
Cadmium inhibits acid secretion in stimulated frog gastric mucosa
8
Cadmium inhibits acid secretion in stimulated frog gastric mucosa Andrea Gerbino a, ⁎, Lucantonio Debellis a , Rosa Caroppo a , Silvana Curci b , Matilde Colella a a Dept. of General and Environmental Physiology, University of Bari, 70126 Bari, Italy b VA Boston Healthcare System and the Department of Surgery, Harvard Medical School, and Brigham and Women's Hospital, 1400 VFW Parkway, West Roxbury MA 02132 USA abstract article info Article history: Received 29 January 2010 Revised 12 March 2010 Accepted 15 March 2010 Available online 20 March 2010 Keywords: Cadmium Gastric acid secretion Short circuit current Omeprazole H + /K + -ATPase Cadmium, a toxic environmental pollutant, affects the function of different organs such as lungs, liver and kidney. Less is known about its toxic effects on the gastric mucosa. The aim of this study was to investigate the mechanisms by which cadmium impacts on the physiology of gastric mucosa. To this end, intact amphibian mucosae were mounted in Ussing chambers and the rate of acid secretion, short circuit current (I sc ), transepithelial potential (V t ) and resistance (R t ) were recorded in the continuous presence of cadmium. Addition of cadmium (20 μM to 1 mM) on the serosal but not luminal side of the mucosae resulted in inhibition of acid secretion and increase in NPPB-sensitive, chloride-dependent short circuit current. Remarkably, cadmium exerted its effects only on histamine-stimulated tissues. Experiments with TPEN, a cell-permeant chelator for heavy metals, showed that cadmium acts from the intracellular side of the acid secreting cells. Furthermore, cadmium-induced inhibition of acid secretion and increase in I sc cannot be explained by an action on: 1) H 2 histamine receptor, 2) Ca 2+ signalling 3) adenylyl cyclase or 4) carbonic anhydrase. Conversely, cadmium was ineffective in the presence of the H + /K + -ATPase blocker omeprazole suggesting that the two compounds likely act on the same target. Our findings suggest that cadmium affects the functionality of histamine-stimulated gastric mucosa by inhibiting the H + /K + -ATPase from the intracellular side. These data shed new light on the toxic effect of this dangerous environmental pollutant and may result in new avenues for therapeutic intervention in acute and chronic intoxication. © 2010 Elsevier Inc. All rights reserved. Introduction Cadmium is a highly toxic, non-essential heavy metal that displays significant threats to living organisms by interacting with vital cellular functions. The effects induced by acute and/or chronic exposure of target organs and processes (kidneys, liver, lungs, cardiovascular, immune and reproductive systems) to cytotoxic concentrations of cadmium have been intensely investigated (Jarup and Akesson, 2009). These studies defined diverse mechanisms of cadmium uptake, accumulation and toxicity that strongly depend on cell type and condition/duration of exposure. Gastrointestinal ingestion through contaminated food and drink- ing water is considered to be the main source of cadmium for non- occupationally exposed, non-smokers (Jarup and Akesson, 2009). Gastric mucosa is therefore the epithelium that is first exposed to food and water containing high levels of cadmium. Previous studies concerning the chronic effect of cadmium on rat gastric mucosa showed that a 30 day exposure leads to significant accumulation of the heavy metal in the mucosa and significant reduction in the basal acid output (Asar et al., 2000; Oner et al., 1994). According to Oner et al. the resulting lipid peroxidation would bring about decreased production of gastric mucin and PGE 2 and impairment of endogenous protective mechanisms (Oner et al., 1994). Asar et al. suggest that the significant decline in the recorded basal acid output in response to cadmium may be explained by a loss of parietal cells population (Asar et al., 2000). Moreover, Dupuy and Szabo showed that rats pre-treated with cadmium (2.5 mg/100 g body weight) for 30 min presented a significant inhibition of gastric acid output after 1 h of piloric ligation (Dupuy and Szabo, 1986). These studies were all performed in whole stomach ex vivo preparations where the gastric acid output was measured after titration of the extracted luminal fluid. Therefore, no information regarding the potential effects of acute exposure to this heavy metal on the cellular mechanisms leading to gastric acid secretion is available. Acid secretion by gastric oxyntopeptic/parietal cells is an intricate process that is tightly regulated by paracrine, endocrine and neural factors. Stimulation of acid secretion typically involves an initial elevation in cAMP followed by activation of a cAMP-dependent protein kinase (PKA) that triggers the translocation and insertion of the gastric proton pump (H + /K + -ATPase) together with K + and Cl − conductances into the apical plasma membrane of acid secreting cells. The process ultimately produces net output of protons, chloride ions and water across the apical plasma membrane of these cells (Yao and Forte, 2003; Gerbino et al., 2007). Toxicology and Applied Pharmacology 245 (2010) 264–271 Abbreviations: I sc , short circuit current; V t , transepithelial potential; R t , transepithe- lial resistance; CA, carbonic anhydrase; AC, adenylyl cyclase. ⁎ Corresponding author. University of Bari, Dept of General and Environmental Physiology, Via Amendola 165/A, 70126 Bari, Italy. Fax: + 39 080 5443388. E-mail address: [email protected] (A. Gerbino). 0041-008X/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2010.03.010 Contents lists available at ScienceDirect Toxicology and Applied Pharmacology journal homepage: www.elsevier.com/locate/ytaap
-
Upload
andrea-gerbino -
Category
Documents
-
view
214 -
download
0
Transcript of Cadmium inhibits acid secretion in stimulated frog gastric mucosa

Toxicology and Applied Pharmacology 245 (2010) 264–271
Contents lists available at ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /ytaap
Cadmium inhibits acid secretion in stimulated frog gastric mucosa
Andrea Gerbino a,⁎, Lucantonio Debellis a, Rosa Caroppo a, Silvana Curci b, Matilde Colella a
a Dept. of General and Environmental Physiology, University of Bari, 70126 Bari, Italyb VA Boston Healthcare System and the Department of Surgery, Harvard Medical School, and Brigham and Women's Hospital, 1400 VFW Parkway, West Roxbury MA 02132 USA
Abbreviations: Isc, short circuit current; Vt, transepithlial resistance; CA, carbonic anhydrase; AC, adenylyl cyc⁎ Corresponding author. University of Bari, Dept o
Physiology, Via Amendola 165/A, 70126 Bari, Italy. Fax:E-mail address: [email protected] (A. Gerbin
0041-008X/$ – see front matter © 2010 Elsevier Inc. Adoi:10.1016/j.taap.2010.03.010
a b s t r a c t
a r t i c l e i n f oArticle history:Received 29 January 2010Revised 12 March 2010Accepted 15 March 2010Available online 20 March 2010
Keywords:CadmiumGastric acid secretionShort circuit currentOmeprazoleH+/K+-ATPase
Cadmium, a toxic environmental pollutant, affects the function of different organs such as lungs, liver andkidney. Less is known about its toxic effects on the gastric mucosa. The aim of this study was to investigatethe mechanisms by which cadmium impacts on the physiology of gastric mucosa. To this end, intactamphibian mucosae were mounted in Ussing chambers and the rate of acid secretion, short circuit current(Isc), transepithelial potential (Vt) and resistance (Rt) were recorded in the continuous presence of cadmium.Addition of cadmium (20 µM to 1 mM) on the serosal but not luminal side of the mucosae resulted ininhibition of acid secretion and increase in NPPB-sensitive, chloride-dependent short circuit current.Remarkably, cadmium exerted its effects only on histamine-stimulated tissues. Experiments with TPEN, acell-permeant chelator for heavy metals, showed that cadmium acts from the intracellular side of the acidsecreting cells. Furthermore, cadmium-induced inhibition of acid secretion and increase in Isc cannot beexplained by an action on: 1) H2 histamine receptor, 2) Ca2+ signalling 3) adenylyl cyclase or 4) carbonicanhydrase. Conversely, cadmium was ineffective in the presence of the H+/K+-ATPase blocker omeprazolesuggesting that the two compounds likely act on the same target. Our findings suggest that cadmium affectsthe functionality of histamine-stimulated gastric mucosa by inhibiting the H+/K+-ATPase from theintracellular side. These data shed new light on the toxic effect of this dangerous environmental pollutantand may result in new avenues for therapeutic intervention in acute and chronic intoxication.
