
Biosynthesis of the Peptidoglycan of Bacterial Cell Walls · Biosynthesis of the Peptidoglycan of...
12
THE ~JOURIAL OF Bmmo~ca~ CHE>IISTRY Vol. 2.13, No. 11, Issue of June 10, pp. 3169-3li9, 1968 Pnnbd in U.S.A. Biosynthesis of the Peptidoglycan of Bacterial Cell Walls XII. ISHIBITION OF CROSS-LIxKI-\TG RY PENCILLII\‘S AND CEPHALOSPORISS : STUDIES Ix STAPH- YLOCOCCUS A GRECS IN VIVO* (Received for publication, I)ec:ernber 26, 19G7) DONALD J. TIPPER ASU JACK L. STROMINGER b’lwn the Department of Pharmacology, University of Wisconsin Medical School, Madison, Wisconsin 53706 SUMMARY An uncross-linked monomer (nascent peptidogly’can unit) accumulates in cells of Staphylococcus aureus treated with low concentrations of penicillin G or with ampicillin, methi- cillin, or cephalothin. The uncross-linked monomer has been isolated, and analyses indicate that it represents a prefabricated subunit of the wall bearing both of the o-alanine residues of its pentapeptide precursor as well as an open pentaglycine chain. Pulse labeling experiments indicate that this uncross-linked unit is a direct precursor of the cross- linked peptidoglycan and that its cross-linking is inhibited by penicillin G. At high concentrations of penicillin G, wall synthesis ceases abruptly and no accumulation of the nascent peptidoglycan units is observed. These data have been obtained in support of the hypothesis that penicillins are substrate analogues of the D-alanyl-D-alanine end of the nascent peptidoglycan units and that they acylate the trans- peptidase which catalyzes the cross-linking reaction. The data may also provide an explanation of the paradoxical ob- servation that the killing rate in S. aureus by penicillin G is higher at low concentrations than at high concentrations of the antibiotic. In the presence of low concentrations a weakened wall may be formed, thus rendering the organisms more susceptible to lysis than at high concentrations of penicillin where wall synthesis abruptly ceases. Investigation of the mechanism by which penicillins kill sensit,ive organisms started with the discovery of penicillin in 1929, and culminated in 1957 with the theory that penicillin specifically inhibits the biosynthesis of the cell walls of sensitive organisms (24). This hypothesis was confirmed in several laboratories by direct isotopic measurements of the effects of several penicillins on cell wall synthesis in both gram-positive * This study was supported by Research Grant AI-06247 from the United States Public Health Service and by Grant CR4552 from the Sational Science Foundation. The preceding paper in this series is Reference 1. and gram-negative organisms (see Reference 5 and other refer- ences cited therein). However, although much progress has recently been made in elucidating the complex pathways by which uridine disphosphate acetylmuramyl-pentapeptide and uridine diphosphate X-acetylglucosamine are used for pptido- glycan syntheses by cell-free extracts of penicillin-sensitive, gram- positive organisms (Staphylococcus aureus and Alicrococcus Z~sotikticus) (see References 6 to 10 and the preceding papers in this series), none of the reactions involved was found to be sensitive to penicillin, even at concentrations several orders of magnitude greater than minimum growth inhibitory concentra- tions. The product of these biosynthetic reactions in t&o was a linear peptidoglycan in which uncross-linked peptides, retaining both of t.heir COOH-terminal D-alanine residues, were attached t.o the acetylmuramyl residues of the glycan. In S. aureus the peptide moiety formed in the biosynthetic sequence was a pentapeptidc (L-Ala-D-Glu--GLys-D-Ala--D-Ala) substituted by a pentaglycine chain on the e-amino group of lysine. However, concurrent studies of cell wall structure had led to the conclusion that the final product of the biosynthetic sequence in t&o was a highly cross-linked material rather than a linear polymer. The solubilizcd glycopeptide, obt.ained after treat- ment of cell walls of S. aureus with an acetylmuramidase, contained only 1.1 n-alanine residues per glutamic acid residue and was a high molecular weight polypeptide substituted by a disaccharide derived from the glycan (11-13). Subsequently, degradation of the peptide with peptidases and chemical synthesis showed t,hat the repeating unit of the polypeptide was r,-tlla-D- Glu-L-I,ys-D-Ala, i.e. a tetrapeptide lacking one of the D-alaninc residues of its pentapeptide precursor, and that in S. aureus the pentaglycine chain was substituted at its carboxyl end on the e-amino group of lysine (13-X). The attachment of t.he amino end of the pentaglycine chain to the carboxyl group of the terminal D-alanine residue of the tetrapeptide was shown in two ways. In the first place, several bacteriolytic enzymes were found which specifically catalyzed the cleavage of the n-alanyl- glycine linkage in S. uureus or of the linkage of u-alanine to ot.her bridge amino acids in other microorganisms, thus showing directly that the interpeptide bridges were attached to the D- alanine residue of the tetrapeptide (16-18, 14). The other possible point of attachment was the cY-carboxyl group of D- 3169 by guest on September 5, 2018 http://www.jbc.org/ Downloaded from
Transcript of Biosynthesis of the Peptidoglycan of Bacterial Cell Walls · Biosynthesis of the Peptidoglycan of...

THE ~JOURIAL OF Bmmo~ca~ CHE>IISTRY
Vol. 2.13, No. 11, Issue of June 10, pp. 3169-3li9, 1968
Pnnbd in U.S.A.
Biosynthesis of the Peptidoglycan of Bacterial Cell Walls
XII. ISHIBITION OF CROSS-LIxKI-\TG RY PENCILLII\‘S AND CEPHALOSPORISS : STUDIES Ix STAPH- YLOCOCCUS A GRECS IN VIVO*
(Received for publication, I)ec:ernber 26, 19G7)
DONALD J. TIPPER ASU JACK L. STROMINGER
b’lwn the Department of Pharmacology, University of Wisconsin Medical School, Madison, Wisconsin 53706
SUMMARY
An uncross-linked monomer (nascent peptidogly’can unit) accumulates in cells of Staphylococcus aureus treated with low concentrations of penicillin G or with ampicillin, methi- cillin, or cephalothin. The uncross-linked monomer has been isolated, and analyses indicate that it represents a prefabricated subunit of the wall bearing both of the o-alanine residues of its pentapeptide precursor as well as an open pentaglycine chain. Pulse labeling experiments indicate that this uncross-linked unit is a direct precursor of the cross- linked peptidoglycan and that its cross-linking is inhibited by penicillin G. At high concentrations of penicillin G, wall synthesis ceases abruptly and no accumulation of the nascent peptidoglycan units is observed. These data have been obtained in support of the hypothesis that penicillins are substrate analogues of the D-alanyl-D-alanine end of the nascent peptidoglycan units and that they acylate the trans- peptidase which catalyzes the cross-linking reaction. The data may also provide an explanation of the paradoxical ob- servation that the killing rate in S. aureus by penicillin G is higher at low concentrations than at high concentrations of the antibiotic. In the presence of low concentrations a weakened wall may be formed, thus rendering the organisms more susceptible to lysis than at high concentrations of penicillin where wall synthesis abruptly ceases.
Investigation of the mechanism by which penicillins kill sensit,ive organisms started with the discovery of penicillin in 1929, and culminated in 1957 with the theory that penicillin specifically inhibits the biosynthesis of the cell walls of sensitive organisms (24). This hypothesis was confirmed in several laboratories by direct isotopic measurements of the effects of several penicillins on cell wall synthesis in both gram-positive
* This study was supported by Research Grant AI-06247 from the United States Public Health Service and by Grant CR4552 from the Sational Science Foundation. The preceding paper in this series is Reference 1.
and gram-negative organisms (see Reference 5 and other refer- ences cited therein). However, although much progress has recently been made in elucidating the complex pathways by which uridine disphosphate acetylmuramyl-pentapeptide and uridine diphosphate X-acetylglucosamine are used for pptido- glycan syntheses by cell-free extracts of penicillin-sensitive, gram- positive organisms (Staphylococcus aureus and Alicrococcus Z~sotikticus) (see References 6 to 10 and the preceding papers in this series), none of the reactions involved was found to be sensitive to penicillin, even at concentrations several orders of magnitude greater than minimum growth inhibitory concentra- tions. The product of these biosynthetic reactions in t&o was a linear peptidoglycan in which uncross-linked peptides, retaining both of t.heir COOH-terminal D-alanine residues, were attached t.o the acetylmuramyl residues of the glycan. In S. aureus the peptide moiety formed in the biosynthetic sequence was a pentapeptidc (L-Ala-D-Glu--GLys-D-Ala--D-Ala) substituted by a pentaglycine chain on the e-amino group of lysine.
