
Biotechnological Developments in Sugarca
14
REVIEW ARTICLE Biotechnological Developments in Sugarcane Improvement: An Overview P. Suprasanna • V. Y. Patade • N. S. Desai • R. M. Devarumath • P. G. Kawar • M. C. Pagariya • A. Ganapathi • M. Manickavasagam • K. H. Babu Received: 22 July 2011 / Accepted: 10 October 2011 / Published online: 2 November 2011 Ó Society for Sugar Research & Promotion 2011 Abstract Sugarcane (Saccharum officinarum L.) is one of the most important field crops grown in the tropics and sub-tropics. More than half of the world’s sugar is derived from sugar cane. Conventional methods have greatly con- tributed to crop improvement; however limitations such as complex genome, narrow genetic base, poor fertility, sus- ceptibility to biotic and abiotic stresses and long duration to breed elite cultivars still impose a challenge. Sugarcane, thus, is a suitable candidate for application of biotechnol- ogy and genetic engineering tools. In this direction, in vitro culture systems and related biotechnologies have been developed as novel strategies for sugarcane improvement. Studies have been conducted towards employing in vitro culture combined with radiation/chemical induced muta- genesis for mutant isolation. Advancements in genomics tools have paved the way for a detailed understanding of the mechanism underlying biotic and abiotic stress responses. The potential of the current genomics programs, aimed at elucidating the structure, function, and interac- tions of the sugarcane genes, will revolutionize the appli- cation of biotechnology to crop improvement. Genetically modified sugarcane with increased resistance to agronomic traits including biotic and abiotic stresses, yield and juice could become useful in breeding for better varieties. This review outlines some of the biotechnological developments that are in place and tailored to address important issues related to sugarcane improvement. Keywords Sugarcane Á Biotechnology Á Genomics Á Molecular markers Á Stress tolerance Á In vitro culture Á Mutagenesis Á Transgenic plants Introduction Sugarcane (Saccharum spp.) is an important industrial crop, ranking among the ten most planted crops in the world. Besides being the major sugar contributor with more than 70% of the world’s sugar, sugarcane is important as the raw material for sugar producing and allied industries. India is the largest single producer of sugar including tra- ditional cane sugar sweeteners, khandsari and Gur equiv- alent to 26 million tonnes raw value followed by Brazil in the second place (Indian Sugar 2008). The Saccharum complex includes the agronomically and industrially P. Suprasanna (&) Functional Plant Biology Section, Nuclear Agriculture & Biotechnology Division, Bhabha Atomic Research Centre, Trombay, Mumbai 400085, India e-mail: [email protected] V. Y. Patade Molecular Biology and Genetic Engineering Division, Defence Institute of Bio-Energy Research, Haldwani, Nainital 263 139, Uttarakhand, India N. S. Desai Department of Biotechnology & Bioinformatics, Padmashree Dr. D.Y. Patil University, Sect-15, C.B.D. Belapur, Navi Mumbai 400614, India R. M. Devarumath Á P. G. Kawar Á M. C. Pagariya Á K. H. Babu Molecular Biology & Genetic Engineering Division, Vasantdada Sugar Institute, Manjari (Bk.), Pune 412307, Maharashtra, India A. Ganapathi Department of Biotechnology, School of Life Sciences, Bharathidasan University, Tiruchirapalli 620024, Tamil Nadu, India M. Manickavasagam Department of Bioinformatics, School of Life Sciences, Bharathidasan University, Tiruchirapalli 620024, Tamil Nadu, India 123 Sugar Tech (December 2011) 13(4):322–335 DOI 10.1007/s12355-011-0103-3
-
Upload
kambaska-kumar-behera -
Category
Documents
-
view
226 -
download
1
description
na
Transcript of Biotechnological Developments in Sugarca

REVIEW ARTICLE
Biotechnological Developments in Sugarcane Improvement:An Overview
P. Suprasanna • V. Y. Patade • N. S. Desai • R. M. Devarumath •
P. G. Kawar • M. C. Pagariya • A. Ganapathi • M. Manickavasagam •
K. H. Babu
Received: 22 July 2011 / Accepted: 10 October 2011 / Published online: 2 November 2011! Society for Sugar Research & Promotion 2011
Abstract Sugarcane (Saccharum officinarum L.) is oneof the most important field crops grown in the tropics and
sub-tropics. More than half of the world’s sugar is derived
from sugar cane. Conventional methods have greatly con-tributed to crop improvement; however limitations such as
complex genome, narrow genetic base, poor fertility, sus-
ceptibility to biotic and abiotic stresses and long duration tobreed elite cultivars still impose a challenge. Sugarcane,
thus, is a suitable candidate for application of biotechnol-
ogy and genetic engineering tools. In this direction, in vitro
culture systems and related biotechnologies have beendeveloped as novel strategies for sugarcane improvement.
Studies have been conducted towards employing in vitro
culture combined with radiation/chemical induced muta-genesis for mutant isolation. Advancements in genomics
tools have paved the way for a detailed understanding of
the mechanism underlying biotic and abiotic stressresponses. The potential of the current genomics programs,
aimed at elucidating the structure, function, and interac-
tions of the sugarcane genes, will revolutionize the appli-cation of biotechnology to crop improvement. Genetically
modified sugarcane with increased resistance to agronomic
traits including biotic and abiotic stresses, yield and juicecould become useful in breeding for better varieties. This
review outlines some of the biotechnological developments
that are in place and tailored to address important issuesrelated to sugarcane improvement.
Keywords Sugarcane ! Biotechnology ! Genomics !Molecular markers ! Stress tolerance ! In vitro culture !Mutagenesis ! Transgenic plants
Introduction
Sugarcane (Saccharum spp.) is an important industrialcrop, ranking among the ten most planted crops in the
world. Besides being the major sugar contributor with more
than 70% of the world’s sugar, sugarcane is important asthe raw material for sugar producing and allied industries.
India is the largest single producer of sugar including tra-
ditional cane sugar sweeteners, khandsari and Gur equiv-alent to 26 million tonnes raw value followed by Brazil in
the second place (Indian Sugar 2008). The Saccharumcomplex includes the agronomically and industrially
P. Suprasanna (&)Functional Plant Biology Section, Nuclear Agriculture &Biotechnology Division, Bhabha Atomic Research Centre,Trombay, Mumbai 400085, Indiae-mail: [email protected]
V. Y. PatadeMolecular Biology and Genetic Engineering Division, DefenceInstitute of Bio-Energy Research, Haldwani, Nainital 263 139,Uttarakhand, India
N. S. DesaiDepartment of Biotechnology & Bioinformatics, Padmashree Dr.D.Y. Patil University, Sect-15, C.B.D. Belapur, Navi Mumbai400614, India
R. M. Devarumath ! P. G. Kawar ! M. C. Pagariya ! K. H. BabuMolecular Biology & Genetic Engineering Division, VasantdadaSugar Institute, Manjari (Bk.), Pune 412307, Maharashtra, India
A. GanapathiDepartment of Biotechnology, School of Life Sciences,Bharathidasan University, Tiruchirapalli 620024, Tamil Nadu,India
M. ManickavasagamDepartment of Bioinformatics, School of Life Sciences,Bharathidasan University, Tiruchirapalli 620024, Tamil Nadu,India
123
Sugar Tech (December 2011) 13(4):322–335
DOI 10.1007/s12355-011-0103-3

important sugarcane genotypes obtained from S. officina-rum, S. spontaneum and S. robustum crosses. Conventionalbreeding has greatly contributed to the development of
agronomically improved varieties, however limitations
such as narrow gene pool, complex genome, poor fertility,and the long breeding/selection cycle make it difficult to
undertake further improvement. In addition, modern vari-
eties have a variable chromosome number (2n = 100–120)and rarely flower. Sugarcane is a typical glycophyte and
hence exhibits stunted growth or no growth under salinity,with its yield falling to 50% or less than its true potential.
To sustain sugarcane production and to improve the pro-
ductivity, tolerance to biotic and abiotic stresses, nutrientmanagement, and improved sugar recovery are some of the
challenges. Both the conventional and biotechnological
methods need to be integrated in solving some of theseconstraints.
Agronomically improved sugarcane varieties endowed
with tolerance to biotic and abiotic stresses are highlybeneficial, as unfavourable environmental factors can
challenge cultivation and crop productivity. Although
crops tolerant to biotic and abiotic stresses have beenselected by traditional breeding programs, speeding up the
pace is essential in developing improved varieties. Salinity
in the root zones of sugarcane decreases the sucrose yield,through its effect on both biomass and juice quality. The
complexity and polygenic nature of salinity tolerance has
further limited the efforts to develop the tolerant cropvarieties through conventional breeding practices. In this
regard, biotechnological approaches including somaclonal
variation, in vitro mutagenesis and selection are beingapplied for the isolation of agronomically useful mutants
(Jain 2005). In vitro mutagenesis and selection has been
successfully used for improvement in agronomic traits likesalinity and drought tolerance in different crop plants
advocating that tissue culture selection is useful to select
stress tolerant clones. Many examples related to differentvegetatively propagated plants, show that the combination
of in vitro culture with selection is relatively inexpensive,
simple and efficient. Studies using in vitro cultures com-bined with radiation induced mutagenesis have been con-
ducted to isolate mutant clones in sugarcane (Patade and
Suprasanna 2008).There has been a tremendous success in gene transfer
from a wide variety of plant and non-plant sources to
sugarcane. With the availability of efficient transformationsystems, selectable marker genes and genetic engineering
tools, it should also be possible to clone and characterize
useful genes and improve commercially important traitsinto elite germplasm that subsequently can lead to develop
an ideal plant type of sugarcane (Lakshmanan et al. 2005).
