
气候变化 2014 - ipcc.ch · 气候变化2014: 影响、适应和脆弱性 政府间气候变化专门委员会 第五次评估报告 第二工作组报告 编辑 Christopher

第七章 微生物的遗传变异和育种
复旦大学
2015-11-24
1

遗传:亲代与子代相似
变异:亲代与子代、子代间不同个体不完全相同
遗传(inheritance)和变异(variation)是生命的最本质特性之一
遗传型:
表型(表现型):
生物的全部遗传因子及基因
遗传型 + 环境条件→→→→生长发育→→→→形态等生物学特征的总和
表型是由遗传型所决定,但也和环境有关。2

表型饰变:
表型的差异只与环境有关
特点:暂时性、不可遗传性、表现为全部个体的行为
橘生淮南则为橘,生于淮北则为枳。
遗传型变异(基因变异、基因突变):
遗传物质改变,导致表型改变
特点:遗传性、群体中极少数个体的行为
(自发突变频率通常为10-6-10-9)
3

球形节杆菌
少生物素 的培养基--少细胞壁
诺卡氏菌属
完全培养基
放射土壤杆菌
4
蛋白胨琼脂 脑心浸出液琼脂

微生物是遗传学研究中的明星(模式生物):
微生物细胞结构简单,营养体一般为单倍体,方便建立纯系。
很多常见微生物都易于人工培养,快速、大量生长繁殖。
对环境因素的作用敏感,易于获得各类突变株,操作性强。
5

第一节 遗传的物质基础
第二节 基因突变和诱变育种
第三节 基因重组和杂交育种
第五节 菌种保藏
三个经典实验的原理与方法
遗传物质在微生物细胞内存在的部位和形式
原核生物的质粒
基因突变的规律
三种基因水平转移方式及其应用
微生物菌种保藏的基本方法
突变与育种
第四节 基因工程
6

第一节 遗传的物质基础
一、三个经典实验的原理与方法
二、遗传物质在微生物细胞内存在的部位和形式
原核及真核微生物基因组的基本特征
原核生物的质粒
7

种质连续理论:1883-1889年间Weismann提出。认为遗传物
质是一种具有特定分子结构的化合物。
基因学说:1933年摩尔根(Thomas Hunt Morgan)发现了染
色体,并证明基因在染色体上呈直线排列,提出了基因学说
,使得遗传物质基础的范围缩小到染色体上。
但染色体是由核酸和蛋白质两种长链高分子组成。什么
是遗传物质?


(一)经典转化实验
最早进行转化(transformation)实验的是F. Griffith(1928年),以Streptococcus pneumoniae(肺炎链球菌,旧称“
肺炎双球菌”)作为研究对象。
光滑型(S) 粗糙型(R)
有荚膜 无荚膜
菌落光滑 菌落粗糙
分泌毒素 无 毒致 病 不致病
SⅠ、SⅡ、SⅢ三个血清型RⅠ、RⅡ、RⅢ三个血清
型 10

S型 R型
加热灭菌 热死S菌+活R菌
(1)动物实验
11

(2)细菌培养试验
(3)S型菌的无细胞抽提液试验
活R菌+S菌的无细胞提取液
培养皿培养长出大量R菌和少量S菌
12
实验说明:加热杀死的S型细菌,其细胞内可能存在一种具有遗传转化能力的物质,通过某种方式进入R型细胞,并使R型细菌获得表达 S型荚膜性状的遗传特性。

1944年,O. T. Avery等从热
死的S型S.pneumoniae中提纯
了可能作为转化因子的各种成
分,并在离体条件下进行了更
为精密的转化实验。为Griffith
的转化因子是DNA而不是蛋白
质提供了第一证据。
13
O.T.Avery
(1877-1955)

(1)从活的S菌中抽提各种细胞成分(DNA,蛋白质,
荚膜多糖等)
(2)对各组分进行转化试验
14

只有DNA被酶降解破坏的抽提物无转化活性
DNA是转化所必需的转化因子
DNA纯度越高,转化效率也越高; DNA浓度越高,转化效率也越高。
15
1950年,有人提出应给Avery授予诺贝尔奖,但当时许多科学
家包括诺贝尔奖评委仍然对Avery的转化因子持有异议, 提议
被搁置.

(1)因认为蛋白分子量大,结构复杂,二十种氨基酸
的排列组合将是个天文数字,可作为一种遗传信息。而
DNA分子量小,只含4种不同的碱基,人们一度认为不同
种的有机体的核酸只有微小的差异。
(2)认为转化实验中DNA并未能提得很纯,还附有其它
物质。
(3)认为即使转化因子确实是DNA,但也可能DNA只是
对荚膜形成起着直接的化学效应,而不是充当遗传信息
的载体。

(二)噬菌体感染实验—经典实验之二
1952年,A.D.Hershey和M.Chase发表了证明DNA是噬菌体的遗传物质基础的著名实验
17
(三)植物病毒的重建实验--RNA作为遗传物质
噬菌体转化实验成功后Avery的研究才得到广泛认同。当
诺贝尔奖评选委员会准备为Avery授奖时,这位杰出的科
学家已于1955年去世。由于诺贝尔奖评委们的失误,使
人们对DNA的误解延长了差不多10年。

18
1969年诺贝尔生理学或医学奖
颁奖词:发现了病毒的复制和遗传结构,证明遗传物质是DNA而不是蛋白质

二、遗传物质在微生物细胞内存在的
部位和方式
(一)7个水平 具体内容看书p189-194
19
细胞水平: (单细胞与多细胞、性细胞与体细胞和单核与多核细胞)
细胞核水平:(真核与原核、核内与核外和单核与多核等)
染色体水平:(一份染色体与多份染色体及染色体基因组的大小等)
核酸水平: (是DNA还是RNA、是复合还是露裸及是长还是短等)
基因水平: (依基因功能可分为结构基因及调控基因等)
密码子水平:(由结构密码子及起始与终止密码等)
碱基水平: (突变的最低交换单位有A, T, G, C 等)

1、概念
基因组(genome):
一个物种的单倍体的所有染色体及其所包含的遗传信息的总称
原核生物(如细菌),多为单倍体(在一般情况下只有一条染色体)
真核微生物,多条染色体,例如啤酒酵母有16条染色体。有时为双倍体
原核及真核微生物基因组的基本特征
20

2、微生物基因组结构的特点
1)原核生物(细菌、古生菌)的基因组
染色体为双链环状的DNA分子(单倍体);
基因组上遗传信息具有连续性;
功能相关的结构基因组成操纵子结构;
结构基因的单拷贝及rRNA基因的多拷贝;
基因组的重复序列少而短;
古生菌的基因组在结构上类似于细菌。但是信息传递系统(复制、
转录和翻译)则与细菌不同而类似于真核生物。
21

2、微生物基因组结构的特点
2)真核微生物(啤酒酵母)的基因组
典型的真核染色体结构;
没有明显的操纵子结构;啤酒酵母基因组大小为13.5×106bp,分布在16条染色体中。
有间隔区(即非编码区)和内含子序列;
重复序列多;
22


(二) 原核生物的质粒
质粒(plasmid):
一种独立于染色体外,能进行自主复制的细胞质遗传因子,主要存在于各种微生物细胞中。
转座因子(transposable element):
位于染色体或质粒上的一段能改变自身位置的DNA序列,广泛分布于原核和真核细胞中。
质粒和转座因子是细胞中除染色体以外的另外二类遗传因子
质粒的基本特征及类型√
24

一、质粒的分子结构
1、结构
通常以共价闭合环状的超螺旋双链DNA分子存在于细胞中;
也发现有线型双链DNA质粒和RNA质粒;
质粒分子的大小范围从1kb左右到1000kb;(细菌质粒多在10kb以内)
Covalently closed circular DNA (cccDNA)
25

一、质粒的分子结构
2、质粒的检测
提取所有胞内DNA后电镜观察;
琼脂糖凝胶电泳或超速离心后观察;
对于由于三种构型同时存在时造成的多带现象(提取质粒时造成或自然存在),可以进行特异性单酶切,使其成为一条带。
特定的质粒提取方法和后处理使染色体和RNA均被除掉。
26

一、质粒的分子结构
2、质粒的检测
提取所有胞内DNA后电镜观察;
琼脂糖凝胶电泳或超速离心后观察;
对于实验室常用菌,可用质粒所带的某些特点, 如抗药性初步判断。
27

二、质粒的主要类型
质粒与宿主的关系:
质粒所含的基因对宿主细胞一般是非必需的;
在某些特殊条件下,质粒有时能赋予宿主细胞以特殊的机能,
从而使宿主得到生长优势。
28

二、质粒的主要类型
质粒所编码的功能和赋予宿主的表型效应
致育因子(Fertility factor,F因子)
抗性因子(Resistance factor,R因子)
产细菌素的质粒(Bacteriocin production plasmid)
毒性质粒(virulence plasmid)
代谢质粒(Metabolic plasmid)
隐秘质粒(cryptic plasmid)
29

