
CARACTERIZAÇÃO FLORÍSTICA E ESTRUTURAL DE ONZE … · A análise da composição florística e...
235
i UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE BIOLOGIA CARACTERIZAÇÃO FLORÍSTICA E ESTRUTURAL DE ONZE FRAGMENTOS DE MATA ESTACIONAL SEMIDECIDUAL DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE CAMPINAS – SP Karin dos Santos Orientadora: Profa. Dra. Luiza Sumiko Kinoshita Co-Orientador: Prof. Dr. Flávio A. Maes dos Santos Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas para a obtenção do título de Doutor em Biologia Vegetal 2003
Transcript of CARACTERIZAÇÃO FLORÍSTICA E ESTRUTURAL DE ONZE … · A análise da composição florística e...

i
UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
CARACTERIZAÇÃO FLORÍSTICA E ESTRUTURAL DE ONZE FRAGMENTOS DE MATA ESTACIONAL SEMIDECIDUAL
DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE CAMPINAS – SP
Karin dos Santos
Orientadora: Profa. Dra. Luiza Sumiko Kinoshita
Co-Orientador: Prof. Dr. Flávio A. Maes dos Santos
Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas para a obtenção do título de Doutor em Biologia Vegetal
2003

ii

iii
Data da defesa: 4/11/2003 Luiza Sumiko Kinoshita ___________________________________ Fernando Roberto Martins ___________________________________ Ricardo Ribeiro Rodrigues ___________________________________ Waldir Mantovani ___________________________________ Jean Paul Walter Metzger ___________________________________ Luciana Ferreira Alves ___________________________________ Roseli Buzanelli Torres ___________________________________

iv
Dedico este trabalho à todos aqueles
que preservam ou lutam pela
preservação das florestas brasileiras.
Sem essas pessoas este trabalho
não teria sido possível.

v
AGRADECIMENTOS
Aos meus pais que sempre me apoiaram, incentivaram e estiveram ao meu lado em todos os momentos,
vivenciando comigo a dor e a delícia de todo esse processo. Por serem as pessoas maravilhosas e
brilhantes que são e, com isso me iluminarem o caminho.
À Capes pela bolsa de doutorado concedida através do Curso de Pós-Graduação em Biologia Vegetal da
UNICAMP e pelo auxílio financeiro no primeiro ano de realização deste trabalho.
Ao MMA/PROBIO/CNPq, pelo financiamento de parte do trabalho de campo, através do Projeto “Estudos
de conservação e recuperação de fragmentos florestais da APA do Camanducaia (SP/MG)”
Ao Programa Biota/FAPESP pelo auxílio financeiro que foi imprescindível para a realização deste trabalho.
À Universidade Estadual de Campinas, a “Mãe UNICAMP”, por me abrigar, instruir, apoiar, entreter,
alimentar, promover encontros, enfim, por tudo o que ela representa e proporciona.
À SCPG – Biologia Vegetal, pelo apoio à realização desta tese.
À Prof. Dra. Luiza Sumiko Kinoshita pela orientação, confiança, compreenção, apoio e amizade em todos
esses anos. Agradeço também por seu espírito empreendedor, que fez com que nunca faltassem recursos
para nossos projetos.
Ao Prof. Dr. Flavio A. Maës dos Santos, o Flavião, por ter sido muito mais que um co-orientador. Agradeço
por sua dedicação, interesse, paciência, visão e brilhantismo, os quais muito me motivaram nesse tempo
todo e por finalmente me fazer entender porque é tão importante saber “qual é a perrrgunta”. Seus
questionamentos e reflexões dão nó na cabeça de qualquer um, mas me sinto privilegiado por ter
partilhado suas idéias comigo, que foram fundamentais para a realização deste trabalho.
À Bióloga Giselda Person da EcoForça de Campinas, pela imprescindível ajuda em localizar as matas e
seus proprietários e a convecê-los a me deixar entrar em suas terras. Também pelas fotos áereas, mapas e
pela amizade, apoio e interesse em várias fases deste trabalho, sempre com muito carinho e bom humor.
À todos os fazendeiros, proprietários das matas deste estudo por terem permitido meu acesso aos
fragmentos e por serem os principais preservacionistas daquela região.
Ao Renato Belinello, meu ajudante de campo, pelo trabalho de touro. Sem sua ajuda esse trabalho teria
demorado o triplo. Formávamos uma dupla quase militar, desbravadora e no fim do dia éramos os
verdadeiros “survivors” da floresta! Valeu a pena. Obrigada!

vi
Ao Dr. Marcos Sobral (UFRGS), botânico brilhante e de grande simpatia, pela confirmação e identificação
das espécies de Myrtaceae.
Ao Dr. João Batista Baitello (IF, SMA-SP) por, novamente, me ajudar com as Lauraceae e por me encorajar
no estudo dessa família maravilhosa.
À Prof. Dra. Ana Maria de A. G. Tozzi (UNICAMP) pela ajuda com as Fabaceae, pelo eterno bom humor,
risadas fantásticas e por facilitar a vida dos alunos, a minha em especial, como uma coordenadora de pós
desatadora de nós. Com ela tudo é possível!
Ao Prof. Jorge Y. Tamashiro (UNICAMP) pelo auxílio com as Mimosaceae e por seu mau humor adorável!
Ao Prof. Dr. João Semir (UNICAMP) pela ajuda com as Asteraceae e pela convivência deliciosa no
laboratório de taxonomia.
Ao Dr. João A. Pastori (IF, SMA-SP) pela confirmação das identificações de Meliaceae e por seu bom
humor e interesse pelo material que lhe mostrei.
Ao Dr. Sérgio Romaniuc Neto, Dra. Inês Cordeiro e Dra. Lúcia Rossi do Instituto Botânico (IBT, SMA-SP)
pela confirmação das identificações de Moraceae, Euphorbiaceae e Olacaceae, respectivamente.
À Dra. Sigrid Jung-Mendaçoli (IAC) pelo auxílio com as Rubiaceae.
Às queridas Márcia Rocca e Michele de Sá pela ajuda na realização das fotografias hemisféricas em
campo, sempre com muita garra e bom humor. O trabalho, às vezes estenuante, na companhia delas, se
tornava divertido.
Aos colegas e amigos de Unicamp que me ajudaram em campo e compartilharam um pouco do perrengue:
Pricila, Dani, Lica, Carla, Giselda, Paula, Rossano, Aneliza, Evandro, Sílvia e Fabiana.
Ao Dr. Pedro Luís R. de Moraes pelas sugestões e auxílio na estatística.
Aos membros de pré-banca Fernando Roberto Martins, Sergius Gandolfi e Marcelo Tabarelli, pela leitura
cuidadosa, críticas e importantes sugestões.
Aos amigos Leonardo Meirelles, Lidyane Aona e Cristiano Verola pela companhia na fase de escrita desta
tese, durante as férias desérticas do departamento de Botânica-UNICAMP.

vii
SUMÁRIO Resumo geral.................................................................................................................................................... 1
Abstract ............................................................................................................................................................. 2
Introdução geral ................................................................................................................................................ 3
Referências bibliográficas................................................................................................................................. 6
CAPÍTULO 1 - Avaliação de aspectos associados ao estado de preservação de onze fragmentos de
floresta estacional semidecídua na região da Área de Proteção Ambiental do município de
Campinas, SP. ................................................................................................................................................. 8
Resumo......................................................................................................................................................... 9
Introdução................................................................................................................................................... 10
Material e Métodos ..................................................................................................................................... 12
Área de estudo....................................................................................................................................... 12
Amostragem........................................................................................................................................... 15
Aspectos qualitativos ............................................................................................................................. 15
Histórico dos fragmentos .................................................................................................................... 15
Qualidade do dossel ........................................................................................................................... 15
Abertura de dossel .............................................................................................................................. 16
Infestação por lianas........................................................................................................................... 18
Análise dos dados.................................................................................................................................. 18
Resultados.................................................................................................................................................. 19
Histórico dos fragmentos ....................................................................................................................... 19
Qualidade do dossel .............................................................................................................................. 23
Abertura de dossel................................................................................................................................. 25
Infestação por lianas.............................................................................................................................. 28
Discussão ................................................................................................................................................... 30
Referências bibliográficas .......................................................................................................................... 35

viii
CAPÍTULO 2 - Composição florística e similaridade de onze fragmentos de floresta estacional
semidecídua na região da Área de proteção Ambiental do município de Campinas, SP ............... 40
Resumo....................................................................................................................................................... 41
Introdução................................................................................................................................................... 42
Material e Métodos ..................................................................................................................................... 45
Área de estudo....................................................................................................................................... 45
Amostragem........................................................................................................................................... 47
Síndromes de dispersão........................................................................................................................ 48
Tratamento dos dados........................................................................................................................... 48
Resultados.................................................................................................................................................. 49
Aspectos florísticos ................................................................................................................................ 49
Síndromes de dispersão........................................................................................................................ 51
Similaridade ........................................................................................................................................... 54
Discussão ................................................................................................................................................... 58
Referências bibliográficas .......................................................................................................................... 64
Anexo I........................................................................................................................................................ 72
CAPÍTULO 3 - Riqueza e diversidade de espécies arbóreas em onze fragmentos de floresta
estacional semidecídua na região da Área de Proteção Ambiental do município de Campinas, SP ....
......................................................................................................................................................................... 82
Resumo....................................................................................................................................................... 83
Introdução................................................................................................................................................... 84
Material e Métodos ..................................................................................................................................... 86
Área de estudo....................................................................................................................................... 86
Amostragem........................................................................................................................................... 86
Tratamento dos dados........................................................................................................................... 87
Resultados.................................................................................................................................................. 89
Discussão ................................................................................................................................................... 96
Referências bibliográficas ........................................................................................................................ 101

ix
Anexo I...................................................................................................................................................... 106
CAPÍTULO 4- Estrutura, abundância e distribuição de espécies em onze fragmentos de floresta
estacional semidecídua na região da Área de Proteção Ambiental do município de Campinas, SP ....
....................................................................................................................................................................... 107
Resumo..................................................................................................................................................... 108
Introdução................................................................................................................................................. 109
Material e Métodos ................................................................................................................................... 111
Área de estudo..................................................................................................................................... 111
Amostragem......................................................................................................................................... 112
Estrutura............................................................................................................................................... 112
Análise dos dados................................................................................................................................ 113
Resultados................................................................................................................................................ 114
Padrões de abundância....................................................................................................................... 115
Valor de importância ............................................................................................................................ 122
Classes de diâmetro ............................................................................................................................ 129
Distribuição vertical de alturas............................................................................................................. 132
Discussão ................................................................................................................................................. 136
Referências bibliográficas ........................................................................................................................ 144
Anexo I...................................................................................................................................................... 151
CAPÍTULO 5 - Utilização de diferentes critérios de inclusão em inventários florísticos e de estrutura
de florestas estacionais semidecíduas e suas implicações sobre os resultados – um estudo de caso.
....................................................................................................................................................................... 165
Resumo..................................................................................................................................................... 166
Introdução................................................................................................................................................. 167
Material e Métodos ................................................................................................................................... 168
Área de estudo..................................................................................................................................... 168
Amostragem......................................................................................................................................... 168

x
Aspectos qualitativos .......................................................................................................................... 170
Tratamento dos dados......................................................................................................................... 170
Resultados................................................................................................................................................ 173
Composiçâo florística........................................................................................................................... 173
Diversidade .......................................................................................................................................... 176
Riqueza ................................................................................................................................................ 177
Similaridade ......................................................................................................................................... 179
Estrutura............................................................................................................................................... 182
Aspectos qualitativos ........................................................................................................................... 193
Discussão ................................................................................................................................................. 197
Referências bibliográficas ........................................................................................................................ 201
Anexo I...................................................................................................................................................... 204
Anexo II..................................................................................................................................................... 215

1
RESUMO GERAL
O processo de fragmentação de florestas tem sido freqüentemente relacionado a efeitos deletérios
sobre as comunidades bióticas. Tais efeitos seriam: perda de espécies, alterações de estrutura e
composição da vegetação, aumento da proporção de árvores mortas ou danificadas, maior ocorrência
de árvores infestadas por lianas. A magnitude de tais alterações tem sido associada principalmente
ao tamanho dos remanescentes florestais. Afirma-se que fragmentos pequenos apresentariam um
número menor de espécies, pois muitas não sobreviveriam em áreas reduzidas. Além disso, florestas
pequenas estariam mais expostas ao efeito de borda, devido sua alta razão borda:interior. Este
trabalho teve como principal objetivo investigar questões associadas à fragmentação de
ecossistemas através da análise da composição florística, estrutura da vegetação e análises de
aspectos qualitativos como estado de dossel e grau de infestação por lianas em 11 fragmentos
florestais na região da Área de Proteção Ambiental - APA - do município de Campinas, SP, (22°45’ a
23°00’ S, 47°00’ a 47°12’ O). Dez fragmentos têm área entre 10 e 65 hectares (ha) e um fragmento,
244,9ha. Os fragmentos com área inferior à 65ha foram agrupados em três classes de tamanho: 10-
15ha (pequenos), 16-25ha (médios) e 40-65ha (grandes). A amostragem foi através do método de
pontos quadrantes. Em cada um dos fragmentos foram amostrados 125 pontos quadrantes. No
fragmento de 244,9ha foram feitas três amostragens de 125 pontos em três áreas distintas. Em cada
ponto, foram incluídos os indivíduos arbóreos com DAP ≥ 10cm. Para avaliar os aspectos qualitativos
cada indivíduo amostrado foi categorizado em árvore do futuro (indivíduos jovens e/ou em
crescimento), árvore do presente (indivíduos adultos) ou árvore do passado (indivíduos danificados
ou morrendo). Os indivíduos foram classificados quanto ao grau de infestação de sua copa por lianas.
O índice de cobertura de dossel foi estimado através de fotografias hemisféricas. Com isso
pretendeu-se avaliar se a ocorrência de árvores danificadas ou morrendo (árvores do passado),
árvores muito infestadas por lianas e baixa cobertura de dossel estariam associadas ao tamanho dos
fragmentos. A análise da composição florística e estrutural dos fragmentos, sua diversidade e
similaridade permitiu investigar se fragmentos pequenos suportariam um menor número de espécies
do que fragmentos grandes e se apresentariam estrutura e composição diferente de fragmentos
grandes. Ao todo foram coletados 6500 indivíduos pertencentes a 248 espécies e 55 famílias. O
número de espécies variou de 47 a 110 entre as amostras. As análises de agrupamento mostraram
que fragmentos de mesmo tamanho podem ser tão heterogêneos entre si quanto áreas distintas
dentro de um mesmo fragmento, indicando que as variações na composição florística não estariam
relacionadas ao tamanho dos fragmentos. A maior parte das variações observadas em todos os
aspectos analisados não pareceu estar associada ao tamanho dos fragmentos. Dentre os fatores que
parecem estar influenciando as variações observadas estão a heterogeneidade ambiental e a
ocorrência de fatores de perturbação como queimadas. Todos os fragmentos pareceram exibir
alguma particularidade, seja do ponto de vista estrutural, florístico, de riqueza ou qualitativo que
ressalta a importância de sua preservação e de recuperação.

2
ABSTRACT The main purpose of this work is to deal with some questions referring to the ecosystem
fragmentation, which will be made by means of an analysis of the floristic composition, forest structure
and the distribution of tree species in eleven forest fragments located in Campinas (22°45’ to 23°00’ S,
47°00’ to 47°12’ O), state of São Paulo, SE Brazil. The study area comprises the Environmental
Protection Area of Sousas and Joaquim Egídio. Currently, forest fragmentation is said to be related to
deleterious effects on biotic communities. Such effects would be: species loss, changes in species
composition and structure, increasing rates of damaged or dead trees and a proliferation of lianas.
The extent of these alterations has been mainly associated to the size of the forest remnants. It has
been said that small fragments often have fewer species recorded for the same effort of observation
than large fragments or areas of continuous forest. In order to investigate a few of the questions raised
in this discussion, eleven fragments of the semideciduous forests, physionomically similar, were
chosen: ten of the fragments are between 10 and 65 hectares (ha) in area and a 244,9ha forest. The
fragments smaller than 65ha were grouped into three classes: 10-15ha (small), 16-25ha (medium) e
40-65ha (large). A tree community survey was performed using the point-centered quarter method.
For each sampling point, trees with a diameter at breast height (DBH) ≥ 10cm were included. In each
fragment, 125 sampling points were surveyed. In the >200ha fragment three surveys of 125 sampling
points were made, in three distinct areas. Each sampled tree was classified as trees of the future (or
potential trees, which are young and/or growing individuals), trees of the present (adult individuals) or
trees of the past (damaged or dying individuals). Moreover, it was recorded wether the canopy was
infested by lianas or vines for each sampled tree. A total of 248 species were found, distributed in 58
families. The number of species for each sample of 500 individuals ranged from 47 to 110 species.
Two samples showed a much lower richness: one of them, containing 47 species, was a small
fragment the other, containing 65, was a large one. The number of individuals infested by lianas was
also similar for most samples. However, in the smallest fragment, the percentage of infested trees was
equivalent to half of that shown in the other samples. It is important to stress that this fragment
revealed species richness similar to the others. In most samples the proportion of dead or damaged
individuals (trees of the past) varied between 30 and 40%. A single fragment, the biggest one,
presented twice the percentage observed in the other samples. Considering the analysed data, the
variation found among samples did not seem to be related to the size of the fragments. Indeed, other
factors may be associated to the observed variations, such as: history of the preservation, spatial
species distribution and preferences for specific environmental conditions. In addition, some species
occurred exclusively in one fragment. This fact associated with the differences of the species richness
among the samples, emphasizes the importance of preserving all the existent semideciduous forest
remnants, since a relevant portion of the biodiversity of this ecosystem might be spread through small
isolated fragments, such as these.

3
INTRODUÇÃO GERAL Fragmentação florestal é a substituição de amplas áreas de floresta nativa por outros
ecossistemas, deixando uma série de manchas remanescentes ou fragmentos de mata entremeadas
por uma matriz de vegetação diferenciada e/ou de usos diversos. Alteram-se assim os fluxos de
vento, radiação e água ao longo da paisagem. Todos os remanescentes de vegetação estão
expostos a essas mudanças, em maior ou menor grau, mas seus efeitos são modificados pelo
tamanho, forma e posição na paisagem de cada fragmento em particular (Saunders et al. 1991).
A interação entre estes dois ecossistemas adjacentes, o fragmento florestal e a matriz, se dá
através de uma transição abrupta, a região da borda do fragmento, e é denominada “efeito de borda”
(Murcia 1995). Nas bordas ocorreriam alterações bióticas e abióticas. A distância da penetração de
tais alterações em direção ao interior da mata variaria com a idade, orientação, fisionomia, matriz e
histórico do fragmento, da borda e da matriz (Murcia1995).
Dentre as mudanças abióticas estariam: aumento na penetração de luz, elevação da
temperatura e redução da umidade relativa do ar e do solo (Kapos 1989, Matlack 1993, Didhan &
Lawton 1999). Os efeitos bióticos seriam perda de diversidade (Diamond & May 1976, Harris 1984),
mudanças na composição (Tabarelli et al. 1999), alterações nos padrões de distribuição e
abundância dos organismos (Laurance et al. 2001, Willians-Linera 1990) e aumento da proporção de
árvores mortas ou danificadas, o que gerariam uma maior proporção de interrupções no dossel e
formação de clareiras (Laurance 1997, Ferreira & Laurance 1997).
A perda de espécies não se daria simplesmente por um efeito de redução de área ou perda de
heterogeneidade ambiental. Algumas espécies não persistiriam em fragmentos devido às alterações
das condições microclimáticas em relação à floresta contínua, resultantes do efeito de borda (Lovejoy
et al. 1996, Kapos et al. 1997). Outras se extinguiriam nos fragmentos devido a eventos genéticos ou
demográficos (Shafer 1981), perda de variabilidade genética (Lande 1988) e declínio da reprodução
devido a perda de polinizadores (Aizen & Feinsinger 1994).
A alteração da composição original ocorreria porque muitas espécies se beneficiariam das
condições do ambiente fragmentado. Espécies associadas às atividades antrópicas e espécies
características de ambientes perturbados, como lianas e espécies dos estádios sucessionais iniciais,
teriam suas capacidades de colonização e expansão aumentadas (Tabarelli et al. 1999, Laurance et
al. 2001). As espécies dispersas pelo vento (anemocóricas) também poderiam ser beneficiadas em
paisagens fragmentadas. Em ambientes mais abertos (matriz), suas chances de atingirem longas
distâncias são maiores (Howe & Smallwood 1982), enquanto para as espécies zoocóricas, a matriz,
muitas vezes, constitui um obstáculo intransponível. A penetração e/ou aumento excessivo da
abundância dessas espécies alteraria gradualmente a composição original das florestas
fragmentadas e também influenciaria a abundância e persistência de espécies da composição original
(Terborgh 1992).

4
O efeito de borda também estaria associado à alteração na dinâmica das populações em
fragmentos, pois ocasionaria aumento das taxas de mortalidade e recrutamento nas áreas de borda
(Laurance et al. 1998a, 1998b, Willians-Linera 1990), resultando em alterações nos padrões de
abundância das espécies florestais (Murcia 1995).
A magnitude de todas essas alterações tem sido associada, principalmente, ao tamanho dos
remanescentes florestais (Turner 1996). Fragmentos pequenos englobariam menor heterogeneidade
ambiental do que fragmentos grandes e, devido à sua área reduzida, suportariam um menor número
de espécies, as populações seriam menores e portanto, mais sujeitas à extinção (Hill & Curran 2001).
Além disso, fragmentos pequenos seriam mais afetados pelo efeito de borda devido à sua maior
razão borda:interior (Kapos 1989). Nessas áreas, a proporção de árvores mortas e danificadas seria
maior, a proporção de aberturas no dossel e ocorrência de clareiras aumentaria, haveria também uma
maior proporção de árvores infestadas por lianas e, com o aumento das taxas de mortalidade e
recrutamento a densidade de indivíduos seria aumentada.
Segundo o exposto acima, variações na diversidade, composição e estrutura de fragmentos
estariam associadas ao tamanho dos mesmos, embora não sejam apenas um efeito de redução de
área. Face a isso, seria esperado que se amostrássemos uma determinada área dentro de um
fragmento pequeno e a mesma área dentro de um fragmento grande, essas duas amostras, de
mesmo tamanho, deveriam apresentar diferenças quanto aos parâmetros analisados. Ou seja, 1ha
dentro de um fragmento pequeno apresenta, por exemplo, menos espécies do que 1ha dentro de um
fragmento grande? Segundo as proposições da literatura a resposta deveria ser afirmativa.
Fragmentos de mesmo tamanho estariam expostos de maneira similar ao efeito de borda, devido a
isso seria esperado que amostras feitas em fragmentos pequenos isolados ao longo da paisagem
deveriam ser mais homegêneos entre si do que amostras feitas em diferentes áreas de um fragmento
grande ou floresta contínua. Baseando-se nestes pressupostos, o objetivo deste trabalho foi verificar
se as variações de composição, diversidade, estrutura e de aspectos qualitativos como infestação por
lianas, proporção de árvores danificadas e porcentagem de abertura do dossel, estavam associadas
à área total de onze fragmentos de floresta estacional semidecídua.
O quarto capítulo tenta relacionar aspectos qualitativos, geralmente associados a fatores de
perturbação como grau de infestação por lianas, qualidade de dossel, abertura de dossel e histórico
de preservação, com o tamanho dos fragmentos. A principal questão investigada foi se fragmentos
menores apresentavam uma maior proporção de árvores danificadas, árvores infestadas por lianas e
maiores taxas de abertura de dossel do que fragmentos maiores.
O primeiro capítulo enfoca as variações de composição florística entre os fragmentos do estudo.
As principais questões abordadas são: como variam amostras de mesmo tamanho tomadas em
fragmentos de diferentes tamanhos e em regiões distintas de uma área contínua quanto à
composição florística? Fragmentos pequenos são mais semelhantes entre si, do ponto de vista
florístico, do que amostras tomadas em diferentes regiões de um fragmento grande?

5
O segundo capítulo trata dos aspectos da diversidade como número de espécies coletadas,
padrões de riqueza, similaridade e diversidade. Pretendeu-se responder às seguintes perguntas:
fragmentos grandes apresentam maior riqueza de espécies arbóreas do que fragmentos pequenos?
Fragmentos são mais similares entre si em termos de composição e abundância de espécies do que
áreas distintas dentro de um fragmento grande?
O terceiro capítulo aborda a estrutura da vegetação nos fragmentos, visando responder às
questões: amostragens feitas em diferentes pontos de uma área contínua são mais heterogêneas
entre si, do ponto de vista estrutural, do que com fragmentos isolados ao longo da paisagem? As
variações na estrutura da vegetação de fragmentos de tamanhos diversos poderiam ser atribuídas à
área total dos mesmos? Fragmentos menores apresentam maior densidade de indivíduos do que
fragmentos grandes ou uma área contínua?
Este trabalho está inserido dentro do Programa Biota/Fapesp, cujas principais metas são
promover uma melhor integração entre os estudos de diversidade realizados com os mais variados
organismos e gerar um banco de dados de biodiversidade. Visando atingir tais objetivos, a
padronização de metodologia entre os projetos que financia é uma de suas principais diretrizes.
Na época da inclusão do presente trabalho no Programa Biota/Fapesp havia uma ampla
discussão entre os projetos integrantes sobre qual critério de inclusão deveria ser adotado nos
estudos de florística e fitossociologia. Naquela ocasião, foi sugerido que todos os projetos adotassem
diâmetro à altura do peito (DAP) maior ou igual a cinco centímetros.
A falta de padronização metodológica entre estudos de florística e fitossociologia realizados não
só no estado de São Paulo, ou Brasil, mas também em outros lugares do mundo, é apontada por
muitos autores como uma dificuldade para a realização de comparações seguras que contribuiriam
muito para um melhor entendimento dos padrões observados (Ferreira & Prance 1998, Duringan et
al. 2000, Dislich et al. 2001). Dentre as metodologias empregadas um dos aspectos que mais variam
é o critério de inclusão. Segundo Dislich et al. (2001), até mesmo estudos realizados dentro de uma
mesma floresta, usando critérios de inclusão diferentes, levariam a resultados distintos. Os autores
chamaram a atenção para a necessidade de realização de estudos que explorem a influência da
variação dos critérios de inclusão adotados sobre os resultados obtidos.
É dentro desse contexto que se desenvolve o tema do quinto e último capítulo do presente
trabalho. Procuramos analisar as variações observadas quando se utilizam dois critérios de inclusão
diferentes. Analisamos todos os aspectos abordados nos capítulos anteriores, comparando dois
conjuntos de dados obtidos em cada fragmento, num deles o critério de inclusão foi DAP ≥ 5cm e no
outro DAP ≥ 10cm.

6
Referências bibliográficas AIZEN, M.A. & FEINSINGER, P. 1994. Forest fragmentation, pollination, and plant reproduction in a
chaco dry forest, Argentina. Ecology 75: 330-351.
DIAMOND, J.M. & MAY, R.M. 1976. Island biogeography and the design of natural reserves. Pp 163-
186. In: R.M. May (ed.) Theoretical ecology: principles and applications. Saunders.
Philadelfia.
DIDHAM, R.K. & LAWTON, J.H. 1999. Edge structure determines the magnitude of changes in
microclimate and vegetation structure in tropical forest fragments. Biotropica 31: 17-30.
DISLICH, R., CERSÓSIMO, L. & MANTOVANI, W. 2001. Análise da estrutura de fragmentos
florestais no planalto paulistano - SP. Revista Brasileira de Botânica 24: 321-332.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista Brasileira de Botânica 23: 371-383.
FERREIRA, L.V. & LAURANCE, W 1997. Effects of forest fragmentation on mortality and damage of
selected trees in Central Amazonia. Conservation Biology 11: 797-801.
FERREIRA, L.V. & PRANCE, G.T. 1998. Species richness and floristic composition in four hectares in
the Jaú National Park in upland forests in Central Amazonia. Biodiversity and Conservation 7:
1349-1364.
HARRIS, L.D. 1984. The fragmented forest: island biogeography theory and the preservation of biotic diversity. University of Chicago Press. Chicago.
HILL, J.L. & CURRAN, P.J. 2001. Species composicion in fragmented forests: conservation
implications of changing forest area. Applied Geography 21: 157-174.
HOWE, H.F. & SMALLWOOD, J. 1982. Ecology of seed dispersal. Annual Review of Ecology and Systematics 13: 201-228.
KAPOS, V. 1989. Effects of isolation on the water status of forest patches in the brazilian Amazon.
Journal of Tropical Ecology 5: 173-185
KAPOS, V., WANDELLI, E., CAMARGO, J.L. & GANADE, G. 1997. Edge-related changes in
environment and plant responses due to forest fragmentation in Central Amazonia. Pp 33-44. In:
W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: ecology, management, and conservation of forest communities. University of Chicago Press. Chicago.
LANDE, R. 1988. Genetics and demography in biological conservation. Science 242: 1455-1460.
LAURANCE, W.F. 1997. Hyper-disturbed parks: edge effects and the ecology of isolated rainforest
reserves in tropical Australia. Pp 71-83. In: W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: ecology, management, and conservation of forest communities. University
of Chicago Press. Chicago.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M. & LAURANCE, S.G. 1998a. Rain
forest fragmentation and the dynamics of amazoniam tree communities. Ecology 79: 2032-2040.

7
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M., LAURANCE, S.G., HUTCHINGS,
R.W. & LOVEJOY, T.E. 1998b. Effects of forest fragmentation on recruitment patterns in
amazonian tree communities. Conservation Biology 12: 460-464.
LAURANCE, W.F., PÉREZ-SALICRUP, D., DELAMÔNICA, P., FEARNSIDE, P.M., D’ANGELO, S.,
JEROLINSKI, A ., POHL, L. & LOVEJOY, T.E. 2001. Rain forest fragmentation and structure of
amazonian liana communities. Ecology 82: 105-116.
LOVEJOY, T.E., BIERREGAARD, R.O., RYLANDS , A.B., MALCOLM, J.R., QUINTELA, C.E.,
HARPER, L.H., BROWN, K.S., POWELL, A.H., POWELL, G.V.N., SCHUBART, H.O.R. & HAYS
.B. 1996. Edge and other effects of isolation on Amazon forest fragments. Pp 257-285. In: M.E.
Soule (ed.). Conservation Biology: the science of scarcity and diversity. Sinauer, Suderland.
USA
MATLACK, G.R. 1993. Microenvirnment variation within and among forest edge sites in the eastern
United States. Biological Conservation 66: 185-194.
MURCIA, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends in
Ecology and Evolution 10: 58-62.
SAUNDERS, D.A., HOBBS, R.J. & MARGULES, C.R. 1991. Biological consequences of ecossystem
fragmentation: a review. Conservation Biology 5: 18-32.
SHAFER, M.L. 1981. Minimum popuations sizes for species conservation. Bioscience 31: 131-134.
TABARELLI, M. & MANTOVANI, W. 1999. A riqueza de espécies arbóreas na floresta atlântica de
encosta no estado de São Paulo (Brasil). Revista brasileira de Botânica 22: 217-223.
TERBORGH, J., 1992. Maintenance of diversity in tropical forests. Biotropica 24: 283-292.
TURNER, I.M. 1996. Species loss in fragments of tropical rain forest: a review of evidence. Journal of Applied Ecology 33: 200-209.
WILLIANS-LINERA, G. 1990. Vegetation structure and enviromental conditions of forest edges in
Panama. Journal of Ecology 78: 356-373.

8
CAPÍTULO 1
AVALIAÇÃO DE ASPECTOS ASSOCIADOS AO ESTADO DE PRESERVAÇÃO DE ONZE FRAGMENTOS DE
FLORESTA ESTACIONAL SEMIDECÍDUA NA REGIÃO DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE
CAMPINAS, SP

9
Capítulo 1
Avaliação de aspectos associados ao estado de preservação de onze fragmentos de floresta estacional semidecídua na região da Área de Proteção Ambiental do município de Campinas,
SP
Karin dos Santos Pós-Graduação em Biologia Vegetal – Universidade Estadual de Campinas – Parte da tese de doutorado,
Luiza Sumiko Kinoshita Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Flavio A. M. dos Santos Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Resumo A fragmentação florestal tem sido associada a alterações deletérias sobre as comunidades florestais
remanescentes devido a mudanças bióticas e abióticas introduzidas pelo efeito de borda. Essas
alterações seriam mais drásticas em fragmentos menores devido à sua razão borda:interior. O
objetivo deste estudo foi avaliar se o estado de conservação de fragmentos varia com sua área total
O estudo foi desenvolvido na região da Área de Proteção Ambiental do município de Campinas, SP.
Foram escolhidos onze fragmentos de floresta estacional semidecidua. Dez com áreas inferiores a
100ha e um fragmento de 244,9 ha. Em cada fragmento foram amostrados 125 pontos quadrantes,
onde foram incluídos indivíduos arbóreos com DAP ≥ 10cm. No fragmento com área > 200ha foram
feitas 3 amostragens em locais diferentes da floresta. O estado de conservação foi avaliado, em cada
ponto, analisando-se: qualidade do dossel (avaliado através da categorização das árvores
amostradas em árvores do presente, árvores do passado e árvores do futuro); abertura de dossel
(avaliado através de fotografias hemisféricas) e grau de infestação por lianas. Todas as amostras
apresentaram proporção semelhante de árvores infestadas por lianas, com exceção de um fragmento
pequeno, onde a proporção foi a metade da exibida pelas outras áreas. Um fragmento grande
apresentou maior proporção de árvores danificadas (árvores do passado). As áreas ao longo da
floresta > 200ha, diferiram das demais, exibindo maiores proporções de árvores do futuro e menores
proporções de abertura total de dossel. A menor proporção de árvores do futuro em fragmentos
pequenos e as maiores aberturas de dossel nessas áreas pode estar indicando que a regeneração
possa ser reduzida, com riscos para a manutenção da estrutura e composição dessas matas e que a
estrutura do dossel é menos uniforme vertical ou horizontalmente nos fragmentos pequenos.

10
Introdução Fragmentação florestal é a substituição de amplas áreas de floresta nativa por outros
ecossistemas, deixando pequenos fragmentos de mata isolados e expostos a diferentes condições
dos ecossistemas que circundam o remanescente (a matriz). O resultado da interação entre esses
dois ecossistemas adjacentes, que estão separados por uma transição abrupta (a borda), é o
chamado “efeito de borda” (Murcia 1995).
Nas áreas de borda dos remanescentes, ocorreriam alterações abióticas e bióticas. Dentre as
abióticas podemos citar: aumento na penetração de luz e da temperatura e redução da umidade
relativa do ar e do solo (Kapos 1989, Matlack 1993, Didhan & Lawton 1999). Os efeitos bióticos
referem-se à mudanças na composição, distribuição e abundância dos organismos (Lovejoy et al.
1996, Willians-Linera 1990, Chen et al. 1992).
A elevação das taxas de mortalidade e de danos à estrutura das árvores, levando a mudanças
na comunidade florestal também tem sido associada ao efeito de borda (Laurance 1991, 1997,
Ferreira & Laurance 1997). A maior incidência de forças externas, como ventos e queimadas, nas
áreas de borda aumentariam as taxas de queda, quebra e morte de árvores (Putz & Milton 1982,
Chen et al. 1992). As taxas de mortalidade também aumentariam porque as plantas sofreriam maior
estresse hídrico, sendo que espécies mais susceptíveis ao ressecamento não sobreviveriam (Kapos
et al. 1997).
A morte de árvores cria interrupções no dossel e, quanto menor a superposição de copas, maior
a incidência de luz nos estratos inferiores (Duringan et al. 2000). Algumas plantas beneficiam-se
dessa situação como lianas e espécies dos estádios iniciais de sucessão (Turner & Tan 1992,
Malcolm 1994, Tabarelli et al. 1999).
Fragmentos freqüentemente apresentam uma proliferação excessiva de lianas (Laurance 1991,
1997, Oliveira-Filho et al. 1997, Tabanez et al. 1997, Viana et al. 1997, Laurance et al. 2001). A
maioria delas necessita de muita luz para crescer (Putz 1984) e são, em sua maioria, anemocóricas,
beneficiando-se de ambientes mais abertos (Laurance 1991). Além disso, sua taxa de crescimento é
bem maior que a de árvores, pois investem menos em tecidos de sustentação (Peñalosa 1985), o que
seria uma vantagem em bordas e clareiras recém-formadas.
Em relação às espécies arbóreas, tem sido mostrado que as lianas, além de competirem por
luz, umidade e nutrientes do solo - o que contribuiria para a redução do crescimento e reprodução de
árvores (Putz 1984, Whigham 1984, Stevens 1987, Pérez-Salicrup 1998) – causariam estresse
mecânico sobre a estrutura da árvore hospedeira, ocasionando quebra ou queda, principalmente no
caso de árvores jovens (Lowe & Walker 1977, Putz 1980, 1984). Foi também observado que, em
algumas clareiras, as lianas chegam a formar um tapete de até um metro de altura, compondo
emaranhados que impossibilitam qualquer regeneração de árvores ou arvoretas (Tabanez et al.
1997). Assim, de acordo com Putz (1980), a proliferação excessiva de lianas poderia retardar a

11
regeneração das espécies arbóreas por anos ou décadas ou mesmo impedi-la (Nicholson 1958,
Dawkins 1961).
Devido a esses fatores, as lianas têm sido consideradas um problema para a conservação da
biodiversidade de florestas naturais, que atualmente depende da preservação e manejo de pequenos
remanescentes isolados da vegetação (Saunders et al. 1991).
As condições microclimáticas no interior de fragmentos tendem a ser mais amenas do que nas
bordas (Kapos 1989), entretanto, a distância da penetração das alterações associadas ao efeito de
borda varia com a idade, orientação, fisionomia, matriz e histórico do fragmento, da borda e da matriz
(Murcia 1995). Segundo Kapos (1989), essas alterações podem ser detectadas a até 40 ou 60 metros
de distância da margem do fragmento, mas, segundo Laurance et al. (1998a), podem atingir
distâncias de até 300 metros, embora sejam mais facilmente detectáveis a até 100 metros da borda.
O aumento da mortalidade de árvores se estenderia portanto, a regiões distantes da borda, o que
elevaria as taxas de abertura de dossel e a proporção de áreas ocupadas por clareiras. De qualquer
forma, fica evidente que fragmentos pequenos, cuja razão borda:interior é maior, seriam mais
afetados do que fragmentos grandes, pois o efeito de borda poderia influenciar quase todo o
fragmento (Kapos 1989). Para Lovejoy et al. (1996), este seria o caso de fragmentos de até 10ha.
Face ao exposto, fragmentos menores deveriam apresentar uma maior proporção de árvores
danificadas, árvores infestadas por lianas e maiores taxas de abertura de dossel do que fragmentos
maiores. O presente trabalho pretende investigar se estas proposições se aplicam aos fragmentos de
floresta estacional semidecídua na região da Área de Proteção Ambiental - APA - do município de
Campinas, SP.

12
Material e métodos Área de estudo
O estudo foi realizado predominantemente na região da Área de Proteção Ambiental - APA - do
município de Campinas, SP (22°45’ a 23°00’ S, 47°00’ a 47°12’ W) e no município de Itatiba, SP
(23°10’ a 23°15’ S, 45°45’ a 46°45’ W) em áreas fronteiriças à APA. A APA localiza-se na porção
nordeste do município de Campinas e está compreendida entre os rios Atibaia, que determina sua
fronteira à oeste, e Jaguari, que sinaliza a fronteira a leste da APA e também do município de
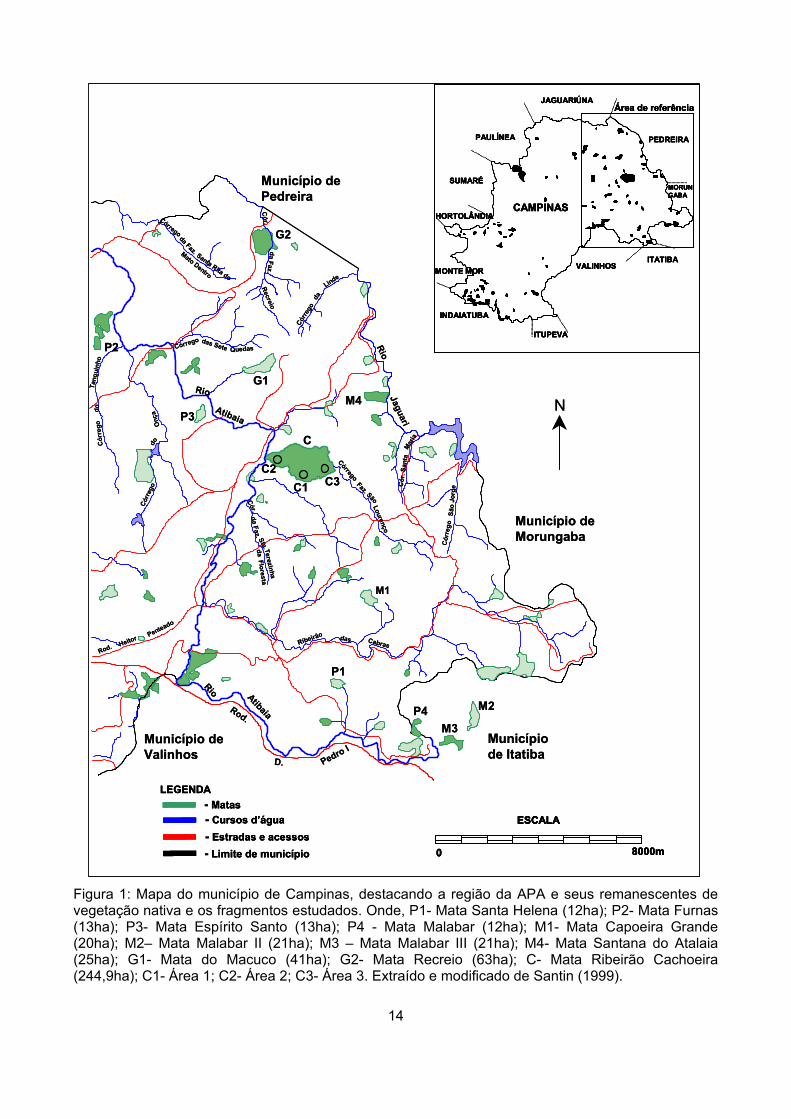
Campinas (Figura 1). Foram escolhidos dez fragmentos com áreas entre 12 e 63 hectares (ha) e um
fragmento com área de 244,9 ha (Figura 1). Os fragmentos menores do que 65ha foram
categorizados em três classes de tamanho: sendo quatro pequenos, com área entre 12 e 13,5ha (P1,
P2, P3 e P4), quatro médios, com área entre 19 e 25,5ha (M1, M2, M3 e M4) e dois grandes, G1 e G2
com áreas de 41,8 e 63,3ha, respectivamente.
O clima da região, segundo Köppen (1948), é Cwag'- temperado, moderadamente chuvoso, de
inverno seco não rigoroso, com precipitação média anual de 1409 mm (Mello et al. 1994). Os solos
que ocorrem na área de estudo são Podzólico Vermelho-Amarelo e Latossolo Vermelho. Os relevos
podem ser suave ondulados, forte ondulados e montanhosos ou ondulados e suave ondulados
(Oliveira et al. 1999). A vegetação dos fragmentos pode ser classificada como floresta estacional
semidecídua, nomenclatura proposta por Veloso et al. (1991). A matriz em que os fragmentos estão
inseridos constitui-se de um mosaico de culturas anuais e perenes, pastagens, reflorestamentos e
pequenos assentamentos humanos. A agricultura é caracterizada por pequenas áreas de café,
laranja, cana de açúcar e culturas de subsistência. Os reflorestamentos com eucalipto e pastagens
ocupam grandes extensões (Miranda 1996) (Tabela 1).
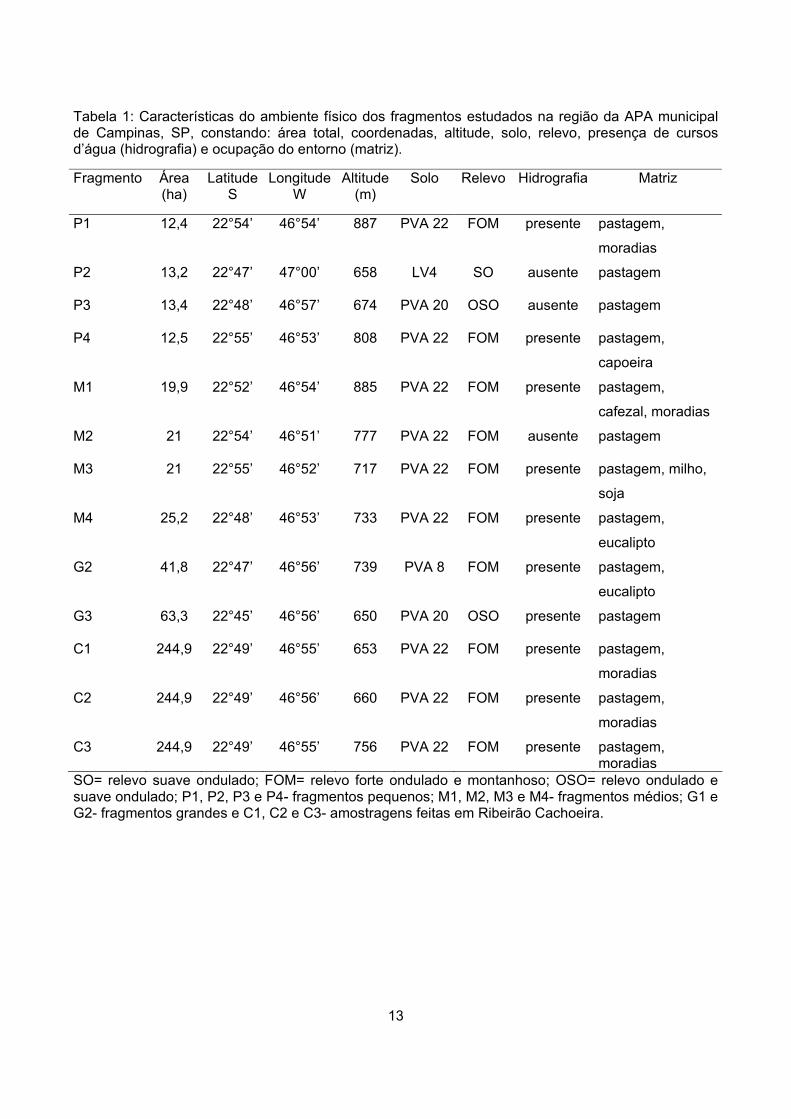
13
Tabela 1: Características do ambiente físico dos fragmentos estudados na região da APA municipal de Campinas, SP, constando: área total, coordenadas, altitude, solo, relevo, presença de cursos d’água (hidrografia) e ocupação do entorno (matriz). Fragmento Área
(ha) Latitude
S Longitude
W Altitude
(m) Solo Relevo Hidrografia Matriz
P1 12,4 22°54’ 46°54’ 887 PVA 22 FOM presente pastagem,
moradias
P2 13,2 22°47’ 47°00’ 658 LV4 SO ausente pastagem
P3 13,4 22°48’ 46°57’ 674 PVA 20 OSO ausente pastagem
P4 12,5 22°55’ 46°53’ 808 PVA 22 FOM presente pastagem,
capoeira
M1 19,9 22°52’ 46°54’ 885 PVA 22 FOM presente pastagem,
cafezal, moradias
M2 21 22°54’ 46°51’ 777 PVA 22 FOM ausente pastagem
M3 21 22°55’ 46°52’ 717 PVA 22 FOM presente pastagem, milho,
soja
M4 25,2 22°48’ 46°53’ 733 PVA 22 FOM presente pastagem,
eucalipto
G2 41,8 22°47’ 46°56’ 739 PVA 8 FOM presente pastagem,
eucalipto
G3 63,3 22°45’ 46°56’ 650 PVA 20 OSO presente pastagem
C1 244,9 22°49’ 46°55’ 653 PVA 22 FOM presente pastagem,
moradias
C2 244,9 22°49’ 46°56’ 660 PVA 22 FOM presente pastagem,
moradias
C3 244,9 22°49’ 46°55’ 756 PVA 22 FOM presente pastagem, moradias
SO= relevo suave ondulado; FOM= relevo forte ondulado e montanhoso; OSO= relevo ondulado e suave ondulado; P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
14
Figura 1: Mapa do município de Campinas, destacando a região da APA e seus remanescentes de vegetação nativa e os fragmentos estudados. Onde, P1- Mata Santa Helena (12ha); P2- Mata Furnas (13ha); P3- Mata Espírito Santo (13ha); P4 - Mata Malabar (12ha); M1- Mata Capoeira Grande (20ha); M2– Mata Malabar II (21ha); M3 – Mata Malabar III (21ha); M4- Mata Santana do Atalaia (25ha); G1- Mata do Macuco (41ha); G2- Mata Recreio (63ha); C- Mata Ribeirão Cachoeira (244,9ha); C1- Área 1; C2- Área 2; C3- Área 3. Extraído e modificado de Santin (1999).
Município de Morungaba
Município de Pedreira
Município de Valinhos
ESCALA
0 8000m
LEGENDA- Matas- Cursos d’água- Estradas e acessos- Limite de município
N
Rio
CórregoFaz. São
Lourenço
Cór
rego
São
Jorg
e
RioJaguari
Cór
. San
taM
aria
Ribeirão das Cabras
Córre
goda
Linde
RecreioC
ór.da Faz.
Córregoda Faz. Santa Rita do
Mato Dentro
Córrego das Sete Quedas
Rio
Atibaia
Córr
ego
doO
nça
Cór
rego
doTa
nqui
nho
Cór. da Faz. Sta. Terezinha
Florestada
Atibaia
Pedro I
Rod.
D.
Rod. Heitor Penteado
M4
C
C3C1C2
G1
P3
G2
P2
M1
P1
P4M3
M2
HORTOLÂNDIA
MONTE MOR
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
Município de Itatiba
Município de Morungaba
Município de Pedreira
Município de Valinhos
ESCALA
0 8000m
ESCALAESCALA
0 8000m0 8000m
LEGENDA- Matas- Cursos d’água- Estradas e acessos- Limite de município
LEGENDA- Matas- Cursos d’água- Estradas e acessos- Limite de município
N
Rio
CórregoFaz. São
Lourenço
Cór
rego
São
Jorg
e
RioJaguari
Cór
. San
taM
aria
Ribeirão das Cabras
Córre
goda
Linde
RecreioC
ór.da Faz.
Córregoda Faz. Santa Rita do
Mato Dentro
Córrego das Sete Quedas
Rio
Atibaia
Córr
ego
doO
nça
Cór
rego
doTa
nqui
nho
Cór. da Faz. Sta. Terezinha
Florestada
Atibaia
Pedro I
Rod.
D.
Rod. Heitor Penteado
M4
C
C3C1C2
G1
P3
G2
P2
M1
P1
P4M3
M2
HORTOLÂNDIA
MONTE MOR
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
HORTOLÂNDIA
MONTE MOR
HORTOLÂNDIA
MONTE MOR
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
Município de Itatiba

15
Amostragem
Para a amostragem da comunidade arbórea foi adotado o método de pontos quadrantes
proposto por Cottam & Curtis (1956). Os pontos foram dispostos em intervalos regulares de 15m ao
longo de transecções que percorriam área semelhante em fragmentos grandes e pequenos, evitando
apenas áreas de bordas e clareiras. As amostragens foram feitas de forma a incluir áreas
semelhantes, tanto do ponto de vista da fisionomia da vegetação, área percorrida e ambiente físico
(condições de altitude, solo e drenagem).
Em cada fragmento <65ha foram amostrados 125 pontos quadrantes, onde foram incluídos
indivíduos arbóreos com diâmetro à altura do peito (DAP) maior ou igual a 10cm, totalizando 500
indivíduos por fragmento. Devido à inexistência de três fragmentos maiores que 200ha como é o caso
da mata Ribeirão Cachoeira, foram feitas três amostragens de 125 pontos em três áreas diferentes
desta mata. Nos referiremos a estas amostras, a partir daqui, como amostras C1, C2 e C3. Com isso
pretendia-se verificar se áreas distintas dentro de um fragmento grande seriam mais ou menos
similares entre si do que com os fragmentos isolados ao longo da paisagem. Pretendia-se também
avaliar as variações no estado de conservação entre fragmentos de diversos tamanhos.
Aspectos qualitativos
Histórico dos fragmentos
Para remontar à história de cada fragmento estudado, foram consultadas fotografias aéreas da
região, realizadas desde a década de 1960 (Instituto Agronômico; Instituto Base – Aerofotometria e
Projetos/SA). Foram consultados também, os trabalhos de Santin (1999), Miranda (1996) e Mattos
(1996), que fornecem algumas informações sobre o histórico da região do estudo e ocorrência de
fatores de perturbação em alguns fragmentos. Além disso, foram feitas observações de campo para
tentar identificar sinais da ocorrência, atual ou recente, de fatores de perturbação como penetração
de gado, extração de madeira, ocorrência de queimadas, atividades de caça. Os proprietários dos
fragmentos, bem como moradores da região foram entrevistados sobre a ocorrência de tais fatores na
região e nos fragmentos do estudo. Entretanto, as informações obtidas nas entrevistas foram
utilizadas somente para conferir ou reforçar as observações de campo e informações colhidas na
literatura como mencionado acima, pois havia a possibilidade de proprietários ou funcionários não
serem sinceros em suas respostas, principalmente em relação a fatores como extração seletiva de
madeira, caça e fogo proposital.
Qualidade do dossel
A avaliação da qualidade do dossel foi feita através da categorização das árvores amostradas
em: árvores do futuro, árvores do presente e árvores do passado, segundo as definições propostas
por Oldeman (1989), onde: a) árvores do futuro (AF) seriam indivíduos jovens, que teriam potencial
para crescer em altura e sofrer expansão de copa; b) árvores do presente (APR) seriam indivíduos

16
adultos, mas que ainda teriam potencial para extensão e expansão e, mesmo que seu tamanho fosse
máximo em um determinado local, sua sustentabilidade seria aparentemente durável devido à sua
capacidade de repor partes perdidas; c) árvores do passado (APS) seriam aquelas muito danificadas,
ou seja, com vários galhos quebrados, árvores morrendo, em decadência ou uma combinação de
todos esses estados. Entretanto, não foi possível a categorização de alguns indivíduos em algumas
amostras, por isso o número total de indivíduos nesta análise difere entre as amostras. De acordo
com as proposições da literatura, mencionadas na introdução, seria esperado que fragmentos
pequenos tivessem maiores proporções de árvores danificadas (APS) e também de árvores do futuro
do que fragmentos grandes ou as amostras de Ribeirão Cachoeira.
Abertura de dossel
A porcentagem de abertura de dossel foi obtida através da análise de fotografias hemisféricas.
As fotografias foram obtidas utilizando-se câmera Digital Nikon Coolpix 950 e lente olho de peixe –
autofocus Nikon 8mm com ângulo de 180°. A câmera foi fixada sobre um tripé com a lente voltada
para o dossel a cerca de 1 metro de altura do solo, alinhada com auxílio de um nível e orientada para
o norte magnético com uma bússola.
As fotos foram obtidas nos pontos de amostragem dos fragmentos. Para se evitar sobreposição
de imagens, as fotos foram realizadas em pontos alternados, até que se totalizasse 50 fotos em cada
área. O mesmo procedimento foi feito em duas estações do ano: estação chuvosa (janeiro-fevereiro)
e estação seca (julho-agosto), durante o ano de 2002. Nos fragmentos M1, M2 e M4, essas imagens
não puderam ser obtidas devido à impossibilidade de retorno aos pontos de amostragem em ambos
os períodos mencionados. Na mata M1 os pontos foram retirados por terceiros, a mata M4 foi atingida
por uma forte queimada, destruindo os pontos e, o proprietário da mata M2 proibiu o acesso à área.
Para evitar problemas de reflexo dos raios solares pela folhagem ou diretamente na lente, as
fotografias foram feitas em dias de céu encoberto por nuvens, seguindo sugestão de Whitmore et al.
(1993), ou nos horários de 6 às 10 horas e de 16 às 18 horas.
As imagens digitais foram analisadas, utilizando-se o programa Gap Light Analyser versão 2.0
(Frazer et al. 1999), que converte os pontos escuros da foto em preto e os pontos claros em branco.
Os pontos pretos são interpretados como dossel e os pontos brancos, como aberturas de dossel
(figura 2). O programa analisa fotos coloridas e converte tonalidades escuras em pontos pretos e
tonalidades claras em pontos brancos, sendo possível regular o limiar de tonalidade, expresso
numericamente, acima do qual as imagens serão convertidas em pontos pretos pelo programa. Como
isso envolve uma certa subjetividade, que poderia influenciar nos resultados, todas as fotos foram
analisadas por três pessoas diferentes. Cada pessoa estabeleceu um limiar de tonalidade para cada
foto, em seguida foi calculada a média desses valores. O valor médio calculado para cada foto foi
adotado para a transformação das fotos em preto e branco na análise da abertura do dossel.

17
Figura 2: Comparação entre uma fotografia hemisférica (A) e a mesma imagem digitalizada e transformada em preto e branco (B)
A
B
A
B

18
O cálculo da abertura de dossel da imagem fotográfica é feito através da relação entre o
somatório dos pixels classificados como abertura dentro do hemisfério total da foto. Assim, cada
ponto quadrante fotografado teve um valor de abertura de dossel, ou seja, uma porcentagem de
espaços abertos no hemisfério total da fotografia obtida de seu dossel. Este valor tende a aumentar
de fotografias obtidas sob dossel fechado para fotografias obtidas em clareiras.
Segundo as proposições da literatura seria esperado que fragmentos pequenos, mais expostos
à perturbações e ao ressecamento, apresentassem maiores porcentagens de abertura de dossel e
também maior amplitude de variação entre as duas estações do ano.
Infestação por lianas
Para cada indivíduo arbóreo amostrado foi estimada a porcentagem de sua copa que se
encontrava infestada por lianas. Foram utilizadas cinco classes de porcentagem de infestação (0, 1-
25%, 26-50%, 51-75% e 76 a 100%). Neste trabalho, foi considerada liana qualquer planta que
mantivesse contato com o solo e necessitasse de um suporte para seu desenvolvimento (Müller-
Dombois & Ellenberg 1974). No caso da infestação por lianas, seria esperado que fragmentos
menores apresentassem uma maior proporção de árvores infestadas por lianas do que fragmentos
maiores e áreas de Ribeirão Cachoeira.
Análise dos dados
Para avaliar se a proporção de árvores categorizadas em árvores do presente, passado e futuro
diferia entre os diferentes fragmentos, foi construída uma tabela de contingência do número de
indivíduos em cada categoria. Foram feitas comparações, através do teste de Qui-quadrado com
correção de Yates (Zar 1989), entre os número esperados, obtidos na tabela de contingência, e
observados em cada categoria nos diferentes fragmentos. O mesmo procedimento foi realizado para
verificar se havia diferença entre a proporção de árvores infestadas por lianas nos diversos
fragmentos. Os indivíduos foram separados em infestados (por lianas) ou não infestados,
independentemente do grau de infestação de cada indivíduo. Isto foi feito para facilitar a
apresentação e análise dos resultados.
Para verificar se a proporção de indivíduos infestados por lianas, bem como a proporção de
árvores categorizadas como AF, APR e APS, estava relacionada ao tamanho dos fragmentos, foi feita
uma análise de correlação de Pearson, utilizando o programa BioEstat (Ayres et al. 1998).
Para verificar se havia diferenças no total de aberturas do dossel entre as áreas, as
porcentagens de abertura de dossel obtidas em cada fragmento foram comparadas através da
análise de Kruskall-Wallis. Optamos por um teste não-paramétrico porque os dados se mostraram
heterocedásticos através do teste de Bartlett e não apresentaram distribuição normal. A análise de
correlação de Pearson foi utilizada para verificar se havia correlação entre a área dos fragmentos e a
porcentagem de abertura nas duas estações do ano.

19
Para verificar se havia relação entre as variações da abertura de dossel entre as duas estações
do ano e a proporção de plantas decíduas em cada área, foi calculada a proporção de plantas
decíduas, semidecíduas ou perenifólias de cada fragmento. A classificação das plantas foi feita
através de informações da literatura. Foi feita uma tabela de contingência do número de indivíduos
em cada categria e os resultados foram comparados através do teste de qui-quadrado com correção
de Yates. As relações entre os dois parâmetros (variação na abertura e proporção de indivíduos
decíduos ou semidecíduos) foi analisada através da análise de correlação de Pearson. Seria
esperado que em áreas onde houvesse predominância de árvores perenifólias, a variação de
abertura de dossel entre as duas estações fosse menor.
Quando necessário, valores em porcentagem foram convertidos em arcoseno da raiz quadrada.
Nas análises de correlação consideramos apenas um valor médio das amostras de Ribeirão
Cachoeira, pois se tratava de três amostras realizadas em um mesmo fragmento.

20
Resultados Histórico dos fragmentos
É muito difícil precisar exatamente quando e em que velocidade as florestas da região da área
de estudo foram substituídas pela agricultura. Segundo Mattos (1996), a penetração humana na
região de Campinas foi iniciada no século XVIII e foi sempre predatória. Em 1796 a população era de
2100 habitantes e se concentrava nas zonas norte e leste de Campinas, englobando a área hoje
delimitada pela APA (Miranda 1996). A cultura de cana de açúcar teve início nesta época e foi
bastante próspera até 1838, época em que as plantações de café já ocorriam em grande extensão
(Santin 1999, Miranda 1996). Entretanto, em 1818, o naturalista Saint-Hilaire, ao passar pela região,
registrou que “a cidade de Campinas era totalmente rodeada de matas” (Saint-Hilaire 1976 apud
Mattos 1996).
Através das fotografias aéreas, observamos que a maioria dos fragmentos mantém sua forma e
tamanho atuais há pelo menos 40 anos, com exceção do fragmento P4, que vem aumentando sua
área total nos últimos anos. Segundo informações dos proprietários, essa área é uma floresta
secundária que se desenvolveu numa área antes utilizada para pastagens.
Dentre os fatores de perturbação analizados, a penetração de gado parece ser o mais comum
(Tabela 2), tendo sido observada na maioria das áreas do estudo. Apesar de freqüente, a maioria dos
proprietários expressam o desejo de acabar com essa prática, pois ela dificulta o controle do rebanho.
Entretanto, a instalação e manutenção de cercas é muito onerosa, segundo eles.
A ocorrência de fogo na região também parece ser freqüente, segundo relatos de habitantes
locais e demais informações obtidas. Várias áreas do estudo já foram atingidas total ou parcialmente
(Tabela 2). Apesar disso, sinais evidentes da passagem de fogo pelas matas estudadas foram
observados somente no fragmento G2 e área C3 de Ribeirão Cachoeira. Os proprietários da mata G2
relataram a ocorrência de uma grande queimada em 1996, que atingiu a área quase por completo.
Moradores próximos à área C3 de Ribeirão Cachoeira e da mata G1 também relataram que essas
áreas foram parcialmente atingidas por queimadas em 1994. As queimadas no entorno da mata P2
parecem ser freqüentes, como observado em campo, e os proprietários admitem que o fogo já atingiu
a mata parcialmente, mas não conseguiram precisar o ano. Segundo Santin (1999), o fogo também já
atingiu parcialmente a área M1. Além disso, a mata M4 foi completamente atingida por uma queimada
logo após o término dos trabalhos de campo deste estudo.
Em nenhuma das áreas foram observados sinais evidentes de caça recente ou extração de
madeira. A maioria dos proprietários nega a realização dessas práticas atualmente e também a
introdução de espécies exóticas nas matas. Apenas o proprietário do fragmento P1 admitiu retirada
de madeira no passado (Tabela 2). O mesmo proprietário relatou que, no passado, foram realizadas
atividades de controle de lianas na área. Essas plantas foram extraídas por vários anos e, segundo
ele, essa prática auxiliou no desenvolvimento da mata.

21
Os principais usos a que essas áreas são ou foram submetidos, segundo os proprietários e
observações de campo, são: coleta de plantas medicinais; produção de mel, atividade observada nas
matas P3, M3 e M4; captação de água, observada na maioria das áreas com recursos hídricos como
as matas P1, P4, M1, M3, G1, G2 e C1 de Ribeirão Cachoeira. A extração de mudas de espécies
nativas do interior da mata é praticada por um morador vizinho à mata G1, com a finalidade de
atividades de reflorestamento de áreas adjacentes à mata. Algumas áreas, como as matas P1, M4,
G1 e a mata Ribeirão Cachoeira são utilizadas para atividades de lazer e de educação ambiental.
A maior parte dos entrevistados considera que a região sofreu poucas alterações em relação à
hidrografia e à infraestrutura local, mas relatam que a região tem sofrido uma forte pressão imobiliária
nos últimos anos e corre sério risco com a expansão dos limites urbanos. Durante a realização desse
trabalho, por exemplo, a fazenda a que pertencia o fragmento M1, foi vendida para o estabelecimento
de um condomínio rural. Apesar disso, os proprietários das áreas P1, P4 e M3 acham que as matas
de sua propriedade aumentaram em área e exibem um aspecto mais “exuberante” nos últimos dez
anos.
O desejo de preservação dos fragmentos é demostrado pela maioria dos moradores e
proprietários, que reconhecem seu valor paisagístico e para a preservação da fauna. Os animais mais
citados por eles, e cuja sobrevivência gostariam de assegurar, são tucanos e aves em geral, bugios,
micos-estrela, lontras, capivaras e veados campeiros. Entretanto, muitos se queixam da falta de apoio
das autoridades competentes no controle de queimadas e da falta de incentivos fiscais para a
preservação dessas áreas.

22
Tabela 2: Características do histórico dos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: 0= não ocorrência de queimadas nos últimos dez anos; 1=ocorrência de queimadas no entorno da mata; 2= queimadas atingiram parcialmente a mata; 3= queimadas atingiram toda a extensão da mata; - = sem informações. Locais Idade Penetração de
gado* Extração de
madeira Ocorrência de
fogo*
P1 ≥40 sim sim 0
P2 ≥40 sim não 1/2
P3 ≥40 sim não -
P4 <30 sim não 0
M1 ≥40 não não 2
M2 ≥40 sim não 0
M3 ≥40 sim não 0
M4 ≥40 não não 3**
G1 ≥40 sim não 2
G2 ≥40 não não 3
C1 ≥40 não não 0
C2 ≥40 não não 0
C3 ≥40 sim não 2
P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira. * informações referentes aos últimos 10 anos. ** ocorrência de queimada após a finalização da coleta de dados.

23
Qualidade do dossel
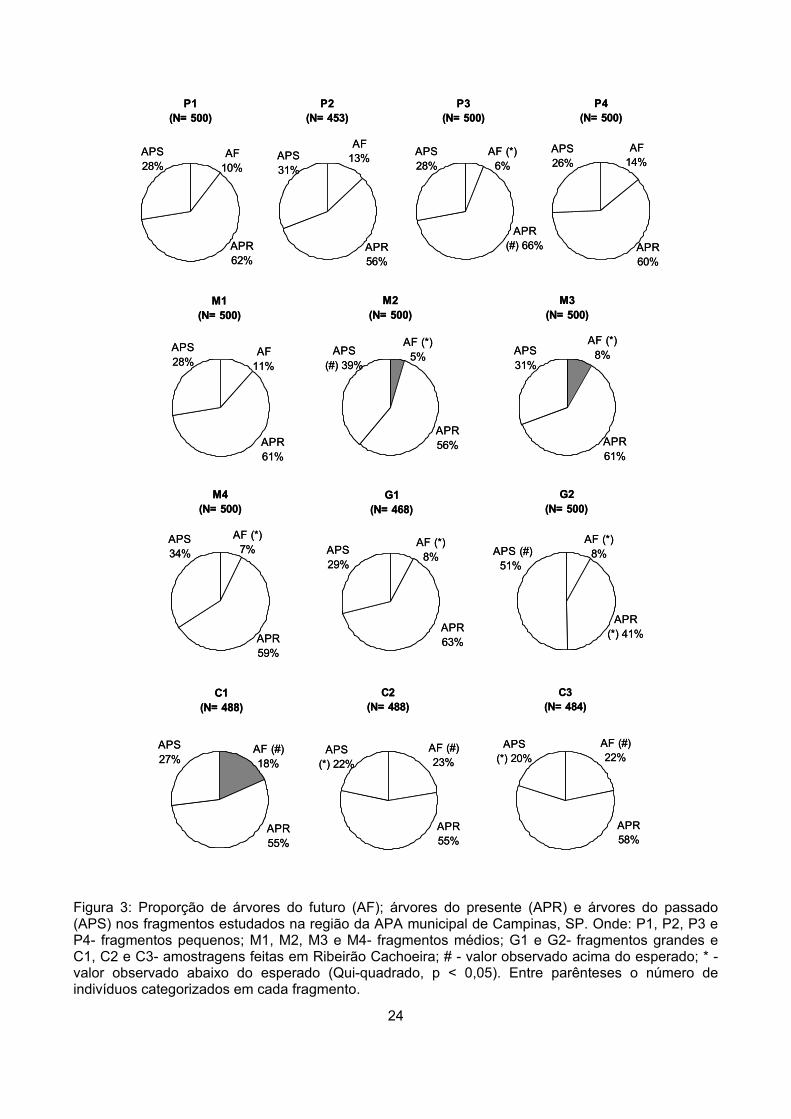
Considerando todos os indivíduos categorizados de todas as amostras temos 58% de árvores
do presente (APR), 30% de árvores do passado (APS) e 12% de árvores do futuro (AF). A proporção
de APR foi maior do que as proporções de APS e AF em todos os fragmentos, exceto em G2. Em
todos os fragmentos <100ha as proporções de APS foram o dobro, o triplo ou até o quíntuplo da
proporção de AF, indicando um desequilíbrio entre a proporção de indivíduos que estariam deixando
o sistema (APS) e de indivíduos que estariam entrando no sistema (AF) (Figura 3). Nas áreas de
Ribeirão Cachoeira – C1, C2 e C3 - as proporções de APS e de AF foram bastante semelhantes,
indicando um maior equilíbrio entre a proporção de plantas que estariam deixando o sistema e que
estariam penetrando no sistema (Figura 3).
A proporção de APR diferiu entre os fragmentos (χ2 = 40,2, gl = 12, p < 0,0001), variando de 41
a 66%. O fragmento P3 apresentou proporção superior de APR (χ2 = 6,4, gl = 1, p = 0,01), enquanto
em G2 essa proporção foi significativamente inferior (χ2 = 21,5, gl = 1, p < 0,001) (Figura 3).
A proporção de APS também diferiu entre as áreas (χ2 = 127,4, gl = 12, p < 0,0001) e oscilou
entre 20 e 51%. Nos fragmentos G2 e M2 essa proporção foi significativamente superior (G2- χ2 =
72,9, gl = 1, p < 0,0001; M2- χ2 = 14,9, gl = 1, p < 0,005), enquanto nas amostras C2 e C3 foi inferior
às demais (C2- χ2 = 11,4, gl = 1, p = 0,001; C3- χ2 = 16,9, gl = 1, p < 0,0001) (Figura 3).
As maiores variações, entretanto, ocorreram na proporção de AF, que também diferiu entre os
fragmentos (χ2 = 165,3, gl = 12, p < 0,0001). Observando as proporções obtidas seria possível
separar as amostras em três grupos. O primeiro, onde a proporção de AF variou entre 5 e 8%,
reuniria o fragmento pequeno P3, os fragmentos médios M2, M3 e M4, os fragmentos grandes G1 e
G2. Em todas essas amostras a proporção de AF foi significativamente inferior às demais (P3- χ2 =
12,0, gl = 1, p = 0,001; M2- χ2 = 20,4, gl = 1, p < 0,0001; M3- χ2 = 5,2, gl = 1, p < 0,05; M4- χ2 = 8,7, gl
= 1, p < 0,005; G1- χ2 = 8,7, gl = 1, p < 0,005; G2- χ2 = 5,2, gl = 1, p < 0,05). O segundo grupo seria
composto pelos fragmentos pequenos, P1 e P2 e pelos médios M1 e P4, onde a proporção de AF
oscilou entre 10 e 14%, entretanto a proporção de AF nessas áreas não difere significativamente das
demais. No terceiro grupo estariam somente as amostras de Ribeirão Cachoeira (C1, C2, C3) que
apresentaram proporções entre 18% e 23%. Essas amostras apresentaram proporções superiores às
demais amostras do estudo – C1 (χ2 = 17,2, gl = 1, p < 0,0001), C2 (χ2 = 46,0, gl = 1, p < 0,0001) e
C3 (χ2 = 39,1, gl = 1, p < 0,0001) (Figura 3).
Apesar da grande variação entre os fragmentos quanto a essas proporções, apenas a
proporção de AF apresentou correlação com a área dos fragmentos (r = 0,668, p = 0,02).
24
Figura 3: Proporção de árvores do futuro (AF); árvores do presente (APR) e árvores do passado (APS) nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira; # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05). Entre parênteses o número de indivíduos categorizados em cada fragmento.
M2(N= 500)
APR56%
AF (*) 5%APS
(#) 39%
G1(N= 468)
APS29%
APR63%
AF (*)8%
P1(N= 500)
APS28%
APR62%
AF10%
P2 (N= 453)
APS31%
APR56%
AF13%
P3(N= 500)
APS28%
APR(#) 66%
AF (*)6%
P4(N= 500)
AF14%
APS26%
APR60%
M1(N= 500)
APS28%
APR61%
AF11%
M3(N= 500)
APS31%
APR61%
AF (*)8%
M4(N= 500)
APR59%
APS34%
AF (*)7%
G2(N= 500)
APR(*) 41%
APS (#) 51%
AF (*)8%
C1(N= 488)
APS27%
APR55%
AF (#)18%
C2(N= 488)
APS(*) 22%
AF (#)23%
APR55%
C3(N= 484)
APS(*) 20%
AF (#)22%
APR58%
M2(N= 500)
APR56%
AF (*) 5%APS
(#) 39%
G1(N= 468)
APS29%
APR63%
AF (*)8%
P1(N= 500)
APS28%
APR62%
AF10%
P2 (N= 453)
APS31%
APR56%
AF13%
P3(N= 500)
APS28%
APR(#) 66%
AF (*)6%
P4(N= 500)
AF14%
APS26%
APR60%
M1(N= 500)
APS28%
APR61%
AF11%
M3(N= 500)
APS31%
APR61%
AF (*)8%
M4(N= 500)
APR59%
APS34%
AF (*)7%
G2(N= 500)
APR(*) 41%
APS (#) 51%
AF (*)8%
C1(N= 488)
APS27%
APR55%
AF (#)18%
C2(N= 488)
APS(*) 22%
AF (#)23%
APR55%
C3(N= 484)
APS(*) 20%
AF (#)22%
APR58%

25
Abertura do dossel
Houve grande variação entre as médias de abertura de dossel entre os fragmentos, nas duas
estações do ano. A porcentagem de abertura diferiu entre as duas estações do ano em todos os
fragmentos, sendo maior durante a estação seca. A heterogeneidade entre os fragmentos quanto à
abertura de dossel pareceu ser mais acentuada durante a estação úmida (Tabela 3). A variabilidade
da abertura entre os pontos de amostragem foi maior nos fragmentos P4 e G2 e durante a estação
seca para todas as áreas (Figura 4).
As áreas de Ribeirão Cachoeira apresentaram as menores médias de abertura de dossel,
menor variabilidade entre os pontos de amostragem e menor variação na abertura entre as duas
estações do ano. Os fragmentos P4 e G2 exibiram as maiores médias de abertura, em ambas as
estações do ano. Essas áreas, juntamente com o fragmento P3, apresentaram também maior
amplitude de variação de abertura de dossel entre as duas estações do ano (Tabela 3, Figura 4).
Somente durante a estação chuvosa houve uma correlação negativa entre a área dos
fragmentos e a porcentagem de abertura de dossel (r= -0,738, p= 0,037).
A proporção de árvores decíduas não variou entre os fragmentos, exceto na área C3 de
Ribeirão cachoeira onde foi significativamente superior às demais (χ2 = 6,9, gl = 1, p < 0,05). O
fragmento P3 foi o único que apresentou proporção de árvores semidecíduas significativamente
inferior aos demais (χ2 = 86,7, gl = 1, p < 0,001), enquanto no fragmento P2 essa proporção foi
superior à dos demais fragmentos (χ2 = 21,8, gl = 1, p < 0,001). A proporção de árvores perenifólias
foi superior às demais amostras nos fragmentos P3 (χ2 = 73,8, gl = 9, p < 0,001) e G3 (χ2 = 6,9, gl = 1,
p < 0,005), enquanto nas amostras P2, M3, G1 e C3 essa proporção foi inferior às demais (P2- χ2
=6,2, gl=1, p < 0,05; M3- χ2 =6,6, gl=1, p < 0,05; G1- χ2 =6,2, gl=1, p < 0,05; C3- χ2 =7,3, gl=1, p <
0,05) (Tabela 4). Ao contrário do que seria esperado, houve uma correlação positiva entre a
proporção de árvores perenifólias nos fragmentos e a variação de abertura nas duas estações do ano
(r= 0,704, p= 0,023).

26
Tabela 3: Médias da porcentagem de abertura de dossel em duas estações do ano: verão e inverno, nos fragmentos estudados na região da APA municipal de Campinas, SP. Valores entre parênteses: desvio padrão.
P1 P2 P3 P4 M3 G1 G2 C1 C2 C3
Verão 7,234abc
(+1,3)
(-1,2)
7,315ab
(+0,99)
(-0,93)
6,827bc
(+1,19)
(-1,11)
7,655a
(+1,79)
(-1,61)
7,124abc
(+1,34)
(-1,23)
6,746c
(+1,36)
(-1,25)
7,838a
(+2,15)
(-1,91)
5,799de
(+0,89)
(-0,83)
6,198d
(+0,93)
(-0,87)
5,554e
(+0,91)
(-0,84)
Inverno 9,308b
(+1,57)
(-1,45)
8,746bc
(+1,49)
(-1,39)
10,504a
(+1,74)
(-1,62)
10,998a
(+2,81)
(-2,53)
9,448b
(+2,14)
(-1,94)
8,805bc
(+2,65)
(-2,33)
10,935a
(+3,36)
(-2,96)
6,89d
(+1,18)
(-1,09)
8,353c
(+1,51)
(-1,39)
7,427d
(+0,83)
(-0,79)
Onde: P1, P2, P3 e P4- fragmentos pequenos; M3- fragmento médio; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira. Obs.: letras diferentes na mesma linha indicam médias significativamente diferentes (Kruskal-Wallis, p< 0,05).
Tabela 4: Porcentagem de indivíduos arbóreos quanto a deciduidade nos fragmentos estudados na região da APA municipal de Campinas, SP. P1 P2 P3 P4 M3 G1 G2 C1 C2 C3
Decíduos 28,4 26,8 32,8 31,6 32,7 32 27,9 30,3 31,9 37,8#
Semidecíduos 28 40,4# 6,2* 31 34,2 34,9 24,9 34,6 25,4 30,1
Perenifólios 43,6 32,8* 61,0# 37,4 33,1* 33,1* 47,2# 35,1 42,7 32,1*
Onde: # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05). Para símbolos de fragmentos ver rodapé da tabela 3.
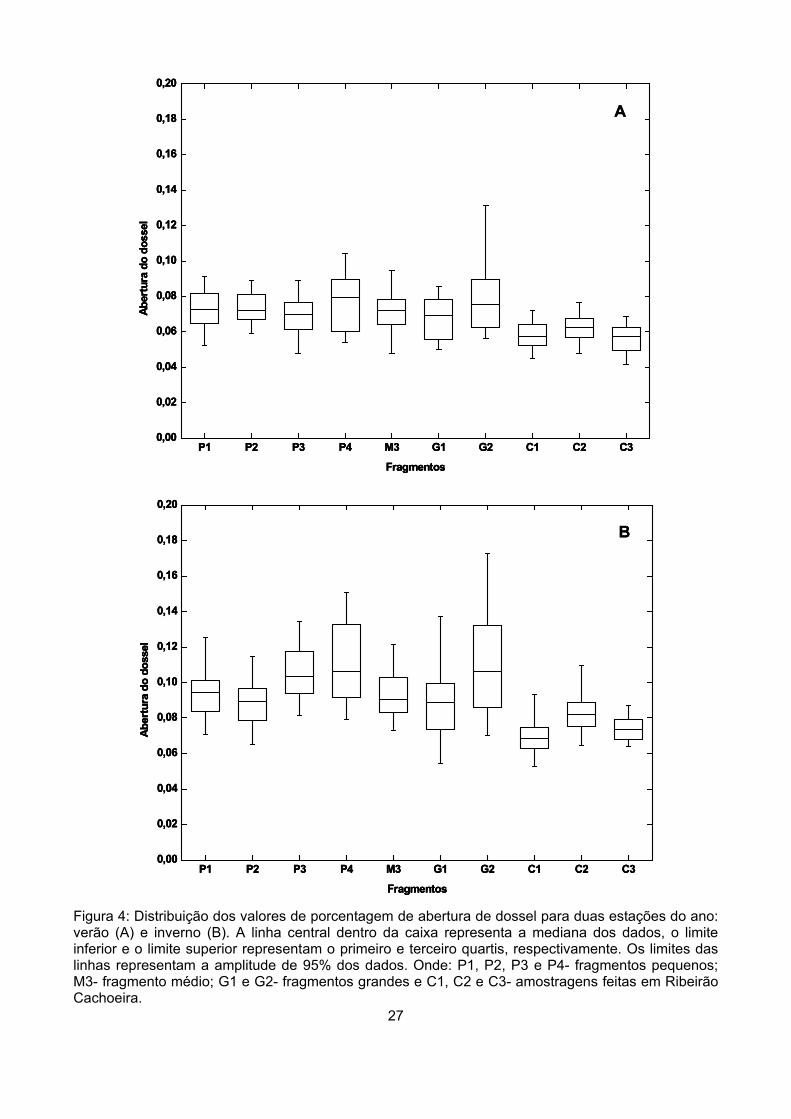
27
Figura 4: Distribuição dos valores de porcentagem de abertura de dossel para duas estações do ano: verão (A) e inverno (B). A linha central dentro da caixa representa a mediana dos dados, o limite inferior e o limite superior representam o primeiro e terceiro quartis, respectivamente. Os limites das linhas representam a amplitude de 95% dos dados. Onde: P1, P2, P3 e P4- fragmentos pequenos; M3- fragmento médio; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
A
B
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
P1 P2 P3 P4 M3 G1 G2 C1 C2 C30,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Aber
tura
do
doss
el
Fragmentos
A
B

28
Infestação por lianas
Dentre os 6500 indivíduos analisados, 4274 (65,8%) apresentavam algum grau de infestação
por lianas. A proporção de árvores infestadas variou de 34 a 86% entre os fragmentos, diferindo entre
as áreas (χ2 = 144,2, gl = 12, p < 0,0001) (Figura 5).
Dois fragmentos pequenos, P1 e P4 e um fragmento grande G1 apresentaram proporção de
árvores infestadas por lianas significativamente inferior às demais áreas, sendo P4 (χ2 = 7,1, gl = 1, p
= 0,008) e G1 (χ2 = 5,1, gl = 1, p < 0,05). O fragmento P1, o de menor área do estudo (12,4ha),
apresentou a menor proporção de árvores infestadas por lianas equivalente à metade da exibida
pelas outras áreas (χ2 = 79,1, gl = 1, p < 0,0001). Os fragmentos P2, M1 e M2, por outro lado,
apresentaram proporções significativamente superiores às demais áreas, sendo P2 (χ2 = 34,1, gl = 1,
p = 0,008) e M2 (χ2 = 5,2, gl = 1, p < 0,05). O fragmento médio M1 apresentou proporção de árvores
infestadas cerca de 30% superior às outras áreas (χ2 = 30,9, gl = 1, p < 0,0001) (Figura 5).
Não houve relação entre a proporção de indivíduos infestados por lianas e o tamanho dos
fragmentos (r = -0,047, p = 0,891).

29
Figura 5: Proporção de árvores infestadas por lianas (área sombreada) nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira; # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05). Entre parênteses o número de indivíduos categorizados em cada fragmento.
M2 (N=500)
26%
74%(#)
M3 (N=500)
73%
27%
G1 (N=500)
42%
58%(*)
G2 (N=500)
65%
35%
P1 (N=500)
66%
34% (*)
P2 (N=500)
25%
75%(#)
P3 (N=500)
70%
30%
P4 (N=500)
44%
56%(*)
M1 (N=500)
14%
86% (#)
M4 (N=500)
28%
72%
C1 (N=500)
66%
34%
C2 (N=500)
63%
37%
C3 (N=500)
62%
38%
M2 (N=500)
26%
74%(#)
M3 (N=500)
73%
27%
G1 (N=500)
42%
58%(*)
G2 (N=500)
65%
35%
P1 (N=500)
66%
34% (*)
P2 (N=500)
25%
75%(#)
P3 (N=500)
70%
30%
P4 (N=500)
44%
56%(*)
M1 (N=500)
14%
86% (#)
M4 (N=500)
28%
72%
C1 (N=500)
66%
34%
C2 (N=500)
63%
37%
C3 (N=500)
62%
38%

30
Discussão A heterogeneidade espacial das florestas tropicais dentro de uma escala local foi primeiramente
reconhecida por Aubréville (1938), para o qual a floresta é um mosaico de manchas de vegetação em
estádios sucessionais distintos, em que os fatores de perturbação exercem um papel preponderante
na determinação de sua estrutura (Hartshorn 1978, Whitmore 1984). A heterogeneidade espacial da
vegetação ocorre usualmente em duas dimensões: na vertical, correspondente à estratificação da
vegetação e na horizontal determinada pela heterogeneidade de fatores abióticos, condições
microclimáticas, edáficas, topográficas, etc (Barberis et al. 2002, Decocq 2002). A fragmentação
florestal “espalhou” esse mosaico ao longo da paisagem e assim a variabilidade entre essas áreas
remanescentes tenderia a ser maior. Entretanto, como os fragmentos pequenos estariam mais
expostos à influencia das condições microclimáticas do ambiente circundante, seria esperada maior
homogeneidade entre os aspectos analisados entre esses fragmentos.
Os fragmentos estudados apresentaram grande variabilidade quanto às suas características
abióticas e bióticas. Existem variações de solo, relevo, hidrografia, tamanho e também quanto à
freqüência e intensidade de fatores de perturbação. Todos esses fatores agem sinergisticamente, por
isso seria ingenuidade atribuir as variações observadas somente à área total dos fragmentos ou a
efeitos da fragmentação. Além disso, nem todos os remanescentes aqui estudados constituem
remanescentes de uma floresta outrora contínua. O fragmento P4, por exemplo, é uma floresta
secundária, que se estabeleceu em uma área de pastagem abandonada.
A maioria dos aspectos qualitativos analisados não variaram apenas com a área dos
fragmentos. Por exemplo, as maiores médias de abertura de dossel, assim como a maior
variabilidade, foram encontradas em um fragmento grande (G2) e em um fragmento pequeno (P4). A
mata G2 apresentou também uma maior proporção de árvores danificadas, o que está de acordo com
o proposto por Trichon et al. (1998), segundo os quais as aberturas no dossel podem se formar não
só pela ocorrência de clareiras, mas também por queda ou quebra de galhos. Essas duas
observações nesse fragmento parecem estar associadas a seu histórico de perturbação, com a
ocorrência de uma queimada recente que atingiu quase toda a mata. Para Bianchini et al. (2001) a
freqüência e a intensidade de perturbações no dossel contribuem para a maior variação nas taxas de
abertura dentro de uma mesma área e entre áreas.
No caso do fragmento P4, é possível que o dossel apresente maior proporção de abertura por
se tratar de uma floresta mais recente, em que a maioria das unidades do mosaico florestal seria
constituída pelos estádios sucessionais iniciais. A proporção de abertura no dossel de uma floresta
pode ser influenciada por muitos aspectos de sua estrutura, como o arranjo espacial das plantas, a
arquitetura das copas e a área foliar (Bianchini et al. 2001). Florestas de estádios sucessionais
iniciais, onde não existem três estratos de vegetação bem estabelecidos e áreas que sofreram
perturbações recentes tendem a apresentar maiores porcentagens de abertura de dossel (Trichon et
al. 1998).

31
São raros os trabalhos que utilizam fotografias hemisféricas para avaliar a abertura do dossel
em florestas semidecíduas. Isso dificulta a interpretação e a comparação dos resultados aqui obtidos.
Entretanto, Trichon et al. (1998), estudaram a estrutura de uma floresta tropical na Indonésia através
de fotografias hemisféricas e observaram que porcentagens de abertura inferiores a 7%
correspondiam a florestas mais bem estruturadas ou “maduras”. Apesar de as comparações entre os
dois ecossistemas exigirem cautela por serem distintos, observamos que apenas as áreas de
Ribeirão Cachoeira e o fragmento G1 apresentaram porcentagens de abertura inferiores a esse valor.
Assim, os resultados obtidos em relação a este parâmetro concordam em parte com o que seria
esperado: as áreas dentro de um fragmento grande apresentaram menores taxas de abertura de
dossel do que fragmentos isolados ao longo da paisagem. Entretanto, essa relação não parece estar
associada simplesmente ao tamanho dos fragmentos, uma vez que um dos maiores fragmentos do
estudo apresentou a maior abertura. Outros fatores parecem estar influenciando, como a idade do
fragmento e a ocorrência de fatores de perturbação, como queimadas.
A variação das taxas de abertura do dossel entre as estações do ano também foi menor nas
áreas de Ribeirão Cachoeira e maior nos demais fragmentos, atingindo as maiores amplitudes em G2
e P4. Apesar de não ter sido observada relação entre essas variações e a proporção de árvores
decíduas ou semidecíduas nessas áreas, a deciduidade pode ser um dos principais fatores nas
variações observadas. Plantas decíduas podem perder mais ou menos folhas dependendo das
condições ambientais (Bianchini et al. 2001). Em ambientes mais abertos, como é o caso das áreas
G2 e P4, a umidade relativa do ar tende a ser menor e a transpiração maior (Heich 1995). Além disso,
fragmentos menores sofreriam maior estresse hídrico e ressecamento do que fragmentos grandes,
por estarem mais expostos a ventos e penetração de luz, com conseqüente aumento da temperatura
(Lovejoy et al. 1996, Murcia 1995). Nesses ambientes as plantas decíduas ou semidecíduas
tenderiam a perder mais folhas do que em ambientes mais úmidos (Heich 1995, Popma et al. 1988).
A deciduidade também pode ser influenciada pelo custo da produção e manutenção de folhas em
determinados ambientes (Kikuzawa 1991). O custo de produção de folhas novas pode ser limitante
em ambientes mais sombreados, como nas áreas de Ribeirão Cachoeira, e pode ser restringido a
indivíduos emergentes ou que compõem as camadas superiores do dossel.
O aumento de lianas em ambientes perturbados, como fragmentos, tem sido associado às
condições microclimáticas. Fragmentos pequenos sofreriam maior influência das condições externas,
como mencionado acima, e tais condições poderiam favorecer a proliferação de lianas nesses
ambientes (Laurance 1991, 1997, Laurance et al. 2001, Tabanez et al. 1997, Viana et al. 1997).
Entretanto, nos resultados encontrados sobre a infestação por lianas, não foi observado aumento de
infestação com a diminuição da área dos fragmentos, como seria esperado segundo Viana &
Tabanez (1996). Ao contrário, dois fragmentos pequenos (P1 e P4) apresentaram as menores
proporções de árvores infestadas.

