
biological control of pest populations - Texas A&M AgriLIFE
36
OFFPRINTS FROM INTEGRATED PEST MANAGEMENT SYSTEMS AND COTTON PRODUCTION, IED Edited by Raymond E Fnsbie. Kamal M El-Zik. and L Ted Wilson Copyright O 1989 by John Wiley & Sons. Inc 7 BIOLOGICAL CONTROL OF PEST POPULATIONS I W. L. Sterling ~ Department of Entomology Texas A&M University, College Station, Texas K. M. El - Zik Department of Soil and Crop Sciences Texas A&M University, College Station, Texas L. T. Wilson Department of Entomology University of California, Davis, California Natural Control of Arthropod Pests Current Practices in Cotton Arthropod Management Nonintervention as a Tactic Key Predators Inaction Levels Natural Control of Selected Arthropod Pests Boll Weevil Heliothis spp. Cotton Flea hopp er Lygus Species Pink Bollworm Spider Mites Other Pests Arthropod Control with Microbials 155
Transcript of biological control of pest populations - Texas A&M AgriLIFE

OFFPRINTS FROM INTEGRATED PEST MANAGEMENT SYSTEMS AND COTTON PRODUCTION, IED Edited by Raymond E Fnsbie. Kamal M El-Zik. and L Ted Wilson Copyright O 1989 by John Wiley & Sons. Inc
7 BIOLOGICAL CONTROL OF PEST POPULATIONS
I W . L. Sterling ~
Department of Entomology Texas A&M University, College Station, Texas
K . M . El-Zik
Department of Soil and Crop Sciences Texas A&M University, College Station, Texas
L. T . Wilson
Department of Entomology University of California, Davis, California
Natural Control of Arthropod Pests Current Practices in Cotton Arthropod Management Nonintervention as a Tactic
Key Predators Inaction Levels
Natural Control of Selected Arthropod Pests Boll Weevil Heliothis spp. Cotton Flea hopp er Lygus Species Pink Bollworm Spider Mites Other Pests
Arthropod Control with Microbials 155

156 Biological Control of Pest Populations
Natural Control of Plant Pathogens Concepts and Mechanisms Approaches to Biological Control of Cotton Pathogens Current Status of Biological Control of Plant Pathogens
Natural Control of Weeds Toward Classical Biological Control
Augmentation of Natural Enemies Restoration Ecology Classical Biological Control
Conclusion References
In this chapter biological control is defined as applied natural control. Thus any way that man intervenes to improve the efficacy of the natural enemies of pest species, whether by introductions, conservation, or augmentation, is classified as biological control. Suppression, maintenance, or regulation of organisms by natural enemies or the physical environment in the absence of human intervention to manipulate them constitutes natural control.
The following sections cover the biological control of arthropods, plant pathogens, and weeds. For the arthropod sections, emphasis is placed on naturally occurring predators, with no intent of minimizing the importance of naturally occurring parasites, pathogens, or physical mortality. Although less developed in cotton, we provide evidence recognizing the importance of natural enemies in effecting the regulation of pathogen and weed pests.
NATURAL CONTROL OF ARTHROPOD PESTS
The total complex of natural enemies, including predators, parasites, and pathogens, plays an important role in preventing most potential arthropod pests in most cotton-growing regions of the United States from causing eco- nomic loss. Native natural enemies usually dominate the dynamics of most pests, so that management intervention is rarely needed to prevent economic losses. Exceptions include certain key, induced, or occasional pests that temporarily escape control, or induced pests in agroecosystems continuously perturbed with agrichemicals. The boll weevil (Anthonomus grandis) and the pink bollworm (Pectinophora gossypiella), which have few effective natural enemies in some parts of the United States, may be exceptions in cotton agroecosystems.
Some of the initial evidence supporting the foregoing statement was pro- vided by examples of natural enemy perturbations with insecticides. For example, in the San Joaquin Valley of California, the treatment of Lygus
?

Natural Control of Arthropod Pests 157
with organophosphate insecticides had a severe impact on the natural ene- mies that normally keep Heliothis spp., beet armyworm, and cabbage loop- ers in check (Falcon et al., 1971; van den Bosch and Hagen, 1966). In the absence of natural enemies, populations of these pests build rapidly. During the late 1960s and early 1970s, San Joaquin Valley cotton received as many as 6 to 14 sprays per season, with most treatments directed at bollworm.
This scenario has been repeated in Peru (Smith and van den Bosch, 1967), Western Australia (Sterling, 1984), south Texas (Adkisson, 1971), Egypt, Mexico, and South and Central America (van den Bosch, 1978), and will probably be repeated whenever unilateral insecticidal panaceas are attempted.
In the past, treatments of insecticides for the control of boll weevil and fleahoppers have decimated natural enemies of the bollworm and the tobacco budworm, resulting in tremendous outbreaks in Texas (Adkisson, 1973). Where insecticidal control of the boll weevil or fleahopper is avoided in early or midseason, widespread outbreaks of the bollworm and budworm seldom occur, and they may be relegated to the status of secondary pests.
Current Practices in Cotton Arthropod Management
Native natural enemies and other natural control factors are primarily re- sponsible for the control of cotton insect pests in Texas, in California, and probably in most other cotton-growing regions where they are not decimated by repeated applications of broad-spectrum insecticides. For example, using the most recent data available, 66% or 2.28 million acres of the cotton acreage in Texas received no foliar applications of insecticides in 1983 (Anon., 1985). Cotton on 1.13 million acres received 2.6 foliar applications, and 1.12 million acres received 1 seed application in 1983. The overlap be- tween acres treated with foliar or seed application is not known. Conser- vatively assuming that each application of insecticide has some residual efficiency for 7 days, the foliar-treated acres are protected by insecticides for about 19 days during the growing season. It is assumed that seed treat- ment provides some protection from insect pests for 14 days after emergence. Based on these data and assumptions, we calculate that about 95% of the time, cotton in Texas is growing essentially free of insecticides and is de- pending on the natural control of pests (Table 7.1). Similar statistics are available for California, where an average of between 1 and 2 sprays are applied annually, in addition to 50% of the 0.9 to 1.3 million acres also receiving a prophylactic insecticidal seed treatment. Absence of pests in a cotton field may also be attributed largely to natural control of the pests on their reservoir host plants.
The data provided in Table 7.1 are based on the best estimates of several extension personnel from different areas of Texas. It seems fair to conclude that a majority of cotton in Texas and California is grown using a “low input” system where chemical insecticides are used infrequently or not at all. On

158 Biological Control of Pest Populations
Table 7.1 Comparison of Cotton Growing Season (1983) When the Crop Was Protected by Chemicals (CHEM) as Compared to Protection by Natural Control “T)
Acres Average Cumulative Days Days Percent Treatment ( x lo6) Treatments Acres CHEM NAT NAT
Foliar 1.13 2.6 2.98 19 141 88 Seed 1.12 1 .o 1.12 14 146 91 Untreated 2.28 0.0 2.28 O 160 100
95.2
the other hand, some growers choose a “high input” system and may use 15 or more applications of insecticides during the growing season. Growers that spray frequently for cotton insect pests tend to prescribe to the notion that they are minimizing the risk of insect damage by using insecticides rather than by depending on natural control. Other growers, who use limited amounts or no insecticides, apparently feel that it is to their advantage to depend on natural control. It is likely that exclusive dependence on either insecticides or natural enemies is not the optimal solution to all pest prob- lems. Sometimes too few natural enemies are available to control outbreaks of pests. On the other hand, to use insecticides when they are not needed is also not optimal. Growers who lean toward insecticidal solutions will tend to spray when in doubt, whereas growers who lean toward natural control solutions may tend to refrain from spraying when in doubt. Both groups of growers are probably making erroneous economic and ecological decisions on occasion. What is needed is a means of increasing the confidence of decisions made regardless of the propensity of growers to chose one strategy over the other. Before growers can make decisions with confidence, the efficacy and impact of natural enemies must be highly predictable. To be able to predict their impact, it is essential to know which natural enemies are the most important and how many are needed to control pest populations.
Nonintervention as a Tactic
It is clear from the discussion above that if cotton is not being protected by chemicals during 95% of the growing season, the dominant decision being made by the growers is not to intervene in the affairs of natural enemies or pests in cotton fields. Often, the best decision that a farmer can make is to do nothing about the pests except to let the natural enemies do their job. The key decision is to determine whether there is an economic and envi- ronmental justification for intervening in the affairs of predators, parasites, or pathogens as they go about controlling pests.
Key Predators If we are to take maximum advantage of natural enemy

