
Carbohydrates - Marquette University · Carbohydrates Objectives: I. Define carbohydrates on the...
20
Carbohydrates Objectives: I. Define carbohydrates on the basis of the organic functional groups present on the molecule. II. Define carbohydrates on the basis of size A. Monosaccharides B. Disaccharides C. Oligosaccharides D. Polysaccharides III. Classify the important monosaccharides on the basis of: A. The number of carbons in the molecule. B. Whether it is an aldehyde or a ketone. C. Both IV. Describe the biochemically most important monosaccharides. A. Aldoses (6) versus ketoses (4) B. Significance of stereoisomerism on carbohydrate structure and function. C. Recognize and name the common monosaccharides from their Fischer projection D. From a Fischer projection identify a carbohydrate as a D or L sugar. E. Recognize and name the enantiomer pairs and/or diastereomers of the common monosaccharides from their Fischer projections. V. Describe the differences and similarities in structure and function among glucose, galactose, mannose, fructose, and ribose. VI. What are epimers? A. Which of the monosaccharides are epimers of each other? VII. Cyclic forms of the monosaccharides A. What organic functional group form when an aldose cyclizes? B. What organic functional group form when a ketose cyclizes? C. What are anomers? D. What is the anomeric carbon? E. Which of the monosaccharides are anomers of each other? F. What is Mutarotation? G. Recognize the monosaccharide when given in the Haworth projection of its α- and/or β-cyclic forms. H. Be able to convert the Fischer projection of a monosaccharide into its corresponding Haworth projection or a Haworth projection into its corresponding Fischer projection. VIII. Recognize some of the chemical properties of carbohydrates and relate them to the chemistry of alcohols and carbonyl compounds. A. Oxidation of aldehyde function group to form aldonic acids B. Reduction of carbonyl group to form sugar alcohols C. Oxidation of terminal hydroxyl group to form uronic acids D. Esterification E. Reduction of hydroxyl group to form deoxy sugars F. Substitution of hydroxyl group by an amino group G. Reaction of amino group with a carboxylic acid - amide formation H. Non-enzymatic glycation reactions ©Kevin R. Siebenlist, 2019 1
Transcript of Carbohydrates - Marquette University · Carbohydrates Objectives: I. Define carbohydrates on the...

Carbohydrates
Objectives: I. Define carbohydrates on the basis of the organic functional groups present on the molecule. II. Define carbohydrates on the basis of size
A. Monosaccharides B. Disaccharides C. Oligosaccharides D. Polysaccharides
III. Classify the important monosaccharides on the basis of: A. The number of carbons in the molecule. B. Whether it is an aldehyde or a ketone. C. Both
IV. Describe the biochemically most important monosaccharides. A. Aldoses (6) versus ketoses (4) B. Significance of stereoisomerism on carbohydrate structure and function. C. Recognize and name the common monosaccharides from their Fischer projection D. From a Fischer projection identify a carbohydrate as a D or L sugar. E. Recognize and name the enantiomer pairs and/or diastereomers of the common
monosaccharides from their Fischer projections. V. Describe the differences and similarities in structure and function among glucose, galactose,
mannose, fructose, and ribose. VI. What are epimers?
A. Which of the monosaccharides are epimers of each other? VII. Cyclic forms of the monosaccharides
A. What organic functional group form when an aldose cyclizes? B. What organic functional group form when a ketose cyclizes? C. What are anomers? D. What is the anomeric carbon? E. Which of the monosaccharides are anomers of each other? F. What is Mutarotation? G. Recognize the monosaccharide when given in the Haworth projection of its α- and/or β-cyclic
forms. H. Be able to convert the Fischer projection of a monosaccharide into its corresponding Haworth
projection or a Haworth projection into its corresponding Fischer projection. VIII.Recognize some of the chemical properties of carbohydrates and relate them to the chemistry of
alcohols and carbonyl compounds. A. Oxidation of aldehyde function group to form aldonic acids B. Reduction of carbonyl group to form sugar alcohols C. Oxidation of terminal hydroxyl group to form uronic acids D. Esterification E. Reduction of hydroxyl group to form deoxy sugars F. Substitution of hydroxyl group by an amino group G. Reaction of amino group with a carboxylic acid - amide formation H. Non-enzymatic glycation reactions
©Kevin R. Siebenlist, 20191

IX. Recognize whether a monosaccharide is a reducing sugar or a non reducing sugar A. Discuss the use of the Tollen’s reagent and/or Benedict’s reagent to determine whether an
aldose is present. X. Describe the reaction by which monosaccharides are joined to form disaccharides and
polysaccharides. A. Describe the glycosidic bond.
1. What type of organic group is formed when a glycosidic bond forms? B. Describe the involvement of the anomeric carbon / anomeric hydroxyl group in the formation
of a glycosidic bond. XI. Discuss the structural, chemical, and biochemical properties of the three common disaccharides.
A. Describe the nature of the glycosidic bond(s) in the common disaccharides. B. Identify the disaccharide as reducing or non reducing.
XII. Homopolysaccharides A. Discuss the structural, chemical, and biochemical properties of the homopolysaccharides
produced by bacteria. B. Discuss the structural, chemical, and biochemical properties of the three common
homopolysaccharides in eukaryotes. 1. Describe the nature of the glycosidic bond(s) in the common eukaryotic polysaccharides.
XIII.Heteropolysaccharides A. With repeating disaccharide units - The Glycosaminoglycans
1. Discuss biochemical properties of the common glycosaminoglycans. a) Chondroitin sulfate.
(1) D-Glucuronate (D-Glucuronic acid) and N-Acetyl-D-galactosamine. (2) C-4 or C-6 of N-acetyl galactosamine is often esterified with sulfate. (3) found in tendons, cartilage and bone where it modulates the compressibility of
these tissues. b) Dermatan sulfate.
(1) L-Iduronate (L-Iduronic acid) and N-Acetyl-D-galactosamine. (2) L-Iduronate is the C-5 epimer of D-Glucuronate (D-Glucuronic acid). (3) N-acetyl-D-galactosamine is often sulfated. (4) found in most tissues, but is especially rich in skin.
c) Heparin. (1) D-Glucuronate (D-Glucuronic acid) and D-Glucosamine. (2) both are sulfated. (3) found in all cells, particularly rich Mast Cells where it has anticoagulant activities.
d) Hyaluronic acid. (1) D-Glucuronate (D-Glucuronic acid) and N-Acetyl-D-glucosamine. (2) molecule Is Not Sulfated. (3) found in synovial fluid, vitreous humor, loose connective tissue, and cartilage. (4) plays a role in directing cell migration through the extracellular matrix and it
modulates the compressibility of cartilage. e) Keratan sulfate.
(1) N-Acetyl-D-glucosamine and D-Galactose. (2) N-acetyl-D-glucosamine is often sulfated. (3) found in the cornea of the eye where it plays a role in corneal transparency and
©Kevin R. Siebenlist, 20192