elial potential; Rt, transepithe-lase.f General and Environmental+39 080 5443388.o).
ll rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Introduction
Cadmium is a highly toxic, non-essential heavy metal that displayssignificant threats to living organisms by interacting with vital cellularfunctions. The effects induced by acute and/or chronic exposure oftarget organs and processes (kidneys, liver, lungs, cardiovascular,immune and reproductive systems) to cytotoxic concentrations ofcadmiumhave been intensely investigated (Jarup and Akesson, 2009).These studies defined diverse mechanisms of cadmium uptake,accumulation and toxicity that strongly depend on cell type andcondition/duration of exposure.
Gastrointestinal ingestion through contaminated food and drink-ing water is considered to be the main source of cadmium for non-occupationally exposed, non-smokers (Jarup and Akesson, 2009).Gastric mucosa is therefore the epithelium that is first exposed to foodand water containing high levels of cadmium. Previous studiesconcerning the chronic effect of cadmium on rat gastric mucosashowed that a 30 day exposure leads to significant accumulation ofthe heavy metal in the mucosa and significant reduction in the basal
acid output (Asar et al., 2000; Oner et al., 1994). According to Oner etal. the resulting lipid peroxidation would bring about decreasedproduction of gastric mucin and PGE2 and impairment of endogenousprotective mechanisms (Oner et al., 1994). Asar et al. suggest that thesignificant decline in the recorded basal acid output in response tocadmiummay be explained by a loss of parietal cells population (Asaret al., 2000). Moreover, Dupuy and Szabo showed that rats pre-treatedwith cadmium (2.5 mg/100 g body weight) for 30 min presented asignificant inhibition of gastric acid output after 1 h of piloric ligation(Dupuy and Szabo, 1986). These studies were all performed in wholestomach ex vivo preparations where the gastric acid output wasmeasured after titration of the extracted luminal fluid. Therefore, noinformation regarding the potential effects of acute exposure to thisheavy metal on the cellular mechanisms leading to gastric acidsecretion is available.
Acid secretion by gastric oxyntopeptic/parietal cells is an intricateprocess that is tightly regulated by paracrine, endocrine and neuralfactors. Stimulation of acid secretion typically involves an initialelevation in cAMP followed by activation of a cAMP-dependentprotein kinase (PKA) that triggers the translocation and insertion ofthe gastric proton pump (H+/K+-ATPase) together with K+ and Cl−
conductances into the apical plasmamembrane of acid secreting cells.The process ultimately produces net output of protons, chloride ionsand water across the apical plasma membrane of these cells (Yao andForte, 2003; Gerbino et al., 2007).

265A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
According to available literature cadmium might theoreticallyaffect gastric acid secretion at several different levels: i) cadmium caninterfere with the activity of different plasma membrane receptors(Coddou et al., 2005; Giridhar et al., 1992; Stoica et al., 2000), hencethe heavy metal might interact with the histamine receptor (H2) and/or with transporters located on the basolateral membrane andinvolved in acid secretion; ii) it can interfere with essentialintracellular messenger cascades, Ca2+ and cAMP signalling, (see(Thevenod, 2009); iii) cadmium is known to inhibit carbonicanhydrase by displacing key metal components such as zinc fromthe proper binding site, thus it may affect the secretory processes thatdepend on HCO3
− availability in the gastric mucosa; and iv) it couldpotentially block gastric (H+/K+)-ATPase activity. Regarding thelatter possibility, it has already been shown that cadmium impacts onthe activity of most members of this class of P-ATPases (Hechtenbergand Beyersmann, 1991; Herak-Kramberger et al., 2000; Kinne-Saffranet al., 1993; Verbost et al., 1988; Zhang et al., 1990) including thegastric proton pump (Hongo et al., 1990).
The purpose of this paper was to determine the mechanism bywhich micromolar concentrations (ranging from 20 to 100 µM) ofcadmium can affect acid secretion in amphibian gastric mucosa.Although plasma cadmium levels in the general population aretypically less than 0.02 µM, we used this range of concentrationsbecause significantly higher levels are found in smokers and peoplewith occupational exposure (Tsalev and Zaprianov, 1984).
Here, by using a classic, straightforward electrophysiologicaltechnique on a well characterized experimental model such as theintact amphibian gastric mucosa (Gerbino et al., 2007; Caroppo et al.,2004, 2001), we found that exposure to serosal cadmium producesinhibition of histamine-stimulated acid secretion. This in turn causesan elevation of short circuit current, which is correlated with theelectrogenic Cl− flux known to be regulated by the proton pump(Reenstra et al., 1987).
Materials and methods
Tissue preparation and solutions. Experiments were performed ongastric fundus mucosa of Rana esculenta in accordance with the Italianguidelines for animal experiments. The frogs were kept in an aquariumat room temperature and killed by decapitation and destruction of thespinal cord. The stomach was isolated and the muscle layer andconnective tissue were removed by blunt dissection. Where needed,tissues were kept in resting state by adding 100 µM cimetidine. Tostimulate acid secretion a maximal concentration of histamine(500 µM) was added to the serosal solution and maximal rate of acidsecretion was reached within 60–90 min. After stable secretion valueswere reached, cadmium (20–100 µM) and/or other drugs were addedto the serosal and/or luminal solution. All chemicals were of reagentgrade and purchased from Farmitalia Carlo Erba (Milan, Italy), Sigmaand Fluka Chemie AG (Buchs, Switzerland), Alexis Biochemicals(Lausen, Switzerland).
Acid secretion measurements. Tissues were mounted vertically be-tween twohalves of a Lucite chamberhaving an exposed area of 0.64 cm2.Eachhalf-chamber consistedof a circularfluid canal of 2.5 ml total volumefilledwithmodified Ringer's solution thatwas constantly re-circulated bymeans of a bubble lift. The control Ringer's solution on the serosal sidecontained (inmM): 102.4 Na+, 4.0 K+, 1.4 Ca2+, 0.8 Mg2+, 91.4 Cl−, 17.8HCO3
− and 11 D-glucose. Solutionswere gassed continuallywith 5%CO2 inO2 (pH 7.36).
