
Bothrops jararaca and Crotalus durissus terrificus venoms elicit distinct responses regarding to...
10
Toxicon 49 (2007) 615–624 Bothrops jararaca and Crotalus durissus terrificus venoms elicit distinct responses regarding to production of prostaglandins E 2 and D 2 , and expression of cyclooxygenases Vanessa Moreira a,1 , Stella Regina Zamuner a,b,1 , John L. Wallace b , Catarina de Fa´tima Pereira Teixeira a, a Laboratory of Pharmacology, Butantan Institute, Ave Vital Brazil, 1500, 05503-900 Sao Paulo, Brazil b Mucosal Inflammation Research Group, University of Calgary, Calgary, Alberta, Canada T2N 4N1 Received 27 May 2006; received in revised form 4 September 2006; accepted 11 September 2006 Available online 16 September 2006 Abstract Prostaglandins (PGs), synthesized by cyclooxygenases, play important roles in many pathophysiological processes including inflammation and hyperalgesia. In this study the profiles of PGE 2 and PGD 2 production secondary to injection of Bothrops jararaca venom (BjV), with inflammatory activity or Crotalus durissus terrificus venom (CdtV), with anti- inflammatory and antinociceptive properties, into mice were evaluated, and the ability of these venoms to induce expression of cyclooxygenases-1 (COX-1) and -2 (COX-2) was investigated. Intraperitoneal injection of BjV but not of CdtV induced the release and PGD 2 at 30 min and of PGE 2 from 3 up to 12 h after injection. Moreover, BjV up-regulated expression of COX-2 but not of the constitutive COX-1, suggesting that expressed COX-2 provides more substrate for synthesis of PGs by the respective terminal synthases, being the critical enzyme for PGs production in the late periods of BjV effect. In contrast, CdtV does not have any effect on constitutive COX-1 and do not induce expression of COX-2. Therefore, differences between BjV and CdtV in the ability to regulate PGs synthesis can account for their distinct effects with regard to inflammation. Moreover, inhibition of COX-2 by selective drugs may be of value to counteract the severe local inflammation induced by BjV in the victims. r 2006 Elsevier Ltd. All rights reserved. Keywords: Snake venom; Prostaglandins; Cyclooxygenases 1. Introduction Bothrops jararaca and Crotalus durissus terrificus snakes are responsible for the majority of snakebites in Brazil (Rosenfeld, 1971). A complex and promi- nent inflammatory reaction is developed at the site of B. jararaca venom injection in humans and experimental animals (Rosenfeld, 1971; Trebien and Calixto, 1989, Cardoso et al., 1993). This reaction comprises oedema, hyperalgesia and infiltration of leukocytes into tissues (Trebien and Calixto, 1989; Teixeira et al., 1994; Farsky et al., 1997; Zamuner et al., 2001). On the contrary, C. durissus terrificus ARTICLE IN PRESS www.elsevier.com/locate/toxicon 0041-0101/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2006.09.006 Corresponding author. Tel.: +55 11 37267222; fax: +55 11 37261505. E-mail address: [email protected] (C.F.P. Teixeira). 1 These authors contributed equally to this work.
-
Upload
vanessa-moreira -
Category
Documents
-
view
212 -
download
0
Transcript of Bothrops jararaca and Crotalus durissus terrificus venoms elicit distinct responses regarding to...

ARTICLE IN PRESS
0041-0101/$ - see
doi:10.1016/j.tox
�Correspondifax: +5511 372
E-mail addre1These author
Toxicon 49 (2007) 615–624
www.elsevier.com/locate/toxicon
Bothrops jararaca and Crotalus durissus terrificus venoms elicitdistinct responses regarding to production of prostaglandins
E2 and D2, and expression of cyclooxygenases
Vanessa Moreiraa,1, Stella Regina Zamunera,b,1, John L. Wallaceb,Catarina de Fatima Pereira Teixeiraa,�
aLaboratory of Pharmacology, Butantan Institute, Ave Vital Brazil, 1500, 05503-900 Sao Paulo, BrazilbMucosal Inflammation Research Group, University of Calgary, Calgary, Alberta, Canada T2N 4N1
Received 27 May 2006; received in revised form 4 September 2006; accepted 11 September 2006
Available online 16 September 2006
Abstract
Prostaglandins (PGs), synthesized by cyclooxygenases, play important roles in many pathophysiological processes
including inflammation and hyperalgesia. In this study the profiles of PGE2 and PGD2 production secondary to injection
of Bothrops jararaca venom (BjV), with inflammatory activity or Crotalus durissus terrificus venom (CdtV), with anti-
inflammatory and antinociceptive properties, into mice were evaluated, and the ability of these venoms to induce
expression of cyclooxygenases-1 (COX-1) and -2 (COX-2) was investigated. Intraperitoneal injection of BjV but not of
CdtV induced the release and PGD2 at 30min and of PGE2 from 3 up to 12 h after injection. Moreover, BjV up-regulated
expression of COX-2 but not of the constitutive COX-1, suggesting that expressed COX-2 provides more substrate for
synthesis of PGs by the respective terminal synthases, being the critical enzyme for PGs production in the late periods of
BjV effect. In contrast, CdtV does not have any effect on constitutive COX-1 and do not induce expression of COX-2.
Therefore, differences between BjV and CdtV in the ability to regulate PGs synthesis can account for their distinct effects
with regard to inflammation. Moreover, inhibition of COX-2 by selective drugs may be of value to counteract the severe
local inflammation induced by BjV in the victims.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Snake venom; Prostaglandins; Cyclooxygenases
1. Introduction
Bothrops jararaca and Crotalus durissus terrificus
snakes are responsible for the majority of snakebites
front matter r 2006 Elsevier Ltd. All rights reserved
icon.2006.09.006
ng author. Tel.: +5511 37267222;
61505.
ss: [email protected] (C.F.P. Teixeira).
s contributed equally to this work.
in Brazil (Rosenfeld, 1971). A complex and promi-nent inflammatory reaction is developed at the siteof B. jararaca venom injection in humans andexperimental animals (Rosenfeld, 1971; Trebien andCalixto, 1989, Cardoso et al., 1993). This reactioncomprises oedema, hyperalgesia and infiltration ofleukocytes into tissues (Trebien and Calixto, 1989;Teixeira et al., 1994; Farsky et al., 1997; Zamuneret al., 2001). On the contrary, C. durissus terrificus
.