二、质粒的主要类型
1、致育因子(Fertility factor,F因子)
又称F质粒,其大小约100kb,这是最早发现的一种与大肠杆菌的有性生殖现象(接合作用)
有关的质粒。
携带F质粒的菌株称为F+菌株(相当于雄性),无F质粒的菌株称为F-菌株(相当于雌性)。
F因子能以游离状态(F+)和以与染色体相结合的状态
(Hfr)存在于细胞中,所以
又称之为附加体(episome)。
有关内容在讲细菌的接合作用(conjugation)时具体介绍
!自身特性的体现
30

二、质粒的主要类型
2、抗性因子(Resistance factor,R因子)
包括抗药性和抗重金属二大类,简称R质粒。
R100质粒(89kb)可使宿主对
下列药物及重金属具有抗性:
汞(mercuric ion ,mer)四环素(tetracycline,tet )链霉素(Streptomycin, Str)、磺胺(Sulfonamide, Su)、氯霉素(Chlorampenicol, Cm)夫西地酸(fusidic acid,fus)
并且负责这些抗性的基因是成簇地存在于抗
性质粒上。
抗性质粒在细菌间的传递是细菌产生抗药性的重要原因之一。
!威力的体现
31
大环质粒:脆弱类杆菌,抗克林霉素。小质粒,大肠杆菌,四环素抗性

二、质粒的主要类型
3、产细菌素的质粒(Bacteriocin production plasmid)
细菌素:能抑制或杀死近缘、甚至
同种不同株的细菌的因子(细菌蛋白),
!同类竞争的武器
32

细 菌 素 抗 生 素
抑 制 或 杀 死 近 缘 , 甚 至 同 种 不 同
株 的 细 菌
较 广 的 抗 菌 谱
通 过 核 糖 体 直 接 合 成 的 多 肽 类 物
质
一 般 是 次 级 代 谢 产 物
编 码 细 菌 素 的 结 构 基 因 及 相 关 的
基 因 一 般 位 于 质 粒 或 转 座 子 上
一 般 无 直 接 的 结 构 基 因 , 相 关 酶
的 基 因 多 在 染 色 体 上
二、质粒的主要类型
3、产细菌素的质粒(Bacteriocin production plasmid)
细菌素(bacteriocin) 1.细菌产生的一种抗生代谢产物,对同源种或近似种才有拮抗作用; 2.蛋白质是主要成分; 3.有一定的作用机制(杀菌模式); 4.由质粒控制。
33

二、质粒的主要类型
3、产细菌素的质粒(Bacteriocin production plasmid)
细菌素一般根据产生菌的种类进行命名:
大肠杆菌(E. coli)产生的细菌素为colicins(大肠杆菌素),
而质粒被称为Col质粒。
由G+细菌产生的细菌素或与细菌素类似的因子与colicins有所不同,但通常也是由质粒基因编码,有
些甚至有商业价值,例如一种乳酸细菌产生的细菌素NisinA能强烈抑制某些G+细菌的生长,而被用于
食品工业的保藏。
34

产嗜盐菌素嗜盐古菌菌株筛选
某些极端嗜盐菌(古生菌)也被发现能产生具有广谱杀菌效果的嗜盐菌素(耐热、稳定)
35

二、质粒的主要类型
4、毒性质粒(virulence plasmid)
许多致病菌的致病性是由其所携带的质粒引起的,这些质粒具有
编码毒素的基因,其产物对宿主(动物、植物)造成伤害。
编码肠毒素的质粒(产毒素大肠杆菌:引起人类和动物腹泻)
编码δ内毒素(伴孢晶体中)的质粒(苏云金杆菌)
Ti质粒(根癌土壤杆菌):双子叶植物冠瘿瘤
36

二、质粒的主要类型
5、代谢质粒(Metabolic plasmid)
质粒上携带有有利于微生物生存的基因,如能降解某些基质的酶,进行共生固氮,或产生抗生素(某些放线菌)等
将复杂的有机化合物降解成能被其作为碳源和能源利用的简单形式,
环境保护方面具有重要的意义。
假单胞菌:具有降解一些有毒化合物,如芳香簇化合物(苯)、农药(2,4 dichlorophenoxyacetic acid)、辛烷
和樟脑等的能力。
亦称降解质粒:
!人类的公仆
37

二、质粒的主要类型
6、隐秘质粒(cryptic plasmid)
隐秘质粒不显示任何表型效应,它们的存在只有通过物理的方法,例如用凝胶电泳检测
细胞抽提液等方法才能发现。
他们存在的生物学意义,目前几乎不了解。
在应用上,很多隐秘质粒被加以改造作为基因工程的载体(一般加上抗性基因)
38

二、质粒的主要类型
高拷贝数(high copy number)质粒(每个宿主细胞中可以有10-100个拷贝)
———————松弛型质粒(relaxed plasmid)
低拷贝数(low copy number)质粒(每个宿主细胞中可以有1-4个拷贝)
———————严谨型质粒(stringent plasmid)
窄宿主范围质粒(narrow host range plasmid)(只能在一种特定的宿主细胞中复制)
广宿主范围质粒(broad host range plasmid)(可以在许多种细菌中复制)
39

三、质粒的不亲和性
质粒之间的不亲和性、以及质粒拷贝数的多少、能适应的
宿主范围的宽窄等特性均与质粒的复制控制类型有关,欲
了解详细内容请选参考“微生物遗传学”。
40

第二节 基因突变和诱变育种
一个基因内部遗传结构或DNA序列的任何改变基因突变:
微生物的基因突变及其应用
41

突变
自发突变
诱变
环境因素的影响,DNA复制过程的偶然错误等而导致,
一般频率较低,通常为10-6-10-9 。
某些物理、化学因素对生物体的DNA进行直接作用,
突变以较高的频率产生。
基因突变: 一个基因内部遗传结构或DNA序列的任何改变
诱变育种
灭菌处理
危害健康(辐射、化学等的致癌作用)
第二节 基因突变和诱变育种
42

一个基因内部遗传结构或DNA序列的任何改变基因突变:
基因突变是重要的生物学现象,它是一切生物变化的根源,连同基因转移、重组一起提供了
推动生物进化的遗传多变性。
基因突变DNA损伤修复机制
DNA复制→→真正的突变;
损伤修复→→原来的结构;
第二节 基因突变和诱变育种
43

1、特点
1)自发性
2)非对应性-突变性状与引起该突变的原因
3)稀有性
4)独立性
5)遗传和回复性
6)可诱变性
一、基因突变
﹖
第二节 基因突变和诱变育种
44

抗性突变引起的争论和斗争
观点之一: “驯化”或“驯养”论 突变是生物对某特定环境的适应而产生的,这种
环境正是突变的诱因,所产生的抗性性状是与该环境
因素相对应的,并认为这就是“定向变异”。
观点之二: 自发突变论 基因突变是自发的,即使诱发产生,其产生的性状与诱变
因素间也是不对应的,即最终适应了的化学药物等不良因
素并非诱变因素,而仅仅是一种用于筛选的环境而已。
45

证明突变的性状与引起突变的原因间无直接对应关系!
如何证明基因突变的非对应性?
三个经典实验
变量实验、涂布实验、影印实验
一、基因突变的特点
2、实验证据
46

变量实验(fluctuation analysis),又称波动试验或彷徨试验;Salvador Luria and Max Delbruck(1943)根据统计学的原理设计
47
抗噬菌体抗性产生并非由所抗的环境因素—T1诱导出来的,而是在接触T1之前,在某次细胞分裂过程中自发产生的;发生越早,抗性菌落越多;T1只是起淘汰和筛选作用

变量实验(fluctuation analysis)Salvador Luria and Max Delbruck(1943)
Salvador Luria Max Delbruck
The Nobel Prize in Physiology or Medicine 1969
48

Newcombe的涂布实验(1949)
√NATURE,实验方法更简单
49353/28

影印实验(replica plating )Joshua Lederberg and Esther Lederberg(1952)
J. Lederberg is awarded the Noble Prize in Medicine and Physiology in 1958
证明微生物的抗药性在未接触药物前自发产生,突变与相应的药物环境毫不相干
50

突变真的与选择压力无关吗?
适应性突变(adaptive mutability)和高频突变(hypermutation )Benson S.Adaptive mutation: a general phenomenon or special case?Bioessays. 1997 Jan;19(1):9-11. Review.
Galitski T, Roth JR. A search for a general phenomenon of adaptive mutability.Genetics. 1996 Jun;143(2):645-59.
Dan I. Andersson, E. Susan Slechta, and John R. Roth Evidence That Gene Amplification Underlies Adaptive Mutability of the Bacterial lac Operon。 Science 1998 November 6; 282: 1133-1135.
Godoy VG, Fox MS. Transposon stability and a role for conjugational transfer in adaptive mutability.Proc Natl Acad Sci U S A. 2000 Jun 20;97(13):7393-8.
Bull HJ, McKenzie GJ, Hastings PJ, Rosenberg SM. Evidence that stationary-phase hypermutation in the Escherichia coli chromosome is promoted by recombination. Genetics. 2000 Apr;154(4):1427-37.
51