However, concurrent studies of cell wall structure had led to the conclusion that the final product of the biosynthetic sequence in t&o was a highly cross-linked material rather than a linear polymer. The solubilizcd glycopeptide, obt.ained after treat- ment of cell walls of S. aureus with an acetylmuramidase, contained only 1.1 n-alanine residues per glutamic acid residue and was a high molecular weight polypeptide substituted by a disaccharide derived from the glycan (11-13). Subsequently, degradation of the peptide with peptidases and chemical synthesis showed t,hat the repeating unit of the polypeptide was r,-tlla-D- Glu-L-I,ys-D-Ala, i.e. a tetrapeptide lacking one of the D-alaninc residues of its pentapeptide precursor, and that in S. aureus the pentaglycine chain was substituted at its carboxyl end on the e-amino group of lysine (13-X). The attachment of t.he amino end of the pentaglycine chain to the carboxyl group of the terminal D-alanine residue of the tetrapeptide was shown in two ways. In the first place, several bacteriolytic enzymes were found which specifically catalyzed the cleavage of the n-alanyl- glycine linkage in S. uureus or of the linkage of u-alanine to ot.her bridge amino acids in other microorganisms, thus showing directly that the interpeptide bridges were attached to the D-
alanine residue of the tetrapeptide (16-18, 14). The other possible point of attachment was the cY-carboxyl group of D-
3169
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

31iO Riosynthesis of Peptidoglycan oJ Bacterial Cell Walls. XII Vol. 243, So. 11
glutamic acid, but it was shown that this carboxyl group was present as an amide in the cell walls of S. nureus (19, 20). The y-carboxyl group of glutamic acid had previously been shown to he attached IO the a-amino group of lysine in t.hc main peptide chain (21, 22). Thus, the structural studies made it clear that the terminal n-alaninc residue of the pentapeptide present in the biosynthetic precursors of the prptidoglycan must be eliminated in the course of cell wall synthesis and that the point of elimina- tion was also the point. of attachment of the interpeptide bridge. Therefore, it seemed likely that this terminal reaction in cell wall synthesis was a transpeptidation.
The hypothesis t,hat penicillin must be an inhibitor of the terminal cross-linking reaction in cell wall synthesis was adduced at about the same time in several different laboratories. The rod-shaped cells of Prokus mirabilis are reversibly converted in the presence of penicillin to spherical forms which retain all of the constituents of the cell wall and continue to grow in the form of spheres. Martin suggested that the transformation from the rod to the spherical form could be due to loss of rigidity as a consequence of the absence of cross-linking in the spherical form (23), and carried out some experiments with lysozymc lysates of cell walls of i.he rod and sphere forms which supported this hypothesis (24). Wise and Park (25) carried out double labeling experimrnt.s with cells of S. uureus which indicated that the amount of alanine incorporated into the wall in t.he presence of penicillin was in excess of t.hat. incorporated in its absencr, and they therefore reasoned that penicillin must bc blocking the terminal cross-linking reaction in which n-alanine was eliminated. Tipper and Strominger (19), utilizing techniques which had been developed in the course of structural studies of the cell wall, showed that an uncross-linked monomeric unit of t.he wall, present in small amount. in control culturr~, increased greatly in tho presence of penicillin. This uncross-linked unit containrd a pentapeptide unit with both n-alanine residues of t.he precursor and bore an oprn pentaglycine chain substituted on the e-amino group of lysine.
Moreover, it was suggested that penicillin was an analogue of the D-alanyl-n-alanine end of the pentaprptide in the uncross- linked precursor of the cell wall (19). Penicillin is a cyclic dipeptide of L-cysteine and n-valinc, but molecular models showed a striking resemblance between the antibiotic and the dipeptide, n-alanyl-nalanine. The CO- N bond in the @-lactam ring of penicillin lirs in the same position as the peptide bond involved in the t.ranspeptitlation, and it was therefore proposed that penicillin, acting as a subst,rate analogue of the normal transpoptidation >ubst.rate, acylat.cd the transpeptidase and thereby irreversibly inactivated it.. This view was compati- hle with much earlier studies which indicated that bacterial cells irreversibly bound penicillin, that. the binding might be related to the inhibition of growth by pcuicillin, and that. t.he binding component was a lipoprotein located near the surface of the bacterial cell (2(X%).
The present paper is an amplification of preliminary reports (19, 29) of the occurrrnce of the uncross-linked unit in the cell wall and of its accumulation induced hy penicillin. Further- more, it is shown t.hat t.he uncross-linked unit is a direct precursor of t.he final cross-linked pcptidoglycan of the cell wall.
MATI:RIAIS AND METHODS
Organism-S. aureus, strain Copenhagen, was grown at, 37’ in media consisting of 0.5yc Bactopeptone (Difco Laboratories,
Detroit,), 0.5% yeast extract (Difco), 0.2% glucose, and O.lo/, K2HPO4. Growth was followed by removal of aliquots (0.2 ml), dilution with water (0.8 ml), and measurement of absorbance at 706 rnp in a Zeiss PMQ II spectrophotometer. At. maximum growth ~1;~~ reached about 1.6. X11 experiments were performed in t.he exponential phase of growth at 11;~~ between 0.1 and 0.4. Sterile medium (100 ml) was inoculated from an agar slant of the organism and grown for I6 hours at 37”. Portions (20-m]) of the stationary culture served as inocula for fresh medium (1000 ml). Cultures were incubated at 37” in Gyrotory incu- bators (New Brunsivick Scientific Company, Sew Brunswick, New Jersey).
Enzymes-Trypsin and pancreatic RNase were purchased from Sigma. The B enzyme from Chalaropsis, an acetylmuram- idase (30, 31), was a gift from Dr. John Hash. Bacillus cereus @-lactamase (penicillinase) was a commercial preparation (Neutropen) from Riker Laboratories, Northridge, California. The Ln enzyme from a flavobacterium species (see below) was a gift from Dr. Keijiro Kate.
,4ntibiotics--Penicillin G (6-(phenylacetamido)penic~illanic acid), ampicillin (6-a-aminophcnylacetamido)penicillanic acid), methicillin (6.(2’,6’-dimethoxybenzamido)penicillanic acid), cephalothin (7-(2.thienylacetamido)cephalosporanic acid), rist- ocetin, and bacitracin were gifts from Eli Lilly and Company, Indianapolis, Bristol Laboratories, Inc., Syracuse, New York, and Abbott Laboratories, North Chicago. Potassium 6- (phenylacet-1-W-amido)penicillanate (15.7 mC per mmole) was obtained from Nuclear-Chicago, Des Plaines, Illinois.
Amino Acids and Pc@ides--Glycine-i4C (uniformly labeled; 106 mC per mmole) was obtained from New England Nuclear. n-Alanyl-n-alanine was obtained from Cycle Chemical Corpo- ration, Los Angeles. uL-dlan31-nL-alarline and glycylglycine was obtained from Sigma.
Preparation of Cell Walls and Soluble Glycopeptide-At t.hc end of incubations, cells were recovered by centrifugation for 5 min at 5,000 x g, washed once in water at 2”, and finally re- suspended in water (30 ml) in the capsule of a Nossal cell dis- integrator. They were shaken for a total of 3 min with 25 ml of glass beads (0.2 mm, Fisher Scient.ific) and separated from the beads by a coarse, sintered glass filter. The cell walls were rc- covered by centrifugation for 20 min at 35,000 x g and washed once with water at 2”. After resuspension in water (5 ml), the walls were heated in a boiling water bath for 15 min to inactivate any autolytic enzymes. The suspension was cooled and then treated with t.rypsin (0.5 mg) and RSase (0.5 me;) in 0.1 M K2HP04 (5.5 ml) for 4 hours at. 37”. After thorough washing with water, the cell Falls were resuspended in 0.01 hr acetate buffer, pH 4.5 (2 ml), and treated with theCWaropsis B enzyme (0.08 mg) for 16 hours at. 37”. The entire lysates were applied to columns (0.8 X 15 cm) of Ecteola-cellulose (Cl- form, Cellex-E, Bio-Rad) previously equilibrated with water. The soluble glycopeptides were eluted wiith water (50 ml), and the glycopep- tide-tcichoic acid complexes wcrc subsequently elutcd with 0.3 br LiCl (50 ml). The Chuluropsis B enzyme is an endoacetyl- muramidase devoid of peptidase activit.y (30, 31). It cat.alyzes hydrolysis of all the acet.ylmuramyl linkages in the cell wall of S. aureus. The glycopeptide therefore consists of t.he intact polypcptide of the peptidoglycan, acylated on its calaninc amino termini by the carboxyl group of the disaccharide, 4-O-&11’- acetylglucosaminyl-N-acetylmuramic acid (Fig. I). This glyco- pept.ide us.s measured in the esperiments described below by
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of June 10, 1968 D. J. Tipper and J. I,. Straminger 3171
. . .