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624616
venom does not induce a significant inflammatoryreaction at the site of bite (Brazil, 1934; Amorimand Mello, 1954; Cardoso and Mota, 1997; Land-ucci et al., 1995; Sousa e Silva et al., 1996; Giorgiet al., 1993). This venom exerts severe systemicneurotoxic, nephrotoxic, hepathotoxic and myo-toxic effects (Cupo et al., 1988; Barraviera et al.,1990; 1995), and causes antinociception in experi-mental models (Brigatte et al., 2001; Picolo et al.,1998).
Inflammatory events induced by B. jararaca
venom have been associated to the release of anumber of inflammatory mediators which includeeicosanoids such as prostaglandins (PGs), throm-boxane and leukotrienes (Zamuner and Teixeira,2002; Farsky et al., 1997; Flores et al., 1993). Therole of PGs on oedematogenic response causedby Bothrops sp. snake venom was described forB. asper, B. insularis and B. jararaca venom(Trebien and Calixto, 1989; Chaves et al., 1995;Barbosa et al., 2003). In contrast, production ofPGs under in vivo activity of C. durissus terrificus isstill unclear although the in vitro release of PGE2
from isolated macrophages stimulated by thisvenom has been demonstrated recently (Sampaioet al., 2006).
Prostaglandin E2 (PGE2) is a major member ofPGs family, and plays important regulatory roles inphysiological systems such as gastrointestinal, renaland cardiovascular (Harris et al., 2002; Narumiyaand Fitzgerald, 2001; Kennedy et al., 1999; Warneret al., 1999). PGE2 is also generated in a number ofsettings of inflammation, modulating its evolutionand exerting potent immunomodulatory effects(Nishijima et al., 1985; Portanova et al., 1996;Trebino et al., 2003, Molloy and McCarthy, 2005).In certain instances, PGE2 has been shown to havemultiple apparently opposing functional effects(Hata and Breyer, 2004). In inflammatory processesPGE2 is a potent vasodilator, which markedlyenhances oedema formation (Portanova et al.,1996), and potentiates the pain-inducing activity ofbradykinin (Nakamura and Ferreira, 1987; Cunhaet al., 1991, 1992; Poole et al., 1999). However, thisPG can also suppress the production of theinflammatory cytokines TNF-a and IL-1b (Knund-sen et al., 1986; Kunkel et al., 1986; Phipps et al.,1991; Nataraj et al., 2001; Takayama et al., 2002),phagocytosis by macrophages (Oropeza-Rendonet al., 1980; Kozlov et al., 1990; Davidson et al.,1998), and inhibits T cell proliferation (Natarajet al., 2001).
Prostaglandin D2 (PGD2), another member ofPGs family, mediates various physiological pro-cesses in the central nervous and vascular systems(Giles and Leff, 1988; Ito et al., 1989; Mizoguchiet al., 2001). In addition, PGD2 is released duringacute allergic reactions (Murray et al., 1986; Barret al., 1988) and participates in inflammatoryprocesses promoting vasodilatation and increase ofvascular permeability (Kanaoka and Urade, 2003;Hart, 2001).
The biosynthesis of PGs is initiated by theendoperoxide-synthase or cyclooxygenase (COX)which possesses both fatty acid COX activityconverting free arachidonic acid to the unstableintermediate PGG2, and peroxidase activity con-verting PGG2 to PGH2. This is then converted tothe bioactive end products PGE2, PGD2, PGF2a,PGI2 and thromboxane A2 by the respectiveterminal synthases which exhibit cell- and tissue-specific distributions (Smith et al., 2000; Rocca andFitzgerald, 2002). It is recognized that the COXexists in two isoforms commonly referred to ascyclooxygenase-1 (COX-1) and cyclooxygenase-2(COX-2) which share approximately 60% sequenceidentity (Smith et al., 2000; 1996). COX-1 isexpressed constitutively in most tissues whileCOX-2 is a largely inducible enzyme that isprominent at sites of inflammation although beingstably expressed in the brain, reproductive tissues,kidney and thymus (Smith and Langenbach, 2001).
Given the relevance of the regulatory PGE2 andPGD2 in many pathophysiological processes, andthe differences between B. jararaca and C. durissus
terrificus venoms to cause local inflammation, thepresent study was designed to comparatively eval-uate the profiles of PGE2 and PGD2 productionsecondary to injection of B. jararaca or C. durissus
terrificus venoms into mice, and the ability of thesevenoms to induce the expression of COXs.
2. Materials and methods
2.1. Venom
Lyophilized crude venom of B. jararaca (BjV) andC. durissus terrificus (CdtV) were supplied byHerpetology Laboratory from Butantan Institute,Sao Paulo—Brazil. Venoms were dissolved in0.15M NaCl solution and subsequently filteredthrough sterilizing membranes (0.22 mm pore size;Millipore Ind. Com. Ltd., Brazil) before use.

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624 617
2.2. Animals
Male Swiss mice (19–20 g) were used. Theseanimals were housed in temperature-controlledrooms and received water and food ad libitum untiluse. These studies were approved by the Experi-mental Animal Committee of Butantan Institute(protocol no. 177/2004) in accordance with theprocedures laid down by the Universities Federa-tion for Animal Welfare.
2.3. Quantification of eicosanoid concentrations
Concentrations of PGE2 and PGD2 were mea-sured in peritoneal washes at 30min, 1, 3, 6 and 12 hafter intraperitoneal (i.p.) injections of non lethaldoses of BjV (250 mg/kg) or CdtV (25 mg/kg) orsterile saline, by a specific enzymatic immunoassaypreviously described by Pradelles et al., (1985) usinga commercial kit (Cayman Chemicals, MI, USA),after extraction of eicosanoids on Sep Pak C18columns eluted with ethanol. In brief, 50 mL aliquotsof each extracted sample were incubated with theeicosanoid conjugated with acetylcholinesterase andthe specific rabbit antiserum, in 96-well microtitra-tion plates coated with anti-rabbit IgG mousemonoclonal antibody. After addition of the sub-strate, the absorbances of the samples were recordedat 412 nm in a microplate reader, and the concen-tration of the eicosanoid was estimated fromstandard curves.
