
CAMELET: Salute e campi elettromagnetici -...
42
CAMELET: "Salute e campi elettromagnetici" Meccanismi di interazione M. Liberti , F. Apollonio, G. d’Inzeo, F. Maggio, P. Marracino, C. Merla, A. Paffi Centro InterUniversitario per lo Studio dei Campi Elettromagnetici e i Sistemi Centro InterUniversitario per lo Studio dei Campi Elettromagnetici e i Sistemi Biologici Biologici Dip. Ingegneria Elettronica, Università di Roma“La Sapienza” Dip. Ingegneria Elettronica, Università di Roma“La Sapienza” Convegno finale del progetto CAMELET Roma, 29-30 ottobre 2009
Transcript of CAMELET: Salute e campi elettromagnetici -...

CAMELET: "Salute e campi elettromagnetici"
Meccanismi di interazione
M. Liberti, F. Apollonio, G. d’Inzeo, F. Maggio, P. Marracino, C. Merla, A. Paffi
Centro InterUniversitario per lo Studio dei Campi Elettromagnetici e i Sistemi Centro InterUniversitario per lo Studio dei Campi Elettromagnetici e i Sistemi BiologiciBiologici
Dip. Ingegneria Elettronica, Università di Roma“La Sapienza”Dip. Ingegneria Elettronica, Università di Roma“La Sapienza”
Convegno finale del progetto CAMELETRoma, 29-30 ottobre 2009

Slide 2/42CAMELET,29-30 Ottobre 2009
Il problema scientifico
mec
can
ism
im
ecca
nis
mi
Bassa frequenza (ELF): 50 Hz
Alta frequenza (RF): ~ GHz
safety safety
guidelinesguidelines
medical medical applicationsapplications
EFFETTIBiologici
E, JE, J
E, SARE, SAR

Slide 3/42CAMELET,29-30 Ottobre 2009
Classificazione degli effetti a ELF
EMF-NET Effects of the Exposure to Electromagnetic Fields: From Science to Public Health and Safer Workplace (2004-2008)

Slide 4/42CAMELET,29-30 Ottobre 2009
Classificazione degli effetti a RF
Type of biological system Evidence for effects
(Classification based on studies published in years 2000-2007)
BLOOD BRAIN BARRIER
In vitro studies Inadequate evidence
Short-term in vivo studies Evidence suggesting lack of effects
Long-term in vivo studies Evidence suggesting lack of effects
AUDITORY SYSTEM
In vitro studies Inadequate evidence
In vivo studies Evidence suggesting lack of effects
Human studies Evidence suggesting lack of effect
BEHAVIOUR
In vivo studies: Spatial memory and place learning Limited evidence of an effect
In vivo studies: Effects on immature animals Inadequate evidence
NERVOUS SYSTEM
Human studies: EEG and Neurophysiology Limited evidence of an effect
Human studies: Cognition and Sleep Limited evidence of lack of effecst
In vivo studies Inadequate evidence
CARDIOVASCULAR SYSTEM
Human studies: Blood pressure Limited evidence of an effect
Human studies: ECG and HRV Inadequate evidence
In vivo studies Inadequate evidence
EM
F-N
ET:
Eff
ects
of t
he
Exp
osu
re
to E
lect
rom
agne
tic F
ield
s:
Fro
m S
cien
ce t
o P
ublic
Hea
lth
and
Saf
er W
ork
pla
ce (
200
4-20
08)

Slide 5/42CAMELET,29-30 Ottobre 2009
Exposure of 5-HT1B to B (50 Hz ; 0÷ 2.5 mT)
receptor desensibilization
Massot et al., 1999Espinosa et al., 2006
Exposure of A2A and A3 to pulsed B (75 Hz; 0.2÷ 3.5 mT)
Varani et al., 2002,2003
modification in receptor functionality
In vitro experimental results
Exposure of chicken brain to modulated and ELF fields
Blackman, 1982-1989
increase in Calcium efflux
Azione specifica di ben definiti campi magnetici su alcuni recettori di membrana
Evidenza di ‘effetti a finestra’ in frequenza e ampiezza
frequency
ampl
itude
frequency
Specificità di alcuni effetti

Slide 6/42CAMELET,29-30 Ottobre 2009
Il dibattito internazionale
IPOTESI
PLAUSIBILITA’
TESTABILITA’/VALIDAZIONE SPERIMENTALE
SOGLIASpecificità del segnale applicato
kT/dumping e Rumore biologico (i.e 1/f) (REALISMO)
Def. of Plausibility
- [..] For a mechanism to be plausible it should link electric- and magnetic-field exposure by means of a biophysical mechanism to the beginning of a chain of events of the sort shown in Figure. [..]
- [..] The term “mechanism of interaction” is generally considered to include the full chain of events, although the term is sometimes used to refer only to the initial transduction step or to the initial biological signal after that first step.