32
Lianas são um componente natural da comunidade florestal tropical e contribuem para boa parte
de sua biodiversidade (Schenck 1892 apud Putz 1984, Croat 1978, Peñalosa 1985). Na Reserva de
Santa Genebra, o maior fragmento de floresta estacional semidecídua do município de Campinas, as
lianas representam cerca de 20% das espécies de plantas fanerogâmicas (Leitão Filho 1995).
Essas plantas ocorrem em abundância em muitas florestas. Em 1ha de floresta em Barro
Colorado, Panamá, Putz (1984) observou que 43% das árvores com DAP ≥ 20cm suportavam pelo
menos uma liana. Para árvores com DAP ≥ 10cm, Putz (1983) encontrou 42% de árvores infestadas
em um trecho de floresta amazônica da Venezuela e, na floresta do Parque Nacional de Lambir, na
Malásia, essa proporção variou de 20,8 a 63,3% (Putz & Chai 1987). No presente estudo as
proporções encontradas foram semelhantes às citadas acima ou maiores em alguns casos (35-85%).
Tabanez et al. (1997) encontraram 63% da cobertura de dossel de uma floresta estacional no
interior do estado de São Paulo representado por lianas. Para os autores, tal proporção seria sinal de
degradação da floresta, pois, segundo Peñalosa (1985), lianas são, em sua maioria, espécies
heliófilas que se beneficiam muito de perturbações naturais ou antrópicas. Por outro lado, para Leitão
Filho (1995), a ocorrência de lianas nas florestas estacionais semidecíduas acaba sendo favorecida
em função da deciduidade de um grande número de árvores, o que contribuiria para o aumento da
luminosidade no chão da floresta, favorecendo as condições de germinação destas plantas. Não se
sabe, portanto se a proporção aqui encontrada seria natutral ou reflexo de perturbações externas,
mas observamos que os ambientes com maiores aberturas de dossel ou mais perturbados não
apresentaram maior proporção de infestação do que áreas mais fechadas, tendo ocorrido até mesmo
o inverso no fragmento P4.
Para Putz & Chai (1987) a abundância de lianas pode aumentar em florestas onde as quedas
de árvores são mais freqüentes, como bordas de mata e fragmentos. Mas, para Putz (1984), as
perturbações na estrutura da floresta explicariam apenas parcialmente a abundância de lianas. O
autor argumentou que muitas árvores pioneiras raramente são infestadas por lianas devido a seu
crescimento rápido e monopodial. Isto poderia explicar a menor proporção de lianas no fragmento P4,
o mais recente do estudo. Segundo Putz (1984), a disponibilidade de suporte parece ser um fator
mais importante para que uma liana atinja o dossel da floresta do que a disponibilidade de luz. A
distribuição vertical e horizontal de árvores ou ramos de diâmetros suficientemente pequenos para
servir de suporte para uma liana estariam entre os fatores preponderantes que controlariam sua
abundância no ambiente florestal, que em última análise dependeria da estrutura da floresta. Além
disso, as lianas tendem a ser mais abundantes em solos mais férteis (Putz & Chai 1987).
Tais colocações auxiliam na interpretação dos resultados obtidos aqui. A ausência de relação
entre a área dos fragmentos e a proporção de árvores infestadas por lianas pode indicar que, para os
fragmentos estudados, essa proporção pode ser influenciada por outros fatores que não
simplesmente o tamanho dos fragmentos. No fragmento P1, por exemplo, onde foi encontrada a

33
menor proporção de infestação, foram realizadas atividades silviculturais de retirada de lianas durante
muitos anos, segundo relatos dos proprietários.
Não encontramos também relação entre o tamanho dos fragmentos e a proporção de árvores
danificadas. Entretanto, nas áreas de Ribeirão Cachoeira observamos um maior equilíbrio entre as
proporções de árvores do passado e árvores do futuro do que nos demais fragmentos. Isto indica que
em Ribeirão Cachoeira existe um maior potencial de reposição para as as plantas que estariam
deixando o sistema, como é o caso das árvores do passado. Nos fragmentos <100ha a proporção de
árvores do passado representaram o dobro ou o triplo da proporção de árvores do futuro, indicando
que a regeneração nessas áreas pode estar comprometida. Em todos os fragmentos <100ha a
proporção de árvores do passado que, teoricamente, estariam deixando o sistema, representam o
dobro ou o triplo da proporção de árvores do futuro. A confirmação de tais indicações, no entanto,
prescindem estudos específicos sobre dinâmica e regeneração nestes fragmentos.
Tais resultados diferem do proposto por Laurance et al. (1998b), segundo os quais as taxas de
recrutamento seriam maiores em fragmentos mais sujeitos a perturbações, como os fragmentos
pequenos. Nos fragmentos pequenos (< 100ha) e nas áreas mais perturbadas a proporção de
árvores do futuro ou em regeneração foi menor que nas áreas de Ribeirão Cachoeira (244ha).
A diminuição das taxas de regeneração em determinada área é uma questão importante para
sua sustentabilidade e manutenção. As espécies que não estão regenerando tendem a desaparecer
dessas áreas, comprometendo a manutenção de sua diversidade e composição no futuro. Estudos
aprofundados sobre esta questão e sobre quais fatores a determinam são, portanto, urgentes. Muitos
fatores podem estar envolvidos na diminuição da regeneração de espécies em fragmentos. Ausência
de polinizadores poderia influenciar nas taxas de reprodução, aumentando as taxas de extinção local
e a dificuldade de deslocamento de dispersores ao longo da matriz influenciaria as taxas de
colonização (Aizen & Feinsinger 1994). Ambas as situações são previstas na literatura como os
principais causadores de perda de diversidade em ambientes fragmentados como prevê a teoria de
biogeografia de ilhas (Mac Arthur & Wilson 1967).
As observações do presente estudo indicaram que mais fatores podem estar associados aos
aspectos qualitativos avaliados, além do tamanho dos fragmentos. Não houve uma relação direta
entre a área dos fragmentos e aspectos como infestação por lianas, abertura de dossel e qualidade
de dossel. Fragmentos menores não necessariamente exibiram maiores proporções de infestação por
lianas ou de árvores danificadas do que fragmentos maiores. A proporção de abertura de dossel
também não variou apenas com a área dos fragmentos. Essas proporções parecem variar segundo o
histórico de preservação dessas áreas, como a ocorrência de fogo ou atividades de manejo. Tais
variações também podem estar associadas a características intrínsecas dessas matas que não foram
analisadas aqui, como a estrutura florestal, a arquitetura das plantas e outros aspectos que, em
última análise estão associados à heterogeneidade ambiental.

34
Apesar disso, as áreas de Ribeirão Cachoeira diferiram dos demais fragmentos por
apresentarem menor porcentagem de abertura de dossel em ambas as estações do ano e maior
equilíbrio entre as proporções de APS e AF. As maiores proporções de AF e menores proporções de
APS podem estar se refletindo na menor abertura de dossel e este conjunto de características
indicando que as áreas de Ribeirão Cachoeira estariam mais bem preservadas do que os fragmentos
<100ha. Essa variação pode ter implicações importantes sobre a composição, estrutura e diversidade
dessas áreas ao longo do tempo, tornando urgentes estudos mais aprofundados sobre a dinâmica
das populações nessas áreas, com o intuito de delinear medidas mais eficazes de preservações e de
recuperação de fragmentos <100ha.

35
Referêcias bibliográficas AIZEN, M.A. & FEINSINGER, P. 1994. Forest fragmentation, pollination, and plant reproduction in a
chaco dry forest, Argentina. Ecology 75: 330-351.
AUBRÉVILLE, A. 1938. La forêt coloniale de l'Afrique Occidentale Fançaise. Annales de l'Académie des Sciences Coloniales 9
AYRES, M., AYRES Jr., M., AYRES, D.M. & SANTOS, A.S. 1998. BioEstat: Aplicações estatísticas nas áreas das Ciências Biológicas e Médicas. Sociedade civil Mamirauá. Manaus.
BARBERIS, I.M., BATISTA, W.B., PIRE, E.F., LEWIS, J.P. & LEÓN, R.J.C. 2002. Wood population
distribution and environmental heterogeneity in a Chaco forest, Argentina. Journal of Vegetation Science 13: 607-614.
BIANCHINI, E., PIMENTA, J.A., SANTOS, F.A.M., 2001. Spatial and temporal variation in the canopy
cover in a tropical semideciduous forest. Brazilin Archives of Biology and Technology 44: 269-
276.
CHEN, J., FRANKLIN, J.F. & LOWE, J.S. 1992. Vegetation responses to edge environments in Old-
Growth Douglas-fir forests. Ecological Applications 2:387-396.
COTTAN, G. & CURTIS, J.T. 1956. The use of distance measures in phytossociological sampling.
Ecology 37: 451-460.
CROAT, R.B. 1978. Flora of Barro Colorado Island. Stanford University Press, Stanford, California.
DAWKINS, C. 1961. New methods of improving stand composition in tropical forests. Caribbean Forester 22: 12-20.
DECOCQ, G. D. 2002. Patterns of plant species and community diversity at different organization
levels in a forested riparian landscape. Journal of Vegetation Science 13: 91-106.
DIDHAM, R.K. & LAWTON, J.H. 1999. Edge structure determines the magnitude of changes in
microclimate and Vegetation structure in tropical forest fragments. Biotropica 31: 17-30.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista
Brasileira de Botânica 23: 371-383.
FERREIRA, L.V. & LAURANCE, W 1997. Effects of forest fragmentation on mortality and damage of
selected trees in Central Amazonia. Conservation Biology 11: 797-801.
FRAZER, G.W., CANHAM, C.D. & LERTZMAN, K.P. 1999. Gap Light Analyzer (GLA): Imaging
software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. Simon Frazer University,
Burnaby, British Columbia, and the Institute of Ecossystem Studies, Millbrook, New York.
HARTSHORN, G.S. 1978. Treefalls and tropical forest dynamics. Pp 617-638. In: P.B. Tonlinson &
M.H. Zimmerman (eds.), Tropical trees as living systems. Cambridge University Press, New
York.

36
HEICH, P.B. 1995. Phenology of tropical forests: patterns, causes, and consequences. Cannadian Journal of Botany 73: 164-174.
KAPOS, V. 1989. Effects of isolation on the water status of forest patches in the brazilian Amazon.
Journal of Tropical Ecology 5: 173-185
KAPOS, V., WANDELLI, E., CAMARGO, J.L. & GANADE, G. 1997. Edge-related changes in
environment and plant responses due to forest fragmentation in Central Amazonia. Pp 33-44. In:
W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: Ecology, management, and conservation of forest communities. University of Chicago Press. Chicago.
KIKUZAWA, K. 1991. A cost-benefit analysis of leaf longevity of trees and their geographical pattern.
The American Naturalist 138: 1250-1263.
KÖPPEN, W. 1948. Climatologia. México - Buenos Aires. Ed. Fundo de Cultura Econômica, 478p.
LAURANCE, W.F. 1991. Edge effects in tropical forest fragments: application of a model for the
design of nature reserves. Biological Conservation 57:205-219.
LAURANCE, W.F. 1997. Hyper-disturbed parks: edge effects and the Ecology of isolated rainforest
reserves in tropical Australia. Pp 71-83. In: W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: Ecology, management, and conservation of forest communities. University
of Chicago Press. Chicago.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M. & LAURANCE, S.G. 1998a. Rain
forest fragmentation and the dynamics of amazoniam tree communities. Ecology 79: 2032-2040.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M., LAURANCE, S.G., HUTCHINGS,
R.W. & LOVEJOY, T.E. 1998b. Effects of forest fragmentation on recruitment patterns in
amazonian tree communities. Conservation Biology 12: 460-464.
LAURANCE, W.F., PÉREZ-SALICRUP, D., DELAMÔNICA, P., FEARNSIDE, P.M., D’ANGELO, S.,
JEROLINSKI, A ., POHL, L. & LOVEJOY, T.E. 2001. Rain forest fragmentation and structure of
amazonian liana communities. Ecology 82: 105-116.
LEITÃO FILHO, H.F. 1995. A vegetação da Reserva de Santa Genebra. Pp 19-29. In: L.P.C Morellato
& H.F. Leitão Filho (eds.), Ecologia e preservação de uma floresta tropical urbana - Reserva
de Santa Genebra. Editora Unicamp. Campinas, SP.
LOVEJOY, T.E., BIERREGAARD, R.O., RYLANDS , A.B., MALCOLM, J.R., QUINTELA, C.E.,
HARPER, L.H., BROWN, K.S., POWELL, A.H., POWELL, G.V.N., SCHUBART, H.O.R. & HAYS
.B. 1996. Edge and other effects of isolation on Amazon forest fragments. Pp 257-285. In: M.E.
Soule (ed.). Conservation Biology: the science of scarcity and diversity. Sinauer, Suderland.
USA
LOWE, R.G. & WALKER, P. 1977. Classification of canopy stem crown status and climber infestation
in natural tropical forest in Nigeria. Journal of Applied Ecology 14: 897-903.
MacARTHUR , R.H. & WILSON, E.O. 1967. The theory of island biogeography. Princeton
University Press. Princeton.

37
MALCOLM, J.R. 1994. Edge effects in central amazonian forest fragments. Ecology 75: 2438-2445.
MATLACK, G.R. 1993. Microenvirnment variation within and among forest edge sites in the eastern
United States. Biological Conservation 66: 185-194.
MATTOS, C.O. 1996. Contribuição ao planejamento e gestão da Área de Proteção Ambiental de Sousas e Joaquim Egídio, Campinas/SP. Dissertação de Mestrado. Universidade Estadual de
São Paulo. São Paulo.
MELLO, M.H.A., PEDRO JUNIOR, M.J., ORTOLANI, A. A. & ALFONSI R.R. 1994. Chuva e
Temperatura: cem anos de observações em Campinas. Boletim Técnico 154 . Instituto
Agronômico Campinas, SP.
MIRANDA, Z.A.I. 1996. Plano de gestão da Área de Proteção Ambiental da região de Sousas e
Joaquim Egídio - APA Municipal. Relatório da Secretaria de Planejamento e Meio Ambiente - SEPLAMA. Prefeitura Municipal de Campinas, SP.
MÜLLER-DOMBOIS, D. & ELLENBERG, H. 1974. Aims and methods of vegetation Ecology. New
York , John Wiley & Sons.
MURCIA, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends in Ecology and Evolution 10: 58-62.
NICHOLSON, D.I. 1958. An analysis of logging damage in tropical rain forest, North Borneo. Malayan Forester 21: 235-245.
OLDEMAN R.A.A. 1989. Dynamics in tropical forests. pp 3-21. In: Holm-Nielsen, L.B., Nielsen, I.C. &
Balslev, H. 1989. Tropical Forest: Botanical Dynamics, Speciation and Diversity. Academic
Press.
OLIVEIRA, J.B., MARCELO, N.C., ROSSI, M. & CALDERANO FILHO, B. 1999. Mapa pedológico do
estado de São Paulo. Legenda expandida In: OLIVEIRA, J.B. 1999. Boletim científico n.o 45,
Solos do estado de São Paulo. Embrapa e IAC. Campinas.
OLIVEIRA-FILHO, A. T., MELLO, J.M. & SCOLFARO, J.R.S. 1997. Effects of past disturbance and
edges on tree community structure and dynamics within a fragment of tropical semideciduous
forest in south-eastern Brazil over a five-year period (1987-1992). Plant Ecology 131: 45-66.
PEÑALOSA, J. 1985 Dinamica del crescimiento de lianas. Pp 147-169. In: A. Gomez-Pompa & R.S.
Del Amo (eds.). Invetigaciones sobre la regeneracion de selvas altas em Vera Cruz, Mexico.
Editorial Alhambra Mexicana, Mexico D.F.
PÉREZ-SALICRUP, D.R. 1998. Effects of liana cutting on trees and tree seedlings in a tropical forest in Bolivia. Dissertation. University of Missouri, St. Louis, Missouri.
POPMA, J., BONGERS, F. & CASTILLO, J.M. 1988. Patterns in the vertical struture of the tropical
lowland rain forest of Los Tuxtlas, Mexico. Vegetatio 74: 81-91.
PUTZ, F.E. 1980. Lianas vs. Trees. Biotropica 12: 224-225.
PUTZ, F.E. 1983. Treefall pits and mounds, buried seeds and the importance of soil disturbance to
pioneer trees on Barro Colorado Island, Panama. Ecology 64: 1069-1074.

38
PUTZ, F.E. 1984. The natural history of lianas on Barro Colorado Island, Panama. Ecology 65: 1713-
1724.
PUTZ, F.E. & MILTON, K. 1982. Tree mortality rates on Barro Colorado Island. Pp 95-100. E.G. Leigh,
A.S. Rand & D.M. Windsor (ed.). The Ecology of tropical forest: seasonal rhythms and long-term changes. Smithsonian Press, Washington.
PUTZ, F.E. & CHAI, P. 1987. Ecological studies of lianas in Lambir National Park, Sarawak, Malaysia.
Journal of Ecology 75: 523-531.
SANTIN, D.A. 1999. A vegetação remanescente do município de Campinas (SP): mapeamento, caracterização fisionômica e florística, visando a conservação. Tese de doutorado.
Universidade Estadual de Campinas, SP.
SAUNDERS, D.A., HOBBS, R.J. & MARGULES, C.R. 1991. Biological consequences of ecossystem
fragmentation: a review. Conservation Biology 5: 18-32.
STEVENS, G.C. 1987. Lianas as structural parasites: the Bursera simaruba example. Ecology 68:
77-81.
TABANEZ, A.A.J., VIANA, V.M. & DIAS, A.S. 1997. Conseqüências da fragmentação e do efeito de
borda sobre estrutura, diversidade e sustentabilidade de um fragmento de floresta de planalto de
Piracicaba, SP. Revista Brasileira de Biologia 57: 47-60.
TABARELLI, M. & MANTOVANI, W. 1999. A riqueza de espécies arbóreas na floresta atlântica de
encosta no estado de São Paulo (Brasil). Revista Brasileira de Botânica 22: 217-223.
TRICHON, V., WALTER, J.-M.N. & LAUMONIER, Y. 1998. Identifying spatial patterns in the tropical
rain forest structure using hemispherical photographs. Plant Ecology 137: 227-244.
TURNER, I.M. & TAN, H.T.W. 1992. Ecological impact of alien plant species in Singapore. Pacific
Science 46: 389-390.
VELOSO, H.P., RANGEL FILHO, A .L.R. & LIMA, J.C. 1991. Classificação da vegetação brasileira adaptada a um sistema universal. Rio de Janeiro. Instituto Brasileiro de Geografia e Estatística
(IBGE).
VIANA, V.M. & TABANEZ, A.A.J. 1996. Biology and conservation of forest fragments in the Brazilian
Atlantic moist forest. Pp 151-165. In: Schelhas, J. & Greenberg, R. (ed.). Forest patches in tropical landscapes. Island Press, Washington.
VIANA, V.M., TABANEZ, A.A. & BATISTA, J. 1997. Dynamics and restoration of forest fragments in
the Brazilian Atlantic moist forest. Pp 351-365. In: W.F. Laurance & R.O. Bierregaard (eds.)
Tropical forest remnants: Ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago.
WHIGHAM, D. 1984. The influence of vines on the growth of Liquidambar styraciflua L. (swetgum).
Canadian Journal of Forest Research 14: 37-39.
WHITMORE, T.C. 1984. Tropical rain forest of the Far East. Clarendon Press, Oxford.

39
WHITMORE,T.C., BROWN, N.D., SWAINE, M.D., KENNEDY, D., GOODWINBALEY, C.I. & GONG,
W.K. 1993. Use of hemisferical photographs in forest Ecology: measurement of gap size and
radiation totals in a Borneo tropical rain forest. Journal of Tropical Ecology 9: 131-151.
WILLIANS-LINERA, G. 1990. Vegetation structure and enviromental conditions of forest edges in
Panama. Journal of Ecology 78: 356-373.
ZAR, J.H. 1989. Bioestatistical Analysis. Prentice Hall, New Jersey.

40
CAPÍTULO 2
COMPOSIÇÃO FLORÍSTICA E SIMILARIDADE DE ONZE FRAGMENTOS DE FLORESTA ESTACIONAL SEMIDECÍDUA
NA REGIÃO DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE CAMPINAS, SP

41
Capítulo 2
Composição florística e similaridade de onze fragmentos de floresta estacional semidecídua na região da Área de proteção Ambiental do município de Campinas, SP
Karin dos Santos Pós-Graduação em Biologia Vegetal – Universidade Estadual de Campinas – Parte da tese de doutorado,
Luiza Sumiko Kinoshita Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Flavio A. M. dos Santos Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Resumo As florestas estacionais semidecíduas encontram-se extremamente fragmentadas e representadas
por remanescentes de vegetação isolados e de tamanhos diversos. Fragmentos menores estariam
mais sujeitos a extinções locais de espécies nativas e mais expostos a fatores de perturbação do que
fragmentos grandes, levando a padrões diferenciais de extinção e a alterações da composição
florística. O objetivo deste trabalho foi verificar se a composição florística de fragmentos pequenos
diferia da encontrada em fragmentos grandes. Foram escolhidos onze fragmentos de floresta
estacional semidecídua na Área de Proteção Ambiental do município de Campinas, SP. Dez
fragmentos com áreas < 100ha agrupados em três classes de tamanho e um fragmento com 244,9ha.
Em cada fragmento foi realizada uma amostragem de 125 pontos quadrantes, onde foram
amostrados indivíduos com DAP ≥ 10cm. As amostras de 500 indivíduos foram comparadas com
amostras realizadas nas mesmas condições em três áreas diferentes de um fragmento de 244,9ha.
Foram coletados 6500 indivíduos pertencentes a 248 espécies, 149 gêneros e 55 famílias. O número
de espécies variou pouco entre as amostras. As famílias mais ricas foram Myrtaceae, Lauraceae,
Fabaceae, Euphorbiaceae, Rutaceae, Meliaceae e Moraceae e as espécies mais constantes foram
Astronium graveolens, Centrolobium tomentosum, Piptadenia gonoacantha, Seguieria langsdorffi,
Aspidosperma polyneuron e Copaifera langsdorffii. Cerca de 22% das espécies ocorreram em apenas
um fragmento. Muitas dessas espécies estão sendo citadas pela primeira vez para a região de
Campinas. Fragmentos de mesmo tamanho foram tão heterogêneos entre si quanto áreas distintas
dentro de um mesmo fragmento, indicando que as variações na composição florística não estariam
relacionadas ao tamanho dos fragmentos. As variações de composição pareceram estar mais
associadas à heterogeneidade ambiental do que aos efeitos da fragmentação, embora fatores como
proximidade geográfica e histórico de preservação também pareçam estar influenciando.

42
Introdução As florestas estacionais semidecíduas têm ampla distribuição no interior do estado de São
Paulo. Ocorrem em áreas de clima e solo variados, mas com uma característica climática muito
constante: invernos secos e frios e verões quentes e úmidos (Leitão Filho 1982). É esta sazonalidade
que determina uma de suas características mais marcantes: durante a estação seca, cerca de 50 a
70% de suas espécies perdem folhas (Morellato et al. 1989, Morellato 1995), daí o termo
semidecídua.
Esse tipo florestal foi mais rápida e intensamente devastado no estado de São Paulo. A
devastação ocorreu associada à expansão da fronteira agrícola, pois essas florestas ocupavam os
solos de maior fertilidade do estado e em regiões de relevo favorável à agricultura (Victor 1975).
Restaram poucos fragmentos remanescentes, cuja representatividade e estado de preservação têm
sido questionadas (Fonseca 1981, Tabanez et al. 1997, Durigan et al. 2000). Em Campinas, essas
florestas, originalmente predominantes e contínuas (Christofoletti & Federici 1972), ocupam hoje
apenas 2,55% da área total do município, distribuídos em 197 fragmentos de tamanhos
predominantemente inferiores a 10 hectares (Santin 1999).
Para Nascimento et al. (1999), as florestas estacionais semidecíduas do interior do estado de
São Paulo são um exemplo claro do processo de fragmentação, uma vez que constituem ilhas de
vegetação em meio a uma matriz dominada pela agricultura e grandes centros urbanos. Vale lembrar,
entretanto, que nem todas essas matas representam áreas remanescentes de uma vegetação
original e sim, áreas exploradas, manejadas ou florestas secundárias em diversos estádios do
processo de regeneração.
O estudo das florestas estacionais remanescentes teve significativo incremento nas últimas
duas décadas. Alguns estudos mostraram que essas florestas são bastante heterogêneas (Salis et al.
1995, Torres et al. 1997, Costa & Mantovani 1995, Santos & Kinoshita 2003). As espécies distribuem-
se diferencialmente entre os fragmentos e mesmo dentro dos fragmentos (Assumpção et al. 1982;
Rodrigues et al. 1989). Fatores abióticos como umidade, solo, topografia e altitude influenciam
fortemente a distribuição das espécies (Basnet 1992, Lieberman et al. 1985, Oliveira Filho et al.
1994a, 1994b, 1997, Botrel et al. 2002). Além da heterogeneidade ambiental, para Bertoni & Martins
(1987), os diferentes graus de perturbação antrópica a que essas matas estão submetidas também
contribuem para que algumas espécies ocorram em algumas localidades e sejam raras ou ausentes
em outras.
Grande parte das espécies que compõem esse ecossistema ocorrem com poucos indivíduos
por unidade de área (Cavassan et al. 1984, Matthes et al. 1988, Pagano et al. 1995). Segundo
Terborgh & Winter (1980), espécies que ocorrem em baixas densidades são mais suscetíveis à
extinção pela fragmentação porque formam pequenas populações que não se perpetuariam com a
insularização. A persistência de espécies de árvores em fragmentos pode ser influenciada tanto por
eventos no nível das populações, via eventos genéticos ou demográficos (Shafer 1981), quanto no da

43
comunidade com o declínio da reprodução devido à perda de polinizadores e dispersores de
sementes (Powel & Powell 1987, Aizen & Feinsinger 1994). Além disso, espécies mais sensíveis às
perturbações advindas do entorno dos fragmentos não sobreviveriam nesses ambientes (Lovejoy et
al. 1996, Kapos et al. 1997).
Tais efeitos seriam mais ou menos acentuados de acordo com o grau de isolamento e a área
total do fragmento (Saunders et al. 1991, Turner 1996). A maioria dos organismos têm um poder de
dispersão limitado e, em conseqüência disso seria esperado que as taxas de colonização ou
recolonização decrescessem com o aumento da distância entre fragmentos (Hanski 1994) e que
fragmentos pequenos suportassem apenas populações pequenas, cujo risco de extinção seria maior
(Williamson 1981).
Por outro lado, espécies invasoras e dos estádios iniciais de sucessão (pioneiras) têm suas
oportunidades de expansão e colonização aumentadas nesses fragmentos (Gómez-Pompa 1971;
Brown & Brown 1992, Laurance et al. 1998). Espécies invasoras são aquelas que alcançam
determinadas áreas em consequência de atividades humanas (Pysek 1995 apud Dislich et al. 2002).
Segundo Janzen (1983), Bierregaard et al. (1992) e Gascon et al. (1999), tais espécies viriam,
principalmente, do entorno dos fragmentos, dominado por atividades antrópicas. Espécies pioneiras
são componentes naturais dessas florestas, mas regeneram somente sob altas intensidades de luz,
situação que, em florestas naturais, só ocorrem em grandes clareiras ou bordas de mata (Budowski
1965, Hartshorn 1978, Swaine & Whitmore 1988). A penetração e/ou aumento excessivo da
abundância dessas espécies alteraria gradualmente a composição original das florestas
fragmentadas e também influenciaria a abundância e persistência de espécies da composição original
(Terborgh 1992).
As espécies dispersas pelo vento (anemocóricas) também poderiam ser beneficiadas em
paisagens fragmentadas. Em ambientes mais abertos (matriz), suas chances de atingirem longas
distâncias são maiores (Howe & Smallwood 1982), enquanto para as espécies zoocóricas, a matriz
muitas vezes constitui um obstáculo intransponível. Dessa forma, as plantas anemocóricas teriam
maiores chances de colonização e recolonização de áreas, o que ajudaria tanto em sua manutenção
quanto na ocupação de novas áreas. Por outro lado, as espécies zoocóricas poderiam estar mais
sucetíveis à extinção via eventos demográficos e genéticos nos fragmentos, pois não teriam as
mesmas probabilidades de colonização e recolonização que as espécies dispersas pelo vento.
Tabarelli et al. (1999) observaram que em fragmentos pequenos havia uma redução na riqueza
de espécies das famílias Myrtaceae, Lauraceae, Rubiaceae e Sapotaceae, consideradas as famílias
mais importantes para vertebrados frugívoros na floresta Atlântica (Moraes 1992, Galetti 1996). Para
os autores, a diminuição da proporção de plantas zoocóricas teria um efeito desastroso sobre as
populações de frugívoros, pois ocasionaria uma redução na oferta de frutos comestíveis à fauna local.
A relação entre disponibilidade de frutos e a abundância e riqueza de aves frugívoras foi
observada por Guidon (1995) nas florestas montanas da Costa Rica e também por Kattan & Alvarez-

44
Lópes (1995) nos andes colombianos. Os mesmos estudos apontaram que esta relação era
influenciada pelo tamanho dos fragmentos. Fragmentos menores ofereciam menor disponibilidade de
frutos do que fragmentos maiores.
Como indicado acima, vários estudos sugerem que os padrões de extinção e colonização em
fragmentos seriam diferentes e influenciados por seu tamanho e também pela redução da
heterogeneidade local que acompanha os efeitos da redução de tamanho. Ocorreria portanto, uma
eliminação diferencial de espécies: em fragmentos pequenos sobreviveriam somente as espécies
adaptadas àquelas condições, onde deveriam ser mais freqüentes e abundantes e, o número de
espécies por unidade de área deveria ser menor do que em fragmentos grandes. No entanto, se os
fragmentos pequenos estão mais sujeitos à penetração de plantas invasoras e pioneiras e à
colonização de espécies anemocóricas, talvez não seriam detectadas variações no número de
espécies entre fragmentos e sim na sua composição florística. Análises de similaridade poderiam
apontar tais diferenças e, segundo as proposições acima, seria esperado que fragmentos pequenos
exibissem maior similaridade florística entre si do que com os fragmentos maiores. Isto seria
esperado, porque os efeitos da fragmentação e o grau de exposição às perturbações externas
tenderiam a ocorrer com intensidade e amplitude semelhantes nos fragmentos pequenos. Além disso,
os fragmentos grandes englobariam uma maior heterogeneidade ambiental e os fatores de
perturbação externos os atingiriam em menor proporção. Com isso, pequenas áreas escolhidas ao
acaso dentro de um fragmento grande tenderiam a ser mais heterogêneas entre si.
O presente trabalho teve como objetivo investigar as seguintes questões: Como varia a
composição florística de amostras de mesmo tamanho tomadas em fragmentos de diferentes
tamanhos e em regiões distintas de uma área contínua? Fragmentos pequenos são mais
semelhantes entre si, do ponto de vista florístico, do que amostras tomadas em diferentes regiões de
um fragmento grande? A proporção de espécies zoocóricas, anemocóricas e autocóricas difere entre
fragmentos pequenos e grandes?

45
Material e métodos Área de estudo
O estudo foi realizado predominantemente na região da Área de Proteção Ambiental - APA - do
município de Campinas, SP (22°45’ a 23°00’ S, 47°00’ a 47°12’ W) e município de Itatiba, SP (23°10’
a 23°15’ S, 45°45’ a 46°45’ W), em áreas fronteiriças à APA. A APA localiza-se na porção nordeste
do município de Campinas e está compreendida entre os rios Atibaia, que determina sua fronteira à
oeste, e Jaguari, que sinaliza a fronteira a leste da APA e também do município de Campinas (Figura
1). Os fragmentos estudados pertencem a propriedades particulares. O clima, segundo Köppen
(1948), é Cwag'- temperado, moderadamente chuvoso, de inverno seco não rigoroso, com
precipitação média anual de 1409 mm (Mello et al. 1994).
Os solos ocupados pelos fragmentos em estudo podem ser Podzólicos Vermelho-Amarelos de
três diferentes tipos: PVA 8, PVA 20 e PVA 22 ou Latossolo Vermelho: LV 4. Os relevos podem ser
suave ondulados (SO), forte ondulados e montanhosos (FOM) ou ondulados e suave ondulados
(OSO) (Oliveira et al. 1999) (Tabela 1).
Foram escolhidos dez fragmentos com áreas inferiores a 100ha e um fragmento de 244,9ha, a
Mata Ribeirão Cachoeira, cujo levantamento florístico foi realizado por Santos & Kinoshita (2003). Os
fragmentos menores do que 100ha foram categorizados em três classes de tamanho: quatro
pequenos, com áreas entre 12 e 13,5ha (P1, P2, P3 e P4), quatro médios, com áreas entre 19 e
25,5ha (M1, M2, M3 e M4) e dois grandes, G1 e G2 com áreas de 41,8 e 63,3ha, respectivamente
(Tabela 1). A vegetação dos fragmentos pode ser classificada como floresta estacional semidecídua
(Veloso et al. 1991) e os critérios para a escolha de cada área foram: similaridade fisionômica,
facilidade de acesso e obtenção de autorização dos proprietários para a realização do trabalho.
Para tentar construir um histórico dos fragmentos estudados foram consultadas fotografias
aéreas da região realizadas desde a década de 60 (Instituto Agronômico; Instituto Base –
Aerofotometria e Projetos/SA). Pôde-se perceber que a maioria dos fragmentos mantêm sua forma e
tamanho atuais há pelo menos 40 anos, com exceção da mata P4, que vem aumentando em área
nos últimos 20 anos e, segundo os proprietários, trata-se de uma área onde houve regeneração
natural. Os proprietários dessas áreas forneceram ainda algumas informações sobre a ocorrência de
fatores de perturbação como queimadas e extração de madeira (Tabela 1, ver também Capítulo 1).
A paisagem atual constitui-se de um mosaico de culturas anuais e perenes, pastagens,
reflorestamentos e pequenos assentamentos humanos. A agricultura é caracterizada por pequenas
áreas de café, laranja, cana de açúcar e culturas de subsistência. Os reflorestamentos com eucalipto
e pastagens ocupam grandes extensões (Miranda 1996).
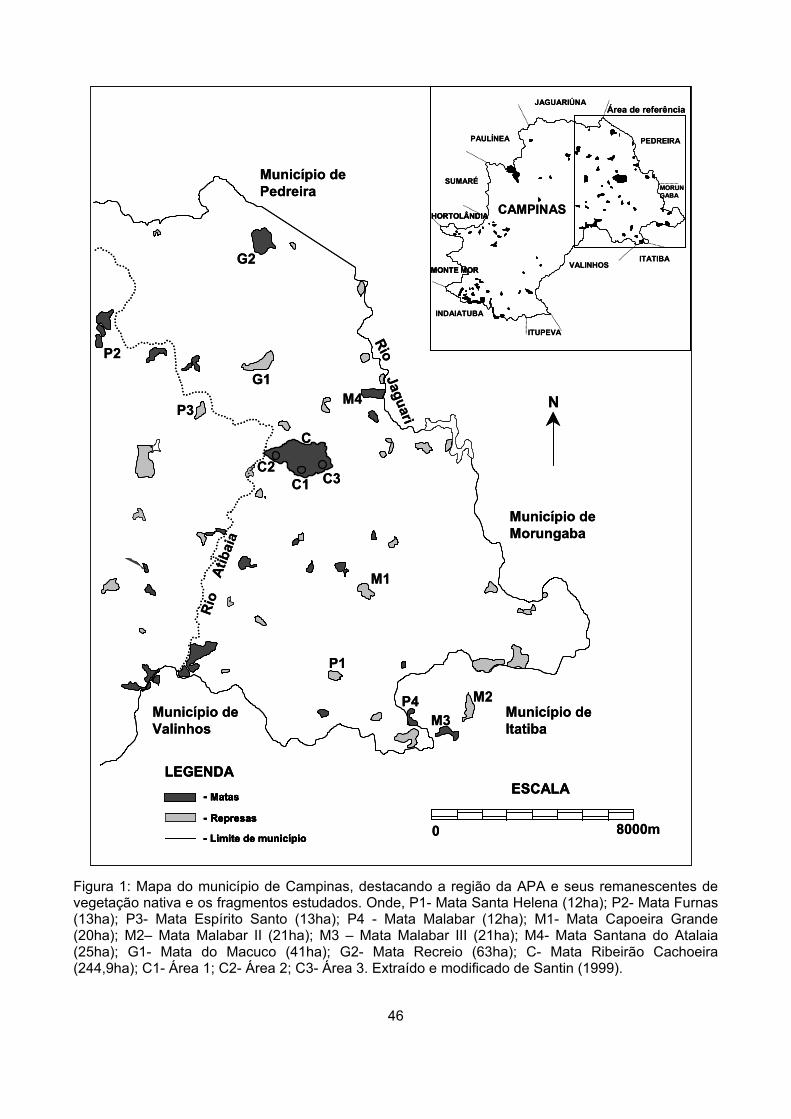
46
Figura 1: Mapa do município de Campinas, destacando a região da APA e seus remanescentes de vegetação nativa e os fragmentos estudados. Onde, P1- Mata Santa Helena (12ha); P2- Mata Furnas (13ha); P3- Mata Espírito Santo (13ha); P4 - Mata Malabar (12ha); M1- Mata Capoeira Grande (20ha); M2– Mata Malabar II (21ha); M3 – Mata Malabar III (21ha); M4- Mata Santana do Atalaia (25ha); G1- Mata do Macuco (41ha); G2- Mata Recreio (63ha); C- Mata Ribeirão Cachoeira (244,9ha); C1- Área 1; C2- Área 2; C3- Área 3. Extraído e modificado de Santin (1999).
Município de Morungaba
Município de Pedreira
Município de Valinhos
ESCALA
0 8000m
N
Rio
Atib
aia
M4
C
C3C1C2
G1
P3
G2
P2
M1
P1
P4M3
M2
HORTOLÂNDIA
MONTE MOR
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
LEGENDA- Matas
- Limite de município
- Represas
Município de Itatiba
Jaguari
Rio
Município de Morungaba
Município de Pedreira
Município de Valinhos
ESCALAESCALA
0 8000m
N
Rio
Atib
aia
M4
C
C3C1C2
G1
P3
G2
P2
M1
P1
P4M3
M2
HORTOLÂNDIA
MONTE MOR
HORTOLÂNDIA
MONTE MOR
SUMARÉ
ITATIBAVALINHOS
CAMPINAS
PEDREIRA
MORUNGABA
Área de referênciaJAGUARIÚNA
PAULÍNEA
ITUPEVA
INDAIATUBA
LEGENDA- Matas
- Limite de município
- Represas
LEGENDA- Matas
- Limite de município
- Represas
- Matas
- Limite de município
- Represas
Município de Itatiba
Jaguari
Rio
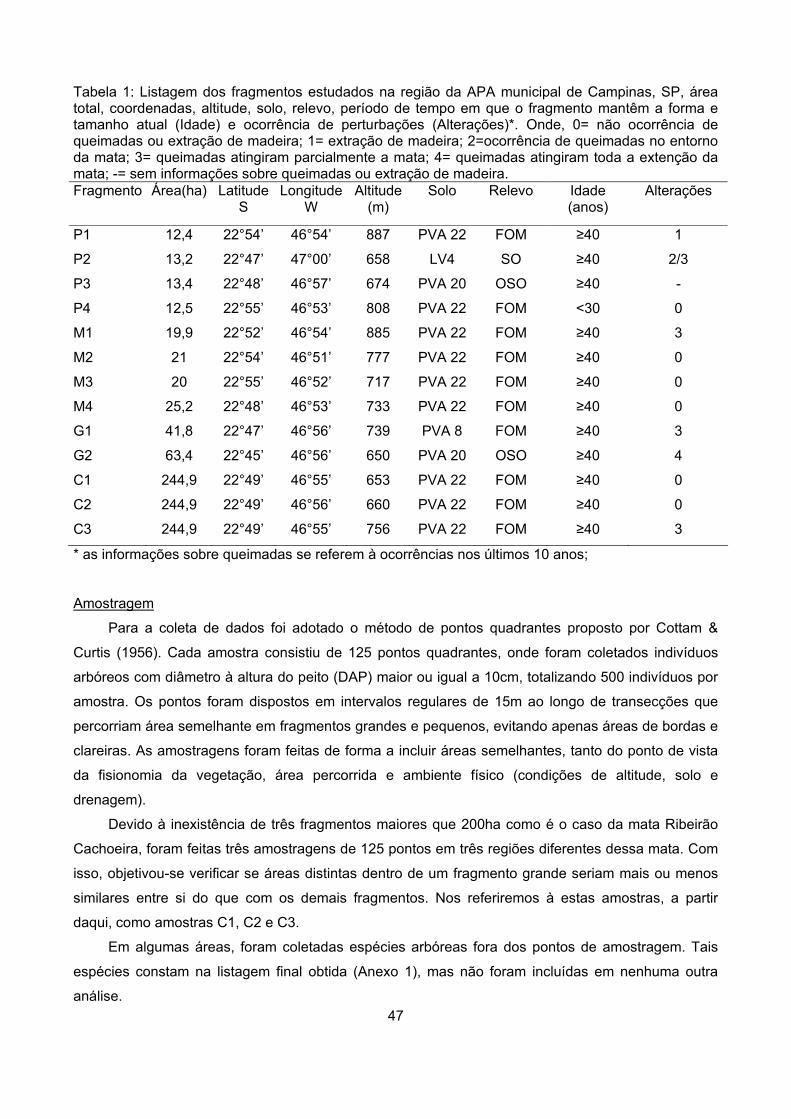
47
Tabela 1: Listagem dos fragmentos estudados na região da APA municipal de Campinas, SP, área total, coordenadas, altitude, solo, relevo, período de tempo em que o fragmento mantêm a forma e tamanho atual (Idade) e ocorrência de perturbações (Alterações)*. Onde, 0= não ocorrência de queimadas ou extração de madeira; 1= extração de madeira; 2=ocorrência de queimadas no entorno da mata; 3= queimadas atingiram parcialmente a mata; 4= queimadas atingiram toda a extenção da mata; -= sem informações sobre queimadas ou extração de madeira. Fragmento Área(ha) Latitude
S Longitude
W Altitude
(m) Solo Relevo Idade
(anos) Alterações
P1 12,4 22°54’ 46°54’ 887 PVA 22 FOM ≥40 1
P2 13,2 22°47’ 47°00’ 658 LV4 SO ≥40 2/3
P3 13,4 22°48’ 46°57’ 674 PVA 20 OSO ≥40 -
P4 12,5 22°55’ 46°53’ 808 PVA 22 FOM <30 0
M1 19,9 22°52’ 46°54’ 885 PVA 22 FOM ≥40 3
M2 21 22°54’ 46°51’ 777 PVA 22 FOM ≥40 0
M3 20 22°55’ 46°52’ 717 PVA 22 FOM ≥40 0
M4 25,2 22°48’ 46°53’ 733 PVA 22 FOM ≥40 0
G1 41,8 22°47’ 46°56’ 739 PVA 8 FOM ≥40 3
G2 63,4 22°45’ 46°56’ 650 PVA 20 OSO ≥40 4
C1 244,9 22°49’ 46°55’ 653 PVA 22 FOM ≥40 0
C2 244,9 22°49’ 46°56’ 660 PVA 22 FOM ≥40 0
C3 244,9 22°49’ 46°55’ 756 PVA 22 FOM ≥40 3
* as informações sobre queimadas se referem à ocorrências nos últimos 10 anos;
Amostragem
Para a coleta de dados foi adotado o método de pontos quadrantes proposto por Cottam &
Curtis (1956). Cada amostra consistiu de 125 pontos quadrantes, onde foram coletados indivíduos
arbóreos com diâmetro à altura do peito (DAP) maior ou igual a 10cm, totalizando 500 indivíduos por
amostra. Os pontos foram dispostos em intervalos regulares de 15m ao longo de transecções que
percorriam área semelhante em fragmentos grandes e pequenos, evitando apenas áreas de bordas e
clareiras. As amostragens foram feitas de forma a incluir áreas semelhantes, tanto do ponto de vista
da fisionomia da vegetação, área percorrida e ambiente físico (condições de altitude, solo e
drenagem).
Devido à inexistência de três fragmentos maiores que 200ha como é o caso da mata Ribeirão
Cachoeira, foram feitas três amostragens de 125 pontos em três regiões diferentes dessa mata. Com
isso, objetivou-se verificar se áreas distintas dentro de um fragmento grande seriam mais ou menos
similares entre si do que com os demais fragmentos. Nos referiremos à estas amostras, a partir
daqui, como amostras C1, C2 e C3.
Em algumas áreas, foram coletadas espécies arbóreas fora dos pontos de amostragem. Tais
espécies constam na listagem final obtida (Anexo 1), mas não foram incluídas em nenhuma outra
análise.

48
A maior parte das coletas foi realizada durante o período de abril a setembro de 2000. As
identificações foram feitas segundo técnicas usuais em taxonomia e os espécimes foram incluídos em
famílias segundo o sistema proposto por Cronquist (1981). A coleção principal de material
herborizado foi incorporada ao herbário UEC e as duplicatas foram enviadas a outros herbários ou
especialistas que ajudaram na identificação de alguns grupos. Os materiais coletados em estado
vegetativo foram conservados como material testemunha, em separado.
Síndromes de dispersão
Para verificar se a proporção de indivíduos e espécies zoocóricas, anemocóricas e autocóricas
diferia entre as amostras e estava relacionado ao tamanho dos fragmentos, as espécies foram
agrupadas por síndromes de dispersão. Foram consideradas espécies zoocóricas aquelas que
produzem frutos carnosos (baga ou drupa), sementes ariladas ou diásporos com adaptações que
permitam dispersão por animais; anemocóricas, as espécies cujos diásporos apresentam formas,
estruturas ou dimensões que permitem a dispersão pelo vento; e autocóricas, as espécies com
diásporos que não apresentam nenhuma das características indicadas acima (Mantovani & Martins
1988). As espécies que não foram encontradas com frutos tiveram o tipo de síndrome de dispersão
determinada através de verificação em herbário e/ou indicações bibliográficas.
Tratamento dos dados
Análises de similaridade servem para detectar relações entre as comunidades e ambientes
através dos agrupamentos formados segundo as variáveis analisadas (Van Tongeren 1987). Para
verificar se a composição florística de fragmentos de mesmo tamanho é mais homogênea entre si do
que entre as demais classes de tamanho foi adotado o índice de similaridade de Jaccard (Pielou
1984). As relações de similaridade entre os fragmentos foram investigadas através da análise de
agrupamento UPGMA, utilizando-se o programa FITOPAC (Shepherd 1988). O índice de Jaccard foi
escolhido por ser um índice simples, amplamente usado e por dar menos peso para as espécies
comuns entre as áreas, enfatizando as diferenças entre elas (Magurran 1988).
Para verificar se a proporção de espécies e indivíduos das diferentes síndromes de dispersão
diferia entre as amostras, as proporções de cada síndrome nas amostras foi comparada por meio da
construção de tabelas de contigência, uma contendo o número de espécies categorizadas por
amostra e outra com as abundâncias das referidas espécies. Foram feitas comparações, através do
teste de Qui-quadrado com correção de Yates (Zar 1989), entre os número esperados e observados
nos diferentes fragmentos. A ocorrência da relação entre o tamanho dos fragmentos e a proporção de
espécies e indivíduos das diferentes sídromes de dispersão foi investigada através de análises de
correlação de Pearson (Zar 1989), utilizando-se o programa BioEstat 2.0 (Ayres et al. 1998). Em tais
análises consideramos apenas um valor médio das amostras de Ribeirão Cachoeira, pois se tratavam
de três amostras realizadas em um mesmo fragmento.

49
Resultados Aspectos florísticos
Dentre os 6500 indivíduos coletados, foram encontradas 248 espécies (incluindo morfo-
espécies), 156 gêneros e 58 famílias (Tabela 2). Dentre as morfo-espécies, 13 táxons foram
identificados até o nível de gênero e um permaneceu sem nenhuma identificação. Duas espécies
permaneceram a conferatum (cf.) por falta de material reprodutivo para a confirmação das
identificações. Considerando apenas as plantas identificadas ao nível de espécie, observou-se um
total de 232 taxa (Anexo I). Mais trinta espécies e morfo-espécies foram coletadas fora dos pontos de
amostragem. Destas, vinte e cinco foram identificadas ao nível de espécie. Somando-se estas últimas
ao total coletado na amostragem temos 257 taxa identificados no nível de espécie (Anexo 1).
As espécies que ocorreram em somente uma amostra totalizaram 53 espécies ou 21,7% do
total de espécies (Tabela 2). Entre elas está incluída Ficus insipida, espécie apontada por Santin
(1999) como “ameaçada de extinção” no município. Quase metade das espécies exclusivas (40%)
raramente figuram em levantamentos florísticos do interior do estado de São Paulo e/ou estão sendo
citadas pela primeira vez para a região de Campinas (Anexo 1). Os fragmentos com maior número de
espécies exclusivas foram M3 com nove espécies e P4, M2 e M4 com sete espécies cada.
Seis espécies, Astronium graveolens, Centrolobium tomentosum, Cariniana legalis, Piptadenia
gonoacantha, Maclura tinctoria e Seguieria langsdorffi ocorreram em todas as amostras,
representando 2,4% do total amostrado.
As famílias mais ricas foram Myrtaceae (25 espécies), Fabaceae (20), Lauraceae (18),
Euphorbiaceae (15), Meliaceae e Rutaceae (12), Moraceae (11), Caesalpiniaceae, Mimosaceae e
Rubiaceae (8) (Tabela 3). As famílias mais ricas contribuíram com a maioria (55,2%) das espécies
coletadas, enquanto a maioria das famílias (73%) contribuíram com uma pequena proporção das
espécies (27%).
As famílias mais ricas entre as amostras foram, em geral, as mesmas (Tabela 3), mas as
amostras dos fragmentos G2 e, principalmente, P3 apresentaram algumas variações. As famílias
Lauraceae e Euphorbiaceae estiveram entre as famílias mais ricas na maioria das amostras, mas não
ocorreram no fragmento P3 e apresentaram apenas duas espécies cada na área G2.

50
Tabela 2: Aspectos florísticos dos fragmentos estudados na região da APA municipal de Campinas, SP e também para o conjunto das áreas. Constam número de indivíduos coletados, número de famílias, gêneros e espécies encontrados e número de espécies que ocorreram em apenas uma amostra (Exclusivas). Locais Indivíduos Famílias Gêneros Espécies Exclusivas
P1 500 39 79 103 6
P2 500 35 68 85 4
P3 500 25 38 47 3
P4 500 40 73 99 7
M1 500 36 72 98 0
M2 500 40 82 107 7
M3 500 36 84 110 9
M4 500 36 73 97 7
G1 500 38 80 104 2
G2 500 33 56 65 1
C1 500 39 75 97 3
C2 500 38 75 98 3
C3 500 40 73 90 1
Total 6500 58 156 248 53
P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
Tabela 3: Número de espécies das famílias mais ricas nos fragmentos estudados na região da APA municipal de Campinas, SP. Família P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 Total
Myrtaceae 7 7 3 8 8 12 11 9 8 6 7 5 6 25
Fabaceae 6 5 6 10 7 7 10 11 8 5 7 9 4 20
Lauraceae 7 8 0 9 5 8 8 7 9 2 7 7 8 18
Euphorbiaceae 9 9 0 4 5 4 7 3 6 2 5 7 8 15
Meliaceae 7 4 4 4 6 5 6 6 7 4 7 8 7 12
Rutaceae 7 5 2 4 7 3 5 5 7 5 8 8 5 12
Moraceae 5 4 2 6 5 3 2 4 3 2 4 3 3 11
Caesalpiniaceae 3 3 4 4 5 5 5 6 4 3 4 4 5 8
Mimosaceae 3 4 2 4 3 5 5 4 5 3 5 4 4 8
Rubiaceae 2 2 1 3 3 6 6 3 3 2 5 1 4 8
P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
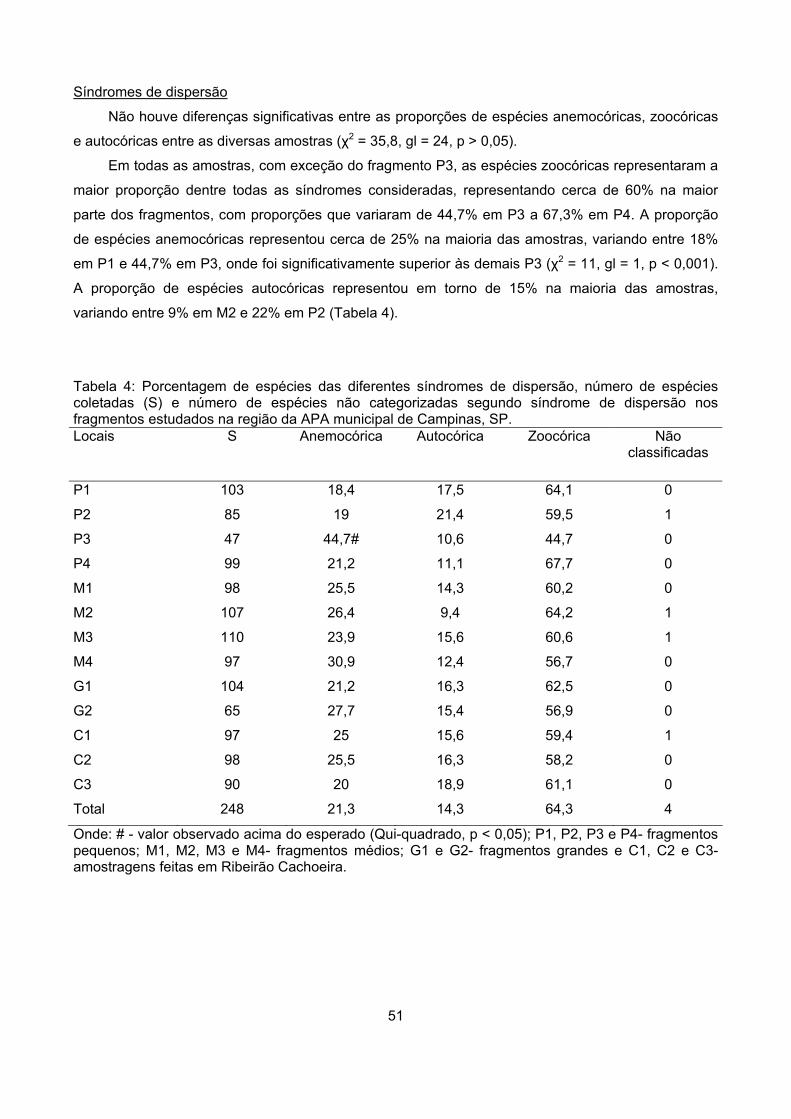
51
Síndromes de dispersão
Não houve diferenças significativas entre as proporções de espécies anemocóricas, zoocóricas
e autocóricas entre as diversas amostras (χ2 = 35,8, gl = 24, p > 0,05).
Em todas as amostras, com exceção do fragmento P3, as espécies zoocóricas representaram a
maior proporção dentre todas as síndromes consideradas, representando cerca de 60% na maior
parte dos fragmentos, com proporções que variaram de 44,7% em P3 a 67,3% em P4. A proporção
de espécies anemocóricas representou cerca de 25% na maioria das amostras, variando entre 18%
em P1 e 44,7% em P3, onde foi significativamente superior às demais P3 (χ2 = 11, gl = 1, p < 0,001).
A proporção de espécies autocóricas representou em torno de 15% na maioria das amostras,
variando entre 9% em M2 e 22% em P2 (Tabela 4).
Tabela 4: Porcentagem de espécies das diferentes síndromes de dispersão, número de espécies coletadas (S) e número de espécies não categorizadas segundo síndrome de dispersão nos fragmentos estudados na região da APA municipal de Campinas, SP. Locais S Anemocórica Autocórica Zoocórica Não
classificadas
P1 103 18,4 17,5 64,1 0
P2 85 19 21,4 59,5 1
P3 47 44,7# 10,6 44,7 0
P4 99 21,2 11,1 67,7 0
M1 98 25,5 14,3 60,2 0
M2 107 26,4 9,4 64,2 1
M3 110 23,9 15,6 60,6 1
M4 97 30,9 12,4 56,7 0
G1 104 21,2 16,3 62,5 0
G2 65 27,7 15,4 56,9 0
C1 97 25 15,6 59,4 1
C2 98 25,5 16,3 58,2 0
C3 90 20 18,9 61,1 0
Total 248 21,3 14,3 64,3 4
Onde: # - valor observado acima do esperado (Qui-quadrado, p < 0,05); P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.

52
Quando consideramos o número de indivíduos das diferentes síndromes observamos grande
variação. As proporções das três síndromes diferiram entre os fragmentos: anemocóricas (χ2 = 82,6,
gl = 12, p < 0,001), autocóricas (χ2 = 296,1, gl = 12, p < 0,001) e zoocóricas (χ2 = 112,6, gl = 12, p <
0,001).
A proporção de indivíduos de espécies zoocóricas variou de 34 a 69% entre as amostras e
representou a maior proporção dos indivíduos em todas elas, com exceção de P2 e C3. Nas
amostras P1, P3 e M3 os indivíduos zoocóricos apresentaram proporção significativamente superior
às demais (P1- χ2 = 8,4, gl = 1, p = 0,004; P3- χ2 = 58,7, gl = 1, p < 0,001; M3- χ2 = 4,3, gl = 1, p =
0,04). Nas amostras P2, G2, C1 e C3 a proporção de indivíduos zoocóricos foi inferior às demais (P2-
χ2 = 14,2, gl = 1, p < 0,001; G2- χ2 = 7,1, gl = 1, p = 0,008; C1- χ2 = 5,3, gl = 1, p < 0,05; C3- χ2 = 9,8,
gl = 1, p = 0,002) (Figura 2).
Os indivíduos anemocóricos representaram de 16 a 37% do total coletado. Nas amostras P4,
G2 e C1 os indivíduos anemocóricos apresentaram proporção significativamente superior às demais
(P4- χ2 = 7,6, gl = 1, p < 0,05; G2- χ2 = 21,8, gl = 1, p < 0,001; C1- χ2 = 5,9, gl = 1, p < 0,05), enquanto
nas amostras P2, M2 e C3 a proporção de indivíduos anemocóricos foi inferior às outras áreas (P2- χ2
= 22,8, gl = 1, p < 0,001; M2- χ2 = 5,9, gl = 1, p < 0,05; C3- χ2 = 6,9, gl = 1, p < 0,05) (Figura 2).
A proporção de indivíduos autocóricos foi a que mais variou entre as amostras, representando
de 1 a 43% dos indivíduos coletados. Nas amostras P2, M4 e C3 a proporção de indivíduos
autocóricos foi significativamente superior, sendo que em P2 e C3 representou quase o dobro da
observada na maioria das amostras (P2- χ2 = 87,4, gl = 1 p < 0,001; C3- χ2 = 41,4, gl = 1, p < 0,001;
M4- χ2 = 4,9, gl = 1, p < 0,05). Já nas amostras P1, P4 e P3 essa proporção foi significativamente
inferior, chegando a representar apenas 1% dos indivíduos em P3 (χ2 = 128,7, gl = 1, p < 0,001), (P1-
χ2 = 8,3, gl = 1, p < 0,001; P4- χ2 = 14,5, gl = 1, p < 0,001) (Figura 2).
As variações observadas quanto ao número de indivíduos das diferentes síndromes não
estiveram relacionadas ao tamanho dos fragmentos – anemocóricos (r = 0,188, p = 0,579);
autocóricos (r = 0,147, p = 0,667); zoocóricos (r = -0,295, p = 0,379).
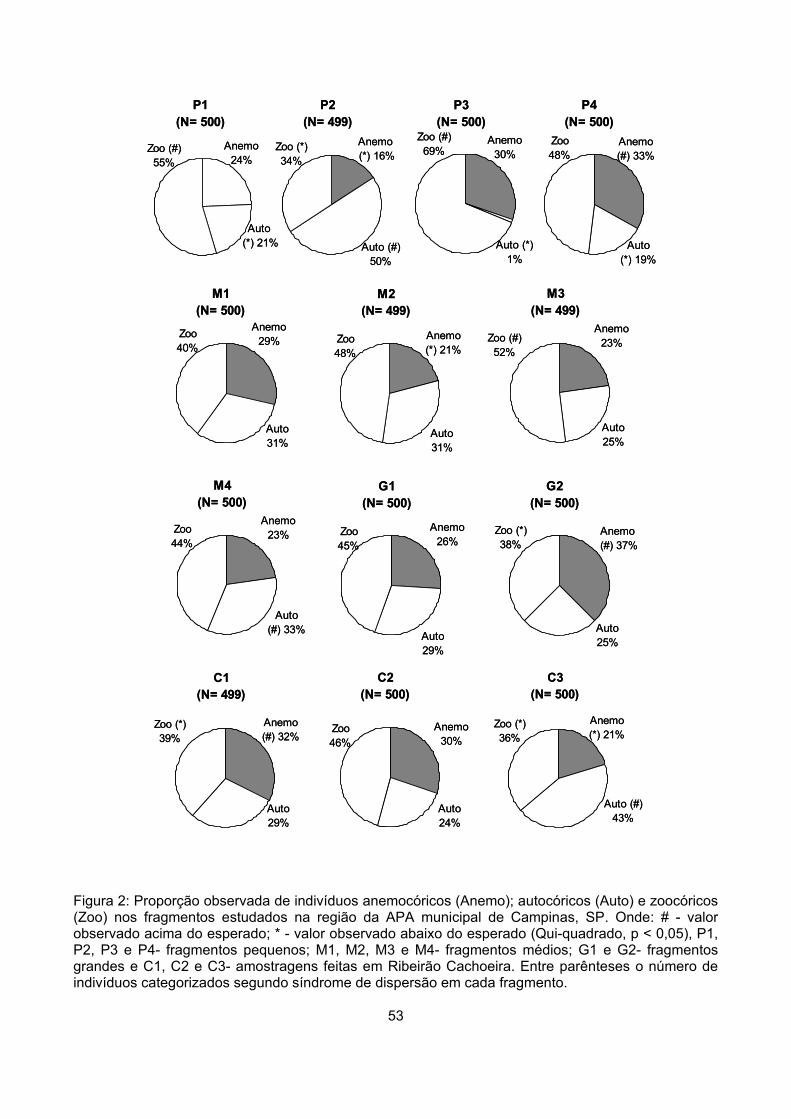
53
Figura 2: Proporção observada de indivíduos anemocóricos (Anemo); autocóricos (Auto) e zoocóricos (Zoo) nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05), P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira. Entre parênteses o número de indivíduos categorizados segundo síndrome de dispersão em cada fragmento.
M3 (N= 499)
Anemo23%
Auto25%
Zoo (#)52%
M2 (N= 499)
Auto31%
Anemo(*) 21%
Zoo48%
G1 (N= 500)
Auto29%
Anemo26%
Zoo45%
G2 (N= 500)
Zoo (*)38%
Auto25%
Anemo (#) 37%
P1 (N= 500)
Auto (*) 21%
Anemo24%
Zoo (#)55%
P2 (N= 499)
Zoo (*)34%
Auto (#)50%
Anemo (*) 16%
P3 (N= 500)
Anemo30%
Zoo (#)69%
Auto (*)1%
P4 (N= 500)
Auto(*) 19%
Anemo(#) 33%
Zoo48%
M1 (N= 500)
Auto31%
Anemo29%
Zoo40%
M4(N= 500)
Anemo23%
Auto (#) 33%
Zoo44%
C1 (N= 499)
Zoo (*)39%
Anemo(#) 32%
Auto29%
C2 (N= 500)
Anemo30%
Zoo46%
Auto24%
C3 (N= 500)
Anemo(*) 21%
Zoo (*)36%
Auto (#)43%
M3 (N= 499)
Anemo23%
Auto25%
Zoo (#)52%
M2 (N= 499)
Auto31%
Anemo(*) 21%
Zoo48%
G1 (N= 500)
Auto29%
Anemo26%
Zoo45%
G2 (N= 500)
Zoo (*)38%
Auto25%
Anemo (#) 37%
P1 (N= 500)
Auto (*) 21%
Anemo24%
Zoo (#)55%
P2 (N= 499)
Zoo (*)34%
Auto (#)50%
Anemo (*) 16%
P3 (N= 500)
Anemo30%
Zoo (#)69%
Auto (*)1%
P4 (N= 500)
Auto(*) 19%
Anemo(#) 33%
Zoo48%
M1 (N= 500)
Auto31%
Anemo29%
Zoo40%
M4(N= 500)
Anemo23%
Auto (#) 33%
Zoo44%
C1 (N= 499)
Zoo (*)39%
Anemo(#) 32%
Auto29%
C2 (N= 500)
Anemo30%
Zoo46%
Auto24%
C3 (N= 500)
Anemo(*) 21%
Zoo (*)36%
Auto (#)43%
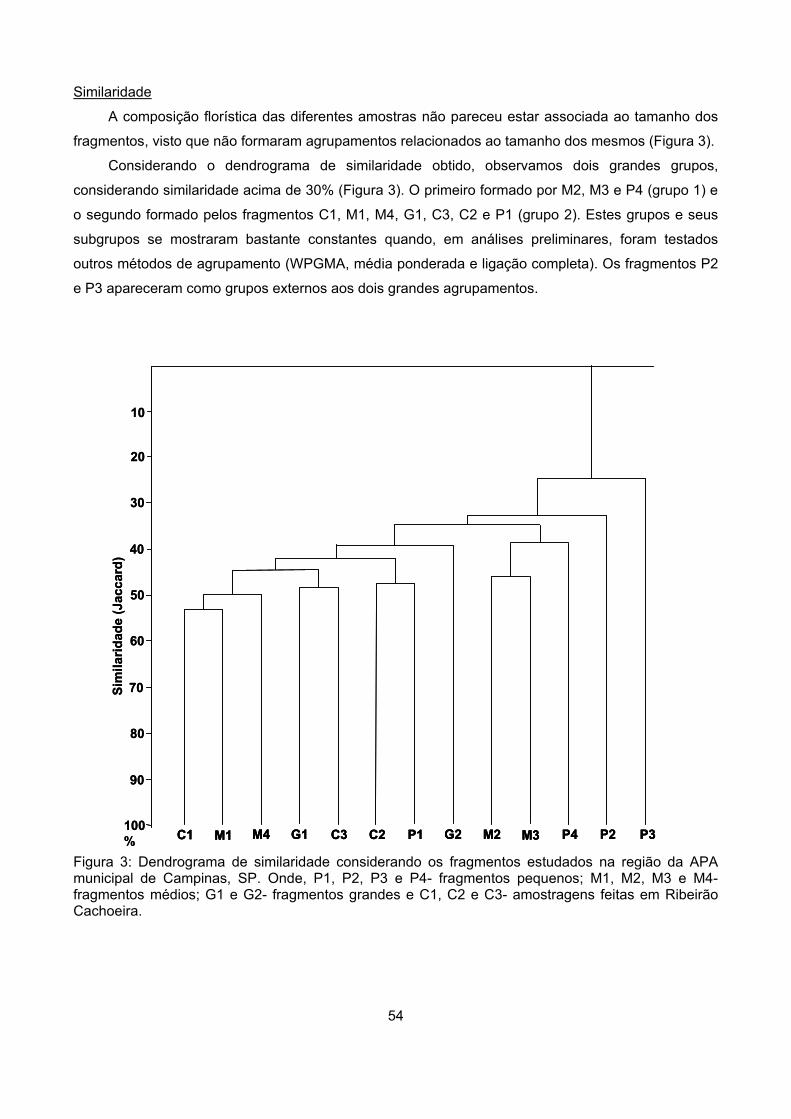
54
Similaridade
A composição florística das diferentes amostras não pareceu estar associada ao tamanho dos
fragmentos, visto que não formaram agrupamentos relacionados ao tamanho dos mesmos (Figura 3).
Considerando o dendrograma de similaridade obtido, observamos dois grandes grupos,
considerando similaridade acima de 30% (Figura 3). O primeiro formado por M2, M3 e P4 (grupo 1) e
o segundo formado pelos fragmentos C1, M1, M4, G1, C3, C2 e P1 (grupo 2). Estes grupos e seus
subgrupos se mostraram bastante constantes quando, em análises preliminares, foram testados
outros métodos de agrupamento (WPGMA, média ponderada e ligação completa). Os fragmentos P2
e P3 apareceram como grupos externos aos dois grandes agrupamentos.
Figura 3: Dendrograma de similaridade considerando os fragmentos estudados na região da APA municipal de Campinas, SP. Onde, P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
Sim
ilarid
ade
(Jac
card
)
100%
20
30
40
50
60
70
80
90
10
C1 M1 M4 G1 C3 M2C2 P1 G2 M3 P4 P2 P3
Sim
ilarid
ade
(Jac
card
)
100%
20
30
40
50
60
70
80
90
10
C1 M1 M4 G1 C3 M2C2 P1 G2 M3 P4 P2 P3C1 M1 M4 G1 C3 M2C2 P1 G2 M3 P4 P2 P3

55
Dentro do grupo 1, as concordâncias variaram entre 37 e 47%. A maior similaridade observada
dentro deste grupo se deu entre as matas M2 e M3 (47%) (Tabela 6). Em geral a similaridade das
matas do grupo 1 com as matas do grupo 2, se manteve inferior a 35%, apesar da uniformidade de
solo e relevo entre estas áreas (Tabela 1), mas alcançou valores iguais ou superiores a 40% com os
fragmentos P1, M1 e C1. A formação do grupo 1 pode estar associada à proximidade geográfica
entre as três matas que compõem o grupo. Corrobora com essa hipótese o fato de as matas P1 e M1
do grupo 2, também bastante próximas geograficamente, serem as que exibiram os maiores graus de
concordância com as matas do grupo 1, chegando a 48% entre M3 e M1 (Tabela 5).
As similaridades observadas entre as amostras do grupo 2 variaram entre 39 e 54%, com as
maiores concordâncias ocorrendo entre M1 e C1, com 54%, a maior de toda a análise, seguida por
M1 e M4 com 51% e, entre C1 e C3 com 50% (Tabela 5). Todas as matas que compõem este
agrupamento apresentam o mesmo tipo de relevo (forte ondulado e montanhoso) e solo (Pva 22),
com exceção de G1 com solo tipo Pva 8 (Tabela 1).
O fragmento G2 apareceu externamente ao grupo 2, com o qual exibe concordâncias entre 37 e
44% e similaridade entre 24 e 30% com as amostras do grupo 1. A mata G2 apresentou cerca de
70% da riqueza média observada nas demais amostras. Além da menor riqueza, que poderia
influenciar nos coeficientes obtidos, fatores como solo, relevo e histórico de preservação difereciam
esta mata das demais pertencentes ao grupo 2. Segundo informações locais, uma grande queimada
atingiu a mata quase por completo em 1996 (Tabela 1).
A mata P3, que figurou como um grupo externo a todos os demais, exibiu similaridades entre 23
e 29% com as amostras do grupo 1, de 18 a 22% com as amostras do grupo 2 e de 18% em relação
ao grupo externo formado por P2. O maior grau de concordância, considerando a mata P3, ocorreu
em relação a amostra G2 (38%). Essas duas matas foram as únicas do estudo a ocuparem relevo
ondulado e suave ondulado e solo tipo PVA 20 (Tabela 1). Além da divergência de solo e relevo com
as matas dos grupos 1 e 2, o número de espécies coletadas em P3 foi cerca da metade coletada nas
demais áreas, o que poderia influenciar nos valores de similaridade obtidos.
A mata P2 também apareceu isolada dos dois grandes grupos formados. Essa mata apresenta
solo e relevo distintos de todos os demais fragmentos, além de histórico de perturbação intenso
(Tabela 1). Segundo informações locais e observações in loco, as queimadas nos arredores da mata
são constantes e já atingiram parcialmente a mata. Ainda assim, a amostra exibiu similaridade
superior a 35% com algumas matas do grupo 2, como M1 e C1 (38%) e M4 (39%). Em relação ao
grupo 1, a similaridade também foi alta, variando entre 32 e 36%., Talvez por isso, se alterarmos o
método de agrupamento, a mata P2 apareça incluída neste grupo.
A proximidade geográfica não explica todos os agrupamentos formados, mas é interessante
apontar que a mata M1, que exibiu similaridades superiores a 40% com a maioria das demais,
encontra-se em um ponto eqüidistante entre a maioria das matas que compõem os dois
agrupamentos (Tabela 5). A proximidade geográfica também poderia estar envolvida no caso das

56
amostras feitas dentro de Ribeirão Cachoeira, que, em alguns casos, exibiram até 50% de
similaridade. Para testar esta hipótese foi feita uma análise de regressão linear para tentar relacionar
a distância entre os fragmentos e a similaridade obtida (os valores do coeficiente de jaccard foram
transformados para arcoseno da raiz quadrada e o valor de p foi ajustado utilizando-se correção de
Bonferroni, pois tratavam-se de comparações múltiplas, Zar 1989). Segundo essa hipótese
fragmentos mais próximos deveriam exibir similaridades mais altas. Observamos uma relação entre a
proximidade geográfica e a similaridade entre os fragmentos (r2 = 0,1449, p < 0,0042) (Figura 4),
indicando que, apesar da variação, quanto maior a proximidade, maior a similaridade entre os
fragmentos.
Tabela 5: Distância (Km) e similaridade florística (Jaccard) entre os fragmentos florestais estudados na região da APA municipal de Campinas, SP. Acima da diagonal constam os valores de similaridade e abaixo as distâncias entre os fragmentos em quilômetros. Locais P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
P1 - 0,28 0,27 0,36 0,48 0,40 0,36 0,35 0,47 0,37 0,39 0,48 0,40
P2 15,70 - 0,18 0,32 0,38 0,34 0,36 0,39 0,32 0,22 0,39 0,33 0,34
P3 11,18 4,53 - 0,18 0,27 0,22 0,22 0,26 0,26 0,38 0,26 0,29 0,23
P4 3,03 18,69 13,94 - 0,39 0,37 0,41 0,31 0,35 0,24 0,32 0,37 0,29
M1 3,24 13,92 9,17 4,44 - 0,43 0,48 0,51 0,47 0,41 0,54 0,45 0,42
M2 4,91 19,91 15,10 1,50 5,19 - 0,47 0,35 0,35 0,30 0,41 0,34 0,31
M3 4,24 20,08 15,37 0,87 5,55 0,60 - 0,37 0,36 0,29 0,40 0,35 0,33
M4 10,84 10,47 6,24 12,02 7,32 12,21 13,13 - 0,41 0,36 0,49 0,36 0,41
G1 12,19 5,46 2,08 14,22 9,31 15,11 15,59 3,78 - 0,44 0,48 0,47 0,49
G2 16,57 6,87 6,18 18,32 13,63 19,03 19,81 6,62 3,79 - 0,40 0,42 0,38
C1 7,70 7,75 2,93 9,76 4,68 10,58 11,01 2,52 3,10 7,53 - 0,46 0,50
C2 8,30 7,05 2,23 10,31 5,38 11,35 11,82 3,19 2,52 6,96 0,70 - 0,39
C3 8,10 8,49 3,21 9,56 4,45 10,31 10,76 2,29 3,26 7,71 0,80 1,50 -
Onde, P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira.
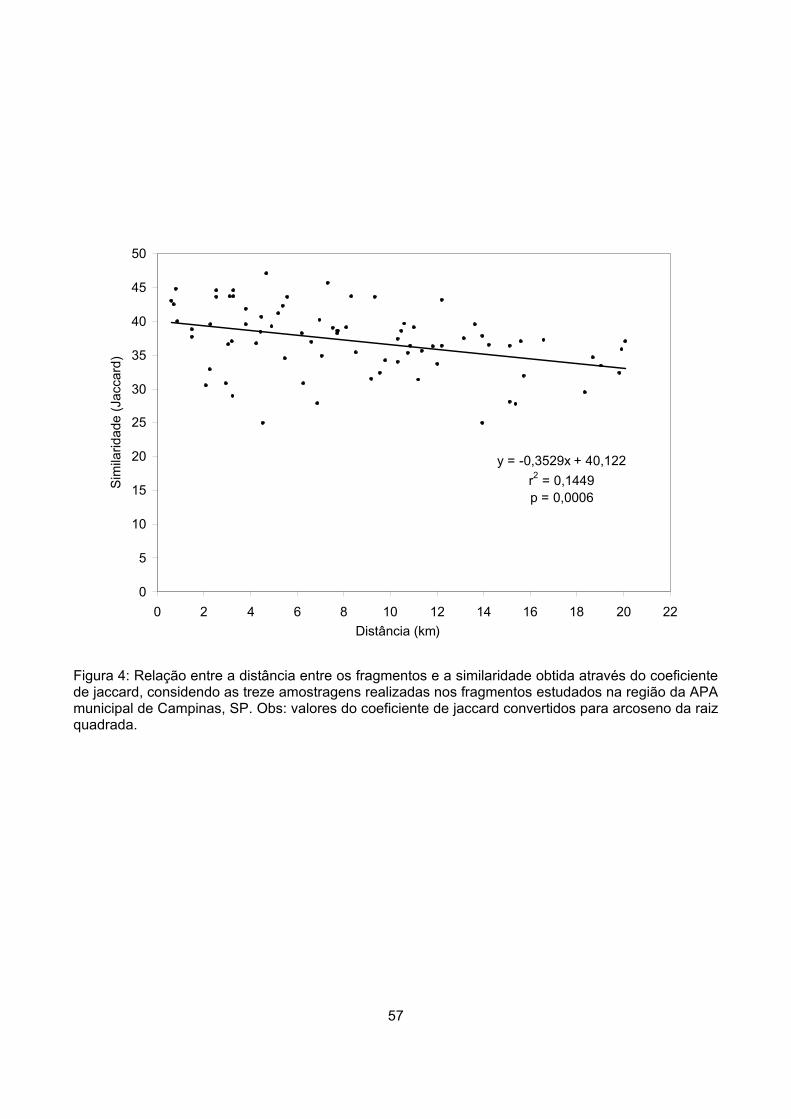
57
Figura 4: Relação entre a distância entre os fragmentos e a similaridade obtida através do coeficiente de jaccard, considendo as treze amostragens realizadas nos fragmentos estudados na região da APA municipal de Campinas, SP. Obs: valores do coeficiente de jaccard convertidos para arcoseno da raiz quadrada.
y = -0,3529x + 40,122r2 = 0,1449p = 0,0006
0
5
10
15
20
25
30
35
40
45
50
0 2 4 6 8 10 12 14 16 18 20 22Distância (km)
Sim
ilarid
ade
(Jac
card
)

58
Discussão A maioria das famílias mais ricas encontradas no presente estudo tem sido apontada como tal
na maioria dos levantamentos florísticos de espécies arbóreas realizados no interior do estado de
São Paulo. A família Myrtaceae, por exemplo, foi mencionada como a mais rica nos trabalhos de
Matthes et al. (1988), Bernacci & Leitão Filho (1996), Costa & Mantovani (1995), Rodrigues et
al.(1989), Gandolfi et al. (1995), Salis et al. (1994), Metzger et al. (1997), Durigan et al. (2000), Cielo
Filho & Santin (2002) e Santos & Kinoshita (2003). As demais famílias também figuram, com alguma
variação de posição, entre as dez mais ricas nos levantamentos acima citados e nos de Cavassan et
al. (1984), Pagano & Leitão Filho (1987) e Bertoni & Martins (1987).
Outro aspecto aqui observado e freqüentemente mencionado por outros estudos deste tipo
florestal é que um pequeno número de famílias contribui com mais de 50% das espécies. Dessa
forma, parece ocorrer uma concentração de riqueza em algumas poucas famílias, que são, em geral,
as mesmas.
Grande parte das famílias observadas no presente estudo é muito constante nos estudos
citados acima e também em outros como os de Assumpção et al. (1982), Baitello et al. (1988),
Grombone et al. (1990) e Silva & Soares (2002). Assim, a composição de famílias aqui encontrada
pode ser considerada similar àquela observada nas demais florestas estacionais semidecíduas do
estado de São Paulo. Isto está de acordo com as observações de Torres et al. (1997), segundo as
quais, a similaridade entre florestas do estado de São Paulo é bastante alta, quando analisada em
nível de família.
A baixa variação entre os diversos estudos no conjunto das famílias mais ricas pode estar
refletindo não apenas que são características desse tipo de ecossistema como apontou Leitão Filho
(1992), mas também a distribuição das espécies em famílias no mundo. Essas famílias são, em sua
maioria, as mais ricas dentre as dicotiledôneas e/ou têm distribuição restrita às regiões tropicais ou
neotropicais, muitas vezes sendo estas regiões o seu centro de dispersão (Mabberley 1997, Marchiori
& Sobral 1997, Mori et al. 1983, Polhill et al. 1981, Ribeiro et al. 1999, Barroso 1986, Marchiori 2000,
Pirani & Skorupa 2002).
As famílias Fabaceae e Mimosaceae, por exemplo, pertencem a um dos grupos de plantas mais
diverso do mundo, as leguminosas, com 18.000 espécies (Polhill et al. 1981). As Fabaceae
compreendem cerca de 12.000 espécies e, apesar de sua ampla distribuição, um de seus principais
centros de diversificação é o Planalto do Brasil (Polhill et al. 1981). As Mimosaceae são
predominantemente pantropicais e apresentam 3.000 espécies (Ribeiro et al. 1999).
As Fabaceae representam 7% do total de espécies do mundo (Polhill et al. 1981) e, neste
estudo, contribuíram com 7% das espécies coletadas. Seria interessante um estudo mais
aprofundado para analisar se esta proporção também se mantém para as outras famílias
freqüentemente citadas como as mais ricas em florestas do estado de São Paulo. Entretanto, deve-se
levar em consideração a forma de vida das plantas em questão. As Fabaceae, por exemplo,

59
compreendem plantas herbáceas, trepadeiras e arbóreas. Por outro lado, as Myrtaceae, que detêm
apenas 2,7% das espécies do mundo (Mabberley 1997) e representaram neste estudo 10% do total
de espécies, são predominantemente arbóreas (Barroso 1986) e, apesar de distribuírem-se por todos
os continentes, à exceção da Antártida, exibem nítida predominância nas regiões tropicais e
subtropicais do mundo (Marchiori & Sobral 1997). Além disso, esta família é apontada por Mori et al.
(1983) como uma das mais importantes nas florestas do leste do Brasil, região que os autores
sugerem ser seu centro de diversificação. Estas divagações suscitam a seguinte questão: Será que
as famílias mais ricas em espécies arbóreas em São Paulo não seriam também as mais ricas em
espécies arbóreas do mundo ou regiões tropicais? ou teriam estas regiões como seus principais
centros de dispersão como as Rutaceae, Euphorbiaceae e Myrtaceae (Pirani & Skorupa 2002,
Marchiori 2000, Mori et al. 1983, Barroso 1986)?
As espécies que ocorreram em todos os fragmentos estão, para Santin (1999), entre “as mais
freqüentes” da região. Salis et al. (1994) apontou algumas delas como características das florestas
estacionais do interior do estado de São Paulo. Em concordância com essas afirmações observamos
que estas espécies figuram na maioria dos estudos florísticos de florestas estacionais realizados no
município de Campinas (Tamashiro et al. 1986, Matthes et al. 1988, Bernacci & Leitão Filho 1996,
Cielo Filho & Santin 2002, Santos & Kinoshita 2003) e de outras localidades como Piracicaba (Costa
& Mantovani 1995), Bauru (Cavassan et al. 1984), Rio Claro (Pagano & Leitão Filho 1987), Santa Rita
do Passa Quatro (Bertoni et al. 1988), Jundiaí (Rodrigues et al. 1989) e Gália (Durigan et al. 2000).
Outras 25 espécies (10% do total coletado) aparecem em pelo menos quinze levantamentos
florísticos realizados no estado de São Paulo de um total de 25 trabalhos consultados, que incluem os
estudos supra citados mais os estudos de Gibbs & Leitão Filho (1978), Tabanez et al. (1987), Rossi
(1987), Bertoni & Martins (1987), Baitello et al. (1988), Meira Neto et al. (1989), Silva (1989), Cezar &
Leitão Filho (1990), Salis et al. (1994), Gandolfi et al. (1995), Metzger et al. (1997), Dislich et al.
(2001) e Silva & Soares (2002). Essas espécies poderiam ser consideradas comuns ou constantes.
Por outro lado, 25% das espécies amostradas (65 espécies) raramente são citadas nesses
mesmos levantamentos. Além disso, dos 257 taxa identificados até o nível de espécie, 35 ou 13,6%
estão sendo citados pela primeira vez para a região de Campinas. Este número representa 7,7% do
total de 453 espécies arbóreas listadas para o município de Campinas por Santin (1999). Vale
lembrar, entretanto, que no referido trabalho, a autora incluiu as matas estacionais semidecíduas e
também os demais tipos vegetacionais que ocorrem no município, como Cerrados (sensu lato), matas
de brejo, matas de transição, florestas de altitude e matas ciliares.
Assim, as florestas estacionais do interior do estado parecem ser compostas por dois grupos de
espécies. O primeiro englobaria um pequeno número de espécies, mas com ampla distribuição, que
suportariam maior amplitude de variações abióticas e por isso ocorreriam em um grande número de
áreas. No segundo grupo estariam aquelas espécies com distribuição restrita a certas regiões ou com
distribuição disjunta e aquelas cuja ocorrência estaria condicionada a um fator biótico ou abiótico

60
específico (ver Brown 1984). Essas últimas possuiriam adaptações restritas a certas condições
específicas, que influenciariam tanto sua ocorrência quanto abundância. A ocorrência das espécies
que se encaixariam no segundo grupo estaria, em última análise, mais relacionada à
heterogeneidade ambiental, sendo que as diferenças florísticas entre áreas poderiam ser
determinadas pela menor ou maior proporção das espécies desse grupo.
Nos trabalhos em florestas estacionais do interior do estado que amostraram um número de
indivíduos igual ou superior a 500, o número de espécies coletado variou entre 60 e 155 espécies
(Metzger et al. 1998, Cavassan et al. 1984, Durigan et al. 2000, Bertoni et al. 1988, Matthes et al.
1988). Assim, a única amostra que apresentou número de espécies que pode ser considerado baixo,
foi a do fragmento P3. Nas demais amostras houve poucas variações no número de espécies
coletadas (ver Capítulo 3).
Esse resultado vai contra à freqüentemente mencionada perda de riqueza de espécies nos
estudos sobre fragmentação de ecossistemas (Curtis 1956, Diamond & May 1976, Harris 1980,
1984). Por outro lado, alguns estudos também mencionam que algumas espécies poderiam se
beneficiar do ambiente fragmentado, pois teriam maiores chances de dispersão e colonização
(Janzen 1983, Denslow 1995, Lovejoy et al. 1996, Laurance 1997, Tabarelli et al. 1999). Neste caso,
poderiam não ser observadas diferenças entre as riquezas das amostras e sim em sua composição.
As proporções de espécies das diferentes síndromes de dispersão variaram pouco entre os
fragmentos e essas variações não pareceram associadas ao tamanho dos mesmos. Na maioria das
amostras as espécies zoocóricas representaram em torno de 60% do total de espécies, as
anemocóricas 25% e as autocóricas 15%. As proporções observadas de espécies zoocóricas
estiveram de acordo com o proposto por Howe & Smallwood (1982), segundo os quais as espécies
dispersas por animais representariam de 50 a 75% do total em florestas tropicais. Proporções
semelhantes também foram encontradas por Martins et al. (1995), Morellato & Leitão Filho (1992) e
Penhalber & Mantovani (1997). Os primeiros avaliaram as espécies arbóreas de uma floresta
atlântica e encontraram 75% de espécies zoocóricas e 18% de anemocóricas. Morellato & Leitão
Filho (1992) analisaram esse aspecto em uma floresta estacional do interior do estado de São Paulo
e observaram que 70% das espécies eram zoocóricas e 22,5%, anemocóricas, e Penhalber &
Mantovani (1997) observaram que 59% das espécies eram zoocóricas e 33% anemocóricas em uma
floresta secundária de São Paulo.
Em apenas um fragmento pequeno (P3) a proporção de espécies anemocóricas foi superior aos
demais e também às citadas acima. Entretanto, este fato parece insuficiente para comprovar as
sugestões de Howe & Smallwood (1982) e Hughes et al. (1994), para os quais a proporção de
espécies anemocóricas seria superior em fragmentos mais perturbados ou mais recentes. Em todas
as outras áreas estudadas, independentemente de seu tamanho, idade ou histórico de perturbação, a
proporção de espécies anemocóricas foi semelhante, sendo que os menores valores ocorreram em
outros dois fragmentos pequenos que, teoricamente, estariam mais expostos à perturbações.