2.4. Western Blot analysis
The presence of COX-1 and COX-2 proteins inperitoneal leukocytes collected 1, 3, 6, 12 and24 h after i.p. injection of BjV (250 mg/kg), CdtV(25 mg/kg) or saline solution (control) was detectedby western blotting. For all western blottingexperiments, the cells were incubated in a lysisbuffer (0.1% Triton X-100, 50 mM Pepstatin-A,0.2mM Leupeptin, 1 mgml�1 aprotinin, 10mgml�1
phenylmethyl sulfonyl fluoride, 50mM Tris, 10mMEDTA). The samples were then centrifuged and theprotein concentration of the supernatant wasdetermined by colorimetric assay (BioRad, Her-cules, CA). About 30 mg of protein were separatedon a 10% polyacrylamide gel and then transferredto a nitrocellulose membrane. The membrane wasincubated for 1 h with blocking buffer (20mM Tris,100mM NaCl, 0.5% Tween 20, and 5% non-fatdried milk) and then probed 1 h with antibody
against COX-2 (1:500; Cayman Chemical, AnnArbor, Michigan, USA), COX-1 (1:500; Santa CruzBiotechnology, California, USA) and b-actin(1:2000; Santa Cruz Biotechnology, California,USA). The membrane was then incubated with anappropriate peroxidase-conjugated secondary anti-body for 1 h at room temperature. The immunor-eactive bands were visualized by using an enhancedchemiluminescence reagent (ECL; Amersham Phar-macia). Densitometry was done using a GS-710Calibrated Imaging Densitometer (Bio-Rad) andanalyzed with Quantity One software (Bio-Rad).
2.5. Statistical analysis
Results are expressed as mean7S.E.M. Differ-ences among groups were analyzed by one-wayanalysis of variance (ANOVA) followed by eitherTukey’s test or Student’s t-test. Values of prob-ability less than 5% (po0.05) were consideredsignificant.
3. Results
3.1. Release of prostanoids in the peritoneal cavity
To investigate the ability of BjV and CdtV torelease prostanoids in the peritoneal cavity of mice,the concentrations of PGE2 and PGD2 in theperitoneal fluids were measured. Levels of PGE2
were significantly elevated above basal valuesbetween 3 and 12 h (Fig. 1A). Changes in PGD2
concentrations after BjV administration followed avery different pattern: a significant increase inPGD2 synthesis was detected 30min after BjVinjection, with a return towards basal levels at 1 hafter its injection (Fig. 2A). PGE2 and PGD2 wereabsent in the exudates harvested between 30minand 12 h after i.p. injection of CdtV (Fig. 1B and2B). This venom is active since the batch of CdtVhad an i.p. LD50 of 1.3 mg/ 20 g mouse (1.0–1.4 mg/20 g; 95% confidence limits) evaluated in the presentexperimental condition.
3.2. COX-2 and COX-1 expression from peritoneal
leukocytes
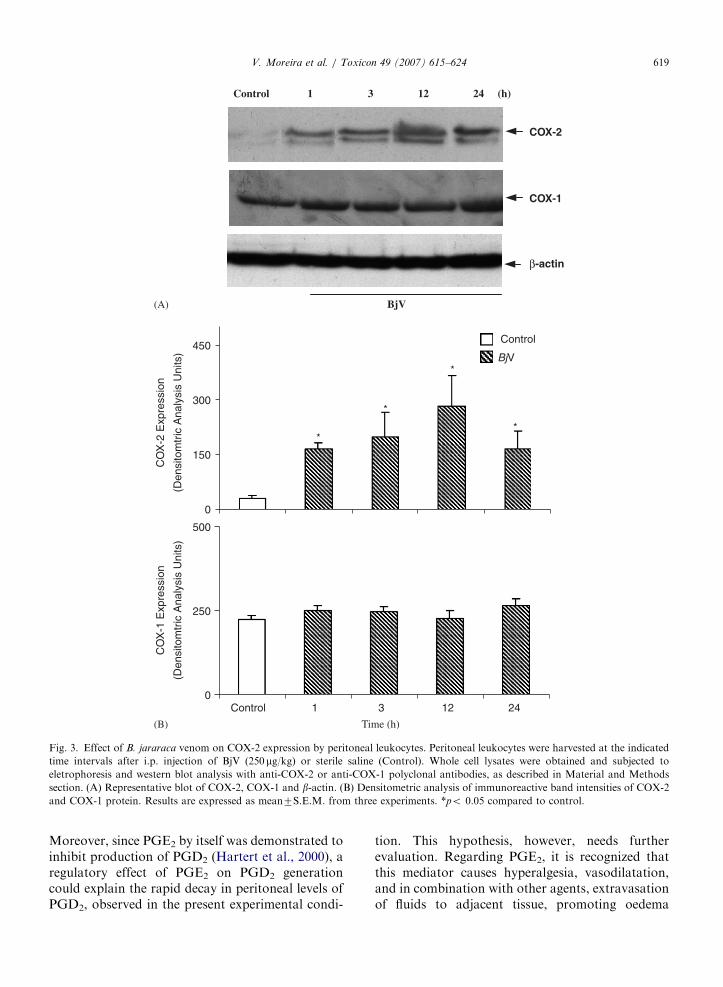
To further explore the mechanism involved invenom-induce inflammatory events, the expressionof COX-1 and COX-2, in the peritoneal leukocytes,was examined by western blotting. Fig. 3 shows asignificant increase in the protein expression of

ARTICLE IN PRESS
0
20000
40000
PG
E2
(pg/
mL)
PG
E2
(pg/
mL)
Control
BjV
*
*
*
0
4500
9000
0.5 1 12
Time (h)
Control
CdtV
3 6
(A)
(B)
Fig. 1. Release of PGE2 induced by i.p. injection of B. jararaca
(A) and C. d. terrificus (B) venoms. Groups of animals were
injected i.p. with BjV (250 mg/kg), CdtV (25mg/kg), or sterile
saline (Control). Concentration of PGE2 was evaluated by
specific EIA in peritoneal washes collected 30min, 1, 3, 6, 12 h
after venom injection. Each bar represents the mean7S.E.M.
from four to five animals. *po0.05 compared to Control.
0
150
300
0.5 1 6 12
Time (h)
Control
CdtV
0
250
500
PG
D2
(pg/
mL)
PG
D2
(pg/
mL)
Control
BjV
*
3
(A)
(B)
Fig. 2. Release of PGD2 induced by i.p. injection of B. jararaca
(A) and C. d. terrificus (B) venoms. Groups of animals were
injected i.p. with BjV (250 mg/kg), CdtV (25mg/kg) or sterile saline(Control). Concentration of PGD2 was evaluated by specific EIA
in peritoneal washes collected 30min, 1, 3, 6, 12 h after venom
injection. Each bar represents the mean7S.E.M. from four to
five animals. *po 0.05 compared to control.