Natural Control of Arthropod Pests 159
species as control agents, the effective species should be identified and some quantitative measure of their efficiency must be established (Roach et al., 1979). This information should greatly aid in our ability to predict the num- bers of pests. Any predator species or stage of a predator species that pro- vides predictive value for forecasting future prey population trends and is capable of providing irreplaceable mortality leading to prey population con- trol may be considered a key predator (Sterling, 1984). Irreplaceable mor- tality is the fraction of the total generation mortality attributed to one natural enemy species so that pest (or prey) survival would increase significantly if the natural enemy species were removed from the ecosystem. An example of irreplaceable mortality (Fig. 7.1) is provided by the ants, Solenopsis in- victa, a key predator of the boll weevil. When ants were selectively removed with Mirex insecticide, boll weevil numbers increased dramatically during both 1974 and 1978.
Removal of a secondary predator would probably have little noticeable impact on prey abundance. Since.over 600 species of predaceous arthropods inhabit cotton fields (Whitcomb and Bell, 1964), it is intuitive that most do not contribute measurable irreplaceable mortality to any particular prey. Definitive evidence supporting the classification of the key predators in- habiting cotton agroecosystems is largely lacking. However, based on avail- able data, estimates have been provided as to those predators that probably satisfy the criterion of a key predator (Johnson et al., 1986; McDaniel and Sterling, 1979,1982; Ridgway and Lingren, 1972; Sterling et al., 1984; Wilson and Gutierrez, 1980).
The treatment of various predaceous arthropod species as if they were equal in effectiveness cannot be justified any more than expecting various pest species to have an equal impact on cotton yield. Each species and developmental stage has a different prey preference, searching rate, re- sponse to prey density, and feeding rate. Although there is some pragmatic justification for “collapsing natural enemy impact information to a common unit” (Tamaki et al., 1974; Wilson, 1985), the predictive value of such units is limited by the degree of realism in the relative weightings assigned to each predator species and age class (Hartstack and Sterling, 1986, 1988). To sim- plify the complexity of dealing with multiple species, Hartstack et al. (1975) suggested that key predators be identified and then treated as independent units in predictive computer models. We discuss these concepts further later in the chapter.
Inaction Levels The density of natural enemies sufficient to maintain pests below the action level (economically damaging levels) is the definition of an inaction level (Fillman and Sterling, 1985; Sterling, 1984). The estab- lishment of inaction levels for natural enemies is suggested as one of the first steps in optimizing pest management decisions so that the impact of mortality factors that impinge on pests can be considered. Management de- cisions based solely on the abundance of pests can result in unneeded ex-

160 Biological Control of Pest Populations
EFFECT OF ANT REMOVAL 100
80
IR n
n
3 60
w 0 4 3 40 a R
20
0 0 2
/
/
/ 0 c3
0 2 cv
2 N
DAY OR YEAR (1974)
EFFECT OF ANT REMOVAL
-+- ANTS REM
+ W/ANTS
8
1
-+- ANTS REM -9- W/ANTS
140 150 160 170 180 190
DAY OF YEAR (1978)
Figure 7.1 Impact of ants on boll weevil populations. After Jones and Sterling (1979) and Sterling et al. (1984).

Natural Control of Arthropod Pests
L 1 I 1 1 . I 1 I
0.8
0.6
TOTAL MORTALITY
0.2 hzbv 1 PARAS1 T I SM
I I , k2 0 0.1 DESICCATION
- - - -*- 0-•
0.1 E G G INFERTILITY 4 -
161
\u-*-*\ L. . - I . I
- 1 2 3 4 5 6 7 8 9
WEEK Figure 7.2 Impact of four mortality factors on the boll weevil. After Fillman and Sterling (1983).
penses for chemical insecticides and harm to the environment. If sufficient numbers of natural enemies are present to control the pests, decisions to use either insecticides or inundative releases of biotic mortality agents would not be cost-effective.
Natural Control of Selected Arthropod Pests
Boll Weevil Although the boll weevil has many natural enemies (Hinds, 1907; Hunter and Pierce, 1912), the impact of these enemies on the dynamics of the boll weevil is generally unknown. One exception is the impact of red imported fire ants. The inaction level of red imported fire ants needed to limit boll weevil abundance has been determined for conditions in east Texas. A density of 0.4 ant per plant terminal will provide boll weevil control about 90% of the time (Fillman and Sterling, 1985). Expected mortality rates over a range of ant densities provides a continuum of inaction levels for use when the impact of other mortality factors, including desiccation, parasitism, dis- ease, and egg inviability, are available. The evidence indicates that in some locations the red imported fire ant functions as a key predator (Fig. 7.2) of the boll weevil (Fillman and Sterling, 1983).
Methods useful in the assessment of boll weevil mortality occurring in the flower buds have been detailed by Agnew and Sterling (1981), Fillman

162 Biological Control of Pest Populations
COMPARISON OF CONTROL TACTICS 3.5
3
s \ 0 W 2.5 J c z
J
m 6 . 4
a 2
2 1.5
1
CHEMICAL
&9 NATURAL
71 72 73 74 76 77 78 79 80 81 82 AVG YEAR
Figure 7.3 Comparison of chemical and natural control for the boll weevil during 11 years in east Texas. After Sterling et al. (1984).
and Sterling (1983), Sterling (1978), and Sturm and Sterling (1986). The major advantage of their methods is that the developing weevils in the decaying flower buds or green bolls remain undisturbed and are not removed from their natural habitat until all mortality factors have had an impact on the preemergent boll weevils. Using the evidence remaining with the flower bud or green boll, age-specific mortality from both biotic and physical causes can be reliably identified, the exception being that mortality of free-living adults is not measured. However, identification of 98% of the seasonal cohort mortality was possible during 1981 using these methods (Fillman and Ster- ling, 1983). On the average, only 2% of each generation survived to the free- living adult stage. Sterling et al. (1984) provided evidence supporting the effectiveness of natural enemies, especially ants, in regulating boll weevil abundance. In an east Texas cotton field, natural control of boll weevils resulted in higher yields than insecticidal control in 7 out of 11 years due primarily to mortality attributed to ants (Fig. 7.3).
Contemporaneous mortality (sensu Morris, 1965; Royama, 1981) can sometimes confuse the assignment of mortality to a single agent. The most commonly observed example of contemporaneous mortality in boll weevil studies has been parasitization by Bracan mellitor and predation by the red imported fire ant. The issue of contemporaneous mortality is confused when the evidence of parasitization is removed by the ant predator. The presence of a parasite cocoon in the old boll weevil pupal cell constitutes acceptable evidence of parasitism. However, when the ants remove the immature boll

Natural Control of Arthropod Pests 163
weevil, they may also remove the parasite pupal cocoon. Thus the incidence of parasitism may often be underestimated when using durable evidence techniques. But the total generation mortality will not be underestimated if contemporary mortality exists. This is an example of replaceable mortality, that is, the boll weevil mortality attributed to either the parasite or predator was replaceable since boll weevils not killed by ants would have been killed by parasites, and vice versa. Of course, both ants and parasites kill some boll weevils that would have otherwise survived, and thus either one has the potential to contribute irreplaceable mortality. Ant predators that kill parasites when feeding on the weevil are viewed as a potential threat to both native and introduced parasitoids (Cate, 1985). However, since no intro- duced natural enemy of the boll weevil has become permanently established in the United States (Cate, 1985), and since the potential for classical bio- logical control of pests of annual crops is low (Simmonds, 1948), the threat of ants to parasitoids may be more of a theoretical interest than of a practical one.
It is generally thought to be far safer in empirical studies to find ways to evaluate the causes of mortality than to depend entirely on correlative data based on samples of natural enemy and pest abundance. Thus greater con- fidence is possible where evidence of the mortality of each individual animal is obtained rather than from extrapolations of population density studies. At the risk of redundancy, the often-quoted admonition that “correlation is not causation” is still appropriate. But we would add that correlative evidence of causation can add increasing confidence to observations of mortality to individual animals. Using both experimental methods should provide more definitive evidence than either one alone if they lead to the same conclusion.
Heliothis spp. In the absence of natural enemy-induced mortality, P. M. Ives and L. T. Wilson (unpubl. data) recorded about 60 to 70% of Heliothis mortality before reaching the more damaging fourth and fifth larval instars. This level of mortality, although substantial, is not sufficient to prevent He- liothis from reaching damaging levels. Natural enemies in combination with physically induced mortality have been observed to contribute up to 100% mortality of the egg and first two larval stages under field conditions (McDaniel et al., 1981; Wilson and Gutierrez, 1980). In just 2 days, Heliothis eggs exposed to native natural enemies experienced an average of 86.6% daily mortality from eight weekly experiments (McDaniel and Sterling, 1982). Losses to cotton from damage inflicted by Heliothis were low when such high levels of mortality were observed. In fact, at this high level of mortality, Heliothis was not even surviving at replacement levels (Le., Ro < 1). Similar high levels of mortality have been observed by others (DeLoach and Peters, 1972, Fletcher and Thomas, 1943; van den Bosch et al., 1969).
Under laboratory conditions, female Heliothis will on average produce between 370 and 1774 eggs, the number depending on the species and tem- perature conditions (Fye and McAda, 1972). Assuming that an average of
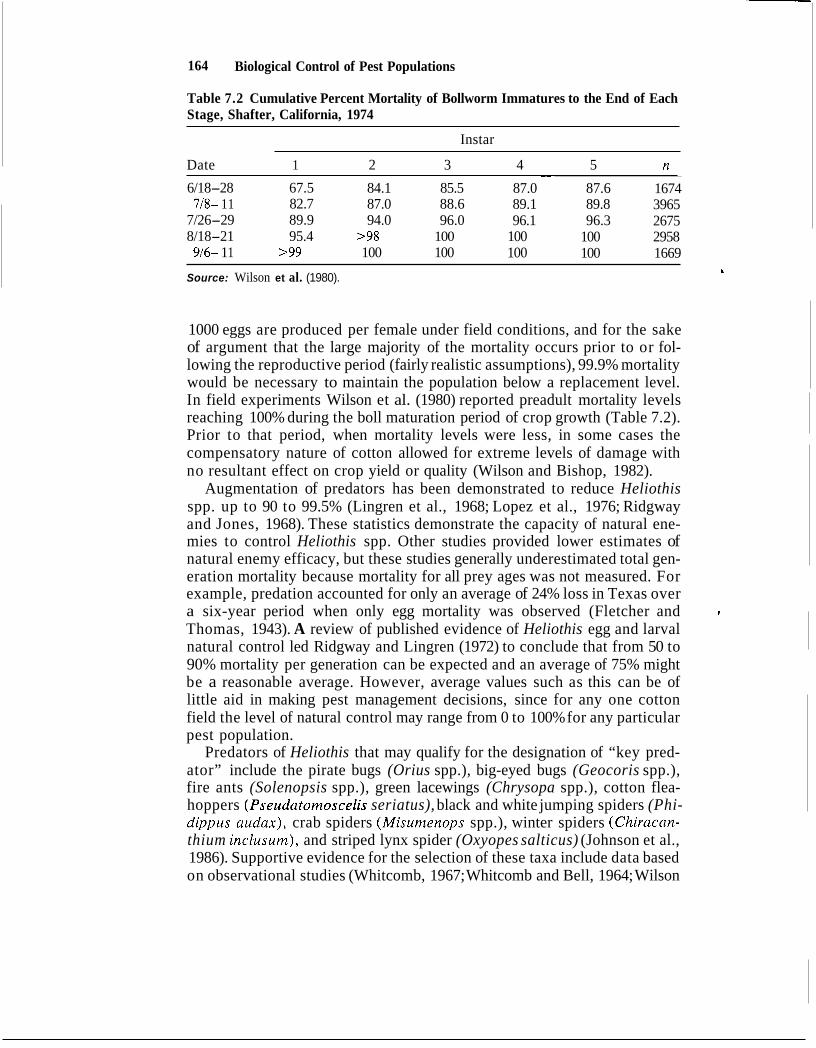
164 Biological Control of Pest Populations
Table 7.2 Cumulative Percent Mortality of Bollworm Immatures to the End of Each Stage, Shafter, California, 1974
Instar
Date 1 2 3 4 5 n ~~
6/18-28 67.5 84.1 85.5 87.0 87.6 1674 718- 1 1 82.7 87.0 88.6 89.1 89.8 3965
7/26-29 89.9 94.0 96.0 96.1 96.3 2675 8/18-21 95.4 >98 100 100 100 2958 916- 1 1 >99 100 100 100 100 1669
Source: Wilson et al. (1980). t
1000 eggs are produced per female under field conditions, and for the sake of argument that the large majority of the mortality occurs prior to or fol- lowing the reproductive period (fairly realistic assumptions), 99.9% mortality would be necessary to maintain the population below a replacement level. In field experiments Wilson et al. (1980) reported preadult mortality levels reaching 100% during the boll maturation period of crop growth (Table 7.2). Prior to that period, when mortality levels were less, in some cases the compensatory nature of cotton allowed for extreme levels of damage with no resultant effect on crop yield or quality (Wilson and Bishop, 1982).
Augmentation of predators has been demonstrated to reduce Heliothis spp. up to 90 to 99.5% (Lingren et al., 1968; Lopez et al., 1976; Ridgway and Jones, 1968). These statistics demonstrate the capacity of natural ene- mies to control Heliothis spp. Other studies provided lower estimates of natural enemy efficacy, but these studies generally underestimated total gen- eration mortality because mortality for all prey ages was not measured. For example, predation accounted for only an average of 24% loss in Texas over a six-year period when only egg mortality was observed (Fletcher and Thomas, 1943). A review of published evidence of Heliothis egg and larval natural control led Ridgway and Lingren (1972) to conclude that from 50 to 90% mortality per generation can be expected and an average of 75% might be a reasonable average. However, average values such as this can be of little aid in making pest management decisions, since for any one cotton field the level of natural control may range from 0 to 100% for any particular pest population.
Predators of Heliothis that may qualify for the designation of “key pred- ator” include the pirate bugs (Orius spp.), big-eyed bugs (Geocoris spp.), fire ants (Solenopsis spp.), green lacewings (Chrysopa spp.), cotton flea- hoppers (Pseudatomoscelis seriatus), black and white jumping spiders (Phi- dippus audax), crab spiders (Misumenops spp.), winter spiders (Chiracan- thium inclusum), and striped lynx spider (Oxyopes salticus) (Johnson et al., 1986). Supportive evidence for the selection of these taxa include data based on observational studies (Whitcomb, 1967; Whitcomb and Bell, 1964; Wilson