61
No fragmento P3 a proporção de espécies zoocóricas foi ainda ligeiramente inferior em relação
aos demais mas, por outro lado, em outros dois fragmentos pequenos (P1 e P4) foram observadas as
maiores proporções de zoocóricas do estudo. Dessa forma, não encontramos redução na proporção
de espécies zoocóricas com a diminuição da área dos fragmentos, como observado por Tabarelli et
al. (1999). Isto pode indicar que, se as alterações de composição estão ocorrendo, elas não estão se
refletindo na proporção de espécies das diferentes síndromes de dispersão.
Essas alterações poderiam, entretanto, ser detectadas quando consideramos a proporção de
indivíduos das diferentes síndromes nos fragmentos. Neste caso, observamos grande variabilidade,
mas essas variações também não estiveram relacionadas ao tamanho dos fragmentos, nem à
ocorrência de fatores de perturbação, como queimadas. Apenas uma das áreas com histórico de
perturbação mais intenso, o fragmento G2, apresentou proporção significativamente maior de
indivíduos anemocóricos como prevêem as proposições da literatura. Nas demais áreas com histórico
de perturbação intenso, como P2 e C3 ocorreu o inverso, isto é, as proporções de indivíduos
anemocóricos foram inferiores aos demais fragmentos. Em P2 e C3 foram os indivíduos autocóricos
que apresentaram proporção significativamente superior aos demais. Isto se deve, possivelmente, ao
fato de que algumas das espécies mais abundantes nessas áreas exibiram esta síndrome (Capítulo
4). Em relação à proporção de indivíduos zoocóricos ocorreu quase o inverso do previsto na
literatura, isto é, diminuição desses indivíduos com a redução da área. Embora não tenha havido
correlação entre essa proporção e a área dos fragmentos, observamos maiores proporções em dois
fragmentos pequenos (P1 e P3) e em um médio (M3) e menores proporções em um fragmento
grande (G2) e em duas áreas de Ribeirão Cachoeira (C1 e C3). A proporção de indivíduos zoocóricos
em P3 foi 50% superior às demais áreas. A grande proporção de indivíduos zoocóricos nessa área se
deve à dominância de Trichilia clausseni, espécie zoocórica que ocorreu com 229 indivíduos nessa
área, quase 50% do total coletado. Boa parte das variações observadas quanto às proporções de
indivíduos das diferentes síndromes pareceu estar mais associada à dominância de determinadas
espécies em cada área, como no caso de P3, P2 e C3, do que a efeitos de fragmentação. A causa
dessas dominâncias diferenciais, entretanto, pode estar relacionada a diversos fatores, cuja
determinação exigiria estudos mais aprofundados e específicos. De qualquer forma, na maioria dos
casos, os resultados estão em desacordo com as hipóteses testadas e parecem mais relacionados
com a heterogeneidade ambiental intrínseca de cada fragmento do que a fatores de perturbação ou
tamanho dos fragmentos.
As áreas também não se agruparam segundo suas classes de tamanho, como seria esperado
de acordo com a hipótese testada. As amostragens feitas ao longo de uma área contínua (C1, C2 e
C3) pareceram tão heterogêneos entre si quanto fragmentos espalhados ao longo da paisagem. As
amostras de Ribeirão Cachoeira mesclaram-se com as amostras de fragmentos pequenos, médios e
grandes. Por outro lado, os fragmentos G2, P2 e P3 apareceram algo isolados em relação aos
demais, indicando riquezas e composição muito distintas. Os fragmentos P3 e G2 apresentaram

62
número de espécies coletadas inferior à média apresentada pelas demais amostras (representando
50% para P3 e 70% no caso de G2). Isto pode influenciar os coeficientes de similaridade obtidos.
A heterogeneidade florística entre os remanescentes de florestas estacionais semidecíduas do
interior do estado tem sido mencionada por diversos autores (Salis et al. 1995, Gandolfi et al. 1995,
Matthes et al. 1988, Bertoni & Martins 1987). Para Bertoni & Martins (1987), as variações florísticas
estariam relacionadas à heterogeneidade ambiental e aos diferentes graus de perturbação a que
essas florestas são submetidas. Torres et al. (1997) apontaram mais especificamente as variações
climáticas, edáficas e de altitude. Para Salis et al. (1995) fatores edáficos e topográficos seriam os
principais fatores envolvidos, mas não necessariamente a proximidade geográfica. Jacquemyn et al.
(2001) observaram que a similaridade florística diminuía com o aumento da distância entre as áreas,
em concordância com as idéias de Condit (1998 apud Cook 1998), segundo o qual a proximidade
geográfica seria o único fator confiável para se prever a similaridade entre áreas. Os resultados
obtidos indicam que todos esses fatores podem estar envolvidos e a proximidade geográfica poderia
ser um aspecto preponderante, uma vez que os fatores físicos tenderiam a ser mais uniformes quanto
mais próximos os remanescentes.
Os fragmentos P2, P3 e G2 foram os que mais diferiram dos demais em relação aos aspectos
analisados: famílias mais ricas, composição e proporção de indivíduos das diferentes síndromes de
dispersão. Estas variações não puderam ser associadas ao tamanho dos fragmentos e parecem estar
mais relacionadas a seu histórico de preservação, como a ocorrência de queimadas, e à
heterogeneidade ambiental, como fatores edáficos e de relevo.
Os padrões de vegetação tem sido associados à heterogeneidade ambiental há muito tempo e
por diversos autores (Cooper 1926, Austin 1985, Barberis 2002). Mas para Condit (1996), seria
simplificar demais achar que a heterogeneidade entre florestas possa ser devida somente a fatores
abióticos. Fatores como pragas ou patógenos espécie-específicos podem ocorrer em um fragmento,
dizimando populações de determinadas espécies naquele local, mas sem capacidade de atingir
outras áreas. Ainda assim, para aquele autor, a heterogeneidade ambiental deve ser considerada e,
no presente estudo, a maior parte das variações de composição entre amostras parece estar muito
mais relacionada à heterogeneidade pré-existente do que às modificações causadas pela
fragmentação.
Existem ainda fatores que podem mascarar os efeitos das alterações ocasionadas pela
fragmentação. As espécies respondem diferentemente ao ambiente e suas modificações (Kellman et
al. 1998, Kellman & Tackeberry 1993) e espécies com longa expectativa de vida, como muitas
espécies de árvores, podem permanecer em certos ambientes por muitos anos, mesmo que as
condições que determinaram sua ocorrência tenham se alterado (Condit 1996, Turner & Colett 1996).
Por isso, é importante termos em mente que a percepção dos diferentes efeitos da fragmentação
sobre a composição de florestas depende da escala temporal adotada, pois a comunidade pode

63
nunca chegar a um equilíbrio na composição de espécies em qualquer escala de tempo ou espaço
(Condit 1996).
Os resultados apresentados aqui, de certa forma, confirmam as idéias de Condit (1996) citadas
acima. Cada fragmento deste estudo exibe uma composição de espécies que parece resultante de
uma série de fatores que variaram diferentemente ao longo do espaço e do tempo. Talvez por isso
seja tão difícil estabelecer padrões considerando apenas as classes de tamanho dessas matas. Essa
dificuldade, entretanto, indica que cada fragmento apresenta um conjunto de características próprias,
o que ressalta sua importância em termos de conservação. A mata P3, por exemplo, apesar do baixo
número de espécies, possivelmente, constitui importante suporte à fauna da região devido à
abundância de plantas zoocóricas. Além disso, todas as áreas estudadas abrigam espécies raras ou
que estão sendo citadas pela primeira vez para a região de Campinas, que já é relativamente bem
estudada do ponto de vista florístico. Vale lembrar ainda que 14 taxa (5%) não puderam ser
identificados até o nível de espécie, um deles permanecendo sem qualquer identificação. Isso mostra
a importância de estudos descritivos como este e indicam a necessidade de mais pesquisa em
taxonomia e florística, como apontado por Prance et al. (2000).
Tais observações estão de acordo com o sugerido por Turner & Colett (1996), para os quais
fragmentos pequenos espalhados em uma paisagem agricultural são fundamentais para a
manutenção da biodiversidade de uma região, pois aumentam a probabilidade de sobrevivência de
diversas espécies e muitas vezes abrigam espécies ameaçadas de extinção.
Em face disso, o questionamento sobre a relevância dos remanescentes atuais na manutenção
da composição original desse ecossistema parece de pouca relevância prática e teórica. A
devastação das florestas da área de estudo teve início no século XVIII. Antes disso, estas matas
eram utilizadas por indígenas, que certamente influenciavam na composição de espécies (Petrone
1995). Assim sendo, é praticamente impossível saber qual seria a composição original dessas
florestas e, se quisermos preservar algo do que seria a composição original só nos resta conservar
esses remanescentes que, pelo apresentado aqui têm importantes contribuições.

64
Referêcias bibliográficas AIZEN, M.A. & FEINSINGER, P. 1994. Forest fragmentation, pollination, and plant reproduction in a
chaco dry forest, Argentina. Ecology 75: 330-351.
ASSUMPÇÃO, C.T., LEITÃO FILHO, H.F. & CESAR, O. 1982 Descrição das matas da Fazenda
Barreiro Rico, Estado de São Paulo. Revista Brasileira de Botânica 5: 53-66.
AUSTIN, M.P. 1985. Continuum concept, ordination methods and niche theory. Annual Review of Ecology and Systematics 16: 39-61.
BAITELLO, J.B., PASTORE J.A., AGUIAR, O.T., SERIO, F.C. & SILVA, C.E.F. 1988. A vegetação
arbórea do parque do Morro do Diabo, Mun. Teodoro Sampaio, estado de São Paulo. Acta Botanica Brasilica 1:220-230.
BARBERIS, I.M., BATISTA, W.B., PIRE, E.F., LEWIS, J.P. & LEÓN, R.J.C. 2002. Wood population
distribution and environmental heterogeneity in a Chaco forest, Argentina. Journal of Vegetation Science 13: 607-614.
BARROSO, G.M. 1991. Sistemática das Angiospermas do Brasil. Volume 3, 2.o impressão. UFV,
Imprensa Universitária, Viçosa.
BASNET, K. 1992. Effect of topography on the pattern of trees in Tabonuco (Dacryodes excelsa)
dominated rain forest of Porto Rico. Biotropica 24: 31-42.
BERNACCI, L.C. & LEITÃO FILHO, H.F 1996. Flora fanerogâmica da floresta da Fazenda São
Vicente, Campinas, SP. Revista Brasileira de Botânica 19: 149-164.
BERTONI, J.E.A., MARTINS, F.R 1987. Composição florística de uma floresta ripária na Reserva
Estadual de Porto Ferreira,SP. Acta Botanica Brasilica 1:17-26.
BERTONI, J.E.A., MARTINS,F.R., MORAES, J.L. & SHEPHERD, G.J. 1988. Composição florística e
estrutura fitossociológica do Parque Estadual de Vaçununga, Santa Rita do Passa Quatro –
Gleba Praxedes, Boletim Técnico do Instituto Florestal de São Paulo 42: 149-170.
BIERREGAARD, R.O., LOVEJOY, T.E., KAPOS, V., SANTOS, A.A. & HUTCHINGS, R.W. 1992. The
biological dynamics of tropical rainforest fragments. Bioscience 42: 859-866.
BOTREL, R.T., OLIVEIRA FILHO, A. T., RODRIGUES, L.A. & CURI, N. 2002. Influência do solo e
topografia sobre as variações na composição florística e estrutura da comunidade arbóreo-
arbustiva de uma floresta estacional semidecidual em Ingaí, MG. Revista Brasileira de Botânica
25: 195-213.
BROWN, J.H. 1984. On the relationship between abundance and distribution of species. The American Naturalist 124: 255-279.
BROWN, K.S.Jr. & BROWN, G.G. 1992. Habitat alteration and species loss in Brazilian forests. Pp
129-142. In: T.C. Whitmore & J.A. Slater (ed.). Tropical deforestation and spcies extinction.
Chapman and Hall, London.
BUDOWSKY, G. 1965. Distribution of tropical american rain forest species in light of successional
process. Turrialba 15: 40-42.

65
CAVASSAN, O., CESAR, O. & MARTINS, F.R. 1984. Fitossociologia da vegetação arbórea da
reserva estadual de Bauru, Estado de São Paulo. Revista Brasileira de Botânica 7: 91-106.
CESAR ,O. & LEITÃO FILHO, H.F 1990. Estudo florístico quantitativo de mata mesófila semidecídua
na fazenda Barreiro Rico, município de Anhembi, SP. Revista Brasileira de Botânica 50: 133-
147.
CHRISTOFOLETTI, A. & FEDERICI, H. 1972. A terra campineira. Indústrias gráficas Mousinho.
CIELO FILHO, R. & SANTIN, D.A. 2002. Estudo florístico e fitossociológico de um fragmento florestal
urbano - Bosque dos Alemães, Campinas, SP. Revista Brasileira de Botânica 25: 291-301.
CONDIT, R. 1996. Defining and mapping Vegetation types in mega-diverse tropical forests. Trends in Ecology and Evolution 11: 4-5.
COOK, S. 1998. A diversity of approaches to the study of species richness. Trends in Ecology and Evolution 13: 340-341.
COOPER, W.S. 1926. The fundamentals of Vegetation change. Ecology 7: 391-413.
COSTA, L.G.S. & MANTOVANI, W. 1995. Flora arbustivo-arbórea de trecho de mata mesófila
semidecídua, no sítio ecológico de Ibicatu, Piracicaba (SP). Hoehnea 22: 47-59.
COTTAN, G. & CURTIS, J.T. 1956. The use of distance measures in phytossociological sampling.
Ecology 37: 451-460.
CRONQUIST, A. 1981. An integrated system of classification of flowering plants. NewYork:
Columbia University Press & The New York Botanical Garden.
CURTIS, J.T. 1956. The modification of mid-latitude grasslands and forests by ants. Pp 721-736. In:
W.L. Thomas (ed.). Man’s role in changing the face of the earth. University os Chicago Press.
Chicago.
DENSLOW, J.S. 1995. Disturbance and diversity in tropical rain forests: The density effect.
Ecological Aplications 5: 962-968.
DIAMOND, J.M. & MAY, R.M. 1976. Island biogeography and the design of natural reserves. Pp 163-
186. In: R.M. May (ed.) Theoretical Ecology: principles and applications. Saunders.
Philadelfia.
DISLICH, R., CERSÓSIMO, L. & MANTOVANI, W. 2001. Análise da estrutura de fragmentos
florestais no planalto paulistano - SP. Revista Brasileira de Botânica 24: 321-332.
DISLICH, R., KISSER, N. & PIVELLO, V.R. 2002. A invasão de um fragmento florestal em São Paulo
(SP) pela palmeira australiana Archontophoenix cunninghamiana H. Wendl. & Drude. Revista
Brasileira de Botânica 25: 55-64.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista Brasileira de Botânica 23: 371-383
FONSECA, G.A.B. 1981. Biogeografia insular aplicada à conservação. Revista Brasileira de Geografia 43: 383-388.

66
GALETTI, M. 1996. Fruits and frugivores in a Brazilian Atlantic forest. Ph.D thesis, University of
Cambridge, Cambridge, UK.
GANDOLFI, S., LEITÃO FILHO, H.F. & BEZERRA, C.L.F. 1995. Levantamento florístico e carater
sucessional das espécies arbustivo-arbóreas de uma floresta mesófila semidecídua no município
de Guarulhos, SP. Revista Brasileira de Biologia 55: 753-767.
GASCON, C. LOVEJOY, T.E., BIERREGAARD Jr., R.O., MALCOLM, J.R. STOUFFER, P.C.,
VASCONCELOS, H.L., LAURANCE, W.F. LAURANCE, ZIMMERMAN, B. TOCHER, M. &
BORGES, S. 1999. Matrix habitat and species richness in tropical forest remnants. Biological Conservation 91: 223-229.
GIBBS, P.E. & LEITÃO FILHO, H.F. 1978. floristic composition of an area of gallery forest, near Mogi
guaçu, State of São Paulo, SE Brasil. Revista Brasileira de Botânica 3: 17-22.
GÓMEZ-POMPA, A. 1971. Posible papel de la vegetación secundária em la evolución de la flora
tropical. Biotropica 3: 125-135.
GROMBONE, M.T., BERNACCI,L.C., MEIRA NETO,J.A.A., TAMASHIRO,J.Y. & LEITÃO FILHO, H.
1990. Estrutura fitossociológica da floresta semidecídua de altitude do Parque Municipal de Grota
Funda (Atibaia-Estado de São Paulo). Acta Botanica Brasilica 4: 47-64.
GUIDON, F.C. 1995. The importance of forest fragments to the maintenance of regional biodiversity in
Costa Rica. Pp 163-186. In: J. Schelhas, & R. Greenberg, (ed.), Forest patches in tropical landscapes. Island Press, Washington, D.C.
HANSKI, I. 1994. Patch-occupancy dynamics in fragmented landscapes. Trends in Ecology and Evolution 9: 131-135.
HARRIS, L.D. 1980. Forest and wildlife dynamics in the southeast. Transactions of the North American wildlife and natural resourses conference 45: 307-322.
HARRIS, L.D. 1984. The fragmented forest; island biogeography theory and the preservation of biotic diversity. University of Chicago Press. Chicago.
HARTSHORN, G.S. 1978. Treefalls and tropical forest dynamics. Pp 617-638. In: P.B. Tonlinson &
M.H. Zimmerman (ed.), Tropical trees as living systems. Cambridge University Press, New York.
HOWE, H.F. & SMALLWOOD, J. 1982. Ecology of seed dispersal. Annual Review of Ecology and Systematics 13: 201-228.
HUGHES, L., DUNLOP, M., FRENCH, K., LEISHMAN, M.R., RICE, B., RODGERSON, L. &
WESTOBOY, M. 1994. Predicting dispersal spectra: a minimal set of hypotheses based on plant
attributes. Journal of Ecology 82: 933-950.
JACQUEMYN, H., BUTAYE, J., DUMORTIER, M., HERMY, M. & LUST, N. 2001. Effects of age and
distance on the composition of mixed deciduous forest fragments in an agricultural landscape.
Journal of Vegetation Science 12: 635-642.
JANSEN, D.H. 1983. No park is a island: increase in interference from outside as park size decreases.
Oikos 41: 402-410.

67
KAPOS, V., WANDELLI, E., CAMARGO, J.L. & GANADE, G. 1997. Edge-related changes in
environent and plant responses due to forest fragmentation in central Amazonia. Pp 33-44. In:
W.F. Laurance & R.O Bierregaard (ed.), Tropical forest remnants: Ecology, management, and conservation of forest communities. University of Chicago Press. Chicago.
KATTAN, G. & ALVAREZ-LÓPEZ, H. 1995. Preservation and management of biodiversity in
fragmented landscapes in the Colombian Andes. Pp 3-18. In: J. Schelhas, & R. Greenberg,
(ed.), Forest patches in tropical landscapes. Island Press, Washington, D.C.
KELLMAN, M. & TACKABERRY, R. 1993. Disturbance and tree species coexistence in tropical
riparian forest fragments. Globbal Ecology and Biogeography Letters 3: 1-9
KELLMAN, M. & TACKABERRY, R. & RIGG, L. 1998. Structure and function in two tropical gallery
forest communities: implications for forest conservation in fragmented systems. Journal of Applied Ecology 35: 195-206.
KÖPPEN, W. 1948. Climatologia. México - Buenos Aires. Ed. Fundo de cultura Econômica, 478p.
LAURANCE, W.F. 1997. Hyper-disturbed parks: edge effects and the Ecology of isolated rainforest
reserves in tropical Australia. Pp 71-83. In: W.F. Laurance & R.O Bierregaard (eds.), Tropical forest remnants: Ecology, management, and conservation of forest communities. University
of Chicago Press. Chicago.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M., LAURANCE, S.G., HUTCHINGS,
R.W. & LOVEJOY, T.E. 1998. Effects of forest fragmentation on recruitment patters in amazonian
tree communities. Conservation Biology 12: 460-464.
LEITÃO FILHO, H.F. 1982. Aspectos taxonômicos das florestas do Estado de São Paulo. Silvicultura em São Paulo 16: 197-206.
LEITÃO FILHO, H.F. 1992. A flora arbórea da Serra do Japi. Pp 40-63. In: L.P.C Morellato (ed.),
História Natural da Serra do Japi. Editora Unicamp/FAPESP. Campinas, SP.
LIEBERMAN, M., LIEBERMAN, D., HARTSHORN, G.S. & PERALTA, R. 1985. Small-scale altitudinal
variation in lowland wet tropical forest Vegetation. Journal of Ecology 73: 505-516.
LOVEJOY, T.E., BIERREGAARD, R.O., RYLANDS , A.B., MALCOLM, J.R., QUINTELA, C.E.,
HARPER, L.H., BROWN, K.S., POWELL, A.H., POWELL, G.V.N., SCHUBART, H.O.R. & HAYS
.B. 1996. Edge and other effects of isolation on Amazon forest fragments. Pp 257-285. In: M.E.
Soule (ed.), Conservation Biology: the science of scarcity and diversity. Sinauer, Suderland.
USA
MABBERLEY, D.J. 1997. The plant book: a portable dictionary of the vascular plants. Cambridge
University Press.
MAGURRAN, A. 1988. Ecological diversity and its measurement. Croom Helm, London
MANTOVANI, W. & MARTINS, F.R. 1988. Variações fenológicas das espécies do cerrado da reserva
biológica de Mogi-Guaçu, estado de São Paulo. Revista Brasileira de Botânica 11: 101-112.

68
MARCHIORI, J.N.C. 2000. Dendrologia das Angiospermas: das bixáceas às rosáceas. UFSM,
Santa Maria.
MARCHIORI, J.N.C. & SOBRAL, M. 1997. Dendrologia das Angiospermas: Myrtales. UFSM,
Santa Maria.
MARTINS, S.E. POMPÉIA, S.L. & ROSSI, L. 1995. Síndromes de dispersão de espécies arbóreas da
Mata Atlântica de encosta no estado de São Paulo. In Resumos do 46o Congresso Nacional de Botânica (Sociedade Botânica do Brasil, ed.) SBB, Ribeirão Preto, SP.
MATTHES, L.A.F., LEITÃO FILHO, H.F. & MARTINS, F.R. 1988. Bosque dos Jequitibás (Campinas,
SP): composição florística e fitossociológica do estrato arbóreo. Anais V congresso da Sociedade Botânica de São Paulo, p. 55-76.
MEIRA NETO, J.A.A., BERNACCI, L.C., GROMBONE, M.T., TAMASHIRO,GJ.Y. & LEITÃO FILHO,
H.F. 1989. Composição florística de mata semidecídua de altitude do Parque Municipal da Grota
Funda (Atibaia - Estado de São Paulo). Acta Botanica Brasilica 3: 51-74.
MELLO, M.H.A., PEDRO JUNIOR, M.J., ORTOLANI, A. A. & ALFONSI R.R. 1994. Chuva e
Temperatura: cem anos de observações em Campinas. Boletim Técnico 154 . Instituto
Agronômico Campinas, SP.
METZGER, J.P., BERNACCI, L.C. & GOLDEMBERG, R. 1997. Pattern of tree species diversity in
riparian forest fragments of different widths (SE Brazi). Plant Ecology 133: 135-152.
METZGER, J.P., GOLDEMBERG, R. & BERNACCI, L.C. 1998. Diversidade e estrutura de fragmentos
de mata de várzea e de mata mesófila semidecídua submontana do rio Jacaré-Pepira (SP).
Revista Brasileira de Botânica 21: 321-330.
MIRANDA, Z.A.I. 1996. Plano de gestão da Área de Proteção Ambiental da região de Sousas e
Joaquim Egídio - APA Municipal. Relatório da Secretaria de Planejamento e Meio Ambiente - SEPLAMA. Prefeitura Municipal de Campinas, SP.
MORAES, P.L.R. 1992. Dispersão de sementes pelo mono-carvoeiro (Brachyteles arachnoides E.
Geoffroy, 1806) no Parque Estadual de Carlos Botelho. Revista Instituto Florestal 4: 1199-1205.
MORELLATO, L.P.C. 1995. Estações do ano na floresta. Pp 37-41. In: L.P.C Morellato & H.F. Leitão
Filho (eds.), Ecologia e preservação de uma floresta tropical urbana - Reserva de Santa Genebra. Editora Unicamp. Campinas, SP.
MORELLATO, L.P.C., RODRIGUES, R.R., LEITÃO FILHO, H.F. & JOLY, C.A. 1989. Estudo
comparativo da fenologia de espécies arbóreas de floresta de altitude e floresta mesófila
semidecídua na Serra do Japi, Jundiaí, São Paulo. Revista Brasileira de Botânica 12: 85-98.
MORELLATO, L.P.C. & LEITÃO FILHO, H.F. 1992. Padrões de frutificação e dispersão na Serra do
Japi. Pp 112-141. In: L.P.C Morellato (ed.), História Natural da Serra do Japi. Editora
Unicamp/FAPESP. Campinas, SP.
MORI, S.A., BOOM, B.M., CARVALHO, A.M.V. & SANTOS, T.S. 1983. Ecological importance of
Myrtaceae in an eastern Brazilian wet forest. Biotropica 15: 68-70.

69
NASCIMENTO, H.E.M., DIAS, A.S., TABANEZ, A.A.J. & VIANA, V.M. 1999. Estrutura e dinâmica de
populações arbóreas de um fragmento de floresta estacional semidecidual na região de
Piracicaba, SP. Revista Brasileira de Biologia 59: 329-342.
OLIVEIRA, J.B., MARCELO, N.C., ROSSI, M. & CALDERANO FILHO, B. 1999. Mapa pedológico do
estado de São Paulo. Legenda expandida In: OLIVEIRA, J.B. 1999. Boletim científico n.o 45, Solos do estado de São Paulo. Embrapa e IAC. Campinas.
OLIVEIRA-FILHO, A.T. , VILELA, E.A., CARVALHO, D.A & GAVILANES, M.L. 1994a. Differentiation
of streamside and upland Vegetation in an area of montane semideciduous forest in southeaster
Brazil. Flora 189: 287-305.
OLIVEIRA-FILHO, A.T. , VILELA, E.A., CARVALHO, D.A. & GAVILANES, M.L. 1994b. Effects of soil
and topography on the distribution of tree species in a tropical riverine forest in south-eastern
Brazil. Journal of Tropical Ecology 10: 483-508.
OLIVEIRA-FILHO, A.T., CURY, N., VILELA, E. A . & CARVALHO, D.A . 1997. Tree species
distribution along soil catenas in a riverside semideciduous forest in southeastern Brazil. Flora
192: 47-64.
PAGANO, S.N. & LEITÃO FILHO, H.F. 1987. Composição florística do estrato arbóreo da mata
mesófila semidecídua no município de Rio Claro (Estado de São Paulo). Revista Brasileira de Botânica 10: 37-47.
PAGANO, S.N.,LEITÃO FILHO, H.F. CAVASSAN, O. 1995. Variação temporal da composição
florística e estrutura fitossociológica de uma floresta mesófila semidecídua – Rio Claro – Estado
de São Paulo. Revista Brasileira de Biologia 55: 241-258.
PENHALBER, E.F. & MANTOVANI, W. 1997. Floração e chuva de sementes em mata secundária em
São Paulo, SP. Revista Brasileira de Botânica 20: 205-230.
PETRONE, P. 1995. Aldeamentos paulistas. Editora da Universidade Estadual de são Paulo, São
Paulo.
PIELOU, E.C. 1984. The interpretation of ecological data. Jonh Wiley & Sons, New York.
PIRANI, J.R. & SKORUPA, L.A. 2002. Rutaceae. Vol. 2, pp 281-308. In: M.G.L. WANDERLEY,
SHEPHERD, G.J., GIULIETTI, A.M., MELHEM, T.S., BITTRICH, V. & KAMEYAMA, C. (eds)
Flora fanerogâmica do estado de São Paulo. FAPESP: Hucitec, São Paulo.
POLHILL, R.M., RAVEN, P.H. & STIRTON, C.H. 1981. Evolution and systematics of the
Leguminosae. Vol 1, pp 1-34. In: R.M. POLHILL & P.H. RAVEN (eds.) Advances in legume systematics. Kew, Royal Botanic Garden.
POWELL, A.H. & POWELL, G.V.N., 1987. Population dynamics of male Euglossine bees in
Amazonian forest fragments. Biotropica 19: 176-179
PRACE, G.T., BEENTJE, H., DRANSFIELD, J. & JOHNS, R. The tropical flora remains
undercollected. Annals of the Missouri Botanical Gardens 87: 67-71.

70
RIBEIRO, J.E.L.S., HOPKINS, M.J.G., VICENTINI, A., SOTHERS, C.A., COSTA, M.A.S., BRITO,
J.M., SOUZA, M.A.D., MARTINS, L.H.P., LOHMANN, L.H.P., ASSUNÇÃO, P.A.C.L., PEREIRA,
E.C., SILVA, C.F., MESQUITA, M.R. & PROCÓPIO, L.C. 1999. Flora da Reserva Ducke: Guia de identificação das plantas vasculares de uma floresta de terra-firme na Amazônia Central. INPA, Manaus.
RODRIGUES, R. R., MORELLATO, L.P.C., JOLY, C.A. & LEITÃO FILHO, H.F. 1989. Estudo florístico
e fitossociológico em um gradiente altitudinal de mata estacional mesófila semidecídua, na Serra
do Japi, Jundiaí, SP. Revista Brasileira de Botânica 12:71-84.
ROSSI, L. 1994. A flora arbórea-arbustiva da mata da reserva da Cidade Universitária “Armando de
Salles Oliveira” (São Paulo, Brasil) Boletim do Instituto de Botânica n.º 9. Instituto de Botânica,
São Paulo.
SALIS, S.M. TAMASHIRO, J.Y. JOLY, C.A. 1994. Florística e fitossociologia do estrato arbóreo de um
remanescente de mata ciliar no Rio Jacaré Pepira, Brotas, SP. Revista Brasileira de Botânica
17: 93-103.
SALIS, S.M., SHEPHERD, G.J. & JOLY, C.A. 1995. Floristic comparison of mesophytic
semideciduous forests of the interior of the state of São Paulo, Southeast Brazil. Vegetatio 119:
155-164.
SANTIN, D.A. 1999. A vegetação remanescente do município de Campinas (SP): mapeamento, caracterização fisionômica e florística, visando a conservação. Tese de doutorado.
Universidade Estadual de Campinas, SP.
SANTOS, K. & KINOSHITA, L.S. (2003) Flora arbustivo-arbórea do fragmento de floresta estacional
semidecidual do Ribeirão Cachoeira, Município de Campinas - SP. Acta Botanica Brasilica 17:
325-341.
SAUNDERS, D.A., HOBBS, R.J. & MARGULES, C.R. 1991. Biological consequences of ecossystem
fragmentation: a review. Conservation Biology 5: 18-32.
SHAFER, M.L. 1981. Minimum popuations sizes for species conservation. Bioscience 31: 131-134.
SHEPHERD, G.J. 1988. FITOPAC- versão preliminar. Manual do usuário. Departamento de
Botânica. Instituto de Biologia, Universidade Estadual de Campinas, Campinas-SP.
SILVA, A.F., 1989. Composição florística e estrutura fitossociológica do estrato arbóreo da Reserva florestal Augusto Ruschi, São José dos Campos, SP. Tese de Doutorado.
Universidade Estadual de Campinas, Campinas.
SILVA, L.A. & SOARES, J.J. 2002. Levantamento fitossociológico em um fragmento de floresta
estacuionla semidecídua, no município de São Carlos, SP. Acta Botanica Brasilica 16: 205-216
SWAINE, M.D. & WITMORE, T.C. 1988. On the definition of ecological species groups in tropical rain
forests. Vegetatio 75: 81-86.

71
TABANEZ, A.A.J., VIANA, V.M. & DIAS, A.S. 1997. Consequências da fragmentação e do efeito de
borda sobre estrutura, diversidade e sustentabilidade de um fragmento de floresta de planalto de
Piracicaba, SP. Revista Brasileira de Biologia 57: 47-60.
TABARELLI, M., MANTOVANI, W. & PERES C.A. 1999. Effects of habitat fragmentation on plant guild
structure in the montane Atlantic forest of southeastern Brazil. Biological Conservation 91: 119-
127.
TAMASHIRO, J.Y.; RODRIGUES, R.R. & SHEPHERD, G.J.1986. Estudo florístico e fitossociológico da Reserva da Mata de Santa Genebra, Campinas-SP. Relatório de
Pesquisa. Fundação de Amparo à Pesquisa de São Paulo.
TERBORGH, J., 1992. Maintenance of diversity in tropical forests. Biotropica 24: 283-292.
TERBORGH, J. & WINTER, B. 1980. Some causes of extinction. Pp 119-133. In: M.E. Soulé & B.A.
Wilcox (ed.), Conservation Biology: an evolutionary-ecological perspective. Sinauer
Associates, Sunderland.
TORRES, R.B., MARTINS, F.R. & KINOSHITA, L.S. 1997. Climate, soil and tree flora relationships in
forests in the state of São Paulo, southeastern Brasil. Revista Brasileira de Botânica 20: 41-51.
TURNER, I.M. 1996. Species loss in fragments of tropical rain forest: a review of evidence. Journal of Applied Ecology 33: 200-209.
TURNER, I.M. & CORLETT, R.T. 1996. The conservation value of small, isolated fragments of
lowland tropical rain forest. Trends in Ecology and Evolution 11: 330-333.
Van TONGEREN, O.F.G. 1987. Cluster analysis. Pp 174-212. In: R.H.G. Jongman, C.J.F. ter Braak &
O.F.R. van Tongeren (eds.) Data analysis in community and landscape Ecology. Pudoc,
Wageningen.
VELOSO, H.P., RANGEL FILHO, A .L.R. & LIMA, J.C. 1991. Classificação da vegetação brasileira adaptado a um sistema universal. Rio de Janeiro. Instituto Brasileiro de Geografia e Estatística
(IBGE).
VICTOR, M.A.M.1975. A devastação florestal. Sociedade Brasileira de Silvicultura. São Paulo.
WILLIAMSON, M. 1981. Island populations. Oxford University Press. Oxford.
ZAR, J.H. 1989. Bioestatistical Analysis. Prentice Hall, New Jersey.

72
ANEXO I
Relação das famílias, espécies e morfo-espécies encontradas nos fragmentos estudados em Campinas e Itatiba, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens de Ribeirão Cachoeira; SD- método de dispersão; NC- número do coletor; X= indica ocorrência da espécie dentro da amostragem de 125 pontos quadrantes; + = espécies coletadas fora dos pontos de amostragem; a= espécies anemocóricas; u= autocóricas; z= zoocóricas; -= não classificada; *= Espécies citadas pela primeira vez para os municípios de Campinas/Itatiba, SP. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Anacardiaceae
Astronium graveolens Jacq. X X X X X X X X X X X X X a 3836
Lithraea molleoides (Vell.) Engl. X z 2587
Tapirira obtusa (Benth.) Mitchell X X X X X X z 609
Annonaceae
Annona cacans Warm. X X X X X X X X X z 413
Duguetia lanceolata A.St.-Hil. X z 2709
Guatteria nigrescens Mart. X X X X X X X + z 4054
Rollinia sylvatica (A.St.-Hil.) Mart. X X + X X X X X X X X z 4155
Xylopia brasiliensis Spreng. X X z 568
Apocynaceae
Aspidosperma cylindrocarpon Müll. Arg. X a 383
Aspidosperma olivaceum Müll. Arg. X X X X X X a 4055
Aspidosperma polyneuron Müll. Arg. X X X X X X X X X X X X a 332
Aspidosperma ramiflorum Müll. Arg. X X X X X X X X X X X a 370
Peschiera catharinensis (A.DC.) Miers + z 1081
Rauvolfia sellowi Müll. Arg. X X X + z 1234
Aquifoliaceae
Ilex taubertianum Reissek* X z 4156
Araliaceae
Dendropanax cuneatum (C.DC.) Decne. et Planch. X X X X z 1683
Didymopanax cf. calvum (Cham.) Decne et Panch. X X z 757
Arecaceae
Syagrus romanzoffiana (Cham.) Glassm. X X X X X X X X X X X z 710
Asteraceae
Gochnatia polymorpha (Less.) Cabrera X X X a 1007
Vernonia diffusa Less. X X X a 1961
Vernonia petiolaris A.DC.* X X X a 2925
Bignoniaceae
Jacaranda micrantha Cham. X X X X X X X X a 4005
Tabebuia serratifolia (Vahl.) Nich. X a 2057
Zeyheria tuberculosa (Vell.) Bureau X X X a 2058
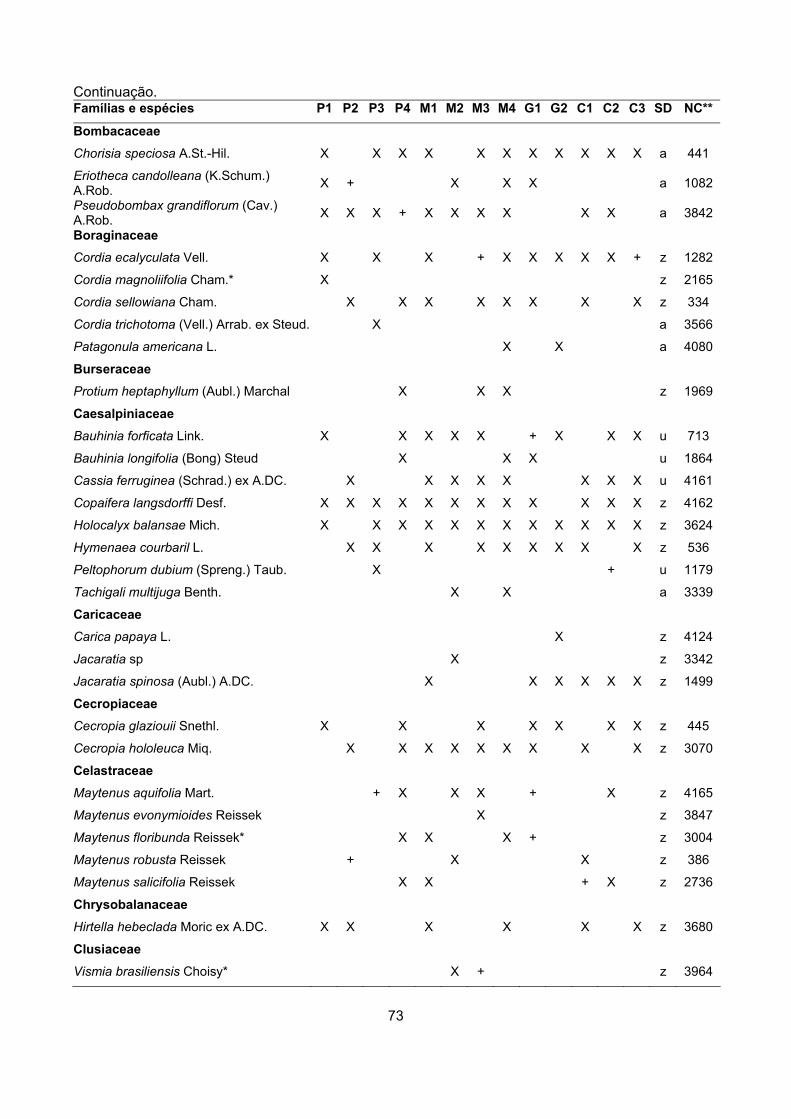
73
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Bombacaceae
Chorisia speciosa A.St.-Hil. X X X X X X X X X X X a 441
Eriotheca candolleana (K.Schum.) A.Rob. X + X X X a 1082
Pseudobombax grandiflorum (Cav.) A.Rob. X X X + X X X X X X a 3842
Boraginaceae
Cordia ecalyculata Vell. X X X + X X X X X + z 1282
Cordia magnoliifolia Cham.* X z 2165
Cordia sellowiana Cham. X X X X X X X X z 334
Cordia trichotoma (Vell.) Arrab. ex Steud. X a 3566
Patagonula americana L. X X a 4080
Burseraceae
Protium heptaphyllum (Aubl.) Marchal X X X z 1969
Caesalpiniaceae
Bauhinia forficata Link. X X X X X + X X X u 713
Bauhinia longifolia (Bong) Steud X X X u 1864
Cassia ferruginea (Schrad.) ex A.DC. X X X X X X X X u 4161
Copaifera langsdorffi Desf. X X X X X X X X X X X X z 4162
Holocalyx balansae Mich. X X X X X X X X X X X X z 3624
Hymenaea courbaril L. X X X X X X X X X z 536
Peltophorum dubium (Spreng.) Taub. X + u 1179
Tachigali multijuga Benth. X X a 3339
Caricaceae
Carica papaya L. X z 4124
Jacaratia sp X z 3342
Jacaratia spinosa (Aubl.) A.DC. X X X X X X z 1499
Cecropiaceae
Cecropia glaziouii Snethl. X X X X X X X z 445
Cecropia hololeuca Miq. X X X X X X X X X z 3070
Celastraceae
Maytenus aquifolia Mart. + X X X + X z 4165
Maytenus evonymioides Reissek X z 3847
Maytenus floribunda Reissek* X X X + z 3004
Maytenus robusta Reissek + X X z 386
Maytenus salicifolia Reissek X X + X z 2736
Chrysobalanaceae
Hirtella hebeclada Moric ex A.DC. X X X X X X z 3680
Clusiaceae
Vismia brasiliensis Choisy* X + z 3964

74
Continuação Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Combretaceae
Terminalia brasiliensis Cambess. X a 2073
Conaraceae
Conarus regnelli G. Schellenb. X X X X X X X z 498
Ebenaceae
Diospyrus inconstans Jacq. X X X X X X z 2741
Elaeocarpaceae
Sloanea monosperma Vell. X X + X X z 3351
Erythroxylaceae
Erythroxylum deciduum A.St.-Hil. + z 1975
Euphorbiaceae
Actinostemon communis (Müll. Arg.) Pax + + + + + + + + u 1088
Actinostemon concolor (Spreng) Müll. Arg. + + + + + u 4015
Actinostemon sp X u 2171
Alchornea glandulosa Poepp. & Endl. X X X X X X X X X X u 4016
Alchornea sidifolia Müll. Arg. X u 4060
Alchornea triplinervea (Spreng.) Müll. Arg. X X X X u 667
Aparisthmium sp + - 3626
Croton floribundus Spreng. X X X X X X X X X X X X u 2881
Croton priscus Croizat X X X u 337
Mabea fistulifera Mart. X u 1089
Margaritaria nobilis L.f. X X X X X X u 341
Pachystroma longifolium (Nees) I.L.Johnston X u 672
Pera glabrata (Schott.) Baill. X X X X z 1695
Sapium glandulatum (Vell.) Pax X X X X z 3753
Savia dictyocarpa Kuhlm. X X X X X X X X X X X u 582
Sebastiania commersoniana (Baill.) L.B. Smith & R.J. Downs X X u 3852
Sebastiania edwaliana Pax & Hoffm. X + X + X + X X X u 317
Tetrorchidium rubrivenium Poepp. & Endl. X X X u 3856
Fabaceae
Andira fraxinifolia Benth. X z 3631
Apuleia sp X - 388
Centrolobium tomentosum Benth. X X X X X X X X X X X X X a 318
Dalbergia foliolosa Benth.* X a 1978
Dalbergia frutecens (Vell.) Britton X X a 2743
Erythrina falcata Benth. X X u 3815
Lonchocarpus campestris Mart ex Benth. X X a 930
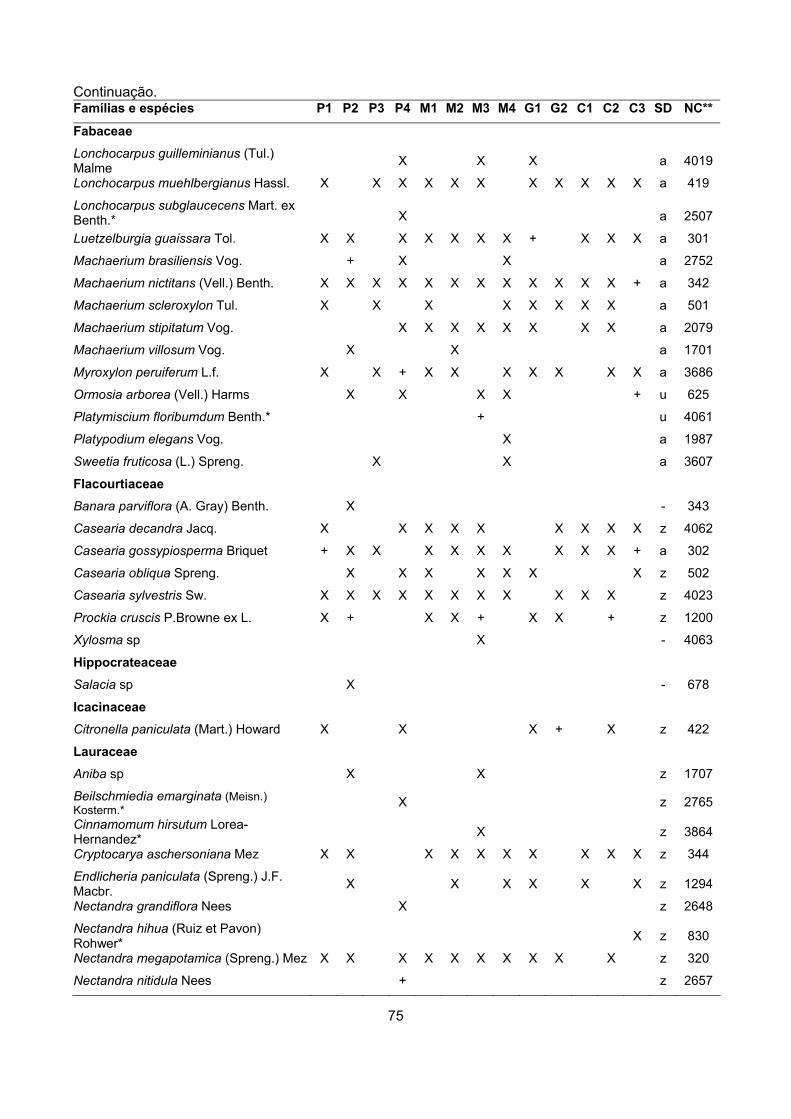
75
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Fabaceae
Lonchocarpus guilleminianus (Tul.) Malme X X X a 4019
Lonchocarpus muehlbergianus Hassl. X X X X X X X X X X X a 419
Lonchocarpus subglaucecens Mart. ex Benth.* X a 2507
Luetzelburgia guaissara Tol. X X X X X X X + X X X a 301
Machaerium brasiliensis Vog. + X X a 2752
Machaerium nictitans (Vell.) Benth. X X X X X X X X X X X X + a 342
Machaerium scleroxylon Tul. X X X X X X X X a 501
Machaerium stipitatum Vog. X X X X X X X X a 2079
Machaerium villosum Vog. X X a 1701
Myroxylon peruiferum L.f. X X + X X X X X X X a 3686
Ormosia arborea (Vell.) Harms X X X X + u 625
Platymiscium floribumdum Benth.* + u 4061
Platypodium elegans Vog. X a 1987
Sweetia fruticosa (L.) Spreng. X X a 3607
Flacourtiaceae
Banara parviflora (A. Gray) Benth. X - 343
Casearia decandra Jacq. X X X X X X X X X z 4062
Casearia gossypiosperma Briquet + X X X X X X X X X + a 302
Casearia obliqua Spreng. X X X X X X X z 502
Casearia sylvestris Sw. X X X X X X X X X X X z 4023
Prockia cruscis P.Browne ex L. X + X X + X X + z 1200
Xylosma sp X - 4063
Hippocrateaceae
Salacia sp X - 678
Icacinaceae
Citronella paniculata (Mart.) Howard X X X + X z 422
Lauraceae
Aniba sp X X z 1707
Beilschmiedia emarginata (Meisn.) Kosterm.* X z 2765
Cinnamomum hirsutum Lorea-Hernandez* X z 3864
Cryptocarya aschersoniana Mez X X X X X X X X X X z 344
Endlicheria paniculata (Spreng.) J.F. Macbr. X X X X X X z 1294
Nectandra grandiflora Nees X z 2648
Nectandra hihua (Ruiz et Pavon) Rohwer* X z 830
Nectandra megapotamica (Spreng.) Mez X X X X X X X X X X z 320
Nectandra nitidula Nees + z 2657
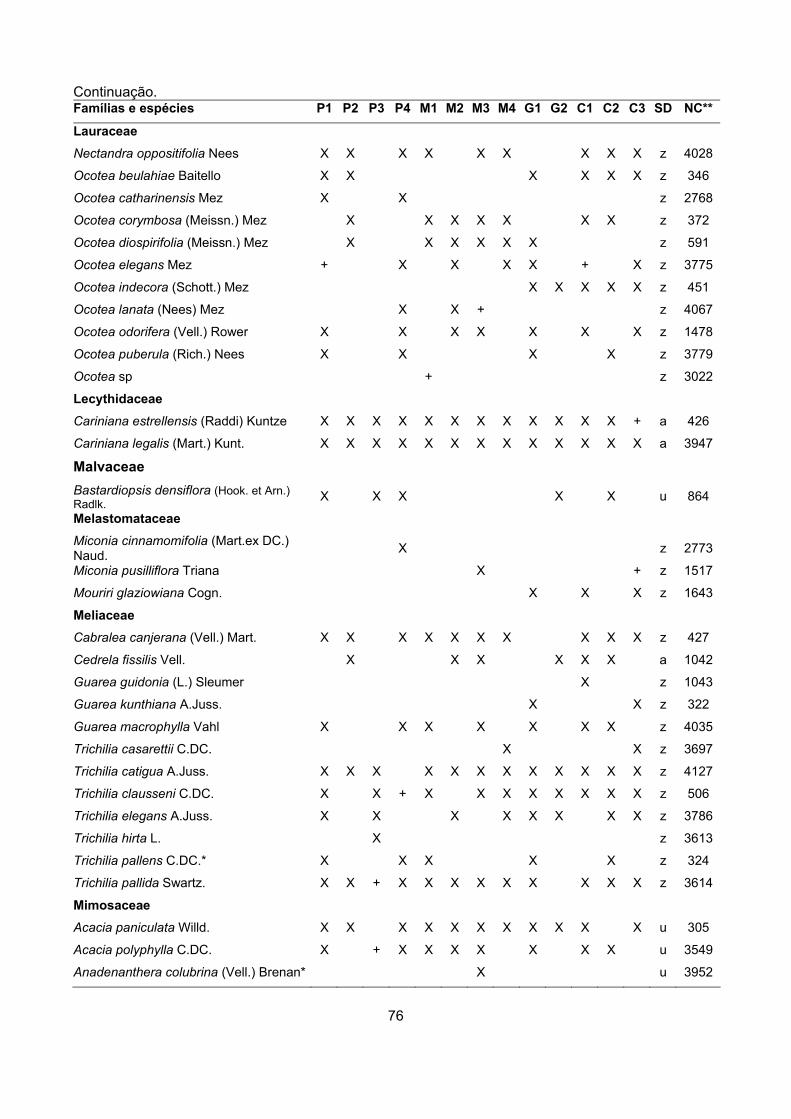
76
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Lauraceae
Nectandra oppositifolia Nees X X X X X X X X X z 4028
Ocotea beulahiae Baitello X X X X X X z 346
Ocotea catharinensis Mez X X z 2768
Ocotea corymbosa (Meissn.) Mez X X X X X X X z 372
Ocotea diospirifolia (Meissn.) Mez X X X X X X z 591
Ocotea elegans Mez + X X X X + X z 3775
Ocotea indecora (Schott.) Mez X X X X X z 451
Ocotea lanata (Nees) Mez X X + z 4067
Ocotea odorifera (Vell.) Rower X X X X X X X z 1478
Ocotea puberula (Rich.) Nees X X X X z 3779
Ocotea sp + z 3022
Lecythidaceae
Cariniana estrellensis (Raddi) Kuntze X X X X X X X X X X X X + a 426
Cariniana legalis (Mart.) Kunt. X X X X X X X X X X X X X a 3947
Malvaceae
Bastardiopsis densiflora (Hook. et Arn.) Radlk. X X X X X u 864
Melastomataceae
Miconia cinnamomifolia (Mart.ex DC.) Naud. X z 2773
Miconia pusilliflora Triana X + z 1517
Mouriri glaziowiana Cogn. X X X z 1643
Meliaceae
Cabralea canjerana (Vell.) Mart. X X X X X X X X X X z 427
Cedrela fissilis Vell. X X X X X X a 1042
Guarea guidonia (L.) Sleumer X z 1043
Guarea kunthiana A.Juss. X X z 322
Guarea macrophylla Vahl X X X X X X X z 4035
Trichilia casarettii C.DC. X X z 3697
Trichilia catigua A.Juss. X X X X X X X X X X X X z 4127
Trichilia clausseni C.DC. X X + X X X X X X X X z 506
Trichilia elegans A.Juss. X X X X X X X X z 3786
Trichilia hirta L. X z 3613
Trichilia pallens C.DC.* X X X X X z 324
Trichilia pallida Swartz. X X + X X X X X X X X X z 3614
Mimosaceae
Acacia paniculata Willd. X X X X X X X X X X X u 305
Acacia polyphylla C.DC. X + X X X X X X X u 3549
Anadenanthera colubrina (Vell.) Brenan* X u 3952
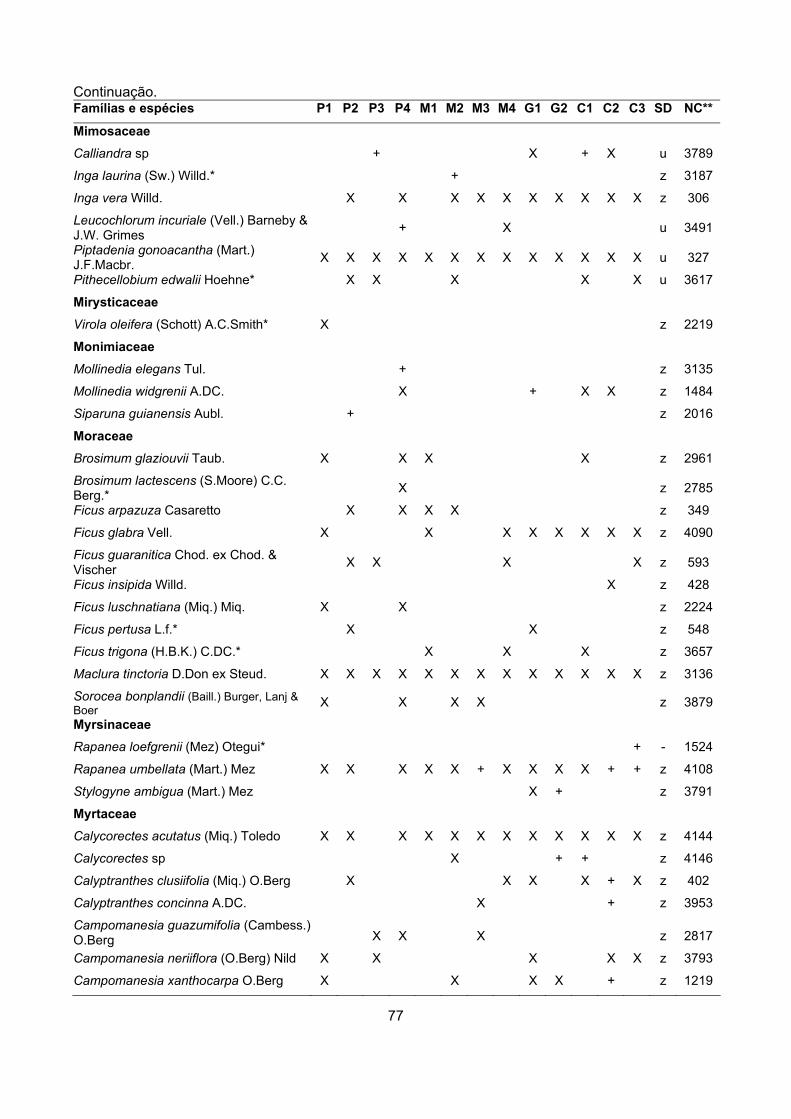
77
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Mimosaceae
Calliandra sp + X + X u 3789
Inga laurina (Sw.) Willd.* + z 3187
Inga vera Willd. X X X X X X X X X X z 306
Leucochlorum incuriale (Vell.) Barneby & J.W. Grimes + X u 3491
Piptadenia gonoacantha (Mart.) J.F.Macbr. X X X X X X X X X X X X X u 327
Pithecellobium edwalii Hoehne* X X X X X u 3617
Mirysticaceae
Virola oleifera (Schott) A.C.Smith* X z 2219
Monimiaceae
Mollinedia elegans Tul. + z 3135
Mollinedia widgrenii A.DC. X + X X z 1484
Siparuna guianensis Aubl. + z 2016
Moraceae
Brosimum glaziouvii Taub. X X X X z 2961
Brosimum lactescens (S.Moore) C.C. Berg.* X z 2785
Ficus arpazuza Casaretto X X X X z 349
Ficus glabra Vell. X X X X X X X X z 4090
Ficus guaranitica Chod. ex Chod. & Vischer X X X X z 593
Ficus insipida Willd. X z 428
Ficus luschnatiana (Miq.) Miq. X X z 2224
Ficus pertusa L.f.* X X z 548
Ficus trigona (H.B.K.) C.DC.* X X X z 3657
Maclura tinctoria D.Don ex Steud. X X X X X X X X X X X X X z 3136
Sorocea bonplandii (Baill.) Burger, Lanj & Boer X X X X z 3879
Myrsinaceae
Rapanea loefgrenii (Mez) Otegui* + - 1524
Rapanea umbellata (Mart.) Mez X X X X X + X X X X + + z 4108
Stylogyne ambigua (Mart.) Mez X + z 3791
Myrtaceae
Calycorectes acutatus (Miq.) Toledo X X X X X X X X X X X X z 4144
Calycorectes sp X + + z 4146
Calyptranthes clusiifolia (Miq.) O.Berg X X X X + X z 402
Calyptranthes concinna A.DC. X + z 3953
Campomanesia guazumifolia (Cambess.) O.Berg X X X z 2817
Campomanesia neriiflora (O.Berg) Nild X X X X X z 3793
Campomanesia xanthocarpa O.Berg X X X X + z 1219

78
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Myrtaceae
Eugenia burkartiana (D.Legrand) D.Legrand X X X X X X X + z 4148
Eugenia cerasiflora Miq. X X X z 2818
Eugenia dodoneifolia Cambess.* X X X X X X + z 3954
Eugenia excelsa O.Berg + + + X X X X + X z 738
Eugenia florida A.DC. + z 3794
Eugenia glazioviana Kiaersk. X X X X X X X + X z 4039
Eugenia ligustrina (O.Berg) Nild + + X + X + X z 1264
Eugenia lucida Cambess.* X z 2140
Eugenia microcarpa O.Berg* X X + z 1265
Eugenia pyriformis Cambess. + X X X + z 3989
Eugenia ramboi D.Legrand* X + X X X X z 739
Eugenia sp 1 X + z 4149
Eugenia sp 2 X z 3990
Eugenia verrucosa D.Legrand + X X X X X + + + z 4133
Gomidesia affinis (Cambess.) D.Legrand + z 3201
Myrcia cf. citrifolia (Poir.) Urb. + z 4074
Myrcia fallax (Rich.) A.DC. X X X X X X X z 3896
Myrcia richardiana A.DC. + X + z 2449
Myrciaria floribunda (West ex Wild.) O.Berg + + + + X X + + + X + z 3992
Myrciaria sp X X z 514
Neomitranthes glomerata (D.Legrand) D.Legrand* + z 3415
Psidium guajava L. + z 4040
Psidium sartorianum (O.Berg) Nild* X X X z 879
Nyctaginaceae
Guapira hirsuta Choisy* + + X z 4041
Guapira opposita (Vell.) Reitz X X X X X X + X X X X X X z 353
Pisonia ambigua Heimerl. X X X X X z 3205
Olacaceae
Heisteria silvianii Schwacke X z 2351
Schoepfia brasiliensis A.DC. X X z 2980
Oleaceae
Chionanthus filiformis (Vell.) P.S. Green X X X z 380
Phytolaccaceae
Gallesia integrifolia (Spreng.) Harms X X X X X X X a 2981
Seguieria langsdorffii Moq. X X X X X X X X X X X X X a 354
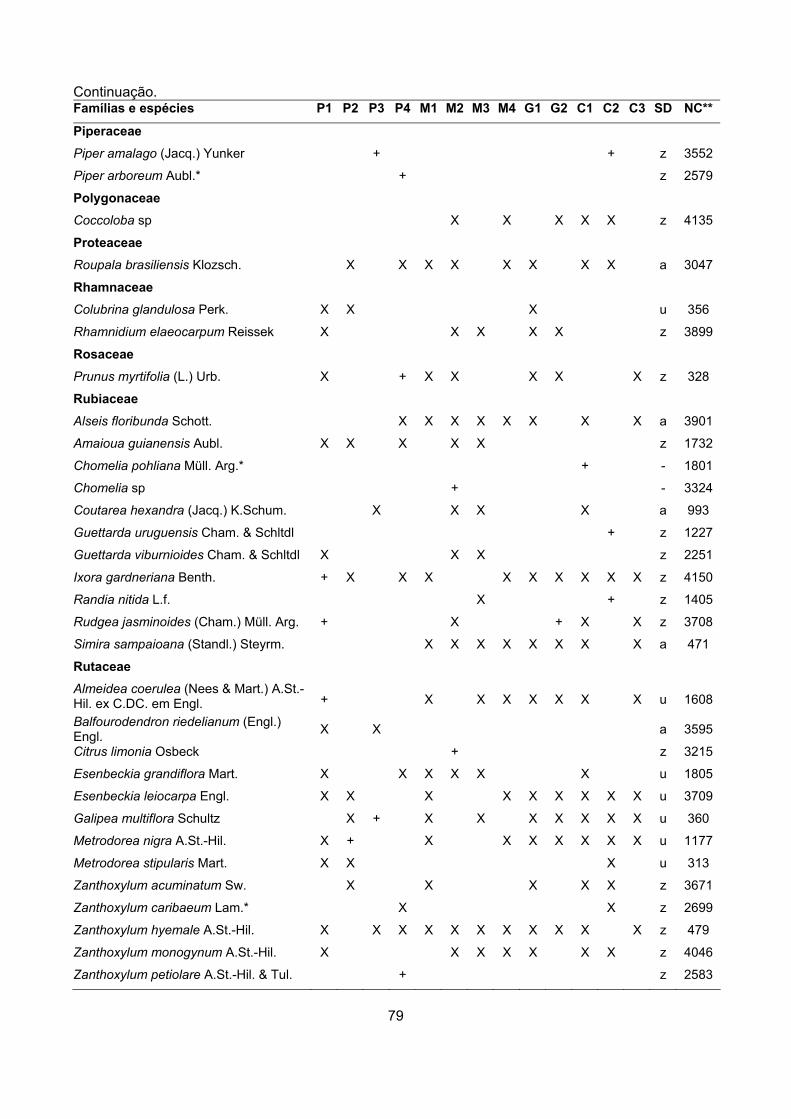
79
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Piperaceae
Piper amalago (Jacq.) Yunker + + z 3552
Piper arboreum Aubl.* + z 2579
Polygonaceae
Coccoloba sp X X X X X z 4135
Proteaceae
Roupala brasiliensis Klozsch. X X X X X X X X a 3047
Rhamnaceae
Colubrina glandulosa Perk. X X X u 356
Rhamnidium elaeocarpum Reissek X X X X X z 3899
Rosaceae
Prunus myrtifolia (L.) Urb. X + X X X X X z 328
Rubiaceae
Alseis floribunda Schott. X X X X X X X X a 3901
Amaioua guianensis Aubl. X X X X X z 1732
Chomelia pohliana Müll. Arg.* + - 1801
Chomelia sp + - 3324
Coutarea hexandra (Jacq.) K.Schum. X X X X a 993
Guettarda uruguensis Cham. & Schltdl + z 1227
Guettarda viburnioides Cham. & Schltdl X X X z 2251
Ixora gardneriana Benth. + X X X X X X X X X z 4150
Randia nitida L.f. X + z 1405
Rudgea jasminoides (Cham.) Müll. Arg. + X + X X z 3708
Simira sampaioana (Standl.) Steyrm. X X X X X X X X a 471
Rutaceae
Almeidea coerulea (Nees & Mart.) A.St.-Hil. ex C.DC. em Engl. + X X X X X X X u 1608
Balfourodendron riedelianum (Engl.) Engl. X X a 3595
Citrus limonia Osbeck + z 3215
Esenbeckia grandiflora Mart. X X X X X X u 1805
Esenbeckia leiocarpa Engl. X X X X X X X X X u 3709
Galipea multiflora Schultz X + X X X X X X X u 360
Metrodorea nigra A.St.-Hil. X + X X X X X X X u 1177
Metrodorea stipularis Mart. X X X u 313
Zanthoxylum acuminatum Sw. X X X X X z 3671
Zanthoxylum caribaeum Lam.* X X z 2699
Zanthoxylum hyemale A.St.-Hil. X X X X X X X X X X X z 479
Zanthoxylum monogynum A.St.-Hil. X X X X X X X z 4046
Zanthoxylum petiolare A.St.-Hil. & Tul. + z 2583

80
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Rutaceae
Zanthoxylum rhoifolium Lam. X X X z 607
Sapindaceae
Allophylus edulis (A.St.-Hil.) Radlk. X X X X X X X X X z 4121
Cupania vernalis Cambess. X + X X X X + z 482
Dianopteryx sorbifolia Radlk. X X a 3434
Matayba cf. guianensis Aubl. + z 3435
Matayba elaeagnoides Radlk. X X X X X X X X X X X z 3915
Sapotaceae
Chrysophyllum gonocarpum (Mart.& Eichler) Engl. X X X X X X X X X X z 4095
Chrysophyllum marginatum (Hook. et Arn.) Radlk. + z 2050
Diploon cuspidatum (Hoehne) Cronquist* X z 2270
Simaroubaceae
Picramnia ramiflora Planch. + z 2471
Picrasma crenata (Vell.) Engl.* X X z 2271
Solanaceae
Acnistus arborescens (L.) Schltdl.* + z 2374
Aureliana sp X + + z 1547
Cestrum intermedium Sendt. X X X X z 3804
Solanum argenteum Dunal + + + z 1231
Solanum bullatum Vell.* + X X X z 3998
Solanum caeruleum Vell.* X z 3225
Solanum granuloso-leprosum Dunal + z 4122
Solanum pseudoquina A.St.-Hil. X X z 377
Solanum sanctae-catharinae Dunal X + X X + + X z 3148
Sterculiaceae
Guazuma ulmifolia Lam. X X X u 315
Styracaceae
Styrax cf. acuminatus Pohl. X X z 3521
Styrax glabratum Spreng. X + X z 843
Styrax longiflorum A.DC. X X X z 4077
Styrax pohlii A.DC. X X z 699
Tiliaceae
Heliocarpus americanus L. + X X X + X X a 4152
Luehea divaricata Mart. X X X X X X a 331
Ulmaceae
Celtis tala Gillies ex Planch. X X X z 1955
Trema micrantha (L.) Blume X X X X + X z 484
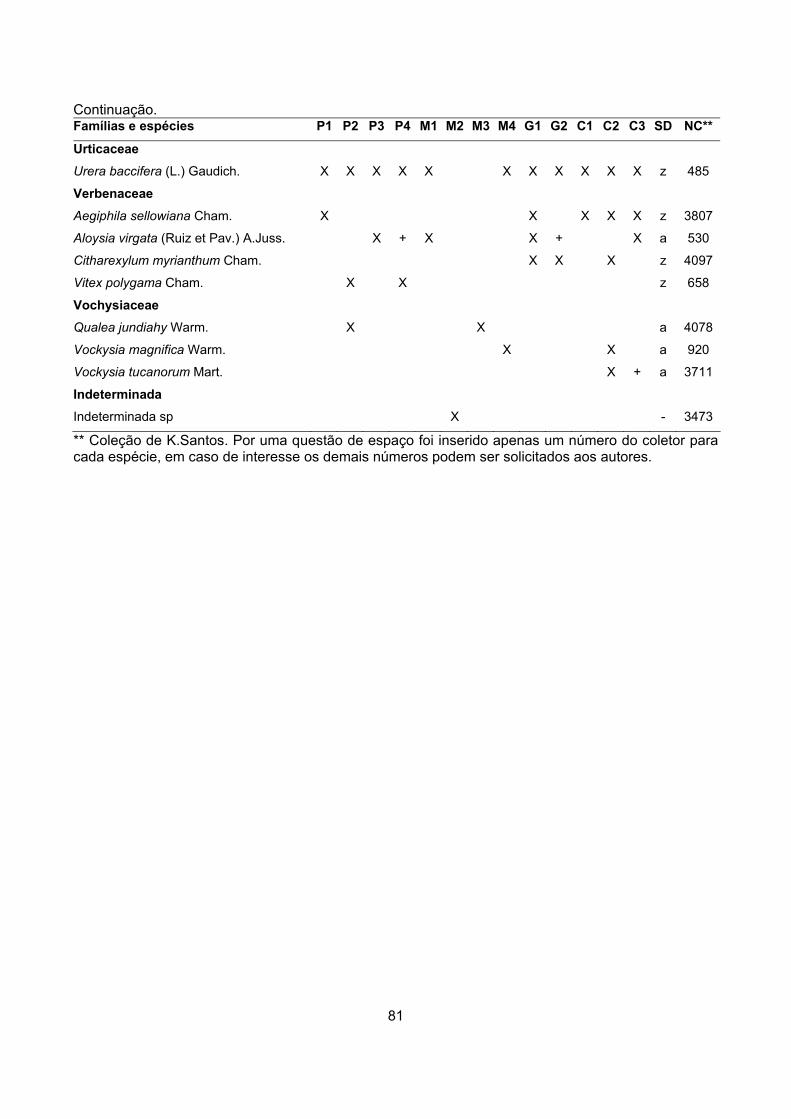
81
Continuação. Famílias e espécies P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 SD NC**
Urticaceae
Urera baccifera (L.) Gaudich. X X X X X X X X X X X z 485
Verbenaceae
Aegiphila sellowiana Cham. X X X X X z 3807
Aloysia virgata (Ruiz et Pav.) A.Juss. X + X X + X a 530
Citharexylum myrianthum Cham. X X X z 4097
Vitex polygama Cham. X X z 658
Vochysiaceae
Qualea jundiahy Warm. X X a 4078
Vockysia magnifica Warm. X X a 920
Vockysia tucanorum Mart. X + a 3711
Indeterminada
Indeterminada sp X - 3473
** Coleção de K.Santos. Por uma questão de espaço foi inserido apenas um número do coletor para cada espécie, em caso de interesse os demais números podem ser solicitados aos autores.

82
CAPÍTULO 3
RIQUEZA E DIVERSIDADE DE ESPÉCIES ARBÓREAS EM ONZE FRAGMENTOS DE FLORESTA ESTACIONAL
SEMIDECÍDUA NA REGIÃO DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE CAMPINAS, SP

83
Capítulo 3
Riqueza e diversidade de espécies arbóreas em onze fragmentos de floresta estacional semidecídua na região da Área de Proteção Ambiental do município de Campinas, SP
Karin dos Santos Pós-Graduação em Biologia Vegetal – Universidade Estadual de Campinas – Parte da tese de doutorado,
Luiza Sumiko Kinoshita Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Flavio A. M. dos Santos Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Resumo A perda de espécies em paisagens fragmentadas tem sido uma grande preocupação devido às suas
implicações sobre a conservação da biodiversidade. Tais perdas seriam maiores em fragmentos
pequenos, pois eles suportariam populações menores e mais sujeitas à extinção e estariam mais
expostos à perturbações advindas do exterior. O objetivo deste trabalho foi verificar se fragmentos
pequenos apresentavam riqueza menor do que fragmentos grandes e se as variações de riqueza
entre fragmentos são maiores do que as que ocorreriam entre áreas distintas ao longo de uma
mesma floresta. Foram escolhidos dez fragmentos com áreas inferiores a 100ha, agrupados em três
classes de tamanho e um fragmento com área de 244,9ha, localizados na região da Área de Proteção
Ambiental do município de Campinas, SP. Em cada área foram amostrados 125 pontos quadrantes,
incluindo somente indivíduos arbóreos com DAP ≥ 10cm. Foram utilizados índices de diversidade,
estimadores de riqueza não-paramétricos, curvas de rarefação e análises de similaridade. Ao todo
foram coletados 6500 indivíduos pertencentes à 248 espécies. Houve pouca variação entre as
amostras quanto ao número de espécies coletadas, diversidade e riqueza estimada. Apenas um
fragmento pequeno diferiu, apresentando cerca da metade do número de espécies coletadas nos
demais. Os estimadores de riqueza indicaram que os locais de maior riqueza seriam um fragmento
pequeno, um médio e uma das áreas do fragmento de 244,9ha. As curvas de rarefação mostraram
que as espécies distribuem-se diferencialmente entre os fragmentos, indicando que algumas áreas
contêm maior proporção do estoque regional de espécies. As variações de riqueza entre as amostras
não estiveram relacionadas ao tamanho dos fragmentos, mas possivelmente à heterogeneidade
ambiental intrínseca e à aspectos do histórico de preservação dessas matas.

84
Introdução Acessar a riqueza de espécies de uma área é uma questão importante, pois o conhecimento da
diversidade biológica muitas vezes direciona as ações de preservação. Geralmente áreas de alta
diversidade e/ou com muitas espécies raras ou endêmicas são priorizadas em tais ações
(Prendergast et al. 1993; Pressey et al. 1993). As medidas de diversidade são freqüentemente vistas
como indicadores do estado de conservação dos sistemas ecológicos (Magurran 1988). Além disso,
também são úteis na determinação da saturação de determinada comunidade local em relação ao
estoque regional de espécies (Cornell 1999).
Atualmente, uma grande proporção dos ecossistemas florestais existentes encontra-se
fragmentada, isto é, reduzida a remanescentes florestais isolados ou fragmentos (Willians-Linera
2002). Muitos autores acreditam que o processo de fragmentação florestal contribui para o declínio da
riqueza de espécies (Curtis 1956, Diamond & May 1976, Harris 1980, 1984).
A perda de espécies não se daria simplesmente por um efeito de redução de área ou perda de
heterogeneidade ambiental. Algumas espécies não persistiriam em fragmentos devido às alterações
das condições microclimáticas em relação à floresta contínua, principalmente nas regiões de borda
(Lovejoy et al. 1996, Kapos et al. 1997). Outras espécies se extinguiriam nos fragmentos devido a
eventos genéticos ou demográficos (Shafer 1981), perda de variabilidade genética (Lande 1988) e
declínio da reprodução devido à perda de polinizadores (Powel & Powell 1987, Aizen & Feinsinger
1994). Haveria ainda uma alteração da composição original, com espécies associadas às atividades
antrópicas e espécies características de ambientes perturbados tendo suas capacidades de
colonização e expansão aumentadas (Laurance et al. 1998a).
Fragmentos pequenos sofreriam reduções ainda maiores, pois suportariam populações
menores e mais sujeitas à extinção e estariam mais suscetíveis a perturbações devido à sua maior
razão borda:interior (Bierregaard et al. 1992, Turner 1996, Willians 1997, Hill & Curran 2001). Tais
perdas se acentuariam principalmente com o tempo e com o grau de isolamento dos fragmentos
(Saunders et al. 1991, Turner et al. 1996). Além disso, os fragmentos pequenos exibiriam composição
de espécies muito distinta da floresta original, por estarem mais expostos à invasão de espécies
advindas de seu entorno (Tabarelli et al. 1999). Com base em tais idéias, muitas pessoas questionam
o valor de fragmentos pequenos (< 100ha) na manutenção da diversidade de espécies nativas
(Turner & Corlett 1996).
A maior parte dos estudos sobre efeito de fragmentação na diversidade foram realizados na
Amazônia brasileira, dentro do projeto “Biological Dynamics of Forest Fragments” (Bierregaard et al.
1992). Esses estudos abordaram aspectos da diversidade e estrutura das comunidades de
fragmentos de diversos tamanhos antes e depois de seu isolamento da floresta contínua. Muitas
publicações abordando a primeira década de isolamento já foram divulgadas (Stouffer & Bierregaard
1995a, Stouffer & Bierregaard 1995b, Laurance et al. 1997, Laurance et al. 1998a, Laurance et al.
1998b, Laurance et al. 2001).

85
Entretanto, para Turner & Corlett (1996), dez anos é um período de tempo curto demais para
acessar os efeitos da fragmentação a longo prazo. Segundo esses autores e também para Williams
(1997), existem poucos estudos sobre riqueza em fragmentos isolados há mais de uma década e dos
poucos existentes a maioria aborda apenas aspectos da fauna (Leck et al. 1993, Willis 1979, Estrada
et al. 1993, Laurance 1994 apud Turner & Corlett 1996). Para Turner et al. (1996) as respostas à
insularização e perda de habitat seriam melhor acessadas em fragmentos mais antigos, onde houve
tempo para que a comunidade atingisse um certo “equilíbrio”. Nesse sentido, os fragmentos da região
da Área de Proteção Ambiental (APA) do município de Campinas constituiriam uma boa área de
estudo. A fragmentação na região teve início no século XVIII e a maior parte dos fragmentos tem
mais de 40 anos de isolamento.
O presente trabalho teve por objetivo investigar a distibuição de riqueza de espécies arbóreas
em fragmentos de floresta estacional semidecídua na região da APA do município de Campinas, SP.
Baseando-se nas proposições acima, seria esperado que em uma área restrita dentro de uma floresta
contínua ou de um fragmento grande ocorressem mais espécies de árvores do que em uma área
equivalente no interior de um fragmento pequeno. Seria esperado também que fragmentos pequenos
apresentassem composição mais homogênea do que fragmentos grandes, pois todos eles estariam
igualmente expostos à perturbações externas. Assim, pretendeu-se responder às seguintes
perguntas: fragmentos grandes apresentam maior riqueza de espécies arbóreas do que fragmentos
pequenos? fragmentos são mais similares entre si em termos de composição e abundância de
espécies do que áreas distintas dentro de um fragmento grande?

86
Material e métodos Área de estudo
O estudo foi realizado predominantemente na região da Área de Proteção Ambiental - APA - do
município de Campinas, SP (22°45’ a 23°00’ S, 47°00’ a 47°12’ W) e município de Itatiba, SP (23°10’
a 23°15’ S, 45°45’ a 46°45’ W) em áreas fronteiriças à APA. Sendo que seis fragmentos localizam-se
dentro dos domínios da APA (P1, M1, M4, G1, G2 e a mata Ribeirão Cachoeira) dois localizam-se em
áreas fronteiriças à APA, dentro do município de Campinas (P3 e P2) e três em áreas fronteiriças à
APA dentro do município de Itatiba (P4, M3 e M2) (ver Figura 1, Capítulo 1). Os fragmentos
estudados pertencem a propriedades particulares. O clima, segundo Köppen (1948), é Cwag'-
temperado, moderadamente chuvoso, de inverno seco não rigoroso, com precipitação média anual de
1409 mm (Mello et al. 1994).
Os solos ocupados pelos fragmentos em estudo podem ser Podzólicos Vermelho-Amarelos de
três diferentes tipos: PVA 8, PVA 20 e PVA ou Latossolo Vermelho: LV 4. Os relevos podem ser
suave ondulados, forte ondulados e montanhosos ou ondulados e suave ondulados (Oliveira et al.
1999). A vegetação dos fragmentos pode ser classificada como floresta estacional semidecídua
(Veloso et al. 1991). A paisagem em que os fragmentos estão inseridos constitui-se de um mosaico
de culturas anuais e perenes, pastagens, reflorestamentos e pequenos assentamentos humanos. A
agricultura é caracterizada por pequenas áreas de café, laranja, cana de açúcar e culturas de
subsistência. Os reflorestamentos com eucalipto e pastagens ocupam grandes extensões (Miranda
1996).
Foram escolhidos dez fragmentos com áreas <100ha e um fragmento grande, a Mata Ribeirão
Cachoeira (244,9 ha), cujo levantamento florístico foi realizado por Santos & Kinoshita (2003). Os
fragmentos menores do que 100ha foram categorizados em três classes de tamanho: quatro
pequenos, com área entre 12 e 13,5ha (P1, P2, P3 e P4), quatro médios, com área entre 19 e 25,5ha
(M1, M2, M3 e M4) e dois grandes, G1 e G2 com áreas de 41,8 e 63,3ha, respectivamente. Os
critérios para a escolha de cada fragmento foram: similaridade fisionômica, facilidade de acesso e
obtenção de autorização dos proprietários para a realização do trabalho.
Amostragem
Para a coleta de dados foi adotado o método de pontos quadrantes proposto por Cottam &
Curtis (1956). Cada amostra consistiu de 125 pontos quadrantes, onde foram coletados indivíduos
arbóreos com diâmetro à altura do peito (DAP) maior ou igual à 10cm, totalizando 500 indivíduos por
amostra. Os pontos foram dispostos em intervalos regulares de 15m ao longo de transecções que
percorriam área semelhante em fragmentos grandes e pequenos, evitando apenas áreas de bordas e
clareiras. As amostragens foram feitas de forma a incluir áreas semelhantes, tanto do ponto de vista

87
da fisionomia da vegetação, área percorrida e ambiente físico (condições de altitude, solo e
drenagem).
Na mata Ribeirão Cachoeira, fragmento com 244,9ha, foram feitas três amostragens de 125
pontos em três regiões diferentes do fragmento (C1, C2 e C3). As áreas C1 e C2 distam
aproximadamente 700m, a distância entre as áreas C1 e C3 é de aproximadamente 800m e entre C2
e C3 em torno de 1500m (ver Figura 1 Capítulo 1). Com isso, pretendia-se verificar se áreas distintas
dentro de um fragmento grande seriam mais similares entre si ou apresentariam mais espécies do
que os demais fragmentos.
As identificações foram feitas segundo técnicas usuais em taxonomia e os espécimes foram
incluídos em famílias segundo o sistema proposto por Cronquist (1981). A coleção principal de
material herborizado foi incorporada ao herbário UEC e as duplicatas foram enviadas a outros
herbários ou especialistas que ajudaram na identificação de alguns grupos. Os materiais coletados
em estado vegetativo foram conservados como material testemunha, em separado.
Tratamento dos dados
Para a análise da diversidade específica foram utilizados o número de espécies coletadas (S), o
índice de diversidade de Shannon-Weaver (H’), calculado na base e (nats/indivíduo) (Ludwig &
Reynolds 1988) e a eqüabilidade (J= H’/Ln S) (Magurran 1988).
Para estimar a riqueza de espécies em cada amostra foram utilizados os estimadores de
riqueza Chao I e Chao II (Chao 1984, 1987). Os estimadores fornecem a riqueza para cada área com
os desvios padrões. As riquezas estimadas em cada fragmento foram comparadas entre si através do
Teste t, com o valor de p ajustado com correção de Bonferroni, pois tratavam-se de comparações
múltiplas (Zar 1989). Dessa forma, os valores de p obtidos seriam significativos somente quando
inferiores à 0,004. Esta análise foi feita com o intuito de identificar áreas de maior riqueza.
Para verificar se havia correlação entre as riquezas estimadas e a área total dos fragmentos foi
realizada análise de correlação de Pearson, utilizando-se o programa BioEstat (Ayres et al. 1998).
Nessa análise consideramos apenas um valor para as amostras de Ribeirão Cachoeira, pois se
tratavam de três amostras realizadas em um mesmo fragmento. O valor considerado foi a média dos
valores obtidos nas três amostras.
Estimadores de riqueza não-paramétricos, como os adotados aqui, utilizam informações sobre a
distribuição de espécies raras na amostra, isto é, aquelas representadas por apenas um (singletons)
ou dois indivíduos (doubletons). Quanto maior o número de espécies raras na amostra, maior é a
probabilidade de que outras espécies que não as representadas na amostra ocorram na área (Gotelli
& Colwell 2001). No caso do estimador Chao I, a riqueza estimada seria igual à observada quando
todas as espécies presentes na amostra exibissem abundância de dois ou mais indivíduos. Para
Chao II, isto ocorreria quando todas as espécies observadas ocorressem em duas ou mais unidades
amostrais (Colwell & Coddington 1994). Assim, riquezas estimadas com valores próximos ao número

88
de espécies coletadas indicam que a maior parte das espécies que possivelmente ocorreriam
naquela área foi coletada. Por outro lado, grande variação entre a riqueza estimada e o número de
espécies coletadas indica que uma pequena parcela das espécies que ocorrem naquela área foi
coletada e que sua riqueza tende a ser elevada.
Para estimar a riqueza por número de indivíduos em cada fragmento e para o total amostrado
em todos os fragmentos, foram feitas curvas de rarefação, utilizando o programa PAST (Hammer et
al. 2001). O número de espécies coletadas por amostra de 500 indivíduos foi comparado ao número
de espécies obtidas na curva de rarefação para este tamanho amostral. Isso foi feito para verificar
como as espécies estavam distribuídas pelos fragmentos. Os fragmentos com número de espécies
coletadas mais próximo ao que seria esperado ao acaso para 500 indivíduos seriam os fragmentos
de maior riqueza ou que detivessem maior proporção do conjunto total de espécies. Estimativas de
riqueza para 250 indivíduos, com os respectivos desvios padrões, foram obtidas para cada fragmento
e comparadas com a estimativa para esse tamanho amostral para o conjunto das áreas, também com
o intuito de identificar áreas de maior riqueza.
Para verificar se fragmentos de mesmo tamanho são mais semelhantes entre si do que entre as
demais classes de tamanho, foram calculados os coeficientes de similaridade de Marczewski-
Steinhaus (Digby & Kempton 1987) que leva em conta a composição de espécies e suas
abundâncias relativas. As relações de similaridade entre os fragmentos foram investigadas através da
análise de agrupamento UPGMA, utilizando o programa FITOPAC (Shepherd 1988).

89
Resultados O número de espécies coletadas por amostra de 500 indivíduos variou entre 47 e 110. As listas
com as espécies coletadas em cada área e número de coleta encontram-se no Anexo 1 do Capítulo
2. As áreas P3 e G2 foram as que apresentaram o menor número de espécies, sendo que em P3
foram coletadas cerca da metade do número de espécies encontradas nas demais áreas. A maioria
das amostras apresentou em torno de 100 espécies, independentemente de sua área total. Os
maiores números de espécies ocorreram nos fragmentos médios M2 e M3 (Tabela 1).
A diversidade variou de 2,37 a 4,06 entre as amostragens e a diversidade para o conjunto das
áreas foi de 4,46. Os valores de equabilidade variaram entre 0,61 e 0,87 e, para o conjunto das áreas
foi de 0,81. Ambos parâmetros variaram pouco entre as amostras, mas o fragmento P3 apresentou
valores menores, que representaram cerca de 60% dos valores obtidos para as outras áreas (Tabela
1).
Tabela 1: Número de indivíduos coletados (N), número de espécies coletadas (S), diversidade (H’), e equabilidade (J’) nos fragmentos estudados na região da APA municipal de Campinas, SP, e também para o conjunto das áreas (Total). Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 Total
N 500 500 500 500 500 500 500 500 500 500 500 500 500 6500
S 103 85 47 99 98 107 110 97 104 65 97 98 90 248
H’ 4,03 3,61 2,37 3,83 3,96 3,98 3,98 3,79 4,06 3,29 3,8 3,98 3,7 4,46
J’ 0,87 0,81 0,61 0,83 0,86 0,85 0,85 0,83 0,87 0,79 0,83 0,87 0,82 0,81
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
A riqueza estimada pelo estimador Chao I variou de 69,6 (±13,91) a 206,6 (±54,82) e por Chao
II entre 95,4 (±30,71) e 231,3 (±69,81). Para o conjunto das áreas a riqueza estimada por Chao I foi
274,6 (±11,77) e por Chao II, 273,9 (± 11,29) (Figuras 1 e 2).
Os valores de riqueza estimada para o conjunto das áreas foram bem próximos ao número de
espécies amostradas. Isto indica que a maior parte das espécies esperadas, segundo o estimador,
foram coletadas para o conjunto das áreas. Na maioria das amostras a proporção entre o número de
espécies amostradas e estimadas ficou entre 60 e 70%, chegando a 78 e 87% nas amostras C1 e P1,
respectivamente. Na amostra C2, entretanto, a riqueza estimada foi quase o dobro do total de
espécies coletado na área, indicando alta riqueza neste fragmento.
Através da comparação entre os desvios padrões das estimativas de Chao I, observamos que
ocorre sobreposição na maioria dos casos, indicando que a riqueza tende a ser semelhante entre os
fragmentos, independentemente de sua classe de tamanho (Figura 1). Os desvios da riqueza

90
estimada para a mata P2, por exemplo, se sobrepõem aos de todas as outras áreas, com exceção de
P3 e G2. No caso dessas duas áreas não ocorre sobreposição dos desvios com nenhuma outra área,
indicando que esses fragmentos apresentam riquezas um pouco inferiores aos demais. Entretanto,
através do Teste t observamos que nenhuma das áreas apresentou riqueza estimada
significativamente diferente das demais (p > 0,004) (valores de t e de p na Tabela 1 do Anexo I).
Algo semelhante ocorreu com as riquezas estimadas por Chao II, embora estas tenham variado
menos entre as áreas do que no caso de Chao I. Observamos que a sobreposição dos desvios
padrões das estimativas ocorrem em um maior número de casos para Chao II (Figura 2). A área P3,
por exemplo, apresentou riqueza que pode ser considerada semelhante às de várias outras amostras.
Ainda assim, os padrões gerais se mantiveram, com P3 e G2 figurando entre as áreas de menor
riqueza e C2 como a área mais rica. Apesar disso, da mesma forma que para Chao I, não houve
diferenças significativas entre as riquezas estimadas das diversas amostras (Teste t, p > 0,004) (ver
valores de t e de p na Tabela 2 do Anexo I)
Não houve correlação entre a área dos fragmentos e as riquezas estimadas por Chao I (r =
0,181, p = 0,594) ou por Chao II (r = 0,275, p = 0,414).