Slide 7/42CAMELET,29-30 Ottobre 2009
La scatola nera dei meccanismi?
Campi EM Effetticome?
Sistema Biologico
mec
can
ism
im
ecca
nis
mi

Slide 8/42CAMELET,29-30 Ottobre 2009
Complessità biologica? (1967)
“Biologists can be divided into two classes: experimentalists who observe things that cannot be explained, and theoreticians who explain things that cannot be observed.”
A. Katchalsky, Noneq. Proc. in Bioph., 1967
Uguale per il Bioelettromagnetismo?

Slide 9/42CAMELET,29-30 Ottobre 2009
Complessità biologica (2000)
There is now a golden opportunity for system level analysis to be grounded in molecular-level understanding, resulting in a continuous spectrum of
knowledge.H. Kitano, Science, 2002

Slide 10/42CAMELET,29-30 Ottobre 2009
Complessità biologica (2003-2008)
Hunter & Borg, Nature Rev., 2003

Slide 11/42CAMELET,29-30 Ottobre 2009
Uguale per il Bioelettromagnetismo?

Slide 12/42CAMELET,29-30 Ottobre 2009
L’approccio dell’OMS (1997)

Slide 13/42CAMELET,29-30 Ottobre 2009
Uguale per il Bioelettromagnetismo?
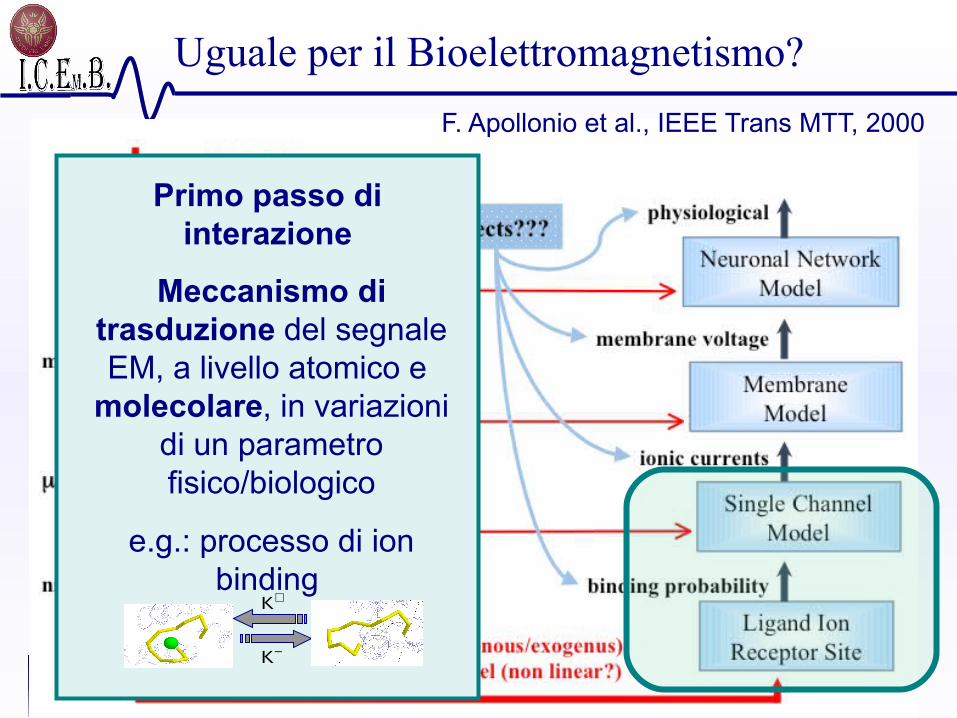
Transduzione del campo EM
(comprensione del livello molecolare)
F. Apollonio et al., IEEE Trans MTT, 2000
Analisi al livello del sistema
Slide 14/42CAMELET,29-30 Ottobre 2009
Uguale per il Bioelettromagnetismo?
F. Apollonio et al., IEEE Trans MTT, 2000
Primo passo di interazione
Meccanismo di trasduzione del segnale EM, a livello atomico e
molecolare, in variazioni di un parametro fisico/biologico
e.g.: processo di ion binding
+K
−K

Slide 15/42CAMELET,29-30 Ottobre 2009
Uguale per il Bioelettromagnetismo?
F. Apollonio et al., IEEE Trans MTT, 2000
Catena/successione di modelli biofisici che
includono la presenza di un campo
elettromagnetico
Ad ogni livello di complessità, i modelli sono testabili sperimentalmente
e contengono sufficiente realismo per studiare
l’interazione con il campo EM