突变
诱因
基因突变
染色体畸变:断裂、重排等
自发突变
诱发突变
复制错误
化学错误
碱基错配
DNA复制跳格
脱嘌呤
脱氨基
氧化损伤
放射线-----产生嘌呤二聚体
化学诱变剂碱基类似物:5BU,2AP
碱基修饰物:NA,HA,烷化剂
DNA插入剂:吖啶类
3、基因突变及其机制多样性
第二节 基因突变和诱变育种

4、紫外线对DNA的损伤及其修复
嘧啶(敏感性)>嘌呤
紫外线(ultraviolet ray,UV)
嘧啶二聚体(TT,TC,CC)和水合物
53
第二节 基因突变和诱变育种

经UV照射后的微生物立即暴露于可见光下时,可明显降低其死亡率的现象。
1)光复活作用(photoreactivation photorestoration)
E.coli的实验
① 对照:8×106个/mL E.coli → 100个/mL
② 试验:8×106个/mL E.coli →----→2×106个/mL
UV
UV 360~490 nm
可见光,30 min
54
注 意:在一般的微生物中都存在光复活作用,
在利用UV进行诱变育种等工作时,应在红光下进行
照射和后续操作,并放置在黑暗条件下培养。

使二聚体(dimer)重新分解成单体(monomer)
带有嘧啶二聚体的DNA分子
DNA分子
UV照射
黑暗下结合
光激活酶——光解酶
(光裂合酶,photolyase)
复合物
300~500 nm可见光
获得光能而激活释放光解酶
55

修复
DNA Pol 3'→5'外切酶活性
光复活—光裂合酶直接修复
复制后修复
AP内切酶
糖基酶
GO系统-MutM,MutY,MutT
错配修复-Dam, MutL,MutH,UvrD
重组修复-RecA
SOS修复-RecA,LexA,UvrAB,UmuC,HimA
双链断裂的修复
切除修复
一般切除修复——UvrABC系统
特殊切除修复
同源重组修复
非同源重组修复
2015年诺贝尔化学奖授予3位研究DNA损伤与修复的科学家

5、常见的微生物突变类型
表型基因型
这里主要介绍几种常用的由于基因突变而造成微生物表型
变化的突变型及其分离
第二节 基因突变和诱变育种
57
毒力、生产某种代谢产物的发酵能力的变化等在实际应用
中具有重要意义突变类型一般都不具有很明显或可直接检
测到的表型。其突变株的获得往往需要较大的工作量。

1)营养缺陷型(auxotroph)
第二节 基因突变和诱变育种
58
与营养缺陷型相关的三类遗传型个体: 野生型:自然界中分离的原始菌株, 营养缺陷型:诱变剂处理后产生 原养型:营养缺陷型菌株回复突变或重组后产生
缺乏合成其生存所必须的营养物(包括氨基酸、维生素、碱基等)的突变型,
只有从周围环境或培养基中获得这些营养或其前体物(precursor)才能生长。
5、常见的微生物突变类型
回复突变(reverse mutation或back mutation): 突变体失去的野生型性状,可以通过第二次突变得到恢复,这种第二次突
变称为回复突变。

基本培养基( minimal medium,MM,符号为[-])
与筛选营养缺陷型突变株有关的3类培养基
完全培养基( CM,符号为[+])
补充培养基( SM,符号为[A]或[B]等)
仅能满足某微生物的野生型菌株生长所需的最低成分的组合培养基。
凡可满足一切营养缺陷型菌株营养需要的天然或半组合培养基
一般可在基本培养基中加入一些富含氨基酸、维生素、核苷酸和碱基之类的
天然物质,如蛋白胨或酵母膏等配制而成。
凡只能满足相应的营养缺陷型突变株生长需要的组合或半组合培养基。
59

1)营养缺陷型 微生物遗传学研究中重要的选择标记和育种的重要手段
表型判断的标准:在基本培养基上能否生长
第二节 基因突变和诱变育种
60
特点: 在选择培养基(一般为基本培养基)上不生长
负选择标记
(突变株不能通过选择平板直接获得)
5、常见的微生物突变类型

负筛选模拟实验
61

营养缺陷型的表示方法:
1)营养缺陷型(auxotroph)
基因型:所需营养物的前三个英文小写斜体字母表示:hisC(组氨酸缺陷型,大写字母C代表同一表型中不同基因的突变)
表型: 同上,但第一个字母大写,且不用斜体:HisC
在具体使用时多用hisC-和hisC+,分别表示缺陷型和野生型。
第二节 基因突变和诱变育种
62
5、常见的微生物突变类型

2)抗药性突变型(resistant mutant)
基因突变使菌株对某种或某几种药物,特别是抗生素,产生抗性。
特点:正选择标记
(突变株可直接从抗性平板上获得-----在加有相应抗生素的
平板上,只有抗性突变能生长。所以很容易分离得到。)
表示方法: 所抗药物的前三个小写斜体英文字母加上“r”表示
strr 和 strs 分别表示对链霉素的抗性和敏感性
第二节 基因突变和诱变育种
63
5、常见的微生物突变类型

3)条件致死突变型(conditional lethal mutant)
在某一条件下具有致死效应,而在另一条件下没有致死效应的突变型。
温度敏感突变:(temperature sensitive, ts ) : 在高温下(原来可以生活的温度如42℃)致死的;
在低温下(如25-30℃)生活正常;
特点: 负选择标记
这类突变型常被用来分离生长繁殖必需的突变基因
64
5、常见的微生物突变类型

4)形态突变型(morphological mutant)
造成形态改变的突变型
特点: 非选择性(非致死性)突变突变株和野生型菌株均可生长,但可从形态特征上进行区分。
举例: 产蛋白酶缺陷突变株的筛选
菌落颜色变化
β半乳糖苷酶基因的插入失活,使重组子菌落为白色而与蓝色的非重组子分开。
第二节 基因突变和诱变育种
65
形成芽胞缺陷菌株
细胞水平上的形态突变,突变株的检出更加困难。
5、常见的微生物突变类型

(一)自发突变与育种 1. 从生产中育种:生产第一线易获得较优良的生产菌株
2. 定向培育优良菌株是一种利用微生物自发突变,并采用
特定的选择条件,通过对微生物群体不断移植以选育出较
优良菌株的古老方法。卡介苗的筛选 230代
从污染噬菌体的发酵液中分离到抗噬菌体的自发突变株
66
二、突变与育种

筛选
诱变诱变育种 定向的——更重要
随机的
(二)诱变育种
获得各种菌株;提高产品质量、有用代谢产物的产量等;
诱变育种具有重大的实践意义:
诱变育种的特点:高效(突变率)、广谱(多性状)
67

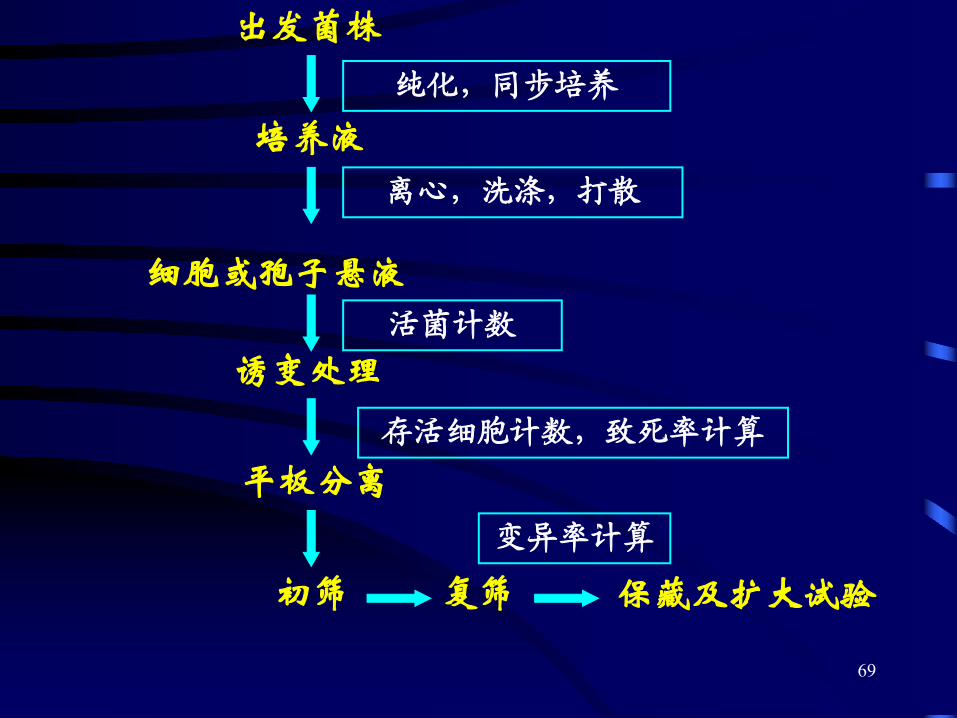
1. 诱变育种的基本环节
68
出发菌株
纯化,同步培养
培养液
离心,洗涤,打散
细胞或孢子悬液
诱变处理
平板分离
初筛 复筛 保藏及扩大试验
活菌计数
存活细胞计数,致死率计算
变异率计算
69

