Chapter 7 · An overview of six major functions of membrane proteins Figure 7.9 Transport. (left) A...
71
Chapter 7 Membrane Structure and Function
Transcript of Chapter 7 · An overview of six major functions of membrane proteins Figure 7.9 Transport. (left) A...

Chapter 7Membrane Structure and Function

CELL MEMBRANE:
Basics:
Regulates flow in and out of the cell
Composed of phospholipids and proteins -some carbohydrates and lipids
SELECTIVELY PERMEABLE
- allows some things across more easily than others
Structure varies from one cell type to another
determines function of the membrane


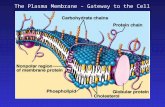
Membrane Structure
Cellular membranes are fluid mosaics of lipids and proteins
Phospholipids
Are the most abundant lipid in the plasma membrane
Are amphipathic, containing both hydrophobic and hydrophilic regions
Organized in a bilayer

Hydrophilic
head
Hydrophobic
tail
WATER
WATER

Membrane Structure
States that a membrane is a fluid structure with a “mosaic” of various proteins embedded in it
Proteins are either integral or peripheral
Integral – embedded in the membrane and traverse the phospholipid bilayer
- transmembranal
Peripheral – on the surface
- on both the inner and outer surfaces
- surfaces differ – membrane is bifacial or asymmetric


Singer and Nicolson Model Proposed that membrane proteins are
dispersed and individually inserted into the phospholipid bilayer
Figure 7.3
Phospholipid
bilayer
Hydrophobic
region
of protein
Hydrophobic region of protein

Freeze-fracture studies of the plasma membrane
Supported the fluid mosaic model of membrane structure
• A cell is frozen and fractured with a knife. The fracture plane often follows the hydrophobic interior of a membrane, splitting the phospholipid bilayer into two separated layers. The membrane proteins go wholly with one of the layers.
Coat the split layers with a metal, usually gold or silver, and examine it under an electron scanning microscope

APPLICATION A cell membrane can be split into its two layers, revealing the ultrastructure of the membrane’s interior.
TECHNIQUE
Extracellular
layer
Proteins
Cytoplasmic layer
Knife
Plasma
membrane
These SEMs show membrane proteins (the “bumps”) in the two layers,
demonstrating that proteins are embedded in the phospholipid bilayer. RESULTS

Chemical Nature of Bilayer
Amphipathic Parts Match Up
- non-polar portions of proteins embed in the hydrophobic tails of the bilayer
- polar portions of proteins embed in the hydrophilic heads or jut into the interior or exterior of the cell

The Fluidity of Membranes
Membrane parts are held together by hydrophobic interactions - weak bonds so phospholipids move around laterally(Side to side)
very rare for a phospholipid to flip-flopfrom one side of the membrane to the other

The Fluidity of Membranes
Figure 7.5 A
Lateral movement
(~107 times per second) Flip-flop(~ once per month)
(a) Movement of phospholipids

Movement of Molecules
phospholipids move quickly
Slows with temperature decrease until freezes
Unsaturated tails - more flowing
Saturated tails - more viscous

Figure 7.5 B
Fluid Viscous
Unsaturated hydrocarbontails with kinks
Saturated hydro-
Carbon tails
(b) Membrane fluidity

Role of cholesterol
wedges between phospholipids and stabilizes structure
hinders close packing decreases freezing point of membrane
Figure 7.5 (c) Cholesterol within the animal cell membrane
Cholesterol

Proteins move slowly
larger
anchored to the cytoskeleton
some travel along the cytoskeleton
some immobile

Movement of Proteins in the Plasma Membrane
EXPERIMENT Researchers labeled the plasma mambrane proteins of a mouse
cell and a human cell with two different markers and fused the cells. Using a microscope,
they observed the markers on the hybrid cell.
Membrane proteins
Mouse cell
Human cell
Hybrid cell
Mixed
proteins
after
1 hour
RESULTS
CONCLUSION The mixing of the mouse and human membrane proteins
indicates that at least some membrane proteins move sideways within the plane
of the plasma membrane.Figure 7.6
+

TYPES OF STRUCTURES ATTACHED TO MEMBRANES1) Proteins:
Integral - transmembranal - interior and exterior end
Peripheral - surface - on outside attached to membrane but not imbedded
- attached to extracellular matrix and cytoskelton
2) Carbohydrates –
glycocalyx
glycoproteins
glycolipids
FUNCTIONS:
cell stickyness
cell communication and recognition

Figure 7.7
Glycoprotein
Carbohydrate
Microfilaments
of cytoskeleton Cholesterol Peripheral
proteinIntegral
proteinCYTOPLASMIC SIDE
OF MEMBRANE
EXTRACELLULAR
SIDE OF
MEMBRANE
Glycolipid
Membrane Proteins and Their Functions
Fibers of
extracellular
matrix (ECM)