P /
GlcNAc
MurNAc
MurNAc
MurNAc
MurNAc P
P GlcNAc
- L-I@ P
I GIcNAc
D-+la (<lY& - I--w I
Nil2 a D- a I
Flu. 1. Representation of the struct.ure of the cell wall of S. nureus showiug cross-linked units (above) and naaccut, uncross-linked units (below). The points of cleavage of the glycan by the acetylmuramidase to yield disaccharide unit.s substituted on the peptides arc indicated by the small arrows. ~Mur, muramyl.
determination of its reducing power. Its reduciug equivalent is 1.5 per mole of disaccharide, relative to the reducing power of IV-acetylglucosamine (32). :1nalyses of many of the glycopcp- tide fractions described in this paper have confirmed this value for the reducing power (see Table III, below).
Analytical Procedures--Most of the procedures used have been completely described (33). Total amino acids and amino sugars were also drtermined after hydrolysis for 8 hours in 40 ~1 of 4 .u HCl at 100”. The samples were then diluted directly with 1 ml of buffer and applied to the column of a Beckman-Spinro amino acid analyzer. Lyophilization of hydrolysatcs was avoided since it causes nonreproducible losses of amino sugars, particu- larly of muramic acid. Known amounts of amino sugars were hydrolyzed under the standard conditions for the estimation of losses in the procedure used. l~ound ammonia was also measured on the amino acid analyzer as the increase in free ammonia after hydrolysis for 3 hours in 4 N HCl at lOO”, and by a calorimetric procedure used previously (20). O-Acyl groups were determined by hydroxamate formation as described previously (12).
Radioactivity was determined with a Packard Tri-Carb liquid scintillation spectrometer by one of three procedures. (a) Aliquots (O.l- to 0.5.ml) of aqueous solutions were mixed in counting vials wit.h 10 ml of dioxanc containing 100 g of napht.halene, 5 g of 2,5-diphcnyloxazole (PPO), and 125 mg of I ,4-bis[2(4-methyl-5-phenyloxazolyl)]bcnzene (dimethyl I’O- POP) per liter. (b) Aliquots (O.Ol- to O.l-ml) of aqueous solu- tions were spotted on I-inch squares of Whatman No. 31UvI filter paper, dried, and immersed in 15 ml of toluene containing 4 g of
2,5-diphenyloxazolc and 300 mg of 1,4-bis[2(4-methyl-j- phenyloxazolyl)]benzene per liter. (c) ltadioact.ive zones, detected on thin layer chromatograms by radioautography, were transferred directly to counting vials by a suction device and dispersed in 10 ml of the dioxane solution described above, which in addition contained 50 ml of water per liter.
Gel ZUration-Columns of Sephadex G-50 and G-25, fine grade, bead form (Pharmacia, IJppsala, Sweden), were packed in water and cluted with water at room temperature The void volume (I’,) was determined from the elution volume of blue dextran (Pharmacia), and the sum of void and interior volumes (V, + Vi) was determined from the elution volume of T\‘aCl (detected with 11gp1’03 solution). Kd = (V, - V,)/Vi, and is a measure of the fraction of t.he interior volume available to a solute cluted by a volume (V,) of elucnt.
RESULTS
Accumulation of Uncross-linked Monomer in Presence of Penicillin-horn preliminary experiments it became obvious that accumulation of the uncross-linked monomer could be observed only when growth continued at a normal or near normal rate for some period aft.cr t.he addition of penicillin. This occurred at relatively low penicillin concentrations, under conditions similar to those used previously in which growth did not ceaSe for 1 hour after the addition of penicillin (34; see Fig. Ii). At higher penicillin concentrations growth was inhibited more rapidly and littlc or no accumulation of the uncross-linked monomer was observed.
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3172 Biosynthesis of Peptidoglycan of Bacterial Cell Walls. XZZ Vol. 243, No. 11
Cultures of S. uureus, strain Copenhagen (500 ml), were pre- pared and, when turbidity was 2Oc/, of maximum, the cult.ures were harvested by centrifugation in the cold and resuspended in 500 ml of one-tenthstrength medium (diluted with water) contain- ing penicillin G at 0.086,0.15,0.3, or 1 pg per ml. An additional flask served as a control. After 5 min, each culture received X’-glycinc (6.8 PC). Dilute medium was used t.o minimize dilution of the label while permitting growth under near normal conditions for a short period. The cultures were then quickly warmed to 37” by swirling in a 60” water bath and were reincu- bated at 37” for 30 min. Growth in all cultures was exponential during this period, with no detectable lag. The rat,c of growth
1.5
1.4
1.3
1.2
I.1
1.0
9 0.9 .
5
a6
0.7
2 0.6
4 0.5
g 0.4
8 0.3
0.1
7
2’
,
A
0
control
/
m
B
250,000
3b 4i
ML
FIG. 2. Filtration on a column of Sephadex G-25 of the soluble glycopcptides obtained from a control culture and from a culture treated with 0.086 rg of penicillin G per ml by lysis of cell walls with acetylmuramidase. See text for details. ?vleasurcmcnts of reducing power (A) and of radioactivity incorporated from W- glycine (B) are shown.
was about 75% of that in normal, undiluted medium. Previous experiments had shown that growth in the cultures containing penicillin began t.o deviate from normal at 35 min, and all cultures were therefore harvested at this time.
Cell walls were prepared from each of the cultures. Examina- tion of the products of hydrolysis in 6 N HCI indicated that all of the incorporated radioact.ivity was 14C-glycine. The walls were solubilized with the 13 enzyme from Chalaropsis. The solubilized glycopeptide was then separated from the teichoic acid-glyco- peptide complex as described in “Materials and Methods” on a column of Ectcola-cellulose. The solubilized glycopeptides, in 2 ml of water, were applied separately t.o a column of Sephadex G-25 (95 x 1 cm; V0 = 24 ml; V0 + V; = 50 ml). The column was developed with water at a rate of 0.3 ml per min. Fractions of 1.1 ml were collected and analyzed for total reducing power and for radioactivity. The reducing power determination was a measure of all of the mat.erial present., including both that synthesized before the addition of penicillin and t.hat synthesized after its addition, while the radioactivity measured only the material synthesized after addition of penicillin. The solubilized glycopeptide was separated by the column into three fractions (Fig. 2), which will be referred to as oligomer (Kd = O.O), dimer (K,j = 0.2), and monomer (& = 0.65) (for analyses, see below). ‘The reducing power measurements indicated that addition of penicillin at low concentrations rtqulted in a sharp redistribution of the material present., with a marked decreme in the amount of the oligomer fraction and a marked increase in the amount of the monomer fraction (Fig. 2A). Much of the oligorner fraction was undoubtedly material synthesized during the period before addition of penicillin, because the radioactivity measurements indicated an even sharper decrease in the amount of the oligomer fraction at all penicillin concentrations (Fig. 2B). Iindcr the condit.ions of the experiment the total amount of radioactivity incorporated into all fractions was 507, of the control value at 0.086 pg of penicillin G per ml, and it decrcasd further at higher penicillin concentrations. Comparison of data obtained at the four penicillin concentrations (Table I) indicated that the greatest accumulation of the monomer was observed at the lowest penicillin concentration used and that &$ the penicillin concen- tration was increased the total incorporation was suppressed, with a corresponding decrease in the amount of the monomer
TABLE I
Eflecls of penicillin G, bacilrocin, and vancomycin on synthesis of glycopeplide oligomers
These are the quantitative data obtained from the experiments illustrated in Figs. 2 and 3. Data are expressed as the percentage of reducing power or radioactivity (W-glycine incorporated) in the control fractions. The distribution of (a) reducing power and (b) radioactivity in the control oligomcr, dimer, and monomer fractions was (a) 75%,, 17'yJoo, and S%, and (b) 76%, 1470, and lo’%, respec- tively. -- _.. -. -.
Reducing power Radioactivity incorporated
Antibiotic added .- ___...
Total 1 Oligomer 1 Dimer ’ Monomer Total Oligomer Dimer Monomer -._ _. -.
at/ml % con1roI % conlrol
Penicillin G 0.086....... . . . . 72 0.15.. 57
I ! I
I I
loo 264 50 20 93 212 I 35 ' 81 206 35 10 cil 183
0.3.. -
~fp~&&~n+~~:.~.~: 1-1 :I i 1 % 1
-
i” 1 ‘!j 1 i l 1 4: ‘Zj
I 0 These values are probably artificially high. See the high base-line in Fig. 3.