also found in cartilage and tendons. 2. Assembly of glycosaminoglycans into proteoglycans
B. With a “random” structure. 1. O-Linked vs N-Linked 2. Lectins & Selectins 3. A, B, O Blood group heteropolysaccharides (A, B, O Blood group antigens)
kkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkkk
General Considerations
The CARBOHYDRATES are widely distributed in nature, and by mass they are the most abundant biomolecules on earth. Carbohydrates are also called SACCHARIDES or SUGARS.
In terms of organic functional groups present on the molecule, carbohydrates are polyhydroxyl aldehydes or ketones, or molecules that yield polyhydroxyl aldehydes or polyhydroxyl ketones upon hydrolysis.
MONOSACCHARIDES are the monomeric units of carbohydrates. They all share the general formula, Cn(H2O)n, where n ≥ 3. From this formula the origin of the name “carbohydrate” is obvious - Carbon + Water.
DISACCHARIDES contain two monomeric units covalently linked.
OLIGOSACCHARIDES are polymers of monosaccharides containing ≤ 20 monomeric units.
POLYSACCHARIDES are polymers of monosaccharides containing > 20 monomeric units.
GLYCOCONJUGATES are carbohydrate derivatives in which oligosaccharides or polysaccharides are covalently linked to peptides, proteins, or lipids. Glycoconjugates include the:
• PEPTIDOGLYCANS and PROTEOGLYCANS- more sugar than peptide/protein by mass. • GLYCOPROTEINS - more protein than sugar by mass. • GLYCOLIPIDS - more lipid than sugar by mass.
The Monosaccharides
The monosaccharides are all polyhydroxyl aldehydes or ketones. They can be classified by the number of carbon atoms they contain -TRIOSE, TETROSE, PENTOSE, HEXOSE, etc. NONOSES, monosaccharides with 9 carbons, are the largest monosaccharides found in nature. Monosaccharides can be classified by the type / location of carbonyl group they contain; an aldehyde group - ALDOSE; or a ketone group - KETOSE. A combination of these terms is can be used to describe monosaccharides more precisely - e.g., a KETOTETROSE has 4 carbons and a ketone functional group. Aldoses and ketoses with the same number of carbon atoms are isomers of each other. They have the same empirical formula Cn(H2O)n, but different structural formulas. Most monosaccharides end in the suffix “OSE”.
The two simplest monosaccharides are GLYCERALDEHYDE, an aldotriose, and DIHYDROXYACETONE, a
©Kevin R. Siebenlist, 20193

ketotriose.
Glyceraldehyde is a chiral molecule. The second carbon atom of this molecule is a chiral (asymmetric) carbon. Since glyceraldehyde contains a chiral carbon it can exist as a pair of enantiomers, a D-form (R) and an L-form (S). All of the aldoses and the ketoses with four or more carbons contain at least one chiral carbon and therefore can exist as at least one pair of enantiomers. Like the amino acids, only one enantiomeric form of the carbohydrates is primarily used by cells. The D-form of the carbohydrates is the biologically active form. L-forms are used to a limited extent in specialized structures or for specialized functions.
The larger aldoses and ketoses can be modeled as extensions of glyceraldehyde or dihydroxyacetone. A secondary alcohol functional group H-C-OH or its mirror image HO-C-H is inserted between the carbonyl carbon and the adjacent secondary alcohol group. When these groups are inserted, two possible stereoisomers result; one with the -OH group pointing to the right the other with the -OH group pointing left. Of the 15 possible D-form aldoses with 6 or fewer carbons the ones most often encountered in living organisms are: D-GLYCERALDEHYDE is a aldotriose, D-ERYTHROSE is a aldotetrose, D-RIBOSE (Rib) is a aldopentose, and D-GLUCOSE (Glc), D-GALACTOSE (Gal), & D-MANNOSE (Man) are aldohexoses.
C
C HHO
CH2OH
OHC
C OHH
CH2OH
OHCH2OH
C O
CH2OHL-Glyceraldehyde D-Glyceraldehyde Dihydroxyacetone
D-Glucose
H
C
C OH
C HHO
C OHH
C OHH
CH2OH
OH
H
C
C OH
C HHO
C HHO
C OHH
CH2OH
OH
D-Galactose
HO
C
C H
C HHO
C OHH
C OHH
CH2OH
OH
D-Mannose
C
C OHH
CH2OH
OH
D-Glyceraldehyde
C
C OHH
C OHH
CH2OH
OH
D-Erythrose
C
C OHH
C OHH
C OHH
CH2OH
OH
D-Ribose
©Kevin R. Siebenlist, 20194