Integral proteins
Penetrate the hydrophobic core of the lipid bilayer
Are often transmembrane proteins, completely spanning the membrane
EXTRACELLULAR
SIDE
Figure 7.8
N-terminus
C-terminus
a HelixCYTOPLASMIC
SIDE

Peripheral proteins
Are appendages loosely bound to the surface of the membrane

An overview of six major functions of membrane proteins
Figure 7.9
Transport. (left) A protein that spans the membrane
may provide a hydrophilic channel across the
membrane that is selective for a particular solute.
(right) Other transport proteins shuttle a substance
from one side to the other by changing shape. Some
of these proteins hydrolyze ATP as an energy ssource
to actively pump substances across the membrane.
Enzymatic activity. A protein built into the membrane
may be an enzyme with its active site exposed to
substances in the adjacent solution. In some cases,
several enzymes in a membrane are organized as
a team that carries out sequential steps of a
metabolic pathway.
Signal transduction. A membrane protein may have
a binding site with a specific shape that fits the shape
of a chemical messenger, such as a hormone. The
external messenger (signal) may cause a
conformational change in the protein (receptor) that
relays the message to the inside of the cell.
(a)
(b)
(c)
ATP
Enzymes
Signal
Receptor

Cell-cell recognition. Some glyco-proteins serve as
identification tags that are specifically recognized
by other cells.
Intercellular joining. Membrane proteins of adjacent cells
may hook together in various kinds of junctions, such as
gap junctions or tight junctions (see Figure 6.31).
Attachment to the cytoskeleton and extracellular matrix
(ECM). Microfilaments or other elements of the
cytoskeleton may be bonded to membrane proteins,
a function that helps maintain cell shape and stabilizes
the location of certain membrane proteins. Proteins that
adhere to the ECM can coordinate extracellular and
intracellular changes (see Figure 6.29).
(d)
(e)
(f)
Glyco-
protein
Figure 7.9

Synthesis and Sidedness of Membranes
Secreted proteins are on the inside of secretory vesicles
Proteins on the outside of the cell membraneare on the inside of a secretory vesicle.
Proteins on the inside of the cell membraneare on the outside of the secretory vesicle.

ER
Figure 7.10
Transmembrane
glycoproteins
Secretory
protein
Glycolipid
Golgi
apparatus
Vesicle
Transmembrane
glycoprotein
Membrane glycolipid
Plasma membrane:
Cytoplasmic face
Extracellular face
Secreted
protein
4
1
2
3

Traffic Across Membranes
movement of materials is dependent upon the make up of the membrane
certain substances move more easily
certain substances move more quickly

The Permeability of the Lipid BilayerHydrophobic molecules
Are lipid soluble and can pass through the membrane rapidly
Polar molecules
Do not cross the membrane rapidly
Transports:
hydrocarbons
carbon dioxide
oxygen
small polar molecules with a neutral charge (H2O)
Impedes:
ions
polar molecule
large, uncharged polar molecules - glucose

Transport Proteins:
Create channels for ions and polar molecules to pass through the lipid bilayer
Selectively transport like enzymes - substrate specific
each protein transports one specific molecule
Methods:
hydrophilic channel
bind substance and move it across the bilayer

Types of Transport Across MembranePassive vs. Active
Passive: does not require energy
downhill process
moves from high concentration to low concentration with the concentration gradient (comparison of concentration levels across a membrane)
Active: Requires energy
uphill process
moves from low concentration to high concentration
against the concentration gradient
requires transport proteins

Types of Passive Transport
Diffusion
Osmosis
Facilitated Diffusion

Diffusion
based on thermal kinetics – particles move throughout a space until they are distributed evenly
movement of particles from high concentration to low concentration until net flow is zero
NET FLOW: change in concentrations due to flow across membrane -
each substance moves independently of the others
continues until equilibrium is reached

Diffusion
Figure 7.11 A
Diffusion of one solute. The membrane
has pores large enough for molecules
of dye to pass through. Random
movement of dye molecules will cause
some to pass through the pores; this
will happen more often on the side
with more molecules. The dye diffuses
from where it is more concentrated
to where it is less concentrated
(called diffusing down a concentration
gradient). This leads to a dynamic
equilibrium: The solute molecules
continue to cross the membrane,
but at equal rates in both directions.
Molecules of dye Membrane (cross section)
Net diffusion Net diffusion Equilibrium
(a)

Figure 7.11 B
Diffusion of two solutes. Solutions of
two different dyes are separated by a
membrane that is permeable to both.
Each dye diffuses down its own concen-
tration gradient. There will be a net
diffusion of the purple dye toward the
left, even though the total solute
concentration was initially greater on
the left side.
(b)
Net diffusion
Net diffusion
Net diffusion
Net diffusion Equilibrium
Equilibrium