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of June 10, 1968 L). J. Tipper and J. IL. Strominge?
fraction. However, the proportion of the total radioactivity incorporated into monomer was 50 to 60% in all penicillin- containing cultures, compared t.0 only 10yO in the control incubation. On the other hand, 75yc of the material synthesized in the control incubation was oligomer, while only 20 to 307, w&s oligomer in the penicillin-treated cultures.
Eflects of Bncitracin and Vuncomycin-Similar experiments were carried out with the cells exposed to concent.rations of vancomycin (7.8 I.rg per ml) or bacitracin (78 c(g per ml) near the minimum growth-inhibitory concentrations for these antibiotics. Vancomycin induced a nearly complete cessation of incorporation of isotope, while a less complete diminution was induced by bacitracin (Fig. 3 and Table I). These data are compat.ible with the effects observed during studies of the inhibition of the polymerization reaction in vitro. Examination of the total material present (by reducing power measurements) indicated that both in t.hc presence of vancomycin and in the presence of bacit.racin the relative amount of the monomer had decreased, rather than increaTed as in the presence of penicillin, and that there was 110 decrease in the amount of oligomer (Fig. 3 and Table 1). Thus, although bacitracin and vancomycin inhibited the incorporation of isotope into the monomer fraction as well as into the other fractions, the previously existing monomer fraction was utilized to form a cross-linked peptidoglycan.
These isotopic dat.a arc compatible with enzymatic studies, which have indicated that van&mycin inhibits the transfer of disac!c:haride-pcl,tide units from the lipid intermediate to a cell wall acceptor, while bacitracin inhibits the dephosphorylation of lipid pyrophosphate, the next step in the same reaction cycle
(34-36). h sample of soluble glycopeptide was also treated with the
L1, enzyme prior to filtration on Sephadex G-25 in order to obtain materials for reference. This enzyme catalyzes the hydrolysis of glycyl-glycine and n-alanyl-glycine linkages in the interpeptide bridges of the soluble glycopeptide (16, 17). The reaction was stopped when 65cj, of the bridges had been hy- drolyzed so that all fractions would remain in the lysate. Fil- tration on Sephadex G-26 revealed the expected dist.ribution of fragments, with a marked increase in the amount of the monomer fract.ion and a marked decrease in the amount of the polymer fraction, and provided a standard for the position of the monomer on the Sephadex column (Fig 3).
Pulse Labeling l?xperimenls: Evidence that Uncross-linked Monomer is Precursor of Cross-linked Peptidoglycan-Further experiments of the type described above indicated that, a b(ltter fractionation of the glycopeptide oligomers could be obtained if the filtrations were carried out on a column of Sephadex G-50 operated in series with a column of Sephadex G-25. The pulse labeling experiments described here were carried out with this technique. Early log phase cells (21) were labeled with W- glycine for 30 sec. The suspension was rapidly cooled. The cells were recovered by ccntrifugation and washed. One sample was saved for analysis, a second was incuba.ted for 3 min at 37”, a third was incubated for 20 min, and a fourth was incubated fol 20 min in the presence of 120 pg of penicillin G per tnl. Aft.er this treatment the cell walls were prepared and solubilized with the 1{ enzyme from Chalaropsis. The soluble glycopeptide was separated from t.he t.richoic acid-glycopeptidc complex and then filtered OJI the G-50 and G-25 columns (Fig. 4) Under the conditions used the total amount of cell wall did not increase greatly and t.he distribution of monomer, dimer, trimer, tetramer,
2 0.7
2 5 0.6 w g 0.5
$ 0.4
P u 0.3
2 g 0.2
ln y 0.1
e A ‘
20
!46wO
80 o&25,000
70 T-T C
10
t 20
ML
FIG. 3. Yiltration on a column of Sephudex (i-25 of the soluble glycopeptides obtained from cells treated with bacitracin (i8 rg per ml) or vuncomycin (7.8pg per ml) by lysis of isolated cell walls with ncetylmuramidase. Experiments were carried out in a manner similar to those described for penicillin-treated cultures in Fig. 2 (see text). Lefl (A nnd H), reducing power measure- ments; righl (C), radioactivity measurements. Polymer was eluted between 20 and 25 ml, dimer between 27 and 30 ml, and monomer between 32 and 37 ml. Also indicated is the fractiona- tion on the same column of a soluble glycopeptidc partially hy- drolyzed with the LII enzyme from Pluuobacterium (kindly given by Dr. K. Kato).
and higher oligomcrs (which were separated by the Sephadex columns) did not change greatly (measured by reducing power, Fig. 4).
After 30 SW of labeling with 1%.glycine, most of the radio- activity in the cell wall was found in t.he monomer fraction and small amounts were found in each of the higher oligomers (Fig. 411). After incubation for 3 min the total amount of radio- activity in the cell wall had increased lo-fold by incorporation of “C-glycine from precursors and endogenous pools which were not removed by the washing procedure, and by this time radio- activity WLLS uniformly distributed among the various oligomers (Fig. 4A). r\fter 20 min of incubat.ion the radioactivity had redistributed itself so that now most of the radioactivity was present in the highest oligomers, although small amounts re- mained in the monomer (Fig. 4B). The distribution of radio- activity was now similar to the distribution of reducing power, which represents t.he total previously synthesized glycopeptide. On the other hand, if penicillin G wm present from the beginning of the second incubation, the increase of total glycine incorpora-
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Biosynthesis of Peptidoglycan of Bacterial Cell Walls. XII Vol. 243, No. 11
zQ* l -
L-7
FIG. 4. Filtration on tandem columns of Sephadex G-50 and G-25 of the solnble glycopeptides obtained from cells pulse labeled with W-glycine for 30 set and then incubated for various periods in the presence or absence of penicillin. A culture (21) of S. aureus, strain Copenhagen, at 15% of maxima1 growth was har- vested and resuspended in 100 ml of medium at one-tenth of nor- mnl strength nt 2’. The suspension was rapidly warmed to 37”, “C-glycine (14.4 pC) was added, and 30 set later the culture was rapidly iced and the cells were recovered by centrifugation. After one wash with water at 2” the cultures were resuspended in 100 ml of full strength growth medium. One sample (16 ml) was immedi- ately iced and recovered by centrifugation (zero time control sample). A second sample was rapidly brought to 37” and the cells were recovered after 3 min of incubation. A third sample of 16 ml was mixed with 160 ml of full strength growt.h medium and incubated for 20 min prior to centrifugation, and a fourth sample was similarly treated except that it contained penicillin G (120 pg per ml). The cell walls and soluble glycopeptides were prepared as described in “Mat.erials and Methods.” The various samples were then filtered on a column of Sephadex G-50 (116 X 1.5 cm, V0 = 105 ml, V0 + Vi = 218 ml) connected by a plastic capillary tube directly to a column of Sephadex G-25 (95 X 1 cm, Vo = 25 ml, Vo + Vi = 50 ml). Fractions of 3 ml were collected every 20 min, and aliquots were analyzed for redncing power (52 ~1) and radioactivity (825 ~1). Monomer is the peak with maximal concentration at 215 to 225 ml, dimer at 135 to 195 ml, trimer at. 165 to 175 ml, and tetramer at 155 to 160 ml; oligomer is all ma- terial elated before tetramcr.
TABLE II
Incorporation of rudioaclivily into glycopeplide oligomers ajte~ incubation for various times following SO set of pulse labeling
u&h WY-glycine
These are t.he qunntitative data obtained from the experiment described in the legend to Fig. 4 and in the text. Data arc ex- pressed as the percentage in each fraction of the total rndioac- tivity incorporated into glycopeptide at each time. The total amounts of radioactivity incorporated in each experiment, ex- pressed as percentage of the amount incorporated in the 20.min control, were zero time, 107”; 3 min, MO/,; and 20 min plus peni- cillin G, 59%.
I Time of second incubation
-1 hIonomerl Dimer
I I Trimer Tetramer’ Oligomer
I ,_-.- I min ‘;G Ida1 incorporoled info glycopcptidc
0 40 : 3 3 23
20 ! 11 :I :i : ~ E I ’ ’ ~
44
20, + penicillin G ! A6 / 10 1 ’ _-.._
2 ! 2 / 20
t,ion from the internal pool still occurred but all of the in- corporated glycinr remained in the monomer fraction and little of it was redistributed into the oligorner fract.ion (Fig. 4B). The extremely small amount of radioactivity which entered the oligomer fractions is particularly evident. in quantitative data obtained by pooling and counting the various fractions (Table II). Thus, t.his pulse labeling experiment showed that. radioactivity was initially incorporated as the monomer and that on continued incubation this fraction became incorporated into an oligomer fraction. Moreover, penicillin virtually completely blocked the transformation of monomer into the cross-linked oligomers.