The luminal solution was unbuffered and had the following compo-sition (in mM): 102.4 Na+, 4.0 K+, 91.4 Cl−, 15 isethionate, 7 mannitol,and 11 D-glucose. To prevent accumulation of CO2, this solution wasgassedwith 100%O2 thatwas passed through abottle containingBa(OH)2solution (50 mM). Acid secretionwasmeasuredwith the pH-statmethod
(Radiometer, Copenhagen, Denmark). The titration procedure wasactivated every 10 min using 5 mMNaOH to titrate the mucosal solutionto a constant pH of 5.50. The transepithelial potential (Vt) was contin-uouslymonitoredwithavoltmeterusing twocalomelhalf-cells connectedto each bath solution via agar-Ringer's bridges. Vt is reported as that of themucosal solution with respect to the serosal side and the short circuitcurrent (Isc) was measured using a voltage clamp device as originallydescribed byUssing andZerahn (1951). The transepithelial resistance (Rt)was calculated using the open circuit potential difference that developed1 s after brief interruption of constant short-circuiting conditions.
Data analysis and statistics. Mean values are expressed ±S.E.M. of nindividual experiment performed. The significance of the observa-tions was evaluated by Student's t test for paired or unpaired data asappropriate and pb0.05 denoted a statistical difference.
Results
Serosal but not luminal cadmium inhibits histamine-stimulated acidsecretion
Our study was performed on amphibian gastric mucosa. Thisepithelium has been used as a model for the study of the function ofthe gastric mucosa in many laboratories, and its secretory and iontransport properties have already been very well characterized(Carlisle et al., 1978; Debellis et al., 1990, 1992, 1998; Forte et al.,1967; Kasbekar and Durbin, 1965; Machen and McLennan, 1980; Ruizet al., 1993; Shoemaker and Sachs, 1972; Silen et al., 1975; Supplissonet al., 1991).
Ingestion of contaminated food or water is one of the main causesof cadmium intoxication. Due to the extremely low intra-luminal pHrecorded in stimulated stomach, it is reasonable to assume that theapical side of the gastric mucosa can be exposed to cadmium ionsderived by degradation of Cd–protein complexes (with metallothio-nein or glutathione) induced by acidic pH. Therefore, we firstevaluated the effect of exposure to luminal cadmium in epitheliastimulated with high histamine concentrations (500 µM). The rate ofacid secretion, measured at 10 min intervals, reached a steady valueusually 60–90 min after stimulation and was constant up to 2 h (datanot shown). When CdCl2 was added to the Ringer's solution perfusingthe luminal side of histamine-stimulated mucosae, the rate of acidsecretion did not change significantly even after prolonged treatment(2 h) at all the concentrations used (Fig. 1A, 20–100 µM and 1 mM,respectively).
Since it is well established that chronic cadmium intoxicationresults in increased blood levels of this heavy metal (Tsalev andZaprianov, 1984; Asar et al., 2000) we next assessed the effect of Cd2+
addition to the serosal side of the epithelium. These experiments,summarized in Fig. 1B, showed that basolateral incubation ofstimulated mucosa with cadmium (20, 50 and 100 µM) resulted in asignificant, irreversible (data not shown), inhibition of acid secretion.While the final inhibitory effect (i.e. the final value of acid secretionrate reached after addition of cadmium) was similar at all theconcentrations used, the effect of 100 µM cadmium was faster. Itstarted earlier and reached ∼70% of maximal inhibition two timesfaster than the other concentrations used (40 min vs 80 min). Asshown in Table 1, the cadmium-induced decrease in acid secretionrate was associated to significant elevations in potential difference(Vt) and transepithelial resistance (Rt). The increase in Rt mightreasonably exclude harmful effects of cadmium on cell–cell adhesions(tight junctions) reported by others (Prozialeck and Niewenhuis,1991; Prozialeck et al., 1993).
The lack of effect of cadmium from the apical side of the epitheliummaybeexplainedbyoneormore of these factors: 1) lowpermeability ofthe apicalmembrane, likelymore tight to cadmium than the basolateral
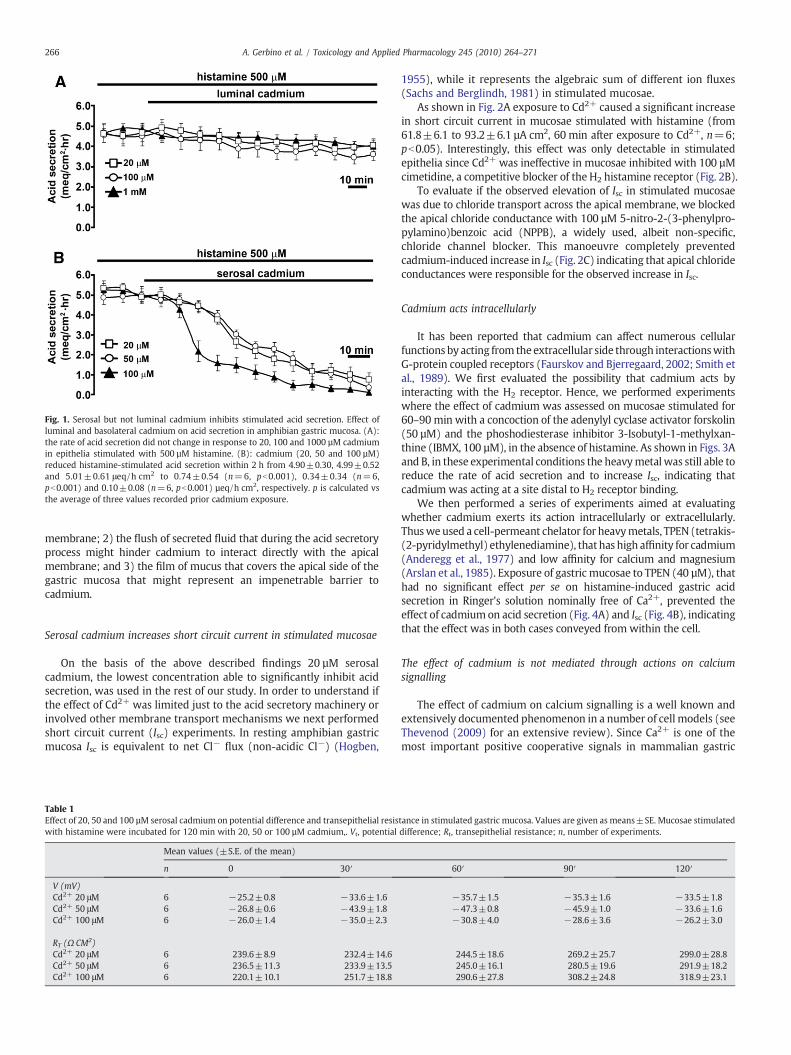
Fig. 1. Serosal but not luminal cadmium inhibits stimulated acid secretion. Effect ofluminal and basolateral cadmium on acid secretion in amphibian gastric mucosa. (A):the rate of acid secretion did not change in response to 20, 100 and 1000 µM cadmiumin epithelia stimulated with 500 µM histamine. (B): cadmium (20, 50 and 100 µM)reduced histamine-stimulated acid secretion within 2 h from 4.90±0.30, 4.99±0.52and 5.01±0.61 μeq/h cm2 to 0.74±0.54 (n=6, pb0.001), 0.34±0.34 (n=6,pb0.001) and 0.10±0.08 (n=6, pb0.001) µeq/h cm2, respectively. p is calculated vsthe average of three values recorded prior cadmium exposure.