91
Figura 1: Riqueza estimada por Chao I com os respectivos desvios padrões em relação ao número de espécies amostradas (*) nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
Figura 2: Riqueza estimada por Chao II com os respectivos desvios padrões em relação ao número de espécies amostradas (*) nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
P1
P2
P3
P4M1 M2
M3
M4
G1
G2
C1
C2
C3
0
50
100
150
200
250
300
Fragmentos
N.o
de
espé
cies
P1
P2
P3
P4M1 M2 M3
M4G1
G2
C1
C2
C3
0
50
100
150
200
250
300
350
Fragmentos
N.o
de
espé
cies

92
A riqueza de espécies estimada para cada conjunto de 500 indivíduos foi maior quando
consideradas todas as áreas em conjunto do que o número de espécies encontradas por amostra em
qualquer uma das áreas separadamente (Figura 3). As espécies pareceram não estar
homogeneamente distribuídas entre as áreas. Se isto ocorresse, seria esperado que o número de
espécies coletadas por amostra de 500 indivíduos não diferisse da estimativa para o geral. As áreas
que mais se aproximaram do valor estimado foram os fragmentos M2 e M3, indicando que essas
áreas detêm uma maior proporção do total de espécies coletadas e poderiam ser consideradas as
mais ricas. As áreas P3 e G2, por outro lado, apresentaram um baixo número de espécies em relação
ao que seria esperado para este número de indivíduos. Esses fragmentos, portanto detiveram a
menor proporção do total de espécies coletadas no estudo. A maioria das áreas, entretanto, aparece
em uma posição intermediária, muitas vezes se sobrepondo. Este conjunto inclui desde fragmentos
pequenos (P1 e P4) até áreas de Ribeirão Cachoeira (C1 e C2), indicando que, independentemente
de tamanho, essas áreas detiveram proporções semelhantes do conjunto total de espécies.
Figura 3: Curva de rarefação para o total de indivíduos amostrados e número de espécies coletadas por amostra de 500 indivíduos nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
0
50
100
150
200
250
0 500 1000 1500 2000 2500 3000 3500 4000 4500 5000 5500 6000 6500
Indivíduos
Espé
cies
N.o esperado de espéciesP1P2P3P4M1M2M3M4G1G2C1C2C3

93
As estimativas para 250 indivíduos (Figura 4) indicaram que nenhuma das áreas apresentou
número de espécies similar ao que seria esperado ao acaso, considerando-se o total de espécies e
indivíduos. Isto confirma, a observação anterior de que as espécies não estão distribuídas
homogêneamente pelos fragmentos estudados. Neste caso, as áreas que poderiam ser consideradas
as mais ricas seriam M2, M3, P1 e G1 e, as áreas de menor riqueza P3 e G2.
Figura 4: Riqueza estimada para um conjunto de 250 indivíduos em cada um dos fragmentos estudados na região da APA municipal de Campinas, SP e, também para o conjunto das áreas (Todas). Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
P1
P2
P3
P4M1
M2 M3M4
G1
G2
C1 C2C3
TODAS
0
20
40
60
80
100
120
Fragmentos
N.o
de
espé
cies
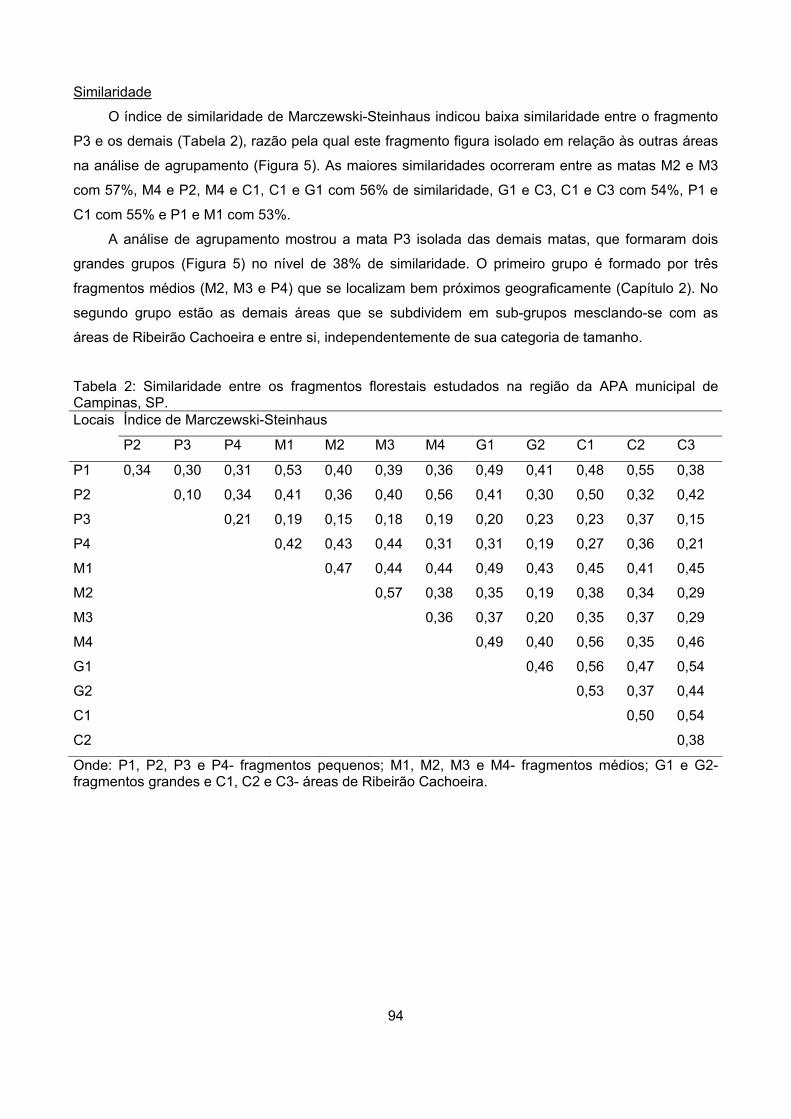
94
Similaridade
O índice de similaridade de Marczewski-Steinhaus indicou baixa similaridade entre o fragmento
P3 e os demais (Tabela 2), razão pela qual este fragmento figura isolado em relação às outras áreas
na análise de agrupamento (Figura 5). As maiores similaridades ocorreram entre as matas M2 e M3
com 57%, M4 e P2, M4 e C1, C1 e G1 com 56% de similaridade, G1 e C3, C1 e C3 com 54%, P1 e
C1 com 55% e P1 e M1 com 53%.
A análise de agrupamento mostrou a mata P3 isolada das demais matas, que formaram dois
grandes grupos (Figura 5) no nível de 38% de similaridade. O primeiro grupo é formado por três
fragmentos médios (M2, M3 e P4) que se localizam bem próximos geograficamente (Capítulo 2). No
segundo grupo estão as demais áreas que se subdividem em sub-grupos mesclando-se com as
áreas de Ribeirão Cachoeira e entre si, independentemente de sua categoria de tamanho.
Tabela 2: Similaridade entre os fragmentos florestais estudados na região da APA municipal de Campinas, SP. Locais Índice de Marczewski-Steinhaus
P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
P1 0,34 0,30 0,31 0,53 0,40 0,39 0,36 0,49 0,41 0,48 0,55 0,38
P2 0,10 0,34 0,41 0,36 0,40 0,56 0,41 0,30 0,50 0,32 0,42
P3 0,21 0,19 0,15 0,18 0,19 0,20 0,23 0,23 0,37 0,15
P4 0,42 0,43 0,44 0,31 0,31 0,19 0,27 0,36 0,21
M1 0,47 0,44 0,44 0,49 0,43 0,45 0,41 0,45
M2 0,57 0,38 0,35 0,19 0,38 0,34 0,29
M3 0,36 0,37 0,20 0,35 0,37 0,29
M4 0,49 0,40 0,56 0,35 0,46
G1 0,46 0,56 0,47 0,54
G2 0,53 0,37 0,44
C1 0,50 0,54
C2 0,38
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
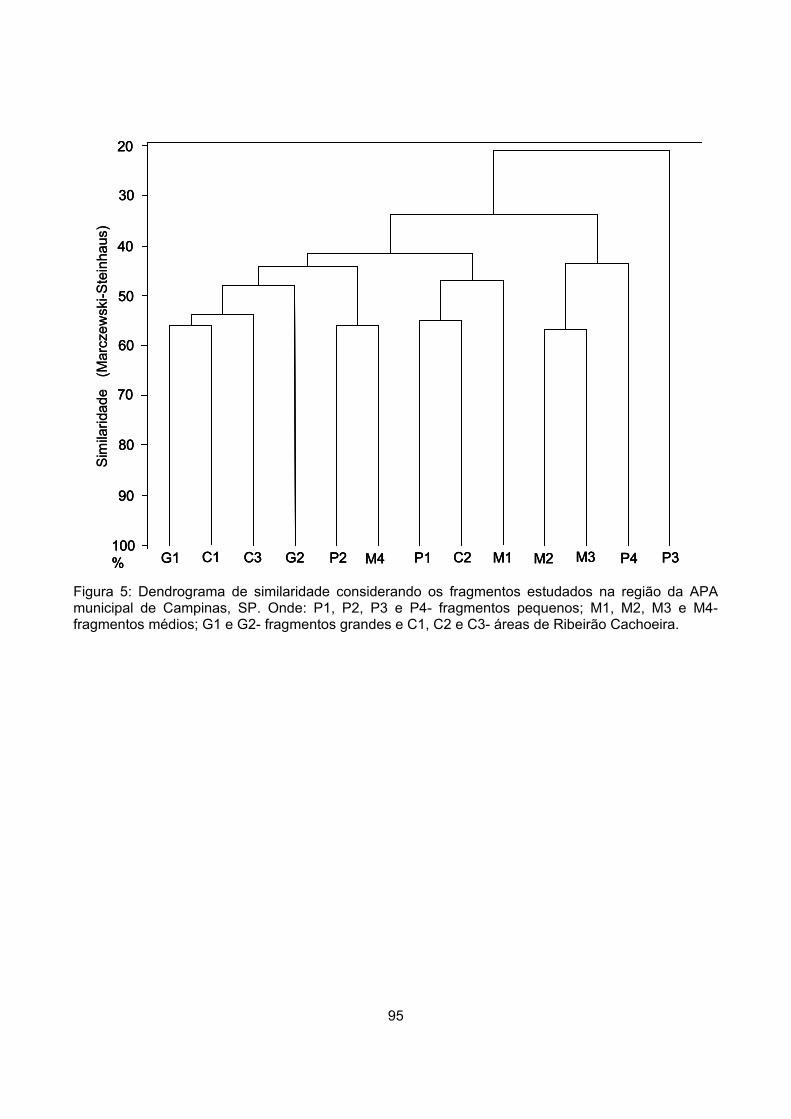
95
Figura 5: Dendrograma de similaridade considerando os fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
Sim
ilarid
ade
(M
arcz
ewsk
i-Ste
inha
us)
100%
20
30
40
50
60
70
80
90
G1 C1 C3 G2 P2 M1M4 P1 C2 M2 M3 P4 P3
Sim
ilarid
ade
(M
arcz
ewsk
i-Ste
inha
us)
100%
20
30
40
50
60
70
80
90
G1 C1 C3 G2 P2 M1M4 P1 C2 M2 M3 P4 P3100%
20
30
40
50
60
70
80
90
G1 C1 C3 G2 P2 M1M4 P1 C2 M2 M3 P4 P3

96
Discussão O número de espécies presentes em uma comunidade é uma porção do estoque regional de
espécies (Pärtel et al. 1996). Rickefs & Shluter (1993) defenderam que é impossível separar a
diversidade local da regional, devendo-se considerar ambas como expressões diferentes de um
sistema integrado em cada região. Entretanto, áreas de alta diversidade e/ou com muitas espécies
raras ou endêmicas são priorizadas para a adoção de medidas de conservação (Prendergast et al.
1993, Pressey et al. 1993). Dentro desse contexto, fragmentos pequenos são geralmente
desconsiderados em programas de conservação porque, segundo a idéia corrente, estes
remanescentes apresentariam menor riqueza de espécies, tanto pelo efeito de redução de área,
quanto por sua maior exposição a fatores de perturbação (Saunders et al. 1991, Turner 1996, Brokaw
1998).
O número de espécies encontradas no presente estudo correspondeu a mais de 50% do total
listado para o município de Campinas por Santin (1999), lembrando que, apesar de as matas
estacionais semidecíduas constituírem mais de 90% da vegetação remanescente no município, a
autora incluiu em sua amostra todos os demais tipos vegetacionais ocorrentes no município, como
cerrados (sensu lato), matas de brejo, matas de transição, florestas de altitude e matas ciliares. Isso
indica a importância dos fragmentos estudados na manutenção do estoque regional de espécies.
Todas as áreas, com exceção dos fragmentos P3 e G2, contiveram cerca de 40% do total de
espécies coletadas, independentemente de sua área. Alguns fragmentos médios (~20ha), como as
áreas M2 e M3, pareceram deter uma maior proporção da diversidade regional do que outros, como
apontado na comparação entre o número de espécies coletadas e esperadas por conjunto de 500
indivíduos na curva de rarefação. Esta análise mostrou ainda que fragmentos pequenos ou grandes
podem deter a mesma proporção do total de espécies de árvores do total amostrado, considerando a
mesma base amostral. Além disso, várias espécies ocorreram em um único fragmento, reforçando a
idéia de que as espécies estão desigualmente distribuídas entre as áreas (Capítulos 2 e 4).
Fragmentos médios, grandes, pequenos e áreas dentro de um mesmo fragmento apresentaram
números de espécies similares. Apenas um fragmento pequeno (P3) e um fragmento grande (G2)
apresentaram valores menores. No caso de P3, o número de espécies coletadas foi cerca da metade
das coletadas nas demais áreas e, no caso de G2, esse valor equivaleria a 70% das coletadas nas
outras amostras. Dessa forma, as variações no número de espécies coletadas entre as áreas não
pareceu associada ao tamanho dos fragmentos.
A riqueza estimada foi semelhante entre a maioria dos fragmentos, independentemente de sua
área. Não houve diferenças significativas entre os valores obtidos. Apesar disso, observamos que
novamente as áreas P3 e G2 apresentaram riqueza estimada um pouco inferior às demais áreas. Na
área C2 de Ribeirão Cachoeira, por outro lado, ocorreu o inverso, o que indicou alta riqueza para
essa área. As variações de riqueza, portanto, também não pareceram associadas ao tamanho dos

97
fragmentos. Para reforçar essa hipótese, observamos que não houve correlação entre as riquezas
estimadas e o tamanho dos fragmentos.
Os valores de diversidade (H’) encontrados para os fragmentos foram muito semelhantes aos
encontrados por outros autores em outros remanescentes de mata estacional semidecídua do interior
do estado de São Paulo. No município de Campinas, Matthes et al. (1988) encontraram H’ igual a
3,71nats/ind no Bosque dos Jequitibás e Bernacci (1992) encontrou 3,53nats/ind na mata da Fazenda
São Vicente. Em Bauru, Cavassan et al. (1984) encontraram 3,50nats/ind, em Rio Claro Pagano et al.
(1987) encontraram 4,29nats/ind, Rodrigues et al. (1989) encontraram 3,94nats/ind em Jundiaí,
Bertoni et al. (1988), 3,6nats/ind em Santa Rita do Passa Quatro, Cesar & Leitão Filho (1990)
3,56nats/ind em Barreiro Rico e Metzger et al. (1998) encontraram valores entre 3,03 e 4,3nats/ind
nos vários fragmentos que estudaram na região do rio Jacaré Pepira. Assim, comparado a esses
estudos, os valores de diversidade de Shannon encontrados podem ser considerados similareas
àqueles encontrados nesse tipo de formação florestal. Entretanto, o valor encontrado para o
fragmento P3 (2,37nats/ind) pode ser considerado baixo. Durigan et al. (2000) encontraram valor
semelhante (2,41nats/ind) na floresta da Estação Ecológica de Caetetus, município de Gália. Os
autores consideraram esse valor baixo para esse tipo de floresta (estacional semidecídua).
Aparentemente, na maioria das matas os valores de diversidade variam entre três e quatro, mas
alguns remanescentes exibem valores um pouco inferiores, como é o caso de P3, Caetetus e Ibicatu
(Costa 1992). Existe, portanto uma variação entre os valores de diversidade de Shannon encontrados
em florestas estacionais e esta variação possivelmente não está correlacionada ao tamanho nem às
perturbações sofridas pelos fragmentos.
A análise de agrupamento indicou que as variações de composição e abundância relativa das
espécies nas diversas áreas não parecem associadas ao tamanho dos fragmentos, pois os
remanescentes não se agruparam segundo suas classes de tamanho. Essa análise também mostrou
que áreas distintas dentro de um fragmento grande podem variar tanto entre si, quanto em relação a
áreas isoladas pela fragmentação. Isto indica que essas variações podem ser conseqüência da
heterogeneidade ambiental ao longo da paisagem e não simplesmente um efeito da fragmentação.
Todas as análises realizadas no presente estudo indicaram que a riqueza e a diversidade de
espécies arbóreas nos fragmentos estudados não parecem estar relacionadas à área total desses
fragmentos. É possível que não encontramos a relação esperada devido à idade dos fragmentos ou
tempo de fragmentação. Butaye et al. (2002) encontraram relação entre área e riqueza somente em
fragmentos com idades superiores a 50 anos. Segundo Jacquemyn et al. (2001), não seria possivel
encontrar uma relação significativa entre área e riqueza de espécies em fragmentos isolados há
menos de 35 anos. Todos os fragmentos estudados aqui, com exceção da mata P4, exibem a mesma
forma e tamanho há pelo menos 40 anos (Capítulo 1). Apesar disso, é possível que muitas das
espécies encontradas nesses fragmentos tenham permanecido nessas áreas simplesmente por sua
longevidade (Condit 1996). Para Turner & Colett (1996), a área de um fragmento pode restringir o

98
tamanho das populações, mas até hoje não se sabe como determinar o tamanho populacional
mínimo para garantir a manutenção de uma espécie em uma área. Dessa forma, pode ser que
algumas dessas espécies estejam regenerando somente nos fragmentos onde restaram populações
viáveis e isso certamente terá influência sobre a riqueza de espécies no futuro. É possível também
que muitas espécies já tenham desaparecido dos fragmentos ao longo do tempo, mas essa é uma
hipótese difícil de ser testada, dada a ausência de estudos anteriores à fragmentação.
As diferenças de riqueza entre florestas para Sheil (1999), podem estar associadas a uma série
de fatores, como estádio sucessional, contexto ambiental, eventos estocásticos e também aos
métodos e escalas empregados na medição. Para Rey Benayas & Scheiner (2002), as condições
ambientais e biogeográficas seriam os fatores preponderantes sobre a diversidade de plantas.
Enquanto para Cadotte et al. (2002) essas diferenças estariam relacionadas a fatores edáficos,
condições microclimáticas e freqüência e intensidade de perturbações antrópicas. No caso dos
fragmentos do presente estudo, as variações na riqueza parecem estar associadas à heterogenidade
ambiental intrínseca da região e à ocorrência e intensidade de fatores de perturbação. O fragmento
G2, por exemplo, apesar de ser o de maior área entre os fragmentos menores que 100ha, apresentou
uma das menores riquezas e número de espécies coletadas do estudo. A área G2 apresenta histórico
de perturbação bastante intenso. Segundo informações locais, houve uma grande queimada em
1996, que atingiu a área quase por completo. Isto pode estar relacionado à baixa riqueza na área
(Capítulos 1 e 2). Além disso, as variações observadas podem ser resultado da combinação de
diversos eventos, pois os fragmentos apresentam diferentes combinações de condições abióticas e
históricos de preservação bastante variáveis (Capítulo 1). Por exemplo, algumas dessas áreas podem
ser remanescentes de uma floresta outrora contínua, enquanto outras podem ser áreas reflorestadas
naturalmente, isto é, áreas anteriormente utilizadas pelo homem, que foram abandonadas e sofreram
regeneração. O fragmento P4 é um exemplo disso (Capítulo 1).
O fragmento P3 exibiu os menores índices de similaridade com as demais amostras. Esta mata
apresenta solo e relevo distintos dos demais fragmentos (com exceção de G2 – ver Capítulos 1 e 2).
O baixo número de espécies dessa área certamente a torna distinta das demais, daí os índices de
similaridade obtidos. Entretanto, das 47 espécies coletadas na área, 27 ocorreram em pelo menos
outros nove fragmentos, reforçando a hipótese de que seria o baixo número de espécies que a torna
distinta das demais áreas. Outra particularidade desta mata é que dentre os 500 indivíduos coletados,
229 pertenceram a uma única espécie, Trichilia clausseni. Essa espécie ocorreu em várias outras
amostras, mas com 52 indivíduos, no máximo. A grande dominância dessa espécie na área pode
estar restringindo a ocorrência de outras espécies e talvez por isso a riqueza e a diversidade foram
inferiores às demais amostras. Segundo Connel & Lowman (1989), a diversidade de árvores
decresce com o aumento da dominância de uma única espécie. Os autores estudaram florestas de
baixa diversidade em regiões em que o estoque regional de espécies era bastante elevado e
observaram que a ocorrência de florestas dominadas por uma única espécie (monodominância) não é

99
rara, ao contrário, essas florestas são encontradas em cada uma das principais regiões de domínio
das florestas tropicais. Segundo os autores, existem dois mecanismos através dos quais uma espécie
pode se tornar dominante em uma área. Um deles seria colonizando uma ampla área aberta e lá
persistindo por mais de uma geração, o outro seria através da reposição árvore a árvore, com a
espécie tornando-se dominante gradualmente. A monodominância, para Cadotte et al. (2002), pode
ser conseqüência da remoção de espécies competidoras ou da redução do número ou abundância de
predadores naturais. Entretanto, para Torti et al. (2001), não deve estar relacionada simplesmente a
um único fator e sim a uma combinação das condições ambientais e particularidades do histórico do
desenvolvimento daquela condição. De qualquer forma, tais particularidades exibidas pela mata P3
aumentam sua importância em termos de conservação.
No presente trabalho, observamos que os fragmentos maiores não são necessariamente mais
ricos do que fragmentos menores e que áreas distintas dentro de um fragmento grande podem conter
tantas espécies quanto um fragmento pequeno. Além disso, a riqueza em áreas distintas dentro de
um fragmento grande pode variar tanto entre si quanto em relação a áreas isoladas pela
fragmentação. Isto indica que essas variações podem ser conseqüência da heterogeneidade
ambiental ao longo da paisagem e não simplesmente a um efeito da fragmentação ou da área do
fragmento remanescente. Como mencionado acima, existem muitos fatores que podem estar
envolvidos na perda de riqueza em fragmentos e para Willians-Linera (2002) todos eles agem
sinergisticamente.
Segundo Leitão Filho (1982), as florestas estacionais semidecíduas ocupam uma ampla área no
interior do estado de São Paulo, percorrendo diversos tipos de solo e relevo e encontra-se
entremeado por outros tipos vegetacionais, como matas ciliares, cerrados e cerradões. Para o autor,
essas cacaracterísticas contribuiriam para a heterogeneidade entre os remanescentes atuais. No
passado, as florestas na região da APA eram praticamente contínuas (Christofoletti & Federici 1972,
Mattos 1996) e essas observações, aliadas aos resultados obtidos aqui poderiam sugerir que,
anteriormente à fragmentação, áreas da floresta tomadas ao acaso, ao longo daquele contínuo,
seriam tão diferentes entre si quanto são hoje em dia, após a fragmentação. Isto não quer dizer que a
fragmentação não tenha efeitos sobre a riqueza de espécies arbóreas nesses fragmentos. Como
mencionado acima, a perda de espécies pode ocorrer em uma escala de tempo difícil de ser
acessada e isso aumenta a importância desses fragmentos para a preservação. Estudos sobre a
dinâmica das populações nessas áreas são imprescindíveis para o melhor entendimento das
observações do presente estudo e também para a adoção de medidas de preservação e recuperação
seguras.
Nossos resultados concordaram com o proposto por Turner & Corlett (1996), para os quais
fragmentos pequenos (<100ha) podem ser capazes de suportar um grande número de espécies. Isto
mostra a importância desses fragmentos para a conservação do estoque regional de espécies.
Entretanto, devemos lembrar que os fragmentos grandes (>100ha), como a Mata Ribeirão Cachoeira,

100
também são fundamentais nesse sentido. Fragmentos grandes englobam maior heterogeneidade
ambiental e com isso sua contribuição para a riqueza regional acaba sendo maior. As três amostras
de Ribeirão Cachoeira, por exemplo, exibiram juntas 151 espécies, mais da metade do total coletado
no presente estudo. De qualquer forma, florestas contínuas e fragmentos estão sujeitos a condições e
processos diferentes. Por isso, fragmentos se tornaram novos tipos de comunidades e a conservação
de espécies em seu interior pode depender da minimização do impacto dessas alterações ao longo
do tempo (Brokaw 1998).

101
Referêcias bibliográficas AIZEN, M.A. & FEINSINGER, P. 1994. Forest fragmentation, pollination, and plant reproduction in a
chaco dry forest, Argentina. Ecology 75: 330-351.
AYRES, M., AYRES Jr., M., AYRES, D.M. & SANTOS, A.S. 1998. BioEstat: Aplicações estatísticas nas áreas das Ciências Biológicas e Médicas. Sociedade civil Mamirauá. Manaus.
BERNACCI, L.C. & LEITÃO FILHO, H.F 1996. Flora fanerogâmica da floresta da Fazenda São
Vicente, Campinas, SP. Revista Brasileira de Botânica 19: 149-164.
BERTONI, J.E.A., MARTINS,F.R., MORAES, J.L. & SHEPHERD, G.J. 1988. Composição florística e
estrutura fitossociológica do Parque Estadual de Vaçununga, Santa Rita do Passa Quatro –
Gleba Praxedes, Boletim Técnico do Instituto Florestal de São Paulo 42: 149-170.
BIERREGAARD, R.O., LOVEJOY, T.E., KAPOS, V., SANTOS, A.A. & HUTCHINGS, R.W. 1992. The
biological dynamics of tropical rainforest fragments. Bioscience 42: 859-866.
BROKAW, N. 1998. Fragments past, present and future. Trends in Ecology and Evolution 13: 382-
383.
BUTAYE, J., JACQUEMYN, H., HONNAY, O. & HERMY, M. 2002. The species pool concept applied
to forests in a fragmented landscape: dispersal limitation versus habitat limitation. Journal of Vegetation Science 13: 27-34.
CADOTTE, M.W., FRANCK, R., REZA, L. & LOVETT-DOUST, J. 2002. Tree and shrub diversity and
abundance in fragmented littiral forest of southeastern Madagascar. Biodiversity and Conservation 11: 1417-1436.
CAVASSAN, O., CESAR, O. & MARTINS, F.R. 1984. Fitossociologia da vegetação arbórea da
reserva estadual de Bauru, Estado de São Paulo. Revista Brasileira de Botânica 7: 91-106.
CESAR ,O. & LEITÃO FILHO, H.F 1990. Estudo florístico quantitativo de mata mesófila semidecídua
na fazenda Barreiro Rico, município de Anhembi, SP. Revista Brasileira de Botânica 50: 133-
147.
CHAO, A. 1984. Non-parametric estimation of the number of classes in a population. Scandinavian
Journal of Statistics 11: 265-270.
CHAO, A. 1987. Estimating the population size for capture-recapture data with inequal catchability.
Biometrics 43: 783-791.
CHRISTOFOLETTI, A. & FEDERICI, H. 1972. A terra campineira. Indústrias gráficas Mousinho.
COLWELL, R. K. & CODDINGTON, J. A. 1994. Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions: Biological Sciences 345: 101-118.
CONDIT, R. 1996. Defining and mapping Vegetation types in mega-diverse tropical forests. Trends in Ecology and Evolution 11: 4-5.
CONNELL, J.H. & LOWMAN, M.D. 1989. Low-diversity tropical rain forests: some possible
mechanisms for their existence. The American Naturalist 134: 88-119

102
CORNELL, H.V. 1999. Unsaturation and regional influences on species richness in ecological
communities:a review of the evidence. Ecoscience 6: 306-315.
COSTA, L.G.S. 1992. Estrutura e dinâmica de trecho de mata mesófila semidecídua na Estação Ecológica de Ibicatú, Piracicaba, SP. Dissertação de Mestrado. Universidade de São Paulo,
São Paulo.
COTTAN, G. & CURTIS, J.T. 1956. The use of distance measures in phytossociological sampling.
Ecology 37: 451-460.
CRONQUIST, A. 1981. An integrated system of classification of flowering plants. NewYork:
Columbia University Press & The New York Botanical Garden.
CURTIS, J.T. 1956. The modification of mid-latitude grasslands and forests by ants. Pp 721-736. In:
W.L. Thomas (ed.). Man’s role in changing the face of the earth. University os Chicago Press.
Chicago.
DIAMOND, J.M. & MAY, R.M. 1976. Island biogeography and the design of natural reserves. Pp 163-
186. In: R.M. May (ed.) Theoretical Ecology: principles and applications. Saunders.
Philadelfia.
DIGBY, P.G.N. & KEMPTON, R.A. 1987. Multivariate analysis of ecological communities.
Chapman & Hall. Londres.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista Brasileira de Botânica 23: 371-383.
GOTELLI, N. J. & COWELLl, R. K. 2001. Quantifying biodiversity: procedures and pitfalls in the
measurement and comparison of species richness. Ecological Letters 4: 379-391.
HAMMER, O., HARPER, D.A.T. & RYAN, P.D. 2001. PAST: Paleontological Statistics Software
Package for Education and Data Analysis. Paleontologia Electronica 4(1): 9pp. http://palaeo-
electronica.org/2001_1/past/issue1_01.htm
HARRIS, L.D. 1980. Forest and wildlife dynamics in the southeast. Transactions of the North
American wildlife and natural resourses conference 45: 307-322.
HARRIS, L.D. 1984. The fragmented forest: island biogeography theory and the preservation of biotic diversity. University of Chicago Press. Chicago.
HILL, J.L. & CURRAN, P.J. 2001. Species composicion in fragmented forests: conservation
implications of changing forest area. Applied Geography 21: 157-174.
JACQUEMYN, H., BUTAYE, J., & HERMY, M. 2001. Forest plant species richiness in small,
fragmented mixed deciduous forest patches: the role of area, time and dispersal limitations.
Journal of Biogeography 28: 801-812.
KAPOS, V., WANDELLI, E., CAMARGO, J.L. & GANADE, G. 1997. Edge-related changes in
environment and plant responses due to forest fragmentation in Central Amazonia. Pp 33-44. In:

103
W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: Ecology, management, and conservation of forest communities. University of Chicago Press. Chicago.
KÖPPEN, W. 1948. Climatologia. México - Buenos Aires. Ed. Fundo de Cultura Econômica, 478p.
LANDE, R. 1988. Genetics and demography in biological conservation. Science 242: 1455-1460.
LAURANCE, W.F. 1997. Hyper-disturbed parks: edge effects and the Ecology of isolated rainforest
reserves in tropical Australia. Pp 71-83. In: W.F. Laurance & R.O Bierregaard (ed.). Tropical forest remnants: Ecology, management, and conservation of forest communities. University
of Chicago Press. Chicago.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M., LAURANCE, S.G., HUTCHINGS,
R.W. & LOVEJOY, T.E. 1998a. Effects of forest fragmentation on recruitment patterns in
amazonian tree communities. Conservation Biology 12: 460-464.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M. & LAURANCE, S.G. 1998b. Rain
forest fragmentation and the dynamics of amazoniam tree communities. Ecology 79: 2032-2040.
LAURANCE, W.F., PÉREZ-SALICRUP, D., DELAMÔNICA, P., FEARNSIDE, P.M., D’ANGELO, S.,
JEROLINSKI, A., POHL, L. & LOVEJOY, T.E. 2001. Rain forest fragmentation and structure of
amazonian liana communities. Ecology 82: 105-116.
LEITÃO FILHO, H.F. 1982. Aspectos taxonômicos das florestas do Estado de São Paulo. Silvicultura em São Paulo 16: 197-206.
LOVEJOY, T.E., BIERREGAARD, R.O., RYLANDS , A.B., MALCOLM, J.R., QUINTELA, C.E.,
HARPER, L.H., BROWN, K.S., POWELL, A.H., POWELL, G.V.N., SCHUBART, H.O.R. & HAYS
.B. 1996. Edge and other effects of isolation on Amazon forest fragments. Pp 257-285. In: M.E.
Soule (ed.). Conservation Biology: the science of scarcity and diversity. Sinauer, Suderland.
USA
LUDWIG, J.A. & REYNOLDS, J.F. 1988. Statistical Ecology. A primer on methods and computing. J. Wiley & Sons, New York.
MAGURRAN, A. 1988. Ecological diversity and its measurement. Croom Helm, London
MATTHES, L.A.F., LEITÃO FILHO, H.F. & MARTINS, F.R. 1988. Bosque dos Jequitibás (Campinas,
SP): composição florística e fitossociológica do estrato arbóreo. Anais V congresso da Sociedade Botânica de São Paulo, p. 55-76.
MATTOS, C.O. 1996. Contribuição ao planejamento e gestão da Área de Proteção Ambiental de Sousas e Joaquim Egídio, Campinas/SP. Dissertação de Mestrado. Universidade Estadual de
São Paulo. São Paulo.
MELLO, M.H.A., PEDRO JUNIOR, M.J., ORTOLANI, A. A. & ALFONSI R.R. 1994. Chuva e
Temperatura: cem anos de observações em Campinas. Boletim Técnico 154 . Instituto
Agronômico Campinas, SP.
METZGER, J.P., BERNACCI, L.C. & GOLDEMBERG, R. 1997. Pattern of tree species diversity in
riparian forest fragments of different widths (SE Brazi). Plant Ecology 133: 135-152.

104
MIRANDA, Z.A.I. 1996. Plano de gestão da Área de Proteção Ambiental da região de Sousas e
Joaquim Egídio - APA Municipal. Relatório da Secretaria de Planejamento e Meio Ambiente - SEPLAMA. Prefeitura Municipal de Campinas, SP.
OLIVEIRA, J.B., MARCELO, N.C., ROSSI, M. & CALDERANO FILHO, B. 1999. Mapa pedológico do
estado de São Paulo. Legenda expandida In: OLIVEIRA, J.B. 1999. Boletim científico n.o 45,
Solos do estado de São Paulo. Embrapa e IAC. Campinas.
PAGANO, S.N.,LEITÃO FILHO, H.F. CAVASSAN, O. 1995. Variação temporal da composição
florística e estrutura fitossociológica de uma floresta mesófila semidecídua – Rio Claro – Estado
de São Paulo. Revista Brasileira de Biologia 55: 241-258.
PÄRTEL, M. ZOBEL, M. ZOBEL, K. & van der MAAREL, E. 1996. The species pool and its relation to
species richiness: evidence from Estonian plant communities. Oikos 75: 111-117.
POWELL, A.H. & POWELL, G.V.N., 1987. Population dynamics of male Euglossine bees in
Amazonian forest fragments. Biotropica 19: 176-179
PRENDERGAST, J.R., QUINN, R.M., LAWTON, J.H., EVERSHAM, B.C. & GIBBONS, DW. 1993.
Rare species, the coincidence of diversity hotspots and conservation strategies. Nature 365: 335-
367
PRESSEY, R.L., HUMPHRIES, C.J., MARGULES, C.R., VANE-WRIGHT, R.I. & WILLIAMS, P.H.
1993. Beyond the opportunism: key principles for systematic reserve selection. Trends in Ecology and Evolution 8: 124-128.
REY BENAYAS, J.M. & SCHEINER, S.M. 2002. Plant diversity, biogeography and environment in
Iberia: patterns and possible causal factors. Journal of Vegetation Science 13: 245-258.
RICKLEFS, R.E. & SCHLUTER, D. 1993. Species diversity: regional and historical influences. Pp 350-
363. In: R.E. RICKLEFS & D. SCHLUTER (ed.) Species diversity in ecological communities.
University of Chicago Press. Chicago.
RODRIGUES, R. R., MORELLATO, L.P.C., JOLY, C.A. & LEITÃO FILHO, H.F. 1989. Estudo florístico
e fitossociológico em um gradiente altitudinal de mata estacional mesófila semidecídua, na Serra
do Japi, Jundiaí, SP. Revista Brasileira de Botânica 12: 71-84.
SANTIN, D.A. 1999. A vegetação remanescente do município de Campinas (SP): mapeamento, caracterização fisionômica e florística, visando a conservação. Tese de doutorado.
Universidade Estadual de Campinas, SP.
SANTOS, K. & KINOSHITA, L.S. 2003. Flora arbustivo-arbórea do fragmento de floresta estacional
semidecidual do Ribeirão Cachoeira, Município de Campinas - SP. Acta Botanica Brasilica 17:
325-341.
SAUNDERS, D.A., HOBBS, R.J. & MARGULES, C.R. 1991. Biological consequences of ecossystem
fragmentation: a review. Conservation Biology 5: 18-32.
SHAFER, M.L. 1981. Minimum popuations sizes for species conservation. Bioscience 31: 131-134.

105
SHEIL, D. 1999. Tropical forest diversity, environmental change and species augmentation: after the
intermediate disturbance hypothesis. Journal of Vegetation Science 10: 851-860.
SHEPHERD, G.J. 1988. FITOPAC- versão preliminar. Manual do usuário. Departamento de
Botânica. Instituto de Biologia, Universidade Estadual de Campinas, Campinas-SP.
STOUFFER, P.C. & BIERREGAARD, R.O. 1995a. Effects of forest fragmentation on understory
hummingbirds in Amazon Brazil. Conservation Biology 9: 1085-1094
STOUFFER, P.C. & BIERREGAARD, R.O. 1995b. Use of Amazonian forest fragments by
insectivorous birds. Ecology 76: 2429-2445.
TABARELLI, M., MANTOVANI, W. & PERES C.A. 1999. Effects of habitat fragmentation on plant guild
structure in the montane Atlantic forest of southeastern Brazil. Biological Conservation 91: 119-
127.
TORTI, S.D., COLEY, P.D. & KURSAR, T.A. 2001. Causes and consequences of monodominance in
tropical lowland forests. The American Naturalist 157: 141-153.
TURNER, I.M. 1996. Species loss in fragments of tropical rain forest: a review of evidence. Journal of
Applied Ecology 33: 200-209.
TURNER, I.M. & CORLETT, R.T. 1996. The conservation value of small, isolated fragments of
lowland tropical rain forest. Trends in Ecology and Evolution 11: 330-333.
TURNER, I.M., CHUA, K.S., ONG,J., SOONG, B. & TAN, H. 1996. A century of plant species loss
from a isolated fragment of lowland tropical rainforest. Conservation Biology 10: 1229-1244.
VELOSO, H.P., RANGEL FILHO, A .L.R. & LIMA, J.C. 1991. Classificação da vegetação brasileira adaptada a um sistema universal. Rio de Janeiro. Instituto Brasileiro de Geografia e Estatística
(IBGE).
WILLIANS, N. 1997. Rain forest fragments fare poorly. Science 278: 1016.
WILLIANS-LINERA, G. 2002. Tree species richness complementarity, disturbance and fragmentation
in Mexican tropical montane cloud forest. Biodiversity and Conservation 11: 1825-1843.
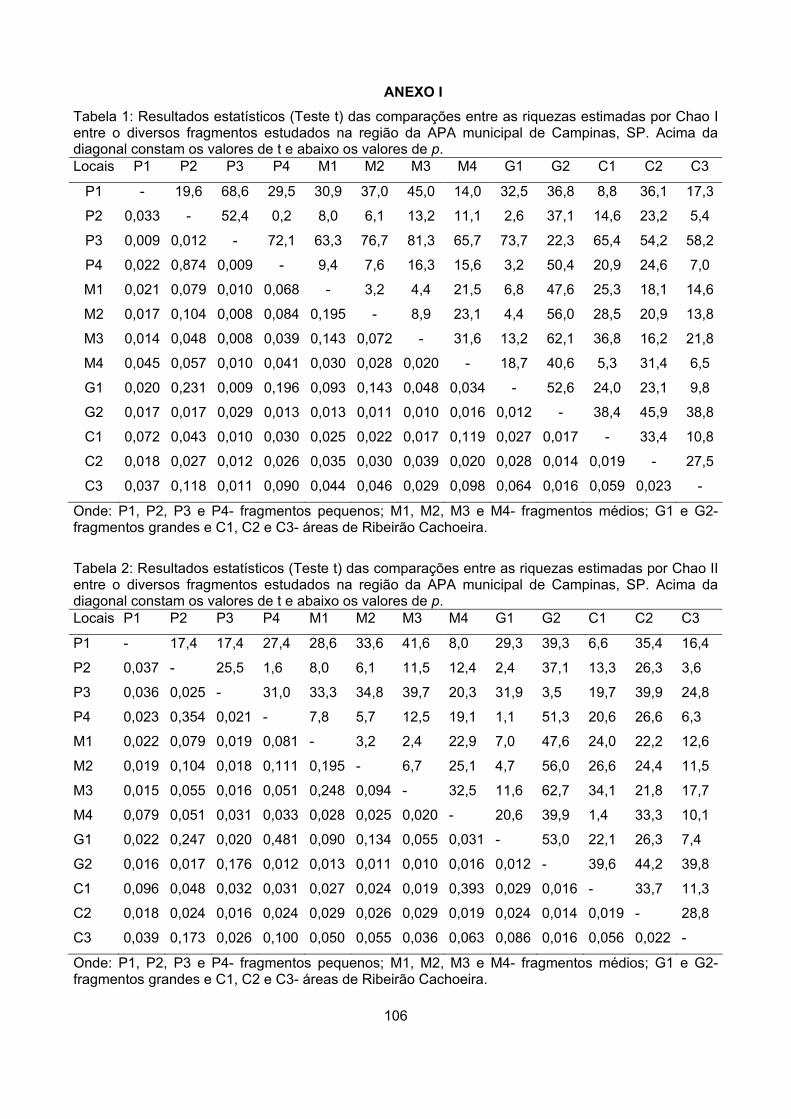
106
ANEXO I Tabela 1: Resultados estatísticos (Teste t) das comparações entre as riquezas estimadas por Chao I entre o diversos fragmentos estudados na região da APA municipal de Campinas, SP. Acima da diagonal constam os valores de t e abaixo os valores de p. Locais P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
P1 - 19,6 68,6 29,5 30,9 37,0 45,0 14,0 32,5 36,8 8,8 36,1 17,3
P2 0,033 - 52,4 0,2 8,0 6,1 13,2 11,1 2,6 37,1 14,6 23,2 5,4
P3 0,009 0,012 - 72,1 63,3 76,7 81,3 65,7 73,7 22,3 65,4 54,2 58,2
P4 0,022 0,874 0,009 - 9,4 7,6 16,3 15,6 3,2 50,4 20,9 24,6 7,0
M1 0,021 0,079 0,010 0,068 - 3,2 4,4 21,5 6,8 47,6 25,3 18,1 14,6
M2 0,017 0,104 0,008 0,084 0,195 - 8,9 23,1 4,4 56,0 28,5 20,9 13,8
M3 0,014 0,048 0,008 0,039 0,143 0,072 - 31,6 13,2 62,1 36,8 16,2 21,8
M4 0,045 0,057 0,010 0,041 0,030 0,028 0,020 - 18,7 40,6 5,3 31,4 6,5
G1 0,020 0,231 0,009 0,196 0,093 0,143 0,048 0,034 - 52,6 24,0 23,1 9,8
G2 0,017 0,017 0,029 0,013 0,013 0,011 0,010 0,016 0,012 - 38,4 45,9 38,8
C1 0,072 0,043 0,010 0,030 0,025 0,022 0,017 0,119 0,027 0,017 - 33,4 10,8
C2 0,018 0,027 0,012 0,026 0,035 0,030 0,039 0,020 0,028 0,014 0,019 - 27,5
C3 0,037 0,118 0,011 0,090 0,044 0,046 0,029 0,098 0,064 0,016 0,059 0,023 -
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
Tabela 2: Resultados estatísticos (Teste t) das comparações entre as riquezas estimadas por Chao II entre o diversos fragmentos estudados na região da APA municipal de Campinas, SP. Acima da diagonal constam os valores de t e abaixo os valores de p. Locais P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
P1 - 17,4 17,4 27,4 28,6 33,6 41,6 8,0 29,3 39,3 6,6 35,4 16,4
P2 0,037 - 25,5 1,6 8,0 6,1 11,5 12,4 2,4 37,1 13,3 26,3 3,6
P3 0,036 0,025 - 31,0 33,3 34,8 39,7 20,3 31,9 3,5 19,7 39,9 24,8
P4 0,023 0,354 0,021 - 7,8 5,7 12,5 19,1 1,1 51,3 20,6 26,6 6,3
M1 0,022 0,079 0,019 0,081 - 3,2 2,4 22,9 7,0 47,6 24,0 22,2 12,6
M2 0,019 0,104 0,018 0,111 0,195 - 6,7 25,1 4,7 56,0 26,6 24,4 11,5
M3 0,015 0,055 0,016 0,051 0,248 0,094 - 32,5 11,6 62,7 34,1 21,8 17,7
M4 0,079 0,051 0,031 0,033 0,028 0,025 0,020 - 20,6 39,9 1,4 33,3 10,1
G1 0,022 0,247 0,020 0,481 0,090 0,134 0,055 0,031 - 53,0 22,1 26,3 7,4
G2 0,016 0,017 0,176 0,012 0,013 0,011 0,010 0,016 0,012 - 39,6 44,2 39,8
C1 0,096 0,048 0,032 0,031 0,027 0,024 0,019 0,393 0,029 0,016 - 33,7 11,3
C2 0,018 0,024 0,016 0,024 0,029 0,026 0,029 0,019 0,024 0,014 0,019 - 28,8
C3 0,039 0,173 0,026 0,100 0,050 0,055 0,036 0,063 0,086 0,016 0,056 0,022 -
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.

107
CAPÍTULO 4
ESTRUTURA E ABUNDÂNCIA DE ESPÉCIES EM ONZE FRAGMENTOS DE FLORESTA ESTACIONAL
SEMIDECIDUA NA REGIÃO DA ÁREA DE PROTEÇÃO AMBIENTAL DO MUNICÍPIO DE CAMPINAS, SP

108
Capítulo 4
Estrutura e abundância de espécies em onze fragmentos de floresta estacional semidecídua na região da Área de Proteção Ambiental do município de Campinas, SP
Karin dos Santos Pós-Graduação em Biologia Vegetal – Universidade Estadual de Campinas – Parte da tese de doutorado,
Luiza Sumiko Kinoshita Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Flavio A. M. dos Santos Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Resumo O processo de fragmentação florestal e a conseqüente exposição das comunidades a fatores de
perturbação têm sido associados a alterações na dinâmica das comunidades, modificando sua
estrutura original. O objetivo deste trabalho foi investigar se as variações na estrutura florestal de
onze fragmentos de mata estacional semidecídua estavam relacionadas à sua área total. Segundo as
proposições da literatura, seria esperado que fragmentos pequenos apresentassem maior densidade
de indivíduos do que fragmentos grandes, havendo ainda diferenças na distribuição e abundância das
espécies, pois muitas delas não sobreviverem em áreas reduzidas. Na região da Área de Proteção
Ambiental do município de Campinas, SP, foram escolhidos dez fragmentos com áreas entre 12 e
65ha e um com 244,9ha. Em cada fragmento <65ha foi realizada uma amostragem de 125 pontos
quadrantes, onde foram incluídos indivíduos arbóreos com DAP ≥ 10cm. As amostras de 500
indivíduos foram comparadas com amostras feitas nas mesmas condições em três locais diferentes
do fragmento Ribeirão Cachoeira, de 244,9ha. Ao todo, foram amostrados 6500 indivíduos
pertencentes a 248 espécies, 149 gêneros e 55 famílias. A densidade de indivíduos foi 40% menor
nos fragmentos grandes e nas áreas de Ribeirão Cachoeira, mas a área basal não diferiu. As famílias
e espécies de maior VI foram as mesmas comumente apontadas como tal em outros estudos
realizados em remanescentes deste ecossistema. A distribuição das abundâncias das espécies
variou bastante entre as amostras, mas em todas elas houve concentração de abundância, com
poucas espécies exercendo grande dominância em sua área de ocorrência. Não houve correlação
entre a área dos fragmentos e as densidades totais e os demais parâmetros de estrutura e
abundância analisados. Fatores como ocorrência de perturbação e heterogeneidade ambiental
podem estar envolvidos nas variações observadas.

109
Introdução No estado de São Paulo muitos remanescentes florestais foram estudados do ponto de vista
florístico e estrutural nas últimas duas décadas (ver Meira Neto et al. 1989, Torres et al. 1997, Dislich
et al. 2001, Santos & Kinoshita 2003). A heterogeneidade na estrutura arbórea de florestas
estacionais semidecíduas pode ser reconhecida através de uma rápida análise dos estudos
realizados nos diversos remanescentes de mata do interior do estado de São Paulo (Cavassan et al.
1984, Matthes et al. 1988, Bertoni et al. 1988, Rodrigues et al. 1989, Grombone et al. 1990, Martins
1991, Baitello et al. 1992, Salis et al. 1994, Tabanez et al. 1997, Duringan et al. 2000, Cielo Filho &
Santin 2002, Silva & Soares 2002). Parâmetros como densidade e área basal variam enormemente,
mesmo entre estudos que adotaram os mesmos métodos de amostragem e critérios de inclusão.
As variações na estrutura de florestas tropicais podem estar associadas ao tipo de solo
(Oliveira-Filho et al. 1997, 1994a, Clark & Clark 2000), fertilidade do solo (Laurance et al. 1999),
perturbações naturais (Denslow 1995, Lugo & Scatena 1996), topografia (Clark & Clark 2000), altitude
(Lieberman et al. 1985) e perturbações antrópicas (Brown et al. 1995, Laurance et al. 1997). Além
disso, as variações também ocorrem ao longo do tempo, influenciadas pela dinâmica da comunidade
e das diferentes espécies que a compõem (Rankin-de-Merona et al. 1990, Felfili 1995, Lieberman et
al. 1985, Santos et al. 1996).
Variações estruturais podem ocorrer ao longo de distâncias relativamente curtas (Popma et al.
1988). Em função disso, Santos et al. (1998) propuseram o seguinte questionamento: as diferenças
encontradas entre levantamentos feitos em diferententes localidades também ocorreriam se várias
amostras feitas ao longo de uma mesma floresta fossem comparadas? Para os autores, a ausência
de réplicas nos levantamentos produzidos em cada localidade dificultam a elucidação dessa questão.
Com a fragmentação de florestas outrora contínuas, surgiram novos componentes de variação,
como o efeito de borda, resultante da interação entre a floresta e o ecossistema circundante (Murcia
1995). Esta interação ocasionaria uma série de alterações bióticas e abióticas nos remanescentes
que levariam a um aumento das taxas de mortalidade e de recrutamento de indivíduos arbóreos,
principalmente nas regiões de borda e em fragmentos pequenos (Willians-Linera 1990, Laurance et
al. 1998a, Mesquita et al. 1999) e resultariam em alterações nos padrões de abundância das
espécies florestais (Murcia 1995). Além disso, espécies características dos estádios iniciais de
sucessão poderiam ser favorecidas nas novas condições microclimáticas da floresta fragmentada
(Tabarelli et al. 1999). Willians-Linera (1990) observou que áreas de borda de fragmentos, por
estarem mais expostas às perturbações externas, apresentavam maior densidade de indivíduos e
área basal em relação às áreas de interior da floresta.
A amplitude e intensidade do efeito de borda são influenciadas pela área e forma dos
fragmentos, idade e orientação espacial da borda e outros fatores (Kapos 1989, Matlack 1994, Murcia
1995, Didham & Lawton 1999). Para alguns autores, o tamanho dos fragmentos exerceria um papel
preponderante, pois quanto menor o fragmento maior seria a influência do efeito de borda, devido a

110
sua maior razão borda:interior (Saunders et al. 1991, Turner 1996, Laurance et al. 1998a, 1998b).
Laurance et al. (1998a) mostraram que em fragmentos florestais as taxas de recrutamento de
indivíduos arbóreos aumentam com a redução da área total do fragmento, levando a um aumento da
densidade nessas áreas.
Estudos quantitativos de comunidades florestais são uma forma de descrever sua estrutura
através do número de indivíduos por unidade de área, número de indivíduos por espécie ou da área
basal que determinada espécie ocupa na comunidade. Um de seus objetivos é detectar padrões e
tentar relacioná-los a processos ou aspectos do ambiente físico ou biótico que estariam influenciando
a estrutura observada. Por exemplo, o modo como uma população ocupa o espaço de uma
comunidade pode indicar a maneira como ela responde à distribuição espacial de recursos ou ser
resultado da ação de processos diferenciados ao longo do tempo (Condit et al. 1996).
Processos diferentes produziriam efeitos diferentes sobre as comunidades (Burslem & Whitmore
1999). Por exemplo, florestas mais expostas a fatores de perturbações, como fragmentos pequenos,
fatalmente exibiriam diferenças de composição e de estrutura em relação a florestas menos expostas,
como fragmentos maiores ou florestas contínuas. Dessa forma, seria esperado que fragmentos
pequenos fossem estruturalmente mais homogêneos entre si, pois estariam expostos de forma similar
às alterações relacionadas ao efeito de borda mencionado acima.
O presente trabalho teve por objetivo investigar as variações na estrutura da vegetação arbórea
de onze fragmentos de floresta estacional semidecídua, visando responder às seguintes perguntas:
amostragens feitas em diferentes pontos de uma área contínua são mais heterogêneas entre si, do
ponto de vista estrutural, do que entre fragmentos isolados ao longo da paisagem? As variações na
estrutura da vegetação de fragmentos de tamanhos diversos poderiam ser atribuídas à área total dos
mesmos? Fragmentos menores apresentam maior densidade de indivíduos do que fragmentos
grandes ou uma área contínua?

111
Materiais e métodos Área de estudo
Os fragmentos estudados localizam-se, predominantemente na região da Área de Proteção
Ambiental (APA) de Sousas e Joaquim Egídio, município de Campinas (22°45’ a 23°00’ S, 47°00’ a
47°12’ W) e no município de Itatiba (23°10’ a 23°15’ S, 45°45’ a 46°45’ W). O clima, segundo Köppen
(1948), é Cwag'- temperado, moderadamente chuvoso, de inverno seco não rigoroso, com
precipitação média anual de 1409 mm (Mello et al. 1994). Os solos ocupados pelos fragmentos em
estudo podem ser Podzólicos Vermelho-Amarelos de três diferentes tipos: PVA 8, PVA 20 e PVA ou
Latossolo Vermelho: LV 4 (Oliveira et al. 1999). Os relevos podem ser suave ondulados, forte
ondulados e montanhosos ou ondulados e suave ondulados (Oliveira et al. 1999). As altitudes variam
de 650 a 890m (Capítulo 1). A vegetação dos fragmentos pode ser classificada floresta estacional
semidecídua, nomenclatura proposta por Veloso et al. (1991).
Foram escolhidos dez fragmentos pequenos com áreas entre 12 e 63 hectares (ha) e um
fragmento grande - Mata Ribeirão Cachoeira - (244,9 ha), cujo levantamento florístico foi realizado
por Santos & Kinoshita (2003). Os fragmentos menores do que 100ha foram categorizados em três
classes de tamanho: quatro pequenos, com áreas entre 12 e 13,5ha (P1, P2, P3 e P4), quatro
médios, com áreas entre 19 e 25,5ha (M1, M2, M3 e M4) e dois grandes com áreas de 41,8 e 63,3ha
(G1 e G2). Os critérios para a escolha de cada fragmento foram: similaridade fisionômica, facilidade
de acesso e obtenção de autorização dos proprietários para a realização do trabalho.
Na região da Área de Proteção Ambiental do município de Campinas a fragmentação de
florestas teve início com o desenvolvimento da agricultura, durante o ciclo da cana de açúcar, no
século XVIII (Miranda 1996, Mattos 1996). Desde aquela época, a vegetação vem sendo
extensivamente modificada e as florestas estacionais, que recobriam toda a APA e grande parte do
município de Campinas, hoje representam somente 2,42% da área municipal (Santin 1999). Todos os
fragmentos do estudo apresentam aproximadamente a mesma forma e tamanho atuais há pelo
menos 40 anos, com exceção da mata P4 que vem aumentando em área nos últimos 20 anos (ver
Capítulo 1). Os fragmentos que mais sofreram perturbações como a ocorrência de queimadas foram:
o fragmento P2, atingido parcialmente pelo fogo há alguns anos e sujeito à queimadas freqüentes na
região de entorno; o fragmento G2, atingido quase totalmente por uma queimada bastante intensa no
ano de 1996; o fragmento G1 e a área C3 da mata Ribeirão Cachoeira, atingidos parcialmente por
queimadas no ano de 1994 e o fragmento M1 atingido parcialmente por queimadas há menos de dez
anos (ver Capítulo 1).
A paisagem em que os fragmentos estão inseridos constitui-se de um mosaico de culturas
anuais e perenes, pastagens, reflorestamentos e pequenos assentamentos humanos. A agricultura é
caracterizada por pequenas áreas de café, laranja, cana de açúcar e culturas de subsistência. Os
reflorestamentos com eucalipto e pastagens ocupam grandes extensões (Miranda 1996).

112
Amostragem
Para a coleta de dados foi adotado o método de pontos quadrantes proposto por Cottam &
Curtis (1956). Cada amostra consistiu de 125 pontos quadrantes, onde foram coletados indivíduos
arbóreos com diâmetro à altura do peito (DAP) maior ou igual a 10cm, totalizando 500 indivíduos
coletados por amostra. Os pontos foram dispostos em intervalos regulares de 15m ao longo de
transecções que percorriam área semelhante em fragmentos grandes e pequenos, evitando apenas
áreas de bordas e clareiras. As amostragens foram feitas de forma a incluir áreas semelhantes, tanto
do ponto de vista da fisionomia da vegetação, área percorrida e ambiente físico (condições de
altitude, solo e drenagem).
No Ribeirão Cachoeira, fragmento >200ha, foram feitas três amostragens de 125 pontos em três
regiões diferentes do fragmento (C1, C2 e C3). As áreas C1 e C2 distam aproximadamente 700m, a
distância entre as áreas C1 e C3 é de aproximadamente 800m e entre C2 e C3 em torno de 1500m
(ver Figura 1 Capítulo 1). Com isso pretendia-se verificar se áreas distintas dentro de um fragmento
grande seriam mais similares entre si do que entre os demais fragmentos.
Cada indivíduo teve sua altura estimada através da comparação com a tesoura de poda alta,
cujo cabo extensível media de 3 a 15m.
As espécies foram identificadas segundo técnicas usuais em taxonomia e os espécimes foram
incluídos em famílias segundo o sistema proposto por Cronquist (1981). A coleção principal de
material herborizado foi incorporada ao herbário UEC e as duplicatas foram enviadas a outros
herbários ou especialistas que ajudaram na identificação de alguns grupos. Os materiais coletados
em estado vegetativo foram conservados como material testemunha, em separado.
Estrutura
A estrutura da comunidade arbórea de cada fragmento foi caracterizada em termos de
abundância relativa, área basal, densidade, dominância das espécies e importância de cada espécie
em cada fragmento, expressa através do índice de valor de importância (VI, Curtis & McIntosh 1951).
Os parâmetros estruturais da vegetação foram calculados seguindo as fórmulas indicadas em Martins
(1991). Os cálculos foram efetuados com auxílio do programa FITOPAC (Shepherd 1988).
A distribuição de abundância e de área basal foi comparada usando curva de Lorenz (Weiner &
Solbrig 1984) que representa desigualdades nas abundâncias ou área basal das espécies (Martins &
Santos 1999). A curva de Lorenz obtida em cada área foi comparada às demais através do
coeficiente de Gini médio corrigido (G) estimado através do método de bootstrap, usando 3000
repetições (Weiner & Solbrig 1984). O mesmo procedimento foi utilizado para se avaliar a
concentração dos indivíduos em espécies e da área basal em espécies. Isto foi feito para verificar se
a concentração de indivíduos em espécies e da área basal em espécies diferia entre os fragmentos.
Segundo as proposições da literatura, seria esperado que em fragmentos pequenos a concentração
de abundância e de área basal fosse mais acentuada do que nos fragmentos grandes, pois nessas

113
áreas um menor número de espécies tenderiam a se tornar dominantes, uma vez que poucas
espécies sobreviveriam em áreas reduzidas (Lovejoy et al. 1986).
Para verificar se havia estratificação da comunidade tanto vertical quanto horizontalmente e
como estes aspectos variavam entre os fragmentos, foram construídos, para cada área, histogramas
de freqüência considerando classes de altura e de diâmetro.
A construção de diagramas de altura, segundo Popma et al. (1988), apresenta alguns
problemas, pois os intervalos considerados devem ser suficientemente grandes para eliminar
pequenas irregularidades e com isso facilitar a identificação de padrões, mas também devem ser
suficientemente pequenos para que os padrões possam ser reconhecidos em suficiente detalhe.
Também existe a probabilidade de que a escolha de determinado intervalo na construção dos
diagramas influencie significativamente os resultados. Em função disso, foram construídos
histogramas de classes de altura considerando intervalos de 1m e considerando intervalos de 5m,
dividindo os indivíduos em cinco classes de altura: Altura(A)≤5m, 5<A≤10m, 10<A≤15m, 15<A≤20m,
A>20m. As classes de diâmetro também foram dispostas considerando dois intervalos diferentes: de
5cm, na forma de histogramas e de 20cm. Segundo as proposições da literatura, seria esperado que
fragmentos pequenos apresentassem uma maior proporção de indivíduos nas classes de menor
diâmetro e menor altura, pois nessas áreas as taxas de mortalidade e recrutamento seriam mais
elevadas (Laurance et al. 1998a, 1998b) e portanto esperaria-se uma maior proporção de indivíduos
nas referidas classes.
Análise dos dados
Para verificar se os parâmetros analisados de cada fragmento estava relacionada ao tamanho
dos mesmos foi feita uma análise de correlação de Pearson, usando o programa BioEstat (Ayres et
al. 1998). Em tais análises consideramos apenas um valor médio das amostras de Ribeirão
Cachoeira, pois se tratava de três amostras realizadas em um mesmo fragmento.
Para os cálculos do coeficiente de Gini utilizou-se o programa Wingini (Santos 1996). O valor de
G varia de zero a um, sendo que quanto mais próximo de um, mais desigual ou concentrada é a
amostra.
Para comparar a proporção de indivíduos das diversas classes de diâmetro e altura foram
construídas tabelas de contingência do número de indivíduos em cada classe, nos diversos
fragmentos. Foram feitas comparações por meio do teste de Qui-quadrado com correção de Yates
(Zar 1989), entre os número esperados, obtidos na tabela de contingência, e observados em cada
classe nos diferentes fragmentos.
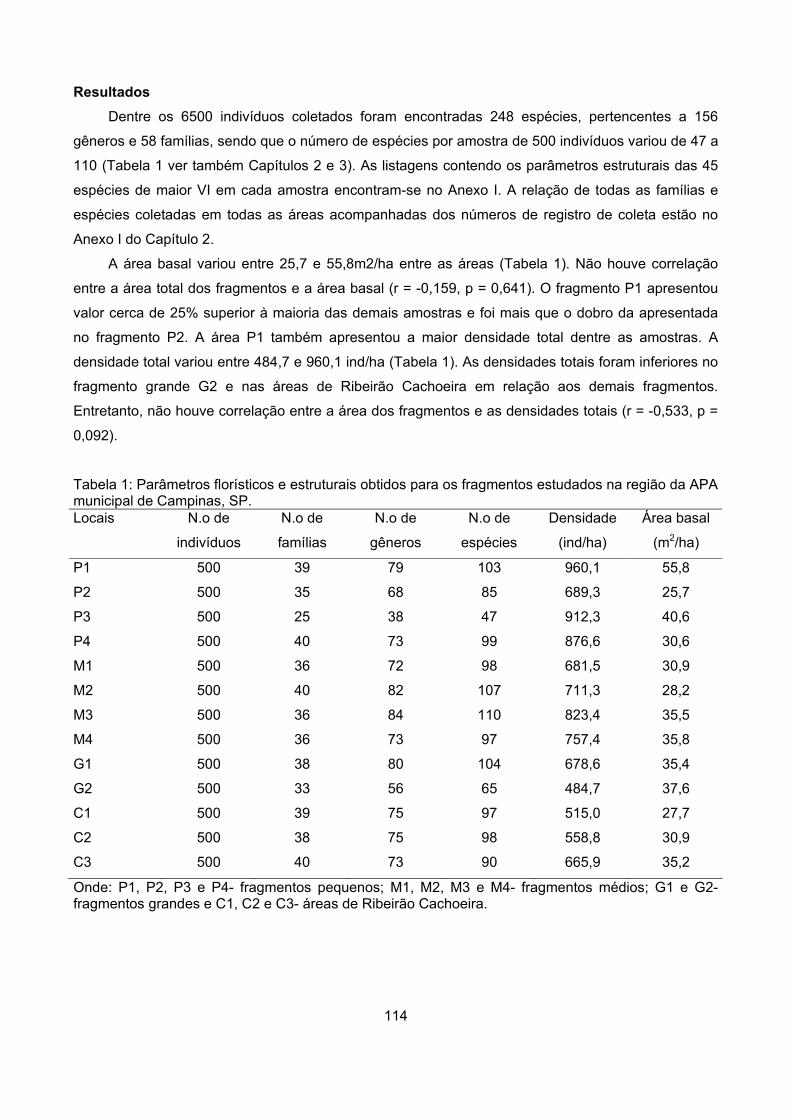
114
Resultados Dentre os 6500 indivíduos coletados foram encontradas 248 espécies, pertencentes a 156
gêneros e 58 famílias, sendo que o número de espécies por amostra de 500 indivíduos variou de 47 a
110 (Tabela 1 ver também Capítulos 2 e 3). As listagens contendo os parâmetros estruturais das 45
espécies de maior VI em cada amostra encontram-se no Anexo I. A relação de todas as famílias e
espécies coletadas em todas as áreas acompanhadas dos números de registro de coleta estão no
Anexo I do Capítulo 2.
A área basal variou entre 25,7 e 55,8m2/ha entre as áreas (Tabela 1). Não houve correlação
entre a área total dos fragmentos e a área basal (r = -0,159, p = 0,641). O fragmento P1 apresentou
valor cerca de 25% superior à maioria das demais amostras e foi mais que o dobro da apresentada
no fragmento P2. A área P1 também apresentou a maior densidade total dentre as amostras. A
densidade total variou entre 484,7 e 960,1 ind/ha (Tabela 1). As densidades totais foram inferiores no
fragmento grande G2 e nas áreas de Ribeirão Cachoeira em relação aos demais fragmentos.
Entretanto, não houve correlação entre a área dos fragmentos e as densidades totais (r = -0,533, p =
0,092).
Tabela 1: Parâmetros florísticos e estruturais obtidos para os fragmentos estudados na região da APA municipal de Campinas, SP. Locais N.o de
indivíduos
N.o de
famílias
N.o de
gêneros
N.o de
espécies
Densidade
(ind/ha)
Área basal
(m2/ha)
P1 500 39 79 103 960,1 55,8
P2 500 35 68 85 689,3 25,7
P3 500 25 38 47 912,3 40,6
P4 500 40 73 99 876,6 30,6
M1 500 36 72 98 681,5 30,9
M2 500 40 82 107 711,3 28,2
M3 500 36 84 110 823,4 35,5
M4 500 36 73 97 757,4 35,8
G1 500 38 80 104 678,6 35,4
G2 500 33 56 65 484,7 37,6
C1 500 39 75 97 515,0 27,7
C2 500 38 75 98 558,8 30,9
C3 500 40 73 90 665,9 35,2
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.

115
Padrões de abundância
As dez famílias mais abundantes, considerando todos os indivíduos coletados, representaram
71% do total de indivíduos.
A relação entre abundância e freqüência mostrou que as espécies mais abundantes tenderam a
possuir maiores valores de frequência (Figura 1). Entretanto, houve grande variação, mesmo quando
consideramos as espécies mais abundantes e/ou mais freqüentes (Figura 2). Considerando o total de
indivíduos coletados por espécie, observamos que algumas espécies que apresentaram abundâncias
iguais ou superiores à 50 indivíduos ocorreram em apenas uma ou duas áreas, enquanto outras, com
a mesma abundância, ocorreram em todas as áreas. Das seis espécies que ocorreram em todas as
amostras, três apresentaram abundâncias inferiores a 50 indivíduos. Mabea fistulifera, por outro lado,
apresentou 30 indivíduos no total, mas ocorreu em apenas uma área. Em geral, as espécies foram
muito abundantes em um fragmento e pouco abundantes ou ausentes em outros. Algumas espécies
que figuraram entre as mais abundantes no total de coletas também exibiram esse padrão (Tabela 2).
Esenbeckia leiocarpa, por exemplo, a mais abundante para o total de indivíduos coletados e também
para a maioria das áreas, nem mesmo ocorreu em algumas amostras (Tabela 2). Croton floribundos,
a segunda espécie mais abundante do estudo não ocorreu no fragmento P3 e apresentou somente 5
indivíduos no fragmento G2. Piptadenia gonoacantha, também uma das espécies mais abundantes,
ocorreu com apenas 1 indivíduo em P3 e apenas 3 e 4 indivíduos nas amostras G2 e C3,
respectivamente. Trichilia clausseni representou quase 50% dos indivíduos coletados no fragmento
P3, mas não figurou entre as espécies mais abundantes na maioria das demais áreas. Essas
variações também são perceptíveis quando analisamos outros parâmetros como densidade absoluta
e VI (Anexo I).
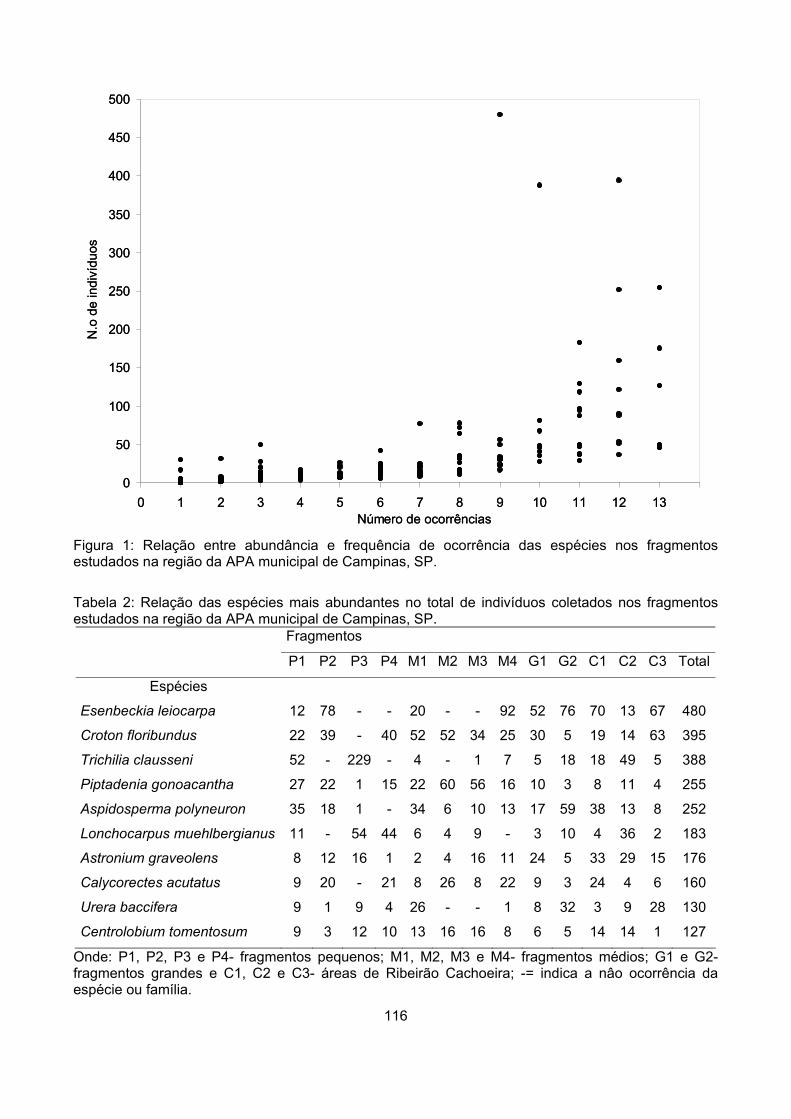
116
Figura 1: Relação entre abundância e frequência de ocorrência das espécies nos fragmentos estudados na região da APA municipal de Campinas, SP.
Tabela 2: Relação das espécies mais abundantes no total de indivíduos coletados nos fragmentos estudados na região da APA municipal de Campinas, SP. Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 Total
Espécies
Esenbeckia leiocarpa 12 78 - - 20 - - 92 52 76 70 13 67 480
Croton floribundus 22 39 - 40 52 52 34 25 30 5 19 14 63 395
Trichilia clausseni 52 - 229 - 4 - 1 7 5 18 18 49 5 388
Piptadenia gonoacantha 27 22 1 15 22 60 56 16 10 3 8 11 4 255
Aspidosperma polyneuron 35 18 1 - 34 6 10 13 17 59 38 13 8 252
Lonchocarpus muehlbergianus 11 - 54 44 6 4 9 - 3 10 4 36 2 183
Astronium graveolens 8 12 16 1 2 4 16 11 24 5 33 29 15 176
Calycorectes acutatus 9 20 - 21 8 26 8 22 9 3 24 4 6 160
Urera baccifera 9 1 9 4 26 - - 1 8 32 3 9 28 130
Centrolobium tomentosum 9 3 12 10 13 16 16 8 6 5 14 14 1 127
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira; -= indica a nâo ocorrência da espécie ou família.
0
50
100
150
200
250
300
350
400
450
500
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14Número de ocorrências
N.o
de
indi
vídu
os
0
50
100
150
200
250
300
350
400
450
500
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14Número de ocorrências
N.o
de
indi
vídu
os
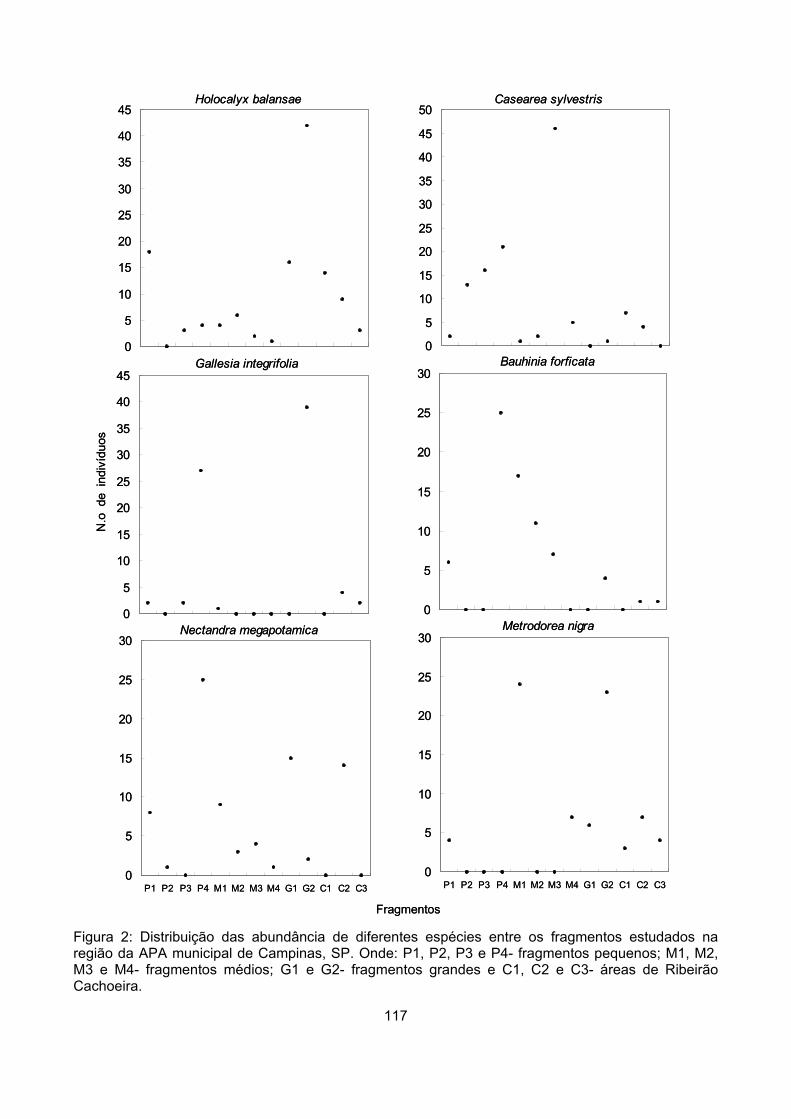
117
Figura 2: Distribuição das abundância de diferentes espécies entre os fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
Fragmentos
N.o
de
indi
vídu
os
Holocalyx balansae
0
5
10
15
20
25
30
35
40
45Casearea sylvestris
0
5
10
15
20
25
30
35
40
45
50
Gallesia integrifolia
0
5
10
15
20
25
30
35
40
45Bauhinia forficata
0
5
10
15
20
25
30
Nectandra megapotamica
0
5
10
15
20
25
30
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
Metrodorea nigra
0
5
10
15
20
25
30
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
Fragmentos
N.o
de
indi
vídu
os
Holocalyx balansae
0
5
10
15
20
25
30
35
40
45Casearea sylvestris
0
5
10
15
20
25
30
35
40
45
50
Gallesia integrifolia
0
5
10
15
20
25
30
35
40
45Bauhinia forficata
0
5
10
15
20
25
30
Nectandra megapotamica
0
5
10
15
20
25
30
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
Metrodorea nigra
0
5
10
15
20
25
30
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3

118
A proporção de espécies que ocorreu com apenas um indivíduo nos diversos fragmentos variou
de 28,2 a 42,4%. As menores proporções ocorreram nas áreas P1 e G2 e a maior no fragmento P4.
Apesar de o número de indivíduos por espécie ter variado bastante entre as áreas, houve uma
concentração de abundância em todas as amostras, isto é, poucas espécies representaram mais da
metade dos indivíduos. As dez espécies mais abundantes (c.a. 20%) em cada amostra contribuíram
com cerca de 41 a 82% dos indivíduos amostrados. Por outro lado 50% das espécies contribuíram
com menos de 20% do total de indivíduos. Essa assimetria também ocorreu quando analisamos
outros parâmetros como a área basal total. Em todas as áreas um pequeno número de espécies
compuseram mais da metade do valor total obtido. Entretanto, a concentração de abundância ou de
área basal variou entre as áreas, sendo mais ou menos acentuada em cada uma delas.
Nas amostras P3 e G2 a concentração de abundância foi mais acentuada do que nas demais
(Figura 3). Em P3 mais da metade dos indivíduos coletados pertenciam a apenas duas espécies,
enquanto em G2, seis espécies (9,2% do total coletado) totalizaram mais de 50% dos indivíduos. Nas
amostras P1 e G1 a concentração de abundância foi menos acentuada, foram necessárias 14 e 15
espécies, respectivamente (cerca de 15%) para somar mais de 50% indivíduos (Figura 3). Os valores
do coeficiente de Gini médio corrigido (G) variaram de 0,552 a 0,730 entre as amostras (Tabela 3).
Os maiores valores ocorreram nas amostras P3 e G2, indicando maior concentração. Nas áreas P1 e
G1 obtivemos os menores valores de G: 0,552 e 0,554, respectivamente (Tabela 3). Apesar dessas
variações, não houve diferença significativa entre as amostras quando comparamos os valores de G
(Tabela 3).
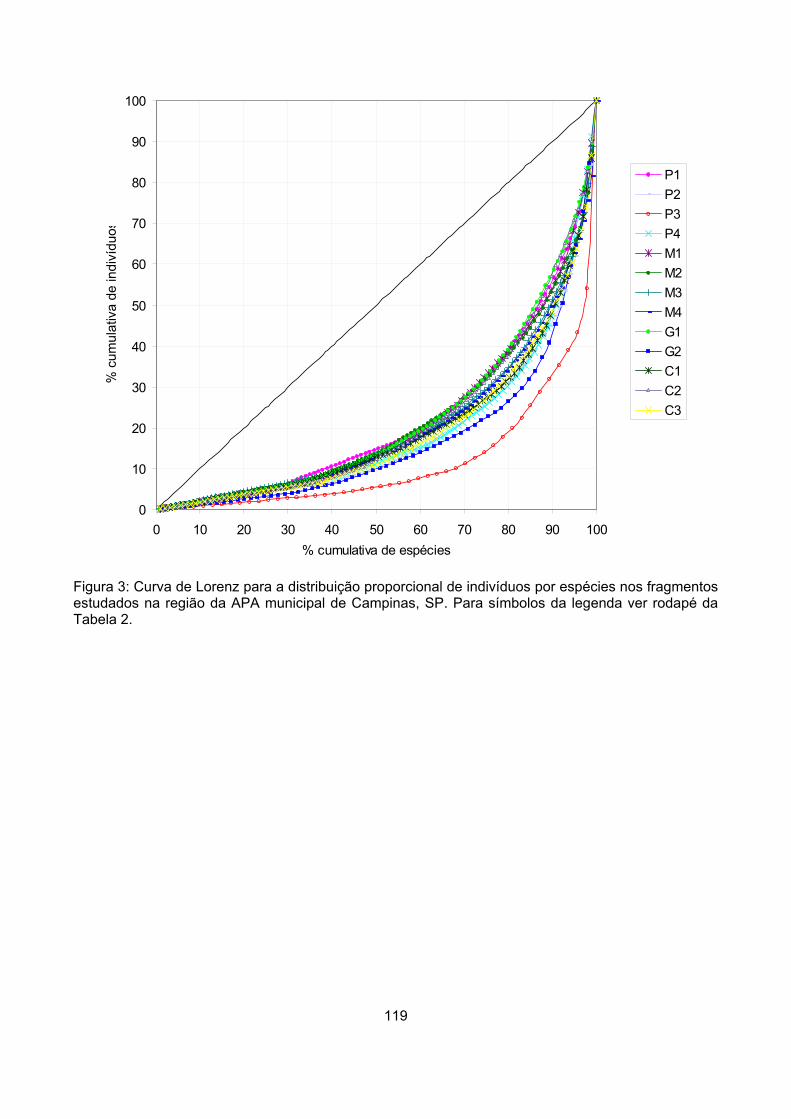
119
Figura 3: Curva de Lorenz para a distribuição proporcional de indivíduos por espécies nos fragmentos estudados na região da APA municipal de Campinas, SP. Para símbolos da legenda ver rodapé da Tabela 2.
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60 70 80 90 100% cumulativa de espécies
% c
umul
ativ
a de
indi
vídu
os
P1P2P3P4M1M2M3M4G1G2C1C2C3
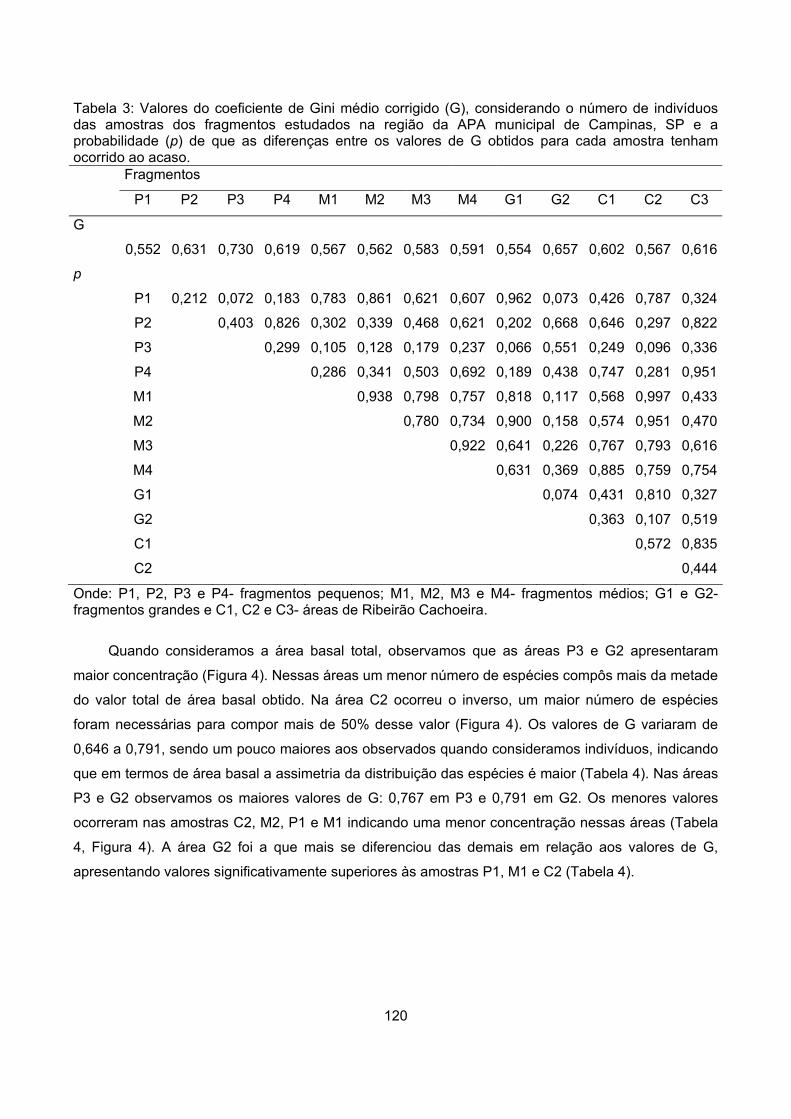
120
Tabela 3: Valores do coeficiente de Gini médio corrigido (G), considerando o número de indivíduos das amostras dos fragmentos estudados na região da APA municipal de Campinas, SP e a probabilidade (p) de que as diferenças entre os valores de G obtidos para cada amostra tenham ocorrido ao acaso. Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
G
0,552 0,631 0,730 0,619 0,567 0,562 0,583 0,591 0,554 0,657 0,602 0,567 0,616
p
P1 0,212 0,072 0,183 0,783 0,861 0,621 0,607 0,962 0,073 0,426 0,787 0,324
P2 0,403 0,826 0,302 0,339 0,468 0,621 0,202 0,668 0,646 0,297 0,822
P3 0,299 0,105 0,128 0,179 0,237 0,066 0,551 0,249 0,096 0,336
P4 0,286 0,341 0,503 0,692 0,189 0,438 0,747 0,281 0,951
M1 0,938 0,798 0,757 0,818 0,117 0,568 0,997 0,433
M2 0,780 0,734 0,900 0,158 0,574 0,951 0,470
M3 0,922 0,641 0,226 0,767 0,793 0,616
M4 0,631 0,369 0,885 0,759 0,754
G1 0,074 0,431 0,810 0,327
G2 0,363 0,107 0,519
C1 0,572 0,835
C2 0,444
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
Quando consideramos a área basal total, observamos que as áreas P3 e G2 apresentaram
maior concentração (Figura 4). Nessas áreas um menor número de espécies compôs mais da metade
do valor total de área basal obtido. Na área C2 ocorreu o inverso, um maior número de espécies
foram necessárias para compor mais de 50% desse valor (Figura 4). Os valores de G variaram de
0,646 a 0,791, sendo um pouco maiores aos observados quando consideramos indivíduos, indicando
que em termos de área basal a assimetria da distribuição das espécies é maior (Tabela 4). Nas áreas
P3 e G2 observamos os maiores valores de G: 0,767 em P3 e 0,791 em G2. Os menores valores
ocorreram nas amostras C2, M2, P1 e M1 indicando uma menor concentração nessas áreas (Tabela
4, Figura 4). A área G2 foi a que mais se diferenciou das demais em relação aos valores de G,
apresentando valores significativamente superiores às amostras P1, M1 e C2 (Tabela 4).

121
Figura 4: Curva de Lorenz para a distribuição proporcional de área basal por espécies nos fragmentos estudados na região da APA municipal de Campinas, SP. Para símbolos da legenda ver rodapé da Tabela 2.
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60 70 80 90 100% cumulativa de espécies
% c
umul
ativ
a de
áre
a ba
sa
P1P2P3P4M1M2M3M4G1G2C1C2C3

122
Tabela 4: Valores do coeficiente de Gini médio corrigido (G), considerando a área basal nas amostras dos fragmentos estudados na região da APA municipal de Campinas, SP e a probabilidade (p) de que as diferenças entre os valores de G obtidos para cada amostra tenham ocorrido ao acaso. Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3
G
0,679 0,702 0,765 0,702 0,698 0,669 0,704 0,704 0,710 0,791 0,714 0,646 0,714
p
P1 0,741 0,202 0,649 0,707 0,901 0,688 0,625 0,528 0,042* 0,499 0,480 0,514
P2 0,398 0,995 0,954 0,697 0,972 0,974 0,906 0,162 0,856 0,358 0,857
P3 0,287 0,278 0,263 0,383 0,316 0,360 0,675 0,406 0,069 0,440
P4 0,931 0,674 0,971 0,948 0,845 0,051 0,762 0,149 0,791
M1 0,708 0,906 0,900 0,790 0,046* 0,731 0,204 0,755
M2 0,664 0,653 0,602 0,091 0,572 0,775 0,567
M3 0,999 0,918 0,127 0,881 0,279 0,870
M4 0,886 0,069 0,837 0,173 0,847
G1 0,084 0,924 0,121 0,927
G2 0,100 0,002* 0,129
C1 0,102 0,996
C2 0,145
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira; * = diferença significativa.
Valor de importância
Algumas famílias de maior VI em algumas amostras nem mesmo ocorreram em outras,
refletindo as variações nos padrões de abundância observados entre as amostras. Algumas famílias
e espécies estiveram entre as de maior VI nas poucas áreas em que ocorreram, enquanto outras, que
ocorreram com maior constância entre os fragmentos, raramente atingiram altos VIs.
Vinte e uma famílias figuraram entre as 10 primeiras em VI nas treze amostras. Nenhuma
família figurou entre as de maior VI em todas as amostras, entretanto houve pouca variação entre os
fragmentos. Em geral, as mesmas famílias figuraram entre as dez primeiras em VI nas amostras,
variando apenas de posição nesse ranking de uma área para outra. Entretanto, as áreas C3 e,
principalmente, P3 exibiram poucas famílias em comum nesse ranking com as outras áreas (Tabela
5). As famílias que estiveram entre as de maior VI na maioria das amostras foram Apocynaceae,
Caesalpiniaceae, Euphorbiaceae, Fabaceae, Meliaceae, Mimosaceae e Rutaceae. A família
Rutaceae foi a maior em VI em cinco amostras P2, M4, G2, G3 e C1 e, Meliaceae foi a primeira em VI
nas amostras P1, P3 e C2.

123
Apesar de o conjunto das famílias de maior VI nas diferentes áreas ser semelhante,
observamos que em algumas áreas os valores de VI são muito similares entre essas famílias,
enquanto em outras as primeiras colocadas apresentam valores muito superiores em relação às
demais. Nas amostras P1, M1 e nas áreas C1 e C2 de Ribeirão Cachoeira os valores de VI
decresceram de forma contínua da primeira para a décima colocada, indicando que nenhuma dessas
famílias é desproporcionalmente dominante em relação às demais. Nas demais áreas ocorreu uma
descontinuidade muito grande entre os valores apresentados ao longo do ranking, onde a primeira
em VI apresentou valores equivalentes ao dobro ou triplo do da segunda colocada, como nas
amostras P4, M1, M3 e G1. Em outras áreas essa concentração de dominância foi ainda maior com
uma ou duas famílias chegando a compor mais de um terço do valor total do VI. A amostra P3, foi o
caso mais extremo, onde uma única família (Meliaceae) representa mais de um terço do valor total de
VI. Essas observações indicam diferenças estruturais entre os fragmentos.