Analyses of Mycopeptide Oligomers---Initially, materials ob- tained from the cslicriments shown in Fig. 2 were analyzed. Comparison of the monomer fractions from the penicillin- treated and control cultures (Table III) revealed that both contained 3 alaninc residues, 2 of which were n-alanine residues, 1 of which was COOH-terminal. The presence of the COOH- terminal D-alanine residue was accompanied in each case by the presence of an NHz-terminal glycinc residue. By contrast, the polymer fraction had 2 rather than 3 alanine residues, only 1 n-alanine residue, and contained dy trace amounts of any COOH-terminal or NHz-terminal groups. All fractions analyzed contained an amide group. The presence of this amide indicates that the introduction of the amide residue must precede bridge closure and that t.his reaction is not inhibited by penicillin. Recent studies have indicated that Iipid-P-P-disaccharide- pentapept~ide is the substrate for the amidation reaction (1).
Electrophoresis of the monomer fractions from the prnicillin- treated and control cultures indicated that each was a single com- pound, positively charged at pH 1.9 (0.3 nr formic acid, mobility 10 cm during 2 hours of electrophor&s at 35 volts per cm on Whatman No. 3MhI filter paper), and was neutral at pH 6.4 in pyridinium-acetate buffer. There was no separat.ion of tlisac- charide and pept.idc at eit.her pH value. Thus, these fractions had the expected charge of the amidated monomer. Moreover, both monomers were separated into two romponents on paper chromatography in isobutyric acid-l s SII,OH (5:3) wit.h mobilities of 0.54 and 0.67. Similarly, the monomer fragment obtained by hydrolysis with the Lii enzyme (Fig. 3) was sep-
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of June 10, 1968 D. J. l’ipper and J. L. Xtrominge? 3175
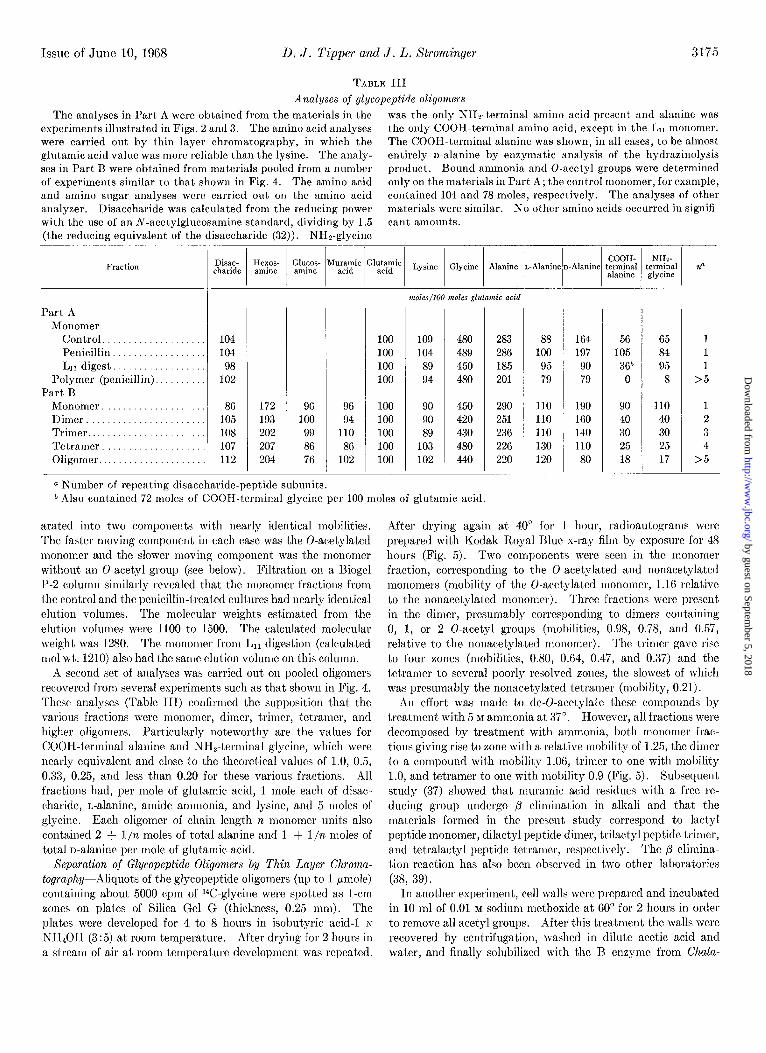
TABLE III
Analyses of glycopeptide oligomers
The analyses in Part A were obtained from the materials in the was the only NHz-terminal amino acid present and alanine was
experiments illustrated in Figs. 2 and 3. The amino acid analyses the only COOH-terminal amino acid, except in the Ln monomer. were carried out by thin layer chromatography, in which the The COOH-terminal alanine was shown, in all cases, to be almost glutamic acid value was more reliable than the lysine. The analy- entirely n-alanine by enzymatic analysis of the hydrazinolysis
ses in Part B were obtained from materials pooled from a number product. Bound ammonia and 0-acetyl groups were determined of experiments similar to that shown in Fig. 4. The amino acid only on the materials in Part A; the control monomer, for example,
and amino sugar analyses were carried out on the amino acid contained 104 and 78 moles, respectively. The analyses of other
analyzer. Disaccharide was calculated from the reducing power materials were similar. No other amino acids occurred in signifi- with the use of an A-acetylglucosamine standard, dividing by 1.5 cant amounts. (the reducing equivalent of the disaccharide (32)). NH*-glgcine
Fraction
Part A Monomer
Control.
Penicillin. L,r digest..................
Polymer (penicillin). Part B
Monomer.
Dimer Trimer
Tetramer. .._........... Oligomer
172 96 96 193 100 94 202 99 110 207 86 86 204 76 102
100
100 100 100
100 100
100 100 100
109
104 89 94
90 90
89 103 102
480 283
489 28F 450 185 480 201
450 290 420 251
430 236 480 226 440 220
88 100
95 79
110 110
110 130 120
164 56 65 197 105 84
90 36r’ 95 79 0 8
190 90 110 160 40 40
140 30 30 110 25 25
80 18 17
1 1
1
>5
1 2
3 4
>5
a Number of repeating disaccharide-peptide subunits.
* Also contained 72 moles of COOII-terminal glycine per 100 moles of glutamic acid.
arated into two components with nearly identical mobilities. The faster moving component in each case was the 0-acetylated monomer and the slower moving component was the monomer without an 0-acetyl group (see below). Filtration on a Biogel P-2 column similarly revealed that the monomer fractions from the control and the penicillin-treated cultures had nearly identical elution volumes. The molecular weights estimated from the elution volumes mere 1100 to 1500. The calculated molecular weight was 1280. The monomer from L1i digestion (calculated mol wt. 1210) also had the same elution volume on this column.
A second set of analyses was carried out on pooled oligomers recovered from several experiments such as that shown in Fig. 4. These analyses (Table III) confirmed the supposition that the various fractions were monomer, dimer, trimer, tetramer, and higher oligomers. Particularly noteworthy are the values for COOH-terminal alanine and NH&erminal glycine, which were nearly equivalent and close to the theoretical values of 1.0, 0.5, 0.33, 0.25, and less than 0.20 for these various fractions. All fractions had, per mole of glutamic acid, 1 mole each of disac- charide, L-alanine, amide ammonia, and lysine, and 5 moles of glycine. Each oligomer of chain length n monomer units also contained 2 + l/n moles of total alanine and 1 + l/n moles of total n-alanine per mole of glutamic acid.
Separation of Glycopeptide Oligomers by Thin Layer Chroma-
tography-Aliquots of the glycopeptide oligomers (up to 1 pmole) containing about 5000 cpm of W-glycine were spotted as l-cm zones on plates of Silica Gel G (thickness, 0.25 mm). The plates were developed for 4 to 8 hours in isobutyric acid-l N NH,OH (3:5) at room temperature. After drying for 2 hours in a stream of air at room temperature development was repeated.
After drying again at 40” for 1 hour, radioautograms were prepared with Kodak Royal Blue x-ray film by exposure for 48 hours (Fig. 5). Two components were seen in the monomer fraction, corresponding to the 0-acetylated and nonacetylated monomers (mobility of the 0-acet,ylated monomer, 1.16 relative to the nonacetylated monomer). Three fractions were present in the dimer, presumably corresponding to dimers containing 0, 1, or 2 0-acetyl groups (mobilities, 0.98, 0.78, and 0.57, relative to the nonacetylated monomer). The trimcr gave rise to four zones (mobilities, 0.80, 0.64, 0.47, and 0.37) and the tetramer to several poorly resolved zones, the slowest of which was presumably the nonacet’ylated tetramer (mobility, 0.21).