Of the 8 possible D-form ketoses with 6 or fewer carbon atoms the ones most often encountered are: DIHYDROXYACETONE is a ketotriose, D-RIBULOSE & D-XYLULOSE are ketopentoses, and D-FRUCTOSE (Fru) is a ketohexose. While the ketone functional group can be on any interior carbon of the molecule, all of the naturally occurring ketoses have the carbonyl group on carbon two.
Except for dihydroxyacetone all of the monosaccharides depicted here contain one or more chiral carbon atoms. All of the sugars presented here are the D-enantiomeric form. When more than one chiral carbon is present in the molecule how is the D-form determined? If the configuration of the chiral carbon furthest from the carbonyl carbon; if the configuration of the last chiral carbon is identical to that of D-glyceraldehyde (i.e., it points to the right in the Fischer Projection), it is a D-monosaccharide. If this carbon has a configuration opposite from the configuration of D-glyceraldehyde (i.e., the –OH group points to the left in the Fischer Projection); if it matches L-glyceraldehyde, it is a L-sugar.
The aldohexoses (Glc, Gal, & Man) each contain four chiral carbons. Comparing the structure of D-glucose with D-mannose; notice that they differ from one another by the configuration about only one of the four chiral carbons. D-glucose differs from D-mannose at carbon two. Comparing the structures of D-glucose and D-galactose; notice that D-glucose differs from D-galactose by the configuration at carbon four. Molecules with more than one chiral carbon having identical configurations around all of the chiral carbons except for one are called EPIMERS. D-glucose & D-mannose are epimers of each other, as are D-glucose & D-galactose and the D-xylulose & D-ribulose pair. Molecules with more than one chiral carbon and related configurations, other than an enantiomeric relationship or an epimeric relationship, are DIASTEREOMERS. D-Mannose & D-galactose are diastereomers or each other.
Cyclization - Intramolecular Hemiacetal or Hemiketal Formation
Alcohols react with aldehydes to form Hemiacetals
Alcohols react with ketones to form Hemiketals.
CH2OH
C O
C HHO
C OHH
C OHH
CH2OH
D-Fructose
C
C HHO
C OHH
CH2OH
CH2OH
O
D-Xylulose
C
C OHH
C OHH
CH2OH
CH2OH
O
D-Ribulose
CH2OH
CH2OH
C O
Dihydroxyacetone
C OH C RH
OC O C
OH
H
R+
Hemiacetal
©Kevin R. Siebenlist, 20195
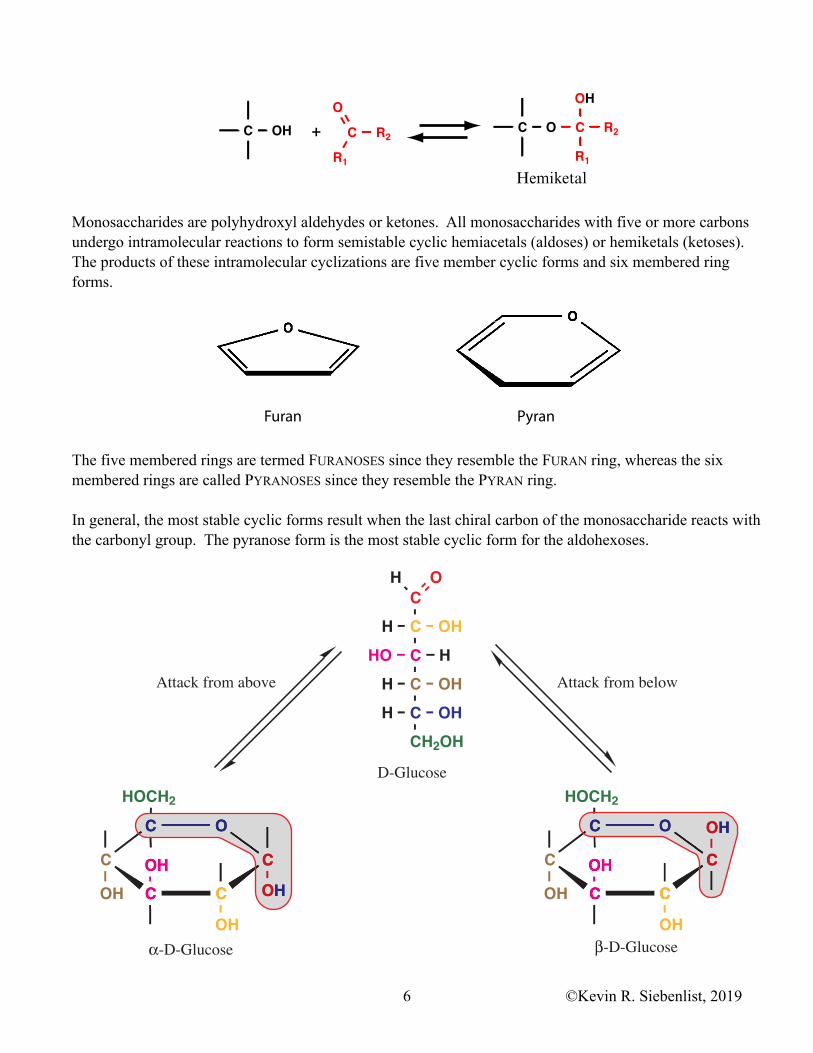
Monosaccharides are polyhydroxyl aldehydes or ketones. All monosaccharides with five or more carbons undergo intramolecular reactions to form semistable cyclic hemiacetals (aldoses) or hemiketals (ketoses). The products of these intramolecular cyclizations are five member cyclic forms and six membered ring forms.
The five membered rings are termed FURANOSES since they resemble the FURAN ring, whereas the six membered rings are called PYRANOSES since they resemble the PYRAN ring.
In general, the most stable cyclic forms result when the last chiral carbon of the monosaccharide reacts with the carbonyl group. The pyranose form is the most stable cyclic form for the aldohexoses.
C OH +
Hemiketal
C R2R1
OC O C
OH
R1
R2
Furan Pyran
C
C
C C
C
O
OH OH
OH
OH
HOCH2
HCC OHC HHOC OHHC OHHCH2OH
OH
C
C
C C
C
O
OH
OH
OH
OH
HOCH2
α-D-Glucose
D-Glucose
β-D-Glucose
Attack from belowAttack from above
©Kevin R. Siebenlist, 20196
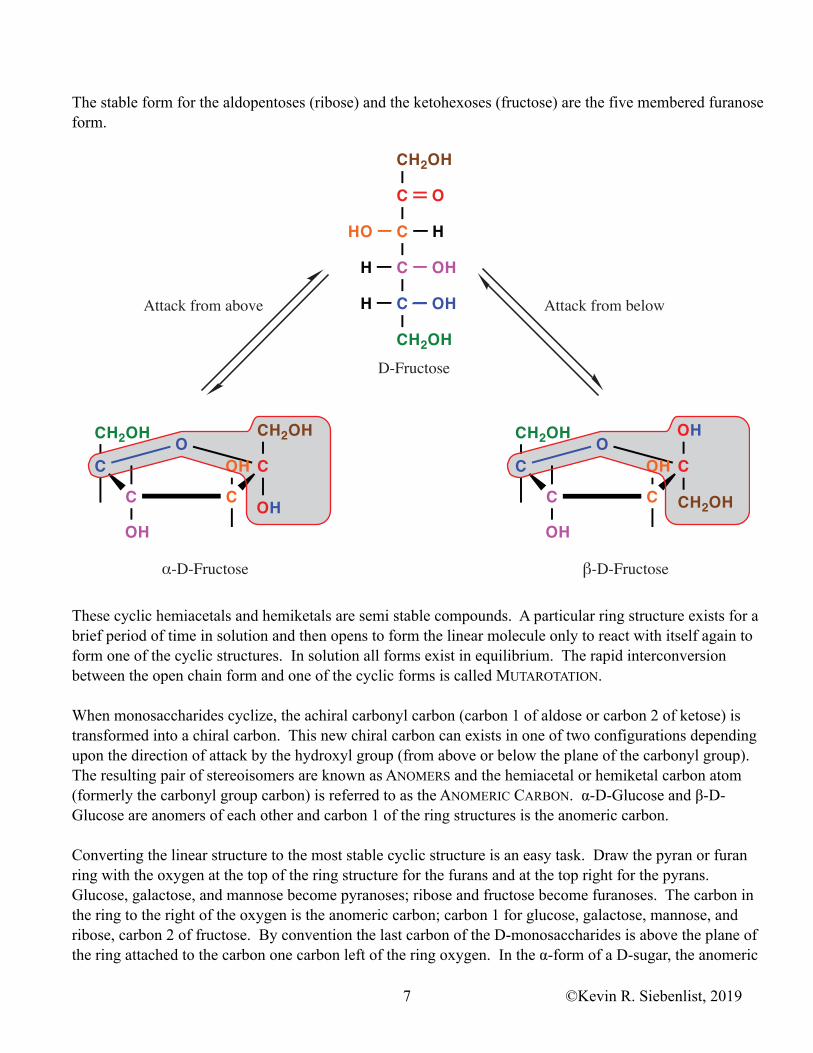
The stable form for the aldopentoses (ribose) and the ketohexoses (fructose) are the five membered furanose form.
These cyclic hemiacetals and hemiketals are semi stable compounds. A particular ring structure exists for a brief period of time in solution and then opens to form the linear molecule only to react with itself again to form one of the cyclic structures. In solution all forms exist in equilibrium. The rapid interconversion between the open chain form and one of the cyclic forms is called MUTAROTATION.
When monosaccharides cyclize, the achiral carbonyl carbon (carbon 1 of aldose or carbon 2 of ketose) is transformed into a chiral carbon. This new chiral carbon can exists in one of two configurations depending upon the direction of attack by the hydroxyl group (from above or below the plane of the carbonyl group). The resulting pair of stereoisomers are known as ANOMERS and the hemiacetal or hemiketal carbon atom (formerly the carbonyl group carbon) is referred to as the ANOMERIC CARBON. α-D-Glucose and β-D-Glucose are anomers of each other and carbon 1 of the ring structures is the anomeric carbon.
Converting the linear structure to the most stable cyclic structure is an easy task. Draw the pyran or furan ring with the oxygen at the top of the ring structure for the furans and at the top right for the pyrans. Glucose, galactose, and mannose become pyranoses; ribose and fructose become furanoses. The carbon in the ring to the right of the oxygen is the anomeric carbon; carbon 1 for glucose, galactose, mannose, and ribose, carbon 2 of fructose. By convention the last carbon of the D-monosaccharides is above the plane of the ring attached to the carbon one carbon left of the ring oxygen. In the α-form of a D-sugar, the anomeric
Attack from belowAttack from above
CH2OH
C
C
C
C
CH2OH
O
HHO
H
OHH
OH
C CH2OH
OHCH2OH
OH
CCCOH
O
C OH
CH2OHCH2OH
OH
CCC
OOH
©Kevin R. Siebenlist, 20197