V. Moreira et al. / Toxicon 49 (2007) 615–624618
COX-2 induced by BjV from 3 up to 24 h after itsinjection, with maximum at 12 h. However, thelevels of COX-1 were not modified in peritonealcells obtained from mice injected with this venom incomparison to controls (Fig. 3). With regard toCdtV, this venom did not modify the basal proteinexpression of COX-1 (Fig. 4) and did not induceCOX-2 protein expression in leukocytes collectedfrom 1h up to 24 h after its injection.
4. Discussion
PGs, in addition to playing important regulatoryroles in a broad range of physiological systems alsomodulate several pathophysiological processes suchas inflammation. In this study we have analyzed therelease of PGE2 and PGD2 and their relationship toCOX-1 and COX-2 expression in peritoneal
leukocytes following intraperitoneal injection ofB. jararaca and C. durissus terrificus venoms.
We herein demonstrate that intraperitoneal injec-tion B. jararaca venom into mice induced the releaseof PGD2 at 30min followed by release of PGE2
from 3 up to 12 h with maximal response at 3 h.These results are in accordance with previousreports where local inflammatory events inducedby venoms from Bothrops genus involve participa-tion of eicosanoids derived from arachidonic acidmetabolism (Trebien and Calixto, 1989; Chaveset al., 1995; Zamuner and Teixeira, 2002), and thathyperalgesia and edema caused by BjV weremediated, at least in part, by PGs (Teixeira et al.,1994). It has been shown that PGD2 is involved ininflammatory processes promoting vasodilatationand increase of vascular permeability (Kanaoka andUrade, 2003; Herschman, 1999; Ito et al., 1989).Accordingly, our data showing a rapid onset forPGD2 release suggest a role for this mediator inthe early formation of edema elicited by BjV.
ARTICLE IN PRESS
Time (h)
0
150
300
450
CO
X-2
Exp
ress
ion
(Den
sito
mtr
ic A
naly
sis
Uni
ts)
CO
X-1
Exp
ress
ion
(Den
sito
mtr
ic A
naly
sis
Uni
ts)
*
*
*
*
0
250
500
Control 3 12 24
Control
BjV
COX-1
β-actin
COX-2
Control 1231 24 (h)
BjV
1
(B)
(A)
Fig. 3. Effect of B. jararaca venom on COX-2 expression by peritoneal leukocytes. Peritoneal leukocytes were harvested at the indicated
time intervals after i.p. injection of BjV (250 mg/kg) or sterile saline (Control). Whole cell lysates were obtained and subjected to
eletrophoresis and western blot analysis with anti-COX-2 or anti-COX-1 polyclonal antibodies, as described in Material and Methods
section. (A) Representative blot of COX-2, COX-1 and b-actin. (B) Densitometric analysis of immunoreactive band intensities of COX-2
and COX-1 protein. Results are expressed as mean7S.E.M. from three experiments. *po 0.05 compared to control.
V. Moreira et al. / Toxicon 49 (2007) 615–624 619
Moreover, since PGE2 by itself was demonstrated toinhibit production of PGD2 (Hartert et al., 2000), aregulatory effect of PGE2 on PGD2 generationcould explain the rapid decay in peritoneal levels ofPGD2, observed in the present experimental condi-
tion. This hypothesis, however, needs furtherevaluation. Regarding PGE2, it is recognized thatthis mediator causes hyperalgesia, vasodilatation,and in combination with other agents, extravasationof fluids to adjacent tissue, promoting oedema

ARTICLE IN PRESS
Control 1231 24 (h)
β-actin
COX-1
COX-1
CdtV
0
100
200
Control 3 12 24
ControlCdtV
1
(A)
(B)
200
0
100
CO
X-2
Exp
ress
ion
(Arb
itrar
y U
nits
)C
OX
-1 E
xpre
ssio
n(A
rbitr
ary
Uni
ts)
Fig. 4. Effect of C. d. terrificus venom on COXs expression by
peritoneal leukocytes. Peritoneal leukocytes were harvested at the
indicated time intervals after i.p. injection of CdtV (25mg/kg) orsterile saline (Control). Whole cell lysates were obtained and
subjected to eletrophoresis and western blot analysis with anti-
COX-2 or anti-COX-1 polyclonal antibodies, as described in
Material and Methods section. (A) Representative blot of COX-
2, COX-1 and b-actin. (B) Densitometric analysis of immunor-
eactive band intensities of COX-2 and COX-1 protein. Results
are expressed as mean7S.E.M. from three experiments.
V. Moreira et al. / Toxicon 49 (2007) 615–624620
formation (James et al., 2001). Therefore, from thepresent findings, it is plausible that PGE2 is a criticalmediator for local edema and hyperalgesia inducedby B. jararaca venom, probably synergizing withother mediators, such as histamine and bradykinin.
We next examined whether the observed BjV-induced increases in PGD2 and PGE2 productionwere dependent on protein synthesis. Throughwestern blot analysis, an up-regulation of COX-2protein was detected since 1 h after BjV injection,
and under the same experimental conditions, COX-1 expression remained unchanged. This suggeststhat COX-2 is the critical enzyme to provide moresubstrate (PGH2) for PGs production in peritonealleukocytes in the later periods of venom action. Thisis the first demonstration of the stimulatory effect ofBothrops snake venoms on COX-2 protein expres-sion. However, the mechanisms involved in theability of BjV to induce COX-2 expression were notpresently studied and need more investigation.Cytokine-induced gene expression of COX-2 havebeen reported by several authors using distinctexperimental models, and the regulatory roles ofTNF-a and IL-1 on COX -2 expression have beendemonstrated (Posadas et al., 2000; Smith et al.,2000; Di Rosa et al., 1996). Considering that BjV isable to induce local and systemic release of TNF-and IL-1 (Zamuner et al., 2001; Petricevich et al.,2000), it is herein suggested that these cytokines areimportant messengers for BjV-induced COX-2expression and the consequent increase of PGslevels.