Slide 16/42CAMELET,29-30 Ottobre 2009
++
+
+
-+
+
+ - -
-
-
-
-
+
+
-
k12
k21
k23
k32
S + E ES E + P
k12
k21
k23
k32
S + E ES E + P
Chiabrera et al. 1984, Weaver et al. 2000
ki ⇐ campi EM
Moto e [ ] ligandi ⇐ campi EM
Binding ⇐ campi EM
I ions⇐ campi EM
Funzioni celluari ⇐ campi EM
Trasduzione del campo EM
Reazioni enzimatiche nell’ambiente extracellulare:
Binding degli ioni al loro specifico sito recettore
Prodotti extracellulari (i.e. ioni liberi)
Flussi ionici attraverso la membrana
Reazioni enzimatiche interne alla cellula

Slide 17/42CAMELET,29-30 Ottobre 2009
Lo ione viene considerato
come una particella
puntiforme polarizzabile di
massa M e carica Q.
Lo ione viene considerato
come una particella
puntiforme polarizzabile di
massa M e carica Q.1) Leggi della fisica classica
2) Meccanica quantistica
In passato… target biologico semplificato
Quando sono coinvolte reazionichimiche è necessario
considerare la distribuzione elettronica
del target. Modelli semplificati potrebbero
non essere adeguati

Slide 18/42CAMELET,29-30 Ottobre 2009
QM (Meccanica Quantistica)
• DFT methods (Density Functional Theory)• post HF methods
• HF (Hartree Fock theory)
• semiempirical methods
ab initio
Estremamente accurati ma in grado di gestire sistemi semplici
dinamica del sistema
• SD (Stochastic Dynamic)
• BD (Brownian Dynamic)
• MD (Molecular Dynamic)
MM(Meccanica Molecolare)
Meno accurati ma in grado di gestire sistemi complessi
Metodi per l’analisi di strutturemolecolari: chimica computazionale

Slide 19/42CAMELET,30-29 Ottobre 2009
Calcolo delle proprietà elettroniche del sistema attraverso la diagonalizzazione della matrice
Hamiltoniana associata all’ambiente perturbato(es., campo elettrico)
Metodi Ibridi (QM/MM)
Perturbed Matrix Method (PMM*)combinano accuratezza e complessità
*Aschi et al. Chem. Phys. Lett. 2002, Spezia et al., Chem. Phys. Lett. 2002, Aschi et al., J.Comp. Chem. 2004
Metodi: chimica computazionale

Slide 20/42CAMELET,29-30 Ottobre 2009
Sito attivo
Dove avviene la reazione
Intera Proteina
Rappresenta una perturbazione del
sito attivo
myoglobin ~ 1360 atoms
PMM MethodPMM Method
Campo EM
Rappresenta una perturbazione del sito
attivo
Metodi: chimica computazionale

Slide 21/42CAMELET,29-30 Ottobre 2009
** Daidone et al., JACS, 2005; Schmidt et al., PNAS, 2005.
Intensità molecolari: Intensità molecolari: 108-109 V/m
Centro di reazioneCentro di reazioneTransizioni stati
elettronici:
(<<fs, sub-fs) (<<fs, sub-fs) *
Intera proteinaIntera proteinaVariazioni lente delle
catene aminoacidi
(da 100 ps a ns) (da 100 ps a ns) *
Azione indiretta di un campo campo MWMW sul sito di reazione
attraverso un accoppiamento con i moti quasi diffusivi
della proteina
Ipotesi di accoppiamento con MW
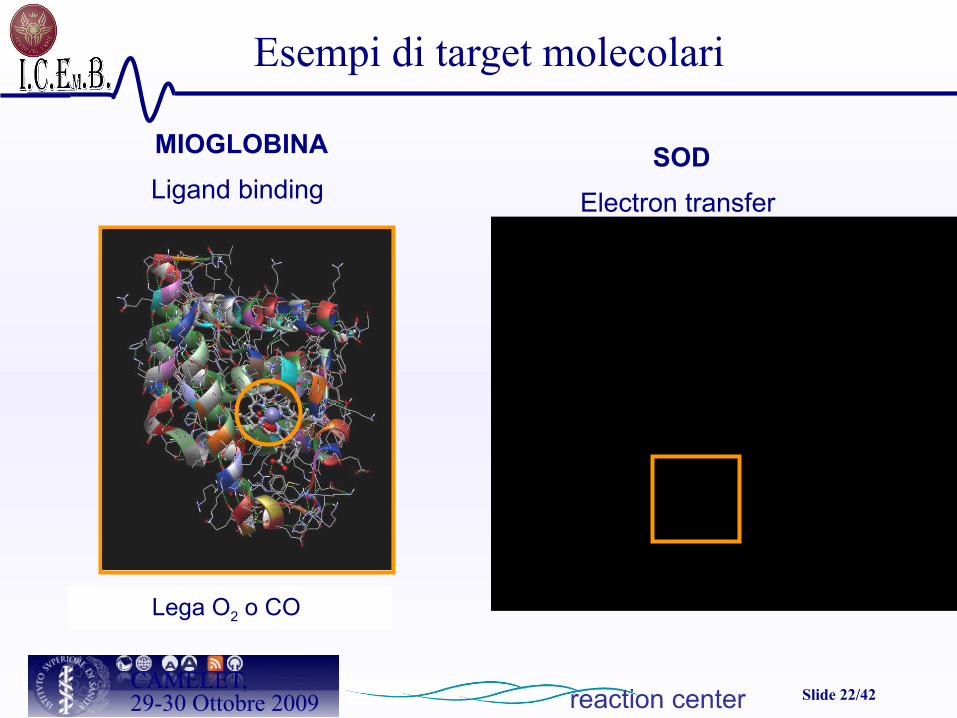
Slide 22/42CAMELET,29-30 Ottobre 2009
MIOGLOBINA
Ligand binding
Lega O2 o CO
Esempi di target molecolari
reaction center
QuickTimeᆰ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
SOD
Electron transfer