Figure 1 presents areas of contemporary research interestand progress. The range of potential applications being
developed through transgenic plants in sugarcane include,
insect resistance, resistance to viruses, altered sucrosecontent, lignin modification, sucrose accumulation and
herbicide resistance. Sustained efforts are being made to
engineer sugarcane varieties that can produce high-valuecompounds such as pharmaceutically important proteins,
functional foods and nutraceuticals, biopolymers and pre-
cursors and enzymes and biopigments. This will go a longway in launching sugarcane as a biofactory (Brumbley
et al. 2008).
Sugarcane Fingerprinting and Marker AssistedSelection
DNA markers have contributed greatly, for fingerprintingof elite genetic stocks, assessing of genetic diversity,
increasing the efficiency of trait selection, and diagnostics.
Various molecular marker systems including restrictionfragment length polymorphism (Lu et al. 1994), ribosomal
RNA (Glaszmann et al. 1990), mitochondria and chloro-
plast genes (D’Hont et al. 1993), RAPDs (Nair et al. 1999;Kawar et al. 2009), and simple sequence repeats (SSR;
Cordeiro et al. 2003; Selvi et al. 2003) have shown use-
fulness for differentiating the genera and to assess germ-plasm diversity within the genus Saccharum.
Fig. 1 Sugarcane biotechnology: research areas of contemporaryprogress
Sugar Tech (December 2011) 13(4):322–335 323
123

RAPD markers have been used to assess the genetic
diversity in elite and exotic sugarcane germplasm (Kawaret al. 2009; Srivastava and Gupta 2006; Nair et al. 1999,
2002) and construct genetic maps (Mudge et al. 1996). The
potential of ISSR markers for molecular profiling wasassessed in sugarcane using 42 varieties of subtropical
India (Srivastava and Gupta 2008). Virupakshi and Naik
(2008) used organellar genome inter-simple sequencerepeat markers (cplSSR and mtISSR) to analyze red rot
disease resistant/moderately resistant and susceptible elitesugarcane (Saccharum spp. hybrid) genotypes. The results
indicated that these markers may be used as a new tool for
the identification of the disease resistant varieties. In sug-arcane AFLP markers have been used to study diversity
existing among tropical and subtropical Indian sugarcane
cultivars (Selvi et al. 2005), and Saccharum complex andErianthus (Selvi et al. 2006; Besse et al. 1998).
One of the major ways sugarcane industries have
already benefited from molecular markers is the use ofSSRs for cultivar identification. SSR markers have been
used to fingerprint 180 sugarcane varieties and the data
stored in a database which could provide a source ofinformation to identify varieties of unknown or disputed
origin and as additional information in Plant Breeding
Rights applications and for quality assurance for deliveryof new cultivars to the industry (Piperidis et al. 2004).
Target region amplification polymorphism (TRAP) has
also been used to characterize the germplasm from thegenera Saccharum, Miscanthus, and Erianthus with the
help of six primers designed using sucrose- and cold tol-
erance-related expressed sequence tags (EST) sequences(Alwala et al. 2006). Development of new high-throughput
marker systems like single nucleotide polymorphisms
(SNPs) and diversity array technology (DArT) markers areexpected to have a major impact in the future for sugarcane
improvement.
Sugarcane Functional Genomics
In spite of immense economic importance, sugarcane
genetics has received relatively little attention as compared
to other crops, mainly due to its highly heterozygous,polyploid and frequently aneuploid nature, complex gen-
ome, poor fertility, and the long breeding/selection cycle
(Gupta et al. 2010). However advancement in moderntechnologies, including development of highly efficient
DNA sequencing techniques, identification of SNPs and
genome mapping, DNA microarray technologies for geneexpression analysis, RNAi (RNA interference) technology
and the rapid improvement in data mining tools, can have a
major influence on future sugarcane crop improvementprograms. At present, both micro- and macro arrays are
being used for the identification of genes expressed spe-
cifically in stems, disease resistance genes, and thoseinvolved in carbohydrate metabolism (Ulian 2000; Grivet
et al. 2001; Casu et al. 2005). Moreover the sequencing of
sugarcane ESTs greatly contributed to the gene discoveryprocess (Prabu et al. 2010). Prior to June 1996, the public
databases of DNA sequences had only 28 sequences from
sugarcane compared to the 270,000 sugarcane sequencesthat are deposited to date (P.G. Kawar, unpublished).
Another investigation using SSH approach has providedinsights into identification of salt induced genes in sugar-
cane leaves specifically to target rare transcripts, such as
those participating in cell signaling and the regulation ofgene expression (Patade et al. 2011b). The validated
expression data by real-time PCR can aid in assigning
function for the sugarcane genes and characterization ofregulatory sequences in sugarcane.
Genomics for Biotic Stress Tolerance
Sugarcane being a long standing crop is constantly chal-lenged by herbivorous insects, nematodes, fungi, bacteria,
and viruses. Plant defence responses to such perturbations
are largely mediated by phytohormones through triggeringconserved defence mechanisms, each with an intricate
signaling pathway leading to plant protection. It has been
shown that both the ethylene and jasmonic acid signalingpathways act synergistically in plant defence.
In a study to identify red-rot-related genes, Gupta et al.
(2010) used enriched subtractive cDNA library preparedfrom the C. falcatum challenged stem of sugarcane variety
(Co 1148) and reported at least 85 red-rot-specific clusters,
unique and not reported in the database previously. Inanother study cDNA-SSH library was constructed and
analyzed to identify the up-regulated genes in sugarcane
under SCGS infection condition. Subtracted library highlyrepresented genes potentially involved in cell rescue,
defence, ageing and apoptosis (13.1%). The forward SSH
approach implemented, allowed to explicate the transcrip-tional regulatory mechanisms of sugarcane in response to
SCGS infection and isolated the R2R3-MYB (SoMYB18)
gene, a potential candidate playing important roles in theregulation of secondary metabolism, signal transduction
during biotic, abiotic and other environmental stresses. In
an attempt towards studying the host–pathogen interactionand decipher the molecular basis of virulence of sugarcane
SCGS disease, Kawar et al. (2010a) isolated partial gen-
ome of first Asiatic strain of phytoplasma (SCGS) bygenomic-SSH. The library yielded 83 SCGS specific
fragments representing approximately 42% of the chro-
mosome of Sugarcane grassy shoot phytoplasma, com-prising approximately 85 predicted partial phytoplasmal
324 Sugar Tech (December 2011) 13(4):322–335
123

CDS. Further, a species specific detection method was
developed for early detection of SCGS infection (Kawaret al. 2010b).
Genomics for Abiotic Stress Tolerance
Salinity and drought are important environmental factorsthat limit crop productivity. Sugarcane, being a typical
glycophyte, exhibits stunted growth or no growth undersalinity, with its yield falling to 50% or even more as
compared to its true potential (Akhtar et al. 2003; Wie-
denfeld 2008). High salt environment adversely affectsplant growth due to alterations in water relations, ionic and
metabolic perturbations, generation of reactive oxygen
species (ROS), and tissue damage (Patade et al. 2011a),enzymes involved in sugar metabolism (Gomathi and
Thandapani 2005) and respond with an altered expression
of stress responsive genes, which may ameliorate the det-rimental effects of salinity. Therefore construction of
cDNA libraries enriched for differentially expressed tran-
scripts is an important first step in attempting to studystress responsive genes. Patade et al. (2011b) constructed a
forward subtracted cDNA library from sugarcane plants
stressed with NaCl (200 mM) for 0.5–18 h to find mRNAspecies that are differentially expressed in sugarcane in
response to salinity stress. Sequencing the differentially
expressed few cDNAs clones led to the identification ofsalinity induced shaggy-like kinase (designated as sugar-
cane shaggy like protein kinase-SuSk). The expression was
induced by salt as well as PEG stress indicating that theinduction of this gene probably occurred in response to the
osmotic component of salt stress rather than the ionic
component.Gene expression profiling is an important tool to
investigate responses to environmental changes at the
transcriptional level. Transcriptomic study of short-term(up to 24 h) salt (NaCl, 200 mM) or iso-osmotic polyeth-
ylene glycol-PEG 8000 (20% w/v) stress has revealed
altered expression of representative stress responsive genesin sugarcane leaves (Patade et al. 2011c). Efficient
sequestration of Na? to vacuole, which reduces the cyto-
solic Na? concentration, is an important aspect of tissuetolerance to salinity. The authors reported down regulation
of a sugarcane homologue of NHX belonging to the family
of Na?/H? and K?/H? antiporters in response to the saltstress. Though the NHX transcript levels increased tran-
siently at 2 h PEG treatment, expression of NHX was
repressed in response to salt or PEG stress subsequently upto 24 h and in sugarcane plants stressed with salt or PEG
for 15 days and correlated to growth inhibition (Patade
2009). The transcript levels of P5CS, an important gene of
the proline biosynthesis pathway, did not alter significantly
upon exposure to salt stress, but were severely reduced inresponse to PEG stress. On the other hand, expression of
PDH, which plays a role in proline catabolism, was
inhibited in response to salt stress and induced under PEGstress treatments (Patade et al. 2011c). On long-term
exposure to salt or PEG stress the steady state levels of
both P5CS and PDH gene expression increased (Patade2009), which also correlated to proline accumulation under
these stress conditions (Patade et al. 2011a). The steadystate transcript levels of CAT2 were also lower in non-
primed stressed sugarcane plants in response to iso-osmotic
salt or PEG stress for 15 days however, the expressionlevel was maintained in priming induced stress tolerant
plants under the stressed conditions (Patade et al. 2009).