The lack of BjV venom effect on COX-1expression in peritoneal leukocytes does not excludethe possibility of BjV to stimulate, direct orindirectly, the enzymatic activity of COX-1. Thishypothesis could explain the increased productionof PGD2 and PGE2 in the early periods after BjVintraperitoneal injection and deserves further in-vestigation. In this context, an actual line of activediscussion is the functional coupling of COX-1 andCOX-2 to distinct prostanoid synthases. Literatureshows evidence that depending on cell types ordifferent physiological/pathological stimuli, the for-mation of PGE2 is linked predominantly to COX-2whereas the formation of PGD2 is linked to COX-1(Matsumoto et al., 1997; Reddy et al., 1997;Morham et al., 1995). In addition, it is now believedthat the constitutive COX-1 is mainly utilized in theimmediate PG biosynthesis, which occurs withinseveral minutes after stimuli whereas the inducibleCOX-2 is an absolute requirement for delayed PGbiosynthesis, which lasts for several hours followingvarious stimuli (James et al., 2001; Naraba et al.,1998). Taking this information in account, theobserved switch from PGD2 to PGE2 productionin leukocytes three hours after BjV intraperitonealinjection may be at least in part related to an initialstimulatory effect on enzymatic activity of COX-1followed by induction of COX-2 expression, owingto differential conversion of the common precursorPGH2 into PGD2 or PGE2, respectively.

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624 621
In contrast to the results obtained with BjV, weobserved that i.p. injection of CdtV neither modifythe basal levels of PGD2 and PGE2 synthesis nor theconstitutive expression of COX-1. Also, no expres-sion of COX-2 was observed. Differences of effectsbetween both venoms cannot be explained bydifferences in the doses used because in similardoses BjV and CdtV induced inflammatory andanti-inflammatory effects, respectively (Zamuneret al., 2001; Sousa e Silva et al., 1996). It has beenshowed by Chisari et al. (1998) and Cardoso et al.(2001) that CdtV is able to stimulate in vivo therelease of IL-10, a potent anti-inflammatory cyto-kine, which blocks the induction of inflammatorycytokines in human monocytes (de Waal Malefyt etal., 1991) and mouse peritoneal macrophages(Fiorentino et al., 1991), and inhibits COX-2expression (Berg et al., 2001; Niiro et al., 1998).Therefore, it is possible that, in our experimentalmodel, this anti-inflammatory cytokine, producedafter administration of CdtV, can impair theexpression of COX-2 after CdtV injection.
Plasma levels of glucocorticoids are significantlyincreased after CdtV injection by stimulation ofhypothalamus-pituitary-adrenal (HPA) (Chisariet al., 1998). These hormones downregulate variousinflammatory events (Garcia Leme and Schapoval,1975; Garcia Leme and Farsky, 1993; Chrousos,1995; Fantuzzi et al., 1995; Farsky et al., 1995)including COX-2 expression, COX-1 activity andprostanoid secretion (Newton et al., 1997; Fernan-dez-Morata et al., 2000). Therefore it is plausible toestablish a correlation between increased levels ofglucocorticoids and the lack of release of PGs andCOX-2 expression under CdtV stimuli. In agree-ment with this hypothesis is a report showing thatdifferently from CdtV, BjV did not stimulate theHPA axis (Cury et al., 1997), a circumstance whichmight be the responsible for maintenance of theinflammatory response induced by Bothrops snakevenoms. Thus the release of specific cytokines, suchas IL-10, and glucocorticoids, as in the case ofCdtV, may explain the presently observed differ-ences between BjV- and CdtV-induced effects onproduction of prostanoids and COXs expression.
In summary, we provide the first demonstrationthat BjV is able to up regulate the synthesis ofPGD2 and PGE2, as well as COX-2 proteinexpression, but does not affect COX-1 expression.It is likely that expressed COX-2 provides moresubstrate for synthesis of PGs by the respectiveterminal synthases, being the critical enzyme
responsible for PGs production in the late periodsof BjV effect. Therefore, these results suggest thattherapeutic use of selective COX-2 inhibitors maybe of value to avoid the progress of the severe localinflammation induced by BjV in the victims. Incontrast, CdtV neither stimulate the synthesis ofprostanoids nor induce COX-2 expression. There-fore, differences between BjV and CdtV in theability to produce PGs and to activate COXpathway can account for their distinct effects withregard the inflammatory reaction.
Acknowledgements
The authors thank to Maria Zelma da Silva andWanda Carrela for technical assistance. Thisinvestigation was supported by research grant fromFundac- ao de Amparo a Pesquisa do Estado de SaoPaulo (FAPESP) Brazil (grants 02/13863-2).C.F.P.T. is recipient of CNPq—PQ grant andV.M. is recipient of PhD fellowship from FAPESP(Grant 02/13458-0).
References
Amorim, M.F., Mello, R.F., 1954. Intermediate nephron
nephrosis from snake poisoning in man; histopathologic
study. Am. J. Pathol. 30, 479–499.
Barbosa, A.M., do Amaral, R.O., Teixeira, C.F., Hyslop, S.,
Cogo, J.C., 2003. Pharmacological characterization of mouse
hind paw oedema induced by Bothrops insularis (jararaca
ilhoa) snake venom. Toxicon 42, 515–523.
Barr, R.M., Koro, O., Francis, D.M., Black, A.K., Numata, T.,
Greaves, M.W., 1988. The release of prostaglandin D2 from
human skin in vivo and in vitro during immediate allergic
reactions. Br. J. Pharmacol. 94, 773–780.
Barraviera, B., 1990. Curso sobre acidentes por animais pec-
onhentos: Acidentes por serpentes do genero Crotalus. Erq.
Bras Med. 64, 14–20.
Barraviera, B., Lomonte, B., Tarkowiki, A., Hanson, L.A.,
Meira, D.A., 1995. Acute phase reactions including cytokines,
in patients bitten by Bothrops spp. and Crotalus durissus
terrificus in Brazil. J. Venom Anim. Toxins. 1, 11–22.
Berg, D.J., Zhang, J., Lauricella, D.M., Moore, S.A., 2001. Il-10
is a central regulator of cyclooxygenase-2 expression and
prostaglandin production. J. Immunol. 166, 2674–2680.
Brazil, V., 1934. Do emprego da pec-onha em terapeutica. Biol.
Med. 1, 7–21.
Brigatte, P., Hoffmann, F.A., Bernardi, M.M., Giorgi, R.,
Fernandes, I., Takehara, H.A., Barros, S.B.M., Almeida,
M.G., Cury, Y., 2001. Tolerance to the antinociceptive effect
of Crotalus durissus terrificus snake venom in mice is mediated
by pharmacodynamic mechanisms. Toxicon 39, 1399–1440.
Cardoso, D.F., Mota, I., 1997. Effect of Crotalus venom on the
humoral and cellular immune response. Toxicon 35, 607–612.