Slide 23/42CAMELET,29-30 Ottobre 2009
unbindingenergy barrier
bindingenergy barrier
dFe-CO
Fe2+ CO
Osservabile: barriere di energia libera

Slide 24/42CAMELET,29-30 Ottobre 2009 *(unpaired t-student test, hyp of Gaussian distributed noise)
ΔABinding
(kJ/mol)ΔAUnbinding
(kJ/mol)
mean std.dev.
mean std.dev.
Unexposed(n=5)
42.9 0.83 73.8 0.51
Exp. 103 V/m (n=7) 43.2 1.62 73.7 0.69
Exp. 104 V/m (n=3) 42.9 0.93 73.8 0.58
Nessuna differenza significativa tra i non esposti e gli esposti (CW, 1 GHz, 103 e 104 V/m) *
Risultati CO-Mioglobina
Statistiche
Apollonio et al, 2008

Slide 25/42CAMELET,29-30 Ottobre 2009
Length (ns)
Free Energy (kJ/mol)
Residual dipole (Cu-O2 bond) ( 10-19 C£チ )
Residual dipole (Cu-His48 bond) ( 10-19 C£チ )
unexp # 1 30 -60.2 2.18 -2.4 unexp # 2 30 -50.1 2.33 -2.77 unexp # 3 30 -65.3 2.23 -2.6 unexp # 4 30 -73.9 2.15 -2.7 unexp # 5 20 -72.5 2.68 -1.22 10^3 # 1 30 -73.2 2.21 -2.96 10^3 # 2 30 -73.17 2.17 -2.87 10^3 # 3 43 -18.6 2.16 -2.75 10^4 # 1 30 -58.8 2.32 -2.4 10^4 # 2 30 -70.59 2.2 -2.9 10^4 # 3 20 -48.48 2.18 -2.38 10^4 # 4 20 -76.84 2.38 -1.47 10^5 # 1 30 -57.82 2.14 -1.29 10^5 # 2 36 -60.4 2.17 -2.88 10^5 # 3 26 -60.69 2.14 -2.26 10^6 # 1 30 -55.19 2.2 -2.96 10^6 # 2 30 -89 2.55 -2 10^6 # 3 24 -66.6 2.18 -1.99 10^6 # 4 27 -40.7 2.2 -1.54
E field (V/m)f=1 GHz
E field=0
Risultati sulla reazione SOD
Apollonio et al, 2009

Slide 26/42CAMELET,29-30 Ottobre 2009
Risultati livello molecolare
(International School of Bioelectromagnetics on Interaction Mechanisms, Erice, 2006)
Campi CW ad 1 GHz Campi CW ad 1 GHz non perturbanonon perturbano né il né il processo di binding CO-Fe binding nella processo di binding CO-Fe binding nella
mioglobina né il processo di electron transfer mioglobina né il processo di electron transfer nella SOD in una scala temporale dei nsnella SOD in una scala temporale dei ns
Suggerimenti per studi futuri:Valutare l’azione di campi EM su proteine inserite in membrana attraverso simulazioni molecolari
Apollonio et al, 2008-2009