Natural Control of Arthropod Pests 165
and Gutierrez, 1980, Wilson et al., 1980), radiolabeling studies (McDaniel and Sterling, 1979, 1982; McDaniel et al., 198l), and cage studies (Lopez et al., 1976). Other predators that may qualify as key predators in certain areas include damsel bugs (Nabis spp.), Collops spp., assassin bugs (Zelus and Sinea spp.), lady beetles (Hippodamia and Coleomegilla spp.), star-bellied orb weavers (Acanthepeira stel lafa), grey dotted spiders (Aysha gracilis), long jawed orb weavers ( Tetragnatha laboriosa), and ridgefaced crab spiders (Misumenoides formosipes) (Sterling, 1983). The age structure of predator and prey may be critical in determining which of the predators above can be classified as a key predator. Stimac and O'Neil (1985) have suggested von Forester equations for dealing with age changes in time. Most predators of Heliothis eggs and small larvae are small predators such as ants, minute pirate bugs, big-eyed bugs, cotton fleahoppers, and immature spiders such as black and white jumping spiders and winter spiders. Larger predators or social hunters are needed to overcome the defenses of Heliothis larvae. Adult leafhopper assassin bugs, fire ants, large winter spiders, celer crab spiders, grey dotted spiders, black and white jumping spiders, and some earwig spe- cies can successfully attack and kill full-grown larvae.
In studying Heliothis predation by four generalist predators, R. E. Jones et al. (unpubl. data) found that each predator differed in its response to temperature, prey availability, and prey density. Geocoris, for example, appeared most sensitive to high temperatures, with peak activity occurring at about 26°C. At first this may seem surprising, since big-eyed bugs can usually be found on cotton plants during the heat of the day. Closer ex- amination, however, revealed that many of the Geocoris have moved down the plant or onto the ground during the hotter hours of the day. At the other extreme, Chrysopa larvae appear very insensitive to high temperatures, with their activity being highest at 39"C, the highest temperature for which data were available. Although all four of these predators are generalists, and feed upon practically any available prey not too large, fast, or equipped with sufficient defenses, minute pirate bug and Chrysopa appear better able to adjust to changes in numbers, types, and distribution of prey. These two predators demonstrated a much greater ability to adjust their distribution to match the distribution of the more readily available prey, while Collops and Geocoris continued to spend the majority of their time feeding on the bottom surface of leaves, even when Heliothis eggs were distributed equally on both surfaces. Each of these predators differ, however in their vertical and struc- tural distribution (Wilson and Gutierrez, 1980) and in their seasonality, and were differentially affected by the above-mentioned factors. As a result, the conclusion of the results from these and related studies is that the predator complex is more effective considering the season as a whole than even the most voracious of the species could be if it were acting alone.
Inaction levels suggested to control Heliothis species include those of McDaniel and Sterling (1982), who proposed that a ratio of one key egg predator to one Heliothis egg was sufficient to control the pest. Hartstack

166 Biological Control of Pest Populations
Table 7.3 Grouping and Efficacy of the Guild of Predators Known to Prey on the Cotton Fleahopper
Efficiency Index
1 .o Species or Group (OEi) Striped lynx spider (Oxyopes salticus) Jumping spiders (Salticidae) 1 .o Crab spiders (Thomisidae) 1 .o Green lynx (Peucetia viridans) 0.7 Star-bellied orb weaver (Acanthepeira stellafa) 0.7 Spotted lady beetle (Coleomegilla maculata) 0.3 Big-eyed bug (Geocoris punctipes) 0.3
et al. (1975) suggested a ratio of one effective natural predator per two He- liothis eggs. Other inaction levels have been proposed by Herrera Aranguena (1965), who concluded that if 15 to 20% of cotton terminals are infested with Rhinacloa spp., the rate of predation on Heliothis larvae should be approx- imately 80 to 100%. In Arkansas, farmers were advised to depend on natural enemies if “beneficials” were present at 20 or more per 56-feet row of cotton (Barnes et al., 1977). Fye (1979) suggested that 35,000 to 50,000 predators per acre were sufficient to remove Heliothis daily. All of these estimates are crude. As new evidence becomes available concerning the identification of the “key predators,” predator-prey age-structure relationships, and many other factors, the prediction of natural enemy efficiency should be- come sufficiently precise for use in pest management programs.
Cotton Fleahopper Several generalist predators have been observed to prey on cotton fleahoppers. Certain species of spiders have been reported preying on fleahoppers (Dean et al. 1987; Reinhard, 1926; Whitcomb and Bell, 1964), and rates of parasitism up to 25% have been recorded (Ewing and Crawford, 1939). Mussett et al. (1979) obtained a correlation (r = 0.62) between the abundance of a complex of predators and the abundance of cotton fleahoppers, indicating that fleahoppers explain about 38% of the variation in predator abundance. These data suggest that fleahoppers are an attractive source of energy for some predaceous arthropods.
To predict the impact of a guild of predators on the cotton fleahopper, the guild is divided into seven groups and weights (OEi) are given to each group according to efficiency (Table 7.3) (Hartstack and Sterling, 1986, 1988). Oxyopes salticus is assumed to be the most efficient predator of the cotton fleahopper, so it is weighted with an efficiency index of 1. Other predators are compared in efficiency to Oxyopes and their intraspecific- group efficiency is weighted to values of 1 or less Oxyopes equivalents (OEi). The total Oxyopes equivalents per unit area (OE,) is estimated by Eq. (7.1), where Ni is the number of individuals of species group i per unit (e.g., per
,
,