An effort was made to de-O-acetylate t’hese compounds by treatment with 5 M ammonia at 37”. However, all fractions were decomposed by treatment with ammonia, both monomer frac- tions giving rise to zone with a relative mobility of 1.25, the dimer to a compound with mobility 1.06, trimer to one with mobility 1.0, and tetramer to one with mobility 0.9 (Fig. 5). Subsequent study (37) showed that muramic acid residues with a free re- ducing group undergo /!I elimination in alkali and that the materials formed in the present study correspond to lactyl peptide monomer, dilactyl peptide dimer, trilactyl peptide trimer, and tetralactyl peptide tetramer, respectively. The p elimina- tion reaction has also been observed in two other laboratories (38, 39).
In another experiment, cell walls were prepared and incubated in 10 ml of 0.01 M sodium methoxide at 60” for 2 hours in order to remove all acetyl groups. After this treatment the walls were recovered by centrifugation, washed in dilute acetic acid and water, and finally solubilized with the R enzyme from Chala-
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3176 Biosynthesis of Peptidoglycan of Bacterial Cell Walls. XII Vol. 243, No. 11
Untreated Alkali-treated
I I I I I & & z E b E E is ii5 z E E z % f 0 f I- s
FIG. 5. Separation of glycopeptide oligomers by thin layer chromatography. See the text for details. Lejt, untreated oli- gomers; right, oligomers treated with 5 M ammonia at 37”. A radioautogram of the thin layer plate is shown. It is evident that
ropsis. The oligomer fractions were then separated as described above and subjected to thin layer chromatography. Now each of t,he fractions gave only a single zone, with the relative mobili- ties 1.0 (monomer), 0.62 (dimer),0.34 (trimer),and0.17 (tetramer). The materials eluted from t’he Sephadex column prior to tet)ramer had relative mobilities bet’ween 0.00 and 0.17. Treatment of each of these zones with 5 BI ammonia at 37” for 1 hour gave rise to the lactyl peptides with the mobilities described above.
Eect of Penicillin Concentration on Incorporation of ‘V- Glycine into Glycopeptide Oligonzers and Irreparable Nature of Lesion in Cell Wall Synthesis Induced by Penicillin-With the technique of examining the various oligomers by thin layer chromatography, it was possible to carry out a number of ex- periments relatively easily which would have been exceedingly difficult to carry out wit,h the Sephadex columns. In order to examine t’he effect of different concentrations of penicillins on accumulation of the monomer, cells were incubated with con- centrations of penicillin G between 0.05 and 20 pg per ml during 20 min with 1°Cglycine present in the medium. At the end of this time cell walls were prepared, de-O-acetylated, and solu- bilized with t,he 13 enzyme from Chalaropsis. After separation from teichoic acid-glycopeptide complex, the glycopeptide fragments were then separated by thin layer chromatography. After radioautography, areas corresponding to the various
some byproducts are also formed in the alkali treatment. See Fig. 8 for separation of glycopeptide oligomers obtained after cell walls had been treated wit’h sodium methoxide at 60” to remove 0-acetyl groups prior to lysis with the acetylmuramidase.
oligomers mere scraped into vials for counting in the liquid scintillation spectrometer. This experiment (Fig. 6d) confirmed the earlier indications that accumulation of the monomer occurred only at low penicillin concentrations. At concen- trations above 0.8 pg per ml accumulation decreased, and at 20 /~g per ml there was an actual decrease in the amount of monomer formed rather than an increase.’ The amount of the higher
1 Some of the decrease in incorporation into monomer at high penicillin concentrations could be due to the formation of soluble peptidoglycan products containing a large proportion of monomer (compare Reference 40), which might not be attached to the cell wall and hence would be lost during the preparation of the walls. In recent, experiments, the amount of surh soluble products (ex- pressed as percentage of the total 14C-glycine incorporated into control cell walls) was 7% in control cells and 12 to 139& in cells labeled in the presence of 0.1, 1.0, or 10 rg of penicillin G per ml. This soluble material obtained from penicillin-treated cells was almost completely degraded to monomer by treatment with the Chalaropsis B enzyme. Conversion to monomer of the material obtained from control cells was much less complete. Thus. al- though formation of soluble peptidoglycan was increased by pen- cillin, the amount formed was not large and was not increased by increasing penicillin concentrations. It cannot, therefore, pro- vide an explanation of the much larger decrease in formation of total glycopeptide or of the decreasing increment of monomer ob- served at increasing penicillin G concentrations (Table I and also Fig. 6).
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of June 10, 1968 D. J. Tipper and .I. I,. Strominge~ 3177
oligomel~s formed wa,j dcc*rca~rtl at all Iicnicillin concentration+ used, and at collc’rnt,rat.ions above 0.8 ccg prr ml the formation of oligomc~rs IV&$ practically eliminated.
.I duplicate set of cell* wax washed frrtt of prnicillin md IX’-
B
After subsequent 70 min incubation in the abwnce of %Glycina and penicillins
0
0.05
0.15 0.77 4.0 20.0
Chaln Length
FIG. (i. Incorporation of “C-glycine into glgcopeptide oligomers in the presence of various concentrations of penirillin G, and the effect of subsequent. incubation in the absence of penicillin (:. A culture (2000 ml) of S. aure~s, st,rain Copenhagen, was harvested in exponential growth phase at 20clL of maximum growth, resus- pended in medium diluted to one-tenth strength (1050 ml) at 2”, and quickly warmed to 37”. Aliquots (170 ml) were transferred to 25@ml flasks containing penicillin G at final concentrations of 0.05, 0.15, 0.8, 4, and 20 pg per ml. A control flask contained no penicillin. After 5 min of shaking at 37”, “C-glycine (1.5 &) was added t,o ouch flask. The cells were harvested after a further 20 min and cell walls were prepared and de-0-acetylated as described in the text. The soluble glycopeptides were prepared with the n enzyme from Chalaropsis. The specific radioactivities of the sol- uble glycopcptides relative to the control (IOOGjc) were 86, 80, 57, ‘2.8, and Oc>i: at 0.05, 0.15, 0.77, 4.0, and 20 pg of penicillin G per ml, respectively. Aliquots (5O-~1) of the solutions (1 ml) were sub- jected to t,hin layer chromatography. The zones corresponding t.o the oligomers of the glycopeptides were detected by radio- autography, eluted, and counted. Recovery of total radioactivit) was 100 f 3(;7c. The total radioact,ivity in each fmction is plotted as the percentage of the corresponding control fraction in A. In R the etrect of a second incubation in the absence of penicillin is depicted. At the end of the first incubation, 17 ml of IO-fold con- centrated medium containing 0.2 M glycine were ndded to a dupli- cate of each flask. The final concent.ration of medium was there- fore normal strength, but each flask contained 0.02 M glycine in addition. This amount of glycine resnlt.ed in a dilution of the “C-glycine by at least lOO&fold. After 5 min of further incnba- tion at 37” to allow ntilization of intracellular glgcine-containing intermediates, penicillinasr (0.12 mg, 4000 llnits; an amount suffl- cient to destroy I he highest concentration of penicillin G in 1 min) was added to each flask. Tncuhat ion wns continued for a further 70 min. Growth was followed turbidimet rirallg dllring the 70.min incubation, and was exponential in all cultures. The growth rates of the cldtures which had been exposed to 0.05, 0.15, 0.8,4.0, and 20rg of penicillin G per ml for 30 min were 92, 93, 91, 74, and 415;,, rcspectivoly, of that of the control, prcsnmably reflecting the nnmber of residual viable cells in each case. After the cells were harvested, the solnble glycopcptides were prepared from the cell walls and treated as described nhove.