266 A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
membrane; 2) the flush of secreted fluid that during the acid secretoryprocess might hinder cadmium to interact directly with the apicalmembrane; and 3) the film of mucus that covers the apical side of thegastric mucosa that might represent an impenetrable barrier tocadmium.
Serosal cadmium increases short circuit current in stimulated mucosae
On the basis of the above described findings 20 µM serosalcadmium, the lowest concentration able to significantly inhibit acidsecretion, was used in the rest of our study. In order to understand ifthe effect of Cd2+ was limited just to the acid secretory machinery orinvolved other membrane transport mechanisms we next performedshort circuit current (Isc) experiments. In resting amphibian gastricmucosa Isc is equivalent to net Cl− flux (non-acidic Cl−) (Hogben,
Table 1Effect of 20, 50 and 100 µM serosal cadmium on potential difference and transepithelial resiswith histamine were incubated for 120 min with 20, 50 or 100 µM cadmium,. Vt, potential
Mean values (±S.E. of the mean)
n 0 30′
V (mV)Cd2+ 20 μM 6 −25.2±0.8 −33.6±1.6Cd2+ 50 μM 6 −26.8±0.6 −43.9±1.8Cd2+ 100 μM 6 −26.0±1.4 −35.0±2.3
RT (Ω CM2)Cd2+ 20 μM 6 239.6±8.9 232.4±14.6Cd2+ 50 μM 6 236.5±11.3 233.9±13.5Cd2+ 100 μM 6 220.1±10.1 251.7±18.8
1955), while it represents the algebraic sum of different ion fluxes(Sachs and Berglindh, 1981) in stimulated mucosae.
As shown in Fig. 2A exposure to Cd2+ caused a significant increasein short circuit current in mucosae stimulated with histamine (from61.8±6.1 to 93.2±6.1 µA cm2, 60 min after exposure to Cd2+, n=6;pb0.05). Interestingly, this effect was only detectable in stimulatedepithelia since Cd2+ was ineffective in mucosae inhibited with 100 µMcimetidine, a competitive blocker of the H2 histamine receptor (Fig. 2B).
To evaluate if the observed elevation of Isc in stimulated mucosaewas due to chloride transport across the apical membrane, we blockedthe apical chloride conductance with 100 µM 5-nitro-2-(3-phenylpro-pylamino)benzoic acid (NPPB), a widely used, albeit non-specific,chloride channel blocker. This manoeuvre completely preventedcadmium-induced increase in Isc (Fig. 2C) indicating that apical chlorideconductances were responsible for the observed increase in Isc.
Cadmium acts intracellularly
It has been reported that cadmium can affect numerous cellularfunctions by acting from the extracellular side through interactionswithG-protein coupled receptors (Faurskov and Bjerregaard, 2002; Smith etal., 1989). We first evaluated the possibility that cadmium acts byinteracting with the H2 receptor. Hence, we performed experimentswhere the effect of cadmium was assessed on mucosae stimulated for60–90 min with a concoction of the adenylyl cyclase activator forskolin(50 µM) and the phoshodiesterase inhibitor 3-Isobutyl-1-methylxan-thine (IBMX, 100 µM), in the absence of histamine. As shown in Figs. 3Aand B, in these experimental conditions the heavymetalwas still able toreduce the rate of acid secretion and to increase Isc, indicating thatcadmium was acting at a site distal to H2 receptor binding.
We then performed a series of experiments aimed at evaluatingwhether cadmium exerts its action intracellularly or extracellularly.Thusweused a cell-permeant chelator for heavymetals, TPEN (tetrakis-(2-pyridylmethyl) ethylenediamine), that has high affinity for cadmium(Anderegg et al., 1977) and low affinity for calcium and magnesium(Arslan et al., 1985). Exposure of gastric mucosae to TPEN (40 µM), thathad no significant effect per se on histamine-induced gastric acidsecretion in Ringer's solution nominally free of Ca2+, prevented theeffect of cadmiumon acid secretion (Fig. 4A) and Isc (Fig. 4B), indicatingthat the effect was in both cases conveyed from within the cell.
The effect of cadmium is not mediated through actions on calciumsignalling
The effect of cadmium on calcium signalling is a well known andextensively documented phenomenon in a number of cell models (seeThevenod (2009) for an extensive review). Since Ca2+ is one of themost important positive cooperative signals in mammalian gastric
tance in stimulated gastric mucosa. Values are given as means±SE. Mucosae stimulateddifference; Rt, transepithelial resistance; n, number of experiments.
60′ 90′ 120′
−35.7±1.5 −35.3±1.6 −33.5±1.8−47.3±0.8 −45.9±1.0 −33.6±1.6−30.8±4.0 −28.6±3.6 −26.2±3.0
244.5±18.6 269.2±25.7 299.0±28.8245.0±16.1 280.5±19.6 291.9±18.2290.6±27.8 308.2±24.8 318.9±23.1

Fig. 2. Effect of serosal cadmium on short circuit current (Isc) in stimulated or inhibitedgastric mucosa. (A): addition of cadmium (20 µM) increased significantly Isc from 61.8±6.1 to 93.2±6.1 µA cm2 (n=6, pb0.05 vs peak at 60 min) in secreting mucosae stim-ulated with histamine. (B): addition of cadmium had no effect on Isc in “resting”mucosae(exposed to the H2 receptor inhibitor cimetidine; 100 µM). (C): effect of cadmium on Isc instimulated gastric mucosa in the continuous presence of 100 µM NPPB. Perfusion withluminal NPPB in stimulated gastric mucosa inhibited completely the cadmium-inducedincrease in Isc.
267A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
acid secretion (Urushidani and Forte, 1997), we sought to determine ifthe reduction in the rate of acid secretion induced by cadmiummightresult from dysfunctional calcium signalling also in our experimentalmodel.
Fig. 3. Cadmium does not affect the function of the histamine receptor. (A): forskolin(50 µM)-and IBMX (100 µM)-induced acid secretion was inhibited by serosal additionof cadmium (from 4.9±0.6 to 0.6±0.5 μeq/h cm2, n=6, pb0.001). (B): addition ofcadmium to epithelia stimulated with forskolin/IBMX increased Isc from 59.3±4.7 to98.7±2.8 µA cm2, (n=6, pb0.01 vs peak at 60 min).
The effect of cadmiumwas first evaluated in the absence of externalCa2+ to exclude any effect of cadmium on Ca2+ entry from theextracellular space. These experiments showed that the cadmium-induced effects on acid secretion rate and Iscwereunaffectedby removalof extracellular Ca2+ (Figs. 5A and B, see for comparison Fig. 1B).