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624622
Cardoso, D.F., Lopes-Ferreira, M., Faquim-Mauro, E.L.,
Macedo, M.S., Farsky, S.H., 2001. Role of crotoxin, a
phospholipase A2 isolated from Crotalus durissus terrificus
snake venom, on inflammatory and immune reactions.
Mediators Inflamm. 10, 125–133.
Cardoso, J.L., Fan, H.W., Franca, F.O., Jorge, M.T., Leite, R.P.,
Nishioka, S.A., Avila, A., Sano-Martins, I.S., Tomy, S.C.,
Santoro, M.L., 1993. Randomized comparative trial of three
antivenoms in the treatment of envenoming by lance-headed
vipers (Bothrops jararaca) in Sao Paulo, Brazil. Q. J. Med. 86,
315–325.
Chaves, F., Barbosa, M., Gutierrez, J.M., 1995. Pharmacological
study of edema induced by venom of the snake Bothrops asper
(Terciopelo) in mice. Toxicon 33, 31–39.
Chisari, A., Spinedi, E., Voirol, M.J., Giovambattista, A.,
Gaillard, R.C., 1998. A phospholipase A2-related snake
venom (from Crotalus durissus terrificus) stimulates neuroen-
docrine and immune functions: determination of different
sites of action. Endocrinology 139, 617–625.
Chrousos, G.P., 1995. The hypothalamic-pituitaqry-adrenal axis
and immune-mediated inflammation. N. Engl. J. Med. 332,
1351–1362.
Cunha, F.Q., Lorenzetti, B.B., Poole, S., Ferreira, S.H., 1991.
Interleukin-8 as a mediator of sympathetic pain. Br. J.
Pharmacol. 104, 765–767.
Cunha, F.Q., Poole, S., Lorenzetti, B.B., Ferreira, S.H., 1992.
The pivotal role of tumour necrosis factor alpha in the
development of inflammatory hyperalgesia. Br. J. Pharmacol.
107, 660–664.
Cupo, P., Azevedo-Marques, M.M., Hering, S.E., 1988. Clinical
and laboratory features of South American rattlesnake
(Crotalus durissus terrificus) envenomation in children. Trans.
R. Soc. Trop. Med. Hyg. 82, 924–929.
Cury, Y., Teixeira, C.F.P., Farsky, S.H.P., 1997. Lack of effect of
endogenous corticosteroids on the acute inflammatory reac-
tion (edema) induced by Bothrops jararaca venom (BJV) in
rats. Toxicon 35, 773–776.
Davidson, J., Kerr, A., Guy, K., Rotondo, D., 1998. Prosta-
glandin and fatty acid modulation of Escherichia coli O157
phagocytosis by human monocytic cells. Immunology 94,
228–234.
de Waal Malefyt, R., Abrams, J., Bennett, B., Figdor, C.G., de
Vries, J.E., 1991. Interleukin 10 (IL-10) inhibits cytokine
synthesis by human monocytes: an autoregulatory role of IL-
10 produced by monocytes. J. Exp. Med. 174, 1209–1220.
Di Rosa, M., Ialenti, A., Ianaro, A., Sautebin, L., 1996.
Interaction between nitric oxide and cyclooxygenase path-
ways. Prostaglandins Leukot. Essent. Fatty Acids 54,
229–238.
Fantuzzi, G., Di Santo, E., Sacco, S., Benigni, F., Ghezzi, P.,
1995. Role of the hypothalamus-pituitary-adrenal axis in the
regulation of TNF production in mice. Effect of stress and
inhibition of endogenous glucocorticoids. J. Immunol. 155,
3552–3555.
Farsky, S.H.P., Costa-Cruz, J.W.M., Cury, Y., Teixeira, C.F.P.,
1997. Leokocyte response induced by Bothrops jararaca
crude venom. In vivo and in vitro studies. Toxicon 35,
85–93.
Farsky, S.P., Sannomiya, P., Garcia-Leme, J., 1995. Secreted
glucocorticoids regulate leukocyte-endothelial interactions in
inflammation. A direct vital microscopic study. J. Leukoc.
Biol. 57, 379–386.
Fernandez-Morata, J.C., Mullol, J., Fuentes, M., Pujols, L.,
Roca-Ferrer, J., Perez, M., Xaubet, A., Picado, C., 2000.
Regulation of cyclooxygenase-1 and -2 expression in human
nasal mucosa. Effects of cytokines and dexamethasone. Clin.
Exp. Allergy 30 (9), 1275–1284.
Fiorentino, D.F., Zlotnik, A., Mosmann, T.R., Howard, M.,
O’Garra, A., 1991. IL-10 inhibits cytokine production by
activated macrophages. J. Immunol. 147, 3815–3822.
Flores, C.A., Zappellini, A., Prado-Franceschi, J., 1993. Lypox-
ygenase-derived mediators may be involved in in vivo
neutrophil migration induced by Bothrops erytromelas and
Bothrops alternatus venom. Toxicon 31, 1551–1559.
Garcia Leme, J., Farsky, S.P., 1993. Hormonal control of
inflammatory responses. Mediat. Inflamm. 2, 181–198.
Garcia Leme, J., Schapoval, E.E.S., 1975. Stimulation of the
hypothalamus-pituitary-adrenal axis by compounds formed
in inflamed tissue. Br. J. Pharmac. 53, 75–83.
Giles, H., Leff, P., 1988. The biology and pharmacology of
PGD2. Prostaglandins 35, 277–300.
Giorgi, R., Bernardi, M.M., Cury, Y., 1993. Analgesic effect
evoked by low molecular weight substances extracted from
Crotalus durissus terrificus venom. Toxicon 31, 1257–1265.
Harris, S.G., Padilla, J., Koumas, L., Ray, D., Phipps, R.P.,
2002. Prostaglandins as modulators of immunity. Trends
Immunol. 23, 144–150.
Hart, P.H., 2001. Regulation of the inflammatory response in
asthma by mast cell products. Immunol. Cell Biol. 79,
149–153.
Hartert, T.V., Dworski, R.T., Mellen, B.G., Oates, J.A., Murray,
J.J., Sheller, J.R., 2000. Prostaglandin E(2) decreases allergen-
stimulated release of prostaglandin D(2) in airways of subjects
with asthma. Am. J. Respir. Crit. Care Med. 162, 637–640.
Hata, A.N., Breyer, R.M., 2004. Pharmacology and signaling of
prostaglandin receptors: multiple roles in inflammation and
immune modulation. Pharmacol. Therap. 103, 147–166.