Natural Control of Arthropod Pests 167
hectare) area and OEi is the intraspecific-group weighted efficiency of in- dividuals of species group i.
P = 1 - exp [-0.6930E,/(OEsoS)] (7.2)
The percent daily mortality of fleahoppers (P) is estimated with Eq. (7.2), where -0.693 is a correction factor to adjust the exponential curve at p = 0.5, represents the OEs needed to kill 50% of the fleahoppers, and S is the searching area (ranging from 1 to 7, depending on plant size).
Lygus Species Lygus spp. is another example of a “pest” that also func- tions as an entomophage. Wheeler (1976) has reviewed observations on the predacious tendencies of lygus bugs which feed on soft-bodied arthropods. Predation on the eggs of the beet armyworm (Spodoptera exigua) in Cali- fornia (Eveleens et al., 1973) by Lygus hesperus suggests that members of this genus may also feed on the eggs of other pests, such as Heliothis spp. and the pink bollworm. Reports of Lygus as important natural enemies of crop pests have all involved egg predation (Wheeler, 1976). The impact of this predation in cotton fields is not yet clear.
Lygus are generally conceived to be of importance as a pest more than as a predator of other pests. Thus the natural enemies of this genus have been evaluated. The natural predator complex of cotton fields in southern California provided 53 to 76% control of L. hesperus in the egg and nymphal stages (Leigh and Gonzalez, 1976). Geocoris pallens was a very effective predator, while Nabis americoferus was effective in sleeve cages but not in larger cages. Early stages of Nabis alternutus were able to prey successfully on the first three instars of L. hesperus, while large N . alternatus nymphs were able to attack and kill all Lygus instars (Perkins and Watson, 1972). Chrysopa carnea was ineffective against L. hesperus.
The mymarid Anaphes ovijentatus and the braconid Euphoriana uniformis are the dominant parasites in California (Clancy and Pierce, 1966). In alfalfa fields the average rate of parasitization was 46.6% over 14 counties in Cal- ifornia. However, A. ovijentatus may be injurious as well as beneficial since they also parasitize the eggs of Nabis americoferus. This suggests the need for great caution in importing parasites of mirid pests that may also parasitize native natural enemies.
PinkBollworm The pink bollworm (Pectinophora gossypiella), is exposed to predators only during the egg stage, part of the first larval stage, the last- stage larvae, and as an adult. Orphanides et al. (1971) reported one neu- ropteran, one dermapteron, five Hemiptera, four Coleoptera, and nine spi- ders feeding on the pink bollworm in southern California. All except spiders fed on eggs, while Orius tristicolor and Geocoris punctipes preferred first-
I I

168 Biological Control of Pest Populations
stage larvae over eggs. Only larger predators and spiders were able to over- come and consume larger larvae. Based on laboratory studies, the following egg predators, arrayed in order of effectiveness, were listed: second-stage larvae of Chrysopa carnea, adults of Collops marginellus, Geocoris punc- tipes, adult Notoxus calcaratus, Nabis americoferus, and 0. tristicolor. In field cage studies, Irwin et al. (1974) concluded that N . calcaratus and Spana- gonicus albofasciatus adults were ineffective predators of pink bollworm eggs. The most promising predators were N . americoferus and G. pallens, which reach natural field densities of one and three per plant, respectively. However, C. carnea, although an efficient predator rarely reached field den- sities above one per plant. Henneberry and Clayton (1985) added Collops vittatus and Hippodamia convergens to the list. This guild of predators is capable of removing more than 90% of pink bollworm eggs placed artificially on plants in the field (Henneberry and Clayton, 1982).
Unfortunately, with the current “scorched earth” insecticide control strategy used in insect management over a large percentage of the south- western cotton regions, predator-induced pink bollworm mortality is of little more than academic interest. The above-mentioned generalist predators in such areas are not able to survive in sufficient numbers through the gauntlet of insecticide sprays to enable them to inflict sufficient mortality to prevent economic loss.
Spider Mites Three spider mite species are responsible for the majority of mite damage to cotton in the United States. Strawberry spider mite, Te- tranychus turkestani, twospotted spider mite, T. urticae, and Pacific spider mite, T. pacificus, each predominate at different times of the season; how- ever, any one or all three can be found throughout the crop’s growth and development. Although primarily a pest of more arid regions of California’s San Joaquin Valley, the relatively recent trend to an increased use of syn- thetic pyrethroids has exacerbated the severity and importance of spider mites not only in California but over much of the world. Spider mites, unlike most other cotton pests, have an ability to literally explode in numbers. Were physical and biotic mortality factors not acting, a single-mated female emerging from the soil or blowing into a 40-ha field early in the season could result in sufficient numbers of mites to cause economic yield loss (L. T. Wilson, unpubl. data). Normally, however, a complex of generalist predators is able to contain spider mite densities below economically damaging levels.
Our understanding of the factors primarily responsible for mite abundance was until recently, extremely limited. Gonzalez et al. (1982) reported that big-eyed bug and minute pirate bug abundance were both very closely as- sociated with mite abundance, However, neither of these predators appear to reach densities sufficiently high early in the season to prevent spider mites from rapidly reaching economically damaging levels. Gonzalez et al. (1982) and Gonzalez and Wilson (1982) also reported the immature stages of the western flower thrips, Frankliniella occidentalis, “actively feeding on Te-

Natural Control of Arthropod Pests 169
PHYSIOLOGICAL TIME (DEGREE DAYS > 12°C)
Figure 7.4 Seasonal abundance of western flower thrips in San Joaquin Valley cotton. L. T. Wilson and D. Gonzalez (unpubl. data).
tranychus eggs, wherever the latter was congregated.” In studies conducted from 1978 through 1984, Frankliniella was consistently the most abundant arthropod found in the cotton, with its greatest abundance often coinciding with the presquaring and early squaring periods of crop growth. This period of colonization is particularly relevant in that none of the other generalist or opportunistic predators are yet in abundance. D. Gonzalez (unpubl. data) and Trichilo (1986) found further that flower thrips are an extremely vora- cious mite predator, with third instar nymphs able to damage over 70 mite eggs over a 24-hour period under laboratory conditions.
High thrips densities (Fig. 7.4) combined with a high feeding potential results in this species being unequaled in its ability to suppress early- and midseason mite populations. Large-scale predator suppression experiments conducted in California from 1981 to 1984 demonstrate that the use of a broad-spectrum insecticide reduces the abundance of thrips and other pred- ators and inevitably resulted in the initiation of nearly exponential mite pop- ulation growth about 2 to 3 weeks hence (D. Gonzalez and L. T. Wilson, unpubl. data). This set of experiments has provided useful information for developing economic thresholds for spider mites on cotton (Ellington et al., 1984; Wilson et al., 1983; Wilson, 1986), but equally important, they have enabled the control potential of natural enemies, particularly thrips, to be quantified, One of the most difficult aspects of pest management is predicting

170 Biological Control of Pest Populations
I 1981 Cotton, Shafter I .o
I P' I
1 I
I I
I I
I p4' I
I I
I I
\ \ I I \ I
b -. . \ -.
I -c. .-
01 1 I I I I I
1350 1525 1700 I€ 650 825 1000 1175
PHYSIOLOGICAL TIME OD>12OC (from planting)
5
Figure 7.5 Several patterns of spider mite infestation in San Joaquin Valley cotton. L. T. Wilson and D. Gonzalez (unpubl. data)
whether a pest species will cause economic loss in the future, Heliothis spp. or Lygus, for example, are both highly mobile, have a wide host range, and within an individual field do not necessarily have an easily forecasted pop- ulation pattern. Spider mites, on the other hand, have a monotonically in- creasing, then decreasing population pattern (Fig. 7 . 3 , lending itself to ac- curate prediction of future spider mite abundance (see Chapter 5, Fig. 5.12). Wilson et al. (1985) reported that by using the early season rate of spider mite increase they were able to predict with a high degree of accuracy up to 5 weeks in advance when an economic infestation would develop. This relatively simple regression procedure enables the combined impact of all mortality factors to be evaluated without the complexity inherent in predator efficacy weightings as necessary for evaluating the impact of generalist pred- ators on some of the pests discussed above. If the rate of spider mite pop- ulation increase exceeds a damage trajectory (Wilson, 1986), this implies that the mortality agents are not sufficient and that economic loss will prob- ably result at some future predicted date unless appropriate management actions are initiated (Fig. 7.5).
Other Pests Observations of the natural enemies of pests such as cotton aphids (Aphis gossypii), cotton leafworm (Alabama argillacea), and thrips have been reported by Whitcomb and Bell (1964). Empirical evidence of the impact of natural enemies on some secondary pests of cotton include the impact of Telenomus mesillae and Collops marginellus to a stink bug. These

Natural Control of Plant Pathogens 171
natural enemies contributed an average of 61.4% mortality to the pentatomid Euschistus impictiventris in Arizona cotton fields (Clancy, 1946). Life table studies by Ehler et al. (1973) and Eveleens et al. (1973) suggested that several of the same species of predators of Heliothis inflict major mortality on the cabbage looper (Trichoplusia ni) and the beet armyworm (Spodoptera ex- igua). These predators include 0. tristicolor, G . pallens, N . americoferus, and C. carnea. Predation rates on the cotton leafworm after a 48-hour ex- posure to the native predator guild was 88.7 and 88.4% for eggs and small larvae, respectively (Gravena and Sterling, 1983). The predator guild of the cotton leafworm was similar to the Heliothis guild and other lepidopterous inhabitants of cotton agroecosystems. However, spiders were the dominant predators of cotton leafworm larvae, whereas the above-mentioned Hem- iptera were dominant predators of eggs.
Arthropod Control with Microbials
Pests of cotton are attacked by several naturally occurring microbials, in- cluding bacteria, protozoa, fungi, and rickettsiae (Falcon, 1971). Although these microbials probably play a role in pest control, their density is not generally monitored to assist in making pest management decisions. How- ever, there is some dependence on the cabbage looper virus in pest man- agement programs. Generally, loopers are controlled by this virus before economic loss occurs (National Apademy of Sciences, 1975).
Microbials such as Bacillus thuringiensis and the nuclear polyhedrosis virus, applied to the field as insecticides, have been used in the management of Heliothis species in many areas of the world. The major advantage of using microbial pathogens to control pests is that they inflict little or no harm to other natural enemies. But their impact has generally been sufficiently sporadic (Bell, 1981) that interest in their use seems to be waning. Emphasis in the past has been placed on efforts to increase the effectiveness of ar- thropod pathogens by developing or identifying new, more virulent strains of pathogens; improving spray formulations; and developing new methods of applying or disseminating pathogens (Bell, 1983). None of these efforts has met with notable success. One method of using microbials that seems to have been largely overlooked is to find ways to monitor natural popu- lations of pathogens as a management tool for predicting their impact on arthropod pest population.
NATURAL CONTROL OF PLANT PATHOGENS
Since ancient times, man has practiced biological control of plant pathogens through cultural practices such as the use of legumes in crop rotation, clean tillage, and the use of organic manurial amendments. These practices pro- vided for the biological destruction and/or suppression of disease organisms.