hydroxyl group is below the plane of the ring, on the side of the ring opposite from the terminal carbon. In the β-form the anomeric hydroxyl group is above the plane of the ring. For fructose, carbon 1 is attached to the anomeric carbon, opposite from the anomeric hydroxyl group. The hydroxyl group oxygen of the last chiral carbon is the oxygen in the ring. For the remaining hydroxyl groups, if it is pointing to the left in the linear structure it is above the plane of the ring in the cyclic structure, if it is pointing to the right in the linear structure it is below the ring.
��- D - G l u c o s e D - G l u c o s e ��- D - G l u c o s e
O H C
C
C C
C
O
O H
O H
O H
O H C
C
C C
C
O O H
O H
O H
H
C
C O H
C H H O
C H H O
C O H H
C H 2 O H
O H
��- D - G a l a c t o s e D - G a l a c t o s e ��- D - G a l a c t o s e
C
C
C C
C
O
O H O H
O H O H
C
C
C C
C
O
O H
O H
O H O H
H O
C
C H
C H H O
C O H H
C O H H
C H 2 O H
O H
��- D - M a n n o s e D - M a n n o s e ��- D - M a n n o s e
C
C
C C
C
O
O H O H
O H
O H
C
C
C C
C
O
O H
O H
O H
O H
H
C
C O H
C H H O
C O H H
C O H H
C H 2 O H
O H
C H 2 O H
C H 2 O H C H 2 O H
C H 2 O H
C H 2 O H C H 2 O H
©Kevin R. Siebenlist, 20198
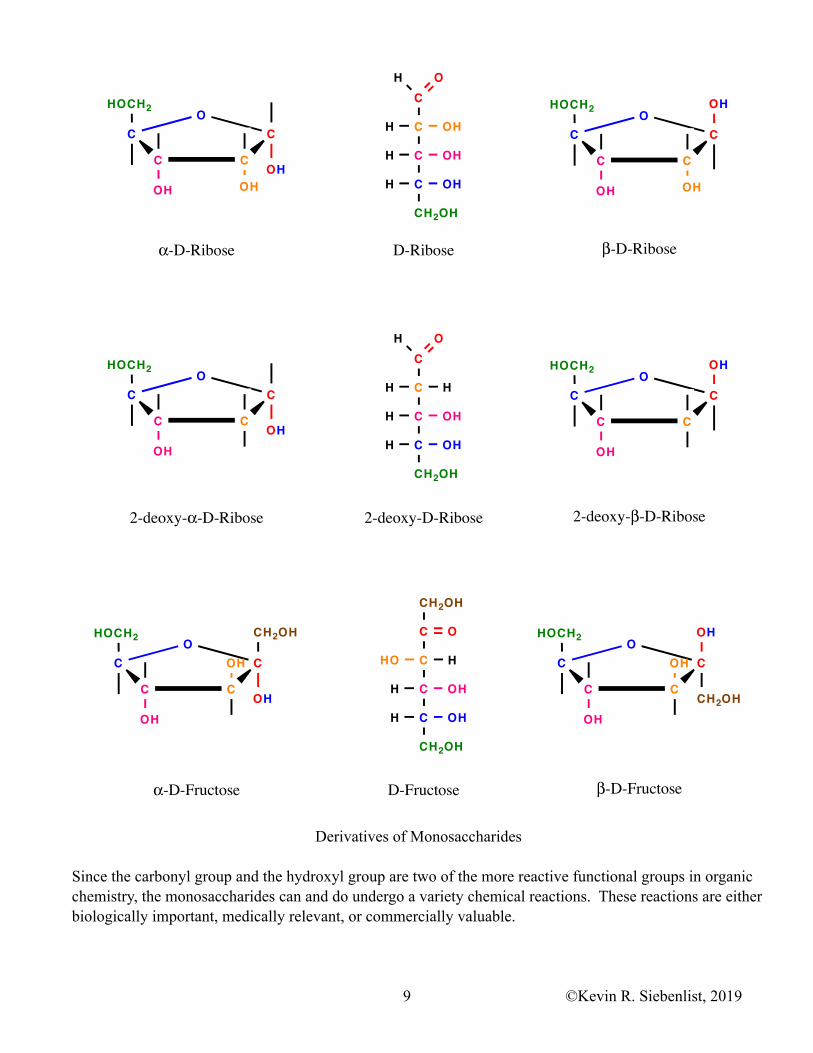
Derivatives of Monosaccharides
Since the carbonyl group and the hydroxyl group are two of the more reactive functional groups in organic chemistry, the monosaccharides can and do undergo a variety chemical reactions. These reactions are either biologically important, medically relevant, or commercially valuable.
C
C OHH
C OHH
C OHH
CH2OH
OH
HOCH2
COH
OH OH
CC
C
OHOCH2
C
OH
OH OH
CC
C
O
�-D-Ribose D-Ribose �-D-Ribose
C
C HH
C OHH
C OHH
CH2OH
OH
HOCH2
COH
OH
CC
C
OHOCH2
C
OH
OH
CC
C
O
2-deoxy-�-D-Ribose 2-deoxy-D-Ribose 2-deoxy-�-D-Ribose
CH2OH
C O
C HHO
C OHH
C OHH
CH2OH
HOCH2
COH
CH2OH
OH
CC
C
OHO
HOCH2
CCH2OH
OH
OH
CC
C
OHO
D-Fructose�-D-Fructose �-D-Fructose
©Kevin R. Siebenlist, 20199