2. 诱变育种几个原则
(1)选择简便有效的诱变剂
(2)挑选优良的出发菌株
(3)处理单细胞或单孢子悬液
(4)选用最适的诱变剂量
(5)充分利用复合处理的协同效应
(6)利用和创造形态、生理与产量间的相关指标
(7)设计高效筛选方案
(8)创造新型筛选方法
70

(3)处理单细胞或单孢子悬液
因为: 分散的细胞可均匀地接触诱变剂
诱变剂一般只作用于DNA双链中的某一条单链,故
某一突变无法在当代的表型上反映。只有经过DNA的复
制和细胞分裂后,这一变异才会在表型上表达出来,
这就叫表型延迟(phenotype lag)。
71

表型延迟(phenotype lag)
用于诱变育种的细胞应尽量选用单核单细胞72

提高育种时初筛的效率,必须进行大量的分析测定和统计
工作,找到形态、生理变异与产量变异两者间的相关性。
(6)利用和创造形态、生理与产量间的相关指标
产量性状转化为与之相平行的肉眼可见的性状。
快速平板检出法
73

蛋白酶水解圈
淀粉酶变色圈(用碘液使淀粉显色)
氨基酸显色圈(菌落转移到滤纸上,再用茚三酮试剂显色)
抗生素抑制圈
指示菌生长圈(测定生长因子产量)
纤维素酶对纤维素水解圈(用刚果红染色)
外毒素的沉淀反应圈
均可为初筛工作中估计某突变株代谢产物量的指标。
74

3. 3类突变株的筛选方法
(1)产量突变株的筛选
(2)抗药性突变株的筛选
(3)营养缺陷型突变株的筛选
75

(1)产量突变株的筛选
1971年,国外有人报道了筛选春日霉素
(kasugamycin)高产菌时所采用琼脂块培
养法,一年内曾使该抗生素产量提高10倍。
76

此法的关键是用打
孔器取出含有一个小菌
落的琼脂块作分别培养
。这样,各琼脂块所含
养料和接触空气面积基
本相同,且产生的抗生
素等代谢产物不致扩散
出琼脂块外。
数据精确,效率高77

(2)抗药性突变株的筛选
正选择标记:(突变株可直接从抗性平板上获得——
在加有相应抗生素的平板上,只有抗性突变能生长。)
78
梯度平板法(gradient plate):定向筛选抗药性突变株
制备药物浓度梯度的平板;
涂布诱变处理后的细胞悬液;
培养---选择抗药性菌落;

4个环节
第一步,诱变剂处理
第二步,淘汰野生型
第三步,检出缺陷型
第四步,鉴定缺陷型
79
(3)营养缺陷型突变株(auxotrophic mutant)的筛选-
--用于理论、应用、生产

第二步,淘汰野生型, 浓缩营养缺陷性
菌丝过滤法
抗生素法青霉素法(细菌)
制霉菌素法(真菌)
MM :野生型生长,缺陷型不长
孢子 菌丝体(过滤、离心)
野生型生长
缺陷型不生长
•MM+青霉素/制霉菌素
•死亡
•幸存
80

夹层培养法----1个培养皿
限量补充培养法----1个培养皿
逐个检出法----2个培养皿
影印平板法----2个培养皿
第三步,检出缺陷型
81

夹层培养法(layer plating method)
82
限量补充培养法:MM + 微量CM
野生型 迅速生长,大菌落缺陷型 营养有限,小菌落

逐个检出法
83
影印平板法

生长谱法(auxanography)第四步,鉴定缺陷型
84
在混有供试菌的基本培养基表面,点加微量营养物

三、诱变剂与致癌物质——Ames试验
a)利用各种诱变剂获得各类遗传突变,进行诱变育种;
c)危害人类自身的健康;
b)对有害微生物进行控制;
很多种化学物质,能以各种机制导致DNA的突变
第二节 基因突变和诱变育种
85

“生物化学统一性”法则:
人和细菌在DNA的结构及特性方面是一致的,能使微生物发生突变的诱变剂
必然也会作用于人的DNA,使其发生突变,最后造成癌变或其他不良的后果
所有生物的DNA在结构及特性上具有一致性
86
三、诱变剂与致癌物质——Ames试验
诱变剂共性原则:化学药剂对细菌的诱变率与其对动物的致癌性成正比
超过95%的致癌物质对微生物有诱变作用90%以上的非致癌物质对微生物没有诱变作用

美国加利福尼亚大学的Bruce Ames教授于
1966年发明,因此称为Ames试验。
具体操作:检测鼠伤寒沙门氏菌(Salmonella typhmurium) 组氨
酸营养缺陷型菌株(his-)的回复突变率
87
利用细菌营养缺陷型的回复突
变来检测环境和食品中是否存
在化学致癌剂的简便有效方法
三、诱变剂与致癌物质——Ames试验
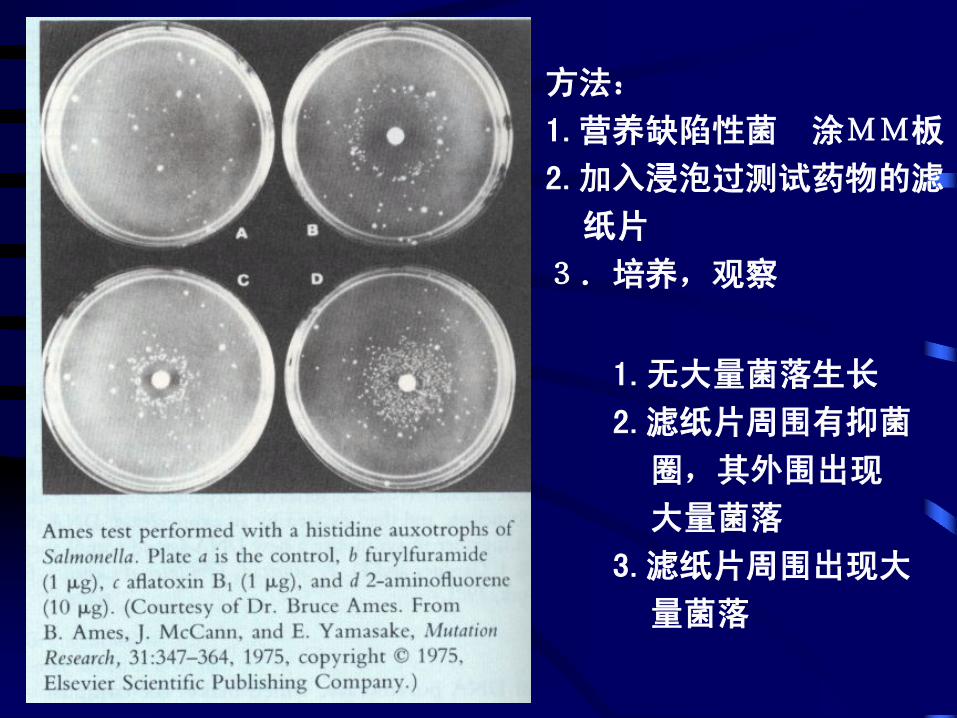
1.无大量菌落生长
2.滤纸片周围有抑菌
圈,其外围出现
大量菌落
3.滤纸片周围出现大
量菌落
方法:
1.营养缺陷性菌 涂MM板
2.加入浸泡过测试药物的滤
纸片
3.培养,观察

证明Ames试验重要性的应用实例:
国外曾开发了一种降低妇女妊娠反应的药物“反应停”,由
于其药效显著,在60-70年代十分流行,
但随后人们就发现(1万余名无臂)畸形儿的出生率明显增高,而且
生产畸形儿的妇女大多曾服用“反应停”,后来采用Ames试验发现这种
物质的确具有很强的致突变作用,因此这种药物被禁止使用
但如果能在这种药物上市之前就进行Ames试验检测,那么这种大量
出生畸形儿的悲剧完全可以避免。
89

第三节 基因重组和杂交育种
本节重点:三种基因水平转移方式及其应用
90

基因的水平转移:
个体A 个体B遗传物质
通过基因的重组可使后者(个体B)及其后代的性状发生改变。
细菌所特有的基因重组形式,有别于一般有性过程中
的亲本和其后代之间遗传信息的垂直传递形式。 91

水平基因转移(Horizontal gene transfer, HGT) ,不
仅发生在不同的微生物细胞之间,而且也发生在微生物与高
等动植物之间。
生物间基因的转移和交换是普遍存在的,是生物进化的
重要动力之一。
92

细菌水平基因转移和重组:
基因工程生物的生态安全
病原菌的抗药性
基因工程技术
93

细菌的三种水平基因转移形式
接合(Conjugation)
转导(Transduction)
转化(Transformation)
第三节 基因重组和杂交育种
94

一、细菌的接合作用(conjugation)
通过细胞与细胞的直接接触而产生的遗传信息的转移和重组过程:接合作用
( conjugation )
1.实验证据
1946年,Joshua Lederberg 和Edward L.Taturm
大肠杆菌的多重营养缺陷型杂交实验
95

中间平板上长出的原养型菌落是两菌株之间发生了遗传交换和重组所致!
为何采用多重营养缺陷型菌株?
尽可能地排除回复突变对实验结果的干扰!
96

97

证实接合过程需要细胞间的直接接触的“U”型管实验:
98
接合的机制?