To understand the molecular basis of salt (NaCl 2%)stress response, Pagariya et al. (2010) carried out cDNA-
RAPD-based gene expression at early growth stage in
tolerant sugarcane variety Co 62175. Among 335 differ-entially expressed transcript-derived fragments (TDFs),
156 up- and 85 down-regulated were sequenced. The 17%
TDFs representing potential transcripts involved in stresstolerance and plant defense in sugarcane were reported.
Further, Pagariya et al. (unpublished) identified 137 salin-
ity tolerant candidate cDNAs from sugarcane, 20% ofwhich were novel sugarcane genes. These unique sequen-
ces that have so far not been reported to be stress related
might provide further understanding on the perception,response and adaptations mechanisms of non model plant
like sugarcane to salinity stress.
In the study of various tissue-specific EST librariessequence data of Indian subtropical sugarcane variety (CoS
767), 25 water-deficit stress-related clusters showed greater
than twofold relative expression during 9 h dehydrationstress (Gupta et al. 2010). Further, recently Prabu et al.
(2010) based on sqRT-PCR analysis showed higher tran-
script expression of WRKY, 22-kDa drought inducedprotein, MIPS and Ornithine-oxo-acid amino transferase at
initial stages of stress induction with a gradual decrease in
advanced stages. Analysis of the expression of these stress-responsive genes in sugarcane plants under water deficit
stress revealed a different transcriptional profile compared
with sucrose accumulation. Prabu et al. (2010) identifieddifferentially expressed transcripts in response to water
deficiency stress in sugarcane cv. Co740 using PCR-based
cDNA suppression subtractive hybridization technique. Ofthe sequenced 158 cDNA clones based on Dot blot, 62%
showed similarity with known functional genes, 12% with
hypothetical proteins of plant origin, while 26% repre-sented new unknown sequences. Annotation of these dif-
ferentially ESTs indicated their possible function in
cellular organization, protein metabolism, signal trans-duction, and transcription.
Sugar Tech (December 2011) 13(4):322–335 325
123

Involvement of Sugarcane miRNA in Abiotic StressResponses
MicroRNAs (miRNAs) are small single stranded, non-
coding, naturally occurring, highly conserved families oftranscripts (18–25 nt in length). Several miRNAs are either
up regulated or down regulated by abiotic stresses, sug-
gesting that they may be involved in stress-responsive geneexpression and stress adaptation (Sunkar and Zhu 2004).
The involvement of miRNAs in abiotic stress has been
studied in plants in response to dehydration or NaCl byusing expression analysis, suggesting stress specific regu-
lation of expression of miRNA (Patade and Suprasanna
2010) in sugarcane. In response to long term (15 days) iso-osmotic (-0.7 MPa) NaCl or PEG stress, no change in
mature transcript level of miR159 over the control was
detected. However, under the short-term (up to 24 h) saltstress, transcript level of the mature miRNA increased to
112% of the control at 16 h treatment. The mature tran-
script level of miR159 was higher under all the PEGinduced osmotic stress treatments as compared to the
control, and it progressively increased with stress exposure
period (1.3 fold at 8 h treatment). This indicated thatexpression of miR159 gene was more responsive to osmotic
stress than ionic stress. The authors studied expression of
one of the predicted target MYB under the same stress(NaCl or PEG) conditions to study the changes in target
gene expression in response to over or under expression ofmiR159. The results on the expression of specific miR159and its targets could be useful in developing appropriate
markers for selection of tolerant cultivars in sugarcane.
In Vitro Culture Systems
In vitro culture methods in sugarcane have had a great
impact both on basic research and applied commercialinterest. These include micropropagation of elite clones,
production of disease-free planting material, generation of
agronomically superior somaclones, screening methods forbiotic and abiotic stress tolerance, and conservation of
novel and useful germplasm. In vitro techniques for the
mass propagation of healthy sugarcane plantlets via directand indirect regeneration pathways are well established and
are critical in numerous ongoing efforts to improve sug-
arcane germplasm through genetic engineering (Snymanet al. 2011). In direct morphogenesis, plants are regener-
ated directly from tissues such as immature leaf roll discs
and also from shoot tip culture, by which sugarcane ispropagated commercially (Hendre et al. 1983). Indirect
morphogenesis involves initial culturing of leaf roll sec-
tions or inflorescences on an auxin-containing medium toproduce an undifferentiated mass of cells, or callus.
Somatic embryogenesis has been useful for propagating
large number of uniform plants in less time, for obtainingvirus resistant plants through somaclonal variation, muta-
genesis and in vitro selection and developing transgenic
plants (Suprasanna and Bapat 2006; Suprasanna et al.2007a). The in vitro conservation methods can be useful in
order to obviate problems of viability during storage for
extended periods, manpower requirements and need forlarge growth facilities. These methods can also facilitate
maintenance of elite lines, transgenic material and mutantstill their field establishment and/or approval. In vitro
preservation of sugarcane germplasm has been explored
using slow growth (Chandran 2010) and cryopreservationtechniques (Gonzales-Arnao and Engelmann 2006).
Direct adventitious plant regeneration in sugarcane has
been achieved from immature inflorescence tissues (Desaiet al. 2004a), immature leaf thin cell layers (Lakshmanan
et al. 2006) and leaf segments (Manickavasagam and
Ganapathi 1998; Gill et al. 2006) and leaf midrib explants(Franklin et al. 2006). Table 1 presents successes achieved
in the area of in vitro plant regeneration from Indian sug-
arcane cultivars and Saccharum species. The direct somaticembryogenesis (DSEM) system is useful for cost-effective,
large-scale clonal propagation besides providing a
new target explant source for genetic transformation(Suprasanna et al. 2007a). For an efficient application, it is
also essential to ensure the rapid development of embryos
from cultured explants with subsequent regeneration intocomplete plants. The plants derived through DSEM have
been found to be uniform in growth pattern with more
vigour compared to plants derived through indirect somaticembryogenesis pathway and exhibited no variation at the
molecular level (Suprasanna et al. 2006b). This method
also yielded a large number of plants (7–8 per explant) in ashort span of 7 weeks. Assuming an average of 24 seg-
ments per inflorescence, the total number of plants that can
be generated could be around 185–200 (Desai et al. 2004a).Subsequently this technique of direct somatic embryogen-
esis has been extended to several other Indian sugarcane
cultivars suggesting wide adaptability.Studies have been made to refine and improve frequency
of callus induction, proliferation and plant regeneration by
using different media, growth regulators and other addi-tives (Snyman et al. 2011). Alternative strategies such as
partial desiccation (Desai et al. 2004b), silver nitrate and
copper sulphate have also been attempted for improvingregeneration response (Patel et al. 2007). Cefotaxime has
been found to be beneficial for improving frequency of
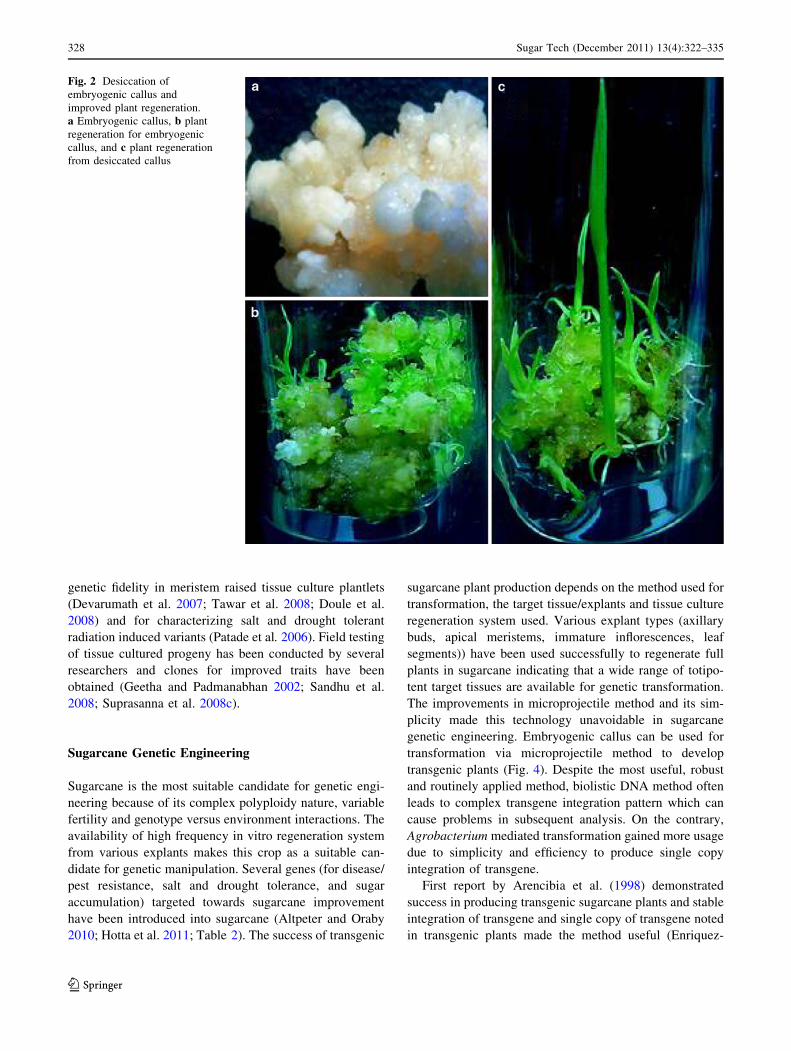
shoot multiplication and elongation (Kaur et al. 2008)Partial desiccation of the immature inflorescence derived-
embryogenic callus was found to enhance plant regenera-
tions frequency, faster regeneration response and vigorousgrowth of the plants (Fig. 2). Sugarcane propagation
326 Sugar Tech (December 2011) 13(4):322–335
123

through mature sugarcane stems is expensive as it incurslabor costs and biologically the method has problems of
spreading viral, bacterial and fungal diseases. Synthetic
seed technology is emerging as a novel tool in plant biol-ogy (Suprasanna et al. 2006a) and synthetic seeds are
potential delivery systems and provide an alternative to
current high-cost vegetative propagation and for storage ofnovel and important germplasm. In sugarcane, there have
been very few studies demonstrating the synthetic seed
research. The somatic embryos derived through directsomatic embryogenesis of sugarcane (Desai et al. 2004a)
were encapsulated in sodium alginate and the beads
showed maximum percentage of germination (73%) onhalf strength MS media (Fig. 3). Synchronous somatic
embryo production combined with good plant regeneration
can be useful for synthetic seed technology in sugarcane.