Slide 27/42CAMELET,29-30 Ottobre 2009
Su cosa si sta lavorandoSu cosa si sta lavorando
~80.000 atoms
Proteine di membrana
N,N Dimetil-Dodecyl-Oxide: zwitterionic micelle
Strutture micellari
5 HT - 1B
Slide 28/42CAMELET,29-30 Ottobre 2009
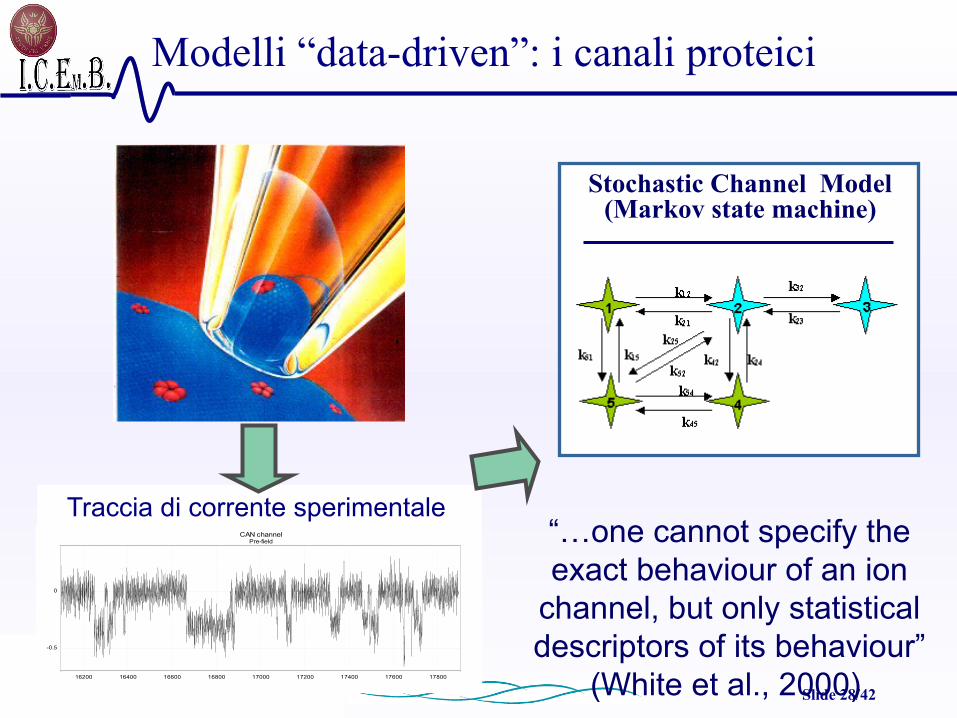
Modelli “data-driven”: i canali proteici
CAN channelPre-field
Time (ms)16200 16400 16600 16800 17000 17200 17400 17600 17800
Ionic current (pA)
-0.5
0
Traccia di corrente sperimentale“…one cannot specify the exact behaviour of an ion
channel, but only statistical descriptors of its behaviour”
(White et al., 2000)
Stochastic Channel Model (Markov state machine)

Slide 29/42CAMELET,29-30 Ottobre 2009
Voltage-gated K+ channel
Modelli di canali ionici di membranaModelli di canali ionici di membrana
'T80
60Vexp125.0n �
���
�� −−=β
( )'T
110
50Vexp
50V01.0n
-
│₩ --
--=a
( )'T
110
35Vexp
35V1.0m
-
│₩ --
--=a 'T
18
60Vexp0.4m �
���
�� −−=β
'T20
60Vexp07.0h �
���
�� −−=α 'T
110
30Vexp
11.0h
+����
�� −−
=β
Hodgkin & Huxley, 1952Voltage-gated Na+ channel
Voltage-gated Ca2+ L-type channel
( )1
10
V5.81exp
V5.8169.15e
-
│₩ -
-=a �
���
�� −=
86.10
Vexp29.0eb
Aradi & Holmes, 1999
Hodgkin & Huxley, 1952
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-----------
-----------
----
----
K +
Na +
Ca++
+-
External medium
Internal medium
K +
Na +
Ca++
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-----------
-----------
----
----
K +
Na +
Ca++
K +
Na +
Ca++
+-
External medium
Internal medium
K +
Na +
Ca++
K +
Na +
Ca++
EX
TR
AC
EL
LU
LA
R
SPA
CE
INT
RA
CE
LL
UL
AR
SPA
CE

Slide 30/42CAMELET,30-29 Ottobre 2009
Osservabili a livello di canaleOsservabili a livello di canale
Spettro di densità di potenza delle tracce di corrente
cross-correlazione
Nel tempo: tracce di corrente come da registrazioni sperimentali
In frequenza:
I risultati sono direttamente testabili:
esiste una corrispondenza 1:1 con l’osservabile
sperimentale

Slide 31/42CAMELET,29-30 Ottobre 2009
I(t) sin(2⋅ � ⋅ 50⋅ t)**
Pellegrino et al., BEMS 2006
Voltage-gated Na+ channel
Vm=-40mV , duration=60 s, n° runs=8, CW (1mV, 50Hz)
CROSS-CORRPSD
Risultati a 50 HzRisultati a 50 Hz