Herschman, H.R., 1999. Function and regulation of prostaglan-
din synthase 2. Adv. Exp. Med. Biol. 469, 3–8.
Ito, S., Narumiya, S., Hayaishi, O., 1989. Prostaglandin D2: a
biochemical perspective. Prostaglandins Leukot. Essent.
Fatty Acids 37, 219–234.
James, M.J., Penglis, P.S., Caughey, G.E., Demasi, M., Clenland,
L.G., 2001. Eicosanoids production by human monocytes:
does COX-2 contribute to a self-limiying inflammatory
response? Inflamm. Res. 50, 249–253.
Kanaoka, Y., Urade, Y., 2003. Hematopoietic prostaglandin D
synthase. Prostaglandins Leukot. Essent. Fatty Acids 69,
163–167.
Kennedy, C.R., Zhang, Y., Brandon, S., Guan, Y., Coffe, K.,
Funk, C.D., Magnuson, M.A., Oates, J.A., Breyer, M.D.,
Breyer, R.M., 1999. Salt-sensitive hypertension and reduced
fertility in mice lacking the prostaglandin EP2 receptor. Nat.
Med. 5, 217–220.
Knundsen, P.J., Dinarello, C.A., Strom, T.B., 1986. Prostaglan-
dins post-transcriptionally inhibit monocyte expression of
interleukin-1 activity by increasing intracellular cyclic adeno-
sine monophosphate. J. Immunol. 137, 3189–3194.
Kozlov, V., Poveshchenko, A., Gromykhina, N., 1990. Some
mechanisms involved in the prostaglandin E2 immunosup-
pressive effect in (CBA�C57BL)F1 mice in vivo. Cell.
Immunol. 128, 242–249.
Kunkel, S.L., Wiggins, R.C., Chensue, S.W., Larrick, J., 1986.
Regulation of macrophage tumor necrosis factor production

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624 623
by prostaglandin E2. Biochem. Biophys. Res. Commun. 137,
404–410.
Landucci, E.C., Antunes, E., Donato, J.L., Faro, R., Hyslop, S.,
Marangoni, S., Oliveira, B., Cirino, G., de Nucci, G., 1995.
Inhibition of carrageenin-induced rat paw oedema by
crotapotin, a polypeptide complexed with phospholipase
A2. Br. J. Pharmacol. 114, 578–583.
Matsumoto, H., Naraba, H., Murakami, I., Kudo, I., Yakami,
K., Ueno, A., Oh-Ishi, S., 1997. Concordant induction of
prostaglandin E2 synthase with cyclooxygenase-2 leads to
preferred production of prostaglandin E2 over thromboxane
and prostaglandin D2 in lipopolysaccharide-stimulated rat
peritoneal macrophages. Biochem. Biophys. Res. Commun.
230, 110–114.
Mizoguchi, A., Eguchi, N., Kimura, K., Kiyohara, Y., Qu,
W.M., Huang, Z.L., Mochizuki, T., Lazarus, M., Kobayashi,
T., Kaneko, T., Narumiya, S., Urade, Y., Hayaishi, O., 2001.
Dominant localization of prostaglandin D receptors on
arachinoid trabecular cells in mouse basal forebrain and their
involvement in the regulation of non-rapid eye movement
sleep. Proc. Natl. Acad. Sci. USA 98, 11674–11679.
Molloy, E.S., McCarthy, G.M., 2005. Eicosanoids, osteoarthritis,
and crystal deposition diseases. Curr. Opin. Rheumatol. 17,
346–350.
Morham, S.G., Langenbach, R., Loftin, C.D., Tiano, H.F.,
Vouloumanos, N., Jennette, J.C., Mahler, J.F., Kluckman,
K.D., Ledford, A., Lee, C.A., Smithies, O., Smithies, O.,
1995. Prostaglandin synthase 2 gene disruption causes severe
renal pathology in the mouse. Cell 83, 473–482.
Murray, J.J., Tonnel, A.B., Brash, A.R., Roberts II, L.J., Gosset,
P., Workman, R., Capron, A., Oates, J.A., 1986. Release of
prostaglandin D2 into human airways during acute antigen
challenge. N. Engl. J. Med. 315, 800–804.
Nakamura, M., Ferreira, S.H., 1987. A peripheral sympathetic
component in inflammatory hyperalgesia. Eur. J. Pharmacol.
135, 145–153.
Naraba, H., Murakami, M., Matsumoto, H., Shimbara, S.,
Ueno, A., Kudo, I., Oh-Ishi, S., 1998. Segregated coupling of
phospholipases A2, cyclooxygenases, and terminal prostanoid
synthases in different phases of prostanoid biosynthesis in rat
perotoneal macrophages. J. Immunol. 160, 2974–2982.
Narumiya, S., Fitzgerald, G.A., 2001. Genetic and pharmacolo-
gical analysis of prostanoid receptor function. Clin. Invest.
108, 25–30.
Nataraj, C., Thomas, D.W., Tilley, S.L., Nguyen, M.T.,
Mannon, R., Koller, B.H., Coffman, T.M., 2001. Receptors
for prostaglandin E(2) that regulate cellular immune re-
sponses in the mouse. J. Clin. Invest. 108, 1229–1235.
Newton, R., Kuitert, L.M., Slater, D.M., Adcock, I.M., Barnes,
P.J., 1997. Cytokine induction of cytosolic phospholipase A2
and cyclooxygenase-2 mRNA is suppressed by glucocorti-
coids in human epithelial cells. Life Sci. 60, 67–78.
Niiro, H., Otsuka, T., Ogami, E., Yamaoka, K., Nagano, S.,
Akahoshi, M., Nakashima, H., Arinobu, Y., Izuhara, K.,
Niho, Y., 1998. MAP kinase pathways as a route for
regulatory mechanisms of IL-10 and IL-4 which inhibit
COX-2 expression in human monocytes. Biochem. Biophys.
Res. Commun. 250, 200–205.
Nishijima, M., Amano, F., Akamatsu, Y., Akagawa, K.,
Tokunaga, T., Raetz, C.R., 1985. Macrophage activation by
monosaccharide precursors of Escherichia coli lipid A. Proc.
Natl. Acad. Sci. USA 82, 282–286.
Oropeza-Rendon, R.L., Bauer, H.C., Fischer, H., 1980. Effect of
prostaglandin E1 on the level of cAMP in bone marrow
macrophages. Inhibition of phagocytosis and cell shape
changes. J. Immunopharmacol. 2, 133–147.