Broadening Genetic Variability Through In VitroCulture and Mutagenesis
Somaclonal variation has emerged as an important in vitroculture tool for crop improvement. This system has been
adopted for improving the quality and production of
sugarcane and somaclones for yield, sugar recovery,disease resistance, drought tolerance, and maturity have
been isolated. Sugarcane was amongst the first plants
where somaclonal variation was reported (Heinz and Mee1969; Larkin and Scowcroft 1981). Physical and chemical
mutagens have been applied to in vitro cultures so as to
enhance the frequency of genetic variation and obtainbeneficial modifications in cultivars (Patade and Supra-
sanna 2008; Suprasanna et al. 2010a). Physical (gamma
rays, ion beams) and chemical (ethyl methanesulfonate(EMS), sodium azide and sodium nitrite) mutagens have
been used successfully and their optimum mutagenic
treatments have been devised (Patade et al. 2008; Ali et al.2008; Kenganal et al. 2008; Koch et al. 2010). For
example, the LD50 for gamma radiation was around 20 Gy(Saif-Ur-Rasheed et al. 2001; Patade et al. 2006), while the
ideal concentration and exposure time were reported as
40 mM for 2.5 h and 16 mM for 4 h for EMS (Kenganalet al. 2008 and Koch et al. 2010).
In vitro selection at the cellular level has been successful
in isolation of mutants for desirable traits by imposing invitro selection pressure either by incorporating fungal
pathotoxins or fungal culture filtrates for selecting disease
resistance (Rai et al. 2011) or incorporation of sodiumchloride, polyethylene glycol, mannitol for selecting salt or
drought tolerance (Suprasanna et al. 2008a). In sugarcane,
somaclonal variant lines resistant to eye spot diseasecaused by Helminthosporium sacchari were selected
(Larkin and Scowcroft 1983). Various researchers have
used mutagenesis and selection to isolate embryogeniccells and plants tolerant to the causal agent of red rot (Ali
et al. 2008; Singh et al. 2008; Sengar et al. 2009).
Mutation induction of in vitro cultures will require thatmeristematic cells or tissues and mitotically active cells are
cultured to prepare sufficient material for mutagenic
treatment. Intrasomatic competition discriminating muta-gen affected cells and causing loss of their cell progenies
may be controlled by modifying in vitro conditions
resulting in a better competitiveness of mutant cells. Insugarcane, partial desiccation has been used successfully to
stimulate and enhance somatic embryo differentiation
(Desai et al. 2004b) and enhance regeneration response ofgamma-irradiated embryogenic callus cultures (Suprasanna
et al. 2008b). Partial desiccation treatment can offer as a
simple and novel method in stimulating regenerationresponse of higher dose gamma- irradiated cultures.
Assessment of genetic fidelity among micropropagated
plants is important especially in polyploid plants likesugarcane. Besides morphological analysis, molecular
markers have been used for detecting genetic change in
tissue cultured raised plants (Jain et al. 2005; Lal et al.2008; Suprasanna et al. 2007a, b, 2010b), for detecting
Table 1 Development of in vitro regeneration systems in Indian cultivars of sugarcane
Cultivar(s) Explant Mode of regeneration Reference
Co94012, VSI 434 Shoot tips Direct regeneration Tawar et al. 2008
CoC-671 Shoot tips Direct regeneration Biradar et al. 2009
Co 91010, CoC 671 Inflorescence segments Indirect somatic embryogenesis Suprasanna and Bapat 2006
CoC 671 Immature Inflorescence Direct Somatic embryogenesis Desai et al. 2004a
Co Si 95071 Immature leaf roll Direct Somatic embryogenesis Geetha and Padmanabhan 2001
Co.J. 83, Co.J. 86 Inner leaf whorls Indirect somatic embryogenesis Gill et al. 2004
CoS96268 Young leaf rolls Direct adventitious regeneration Pandey et al. 2011
CoS 96268, CoS 95255 Shoot tips Direct regeneration Ramanand et al. 2007
Co J 64, Co J 83, Co J 86 Young leaf segments Direct regeneration Gill et al. 2006
Saccharum edule Immature Inflorescence Indirect somatic embryogenesis Chandran 2011
Sugar Tech (December 2011) 13(4):322–335 327
123
genetic fidelity in meristem raised tissue culture plantlets
(Devarumath et al. 2007; Tawar et al. 2008; Doule et al.
2008) and for characterizing salt and drought tolerantradiation induced variants (Patade et al. 2006). Field testing
of tissue cultured progeny has been conducted by several
researchers and clones for improved traits have beenobtained (Geetha and Padmanabhan 2002; Sandhu et al.
2008; Suprasanna et al. 2008c).
Sugarcane Genetic Engineering
Sugarcane is the most suitable candidate for genetic engi-
neering because of its complex polyploidy nature, variable
fertility and genotype versus environment interactions. Theavailability of high frequency in vitro regeneration system
from various explants makes this crop as a suitable can-
didate for genetic manipulation. Several genes (for disease/pest resistance, salt and drought tolerance, and sugar
accumulation) targeted towards sugarcane improvement
have been introduced into sugarcane (Altpeter and Oraby2010; Hotta et al. 2011; Table 2). The success of transgenic
sugarcane plant production depends on the method used for
transformation, the target tissue/explants and tissue culture
regeneration system used. Various explant types (axillarybuds, apical meristems, immature inflorescences, leaf
segments)) have been used successfully to regenerate full
plants in sugarcane indicating that a wide range of totipo-tent target tissues are available for genetic transformation.
The improvements in microprojectile method and its sim-
plicity made this technology unavoidable in sugarcanegenetic engineering. Embryogenic callus can be used for
transformation via microprojectile method to develop
transgenic plants (Fig. 4). Despite the most useful, robustand routinely applied method, biolistic DNA method often
leads to complex transgene integration pattern which can
cause problems in subsequent analysis. On the contrary,Agrobacterium mediated transformation gained more usage
due to simplicity and efficiency to produce single copy
integration of transgene.First report by Arencibia et al. (1998) demonstrated
success in producing transgenic sugarcane plants and stable
integration of transgene and single copy of transgene notedin transgenic plants made the method useful (Enriquez-
Fig. 2 Desiccation ofembryogenic callus andimproved plant regeneration.a Embryogenic callus, b plantregeneration for embryogeniccallus, and c plant regenerationfrom desiccated callus
328 Sugar Tech (December 2011) 13(4):322–335
123

Obregon et al. 1998; Manickavasagam et al. 2004).
Transgenic plant production requires selectable marker
genes that enable the selection of transformed cells, tissueand plants. The most routinely practiced are those that
exhibit resistance to antibiotics or herbicides. Since thereare perceived risks in the deployment of transgenic plants
containing these markers, alternate selection systems
referred to as positive selection and marker-free systemshave become useful (Suprasanna and Ganapathi 2010). In
sugarcane, Jain et al. (2007) used mannose for the selection
of embryogenic callus and found that increased mannoseimproved the overall transformation efficiency by reducing
the number of selection escapes.
Since the first report of successful transformation, sig-nificant progress (Fig. 5) has been made towards the
development of transgenic sugarcane endowed with resis-
tance to biotic stresses, particularly diseases and insectpests (Srikanth et al. 2011; Altpeter and Oraby 2010), viral
diseases, metabolic engineering and towards making sug-
arcane as a biofactory. Genes from bacteria such asBacillus thuringiensis (Bt) and Bacillus sphaericus, prote-
ase inhibitors, plant lectins, ribosome inactivating proteins,
secondary plant metabolites, and small RNA viruses havebeen used alone or in combination with conventional host
plant resistance to develop crop cultivars that suffer less
damage from insect pests (Srikanth et al. 2011). Transgenic
sugarcane for borer resistance was also reported using Cry
1Aa3 gene (Kalunke et al. 2009). Recently the efficacy of
native Cry1Aa, Cry1Ab and Cry1Ac against C. infusca-tellus in in vitro bioassays through diet-surface contami-
nation method and observed that the Cry1Ab as the mosttoxic among the three compounds (Arvinth et al. 2010).
Christy et al. (2009) transferred aprotinin genes to sugar-
cane cultivars. The in vivo bioassay studies showed thatlarvae of top borer Scirpophaga excerptalis Walker
(Lepidoptera: Pyralidae) fed on transgenics showed sig-
nificant reduction in weight and impairment of larvaldevelopment. Sucrose content is a highly desirable trait in
sugarcane. Sugarcane cultivars differ in their capacity to
accumulate sucrose and breeding programs routinely per-form crosses to identify genotypes able to produce more
sucrose. In this regard, transgenic approaches to manipu-
late native genes that influence metabolism may havesignificant application.