124
Tabela 5: Comparação entre as famílias de maior valor de importância (VI) nos fragmentos estudados na região da APA municipal de Campinas, SP. P1 P2 P3 P4 Famílias VI Famílias VI Famílias VI Famílias VI Meliaceae 33,8 Rutaceae 76,1 Meliaceae 115,2 Fabaceae 67,5 Apocynaceae 27,2 Euphorbiaceae 53,7 Fabaceae 52,0 Lauraceae 26,5 Euphorbiaceae 25,9 Mimosaceae 23,1 Bombacaceae 45,1 Myrtaceae 22,8 Mimosaceae 25,8 Lauraceae 16,3 Moraceae 12,9 Euphorbiaceae 22,8 Fabaceae 24,2 Myrtaceae 15,9 Flacourtiaceae 11,6 Caesalpiniaceae 19,8 Moraceae 18,6 Flacourtiaceae 12,0 Anacardiaceae 10,7 Phytollacaceae 18,3 Rutaceae 17,5 Fabaceae 11,3 Caesalpiniaceae 8,2 Mimosaceae 17,1 Phytollacaceae 14,8 Apocynaceae 10,5 Urticaceae 5,5 Flacourtiaceae 12,8 Lauraceae 14,7 Anacardiaceae 10,4 Ebenaceae 5,5 Meliaceae 9,0 Caesalpiniaceae 14,3 Caesalpiniaceae 9,4 Lecythidaceae 5,5 Apocynaceae 6,8
Total 216,7 Total 238,5 Total 272,2 Total 223,4 Restante 83,3 Restante 61,5 Restante 27,8 Restante 76,6
M1 M2 M3 M4 Famílias VI Famílias VI Famílias VI Famílias VI Euphorbiaceae 35,0 Mimosaceae 58,2 Mimosaceae 53,2 Rutaceae 51,9 Rutaceae 25,7 Euphorbiaceae 35,1 Fabaceae 30,4 Euphorbiaceae 23,5 Apocynaceae 25,1 Fabaceae 28,8 Euphorbiaceae 25,2 Meliaceae 22,9 Mimosaceae 23,8 Myrtaceae 28,7 Flacourtiaceae 25,1 Caesalpiniaceae 22,9 Fabaceae 19,0 Caesalpiniaceae 22,3 Lauraceae 22,6 Fabaceae 22,0 Caesalpiniaceae 18,7 Lauraceae 15,0 Meliaceae 20,8 Myrtaceae 20,0 Myrtaceae 14,6 Rubiaceae 9,4 Caesalpiniaceae 16,8 Mimosaceae 18,1 Moraceae 14,1 Meliaceae 9,0 Anacardiaceae 12,4 Bombacaceae 15,9 Urticaceae 13,3 Apocynaceae 8,5 Myrtaceae 11,3 Apocynaceae 15,4 Lecythidaceae 12,8 Annonaceae 7,9 Apocynaceae 11,0 Lecythidaceae 9,6
Total 202,1 Total 222,9 Total 228,7 Total 222,1 Restante 97,9 Restante 77,1 Restante 71,3 Restante 77,9
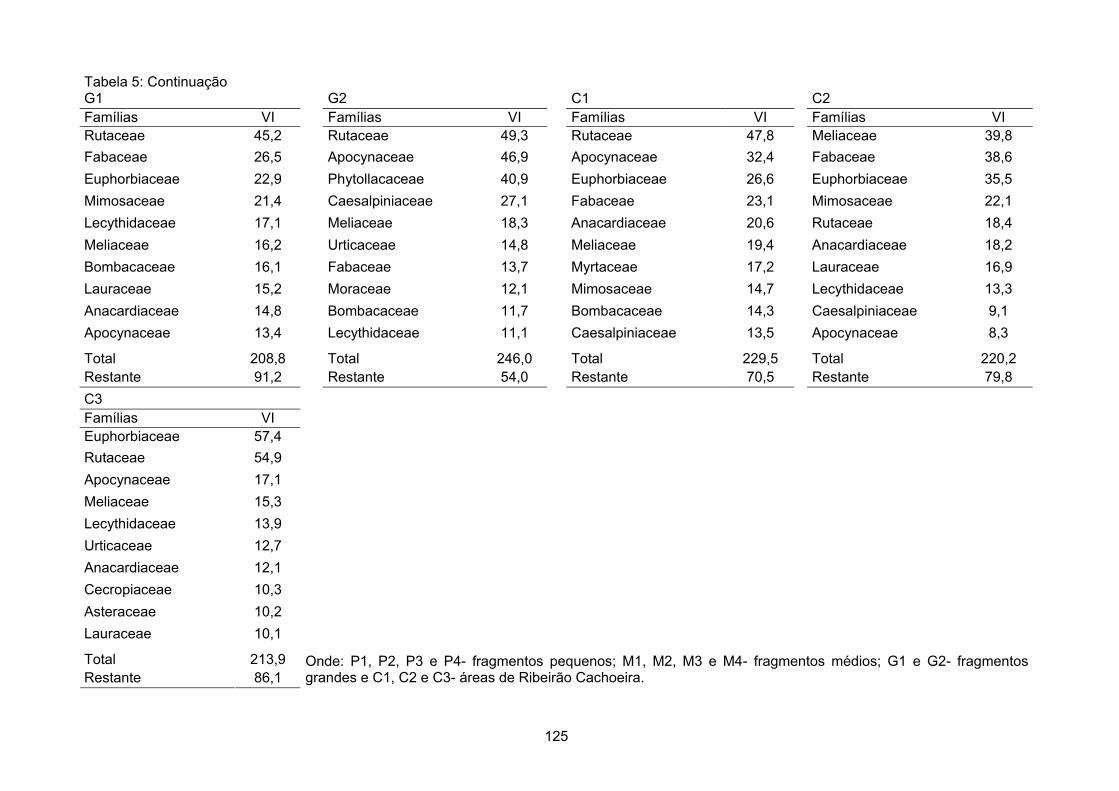
125
Tabela 5: Continuação G1 G2 C1 C2 Famílias VI Famílias VI Famílias VI Famílias VI Rutaceae 45,2 Rutaceae 49,3 Rutaceae 47,8 Meliaceae 39,8 Fabaceae 26,5 Apocynaceae 46,9 Apocynaceae 32,4 Fabaceae 38,6 Euphorbiaceae 22,9 Phytollacaceae 40,9 Euphorbiaceae 26,6 Euphorbiaceae 35,5 Mimosaceae 21,4 Caesalpiniaceae 27,1 Fabaceae 23,1 Mimosaceae 22,1 Lecythidaceae 17,1 Meliaceae 18,3 Anacardiaceae 20,6 Rutaceae 18,4 Meliaceae 16,2 Urticaceae 14,8 Meliaceae 19,4 Anacardiaceae 18,2 Bombacaceae 16,1 Fabaceae 13,7 Myrtaceae 17,2 Lauraceae 16,9 Lauraceae 15,2 Moraceae 12,1 Mimosaceae 14,7 Lecythidaceae 13,3 Anacardiaceae 14,8 Bombacaceae 11,7 Bombacaceae 14,3 Caesalpiniaceae 9,1 Apocynaceae 13,4 Lecythidaceae 11,1 Caesalpiniaceae 13,5 Apocynaceae 8,3
Total 208,8 Total 246,0 Total 229,5 Total 220,2 Restante 91,2 Restante 54,0 Restante 70,5 Restante 79,8 C3 Famílias VI Euphorbiaceae 57,4 Rutaceae 54,9 Apocynaceae 17,1 Meliaceae 15,3 Lecythidaceae 13,9 Urticaceae 12,7 Anacardiaceae 12,1 Cecropiaceae 10,3 Asteraceae 10,2 Lauraceae 10,1
Total 213,9 Restante 86,1
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.

126
Algumas espécies ocuparam os primeiros lugares no ranking de VI em algumas áreas e nem
mesmo ocorreram em outras. Foi o caso, por exemplo, de Trichilia casaretti, Vernonia diffusa e
Metrodorea stipularis. No fragmento M4, por exemplo, Trichilia casaretti foi a quinta espécie em VI,
mas ocorreu somente neste fragmento e na área C3 de Ribeirão Cachoeira com apenas 2 indivíduos,
onde ocupou a 58º posição no ranking de VI. Vernonia diffusa ocorreu em apenas 3 áreas e foi a
sétima em VI na área C3 de Ribeirão Cachoeira, mas figurou com apenas 2 e 3 indivíduos nas
demais áreas onde ocorreu. Algo semelhante ocorreu com Metrodorea stipularis, a terceira em VI em
P2, mas que só ocorreu em outras duas amostras, com apenas seis e um indivíduo, respectivamente.
O número de espécies necessárias para compor mais da metade do valor total de VI (300) em
cada área variou de 3 a 15 espécies, representando 3,4 a 15% das espécies coletadas,
respectivamente (Tabela 6). Isto quer dizer que em algumas áreas, poucas espécies são dominantes,
enquanto em outras a concentração dessa dominância é menor. Nas áreas P2, P3 e G2, essa
concentração foi mais acentuada do que nas demais áreas, isto é, um menor número de espécies foi
necessário para ultrapassar a metade do valor total de VI. Reforçando essa hipótese observamos
que, assim como no ranking de VI para famílias, em algumas áreas os valores de VI são muito
semelhantes entre as espécies que compõem esse ranking, decrescendo de forma contínua,
enquanto em outras, as primeiras colocadas apresentaram valores equivalentes ao dobro ou triplo da
segunda colocada. Algumas espécies estão entre as dez de maior VI em várias amostras, mas
compõem proporções bem diferentes do valor total de VI em cada área. Trichilia clausseni, por
exemplo, figura entre as de maior VI em cinco amostras, mas em P3 representou um terço do valor
total de VI, indicando grande dominância. Esenbeckia leiocarpa ocupou as primeiras posições do
ranking de VI em sete amostras e, em algumas áreas em que foi primeira colocada exibiu valor
equivalente ao dobro do da segunda colocada como em P2, M4, G1 e C3, indicando maior
dominância nessas áreas. Em outras amostras os valores de VI apresentados por essa espécie são
semelhantes aos exibidos pelas demais como em M1, G2 e C1. Tais diferenças denotam diferenças
na estrutura dos fragmentos, embora não tenha sido possível associá-las à área total dos mesmos.
Somando-se as dez espécies de maior VI em cada área, temos um total de 46 espécies (Tabela
6). Nenhuma espécie figurou entre as dez mais importantes em todas as amostras. Apenas 4
espécies ocuparam as primeiras colocações desse ranking em pelo menos 50% das amostras:
Aspidosperma polyneuron, Astronium graveolens, Croton floribundus, Esenbeckia leiocarpa e
Piptadenia gonoacantha. Por outro lado, quase 50%, ou 22 espécies atingiram altos VIs em apenas
uma amostra.

127
Tabela 6: Comparação entre as espécies de maior valor de importância (VI) nos fragmentos estudados na região da APA municipal de Campinas, SP e de outros três levantamentos fitossociológicos realizados no interior do estados de São paulo que adotaram DAP ≥ 10cm como critério de inclusão. P1 P2 P3 P4 Espécies VI Espécies VI Espécies VI Espécies VI Piptadenia gonoacantha 22,4 Esenbeckia leiocarpa 48,3 Trichilia clausseni 99,9 Lonchocarpus muehlbergianus 25,3 Trichilia clausseni 22,1 Croton floribundus 25,3 Chorisia speciosa 39,5 Machaerium nictitans 20,8 Aspidosperma polyneuron 19,4 Metrodoria stipularis 25,0 Lonchocarpus muehlbergianus 35,1 Croton floribundus 20,6 Croton floribundus 13,1 Piptadenia gonoacantha 16,7 Maclura tinctoria 11,6 Gallesia integrifolia 17,1 Ficus glabra 11,6 Mabea fistulifera 16,6 Astronium graveolens 10,2 Nectandra megapotamica 16,0 Holocalyx balansae 10,5 Aspidosperma polyneuron 10,2 Trichilia hirta 9,6 Bauhinia forficata 13,3 Gallesia integrifolia 9,8 Calycorectes acutatus 9,1 Trichilia catigua 8,7 Calycorectes acutatus 10,8 Myroxylon peruiferum 7,2 Astronium graveolens 7,2 Casearia sylvestris 8,2 Casearia sylvestris 10,6 Lonchocarpus muehlbergianus 6,6 Cryptocarya aschersoniana 6,9 Centrolobium tomentosum 7,6 Piptadenia gonoacantha 9,3 Aspidosperma ramiflorum 6,5 Casearia sylvestris 6,3 Myroxylon peruiferum 6,5 Lonchocarpus cultratus 8,9
Total 129,2 Total 171,5 Total 236,8 Total 152,5 Restante 170,8 Restante 128,6 Restante 63,2 Restante 147,5
M1 M2 M3 M4 Espécies VI Espécies VI Espécies VI Espécies VI Croton floribundus 30,6 Piptadenia gonoacantha 45,0 Piptadenia gonoacantha 40,1 Esenbeckia leiocarpa 46,7 Aspidosperma polyneuron 18,8 Croton floribundus 26,2 Casearia sylvestris 21,9 Aspidosperma polyneuron 14,2 Piptadenia gonoacantha 18,4 Calycorectes acutatus 13,0 Croton floribundus 19,9 Chorisia speciosa 14,0 Urera baccifera 13,2 Copaifera langsdorffi 9,6 Cabralea canjerana 12,8 Croton floribundus 13,6 Esenbeckia leiocarpa 10,4 Centrolobium tomentosum 8,2 Astronium graveolens 10,1 Trichilia casarettii 12,5 Metrodoria nigra 10,2 Sebastiania edwaliana 7,6 Centrolobium tomentosum 8,5 Piptadenia gonoacantha 12,0 Cariniana legalis 9,9 Myroxylon peruiferum 7,3 Acacia paniculata 7,8 Calycorectes acutatus 11,0 Bauhinia forficata 9,2 Machaerium nictitans 7,2 Copaifera langsdorffi 7,2 Hymenaea courbaril 10,0 Jacaranda micrantha 8,5 Syagrus romanzoffiana 6,3 Machaerium nictitans 6,6 Copaifera langsdorffi 8,8 Seguieria langsdorffi 7,5 Bauhinia forficata 5,8 Matayba elaeagnoides 6,2 Savia dictyocarpa 8,5
Total 136,5 Total 136,2 Total 141,0 Total 151,2 Restante 163,5 Restante 163,8 Restante 159,0 Restante 148,8

128
Tabela 6: Continuação. G1 G2 C1 C2 Espécies VI Espécies VI Espécies VI Espécies VI Esenbeckia leiocarpa 30,4 Gallesia integrifolia 39,2 Esenbeckia leiocarpa 36,2 Trichilia clausseni 22,6 Croton floribundus 14,9 Esenbeckia leiocarpa 37,3 Aspidosperma polyneuron 23,8 Lonchocarpus muehlbergianus 19,7 Astronium graveolens 14,5 Aspidosperma polyneuron 35,1 Astronium graveolens 20,2 Astronium graveolens 17,4 Chorisia speciosa 14,4 Holocalyx balansae 21,2 Savia dictyocarpa 13,8 Croton priscus 12,6 Cariniana legalis 14,1 Urera baccifera 14,4 Calycorectes acutatus 12,4 Piptadenia gonoacantha 10,4 Aspidosperma polyneuron 10,4 Aspidosperma ramiflorum 13,0 Chorisia speciosa 10,4 Acacia polyphylla 9,3 Piptadenia gonoacantha 8,5 Chorisia speciosa 11,5 Trichilia clausseni 9,5 Cariniana estrellensis 8,0 Galipia multiflora 7,7 Metrodoria nigra 10,1 Centrolobium tomentosum 9,5 Centrolobium tomentosum 8,0 Holocalyx balansae 7,5 Ficus glabra 9,9 Croton floribundus 9,0 Croton floribundus 7,9 Nectandra megapotamica 7,4 Trichilia clausseni 8,3 Holocalyx balansae 8,5 Aspidosperma polyneuron 7,1
Total 129,7 Total 200,0 Total 153,3 Total 122,9 Restante 170,3 Restante 100,0 Restante 146,7 Restante 177,1 C3 Matthes 1980* Cavassan et al. 1984 Bertoni et al. 1988 Espécies VI Espécies VI Espécies VI Espécies VI Esenbeckia leiocarpa 41,8 Trichilia clausseni 45,4 Acacia polyphylla 31,1 Metrodorea nigra 22,5 Croton floribundus 29,7 Machaerium nictitans 8,6 Chorisia speciosa 21,8 Esenbeckia febrifuga 16 Savia dictyocarpa 15,2 Centrolobium 7,4 Aspidosperma polyneuron 20,8 Gallesia integrifolia 12,6 Cariniana legalis 13,7 Cabralea canjerana 6,9 Machaerium stipitatum 20,0 Urera baccifera 12,6 Urera baccifera 12,3 Aspidosperma polyneuron 5,5 Prapiptadenia rigida 16,7 Trichilia catigua 11,6 Astronium graveolens 11,8 Cariniana estrellensis 4,6 Mortas 16,6 Centrolobium tomentosum 10,8 Vernonia diffusa 9,9 Seguieria langsdorffii 3,9 Centrolobium tomentosum 15,5 Acacia polyphylla 10,7 Cecropia hololeuca 9,1 Mortas 3,7 Metrodorea nigra 11,3 Croton priscus 10,6 Chorisia speciosa 8,1 Myroxylon peruiferum 3,6 Gallesia integrifolia 10,8 Jaracatia dodecaphylla 10,4 Aspidosperma ramiflorum 8,0 Piptadenia gonoacantha 3,4 Holocalyx balansae 10,5 Machaerium stipitatum 10,3 Total 159,7 Total 93 Total 175,1 Total 128,1 Restante 140,4 Restante 107 Restante 124,9 Restante 171,9 Para símbolos de fragmentos ver rodapé da tabela 5. * o autor usou valor máximo de VI 200, nos demais levantamentos o valor máximo de VI é 300

129
Classes de diâmetro
Na distribuição das classes de diâmetro observamos, em todas as áreas, o padrão de
distribuição em forma de J invertido, isto é, houve uma concentração de indivíduos nas classes de
menores diâmetros (Figura 5). Apesar do mesmo padrão de distribuição, houve grande variação entre
os fragmentos quanto ao número de indivíduos compreendidos em cada classe (Tabela 7). As
maiores variações ocorreram nas classes de maior diâmetro e os fragmentos que mais se
diferenciaram em relação aos demais foram P3 e G2. No primeiro ocorreu uma maior concentração
de indivíduos nas classes de menor diâmetro e no segundo ocorreu o inverso.
Nas classes de menor diâmetro (10<DAP≤20cm) ocorreu a maior parte do total de indivíduos
coletados: 60,4%. Entre as áreas essa proporção variou de 54,2 a 68%, mas não diferiu
significativamente entre elas (χ2 = 13,7, gl = 12, p = 0,323). As menores proporções ocorreram nas
amostras C1 e C2 e as maiores nos fragmentos P3 e P4. A área P3 foi a única que apresentou
proporção significativamente superior de indivíduos nas classes de menor diâmetro em relação às
demais (χ2 = 4,6, gl = 1, p = 0,032) (Tabela 7).
Nas classes de diâmetro intermediárias (20<DAP≤40cm), ocorreram 32,1% do total de
indivíduos. Não houve diferença significativa entre as áreas quanto a proporção de indivíduos
compreendidos nessa classe (χ2 = 13,6, gl = 12, p = 0,325), embora, no fragmento P3 essa proporção
tenha sido inferior aos demais (χ2 = 5,3, gl = 1, p = 0,022) (Tabela 7).
Dentre as classes de maior diâmetro (DAP>40cm) as variações foram ainda maiores. Os
indivíduos com diâmetros entre 40 e 60cm representaram 5,5% do total e sua proporção diferiu entre
as áreas (χ2 = 44,4, gl = 12, p < 0,001). O fragmento G2 e as áreas C1 e C2 de Ribeirão Cachoeira
apresentaram maior proporção de indivíduos com diâmetros entre 40 e 60cm em relação às demais
amostras (G2- χ2 = 14,3, gl = 1, p < 0,001; C1- χ2 = 5,1, gl = 1, p = 0,024; C2- χ2 = 4,3, gl = 1, p =
0,038). Nas áreas P3 e M1, por outro lado, a proporção desses indivíduos foi significativamente
inferior às demais (P3- χ2 = 4,5, gl = 1, p = 0,035; M1- χ2 = 5,3, gl = 1, p = 0,021) (Tabela 7).
Os indivíduos que atingiram diâmetros superiores a 60cm representaram apenas 1,9% do total
coletado. Essa porcentagem variou de 0,2 a 4,6% entre as áreas, que diferiram significativamente
entre si (χ2 = 47,7, gl = 12, p < 0,001). Alguns fragmentos pequenos e médios como P2, P4, M2 e M3
apresentaram apenas 0,2 a 0,6% de indivíduos com diâmetros maiores que 60cm. Essas proporções
foram inferiores às demais áreas (P2- χ2 = 8,6, gl = 1, p = 0,003; P4- χ2 = 6,8, gl = 1, p = 0,009; M2- χ2
= 5,3, gl = 1, p = 0,022; M3- χ2 = 5,3, gl = 1, p = 0,022). Nas áreas de Ribeirão Cachoeira e nos
fragmentos G1, M4, M1, P1 e P3 essa proporção ficou em torno de 2%. O fragmento G2 foi o que
apresentou a maior proporção de indivíduos nas classes de maiores diâmetros, que representaram
4,6% do total coletado, diferindo dos demais (χ2 = 17,3, gl = 1, p < 0,001) (Tabela 7).
As variações observadas apesar de refletirem diferenças estruturais entre os fragmentos não
pareceram estar associadas à área total dos mesmos. Não houve correlação entre o número de
indivíduos compreendidos em cada classe de diâmetro nos diferentes fragmentos e sua área total:
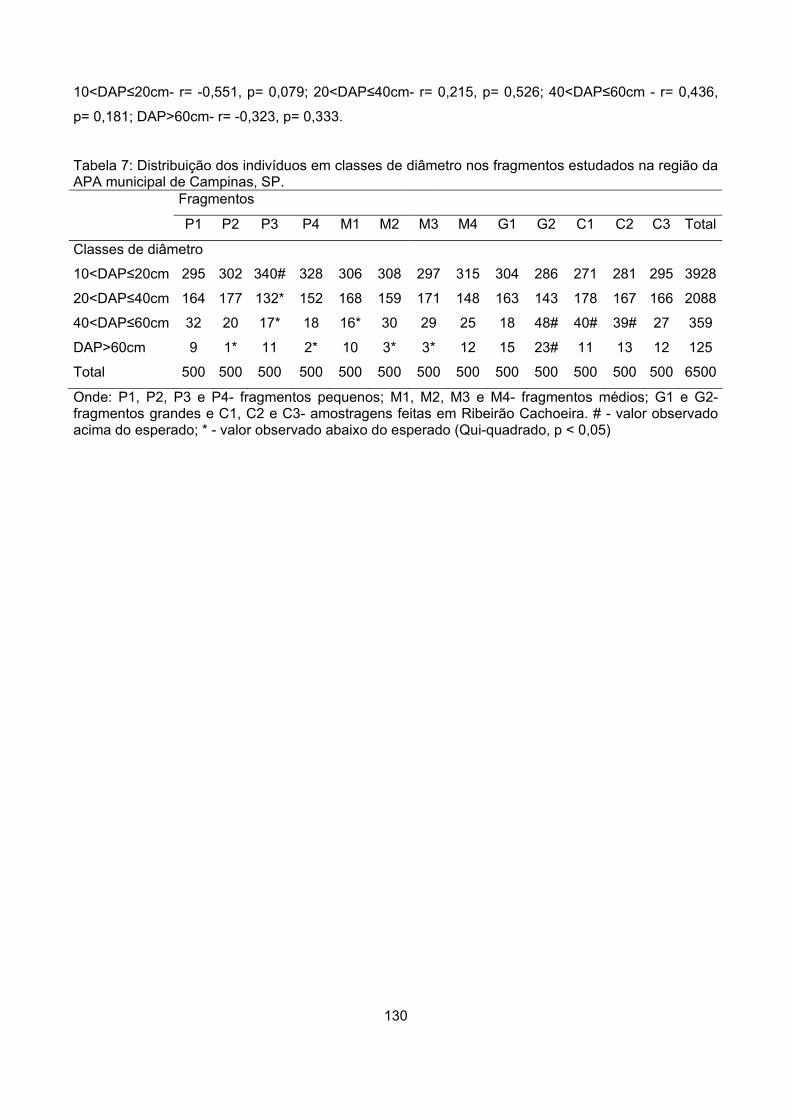
130
10<DAP≤20cm- r= -0,551, p= 0,079; 20<DAP≤40cm- r= 0,215, p= 0,526; 40<DAP≤60cm - r= 0,436,
p= 0,181; DAP>60cm- r= -0,323, p= 0,333.
Tabela 7: Distribuição dos indivíduos em classes de diâmetro nos fragmentos estudados na região da APA municipal de Campinas, SP. Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 Total
Classes de diâmetro
10<DAP≤20cm 295 302 340# 328 306 308 297 315 304 286 271 281 295 3928
20<DAP≤40cm 164 177 132* 152 168 159 171 148 163 143 178 167 166 2088
40<DAP≤60cm 32 20 17* 18 16* 30 29 25 18 48# 40# 39# 27 359
DAP>60cm 9 1* 11 2* 10 3* 3* 12 15 23# 11 13 12 125
Total 500 500 500 500 500 500 500 500 500 500 500 500 500 6500
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- amostragens feitas em Ribeirão Cachoeira. # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05)
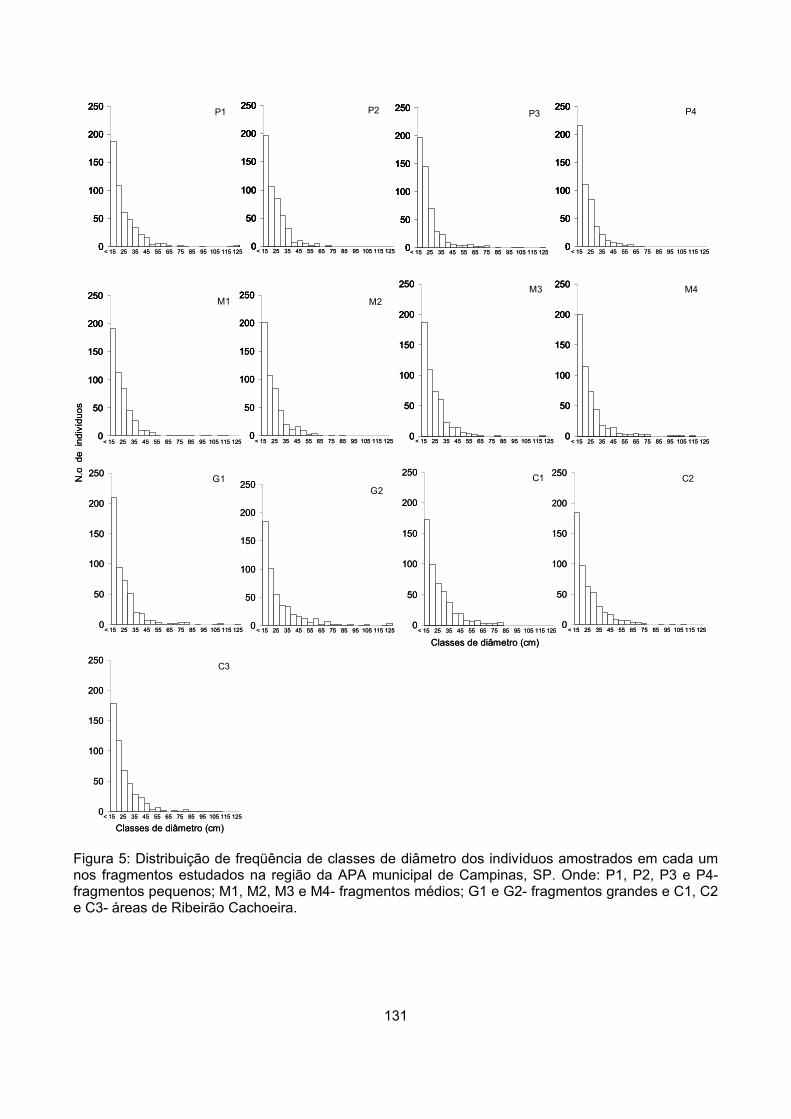
131
Figura 5: Distribuição de freqüência de classes de diâmetro dos indivíduos amostrados em cada um nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
P1
0
50
100
150
200
250 P2
0
50
100
150
200
250P3
0
50
100
150
200
250 P4
0
50
100
150
200
250
M1
0
50
100
150
200
250 M2
0
50
100
150
200
250 M3
0
50
100
150
200
250 M4
0
50
100
150
200
250
C3
0
50
100
150
200
250
G1
0
50
100
150
200
250
G2
0
50
100
150
200
250C1
0
50
100
150
200
250 C2
0
50
100
150
200
250
< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
Classes de diâmetro (cm)
Classes de diâmetro (cm)
N.o
de
indi
vídu
os
P1
0
50
100
150
200
250 P2
0
50
100
150
200
250P3
0
50
100
150
200
250 P4
0
50
100
150
200
250
M1
0
50
100
150
200
250 M2
0
50
100
150
200
250 M3
0
50
100
150
200
250 M4
0
50
100
150
200
250
P1
0
50
100
150
200
250 P2
0
50
100
150
200
250P3
0
50
100
150
200
250 P4
0
50
100
150
200
250P1
0
50
100
150
200
250 P2
0
50
100
150
200
250P3
0
50
100
150
200
250 P4
0
50
100
150
200
250
M1
0
50
100
150
200
250 M2
0
50
100
150
200
250 M3
0
50
100
150
200
250 M4
0
50
100
150
200
250
C3
0
50
100
150
200
250
G1
0
50
100
150
200
250
G2
0
50
100
150
200
250C1
0
50
100
150
200
250 C2
0
50
100
150
200
250
< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125< 15 25 35 45 55 65 75 85 95 105 115 125 < 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
< 15 25 35 45 55 65 75 85 95 105 115 125
Classes de diâmetro (cm)
Classes de diâmetro (cm)
N.o
de
indi
vídu
os

132
Distribuição vertical de alturas
A mediana das alturas das árvores nos fragmentos variou entre 10 e 13m com intervalos de
confiança de 0,25 e 0,42m, respectivamente (Tabela 8). Comparando esses valores através dos
intervalos de confiança, observamos que as amostras P3, P4, M1 e G1 apresentaram os menores
valores, as amostras M3, G2 e C1 apresentaram os maiores valores e as demais valores
intermediários (Figura 6). Não houve correlação entre os valores de mediana das alturas e a área
total dos fragmentos (r= 0,264, p= 0,434).
Nos fragmentos P1, G2 e área C1 de Ribeirão Cachoeira a variabilidade entre as alturas dos
indivíduos foi aparentemente maior, enquanto nas áreas P2, P4 e G1 essa variabilidade foi menor
(Figura 6). Observando a Figura 7, percebemos que nas áreas P4, G1 e, principalmente P2 a maior
parte dos indivíduos esteve concentrada em algumas poucas classes de altura e apresentaram
poucos indivíduos nas classes de maior altura (Tabela 8). Nas áreas P1, M3, M4, G2 e área C1 de
Ribeirão Cachoeira os indivíduos distribuíram-se mais uniformemente pelas diversas classes (Figura
7).
A distribuição dos indivíduos em classes de altura foi semelhante em todos os fragmentos, com
a maioria dos indivíduos (70,6 a 93% dos indivíduos) concentrando-se entre 5 e 15m de altura
(Tabela 8). Entretanto, não foi possível determinar quais ou quantos estratos existem em cada área
(Figura 7).
Houve grande variação entre as áreas quanto a proporção de indivíduos em cada classe de
altura (Tabela 8). Na primeira classe (Altura (A) ≤ 5m), duas amostras se diferenciaram das demais:
G2 que apresentou menor proporção de indivíduos (χ2 = 7,2, gl = 1, p = 0,007) e C3 com proporção
superior às outras amostras (χ2 = 3,9, gl = 1, p < 0,05). Na classe de altura entre 5 e 10m os
fragmentos P3, M1 e G1 apresentaram maior proporção de indivíduos do que os demais (P3- χ2 =
12,6, gl = 1, p < 0,001; M1- χ2 = 6,7, gl = 1, p < 0,05; G1- χ2 = 11,1, gl = 1, p = 0,001), enquanto nos
fragmentos M3, G2 e área C1 de Ribeirão Cachoeira ocorreu o inverso (M3- χ2 = 4,1, gl = 1, p < 0,05;
G2- χ2 = 16,1, gl = 1, p < 0,001; C1- χ2 = 17,1, gl = 1, p < 0,001). Os fragmentos P1, P3 e G1 também
se diferenciaram dos demais por apresentarem uma menor proporção de indivíduos com alturas entre
10 e 15m (P1- χ2 = 9,7, gl = 1, p = 0,002; P3- χ2 = 17,7, gl = 1, p < 0,001; G1- χ2 = 4,4, gl = 1, p <
0,05), no fragmento P2, por outro lado, os indivíduos nessa classe de altura ocorreram em maior
proporção do que nas demais áreas (χ2 = 16,4, gl = 1, p < 0,001). Os indivíduos com alturas entre 15
e 20m ocorreram em maior proporção em M3, M4, G2 e C1 (M3- χ2 = 11,8, gl = 1, p = 0,001; M4- χ2 =
10,2, gl = 1, p = 0,001; G2- χ2 = 32,3, gl = 1, p < 0,001; C1- χ2 = 12,7, gl = 1, p < 0,001), enquanto nas
amostras P2, P4, M1, G1 e C1 ocorreu o inverso (P2- χ2 = 30,9, gl = 1, p < 0,001; P4- χ2 = 4,8, gl = 1,
p < 0,05; M1- χ2 = 5,3, gl = 1, p < 0,05; G1- χ2 = 9,4, gl = 1, p = 0,002; C3- χ2 = 8,6, gl = 1, p = 0,003).
Os indivíduos com alturas superiores a 20m ocorreram em maior proporção nas amostras P1, G2 e
C1 (P1- χ2 = 4,3, gl = 1, p < 0,05; G2- χ2 = 7,6, gl = 1, p = 0,006; C1- χ2 = 10,2, gl = 1, p = 0,001) e em
menor proporção no fragmento P2 (χ2 = 15,5, gl = 1, p < 0,001) (Tabela 8).
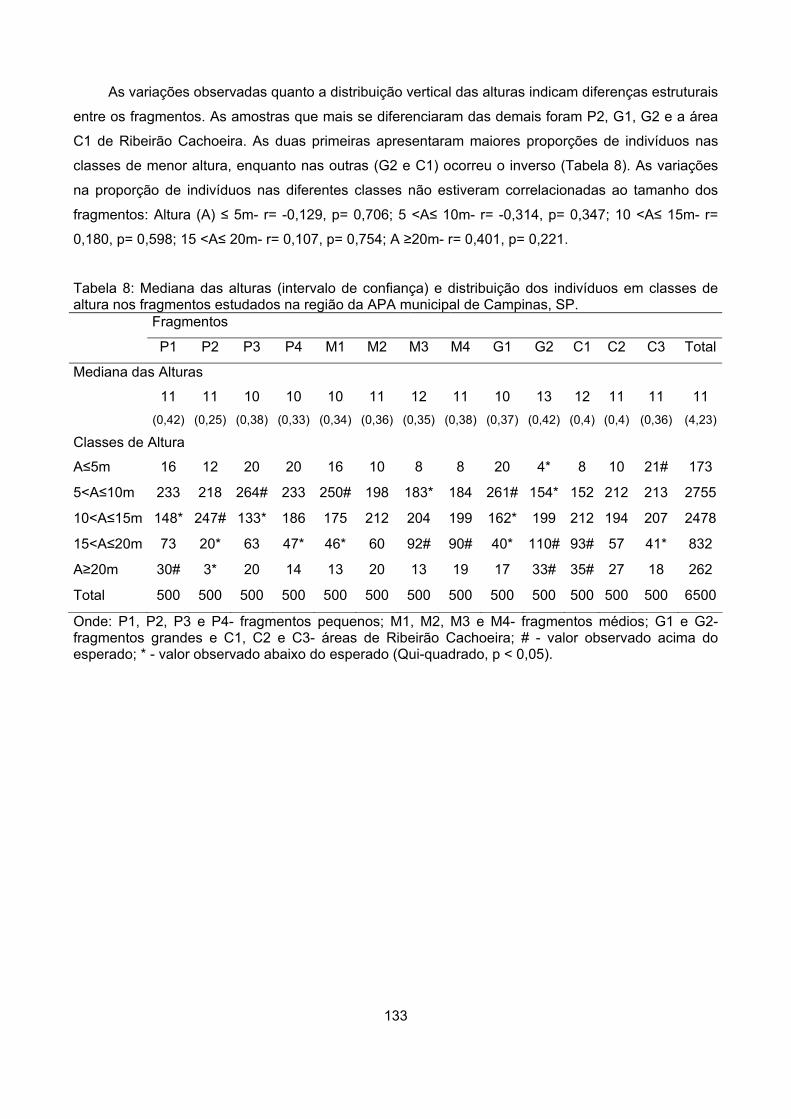
133
As variações observadas quanto a distribuição vertical das alturas indicam diferenças estruturais
entre os fragmentos. As amostras que mais se diferenciaram das demais foram P2, G1, G2 e a área
C1 de Ribeirão Cachoeira. As duas primeiras apresentaram maiores proporções de indivíduos nas
classes de menor altura, enquanto nas outras (G2 e C1) ocorreu o inverso (Tabela 8). As variações
na proporção de indivíduos nas diferentes classes não estiveram correlacionadas ao tamanho dos
fragmentos: Altura (A) ≤ 5m- r= -0,129, p= 0,706; 5 <A≤ 10m- r= -0,314, p= 0,347; 10 <A≤ 15m- r=
0,180, p= 0,598; 15 <A≤ 20m- r= 0,107, p= 0,754; A ≥20m- r= 0,401, p= 0,221.
Tabela 8: Mediana das alturas (intervalo de confiança) e distribuição dos indivíduos em classes de altura nos fragmentos estudados na região da APA municipal de Campinas, SP. Fragmentos
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3 Total
Mediana das Alturas
11 (0,42)
11 (0,25)
10 (0,38)
10 (0,33)
10 (0,34)
11 (0,36)
12 (0,35)
11 (0,38)
10 (0,37)
13 (0,42)
12 (0,4)
11 (0,4)
11 (0,36)
11 (4,23)
Classes de Altura
A≤5m 16 12 20 20 16 10 8 8 20 4* 8 10 21# 173
5<A≤10m 233 218 264# 233 250# 198 183* 184 261# 154* 152 212 213 2755
10<A≤15m 148* 247# 133* 186 175 212 204 199 162* 199 212 194 207 2478
15<A≤20m 73 20* 63 47* 46* 60 92# 90# 40* 110# 93# 57 41* 832
A≥20m 30# 3* 20 14 13 20 13 19 17 33# 35# 27 18 262
Total 500 500 500 500 500 500 500 500 500 500 500 500 500 6500
Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira; # - valor observado acima do esperado; * - valor observado abaixo do esperado (Qui-quadrado, p < 0,05).
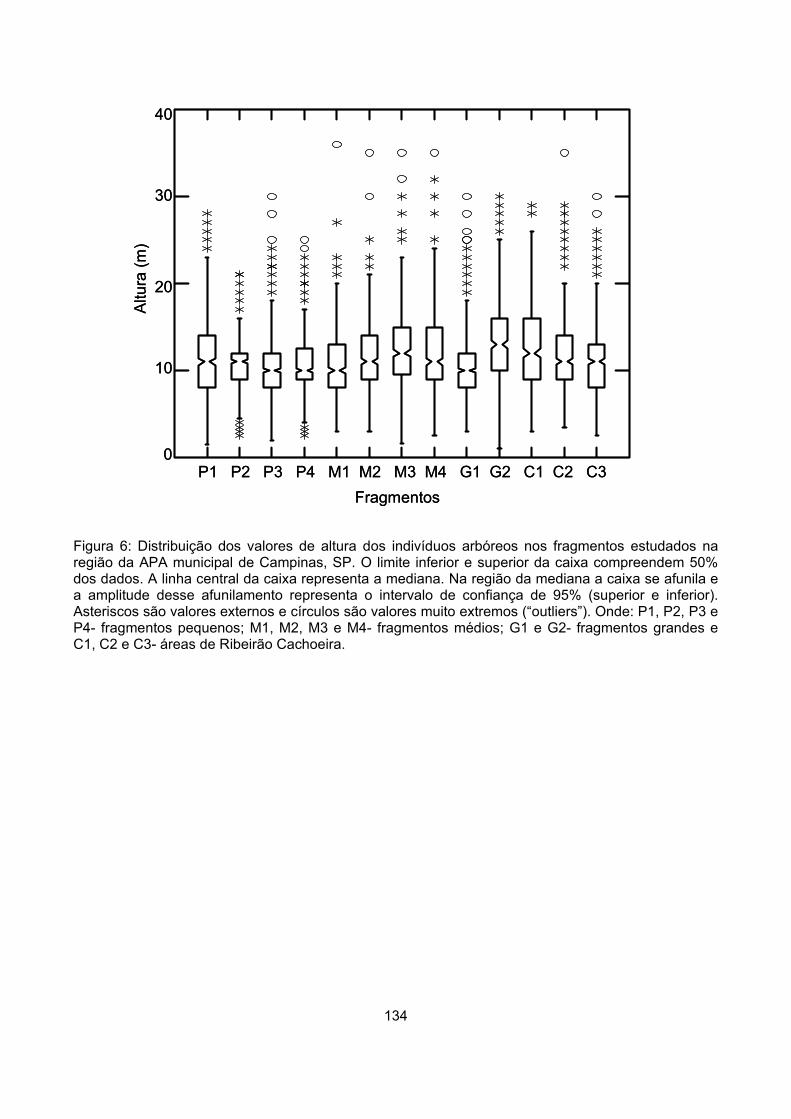
134
Figura 6: Distribuição dos valores de altura dos indivíduos arbóreos nos fragmentos estudados na região da APA municipal de Campinas, SP. O limite inferior e superior da caixa compreendem 50% dos dados. A linha central da caixa representa a mediana. Na região da mediana a caixa se afunila e a amplitude desse afunilamento representa o intervalo de confiança de 95% (superior e inferior). Asteriscos são valores externos e círculos são valores muito extremos (“outliers”). Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3Fragmentos
40
30
20
10
0
Altu
ra (m
)
P1 P2 P3 P4 M1 M2 M3 M4 G1 G2 C1 C2 C3Fragmentos
40
30
20
10
0
Altu
ra (m
)
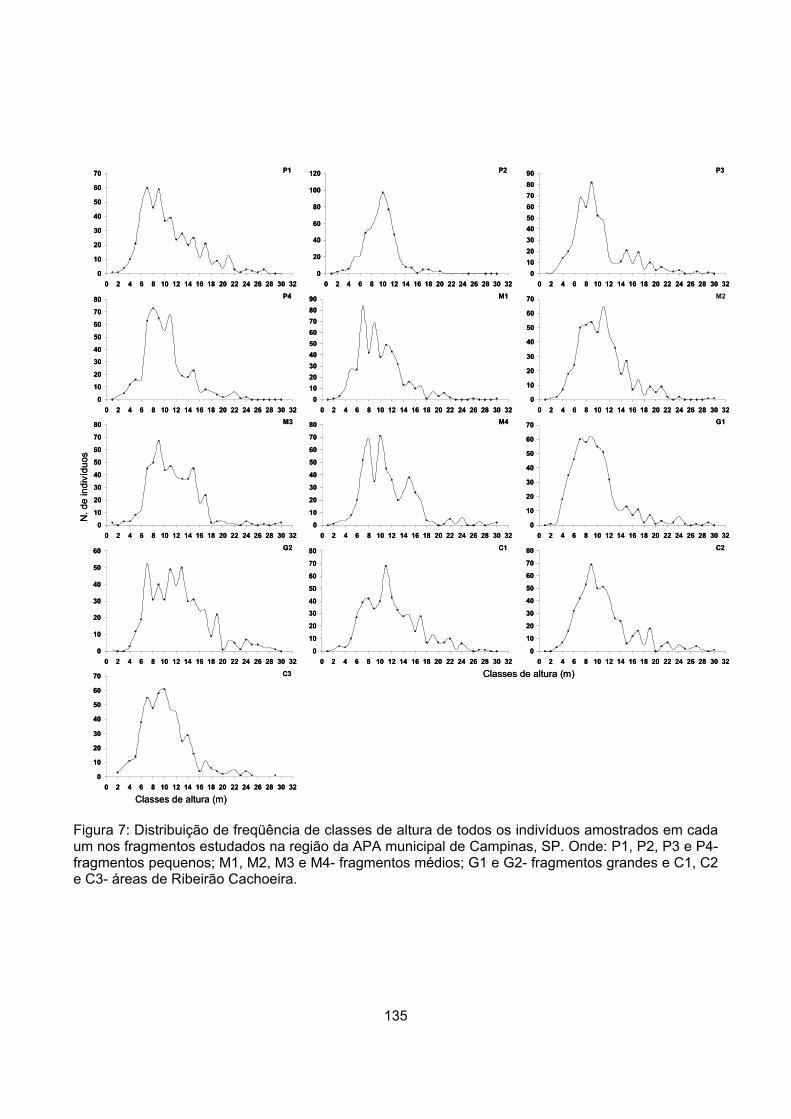
135
Figura 7: Distribuição de freqüência de classes de altura de todos os indivíduos amostrados em cada um nos fragmentos estudados na região da APA municipal de Campinas, SP. Onde: P1, P2, P3 e P4- fragmentos pequenos; M1, M2, M3 e M4- fragmentos médios; G1 e G2- fragmentos grandes e C1, C2 e C3- áreas de Ribeirão Cachoeira.
M2
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
N. d
e in
diví
duo s
Classes de altura (m)
Classes de altura (m)
P1
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P2
0
20
40
60
80
100
120
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P3
0102030405060708090
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
M1
0102030405060708090
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32M3
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
M4
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
G1
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
G2
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P4
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C1
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C2
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C3
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
M2
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
N. d
e in
diví
duo s
Classes de altura (m)
Classes de altura (m)
P1
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P2
0
20
40
60
80
100
120
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P3
0102030405060708090
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
M1
0102030405060708090
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32M3
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
M4
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
G1
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
G2
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
P4
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C1
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C2
0
10
20
30
40
50
60
70
80
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
C3
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32

136
Discussão No presente trabalho as densidades variaram de 515 a 960 ind/ha. Em outros trabalhos
realizados em remanescentes de floresta estacional semidecídua do estado de São paulo que
adotaram DAP ≥ 10cm a densidade de indivíduos variou de 624,4 a 793,2ind/ha (Cavassan et al.
1984, Matthes et al. 1988, Bertoni et al. 1988, Baitello et al. 1992). Comparado a esses trabalhos,
somente as densidades encontradas nos fragmentos pequenos P1, P3 e P4 poderiam ser
consideradas altas e as encontradas no fragmento grande G2 e áreas C1 e C2 de Ribeirão Cachoeira
poderiam ser consideradas baixas. Entretanto, não houve correlação entre a densidade e a área dos
fragmentos como previsto nas proposições da literatura, segundo as quais fragmentos pequenos
estariam mais sujeitos à perturbações, o que aumentaria as taxas de mortalidade e reposição,
levando a um aumento da densidade (Laurance et al. 1998a, Willians-Linera 1990).
A relação entre perturbações e densidade pode não ser tão simples assim. O fragmento que
sofreu perturbações mais rescentes e intensas (G2- ver Capítulos 1 e 2) apresentou a menor
densidade total e também um baixo número de espécies (Tabela 1). Essas observações parecem
estar de acordo com o encontrado por Cadotte et al. (2002) e Chittibabu & Parthasarathy (2000), em
que a densidade, assim como a diversidade, diminuiu com a intensidade das perturbações. É
possível que o aumento da densidade ocorra em resposta a perturbações menos intensas. A
ocorrência de perturbações muito drásticas provocaria alterações muito profundas, que impediriam o
aumento da regeneração em resposta. Fatores de perturbação de diferentes intensidades e
amplitudes fatalmente produzirão efeitos diferentes sobre as comunidades (Burslem & Whitmore
1999).
Baixos valores de área basal também têm sido associados à ocorrência de perturbações (Silva
& Nascimento 2001). Entretanto, comparado a outros estudos de floresta estacional semidecídua que
adotaram DAP ≥ 10cm, nenhum dos valores obtidos aqui pode ser considerado baixo ou abaixo da
variação encontrada para esse ecossistema. Bertoni et al. (1988) encontraram 18,28m2/ha, Cavassan
et al. (1984), 24,96 m2/ha e Baitello et al. (1992), 47,86 m2/ha.
Para Willians-Linera (2002), parâmetros como área basal e densidade variam muito em florestas
neotropicais e por isso não seriam bons indicadores de perturbação. Segundo a autora, a densidade
de árvores (DAP ≥ 5cm) em tais florestas pode variar entre 500 a mais de 1000ind/ha. Em
remanescentes florestais do interior do estado de São Paulo, a densidade variou entre 968 e
2577ind/ha em trabalhos que adotaram DAP ≥ 5cm (Pinto 1989, Martins 1991, Catharino 1989,
Bernacci 1992, Duringan et al. 2000, Cielo Filho & Santin 2002). No estudo de Metzger et al. (1998),
que amostrou oito fragmentos de floresta estacional semidecídua na região do Rio Jacaré Pepira e
adotou DAP ≥ 3cm a densidade variou de 2040 a 5479ind/ha. As variações observadas entre tais
estudos corroboram a observação de Willians-Linera (2002) e assim devemos ter cautela quando
tentamos associar variações na densidade ou área basal a quaisquer aspectos bióticos ou abióticos.

137
Dessa forma, a variação observada na densidade entre os fragmentos estudados pode estar
associada a outros fatores, além da área total e ocorrência de perturbações.
A concentração de abundância, observada em todas as áreas, é muito comum (Hughes 1996).
Nas comunidades de plantas a maioria dos indivíduos geralmente pertencem a poucas espécies,
enquanto a maioria das espécies encontra-se representada por um pequeno número de indivíduos.
Embora a maioria dos autores não comente a ocorrência desse padrão, uma rápida análise das
listagens dos parâmetros fitossociológicos fornecidas por trabalhos similares permite-nos observar
que a concentração de abundância ocorre na maioria dos estudos (Cavassan et al. 1984, Matthes et
al. 1988, Bertoni et al. 1988, Baitello et al. 1992, entre outros). Apesar de ser um padrão comum,
observamos que em alguns fragmentos, como P3 e G2, essa concentração foi mais acentuada. A
distribuição de abundância das espécies de uma amostra fornece um tipo de medida de diversidade,
a equabilidade. Quanto mais equilibrada é a distribuição dos indivíduos entre as espécies, isto é,
quanto menor a concentração da abundância, maior a equabilidade, o que é convencionalmente
entendido como alta diversidade (Magurran 1988). Dentro desse contexto poderíamos dizer que os
fragmentos P3 e G2 tendem a ser os menos diversos do estudo (ver Capítulo 3). A grande
concentração de abundância observada nessas amostras também se reflete em outros parâmetros
analisados, como área basal e VI. Em outras áreas, como P1, G1 e na área C2 de Ribeirão
Cachoeira, ocorreu o inverso: a concentração de abundância foi menor, sendo necessário um maior
número de espécies para compor mais da metade dos valores de área basal e VI. As variações
observadas quanto a esses quesitos, portanto, não pareceram estar associados às categorias de
tamanho dos fragmentos, uma vez que concentrações maiores ou menores ocorreram em fragmentos
de diversos tamanhos.
De forma geral, houve uma relação positiva entre a abundância e a amplitude de distribuição
das espécies, como previsto por Gaston (1994). Espécies com distribuição geográfica mais ampla
tendem a ser mais abundantes localmente (Brown 1984, 1995). Entretanto, houve grande variação,
com algumas espécies abundantes localmente, enquanto outras ocorrendo em várias áreas, mas
com baixo número de indivíduos. Várias espécies foram muito abundantes em um fragmento e pouco
abundantes ou ausentes em outros. Exemplos desse padrão ocorreram em todas as amostras, com
exceção dos fragmentos P1 e M2 (Figura 2). O fragmento no qual um maior número de espécies
apresentou abundância relativa equivalente a pelo menos duas vezes a apresentada nas demais
áreas de ocorrência foi P4, com as espécies Bauhinia forficata, Machaerium nictitans, Nectandra
megapotamica e Gallesia integrifolia.
Para Brown (1984), isso ocorreria porque as espécies teriam ambientes preferenciais e sua
abundância seria máxima nos locais em que a maior parte de seus requerimentos bióticos e abióticos
fosse satisfeita. Quando o ambiente preferencial ocorresse em manchas, ocorreriam múltiplas modas
na distribuíção de abundância da espécie. Essa última observação poderia explicar por que algumas
espécies, que figuraram entre as mais abundantes, considerando o total coletado, nem mesmo

138
ocorreram em algumas áreas, como é o caso de Esenbeckia leiocarpa, Trichilia clausseni e Croton
floribundus. Tais padrões de distribuição das espécies seriam, em última análise, determinados pela
heterogeneidade ambiental.
Uma comparação entre as dez famílias mais importantes (VI) em cada uma das 13 amostras e
outros levantamentos fitossociológicos realizados no interior do estado de São Paulo indicou que as
famílias Apocynaceae, Euphorbiaceae, Fabaceae, Caesalpiniaceae, Mimosaceae, Lauraceae,
Meliaceae, Moraceae, Myrtaceae e Rutaceae estão entre as de maior VI na maioria dos inventários
(Silva & Leitão Filho 1982, Cavassan et al. 1984, Bertoni et al. 1988, Matthes et al. 1988, Rodrigues et
al. 1989 e Grombone et al. 1990). Tais famílias foram citadas por Leitão Filho (1982) como
caracteristicamente abundantes nas matas do interior do estado de São Paulo e caracterizadoras
deste tipo de formação. Entretanto, existe uma variação de posição entre essas famílias no ranking
de VI e da proporção do valor total de VI que cada família representa, tanto entre as amostras do
presente estudo quanto em relação aos trabalhos comparados. Além disso, a maioria das amostras
do presente estudo apresentou famílias entre as dez de maior VI que raramente figuraram como tal
em outra amostra ou levantamento comparado, como Asteraceae, Cecropiaceae e Ebenaceae. Tais
variações não pareceram associadas ao tamanho dos fragmentos. Ferreira & Prance (1998),
estudando quatro trechos de floresta Amazônica de terra firme, encontraram variações semelhantes
às do presente estudo quanto ao ranking de VI de famílias, tanto entre os trechos analisados quanto
entre outros levantamentos fitossociológicos realizados na região. Para os autores, essas variações
reforçam a idéia de que aquelas florestas são um mosaico de comunidades florística e
estruturalmente distintas, resultantes da variação de condições edáficas, topográficas, geológicas e
climáticas. É possível que o mesmo esteja ocorrendo nas florestas de São Paulo e com os fragmento
deste estudo, pois, como comentou Leitão Filho (1982), as florestas estacionais semidecíduas
ocupam uma área bastante expressiva do sudeste do Brasil, em áreas de clima e solo variados e
muitas vezes entremeadas com outras formações vegetais, como cerrados, matas ciliares e campos
rupestres. Todos estes fatores podem contribuir para a heterogeneidade entre os remanescentes
atuais.
Nas comparações com outros trabalhos observamos ainda que algumas famílias que atingiram
altos VIs em várias amostras do presente estudo raramente figuraram como tal nos trabalhos
supracitados, como por exemplo Lecythidaceae e Urticaceae. A família Lecythidaceae apresenta
altos VIs apenas no estudo de Salis et al. (1994), mas no presente estudo figurou entre as 10 mais
importantes em sete amostras (Tabela 5). As espécies desta família são consideradas características
dos estádios finais do processo sucessional, o que significa que são mais abundantes em florestas
maduras ou bem preservadas (Salis et al. 1994, Gandolfi et al. 1995, Bernacci & Leitão Filho 1996 e
Sanchez et al. 1999).
As famílias Urticaceae e Cecropiaceae também raramente apresentam altos VIs nos trabalhos
consultados. Urticaceae figurou entre as mais importantes nas amostras M1, G2 e C3, enquanto

139
Cecropiaceae foi a oitava no ranking de VI na área C3 de Ribeirão Cachoeira. Para Bertoni et al.
(1988), as espécies de Urticaceae constituem um componente normal deste tipo de mata, mas sua
ocorrência com altos VIs poderia indicar maior ocorrência de fatores de perturbação. Todas as áreas
onde Urticaceae apresentou altos VIs, principalmente G2, foram expostas a fatores de perturbação,
como queimadas recentes, o que corrobora a hipótese de Bertoni et al. (1988). Entretanto, é
interessante notar que Urticaceae, Cecropiaceae e Lecythidaceae atingiram altos VIs nos mesmos
fragmentos.
Tais observações corroboram a idéia de que as formações florestais seriam naturalmente um
mosaico de manchas de vegetação em estádios sucessionais distintos (Hartshorn 1978, Whitmore
1984), e que mesmo as florestas mais bem preservadas apresentam ao longo do tempo variações
nas proporções das espécies típicas de cada estádio sucessional, pela abertura de clareiras,
eventuais deslizamentos, incêncios, ou seja, pela própria dinâmica da floresta e por fatores
intrínsecos da biologia de cada espécie, como a longevidade (Spurr 1952, Whitmore 1983).
As espécies que figuraram entre as de maior VI na maioria dos fragmentos deste estudo foram
Aspidosperma polyneuron, Astronium graveolens, Centrolobium tomentosum, Chorisia speciosa,
Croton floribundus, Esenbeckia leiocarpa e Piptadenia gonoacantha. Tais espécies são comumente
citadas com as de maior VI em estudos fitossociológicos realizados no interior do estado de São
Paulo (Cavassan et al. 1985, Bertoni et al. 1988, Matthes et al. 1988, Rodrigues et al. 1989,
Grombone et al. 1990, Salis et al. 1994, Tabanez et al. 1997, Duringan et al. 2000, Cielo Filho 2001,
Silva & Soares 2002, Cielo Filho & Santin 2002). Isto pode indicar que os resultados encontrados
poderiam ser considerados o esperado para esse tipo de ecossistema. Entretanto, 50% das plantas
que apresentaram altos VIs neste estudo não são citadas como tal em nenhum dos estudos
supracitados. Como exemplo, podemos citar Cariniana legalis, Calycorectes acutatus, Holocalyx
balansae, Hymenaea courbaril e Ficus glabra, que atingiram altos VIs em diversos fragmentos,
independentemente de sua área total (Tabela 6). A maioria dessas espécies pode ser considerada
comum, pois figuram freqüentemente em listagens florísticas de florestas semidecíduas do estado
(Tamashiro et al. 1986, Bertoni & Martins 1987, Pagano & Leitão Filho 1987, Matthes et al. 1988,
Costa & Mantovani 1995, Bernacci & Leitão Filho 1996, Metzger et al. 1997, Santos & Kinoshita
2003). Apesar disso, aparentemente só atingiram altos VIs no presente estudo. Algumas dessas
espécies são de grande porte, consideradas por muitos autores espécies características dos estadios
finais de sucessão (Salis et al. 1994, Gandolfi et al. 1995, Penhalber & Mantovani 1997, Metzger et al.
1997, Dislich et al. 2001). Isto pode indicar que nos fragmentos em estudo as condições para a
ocorrência dessas espécies podem ser mais favoráveis à sua ocorrência e dominância. Dessa forma,
as áreas em que essas espécies atingiram altos VIs poderiam ser consideradas importantes para a
manutenção dessas espécies.
A maior parte das variações entre os rankings de VIs de espécies entre as áreas se refere à
posição que as espécies ocupam no ranking nas diferentres áreas e à proporção do valor total de VI

140
que cada espécie representa. Algumas espécies estão entre as dez de maior VI em várias amostras,
mas em determinada área representaram quase um terço do valor total de VI, indicando grande
dominância naquela área. Isto ocorreu principalmente nas áreas P2, P3 e G2. Além disso, a maioria
dos fragmentos apresentou uma, duas ou três espécies que atingiram altos VIs somente naquela
área. Algumas espécies, como mencionado anteriormente, tiveram ocorrência restrita àqueles
fragmentos em que atingiram um dos maiores VIs da amostra. São exemplos Trichilia casaretti,
Mabea fistulifera e Metrodorea stipularis. Várias espécies atingiram altos VIs em alguns fragmentos e
nem mesmo ocorreram em outros. Os fragmentos que mais se diferenciaram dos demais em relação
à composição do ranking de VI para espécies foram P2, M3 e C2. Tais observações indicam
diferenças estruturais entre os fragmentos, embora não pareçam associadas à sua área total.
A concentração de indivíduos nas classes de menor diâmetro, formando curva de distribuição
em forma de J invertido foi observada em todas as amostras. Segundo Uutera et al. (2000), este é um
padrão freqüentemente observado em florestas tropicais, tendo sido observado em florestas
estacionais semidecíduas por Cavassan et al. (1984), Matthes et al. (1988), Bertoni et al. (1988) e
Lopes et al. (2002), em outras florestas tropicais por Silva & Nascimento (2001), Ferreira & Prance
(1998), Willians-Linera (2002) e Uutera et al. (2000) e no Cerrado por Felfili & Silva Júnior (1988) e
Silva Júnior e Silva (1988). Apesar do mesmo padrão a proporção de indivíduos em cada classe de
diâmetro variou bastante entre as áreas. Os fragmentos que mais se diferenciaram dos demais foram
P3 e G2 (Tabela 7). No primeiro ocorreu uma menor proporção de indivíduos nas classes de maior
diâmetro, enquanto em G2 ocorreu o inverso. As variações observadas não pareceram estar
associadas ao tamanho dos fragmentos e sim a fatores como densidade e composição. Em P3 quase
50% dos indivíduos pertencem a uma única espécie dos quais 83% ocorreram com diâmetros
inferiores a 20cm. Dessa forma, a maior porporção de indivíduos nas classes de menor diâmetro em
P3 pode estar associada a dominância dessa espécie. O fragmento G2 apresentou a menor
densidade de indivíduos do estudo e a maior proporção de indivíduos nas classes de maior diâmetro.
Segundo Denslow (1995), existe uma correlação inversa entre densidade e o diâmetro médio dos
indivíduos. Assim, seria esperado que nas áreas de menor densidade a proporção de indivíduos
grandes fosse maior. Essa relação pareceu válida para a maioria dos fragmentos. A baixa densidade
em G2, por sua vez, pode estar associada a ocorrência e intensidade de fatores de perturbação na
área como mencionada anteriormente. Além disso, fatores edáficos, topográficos e de relevo também
podem influenciar no tamanho dos indivíduos.
Assim como a proporção de indivíduos nas várias classes de diâmetro, houve grande variação
entre os fragmentos quanto à proporção de indivíduos em cada classe de altura e também quanto
aos valores de mediana das alturas. Novamente, as variações observadas não estiveram associadas
ao tamanho dos fragmentos. Para Montagnini & Jordan (2002), fatores edáficos, como ausência ou
excesso de alguns nutrientes no solo, podem limitar o crescimento das plantas e dessa forma
influenciar na altura máxima que os indivíduos atingiriam em determinadas áreas. Além disso, Basnet

141
(1992) e Bellinghan & Tanner (2000) observaram que a proporção de árvores grandes era maior em
trechos de mata menos acidentados. Algumas matas que apresentaram valores de mediana de altura
menores do que as demais amostras apresentam topografia muito acidentada e declividade bastante
acentuada em alguns trechos, como P4, M1 e G1. A mata G2, por outro lado, apresenta relevo
menos acidentado que as demais áreas (Capítulos 1 e 2). Nessa área o valor da mediana das alturas
foi o mais elevado dentre os fragmentos, além disso este, juntamente com o fragmento P1 e a área
C1 apresentou maior proporção de indivíduos nas classes de maior altura. É possível que
características do solo, do relevo e da topografia estejam associados aos padrões observados nas
referidas áreas, entretanto tais aspectos não foram devidamente investigados no presente estudo,
sendo necessários estudos específicos para comprovar tal hipótese.
Assim como no presente estudo, Popma et al. (1988) também não conseguiram determinar
estratos verticais na vegetação através de diagramas de classes de altura em uma floresta tropical do
México. A mesma dificuldade foi mencionada por Lopes et al. (2002) em uma floresta estacional
semidecídua de Minas Gerais. Para Popma et al. (1988), esta dificuldade decorre de a distribuição
das copas ao longo do plano vertical ser contínua, ao invés de descontínua, com camadas bem
definidas. Segundo os autores, se essas camadas realmente ocorrem, então suas alturas devem
variar ao longo da floresta, pois as variações na estrutura da vegetação tanto horizontal quanto
verticalmente, pareciam ocorrer ao longo de distâncias relativamente curtas. Algo semelhante pode
estar ocorrendo nos fragmentos em estudo.
Os fragmentos que se diferenciaram dos demais em relação à maior parte dos parâmetros
analisados foram P2, P3 e G2, dois fragmentos pequenos e um grande. O fragmento P2 apresentou
diferenças em relação a estratificação vertical e os rankings de VI de espécies foram muito
diferenciados em relação às demais amostras. Tais características podem estar associadas a seu
histórico de preservação: a mata já foi parcialmente atingida por queimadas e a ocorrência de fogo
em seu entorno é algo freqüente. Além disso, essa mata ocupa solo e relevo diferente de todas as
demais (Capítulos 1 e 2).
No caso de P3, grande parte das variações observadas, pode estar associada à
monodominância de Trichilia clausseni na área. Essa espécie representou quase 50% do total de
indivíduos coletados, 25% da área basal e um terço do VI da área. Talvez devido à monodominância
dessa espécie, algumas famílias e espécies abundantes e importantes nas demais áreas não figurem
como tal no fragmento P3. Trichilia clausseni é uma espécie de médio porte, que dificilmente atinge
diâmetros superiores a 40cm. Talvez por isso a área P3 tenha apresentado um número menor de
indivíduos nas classes de maior diâmetro em relação às outras áreas, como mencionado
anteriormente. Em outro fragmento do município de Campinas, estudado por Matthes et al. (1988),
essa espécie também foi desproporcionalmente abundante, representando quase um terço do total
de indivíduos coletados e 25% do índice de valor de cobertura (área basal). É possível que essa
espécie se torne dominante em áreas onde encontre condições ambientais ótimas para seu

142
estabelecimento. Estudos sobre a estrutura populacional e distribuição espacial dessa espécie seriam
muito interessantes para se tentar compreender melhor os aspectos envolvidos na ocorrência dessa
espécie na condição de monodominante.
No fragmento G2, a maior parte das diferenças observadas em relação às outras áreas pode
estar relacionada a seu histórico de perturbação, marcado pela ocorrência de uma queimada
bastante intensa que atingiu a mata quase por completo. É possível que apenas as árvores maiores e
mais altas de determinadas espécies tenham sobrevivido à essa queimada e, por isso, a mata exiba
maior proporção de árvores grandes, altas e maior concentração de indivíduos e área basal por
espécie.
Como mencionado acima, os fragmentos P2, P3 e G2 diferiram dos demais em quase todos os
aspectos analisados. Entretanto, todos os fragmentos pareceram exibir alguma particularidade do
ponto de vista estrutural que o distingue dos demais, dependendo do parâmetro considerado. No
fragmento P4 ocorreu o maior número de espécies representadas por apenas um indivíduo. Os
fragmentos P1 e G1 exibiram a menor concentração de indivíduos por espécie, indicando maior
diversidade. As áreas C3 e M2 foram as que exibiram o maior número de famílias com altos VIs
somente nessas áreas. As áreas P2, M3 e C2 exibiram a composição de espécies de maior VI mais
diferenciado em relação às demais amostras. Além disso, as áreas de Ribeirão Cachoeira pareceram
ser tão heterogêneas entre si quanto os fragmentos isolados ao longo da paisagem. Não foi possível,
portanto, associar todas essas variações simplesmente ao tamanho dos fragmentos. Possivelmente,
as variações observadas se devem a um conjunto de características bióticas e abióticas específicas
de cada fragmento. É importante lembrar que variações na estrutura da vegetação podem ocorrer
tanto em função de perturbações naturais (Lugo & Scatena 1996), antrópicas (Laurance et al. 1997),
em decorrência da fragmentação (Laurance et al. 1998a e 1998b), efeito de borda (Willians-Linera
1990), mas também ao longo de distâncias relativamente curtas, influenciadas por condições
abióticas como solo (Oliveira-Filho et al. 1994a, Botrel et al. 2002) , umidade (Oliveira-Filho et al.
1994b) e topografia (Lieberman et al. 1985, Ashton & Hall 1992, Basnet 1992, Bellinghan & Tanner
2000, Clark & Clark 2000). Dessa forma, numerosos fatores bióticos e abióticos, bem como as
interações entre eles, podem estar influenciando as variações estruturais observadas entre os
fragmentos do presente estudo e seriam necessários estudos mais específicos para determinar quais
fatores seriam preponderantes em cada área.
Portanto, as variações nos aspectos estruturais analisados não pareceram estar associadas
somente ao tamanho dos fragmentos. Esses fragmentos estão expostos a perturbações de tamanho,
amplitude e intensidade diferentes e, além das diferenças de tamanho, ocupam diversos tipos de
solo, relevo, hidrografia e tipo de ocupação do entorno (Capítulo 1). Assim, as variações observadas
podem ser resultantes de todos esses fatores em conjunto e não apenas relacionadas à área dos
fragmentos. Essas variações, aliadas às variações nos padrões de distribuição e importância das
espécies nos diferentes fragmentos, ressaltam sua importância em termos de conservação, pois é

143
possível que muitas dessas áreas constituam os últimos refúgios na região para determinadas
espécies.

144
Referências bibliográficas
ASHTON, P.S. & HALL, P. 1992. Comparisons of structure among mixed Dipterocarp forest of North-
Western Borneo. Journal of Ecology 80: 459-481.
AYRES, M., AYRES Jr., M., AYRES, D.M. & SANTOS, A.S. 1998. BioEstat: Aplicações estatísticas nas áreas das Ciências Biológicas e Médicas. Sociedade civil Mamirauá. Manaus.
BAITELLO, J.B., AGUIAR, O.T., ROCHA, F.T., PASTORE J.A., & ESTEVES, R. 1992. Florística e
fitossociologia do estrato arbóreo de um trecho da serra da Cantareira (Núcleo Pinheirinho) - SP
Anais II Congresso Nacional sobre Essências Nativas 291-297.
BASNET, K. 1992. Effect of topography on the pattern of trees in Tabonuco (Dacryodes excelsa)
dominated rain forest of Porto Rico. Biotropica 24: 31-42
BELLINGHAM, P.J. & TANNER, V.J. 2000. The influence of topography on tree growth, mortality, and
recruitment in a tropical montane forest. Biotropica 32: 378-384.
BERNACCI, L.C. & LEITÃO FILHO, H.F 1996. Flora fanerogâmica da floresta da Fazenda São
Vicente, Campinas, SP. Revista Brasileira de Botânica 19: 149-164.
BERNACCI, L.C. 1992. Estudo florístico e fitossociológico de uma floresta no município de Campinas, com ênfase nos componentes herbáceo e arbustivo. Dissertação de Mestrado.
Universidade Estadual de Campinas. Campinas.
BERTONI, J.E.A., MARTINS, F.R 1987. Composição florística de uma floresta ripária na Reserva
Estadual de Porto Ferreira,SP. Acta Botanica Brasilica 1:17-26.
BERTONI, J.E.A., MARTINS,F.R., MORAES, J.L. & SHEPHERD, G.J. 1988. Composição florística e
estrutura fitossociológica do Parque Estadual de Vaçununga, Santa Rita do Passa Quatro –
Gleba Praxedes, Boletim Técnico do Instituto Florestal de São Paulo 42: 149-170.
BOTREL, R.T., OLIVEIRA FILHO, A. T., RODRIGUES, L.A. & CURI, N. 2002. Influência do solo e
topografia sobre as variações na composição florística e estrutura da comunidade arbóreo-
arbustiva de uma floresta estacional semidecidual em Ingaí, MG. Revista Brasileira de Botânica
25: 195-213.
BROWN, I.F., MARTINELLI, L.A., THOMAS, W.W., MOREIRA, M.Z., CID FERREIRA, C.A. &
VICTORIA, R.A. 1995. Uncertainty in the biomass of Amazonian forests: an example from
Rondonia, Brazil. Forest Ecology and Management 75: 175-189.
BROWN, J,H. 1995. Macroecology. The University Chicago Press, Chicago.
BROWN, J.H. 1984. On the relationship between abundance and distribution of species. The American Naturalist 124: 255-279.
BURSLEM, D.F.R.P. & WHITMORE, T.C. 1999. Species diversity, suscetibility to disturbance and tree
population dynamics in tropical rain forest. Journal of Vegetation Science 10: 767-776.
CADOTTE, M.W., FRANCK, R., REZA, L. & LOVETT-DOUST, J. 2002. Tree and shrub diversity and
abundance in fragmented littiral forest of southeastern Madagascar. Biodiversity and Conservation 11: 1417-1436.

145
CATHARINO, E.L.M. 1989. Estudo fisionômico, florístico e fitossociológico em matas residuais secundárias do município de Piracicaba, SP. Dissertação de Mestrado. Universidade Estadual
de Campinas. Campinas, SP.
CAVASSAN, O., CESAR, O. & MARTINS, F.R. 1984. Fitossociologia da vegetação arbórea da
reserva estadual de Bauru, Estado de São Paulo. Revista Brasileira de Botânica 7: 91-106.
CHITTIBABU, C.V. & PARTHASARATHY, N 2000. Attenuated tree species diversity in huma-
impacted tropical evergreen forest sites at Kolli hills, Eastern Ghats, India. Biodiversity and Conservation 9: 1493-1519.
CIELO FILHO, R. & SANTIN, D.A. 2002. Estudo florístico e fitossociológico de um fragmento florestal
urbano - Bosque dos Alemães, Campinas, SP. Revista Brasileira de Botânica 25: 291-301.
CIELO FILHO, R. 2001. Estrutura e abundância de um trecho de floresta estacional semidecídua no município de Campinas, estado de São Paulo: Mata Ribeirão Cachoeira.
Dissertação de Mestrado. Universidade Estadual de Campinas. Campinas, SP.
CLARK, D.B. & CLARK, D.A. 2000. Landscape-scale variation in forest structure and biomass in a
tropical rain forest. Forest Ecology and Management 137: 187-198.
CONDIT, R., HUBBELL, S.P. & FOSTER, R.B. 1996. Changes in tree species abundance in a
neotropical forest: impact of climat change. Journal of Tropical Ecology 12: 231-256.
COSTA, L.G.S. & MANTOVANI, W. 1995. Flora arbustivo-arbórea de trecho de mata mesófila
semidecídua, no sítio ecológico de Ibicatu, Piracicaba (SP). Hoehnea 22: 47-59.
COTTAN, G. & CURTIS, J.T. 1956. The use of distance measures in phytossociological sampling.
Ecology 37: 451-460.
CRONQUIST, A. 1981. An integrated system of classification of flowering plants. NewYork:
Columbia University Press & The New York Botanical Garden.
CURTIS, J.T. & McINTOSH, R.P. 1951. An upland forest continuum in the prairie-forest border region
of Wiscosin. Ecology 32: 476-496.
DENSLOW, J.S. 1995. Disturbance and diversity in tropical rain forests: The density effect.
Ecological Aplications 5: 962-968.
DIDHAM, R.K. & LAWTON, J.H. 1999. Edge structure determines the magnitude of changes in
microclimate and Vegetation structure in tropical forest fragments. Biotropica 31: 17-30.
DISLICH, R., CERSÓSIMO, L. & MANTOVANI, W. 2001. Análise da estrutura de fragmentos
florestais no planalto paulistano - SP. Revista Brasileira de Botânica 24: 321-332.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista Brasileira de Botânica 23: 371-383.
FELFILI, J.M. 1995. Growth, recruitment and mortality in the Gama gallery forest in central Brazil over
a six-year period. Journal of Tropical Ecology 11: 67-83.

146
FELFILI, J.M. & SILVA JÚNIOR, M.C. 1988. Distribuição dos diâmetros numa faixa de cerrado na
Fazenda Água Limpa (FAL) em Brasília-DF. Acta Botanica Brasilica 2: 85-104.
FERREIRA, L.V. & PRANCE, G.T. 1998. Species richness and floristic composition in four hectares in
the Jaú National Park in upland forests in Central Amazonia. Biodiversity and Conservation 7:
1349-1364.
GANDOLFI, S., LEITÃO FILHO, H.F. & BEZERRA, C.L.F. 1995. Levantamento florístico e carater
sucessional das espécies arbustivo-arbóreas de uma floresta mesófila semidecídua no município
de Guarulhos, SP. Revista Brasileira de Biologia 55: 753-767.
GASTON, K.J., 1994. Rarity. Chapman & Hall, London.
GROMBONE, M.T., BERNACCI,L.C., MEIRA NETO,J.A.A., TAMASHIRO,J.Y. & LEITÃO FILHO, H.
1990. Estrutura fitossociológica da floresta semidecídua de altitude do Parque Municipal de Grota
Funda (Atibaia-Estado de São Paulo). Acta Botanica Brasilica 4: 47-64.
HARTSHORN, G.S. 1978. Treefalls and tropical forest dynamics. Pp 617-638. In: P.B. Tonlinson &
M.H. Zimmerman (eds.). Tropical trees as living systems. Cambridge University Press, New
York.
HUGHES, R.G. 1986. Theories and models of species abundance. The American Naturalist 128:
879-899.
KAPOS, V. 1989. Effects of isolation on the water status of forest patches in the brazilian Amazon.
Journal of Tropical Ecology 5: 173-185
KÖPPEN, W. 1948. Climatologia. México - Buenos Aires. Ed. Fundo de Cultura Econômica, 478p.
LAURANCE, W.F., LAURANCE, S.G., FERREIRA, L.V., RANKIN-DE-MERONA, J.M. GASCON, C. &
LOVEJOY, T.E. 1997. Biomass collapse in Amazonian forest fragments. Science 278: 1117-
1118.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M., LAURANCE, S.G., HUTCHINGS,
R.W. & LOVEJOY, T.E. 1998a. Effects of forest fragmentation on recruitment patterns in
amazonian tree communities. Conservation Biology 12: 460-464.
LAURANCE, W.F., FERREIRA, L.V., RANKIN-DE-MERONA, J.M. & LAURANCE, S.G. 1998b. Rain
forest fragmentation and the dynamics of amazoniam tree communities. Ecology 79: 2032-2040.
LEITÃO FILHO, H.F. 1982. Aspectos taxonômicos das florestas do Estado de São Paulo. Silvicultura em São Paulo 16: 197-206.
LIEBERMAN, D., LIEBERMAN, M., PERALTA, R. & HARTSHORN, G.S. 1985. Mortality patterns and
stand turnover rates in a wet tropical forest in Costa Rica. Journal of Ecology 73: 915-924.
LOPES, W.P., SILVA, A.F., SOUZA, A.L. & MEIRA NETO, J.A.A. 2002. Estrutura fitossociológica de
um trecho de vegetação arbórea no Parque Estadual do Rio Doce - Minas Gerais, Brasil. Acta Botanica Brasilica 16: 443-456.
LOVEJOY, T.E., BIERREGAARD, R.O., RYLANDS , A.B., MALCOLM, J.R., QUINTELA, C.E.,
HARPER, L.H., BROWN, K.S., POWELL, A.H., POWELL, G.V.N., SCHUBART, H.O.R. & HAYS

147
.B. 1996. Edge and other effects of isolation on Amazon forest fragments. Pp 257-285. In: M.E.
Soule (ed.). Conservation Biology: the science of scarcity and diversity. Sinauer, Suderland.
USA
LUGO, A.E. & SCATENA, F.N. 1996. Background and catastropic tree mortality in tropical moist, wet,
and rain forests. Biotropica 28: 585-599.
MAGURRAN, A. 1988. Ecological diversity and its measurement. Croom Helm, London.
MARTINS, F.R. 1991. Estrutura de uma floresta mesófila. Editora da Universidade Estadual de
Campinas. Campinas.
MARTINS, F.R. SANTOS, F.A.M. 1999. Técnicas usuais de estimativas da biodiversidade. Revista
Holos 1: 236-267.
MATLACK, G.R. 1994. Vegetation dynamics of the edge - trends in space and successional time.
Journal of Ecology 82: 113-123.
MATTHES, L.A.F. 1980. Composição florística, estrutura e fenologia de uma floresta residual do planalto paulista: Bosque dos Jequitibás (Campinas, SP). Dissertação de Mestrado.
Universidade Estadual de Campinas, Campinas, SP.
MATTHES, L.A.F., LEITÃO FILHO, H.F. & MARTINS, F.R. 1988. Bosque dos Jequitibás (Campinas,
SP): composição florística e fitossociológica do estrato arbóreo. Anais V congresso da Sociedade Botânica de São Paulo, p. 55-76.
MATTOS, C.O. 1996. Contribuição ao planejamento e gestão da Área de Proteção Ambiental de Sousas e Joaquim Egídio, Campinas/SP. Dissertação de Mestrado. Universidade Estadual de
São Paulo. São Paulo.
MEIRA NETO, J.A.A., BERNACCI, L.C., GROMBONE, M.T., TAMASHIRO,GJ.Y. & LEITÃO FILHO,
H.F. 1989. Composição florística de mata semidecídua de altitude do Parque Municipal da Grota
Funda (Atibaia - Estado de São Paulo). Acta Botanica Brasilica 3: 51-74.
MELLO, M.H.A., PEDRO JUNIOR, M.J., ORTOLANI, A. A. & ALFONSI R.R. 1994. Chuva e
Temperatura: cem anos de observações em Campinas. Boletim Técnico 154. Instituto
Agronômico Campinas, SP.
MESQUITA, R.C.G., DELAMÔNICA, P. & LAURANCE, W.F. 1999. Effect of surrounding Vegetation
on edge-related tree mortality in Amazonian forest fragments. Biological Conservation 91: 129-
134.
METZGER, J.P., BERNACCI, L.C. & GOLDEMBERG, R. 1997. Pattern of tree species diversity in
riparian forest fragments of different widths (SE Brazi). Plant Ecology 133: 135-152.
METZGER, J.P., GOLDEMBERG, R. & BERNACCI, L.C. 1998. Diversidade e estrutura de fragmentos
de mata de várzea e de mata mesófila semidecídua submontana do rio Jacaré-Pepira (SP).
Revista Brasileira de Botânica 21: 321-330.

148
MIRANDA, Z.A.I. 1996. Plano de gestão da Área de Proteção Ambiental da região de Sousas e
Joaquim Egídio - APA Municipal. Relatório da Secretaria de Planejamento e Meio Ambiente - SEPLAMA. Prefeitura Municipal de Campinas, SP.
MONTAGNINI, F. & JORDAN, C.F. 2002. Reciclaje de nutrientes. Pp 167-191. In: M.R. Guariguata &
G.H. Kattan, (eds.), Ecología y conservación de bosques neotropicales. Libro Universitario
Regional.
MURCIA, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends in Ecology and Evolution 10: 58-62.
OLIVEIRA, J.B., MARCELO, N.C., ROSSI, M. & CALDERANO FILHO, B. 1999. Mapa pedológico do
estado de São Paulo. Legenda expandida In: OLIVEIRA, J.B. 1999. Boletim científico n.o 45,
Solos do estado de São Paulo. Embrapa e IAC. Campinas.
OLIVEIRA-FILHO, A.T. , VILELA, E.A., CARVALHO, D.A. & GAVILANES, M.L. 1994a. Effects of soil
and topography on the distribution of tree species in a tropical riverine forest in south-eastern
Brazil. Journal of Tropical Ecology 10: 483-508.
OLIVEIRA-FILHO, A.T. , VILELA, E.A., CARVALHO, D.A & GAVILANES, M.L. 1994b. Differentiation
of streamside and upland Vegetation in an area of montane semideciduous forest in southeaster
Brazil. Flora 189: 287-305.
OLIVEIRA-FILHO, A.T., CURY, N., VILELA, E. A . & CARVALHO, D.A . 1997. Tree species
distribution along soil catenas in a riverside semideciduous forest in southeastern Brazil. Flora
192: 47-64.
PAGANO, S.N. & LEITÃO FILHO, H.F. 1987. Composição florística do estrato arbóreo da mata
mesófila semidecídua no município de Rio Claro (Estado de São Paulo). Revista Brasileira de Botânica 10: 37-47.
PENHALBER, E.F. & MANTOVANI, W. 1997. Floração e chuva de sementes em mata secundária em
São Paulo, SP. Revista Brasileira de Botânica 20: 205-230.
PINTO, M.M. 1989. Levantamento fitossociológico de uma mata residual: Campus de Jaboticabal da UNESP. Dissertação de Mestrado. Universidade Estadual Paulista, Jaboticabal.
POPMA, J., BONGERS, F. & CASTILLO, J.M. 1988. Patterns in the vertical struture of the tropical
lowland rain forest of Los Tuxtlas, Mexico. Vegetatio 74: 81-91.
RANKIN-DE-MERONA, J.M., HUTCHINS, R.W. & LOVEJOY, T.E. 1990. Tree mortality and
recruitment over a five year period in undisturbed upland rainforest of central Amazon. In: A.H.
Gentry (ed.) Four neotropical rainforests. Yale University Press.
RODRIGUES, R. R., MORELLATO, L.P.C., JOLY, C.A. & LEITÃO FILHO, H.F. 1989. Estudo florístico
e fitossociológico em um gradiente altitudinal de mata estacional mesófila semidecídua, na Serra
do Japi, Jundiaí, SP. Revista Brasileira de Botânica 12: 71-84.

149
SALIS, S.M. TAMASHIRO, J.Y. JOLY, C.A. 1994. Florística e fitossociologia do estrato arbóreo de um
remanescente de mata ciliar no Rio Jacaré Pepira, Brotas, SP. Revista Brasileira de Botânica
17: 93-103.
SANCHEZ, M., PEDRONI, F., LEITÃO FILHO, H. F. & CESAR, O. 1999. Composição florística de um
trecho de floresta ripária na Mata Atlântica em Picinguaba, Ubatuba, SP. Revista Brasileira de Botânica 22: 31-42.
SANTIN, D.A. 1999. A vegetação remanescente do município de Campinas (SP): mapeamento, caracterização fisionômica e florística, visando a conservação. Tese de doutorado.
Universidade Estadual de Campinas, SP.
SANTOS, F.A.M. 1996. Programa para cálculo do coeficiente de Gini. Departamento de Botânica.
Instituto de Biologia, Universidade Estadual de Campinas, Campinas-SP.
SANTOS, F.A.M., TAMASHIRO, J.Y., RODRIGUES, R.R. & SHEPHERD, G.J. 1996. The dynamics of
tree populations in a semideciduous forest at Santa Genebra Reserve, Campinas SE Brazil.
Supplement to Bulletin of the Ecologiacl Society of America. 77: 389-389.
SANTOS, F.A.M., PEDRONI, F., ALVES, L.F. & SANCHEZ, M. 1998. Structure and dynamics of tree
species of the Atlantic Forest. Anais da Academia Brasileira de Ciências 70: 874-880.
SANTOS, K. & KINOSHITA, L.S. 2003. Flora arbustivo-arbórea do fragmento de floresta estacional
semidecidual do Ribeirão Cachoeira, Município de Campinas - SP. Acta Botanica Brasilica 17:
325-341.
SAUNDERS, D.A., HOBBS, R.J. & MARGULES, C.R. 1991. Biological consequences of ecossystem
fragmentation: a review. Conservation Biology 5: 18-32.
SHEPHERD, G.J. 1988. FITOPAC- versão preliminar. Manual do usuário. Departamento de
Botânica. Instituto de Biologia, Universidade Estadual de Campinas, Campinas-SP.
SILVA, A .F. & LEITÃO FILHO, H.F. 1982. Composição florística e estrutura de um trecho de mata
atlântica de encosta no município de Ubatuba (São Paulo, Brasil). Revista Brasileira de Botânica 5: 43-52.
SILVA, G.C. & NASCIMENTO, M.T. 2001. Fitossociologia de um remanescente de mata sobre
tabuleiros no norte do estado do Rio de Janeiro (Mata do Carvão). Revista Brasileira de Botânica 24: 51-62.
SILVA, L.A. & SOARES, J.J. 2002. Levantamento fitossociológico em um fragmento de floresta
estacuionla semidecídua, no município de São Carlos, SP. Acta Botanica Brasilica 16: 205-216
SILVA JÚNIOR, M.C. & SILVA, A.F. 1988. Distribuição dos diâmetros dos troncos das espécies mais
importantes do cerrado na Estação Florestal de Experimentação de Paraopeba (EFLEX)-MG.
Acta Botanica Brasilica 2: 107-126.
SPURR, S.H. 1952. Origin of the concept of forest sucession. Ecology 33: 426-27.

150
TABANEZ, A.A.J., VIANA, V.M. & DIAS, A.S. 1997. Conseqüências da fragmentação e do efeito de
borda sobre estrutura, diversidade e sustentabilidade de um fragmento de floresta de planalto de
Piracicaba, SP. Revista Brasileira de Biologia 57: 47-60.
TABARELLI, M. & MANTOVANI, W. 1999. A riqueza de espécies arbóreas na floresta atlântica de
encosta no estado de São Paulo (Brasil). Revista Brasileira de Botânica 22: 217-223.
TAMASHIRO, J.Y.; RODRIGUES, R.R. & SHEPHERD, G.J.1986. Estudo florístico e fitossociológico da Reserva da Mata de Santa Genebra, Campinas-SP. Relatório de
Pesquisa. Fundação de Amparo à Pesquisa de São Paulo.
TORRES, R.B., MARTINS, F.R. & KINOSHITA, L.S. 1997. Climate, soil and tree flora relationships in
forests in the state of São Paulo, southeastern Brasil. Revista Brasileira de Botânica 20: 41-51.
TURNER, I.M. 1996. Species loss in fragments of tropical rain forest: a review of evidence. Journal of Applied Ecology 33: 200-209.
UUTERA, J., TOKOLA, T. & MALTAMO, M. 2000. Diferences in the structure of primary and managed
forests in East Kalimantan, Indonesia. Forest Ecology and Management 129: 63-74.
VELOSO, H.P., RANGEL FILHO, A .L.R. & LIMA, J.C. 1991. Classificação da vegetação brasileira adaptada a um sistema universal. Rio de Janeiro. Instituto Brasileiro de Geografia e Estatística
(IBGE).
WEINER, J. & SOLBRIG, O.T. 1984. The meaning and measurement of size hierachies in plant
populations. Oecologia 61: 334-336.
WHITMORE, T.C. 1983. Secondary sucession from seed in tropical rain forest. Forestry Abstracts
44: 767-779.
WHITMORE, T.C. 1984. Tropical rain forest of the Far East. Clarendon Press, Oxford.
WILLIANS-LINERA, G. 1990. Vegetation structure and enviromental conditions of forest edges in
Panama. Journal of Ecology 78: 356-373.
WILLIANS-LINERA, G. 2002. Tree species richness complementarity, disturbance and fragmentation
in Mexican tropical montane cloud forest. Biodiversity and Conservation 11: 1825-1843.
ZAR, J.H. 1989. Bioestatistical Analysis. Prentice Hall, New Jersey.