Nextwe analysed the possibility that cadmiummight induce releaseof Ca2+ form the internal stores. Thus, gastric mucosae were exposed tothe Sarco-Endoplasmic Reticulum Ca2+-ATPase (SERCA) pump inhib-itor cyclopiazonic acid (CPA, 40 µM, for 60 min). Blockade of the SERCApump in the absence of extracellular Ca2+ causes a slow calcium leakfrom Ins(1,4,5)P3-sensitive calcium stores and consequently increase incytoplasmic calcium(Demaurex et al., 1992). As shown in Figs. 5C andDCPA, which per se did not elicit any significant effect on gastric acidsecretion, caused a small albeit significant increase in Isc (from69.3±3.9to 80.8±4.4 µA cm2, 60 min after exposure to cadmium, n=6;pb0.05). However the heavy metal was still able to reduce histamine-induced acid secretion rate and to increase Isc even in the continuouspresence of CPA.
These findings suggest that neither intracellular nor extracellularCa2+ mediate the effects induced by cadmium in amphibian gastricmucosa.
Cadmium does not affect the enzymatic activity of adenylyl cyclase andcarbonic anhydrase
It has been reported that cadmium can perturb cAMP levels actingdirectly on the activity of the enzyme that generates cAMP, adenylylcyclase (AC) (Fabbri et al., 2003; Merali and Singhal, 1975; Mgbonyebiet al., 1994). Therefore, we considered the possibility that cadmiummight exert its action by reducing basal cellular levels of cAMP andconsequently acid secretion.We therefore performed experimentswiththe adenylyl cyclase inhibitor 9-(tetrahydro-2-furyl)-9H-purin-6-amine (SQ 22,536 (Harris et al., 1979)). We have reported previouslythat treatment of cimetidine-inhibited gastric mucosaewith 100 µMSQ22,536 results in gland lumenalkalinization (Caroppoet al., 2004). Here,exposure to SQ 22,536 for 2 h caused a slight inhibition of acid secretionrate significantly different from the one elicited by cadmium under thesame experimental conditions (Fig. 6A). Furthermore, exposure to
Fig. 4. Cadmium affects acid secretion and short circuit current acting from theintracellular side. (A): exposure to 40 µM TPEN in the absence of Ca2+prevented theinhibition of acid secretion rate by cadmium in epithelia stimulated with histamine,(B): cadmium had no effect on Isc in stimulated mucosae exposed to TPEN.

Fig. 5. The effect of cadmium on acid secretion rate and short circuit current is not mediated by calcium signalling. When calcium was removed from the bathing solution cadmiumwas still able to (A) completely inhibit the rate of acid secretion (from 4.8±0.3 to 0.4±0.2 µeq/h cm2, n=8, pb0.001) and to (B) increase Isc from 65.9±4.5 to 96.8±6.9 µA cm2,(n=8, pb0.01 vs peak at 40 min). (C) After 60 min treatment with CPA addition of cadmium to stimulated mucosae reduced acid secretion rate (from 3.5±0.2 to 0.5±0.2 μeq/h cm2, n=6, pb0.01) and (D) increased Isc from 80.8±4.4 to 103.1±4.8 µA cm2, (n=6, pb0.05 vs peak at 60 min).
268 A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
cadmium in the continuous presence of SQ 22,536 promptly inhibitedthe rate of acid secretion (Fig. 6B) indicating that the heavy metal wasacting via a different mechanism. Finally, although perfusion with SQ22,536 inmucosae stimulatedwith histamine caused a small increase inshort circuit current, subsequent exposure to cadmiumwas still able tocause a rapid increase in Isc (Fig. 6C) not significantly different from theone obtained in control conditions.
We subsequently evaluated the effect of cadmium intoxication oncarbonic anhydrase (CA) that, by catalyzing the rapid conversion ofcarbon dioxide to bicarbonate and protons, sustains acid secretion byproviding H+ for the proton pump (Hersey and High, 1971). Wetherefore used the CA inhibitor acetazolamide (100 µM). Although wecould not evaluate the effect of cadmium on acetazolamide-treatedepithelia because, as expected, the enzyme reached ∼80% of maximalinhibition four times faster than20 µMcadmium(30 minvs 120 min, seeFigs. 7A and 1B for comparison), the fact that acetazolamide inhibited,rather than increased Isc (Fig. 7B) was a strong indication that cadmiumand acetazolamide act on different targets.
Fig. 6. Cadmium does not affect adenylyl cyclase activity. (A): addition of 100 µM SQ22,536 elicited a small albeit significant reduction of acid secretion rate in tissuestimulated with histamine (from 6.6±0.3 to 5.4±0.4 μeq/h cm2, n=4, pb0.05).Under these experimental conditions, cadmium was still able to (B) completely reducethe rate of acid secretion (from 5.3±0.4 to 0.6±0.4 µeq/h cm2, n=4, pb0.001) and to(C) increase Isc from 58.8±3.2 to 109.4±2.0 μA cm2, (n=4, pb0.01 vs peak at 50 min).
Possible involvement of the proton pump
The cellular mechanism that could most likely explain the resultsobtained so far is an interaction of cadmium with the gastric protonpump, as previously proposed by others (Hongo et al., 1990). It hasbeen shown that in the presence of the well known H+/K+-ATPaseblocker, omeprazole, chloride secretion continues and that thisuncoupled chloride secretion becomes electrogenic (McGreevy etal., 1986; Starlinger et al., 1986; Reenstra et al., 1987). Also in ourexperimental model, exposure of epithelia stimulated with histamineto serosal omeprazole increased Isc (Fig. 8A). This effect wasquantitatively comparable to the one elicited by cadmium in thesame experimental conditions and pre-treatment with omeprazoleprevented cadmium from exerting its effect on short circuit current(Fig. 8B). Conversely, same results were obtained when omeprazolewas added after cadmium, indicating that the two compounds wereacting with the same mechanism (Fig. 8C). Thus, the changes in Iscobserved during omeprazole and cadmium treatments most probably
represent the uncoupled component of chloride secretion unmaskedby proton pump inhibition.
Discussion
The present study shows that acute, basolateral exposure of gastricoxyntopeptic cells to cadmium largely and significantly inhibits acidsecretion. Although this effect could be the result of actions at differentfunctional levels in the acid secreting cells, our findings indicate aspecific action of the heavy metal on the gastric proton pump.

Fig. 7. Cadmium does not affect carbonic anhydrase activity. (A): 100 µM acetazolamideelicited a significant reduction of the rate of the acid secretion (from 4.8±0.5 to 1.3±0.5 μeq/h cm2, n=6, pb0.001) in epithelia stimulated with histamine addition.(B): under these experimental conditions cadmium reduced Isc from 57.5±6.9 to44.1±6.0 µA cm2, (n=6, pb0.01 vs peak at 30 min).