Most of the field trials of transgenic sugarcane are
related to the transgene expression for agronomic traits andare being undertaken in Brazil and Australia. The agro-
nomic traits like height, diameter and the number of stalks,
fibre content, disease resistance, and yield of transgenicclones were not significantly different from that of
untransformed sugarcane plants. However the field trials
of insect resistant transgenic sugarcane revealed some
Fig. 3 Synthetic seedsdeveloped using directsomatic embryos of sugarcanecv. CoC671. a Sodiumalginate beads with somaticembryos, b magnified view ofthe bead, and c germination ofembryos
Sugar Tech (December 2011) 13(4):322–335 329
123

morphological, physiological and phytopathological vari-
ations (Arencibia et al. 1999). Sugarcane has a long historyof use as human food (as a source of sugar), molasses,
animal feed and for energy purpose. Possible exposure to
the GM sugarcane plants or pollen can be through workingwith the plants while conducting field trials or post harvest
analyses, and/or living near the area where the GM sug-
arcane plants are grown (Anonymous 2011). Commercialsugarcane cultivars rarely flower or produce seed in the
field, and exposure to non-GM sugarcane has not been
associated with any reports of allergic responses in Aus-tralia (Anonymous 2011). The report suggests that some
measures can be imposed to minimize exposure include
harvesting the GM sugarcane plants before flowering orremoving flower heads before anthesis.
Sugarcane (Saccharum hybrids) is an attractive candi-
date for metabolic engineering aimed at sustainable pro-duction of value-added biomaterials and feedstocks,
Table 2 Genetic engineering of sugarcane for different traits (modified from Hotta et al. 2011, and references therein)
Trait Gene Transformation method Reference
Reporter and Selection system
Neopmycin phosphotransferase npt-II Microprojectile Bower and Birch 1992
B-Glucuronidase uid-A Microprojectile Bower and Birch 1992
Hygromycin phosphotransferase hpt Agrobacterium Arencibia et al. 1998
Green fluorescent protein gfp Agrobacterium Elliott et al. 1998
Phosphinothricin acetyl transferase bar Agrobacterium Manickavasagam et al. 2004
Phosphomannose isomerase manA Microprojectile Jain et al. 2007
Herbicide resistance
Bialophos bar Microprojectile Gallo-Meagher and Irvin 1996
Phosphinothricine bar Agrobacterium Enriquez-Obregon et al. 1998
Glufosinate ammonium pat Microprojectile Leibbrandt and Snyman 2003
Disease resistance
SCMV SCMV-CP Microprojectile Joyce et al. 1998
Sugarcane leaf scald albD Microprojectile Zhang et al. 1999
SrMV SrMV-CP Microprojectile Ingelbrecht et al. 1999
Puccinia melanocephala Glucanase, chitanase & ap24 Agrobacterium Enriquez et al. 2000
SCYLV SCYLV-CP Microprojectile Gilbert et al. 2005
Fiji leaf gall FDVS9 ORF 1 Microprojectile McQualter et al. 2004
Pest resistance
Sugarcane stem borer cry1A Microprojectile Arencibia et al. 1999
Sugarcane stem borer cry1Ab Microprojectile Braga et al. 2003
Sugarcane stem borer cry1Ab Microprojectile Arvinth et al. 2010
Sugarcane stem borer cry1Aa3 Agrobacterium Kalunke et al. 2009
Proceras venosatus Modified cry1Ac Microprojectile Weng et al. 2010
Sugarcane canegrub gna Microprojectile Legaspi and Mirkov 2000
Mexican rice borer gna Microprojectile Setamou et al. 2002
Ceratovacuna lanigera gna Agrobacterium Zhangsun et al. 2007
Scirpophaga excerptalis Aprotinin Microprojectile Christy et al. 2009
Metabolic engineering/alternative products
Sucrose accumulation Antisense soluble acid invertase Microprojectile Ma et al. 2004
Fructo oligosaccharide IsdA Agrobacterium Enriquez et al. 2000
Polyphenol oxidase ppo Microprojectile Vickers et al. 2005
Polyhydroxybutyrate phaA, phaB, phaC Microprojectile Brumbley et al. 2007
p-Hydroxybenzoic acid hch1 and cp1 Microprojectile McQualter et al. 2004
Mannose manA Microprojectile Jain et al. 2007
Isomaltulose SI Microprojectile Wu and Birch 2007
Proline overproduction P5CS Microprojectile Molinari et al. 2008
330 Sugar Tech (December 2011) 13(4):322–335
123

particularly those derived from sucrose (a-D-glucopyrano-
syl-1,2-D-fructofuranose), the major storage product in
sugarcane (Birch 2007). Successful example of
establishing biorefineries at sugar mills to produce biofueland bioplastics and, engineering sugarcane to make new
bioplastics and alternative sugars could demonstrate that
this crop has the potential to contribute to BioEconomy(Brumbley et al. 2007).
Concluding Remarks
Sugarcane is a source of food and fuel, and biotechnologycan contribute to substantially increase the utility of this
crop. The successful application of biotechnological tools
will require reliable, high levels of transgene expressionand their stability over generations. The availability of
cellular and molecular toolbox has opened up a plethora of
prospects. Innovative in vitro culture systems have becomeavailable with potential for rapid propagation and gener-
ating novel germplasm with desirable traits. A greater
understanding of the crop using functional genomics andcellular methods will accelerate understanding responses to
biotic and abiotic stresses and their management. Profiling
of gene expression under conditions that affect crop yield
Fig. 4 Steps in genetic transformation using embryogenic callus in sugarcane. a Gus expression, b selection of callus on hygromycin medium,c regeneration on selection medium, d regeneration on selection medium, and e selection and regeneration
Fig. 5 Chronological events in the developments of GM sugarcane
Sugar Tech (December 2011) 13(4):322–335 331
123

can aid in building up an ‘expression panel’ for sugarcane
cultivars which should become invaluable in target geneselection. Gene silencing is being used in transgenic
research aimed at down-regulation of endogenous genes in
sugarcane. Some of the important challenges include genediscovery, transgenics and controlled transgene expression,
sucrose metabolism and photosynthesis. The advances in
sugarcane biotechnology could become remarkable in thecoming years, both in terms of improving productivity as
well as substantially increasing the value and utility of thiscrop.
References
Akhtar, S., A. Wahid, and E. Rasul. 2003. Emergence, growth andnutrient composition of sugarcane sprouts under NaCl salinity.Biology of Plants 46: 113–117.
Ali, A., S. Naz, F.A. Siddiqui, and J. Iqbal. 2008. An efficientprotocol for large scale production of sugarcane throughmicropropagation. Pakistan Journal of Botany 40(1): 139–149.
Altpeter, F., and H. Oraby. 2010. Sugarcane. In Genetic modificationof plants. Biotechnology in agriculture and forestry, vol. 64, ed.F. Kempken, and C. Jung, 453–472. New York: Springer.
Alwala, S., A. Suman, J.A. Arro, J.C. Veremis, and C.A. Kimbeng.2006. Target region amplification polymorphism (TRAP) forassessing genetic diversity in sugarcane germplasm collections.Crop Science 46: 448–455.
Anonymous. 2011. Biology of sugarcane. Report by the Office of theGene Technology Regulator. www.ogtr.gov.au/internet/ogtr/publishing…/sugarcane…/biologysugarcane. Accessed 29 Sep2011.
Arencibia, A., E. Carmona, M.T. Cornide, S. Castiglione, J. O’Relly,A. Cinea, P. Oramas, and F. Sala. 1999. Somaclonal variation ininsect resistant transgenic sugarcane (Saccharum hybrid) plantsproduced by cell electroporation. Transgenic Research 8:349–360.
Arencibia, A., E. Carmona, P. Tellez, M.T. Chan, S.M. Yu, L.Trujillo, and P. Oramas. 1998. An efficient protocol forsugarcane (Saccharum spp.) transformation mediated by Agro-bacterium tumefaciens. Transgenic Research 7: 213–222.
Arvinth, S., S. Arun, R.K. Selvakesavan, J. Srikanth, N. Mukunthan,P. Ananda Kumar, M.N. Premachandran, and N. Subramonian.2010. Genetic transformation and pyramiding of aprotinin-expressing sugarcane with cry1Ab for shoot borer (Chiloinfuscatellus) resistance. Plant Cell Reports 29(4): 383–395.
Besse, P., G. Taylor, B. Carrol, N. Berding, D. Burner, and C.L. McIntyre. 1998. Assessment of genetic diversity in a sugarcanegermplasm collection using an automated AFLP analysis.Genetica 104: 143–153.
Biradar, S., D.P. Biradar, V.C. Patil, S.S. Patil, and N.S. Kambar.2009. In vitro plant regeneration using shoot tip culture incommercial cultivar of sugarcane. Karnataka Journal of Agri-cultural Sciences 22(1): 21–24.
Birch, R.G. 2007. Metabolic engineering in sugarcane: Assisting thetransition to a bio-based economy. In: Applications of plantmetabolic engineering, ed. R. Verpoorte, A.W. Alfermann, andT.S. Johnson, 249–281. Berlin: Springer.
Bower, R., and R.G. Birch. 1992. Transgenic sugarcane plants viamicroprojectile bombardment. Plant Journal 2: 409–416.