I= 024 2= 1.07 3= 123
I 2 3 4.4 I2 3 4>4 I2 3 4 >4 Chain Lmgth
FIG. 7. Incorporation of “C-glgcine into glycopeptide oligomers in the presence of various concentrations of ampicillin, methicillin, and ccphalothin. A clllture (3000 ml) of S. aurcus, strain Copen- hagen, was harvested in cxpnllcntial gro\vth phase at 22% of mnxi- mum growth, resuspended in 1500 ml of lo-fold dilllted medillm at ‘Lo, and warmed rapidly t,o 37”. Aliquots (K?O-ml) were transferred to 250-1111 flasks containing methicitlin, ampicillin, or ccphatothin at various concentrations. One flask without antibiotic served as control. After 5 min of shaking, laC-glycine (1.5 PC) was added t.o each flask; cells were harvested ‘20 min later. The soluble glycopeptides were prepared from the de-0-acctylated cell walls as described in the text. Clycopeptides in water (1011 PI) were analyzed for radioactivit.y and reducing power. The specific radioactivities of the soluble glycopeptides relative to the control were 03, 80, and 49% at 1, 4, and 42pg of methic:illin per ml; 86, 85, 47, and 40y0 at. 0.15, 0.5, 2.2, alld 25.2 pg of ampicillin per ml; and 88, 5’2, and 40y0 at 0.‘24, 1.1, and 12.3 rg of cephalothin per ml, respectively. Aliqllots, rontaining the same number of counts per min and therefore of unequal size, were suhjec~ted to thin layer chromatography. Darn are expressed as dhe amount of radioactivity in each fraction relative to the amorlnt in thrcquiv- ulenl control fraction.
glycine after the 20.min incubation and then incubated for 70 min in normal medium. The cell wall oligorners were then prepared and fractionated as described above. During the 70- min incubation the cells began to grow after a short lag, and t.urbidity doubled during this second incubation. Nevertheless, there was little change in the distribution of the lGoligon~ers synthesized during the earlier, 20.min incubation in thr presence of penicillin (Fig. 6B). In particular, the accumulatrd lrC- monomer remained uncross-linked during the second incubation period. Thus, it seemed clear that monomer introduced in large amount. into t.he cell wall in the presence of penicillin was irreversibly uncross-linked and subsequent, incubation of the cells in the absence of penicillin under conditions in which growth occurred did not result in cros-linking of the previously intro- duced monomer units.
EJects oj Other I-‘enicillins on Formation of Cell Il*all Oli- gomers--Similar exlleriments were carried out with methicillin, ampicillin, and cephalothin. With these substances accumula- tion of the monomer was also readily shown, together with a decrease in t.hc formation of higher oligomers at all concentratinns tested (Figs. 7 and 8). Total glycopeptide synthesis was reducrtl to a minimum of NC/, of control levels (see legend to Fig. 7), and no decrease in accumulation of the monomer occurred at the highest. conccntrat.ions of antibiotics c>xamined in these clsperi- men&
Failure of I’henacetyl Dipeptides to Inhibit Growth ojS. aureus- Since penicillin G is presumed to bc acting &L; an analogue of acyl-n-alanyl-rl-alaninc in inhibiting the cros+linking reaction, it
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3178 Biosynthesis of Peptidoglycan OJ” Bacterial Cell Walls. XII Vol. 243, No. 11
seerned possible that phenacetyl dipeptides might also inhibit growth to some extent. Phenarrtylglycylgl~cine, phenacetyl- 1)L-alallyl-nL-alatline, and phenacetyl-n-alanyl-n-alanine were prepared with the u*e of phenacetyl chloride to arylate the corre- qonding dipeptides. The growth of S. ~LUTBUS, strain (‘open- hagrn, was measured in flasks which contained 2 7~1~ concentra- tions of each of the peljtidr derivative>, in a flask containing 0.4 pg of penicillin G per ml (i.e. about 0.001 MM), and in a flask containing both peuicillin G and phenacetyl-n-alanyl-n-alanine. -l-o deviation of growth from the normal rate was observed in the flask containing the phenacetyl dipeptides, and the presence of l)hei~acetyl-1J-alan?-l-rJ-alanine together with penicillin G did not rcversr the inhibition of growth induced by this antibiotic. Similar results have recently been reported for studies in which various l)hellacet!-lglvcyl-amino acid derivatives structurall) related to penicillin G were used (41).
Failure 0s Penicillinase to Catalyze Ilydrolysis of Phenacetyl Uipeptides-It has been suggehtetl (19) that penicillinase may be the transpeptidahe released from its site of membrane attach- ment and differing f’ro~~~ transl~cptidasr in that the prnicilloyl euzyme whicsh is formed can further leact with water so that a hydrolysis of penicillin is the end result. It was therefore of
-4
-3
-2
-I
-0
0 0.15 0.50 2.2 25.2
pg/ml Ampicillin
Fro. 8. Incorporation of ‘4C-glvcine into glycopeptidc oligomers ., ~II t,hr presence of various rollcentratiolls of ampicillin. See the legend to Fig. i for details. The, cell walls were treated with sodillm methoxide at 60” to remove O-ncetyl groups and then lysed with t.hr :tcetylmllramid:lse. A radioalltogram of the sepa- rat ion of t,he glycopeptide oligomers by thin layer chromatography is shown. This figure is included to illllst,rate t,he separat’ions which were the basis of the data presented in Figs. 6 and 7, and is a particularly graphic illust,rai ion of the accumulation of monomer induced hy a penicillin. 0, origin; 1, tetramer; 2, trimer; 3, tlimer; 4, monomer.
interest to examine the possibility that penicillinase rnight also catalyze the hydrolysis of peptides related to the natural cell wall substrate. However, no hydrolysis of phenacctylglycylglycine, phenacetyl-r,L-alanyl-oL-alanine, or phenacetyl-n-alanyl-u-ala- nine was observed during a a-hour incubation with an amount of penicillinase which would catalyze the hydrolysis of 8.4 pmoles of penicillin G in less than 1 min. Similarly, penicillinase did not catalyze the hydrolysis of the terminal r>-alanine residue of UDP-acet~lmuramyl-pentai,el)tidr labeled with 1°Cn-alanyl- X’-n-alanine. No inhibition of the penicillinase-catalyzed hy- drolysis of penicillin G by the ljhenacetyl dipeptides was ob- served.
Failure oj YT-Penicillin G to be Incorporated into Cell IITall I’eptidoglycan-Another possibility which needed to be excluded was that the penicilloyl transpeptidase might transfer the pmicilloyl residue to the amino end of the pentaglycine bridge and thereby prevent cross-linking. The isolation of the uncross- linked monomer containing glycine chains with free amino ends indicated that no considerable penicilloylation could have occurred, and the following experiment established that no 1Jenicilloplation could be detected even when W-penicillin G was used. The culture (1000 ml) was incubated with 14C- penicillin G (103 pg, 0.28 pmole; specific activity, 15.7 /.LC per pmole). The penicillin G wab added during exponential growth, and incubation was continued for 1 hour. The cell walls prepared from this culture contained only 1100 cpm (i.e. about 70 ppmoles of V-penicillin G). The cell walls were solubilized with the 13 enzyme from Chalaropsis and fractionated on a column of Sephadex G-25 as described in the legend to Fig. 2. The uncross-linked monomer (10 pmoles) represented 327; of the total glycopeptidr (31 pmoles) but contained not more than 2% of the radioactivity which had been present in the cell wall (i.e. less than 1 ppmole). The remainder of the radioactivity was associated with the high molecular weight material obtained in the fractionation. The extremely small amount of 14C- penicillin G present may be ascribed to some nonspecific reaction, and the absence of radioactivity in the monomer fraction escludcs the possibility that a significant transfer of penicilloyl residues to l~eptidoglycan occurred in this organism.
DISCUSSIOS
Thus, it is clear that in the prchencr of low concentrations of penicillin G an uncross-linked monomer, which may be referred to as a nascent peptidoglgcan unit, accumulates. Pulse labeling experiments have clearly shown that this uncross-linked unit is a precursor of the final cross-linked product and that penicillin blocks its subsequent cross-linking. The accumulation of the unrross-linked material is also induced by ampicillin, cel)halo- thin, and methicillin. .\t higher concentrations of penicillin C cell wall synthesis is brought to a more abrupt halt and no accumulation of the uncross-linked monomer is observed. It has also been established that the uncross-linked unit, once it is present in the wall to a large extent, cannot subsequently be cross-linked when the penicillin is removed from the culture and growth is allowed to ljroceed again. We suggest the following interpretation of these findings. Sormally, uncross-linked units are introduced into a growing prptidoglycan at sites on the exterior of the membrane, and may be immediately cross-linked (presumably to a peptide on a second newly introduced pep tidoglycan unit at a neighboring site for glyran polymerization) before a second unit is introduced. At low concentrations of
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of June 10, 19fi8 Il. J. Tipper and J. L. Stromingel 3179
penicillin it is possible that cell wall synthesis ‘Wrtters,” that is, cram-linking is partially inhibited and cell wall synthesis con- tinues for a time with the introduction of a certain number of cross-linked as well as uncross-linked units. However, in the process of this abnormal type of cell wall synthesis, the uncross- linked units becon~c spatially removed from the mrmbmne- bound enzymes which catalyze cross-linking and therefore, on re-establishment of normal growth and cell wall synthesis by removal of penicillin, these units can no longer become cross- linked. At high concentrations of penicillin G, when cross- linking is totally inhibited, cell wall synthesis is brought t.o an abrupt stop for reasons which are not clear, no stuttering occurs, and hence little or no accumulation of the uncross-linked units can be observed.