α
α α
©Kevin R. Siebenlist, 201910

1. The aldehyde group of the aldoses can by chemically or enzymatically oxidized to a carboxyl group forming ALDONIC ACIDS. Aldonic acids are formed when aldoses are reacted with mild oxidizing agents such as the silver (Ag+) ion of TOLLEN’S REAGENT or the cupric (Cu+2) ion of BENEDICT’S REAGENT can oxidize aldehydes. When Ag+ is the oxidizing agent it is reduced to metallic silver. Oxidation by Cu+2 results in a red precipitate of Cu2O. All of the aldoses can be oxidized by these mild oxidizing agents. The aldoses are called REDUCING SUGARS. The Tollen’s test and Benedict’s test were early analytical procedures used to test for sugars in bodily fluids. {Ketones cannot be easily oxidized and on that basis ketoses are usually not considered reducing sugars. However, under strongly alkaline conditions, α-hydroxy ketones will isomerize to β-hydroxy aldehydes. The ketoses contain an α-hydroxy group and under the alkaline conditions of TOLLEN’S TEST or BENEDICT’S TEST they isomerize to aldoses that subsequently react.}
2. The last carbon of the monosaccharides (especially the hexoses) can be enzymatically oxidized by cells to form URONIC ACIDS.
3. The carbonyl group on the sugars can be reduced to a hydroxyl group forming SUGAR ALCOHOLS. Glucitol (Sorbitol), Mannitol, Xylitol, and Erythritol are all used as artificial sweeteners.
4. The hydroxyl groups of monosaccharides, disaccharides, and/or polysaccharides can be esterified with a variety of organic acids, with phosphate, or with sulfate.
5. The hydroxyl groups on monosaccharides can be enzymatically or chemically reduced to form DEOXYSUGARS. 2-Deoxyribose found in the deoxyribonucleotides is an important deoxysugar.
6. Aldoses undergo an enzyme catalyzed substitution reaction in which one of the hydroxyl groups is replaced by an amino group. The product is an AMINO SUGAR. The hydroxyl group on carbon two of glucose, galactose, or mannose is the group usually replaced. These modified sugars are named by dropping the ‘e’ from the name and adding ‘amine’.
7. The amino group on the amino sugars is often modified by the addition of an acetate group in amide linkage. Muramic Acid is a component of bacterial cell walls. It is glucosamine with lactic acid in ether linkage at carbon three. N-Acetylneuraminic Acid (Sialic Acid) is an component of many eukaryotic glycoconjugates.
Non-Enzymatic Glycation
HCN C
OH
CH2CH2CH2CH2N
C
HC
CH
HC
HC
H2COH
H
OH
HO
OH
OH
HCN C
OH
CH2CH2CH2CH2NH
C
C
CH
HC
HC
H2COH
H
OH
HO
OH
OH
HCN C
OH
CH2CH2CH2CH2NH
HC
C
CH
HC
HC
H2COH
H
O
HO
OH
OH
Schiff BaseGlycation Adduct
AmadoriProduct
Amadori
Rearrangement
LysSide
Chain
HCN C
OH
CH2CH2CH2CH2NH3
Glc
C
HC
CH
HC
HC
H2COH
OH
HO
OH
OH
O H
+
Amadori
Rearrangement
©Kevin R. Siebenlist, 201911

The aldehyde group of monosaccharides will react slowly and non-enzymatically with amino groups. A lysine side chain of cellular proteins are the usual source of the amino groups. Initially a Schiff Base is formed that subsequently rearranges to form a stable keto-amine called an Amadori product. The rate of this reaction is temperature and monosaccharide dependent. After these GLYCATION ADDUCTS form they under go additional reactions (e.g., oxidations, rearrangements, dehydrations, cyclizations, etc.) resulting in protein-bound compounds collectively called ADVANCED GLYCATION END products (AGEs). Glycation of long-lived proteins like collagen or elastin disrupts the structure and function of vascular and connective tissue. The build up of AGEs has been linked to vascular and neurodegenerative diseases, arthritis, and atherosclerosis of aging and diabetes mellitus.
The Disaccharides
The Anomeric Hydroxyl Group, the new hydroxyl group formed when a sugar forms a cyclic hemiacetal or hemiketal, is a very reactive hydroxyl group. It is much more reactive than the other hydroxyl groups present on the monosaccharides. The hydroxyl group attached to the Anomeric Carbon reacts readily with other alcohols to form ACETALS or KETALS.
Acetals and ketals are much more stable than hemiacetals and hemiketals. When the reaction is between the anomeric hydroxyl group on one sugar and a hydroxyl group on a second sugar the resulting bond is called a GLYCOSIDIC BOND. Glycosidic bonds are the chemical bonds (acetal or ketal functional groups) formed when monosaccharides react to produce disaccharides, oligosaccharides, and polysaccharides. It is important to realize that glycosidic bonds always involve the anomeric hydroxyl group of one sugar reacting with a hydroxyl group on the second sugar.
There are four disaccharides found in high abundance in nature; three of which are of major importance to the biochemistry of mammals and the human animal. These three disaccharides serve as a source of monosaccharide precursors. These disaccharides are MALTOSE, LACTOSE, and SUCROSE.
MALTOSE is a disaccharide formed by the partial hydrolysis of the polysaccharide, starch. When grains are “malted” the starch they contain is broken down to maltose. Maltose is composed of two glucose molecules linked by a glycosidic bond. The anomeric hydroxyl group in the α-configuration on the first glucose molecule is covalently linked by the hydroxyl group on carbon four of the second glucose. Maltose is the common name for this disaccharide. The biochemically proper name for maltose is α-D-
HO C
HO C
+
+
Hemiacetal Acetal
Hemiketal Ketal
+
+
HOH
HOHC O C
R1
R2
OH
C O C
R
H
OH C O C
R
H
O C
C O C
R1
R2
O C
©Kevin R. Siebenlist, 201912