269A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
We could in fact exclude an effect of the heavy metal on thehistamine H2 receptor since experiments with forskolin plus IBMX(Fig. 3), strongly suggests that cadmium is acting at site distal to H2
receptor binding/activation of adenylyl cyclase. Also, cadmium does
Fig. 8. Cadmium affects acid secretion: possible involvement of the H+/K+ATPase.(A): addition of 50 µM omeprazole increased Isc (from 58.8±3.2 to 109.4±2.0 µA cm2,n=4, pb0.01 vs peak at 30 min) in tissues stimulated with histamine. (B): addition ofcadmium after 60 min treatment with omeprazole had no effect on Isc. (C): exposure toomeprazole after 60 min treatment with cadmium, failed to induce any further increaseof the Isc.
not appear to perturb the activity of two metal-containing enzymes,like adenylyl cyclase (AC) and carbonic anhydrase (CA), both playinga key role in gastric acid secretion.
Although cadmium-induced reduction of AC activity has beenreported previously (Fabbri et al., 2003; Merali and Singhal, 1975;Mgbonyebi et al., 1994), we found that the action of cadmium is notaffected by inhibition of AC. While cadmium completely blocked acidsecretion, inhibition of AC resulted only in a modest decline of acidsecretion rate (Fig. 6A). This is not surprising since the ability of thisdrug to reduce cAMP is contrasted by the continuous production ofcAMP induced by histamine.
Furthermore, experiments with the CA inhibitor acetazolamideseem to exclude an effect of cadmium on this enzyme since theinhibitory action of acetazolamide on acid secretion rate is much fasterthan the one of cadmium and its effect on Isc is opposite to the one ofcadmium (Fig. 7B). Finally, we believe that we can rule out an action ofcadmium on basolateral ion transporters like the Na+/H+ exchanger,the Na+/K+-ATPase, or the Na+-K+-2Cl− since inhibition of thesetransporters has little effect on stimulated acid secretion in frog gastricmucosa (Horie et al., 1992; Hagen et al., 2000; Reenstra et al., 1987).
Our findings can be explained by a specific inhibitory action ofcadmium on the H+/K+-ATPase that is accompanied by a significantincrease in chloride-dependent short circuit current. While reduction ofthe acid secretion rate is themost predictable result following inhibitionof the gastric proton pump, the effect of cadmiumon Isc is less obvious. Itis known that in resting conditions, amphibian stomach generates acurrent that is equivalent to chloride ion secretion (non-acidic chloride),as measured by short circuit current (Hogben, 1955). Under stimulatedconditions, Isc becomes the algebraic sum of different ionic transportersmainly represented by non-acidic chloride-current, the current carriedbyCl− ions exiting the cells via channels openedon the apicalmembraneupon histamine stimulation (acidic chloride), and protons pumped bythe H+/K+ ATPase. Na+ is excluded because in amphibian gastricmucosa the Na+ net flux is negligible (Hogben, 1955; Sachs andBerglindh, 1981).
We suggest that the cadmium-induced increase in Isc, which isobserved only in stimulated conditions, represents the rheogenicchloride secretion (acidic chloride) that is normally coupled to H+
transport. This process is unmasked by the cadmium-induced inhibitionof the proton pump. This conclusion is strengthened by the fact that theinhibitor of apical chloride conductance, NPPB, completely blocks theincrease in Isc (Fig. 2).
Ourhypothesis of a specific inhibitory action of cadmiumonH+/K+-ATPase is supported by the observation that cadmium (IC50 100 µM)inhibits the gastric proton pump activity in H+/K+ ATPase-enrichedmembrane fractions derived from hog gastric mucosa (Hongo et al.,1990). Furthermore, several studies report that also other ATPases, suchas the Ca2+-ATPase (Hechtenberg and Beyersmann, 1991, 1994;Verbost et al., 1988; Zhang et al., 1990), the Na+/K+-ATPase (Kinne-Saffran et al., 1993), and the vacuolar H+-ATPase (Herak-Krambergeret al., 2000) are blocked by cadmium. Importantly it has been suggestedthat Cd2+ exerts its inhibitory action on Ca2+-ATPase by binding to thiolgroups (Zhang et al., 1990) which are crucial for the enzymatic reaction(MacLennan, 1970). The ability of cadmium to bind –SH groups wouldthus explain also its effect on the gastric proton pump that containsfunctionally important thiol groups which are the target of omeprazoleaction (Dupuy and Szabo, 1986; Shin et al., 2009).
Theomeprazole-like action exerted by cadmium is also supportedbystudies on the morphology of oxyntopeptic cells. It is well known thatstimuli eliciting acid secretion induce an enormous expansion of thesecretory canaliculi of oxyntopeptic cells, necessary to increase thesecretory surface area. When acid secretion stops, the morphology ofacid secreting cells returns to the resting type (Helander and Sundell,1984; Helander, 1984). Interestingly, despite the complete inhibition ofacid secretion obtained by treatment with H+/K+-ATPase inhibitors, apersistent dilation of the secretory canaliculi has been demonstrated

270 A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
both in rat and rabbit gastric mucosae (Fryklund et al., 1988; Helander,1984). A similar persistent dilation of the secretory canaliculi wasobserved in oxyntopeptic cells of cadmium-treated rats (Asar et al.,2000).
The similarity of action shared by cadmium and omeprazole couldalso explain the lack of reversal in the effect of cadmium on gastricacid secretion rate. Omeprazole is an acid-activated pro-drug thatreacts from the luminal face of the H+/K+-ATPase with two cysteinesper mole of phosphoenzyme. The reversal of action with this type ofanti-secretory agent depends on the turnover of the (H+/K+)-ATPase,which has a half-life of 30–48 h, thus much longer than the averagelength of our experimental protocols. Likewise, cadmium mightinterfere with the normal turnover of the gastric proton pump byblocking the recovery of the acid secretion.
In conclusion, we showed here for the first time that acute exposureto serosal cadmium blocks acid secretion likely by inhibiting the gastricproton pump. This effectmay be the result of a direct action of the heavymetal on the H+/K+-ATPase or it can be secondary to the inhibition ofapical transporters functionally coupled with the H+/K+ ATPase, suchas the apical K+ conductance. In fact, we and others have recentlyreported that inhibition of this conductance leads to blockade ofhistamine-stimulated acid secretion (Hagen et al., 2000; Gerbino et al.,2007).
Regarding the clinical aspect of our findings, one could speculatethat the reduction in basal (Asar et al., 2000) and stimulated HClsecretion observed under chronic/acute cadmium exposure mightpotentially lead to a decrease in the antibacterial properties of thestomach that are generally promoted by very low pH. Such conditionsmight thus increase the risk of infections of the digestive tract,especially those mediated by Vibrio or Helicobacter bacteria.
Finally, in light of the fact that amphibians (frogs and toads) areused as bioindicators of environmental conditions and pollution(Dumpert and Zietz, 1984; Lefcort et al., 1998; Sparling, 2000;Venturino et al., 2003) our study possibly provides further insight intothe sensitivity of frogs to heavy metals.