Braga, D.P.V., E.D.B. Arrigoni, M.C. Silva Filho, and E.C. Ulian.2003. Expression of the Cry1Ab protein in genetically modifiedsugarcane for the control of Diatraea saccharalis (Lepidoptera:Crambidae). Journal of New Seeds 5(2/3): 209–221.
Brumbley, S.M., M.P. Purnell, L.A. Petrasovits, L.K. Nielsen, andP.H. Twine. 2007. Developing the sugarcane biofactory for high-value biomaterials. International Sugar Journal 109: 5–15.
Brumbley, S.M., S.J. Snyman, A. Gnanasambandam, P. Joyce, S.R.Hermann, J.A.G. da Silva, R.B. McQualter, M.-L. Wang, B.Egan, A.H. Patterson, H.H. Albert, and P.H. Moore. 2008.Sugarcane. In A compendium of transgenic crop plants, Vol. 7:sugar tuber and fiber crops, ed. C. Kole, and T.C. Hall, 1–58.Oxford: Blackwell.
Casu, R.E., J.M. Manners, G.D. Bonnett, P.A. Jackson, C.L.McIntyre, R. Dunne, S.C. Chapman, A.L. Rae, and C.P.L. Grof.2005. Genomics approaches for the identification of genesdetermining important traits in sugarcane. Field Crops Research92: 137–147.
Chandran, K. 2010. In vitro multiplication and conservation ofSaccharum germplasm. Indian Journal of Plant GeneticResources 23(1): 65–68.
Chandran, K. 2011. In vitro regeneration of Saccharum edule fromimmature inflorescence. Sugar Tech 13(2): 170–173.
Christy, L.A., S. Aravith, M. Saravanakumar, M. Kanchana, N.Mukunthan, J. Srikanth, G. Thomas, and N. Subramonian. 2009.Engineering sugarcane cultivars with bovine pancreatic trypsininhibitor (aprotinin) gene for protection against top borer(Scirpophaga excerptalis Walker). Plant Cell Reports 28:175–184.
Cordeiro, G.M., Y.B. Pan, and R.J. Henry. 2003. Sugarcanemicrosatellites for the assessment of genetic diversity insugarcane germplasm. Plant Science 165: 181–189.
D’Hont, A., Y.H. Lu, P. Feldmann, and J.C. Glaszmann. 1993.Cytoplasmic diversity in sugar cane revealed by heterologousprobes. Sugar Cane 1: 12–15.
Desai, N.S., P. Suprasanna, and V.A. Bapat. 2004a. A simple andreproducible method for direct somatic embryogenesis fromimmature inflorescence segments of sugarcane. Current Science87(6): 764–768.
Desai, N.S., P. Suprasanna, and V.A. Bapat. 2004b. Partial desicca-tion of embryogenic callus improves plant regeneration fre-quency in Sugarcane (Saccharum spp). Journal of PlantBiotechnology 6: 229–233.
Devarumath, R.M., R.B. Doule, P.G. Kawar, S.B. Naikebawane, andY.S. Nerkar. 2007. Field performance and RAPD analysis toevaluate genetic fidelity of tissue culture raised plants vis-a-visconventional setts derived plants of sugarcane. Sugar Tech 9(1):17–22.
Doule, R.B., P.G. Kawar, Y.S. Nerkar, and R.M. Devarumath. 2008.Field performance of promising somaclonal variants and RAPDanalysis to assess genetic variation in sugarcane (Saccharumspp.). Indian Journal of Genetic and Plant Breeding 68(3):301–306.
Elliott, A.R., J.A. Campbell, R.I.S. Bretell, C.P.L. Grof. 1998.Agrobacterium mediated transformation of sugarcane using GFPas a screenable marker. Australian Journal of Plant Physiology25: 739–743.
Enriquez, G.A., L.A. Trujillo, C. Menndez, R.I. Vazquez, K. Tiel, F.Dafhnis, J. Arrieta, G. Selman, and L. Hernandez. 2000.Sugarcane (Saccharum hybrid) genetic transformation mediatedby Agrobacterium tumefaciens: Production of transgenic plantsexpressing proteins with agronomic and industrial value. Devel-opments in Plant Genetics and Breeding 5: 76–81.
Enriquez-Obregon, G.A., P.R.I. Vazquez, S.D.L. Prieto, A.D.L. Riva-Gustavo, and H.G. Selman. 1998. Herbicide resistant sugarcane
332 Sugar Tech (December 2011) 13(4):322–335
123

(Saccharum officinarum L.) plants by Agrobacterium-mediatedtransformation. Planta 206: 20–27.
Franklin, G., S. Arvinth, C.J. Sheeba, M. Kanchana, and N.Subramonian. 2006. Auxin pretreatment promotes regenerationof sugarcane (Saccharum spp. hybrids) midrib segment explants.Plant Growth Regulation 50: 111–119.
Gallo-Meagher, M., and J.E. Irvine. 1996. Herbicide resistanttransgenic sugarcane plants containing the bar gene. CropScience 36: 1367–1374.
Geetha, S., and D. Padmanabhan. 2001. Effect of hormones on directsomatic embryogenesis in sugarcane. Sugar Tech 3(3): 120–121.
Geetha, S., and D. Padmanabhan. 2002. Evaluation of tissue cultureraised sugarcane for yield and quality. Sugar Tech 4: 179–180.
Gilbert, R.A., M. Gallo-Meagher, J.C. Comstock, J.D. Miller, M. Jain,and A. Abouzid. 2005. Agronomic evaluation of sugarcane linestransformed for resistance to sugarcane mosaic virus strain E.Crop Science 45: 2060–2067.
Gill, N.K., R. Gill, and S.S. Gosal. 2004. Factors enhancing somaticembryogenesis and plant regeneration in sugarcane (Saccharumofficinarum L.). Indian Journal of Biotechnology 3: 119–123.
Gill, R., P.K. Malhotra, and S.S. Gosal. 2006. Direct plant regener-ation from cultured young leaf segments of sugarcane. PlantCell, Tissue and Organ Culture 84: 227–231.
Glaszmann, J.C., Y.H. Lu, and C. Lanaud. 1990. Variation of nuclearribosomal DNA in sugarcane. Journal of Genetics and Breeding44: 191–198.
Gomathi, R., and T.V. Thandapani. 2005. Salt stress in relation tonutrient accumulation and quality of sugarcane genotypes. SugarTech 1: 39–47.
Gonzales-Arnao, M.T., and F. Engelmann. 2006. Cryopreservation ofplant germplasm using the encapsulation-dehydration technique:review and the case study on sugarcane. CryoLetters 27:155–168.
Grivet, L., J.C. Glaszmann, and P. Arruda. 2001. Sequencepolymorphism from EST data in sugarcane: a fine analysis of6-phosphogluconate dehydrogenase genes. Genetics and Molec-ular Biology 24: 161–167.
Gupta, V., S. Raghuvanshi, A. Gupta, N. Saini, A. Gaur, M.S. Khan,R.S. Gupta, J. Singh, S.K. Duttamajumder, S. Srivastava, A.Suman, J.P. Khurana, R. Kapur, and A.K. Tyagi. 2010. Thewater-deficit stress- and red-rot-related genes in sugarcane.Functional & Integrative Genomics 10: 207–214.
Heinz, D.J., and G.W.P. Mee. 1969. Plant differentiation from callustissue of Saccharum species. Crop Science 9: 346–348.
Hendre, R.R., R.S. Iyer, M. Kotwal, S.S. Khuspe, and A.F.Mascarenhas. 1983. Rapid multiplication of sugarcane by tissueculture. Sugarcane 3: 5–8.
Hotta, C.T., C.G. Lembke, D.S. Domingues, E.A. Ochoa, G.M.Q.Cruz, D.M. Melotto-Passarin, T.G. Marconi, M.O. Santos, M.Mollinari, G.R.A. Margarido, A.C. Crivellari, W.D. dos Santos,A.P. de Souza, A.A. Hoshino, H. Carrer, A.P. Souza, A.A.F.Garcia, M.S. Buckeridge, M. Menossi, Van Sluys Marie-Anne,and G.M. Souza. 2011. The biotechnology roadmap for sugar-cane improvement. Tropical Plant Biology 3: 75–87.
Indian Sugar. 2008. Sugar Statistics Vii(11): 57 & 70.Ingelbrecht, I.L., J.E. Irvine, and T.E. Mirkov. 1999. Post transcrip-
tional gene silencing in transgenic sugarcane. Dissection ofhomology-dependent virus resistance in a monocot that has acomplex polyploid genome. Plant Physiology 119: 1187–1197.
Jain, S.M. 2005. Major mutation assisted plant breeding programssupported by FAO/IAEA. Plant Cell, Tissue and Organ Culture82: 113–123.
Jain, M., K. Chengalrayan, A. Abouzid, and M. Gallo. 2007.Prospecting the utility of a PMI/mannose selection system for
the recovery of transgenic sugarcane (Saccharum spp. hybrid)plants. Plant Cell Reports 26: 581–590.
Jain, R., S. Srivastava, J. Singh, and P. Gupta. 2005. Assessment ofgenetic purity of micropropagated plants of sugarcane byisozyme and RAPD analysis. Sugar Tech 7: 15–19.
Joyce, P.A., McQualter, R.B., Handley, J.A., Dale, J.L., Harding,R.M., and Smith, G.R. 1998. Transgenic sugarcane resistant tosugarcane mosaic virus. In Proc. Aust. Soc. Sugarcane Technol.,ed. D.M. Hogarth, Vol. 20, 204–210. Townsville, Australia.