151
ANEXO I
Tabelas contendo os parâmetros fitossociológicos analisados nos onze fragmentos estudados
no trabalho “Caracterização florística e estrutural de onze fragmentos de mata estacional
semidecidual da Área de Proteção Ambiental do município de Campinas – SP”.
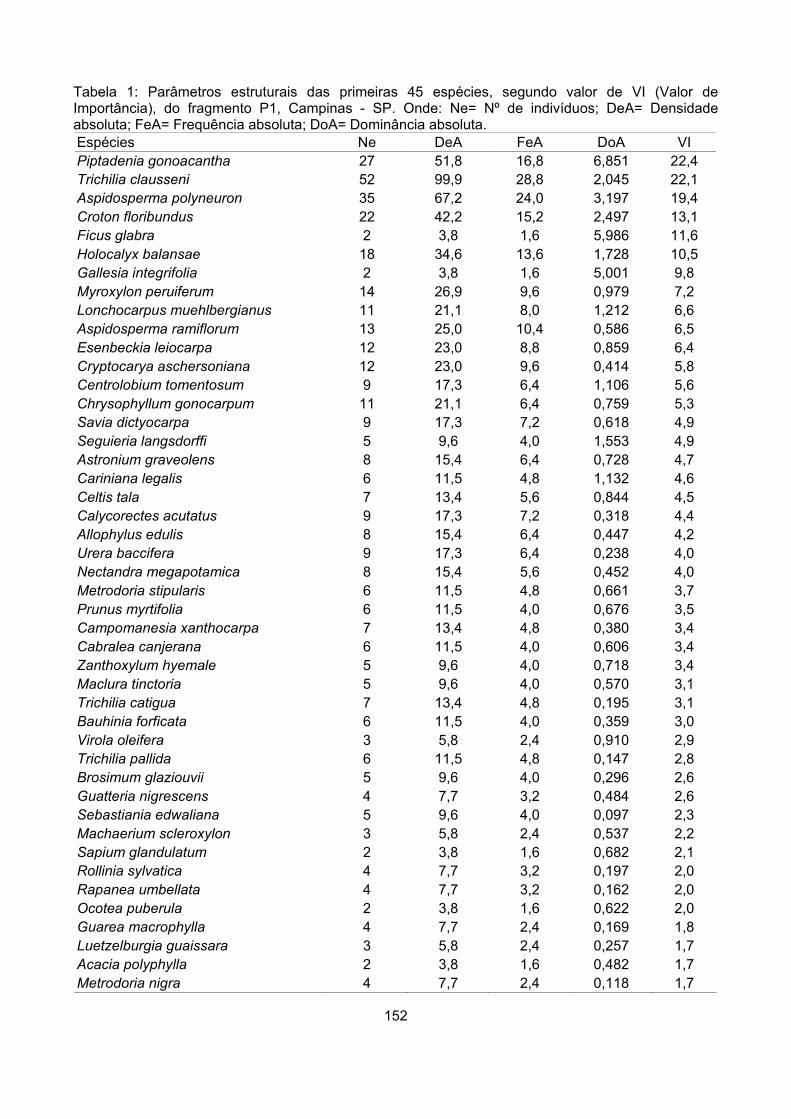
152
Tabela 1: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento P1, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Piptadenia gonoacantha 27 51,8 16,8 6,851 22,4 Trichilia clausseni 52 99,9 28,8 2,045 22,1 Aspidosperma polyneuron 35 67,2 24,0 3,197 19,4 Croton floribundus 22 42,2 15,2 2,497 13,1 Ficus glabra 2 3,8 1,6 5,986 11,6 Holocalyx balansae 18 34,6 13,6 1,728 10,5 Gallesia integrifolia 2 3,8 1,6 5,001 9,8 Myroxylon peruiferum 14 26,9 9,6 0,979 7,2 Lonchocarpus muehlbergianus 11 21,1 8,0 1,212 6,6 Aspidosperma ramiflorum 13 25,0 10,4 0,586 6,5 Esenbeckia leiocarpa 12 23,0 8,8 0,859 6,4 Cryptocarya aschersoniana 12 23,0 9,6 0,414 5,8 Centrolobium tomentosum 9 17,3 6,4 1,106 5,6 Chrysophyllum gonocarpum 11 21,1 6,4 0,759 5,3 Savia dictyocarpa 9 17,3 7,2 0,618 4,9 Seguieria langsdorffi 5 9,6 4,0 1,553 4,9 Astronium graveolens 8 15,4 6,4 0,728 4,7 Cariniana legalis 6 11,5 4,8 1,132 4,6 Celtis tala 7 13,4 5,6 0,844 4,5 Calycorectes acutatus 9 17,3 7,2 0,318 4,4 Allophylus edulis 8 15,4 6,4 0,447 4,2 Urera baccifera 9 17,3 6,4 0,238 4,0 Nectandra megapotamica 8 15,4 5,6 0,452 4,0 Metrodoria stipularis 6 11,5 4,8 0,661 3,7 Prunus myrtifolia 6 11,5 4,0 0,676 3,5 Campomanesia xanthocarpa 7 13,4 4,8 0,380 3,4 Cabralea canjerana 6 11,5 4,0 0,606 3,4 Zanthoxylum hyemale 5 9,6 4,0 0,718 3,4 Maclura tinctoria 5 9,6 4,0 0,570 3,1 Trichilia catigua 7 13,4 4,8 0,195 3,1 Bauhinia forficata 6 11,5 4,0 0,359 3,0 Virola oleifera 3 5,8 2,4 0,910 2,9 Trichilia pallida 6 11,5 4,8 0,147 2,8 Brosimum glaziouvii 5 9,6 4,0 0,296 2,6 Guatteria nigrescens 4 7,7 3,2 0,484 2,6 Sebastiania edwaliana 5 9,6 4,0 0,097 2,3 Machaerium scleroxylon 3 5,8 2,4 0,537 2,2 Sapium glandulatum 2 3,8 1,6 0,682 2,1 Rollinia sylvatica 4 7,7 3,2 0,197 2,0 Rapanea umbellata 4 7,7 3,2 0,162 2,0 Ocotea puberula 2 3,8 1,6 0,622 2,0 Guarea macrophylla 4 7,7 2,4 0,169 1,8 Luetzelburgia guaissara 3 5,8 2,4 0,257 1,7 Acacia polyphylla 2 3,8 1,6 0,482 1,7 Metrodoria nigra 4 7,7 2,4 0,118 1,7
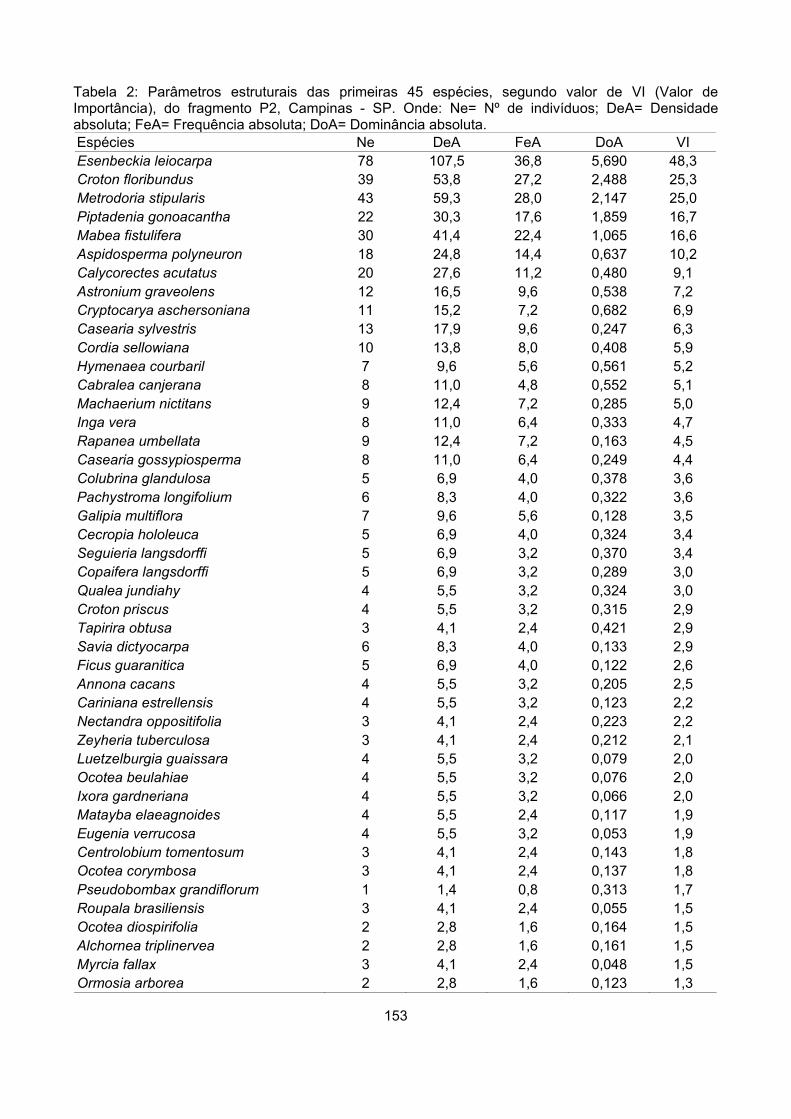
153
Tabela 2: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento P2, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 78 107,5 36,8 5,690 48,3 Croton floribundus 39 53,8 27,2 2,488 25,3 Metrodoria stipularis 43 59,3 28,0 2,147 25,0 Piptadenia gonoacantha 22 30,3 17,6 1,859 16,7 Mabea fistulifera 30 41,4 22,4 1,065 16,6 Aspidosperma polyneuron 18 24,8 14,4 0,637 10,2 Calycorectes acutatus 20 27,6 11,2 0,480 9,1 Astronium graveolens 12 16,5 9,6 0,538 7,2 Cryptocarya aschersoniana 11 15,2 7,2 0,682 6,9 Casearia sylvestris 13 17,9 9,6 0,247 6,3 Cordia sellowiana 10 13,8 8,0 0,408 5,9 Hymenaea courbaril 7 9,6 5,6 0,561 5,2 Cabralea canjerana 8 11,0 4,8 0,552 5,1 Machaerium nictitans 9 12,4 7,2 0,285 5,0 Inga vera 8 11,0 6,4 0,333 4,7 Rapanea umbellata 9 12,4 7,2 0,163 4,5 Casearia gossypiosperma 8 11,0 6,4 0,249 4,4 Colubrina glandulosa 5 6,9 4,0 0,378 3,6 Pachystroma longifolium 6 8,3 4,0 0,322 3,6 Galipia multiflora 7 9,6 5,6 0,128 3,5 Cecropia hololeuca 5 6,9 4,0 0,324 3,4 Seguieria langsdorffi 5 6,9 3,2 0,370 3,4 Copaifera langsdorffi 5 6,9 3,2 0,289 3,0 Qualea jundiahy 4 5,5 3,2 0,324 3,0 Croton priscus 4 5,5 3,2 0,315 2,9 Tapirira obtusa 3 4,1 2,4 0,421 2,9 Savia dictyocarpa 6 8,3 4,0 0,133 2,9 Ficus guaranitica 5 6,9 4,0 0,122 2,6 Annona cacans 4 5,5 3,2 0,205 2,5 Cariniana estrellensis 4 5,5 3,2 0,123 2,2 Nectandra oppositifolia 3 4,1 2,4 0,223 2,2 Zeyheria tuberculosa 3 4,1 2,4 0,212 2,1 Luetzelburgia guaissara 4 5,5 3,2 0,079 2,0 Ocotea beulahiae 4 5,5 3,2 0,076 2,0 Ixora gardneriana 4 5,5 3,2 0,066 2,0 Matayba elaeagnoides 4 5,5 2,4 0,117 1,9 Eugenia verrucosa 4 5,5 3,2 0,053 1,9 Centrolobium tomentosum 3 4,1 2,4 0,143 1,8 Ocotea corymbosa 3 4,1 2,4 0,137 1,8 Pseudobombax grandiflorum 1 1,4 0,8 0,313 1,7 Roupala brasiliensis 3 4,1 2,4 0,055 1,5 Ocotea diospirifolia 2 2,8 1,6 0,164 1,5 Alchornea triplinervea 2 2,8 1,6 0,161 1,5 Myrcia fallax 3 4,1 2,4 0,048 1,5 Ormosia arborea 2 2,8 1,6 0,123 1,3

154
Tabela 3: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento P3, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Trichilia clausseni 229 417,8 88,0 8,921 99,9 Chorisia speciosa 21 38,3 16,0 11,940 39,5 Lonchocarpus muehlbergianus 54 98,5 29,6 5,477 35,1 Maclura tinctoria 16 29,2 12,8 1,515 11,6 Astronium graveolens 16 29,2 12,8 0,945 10,2 Trichilia hirta 17 31,0 12,8 0,618 9,6 Trichilia catigua 19 34,7 9,6 0,584 8,7 Casearia sylvestris 16 29,2 11,2 0,378 8,2 Centrolobium tomentosum 12 21,9 9,6 0,682 7,6 Myroxylon peruiferum 10 18,2 7,2 0,753 6,5 Diospyrus inconstans 11 20,1 5,6 0,429 5,3 Urera baccifera 9 16,4 7,2 0,336 5,3 Pseudobombax grandiflorum 6 10,9 4,8 0,768 4,8 Hymenaea courbaril 1 1,8 0,8 1,487 4,2 Cariniana estrellensis 7 12,8 4,8 0,407 4,2 Rollinia sylvatica 6 10,9 4,8 0,240 3,5 Gallesia integrifolia 2 3,6 1,6 0,843 3,1 Casearia gossypiosperma 3 5,5 2,4 0,566 2,9 Luehea divaricata 3 5,5 2,4 0,388 2,4 Holocalyx balansae 3 5,5 2,4 0,388 2,4 Seguieria langsdorffi 2 3,6 0,8 0,625 2,2 Sweetia fruticosa 3 5,5 2,4 0,159 1,9 Guapira opposita 4 7,3 2,4 0,069 1,8 Cordia trichotoma 2 3,6 1,6 0,166 1,4 Cariniana legalis 1 1,8 0,8 0,265 1,1 Pithecellobium edwalii 2 3,6 1,6 0,059 1,1 Campomanesia neriiflora 2 3,6 0,8 0,162 1,1 Allophylus edulis 2 3,6 1,6 0,033 1,1 Machaerium scleroxylon 2 3,6 1,6 0,028 1,1 Bastardiopsis densiflora 1 1,8 0,8 0,220 1,0 Guazuma ulmifolia 1 1,8 0,8 0,182 0,9 Balfourodendron riedelianum 1 1,8 0,8 0,142 0,8 Peltophorum dubium 1 1,8 0,8 0,139 0,8 Campomanesia guazumifolia 2 3,6 0,8 0,058 0,8 Ficus guaranitica 1 1,8 0,8 0,137 0,8 Zanthoxylum hyemale 1 1,8 0,8 0,128 0,8 Psidium sartorianum 1 1,8 0,8 0,056 0,6 Copaifera langsdorffi 1 1,8 0,8 0,052 0,6 Machaerium nictitans 1 1,8 0,8 0,036 0,6 Aloysia virgata 1 1,8 0,8 0,034 0,6 Coutarea hexandra 1 1,8 0,8 0,032 0,6 Chrysophyllum gonocarpum 1 1,8 0,8 0,028 0,6 Piptadenia gonoacantha 1 1,8 0,8 0,024 0,6 Aspidosperma ramiflorum 1 1,8 0,8 0,023 0,6 Cordia ecalyculata 1 1,8 0,8 0,022 0,6
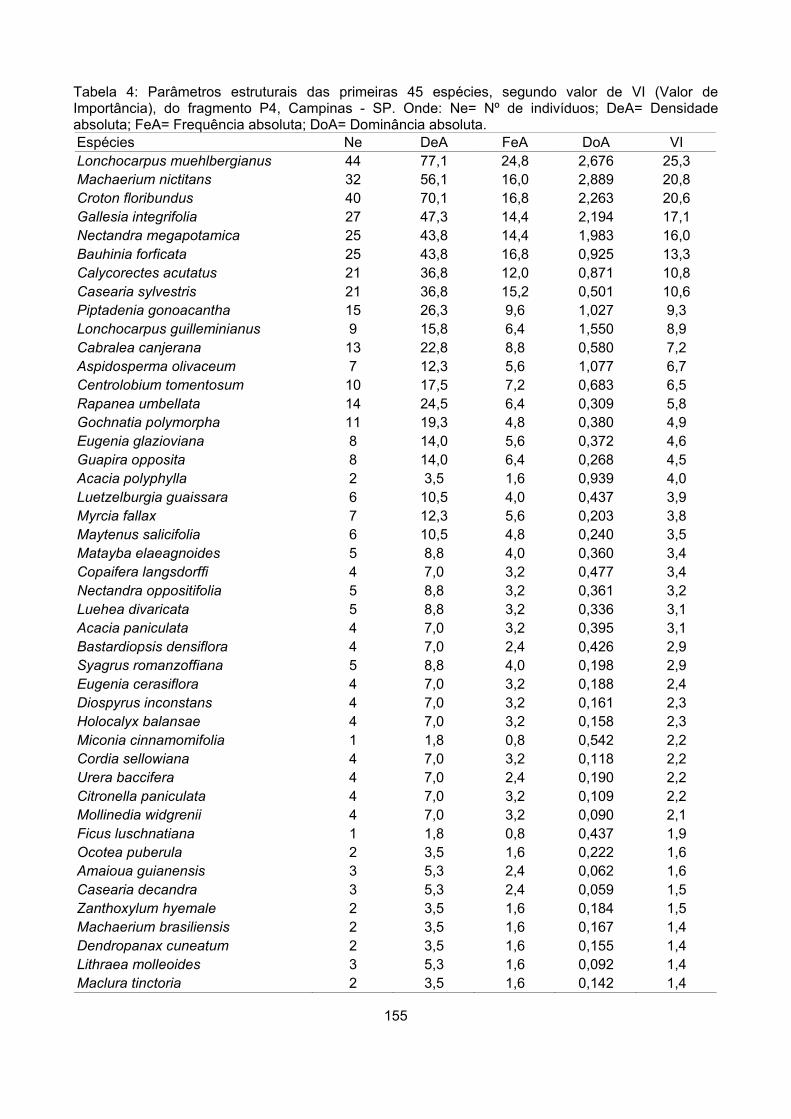
155
Tabela 4: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento P4, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Lonchocarpus muehlbergianus 44 77,1 24,8 2,676 25,3 Machaerium nictitans 32 56,1 16,0 2,889 20,8 Croton floribundus 40 70,1 16,8 2,263 20,6 Gallesia integrifolia 27 47,3 14,4 2,194 17,1 Nectandra megapotamica 25 43,8 14,4 1,983 16,0 Bauhinia forficata 25 43,8 16,8 0,925 13,3 Calycorectes acutatus 21 36,8 12,0 0,871 10,8 Casearia sylvestris 21 36,8 15,2 0,501 10,6 Piptadenia gonoacantha 15 26,3 9,6 1,027 9,3 Lonchocarpus guilleminianus 9 15,8 6,4 1,550 8,9 Cabralea canjerana 13 22,8 8,8 0,580 7,2 Aspidosperma olivaceum 7 12,3 5,6 1,077 6,7 Centrolobium tomentosum 10 17,5 7,2 0,683 6,5 Rapanea umbellata 14 24,5 6,4 0,309 5,8 Gochnatia polymorpha 11 19,3 4,8 0,380 4,9 Eugenia glazioviana 8 14,0 5,6 0,372 4,6 Guapira opposita 8 14,0 6,4 0,268 4,5 Acacia polyphylla 2 3,5 1,6 0,939 4,0 Luetzelburgia guaissara 6 10,5 4,0 0,437 3,9 Myrcia fallax 7 12,3 5,6 0,203 3,8 Maytenus salicifolia 6 10,5 4,8 0,240 3,5 Matayba elaeagnoides 5 8,8 4,0 0,360 3,4 Copaifera langsdorffi 4 7,0 3,2 0,477 3,4 Nectandra oppositifolia 5 8,8 3,2 0,361 3,2 Luehea divaricata 5 8,8 3,2 0,336 3,1 Acacia paniculata 4 7,0 3,2 0,395 3,1 Bastardiopsis densiflora 4 7,0 2,4 0,426 2,9 Syagrus romanzoffiana 5 8,8 4,0 0,198 2,9 Eugenia cerasiflora 4 7,0 3,2 0,188 2,4 Diospyrus inconstans 4 7,0 3,2 0,161 2,3 Holocalyx balansae 4 7,0 3,2 0,158 2,3 Miconia cinnamomifolia 1 1,8 0,8 0,542 2,2 Cordia sellowiana 4 7,0 3,2 0,118 2,2 Urera baccifera 4 7,0 2,4 0,190 2,2 Citronella paniculata 4 7,0 3,2 0,109 2,2 Mollinedia widgrenii 4 7,0 3,2 0,090 2,1 Ficus luschnatiana 1 1,8 0,8 0,437 1,9 Ocotea puberula 2 3,5 1,6 0,222 1,6 Amaioua guianensis 3 5,3 2,4 0,062 1,6 Casearia decandra 3 5,3 2,4 0,059 1,5 Zanthoxylum hyemale 2 3,5 1,6 0,184 1,5 Machaerium brasiliensis 2 3,5 1,6 0,167 1,4 Dendropanax cuneatum 2 3,5 1,6 0,155 1,4 Lithraea molleoides 3 5,3 1,6 0,092 1,4 Maclura tinctoria 2 3,5 1,6 0,142 1,4

156
Tabela 5: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento M1, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Croton floribundus 52 70,9 32,0 3,452 30,6 Aspidosperma polyneuron 34 46,3 20,0 1,974 18,8 Piptadenia gonoacantha 22 30,0 15,2 2,988 18,4 Urera baccifera 26 35,4 15,2 1,139 13,2 Esenbeckia leiocarpa 20 27,3 13,6 0,799 10,4 Metrodoria nigra 24 32,7 14,4 0,413 10,2 Cariniana legalis 6 8,2 4,8 2,270 9,9 Bauhinia forficata 17 23,2 10,4 0,873 9,2 Jacaranda micrantha 12 16,4 8,8 1,125 8,5 Seguieria langsdorffi 12 16,4 9,6 0,727 7,5 Centrolobium tomentosum 13 17,7 8,8 0,537 6,8 Copaifera langsdorffi 3 4,1 2,4 1,484 6,1 Chorisia speciosa 10 13,6 8,0 0,444 5,7 Celtis tala 7 9,5 4,8 0,749 5,2 Acacia paniculata 7 9,5 5,6 0,557 4,8 Nectandra megapotamica 9 12,3 7,2 0,281 4,7 Machaerium nictitans 8 10,9 4,8 0,535 4,7 Brosimum glaziouvii 8 10,9 6,4 0,315 4,4 Cabralea canjerana 6 8,2 4,8 0,520 4,2 Cordia ecalyculata 6 8,2 4,8 0,507 4,2 Ficus glabra 3 4,1 2,4 0,829 4,0 Guapira opposita 8 10,9 6,4 0,168 3,9 Zanthoxylum hyemale 6 8,2 4,8 0,429 3,9 Calycorectes acutatus 8 10,9 5,6 0,220 3,9 Rollinia sylvatica 6 8,2 4,8 0,377 3,8 Simira sampaioana 6 8,2 4,8 0,302 3,5 Aspidosperma olivaceum 4 5,5 3,2 0,539 3,5 Nectandra oppositifolia 5 6,8 4,0 0,375 3,3 Lonchocarpus muehlbergianus 6 8,2 4,8 0,170 3,1 Maclura tinctoria 6 8,2 3,2 0,282 3,0 Cordia sellowiana 5 6,8 4,0 0,230 2,9 Cariniana estrellensis 6 8,2 4,0 0,143 2,8 Eugenia glazioviana 5 6,8 4,0 0,199 2,8 Savia dictyocarpa 6 8,2 4,0 0,103 2,7 Eugenia verrucosa 5 6,8 4,0 0,098 2,4 Pseudobombax grandiflorum 4 5,5 3,2 0,197 2,3 Machaerium scleroxylon 1 1,4 0,8 0,554 2,2 Cecropia hololeuca 4 5,5 3,2 0,155 2,2 Matayba elaeagnoides 4 5,5 3,2 0,143 2,2 Holocalyx balansae 4 5,5 3,2 0,136 2,1 Ficus trigona 1 1,4 0,8 0,516 2,1 Ocotea diospirifolia 4 5,5 3,2 0,114 2,1 Aspidosperma ramiflorum 4 5,5 3,2 0,105 2,0 Pisonia ambigua 3 4,1 2,4 0,199 1,9 Trichilia clausseni 4 5,5 3,2 0,067 1,9
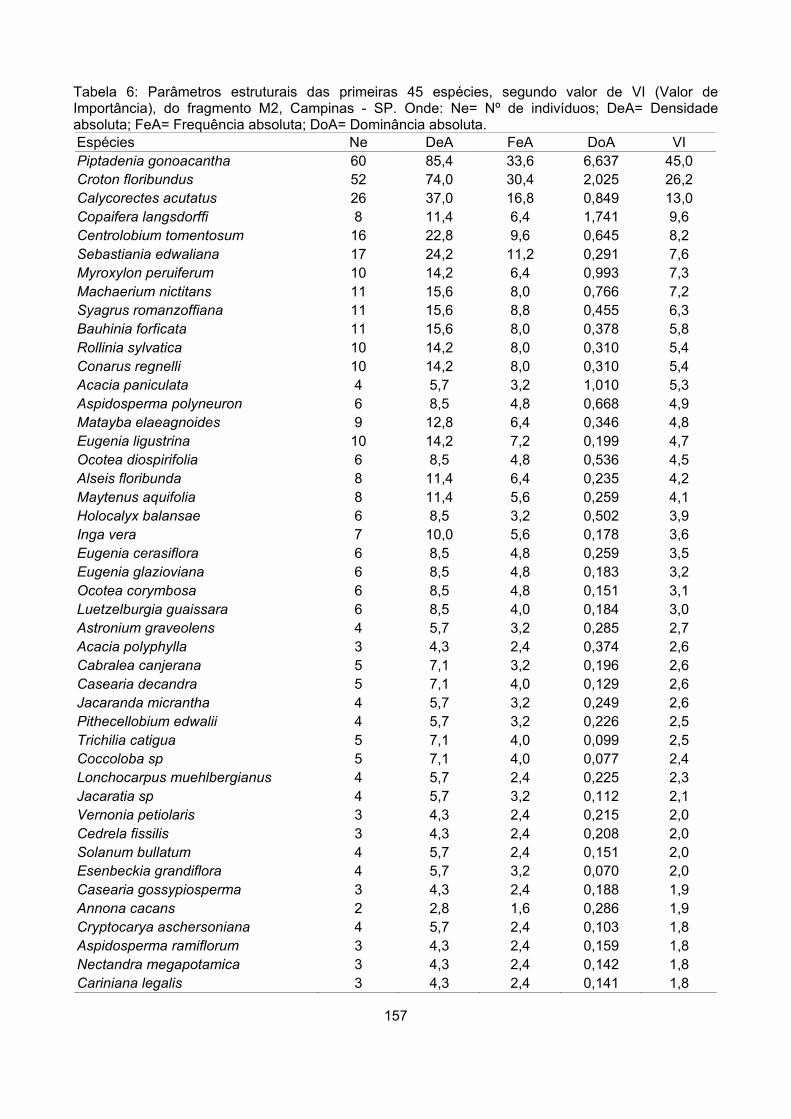
157
Tabela 6: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento M2, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Piptadenia gonoacantha 60 85,4 33,6 6,637 45,0 Croton floribundus 52 74,0 30,4 2,025 26,2 Calycorectes acutatus 26 37,0 16,8 0,849 13,0 Copaifera langsdorffi 8 11,4 6,4 1,741 9,6 Centrolobium tomentosum 16 22,8 9,6 0,645 8,2 Sebastiania edwaliana 17 24,2 11,2 0,291 7,6 Myroxylon peruiferum 10 14,2 6,4 0,993 7,3 Machaerium nictitans 11 15,6 8,0 0,766 7,2 Syagrus romanzoffiana 11 15,6 8,8 0,455 6,3 Bauhinia forficata 11 15,6 8,0 0,378 5,8 Rollinia sylvatica 10 14,2 8,0 0,310 5,4 Conarus regnelli 10 14,2 8,0 0,310 5,4 Acacia paniculata 4 5,7 3,2 1,010 5,3 Aspidosperma polyneuron 6 8,5 4,8 0,668 4,9 Matayba elaeagnoides 9 12,8 6,4 0,346 4,8 Eugenia ligustrina 10 14,2 7,2 0,199 4,7 Ocotea diospirifolia 6 8,5 4,8 0,536 4,5 Alseis floribunda 8 11,4 6,4 0,235 4,2 Maytenus aquifolia 8 11,4 5,6 0,259 4,1 Holocalyx balansae 6 8,5 3,2 0,502 3,9 Inga vera 7 10,0 5,6 0,178 3,6 Eugenia cerasiflora 6 8,5 4,8 0,259 3,5 Eugenia glazioviana 6 8,5 4,8 0,183 3,2 Ocotea corymbosa 6 8,5 4,8 0,151 3,1 Luetzelburgia guaissara 6 8,5 4,0 0,184 3,0 Astronium graveolens 4 5,7 3,2 0,285 2,7 Acacia polyphylla 3 4,3 2,4 0,374 2,6 Cabralea canjerana 5 7,1 3,2 0,196 2,6 Casearia decandra 5 7,1 4,0 0,129 2,6 Jacaranda micrantha 4 5,7 3,2 0,249 2,6 Pithecellobium edwalii 4 5,7 3,2 0,226 2,5 Trichilia catigua 5 7,1 4,0 0,099 2,5 Coccoloba sp 5 7,1 4,0 0,077 2,4 Lonchocarpus muehlbergianus 4 5,7 2,4 0,225 2,3 Jacaratia sp 4 5,7 3,2 0,112 2,1 Vernonia petiolaris 3 4,3 2,4 0,215 2,0 Cedrela fissilis 3 4,3 2,4 0,208 2,0 Solanum bullatum 4 5,7 2,4 0,151 2,0 Esenbeckia grandiflora 4 5,7 3,2 0,070 2,0 Casearia gossypiosperma 3 4,3 2,4 0,188 1,9 Annona cacans 2 2,8 1,6 0,286 1,9 Cryptocarya aschersoniana 4 5,7 2,4 0,103 1,8 Aspidosperma ramiflorum 3 4,3 2,4 0,159 1,8 Nectandra megapotamica 3 4,3 2,4 0,142 1,8 Cariniana legalis 3 4,3 2,4 0,141 1,8
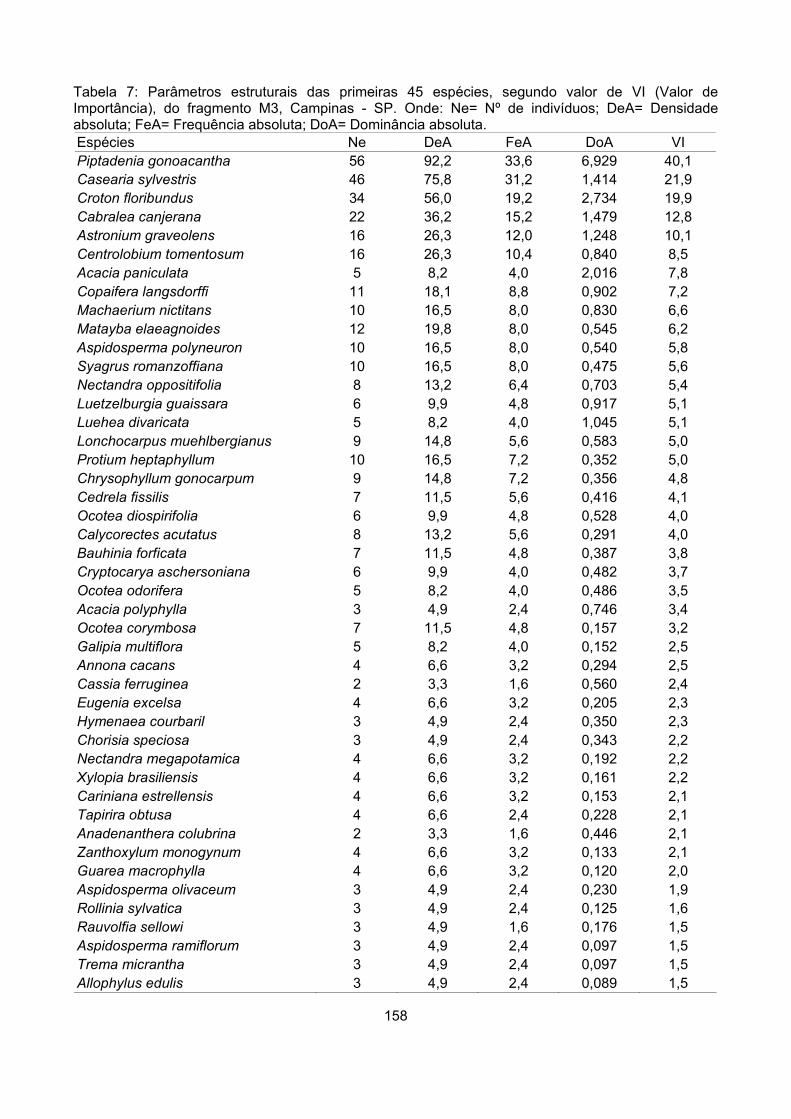
158
Tabela 7: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento M3, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Piptadenia gonoacantha 56 92,2 33,6 6,929 40,1 Casearia sylvestris 46 75,8 31,2 1,414 21,9 Croton floribundus 34 56,0 19,2 2,734 19,9 Cabralea canjerana 22 36,2 15,2 1,479 12,8 Astronium graveolens 16 26,3 12,0 1,248 10,1 Centrolobium tomentosum 16 26,3 10,4 0,840 8,5 Acacia paniculata 5 8,2 4,0 2,016 7,8 Copaifera langsdorffi 11 18,1 8,8 0,902 7,2 Machaerium nictitans 10 16,5 8,0 0,830 6,6 Matayba elaeagnoides 12 19,8 8,0 0,545 6,2 Aspidosperma polyneuron 10 16,5 8,0 0,540 5,8 Syagrus romanzoffiana 10 16,5 8,0 0,475 5,6 Nectandra oppositifolia 8 13,2 6,4 0,703 5,4 Luetzelburgia guaissara 6 9,9 4,8 0,917 5,1 Luehea divaricata 5 8,2 4,0 1,045 5,1 Lonchocarpus muehlbergianus 9 14,8 5,6 0,583 5,0 Protium heptaphyllum 10 16,5 7,2 0,352 5,0 Chrysophyllum gonocarpum 9 14,8 7,2 0,356 4,8 Cedrela fissilis 7 11,5 5,6 0,416 4,1 Ocotea diospirifolia 6 9,9 4,8 0,528 4,0 Calycorectes acutatus 8 13,2 5,6 0,291 4,0 Bauhinia forficata 7 11,5 4,8 0,387 3,8 Cryptocarya aschersoniana 6 9,9 4,0 0,482 3,7 Ocotea odorifera 5 8,2 4,0 0,486 3,5 Acacia polyphylla 3 4,9 2,4 0,746 3,4 Ocotea corymbosa 7 11,5 4,8 0,157 3,2 Galipia multiflora 5 8,2 4,0 0,152 2,5 Annona cacans 4 6,6 3,2 0,294 2,5 Cassia ferruginea 2 3,3 1,6 0,560 2,4 Eugenia excelsa 4 6,6 3,2 0,205 2,3 Hymenaea courbaril 3 4,9 2,4 0,350 2,3 Chorisia speciosa 3 4,9 2,4 0,343 2,2 Nectandra megapotamica 4 6,6 3,2 0,192 2,2 Xylopia brasiliensis 4 6,6 3,2 0,161 2,2 Cariniana estrellensis 4 6,6 3,2 0,153 2,1 Tapirira obtusa 4 6,6 2,4 0,228 2,1 Anadenanthera colubrina 2 3,3 1,6 0,446 2,1 Zanthoxylum monogynum 4 6,6 3,2 0,133 2,1 Guarea macrophylla 4 6,6 3,2 0,120 2,0 Aspidosperma olivaceum 3 4,9 2,4 0,230 1,9 Rollinia sylvatica 3 4,9 2,4 0,125 1,6 Rauvolfia sellowi 3 4,9 1,6 0,176 1,5 Aspidosperma ramiflorum 3 4,9 2,4 0,097 1,5 Trema micrantha 3 4,9 2,4 0,097 1,5 Allophylus edulis 3 4,9 2,4 0,089 1,5
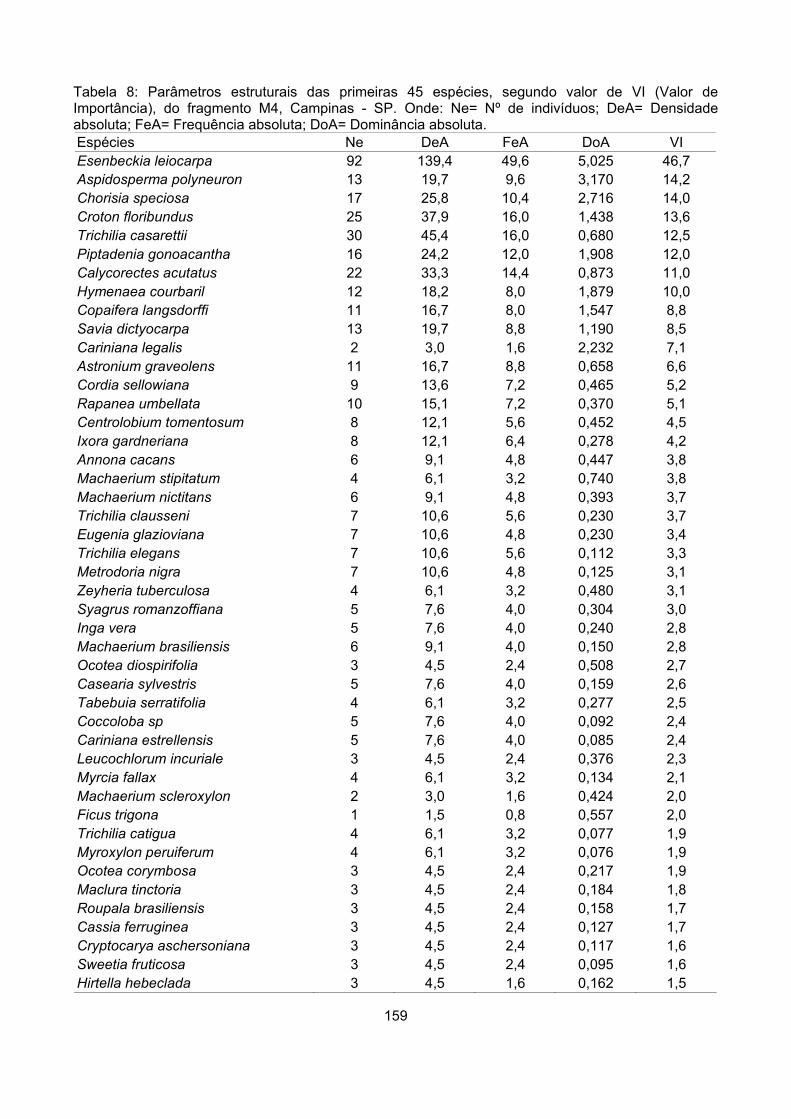
159
Tabela 8: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento M4, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 92 139,4 49,6 5,025 46,7 Aspidosperma polyneuron 13 19,7 9,6 3,170 14,2 Chorisia speciosa 17 25,8 10,4 2,716 14,0 Croton floribundus 25 37,9 16,0 1,438 13,6 Trichilia casarettii 30 45,4 16,0 0,680 12,5 Piptadenia gonoacantha 16 24,2 12,0 1,908 12,0 Calycorectes acutatus 22 33,3 14,4 0,873 11,0 Hymenaea courbaril 12 18,2 8,0 1,879 10,0 Copaifera langsdorffi 11 16,7 8,0 1,547 8,8 Savia dictyocarpa 13 19,7 8,8 1,190 8,5 Cariniana legalis 2 3,0 1,6 2,232 7,1 Astronium graveolens 11 16,7 8,8 0,658 6,6 Cordia sellowiana 9 13,6 7,2 0,465 5,2 Rapanea umbellata 10 15,1 7,2 0,370 5,1 Centrolobium tomentosum 8 12,1 5,6 0,452 4,5 Ixora gardneriana 8 12,1 6,4 0,278 4,2 Annona cacans 6 9,1 4,8 0,447 3,8 Machaerium stipitatum 4 6,1 3,2 0,740 3,8 Machaerium nictitans 6 9,1 4,8 0,393 3,7 Trichilia clausseni 7 10,6 5,6 0,230 3,7 Eugenia glazioviana 7 10,6 4,8 0,230 3,4 Trichilia elegans 7 10,6 5,6 0,112 3,3 Metrodoria nigra 7 10,6 4,8 0,125 3,1 Zeyheria tuberculosa 4 6,1 3,2 0,480 3,1 Syagrus romanzoffiana 5 7,6 4,0 0,304 3,0 Inga vera 5 7,6 4,0 0,240 2,8 Machaerium brasiliensis 6 9,1 4,0 0,150 2,8 Ocotea diospirifolia 3 4,5 2,4 0,508 2,7 Casearia sylvestris 5 7,6 4,0 0,159 2,6 Tabebuia serratifolia 4 6,1 3,2 0,277 2,5 Coccoloba sp 5 7,6 4,0 0,092 2,4 Cariniana estrellensis 5 7,6 4,0 0,085 2,4 Leucochlorum incuriale 3 4,5 2,4 0,376 2,3 Myrcia fallax 4 6,1 3,2 0,134 2,1 Machaerium scleroxylon 2 3,0 1,6 0,424 2,0 Ficus trigona 1 1,5 0,8 0,557 2,0 Trichilia catigua 4 6,1 3,2 0,077 1,9 Myroxylon peruiferum 4 6,1 3,2 0,076 1,9 Ocotea corymbosa 3 4,5 2,4 0,217 1,9 Maclura tinctoria 3 4,5 2,4 0,184 1,8 Roupala brasiliensis 3 4,5 2,4 0,158 1,7 Cassia ferruginea 3 4,5 2,4 0,127 1,7 Cryptocarya aschersoniana 3 4,5 2,4 0,117 1,6 Sweetia fruticosa 3 4,5 2,4 0,095 1,6 Hirtella hebeclada 3 4,5 1,6 0,162 1,5
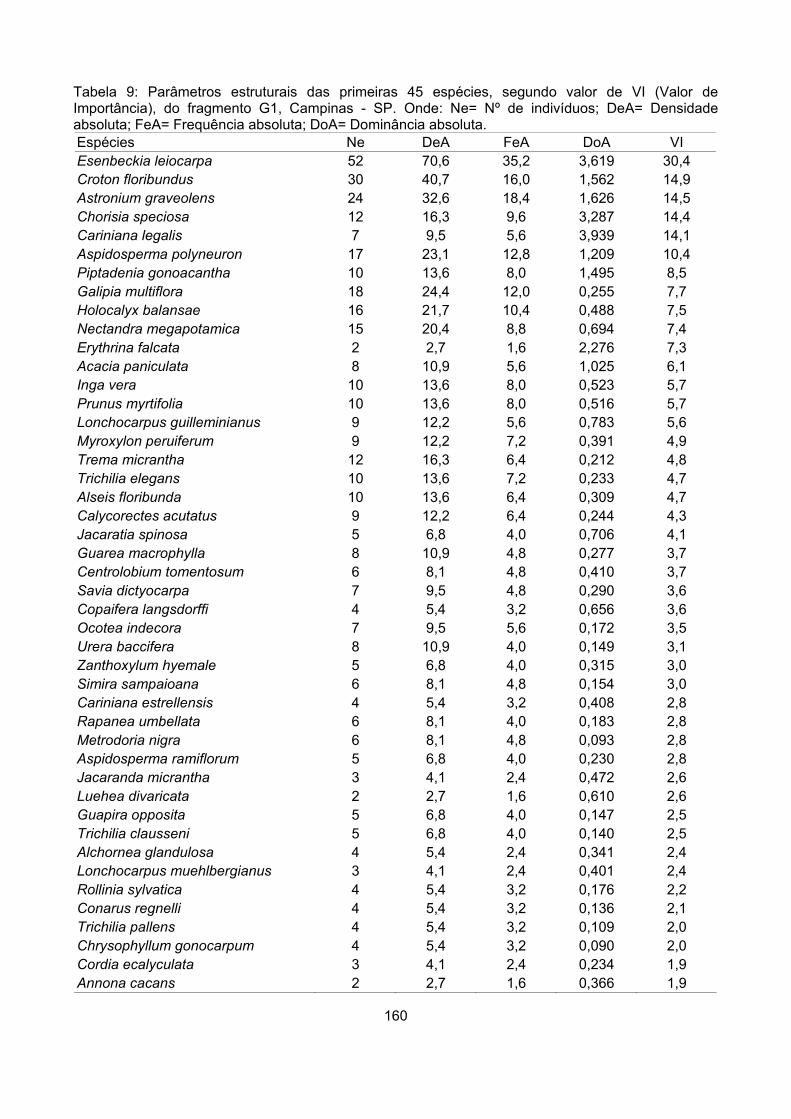
160
Tabela 9: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento G1, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 52 70,6 35,2 3,619 30,4 Croton floribundus 30 40,7 16,0 1,562 14,9 Astronium graveolens 24 32,6 18,4 1,626 14,5 Chorisia speciosa 12 16,3 9,6 3,287 14,4 Cariniana legalis 7 9,5 5,6 3,939 14,1 Aspidosperma polyneuron 17 23,1 12,8 1,209 10,4 Piptadenia gonoacantha 10 13,6 8,0 1,495 8,5 Galipia multiflora 18 24,4 12,0 0,255 7,7 Holocalyx balansae 16 21,7 10,4 0,488 7,5 Nectandra megapotamica 15 20,4 8,8 0,694 7,4 Erythrina falcata 2 2,7 1,6 2,276 7,3 Acacia paniculata 8 10,9 5,6 1,025 6,1 Inga vera 10 13,6 8,0 0,523 5,7 Prunus myrtifolia 10 13,6 8,0 0,516 5,7 Lonchocarpus guilleminianus 9 12,2 5,6 0,783 5,6 Myroxylon peruiferum 9 12,2 7,2 0,391 4,9 Trema micrantha 12 16,3 6,4 0,212 4,8 Trichilia elegans 10 13,6 7,2 0,233 4,7 Alseis floribunda 10 13,6 6,4 0,309 4,7 Calycorectes acutatus 9 12,2 6,4 0,244 4,3 Jacaratia spinosa 5 6,8 4,0 0,706 4,1 Guarea macrophylla 8 10,9 4,8 0,277 3,7 Centrolobium tomentosum 6 8,1 4,8 0,410 3,7 Savia dictyocarpa 7 9,5 4,8 0,290 3,6 Copaifera langsdorffi 4 5,4 3,2 0,656 3,6 Ocotea indecora 7 9,5 5,6 0,172 3,5 Urera baccifera 8 10,9 4,0 0,149 3,1 Zanthoxylum hyemale 5 6,8 4,0 0,315 3,0 Simira sampaioana 6 8,1 4,8 0,154 3,0 Cariniana estrellensis 4 5,4 3,2 0,408 2,8 Rapanea umbellata 6 8,1 4,0 0,183 2,8 Metrodoria nigra 6 8,1 4,8 0,093 2,8 Aspidosperma ramiflorum 5 6,8 4,0 0,230 2,8 Jacaranda micrantha 3 4,1 2,4 0,472 2,6 Luehea divaricata 2 2,7 1,6 0,610 2,6 Guapira opposita 5 6,8 4,0 0,147 2,5 Trichilia clausseni 5 6,8 4,0 0,140 2,5 Alchornea glandulosa 4 5,4 2,4 0,341 2,4 Lonchocarpus muehlbergianus 3 4,1 2,4 0,401 2,4 Rollinia sylvatica 4 5,4 3,2 0,176 2,2 Conarus regnelli 4 5,4 3,2 0,136 2,1 Trichilia pallens 4 5,4 3,2 0,109 2,0 Chrysophyllum gonocarpum 4 5,4 3,2 0,090 2,0 Cordia ecalyculata 3 4,1 2,4 0,234 1,9 Annona cacans 2 2,7 1,6 0,366 1,9
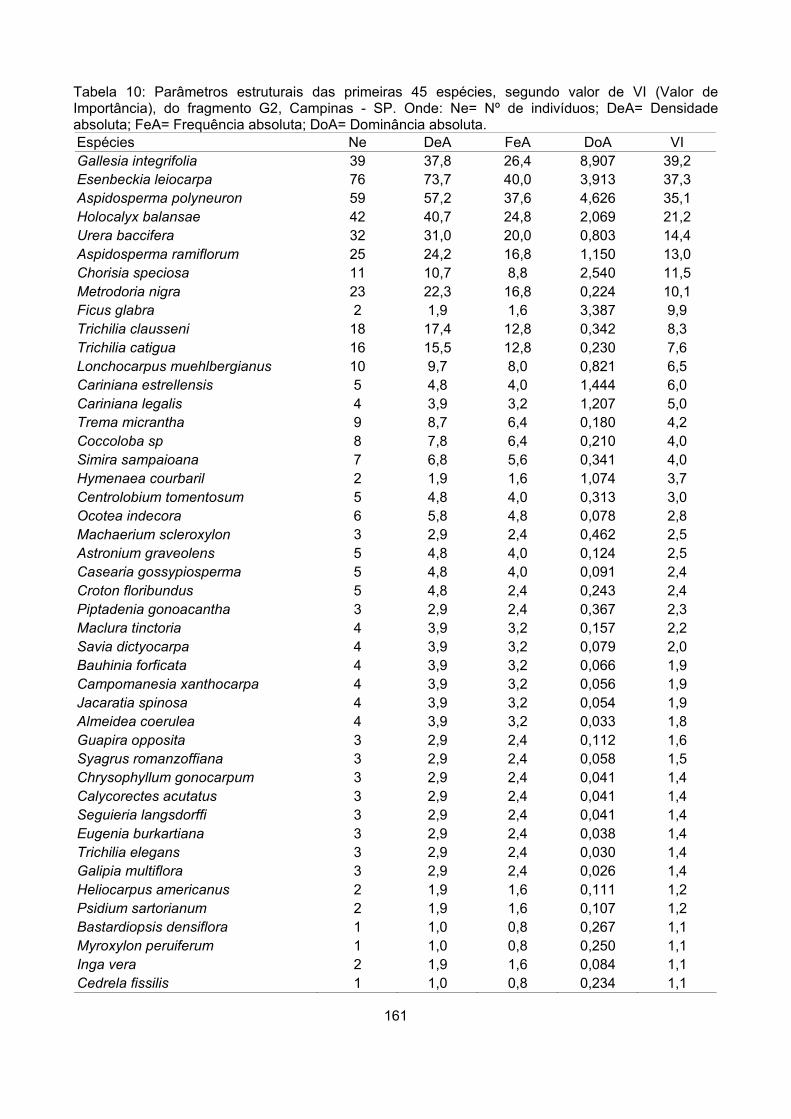
161
Tabela 10: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), do fragmento G2, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Gallesia integrifolia 39 37,8 26,4 8,907 39,2 Esenbeckia leiocarpa 76 73,7 40,0 3,913 37,3 Aspidosperma polyneuron 59 57,2 37,6 4,626 35,1 Holocalyx balansae 42 40,7 24,8 2,069 21,2 Urera baccifera 32 31,0 20,0 0,803 14,4 Aspidosperma ramiflorum 25 24,2 16,8 1,150 13,0 Chorisia speciosa 11 10,7 8,8 2,540 11,5 Metrodoria nigra 23 22,3 16,8 0,224 10,1 Ficus glabra 2 1,9 1,6 3,387 9,9 Trichilia clausseni 18 17,4 12,8 0,342 8,3 Trichilia catigua 16 15,5 12,8 0,230 7,6 Lonchocarpus muehlbergianus 10 9,7 8,0 0,821 6,5 Cariniana estrellensis 5 4,8 4,0 1,444 6,0 Cariniana legalis 4 3,9 3,2 1,207 5,0 Trema micrantha 9 8,7 6,4 0,180 4,2 Coccoloba sp 8 7,8 6,4 0,210 4,0 Simira sampaioana 7 6,8 5,6 0,341 4,0 Hymenaea courbaril 2 1,9 1,6 1,074 3,7 Centrolobium tomentosum 5 4,8 4,0 0,313 3,0 Ocotea indecora 6 5,8 4,8 0,078 2,8 Machaerium scleroxylon 3 2,9 2,4 0,462 2,5 Astronium graveolens 5 4,8 4,0 0,124 2,5 Casearia gossypiosperma 5 4,8 4,0 0,091 2,4 Croton floribundus 5 4,8 2,4 0,243 2,4 Piptadenia gonoacantha 3 2,9 2,4 0,367 2,3 Maclura tinctoria 4 3,9 3,2 0,157 2,2 Savia dictyocarpa 4 3,9 3,2 0,079 2,0 Bauhinia forficata 4 3,9 3,2 0,066 1,9 Campomanesia xanthocarpa 4 3,9 3,2 0,056 1,9 Jacaratia spinosa 4 3,9 3,2 0,054 1,9 Almeidea coerulea 4 3,9 3,2 0,033 1,8 Guapira opposita 3 2,9 2,4 0,112 1,6 Syagrus romanzoffiana 3 2,9 2,4 0,058 1,5 Chrysophyllum gonocarpum 3 2,9 2,4 0,041 1,4 Calycorectes acutatus 3 2,9 2,4 0,041 1,4 Seguieria langsdorffi 3 2,9 2,4 0,041 1,4 Eugenia burkartiana 3 2,9 2,4 0,038 1,4 Trichilia elegans 3 2,9 2,4 0,030 1,4 Galipia multiflora 3 2,9 2,4 0,026 1,4 Heliocarpus americanus 2 1,9 1,6 0,111 1,2 Psidium sartorianum 2 1,9 1,6 0,107 1,2 Bastardiopsis densiflora 1 1,0 0,8 0,267 1,1 Myroxylon peruiferum 1 1,0 0,8 0,250 1,1 Inga vera 2 1,9 1,6 0,084 1,1 Cedrela fissilis 1 1,0 0,8 0,234 1,1
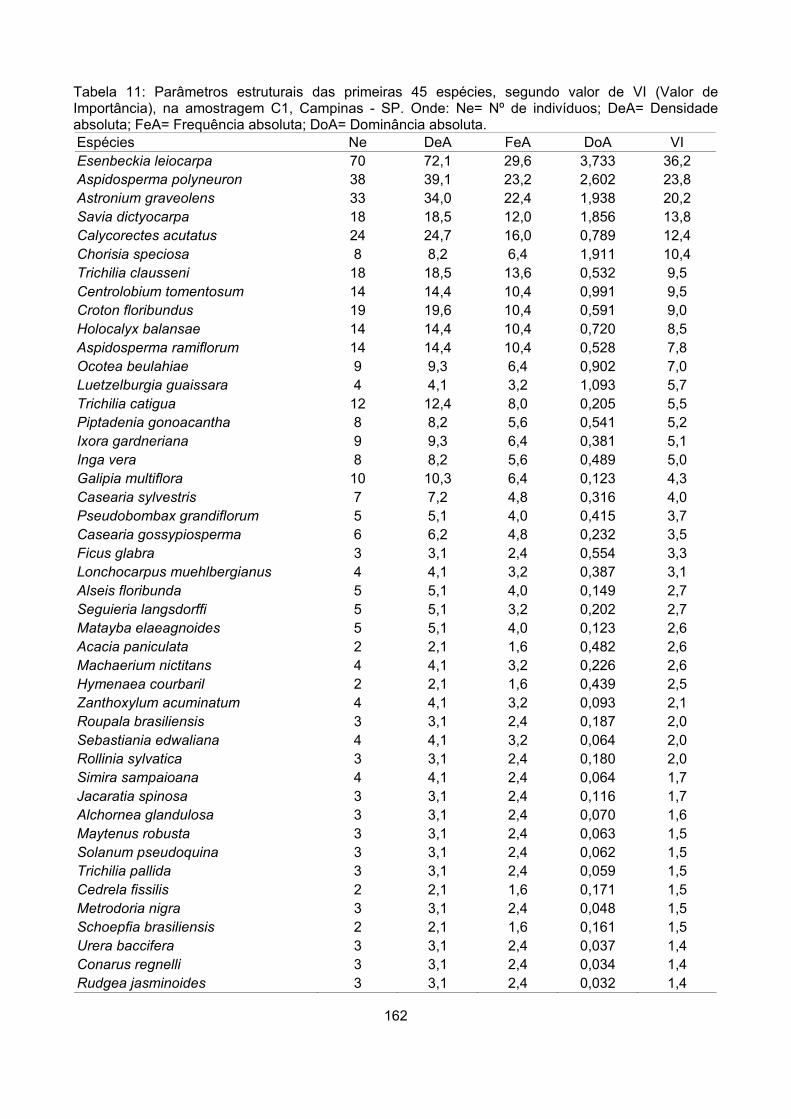
162
Tabela 11: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), na amostragem C1, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 70 72,1 29,6 3,733 36,2 Aspidosperma polyneuron 38 39,1 23,2 2,602 23,8 Astronium graveolens 33 34,0 22,4 1,938 20,2 Savia dictyocarpa 18 18,5 12,0 1,856 13,8 Calycorectes acutatus 24 24,7 16,0 0,789 12,4 Chorisia speciosa 8 8,2 6,4 1,911 10,4 Trichilia clausseni 18 18,5 13,6 0,532 9,5 Centrolobium tomentosum 14 14,4 10,4 0,991 9,5 Croton floribundus 19 19,6 10,4 0,591 9,0 Holocalyx balansae 14 14,4 10,4 0,720 8,5 Aspidosperma ramiflorum 14 14,4 10,4 0,528 7,8 Ocotea beulahiae 9 9,3 6,4 0,902 7,0 Luetzelburgia guaissara 4 4,1 3,2 1,093 5,7 Trichilia catigua 12 12,4 8,0 0,205 5,5 Piptadenia gonoacantha 8 8,2 5,6 0,541 5,2 Ixora gardneriana 9 9,3 6,4 0,381 5,1 Inga vera 8 8,2 5,6 0,489 5,0 Galipia multiflora 10 10,3 6,4 0,123 4,3 Casearia sylvestris 7 7,2 4,8 0,316 4,0 Pseudobombax grandiflorum 5 5,1 4,0 0,415 3,7 Casearia gossypiosperma 6 6,2 4,8 0,232 3,5 Ficus glabra 3 3,1 2,4 0,554 3,3 Lonchocarpus muehlbergianus 4 4,1 3,2 0,387 3,1 Alseis floribunda 5 5,1 4,0 0,149 2,7 Seguieria langsdorffi 5 5,1 3,2 0,202 2,7 Matayba elaeagnoides 5 5,1 4,0 0,123 2,6 Acacia paniculata 2 2,1 1,6 0,482 2,6 Machaerium nictitans 4 4,1 3,2 0,226 2,6 Hymenaea courbaril 2 2,1 1,6 0,439 2,5 Zanthoxylum acuminatum 4 4,1 3,2 0,093 2,1 Roupala brasiliensis 3 3,1 2,4 0,187 2,0 Sebastiania edwaliana 4 4,1 3,2 0,064 2,0 Rollinia sylvatica 3 3,1 2,4 0,180 2,0 Simira sampaioana 4 4,1 2,4 0,064 1,7 Jacaratia spinosa 3 3,1 2,4 0,116 1,7 Alchornea glandulosa 3 3,1 2,4 0,070 1,6 Maytenus robusta 3 3,1 2,4 0,063 1,5 Solanum pseudoquina 3 3,1 2,4 0,062 1,5 Trichilia pallida 3 3,1 2,4 0,059 1,5 Cedrela fissilis 2 2,1 1,6 0,171 1,5 Metrodoria nigra 3 3,1 2,4 0,048 1,5 Schoepfia brasiliensis 2 2,1 1,6 0,161 1,5 Urera baccifera 3 3,1 2,4 0,037 1,4 Conarus regnelli 3 3,1 2,4 0,034 1,4 Rudgea jasminoides 3 3,1 2,4 0,032 1,4
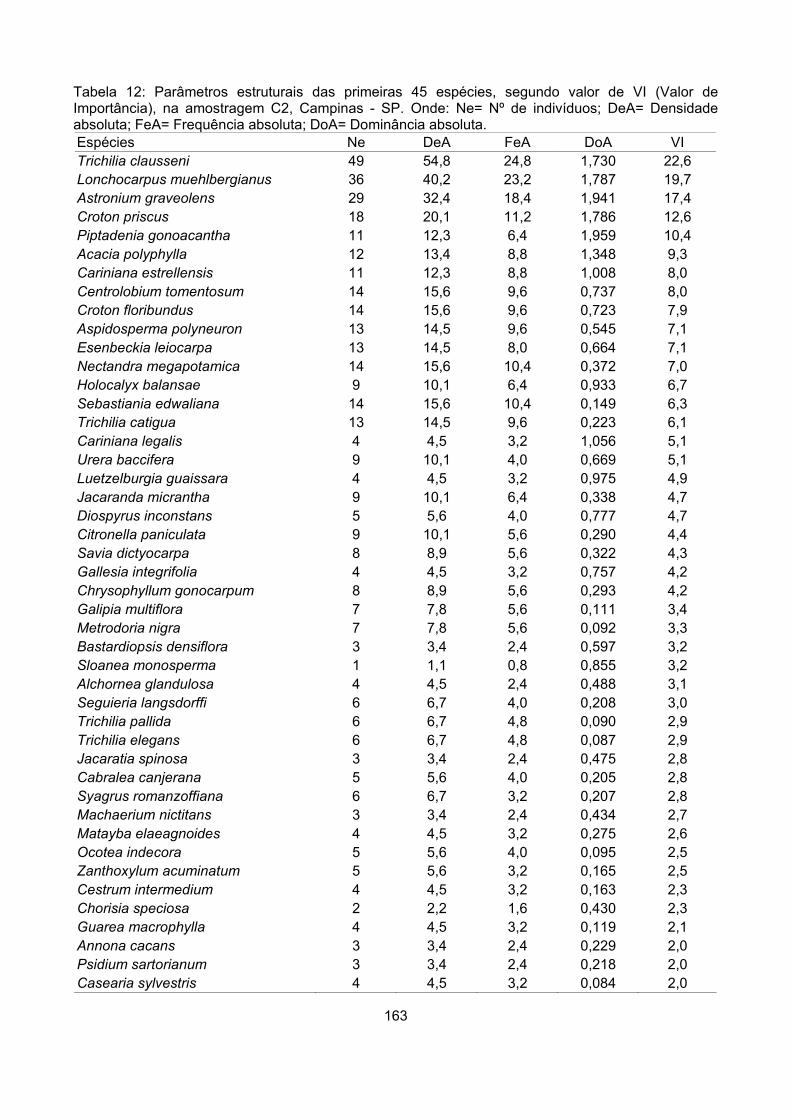
163
Tabela 12: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), na amostragem C2, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Trichilia clausseni 49 54,8 24,8 1,730 22,6 Lonchocarpus muehlbergianus 36 40,2 23,2 1,787 19,7 Astronium graveolens 29 32,4 18,4 1,941 17,4 Croton priscus 18 20,1 11,2 1,786 12,6 Piptadenia gonoacantha 11 12,3 6,4 1,959 10,4 Acacia polyphylla 12 13,4 8,8 1,348 9,3 Cariniana estrellensis 11 12,3 8,8 1,008 8,0 Centrolobium tomentosum 14 15,6 9,6 0,737 8,0 Croton floribundus 14 15,6 9,6 0,723 7,9 Aspidosperma polyneuron 13 14,5 9,6 0,545 7,1 Esenbeckia leiocarpa 13 14,5 8,0 0,664 7,1 Nectandra megapotamica 14 15,6 10,4 0,372 7,0 Holocalyx balansae 9 10,1 6,4 0,933 6,7 Sebastiania edwaliana 14 15,6 10,4 0,149 6,3 Trichilia catigua 13 14,5 9,6 0,223 6,1 Cariniana legalis 4 4,5 3,2 1,056 5,1 Urera baccifera 9 10,1 4,0 0,669 5,1 Luetzelburgia guaissara 4 4,5 3,2 0,975 4,9 Jacaranda micrantha 9 10,1 6,4 0,338 4,7 Diospyrus inconstans 5 5,6 4,0 0,777 4,7 Citronella paniculata 9 10,1 5,6 0,290 4,4 Savia dictyocarpa 8 8,9 5,6 0,322 4,3 Gallesia integrifolia 4 4,5 3,2 0,757 4,2 Chrysophyllum gonocarpum 8 8,9 5,6 0,293 4,2 Galipia multiflora 7 7,8 5,6 0,111 3,4 Metrodoria nigra 7 7,8 5,6 0,092 3,3 Bastardiopsis densiflora 3 3,4 2,4 0,597 3,2 Sloanea monosperma 1 1,1 0,8 0,855 3,2 Alchornea glandulosa 4 4,5 2,4 0,488 3,1 Seguieria langsdorffi 6 6,7 4,0 0,208 3,0 Trichilia pallida 6 6,7 4,8 0,090 2,9 Trichilia elegans 6 6,7 4,8 0,087 2,9 Jacaratia spinosa 3 3,4 2,4 0,475 2,8 Cabralea canjerana 5 5,6 4,0 0,205 2,8 Syagrus romanzoffiana 6 6,7 3,2 0,207 2,8 Machaerium nictitans 3 3,4 2,4 0,434 2,7 Matayba elaeagnoides 4 4,5 3,2 0,275 2,6 Ocotea indecora 5 5,6 4,0 0,095 2,5 Zanthoxylum acuminatum 5 5,6 3,2 0,165 2,5 Cestrum intermedium 4 4,5 3,2 0,163 2,3 Chorisia speciosa 2 2,2 1,6 0,430 2,3 Guarea macrophylla 4 4,5 3,2 0,119 2,1 Annona cacans 3 3,4 2,4 0,229 2,0 Psidium sartorianum 3 3,4 2,4 0,218 2,0 Casearia sylvestris 4 4,5 3,2 0,084 2,0

164
Tabela 13: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância), na amostragem C3, Campinas - SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 67 89,2 38,4 6,104 41,8 Croton floribundus 63 83,9 32,8 2,703 29,7 Savia dictyocarpa 22 29,3 16,0 2,169 15,2 Cariniana legalis 9 12,0 7,2 3,467 13,7 Urera baccifera 28 37,3 18,4 0,508 12,3 Astronium graveolens 15 20,0 12,0 1,879 11,8 Vernonia diffusa 16 21,3 12,8 1,061 9,9 Cecropia hololeuca 18 24,0 13,6 0,558 9,1 Chorisia speciosa 9 12,0 7,2 1,481 8,1 Aspidosperma ramiflorum 15 20,0 7,2 1,042 8,0 Aspidosperma polyneuron 8 10,7 6,4 1,614 8,0 Galipia multiflora 14 18,6 9,6 0,287 6,4 Hirtella hebeclada 8 10,7 6,4 0,581 5,1 Almeidea coerulea 11 14,6 8,0 0,128 4,9 Alchornea glandulosa 9 12,0 6,4 0,366 4,7 Trichilia pallida 9 12,0 7,2 0,274 4,7 Gallesia integrifolia 2 2,7 1,6 1,313 4,6 Ixora gardneriana 5 6,7 4,0 0,542 3,7 Trichilia catigua 8 10,7 5,6 0,148 3,6 Chrysophyllum gonocarpum 7 9,3 5,6 0,126 3,4 Calycorectes acutatus 6 8,0 4,8 0,263 3,3 Alseis floribunda 7 9,3 4,8 0,162 3,2 Margaritaria nobilis 6 8,0 4,8 0,228 3,2 Jacaratia spinosa 6 8,0 4,8 0,173 3,1 Copaifera langsdorffi 3 4,0 1,6 0,705 3,1 Myroxylon peruiferum 4 5,3 3,2 0,381 2,8 Piptadenia gonoacantha 4 5,3 3,2 0,364 2,8 Croton priscus 6 8,0 3,2 0,219 2,7 Trichilia clausseni 5 6,7 4,0 0,190 2,7 Holocalyx balansae 3 4,0 2,4 0,476 2,6 Prunus myrtifolia 4 5,3 3,2 0,316 2,6 Acacia paniculata 1 1,3 0,8 0,626 2,2 Aloysia virgata 5 6,7 3,2 0,091 2,2 Ocotea beulahiae 3 4,0 2,4 0,294 2,1 Trichilia elegans 4 5,3 3,2 0,109 2,0 Eugenia excelsa 4 5,3 3,2 0,081 2,0 Metrodoria nigra 4 5,3 3,2 0,061 1,9 Zanthoxylum hyemale 4 5,3 3,2 0,050 1,9 Cryptocarya aschersoniana 3 4,0 2,4 0,191 1,8 Cordia sellowiana 3 4,0 2,4 0,158 1,7 Seguieria langsdorffi 3 4,0 2,4 0,103 1,6 Eugenia ramboi 3 4,0 2,4 0,098 1,6 Lonchocarpus muehlbergianus 2 2,7 1,6 0,238 1,5 Nectandra oppositifolia 3 4,0 2,4 0,085 1,5 Ocotea indecora 3 4,0 2,4 0,071 1,5

165
CAPÍTULO 5
UTILIZAÇÃO DE DIFERENTES CRITÉRIOS DE INCLUSÃO EM INVENTÁRIOS FLORÍSTICOS E DE ESTRUTURA
DE FLORESTAS ESTACIONAIS SEMIDECÍDUAS E SUAS IMPLICAÇÕES SOBRE OS RESULTADOS –
UM ESTUDO DE CASO.

166
Capítulo 5
Utilização de diferentes critérios de inclusão em inventários florísticos e de estrutura de florestas estacionais semidecíduas e suas implicações sobre os resultados – um estudo de
caso.
Karin dos Santos Pós-Graduação em Biologia Vegetal – Universidade Estadual de Campinas – parte da tese de doutorado,
Luiza Sumiko Kinoshita Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Flavio A. M. dos Santos Universidade Estadual de Campinas, Departamento de Botânica, Caixa Postal 6109, Campinas, SP, 13083-970.
Resumo Muitos autores mencionam que a variação nos critérios de inclusão adotados dificultam as
comparações entre os trabalhos produzidos e poderiam levar a conclusões equivocadas. Este
questionamento é importante, uma vez que em estudos fitossociológicos ou de composição florística
não é amostrada toda a comunidade e sim um conjunto de indivíduos das populações mais
abundantes. Quais indivíduos serão amostrados depende do critério de inclusão adotado. O objetivo
deste trabalho foi investigar a variação nos aspectos qualitativos, florísticos, estruturais e de riqueza
da comunidade arbórea de fragmentos de floresta estacional semidecídua quando se utilizam dois
critérios de inclusão diferentes. Foram escolhidos oito fragmentos florestais na região da APA do
município de Campinas, SP. Em cada fragmento foram amostrados 125 pontos quadrantes,
utilizando-se dois critérios de inclusão: DAP ≥ 5cm e DAP ≥ 10cm. Em um fragmento de 244,9ha
foram feitas três amostragens nas mesmas condições em três áreas diferentes da floresta. Assim, em
cada fragmento foram obtidos dois conjuntos de dados, um contendo os indivíduos com DAP ≥ 5cm e
outro contendo os indivíduos DAP ≥ 10cm. Houve pouca variação entre os dois conjuntos de dados e
a maior parte das variações pareceu estar associada ao porte dos indivíduos, pois um critério de
inclusão menor tende a amostrar mais indivíduos de pequeno porte. Isto ocorreu, principalmente em
relação às variações de composição, de estrutura e dos aspectos qualitativos. Entretanto, parte das
variações também pareceu ter ocorrido ao acaso, principalmente aquelas relativas à composição
florística e parâmetros fitossociológicos como VI. A alta similaridade entre os resultados dos dois
conjuntos de dados, em parte, já era esperada uma vez que eles se sobrepõem. Ainda assim,
sugerem que as variações observadas quando se comparam trabalhos que utilizaram critérios de
inclusão diferentes não poderiam ser atribuídas simplesmente às diferenças nos critérios de inclusão.

167
Introdução O estudo das florestas estacionais remanescentes do estado de São Paulo teve significativo
incremento nas últimas duas décadas. Estes estudos contribuíram principalmente para o melhor
conhecimento florístico deste tipo de formação além de abrirem novas e interessantes perspectivas
para trabalhos futuros. Uma delas é a possibilidade de comparações florísticas, visando a um melhor
entendimento dos padrões florísticos e fitogeográficos, como mostraram os estudos de Salis et al.
(1995), Torres et al. (1997), Costa & Mantovani (1995) e Santos & Kinoshita (2003).
Entretanto, muitos autores mencionaram que a variação nos critérios de inclusão adotados
dificultam as comparações entre os trabalhos produzidos e poderia levar a conclusões equivocadas
(Ferreira & Prance 1998, Duringan et al. 2000, Dislich et al. 2001). Este questionamento é importante,
uma vez que em estudos fitossociológicos ou de composição florística não é amostrada toda a
comunidade e sim um conjunto de indivíduos das populações mais abundantes. Quais indivivíduos
serão amostrados depende do critério de inclusão adotado. Para Dislich et al. (2001), até mesmo
estudos realizados dentro de uma mesma floresta, usando critérios de inclusão diferentes, levariam a
resultados distintos. Os autores chamaram a atenção para a necessidade de realização de estudos
que explorem a influência da variação dos critérios de inclusão adotados sobre os resultados obtidos.
Nos estudos de estrutura e composição florística realizados em remasnescente de floresta
estacional semidecídua do interior do estado de São Paulo o diâmetro à altura do peito (DAP) é a
forma mais comumente usada como critério de inclusão, mas o diâmetro mínimo adotado varia
bastante. Por exemplo, Salis et al. (1994) e Santos & Kinoshita (2003) usaram DAP ≥ 3cm, Duringan
et al. (2000), Grombone et al. (1990), Martins (1991), e Rodrigues et al. 1989 usaram DAP ≥ 5cm e
Matthes et al. (1988), Baitello et al. (1992), Bertoni et al. (1988) e Cavassan et al. (1984) usaram DAP
≥ 10cm.
O Programa Biota/Fapesp, atento a esta problemática e visando promover uma melhor
integração entre os projetos que reúne, após longas discussões entre os projetos integrantes, sugeriu
que todos os projetos sobre estrutura e florística da vegetação arbórea adotassem DAP ≥ 5cm.
O presente trabalho é um dos projetos participantes de tal debate dentro do Programa
Biota/Fapesp. O objetivo deste trabalho foi responder à seguinte pergunta: Qual a variação nos
aspectos qualitativos, florísticos, estruturais e de riqueza da comunidade arbórea de fragmentos de
floresta estacional semidecídua quando adotamos dois critérios de inclusão diferentes?

168
Material e Métodos Área de estudo
Os fragmentos estudados localizam-se, na região da Área de Proteção Ambiental (APA) de
Sousas e Joaquim Egídio, município de Campinas (22°45’ a 23°00’ S, 47°00’ a 47°12’ W) e município
de Itatiba (23°10’ a 23°15’ S, 45°45’ a 46°45’ W) e pertencem à propriedades particulares. Seis
fragmentos localizam-se dentro dos domínios da APA (P1, M1, M4, G1, G2 e a mata Ribeirão
Cachoeira) dois localizam-se em áreas fronteiriças à APA, dentro do município de Campinas (P3 e
P2) e três em áreas fronteiriças à APA dentro do município de Itatiba (P4, M3 e M2) (ver Figura 1,
Capítulo 1). O clima, segundo Köppen (1948), é Cwag'- temperado, moderadamente chuvoso, de
inverno seco não rigoroso, com precipitação média anual de 1409 mm (Mello et al. 1994). Os solos
ocupados pelos fragmentos em estudo podem ser Podzólico Vermelho-Amarelo ou Latossolo
Vermelho. Os relevos podem ser suave ondulados, forte ondulados e montanhosos ou ondulados e
suave ondulados (Oliveira et al. 1999).
Foram escolhidos sete fragmentos com áreas inferiores à 100 hectares (ha) e um fragmento
grande - Mata Ribeirão Cachoeira - (244,9 ha), cujo levantamento florístico foi realizado por Santos &
Kinoshita (2003). A vegetação dos fragmentos pode ser classificada como floresta estacional
semidecídua (Veloso et al. 1991) e os critérios para a escolha de cada área foram: similaridade
fisionômica, facilidade de acesso e obtenção de autorização dos proprietários para a realização do
trabalho.
Amostragem
Para a coleta de dados foi adotado o método de pontos quadrantes proposto por Cottam &
Curtis (1956). Em cada fragmento foi feita uma amostragem de 125 pontos quadrantes, totalizando
500 indivíduos coletados por amostra. Os pontos foram dispostos em intervalos regulares de 15m ao
longo de transecções que percorriam área semelhante em fragmentos grandes e pequenos, evitando
apenas áreas de bordas e clareiras. As amostragens foram feitas de forma a incluir áreas
semelhantes, tanto do ponto de vista da fisionomia da vegetação, área percorrida e ambiente físico
(condições de altitude, solo e drenagem).
Devido à inexistência de pelo menos três fragmentos maiores que 200ha como é o caso da
mata Ribeirão Cachoeira, foram feitas três amostragens de 125 pontos em áreas diferentes desta
mata. Nos referiremos a estas amostras, a partir daqui, como amostras C1, C2 e C3. As áreas C1 e
C2 distam aproximadamente 700m, a distância entre as áreas C1 e C3 é de aproximadamente 800m
e entre C2 e C3 em torno de 1500m (ver Figura 1 Capítulo 1). As amostras realizadas dentro dos
fragmentos menores que 100ha chamaremos F1, F2, F3, F4, F5, F6 e F7.
Foram usados dois critérios de inclusão: diâmetro à altura do peito (DAP) maior ou igual a 5cm
e DAP ≥ 10cm. Em cada ponto foram amostrados de 4 a 8 indivíduos, dependendo do diâmetro do
indivíduo mais próximo encontrado em cada quadrante: sempre que o indivíduo mais próximo do

169
ponto, em determinado quadrante, apresentou DAP ≥ 5cm, porém inferior a 10 cm, foi amostrado um
segundo indivíduo neste mesmo quadrante com DAP ≥ 10cm. No caso de o indivíduo mais próximo
do ponto em um quadrante apresentar DAP ≥ 10cm, somente este indivíduo foi amostrado neste
quadrante. Dessa forma, foram obtidos dois conjuntos de dados contendo 5000 indivíduos cada, o
primeiro contendo os indivíduos com DAP ≥ 5cm e o segundo contendo os indivíduos com DAP ≥
10cm (Figura 1). Isso permite que os dados obtidos sejam comparados tanto com trabalhos cujo
critério de inclusão é DAP ≥ 10cm quanto com àqueles que adotam DAP ≥ 5cm.
Para cada indivíduo foram anotados a altura total, o perímetro do tronco à altura do peito e a
distância do ponto quadrante. As identificações foram feitas segundo técnicas usuais em taxonomia e
os espécimes foram incluídos em famílias segundo o sistema proposto por Cronquist (1981). A
coleção principal de material herborizado foi incorporada ao herbário UEC e as duplicatas foram
enviadas a outros herbários ou especialistas que ajudaram na identificação de alguns grupos. Os
materiais coletados em estado vegetativo foram conservados como material testemunha, em
separado.
Figura 1: Esquema ilustrativo da forma de amostragem, usando pontos quadrantes com dois critérios de inclusão diferentes, DAP ≥ 5cm e DAP ≥ 10cm.
Quadrante 2
Quadrante 3 Quadrante 4
Linha imaginária
Quadrante 1
Transecção
Árvore com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com DAP > 10cm
Árvore com DAP > 10cm
Centro do Ponto
Legenda
Quadrante 2
Quadrante 3 Quadrante 4
Linha imaginária
Quadrante 1
Transecção
Quadrante 2
Quadrante 3 Quadrante 4
Linha imaginária
Quadrante 1
Transecção
Quadrante 2
Quadrante 3 Quadrante 4
Linha imaginária
Quadrante 1
Transecção
Árvore com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com DAP > 10cm
Árvore com DAP > 10cm
Centro do Ponto
Legenda
Árvore com 5 < DAP < 10cmÁrvore com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com 5 < DAP < 10cmConjunto de dados, contendo árvores com 5 < DAP < 10cm
Conjunto de dados, contendo árvores com DAP > 10cmConjunto de dados, contendo árvores com DAP > 10cm
Árvore com DAP > 10cmÁrvore com DAP > 10cm
Centro do PontoCentro do Ponto
Legenda

170
Aspectos qualitativos
Lianas
O grau de infestação por lianas foi estimado usando-se cinco classes de porcentagem de
infestação da copa do indivíduo arbóreo por lianas (0, 1-25%, 26-50%, 51-75% e 76 a 100%). Neste
trabalho, foi considerada liana qualquer planta que mantivesse contato com o solo e necessitasse de
um suporte para seu desenvolvimento (Müller-Dombois & Ellemberg 1974).
Estado do dossel
A avaliação da qualidade do dossel foi feita através da categorização das árvores amostradas
em: árvores do futuro, árvores do presente e árvores do passado segundo as definições propostas
por Oldeman (1989), onde: a) árvores do futuro (AF) seriam indivíduos jovens, aqueles que teriam
potencial para crescer em altura e sofrer expansão de copa; b) árvores do presente (APR) seriam
indivíduos adultos, mas que ainda teriam potencial para extensão e expansão e, mesmo que seu
tamanho fosse máximo em um determinado local, sua sustentação seria aparentemente durável
devido à sua capacidade de repor partes perdidas; c) árvores do passado (APS) seriam aquelas
muito danificadas, ou seja, com vários galhos quebrados, árvores morrendo, em decadência ou uma
combinação de todos esses estados.
Guildas de espécies
Para verificar se a proporção de indivíduos e espécies zoocóricas, anemocóricas e autocóricas
se alterava entre as amostras dos diferentes critérios de inclusão, as espécies foram agrupadas por
síndromes de dispersão. Foram consideradas espécies zoocóricas aquelas que produzem frutos
carnosos (baga ou drupa), sementes ariladas ou diásporos com adaptações que permitam dispersão
por animais; as anemocóricas são diásporos que apresentam formas, estruturas ou dimensões que
permitem a dispersão pelo vento; e autocóricas os diásporos que não apresentam nenhuma das
características indicadas acima (Mantovani & Martins 1988). As espécies que não foram encontradas
com frutos tiveram o tipo de síndrome de dispersão determinada através de verificação em herbário
e/ou indicações bibliográficas.
Tratamento dos dados
Composição florística
Para se verificar se a composição florística obtida em cada fragmento era muito distinta
segundo o critério de inclusão adotado, foram feitas análises de similaridade e agrupamento para
cada conjunto de dados separadamente e os dendrogramas de similaridade obtidos foram
comparados. Se houvesse diferença de composição, segundo o critério de inclusão utilizado, seria
esperado que os dendrogramas obtidos para cada conjunto de dados fossem diferentes. Foram
comparados também os índices de similaridade obtidos entre os fragmentos para cada conjunto de

171
dados. Foi adotado o índice de similaridade de Jaccard (Pielou 1984) e as relações de similaridade
entre os fragmentos foram investigadas através da análise de agrupamento UPGMA, utilizando o
programa FITOPAC (Shepherd 1988).
Diversidade e riqueza
Para a análise da diversidade específica foram utilizados o número de espécies (S), famílias e
gêneros coletados, o índice de diversidade de Shannon-Weaver (H’), calculado na base e
(nats/indivíduo) (Ludwig & Reynolds 1988) e a eqüabilidade J= H’/Ln S (Magurran 1988).
Para estimar a riqueza de espécies em cada amostra foi utilizado o estimador de riqueza Chao I
(Chao 1984, 1987). Estimadores de riqueza não-paramétricos, como os adotados aqui, utilizam
informações sobre a distribuição de espécies raras na amostra, isto é, aquelas representadas por
apenas um (singletons) ou dois indivíduos (doubletons). Quanto maior o número de espécies raras na
amostra, maior é a probabilidade de que outras espécies não representadas na amostra estejam
ocorrendo na área (Gotelli & Colwell 2001). No caso do estimador Chao I a riqueza estimada seria
igual à observada quando todas as espécies presentes na amostra exibissem abundância de dois ou
mais indivíduos. Para Chao II isto ocorreria quando todas as espécies observadas ocorressem em
duas ou mais unidades amostrais (Colwell & Coddington 1994). Assim, riquezas estimadas com
valores próximos ao número de espécies coletadas, indicam que a maior parte das espécies que
possivelmente ocorreriam naquela área foram coletadas. Por outro lado, grande variação entre as
riquezas estimada e o número de espécies coletadas indica que uma pequena proporção das
espécies que ocorrem naquela área foi coletada e que sua riqueza tende a ser elevada.
Para estimar a riqueza por número de indivíduos em cada fragmento e para o total de
fragmentos em cada conjunto de dados, foram feitas curvas de rarefação, utilizando o programa
PAST(Hammer et al. 2001).
Todos esses cálculos foram realizados para cada conjunto de dados separadamente e
posteriormente comparados para verificar se haviam diferenças nesses parâmetros entre os dois
conjuntos de dados. O número de espécies, famílias e gêneros foram comparados através do teste
de qui-quadrado com correção de Yates segundo Zar (1989). A riqueza estimada e as curvas de
rarefação foram comparadas através dos desvios-padrões.
Para verificar a relação existente entre o número de espécies coletadas entre os conjuntos de
dados foi feita uma análise de regressão linear, utilizando-se o programa BioEstat (Ayres et al. 1998).
Estrutura
Os parâmetros estruturais da vegetação foram calculados separadamente para cada conjunto
de dados, segundo as fórmulas recapituladas por Martins (1991). Os cálculos foram efetuados com
auxílio do programa FITOPAC (Shepherd 1988). A estrutura da floresta foi caracterizada em termos
de abundância relativa, área basal, densidade, dominância das espécies e importância de cada