GLUCOPYRANOSYL-(1 → 4)-D-GLUCOPYRANOSE which is usually shortened to GLUCOSE (α1 → 4) GLUCOSE. The notation α1 → 4 indicate the direction of the glycosidic bond - the anomeric hydroxyl of the first monosaccharide in the α configuration is linked to the hydroxyl group on carbon four of the second monosaccharide. The first glucose, the one that donates the anomeric hydroxyl group, is locked in the ring configuration. The second glucose molecule still has a free, unreacted anomeric hydroxyl group that can undergo mutarotation. It can undergo ring opening and closure. When open, when in the straight chain form, there is a free aldehyde group that can be oxidized by Cu++ or Ag+; hence, maltose is a reducing sugar. LACTOSE is the sugar present in mammal’s milk. In lactose the anomeric hydroxyl group of a galactose in the β-configuration is linked to the C-4 hydroxyl group of a glucose. β-D-GALACTOPYRANOSYL-(1 → 4)-D-GLUCOPYRANOSE is the full biochemical name and GALACTOSE (β1 → 4) GLUCOSE is a shortened biochemical name for this molecule. The glucose residue of lactose can still undergo mutarotation since its anomeric hydroxyl group is free to open. Lactose is a reducing sugar.
SUCROSE is the sugar isolated and purified from sugar cane or sugar beets. It is table sugar. In sucrose, the anomeric hydroxyl group of glucose in the α configuration is glycosidically linked to the anomeric hydroxyl group of fructose in the β configuration. The proper biochemical name for sucrose is α-D-GLUCOPYRANOSYL-(1 ↔ 2)-β-D-FRUCTOFURANOSIDE, or β-D-FRUCTOFURANOSYL-(2 ↔ 1)-α-D-GLUCOPYRANOSIDE. The contracted biochemical name that most every one uses is GLUCOSE (α1 ↔ β2) FRUCTOSE. Since the anomeric hydroxyl groups of both sugars are involved in the glycosidic bond, neither of these sugars can undergo mutarotation. Sucrose is not a reducing sugar.
FYI: The fourth disaccharide found in nature is TREHALOSE. Trehalose is the major sugar that circulates in the hemolymph of arthropods {insects and crustaceans}. In mammalian blood the major
©Kevin R. Siebenlist, 201913
Maltose{ -D-glucopyranosyl-(1 → 4)-D-glucopyranose}
{glucose ( 1 → 4) glucose}
O
OH
OH
OH
HOCH2O
OH
OH
OH
HOCH2
O
Lactose{ -D-galactopyranosyl-(1 → 4)-D-glucopyranose}
{galactose ( 1 → 4) glucose}
O
OH
OH
OH
HOCH2
OH O
OH
OH
HOCH2
O
O
OH
OH
OH
HOCH2
HOCH2
CH2OH
O
OH
OHO
Sucrose{ -D-glucopyranosyl-(1 2)- -D-fructofuranoside}{ -D-fructofuranosyl-(2 1)- -D-glucopyranoside}
{glucose ( 1 2) fructose}

circulating sugar is the monosaccharide glucose. Structurally,trehalose is GLUCOSE (α1 → α1) GLUCOSE. Since the anomeric hydroxyl groups of both sugars are involved in the glycosidic bond, neither of these sugars can undergo mutarotation. Trehalose is not a reducing sugar.
The Polysaccharides
Polysaccharides are polymers of twenty or more monosaccharide units. The polysaccharides can be HOMOPOLYMERS, composed of a single type of monosaccharide, or they can be HETEROPOLYMERS, made up from a variety of different monosaccharides and/or modified monosaccharides. In addition they can have linear or branched structures. The glycosidic bonds between aldose monosaccharide units can be 1 → 2, 1 → 3, 1 → 4, or 1 → 6, whereas the bonds between ketose monosaccharide units can be 2 → 1, 2 → 3, 2 → 4, or 2 → 6 with the anomeric carbon in either the α of β configuration.
The Homopolymers
GLUCANS are homopolymers of glucose, MANNANS are homopolymers of mannose and LEVANS are homopolymers of fructose. The homopolymers described below are all glucans.
CELLULOSE is the main structural element of plant cells. It is part of the cell wall and is a major component
O
OH
OH
HOCH2O
OH
OH
HOCH2
O
O
OH
OH
HOCH2O
OH
OH
HOCH2
OO
Cellulosepoly (glucose β1 → 4 glucose)
O
OH
OH
HOCH2
O
OH
OH
HOCH2
O
OH
OH
HOCH2
O
OH
OH
HOCH2O
O
O
O
OH
OH
HOCH2
O
OH
OH
HOCH2
OO
O
OH
OH
HOCH2
O
OH
OH
HOCH2
O
©Kevin R. Siebenlist, 201914

of wood. Cellulose is a linear polymer of glucose linked by β1 → 4 glycosidic bonds. The glucose-(β1 → 4)-glucose glycosidic bonds in cellulose results in a rigid linear polymer and an excellent structural component. The glucose-(β1 → 4)-glucose bonds are very stable, they are difficult or impossible to hydrolyze by chemical means. In addition, higher order multicellular animals do not contain an enzyme capable of hydrolyzing the glucose-(β1 → 4)-glucose glycosidic bond. Only a very few species of bacteria and the wood-rot fungi contain the CELLULASE ENZYME, the enzyme that can hydrolyze the β1 → 4 bond of cellulose. Many of the higher animals that derive their nutrition primarily from plant material utilize these bacteria in some shape, manner, or form to hydrolyze the glucose-(β1 → 4)-glucose bond of cellulose.
STARCH is the storage form of glucose units in plant cells. In plants, seeds, tubers, and/or bulbs the starch is broken down (hydrolyzed) when needed and used for energy and/or carbon precursors. Starch, as isolated from plants, is a mixture of two polymers - AMYLOSE and AMYLOPECTIN. AMYLOSE is a linear polymer of glucose units linked by α1 → 4 glycosidic bonds. AMYLOPECTIN is a branched polymer. It’s backbone is glucose units linked by α1 → 4 glycosidic bonds. Every 24 to 30 residues along a backbone there is a branch point. At the branch points the glucose residues are linked in an α1 → 6 configuration.
GLYCOGEN is a homopolymer of glucose. GLYCOGEN is the storage form of glucose in animal cells. In
O
OH
OH
CH2
OO
O
OH
OH
CH2
O
OH
OH
CH2
OHO
HOHOHO
O
O
OH
OH
CH2
O
OH
OH
CH2
O
HOHO
O
OH
OH
CH2
OO
O
OH
OH
H2C
O
OH
OHO
O
OH
OH
CH2
O
HO
O
O
OH
OH
CH2HO
OH
O
OH
OH
CH2
O
HO
O
OH
OH
CH2
O O
HO
O
OH
OH
CH2
O O
HOH2C
OH
Amylosepoly (glucose α1 → 4 glucose)
Amylopectinpoly (glucose α1 → 4 glucose)
glucose α1 → 6 glucose branches every 24 to 30 residues
Glycogenpoly (glucose α1 → 4 glucose)
glucose α1 → 6 glucose branches every 8 to 12 residues
©Kevin R. Siebenlist, 201915