Slide 32/42CAMELET,29-30 Ottobre 2009
Segnale EMCW, fEM=50Hz
Canale (K+)
Wilcoxon Mann-Whitney test:Wilcoxon Mann-Whitney test: 0.01<P<0.05; 0.001<P<0.01; P<0.001************
Channel type Vm (mV) Minimumion current duration
Minimumnumber of runs
EM signal amplitude ΔVEM
K+ -40 60 s 8 1 mV
Na+ -40 40 s 8 0.8 mV
Ca2+ L-type 0 10 s 4 0.7 mV
Soglia dell’effetto per i canali ioniciSoglia dell’effetto per i canali ionici
Pellegrino et al., BEMS 2006

Slide 33/42CAMELET,29-30 Ottobre 2009
Modello di nerone HH
Questo modello riproduce l’attività di firing di un neurone eccitabile ed è il riferimento fondamentale per la modellistica neuronale
�
Cm dV dt = - gNa V - VL( ) - gK V - VK( ) - gL V - VL( ) + I0
gNa V( ) = � g NA m3h gK V( ) = � g kn4
dm dt = m� - m( ) t m
dh dt = h� - h( ) t h
dn dt = n� - n( ) t n
Hodgkin & Huxley, J Physiol, 1952

Slide 34/42CAMELET,29-30 Ottobre 2009
Modelli HH stocastici
CAN channelPre-field
Time (ms)16200 16400 16600 16800 17000 17200 17400 17600 17800
Ionic current (pA)
-0.5
0
Traccia di corrente sperimentale“…one cannot specify the exact behaviour of an ion
channel, but only statistical descriptors of its behaviour”
(White et al., 2000)
Stochastic Channel Model (Markov state machine)
Introduzione del rumore dei canalinel modello HH
Apollonio et al., 2000-2002
Slide 35/42CAMELET,29-30 Ottobre 2009
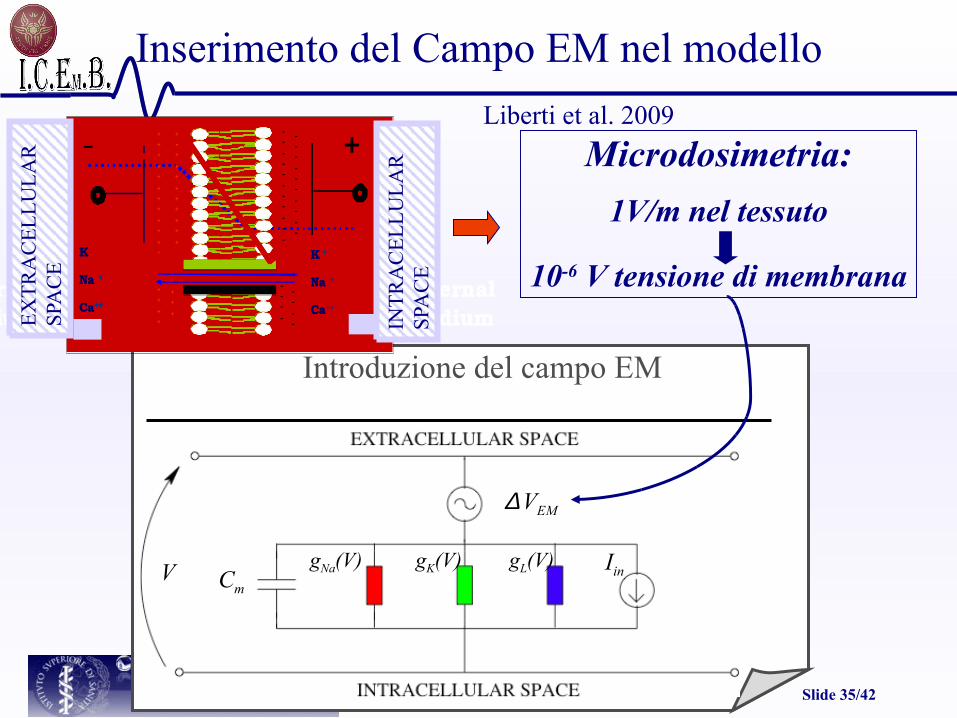
Introduzione del campo EM
Inserimento del Campo EM nel modello
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-----------
-----------
----
----
K +
Na +
Ca++
+-
External medium
Internal medium
K +
Na +
Ca++
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-----------
-----------
----
----
K +
Na +
Ca++
K +
Na +
Ca++
+-
External medium
Internal medium
K +
Na +
Ca++
K +
Na +
Ca++
EX
TR
AC
EL
LU
LA
R
SPA
CE
INT
RA
CE
LL
UL
AR
S
PAC
E
V gNa(V) gK(V) gL(V)Cm
Iin
∆ VEM
Microdosimetria:
1V/m nel tessuto
10-6 V tensione di membrana
Liberti et al. 2009