172
espécie em cada fragmento, expressa através do índice de valor de importância (VI, Curtis &
McIntosh 1951).
Para verificar qual a relação entre a densidade entre os conjuntos de dados foi feita uma análise
de regressão linear, utilizando-se o programa BioEstat (Ayres et al. 1998).
Aspectos qualitativos
Os indivíduos foram separados em infestados por lianas ou não infestados, independentemente
do grau de infestação de cada indivíduo. Isto foi feito para facilitar a apresentação e análise dos
resultados. Para verificar se o número de árvores infestadas por lianas nos diversos fragmentos
diferia entre os dois conjuntos de dados, as amostras foram comparadas através do teste de Qui-
quadrado com correção de Yates segundo Zar (1989).
O número de indivíduos categorizados em árvores do presente, árvores do passado e árvores
do futuro foram expressos em porcentagem, pois não foi possível a categorização de alguns
indivíduos em algumas amostras. Para verificar se a proporção de cada categoria diferia segundo o
critério de inclusão utilizado, as amostras de cada fragmento foram comparadas através do teste de
Qui-quadrado com correção de Yates segundo Zar (1989).
O número de espécies e indivíduos das diferentes síndromes de dispersão também foi expresso
em porcentagem, pois não foi possível determinar a síndrome de algumas morfo-espécies. Para as
comparações entre os dois conjuntos de dados foi adotado o mesmo procedimento descrito no
parágrafo anterior.
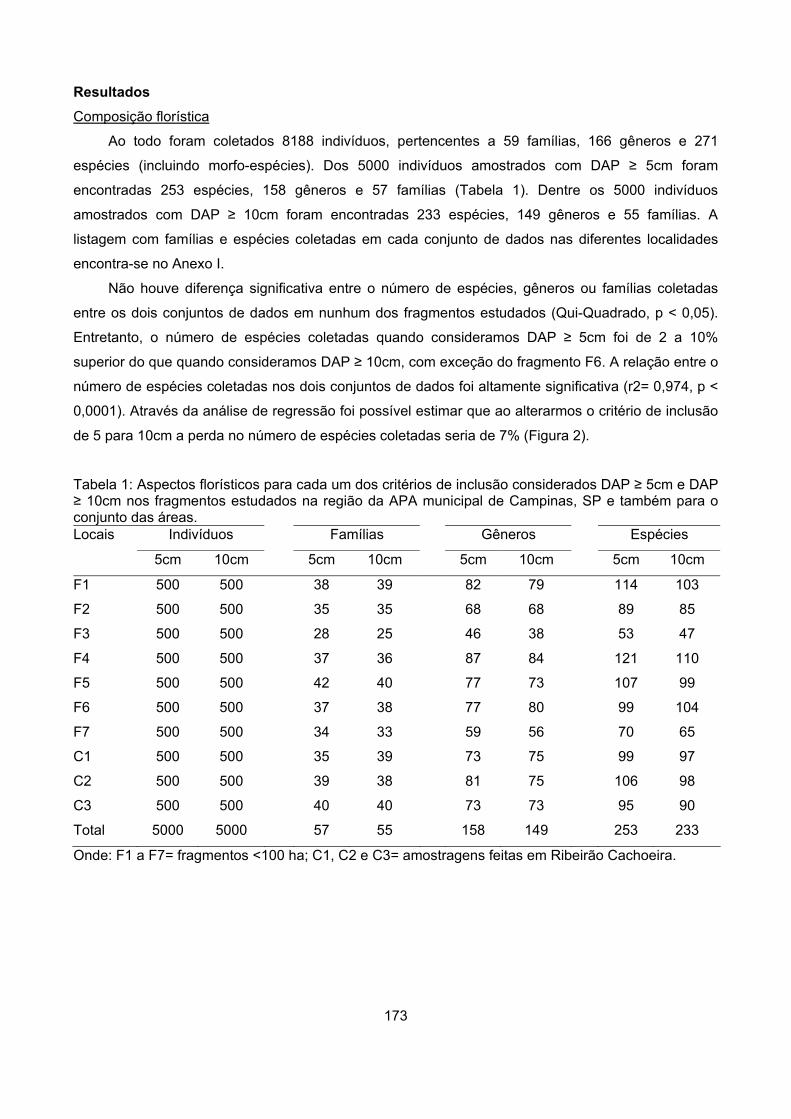
173
Resultados Composição florística
Ao todo foram coletados 8188 indivíduos, pertencentes a 59 famílias, 166 gêneros e 271
espécies (incluindo morfo-espécies). Dos 5000 indivíduos amostrados com DAP ≥ 5cm foram
encontradas 253 espécies, 158 gêneros e 57 famílias (Tabela 1). Dentre os 5000 indivíduos
amostrados com DAP ≥ 10cm foram encontradas 233 espécies, 149 gêneros e 55 famílias. A
listagem com famílias e espécies coletadas em cada conjunto de dados nas diferentes localidades
encontra-se no Anexo I.
Não houve diferença significativa entre o número de espécies, gêneros ou famílias coletadas
entre os dois conjuntos de dados em nunhum dos fragmentos estudados (Qui-Quadrado, p < 0,05).
Entretanto, o número de espécies coletadas quando consideramos DAP ≥ 5cm foi de 2 a 10%
superior do que quando consideramos DAP ≥ 10cm, com exceção do fragmento F6. A relação entre o
número de espécies coletadas nos dois conjuntos de dados foi altamente significativa (r2= 0,974, p <
0,0001). Através da análise de regressão foi possível estimar que ao alterarmos o critério de inclusão
de 5 para 10cm a perda no número de espécies coletadas seria de 7% (Figura 2).
Tabela 1: Aspectos florísticos para cada um dos critérios de inclusão considerados DAP ≥ 5cm e DAP ≥ 10cm nos fragmentos estudados na região da APA municipal de Campinas, SP e também para o conjunto das áreas. Locais Indivíduos Famílias Gêneros Espécies
5cm 10cm 5cm 10cm 5cm 10cm 5cm 10cm
F1 500 500 38 39 82 79 114 103
F2 500 500 35 35 68 68 89 85
F3 500 500 28 25 46 38 53 47
F4 500 500 37 36 87 84 121 110
F5 500 500 42 40 77 73 107 99
F6 500 500 37 38 77 80 99 104
F7 500 500 34 33 59 56 70 65
C1 500 500 35 39 73 75 99 97
C2 500 500 39 38 81 75 106 98
C3 500 500 40 40 73 73 95 90
Total 5000 5000 57 55 158 149 253 233
Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
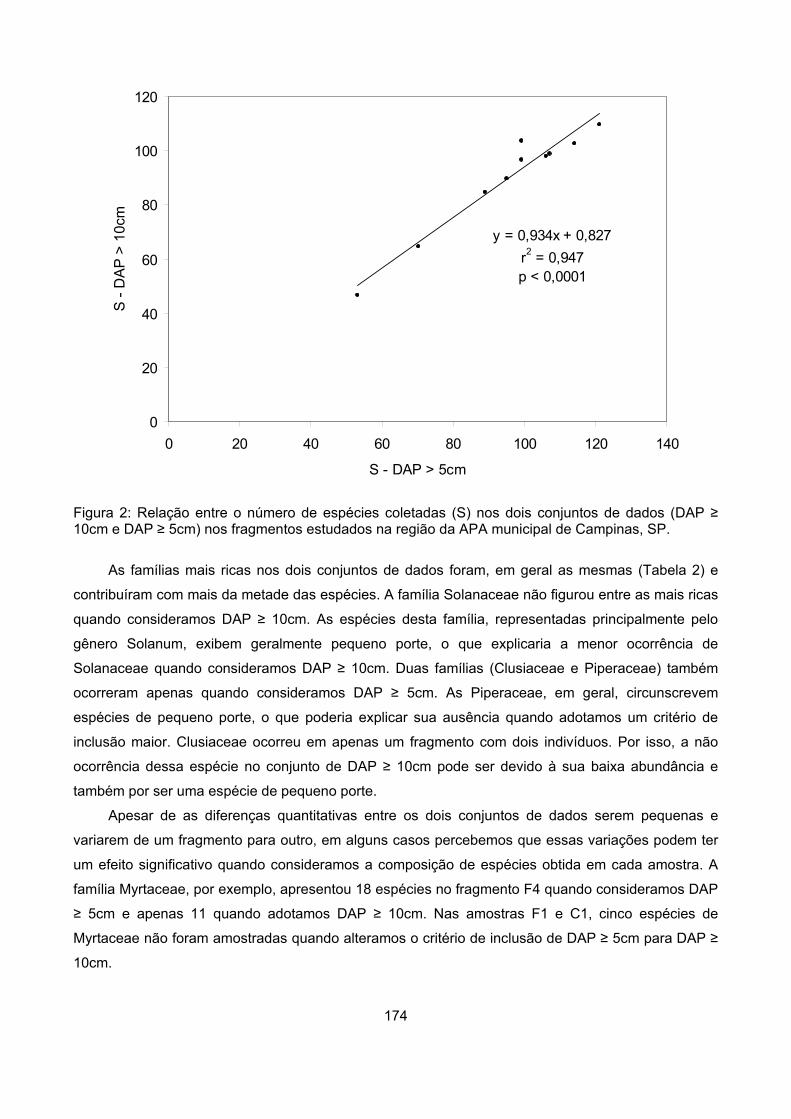
174
Figura 2: Relação entre o número de espécies coletadas (S) nos dois conjuntos de dados (DAP ≥ 10cm e DAP ≥ 5cm) nos fragmentos estudados na região da APA municipal de Campinas, SP.
As famílias mais ricas nos dois conjuntos de dados foram, em geral as mesmas (Tabela 2) e
contribuíram com mais da metade das espécies. A família Solanaceae não figurou entre as mais ricas
quando consideramos DAP ≥ 10cm. As espécies desta família, representadas principalmente pelo
gênero Solanum, exibem geralmente pequeno porte, o que explicaria a menor ocorrência de
Solanaceae quando consideramos DAP ≥ 10cm. Duas famílias (Clusiaceae e Piperaceae) também
ocorreram apenas quando consideramos DAP ≥ 5cm. As Piperaceae, em geral, circunscrevem
espécies de pequeno porte, o que poderia explicar sua ausência quando adotamos um critério de
inclusão maior. Clusiaceae ocorreu em apenas um fragmento com dois indivíduos. Por isso, a não
ocorrência dessa espécie no conjunto de DAP ≥ 10cm pode ser devido à sua baixa abundância e
também por ser uma espécie de pequeno porte.
Apesar de as diferenças quantitativas entre os dois conjuntos de dados serem pequenas e
variarem de um fragmento para outro, em alguns casos percebemos que essas variações podem ter
um efeito significativo quando consideramos a composição de espécies obtida em cada amostra. A
família Myrtaceae, por exemplo, apresentou 18 espécies no fragmento F4 quando consideramos DAP
≥ 5cm e apenas 11 quando adotamos DAP ≥ 10cm. Nas amostras F1 e C1, cinco espécies de
Myrtaceae não foram amostradas quando alteramos o critério de inclusão de DAP ≥ 5cm para DAP ≥
10cm.
y = 0,934x + 0,827r2 = 0,947p < 0,0001
0
20
40
60
80
100
120
0 20 40 60 80 100 120 140
S - DAP > 5cm
S -
DA
P >
10c
m
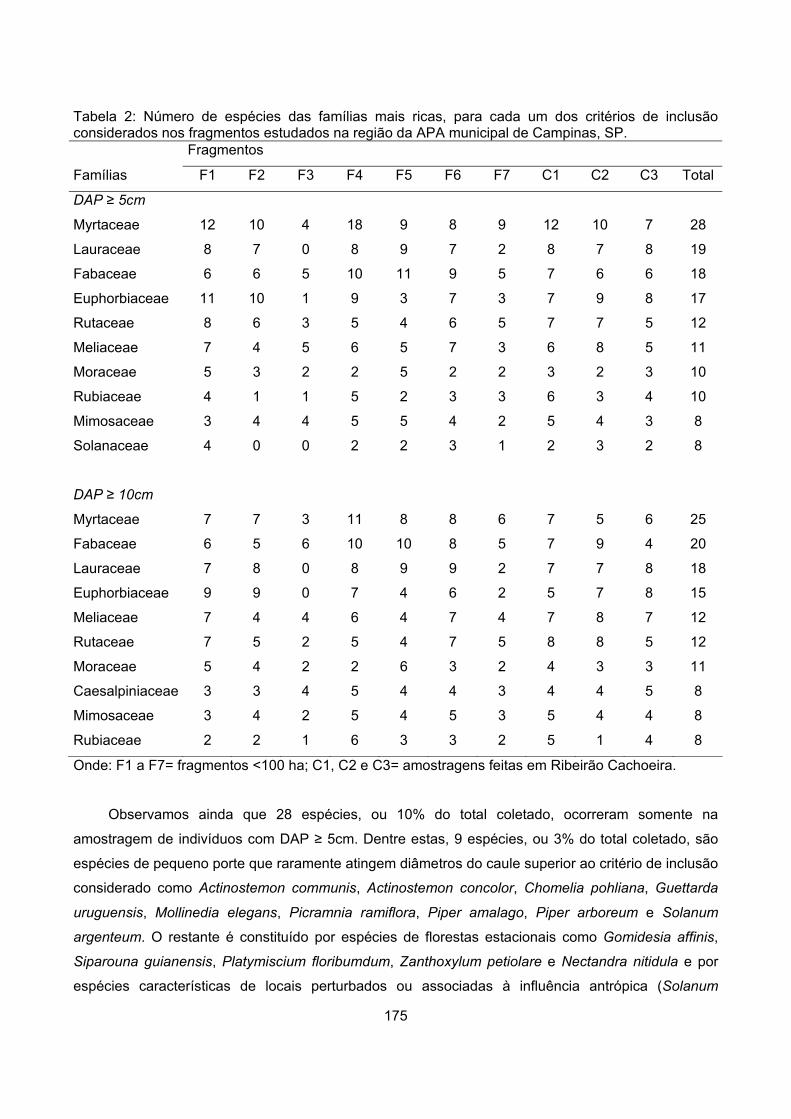
175
Tabela 2: Número de espécies das famílias mais ricas, para cada um dos critérios de inclusão considerados nos fragmentos estudados na região da APA municipal de Campinas, SP. Fragmentos
Famílias F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 Total
DAP ≥ 5cm
Myrtaceae 12 10 4 18 9 8 9 12 10 7 28
Lauraceae 8 7 0 8 9 7 2 8 7 8 19
Fabaceae 6 6 5 10 11 9 5 7 6 6 18
Euphorbiaceae 11 10 1 9 3 7 3 7 9 8 17
Rutaceae 8 6 3 5 4 6 5 7 7 5 12
Meliaceae 7 4 5 6 5 7 3 6 8 5 11
Moraceae 5 3 2 2 5 2 2 3 2 3 10
Rubiaceae 4 1 1 5 2 3 3 6 3 4 10
Mimosaceae 3 4 4 5 5 4 2 5 4 3 8
Solanaceae 4 0 0 2 2 3 1 2 3 2 8
DAP ≥ 10cm
Myrtaceae 7 7 3 11 8 8 6 7 5 6 25
Fabaceae 6 5 6 10 10 8 5 7 9 4 20
Lauraceae 7 8 0 8 9 9 2 7 7 8 18
Euphorbiaceae 9 9 0 7 4 6 2 5 7 8 15
Meliaceae 7 4 4 6 4 7 4 7 8 7 12
Rutaceae 7 5 2 5 4 7 5 8 8 5 12
Moraceae 5 4 2 2 6 3 2 4 3 3 11
Caesalpiniaceae 3 3 4 5 4 4 3 4 4 5 8
Mimosaceae 3 4 2 5 4 5 3 5 4 4 8
Rubiaceae 2 2 1 6 3 3 2 5 1 4 8
Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
Observamos ainda que 28 espécies, ou 10% do total coletado, ocorreram somente na
amostragem de indivíduos com DAP ≥ 5cm. Dentre estas, 9 espécies, ou 3% do total coletado, são
espécies de pequeno porte que raramente atingem diâmetros do caule superior ao critério de inclusão
considerado como Actinostemon communis, Actinostemon concolor, Chomelia pohliana, Guettarda
uruguensis, Mollinedia elegans, Picramnia ramiflora, Piper amalago, Piper arboreum e Solanum
argenteum. O restante é constituído por espécies de florestas estacionais como Gomidesia affinis,
Siparouna guianensis, Platymiscium floribumdum, Zanthoxylum petiolare e Nectandra nitidula e por
espécies características de locais perturbados ou associadas à influência antrópica (Solanum

176
granuloso-leprosum, Psidium guajava). O fato dessas espécies terem apresentado somente
indivíduos com diâmetros entre 5 e 10cm pode ter ocorrido tanto por elas serem pouco abundantes
nessas áreas, quanto por estarem começando a penetrar no sistema ou ainda, devido à estrutura das
populações dessas espécies nessas áreas. Entretanto, seriam necessários estudos mais específicos
para a comprovação de tais afirmações.
Outras 8 espécies (2% do total coletado) ocorreram somente na amostragem de indivíduos com
DAP ≥ 10cm. São, em geral, espécies de médio a grande porte e, a maioria, com excessão de Cordia
trichotoma, Dianopteryx sorbifolia e Guarea guidonea, raramente são citadas nos levantamentos
florísticos do interior do estado de São Paulo. Está incluída entre estas, Ficus insipida espécie
apontada por Santin (1999) como “ameaçada de extinção” no município. O fato de essas espécies
não terem sido encontradas com diâmetros inferiores a 10cm pode ser devido a vários fatores, tais
como: baixa abundância e/ou densidade nessas áreas, ausência de regeneração nos locais onde
foram encontradas ou devido a estrutura de suas populações. A comprovação de tais afirmações,
entretanto, só seria possível através de estudos das populações dessas espécies em cada
fragmento.
O conjunto de DAP ≥ 5cm tendeu a apresentar um número relativamente maior de espécies,
pois incluiu tanto as espécies de pequeno porte, quanto os indivíduos jovens das espécies de grande
porte. O caso do fragmento F6, onde ocorreu o inverso, indica que nem sempre um critério de
inclusão menor resultará em um maior número de espécies coletadas. Neste fragmento é possível
que uma ou mais espécies sejam muito abundantes com diâmetros entre 5 e 10cm, como é o caso de
Urera baccifera e Almeidea coerulea. A alta dominância de tais espécies nessa faixa de diâmetro
poderia dificultar a ocorrência de outras espécies e com isso reduzir o número de espécies coletadas
quando consideramos DAP ≥ 5cm.
As diferenças de composição entre os dois conjuntos de dados também podem ser
consideradas pequenas, pois apenas 3% do total de espécies coletadas dificilmente ocorreriam se
usássemos DAP ≥ 10cm. Dentre as espécies de grande porte que também poderiam ser incluídas se
usássemos um critério de inclusão menor, apenas 2% ocorreram somente no conjunto de DAP ≥
10cm. Tais diferenças de composição também poderiam ocorrer ao acaso.
Diversidade
Os valores obtidos para o índice de diversidade de Shannon e a eqüabilidade para os dois
critérios de inclusão adotados, em cada um dos fragmentos estudados e também para o conjunto das
áreas, foram muito similares (Tabela 3). Os índices de diversidade utilizados levam em conta o
número de espécies e também a abundância relativa das espécies. Assim como o número de
espécies em cada amostra foi muito semelhante, os índices de diversidade também tendem a ser
bastante próximos, independentemente do critério de inclusão considerado.
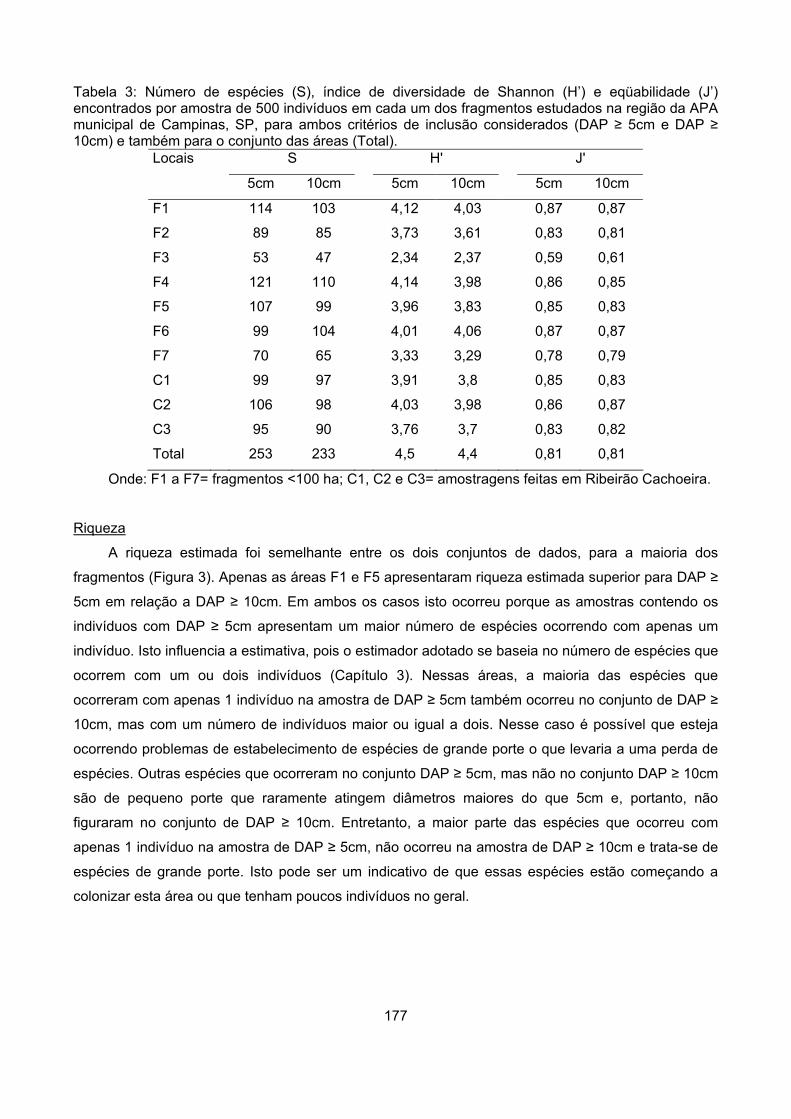
177
Tabela 3: Número de espécies (S), índice de diversidade de Shannon (H’) e eqüabilidade (J’) encontrados por amostra de 500 indivíduos em cada um dos fragmentos estudados na região da APA municipal de Campinas, SP, para ambos critérios de inclusão considerados (DAP ≥ 5cm e DAP ≥ 10cm) e também para o conjunto das áreas (Total).
Locais S H' J'
5cm 10cm 5cm 10cm 5cm 10cm
F1 114 103 4,12 4,03 0,87 0,87
F2 89 85 3,73 3,61 0,83 0,81
F3 53 47 2,34 2,37 0,59 0,61
F4 121 110 4,14 3,98 0,86 0,85
F5 107 99 3,96 3,83 0,85 0,83
F6 99 104 4,01 4,06 0,87 0,87
F7 70 65 3,33 3,29 0,78 0,79
C1 99 97 3,91 3,8 0,85 0,83
C2 106 98 4,03 3,98 0,86 0,87
C3 95 90 3,76 3,7 0,83 0,82
Total 253 233 4,5 4,4 0,81 0,81
Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
Riqueza
A riqueza estimada foi semelhante entre os dois conjuntos de dados, para a maioria dos
fragmentos (Figura 3). Apenas as áreas F1 e F5 apresentaram riqueza estimada superior para DAP ≥
5cm em relação a DAP ≥ 10cm. Em ambos os casos isto ocorreu porque as amostras contendo os
indivíduos com DAP ≥ 5cm apresentam um maior número de espécies ocorrendo com apenas um
indivíduo. Isto influencia a estimativa, pois o estimador adotado se baseia no número de espécies que
ocorrem com um ou dois indivíduos (Capítulo 3). Nessas áreas, a maioria das espécies que
ocorreram com apenas 1 indivíduo na amostra de DAP ≥ 5cm também ocorreu no conjunto de DAP ≥
10cm, mas com um número de indivíduos maior ou igual a dois. Nesse caso é possível que esteja
ocorrendo problemas de estabelecimento de espécies de grande porte o que levaria a uma perda de
espécies. Outras espécies que ocorreram no conjunto DAP ≥ 5cm, mas não no conjunto DAP ≥ 10cm
são de pequeno porte que raramente atingem diâmetros maiores do que 5cm e, portanto, não
figuraram no conjunto de DAP ≥ 10cm. Entretanto, a maior parte das espécies que ocorreu com
apenas 1 indivíduo na amostra de DAP ≥ 5cm, não ocorreu na amostra de DAP ≥ 10cm e trata-se de
espécies de grande porte. Isto pode ser um indicativo de que essas espécies estão começando a
colonizar esta área ou que tenham poucos indivíduos no geral.
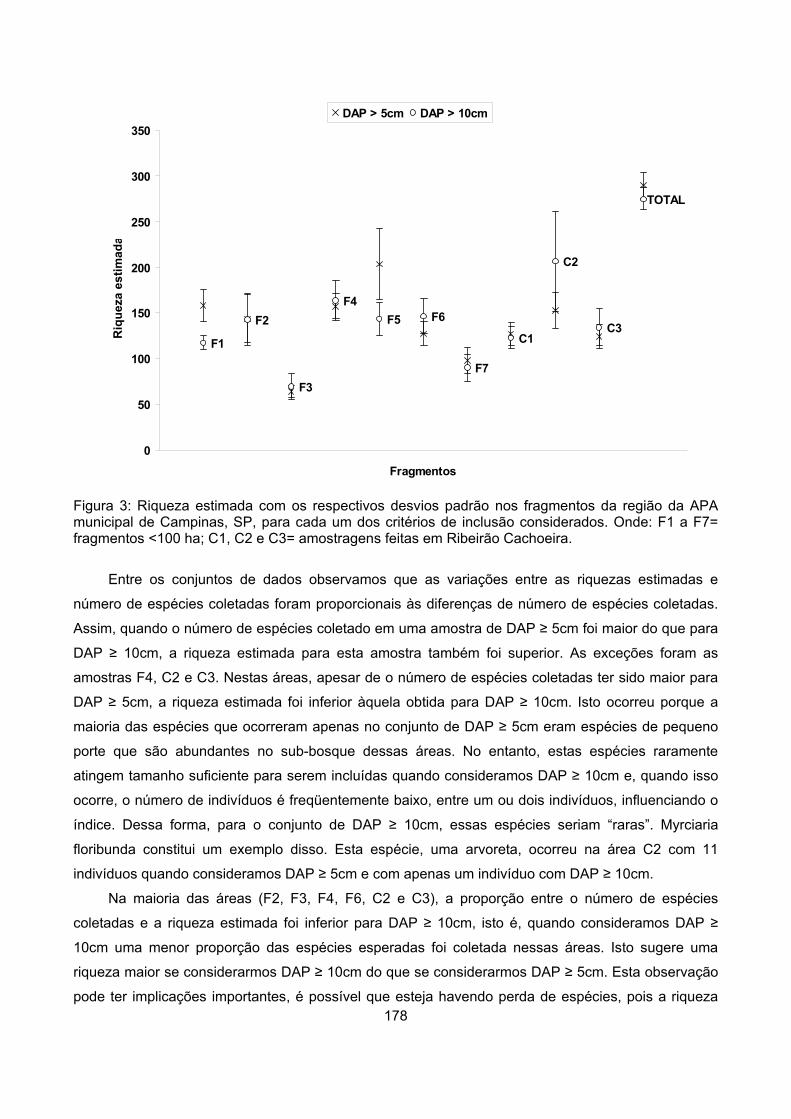
178
Figura 3: Riqueza estimada com os respectivos desvios padrão nos fragmentos da região da APA municipal de Campinas, SP, para cada um dos critérios de inclusão considerados. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
Entre os conjuntos de dados observamos que as variações entre as riquezas estimadas e
número de espécies coletadas foram proporcionais às diferenças de número de espécies coletadas.
Assim, quando o número de espécies coletado em uma amostra de DAP ≥ 5cm foi maior do que para
DAP ≥ 10cm, a riqueza estimada para esta amostra também foi superior. As exceções foram as
amostras F4, C2 e C3. Nestas áreas, apesar de o número de espécies coletadas ter sido maior para
DAP ≥ 5cm, a riqueza estimada foi inferior àquela obtida para DAP ≥ 10cm. Isto ocorreu porque a
maioria das espécies que ocorreram apenas no conjunto de DAP ≥ 5cm eram espécies de pequeno
porte que são abundantes no sub-bosque dessas áreas. No entanto, estas espécies raramente
atingem tamanho suficiente para serem incluídas quando consideramos DAP ≥ 10cm e, quando isso
ocorre, o número de indivíduos é freqüentemente baixo, entre um ou dois indivíduos, influenciando o
índice. Dessa forma, para o conjunto de DAP ≥ 10cm, essas espécies seriam “raras”. Myrciaria
floribunda constitui um exemplo disso. Esta espécie, uma arvoreta, ocorreu na área C2 com 11
indivíduos quando consideramos DAP ≥ 5cm e com apenas um indivíduo com DAP ≥ 10cm.
Na maioria das áreas (F2, F3, F4, F6, C2 e C3), a proporção entre o número de espécies
coletadas e a riqueza estimada foi inferior para DAP ≥ 10cm, isto é, quando consideramos DAP ≥
10cm uma menor proporção das espécies esperadas foi coletada nessas áreas. Isto sugere uma
riqueza maior se considerarmos DAP ≥ 10cm do que se considerarmos DAP ≥ 5cm. Esta observação
pode ter implicações importantes, é possível que esteja havendo perda de espécies, pois a riqueza
F1
F2
F3
F4 F5 F6
F7
C1
C2
C3
TOTAL
0
50
100
150
200
250
300
350
Fragmentos
Riq
ueza
est
imad
aDAP > 5cm DAP > 10cm

179
tendeu a ser maior nas classes de maior diâmetro (DAP ≥ 10cm) do que nas de menor diâmetro (DAP
≥ 5cm). O inverso só ocorreu nos fragmentos F1 e F5. Em outros dois fragmentos esta proporção foi
a mesma, independentemente do critério de inclusão adotado.
As estimativas do número de espécies por número de indivíduos considerando todas as áreas
em conjunto para cada conjunto de dados indicam que a riqueza seria maior para DAP ≥ 5cm,
apenas para um número de indivíduos superior a 1000 (Figura 4).
Figura 4: Curva de rarefação para o total de indivíduos com DAP ≥ 5cm e DAP ≥ 10cm amostrados nos fragmentos em estudo na região da APA municipal de Campinas, SP.
Similaridade
A similaridade entre as áreas variou pouco quando alteramos o critério de inclusão (Tabela 3).
Apenas no caso da amostra F2 a similaridade com as demais áreas foi maior para DAP ≥ 10cm do
que quando consideramos DAP ≥ 5cm. Talvez por isso, nos dendrogramas de similaridade obitidos, a
mata F2 figure isolada das demais para DAP ≥ 5cm, mas apareça agrupada às áreas F4 e F5 para
DAP ≥ 10cm (Figura 5). Ainda assim, os fragmentos formam grupos muito semelhantes e os
principais padrões de agrupamento se mantêm em ambos conjuntos de dados nos dendrogramas de
similaridade obtidos.
Apesar da pouca variação observada, é possível que a similaridade entre áreas possa ser
afetada pelo critério de inclusão adotado, pois como mencionado acima um critério de inclusão menor
0
50
100
150
200
250
300
0 500 1000 1500 2000 2500 3000 3500 4000 4500 5000
Indivíduos
Riq
ueza
est
imad
a
5cm 10cm
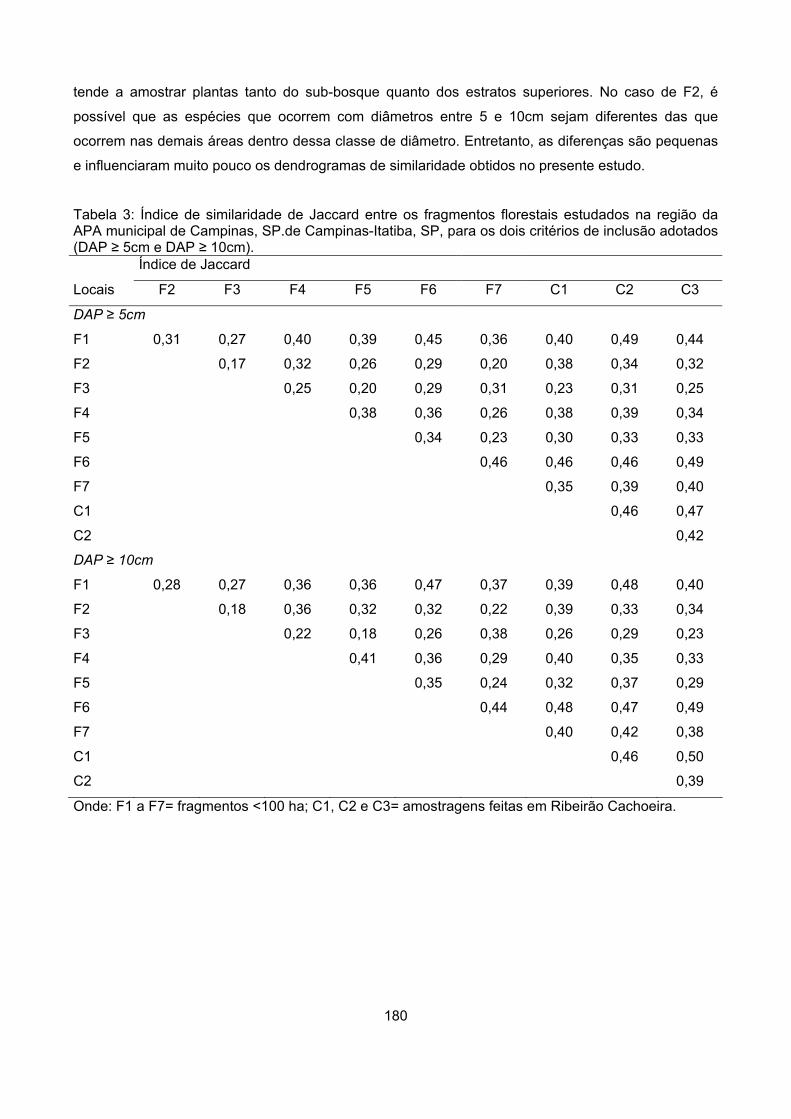
180
tende a amostrar plantas tanto do sub-bosque quanto dos estratos superiores. No caso de F2, é
possível que as espécies que ocorrem com diâmetros entre 5 e 10cm sejam diferentes das que
ocorrem nas demais áreas dentro dessa classe de diâmetro. Entretanto, as diferenças são pequenas
e influenciaram muito pouco os dendrogramas de similaridade obtidos no presente estudo.
Tabela 3: Índice de similaridade de Jaccard entre os fragmentos florestais estudados na região da APA municipal de Campinas, SP.de Campinas-Itatiba, SP, para os dois critérios de inclusão adotados (DAP ≥ 5cm e DAP ≥ 10cm). Índice de Jaccard
Locais F2 F3 F4 F5 F6 F7 C1 C2 C3
DAP ≥ 5cm
F1 0,31 0,27 0,40 0,39 0,45 0,36 0,40 0,49 0,44
F2 0,17 0,32 0,26 0,29 0,20 0,38 0,34 0,32
F3 0,25 0,20 0,29 0,31 0,23 0,31 0,25
F4 0,38 0,36 0,26 0,38 0,39 0,34
F5 0,34 0,23 0,30 0,33 0,33
F6 0,46 0,46 0,46 0,49
F7 0,35 0,39 0,40
C1 0,46 0,47
C2 0,42
DAP ≥ 10cm
F1 0,28 0,27 0,36 0,36 0,47 0,37 0,39 0,48 0,40
F2 0,18 0,36 0,32 0,32 0,22 0,39 0,33 0,34
F3 0,22 0,18 0,26 0,38 0,26 0,29 0,23
F4 0,41 0,36 0,29 0,40 0,35 0,33
F5 0,35 0,24 0,32 0,37 0,29
F6 0,44 0,48 0,47 0,49
F7 0,40 0,42 0,38
C1 0,46 0,50
C2 0,39
Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.

181
Figura 5: Dendrograma de similaridade obtido através do índice de Jaccard, considerando os fragmentos estudados na região da APA municipal de Campinas, SP, quando o critério de inclusão foi (A) DAP ≥ 5cm e (B) DAP ≥ 10cm. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
20
100%
30
40
50
60
70
80
90
F6 C3 C1 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
A
B
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
20
100%
30
40
50
60
70
80
90
F6 C3 C1 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
A20
100%
30
40
50
60
70
80
90
F6 C3 C1 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
20
100%
30
40
50
60
70
80
90
F6 C3 C1 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
20
100%
30
40
50
60
70
80
90
100%
30
40
50
60
70
80
90
F6 C3 C1 F1 C2 F7 F4 F5 F2 F3F6 C3 C1 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
A
B
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
B
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
Sim
ilarid
ade
100%
20
30
40
50
60
70
80
90
100%
20
30
40
50
60
70
80
90
C1 C3 F6 F1 C2 F7 F4 F5 F2 F3C1 C3 F6 F1 C2 F7 F4 F5 F2 F3
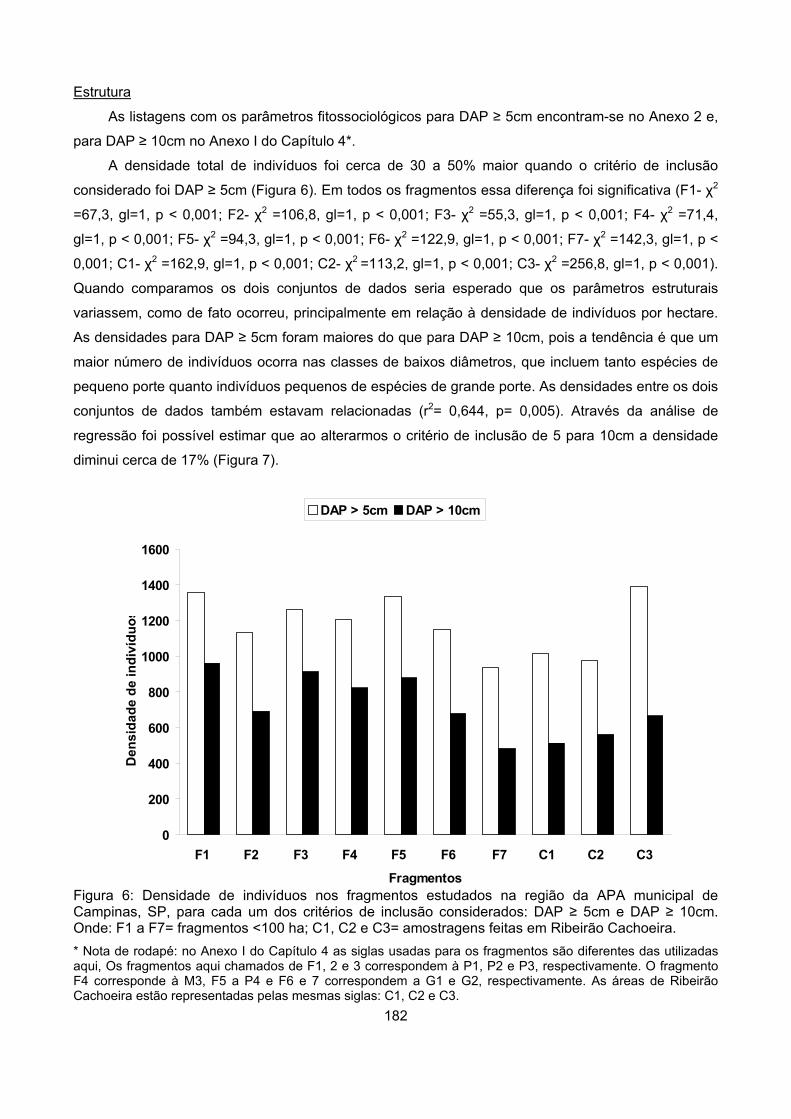
182
Estrutura
As listagens com os parâmetros fitossociológicos para DAP ≥ 5cm encontram-se no Anexo 2 e,
para DAP ≥ 10cm no Anexo I do Capítulo 4*.
A densidade total de indivíduos foi cerca de 30 a 50% maior quando o critério de inclusão
considerado foi DAP ≥ 5cm (Figura 6). Em todos os fragmentos essa diferença foi significativa (F1- χ2
=67,3, gl=1, p < 0,001; F2- χ2 =106,8, gl=1, p < 0,001; F3- χ2 =55,3, gl=1, p < 0,001; F4- χ2 =71,4,
gl=1, p < 0,001; F5- χ2 =94,3, gl=1, p < 0,001; F6- χ2 =122,9, gl=1, p < 0,001; F7- χ2 =142,3, gl=1, p <
0,001; C1- χ2 =162,9, gl=1, p < 0,001; C2- χ2 =113,2, gl=1, p < 0,001; C3- χ2 =256,8, gl=1, p < 0,001).
Quando comparamos os dois conjuntos de dados seria esperado que os parâmetros estruturais
variassem, como de fato ocorreu, principalmente em relação à densidade de indivíduos por hectare.
As densidades para DAP ≥ 5cm foram maiores do que para DAP ≥ 10cm, pois a tendência é que um
maior número de indivíduos ocorra nas classes de baixos diâmetros, que incluem tanto espécies de
pequeno porte quanto indivíduos pequenos de espécies de grande porte. As densidades entre os dois
conjuntos de dados também estavam relacionadas (r2= 0,644, p= 0,005). Através da análise de
regressão foi possível estimar que ao alterarmos o critério de inclusão de 5 para 10cm a densidade
diminui cerca de 17% (Figura 7).
Figura 6: Densidade de indivíduos nos fragmentos estudados na região da APA municipal de Campinas, SP, para cada um dos critérios de inclusão considerados: DAP ≥ 5cm e DAP ≥ 10cm. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
* Nota de rodapé: no Anexo I do Capítulo 4 as siglas usadas para os fragmentos são diferentes das utilizadas aqui, Os fragmentos aqui chamados de F1, 2 e 3 correspondem à P1, P2 e P3, respectivamente. O fragmento F4 corresponde à M3, F5 a P4 e F6 e 7 correspondem a G1 e G2, respectivamente. As áreas de Ribeirão Cachoeira estão representadas pelas mesmas siglas: C1, C2 e C3.
0
200
400
600
800
1000
1200
1400
1600
F1 F2 F3 F4 F5 F6 F7 C1 C2 C3
Fragmentos
Den
sida
de d
e in
diví
duos
DAP > 5cm DAP > 10cm

183
Figura 7: Relação entre a densidade de indivíduos nos dois conjuntos de dados (DAP ≥ 10cm e DAP ≥ 5cm) nos fragmentos estudados na região da APA municipal de Campinas, SP.
As famílias mais abundantes em cada conjunto de dados foram basicamente as mesmas,
variando apenas de posição no ranking de abundância (Tabela 4). Essas famílias representaram
cerca de 70% dos indivíduos em cada conjunto e foram também as de maior densidade absoluta em
ambos os conjuntos.
Tabela 4: Número de indivíduos (N) das famílias mais abundantes para o total de indivíduos amostrados nos fragmentos estudados na região da APA municipal de Campinas, SP, segundo os dois critério de inclusão considerados: DAP ≥ 5cm e DAP ≥ 10cm.
5cm 10cm
Famílias N Famílias N
Rutaceae 746 Rutaceae 606
Meliaceae 694 Meliaceae 635
Euphorbiaceae 524 Euphorbiaceae 493
Fabaceae 387 Fabaceae 452
Myrtaceae 288 Mimosaceae 247
Apocynaceae 249 Apocynaceae 296
Lauraceae 223 Myrtaceae 221
Caesalpiniaceae 195 Lauraceae 239
Mimosaceae 189 Caesalpiniaceae 216
Flacourtiaceae 162 Anacardiaceae 171
Anacardiaceae 153 Flacourtiaceae 157
y = 0,834x - 263,7r2 = 0,644p = 0,005
0
200
400
600
800
1000
1200
0 200 400 600 800 1000 1200 1400 1600Densidade DAP > 5cm
Den
sida
de D
AP
> 1
0cm

184
Apesar de as famílias mais abundantes em cada conjunto de dados serem basicamente as
mesmas (Tabela 4), observamos que outras famílias foram mais abundantes em um determinado
conjunto. As famílias Meliaceae, Rutaceae, Euphorbiaceae, Myrtaceae, Urticaceae, Celastraceae e
Solanaceae, por exemplo, foram muito mais abundantes quando consideramos DAP ≥ 5cm do que
para DAP ≥ 10cm. As famílias Fabaceae, Apocynaceae, Mimosaceae, Lauraceae, Caesalpiniaceae,
Anacardiaceae, Phytollacaceae, Bombacaceae, Moraceae, Lecythidaceae, Annonaceae,
Cecropiaceae e Arecaceae, por outro lado, foram mais abundantes quando o critério de inclusão foi
DAP ≥ 10cm. Esta variação pode estar relacionada ao porte das espécies que compõem estas
famílias, mas também à estrutura de suas populações nessas áreas.
Em todas as amostras ocorreu uma concentração de abundância, isto é, poucas espécies
exerceram grande dominância em sua área de ocorrência. As dez espécies mais abundantes (c.a.
20%) em cada amostra contribuíram com cerca de 41 a 82% dos indivíduos amostrados. Por outro
lado 50% das espécies contribuíram com menos de 20% do total de indivíduos.
A relação entre abundância e freqüência mostra que as espécies mais abundantes tendem a
possuir maiores freqüências de ocorrência, independentemente do critério de inclusão adotado
(Figura 8).
185
Figura 8: Relação entre abundância e freqüência de ocorrência das espécies nos fragmentos estudados na região da APA municipal de Campinas, SP, quando o critério de inclusão foi DAP ≥ 5cm (A) e para DAP ≥ 10cm (B).
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
osA
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
os
B
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
osA
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
osA
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
osA
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
os
B
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 6 7 8 9 10 11
Número de ocorrências
N.o
de
indi
vídu
os
B

186
Houve pouca variação entre as famílias que atingiram os maiores VIs entre os dois conjuntos de
dados. Em geral, quando se comparam as amostras de DAP ≥ 5cm e DAP ≥ 10cm dentro de um
mesmo fragmento, ocorreram apenas variações de posição no ranking de VI (Tabela 5). Dezoito
famílias figuraram entre as dez de maior VI quando consideramos DAP ≥ 5cm e 19 para DAP ≥ 10cm.
As variações entre as amostras de cada critério de inclusão refletem as já mencionadas quanto à
abundância destas famílias.
Considerando as 10 amostras de DAP ≥ 5cm, quarenta e uma espécies figuraram entre as 10
espécies de maior VI. Apenas 5 espécies estiveram entre as 10 de maior VI em pelo menos 50% das
amostras: Esenbeckia leiocarpa, Croton floribundus, Astronium graveolens, Aspidosperma polyneuron
e Piptadenia gonoacantha.
Para DAP ≥ 10cm, observamos que 40 espécies figuraram entre as dez de maior VI. Nenhuma
espécie figurou entre as dez mais importantes em todas as amostras. Sete espécies estiveram entre
as 10 de maior VI em pelo menos 50% das amostras, que são as mesmas mencionadas para DAP ≥
5cm mais Chorisia speciosa e Trichilia clausseni.

187
Tabela 5: Comparação entre as 10 famílias de maior valor de importância (VI) entre os fragmentos estudados na região da APA municipal de Campinas, SP, para os dois critérios de inclusão considerados DAP ≥ 5cm e DAP ≥ 10cm F1 F2
5cm 10cm 5cm 10cm Famílias VI Famílias VI Famílias VI Famílias VI
Meliaceae 37,3 Meliaceae 33,8 Rutaceae 86,0 Rutaceae 76,1
Euphorbiaceae 28,0 Apocynaceae 27,2 Euphorbiaceae 44,4 Euphorbiaceae 53,7
Mimosaceae 26,1 Euphorbiaceae 25,9 Myrtaceae 23,9 Mimosaceae 23,1
Apocynaceae 23,8 Mimosaceae 25,8 Mimosaceae 17,7 Lauraceae 16,3
Fabaceae 22,3 Fabaceae 24,2 Lauraceae 15,4 Myrtaceae 15,9
Moraceae 19,6 Moraceae 18,6 Anacardiaceae 15,2 Flacourtiaceae 12,0
Phytollacaceae 19,1 Rutaceae 17,5 Fabaceae 12,5 Fabaceae 11,3
Rutaceae 17,4 Phytollacaceae 14,8 Flacourtiaceae 10,4 Apocynaceae 10,5
Myrtaceae 16,5 Lauraceae 14,7 Apocynaceae 8,8 Anacardiaceae 10,4
Lauraceae 15,6 Caesalpiniaceae 14,3 Meliaceae 7,7 Caesalpiniaceae 9,4 F3 F4
5cm 10cm 5cm 10cm Famílias VI Famílias VI Famílias VI Famílias VI
Meliaceae 124,6 Meliaceae 115,2 Mimosaceae 37,4 Mimosaceae 53,2
Fabaceae 46,3 Fabaceae 52,0 Fabaceae 30,6 Fabaceae 30,4
Bombacaceae 44,0 Bombacaceae 45,1 Flacourtiaceae 30,2 Euphorbiaceae 25,2
Moraceae 11,3 Moraceae 12,9 Euphorbiaceae 27,1 Flacourtiaceae 25,1
Anacardiaceae 10,8 Flacourtiaceae 11,6 Meliaceae 24,9 Lauraceae 22,6
Caesalpiniaceae 10,7 Anacardiaceae 10,7 Lauraceae 21,7 Meliaceae 20,8
Flacourtiaceae 10,0 Caesalpiniaceae 8,2 Caesalpiniaceae 17,1 Caesalpiniaceae 16,8
Ebenaceae 5,4 Urticaceae 5,5 Myrtaceae 16,2 Anacardiaceae 12,4
Myrtaceae 4,7 Ebenaceae 5,5 Anacardiaceae 12,7 Myrtaceae 11,3
Lecythidaceae 4,1 Lecythidaceae 5,5 Apocynaceae 9,6 Apocynaceae 11,0

188
Tabela 5: Continuação
F5 F6
5cm 10cm 5cm 10cm Famílias VI Famílias VI Famílias VI Famílias VI
Fabaceae 59,3 Fabaceae 67,5 Rutaceae 57,6 Rutaceae 45,2
Lauraceae 29,0 Lauraceae 26,5 Euphorbiaceae 28,5 Fabaceae 26,5
Myrtaceae 22,5 Myrtaceae 22,8 Fabaceae 25,8 Euphorbiaceae 22,9
Euphorbiaceae 18,7 Euphorbiaceae 22,8 Meliaceae 22,8 Mimosaceae 21,4
Caesalpiniaceae 18,6 Caesalpiniaceae 19,8 Mimosaceae 18,7 Lecythidaceae 17,1
Mimosaceae 16,4 Phytollacaceae 18,3 Lauraceae 15,5 Meliaceae 16,2
Phytollacaceae 15,8 Mimosaceae 17,1 Bombacaceae 13,3 Bombacaceae 16,1
Flacourtiaceae 14,5 Flacourtiaceae 12,8 Anacardiaceae 12,6 Lauraceae 15,2
Meliaceae 13,1 Meliaceae 9,0 Caesalpiniaceae 11,3 Anacardiaceae 14,8
Moraceae 8,5 Apocynaceae 6,8 Lecythidaceae 11,2 Apocynaceae 13,4 F7 C1
5cm 10cm 5cm 10cm Famílias VI Famílias VI Famílias VI Famílias VI
Rutaceae 56,3 Rutaceae 49,3 Rutaceae 47,3 Rutaceae 47,8
Apocynaceae 43,2 Apocynaceae 46,9 Euphorbiaceae 33,3 Apocynaceae 32,4
Phytollacaceae 32,3 Phytollacaceae 40,9 Apocynaceae 32,8 Euphorbiaceae 26,6
Caesalpiniaceae 26,0 Caesalpiniaceae 27,1 Meliaceae 25,1 Fabaceae 23,1
Urticaceae 25,3 Meliaceae 18,3 Myrtaceae 19,9 Anacardiaceae 20,6
Meliaceae 17,1 Urticaceae 14,8 Anacardiaceae 18,4 Meliaceae 19,4
Lecythidaceae 13,7 Fabaceae 13,7 Caesalpiniaceae 17,3 Myrtaceae 17,2
Moraceae 13,2 Moraceae 12,1 Fabaceae 16,2 Mimosaceae 14,7
Fabaceae 10,7 Bombacaceae 11,7 Bombacaceae 14,1 Bombacaceae 14,3
Bombacaceae 9,1 Lecythidaceae 11,1 Mimosaceae 13,0 Caesalpiniaceae 13,5
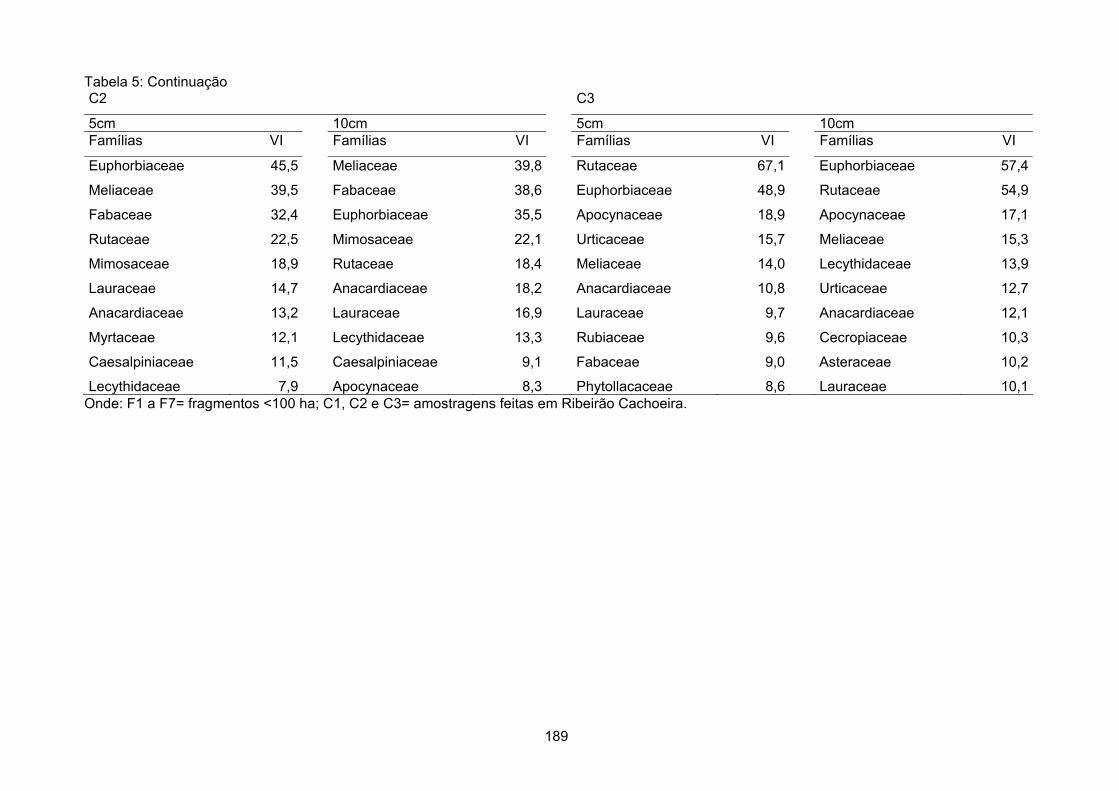
189
Tabela 5: Continuação C2 C3
5cm 10cm 5cm 10cm Famílias VI Famílias VI Famílias VI Famílias VI
Euphorbiaceae 45,5 Meliaceae 39,8 Rutaceae 67,1 Euphorbiaceae 57,4
Meliaceae 39,5 Fabaceae 38,6 Euphorbiaceae 48,9 Rutaceae 54,9
Fabaceae 32,4 Euphorbiaceae 35,5 Apocynaceae 18,9 Apocynaceae 17,1
Rutaceae 22,5 Mimosaceae 22,1 Urticaceae 15,7 Meliaceae 15,3
Mimosaceae 18,9 Rutaceae 18,4 Meliaceae 14,0 Lecythidaceae 13,9
Lauraceae 14,7 Anacardiaceae 18,2 Anacardiaceae 10,8 Urticaceae 12,7
Anacardiaceae 13,2 Lauraceae 16,9 Lauraceae 9,7 Anacardiaceae 12,1
Myrtaceae 12,1 Lecythidaceae 13,3 Rubiaceae 9,6 Cecropiaceae 10,3
Caesalpiniaceae 11,5 Caesalpiniaceae 9,1 Fabaceae 9,0 Asteraceae 10,2
Lecythidaceae 7,9 Apocynaceae 8,3 Phytollacaceae 8,6 Lauraceae 10,1 Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.

190
Tabela 6: Comparação entre as 10 espécies de maior valor de importância (VI) entre os fragmentos estudados na região da APA municipal de Campinas, SP, para os dois critérios de inclusão considerados DAP ≥ 5cm e DAP ≥ 10cm F1 F2
5cm 10cm 5cm 10cm Espécies VI Espécies VI Espécies VI Espécies VI
Piptadenia gonoacantha 22,2 Piptadenia gonoacantha 22,4 Esenbeckia leiocarpa 46,9 Esenbeckia leiocarpa 48,3
Trichilia clausseni 21,2 Trichilia clausseni 22,1 Metrodoria stipularis 20,1 Croton floribundus 25,3
Aspidosperma polyneuron 18,0 Aspidosperma polyneuron 19,4 Galipia multiflora 20,0 Metrodoria stipularis 25,0
Ficus glabra 13,1 Croton floribundus 13,1 Croton floribundus 17,1 Piptadenia gonoacantha 16,7
Gallesia integrifolia 12,8 Ficus glabra 11,6 Calycorectes acutatus 12,0 Mabea fistulifera 16,6
Holocalyx balansae 9,2 Holocalyx balansae 10,5 Mabea fistulifera 12,0 Aspidosperma polyneuron 10,2
Croton floribundus 8,1 Gallesia integrifolia 9,8 Piptadenia gonoacantha 11,5 Calycorectes acutatus 9,1
Myroxylon peruiferum 7,5 Myroxylon peruiferum 7,2 Astronium graveolens 10,4 Astronium graveolens 7,2
Savia dictyocarpa 6,6 Lonchocarpus muehlbergianus 6,6 Aspidosperma polyneuron 8,3 Cryptocarya aschersoniana 6,9
Sebastiania edwaliana 6,5 Aspidosperma ramiflorum 6,5 Casearia sylvestris 5,6 Casearia sylvestris 6,3 F3 F4
5cm 10cm 5cm 10cm Espécies VI Espécies VI Espécies VI Espécies VI
Trichilia clausseni 105,8 Trichilia clausseni 99,9 Piptadenia gonoacantha 28,7 Piptadenia gonoacantha 40,1
Chorisia speciosa 39,6 Chorisia speciosa 39,5 Casearia sylvestris 26,1 Casearia sylvestris 21,9
Lonchocarpus muehlbergianus 32,6 Lonchocarpus muehlbergianus 35,1 Croton floribundus 19,0 Croton floribundus 19,9
Trichilia catigua 11,9 Maclura tinctoria 11,6 Cabralea canjerana 13,8 Cabralea canjerana 12,8
Astronium graveolens 10,3 Astronium graveolens 10,2 Astronium graveolens 11,1 Astronium graveolens 10,1
Maclura tinctoria 10,0 Trichilia hirta 9,6 Copaifera langsdorffi 8,2 Centrolobium tomentosum 8,5
Trichilia hirta 9,8 Trichilia catigua 8,7 Machaerium nictitans 8,1 Acacia paniculata 7,8
Myroxylon peruiferum 6,8 Casearia sylvestris 8,2 Centrolobium tomentosum 7,9 Copaifera langsdorffi 7,2
Casearia sylvestris 6,2 Centrolobium tomentosum 7,6 Ocotea diospirifolia 5,6 Machaerium nictitans 6,6
Hymenaea courbaril 5,5 Myroxylon peruiferum 6,5 Eugenia excelsa 5,4 Matayba elaeagnoides 6,2

191
Tabela 6: Continuação
F5 F6
5cm 10cm 5cm 10cm Espécies VI Espécies VI Espécies VI Espécies VI
Machaerium nictitans 21,4 Lonchocarpus muehlbergianus 25,3 Esenbeckia leiocarpa 23,6 Esenbeckia leiocarpa 30,4
Lonchocarpus muehlbergianus 21,3 Machaerium nictitans 20,8 Almeidea coerulea 16,9 Croton floribundus 14,9
Nectandra megapotamica 20,4 Croton floribundus 20,6 Croton floribundus 15,2 Astronium graveolens 14,5
Croton floribundus 16,8 Gallesia integrifolia 17,1 Astronium graveolens 12,2 Chorisia speciosa 14,4
Gallesia integrifolia 14,6 Nectandra megapotamica 16,0 Chorisia speciosa 12,1 Cariniana legalis 14,1
Casearia sylvestris 11,9 Bauhinia forficata 13,3 Erythrina falcata 11,3 Aspidosperma polyneuron 10,4
Bauhinia forficata 11,6 Calycorectes acutatus 10,8 Galipia multiflora 9,6 Piptadenia gonoacantha 8,5
Calycorectes acutatus 9,4 Casearia sylvestris 10,6 Trichilia elegans 9,6 Galipia multiflora 7,7
Piptadenia gonoacantha 7,6 Piptadenia gonoacantha 9,3 Cariniana legalis 8,7 Holocalyx balansae 7,5
Cabralea canjerana 7,6 Lonchocarpus cultratus 8,9 Nectandra megapotamica 8,4 Nectandra megapotamica 7,4 F7 C1
5cm 10cm 5cm 10cm Espécies VI Espécies VI Espécies VI Espécies VI
Gallesia integrifolia 31,1 Gallesia integrifolia 39,2 Esenbeckia leiocarpa 26,8 Esenbeckia leiocarpa 36,2
Aspidosperma polyneuron 29,9 Esenbeckia leiocarpa 37,3 Aspidosperma polyneuron 22,2 Aspidosperma polyneuron 23,8
Esenbeckia leiocarpa 25,0 Aspidosperma polyneuron 35,1 Astronium graveolens 17,8 Astronium graveolens 20,2
Urera baccifera 24,3 Holocalyx balansae 21,2 Chorisia speciosa 11,3 Savia dictyocarpa 13,8
Holocalyx balansae 21,3 Urera baccifera 14,4 Trichilia clausseni 11,0 Calycorectes acutatus 12,4
Metrodoria nigra 16,2 Aspidosperma ramiflorum 13,0 Holocalyx balansae 10,5 Chorisia speciosa 10,4
Aspidosperma ramiflorum 14,0 Chorisia speciosa 11,5 Sebastiania edwaliana 10,4 Trichilia clausseni 9,5
Almeidea coerulea 13,7 Metrodoria nigra 10,1 Aspidosperma ramiflorum 8,6 Centrolobium tomentosum 9,5
Ficus glabra 12,0 Ficus glabra 9,9 Trichilia catigua 8,6 Croton floribundus 9,0
Chorisia speciosa 8,9 Trichilia clausseni 8,3 Savia dictyocarpa 8,2 Holocalyx balansae 8,5

192
Tabela 6: Continuação
C2 C3
5cm 10cm 5cm 10cm Espécies VI Espécies VI Espécies VI Espécies VI
Lonchocarpus muehlbergianus 18,1 Trichilia clausseni 22,6 Esenbeckia leiocarpa 33,3 Esenbeckia leiocarpa 41,8
Sebastiania edwaliana 17,2 Lonchocarpus muehlbergianus 19,7 Croton floribundus 25,1 Croton floribundus 29,7
Trichilia clausseni 16,9 Astronium graveolens 17,4 Almeidea coerulea 21,9 Savia dictyocarpa 15,2
Croton priscus 13,7 Croton priscus 12,6 Urera baccifera 15,0 Cariniana legalis 13,7
Astronium graveolens 12,9 Piptadenia gonoacantha 10,4 Astronium graveolens 10,4 Urera baccifera 12,3
Trichilia catigua 9,9 Acacia polyphylla 9,3 Savia dictyocarpa 9,7 Astronium graveolens 11,8
Acacia polyphylla 9,0 Cariniana estrellensis 8,0 Aspidosperma polyneuron 9,0 Vernonia diffusa 9,9
Piptadenia gonoacantha 7,9 Centrolobium tomentosum 8,0 Aspidosperma ramiflorum 8,6 Cecropia hololeuca 9,1
Holocalyx balansae 7,7 Croton floribundus 7,9 Galipia multiflora 8,1 Chorisia speciosa 8,1
Croton floribundus 7,5 Aspidosperma polyneuron 7,1 Vernonia diffusa 7,7 Aspidosperma ramiflorum 8,0 Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
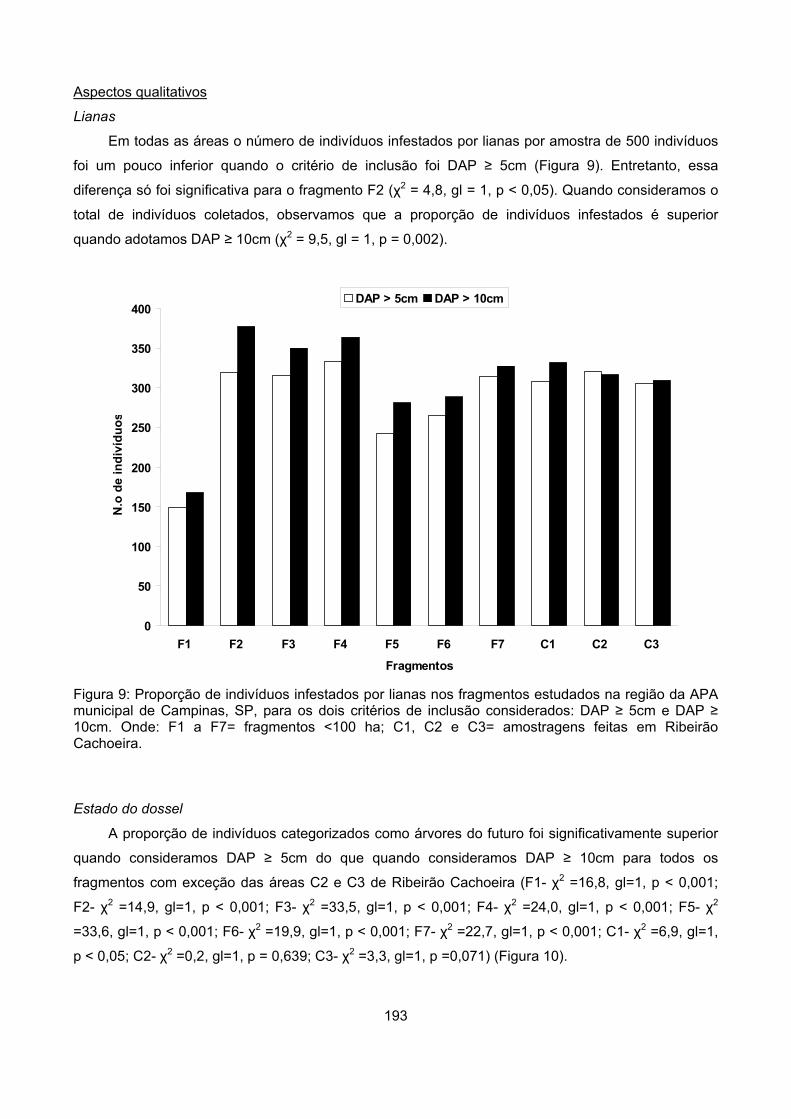
193
Aspectos qualitativos
Lianas
Em todas as áreas o número de indivíduos infestados por lianas por amostra de 500 indivíduos
foi um pouco inferior quando o critério de inclusão foi DAP ≥ 5cm (Figura 9). Entretanto, essa
diferença só foi significativa para o fragmento F2 (χ2 = 4,8, gl = 1, p < 0,05). Quando consideramos o
total de indivíduos coletados, observamos que a proporção de indivíduos infestados é superior
quando adotamos DAP ≥ 10cm (χ2 = 9,5, gl = 1, p = 0,002).
Figura 9: Proporção de indivíduos infestados por lianas nos fragmentos estudados na região da APA municipal de Campinas, SP, para os dois critérios de inclusão considerados: DAP ≥ 5cm e DAP ≥ 10cm. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
Estado do dossel
A proporção de indivíduos categorizados como árvores do futuro foi significativamente superior
quando consideramos DAP ≥ 5cm do que quando consideramos DAP ≥ 10cm para todos os
fragmentos com exceção das áreas C2 e C3 de Ribeirão Cachoeira (F1- χ2 =16,8, gl=1, p < 0,001;
F2- χ2 =14,9, gl=1, p < 0,001; F3- χ2 =33,5, gl=1, p < 0,001; F4- χ2 =24,0, gl=1, p < 0,001; F5- χ2
=33,6, gl=1, p < 0,001; F6- χ2 =19,9, gl=1, p < 0,001; F7- χ2 =22,7, gl=1, p < 0,001; C1- χ2 =6,9, gl=1,
p < 0,05; C2- χ2 =0,2, gl=1, p = 0,639; C3- χ2 =3,3, gl=1, p =0,071) (Figura 10).
0
50
100
150
200
250
300
350
400
F1 F2 F3 F4 F5 F6 F7 C1 C2 C3
Fragmentos
N.o
de
indi
vídu
os
DAP > 5cm DAP > 10cm

194
A proporção de indivíduos categorizados como árvores do presente foi superior quando
consideramos DAP ≥ 10cm do que quando consideramos DAP ≥ 5cm nos fragmentos F3 (χ2 =4,0,
gl=1, p < 0,05), F4 (χ2 =4,1, gl=1, p < 0,05) e F5 (χ2 =7,8, gl=1, p < 0,05). Somente no fragmento F7 a
proporção de árvores do passado foi significativamente superior para DAP ≥ 10cm do que para DAP
≥ 5cm (χ2 =8,7, gl=1, p < 0,05).
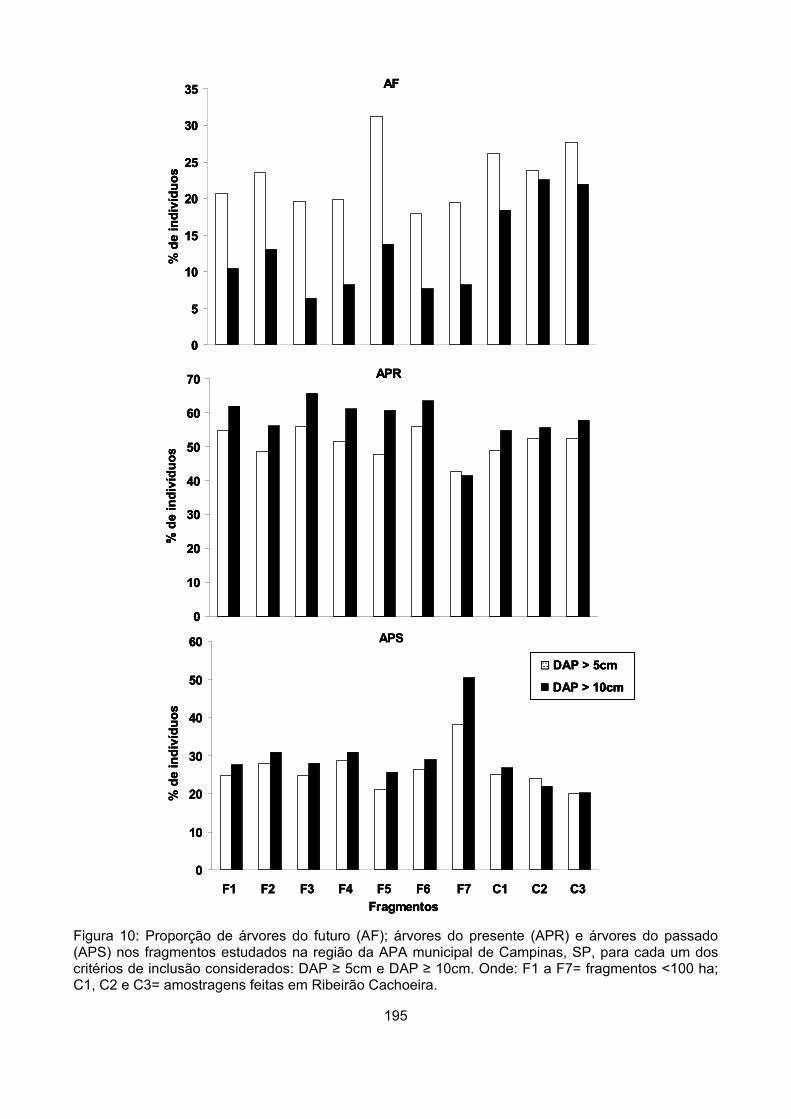
195
Figura 10: Proporção de árvores do futuro (AF); árvores do presente (APR) e árvores do passado (APS) nos fragmentos estudados na região da APA municipal de Campinas, SP, para cada um dos critérios de inclusão considerados: DAP ≥ 5cm e DAP ≥ 10cm. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.
AF
0
5
10
15
20
25
30
35
% d
e in
diví
duos
APR
0
10
20
30
40
50
60
70
% d
e in
diví
duos
APS
0
10
20
30
40
50
60
F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 Fragmentos
% d
e in
diví
duos
. . DAP > 5cm
DAP > 10cm
. .
AF
0
5
10
15
20
25
30
35
% d
e in
diví
duos
APR
0
10
20
30
40
50
60
70
% d
e in
diví
duos
APS
0
10
20
30
40
50
60
F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 Fragmentos
% d
e in
diví
duos
AF
0
5
10
15
20
25
30
35
% d
e in
diví
duos
APR
0
10
20
30
40
50
60
70
% d
e in
diví
duos
APS
0
10
20
30
40
50
60
F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 Fragmentos
% d
e in
diví
duos
. . DAP > 5cm
DAP > 10cm
. .. . DAP > 5cm
DAP > 10cm
. . DAP > 5cm
DAP > 10cm
. .

196
Guildas de espécies
Não foram observadas diferenças significativas entre a proporção de espécies das diferentes
síndromes de dispersão entre os dois conjuntos de dados (Qui-quadrado, p < 0,05) (Tabela 7).
Quando consideramos a proporção de indivíduos das diferentes síndromes, observamos que a
proporção de indivíduos anemocóricos foi superior nos fragmentos F6 (χ2 =9,9, gl=1, p < 0,05), F7 (χ2
=6,5, gl=1, p < 0,05) e área C1 de Ribeirão Cachoeira (χ2 =4,5, gl=1, p < 0,05) quando consideramos
DAP ≥ 10cm. A proporção de indivíduos autocóricos foi maoir para DAP ≥ 5cm nas áreas F6 (χ2 =5,1,
gl=1, p < 0,05) e C1 (χ2 =4,2, gl=1, p < 0,05) (Tabela 7). A proporção de indivíduos zoocóricos não
diferiu entre os dois conjuntos de dados em nenhum dos fragmentos.
Tabela 7: Porcentagem de espécies e indivíduos dos diferentes métodos de dispersão nos fragmentos estudados na região da APA de Sousas e J. Egídio. Campinas. SP, para os dois critérios de inclusão considerados: DAP ≥ 5cm e DAP ≥ 10cm.
Anemocóricas Autocóricas Zoocóricas Locais
5cm 10cm 5cm 10cm 5cm 10cm
Espécies
F1 17,5 18,4 18,4 17,5 64,0 64,1
F2 19,8 19,0 22,1 21,4 59,3 59,5
F3 34,0 44,7 17,0 10,6 49,1 44,7
F4 21,0 23,9 16,0 15,6 63,9 60,6
F5 20,8 21,2 10,4 11,1 69,8 67,7
F6 22,2 21,2 16,2 16,3 61,6 62,5
F7 25,7 27,7 14,3 15,4 60,0 56,9
C1 23,7 25,0 17,5 15,6 59,8 59,4
C2 22,1 25,5 18,3 16,3 60,6 58,2
C3 22,6 20,0 17,2 18,9 62,4 61,1
Indivíduos
F1 22,4 24,4 21,8 21,2 55,8 54,4
F2 15,1 15,6 49,6 50,1 35,3 34,3
F3 25,8 30,0 2,2 1,2 72,0 68,8
F4 20,8 22,8 22,8 25,3 56,3 51,9
F5 29,8 33,0 15,8 19,0 54,4 48,0
F6 17,4 26,2# 37,4# 29,2 45,2 44,6
F7 28,2 37,4# 29,4 25,0 42,4 37,6
C1 25,1 32,3# 36,5# 29,1 38,5 38,7
C2 23,8 30,2 29,9 23,6 46,3 46,2
C3 16,0 20,6 45,8 43,2 38,2 36,2 Onde: #- proporção significativamente superior; F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira.

197
Discussão Os dois conjuntos de dados dos diferentes critérios de inclusão se sobrepõem e por isso seria
esperado que as variações não fossem muito acentuadas. Apesar disso, foi possível detectar que a
maior parte das variações entre os dois conjuntos de dados parece associada ao porte dos indivíduos
e às diferentes abundâncias que as espécies tendem a apresentar quando adotamos um critério de
inclusão maior ou menor. Algumas espécies de sub-bosque tendem a ser muito abundantes quando
adotamos um critério de inclusão menor, mas elas raramente são amostradas se aumentarmos o
critério de inclusão. Observamos a influência desses fatores em quase todos os parâmetros
analisados. Algumas variações também parecem ter ocorrido ao acaso, principalmente aquelas
relativas à composição florística e parâmetros fitossociológicos, como VI.
A maioria das espécies que ocorreram em apenas um dos conjuntos de dados, apresentaram
um ou dois indivíduos, isto é, ocorreram com baixas densidades. Isto indica que as diferenças de
composição entre os dois conjuntos, dadas por essas espécies, ocorreram ao acaso. Algumas
espécies, por outro lado, ocorreram com altas densidades quando consideramos DAP ≥ 5cm, mas
quase não ocorrem ou ocorrem com densidades baixíssimas quando consideramos DAP ≥ 10cm.
Neste caso, a variação observada se deveu ao pequeno porte dessas espécies, que raramente
atingem diâmetros maiores que 10cm.
O número de espécies coletadas variou pouco entre os dois conjuntos, embora esse valor seja,
na maioria dos casos, superior para DAP ≥ 5cm, observamos que a “perda” de espécies quando
alteramos o critério de inclusão de 5 para 10cm é de apenas 7%.
Em relação à riqueza estimada observamos que, na maioria das áreas, a proporção entre o
número de espécies coletadas e a riqueza estimada foi menor quando consideramos DAP ≥ 10cm do
que quando consideramos DAP ≥ 5cm. Como mencionado anteriormente, uma grande diferença
entre a riqueza estimada e número de espécies coletadas indica uma riqueza elevada para aquela
amostra, uma vez que uma pequena proporção do total de espécies esperado foi coletado. Dessa
forma, nas amostras que indicam maior riqueza para DAP ≥ 10cm do que para DAP ≥ 5cm a
heterogeneidade seria maior quando aumentamos o critério de inclusão. Quando aumentamos o
critério de inclusão de 5 para 10cm a tendência é que haja maior espaçamento entre os indivíduos.
Dessa forma, a distância percorrida para se amostrar cada indivíduo seria maior e nos levaria a
englobar uma maior heterogeneidade ambiental dentro da área, levando por fim, a um maior número
de espécies.
A maioria das variações nos aspectos qualitativos entre os dois conjuntos de dados pode ser
atribuída ao porte dos indivíduos e também à composição e abundância das espécies, que variou
entre os dois conjuntos de dados, principalmente no que se refere às síndromes de dispersão das
espécies. A proporção de indivíduos infestados por lianas foi menor para DAP ≥ 5cm, possivelmente,
porque as árvores que compõem o sub-bosque, mais facilmente amostradas quando consideramos
um critério de inclusão menor, são mais raramente infestadas por lianas. Lianas são plantas helíofitas

198
e raramente se estabelecem em locais de baixa luminosidade. Ocorrem principalmente infestando o
dossel de florestas maduras ou em ambientes mais abertos, como bordas de mata e clareiras (Putz
1984, Laurance et al. 2001). Putz (1984) também observou que as lianas tendem a ser mais
abundantes em árvores de diâmetros maiores e que as plantas do sub-bosque são geralmente muito
finas para que as lianas possam utilizá-las como suporte.
Houve maior proporção de indivíduos categorizados como árvores do futuro para DAP ≥ 5cm
em todas as áreas, com exceção de C2 e C3. Isto ocorreu, provavelmente, porque este critério de
inclusão tende a incluir mais indivíduos jovens das espécies que compõem o dossel do que o critério
de inclusão maior. Tais indivíduos, pela definição adotada, seriam categorizados como árvores do
futuro. Nas áreas C2 e C3, possivelmente esses indivíduos jovens foram mais abundantes do que
nas demais áreas, isso faz com que a probabilidade de serem amostrados pelo critério de inclusão
maior seja superior do que nas outras áreas. A proporção de árvores do presente foi superior quando
consideramos DAP ≥ 10cm, nas áreas F3, F4 e F5. Isto ocorreu, possivelmente, porque quando
utilizamos um critério de inclusão maior, a probabilidade de se amostrar indivíduos adultos, que
seriam categorizados como árvores do presente, é maior. A proporção de árvores do passado diferiu
entre os dois conjuntos de dados somente no fragmento F5. Esta área apresenta um histórico de
perturbação bastante intenso, tendo sido atingida em quase toda a sua extensão por uma queimada
no ano de 1996. Possivelmente, quando consideramos DAP ≥ 5cm, grande parte dos indivíduos
amostrados não apresentavam sinais de deterioração, pois se teriam se estabelecido após a
passagem do fogo. O contrário pode ter ocorrido quando consideramos DAP ≥ 10cm, a maior parte
dos indivíduos amostrados, seriam indivíduos adultos, que sobreviveram à queimada, mas
apresentavam sinais de deterioração devido ao ocorrido e com isso foram classificados como árvores
do passado.
A proporção de indivíduos anemocóricos foi superior para DAP ≥ 10cm nas amostras F6, F7 e
C1. Nas áreas F6 e C1 observamos ainda, que a proporção de indivíduos autocóricos foi maior
quando consideramos DAP ≥ 5cm. Como mencionado anteriormente, este critério de inclusão tende a
amostrar mais indivíduos de espécies que compõem o sub-bosque da floresta. Segundo Leitão Filho
(1982) neste estrato, poucas espécies seriam anemocóricas. Para o autor, a anemocoria tende a ser
mais freqüente em espécies que compõem o dossel ou são emergentes. Como não houve diferanças
entre a proporção de espécies das diferentes síndromes entre os dois conjuntos de dados, é provável
que essas variações tenham ocorrido porque nas áreas F6 e C1 as espécies autocóricas do sub-
bosque seriam mais abundantes que nas demais áreas. Da mesma forma, as espécies anemocóricas
que compõem o dossel ou são emergentes, também seriam mais abundantes nessas áreas, assim
como em F7 e, portanto, seriam mais facilmente amostradas quando utilizamos um critério de
inclusão maior.
Quando comparamos os dois conjuntos de dados, seria esperado que os parâmetros estruturais
variassem, principalmente a densidade, porque a tendência é que um maior número de indivíduos

199
ocorra na classe dos baixos diâmetros. A densidade das populações é freqüentemente
negativamente correlacionada ao tamanho dos indivíduos (Hartshorn 1978, Hubbell & Foster 1990,
Rankin-de-Merona et al. 1990). As densidades diminuíram cerca de 17% quando alteramos o critério
de inclusão de 5 para 10cm.
Houve pouca variação entre as espécies de maior VI quando comparamos os dois critérios de
inclusão considerados. Ocorreram variações de posição no ranking de VI e algumas substituições
poderiam ser explicadas, em grande parte dos casos, pelo porte dos indivíduos de cada espécie.
Quando consideramos DAP ≥ 5cm, espécies de grande porte, que tendem a apresentar altas
dominâncias, são substituídas por outras espécies de pequeno porte, que tendem a ser abundantes
quando consideramos um critério de inclusão menor. Quando consideramos DAP ≥ 10cm acontece o
contrário, espécies de pequeno porte são pouco abundantes e acabam substituídas por espécies de
porte maior no ranking de VI (Tabela 6).
É possível que muitas das variações observadas entre os dois conjuntos de dados para as
variações de estrutura e composição tenham ocorrido devido à grande heterogeneidade ambiental
dentro dessas áreas. Amostras distintas feitas dentro de um mesmo fragmento podem ser tão
heterogêneas entre si quanto amostras feitas em fragmentos diferentes. Cielo Filho (2001) realizou
um estudo fitossociológico na mata Ribeirão Cachoeira, na mesma região onde foi realizada a
amostragem C1. O autor usou o método de parcelas e amostrou indivíduos com DAP ≥ 5cm. Ao
compararmos as espécies de maior VI encontradas por Cielo Filho (2001) com os resultados das
amostras de C1 para DAP ≥ 5cm e DAP ≥ 10cm, observamos seis espécies em comum entre as dez
espécies de maior VI, independentemente do critério de inclusão considerado (Tabela 7). As
semelhanças entre a amostra de DAP ≥ 10cm e o estudo de Cielo Filho (2001) podem indicar que
aumentar o critério de inclusão de 5 para 10cm não produz uma variação tão grande nos resultados
obtidos. Por outro lado, as diferenças entre a amostra de DAP ≥ 5cm e o estudo de Cielo Filho (2001)
indicam grande heterogeneidade na distribuição das espécies na área e chamam atenção para se
considerar os parâmetros como estimativas e não como valores absolutos quando se comparam tais
estudos. Essas comparações mostraram que até mesmo levantamentos feitos em uma mesma
floresta, e usando o mesmo critério de inclusão podem chegar a resultados distintos, confirmando em
parte o comentário de Dislich et al. (2001) de que isso aconteceria se fossem realizadas várias
amostragens dentro de uma mesma floresta, mas usando critérios de inclusão diferentes.
Possivelmente as variações observadas quando se altera o critério de inclusão seriam as mesmas
esperadas ao acaso.
No presente estudo, os resultados indicaram que as variações, quando se alterou o critério de
inclusão de DAP ≥ 5cm para DAP ≥ 10cm foram pequenas e podem ter ocorrido tanto porque a
variação de 5 para 10cm é pequena quanto pela sobreposição dos dois conjuntos. Entretanto, através
das comparações com o estudo de Cielo Filho (2001) mencionadas acima, observamos que essas
variações, além de pequenas, podem ocorrer ao acaso, indicando que as variações observadas

200
quando se comparam trabalhos que utilizaram critérios de inclusão diferentes não poderiam ser
atribuídas simplesmente às diferenças de método e sim a fatores relacionados à heterogeneidade de
distribuição das espécies, o que estaria relacionado à heterogeneidade ambiental e histórico de
preservação, por exemplo.
Tabela 7: Comparação entre as dez espécies de maior VI obtidas no presente estudo na área C1 de Ribeirão Cachoeira nos dois critérios de inclusão considerados e o estudo de Cielo Filho(2001), realizado na mesma área, usando DAP ≥ 5cm.
Ribeirão Cachoeira
(C1- presente estudo)
Ribeirão Cachoeira
(Cielo Filho 2001)
Ribeirão Cachoeira
(C1- presente estudo)
5cm 5cm 10cm
Aspidosperma polyneuron Actinostemon klotschii Aspidosperma polyneuron
Aspidosperma ramiflorum Aspidosperma polyneuron Astronium graveolens
Astronium graveolens Astronium graveolens Calycorectes acutatus
Chorisia speciosa Croton floribundus Centrolobium tomentosum
Esenbeckia leiocarpa Esenbeckia leiocarpa Chorisia speciosa
Holocalyx balansae Holocalyx balansae Croton floribundus
Savia dictyocarpa Metrodorea nigra Esenbeckia leiocarpa
Sebastiania edwaliana Ocotea beulahiae Holocalyx balansae
Trichilia catigua Savia dictyocarpa Savia dictyocarpa
Trichilia clausseni Trichilia catigua Trichilia clausseni
Obs.: Espécies em ordem alfabética.

201
Referências bibliográficas AYRES, M., AYRES Jr., M., AYRES, D.M. & SANTOS, A.S. 1998. BioEstat: Aplicações estatísticas
nas áreas das Ciências Biológicas e Médicas. Sociedade civil Mamirauá. Manaus.
BAITELLO, J.B., AGUIAR, O.T., ROCHA, F.T., PASTORE J.A., & ESTEVES, R. 1992. Florística e
fitossociologia do estrato arbóreo de um trecho da serra da Cantareira (Núcleo Pinheirinho) - SP
Anais II Congresso Nacional sobre Essências Nativas 291-297.
BERTONI, J.E.A., MARTINS,F.R., MORAES, J.L. & SHEPHERD, G.J. 1988. Composição florística e
estrutura fitossociológica do Parque Estadual de Vaçununga, Santa Rita do Passa Quatro –
Gleba Praxedes, Boletim Técnico do Instituto Florestal de São Paulo 42: 149-170.
CAVASSAN, O., CESAR, O. & MARTINS, F.R. 1984. Fitossociologia da vegetação arbórea da
reserva estadual de Bauru, Estado de São Paulo. Revista Brasileira de Botânica 7: 91-106.
CHAO, A. 1984. Non-parametric estimation of the number of classes in a population. Scandinavian
Journal of Statistics 11: 265-270.
CHAO, A. 1987. Estimating the population size for capture-recapture data with inequal catchability.
Biometrics 43: 783-791.
CIELO FILHO, R. 2001. Estrutura e abundância de um trecho de floresta estacional semidecídua no município de Campinas, estado de São Paulo: Mata Ribeirão Cachoeira.
Dissertação de Mestrado. Universidade Estadual de Campinas. Campinas, SP.
COLWELL, R. K. & CODDINGTON, J. A. 1994. Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions: Biological Sciences 345: 101-118.
COSTA, L.G.S. & MANTOVANI, W. 1995. Flora arbustivo-arbórea de trecho de mata mesófila
semidecídua, no sítio ecológico de Ibicatu, Piracicaba (SP). Hoehnea 22: 47-59.
COTTAN, G. & CURTIS, J.T. 1956. The use of distance measures in phytossociological sampling.
Ecology 37: 451-460.
CRONQUIST, A. 1981. An integrated system of classification of flowering plants. NewYork:
Columbia University Press & The New York Botanical Garden.
CURTIS, J.T. & McINTOSH, R.P. 1951. An upland forest continuum in the prairie-forest border region
of Wiscosin. Ecology 32: 476-496.
DENSLOW, J.S. 1995. Disturbance and diversity in tropical rain forests: The density effect.
Ecological Aplications 5: 962-968.
DISLICH, R., CERSÓSIMO, L. & MANTOVANI, W. 2001. Análise da estrutura de fragmentos
florestais no planalto paulistano - SP. Revista Brasileira de Botânica 24: 321-332.
DURIGAN, G., FRANCO, G.A.D.C., SAITO, M. & BAITELLO, J.B. 2000. Estrutura e diversidade do
componente arbóreo da floresta na Estação Ecológica dos Caetetus, Gália, SP. Revista Brasileira de Botânica 23: 371-383.

202
FERREIRA, L.V. & PRANCE, G.T. 1998. Species richness and floristic composition in four hectares in
the Jaú National Park in upland forests in Central Amazonia. Biodiversity and Conservation 7:
1349-1364.
GOTELLI, N. J. & COWELLl, R. K. 2001. Quantifying biodiversity: procedures and pitfalls in the
measurement and comparison of species richness. Ecological Letters 4: 379-391.
GROMBONE, M.T., BERNACCI,L.C., MEIRA NETO,J.A.A., TAMASHIRO,J.Y. & LEITÃO FILHO, H.
1990. Estrutura fitossociológica da floresta semidecídua de altitude do Parque Municipal de Grota
Funda (Atibaia-Estado de São Paulo). Acta Botanica Brasilica 4: 47-64.
HAMMER, O., HARPER, D.A.T. & RYAN, P.D. 2001. PAST: Paleontological Statistics Software
Package for Education and Data Analysis. Palaeontologia Electronica 4: 9. http://palaeo-
electronica.org/2001_1/past/issue1_01.htm
HARTSHORN, G.S. 1978. Treefalls and tropical forest dynamics. Pp 617-638. In: P.B. Tonlinson &
M.H. Zimmerman (eds.). Tropical trees as living systems. Cambridge University Press, New
York.
HUBBELL, S.P. & FOSTER, R.B. Diversity of canopy trees in a neotropical forest and implications for
conservation. pp 25-41. In: S.L. Sutton, T.C. Whitmore & A. C. Chadwick, (eds).Tropical Rain forest: Ecology and Management. Blackwell Scientific Publications, Oxford.
KÖPPEN, W. 1948. Climatologia. México - Buenos Aires. Ed. Fundo de Cultura Econômica, 478p.
LAURANCE, W.F., PÉREZ-SALICRUP, D., DELAMÔNICA, P., FEARNSIDE, P.M., D’ANGELO, S.,
JEROLINSKI, A ., POHL, L. & LOVEJOY, T.E. 2001. Rain forest fragmentation and structure of
amazonian liana communities. Ecology 82: 105-116.
LEITÃO FILHO, H.F. 1982. Aspectos taxonômicos das florestas do Estado de São Paulo. Silvicultura em São Paulo 16: 197-206.
LUDWIG, J.A. & REYNOLDS, J.F. 1988. Statistical Ecology. A primer on methods and computing. J. Wiley & Sons, New York.
MAGURRAN, A. 1988. Ecological diversity and its measurement. Croom Helm, London
MANTOVANI, W. & MARTINS, F.R. 1988. Variações fenológicas das espécies do cerrado da reserva
biológica de Mogi-Guaçu, estado de São Paulo. Revista Brasileira de Botânica 11: 101-112.
MARTINS, F.R. 1991. Estrutura de uma floresta mesófila. Editora da Universidade Estadual de
Campinas. Campinas.
MATTHES, L.A.F., LEITÃO FILHO, H.F. & MARTINS, F.R. 1988. Bosque dos Jequitibás (Campinas,
SP): composição florística e fitossociológica do estrato arbóreo. Anais V congresso da Sociedade Botânica de São Paulo, p. 55-76.
MELLO, M.H.A., PEDRO JUNIOR, M.J., ORTOLANI, A. A. & ALFONSI R.R. 1994. Chuva e
Temperatura: cem anos de observações em Campinas. Boletim Técnico 154 . Instituto
Agronômico Campinas, SP.

203
MÜLLER-DOMBOIS, D. & ELLENBERG, H. 1974. Aims and methods of Vegetation Ecology. New
York , John Wiley & Sons.
OLDEMAN R.A.A. 1989. Dynamics in tropical forests. pp 3-21. In: Holm-Nielsen, L.B., Nielsen, I.C. &
Balslev, H. 1989. Tropical Forest: Botanical Dynamics, Speciation and Diversity. Academic
Press.
OLIVEIRA, J.B., MARCELO, N.C., ROSSI, M. & CALDERANO FILHO, B. 1999. Mapa pedológico do
estado de São Paulo. Legenda expandida In: OLIVEIRA, J.B. 1999. Boletim científico n.o 45,
Solos do estado de São Paulo. Embrapa e IAC. Campinas.
PIELOU, E.C. 1984. The interpretation of ecological data. Jonh Wiley & Sons, New York.
PUTZ, F.E. 1984. The natural history of lianas on Barro Colorado Island, Panama. Ecology 65: 1713-
1724.
RODRIGUES, R. R., MORELLATO, L.P.C., JOLY, C.A. & LEITÃO FILHO, H.F. 1989. Estudo florístico
e fitossociológico em um gradiente altitudinal de mata estacional mesófila semidecídua, na Serra
do Japi, Jundiaí, SP. Revista Brasileira de Botânica 12: 71-84.
ROSSI, L. 1994. A flora arbórea-arbustiva da mata da reserva da Cidade Universitária “Armando de
Salles Oliveira” (São Paulo, Brasil) Boletim do Instituto de Botânica n.º 9. Instituto de Botânica,
São Paulo.
SALIS, S.M. TAMASHIRO, J.Y. JOLY, C.A. 1994. Florística e fitossociologia do estrato arbóreo de um
remanescente de mata ciliar no Rio Jacaré Pepira, Brotas, SP. Revista Brasileira de Botânica
17: 93-103.
SALIS, S.M., SHEPHERD, G.J. & JOLY, C.A. 1995. Floristic comparison of mesophytic
semideciduous forests of the interior of the state of São Paulo, Southeast Brazil. Vegetatio 119:
155-164.
SANTOS, K. & KINOSHITA, L.S. 2003. Flora arbustivo-arbórea do fragmento de floresta estacional
semidecidual do Ribeirão Cachoeira, Município de Campinas - SP. Acta Botanica Brasilica 17:
325-341.
SHEPHERD, G.J. 1988. FITOPAC- versão preliminar. Manual do usuário. Departamento de
Botânica. Instituto de Biologia, Universidade Estadual de Campinas, Campinas-SP.
TORRES, R.B., MARTINS, F.R. & KINOSHITA, L.S. 1997. Climate, soil and tree flora relationships in
forests in the state of São Paulo, southeastern Brasil. Revista Brasileira de Botânica 20: 41-51.
VELOSO, H.P., RANGEL FILHO, A .L.R. & LIMA, J.C. 1991. Classificação da vegetação brasileira adaptada a um sistema universal. Rio de Janeiro. Instituto Brasileiro de Geografia e Estatística
(IBGE).

