol. 56: 326-338. Biely, P., Mislovicova, D., and Toman, R. (1985). Soluble chromogenic substrates for the assay of endo-1,4 β- xylanases and endo- 1,4 β-gluconases. Ann. Biochem. 144: 142-146. Chen, Y.C., Eisner, J.D., Kattar, M.M., Rassoulian-Barrett, S.L., Lafe, K., Bui, U., Limaya, A.P., and Cookson, B.T. (2001). Polymorphic internal transcribed spacer region 1 DNA sequences identify medically important yeasts. J. Clin. Microbiol. 39: 4042-4051. Gessesse, A., Rajni, H.K., Berhanu, A.G., and Mattiasson, B.O. (2003). Novel alkaline proteases from alkaliphilic bacteria grown on chicken feather. Enzyme Microb. Technol. 32: 519-524. Ghose, T.K. (1987). International Union of Pure and Applied Chemistry, Measurement of Cellulase activities. Pure & Appl. Chem. 59: 257-268. Gomes, J., Purkarthofer, H., Hyan, M., Kapplmuller, M., Sinner, N., and Steiner, W. (1993). Production of high levels of cellulase- free xylanase by Thermomyces lanuginous in laboratory scale and pilot scale using lignocellulosic materials. Appl. Microbiol. Biotechnol. 39: 700-707. Haq, I.U., Idress, S., and Rajoka, M.I. (2002). Production of lipases by Rhizopus oligosporos by solid state fermentation. Proc. Biochem. 37: 637-641.

Chapter 2 Isolation and screening of xylanase producer
64
Henry, T., Iwen, P.C., and Hinrichs, S.H. (2000). Identification of Aspergillus species using internal transcribed spacer region 1 and 2. J. Clin. Microbiol. 38: 1510-1515. Johnson, L.I., and Curl, E.A. (1972). Methods for Research on Ecology of Soil–bornPathogens. Burgess Pub. Co, Minneapolis. P. 247. Judith, L.S.L., and Nei, P.J. (2002). Influence of some sugars on xylanase production by Aspergillus awamori in Solid State Fermentation. Braz. Arch. Biol. Technol. 45: 431-437. Kapoor, M., Nair, L.M., and Kuhad, R.C. (2008). Cost-effective xylanase production from free and immobilized Bacillus pumilus strain MK001 and its application in saccharification of Prosopis juliflora. Biochem. Eng. J. 38: 88-97. Kenealy, W.R., and Jeffries, T.W. (2003). Rapid 2,2-bicinchoninic-based xylanase assay compatible with high throughput screening. Biotechnol. Lett. 25: 1619-1623. Khan, A., Ul-Haq, I., Butt, W.A., and Ali, S. (2003). Isolation and screening of Aspergillus niger isolate for xylanase biosynthesis. Biotechnol. 2: 185-190. Krisana, A., Rutchadaporn, S., Jarupan, G., Lily, E., Sutipa, T., and Kanyawim, K. (2005). Endo-1,4-β-xylanase from Aspergillus cf. niger BCC14405 isolated in Thailand: purification, characterization and gene isolation. J. Biochem. Mol. Bio. 38: 17-23.
Lee, J.W., Park, J.Y., Kwon, M., and Choi, I. G. (2009). Purification and characterization of thermostable xylanase from the brown-rot fungus Laetiporus sulphurous. J. Biosci. Bioeng. 107: 33-37. Lee, S.T., and Lee, J.J. (1997). Insoluble dye substrate for screening and assay of xylan degrading enzymes. J. Microbiol. Methods. 29: 1-5. Li, Y., Liu, Z., Cui, F., Xu, Y., and Zhao, H. (2007). Production of xylanase from a newely isolated Penicillium sp. ZH-30. World. J. Microbiol. Biotechnol. 23: 837-843. Liu, Z.Q., Hu, Z.Z.C., Zheng, Y.G., and Shen, Y.C. (2008). Optimization of cultivation conditions for the production of 1,3- dihydroxyacetone by Pichia membranifaciens using response surface methodology. Biochem. Eng. J. 38: 285-291.

Chapter 2 Isolation and screening of xylanase producer
65
Liu, Z.Q., Li, Y., Ping, L.F., Xu, Y.Y., Cui, F.G., and Zheng, Y.G. (2007). Isolation and identification of a novel Rhodococcus sp. ML-0004 producing epoxide hydrolase and optimization of enzyme production. Proc. Biochem. 42: 889-894. Lowry, O.H., Rosebrough, N.J., Farr, A.L., and Randall, R.J. (1951). Protein measurements with Folin- phenol reagent. J. Biol. Chem. 193: 265-275. Malarvizhi, K., Murugesan, K., and Kalaichelvan, P.T. (2003). Xylanase production by Ganoderma lucidum on liquid and solid state fermentation. Ind. J. Exp. Biol. 41: 620-626. Mandels, M., and Sternberg, D. (1976). Recent advances in cellular technology. J Ferment. Technol. 54: 267-286. Mandels, M., Hontz, I., and Nystrom, J. (1974). Enzymatic hydrolysis of waste cellulose. Biotechnol. Bioeng. 26: 1471-1493. McCleary, B.V. (1988). Soluble, dye labelled polysaccharides for the assay of endohydrolases. Method. Enz. 160: 74-86. Miller, G.L. (1959). Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31: 426-428. Pandey, A., Soccol, C.R., and Mitchell, D. (2000). New development in Solid State Pryce, T.M., Palladino, S., Kay, I.D., and Coombs, G.W. (2003). Rapid identification of fungi by sequencing the ITS1 and ITS 2 regions using an automated capillary electophoresis system. Med. Mycol. 41: 369-381. Saha, B.C. (2003). Hemicellulose bioconversion: a review. J. Ind. Microbiol. Biotechnol. 30: 279-291. Sunna, A., and Antranikian, G. (1997). Xylanolytic enzymes from fungi and bacteria. Critic. Rev. Biotechnol. 17: 39-67. Ten, L.N., Im, W.T., Kim, M.K., and Lee, S.T. (2005). A plate assay for simultaneous screening of polysaccharide and protein –degrading microorganisms. Lett. App. Microbiol. 40: 92-98. Ten, L.N., Im, W.T., Kim, M.K., Kang, M.S., and Lee, S.T. (2004). Development of a plate technique for screening of polysaccharide-degrading microorganisms by

Chapter 2 Isolation and screening of xylanase producer
66
using a mixture of insoluble cheomogenic substrates. Microbiol. Method. 56: 375-382.
Yang, S.Q., Yan, Q.J., Jiang, Z.Q., Li, L.T., Tian, H.M., and Wang, Y.Z. (2006), High –level of xylanase production by the thermophilic Paecilomyces themophila J18 on wheat straw in solid state fermentation. Biores. Technol. 97: 1794-1800. Yang, V.W., Zhuang, Z., Elegir, G., and Jeffries, T.W. (1995). Alkaline-active xylanase produced by an alkaliphilic Bacillus sp. isolated from kraft pulp. J. Ind. Microbiol. 15: 434-441.


