Petricevich, V.L., Teixeira, C.F.P., Tambourgi, D.V., Gutierrez,
J.M., 2000. Increments in cytokine and nitric oxide serum
levels in mice injected with Bothrops asper and Bothrops
jararaca snake venoms. Toxicon 38, 1253–1266.
Phipps, R.P., Stein, S.H., Roper, R.L., 1991. A new view of
prostaglandin E regulation of the immune response. Immu-
nol. Today 12, 349–352.
Picolo, G., Giorgi, R., Bernardi, M.M., Cury, Y., 1998. The
antinociceptive effect of Crotalus durissus terrificus snake
venom is mainly due to a supraspinally integrated response.
Toxicon 36, 223–227.
Poole, S., Lorenzetti, B.B., Cunha, J.M., Cunha, F.Q., Ferreira,
S.H., 1999. Bradykinin B1 and B2 receptors, tumour necrosis
factor alpha and inflammatory hyperalgesia. Br. J. Pharma-
col. 126, 649–656.
Portanova, J.P., Zhang, Y., Anderson, G.D., Hauser, S.D.,
Masferrer, J.L., Seibert, K., Gregory, S.A., Isakon, P.C.,
1996. Selective neutralization of prostaglandin E2 blocks
inflammation, hyperalgesia, and interleukin 6 production in
vivo. J. Exp. Med. 184, 883–891.
Posadas, I., Terencio, M.C., Guillen, I., Ferrandiz, M.L.,
Coloma, J., Paya, M., Alcaraz, M.J., 2000. Co-regulation
between cyclo-oxygenase-2 and inducible nitric oxide synthase
expression in the time-course of murine inflammation. Arch.
Pharmacol. 361, 98–106.
Pradelles, P., Grassi, J., Mac Louf, J., 1985. Enzyme immunoas-
says of eicosanoids using acetylcholine esterase as label: an
alternative to radioimmunoassay. Analyt. Chem. 57,
1170–1173.
Reddy, S.T., Winstead, M.V., Tischfield, J.A., Herschman, H.R.,
1997. Analysis of the secretory phospholipase A(2) that
mediates prostaglandin production in mast cells. J. Biol.
Chem. 272, 13591–13596.
Rocca, B., Fitzgerald, G.A., 2002. Cyclooxygenases and pros-
taglandins: shaping up the immune response. Int. Immuno-
pharmacol. 2, 603–630.
Rosenfeld, G., 1971. Symptomatology, pathology and treatment
of snakebites in South America. In: Bucherl, W., Buckley, E.
(Eds.), Venomous Animals and their Venoms. Academic
Press, New York, pp. 345–362.
Sampaio, S.C., Alba-Loureiro, T.C., Brigatte, P., Landgraf,
R.G., dos Santos, E.C., Curi, R., Cury, Y., 2006. Lipox-
ygenase-derived eicosanoids are involved in the inhibitory
effect of Crotalus durissus terrificus venom or crotoxin on rat
macrophage pahgocytosis. Toxicon 47, 313–321.
Smith, W.L., Dewitt, D.L., 1996. Prostaglandin endoperoxide H
synthases-1 and -2. Adv. Immunol. 62, 167–215.
Smith, W.L., Dewitt, D.L., Garavito, R.M., 2000. Cyclooxy-
genases: structural, cellular, and molecular biology. Annu.
Rev. Biochem. 69, 145–182.
Smith, W.L., Langenbach, R., 2001. Why there are two
cyclooxygenase isozymes? J. Clin. Invest. 107, 1491–1495.
Sousa e Silva, M.C.C., Gonc-alves, L.R.C., Mariano, M., 1996.
The venom of South American rattlesnakes inhibits macro-
phage functions and is endowed with anti-inflammatory
properties. Med. Inflam. 5, 18–23.
Takayama, K., Garcia-Cardena, G., Sukhova, G.K., Comander,
J., Gimbrone Jr., M.A., Libby, P., 2002. Prostaglandin E2

ARTICLE IN PRESSV. Moreira et al. / Toxicon 49 (2007) 615–624624
supresses chemokine production in human macrophages
though the EP4 receptor. J. Biol. Chem. 277, 44147–44154.
Teixeira, C.F., Cury, Y., Oga, S., Jancar, S., 1994. Hyperalgesia
induced by Bothrops jararaca venom in rats: role of
eicosanoids and platelet activating factor (PAF). Toxicon
32, 419–426.
Trebien, H.A., Calixto, J.B., 1989. Pharmacological evaluation of
rat paw oedema induced by Bothrops jararaca venom. Agents
Actions 26, 292–300.
Trebino, C.E., Stock, J.L., Gibbons, C.P., Naiman, B.M.,
Wachtmann, T.S., Umland, J.P., Pandher, K., Lapointe,
J.M., Saha, S., Roah, M.L., Carter, D., Thomas, N.A.,
Durtschi, B.A., McNeish, J.D., Hambor, J.E., Jakobsson,
P.J., Carty, T.J., Perez, J.P., Audoly, L.P., 2003. Impaired
inflammatory and pain responses in mice lacking an inducible
prostaglandin E synthase. Proc. Natl. Acad. Aci. 100,
9044–9049.
Warner, T.D., Giuliano, F., Vojnovic, I., Bukasa, A., Michell,
J.A., Vane, J.R., 1999. Nonsteroid drug selectivities for cyclo-
oxygenase-1 rather than cyclo-oxigenase-2 are associated with
human gastrointestinal toxicity: a full in vitro analysis. Proc.
Natl. Acad. Sci. USA 95, 7563–7568.
Zamuner, S.R., Gutierrez, J.M., Muscara, M.N., Teixeira, S.A.,
Teixeira, C.F., 2001. Bothrops asper and Bothrops jararaca
snake venoms trigger microbicidal functions of peritoneal
leukocytes in vivo. Toxicon 39, 1505–1513.
Zamuner, S.R., Teixeira, C.F., 2002. Cell adhesion molecules
involved in the leukocyte recruitment induced by venom of
the snake Bothrops jararaca. Mediators Inflamm. 11,
351–357.











![The effect of non-steroidal anti-inflammatory drugs on the ......role of PG in bone tissue, potentially causing a shift in pre-cursor cell action towards bone resorption [6]. Cyclooxygenases](https://static.fdocuments.net/doc/165x107/60f6870aabd189398c680426/the-effect-of-non-steroidal-anti-inflammatory-drugs-on-the-role-of-pg-in.jpg)