质粒的主要类型
质粒所编码的功能和赋予宿主的表型效应
致育因子(Fertility factor,F因子)
抗性因子(Resistance factor,R因子)
产细菌素的质粒(Bacteriocin production plasmid)
毒性质粒(virulence plasmid)
代谢质粒(Metabolic plasmid)
隐秘质粒(cryptic plasmid)
99
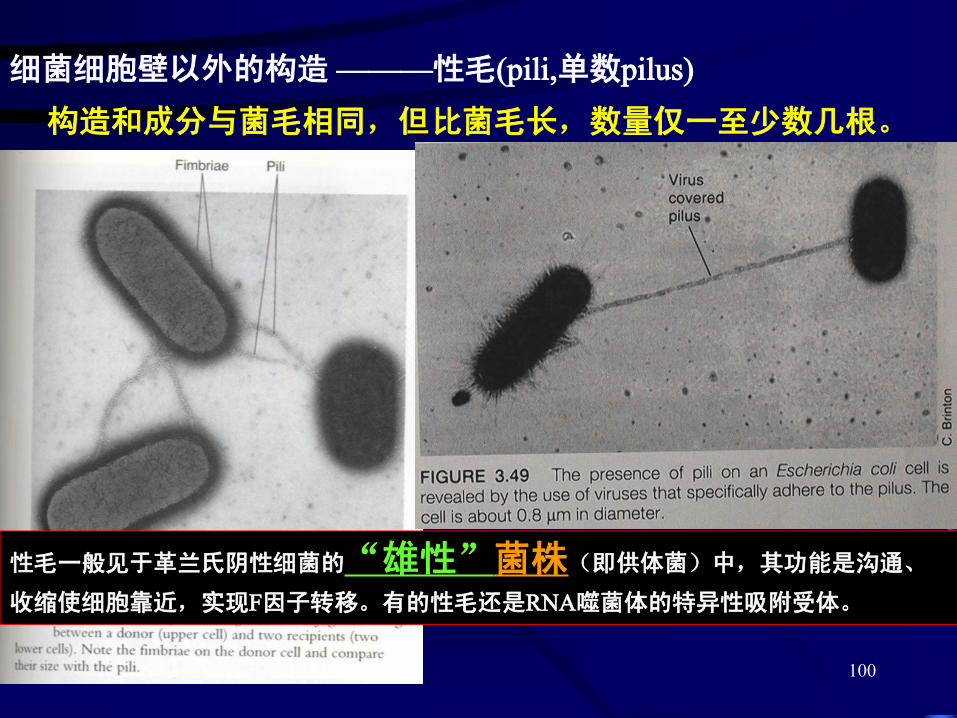
细菌细胞壁以外的构造 ———性毛(pili,单数pilus)
构造和成分与菌毛相同,但比菌毛长,数量仅一至少数几根。
性毛一般见于革兰氏阴性细菌的“雄性”菌株(即供体菌)中,其功能是沟通、
收缩使细胞靠近,实现F因子转移。有的性毛还是RNA噬菌体的特异性吸附受体。
100

2. 大肠杆菌的接合机制
接合作用是由一种被称为F因子的质粒介导;
F因子的分子量通常为5×107,上面有编码细菌产生性菌毛(sex pili)及控制接合过程进行的20多个基因
101
Tn,IS是转座因子,也是整合发生位点
是多位点整合,方向不定

F因子为附加体质粒,既可以脱离染色体在细胞内独立存在,
也可插入(整合)
到染色体上
102

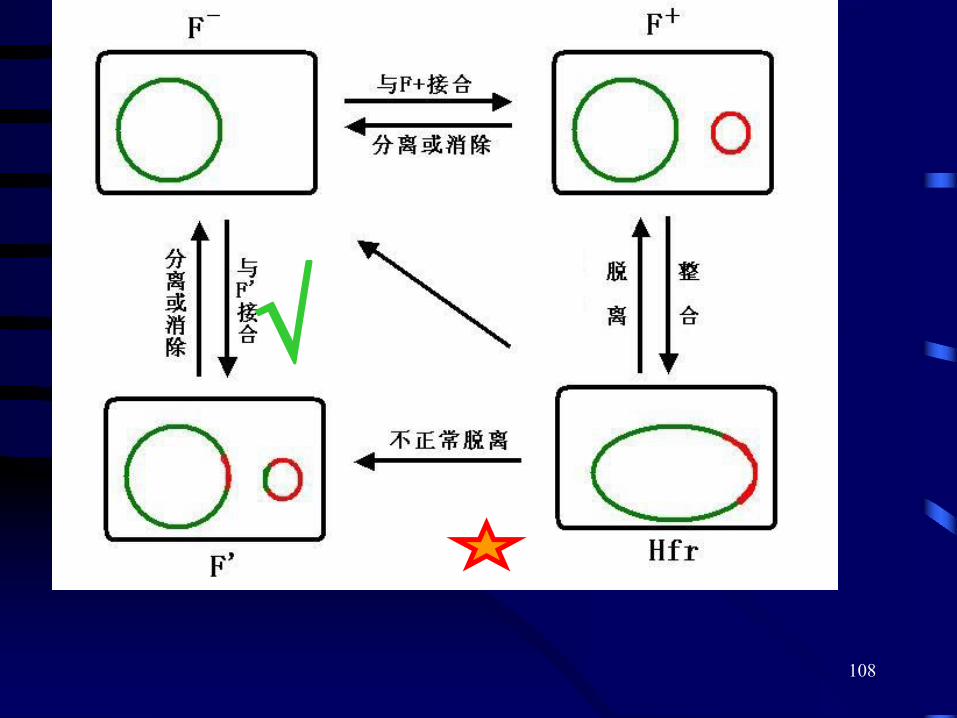
F因子的四种细胞形式
a)F-;
b)F+ ;( F因子独立存在,有性菌毛);
c)Hfr;F因子插入到染色体DNA上,有性菌毛。
d)F′;F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子。 细胞表面同样有性菌毛。
(“雌性”菌株,无性菌毛)
“
雄性”
菌株
103

1) F+×F-杂交F+菌株的F因子向F-细胞转移,但含F因子的宿主细胞的染色体DNA一般不被转移。
104
理化因子可消除F因子
杂交的结果:给体细胞和受体细胞均成为F+细胞

Hfr菌株的F因子插入到染色体DNA上,因此只要发生接合转移过程,就可以把
部分甚至全部细菌染色体传递给F-细胞并发生重组,由此而得名为
高频重组菌株(Hfr)
2)Hfr ×F-杂交
105

染色体上越靠近F因子的先导区的基因,进入的机会就越多,在F-中出现重组子的时间就越早,频率也高。
106
Hfr菌株与F+:都有性菌毛,与F-细胞进行
接合。
区别是:Hfr×F-杂交后的受体细胞(或接合子)大多数仍然是F—。

3)F′×F-杂交
Hfr菌株内的F因子因不正常切割而脱离染色体时,形成游离的但携带一小段染色体基因的F因子,特称为F′因子。
F′×F-与F+×F-的不同:给体的部分染色体基因随F′一起转入受体细胞
a)与染色体发生同源重组;
b)继续存在于F′因子上, 形成一种部分二倍体;
107F′因子传递基因,也叫性导,或F因子转导
√
108

Lederberg coined the term plasmid in the 1950s to describe such apparently extrachromosomal genetic elements, although it did notfind wide usage until the 1970s when infectious drug resistance become a medical problem, and genetic engineering become popular.
109

细菌的三种水平基因转移形式
接合
转导
转化
√
第三节 基因重组和杂交育种
110

二、细菌的转导(transduction)由噬菌体介导的细菌细胞间进行遗传交换的一种方式
一个细胞的DNA通过病毒载体的感染转移到另一个细胞中
能将一个细菌宿主的部分染色体或质粒DNA带到另一个细菌的噬菌体:
转导噬菌体
细菌转导的二种类型:普遍性转导
局限性转导
111

1、普遍性转导(generalized transduction)
噬菌体可以转导给体染色体的任何部分到受体细胞中的转导过程
(1) 意外的发现
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外的其它菌种是
否也存在接合作用,用二株具不同的多重营养缺陷型的鼠伤寒沙门氏菌进行类
似的实验:
用phe-trp-tyr-×met-his-结果获得了10-5的原养型重组子(phe+trp+tyr+met+
his+)。但将两个亲本放在U形管的两臂中,结果也能同样获得重组子,此和
E.coli杂交是不同的
112

沙门氏菌LT22A是携带P22噬菌体的溶源性细菌 另一株是非溶源性细菌
一个表面看起来的常规研究却导致一个惊奇和十分重要发现的重要例证!
基因的传递是由可透过“U”型管滤板的P22噬菌体介导的
(发现了普遍性转导------这一重要的基因转移途径)
Why and How ?
113