Osmosis- special case of diffusion - APPLIES TO WATER ONLY
movement of water across the membrane to balance solute concentration
movement of water continues directionally until solute concentrations are equal
Based on relative comparisons of solute concentration (tonicity)
hypertonic: solution with more solute
hypotonic: solution with less solute
isotonic: solutions with equal concentrations
Movement of water: 2 ways to say the same thing:
from least solute to higher solute
from high water concentration to low water concentration

Figure 7.12
Net Flow

Water Balance of Cells Without Walls
Isotonic
The concentration of solutes is the same as it is inside the cell
There will be no net movement of water

Hypertonic
The concentration of solutes is greater than it is inside the cell
The cell will lose water

Hypotonic
The concentration of solutes is less than it is inside the cell
The cell will gain water

Water balance in cells without walls
Figure 7.13
H2O H2O H2O H2O
Lysed Normal Shriveled
Identify the Tonicity

Water balance in cells without walls
Figure 7.13
Hypotonic solution Isotonic solution Hypertonic solution
H2OH2O H2O H2O
Lysed Normal Shriveled

Water Balance of Cells with Walls
Hypertonic environment
- More solute outside of cell
- Cell loses water
- Becomes plasmolyzed

Water Balance of Cells with Walls
Isotonic
Same level of solute
Net flow of water is zero
No internal pressure – plant cells have no support – flaccid
Plants wilt

Water Balance of Cells with Walls
Hypotonic Environment
More solute inside than out
Positive flow of water into the cell
plant cell is turgid
healthy state in most plants
Cells don’t rupture because of strength of the cell wall
Combination of osmotic pressure and the pressure of the walls is called the cell’s water potential

Water balance in cells with walls
H2OH2OH2OH2O
Figure 7.13
Identify the Tonicity

Water balance in cells with walls
H2OH2OH2OH2O
Turgid (normal) Flaccid Plasmolyzed
Figure 7.13
Hypotonic solution Isotonic solution Hypertonic solution

Water Potential
determines the flow of water
Water flows from high water potential to low water potential
High pressure from the walls raises the water potential (tendency to lose water)
High levels of solute in the cell lower water potential (tendency to gain water)
Water flows into/out of the cell until the water potential reaches zero –pressure from the cell walls prevents any more water from entering the cell.

Water Potential Equation
Ψ = ΨP + Ψπ
ΨP = Pressure Potential
Ψπ = Osmotic Potential
Ψπ = − MiRT or Ψπ = − CiRT
M or C = osmotic molar concentration
R = pressure constant (Gas Laws)
T = temperature (K)
i = ionization constant (with plants the solute is mainly sucrose which has an ionization constant of 1 because sucrose does not ionize)

Facilitated Diffusion
Figure 7.15
EXTRACELLULAR
FLUID
Channel proteinSolute
CYTOPLASM
A channel protein (purple) has a channel through which
water molecules or a specific solute can pass.
(a)

Facilitated Diffusion: Passive Transport Aided by Proteins
Transmembranal proteins speed the movement of molecules across the plasma membrane
passive - no energy required - moves with the concentration gradient
Regulation of Facilitated Diffusion:
protein saturation
can only transport so much at once
proteins are specialized for a specific substance
protein inhibition
similar molecules to normal transport substance bind to protein and block path of transport
THESE ARE ONLY PHYSICAL CHANGES NOT CATALYZED REACTIONS

Mechanism of Transport
Conformational changes - binding and release of substance causes change in protein - moves substance across membrane
gated channels - stimulus causes protein to open and allow the flow of the substance - the stimulus is not the substance transported

Figure 7.15
Carrier proteinSolute
A carrier protein alternates between two conformations, moving a
solute across the membrane as the shape of the protein changes.
The protein can transport the solute in either direction, with the net
movement being down the concentration gradient of the solute.
(b)

Active Transport
pumping of solutes against the concentration gradient
moving uphill - requires ATP or a membrane potential
-membrane potential: electrical charge built up across the cell membrane due to unequal balance of anions and/or cations -

Types of Active Transport
Use of ATP:
phosphorylation reaction - adds phosphate to transport molecule and causes a conformational change
EX: sodium-potassium pump

Na/K Ion Pump
Generates an ELECTROCHEMICAL GRADIENT
Unbalanced concentration of ions
creates a new dynamic for diffusion -ions now move from high concentration and high positive charge
important for Nerve Impulses

Maintenance of Membrane Potential by Ion Pumps
Types of Pumps:
electrogenic pump - pumping of various ions - animal cells
proton pump - pumping H+ - plants, fungi, bacteria