Acknowledgments
This work was supported by Finanziamenti di Ateneo (Bari, Italy)and Fondo per gli Investimenti della Ricerca di Base (FIRB) grantRBIN04PHZ7 (Rome, Italy). We are grateful to Dr. Aldebaran M. Hofer(Harvard University, MA, USA) for critical reading of the manuscript.We wish to thank Claudia Carmone, Claudia Lippolis, Rosa Piazzollaand Michela Sguera for excellent assistance during experiments.
Conflict of interest statement
The authors declare no financial or personal conflict of interest.
References
Anderegg, G., Hubmann, E., Podder, N.G., Wenk, F., 1977. Pyridine derivatives ascomplexing agents XI. Thermodynamics of metal complex formation with bis-, tris-and tetrakis[(2-pyridyl)methyl]-amines. Helv. Chim. Acta 60, 123–140.
Arslan, P., Di Virgilio, F., Beltrame, M., Tsien, R.Y., Pozzan, T., 1985. Cytosolic Ca2+homeostasis in Ehrlich and Yoshida carcinomas. A new, membrane-permeantchelator of heavy metals reveals that these ascites tumor cell lines have normalcytosolic free Ca2+. J. Biol. Chem. 260, 2719–2727.
Asar, M., Kayisli, U.A., Izgut-Uysal, V.N., Oner, G., Polat, S., Kaya, M., 2000. Cadmium-induced changes in parietal cell structure and functions of rats. Biol. Trace Elem.Res. 74, 153–170.
Carlisle, K.S., Chew, C.S., Hersey, S.J., 1978. Ultrastructural changes and cyclic AMP infrog oxyntic cells. J. Cell Biol. 76, 31–42.
Caroppo, R., Gerbino, A., Debellis, L., Kifor, O., Soybel, D.I., Brown, E.M., Hofer, A.M.,Curci, S., 2001. Asymmetrical, agonist-induced fluctuations in local extracellular [Ca(2+)] in intact polarized epithelia. EMBO J. 20, 6316–6326.
Caroppo, R., Gerbino, A., Fistetto, G., Colella, M., Debellis, L., Hofer, A.M., Curci, S., 2004.Extracellular calcium acts as a “third messenger” to regulate enzyme and alkalinesecretion. J. Cell Biol. 166, 111–119.
Coddou, C., Lorca, R.A., Acuna-Castillo, C., Grauso, M., Rassendren, F., Huidobro-Toro, J.P.,
2005. Heavy metals modulate the activity of the purinergic P2X4 receptor. Toxicol.Appl. Pharmacol. 202, 121–131.
Debellis, L., Curci, S., Fromter, E., 1990. Effect of histamine on the basolateral K+conductance of frog stomach oxyntic cells and surface epithelial cells. Am. J.Physiol. 258, G631–636.
Debellis, L., Curci, S., Fromter, E., 1992. Microelectrode determination of oxyntic cell pHin intact frog gastric mucosa. Effect of histamine. Pflugers Arch. 422, 253–259.
Debellis, L., Caroppo, R., Fromter, E., Curci, S., 1998. Alkaline secretion by frog gastric glandsmeasuredwith pHmicroelectrodes in the gland lumen. J. Physiol. 513 (Pt 1), 235–241.
Demaurex, N., Lew, D.P., Krause, K.H., 1992. Cyclopiazonic acid depletes intracellularCa2+ stores and activates an influx pathway for divalent cations in HL-60 cells. J.Biol. Chem. 267, 2318–2324.
Dumpert, K., Zietz, E., 1984. Platanna(Xenopus laevis) as a test organism fordetermining theembryotoxic effects of environmental chemicals. Ecotoxicol. Environ. Saf. 8, 55–74.
Dupuy, D., Szabo, S., 1986. Protection by metals against ethanol-induced gastricmucosal injury in the rat. Comparative biochemical and pharmacologic studiesimplicate protein sulfhydryls. Gastroenterology 91, 966–974.
Fabbri, E., Caselli, F., Piano, A., Sartor, G., Capuzzo, A., 2003. Cd2+ and Hg2+ affectglucose release and cAMP-dependent transduction pathway in isolated eelhepatocytes. Aquat. Toxicol. 62, 55–65.
Faurskov, B., Bjerregaard, H.F., 2002. Evidence for cadmium mobilization of intracellularcalcium through a divalent cation receptor in renal distal epithelial A6 cells. PflugersArch. 445, 40–50.
Forte, J.G., Forte, G.M., Saltman, P., 1967. K+-stimulated phosphatase of microsomesfrom gastric mucosa. J. Cell. Physiol. 69, 293–304.
Fryklund, J., Helander, H.F., Elander, B., Wallmark, B., 1988. Function and structure ofparietal cells after H+-K+-ATPase blockade. Am. J. Physiol. 254, G399–407.
Gerbino, A., Fistetto, G., Colella, M., Hofer, A.M., Debellis, L., Caroppo, R., Curci, S., 2007.Real timemeasurements of water flow in amphibian gastric glands: modulation viathe extracellular Ca2+-sensing receptor. J. Biol. Chem. 282, 13477–13486.
Giridhar, J., Rathinavelu, A., Isom, G.E., 1992. Interaction of cadmium with atrialnatriuretic peptide receptors: implications for toxicity. Toxicology 75, 133–143.
Hagen, S.J., Wu, H., Morrison, S.W., 2000. NH(4)Cl inhibition of acid secretion: possibleinvolvement of an apical K(+) channel in bullfrog oxyntic cells. Am. J. Physiol.Gastrointest. Liver Physiol. 279, G400–410.
Harris, D.N., Asaad, M.M., Phillips, M.B., Goldenberg, H.J., Antonaccio, M.J., 1979.Inhibition of adenylate cyclase in human blood platelets by 9-substituted adeninederivatives. J. Cyclic Nucleotide Res. 5, 125–134.
Hechtenberg, S., Beyersmann, D., 1991. Inhibition of sarcoplasmic reticulum Ca(2+)-ATPase activity by cadmium, lead and mercury. Enzyme 45, 109–115.
Hechtenberg, S., Beyersmann, D., 1994. Interference of cadmium with ATP-stimulatednuclear calcium uptake. Environ. Health Perspect. 102 (Suppl 3), 265–267.
Helander, H.F., 1984. Parietal cell structure during inhibition of acid secretion. Scand. J.Gastroenterol. Suppl. 101, 21–26.
Helander, H.F., Sundell, G.W., 1984. Ultrastructure of inhibited parietal cells in the rat.Gastroenterology 87, 1064–1071.
Herak-Kramberger, C.M., Sabolic, I., Blanusa, M., Smith, P.J., Brown, D., Breton, S., 2000.Cadmium inhibits vacuolar H(+)ATPase-mediated acidification in the rat epidid-ymis. Biol. Reprod. 63, 599–606.
Hersey, S.J., High, W.L., 1971. On the mechanism of acid secretory inhibition byacetazolamide. Biochim. Biophys. Acta 233, 604–609.
Hogben, C.A., 1955. Active transport of chloride by isolated frog gastric epithelium;origin of the gastric mucosal potential. Am. J. Physiol. 180, 641–649.
Hongo, T., Nojima, S., Setaka, M., 1990. Purification and characterization of (H++K+)-ATPase from hog gastric mucosa. Jpn J. Pharmacol. 52, 295–305.