Kalunke, R.M., A.M. Kolge, K.H. Babu, and D.T. Prasad. 2009.Agrobacterium mediated transformation of sugarcane for borerresistance using Cry 1Aa3 gene and one-step regeneration oftransgenic plants. Sugar Tech 11(4): 355–359.
Kaur, A., M. Gill, D. Ruma, and S. Gosal. 2008. Shoot multiplicationand elongation in sugarcane using cefotaxime. Sugar Tech 10(1):60–64.
Kawar, P.G., R.M. Devarumath, and Y.S. Nerkar. 2009. Use of RAPDmarkers for assessment of genetic diversity in sugarcanecultivars. Indian Journal of Biotechnology 8: 67–71.
Kawar, P.G., M.C. Pagariya, G.B. Dixit, and D. Theertha Prasad.2010a. Identification and Isolation of SCGS phytoplasma-specific fragments by riboprofiling and development of specificdiagnostic tool@. Journal of Plant Biochemistry and Biotech-nology 19: 185–194.
Kawar, P.G., M.C. Pagariya, S.R. Patel, G.B. Dixit, and D. TheerthaPrasad. 2010b. An overview of partial genome sequence of firstAsiatic phytoplasma strain (SCGS)-Indian isolate. Asian Journalof Plant Pathology 4(1): 16–19.
Kenganal, M., R.R. Hanchinal, and H.L. Nadaf. 2008. Ethylmethanesulfonate (EMS) induced mutation and selection forsalt tolerance in sugarcane in vitro. Indian Journal of PlantPhysiology 13: 405–410.
Koch, A.C., S. Ramgareeb, S.J. Snyman, M.P. Watt, and R.S.Rutherford. 2010. An in vitro induced mutagenesis protocol forthe production of sugarcane tolerant to imidazolinone herbicides.Proceedings of the International Society for Sugar CaneTechnology 27: 1–5.
Lakshmanan, P., R.J. Geijskes, K.S. Aitken, Christopher L.P. Grof,G.D. Bonnett, and G.R. Smith. 2005. Sugarcane biotechnology:the challenges and opportunities. In Vitro Cellular & Develop-mental Biology—Plant 41: 345–363.
Lakshmanan, P., R.J. Geijskes, L. Wang, A. Elliott, C.P.L. Grof, N.Berding, and G.R. Smith. 2006. Developmental and hormonalregulation of direct shoot organogenesis and somatic embryo-genesis in sugarcane (Saccharum spp. interspecific hybrids) leafculture. Plant Cell Reports 25: 1007–1015.
Lal, M., R. Singh, S. Srivastava, N. Singh, S. Singh, and M. Sharma.2008. RAPD marker based analysis of micropropagated plantletsof sugarcane for early evaluation of genetic fidelity. Sugar Tech10: 99–103.
Larkin, P.J., and W.R. Scowcroft. 1981. Somaclonal variation—anovel source of variability from cell cultures for plant improve-ment. Theoretical and Applied Genetics 60: 197–214.
Larkin, P.J., and W. Scowcroft. 1983. Somaclonal variation andeyespot toxin tolerance in sugarcane. Plant Cell, Tissue andOrgan Culture 2: 111–121.
Legaspi, J.C., and Mirkov, T.E. 2000. Evaluation of transgenicsugarcane against stalk borers. In Proc Int Soc SugCane TechnolSugCane Entomol Workshop, ed. P.G. Allsopp, and W. Suasaard,Vol. 4. 68–71. Khon Kaen, Thailand.
Leibbrandt, N.B., and S.J. Snyman. 2003. Stability of geneexpression and agronomic performance of a transgenic herbi-cide-resistant sugarcane line in South Africa. Crop Science 43:671–678.
Sugar Tech (December 2011) 13(4):322–335 333
123

Lu, Y.H., A. D’Hont, D.I.T. Walker, P.S. Rao, P. Feldmann, and J.C.Glaszmann. 1994. Relationships among ancestral species ofsugarcane revealed with RFLP using single copy maize nuclearprobes. Euphytica 78: 7–18.
Ma, H.M., S. Schulze, S. Lee, M. Yang, E. Mirkov, J. Irvine, P.Moore, and A. Paterson. 2004. An EST survey of the sugarcanetranscriptome. Theoretical and Applied Genetics 108: 851–863.
Manickavasagam, M., and A. Ganapathi. 1998. Direct somaticembryogenesis and plant regeneration from leaf explants ofsugarcane. Indian Journal of Experimental Biology 36: 832–835.
Manickavasagam, M., A. Ganapathi, V.R. Anbazhagan, B. Sudhakar,N. Selvaraj, A. Vasudevan, and S. Kasthurirengan. 2004.Agrobacterium mediated genetic transformation and develop-ment of herbicide resistant sugarcane (Saccharum specieshybrids) using axillary buds. Plant Cell Reports 23: 134–143.
McQualter, R.B., J.L. Dale, R.H. Harding, J.A. McMahon, and G.R.Smith. 2004. Production and evaluation of transgenic sugarcanecontaining a Fiji disease virus (FDV) genome segment S9-derived synthetic resistance gene. Australian Journal of Agri-cultural Research 55: 139–145.
Molinari, H.B.C., C.J. Marur, E. Daros, M.K.P. de Campos, J.F.R.P.de Carvalho, J.C.B. Filho, L.F.P. Pereira, and L.G.E. Vieira.2008. Evaluation of the stress-inducible production of proline intransgenic sugarcane (Saccharum spp.): Osmotic adjustment,chlorophyll fluorescence and oxidative stress. PhysiologiaPlantarum 130(2): 218–229.
Mudge, J., W.R. Anderson, R.L. Kehrer, and D.J. Fairbanks. 1996.A RAPD genetic map of Saccharum officinarum. Crop Science36: 1362–1366.
Nair, N.V., S. Nair, T.V. Sreenivasan, and M. Mohan. 1999. Analysisof genetic diversity and phylogeny in Saccharum and relatedgenera using RAPD markers. Genetic Resources and CropEvolution 46: 73–79.
Nair, N.V., A. Selvi, T.V. Sreenivasan, and K.N. Pushphalatha. 2002.Molecular diversity in Indian sugarcane varieties as revealed byrandomly amplified DNA polymorphisms. Euphytica 127:219–225.
Pagariya, M.C., M. Harikrishnan, P.A. Kulkarni, R.M. Devarumath,and P.G. Kawar. 2010. Physio-biochemical analysis and tran-script profiling of Saccharum officinarum L. submitted to saltstress. Acta Physiologiae Plantarum 33: 1411–1424.
Pandey, R.N., J. Rastogi, M.L. Sharma, and R.K. Singh. 2011.Technologies for cost reduction in sugarcane micropropagation.African Journal of Biotechnology 10(40): 7814–7819.
Patade, V.Y. 2009. Studies on salt stress responses of sugarcane(Saccharum officinarum L.) using physiological and molecularapproaches. PhD Thesis. University of Pune, Pune, Maharashtra,India.
Patade, V.Y., Bhargava, S., and Suprasanna, P. 2011a. Salt anddrought tolerance of sugarcane under iso-osmotic salt and waterstress: Growth, osmolytes accumulation and antioxidant defense.Journal of Plant Interactions. doi:10.1080/17429145.2011.557513.
Patade, V.Y., Bhargava, S., and Suprasanna, P. 2011c. Transcriptexpression profiling of stress responsive genes in response toshort-term salt or PEG Stress in sugarcane leaves. MolecularBiology Reports. doi:10.1007/s11033-011-1100-z.
Patade, V.Y., A.N. Rai, and P. Suprasanna. 2011b. Expressionanalysis of sugarcane shaggy-like kinase (SuSK) gene identifiedthrough cDNA subtractive hybridization in sugarcane (Saccha-rum officinarum L.). Protoplasma 248(3): 613–621.
Patade, V.Y., B. Sujata, and P. Suprasanna. 2009. Haloprimingimparts tolerance in sensitive sugarcane cultivar to salt and PEGinduced drought stress. Agriculture, Ecosystems & Environment134: 24–28.
Patade, V.Y., and P. Suprasanna. 2008. An in vitro radiation inducedmutagenesis-selection system for salinity tolerance in sugarcane.Sugar Tech 11(3): 246–251.
Patade, V.Y., and P. Suprasanna. 2010. Short-term salt and PEGstresses regulate expression of MicroRNA, miR159 in sugarcaneleaves. Journal of Crop Science and Biotechnology 13(3):177–182.
Patade, V.Y., P. Suprasanna, and V.A. Bapat. 2008. Gammairradiation of embryogenic callus cultures and in vitro selectionfor salt tolerance in sugarcane (Saccharum offcinarum L.).Agricultural Sciences in China 7(9): 101–105.
Patade, V.Y., P. Suprasanna, U.G. Kulkarni, and V.A. Bapat. 2006.Molecular profiling using RAPD technique of abiotic stress (saltand drought) tolerant regenerants of sugarcane Cv. Coc-671.Sugar Tech 8: 63–68.
Patel, B.M., Suprasanna, P., Patade, V.Y., and Bapat, V.A. 2007.Simple and novel method of partial desiccation for improvingplant regeneration in gamma-irradiated sugarcane tissue culture.In National Conference on Biotechnology, 2–3 Feb 2007,Mumbai.
Piperidis, G., Rattey, A.R., Taylor, G.O., and Cox, M.C. 2004. DNAmarkers: A tool for identifying sugarcane varieties. In Proc.Aust. Soc. Sugarcane Technol., ed. D.M. Hogarth, vol. 26.Townsville, Australia.