Electrogenic pump
Figure 7.18
EXTRACELLULAR
FLUID
+
H+
H+
H+
H+
H+
H+Proton pump
ATP
CYTOPLASM
+
+
+
+–
–
–
–
–
+

COTRANSPORT
movement of one type of ion out allows for the movement of another substance inwhen the ion diffuses back in through a cotransport protein
Proton pump
Sucrose-H+
cotransporter
Diffusion
of H+
Sucrose
ATP H+
H+
H+
H+
H+
H+
H+
+
+
+
+
+
+–
–
–
–
–
–

Other Types of Active Transport
Use of Energized Electrons To Generate Chemical Gradients
Ex: Kreb’s Cycle and Light Dependent Reaction of Photosynthesis – Drives the production of ATP

Electron Transport Chain In the Kreb’s Cycle and Photosynthesis

Passive and active transport compared
Figure 7.17
Passive transport. Substances diffuse spontaneously
down their concentration gradients, crossing a
membrane with no expenditure of energy by the cell.
The rate of diffusion can be greatly increased by transport
proteins in the membrane.
Active transport. Some transport proteins
act as pumps, moving substances across a
membrane against their concentration
gradients. Energy for this work is usually
supplied by ATP.
Diffusion. Hydrophobic
molecules and (at a slow
rate) very small uncharged
polar molecules can diffuse
through the lipid bilayer.
Facilitated diffusion. Many
hydrophilic substances diffuse
through membranes with the
assistance of transport proteins,
either channel or carrier proteins.
ATP

BULK MOVEMENT
active transport that requires change in the cell membrane
movement of large molecules (proteins and polysaccharides)

Exocytosis
- movement of materials out of the cell transport vesicle fuses with cell membrane dumping contents into extracellular fluid
Examples: insulin
neurotransmitters
glycocalyx

Endocytosis
- movement of materials into the cell
results in the formation of a vacuole or vesicle
Three types
Phagocytosis
Pinocytosis
Receptor Mediated Endocytosis

1) Phagocytosis - cellular eating
cell membrane surrounds and engulfs large particles of food - becomes a food vacuole to be fused with lysosome

2) Pinocytosis - cellular drinking
cell membrane draws in extra cellular fluid and forms a small vesicle
– Non-discriminatory process

EXTRACELLULAR
FLUIDPseudopodium
CYTOPLASM
“Food” or
other particle
Food
vacuole
1 µm
Pseudopodium
of amoeba
Bacterium
Food vacuole
An amoeba engulfing a bacterium via
phagocytosis (TEM).
PINOCYTOSIS
Pinocytosis vesicles
forming (arrows) in
a cell lining a small
blood vessel (TEM).
0.5 µm
In pinocytosis, the cell
“gulps” droplets of
extracellular fluid into tiny
vesicles. It is not the fluid
itself that is needed by the
cell, but the molecules
dissolved in the droplet.
Because any and all
included solutes are taken
into the cell, pinocytosis
is nonspecific in the
substances it transports.
Plasma
membrane
Vesicle
In phagocytosis, a cell
engulfs a particle by
Wrapping pseudopodia
around it and packaging
it within a membrane-
enclosed sac large
enough to be classified
as a vacuole. The
particle is digested after
the vacuole fuses with a
lysosome containing
hydrolytic enzymes.
Three types of endocytosis
Figure 7.20
PHAGOCYTOSIS

3) Receptor Mediated Endocytosis -specific contents are taken into cell -regulated by surface proteins that recognize specific substances called LIGANDS
receptor proteins usually gathered in one area of the cell membrane (coated pits) -concentrates the gathering effort so cell can absorb concentrated amounts of a specific substance

0.25 µm
RECEPTOR-MEDIATED ENDOCYTOSIS
Receptor
Ligand
Coat protein
Coated
pit
Coated
vesicle
A coated pit
and a coated
vesicle formed
during
receptor-
mediated
endocytosis
(TEMs).
Plasma
membrane
Coat
protein
Receptor-mediated endocytosis enables the
cell to acquire bulk quantities of specific
substances, even though those substances
may not be very concentrated in the
extracellular fluid. Embedded in the
membrane are proteins with
specific receptor sites exposed to
the extracellular fluid. The receptor
proteins are usually already clustered
in regions of the membrane called coated
pits, which are lined on their cytoplasmic
side by a fuzzy layer of coat proteins.
Extracellular substances (ligands) bind
to these receptors. When binding occurs,
the coated pit forms a vesicle containing the
ligand molecules. Notice that there are
relatively more bound molecules (purple)
inside the vesicle, other molecules
(green) are also present. After this ingested
material is liberated from the vesicle, the
receptors are recycled to the plasma
membrane by the same vesicle.