172 Biological Control of Pest Populations
Biological control aimed directly at the pathogen or mediated through ad- justments in the host offers unlimited opportunities to reduce losses caused by biotic and abiotic stresses. As in the case of arthropods, biological control of plant pathogens should be regarded as one part of the total integrated pest and crop management system. Its relative importance can be expected to vary with different diseases, dominant in some instances where other mea- sures have failed, but of minor importance where other measures provide inexpensive, effective control.
Biological control of plant pathogens seeks a solution in terms of restoring and maintaining the biological balance within the ecosystem and must be considered part of modern agriculture. Biological control offers a powerful means to improve the health and hence the productivity of plants by suppres- sion or destruction of pathogen inoculum, protection of plants against in- fection, or increasing the ability of plants to resist pathogens.
Concepts and Mechanisms
The mechanisms of biological control of plant pathogens have been grouped into three general categories: (1) reduction of inoculum by microorganisms antagonistic to the target pathogen, (2) protection of host plant surfaces by antagonists, and (3) management of physiological incompatibility between host and pathogen obtained by genetic change in the host or by inoculation with an avirulent or nonpathogenic microorganism (Baker, 1968; Baker and Cook, 1974; and Cook and Baker, 1983).
Antagonists of plant pathogens may be introduced or resident (Garrett, 1965). Introduced antagonists are prepared products applied to the soil, seed, or plant to control a pathogen. Resident antagonists are part of the natural microbiota in soil or on roots, leaves, fruit, or other plant parts.
The equilibria of microorganisms in the agroecosystem are established as a result of antagonistic processes: competition, antibiosis (including fungis- tasis), exploitation, and hyperparasitism. All of these types of antagonism occur commonly, especially in soil, and all can influence the activity of plant pathogenic microorganisms. For example, a soilborne pathogenic fungus may be subject to fungistasis, antibiosis (which can inhibit vegetative growth), competition from other organisms for colonizeable organic matter in soil upon which the fungus is dependent for an energy source to reproduce itself, and parasitism by other microorganisms upon vegetative and survival structures (hyperparasitism) (Papavizas and Lumsden, 1980). Hyperpara- sitism refers to the control of pathogenic microorganisms with other micro- organisms or viruses which parasitize or antagonize the pathogen. The best known cases of hyperparasitism include the bacteriophages, mycoparasites, and nematophagous fungi.
The protection of plant surfaces by antagonists and the management of host pathogen incompatibility have emerged as necessary strategies because plant pathogens commonly exist in close association with their hosts, and
1

Natural Control of Plant Pathogens 173
many complete most or all of their life cycle inside the host plant. Obviously, the host has a biological system of its own which must be intimately involved in the biological control of these pathogens.
Antibiosis is the most widely recognized mechanism that may cause in- activation or destruction of soilborne plant pathogen propagules preventing germination. Antibiosis is used in plant pathology to describe the chemical inhibition of one microorganism by another, which is more parallel to al- lelopathy, whereas usage in entomology describes a mechanism of host plant resistance to a pest.
The specific form of antagonism may be mediated by specific toxic me- tabolites of microbial origin, by soil fungitoxins, and by lytic agents (Jackson, 1965). Antibiosis in soil can also result from elevated fungistasis or liberated inhibitory volatiles. If fungal propagules are not inactivated by antagonism or reduced in numbers below economic threshold levels in the dormant state or during germination, biological control may still be possible by preventing certain processes or functions. These include lysis and mycoparasitism of mycelia, suppression of germling growth and sporulation, hypovirulence and mycoviruses, and cross protection (Garrett, 1965; Baker, 1968; Baker and Cook, 1974; Papavizas and Lumsden, 1980; Cook and Baker, 1983). Viruses or virus-like agents specific for the pathogen may reduce virulence, aggres- siveness, or survival ability of the pathogen (Ghabrial, 1980).
Bird (1982) hypothesized that microorganisms (bacteria, fungi, and ac- tinomycetes) which are natural colonizers of the plant tissues and root sur- faces (symbiotic organisms) have a major role in resistance to diseases and insects, and are under the genetic control of the host. Components of seed and root exudates of Tamcot multi-adversity resistance (MAR) cotton cul- tivars can selectively influence rhizosphere-rhizoplane population levels of bacteria and actinomycetes (Batson, 1972; Tsai and Bird, 1975; Bush, 1980).
Bird et al. (1979) and El-Zik et al. (1985) have shown that the concentra- tions of symbiotic organisms associated with plant parts (leaves, terminals, and squares) were much higher in MAR than in non-MAR cottons (see Chap- ter 8). The predominant symbionts were Bacillus spp., which were desig- nated smooth white (SW) and rough white (RW), based on their colony characteristics when grown on Allen’s soil extract agar. Recently, the SW bacterium was identified as Bacillus megaterium and the RW as B. cereus (Howell et al., 1987). Evidence that symbiotic organisms isolated from Tam- cot CAMD-E influence the response of the host to the bacterial blight path- ogen, Xanthomonas campestris pv malvacearum (Bird et al., 1979; El-Zik
’
.
l
et al., 1983), seedling pathogens and Phymatotrichum omnivorum (Bird et al., 1979, 1980), and boll weevil (Benedict et al., 1979) has been reported.
Baker (1968), Baker and Cook (1974), Papavizas and Lumsden (1980), Cook and Baker (1983), and Baker (1985) have provided extensive reviews on biological control and its principles and mechanisms. The potential for biological control of plant diseases on the phylloplane has recently been

174 Biological Control of Pest Populations
reviewed by Blackeman and Fokkema (1982) and Windels and Lindow (1 985).
Approaches to Biological Control of Cotton Pathogens
One may consider biological control of cotton pathogens by reducing in- oculum density equivalent to biological control of insects: namely, the re- , duction of the insect population to an economically acceptable threshold level. A number of actinomycetes, bacteria, and fungi have been isolated from soil and plant surfaces, and some have shown potential as biological control agents. Under laboratory and controlled growth conditions they per- form quite well; however, a large number fail to control disease consistently under field conditions.
Recent research has focused on utilizing microbes isolated from the cotton rhizosphere and rhizoplane as biological agents to control seed-seedling pathogens. Hagedorn et al. (1985) identified 17 different genera of bacteria from the cotton rhizosphere and rhizoplane. They tested several isolates from these genera on field-grown cotton and obtained biological control against the seed-seedling pathogens Pythium uftimum and Rhizoctonia solani.
Gfiocfadium spp. are resident antagonists in soils, reducing populations of selected fungi. Howell (1982) found that G. virens parasitized R. sofani and inhibited P. uftimum by antibiosis. Damping-off of cotton seedlings was reduced when the antagonist was placed in soil with the seed.
Howell and Stipanovic (1979, 1980) obtained improved emergence of cot- ton seedlings by treating the seed with strains of Pseudomonas jluorescens isolated originally from the rhizosphere of cotton seedlings. Treatment of cotton seed with strain Pf5 of P. fluorescens improved seedling survival from 30 to 79% and 28 to 71% in soil infested with R. sofani and P. uftimum, respectively. Ideally, antagonists should be effective against several path- ogens or strains of a pathogen; however, it has been shown that antagonists can have a high degree of specificity. Howell and Stipanovic (1979, 1980) isolated two chlorinated phenyl pyrrole antibiotics from P. jluorescens. The antibiotic pyrrolnitrin inhibited R . sofani in vitro but was ineffective against P. ultimum, and the antibiotic pyoluteorin was extremely inhibitory to P. uftimum but not to R . sofani. Treatment of cotton seed with the antibiotics alone gave improved emergence and decreased damping-off in soil infested with R. solani and P. uftimum, respectively. Pyrrolnitrin was also effective against Thefaviopsis basicofa, Verticillium dahfiae, and Alternaria spp. Elad et al. (1982) showed that coating cotton seed with Trichoderma hamatum or T. harzianum decreased damping-off of cotton seedlings in field tests.
Yin et al. (1957, 1965) in China selected a Streptomyces sp. from among 4000 isolates of actinomycetes from roots of cotton and alfalfa on the basis of its in vitro antibiosis to R. sofani and V . afbo utrum. This culture, strain
.
\