(2)
转导模型
114

转导噬菌体为什么“错”将宿主的DNA
包裹进去?
噬菌体的DNA包装酶也能识别染色体DNA上类
似pac的位点并进行切割,以“headful”的包装机
制包装进P22噬菌体外壳,形成只含宿主DNA的
转导噬菌体颗粒(假噬菌体)。
因为染色体上的pac与P22 DNA的pac序列不完全相同,利用效率较低,这种“错装”机率一般仅约10-6-10-8 。
在50年代,沙门氏菌的遗传学研究系统尚未完善,所以转导的发现未能得到发展。 1955年Lennox,E.发现了P1噬菌体可在E.coli中转导遗传物质,从而极大地促进了对普遍性转导的研究和应用。
115

形成转导颗粒的噬菌体可以是温和的也可以是烈性的,但必须具有能偶尔
识别宿主DNA的包装机制并在宿主基因组完全降解以前进行包装。
普遍性转导的基本要求:
116

普遍性转导的三种后果:
1)进入受体的外源DNA通过与细胞染色体的重组交换而形成稳定的转导子
2)流产转导(abortive transduction)
转导DNA不能进行重组和复制,但其携带的基因可经过转录而得到表达。
特点:在选择培养基平板上形成微小菌落
1173)外源DNA被降解,转导失败。

2、局限性转导(specialized transduction)温和噬菌体感染
整合到细菌染色体的特定位点上宿主细胞发生溶源化
溶源菌因诱导而发生裂解时,在前噬菌体二侧的少数宿主基因因偶尔发生的不正常切割而连在噬菌体DNA上
部分缺陷的温和噬菌体
把供体菌的少数特定基因转移到受体菌中
118

溶源性感染对细胞的影响:
其它同类噬菌体可以再次感染该细胞,但不增殖,也不导致溶源性细菌裂解。免疫性是由原噬菌体产生的阻遏蛋白的可扩散性质所决定的。
原噬菌体引起的溶源性细菌除免疫性外的其他的表形改变,包括溶源菌细胞表面
性质的改变和致病性转变被称为溶源转变(lysogenic conversion)。
溶源菌中的温和噬菌体基因组:不影响细胞的繁殖功能,
但可能引起其他的细胞变化。
(1)免疫性
(2)溶源转变
白喉杆菌只有在含有特定类型的原噬菌体(带有毒素
基因)时才能产生白喉毒素,引起被感染机体发病。 119

2、局限性转导(specialized transduction) 局限性转导又称特异性转导(restricted transduction),也是由Joshua Lederberg及其学生所发现。
λ噬菌体是否像P22进行普遍性转导?
紫外线诱导野生型E.coli K12 (λ) gal+ 中的原噬菌体,使细胞裂解,收集裂解液去感染非溶源状态的gal— 菌株,结果在基本培养基上选择到10-6 gal+ 的转导子。
①部分转导子具有溶源性细菌所具备的“免疫力”,说明有功
能的噬菌体存在于受体细胞中。
② 上述gal+转导子细胞不能被诱导产生成熟的噬菌体颗粒。 ③gal+ 转导子遗传特点不稳定,有1~10%又变成gal-,表明它
又失去了gal+,因此推论有些gal+是随λ插入到受体的染色
体中,形成部分二倍体,而不是通过双交换而产生的。
新的现象:
120

举例:
λ噬菌体的的溶源性反应:
进入宿主后线状基因组依靠粘性末端环化
121

λ噬菌体的的溶源性反应:
通过特定位点整合(切离)细菌染色体
122
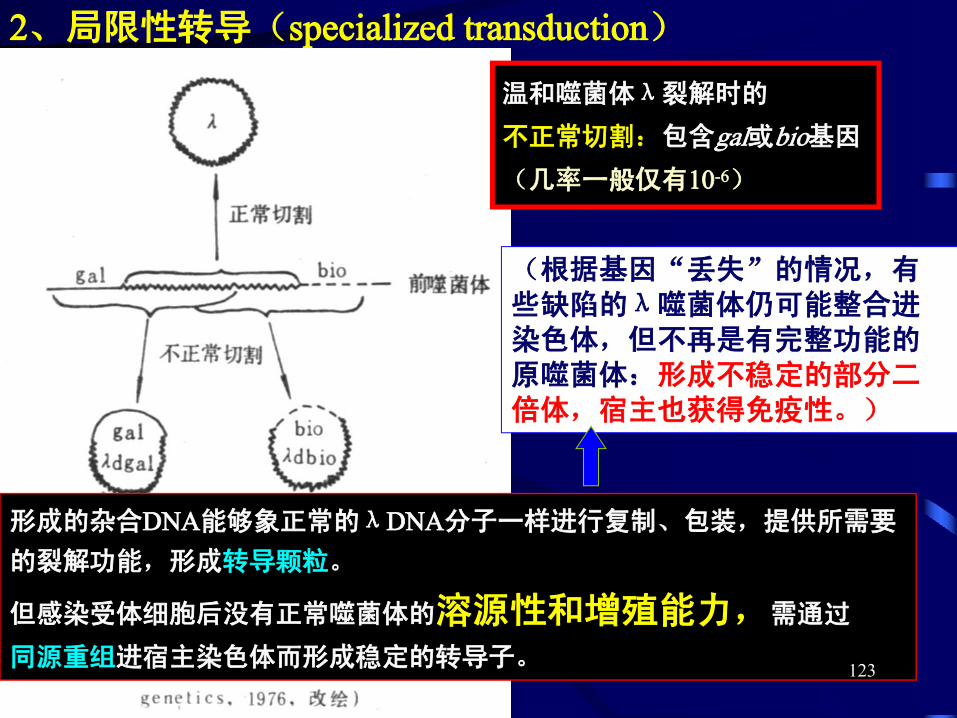
2、局限性转导(specialized transduction)
温和噬菌体λ裂解时的
不正常切割:包含gal或bio基因
(几率一般仅有10-6)
形成的杂合DNA能够象正常的λDNA分子一样进行复制、包装,提供所需要
的裂解功能,形成转导颗粒。
但感染受体细胞后没有正常噬菌体的溶源性和增殖能力,需通过
同源重组进宿主染色体而形成稳定的转导子。
(根据基因“丢失”的情况,有些缺陷的λ噬菌体仍可能整合进染色体,但不再是有完整功能的原噬菌体:形成不稳定的部分二倍体,宿主也获得免疫性。)
123

局限性转导与普遍性转导的主要区别:
a)被转导的基因共价地与噬菌体DNA连接,与噬菌体DNA一起 进行复制、包装以及被导入受体细胞中。
b)局限性转导颗粒携带特定的染色体片段并将固定的个别基因
导入受体,故称为局限性转导。
2、局限性转导(specialized transduction)
普遍性转导:包装的全部是宿主菌的基因
普遍性转导:携带的宿主基因具有随机性
(部分缺陷的噬菌体)
(完全缺陷噬菌体)
124烈性或温和噬菌体;温和噬菌体

溶源转变(lysogenic conversion):
一个与转导相似又不同的现象
温和噬菌体感染细胞后使之发生溶源化,因噬菌体的基因整合到宿主染色体上
,而使后者获得了新性状的现象。
溶源转变与转导的不同?
a)不携带任何供体菌的基因;
b)这种噬菌体是完整的,而不是缺陷的;
125

三、细菌的遗传转化(genetic transformation)游离DNA分子被感受态细胞摄取,并得到表达的水平方向的基因转移过程。
自然感受态:是细胞一定生长阶段的生理状态。 受细菌自身的基因控制(自然遗传转化)
人工感受态:是通过人为诱导获得。 与细菌自身的遗传控制无关(人工转化)
感受态细胞(competent cell):具有摄取外源
DNA能力的细胞;处于最易接受外源DNA片段并能实现转化
的一种生理状态。
126
转化的条件:1)建立了自然感受态的受体细胞
2)外源游离DNA分子

1、自然遗传转化(简称自然转化)
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae)的转化现象
127
Avery在四十年代以更精密的实验设计重复了以上实验
用S型菌细胞抽提物 DNA是转化所必需的转化因子
目前已知有二十多个种的细菌具有自然转化的能力
DNA是遗传物质的确证;
基因工程技术的建立;
重要的微生物遗传学方法;

感受态受遗传控制,但也存在个体差异;
受外界环境因子如腺苷酸(cAMP)及Ca2+等影响;
感受态出现的时间不同(对数期、对数后期,恒定期等);
感受态细胞所占比例和维持时间不同;
1)建立了自然感受态的受体细胞
128
调节感受态的一类特异蛋白称
感受态因子。
膜相关DNA结合蛋白(membrane-associated DNA binding protein)
细胞壁自溶素(autolysin)
几个核酸酶

转化因子(transforming factor):离体的DNA片段。一般只有 ~15 kb。
微生物不同,转化因子的形式不同:dsDNA、ssDNA和质粒DNA
(通常不能与核染色体组发生重组)。
2)外源游离DNA分子
129
转化频率:0.1%~1.0%,最高为20%。
需要的DNA浓度极低:1×10-5 g/mL。