This interpretation may provide an explanation of the zone phenomenon originally described by Eagle (42) and subsequently studied by other investigators (e.g. Reference 43). Eagle observed in S. aureus that the killing rate paradosically was higher at a low penicillin concentration (I pg per ml) than at a high penicillin concentration (100 pg per ml). Lederberg and St. Clair (43) observed t.he same phenomenon with Bscherichiu coli. In plates containing sucrose agar and a gradient of peni- cillin, a zone was observed at a relatively 10~ penicillin concen- tration in which virtually no organism survived either as cells or as protoplasts. At higher penicillin concentrations, however, the colonies of E. coli survived as protoplast colonies. In view of the data presented here these observations could be explained as follows. At low concentrations of penicillin, cell wall synthesis stutters and a weakened wall is synthesized. Under these conditions the organisms are fragile and are more readily killed by various environmental influences. At high penicillin con- centrations, however, cell wall synthesis ceases abruptly. The lxotoplast remains protected by its original wall, which has been only slightly damaged by a small amount of synthesis after the addition of penicillin. It is therefore better able to survive than t.he organism in which a weakened wall has been propagated.
Many facets of the physiology of wall growth remain un- explained. One obvious one is the fact, that cells of P. mirahilis, although converted to spheres by the presence of penicillin, continue to grow and multiply as spheres indefinitely. These cells contain all the normal constituents of the cell wall. How- ever, the units are not cross-linked to each other, thus account.ing for thr transformation of the rigid rod-shaped wall to a less rigid sphere (23,24). On the other hand, the spherical organism, S. uurexs, ceases to grow abruptly in the presence of high concentrations of penicillin when cross-linking is inhibited, and at low concentrations of penicillin it. can continue to grow only for a relatively brief period, during which it synthesizes a partially cross-linked wall. The reason for this striking difference in the growth physiology of these organisms is not apparent.
REFERESCES
SInwEar, G., AND STROWNGER, J. L., .l. Hiol. Chem., 243, 783 (1968).
LEDERRERG, J., hoc. 1Ynt. Acad. Sci. c’. S. A., 42, 574 (1956); J. Bacleriol., 73, 144 (1957).
Pn~w, J. T., AND STIIOJII.I.GER, J. L., Science, 126, 119 (1957). STROYINGEH, J. L., PARK, J. T., .~ND THOYPSON, R. E., J. l?iol.
Chem., 234, 3263 (1959).
5. IIOGERS, II. J., AND JEI..IA~ZE~I(:S, J., Biochem. J., 81, 576 (1961).
6. CI!Arr?xJEx, A. N., AND PARK, J. T., Proc. .Val. Acad. Sci. C. S. A ., 61, 9 (1964).
7. MEADOW, 1’. M., ANDEI~SON, J. S., ANV S~ROMINGER, J. L., Biochem. Riovhus. Res. Commun.. 14, 382 (1964).
8. ASDERSOS, J. ‘S, hIA~l%I-H.4SIII, ar., 11ASI<IN, RI. A., ASD STROMINGER, J. L., Proc. %al. ilcad. hi. u. 8. A., 63, 881 (1965).
9. STRUVE, W. (;., ASD PiEr.liavs, F. C., Hiochem. Biophys. h’es. Commun., 18. 6 (1965).
10. MATSUHASHI, M., I)IE~‘ILICH, C. P., ASD STROMINGER, J. L., I’roc. .Yal. Acad. Sci. I:. S. A., 64, 587 (1965).
11. MANDELWAM, M., AND SmoMINGER, .J. L., Hiochem. Biophys. Xes. Commun., 6, 4G6 (1961).
12. C~HUY~EN, J.-M., AND STROMINGEH, J. L., Biochemistry, 2, 1110 (1963).
13. GHUYSEN, J.-M., TIPPER, I>. J., BIHGE, C. II., AND STROMIN- GER, J. L., Biochr-mistr!/, 4, 2446 (1965).
14. Muloz, E., GHUYSEN, J.-M., LEyH-Bm:ILm, M., PETIT, J.-F., I~EYMANN, H., BRICAS, E., .~NL) LEFIIAIWIXR, P., Biochemis- fry, 6, 3748 (1966).
15. JAWIS, I)., ANV STnohfIh'GErt, J. L., Hiochemisfrv. 6, 2591 (1967).
,,.
IO. K~,ro, K., KOTAXI, S., .\IATSIWARA, T., KnoAur, J., HASHI- MOTO, S., CHIMORI, M., AND KAZAKAWA, I., Biken’s J., 6, 155 (1962).
17. KATO, K., ASD STROYINGER, J. I,., Biochtmisfry, in press. 18. TIPI’ER, I). .J.. S’I‘IIOMINGEI<. J. L.. AND ICNSIGN. J.. Rochemis-
by, 6; 906 (1967). I I
19. TIWXR, 11. J., ASI) STILOHISGER, J. I,., Proc. .\‘a[. Acad. Sci. U. S. A., 64, 1133 (1965).
20. TIPPER, 1). J., KATZ, W., STROMINGEH, J. I~., AND C;IICYSE:S, J.-RI., Biochemistry, 6, Y21 (1967).
21. ITO, E., AND S~ROMINGER, J. L., J. Biol. Chem., a39.210 (1964). 2”. LASZILLOWI, A. b:., BENZ, E., ASD (;nmt.wx, L., J. Amer.
Chum. Sot., 86, 1880 (1964). 23. MARTIN, H. II., J. Gen. Microbial., 36, 441 (1964). 24. MAwIN, II. H., Abslracls Sizlh International Congress of Mo-
chemistry, 1964, ICH 1’01. 32, p. 518, \.I-70. ‘25. WISE, E. M., AND PARK, J. T., Proc. Nnl. Acud. Sci. U. S. A.,
64, 75 (19G5). 26. Conmm, 1’. I>., Bacleriol. Rev., %, 29 (1956). 27. SCHEPARV, S. A., ANV JOHNSON, M. J., J. Bacleriol., 71, 8.1
(1956). 28. DCERKSEN, J. D., Biochim. Biophys. d&a, 8’7, 123 (1964). 29. TIPI~ER, I). J., Fed. Proc., 26, 344 (1966). 30. HASH, J. H., Arch. Hiochem. Biophys., 102, 379 (1963). O’ TIWER. D. J., S’I’ROMINGER, J. L.. ASD (;III:YSEN. J.-M.. .>I.
32.
33.
Science, 146. 781 (1964). TII~PER, L>. J., GIIUYSEN, J.-M., AND STKOMISGER, J. L., Bio-
chemistry, 4, 468 (1965).
34.
GHUYSEN, J.-M., TIPPER, D. J., AND STROMINCIER, .J. L., in S. I’. COLOWICK AND S. 0. KAPLAN (Editors), Me/hods in enzymology, Vol. 8, Academic Press, Sew York, 1967, p. 6&!i.
ANDERSON, .J. S., MA~rsIJIIASIII, M., 11.4SKIN, M. A., ANI) Sl.ROMINGEII, J. I,., Proc. .Val. .4cad. Sci. I’. S. A ., 63, 881 (1965).
35.
36.
MAWUHASIII, M., D~mxIcH, C. P., ANV STanMIrioEa, J. I,., J. Biol. Chem., 244, 3191 (1967).
SIEn.ER.I’, c;., AND SmoMINGEB, J. L., hoc. ,vat. ilcud. Sci. II. S. A., 67, 767 (1967).
37. 38.
39. 40.
41. 4". 43.
TIPPER, I). .J., Biochemistry, 7, 1441 (1968). GIIUYSES, J.-RI., BRICAS, B., ’ LEYH-BOUILLE, RI., LAVIIE, JI.,
ANI) SIIOCKMAN, (;. D., Biochemistry, in press. PERKINS, H. R., Diochem. J., 102, 29C (1967). 1z.41~1, K., ~[ATSI:IIASHI, .\I., ASI) SI’ROHISGER, J. I,., Proc.
Nat. Acad. Sci. II. S. A ., 66, 656 (19GF). ERI.ANGER, B. F., .~SD (GOODE, I,., :\‘alIt?X, 213, 183 (1967). E.4Gm, H., J. Bacleriol., 62, 663 (1951). LEDERRERC;, J., AND ST. CLAIR, .J., J. Bncleriol., 76, 143 (1958).
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Donald J. Tipper and Jack L. StromingerSTUDIES IN STAPHYLOCOCCUS AUREUS IN VIVO
OF CROSS-LINKING BY PENICILLINS AND CEPHALOSPORINS: Biosynthesis of the Peptidoglycan of Bacterial Cell Walls: XII. INHIBITION
1968, 243:3169-3179.J. Biol. Chem.
http://www.jbc.org/content/243/11/3169Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/11/3169.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on September 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