Natural Control of Plant Pathogens 175
5406, has been used on 15 million acres of cotton over a 30 year period, giving increased crop growth (Cook and Baker, 1983).
In recent years, considerable attention has been devoted to mycorrhizal fungi, which have been shown to provide effective protection of feeder roots against soilborne pathogens such as Phytophthora, Pythium, and Fusarium. In addition, mycorrhizal fungi increase the uptake of phosphorus and other nutrients of plants, and generally improve plant growth and health.
Under controlled conditions, Thelaviopsis basicola caused less stunting of cotton and tomato when endomycorrhizae were present in their roots as compared with the same host with roots lacking mycorrhizae. Roncadori and Hussey (1980) have shown that VA-mycorrhizae Gigaspora margarita and Glomus etunicatus are both excellent symbionts on cotton, and in fu- migated low-phosphorus soil may improve early plant growth by as much as 600%. When each mycorrhizal fungus was challenged in greenhouse stud- ies with root-knot nematode, stunting caused by Meloidogyne incognita was completely nullified and there was no effect on root or shoot biomass. Al- though both G. margarita and G. etunicatus nullified stunting caused by M . incognita, their effects on nematode reproduction were different. Only my- corrhizae formed with G. etunicatus significantly reduced the egg popula- tions of M. incognita. Smith et al. (1986) found that G. intraradices and G. margarita can increase host tolerance to the root-knot nematode under field conditions and reduce the severity of yield loss.
Monoculture and suppressive soils have been reported to cause natural disease reduction. Monoculture decline occurs when continuous cropping with a susceptible crop results in a decrease of disease incidence (Shipton, 1977), and soil suppressiveness develops as a natural reduction in disease incidence (Baker and Cook, 1974; Cook and Baker, 1983).
Many groups of organisms have been cited as candidates for the biotic suppressive factor in disease suppressive soil (Hornby, 1983; Papavizas and Lumsden, 1980; Cook and Baker, 1983). King et al. (1934) first showed that heavy application of organic matter controlled Phymatotrichum root rot of cotton. This was confirmed by Clark (1942), who related the rate of break- down of organic matter to the rate of death and decomposition of sclerotia of the pathogen.
Many species of soil microarthropods have been reported to be predom- inantly mycophagous and may have a role in biological control of soil mi- croorganisms. Curl (1979) and Wiggins and Curl (1979) found that two species of collembola (Proisotoma minuta and Onychiurus encarpatus) feed on hypae of R . solani, possibly reducing inoculum density of the pathogen in the root zone. In addition, collembola can transmit and inoculate the cotton seedling root zone with antagonistic and pathogenic sporulating fungi. Curl (1982) reported that effective biological control of pre- and postemergence damping-off of cotton seedlings was obtained by adding collembola to the soil in populations of 909 lb/acre under experimental conditions.
Control of nematode pests by biological agents is a practice which has

176
not been particularly successful to date, although a wide range of predators and parasites attack nematodes. Duddington and Wyborn (1972) have re- ported about 50 known species of predacious fungi that either capture or kill nematodes in the soil. Van Gundy (1972) listed, in addition to the 50 known species of fungi, two protozoans and numerous other small inver- tebrates in the soil that had been reported to destroy or to feed on nematodes.
Mankau (1980, 1981) reviewed biological and microbial control of ne- matode pests. The bacterium Bacillus penetrans has shown promise as a control agent, and has a life cycle remarkably adapted to parasitism of root- knot nematodes (Mankau, 1980, 1981). Recent discovery of several fungal parasites of nematode eggs and cysts has expanded interest in biological agents (Mankau, 1980, 1981).
Biological Control of Pest Populations I
I
Current Status of Biological Control of Plant Pathogens
There are few biocontrol agents of plant pathogens presently employed in agriculture despite long-term research efforts in many public laboratories and the more recent increase in industrial research efforts. A major limitation on the current use of biological control agents has been the inconsistent results achieved when applied under field conditions. Following the initial discovery and testing of biocontrol agents, much work needs to be done concerning the various aspects of industrial microbiological application. The main problem in application is in managing the host, environmental condi- tions, and the agent itself so that the activity of the biocontrol agent is enhanced.
Antagonists of plant pathogens are largely passive and relatively non- mobile, and make accidental contact with pathogens. Therefore, biological control agents of plant pathogens must be applied directly to the locality where infection will occur. Antagonists must be in high concentrations at the infection sites, such as the leaf surface, to control a leaf-infecting path- ogen, or on seeds or roots to control soil pathogens. A major difference between controlling arthropods and plant pathogens with biocontrol agents lies in the mechanisms that affect their populations. Biological control of plant pathogens has been effected principally by competition and antibiosis, and sometimes hyperparasitism, whereas biological control of insects is mainly by predation, parasitism, and disease. Many effective cultural meth- ods currently in use promote and enhance biological control of plant pathogens.
The challenge has been to identify the biological agents and gain a thor- ough understanding of mechanisms and factors, both biotic and abiotic, that affect their behavior. More research is needed on the mechanisms involved in biological control, their interaction with biotic and abiotic agents, and the influence of environment. At present, the problems of application, stability, cost, effective duration of the biological control agent, and other aspects have not been perfected sufficiently to allow commercialization.

Natural Control of Weeds 177
NATURALCONTROLOFWEEDS
The principles and procedures for biological control of weeds are well es- tablished (Huffaker, 1959; Andres, 1977; Templeton et al., 1979; Charudattan and Walker, 1982; Charudattan, 1985; Templeton, 1982). Several successful examples of biological agents for weed control, including phytophagous in- sects and plant pathogens, have been reported; however, many biological control agents are usually highly specific, resulting in a very narrow spec- trum of weeds controlled by a single pest. Although bacterial and viral plant pathogens have been examined, all the microbial weed control agents that are in use or under development in North America are fungal pathogens.
There has been limited success using insects and plant pathogens to con- trol major weeds in cotton. Augmentation of biocontrol agents may be used to overcome some of the limitations. In Mississippi, the native moth Bactra verutana attacks purple nutsedge, Cyperus rotundus, but it is ineffective since too few moths overwinter. By the time the moth population has in- creased, the nutsedge is so vigorous that larval feeding cannot control new shoots. Frick and Chandler (1978) reported that early season inundative release of mass-reared B. verutana larvae and adults in cotton fields allowed larvae to suppress nutsedge, resulting in seed cotton yields equal to those in herbicide-treated or nutsedge-free fields.
Recent advances in biological control of weeds have shown that plant pathogens can control weeds within crops when properly employed. Walker (1980) reported that levels of spurred anoda, Anoda cristata, were reduced 75% with a 100% leaf infection when the fungus Alternaria macrospora was applied as a single foliar spray to seedlings in the two- to three-leaf stage.
Colletotrichum malvarum causes an anthracnose of prickly sida, Sida spinosa. Control of prickly sida with the mycoherbicide, C. malvarum, in the field has been erratic and dependent on environmental conditions at the time of inoculum application. Maximum control (90 to 95%) was achieved when inoculum was applied while cool (24"C), moist conditions prevailed for several days following inoculation (TeBeest, 1981). In field tests, one application of a spore suspension (2 x IO6 spores/mL) of C. malvarum in September at the rate of 378 L/hectare reduced the density of prickly sida by 98%. However, results of identical tests conducted in July did not result in control of prickly sida at any inoculum concentration tested (TeBeest, 1981). The major constraints to the effective use of plant pathogens as bio- control agents are environmental conditions, such as temperature and du- ration of free moisture, required for infection and disease development; and the necessity for multiple inoculum applications to control the repeated emergence of new weed seedlings. On the positive side, mycoherbicides can easily be applied using the same techniques and equipment for herbicides.
Orr (1981) found that silverleaf nightshade is naturally parasitized by the nematode Nothanguina phyllobia. The nematode injures the weed by re- ducing its growth and development, vigor, and seed production. Nematode-

178 Biological Control of Pest Populations
infecting larvae overwinter in the soil or in infested plant tissues, and attack silverleaf nightshade plants at emergence or any time during the growing season when environmental conditions are favorable. After four years, over 80% control of silverleaf nightshade was achieved using N . phyllobiu in two field tests (On, 1981).
Geese have been used successfully for many years and have given eco- nomic control to grasses in cotton. Geese will feed on johnsongrass, bur- mudagrass, nutsedge, and annual grasses; however, they do not control cocklebur or pigweeds. Three to five 6-week-old geese per acre are generally effective (Miller et al., 1977). However, several management problems must be considered when geese are used. These include constant availability of clean drinking water, supplemental feeding, providing shade, protection from predators, and fencing.
The use of insects and plant pathogens in biological control is an exciting and rapidly expanding area in weed science and has potential for broad application. Biological weed control agents may be incorporated and inte- grated with other pest control strategies and crop management systems.
TOWARD CLASSICAL BIOLOGICAL CONTROL
The introduction of exotic agents for the biological control of arthropod and weed pest species in cotton may have little or no chance of success if re- peated applications of broad-spectrum insecticides continue to be used in the cotton agroecosystem. Few predators of insects, pests, or plants can survive and maintain levels necessary for control of insect or weed pests if exposed to repeated insecticide sprays. Thus a prerequisite to the use of classical biological control may be to eliminate broad-spectrum pesticides from the ecosystem; or as a minimum, severely curtail the number of sprays applied during a season. The instant elimination of all insecticides from cot- ton production is not likely to be economically feasible. Some cotton regions, such as California’s lower desert valleys, are essentially addicted to chemical pesticides (van den Bosch, 1978). Were insecticide usage to be instantly stopped in such areas, the resultant damage would probably be catastrophic. Farmers in highly disturbed production systems are dependent on chemicals in order to obtain high yields. Thus if a community of farmers should choose to change their chemical-intensive production approach and instead to con- trol pests with a judicious combination of pesticides and natural biological control, a period of chemical “withdrawal” may be necessary to evolve into a chemically rational system. Under such a system insecticide use must be integrated in a way that will minimize or eliminate the impact on the native natural enemies.
Augmentation of Natural Enemies Theoretically, the augmentation of natural enemies to control cotton pests, particularly the Heliothis spp., should provide a method that would not de-
\