Slide 36/42CAMELET,29-30 Ottobre 2009
Codifica neuronale di un campo EM
SR observedSR not observedSR observedSR not observed
M. Gianni et al., Biol. Cyb., 2006
Un singolo neurone, sfruttando anche il suo rumore intrinseco, è in grado di ottimizzare la codifica di un segnale EM di bassa frequenza
ed elevata intensità (kV/m nel tessuto)
di interesse più delle applicazioni medicali che ai fini protezionistici

Slide 37/42CAMELET,29-30 Ottobre 2009
Nei diversi neuroni il rumore dipende dal numero di canali
pyramidal
Nodo di Ranvier
interneuron
gc
COMPARTIMENTODENDRITICO
Modellizzazione delle diverse forme neuronali
COMPARTIMENTOASSO-SOMATICO
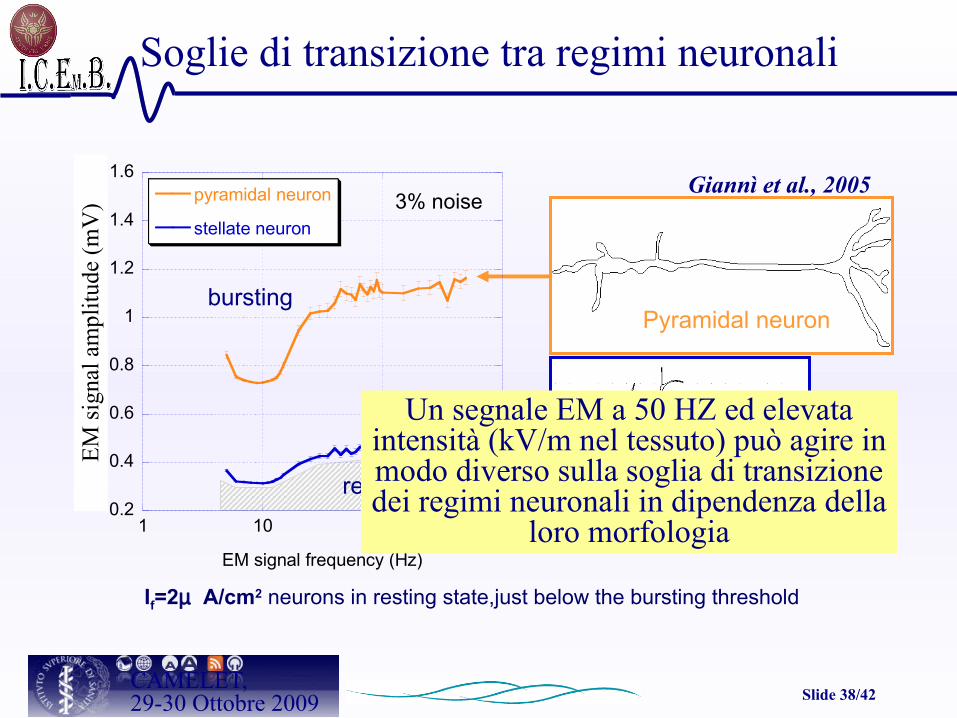
Slide 38/42CAMELET,29-30 Ottobre 2009
If=2µ A/cm2 neurons in resting state,just below the bursting threshold
bursting
3% noise
resting
Giannì et al., 2005
Pyramidal neuron
Stellate neuron
Soglie di transizione tra regimi neuronali
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1 10 100 1000
pyramidal neuron
stellate neuron
EM signal amplitude (mV)
EM signal frequency (Hz)
EM
sig
nal a
mpl
itud
e (m
V)
Un segnale EM a 50 HZ ed elevata intensità (kV/m nel tessuto) può agire in modo diverso sulla soglia di transizione dei regimi neuronali in dipendenza della
loro morfologia

Slide 39/42CAMELET,29-30 Ottobre 2009
Topologie Feedforward •Reti sensoriali•Strati corticali
Topologie Feedforward •Reti sensoriali•Strati corticali
Connessioni sinaptiche
[T ] j = 1mM Vi ≥ -20 mV
[ ]( ) wrgrrT
dt
dr ~1 +--= ba
( )synsynsyn EVrgI -=
Sinapsi stocastiche a dinamica di recettore
Tranquilli et al., 2004
�
C +Tb
� � �
a� � � O
Connessioni tra più cellule