man, glycogen is stored by most tissues with the liver and skeletal muscle being major sites of storage. When energy poor the glycogen is broken down to glucose and the resulting glucose is used for energy and/or precursors. Glycogen is a branched polymer similar to amylopectin. It has a backbone of glucose residues linked in an α1 → 4 configuration and branches that are linked α1 → 6. Glycogen is different from amylopectin in that glycogen is much more highly branched. It contains an α1 → 6 branch every 8 to 13 residues along a backbone.
The branched polysaccharides, AMYLOPECTIN and GLYCOGEN, have only one reducing end and numerous nonreducing ends. All of the biochemical reactions involving these polysaccharides occurs at the numerous nonreducing ends of these molecules.
DEXTRANS are a large and important family of storage polysaccharides in bacteria and yeast. The glycosidic linkage along the backbone of these polysaccharides is α1 → 6. Dextrans are usually branched polysaccharides. The degree of branching and the average branch length depends upon the bacterial species. Likewise, the bonds at the branch points can be 1 → 2, 1 → 3, or 1 → 4 depending on the bacterial species. Bacteria growing on tooth surfaces produce extracellular accumulations of dextrans which are an important component of DENTAL PLAQUE. The dextrans produced by oral bacteria are usually glucans or levans.
The Heteropolysaccharides
Heteropolysaccharides are composed of more than one type of monosaccharide unit. They often contain some of the less common monosaccharides as well as many of the modified monosaccharides. Heteropolysaccharides often contain amino sugars, acetylated amino sugars, sugars that have been oxidized to uronic acids, and sugars that have been esterified with phosphate or sulfate. With this vast array of building blocks (precursor molecules) a very large number of different heteropolysaccharides are possible and a very large number of different heteropolysaccharides have been found in nature. Heteropolysaccharides are usually components of larger biomolecules. They are usually covalently linked to proteins to form a GLYCOPROTEIN or a PROTEOGLYCAN or they can be covalently attached to lipids, forming GLYCOLIPIDS. GLYCOLIPIDS are found in cellular membranes. When compared to homopolysaccharides, heteropolysaccharides have different and unique functions. Heteropolysaccharides:
1. increase the solubility of the molecules to which they are attached. They increase solubility because they contain many hydroxyl and other polar groups that form hydrogen bonds with extracellular water.
2. serve as parts of structural elements within the cell and within the extracellular matrix. 3. orient molecules correctly in cellular membranes. Molecules that make up the cell membrane often
contain covalently attached heteropolysaccharides with the polysaccharide directed extracellularly, directed toward the outside of the cell.
4. are signal molecules. Many extracellular proteins, proteins secreted by the cell, have heteropolysaccharides covalently attached. When attached to the protein they signal to the cell that the protein is to be secreted.
5. are cellular adhesion molecules. Heteropolysaccharides attached to the outside of cells help to hold cells together in tissues and organs by forming numerous hydrogen bonds with molecules in the extracellular matrix and/or with molecules on adjacent cells.
6. are “cell markers”, are “antigenic determinants”. Antigenic determinants are the molecules that
©Kevin R. Siebenlist, 201916

must be “matched” before a transfusion or transplant can be attempted. Heteropolysaccharides covalently attached to other biomolecules on the cell surface are a subset of antigenic determinants that must be matched.
The heteropolysaccharides can be divided into two major types. There are heteropolysaccharides that are composed of repeating (-A-B-) modified disaccharide units. Heteropolysaccharides of this type are the GLYCOSAMINOGLYCANS. The glycosaminoglycans are covalently linked to peptides or proteins to form PEPTIDOGLYCANS or PROTEOGLYCANS. The other type of heteropolysaccharides are composed of an assortment of monosaccharide and/or modified monosaccharide units arranged in a somewhat “random” order, a specific order with no discernible pattern. This later group is often branched.
Glycosaminoglycans
GLYCOSAMINOGLYCANS are unbranched polysaccharides composed of repeating disaccharide units. One of the monosaccharide units is an amino sugar (glycosamino-) and and the other sugar of the repeating unit is most often a Uronic Acid. In the polymer, one or both of the monosaccharide units often contain at least one negatively charged sulfate ion in ester linkage to one of the hydroxyl groups of the modified sugars. There are five major glycosaminoglycans and several minor forms each with a unique pair of repeating monosaccharide units. The five major glycosaminoglycans are: 1. HYALURONAN (HYALURONIC ACID) contains D-Glucuronate
(ionized form of D-Glucuronic Acid) and N-Acetyl-D-glucosamine. This molecule is unique for several reasons. It is the largest of the glycosaminoglycans containing up to 50,000 disaccharide units, it is not linked to protein molecules, and it is not sulfated. It is most often found in cartilage where it modulates compressibility, synovial fluid, loose connective tissue, and vitreous humor. Hyaluronan also plays a role in directing cell migration through the extracellular matrix.
2. CHONDROITIN SULFATE contains D-Glucuronate (ionized form of D-Glucuronic Acid) and N-Acetyl-D-galactosamine. The hydroxyl group on C-4 or C-6 of the N-acetyl-D-galactosamine unit is often esterified with a molecule of sulfate. This molecule is found in tendons, ligaments, cartilage, bone and the aorta where it contributes to the tensile strength and modulates the compressibility of these tissues.
3. DERMATAN SULFATE is composed of L-Iduronate (ionized form of L-Iduronic Acid) and N-Acetyl-D-
O
NH
CH2OH
OO
OH
OH
COO–
C O
CH3
–O3SO
O
D-Glucuronate N-Acetyl-D-Galactosamine-4-sulfate
�� � �
�� �
Chondroitin-4-sulfate
O
NH
CH2OSO3–
OO
OH
OH
COO–
C O
CH3
OH
O
D-Glucuronate N-Acetyl-D-Galactosamine-6-sulfate
�� � �
�� �
Chondroitin-6-sulfate
©Kevin R. Siebenlist, 201917
O
HOOO
OH
OH
COO–
C O
CH3
NH
CH2OH
O
D-Glucuronate N-Acetyl-D-Glucosamine
�� � �
�� �
Hyaluronan (Hyaluronic Acid)