Natural Control of Weeds 177 I
NATURAL CONTROL OF WEEDS
The principles and procedures for biological control of weeds are well es- tablished (Huffaker, 1959; Andres, 1977; Templeton et al., 1979; Charudattan and Walker, 1982; Charudattan, 1985; Templeton, 1982). Several successful examples of biological agents for weed control, including phytophagous in- sects and plant pathogens, have been reported; however, many biological control agents are usually highly specific, resulting in a very narrow spec- trum of weeds controlled by a single pest. Although bacterial and viral plant pathogens have been examined, all the microbial weed control agents that are in use or under development in North America are fungal pathogens.
There has been limited success using insects and plant pathogens to con- trol major weeds in cotton. Augmentation of biocontrol agents may be used to overcome some of the limitations. In Mississippi, the native moth Bactra verutana attacks purple nutsedge, Cyperus rotundus, but it is ineffective since too few moths overwinter. By the time the moth population has in- creased, the nutsedge is so vigorous that larval feeding cannot control new shoots. Frick and Chandler (1978) reported that early season inundative release of mass-reared B . verutana larvae and adults in cotton fields allowed larvae to suppress nutsedge, resulting in seed cotton yields equal to those in herbicide-treated or nutsedge-free fields.
Recent advances in biological control of weeds have shown that plant pathogens can control weeds within crops when properly employed. Walker (1980) reported that levels of spurred anoda, Anoda cristata, were reduced 75% with a 100% leaf infection when the fungus Alternaria macrospora was applied as a single foliar spray to seedlings in the two- to three-leaf stage.
Colletotrichum malvarum causes an anthracnose of prickly sida, Sida spinosa. Control of prickly sida with the mycoherbicide, C. malvarum, in the field has been erratic and dependent on environmental conditions at the time of inoculum application. Maximum control (90 to 95%) was achieved when inoculum was applied while cool (24"C), moist conditions prevailed for several days following inoculation (TeBeest, 1981). In field tests, one application of a spore suspension (2 x IO6 spores/mL) of C. malvarum in September at the rate of 378 L/hectare reduced the density of prickly sida by 98%. However, results of identical tests conducted in July did not result in control of prickly sida at any inoculum concentration tested (TeBeest, 1981). The major constraints to the effective use of plant pathogens as bio- control agents are environmental conditions, such as temperature and du- ration of free moisture, required for infection and disease development; and the necessity for multiple inoculum applications to control the repeated emergence of new weed seedlings. On the positive side, mycoherbicides can easily be applied using the same techniques and equipment for herbicides.
Orr (1981) found that silverleaf nightshade is naturally parasitized by the nematode Nothanguina phyllobia. The nematode injures the weed by re- ducing its growth and development, vigor, and seed production. Nematode-

Toward Classical Biological Control 179
stroy natural enemies which regulate the pests and that could be used to replace insecticides when resistance develops (Ridgway et al., 1973). AI- though technically feasible (Ridgway and Morrison, 1985), the economic feasibility has seldom been demonstrated in the United States. King et al. (1985) reviewed augmentation projects with 29 different species in reaching this conclusion concerning economics. More specifically, they reported that it is not feasible to manage Heliothis spp. in cotton by augmentative releases of Trichogramma pretiosum, one of the most ambitious projects yet at- tempted in the United States (Stinner, 1977). Other reviews of augmentation (Ridgway and Morrison, 1985; Ridgway and Vinson, 1976) indicate the fea- sibility of the augmentation approach in countries, such as in the Soviet Union and China, where labor is considerably cheaper. Although several parasites and predators can be purchased in the United States, federal and state entomologists have generally not recommended their use in cotton. Thus the augmentation approach to biological control of arthropod pests of cotton is currently being used on a limited basis in the United States but largely awaits more efficient natural enemies or cheaper production methods.
I
Restoration Ecology
For a farmer to take advantage of nonchemical means of controlling cotton insect pests, some cotton agroecosystems will require restoration. It is rather simple for a farmer using a “natural control” strategy to change to a chemical strategy. However, returning to a noninsecticidal strategy following heavy use of insecticides may be more difficult. It may take as long as three years for the natural enemy fauna to recover completely following multiple ap- plications of broad-spectrum insecticides (Bartlett, 1964). Restoring the ef- ficiency of the natural enemies may be virtually impossible in a community where several neighbors are using an intensive insecticidal strategy.
Barring resistance to insecticides, it is generally possible to achieve high yields and profits using an intensive insecticide program for pest control. However, if for whatever reason the insecticides lose their effectiveness, growers in some areas can switch to a cotton production system using min- imal applications or no insecticides. A pertinent question is: How is it pos- sible to rely on natural control or increase its effectiveness when needed while minimizing the risk of crop loss? California and Texas growers often rely on natural control almost exclusively; control measures are normally required only during relatively brief periods of the growing season. To aid them in their decision-making process, growers can take periodic samples of pests and natural enemies in cotton fields to determine if sufficient natural enemies are available to prevent unacceptable losses by the pests (Sterling, 1984). This tactic adds the additional cost of sampling to production costs, so for sampling to be a feasible tactic, it should provide economic and eco- logical benefits that exceed the sampling costs. Under current integrated

180 Biological Control of Pest Populations
pest management programs for cotton, lip service is given to the sampling and use of natural enemies. But since few “inaction levels” are available to assist the decision maker, most management decisions are based on the abundance of pests with insufficient regard for the abundance of natural enemies. On the other hand, the decision made by some farmers to grow cotton without insecticides is probably also made “automatically” without regard to the density of natural enemies or pests. This decision is also not likely to be optimal and could probably benefit from better information on the density of natural enemies and pests.
As the true role of natural enemies in the cotton agroecosystem is de- termined, and effective inaction levels are made available for key natural enemies, management decisions should often be improved by counting the natural enemies to predict their potential impact on the pest species. How- ever, this tactic is likely to be unprofitable for those growers who choose either end of the tactical spectrum. Few events in cotton fields can be pre- dicted with a high level of reliability. However, one characteristic of cotton fields and their associated fauna and flora that is highly predictable is that there will be constant change. Plants grow or die, arthropods, plant patho- gens, and weeds increase or decrease in numbers, and weather changes. No two years are the same with regard to plant growth, pest population dynam- ics, or weather. Thus it is unlikely that automatic application of production tactics will be optimal. To achieve optimal decisions, the availability of re- liable information is critical. Thus sampling to obtain reliable information concerning the density of natural enemies should add to the reliability of pest management decisions (see Chapter 5).
I
Classical Biological Control
“No effective exotic natural enemy of the boll weevil has been imported and permanently established in the US” (Cate, 1985). Of about 15 species of entomophagous insects that have been imported and released against He- liothis spp., none have so far been successful (Johnson et al., 1986). The same is essentially true for other key pests of cotton. For example, at least nine parasite species have been introduced into the United States for control of the pink bollworm, and none have become established (Clausen, 1978). Although the potential exists for the classical introductions of foreign bio- logical control agents into cotton for pest control, the risk of failure appears to be fairly high, due largely to the durational instability of the cotton crop and the use of broad-spectrum insecticides for pest control. Success rates for introductions might be increased by restoring the ecosystem to reduce or eliminate the insecticidal contamination in communities where releases are made. According to Beirne (1979, the establishment rate of introduced natural enemies into Canadian annual crops is 16%. Thus it would appear superficially that a similar low probability of successful establishment is possible in a cotton agroecosystem.

References 181
CONCLUSION
All cotton pests would cause greater damage to the crop if natural enemies were absent. It is likely that cotton could not generally be grown profitably without the impact of naturally occurring predators, parasites, and pathogens (van den Bosch and Hagen, 1966). However, the full benefit of natural ene- mies is not being realized on millions of acres of cotton grown in the world that are currently dependent on chemical pesticides for control. The wise use of natural enemies could result in a reduced pesticide load on the en- vironment and greater long-term economic benefit to the farmers.
The advantage of using natural enemies over complete reliance on chem- icals is that native natural enemies voluntarily colonize cotton fields, they are self-multiplying, they can be very effective, they do not pollute the eco- system, and they cost the farmer nothing. To realize the advantage of using natural enemies, there is an urgent need to identify the key natural enemies for each pest and to establish inaction levels for use in pest management decisions. This information, used together with reliable monitoring programs and predictive computer models, should make it possible to determine when not to intervene in the dynamics of pests and their natural enemies with no or limited risk to the cotton farmer.
REFERENCES
Adkisson, P. L. 1971. Objective use of insecticides in agriculture, in J. E. Swift (ed.), Agricultural Chemicals-Harmony or Discord for Food, People, Environment. Univ. Calif. Div. Agric. Sci. Publ. pp. 43-51.
Adkisson, P. L. 1973. The principles, strategies, and tactics of pest control in cotton, in P. W. Geier, L. R. Clark, D. J. Anderson, and H. A. Nix (eds.), Insects: Studies in Population Management. Ecol. SOC. Aust. Mem. 1. Ecological Society of Aus- tralia, Canberra. pp. 274-283.
Agnew, C. W. and W. L. Sterling. 1981. Predation of boll weevils in partially open cotton bolls by the red imported fire ant. Southwest. Entomol. 6:215-219.
Andres, L. A. 1977. The biological control of weeds, in J. D. Fryer and S. Matsunaka (eds.), Integrated Control of Weeds. University of Tokyo Press, Tokyo, pp. 153- 174.
Anon. 1985. 1983 Texas Pesticide Use. Tex. Agric. Exp. Stn. Rep. D-283. 24 pp. Baker, R. 1968. Mechanisms of biological control of soil-borne pathogens. Annu.
Rev. Phytopathol. 6:236-294. Baker, R. 1985. Biological control of plant pathogens: definitions, in M. A. Hoy and
D. C. Herzog (eds.), Biological Control in Agricultural IPM Systems. Academic Press, Inc., Orlando, FL. pp. 25-39.
Baker, K. F. and R. J. Cook. 1974. Biological Control of Plant Pathogens. W.H. Freeman and Company, Publishers, San Francisco, CA. 433 pp.