Horie, S., Yano, S., Watanabe, K., 1992. Effects of drugs acting on Cl(−)-HCO3- and Na(+)-H+ exchangers on acid secretion in the rat gastric mucosa sheet preparation.Eur. J. Pharmacol. 229, 15–19.
Jarup, L., Akesson, A., 2009. Current status of cadmium as an environmental healthproblem. Toxicol. Appl. Pharmacol. 238, 201–208.
Kasbekar, D.K., Durbin, R.P., 1965. An adenosine triphosphatase from frog gastricmucosa. Biochim. Biophys. Acta 105, 472–482.
Kinne-Saffran, E., Hulseweh, M., Pfaff, C., Kinne, R.K., 1993. Inhibition of Na, K-ATPase bycadmium: different mechanisms in different species. Toxicol. Appl. Pharmacol. 121,22–29.
Lefcort, H., Meguire, R.A., Wilson, L.H., Ettinger, W.F., 1998. Heavy metals alter thesurvival, growth, metamorphosis, and antipredatory behavior of Columbia spottedfrog (Rana luteiventris) tadpoles. Arch. Environ. Contam. Toxicol. 35, 447–456.
Machen, T.E., McLennan, W.L., 1980. Na+-dependent H+ and Cl− transport in in vitrofrog gastric mucosa. Am. J. Physiol. 238, G403–413.
MacLennan, D.H., 1970. Purification and properties of an adenosine triphosphatasefrom sarcoplasmic reticulum. J. Biol. Chem. 245, 4508–4518.
McGreevy, J.M., Barton, R.G., Housinger, T., 1986. Chloride transport in bullfrog gastricmucosa. J. Surg. Res. 40, 462–466.
Merali, Z., Singhal, R.L., 1975. Influence of chronic exposure to cadmium on hepatic andrenal cyclic AMP–protein kinase system. Toxicology 4, 207–214.
Mgbonyebi, O.P., Smothers, C.T., Mrotek, J.J., 1994. Modulation of adrenal cell functionsby cadmium salts: 3. Sites affected by CdCl2 during stimulated steroid synthesis.Cell Biol. Toxicol. 10, 35–43.
Oner, G., Izgut-Uysal, V.N., Senturk, U.K., 1994. Role of lipid peroxidation in cadmium-induced impairment of the gastric mucosal barrier. Food Chem. Toxicol. 32, 799–804.
Prozialeck,W.C., Niewenhuis, R.J., 1991. Cadmium (Cd2+) disrupts intercellular junctionsand actin filaments in LLC-PK1 cells. Toxicol. Appl. Pharmacol. 107, 81–97.
Prozialeck, W.C., Wellington, D.R., Lamar, P.C., 1993. Comparison of the cytotoxic effectsof cadmium chloride and cadmium-metallothionein in LLC-PK1 cells. Life Sci. 53,PL337–342.

271A. Gerbino et al. / Toxicology and Applied Pharmacology 245 (2010) 264–271
Reenstra, W.W., Bettencourt, J.D., Forte, J.G., 1987. Mechanisms of active Cl− secretionby frog gastric mucosa. Am. J. Physiol. 252, G543–547.
Ruiz, M.C., Acosta, A., Abad, M.J., Michelangeli, F., 1993. Nonparallel secretion ofpepsinogen and acid by gastric oxyntopeptic cells of the toad (Bufomarinus). Am. J.Physiol. 265, G934–941.
Sachs, G., Berglindh, T., 1981. Physiology of the parietal cell. In: Johnson, L.R.(Ed.), Physiology of the Gastrointestinal Tract. Raven Press, New York, pp.567–602.
Shin, J.M., Munson, K., Vagin, O., Sachs, G., 2009. The gastric HK-ATPase: structure,function, and inhibition. Pflugers Arch. 457, 609–622.
Shoemaker, R.L., Sachs, G., 1972. Microelectrode studies of Necturus gastric mucosa.Gastric Secretion. Academic Press Inc., New York/London, pp. 147–163.
Silen, W., Machen, T.E., Forte, J.G., 1975. Acid–base balance in amphibian gastricmucosa. Am. J. Physiol. 229, 721–730.
Smith, J.B., Dwyer, S.D., Smith, L., 1989. Cadmium evokes inositol polyphosphateformation and calcium mobilization. Evidence for a cell surface receptor thatcadmium stimulates and zinc antagonizes. J. Biol. Chem. 264, 7115–7118.
Sparling, D.W., 2000. Ecotoxicology of organic contaminants to amphibians. In:Sparling, D.W., Linder, G., Bishop, C.A. (Eds.), Ecotoxicology of Amphibians andReptiles. SETAC Press, Pensacola, pp. 461–494.
Starlinger, M.J., Hollands, M.J., Rowe, P.H., Matthews, J.B., Silen, W., 1986. Chloridetransport of frog gastric fundus: effects of omeprazole. Am. J. Physiol. 250,G118–126.
Stoica, A., Katzenellenbogen, B.S., Martin, M.B., 2000. Activation of estrogen receptor-alpha by the heavy metal cadmium. Mol. Endocrinol. 14, 545–553.
Supplisson, S., Loo, D.D., Sachs, G., 1991. Diversity of K+ channels in the basolateralmembrane of resting Necturus oxyntic cells. J. Membr. Biol. 123, 209–221.
Thevenod, F., 2009. Cadmium and cellular signaling cascades: to be or not to be?Toxicol. Appl. Pharmacol. 238, 221–239.
Tsalev, D.L., Zaprianov, Z.K., 1984. Atomic absorption spectrometry in occupational andenvironmental health practice. CRC Press, Boca Raton, FL.
Urushidani, T., Forte, J.G., 1997. Signal transduction and activation of acid secretion inthe parietal cell. J. Membr. Biol. 159, 99–111.
Ussing, H.H., Zerahn, K., 1951. Active transport of sodium as the source of electriccurrent in the short-circuited isolated frog skin. Reprinted from Acta. Physiol.Scand. 23: 110–127, 1951. J. Am. Soc. Nephrol. 10, 2056–2065.
Venturino, A., Rosenbaum, E., Caballero de Castro, A., Anguiano, O.L., Gauna, L., Fonovichde Schroeder, T., Pechen de D'Angelo, A.M., 2003. Biomarkers of effect in toads andfrogs. Biomarkers 8, 167–186.
Verbost, P.M., Flik, G., Lock, R.A., Wendelaar Bonga, S.E., 1988. Cadmium inhibits plasmamembrane calcium transport. J. Membr. Biol. 102, 97–104.
Yao, X., Forte, J.G., 2003. Cell biology of acid secretion by the parietal cell. Annu. Rev.Physiol. 65, 103–131.
Zhang, G.H., Yamaguchi, M., Kimura, S., Higham, S., Kraus-Friedmann, N., 1990. Effectsof heavy metal on rat liver microsomal Ca2(+)-ATPase and Ca2+ sequestering.Relation to SH groups. J. Biol. Chem. 265, 2184–2189.


