(3) 转化过程
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae)
的转化现象,这种G+细菌的转化过程研究得比较深入
130
枯草芽孢杆菌(G+菌)的自然转化过程:dsDNA在细胞外降解,ssDNA进入
某些革兰氏阴性菌的转化模型:dsDNA进入,降解为单链才整合

自然转化过程的特点:
a)对核酸酶敏感;
c)成功与否及效率的高低主要取决于给体和受体菌 株之间的亲源关系;
d)通常情况下质粒的自然转化效率要低得多;
b)不需要活的DNA给体细胞;
131

2、人工转化
用CaCl2处理细胞,电穿孔等是常用的人工转化手段。
自然转化→→→人工转化→→→基因工程
基因工程的奠基石和基础技术
用多种不同的技术处理受体细胞,使其人为地处
于一种可以摄取外源DNA的“人工感受态”。
质粒的转化效率高;
132

噬菌体DNA被感受态细胞摄取并产生有活性的病毒颗粒
转染(transfection):
现在把DNA转移至动物细胞的过程也称转染
提纯的噬菌体DNA以转化的(而非感染)途径进入
细胞并表达后产生完整的病毒颗粒。
特点:
133

自然遗传转化的进行涉及到细菌染色体上几十个基因的功能及彼此间的相
互协调,因此被认为是名副其实 的细菌水平基因转移途径。
目前的研究热点:
1)细菌为什么要耗费如此多的染色体遗传资源来进行自然转化,即该过程 的生物学意义到底何在?它对细菌自身有什么好处? (人们已提出了一些假说,但均不圆满)
2)自然转化过程的很多细节仍不清楚,包括细菌如何协调感受态 的建 立,外源DNA进入和重组的具体过程等等
3)细菌中具有自然转化能力的范围,到底有哪些菌具有自然转化能力?
4)转化DNA的来源和在自然环境中发生的自然转化
134

接合 (conjugation):
由F因子介导(细胞与细胞的直接接触)
转导(transduction):
由噬菌体介导
自然遗传转化(natural genetic transformation):
游离DNA分子 + 感受态细胞
135

“接合” “转导” 及“自然转化”这三种在自然界中存在的细菌遗传重组过程各自的特点:
a)外源DNA的来源及进入途径有差异;
b)决定因素也各有不同;
如何设计实验对三种途径进行区分?
如果二个不同营养缺陷标记(a - b - c + d + 和 a + b + c - d - )的 菌株
经混合后能产生在基本培养基平板上生长的原养型重组菌 株,请设
计一个实验来决定该遗传转移过程是转化、转导还是 接合?
136

四、基因定位和基因组测序
基因定位:遗传重组等手段 → → → 基因在染色体上
的位置及相对距离→ → →遗传图谱。
细菌基因转移的三种方式(接合、转导、转化)都可以用来
进行细菌基因组作图。
基因组测序:测定所有碱基排列顺序→ → → 对遗传信息进行研究和分析
137

四、基因定位和基因组测序1、中断杂交(interrupted mating)技术
利用Hfr×F-的接合过程,在不同时间取样,并把样品猛烈搅拌以分散接合中的细菌,然后分析受体细菌基因型,以时间(分钟)为单位绘制遗传图谱,该图谱是细菌染色体上基因顺序的直接反映。
138

四、基因定位和基因组测序
1、中断杂交技术
大肠杆菌基因组很大(全部转移需要100分钟),其遗传图谱须用多株Hfr菌(F因子整合在不同位置)才能完成。
139

四、基因定位和基因组测序
2、基因连锁
连锁的二基因间的距离与其共转化,或共转导的频率成反比,因此,可根据
二个基因被共转化或共转导的频率判断它们在染色体上的相对距离。
接合作用(中断杂交):
对相隔很近的基因进行定位: 转导、转化
判断相距较远的基因间的相对位置;
140

四、基因定位和基因组测序
3、遗传图谱
对大肠杆菌的染色体已定位了1000多个基因
141

四、基因定位和基因组测序
4、基因组测序
“全基因组鸟枪测序”(whole genome shotgun sequencing)
5、基因组序列的注释
借助计算机对基因组序列进行分析并遗传图谱进行对照
142

真核微生物的基因重组
有性杂交:有减数分裂过程,性细胞,高频率基因重组准性杂交:无减数分裂过程,体细胞,低频率基因重组原生质体融合遗传转化
143
特点:细胞融合或联合,整套染色体参与
原核微生物的基因重组
转化转导原生质体融合接合(性导)
特点:细胞间不接触或暂时沟通,少数基因转移或局部杂合

基因工程(gene engineering)又称遗传工程
(genetic engineering ),是指人们利用分子生物
学的理论和技术,自觉设计、操纵、改造和重建细胞
的遗传核心——基因组,从而使生物体的遗传性状发
生定向变异,以最大限度地满足人类活动的需要。
第四节 基因工程
144

基因工程是指重组DNA技术的产业化设计与应用,包括:
上游技术:基因重组、克隆和表达的设计与构建(即重组DNA)
下游技术:涉及到基因工程菌或细胞的大规模培养以及基因产物
的分离纯化过程。
145
重组DNA技术是指将一种生物体(供体)的基因与载体在
体外进行拼接重组,然后转入另一种生物体(受体)内,使之
稳定遗传并表达出新产物或新性状的DNA体外操作程序,也称
为分子克隆技术。
供体、受体、载体是重组DNA技术的三大基本元件。

基因工程的基本形式
第一代基因工程 蛋白多肽基因的高效表达 经典基因工程
第二代基因工程 蛋白编码基因的定向诱变 蛋白质工程
第三代基因工程 代谢信息途径的修饰重构 途径工程
第四代基因工程 基因组或染色体的转移 基因组工程
146

基因工程的操作过程
三、 转化子的筛选和鉴定(检)
二、 重组DNA分子的转化和扩增(转、增)
一、 DNA的体外重组(切、接)
147

基因工程的主要操作步骤示意图 148

(一)目的基因的取得(3途径)
① 从适当的供体生物中提取;
② 由mRNA合成cDNA;
③ 由化学合成方法合成有特定功能的目的基因。
一、 DNA的体外重组(切、接)
149

作为优良载体的基本要求:
(1)独立的复制子-能够进行独立自主的复制,能在受体细胞内大量增殖;
(2)具有若干限制酶的单一切割位点,便于外源DNA的插入
(3)具有可供选择的遗传标记-便于对阳性克隆的筛选和鉴定
(5)载体DNA须易于分离和实验操作
(4)具有一定的安全性-胞内不重组和转移;胞外不扩散
√√
√
(二)优良载体的选择
150

基因工程的常用载体都来自微生物:
质粒载体
λ噬菌体载体
Cosmid载体
M13噬菌体载体
真核细胞受体
人工染色体
噬菌粒载体
SV40病毒
Ti质粒
原核受体细胞
质粒载体克隆外源DNA片段的大小一般不超过15Kb
容量较大(大约23kb);感染效率高;克隆
容量更大( 31 - 45 kb )
151

1、质粒载体
152

2、λ噬菌体克隆载体
将野生型λ噬菌体DNA进行改造后建成,主要是去掉噬菌体DNA上
过多的常用限制酶的酶切位点,及对非必要基因区域进行改造。
特点:1)分子遗传学背景清楚;
2)容量较大(能容纳大约23kb的外源DNA片段);
3)感染效率高。其感染宿主细胞的效率几乎可达100%, 而质粒DNA的转化率却只有0.1%。
4)具有更为狭窄的寄主范围,更加安全。
153
154

与溶原性相关的基因可以被外源DNA取代而不影响λ噬菌体的
感染、复制和裂解宿主细胞的能力。
插入型载体(insert vector)
取代型载体(replacement vector)155