Slide 40/42CAMELET,30-29 Ottobre 2009
Esperimenti dry (simulazioni)IN
PU
T S
IGN
AL
:E
M f
ield
, 50
Hz,
50
µV
25 primary neurons
25 primary neurons
25 primary neurons
25 gruppo di25 neuroni connessi
2nd layer neurons (25)
25 neuroni secondari connessi
3nd layer neuron
1 neurone terziario con 25 secondari
∆ VEM= 50 Hz, 50 µ VIin=I0+INOISE
⇒ Gaussian process, <Iin>= 4 µ A/cm2,D ∈ [0.2÷60] µ A2/cm4
VgNa(V) gK(V) gL(V)
Cm
Iin
∆ VEM
625 neuroni primari
VgNa(V) gK(V) gL(V)
Cm
Isyn
INOISE = 0,Isyn⇐ 25 pre-synaptic neurons
coincidence neurons(25⇒ 2nd and 1⇒ 3rd layers)
Sequenza di uscita
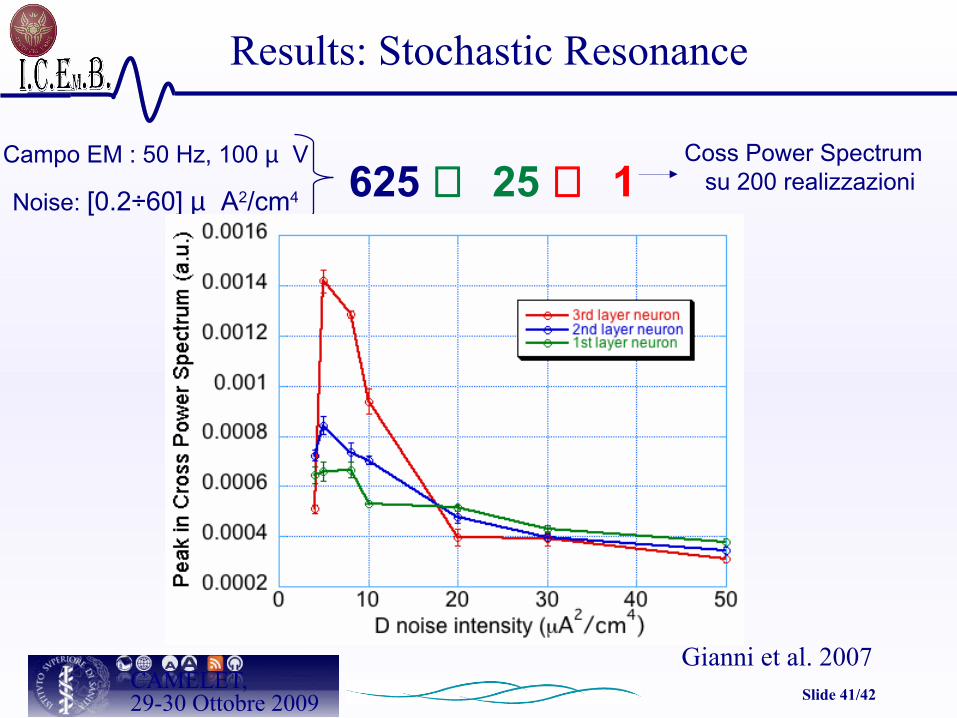
Slide 41/42CAMELET,29-30 Ottobre 2009
625 ⇒ 25 ⇒ 1Campo EM : 50 Hz, 100 µ V
Noise: [0.2÷60] µ A2/cm4
Coss Power Spectrum su 200 realizzazioni
Results: Stochastic Resonance
Gianni et al. 2007

Slide 42/42CAMELET,29-30 Ottobre 2009
Conclusioni
• I meccanismi di interazione devono soddisfare condizioni di plausibilita e testabilità, ed identificare delle soglie.
• I meccanismi devono individuare il passo di trasduzione e tutta la catena di eventi che possa condurre all’identificazione di un effetto.
• Attraverso l’uso di appropriate tecniche (es: simulazioni atomistiche per il livello molecolare) è possibile creare un percorso quantitativo attraverso la scala biologica di complessità.
• Campi CW ad 1 GHz ad intensità inferiori di quelle molecolari non perturbano le reazioni esaminate in una scala temporale dei ns.
• Valutare l’azione di campi EM su proteine inserite in membrana attraverso simulazioni molecolari (ad intensità elevate).
• Un singolo neurone, sfruttando anche il suo rumore intrinseco, è in grado di ottimizzare la codifica di un segnale EM a 50 Hz ed elevata intensità (kV/m nel tessuto).
• Può esistere un effetto cooperativo tra cellule neuronali che abbassa la soglia dei segnali rilevabili (100 V/m nel tessuto).


