galactosamine. L-IDURONATE is the C-5 epimer of D-GLUCURONATE. D-Glucuronate is incorporated into the growing molecule and then enzymatically epimerized into L-Iduronic acid. The N-acetyl-D-galactosamine is often sulfated. This molecule is found in most tissues, but is especially rich in skin and blood vessels where it increases the pliability of these tissues. It is also found in the cornea of the eye where it functions to maintain the transparency.
4. KERATAN SULFATE contains N-Acetyl-D-glucosamine and D-
Galactose. N-acetyl-D-glucosamine is most often sulfated, but both of the sugars can be sulfated. It is most often found in the cornea of the eye where it plays a role in corneal transparency. It is also present in cartilage, tendons and the “dead” tissues of hair, horns, hooves, and claws.
5. HEPARIN is composed of D-Glucuronate (ionized form of D-Glucuronic Acid) and D-Glucosamine. Both monosaccharide units in this polysaccharide are derivatives of glucose and both are sulfated. The glycosidic bonds in heparin are all in the α configuration. Heparin may also contain variable amounts of L-Iduronic acid. This molecule is often found in MAST CELLS where it has anticoagulant activity.
PROTEOGLYCANS are glycosaminoglycans covalently linked to tissue specific core proteins. Proteoglycans are components of the extracellular matrix, the scaffold upon which cells migrate and communicate. As part of the extracellular matrix, proteoglycans interact with collagen and other proteins in the extracellular matrix as well as with proteins on the cell surface to play a role in tissue morphology. The proteoglycans also act as selective filters to regulate the movement of extracellular compounds based upon electrical charge, molecular size or both. They play a role in regulating the activity of growth factors. Proteoglycans attract and bind considerable amounts of water making tissues that contain these molecules in high concentration “spongy”, e.g., cartilage.
“Random” Structured Heteropolysaccharides
While the Glycosaminoglycans sometimes exist as individual functional molecules, especially in the extracellular matrix, the heteropolysaccharides with a more “random” structure rarely, if ever, exist as free molecules. These oligosaccharides and/or polysaccharides are covalently linked to a protein to form a Glycoprotein or to a lipid to form a Glycolipid.
The oligosaccharide/polysaccharide part of a glycoprotein is an information rich structure - THE SUGAR CODE or the GLYCOME. The possibility of forming 1 → 2, 1 → 3, 1 → 4, 1 → 6, 2 → 3, 2 → 4, or 2 → 6 glycosidic bonds between the monosaccharides with the anomeric carbon in either the α of β configuration allows for a great deal of structural diversity (i.e., information) with a limited number of different
©Kevin R. Siebenlist, 201918
O
NH
CH2OH
OO
OH
OH
COO–
C O
CH3
–O3SO
O
L-Iduronate N-Acetyl-D-Galactosamine-4-sulfate
�� �
�� � �
Dermatan sulfate
O
HONH
CH2OSO3–
OOOH
OH
CH2OH
O
C O
CH3
D-Galactose N-Acetyl-D-Glucosamine-6-sulfate
�� � �
�� � �
Keratan sulfate
O
NH
OH
CH2OSO3–
O
O
OH
OSO3–
COO–
O
SO3–D-Glucuronate-2-sulfate N-Sulfo-
D-glucosamine-6-sulfate
�� � � �� � �
Heparin

monosaccharides (words).
LECTINS are proteins found in all organisms that bind carbohydrates with high affinity and specificity. Binding interactions between the oligosaccharide part of the glycoprotein and the lectin mediates the diverse cellular responses brought about by the glycoproteins. The unique structures, the information rich nature of the oligosaccharide allows the lectins to bind with high specificity to the “correct” glycoprotein. SELECTINS are a family of lectins present in the cell membrane. They mediate cell-cell recognition and adhesion in a wide range of cellular processes.
The oligosaccharides or polysaccharides can be linked to the protein by a glycosidic bond involving the hydroxyl group of serine, threonine, or hydroxylysine. Glycoproteins of this type are called O-LINKED SACCHARIDES. In the O-linked glycoproteins, the sugar most often directly attached to the protein is N-ACETYLGALACTOSAMINE. O-linked saccharides can be as small as a single N-acetylgalactosamine or a large heteropolysaccharide terminating in sialic acid residues.
One example of the structural diversity present in O-linked oligosaccharides / polysaccharides is the ABO Blood Group Antigens. These oligosaccharides are cell surface proteins as well as components of glycolipids present in the cell membrane. All three oligosaccharides contain an identical core structure. The presence or absence of one additional sugar moiety, and the nature of the additional sugar determines
MembraneComponent
N-Acetylgalactosamine
Galactose N-Acetylgalactosamine
Fucose
N-Acetylneuraminic Acid
(�1 � 2)
(�1 � 3) (�1 � 3)
(�� � 6)�
Blood Group A Antigen
Blood Group B Antigen
MembraneComponent
Galactose
Galactose N-Acetylgalactosamine
Fucose
N-Acetylneuraminic Acid
(�1 � 2)
(�1 � 3) (�1 � 3)
(�� � 6)�
Blood Group O Antigen
MembraneComponentGalactose N-Acetylgalactosamine
Fucose
N-Acetylneuraminic Acid
(�1 � 2)
(�1 � 3)
(�� � 6)�
©Kevin R. Siebenlist, 201919

whether an individual is Blood Type A, Type B, or Type O.
Alternatively, the polysaccharide can be attached by an N-glycosidic bond to the amide nitrogen of an asparagine side chain in the protein. These are the N-LINKED SACCHARIDES. The N-linked saccharides always have N-ACETYLGLUCOSAMINE in a N-glycosidic bond to the protein and they always contain the core structure:
Attached to this core structure there can be: 1. up to six additional mannose residues (High Mannose Type Polysaccharide) 2. a complex heteropolysaccharide terminating in sialic acids (Complex Type Polysaccharide) 3. a mannose homopolysaccharide on one branch and a heteropolysaccharide on the other branch
(Mixed or Hybrid Type Polysaccharide).
Man �1 � 4 GlcNAc �1 � 4 GlcNAc �1 � N-Asn-Protein
Man
Man
�1�� 3
�1�� 6
©Kevin R. Siebenlist, 201920


