重组噬菌体DNA也可不经过体外包
装 而直接通过转染 方式进入宿主
细胞;
(转化)
经过体外基因操作和包装后形成
重组噬菌体,可以通过正常的感染
途径进 入宿主细胞;
156
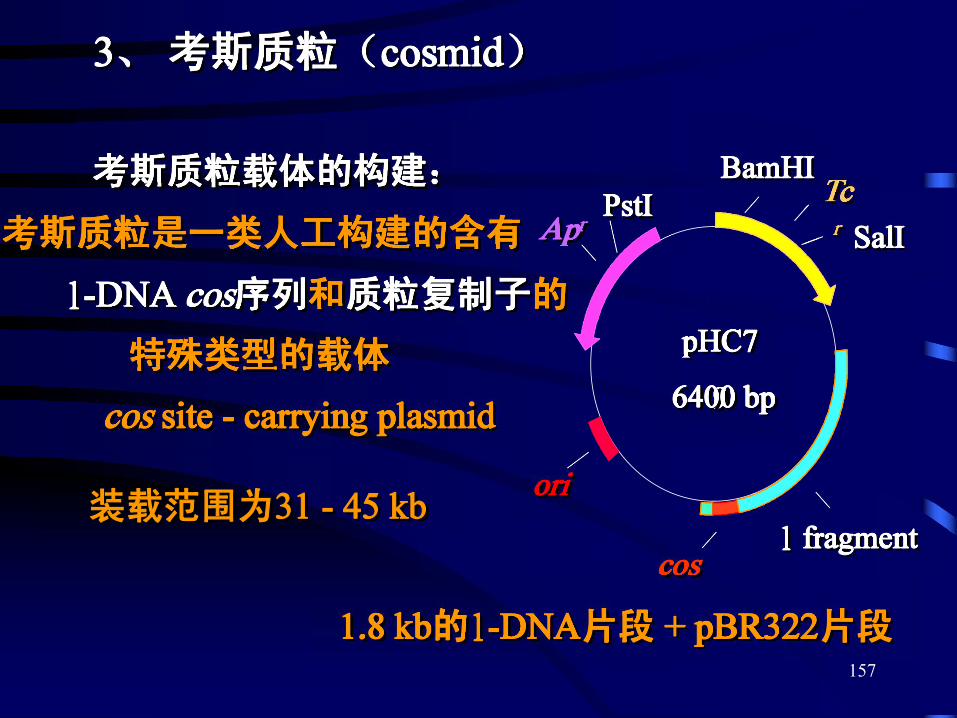
3、 考斯质粒(cosmid)
考斯质粒载体的构建:
考斯质粒是一类人工构建的含有
pHC7
96400 bp
Tcr
l fragmentcos
ori
AprPstI
BamHI
SalI
l-DNA cos序列和质粒复制子的
特殊类型的载体
cos site - carrying plasmid
1.8 kb的l-DNA片段 + pBR322片段
装载范围为31 - 45 kb
157

(三)目的基因 与载体DNA的体外重组
限制性核酸内切酶
粘性末端
退火
连接酶
共价结合
嵌合体
158

限制性核酸内切酶
DNA的3′-末端加上 poly(A) 或 poly(T)
人为处理
目的基因和载体DNA
一个DNA分子产生“榫头”
一个DNA分子产生“卯眼”
互补粘性末端
混合“退火”
共价结合
环状重组载体——嵌合体(chimaera)
连接酶
159

(四)重组载体导入受体细胞
受体细胞种类很多,主要有:
原核生物为主:E. coli、
Bacillus subtilis(枯草杆菌);
真核微生物:Saccharomyces cerevisiae(酿酒酵母) ;
各种高等动、植物的细胞株、组织。
导入受体细胞的途径:
质粒:转化法;
噬菌体或病毒:感染法。 160

(五)重组受体细胞的筛选和鉴定
针对载体携带的标志基因和目的基因设计筛选
方法。
例 抗药性标记选择(插入失活法):将目的
基因插入带ampr和tetr基因载体的tetr基因中,则
tetr基因失活。在分别含有氨苄青霉素和四环素的
两个培养基中培养,进行筛选。
例2 PCR方法
161

发酵罐
生物反应器
(六)“工程菌”或“工程细胞”
的大规模培养
162

微生物和微生物学在基因工程中的重要性:
① 载体;
② 工具酶;
③ 受体;
④ 微生物工程;
⑤ 目的基因的主要供体。
163

三、基因工程的应用
基因工程在工业上的应用
基因工程在农业上的应用
遗传工程在医疗上的应用
基因工程在环境保护中的应用
基因工程与基本理论研究
164

第五节 菌种的衰退、复壮和保藏
性状稳定的菌种是微生物学工作最重要的基本要求,否则
生产或科研都无法正常进行。
影响微生物菌种稳定性的因素:
a)变异;
b)污染;
c)死亡;
微生物菌种保藏的基本方法
165

1. 菌种保藏的原理
(1)挑选典型菌种的优良纯种,
最好用芽孢或分生孢子等休眠体;
(2)创造一个有利于长期休眠的良好环
境条件, 例如,干燥、低温、缺氧、
避光、缺乏营养等;
(3)添加保护剂或酸度中和剂,
避免保藏过程中细胞受伤。166

干燥和低温是菌种保藏中的最重要因素
(1)较低的温度更有利于保藏
液氮(-195℃)>干冰(-70℃)> -20℃ > 4℃
(2)水分在低温下形成冰晶,破坏细胞结构
加入保护剂:0.5 mol/L左右的甘油或二甲亚
砜; 海藻糖、脱脂牛奶、血清白蛋白、糊精或聚乙
烯吡咯烷酮(PVP)等。
167

作用机理:大分子物质如海藻糖、脱脂牛奶、血清白
蛋白、糊精或聚乙烯吡咯烷酮(polyvinylpyrrolidone,
PVP)虽不能透入细胞,但可能是通过与细胞表面结合
的方式而防止细胞膜受冻伤。
作用机理:0.5 mol/L左右的甘油或二甲亚砜
(dimethylsulfoxide, DMSO)可透入细胞,并通过
降低强烈的脱水作用而保护细胞;
(3)介质减少冷冻对细胞损伤的机制
168

在一定时间内使菌种不死、不变、不乱基本要求:
基本方法:生活态
休眠态
培养基传代培养
寄主传代培养
冷冻
干燥
斜面、平板
液氮、低温冰箱
沙土管、冷冻真空干燥
169

菌种保藏
由于微生物的多样性,不同的微生物往往对不同的保藏方法有不同
的适应性,迄今为止尚没有一种方法能被证明对所有的微
生物均适宜。因此,在具体选择保藏方法时必须对被保藏菌株的
特性、保藏物的使用特点及现有条件等进行综合考虑。对于一些比
较重要的微生物菌株,则要尽可能多的采用各种不同的手段进行保
藏,以免因某种方法的失败而导致菌种的丧失。
170

方法名称 主要措施 适宜菌种 保藏期 评价
冰箱保藏法(斜面) 低温(4℃) 各大类 1~6月 简便
冰箱保藏法
(半固体)低温、避氧 细菌、酵母菌 6~12月 简便
石蜡油封藏法 低温、阻氧 各大类 1~2年 简便
甘油悬液保藏法低温(-70℃)、
保护剂细菌、酵母菌 1~10年 较简便
砂土保藏法 干燥、无营养 产孢子微生物 1~10年 简便有效
冷冻干燥保藏法干燥、低温、
无氧、保护剂各大类
>5~15
年繁而高效
液氮保藏法保护剂
超低温(-196℃)各大类 >15年 繁而高效
不同的保藏方法比较
171

第七章思考题
3、 如果二个不同营养缺陷标记(a - b - c + d + 和 a + b + c - d - )的
菌株经混合后能产生在基本培养基平板上生长的原养型重组菌
株,请设计一个实验来决定该遗传转移过程是转化、转导还是
接合?
4、 自然遗传转化与人工转化之间有什么关系?为什么在一般情况下
它们转化质粒的成功率有如此大的差别?5、试比较转化与转染、转导与溶源转换,以及转导与性导间的异同。
2、分析、比较接合、转导、转化这三种基因水平转移方式的主 要特点。
1、试分析在何种情况下涉及营养缺陷型、温度敏感突变型和 抗生素抗性等遗传标记的应用(筛选)需采用负选择策略。
172

173
转导(transduction)由噬菌体将一个细胞的基因传递给另一细胞的过程。它是细菌之间传递遗传物质的方式之一。
普遍性转导:噬菌体能传递供体细菌的任何基因的转导。
转导子: 即接受了噬菌体传递的供 体细胞基因的受体细胞。
由普遍性转导产生的转导子不具溶源性,说明转导噬菌体中不带有完整的噬菌体染色体,却带有噬菌体在繁殖过程中错误包装的供体细菌的基因。
根据噬菌体转导的供体细胞DNA是否整合到受体细胞染色体上,又可将普遍性转导分为完全转导和流产转导。

174
局限性转导:噬菌体只能传递供体染色体上原噬菌体整合位置附近的基因的转导。
(1)低频转导(LFT)诱导溶源性细菌而产生的细胞裂解产物中,除含有正常的噬菌体外,还有极少数(约为10^(-6))部分缺陷噬菌体。用诱导溶源性菌株得来的噬菌体进行转导时的转导频率不过10^(-6) ,称为低频转导。
(2)高频转导(HFT)用高感染复数(m.o.i.) 的LFT裂解物去感染受体菌,λd gal (带有供体菌gal基因的λ缺陷噬菌体)与λ可同时整合在一个受体菌的核染色体组上,这种同时感染有正常噬菌体和缺陷噬菌体的受体菌就称双重溶源菌。
双重溶原菌在紫外辐射等因子的诱导下,原噬菌体容易被切割下来,产生等量的缺陷噬菌体和正常噬菌体,该裂解物称为高频率转导裂解物。再用低m.o.i. 的它去感染另一受体菌,可高频率地(50%)把受体菌转导成转导子。

175
什么是转导?为什么用u.v诱导E.coli K12( λ/λdgal+)双重溶源化细胞所获得的细胞裂解液对E.coli(gal-)可进行高频转导?


