Prabu, G., P.G. Kawar, M.C. Pagariya, and D. Theertha Prasad. 2010.Identification of water deficit stress upregulated genes insugarcane. Plant Molecular Biology Reporter 29(2): 291–304.
Rai, M.K., R.K. Kalia, R. Singh, M.P. Gangola, and A.K. Dhawan.2011. Developing stress tolerant plants through in vitro selec-tion—An overview of the recent progress. Environmental andExperimental Botany 71(1): 89–98.
Ramanand, M., S. Lal, B. Singh, and V.P. Singh. 2007. Optimizationof rooting in micropropagated shoots of sugarcane. Sugar Tech9: 95–97.
Saif-Ur-Rasheed, M., S. Asad, Y. Zafar, and R.A. Waheed. 2001. Useof radiation and in vitro techniques for development of salttolerant mutants in sugarcane and potato. IAEA-TECDOC 1227:51–60.
Sandhu, S., S. Gosal, K. Thind, S. Uppal, B. Sharma, M. Meeta, K.Singh, and G. Cheema. 2008. Field performance of microprop-agated plants and potential of seed cane for stalk yield andquality in sugarcane. Sugar Tech 11: 34–38.
Selvi, A., N.V. Nair, N. Balasundaram, and T. Mohapatra. 2003.Evaluation of maize microsatellite markers for genetic diversityanalysis and fingerprinting in sugarcane. Genome 46(3):394–403.
Selvi, A., Nair, N.V., Mohapatra, T., Kartikprabhu, T., and Sunde-ravelpandian, K. 2006. Identification of molecular markers forred rot resistance in a complex polyploid, sugarcane. InProceedings of Second National Plant Breeding Congress,269–270. March 1–3, Coimbatore: TNAU.
Selvi, A., N.V. Nair, J.L. Noyer, N.K. Singh, N. Balasundaram, K.C.Bansal, K.R. Koundal, and T. Mohapatra. 2005. Genomicconstitution and genetic relationship among the tropical andsub tropical Indian sugarcane cultivars revealed by AFLP. CropScience 45: 1750–1757.
Sengar, A.S., S. Thind, B. Kumar, M. Pallavi, and S.S. Gosal. 2009.In vitro selection at cellular level for red rot resistance insugarcane (Saccharum sp.). Plant Growth Regulation 58(2):201–209.
Setamou, M., J.S. Bernal, J.C. Legaspi, T.E. Mirkov, and B.C.Legaspi. 2002. Evaluation of lectin-expressing transgenic sug-arcane against stalkborers (Lepidoptera: Pyralidae): effects onlife history parameters. Journal of Economic Entomology 95:469–477.
334 Sugar Tech (December 2011) 13(4):322–335
123

Singh, G., S. Sandhu, M. Meeta, K. Singh, R. Gill, and S. Gosal.2008. In vitro induction and characterization of somaclonalvariation for red rot and other agronomic traits in sugarcane.Euphytica 160(1): 35–47.
Snyman, Sandy J., Gwethlyn M. Meyer, Aimee C. Koch, MarzenaBanasiak, and M.Paula Watt. 2011. Applications of in vitroculture systems for commercial sugarcane production andimprovement. In Vitro Cellular & Developmental Biology—Plant 47: 234–249.
Srikanth, J., N. Subramonian, and M.N. Premachandran. 2011.Advances in transgenic research for insect resistance in sugar-cane. Tropical Plant Biology 4: 52–61.
Srivastava, S., and Gupta, P.S. 2006. Low level of genetic diversity insugarcane varieties of India as assessed by RAPD markers. InProc Inter. Symp on ‘‘Technologies to Improve Sugar Produc-tivity in Developing Countries’’, Session 7: Molecular Biology,Biotechnology and Tissue culture in Sugar crops, 574–578.Guilin, P. R. China.
Srivastava, S., and P.S. Gupta. 2008. Inter simple sequence repeatprofile as a genetic marker system in sugar. Sugar Tech 10(1):48–52.
Sunkar, R., and J.K. Zhu. 2004. Novel and stress-regulated microR-NAs and other small RNA from Arabidopsis. The Plant Cell 16:2001–2019.
Suprasanna, P., and Bapat, V.A. 2006. Advances in the developmentof in vitro culture systems and transgenics in sugarcane. In Int.Symp. Technologies to Improve Sugar Productivity in Develop-ing Countries, 629–636. Guilin, China.
Suprasanna, P., N.S. Desai, S. Choudhari Rupali, and V.A. Bapat.2007b. RAPD markers for assessing culture induced variation insomatic embryogenesis-derived plants of sugarcane (SaccharumOfficinarum L.). Sugar Tech 9(4): 284–289.
Suprasanna, P., N.S. Desai, G. Sapna, and V.A. Bapat. 2006b.Monitoring genetic fidelity in plants derived through directsomatic embryogenesis in sugarcane by RAPD analysis. Journalof New Seeds 8(3): 1–9.
Suprasanna, P., and T.R. Ganapathi. 2010. Engineering the plantgenome: prospects of selection systems using non-antibioticmarker genes. Genetically Modified Crops 1(3): 1–9.
Suprasanna, P., Ganapathi, T.R., and Bapat, V.A. 2006a. Syntheticseeds technology, ed. A.S. Basra, 227–267. New York: CRCPress.
Suprasanna, P., Jain, S.M., Ochatt, S.J., Kulkarni, V.M., Predieri, S.2010a. Applications of in vitro techniques in mutation breedingof vegetatively propagated crops. In Plant mutation. ed. Q. Shu,Chap. 28, 369–383. Vienna: IAEA.
Suprasanna, P., B.R. Manjunatha, V.Y. Patade, N.S. Desai, and V.A.Bapat. 2010b. Profiling of culture-induced variation in sugarcaneplants regenerated via direct and indirect somatic embryogenesisby using transposon-mediated polymorphism. Sugar Tech 12(1):26–30.
Suprasanna, P., S. Meenakshi, and V.A. Bapat. 2008a. Integratedapproaches of mutagenesis and in vitro selection for crop
improvement. In Plant tissue culture molecular markers andtheir role in crop productivity, ed. Ashwani Kumar, and N.S.Shekhawat, 73–92. New Delhi: IK International Publishers.
Suprasanna, P., Patade, Y.V., and Bapat, V.A. 2007a. Sugarcanebiotechnology—A perspective on recent developments andemerging opportunities. In Advances in plant biotechnology,ed. G.P. Rao, Zhao Yipeng, V.R. Volodymyr, and S. Bhatnagar,313–342. Hauppauge, NY: Science Publishers.
Suprasanna, P., Patade, V.Y., Vaidya, E.R., and Patil, V.D. 2008c.Radiation induced in vitro mutagenesis, selection for salinitytolerance and characterization in sugarcane. In Induced plantmutations in the genomics era. Proc. International Symposiumon Induced Mutations in Plants, ed. Q.Y. Shu, 159–162. 12–15August 2008, Vienna, IAEA, Austria. Rome: Food and Agri-culture Organization of the United Nations.
Suprasanna, P., C. Rupali, N.S. Desai, and V.A. Bapat. 2008b. Partialdesiccation improves plant regeneration response of gamma-irradiated embryogenic callus in sugarcane (Saccharum Spp.).Plant Cell, Tissue and Organ Culture 92: 101–105.
Tawar, P.N., R.A. Sawant, S.G. Dalvi, A.A. Nikam, P.G. Kawar, andR.M. Devarumath. 2008. An assessment of somaclonal variationin micropropagated plants of sugarcane by RAPD markers.Sugar Tech 10(2): 124–127.
Ulian, E. 2000. Functional genomics for sugar accumulation genediscovery in sugarcane. Brisbane, Australia: Sugarcane Genom-ics Workshop.
Vickers, J.E., C.P.L. Grof, G.D. Bonnett, P.A. Jackson, and T.E.Morgan. 2005. Effects of tissue culture, biolistic transformation,and introduction of PPO and SPS gene constructs a performanceof sugarcane clones in the field. Australian Journal of Agricul-tural Research 56: 57–68.
Virupakshi, S., and G.S. Naik. 2008. ISSR analysis of chloroplast andmitochondrial genome can indicate the diversity in sugarcanegenotypes for red rot resistance. Sugar Tech 10: 65–70.
Weng, L.X., Deng, H.H., Xu, J.L., Li, Q., Zhang, Y.Q., Jiang, Z.D.,Li, Q.W., Chen, J.W., and Zhang, L.H. 2010. Transgenicsugarcane plants expressing high levels of modified cry1Acprovide effective control against stem borers in field trials.Transgenic Research. doi:10.1007/s11248-010-9456-8.
Wiedenfeld, B. 2008. Effects of irrigation water salinity andelectrostatic water treatment for sugarcane production. Agricul-tural Water Management 95: 85–88.
Wu, L.G., and R.G. Birch. 2007. Doubled sugar content in sugarcaneplants modified to produce a sucrose isomer. Plant Biotechnol-ogy Journal 51: 109–117.
Zhang, L., J. Xu, and R.G. Birch. 1999. Engineered detoxificationconfers resistance against a pathogenic bacterium. NatureBiotechnology 17: 1021–1024.
Zhangsun, D.T., S.L. Luo, R.K. Chen, and K.X. Tang. 2007.Improved Agrobacterium-mediated genetic transformation ofGNA transgenic sugarcane. Biologia 62(4): 386–393.
Sugar Tech (December 2011) 13(4):322–335 335
123


