204
ANEXO I
Relação das 60 famílias e 278 espécies e morfo-espécies encontradas nos fragmentos estudados na região da APA municipal de Campinas, SP, nos dois critérios de inclusão adotados. Onde: F1 a F7= fragmentos <100 ha; C1, C2 e C3= amostragens feitas em Ribeirão Cachoeira; SD- síndrome de dispersão; NC- número do coletor; + = indica ocorrência da espécie no conjunto DAP ≥ 5cm; x = indica ocorrência da espécie no conjunto DAP ≥ 10cm; +x = indica a ocorrência da espécie em ambos os conjuntos de dados; o = espécies amostradas em coletas aleatóreas; a = espécies anemocóricas; u = espécies autocóricas; z = espécies zoocóricas; - = não classificada. Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC**
ANACARDIACEAE Astronium graveolens Jacq. +x +x +x +x +x +x +x +x +x +x a 3836
Lithraea molleoides (Vell.) Engl. +x z 2587
Tapirira obtusa (Benth.) Mitchell +x +x +x x z 609
ANNONACEAE Annona cacans Warm. +x +x +x +x +x +x +x z 413
Duguetia lanceolata A.St.-Hil. +x z 2709
Guatteria nigrescens Mart. +x +x +x x +x + z 4054
Rollinia sylvatica (A.St.-Hil.) Mart. +x +x +x + +x +x +x +x z 4155
Xylopia brasiliensis Spreng. +x +x z 568
APOCYNACEAE Aspidosperma cylindrocarpon Müll. Arg. +x a 383
Aspidosperma olivaceum Müll. Arg. +x +x +x +x a 4055
Aspidosperma polyneuron Müll. Arg. +x +x +x +x +x +x +x +x +x a 332
Aspidosperma ramiflorum Müll. Arg. +x +x +x +x +x +x +x +x a 370
Peschiera catharinensis (A.DC.) Miers + z 1081
Rauvolfia sellowi Müll. Arg. x +x + z 1234
AQUIFOLIACEAE Ilex taubertianum Reissek* z 4156
ARALIACEAE Dendropanax cuneatum (C.DC.) Decne. et Planch. x x +x z 1683
Didymopanax cf. calvum (Cham.) Decne et Panch +x x z 757
ARECACEAE Syagrus romanzoffiana (Cham.) Glassm. +x +x +x x +x +x +x +x z 710
Asteraceae Gochnatia polymorpha (Less.) Cabrera +x x a 1007
Vernonia diffusa Less. +x a 1961
Vernonia petiolaris A.D.C.* +x a 2925
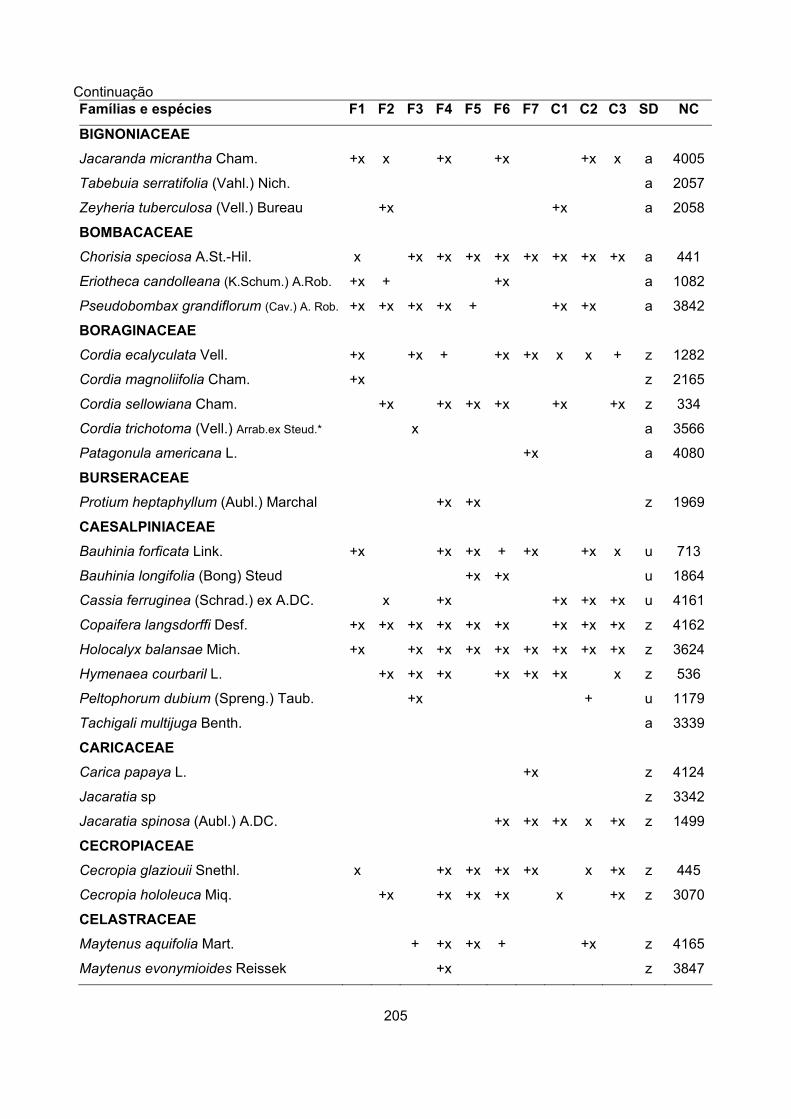
205
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
BIGNONIACEAE Jacaranda micrantha Cham. +x x +x +x +x x a 4005
Tabebuia serratifolia (Vahl.) Nich. a 2057
Zeyheria tuberculosa (Vell.) Bureau +x +x a 2058
BOMBACACEAE Chorisia speciosa A.St.-Hil. x +x +x +x +x +x +x +x +x a 441
Eriotheca candolleana (K.Schum.) A.Rob. +x + +x a 1082
Pseudobombax grandiflorum (Cav.) A. Rob. +x +x +x +x + +x +x a 3842
BORAGINACEAE Cordia ecalyculata Vell. +x +x + +x +x x x + z 1282
Cordia magnoliifolia Cham. +x z 2165
Cordia sellowiana Cham. +x +x +x +x +x +x z 334
Cordia trichotoma (Vell.) Arrab.ex Steud.* x a 3566
Patagonula americana L. +x a 4080
BURSERACEAE Protium heptaphyllum (Aubl.) Marchal +x +x z 1969
CAESALPINIACEAE Bauhinia forficata Link. +x +x +x + +x +x x u 713
Bauhinia longifolia (Bong) Steud +x +x u 1864
Cassia ferruginea (Schrad.) ex A.DC. x +x +x +x +x u 4161
Copaifera langsdorffi Desf. +x +x +x +x +x +x +x +x +x z 4162
Holocalyx balansae Mich. +x +x +x +x +x +x +x +x +x z 3624
Hymenaea courbaril L. +x +x +x +x +x +x x z 536
Peltophorum dubium (Spreng.) Taub. +x + u 1179
Tachigali multijuga Benth. a 3339
CARICACEAE Carica papaya L. +x z 4124
Jacaratia sp z 3342
Jacaratia spinosa (Aubl.) A.DC. +x +x +x x +x z 1499
CECROPIACEAE Cecropia glaziouii Snethl. x +x +x +x +x x +x z 445
Cecropia hololeuca Miq. +x +x +x +x x +x z 3070
CELASTRACEAE Maytenus aquifolia Mart. + +x +x + +x z 4165
Maytenus evonymioides Reissek +x z 3847

206
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
CELASTRACEAE Maytenus floribunda Reissek* +x + z 3004
Maytenus robusta Reissek + +x z 386
Maytenus salicifolia Reissek +x + +x z 2736
CHRYSOBALANACEAE Hirtella hebeclada Moric ex A.DC. x +x x +x z 3680
CLUSIACEAE Vismia brasiliensis Choisy* + z 3964
COMBRETACEAE Terminalia brasiliensis Cambess. a 2073
CONARACEAE Conarus regnelli G.Schellenb. +x +x +x +x +x z 498
EBENACEAE Diospyrus inconstans Jacq. +x +x +x +x z 2741
ELAEOCARPACEAE Sloanea monosperma Vell. +x +x + +x +x z 3351
ERYTHROXYLACEAE Erythroxylum deciduum A.St.-Hil. z 1975
EUPHORBIACEAE Actinostemon communis (Müll. Arg.) Pax + + + + + + + + u 1088
Actinostemon concolor (Spreng.) Müll. Arg. + + + + + u 4015
Actinostemon sp +x u 2171
Alchornea glandulosa Poepp. & Endl. +x x +x +x +x +x +x +x u 4016
Alchornea sidifolia Müll. Arg. x u 4060
Alchornea triplinervea (Spreng.) Müll. Arg. +x +x +x u 667 Aparisthmium sp + - 3626
Croton floribundus Spreng. +x +x +x +x +x +x +x +x +x u 2881
Croton priscus Croizat +x +x +x u 337
Mabea fistulifera Mart. +x u 1089
Margaritaria nobilis L.f. +x x +x +x +x +x u 341
Pachystroma longifolium (Nees) I.L.Johnston +x u 672
Pera glabrata (Schott.) Baill. +x +x +x z 1695
Sapium glandulatum (Vell.) Pax +x +x x z 3753
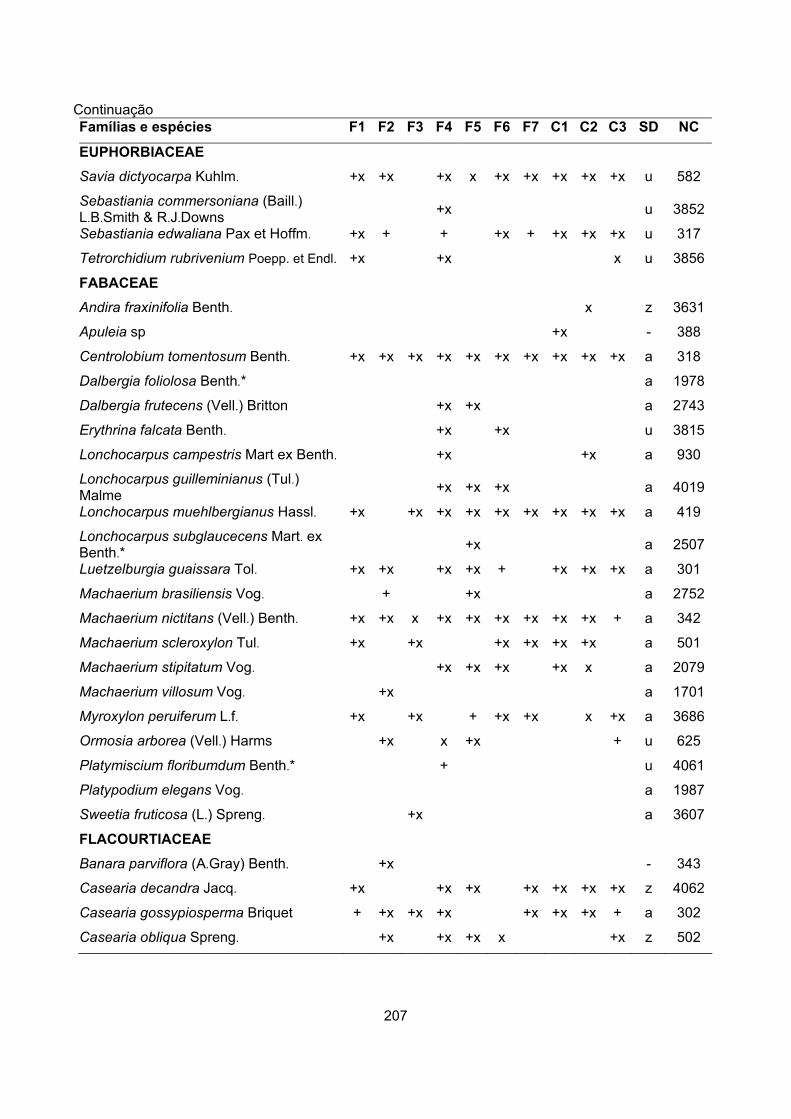
207
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
EUPHORBIACEAE Savia dictyocarpa Kuhlm. +x +x +x x +x +x +x +x +x u 582
Sebastiania commersoniana (Baill.) L.B.Smith & R.J.Downs +x u 3852
Sebastiania edwaliana Pax et Hoffm. +x + + +x + +x +x +x u 317
Tetrorchidium rubrivenium Poepp. et Endl. +x +x x u 3856
FABACEAE Andira fraxinifolia Benth. x z 3631
Apuleia sp +x - 388
Centrolobium tomentosum Benth. +x +x +x +x +x +x +x +x +x +x a 318
Dalbergia foliolosa Benth.* a 1978
Dalbergia frutecens (Vell.) Britton +x +x a 2743
Erythrina falcata Benth. +x +x u 3815
Lonchocarpus campestris Mart ex Benth. +x +x a 930
Lonchocarpus guilleminianus (Tul.) Malme +x +x +x a 4019
Lonchocarpus muehlbergianus Hassl. +x +x +x +x +x +x +x +x +x a 419
Lonchocarpus subglaucecens Mart. ex Benth.* +x a 2507
Luetzelburgia guaissara Tol. +x +x +x +x + +x +x +x a 301
Machaerium brasiliensis Vog. + +x a 2752
Machaerium nictitans (Vell.) Benth. +x +x x +x +x +x +x +x +x + a 342
Machaerium scleroxylon Tul. +x +x +x +x +x +x a 501
Machaerium stipitatum Vog. +x +x +x +x x a 2079
Machaerium villosum Vog. +x a 1701
Myroxylon peruiferum L.f. +x +x + +x +x x +x a 3686
Ormosia arborea (Vell.) Harms +x x +x + u 625
Platymiscium floribumdum Benth.* + u 4061
Platypodium elegans Vog. a 1987
Sweetia fruticosa (L.) Spreng. +x a 3607
FLACOURTIACEAE Banara parviflora (A.Gray) Benth. +x - 343
Casearia decandra Jacq. +x +x +x +x +x +x +x z 4062
Casearia gossypiosperma Briquet + +x +x +x +x +x +x + a 302
Casearia obliqua Spreng. +x +x +x x +x z 502
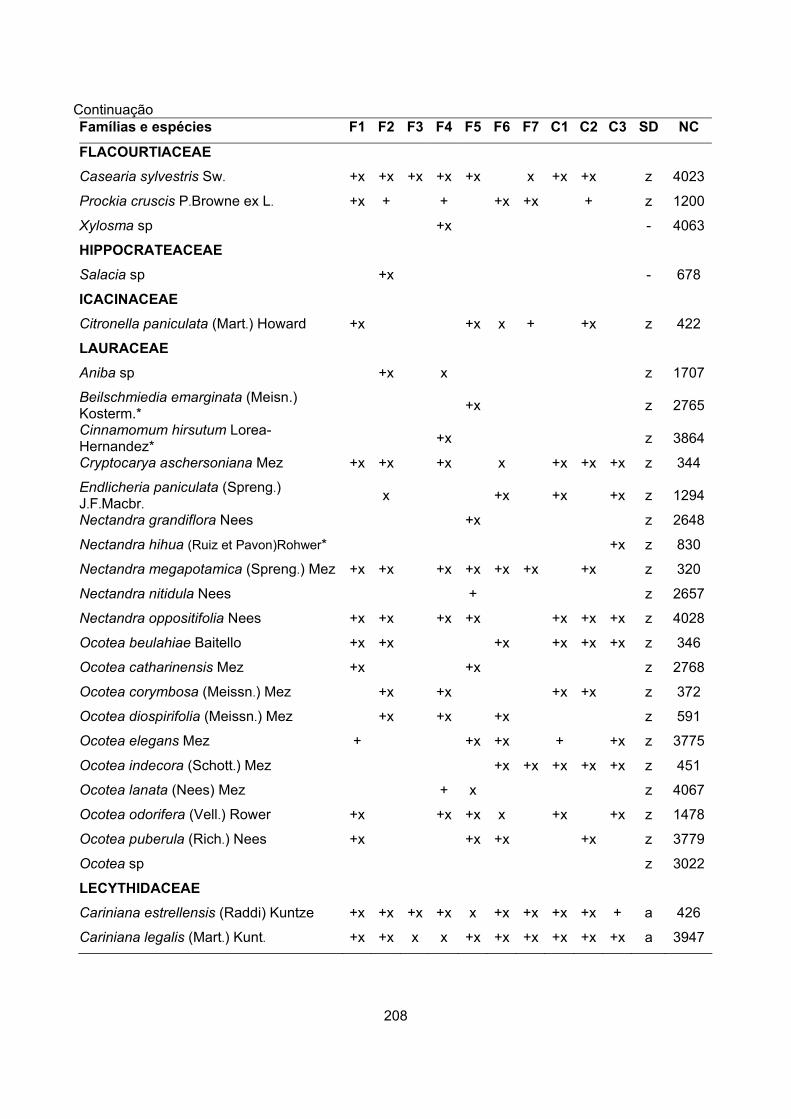
208
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
FLACOURTIACEAE Casearia sylvestris Sw. +x +x +x +x +x x +x +x z 4023
Prockia cruscis P.Browne ex L. +x + + +x +x + z 1200
Xylosma sp +x - 4063
HIPPOCRATEACEAE Salacia sp +x - 678
ICACINACEAE Citronella paniculata (Mart.) Howard +x +x x + +x z 422
LAURACEAE Aniba sp +x x z 1707
Beilschmiedia emarginata (Meisn.) Kosterm.* +x z 2765
Cinnamomum hirsutum Lorea-Hernandez* +x z 3864
Cryptocarya aschersoniana Mez +x +x +x x +x +x +x z 344
Endlicheria paniculata (Spreng.) J.F.Macbr. x +x +x +x z 1294
Nectandra grandiflora Nees +x z 2648
Nectandra hihua (Ruiz et Pavon)Rohwer* +x z 830
Nectandra megapotamica (Spreng.) Mez +x +x +x +x +x +x +x z 320
Nectandra nitidula Nees + z 2657
Nectandra oppositifolia Nees +x +x +x +x +x +x +x z 4028
Ocotea beulahiae Baitello +x +x +x +x +x +x z 346
Ocotea catharinensis Mez +x +x z 2768
Ocotea corymbosa (Meissn.) Mez +x +x +x +x z 372
Ocotea diospirifolia (Meissn.) Mez +x +x +x z 591
Ocotea elegans Mez + +x +x + +x z 3775
Ocotea indecora (Schott.) Mez +x +x +x +x +x z 451
Ocotea lanata (Nees) Mez + x z 4067
Ocotea odorifera (Vell.) Rower +x +x +x x +x +x z 1478
Ocotea puberula (Rich.) Nees +x +x +x +x z 3779
Ocotea sp z 3022
LECYTHIDACEAE Cariniana estrellensis (Raddi) Kuntze +x +x +x +x x +x +x +x +x + a 426
Cariniana legalis (Mart.) Kunt. +x +x x x +x +x +x +x +x +x a 3947

209
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
MALVACEAE Bastardiopsis densiflora (Hook. et Arn.) Radlk. +x +x +x +x +x u 864
MELASTOMATACEAE Miconia cinnamomifolia (Mart.ex A.DC.) Naud. +x z 2773
Miconia pusilliflora Triana +x + z 1517
Mouriri glaziowiana Cogn. +x +x +x z 1643
MELIACEAE Cabralea canjerana (Vell.) Mart. +x +x +x +x +x +x x z 427
Cedrela fissilis Vell. +x +x x +x +x a 1042
Guarea guidonia (L.) Sleumer x z 1043
Guarea kunthiana A. Juss. +x x z 322
Guarea macrophylla Vahl +x +x +x +x +x +x z 4035
Trichilia casarettii C.DC. +x z 3697
Trichilia catigua A.Juss. +x +x +x +x +x +x +x +x +x z 4127
Trichilia clausseni C.DC. +x +x +x + +x +x +x +x +x z 506
Trichilia elegans A.Juss. +x +x +x +x +x +x z 3786
Trichilia hirta L. +x z 3613
Trichilia pallens C.DC.* +x +x +x +x z 324
Trichilia pallida Swartz. +x +x + +x +x +x +x +x +x z 3614
MIMOSACEAE Acacia paniculata Willd. +x +x +x +x +x x +x +x u 305
Acacia polyphylla C.DC. +x + +x +x x x +x u 3549
Anadenanthera colubrina (Vell.) Brenan* +x u 3952
Calliandra sp + +x + +x u 3789
Inga laurina (Sw.) Willd.* z 3187
Inga vera Willd. +x +x +x +x +x +x +x +x z 306
Leucochlorum incuriale (Vell.) Barneby & J.W.Grimes + u 3491
Piptadenia gonoacantha (Mart.) J.F.Macbr. +x +x +x +x +x +x +x +x +x +x u 327
Pithecellobium edwalii Hoehne* +x +x +x x u 3617
MIRYSTICACEAE Virola oleifera (Schott) A.C.Smith* +x z 2219
MONIMIACEAE Mollinedia elegans Tul. + z 3135
Mollinedia widgrenii A.DC. +x + +x +x z 1484
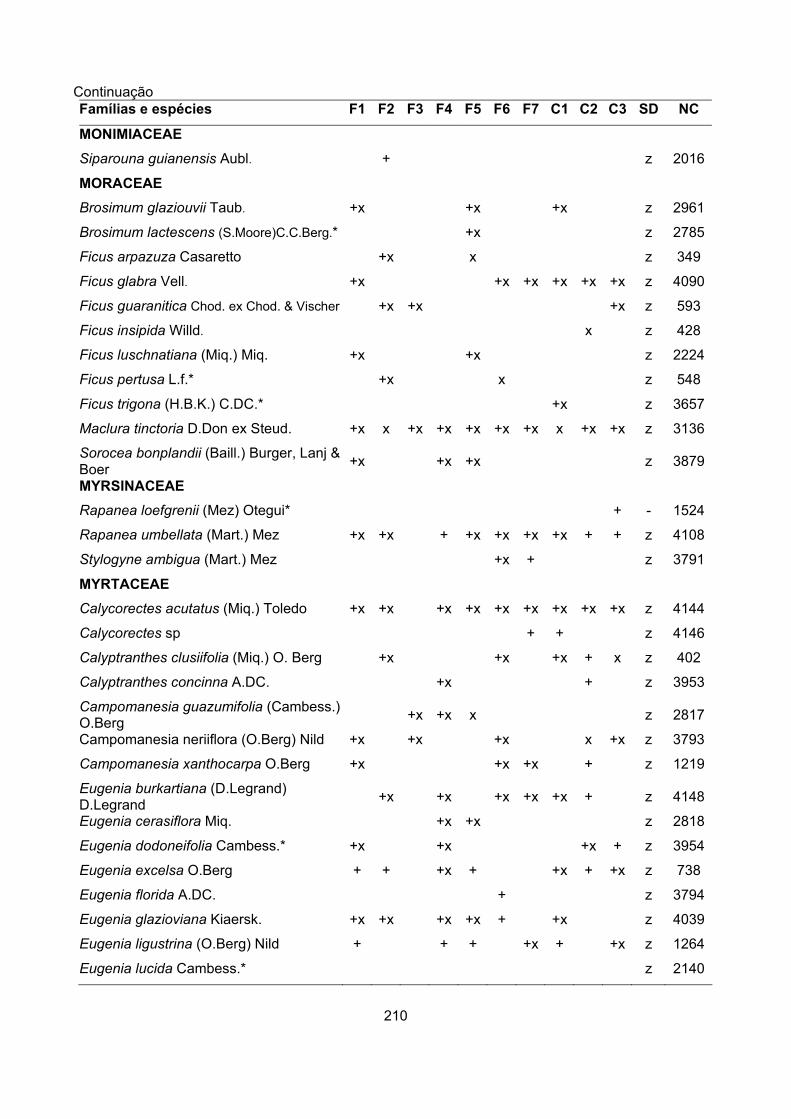
210
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
MONIMIACEAE Siparouna guianensis Aubl. + z 2016
MORACEAE Brosimum glaziouvii Taub. +x +x +x z 2961
Brosimum lactescens (S.Moore)C.C.Berg.* +x z 2785
Ficus arpazuza Casaretto +x x z 349
Ficus glabra Vell. +x +x +x +x +x +x z 4090
Ficus guaranitica Chod. ex Chod. & Vischer +x +x +x z 593
Ficus insipida Willd. x z 428
Ficus luschnatiana (Miq.) Miq. +x +x z 2224
Ficus pertusa L.f.* +x x z 548
Ficus trigona (H.B.K.) C.DC.* +x z 3657
Maclura tinctoria D.Don ex Steud. +x x +x +x +x +x +x x +x +x z 3136
Sorocea bonplandii (Baill.) Burger, Lanj & Boer +x +x +x z 3879
MYRSINACEAE Rapanea loefgrenii (Mez) Otegui* + - 1524
Rapanea umbellata (Mart.) Mez +x +x + +x +x +x +x + + z 4108
Stylogyne ambigua (Mart.) Mez +x + z 3791
MYRTACEAE Calycorectes acutatus (Miq.) Toledo +x +x +x +x +x +x +x +x +x z 4144
Calycorectes sp + + z 4146
Calyptranthes clusiifolia (Miq.) O. Berg +x +x +x + x z 402
Calyptranthes concinna A.DC. +x + z 3953
Campomanesia guazumifolia (Cambess.) O.Berg +x +x x z 2817
Campomanesia neriiflora (O.Berg) Nild +x +x +x x +x z 3793
Campomanesia xanthocarpa O.Berg +x +x +x + z 1219
Eugenia burkartiana (D.Legrand) D.Legrand +x +x +x +x +x + z 4148
Eugenia cerasiflora Miq. +x +x z 2818
Eugenia dodoneifolia Cambess.* +x +x +x + z 3954
Eugenia excelsa O.Berg + + +x + +x + +x z 738
Eugenia florida A.DC. + z 3794
Eugenia glazioviana Kiaersk. +x +x +x +x + +x z 4039
Eugenia ligustrina (O.Berg) Nild + + + +x + +x z 1264
Eugenia lucida Cambess.* z 2140
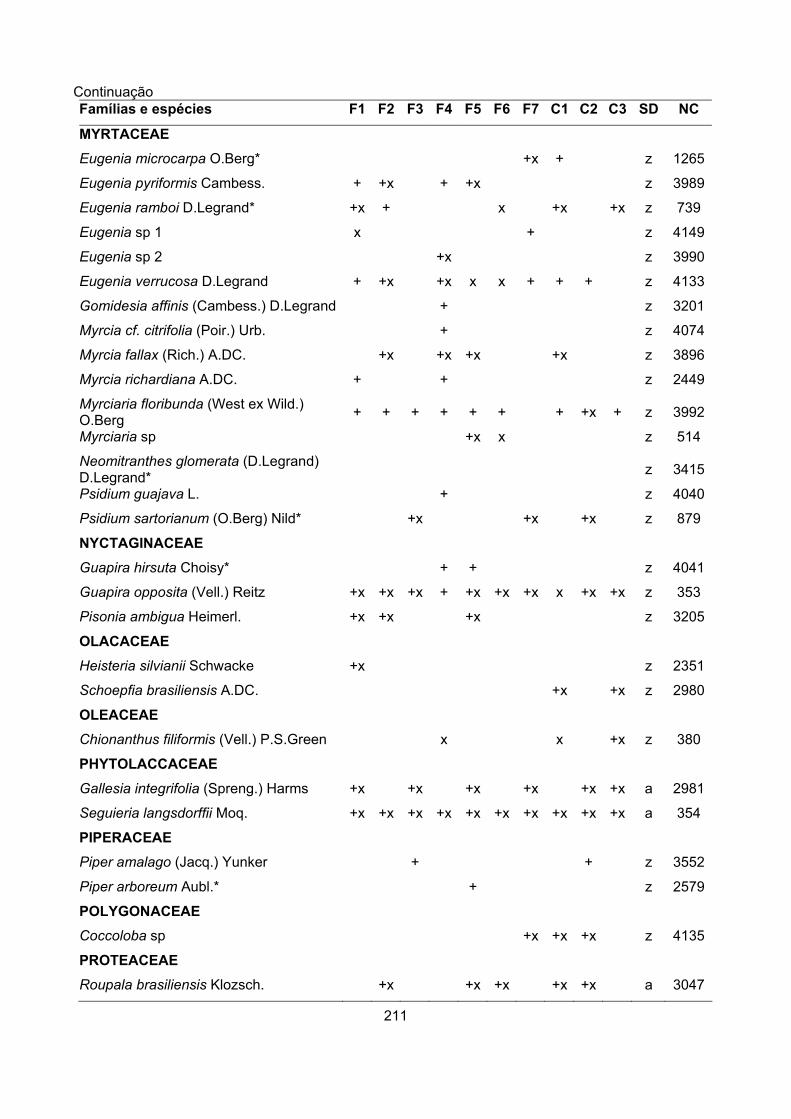
211
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
MYRTACEAE Eugenia microcarpa O.Berg* +x + z 1265
Eugenia pyriformis Cambess. + +x + +x z 3989
Eugenia ramboi D.Legrand* +x + x +x +x z 739
Eugenia sp 1 x + z 4149
Eugenia sp 2 +x z 3990
Eugenia verrucosa D.Legrand + +x +x x x + + + z 4133
Gomidesia affinis (Cambess.) D.Legrand + z 3201
Myrcia cf. citrifolia (Poir.) Urb. + z 4074
Myrcia fallax (Rich.) A.DC. +x +x +x +x z 3896
Myrcia richardiana A.DC. + + z 2449
Myrciaria floribunda (West ex Wild.) O.Berg + + + + + + + +x + z 3992
Myrciaria sp +x x z 514
Neomitranthes glomerata (D.Legrand) D.Legrand* z 3415
Psidium guajava L. + z 4040
Psidium sartorianum (O.Berg) Nild* +x +x +x z 879
NYCTAGINACEAE Guapira hirsuta Choisy* + + z 4041
Guapira opposita (Vell.) Reitz +x +x +x + +x +x +x x +x +x z 353
Pisonia ambigua Heimerl. +x +x +x z 3205
OLACACEAE Heisteria silvianii Schwacke +x z 2351
Schoepfia brasiliensis A.DC. +x +x z 2980
OLEACEAE Chionanthus filiformis (Vell.) P.S.Green x x +x z 380
PHYTOLACCACEAE Gallesia integrifolia (Spreng.) Harms +x +x +x +x +x +x a 2981
Seguieria langsdorffii Moq. +x +x +x +x +x +x +x +x +x +x a 354
PIPERACEAE Piper amalago (Jacq.) Yunker + + z 3552
Piper arboreum Aubl.* + z 2579
POLYGONACEAE Coccoloba sp +x +x +x z 4135
PROTEACEAE Roupala brasiliensis Klozsch. +x +x +x +x +x a 3047

212
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
RHAMNACEAE Colubrina glandulosa Perk. +x +x x u 356
Rhamnidium elaeocarpum Reissek +x x +x +x z 3899
ROSACEAE Prunus myrtifolia (L.) Urb. +x + +x +x +x z 328
RUBIACEAE Alseis floribunda Schott. +x x +x +x +x a 3901
Amaioua guianensis Aubl. +x x +x +x z 1732
Chomelia pohliana Müll. Arg.* + - 1801
Chomelia sp - 3324
Coutarea hexandra (Jacq.) K.Schum. +x +x +x a 993
Guettarda uruguensis Cham. & Schltdl + z 1227
Guettarda viburnioides Cham. & Schltdl +x +x z 2251
Ixora gardneriana Benth. + +x +x +x +x +x +x +x z 4150
Randia nitida L.f. x + z 1405
Rudgea jasminoides (Cham.) Müll. Arg. + + +x +x z 3708
Simira sampaioana (Standl.) Steyrm. +x +x +x +x +x a 471
RUTACEAE Almeidea coerulea (Nees & Mart.) A.St.-Hil. ex C.DC. em Engl. + +x +x +x +x +x u 1608
Balfourodendron riedelianum (Engl.) Engl. +x +x a 3595
Citrus limonia Osbeck z 3215
Esenbeckia grandiflora Mart. +x +x +x +x u 1805
Esenbeckia leiocarpa Engl. +x +x +x +x +x +x +x u 3709
Galipea multiflora Schultz +x + +x +x +x +x +x +x u 360
Metrodorea nigra A.St.-Hil. +x + +x +x +x +x +x u 1177
Metrodorea stipularis Mart. +x +x +x u 313
Zanthoxylum acuminatum Sw. +x x +x +x z 3671
Zanthoxylum caribaeum Lam.* x x z 2699
Zanthoxylum hyemale A.St.-Hil. +x +x +x +x +x +x x +x z 479
Zanthoxylum monogynum A.St.-Hil. +x +x +x +x +x z 4046
Zanthoxylum petiolare A.St.-Hil. & Tul. + z 2583
Zanthoxylum rhoifolium Lam. +x +x +x z 607
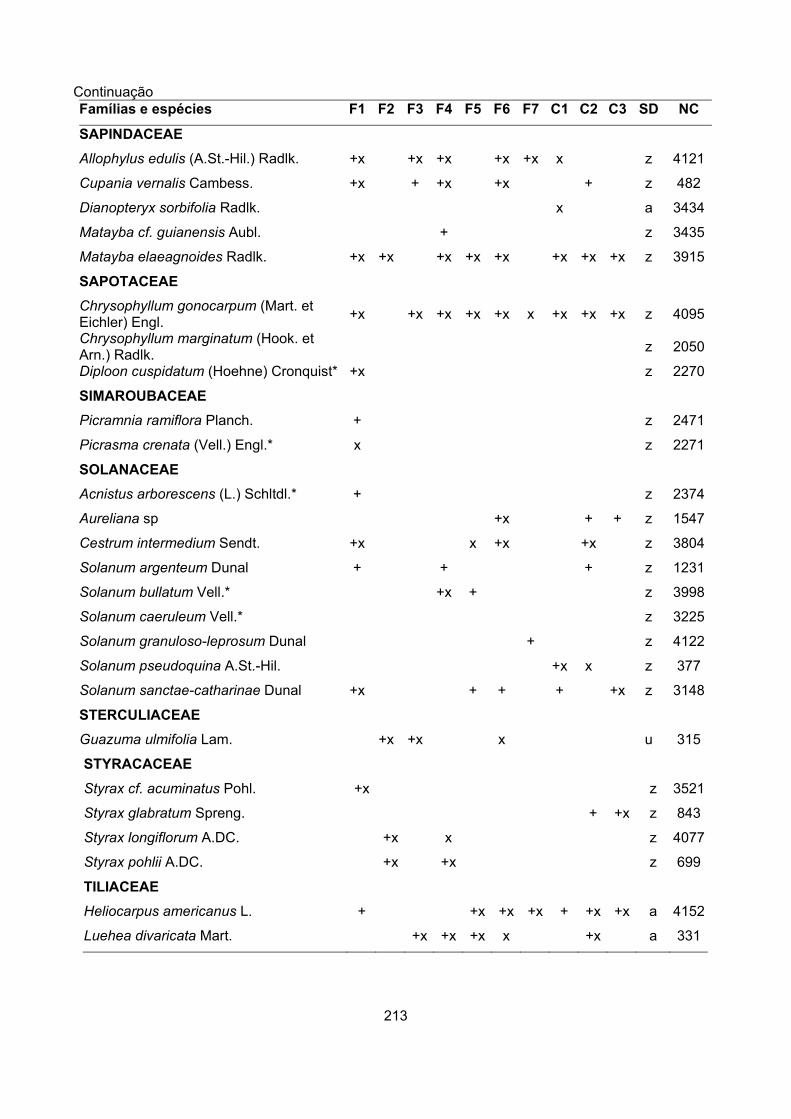
213
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
SAPINDACEAE Allophylus edulis (A.St.-Hil.) Radlk. +x +x +x +x +x x z 4121
Cupania vernalis Cambess. +x + +x +x + z 482
Dianopteryx sorbifolia Radlk. x a 3434
Matayba cf. guianensis Aubl. + z 3435
Matayba elaeagnoides Radlk. +x +x +x +x +x +x +x +x z 3915
SAPOTACEAE Chrysophyllum gonocarpum (Mart. et Eichler) Engl. +x +x +x +x +x x +x +x +x z 4095
Chrysophyllum marginatum (Hook. et Arn.) Radlk. z 2050
Diploon cuspidatum (Hoehne) Cronquist* +x z 2270
SIMAROUBACEAE Picramnia ramiflora Planch. + z 2471
Picrasma crenata (Vell.) Engl.* x z 2271
SOLANACEAE Acnistus arborescens (L.) Schltdl.* + z 2374
Aureliana sp +x + + z 1547
Cestrum intermedium Sendt. +x x +x +x z 3804
Solanum argenteum Dunal + + + z 1231
Solanum bullatum Vell.* +x + z 3998
Solanum caeruleum Vell.* z 3225
Solanum granuloso-leprosum Dunal + z 4122
Solanum pseudoquina A.St.-Hil. +x x z 377
Solanum sanctae-catharinae Dunal +x + + + +x z 3148
STERCULIACEAE Guazuma ulmifolia Lam. +x +x x u 315
STYRACACEAE Styrax cf. acuminatus Pohl. +x z 3521
Styrax glabratum Spreng. + +x z 843
Styrax longiflorum A.DC. +x x z 4077
Styrax pohlii A.DC. +x +x z 699
TILIACEAE Heliocarpus americanus L. + +x +x +x + +x +x a 4152
Luehea divaricata Mart. +x +x +x x +x a 331

214
Continuação Famílias e espécies F1 F2 F3 F4 F5 F6 F7 C1 C2 C3 SD NC
ULMACEAE Celtis tala Gillies ex Planch. +x z 1955
Trema micrantha (L.) Blume +x x +x +x + +x z 484
URTICACEAE Urera baccifera (L.) Gaudich. +x +x +x +x +x +x +x +x +x z 485
VERBENACEAE Aegiphila sellowiana Cham. +x +x +x +x +x z 3807
Aloysia virgata (Ruiz et Pav.) A.Juss. +x + +x + +x a 530
Citharexylum myrianthum Cham. +x +x +x z 4097
Vitex polygama Cham. x +x z 658
VOCHYSIACEAE Qualea jundiahy Warm. +x +x a 4078
Vockysia magnifica Warm. +x a 920
Vockysia tucanorum Mart. +x + a 3711
INDETERMINADA Indeterminada 1 - 3473
* Espécies sendo citadas pela primeira vez para o município de Campinas/Itatiba, SP. ** Série de K.Santos. Por uma questão de espaço foram iseridos apenas um número do coletor para cada espécie, em caso de interesse os demais números podem ser solicitados aos autores.

215
ANEXO II
Tabelas contendo os parâmetros fitossociológicos analisados, considerando DAP ≥ 5cm, nos
onze fragmentos estudados no trabalho “Utilização de diferentes critérios de inclusão em
inventários florísticos e de estrutura de florestas estacionais semidecíduas e suas
implicações sobre os resultados – um estudo de caso”.
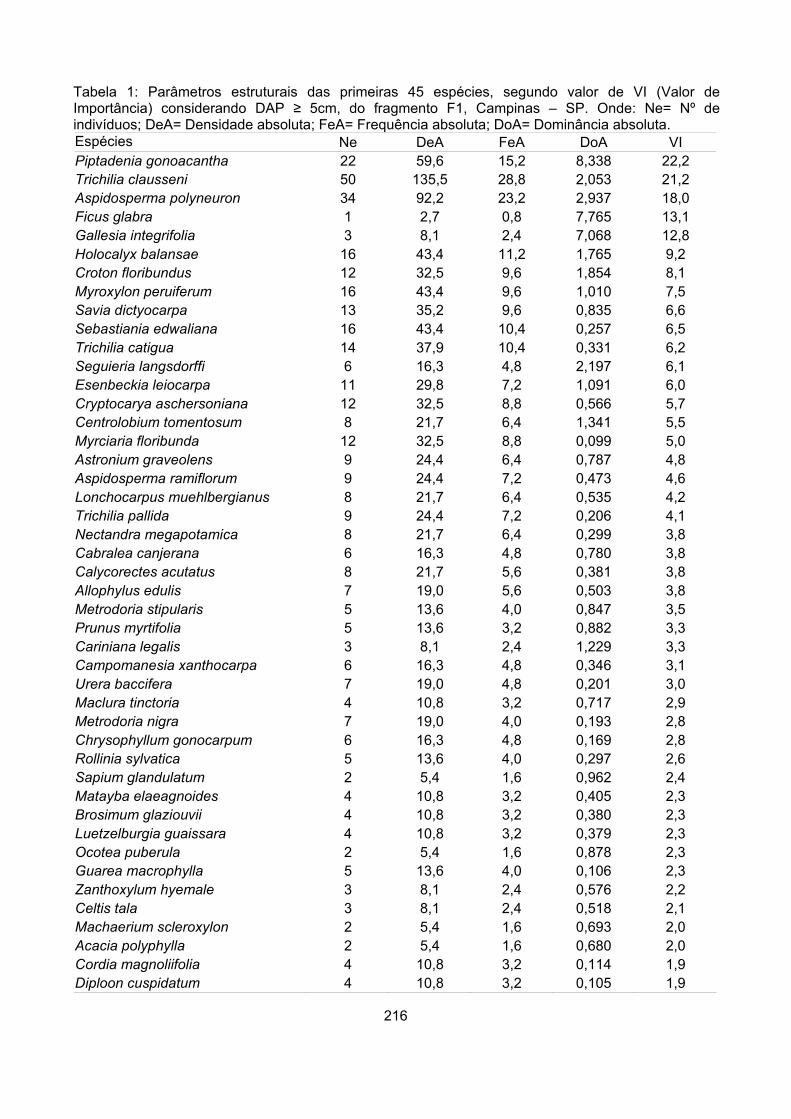
216
Tabela 1: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F1, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Piptadenia gonoacantha 22 59,6 15,2 8,338 22,2 Trichilia clausseni 50 135,5 28,8 2,053 21,2 Aspidosperma polyneuron 34 92,2 23,2 2,937 18,0 Ficus glabra 1 2,7 0,8 7,765 13,1 Gallesia integrifolia 3 8,1 2,4 7,068 12,8 Holocalyx balansae 16 43,4 11,2 1,765 9,2 Croton floribundus 12 32,5 9,6 1,854 8,1 Myroxylon peruiferum 16 43,4 9,6 1,010 7,5 Savia dictyocarpa 13 35,2 9,6 0,835 6,6 Sebastiania edwaliana 16 43,4 10,4 0,257 6,5 Trichilia catigua 14 37,9 10,4 0,331 6,2 Seguieria langsdorffi 6 16,3 4,8 2,197 6,1 Esenbeckia leiocarpa 11 29,8 7,2 1,091 6,0 Cryptocarya aschersoniana 12 32,5 8,8 0,566 5,7 Centrolobium tomentosum 8 21,7 6,4 1,341 5,5 Myrciaria floribunda 12 32,5 8,8 0,099 5,0 Astronium graveolens 9 24,4 6,4 0,787 4,8 Aspidosperma ramiflorum 9 24,4 7,2 0,473 4,6 Lonchocarpus muehlbergianus 8 21,7 6,4 0,535 4,2 Trichilia pallida 9 24,4 7,2 0,206 4,1 Nectandra megapotamica 8 21,7 6,4 0,299 3,8 Cabralea canjerana 6 16,3 4,8 0,780 3,8 Calycorectes acutatus 8 21,7 5,6 0,381 3,8 Allophylus edulis 7 19,0 5,6 0,503 3,8 Metrodoria stipularis 5 13,6 4,0 0,847 3,5 Prunus myrtifolia 5 13,6 3,2 0,882 3,3 Cariniana legalis 3 8,1 2,4 1,229 3,3 Campomanesia xanthocarpa 6 16,3 4,8 0,346 3,1 Urera baccifera 7 19,0 4,8 0,201 3,0 Maclura tinctoria 4 10,8 3,2 0,717 2,9 Metrodoria nigra 7 19,0 4,0 0,193 2,8 Chrysophyllum gonocarpum 6 16,3 4,8 0,169 2,8 Rollinia sylvatica 5 13,6 4,0 0,297 2,6 Sapium glandulatum 2 5,4 1,6 0,962 2,4 Matayba elaeagnoides 4 10,8 3,2 0,405 2,3 Brosimum glaziouvii 4 10,8 3,2 0,380 2,3 Luetzelburgia guaissara 4 10,8 3,2 0,379 2,3 Ocotea puberula 2 5,4 1,6 0,878 2,3 Guarea macrophylla 5 13,6 4,0 0,106 2,3 Zanthoxylum hyemale 3 8,1 2,4 0,576 2,2 Celtis tala 3 8,1 2,4 0,518 2,1 Machaerium scleroxylon 2 5,4 1,6 0,693 2,0 Acacia polyphylla 2 5,4 1,6 0,680 2,0 Cordia magnoliifolia 4 10,8 3,2 0,114 1,9 Diploon cuspidatum 4 10,8 3,2 0,105 1,9

217
Tabela 2: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F2, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 63 142,4 32,8 6,954 46,9 Metrodoria stipularis 36 81,3 23,2 1,766 20,1 Galipia multiflora 49 110,7 30,4 0,454 20,0 Croton floribundus 22 49,7 17,6 2,151 17,1 Calycorectes acutatus 24 54,2 16,8 0,689 12,0 Mabea fistulifera 21 47,5 16,0 0,902 12,0 Piptadenia gonoacantha 12 27,1 9,6 1,762 11,5 Astronium graveolens 18 40,7 12,8 0,875 10,4 Aspidosperma polyneuron 15 33,9 11,2 0,587 8,3 Casearia sylvestris 12 27,1 8,8 0,200 5,6 Cryptocarya aschersoniana 8 18,1 4,8 0,678 5,4 Cordia sellowiana 8 18,1 6,4 0,484 5,2 Savia dictyocarpa 10 22,6 7,2 0,187 4,7 Tapirira obtusa 5 11,3 3,2 0,701 4,4 Inga vera 6 13,6 4,8 0,434 4,1 Rapanea umbellata 8 18,1 6,4 0,196 4,1 Machaerium nictitans 7 15,8 5,6 0,270 4,0 Hymenaea courbaril 4 9,0 3,2 0,487 3,5 Eugenia verrucosa 7 15,8 5,6 0,107 3,4 Colubrina glandulosa 4 9,0 3,2 0,453 3,3 Cabralea canjerana 4 9,0 3,2 0,376 3,1 Qualea jundiahy 3 6,8 2,4 0,453 2,9 Cariniana estrellensis 5 11,3 4,0 0,216 2,9 Myrciaria floribunda 7 15,8 4,8 0,042 2,9 Casearia gossypiosperma 5 11,3 4,0 0,211 2,9 Ixora gardneriana 6 13,6 4,0 0,131 2,8 Trichilia pallida 6 13,6 4,8 0,036 2,7 Actinostemon communis 6 13,6 4,8 0,033 2,7 Luetzelburgia guaissara 5 11,3 4,0 0,142 2,6 Croton priscus 3 6,8 2,4 0,375 2,6 Pachystroma longifolium 4 9,0 2,4 0,290 2,5 Ormosia arborea 4 9,0 3,2 0,219 2,5 Ocotea corymbosa 4 9,0 3,2 0,201 2,4 Roupala brasiliensis 5 11,3 4,0 0,081 2,4 Ocotea diospirifolia 3 6,8 2,4 0,285 2,3 Pseudobombax grandiflorum 1 2,3 0,8 0,514 2,3 Ficus guaranitica 4 9,0 3,2 0,159 2,3 Copaifera langsdorffi 4 9,0 3,2 0,156 2,3 Annona cacans 3 6,8 2,4 0,255 2,2 Ocotea beulahiae 4 9,0 3,2 0,115 2,1 Zeyheria tuberculosa 2 4,5 1,6 0,300 1,9 Conarus regnelli 4 9,0 3,2 0,037 1,8 Sebastiania edwaliana 4 9,0 3,2 0,033 1,8 Cecropia hololeuca 2 4,5 1,6 0,249 1,8 Matayba elaeagnoides 3 6,8 2,4 0,129 1,7

218
Tabela 3: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F3, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Trichilia clausseni 244 614,4 88,8 10,849 105,8 Chorisia speciosa 19 47,8 14,4 14,017 39,6 Lonchocarpus muehlbergianus 47 118,3 25,6 6,259 32,6 Trichilia catigua 25 62,9 13,6 0,818 11,9 Astronium graveolens 16 40,3 12,0 1,192 10,3 Maclura tinctoria 12 30,2 9,6 1,828 10,0 Trichilia hirta 17 42,8 12,8 0,702 9,8 Myroxylon peruiferum 10 25,2 7,2 0,970 6,8 Casearia sylvestris 12 30,2 8,0 0,343 6,2 Hymenaea courbaril 2 5,0 1,6 2,068 5,5 Diospyrus inconstans 10 25,2 5,6 0,481 5,2 Centrolobium tomentosum 7 17,6 5,6 0,504 4,6 Cariniana estrellensis 6 15,1 4,8 0,429 3,9 Pseudobombax grandiflorum 4 10,1 3,2 0,786 3,7 Casearia gossypiosperma 4 10,1 2,4 0,797 3,4 Urera baccifera 5 12,6 4,0 0,329 3,2 Holocalyx balansae 4 10,1 3,2 0,541 3,2 Luehea divaricata 2 5,0 1,6 0,370 1,8 Chrysophyllum gonocarpum 3 7,6 2,4 0,055 1,6 Campomanesia guazumifolia 4 10,1 1,6 0,102 1,6 Rollinia sylvatica 2 5,0 1,6 0,093 1,2 Sweetia fruticosa 2 5,0 1,6 0,089 1,2 Copaifera langsdorffi 2 5,0 1,6 0,085 1,2 Campomanesia neriiflora 2 5,0 0,8 0,223 1,2 Pithecellobium edwalii 2 5,0 1,6 0,082 1,2 Bastardiopsis densiflora 1 2,5 0,8 0,303 1,2 Coutarea hexandra 2 5,0 1,6 0,052 1,1 Allophylus edulis 2 5,0 1,6 0,046 1,1 Aspidosperma ramiflorum 2 5,0 1,6 0,044 1,1 Machaerium scleroxylon 2 5,0 1,6 0,039 1,1 Aloysia virgata 2 5,0 1,6 0,028 1,1 Calliandra sp 2 5,0 1,6 0,021 1,1 Guazuma ulmifolia 1 2,5 0,8 0,251 1,1 Maytenus aquifolia 2 5,0 1,6 0,013 1,0 Myrciaria floribunda 2 5,0 1,6 0,013 1,0 Cupania vernalis 2 5,0 1,6 0,010 1,0 Seguieria langsdorffi 1 2,5 0,8 0,213 1,0 Balfourodendron riedelianum 1 2,5 0,8 0,196 0,9 Peltophorum dubium 1 2,5 0,8 0,192 0,9 Ficus guaranitica 1 2,5 0,8 0,189 0,9 Gallesia integrifolia 1 2,5 0,8 0,185 0,9 Zanthoxylum hyemale 1 2,5 0,8 0,177 0,9 Psidium sartorianum 1 2,5 0,8 0,077 0,7 Piptadenia gonoacantha 1 2,5 0,8 0,034 0,6 Cordia ecalyculata 1 2,5 0,8 0,031 0,6
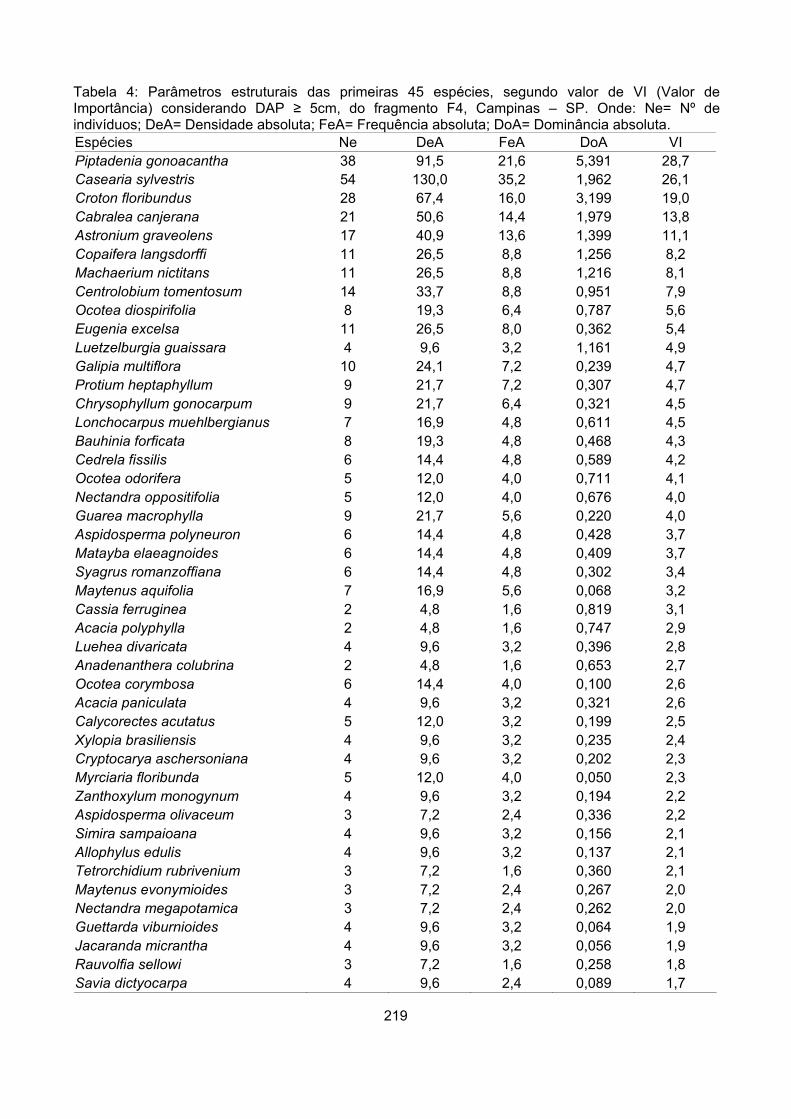
219
Tabela 4: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F4, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Piptadenia gonoacantha 38 91,5 21,6 5,391 28,7 Casearia sylvestris 54 130,0 35,2 1,962 26,1 Croton floribundus 28 67,4 16,0 3,199 19,0 Cabralea canjerana 21 50,6 14,4 1,979 13,8 Astronium graveolens 17 40,9 13,6 1,399 11,1 Copaifera langsdorffi 11 26,5 8,8 1,256 8,2 Machaerium nictitans 11 26,5 8,8 1,216 8,1 Centrolobium tomentosum 14 33,7 8,8 0,951 7,9 Ocotea diospirifolia 8 19,3 6,4 0,787 5,6 Eugenia excelsa 11 26,5 8,0 0,362 5,4 Luetzelburgia guaissara 4 9,6 3,2 1,161 4,9 Galipia multiflora 10 24,1 7,2 0,239 4,7 Protium heptaphyllum 9 21,7 7,2 0,307 4,7 Chrysophyllum gonocarpum 9 21,7 6,4 0,321 4,5 Lonchocarpus muehlbergianus 7 16,9 4,8 0,611 4,5 Bauhinia forficata 8 19,3 4,8 0,468 4,3 Cedrela fissilis 6 14,4 4,8 0,589 4,2 Ocotea odorifera 5 12,0 4,0 0,711 4,1 Nectandra oppositifolia 5 12,0 4,0 0,676 4,0 Guarea macrophylla 9 21,7 5,6 0,220 4,0 Aspidosperma polyneuron 6 14,4 4,8 0,428 3,7 Matayba elaeagnoides 6 14,4 4,8 0,409 3,7 Syagrus romanzoffiana 6 14,4 4,8 0,302 3,4 Maytenus aquifolia 7 16,9 5,6 0,068 3,2 Cassia ferruginea 2 4,8 1,6 0,819 3,1 Acacia polyphylla 2 4,8 1,6 0,747 2,9 Luehea divaricata 4 9,6 3,2 0,396 2,8 Anadenanthera colubrina 2 4,8 1,6 0,653 2,7 Ocotea corymbosa 6 14,4 4,0 0,100 2,6 Acacia paniculata 4 9,6 3,2 0,321 2,6 Calycorectes acutatus 5 12,0 3,2 0,199 2,5 Xylopia brasiliensis 4 9,6 3,2 0,235 2,4 Cryptocarya aschersoniana 4 9,6 3,2 0,202 2,3 Myrciaria floribunda 5 12,0 4,0 0,050 2,3 Zanthoxylum monogynum 4 9,6 3,2 0,194 2,2 Aspidosperma olivaceum 3 7,2 2,4 0,336 2,2 Simira sampaioana 4 9,6 3,2 0,156 2,1 Allophylus edulis 4 9,6 3,2 0,137 2,1 Tetrorchidium rubrivenium 3 7,2 1,6 0,360 2,1 Maytenus evonymioides 3 7,2 2,4 0,267 2,0 Nectandra megapotamica 3 7,2 2,4 0,262 2,0 Guettarda viburnioides 4 9,6 3,2 0,064 1,9 Jacaranda micrantha 4 9,6 3,2 0,056 1,9 Rauvolfia sellowi 3 7,2 1,6 0,258 1,8 Savia dictyocarpa 4 9,6 2,4 0,089 1,7
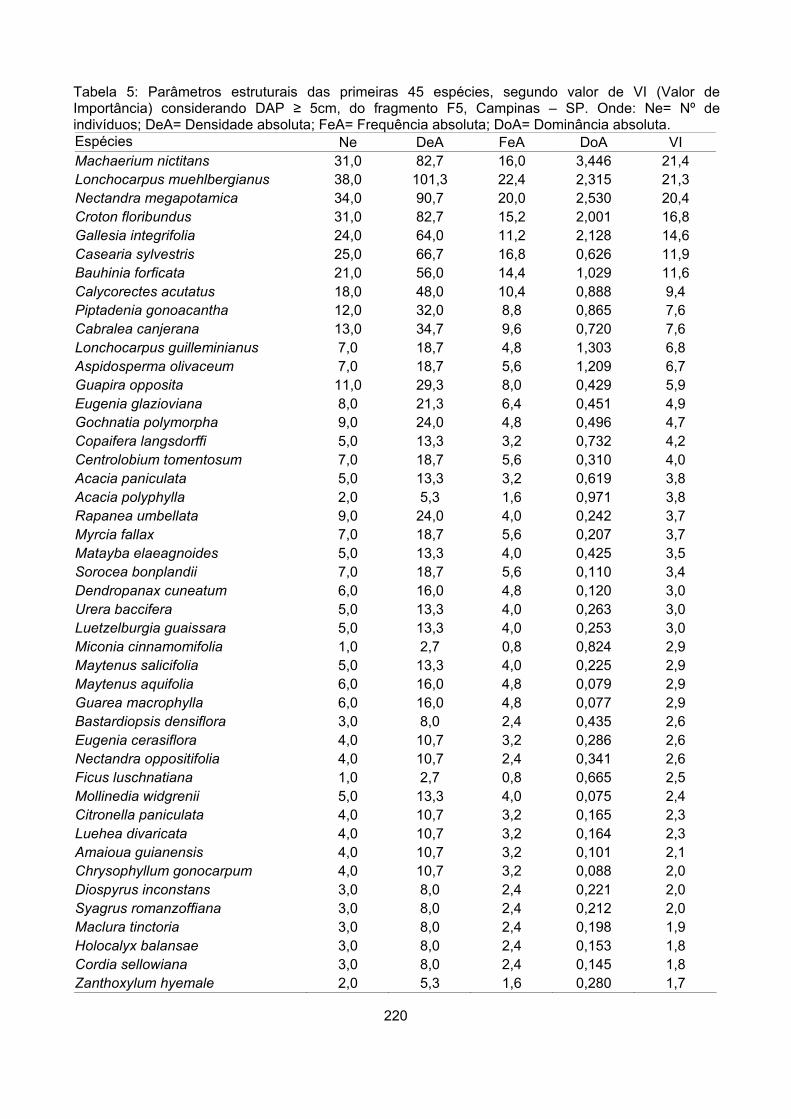
220
Tabela 5: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F5, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Machaerium nictitans 31,0 82,7 16,0 3,446 21,4 Lonchocarpus muehlbergianus 38,0 101,3 22,4 2,315 21,3 Nectandra megapotamica 34,0 90,7 20,0 2,530 20,4 Croton floribundus 31,0 82,7 15,2 2,001 16,8 Gallesia integrifolia 24,0 64,0 11,2 2,128 14,6 Casearia sylvestris 25,0 66,7 16,8 0,626 11,9 Bauhinia forficata 21,0 56,0 14,4 1,029 11,6 Calycorectes acutatus 18,0 48,0 10,4 0,888 9,4 Piptadenia gonoacantha 12,0 32,0 8,8 0,865 7,6 Cabralea canjerana 13,0 34,7 9,6 0,720 7,6 Lonchocarpus guilleminianus 7,0 18,7 4,8 1,303 6,8 Aspidosperma olivaceum 7,0 18,7 5,6 1,209 6,7 Guapira opposita 11,0 29,3 8,0 0,429 5,9 Eugenia glazioviana 8,0 21,3 6,4 0,451 4,9 Gochnatia polymorpha 9,0 24,0 4,8 0,496 4,7 Copaifera langsdorffi 5,0 13,3 3,2 0,732 4,2 Centrolobium tomentosum 7,0 18,7 5,6 0,310 4,0 Acacia paniculata 5,0 13,3 3,2 0,619 3,8 Acacia polyphylla 2,0 5,3 1,6 0,971 3,8 Rapanea umbellata 9,0 24,0 4,0 0,242 3,7 Myrcia fallax 7,0 18,7 5,6 0,207 3,7 Matayba elaeagnoides 5,0 13,3 4,0 0,425 3,5 Sorocea bonplandii 7,0 18,7 5,6 0,110 3,4 Dendropanax cuneatum 6,0 16,0 4,8 0,120 3,0 Urera baccifera 5,0 13,3 4,0 0,263 3,0 Luetzelburgia guaissara 5,0 13,3 4,0 0,253 3,0 Miconia cinnamomifolia 1,0 2,7 0,8 0,824 2,9 Maytenus salicifolia 5,0 13,3 4,0 0,225 2,9 Maytenus aquifolia 6,0 16,0 4,8 0,079 2,9 Guarea macrophylla 6,0 16,0 4,8 0,077 2,9 Bastardiopsis densiflora 3,0 8,0 2,4 0,435 2,6 Eugenia cerasiflora 4,0 10,7 3,2 0,286 2,6 Nectandra oppositifolia 4,0 10,7 2,4 0,341 2,6 Ficus luschnatiana 1,0 2,7 0,8 0,665 2,5 Mollinedia widgrenii 5,0 13,3 4,0 0,075 2,4 Citronella paniculata 4,0 10,7 3,2 0,165 2,3 Luehea divaricata 4,0 10,7 3,2 0,164 2,3 Amaioua guianensis 4,0 10,7 3,2 0,101 2,1 Chrysophyllum gonocarpum 4,0 10,7 3,2 0,088 2,0 Diospyrus inconstans 3,0 8,0 2,4 0,221 2,0 Syagrus romanzoffiana 3,0 8,0 2,4 0,212 2,0 Maclura tinctoria 3,0 8,0 2,4 0,198 1,9 Holocalyx balansae 3,0 8,0 2,4 0,153 1,8 Cordia sellowiana 3,0 8,0 2,4 0,145 1,8 Zanthoxylum hyemale 2,0 5,3 1,6 0,280 1,7
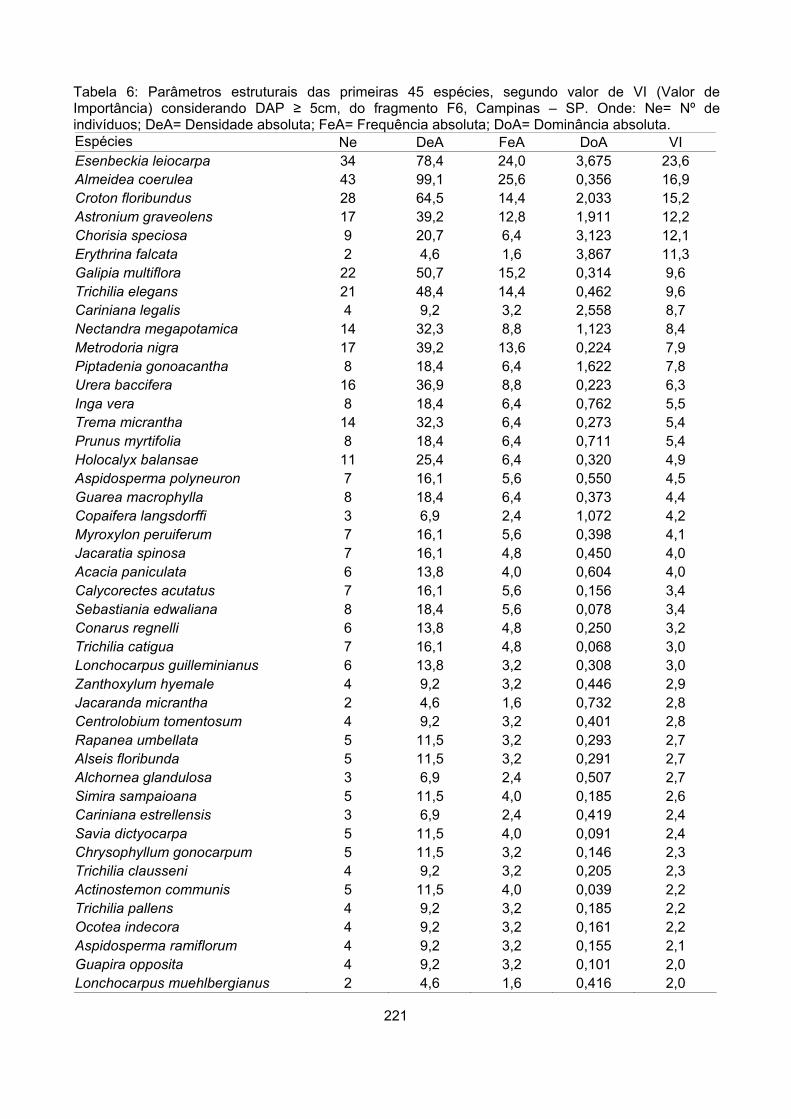
221
Tabela 6: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F6, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 34 78,4 24,0 3,675 23,6 Almeidea coerulea 43 99,1 25,6 0,356 16,9 Croton floribundus 28 64,5 14,4 2,033 15,2 Astronium graveolens 17 39,2 12,8 1,911 12,2 Chorisia speciosa 9 20,7 6,4 3,123 12,1 Erythrina falcata 2 4,6 1,6 3,867 11,3 Galipia multiflora 22 50,7 15,2 0,314 9,6 Trichilia elegans 21 48,4 14,4 0,462 9,6 Cariniana legalis 4 9,2 3,2 2,558 8,7 Nectandra megapotamica 14 32,3 8,8 1,123 8,4 Metrodoria nigra 17 39,2 13,6 0,224 7,9 Piptadenia gonoacantha 8 18,4 6,4 1,622 7,8 Urera baccifera 16 36,9 8,8 0,223 6,3 Inga vera 8 18,4 6,4 0,762 5,5 Trema micrantha 14 32,3 6,4 0,273 5,4 Prunus myrtifolia 8 18,4 6,4 0,711 5,4 Holocalyx balansae 11 25,4 6,4 0,320 4,9 Aspidosperma polyneuron 7 16,1 5,6 0,550 4,5 Guarea macrophylla 8 18,4 6,4 0,373 4,4 Copaifera langsdorffi 3 6,9 2,4 1,072 4,2 Myroxylon peruiferum 7 16,1 5,6 0,398 4,1 Jacaratia spinosa 7 16,1 4,8 0,450 4,0 Acacia paniculata 6 13,8 4,0 0,604 4,0 Calycorectes acutatus 7 16,1 5,6 0,156 3,4 Sebastiania edwaliana 8 18,4 5,6 0,078 3,4 Conarus regnelli 6 13,8 4,8 0,250 3,2 Trichilia catigua 7 16,1 4,8 0,068 3,0 Lonchocarpus guilleminianus 6 13,8 3,2 0,308 3,0 Zanthoxylum hyemale 4 9,2 3,2 0,446 2,9 Jacaranda micrantha 2 4,6 1,6 0,732 2,8 Centrolobium tomentosum 4 9,2 3,2 0,401 2,8 Rapanea umbellata 5 11,5 3,2 0,293 2,7 Alseis floribunda 5 11,5 3,2 0,291 2,7 Alchornea glandulosa 3 6,9 2,4 0,507 2,7 Simira sampaioana 5 11,5 4,0 0,185 2,6 Cariniana estrellensis 3 6,9 2,4 0,419 2,4 Savia dictyocarpa 5 11,5 4,0 0,091 2,4 Chrysophyllum gonocarpum 5 11,5 3,2 0,146 2,3 Trichilia clausseni 4 9,2 3,2 0,205 2,3 Actinostemon communis 5 11,5 4,0 0,039 2,2 Trichilia pallens 4 9,2 3,2 0,185 2,2 Ocotea indecora 4 9,2 3,2 0,161 2,2 Aspidosperma ramiflorum 4 9,2 3,2 0,155 2,1 Guapira opposita 4 9,2 3,2 0,101 2,0 Lonchocarpus muehlbergianus 2 4,6 1,6 0,416 2,0

222
Tabela 7: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento F7, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Gallesia integrifolia 21 39,2 15,2 11,273 31,1 Aspidosperma polyneuron 44 82,2 30,4 6,128 29,9 Esenbeckia leiocarpa 46 85,9 28,8 3,729 25,0 Urera baccifera 63 117,7 32,8 1,090 24,3 Holocalyx balansae 41 76,6 27,2 2,600 21,3 Metrodoria nigra 37 69,1 27,2 0,437 16,2 Aspidosperma ramiflorum 26 48,6 17,6 1,822 14,0 Almeidea coerulea 35 65,4 21,6 0,209 13,7 Ficus glabra 1 1,9 0,8 5,806 12,0 Chorisia speciosa 7 13,1 5,6 2,945 8,9 Trichilia catigua 18 33,6 14,4 0,353 8,5 Cariniana estrellensis 6 11,2 4,8 2,789 8,2 Trichilia clausseni 13 24,3 10,4 0,434 6,5 Cariniana legalis 2 3,7 1,6 2,255 5,4 Galipia multiflora 11 20,5 8,8 0,097 5,0 Lonchocarpus muehlbergianus 6 11,2 4,8 0,836 4,3 Simira sampaioana 6 11,2 4,8 0,557 3,7 Hymenaea courbaril 1 1,9 0,8 1,609 3,7 Jacaratia spinosa 9 16,8 4,8 0,082 3,4 Savia dictyocarpa 6 11,2 4,8 0,161 2,9 Ocotea indecora 6 11,2 4,8 0,114 2,8 Trema micrantha 5 9,3 3,2 0,185 2,3 Coccoloba sp 4 7,5 3,2 0,284 2,3 Centrolobium tomentosum 3 5,6 2,4 0,495 2,3 Sebastiania edwaliana 5 9,3 4,0 0,024 2,2 Machaerium scleroxylon 4 7,5 3,2 0,182 2,1 Croton floribundus 4 7,5 1,6 0,392 2,1 Astronium graveolens 4 7,5 3,2 0,147 2,0 Casearia gossypiosperma 4 7,5 3,2 0,108 1,9 Trichilia elegans 4 7,5 3,2 0,050 1,8 Guapira opposita 3 5,6 2,4 0,181 1,7 Bastardiopsis densiflora 1 1,9 0,8 0,514 1,5 Calycorectes acutatus 3 5,6 2,4 0,068 1,4 Myroxylon peruiferum 1 1,9 0,8 0,482 1,4 Eugenia burkartiana 3 5,6 2,4 0,049 1,4 Psidium sartorianum 2 3,7 1,6 0,205 1,3 Maclura tinctoria 2 3,7 1,6 0,112 1,1 Nectandra megapotamica 2 3,7 1,6 0,092 1,1 Campomanesia xanthocarpa 2 3,7 1,6 0,071 1,0 Seguieria langsdorffi 2 3,7 1,6 0,036 0,9 Syagrus romanzoffiana 2 3,7 1,6 0,027 0,9 Eugenia verrucosa 2 3,7 1,6 0,021 0,9 Citronella paniculata 2 3,7 1,6 0,017 0,9 Heliocarpus americanus 2 3,7 1,6 0,015 0,9 Ixora gardneriana 2 3,7 1,6 0,014 0,9
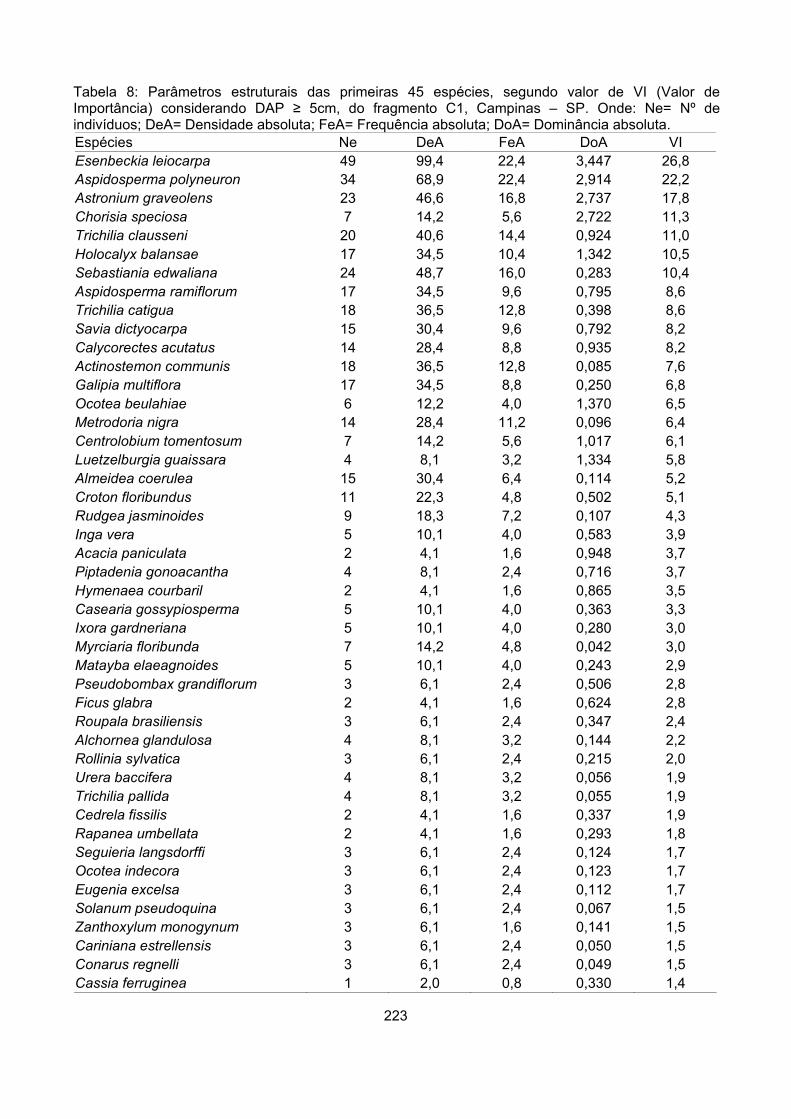
223
Tabela 8: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento C1, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 49 99,4 22,4 3,447 26,8 Aspidosperma polyneuron 34 68,9 22,4 2,914 22,2 Astronium graveolens 23 46,6 16,8 2,737 17,8 Chorisia speciosa 7 14,2 5,6 2,722 11,3 Trichilia clausseni 20 40,6 14,4 0,924 11,0 Holocalyx balansae 17 34,5 10,4 1,342 10,5 Sebastiania edwaliana 24 48,7 16,0 0,283 10,4 Aspidosperma ramiflorum 17 34,5 9,6 0,795 8,6 Trichilia catigua 18 36,5 12,8 0,398 8,6 Savia dictyocarpa 15 30,4 9,6 0,792 8,2 Calycorectes acutatus 14 28,4 8,8 0,935 8,2 Actinostemon communis 18 36,5 12,8 0,085 7,6 Galipia multiflora 17 34,5 8,8 0,250 6,8 Ocotea beulahiae 6 12,2 4,0 1,370 6,5 Metrodoria nigra 14 28,4 11,2 0,096 6,4 Centrolobium tomentosum 7 14,2 5,6 1,017 6,1 Luetzelburgia guaissara 4 8,1 3,2 1,334 5,8 Almeidea coerulea 15 30,4 6,4 0,114 5,2 Croton floribundus 11 22,3 4,8 0,502 5,1 Rudgea jasminoides 9 18,3 7,2 0,107 4,3 Inga vera 5 10,1 4,0 0,583 3,9 Acacia paniculata 2 4,1 1,6 0,948 3,7 Piptadenia gonoacantha 4 8,1 2,4 0,716 3,7 Hymenaea courbaril 2 4,1 1,6 0,865 3,5 Casearia gossypiosperma 5 10,1 4,0 0,363 3,3 Ixora gardneriana 5 10,1 4,0 0,280 3,0 Myrciaria floribunda 7 14,2 4,8 0,042 3,0 Matayba elaeagnoides 5 10,1 4,0 0,243 2,9 Pseudobombax grandiflorum 3 6,1 2,4 0,506 2,8 Ficus glabra 2 4,1 1,6 0,624 2,8 Roupala brasiliensis 3 6,1 2,4 0,347 2,4 Alchornea glandulosa 4 8,1 3,2 0,144 2,2 Rollinia sylvatica 3 6,1 2,4 0,215 2,0 Urera baccifera 4 8,1 3,2 0,056 1,9 Trichilia pallida 4 8,1 3,2 0,055 1,9 Cedrela fissilis 2 4,1 1,6 0,337 1,9 Rapanea umbellata 2 4,1 1,6 0,293 1,8 Seguieria langsdorffi 3 6,1 2,4 0,124 1,7 Ocotea indecora 3 6,1 2,4 0,123 1,7 Eugenia excelsa 3 6,1 2,4 0,112 1,7 Solanum pseudoquina 3 6,1 2,4 0,067 1,5 Zanthoxylum monogynum 3 6,1 1,6 0,141 1,5 Cariniana estrellensis 3 6,1 2,4 0,050 1,5 Conarus regnelli 3 6,1 2,4 0,049 1,5 Cassia ferruginea 1 2,0 0,8 0,330 1,4
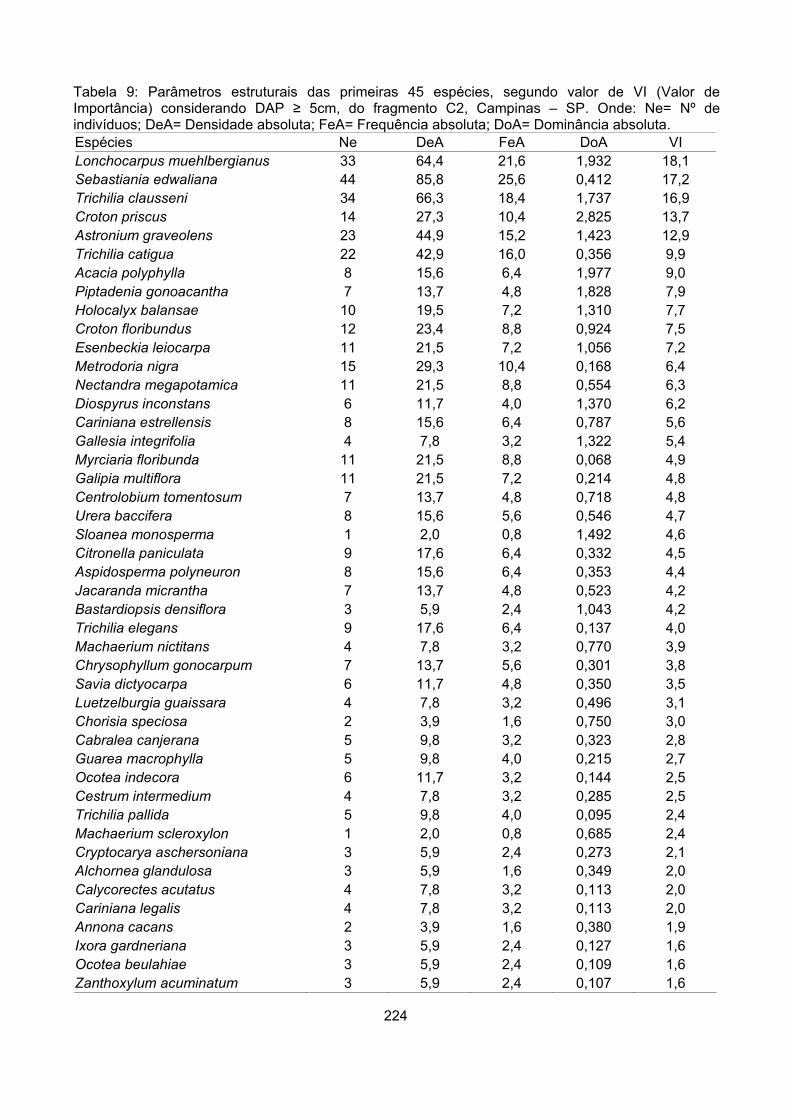
224
Tabela 9: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento C2, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Lonchocarpus muehlbergianus 33 64,4 21,6 1,932 18,1 Sebastiania edwaliana 44 85,8 25,6 0,412 17,2 Trichilia clausseni 34 66,3 18,4 1,737 16,9 Croton priscus 14 27,3 10,4 2,825 13,7 Astronium graveolens 23 44,9 15,2 1,423 12,9 Trichilia catigua 22 42,9 16,0 0,356 9,9 Acacia polyphylla 8 15,6 6,4 1,977 9,0 Piptadenia gonoacantha 7 13,7 4,8 1,828 7,9 Holocalyx balansae 10 19,5 7,2 1,310 7,7 Croton floribundus 12 23,4 8,8 0,924 7,5 Esenbeckia leiocarpa 11 21,5 7,2 1,056 7,2 Metrodoria nigra 15 29,3 10,4 0,168 6,4 Nectandra megapotamica 11 21,5 8,8 0,554 6,3 Diospyrus inconstans 6 11,7 4,0 1,370 6,2 Cariniana estrellensis 8 15,6 6,4 0,787 5,6 Gallesia integrifolia 4 7,8 3,2 1,322 5,4 Myrciaria floribunda 11 21,5 8,8 0,068 4,9 Galipia multiflora 11 21,5 7,2 0,214 4,8 Centrolobium tomentosum 7 13,7 4,8 0,718 4,8 Urera baccifera 8 15,6 5,6 0,546 4,7 Sloanea monosperma 1 2,0 0,8 1,492 4,6 Citronella paniculata 9 17,6 6,4 0,332 4,5 Aspidosperma polyneuron 8 15,6 6,4 0,353 4,4 Jacaranda micrantha 7 13,7 4,8 0,523 4,2 Bastardiopsis densiflora 3 5,9 2,4 1,043 4,2 Trichilia elegans 9 17,6 6,4 0,137 4,0 Machaerium nictitans 4 7,8 3,2 0,770 3,9 Chrysophyllum gonocarpum 7 13,7 5,6 0,301 3,8 Savia dictyocarpa 6 11,7 4,8 0,350 3,5 Luetzelburgia guaissara 4 7,8 3,2 0,496 3,1 Chorisia speciosa 2 3,9 1,6 0,750 3,0 Cabralea canjerana 5 9,8 3,2 0,323 2,8 Guarea macrophylla 5 9,8 4,0 0,215 2,7 Ocotea indecora 6 11,7 3,2 0,144 2,5 Cestrum intermedium 4 7,8 3,2 0,285 2,5 Trichilia pallida 5 9,8 4,0 0,095 2,4 Machaerium scleroxylon 1 2,0 0,8 0,685 2,4 Cryptocarya aschersoniana 3 5,9 2,4 0,273 2,1 Alchornea glandulosa 3 5,9 1,6 0,349 2,0 Calycorectes acutatus 4 7,8 3,2 0,113 2,0 Cariniana legalis 4 7,8 3,2 0,113 2,0 Annona cacans 2 3,9 1,6 0,380 1,9 Ixora gardneriana 3 5,9 2,4 0,127 1,6 Ocotea beulahiae 3 5,9 2,4 0,109 1,6 Zanthoxylum acuminatum 3 5,9 2,4 0,107 1,6
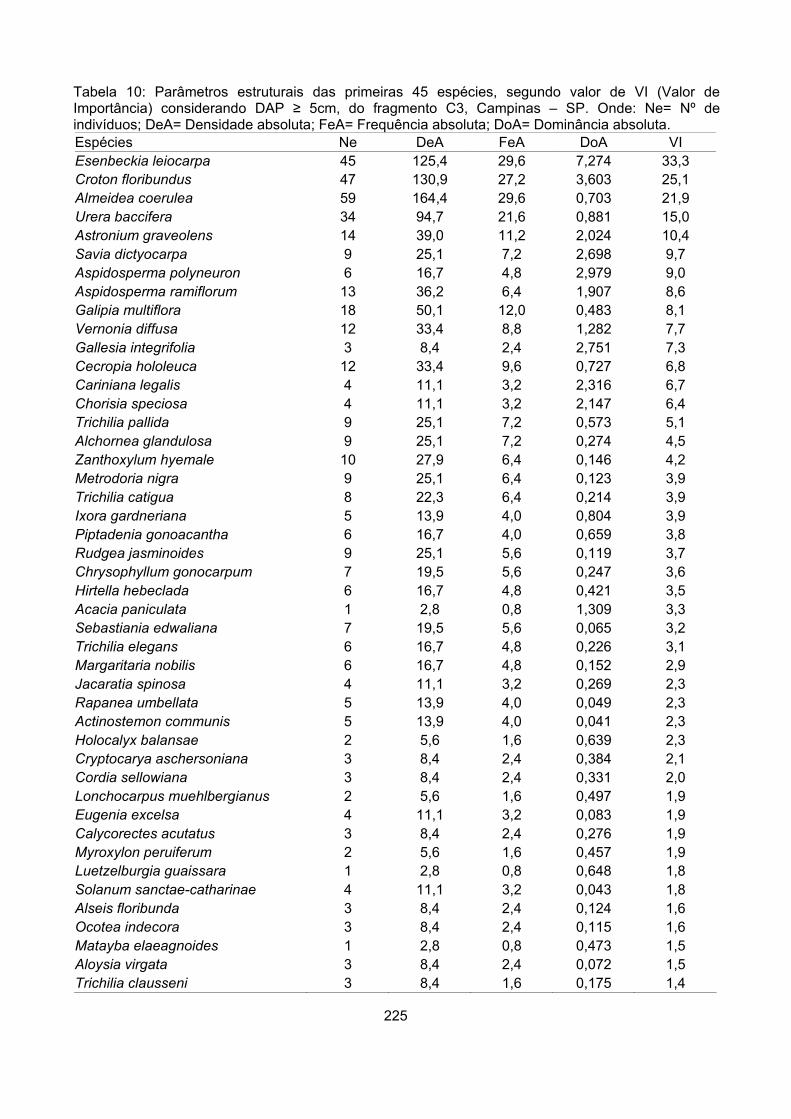
225
Tabela 10: Parâmetros estruturais das primeiras 45 espécies, segundo valor de VI (Valor de Importância) considerando DAP ≥ 5cm, do fragmento C3, Campinas – SP. Onde: Ne= Nº de indivíduos; DeA= Densidade absoluta; FeA= Frequência absoluta; DoA= Dominância absoluta. Espécies Ne DeA FeA DoA VI Esenbeckia leiocarpa 45 125,4 29,6 7,274 33,3 Croton floribundus 47 130,9 27,2 3,603 25,1 Almeidea coerulea 59 164,4 29,6 0,703 21,9 Urera baccifera 34 94,7 21,6 0,881 15,0 Astronium graveolens 14 39,0 11,2 2,024 10,4 Savia dictyocarpa 9 25,1 7,2 2,698 9,7 Aspidosperma polyneuron 6 16,7 4,8 2,979 9,0 Aspidosperma ramiflorum 13 36,2 6,4 1,907 8,6 Galipia multiflora 18 50,1 12,0 0,483 8,1 Vernonia diffusa 12 33,4 8,8 1,282 7,7 Gallesia integrifolia 3 8,4 2,4 2,751 7,3 Cecropia hololeuca 12 33,4 9,6 0,727 6,8 Cariniana legalis 4 11,1 3,2 2,316 6,7 Chorisia speciosa 4 11,1 3,2 2,147 6,4 Trichilia pallida 9 25,1 7,2 0,573 5,1 Alchornea glandulosa 9 25,1 7,2 0,274 4,5 Zanthoxylum hyemale 10 27,9 6,4 0,146 4,2 Metrodoria nigra 9 25,1 6,4 0,123 3,9 Trichilia catigua 8 22,3 6,4 0,214 3,9 Ixora gardneriana 5 13,9 4,0 0,804 3,9 Piptadenia gonoacantha 6 16,7 4,0 0,659 3,8 Rudgea jasminoides 9 25,1 5,6 0,119 3,7 Chrysophyllum gonocarpum 7 19,5 5,6 0,247 3,6 Hirtella hebeclada 6 16,7 4,8 0,421 3,5 Acacia paniculata 1 2,8 0,8 1,309 3,3 Sebastiania edwaliana 7 19,5 5,6 0,065 3,2 Trichilia elegans 6 16,7 4,8 0,226 3,1 Margaritaria nobilis 6 16,7 4,8 0,152 2,9 Jacaratia spinosa 4 11,1 3,2 0,269 2,3 Rapanea umbellata 5 13,9 4,0 0,049 2,3 Actinostemon communis 5 13,9 4,0 0,041 2,3 Holocalyx balansae 2 5,6 1,6 0,639 2,3 Cryptocarya aschersoniana 3 8,4 2,4 0,384 2,1 Cordia sellowiana 3 8,4 2,4 0,331 2,0 Lonchocarpus muehlbergianus 2 5,6 1,6 0,497 1,9 Eugenia excelsa 4 11,1 3,2 0,083 1,9 Calycorectes acutatus 3 8,4 2,4 0,276 1,9 Myroxylon peruiferum 2 5,6 1,6 0,457 1,9 Luetzelburgia guaissara 1 2,8 0,8 0,648 1,8 Solanum sanctae-catharinae 4 11,1 3,2 0,043 1,8 Alseis floribunda 3 8,4 2,4 0,124 1,6 Ocotea indecora 3 8,4 2,4 0,115 1,6 Matayba elaeagnoides 1 2,8 0,8 0,473 1,5 Aloysia virgata 3 8,4 2,4 0,072 1,5 Trichilia clausseni 3 8,4 1,6 0,175 1,4


