182 Biological Control of Pest Populations
Barnes, G., J. J. Kimbrough, and M. L. Wall. 1977. Cotton Insect Management Program. Ark. Agric. Ext. Serv. Leafl. 52.
Bartlett, B. R. 1964. Integration of chemical and biological control, in P. DeBach (ed.), Biological Control of Insect Pests and Weeds. Reinhold Publishing Cor- poration, New York, pp. 489-511.
Batson, W. E., Jr. 1972. Interrelationships among resistances to five major diseases and seed, seedling and plant characters in cotton. Ph.D. dissertation, Texas A&M University, College Station, TX. 125 pp.
Beirne, B. P. 1975. Biological control attempts by introductions against pest insects in the field in Canada. Can. Entomol. 107:225-236.
Bell, M. R. 1981. The potential use of microbials in Heliothis management, in W. Reed and V. Kimble (eds.), International Workshop on Heliothis Management. ICRISAT Publication, Patancheru, India. pp. 137-146.
Bell, M. R. 1983. Microbial agents, in R. L. Ridgway, E. P. Lloyd, and W. H. Cross (eds.), Cotton Insect Management with Special Reference to the Boll Weevil. USDA Agric. Handb. 589. pp. 129-151.
Benedict, J. H., L. S. Bird, and C. Liverman. 1979. Bacterial flora and MAR cottons as a boll weevil resistant character. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 228-230.
Bird, L. S. 1982. The MAR (multi-adversity resistance) system for genetic improve- ment of cotton. Plant Dis. 66: 172-176.
Bird, L. S. , C. Liverman, R, G. Percy, and D. L. Bush. 1979. The mechanism of multi-adversity resistance in cotton: theory and results. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 226- 228.
Bird, L. S. , Liverman, P. Thaxton, and R. G. Percy. 1980. Evidence that micro- organisms in and on tissues have a role in a mechanism of multiadversity resis- tance in cotton. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 283-285.
Blackeman, J. P. and N. J. Fokkema. 1982. Potential for biological control of plant disease on the phylloplane. Annu. Rev. Phytopathol. 20: 167-192.
Bush, D. L. 1980. Variation in root leachate and rhizosphere-rhizoplane microflora among cultivars representing different levels of multiadversity resistance in cot- ton. Ph.D. dissertation. Texas A&M University, College Station, TX. 271 pp.
Cate, J. R. 1985. Cotton: status and current limitations to biological control in Texas and Arkansas, in M. A. Hoy and D. C. Herzog (eds.), Biological Control in Agricultural IPM Systems. Academic Press, Inc., Orlando, FL. pp. 537-556.
Charudattan, R. 1985. The use of natural and genetically altered strains of pathogens for weed control, in M. A. Hoy and D. C. Herzog (eds.), Biological Control in Agricultural IPM Systems. Academic Press, Inc., Orlando, FL. pp. 347-372.
Charudattan, R. and H. L. Walker (eds.). 1982. Biological Control of Weeds with Plant Pathogens. John Wiley & Sons, Inc., New York, 293 pp.
Clancy, D. W. 1946. Natural enemies of some Arizona cotton insects. J . Econ. Entomol. 39:326-328.
Clancy, D. W. and H. D. Pierce. 1966. Natural enemies of some lygus bugs. J . Econ. Entomol. 59:853-858.

References 183
Clark, F. E. 1942. Experiments toward the Control of the Take-All Disease of Wheat and the Phymatotrichum Root Rot of Cotton. U.S. Dep. Agric. Tech. Bull. 835.
Clausen, C. P. (ed.). 1978. Introduced Parasites and Predators of Arthropod Pests and Weeds: A World Review. USDA Agric. Handb. 551 pp.
Cook, R. J. and K. F. Baker. 1983. The Nature and Practice of Biological Control of Plant Pathogens. The American Phytopathological Society, St. Paul, MN. 539 PP.
Curl, E. A. 1979. Effects of mycophagous collembola on Rhizoctonia solani and cotton seedling disease, in B. Schipers and W. Gams (eds.), Soilborne Plant Path- ogens. Academic Press, Inc. (London) Ltd., London. pp. 153-269.
Curl, E. A. 1982. The rhizosphere: relation to pathogen behavior and root disease. Plant Dis. 66:624-630.
Dean, D. A., W. L. Sterling, M. Nyffeler, and R. G. Breene. 1987. Foraging by selected spider predators on the cotton fleahopper and other prey. Southwest. Entomol. 12: 263-270.
DeLoach, C. J. and J. C. Peters. 1972. Effect of strip-planting vs. solid planting on predators of cotton insects in southeastern Missouri, 1969. Environ. Entomol.
Duddington, C. L. and C. H. E. Wyborn. 1972. Recent research on the nemato- phagous Hyphomycetes. Bot. Rev. 38545-565.
Ehler, L. E., K. G. Eveleens, and R. van den Bosch. 1973. An evaluation of some natural enemies of cabbage looper on cotton in California. Environ. Entomol.
Elad, Y., A. Kalfon, and I. Chet. 1982. Control of Rhizoctonia solani in cotton by seed coating with Trichoderma sp. spores. Plant Soil 66:279-281.
Ellington, J., A. G. George, H. M. Kempen, T. A. Kerby, L. Moore, B. Brooks Taylor, and L. T. Wilson (tech. coords.). 1984. Integrated Pest Management for Cotton in the Western Region of the United States. Univ. Calif. Div. Agric. Nat. Resour. Publ. 3305. 144 pp.
El-Zik, K. M., L. S. Bird, C. Liverman, P. Thaxton, G. R. Lazo, and R. G. Percy. 1983. Resistance to bacterial blight caused by treatments of symbiotic bacteria from MAR cottons. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 35-36.
El-Zik, K. M., L. S. Bird, M. Howell, and P. M. Thaxton. 1985. Symbiotic organisms associated with plant parts of multi-adversity resistance (MAR) and non-MAR cottons. Phytopathology 75: 1344.
Eveleens, K. G., R. van den Bosch, and L. E. Ehler. 1973. Secondary outbreak induction of beet armyworm by experimental insecticide applications in cotton in California. Environ. Entomol. 2:497-503.
Ewing, K. P. and H. J. Crawford. 1939. Egg parasites of the cotton flea hopper. J . Econ. Entomol. 32: 303-305.
Falcon, L. A. 1971. Microbial control as a tool in integrated control programs, in C. B. Huffaker (ed.), Biological Control. Plenum Press, New York, pp. 346-364.
Falcon, L. A., R. van den Bosch, J. Gallagher, and A. Davidson. 1971. Investigation
pp. 1-27.
1 : 94- 102.
2~1009-1015.
I

184 Biological Control of Pest Populations
of the pest status of Lygus hesperus in cotton in central California. J. Econ. Entomol. 6456-61.
Fillman, D. A. and W. L. Sterling. 1983. Killing power of the red imported fire ant (Hym.: Formicidae): a key predator of the boll weevil (Col.: Curculionidae). En- tomophaga 28~339-344.
Fillman, D. A. and W. L. Sterling. 1985. Inaction levels for the red imported fire ant, Solenopsis invicta (Hym.: Formicidae): a predator of the boll weevil, An- thonomus grandis (Col.: Curculionidae). Agric. Ecosyst. & Environ. 13:93-102.
Fletcher, R. K. and F. L. Thomas. 1943. Natural control of eggs and first instar larvae of Heliothis armigera. J. Econ. En tomol. 36557-560.
Frick, K. E. and J . M. Chandler. 1978. Augmenting the moth (Bactra verutana) in field plots for early-season suppression of purple nutsedge (Cyperus rotundus). Weed Sci. 26~703-710.
Fye, R. E. 1979. Cotton Insect Populations: Development and Impact of Predators and Other Mortality Factors. U.S. Dep. Agric. Tech. Bull. 1592. 61 pp.
Fye, R. E. and W. C. McAda. 1972. Laboratory Studies on the Development, Lon- gevity, and Fecundity of Six Lepidopterous Pests in Cotton in Arizona. U.S. Dep. Agric. Tech. Bull. 1454. 73 pp.
Garrett, S. D. 1965. Toward biological control of soil-borne plant pathogens, in K. F. Baker and W. C. Snyder (eds.), Ecology of Soil-Borne Plant Pathogens. Uni- versity of California Press, Berkeley, CA. pp. 4-17.
Ghabrial, S. A. 1980. Effects of fungal viruses on their hosts. Annu. Rev. Phyto- pathol. 18:441-461.
Gonzalez, D. and L. T. Wilson. 1982. A food-web approach to economic thresholds: a sequence of pest/predaceous arthropods on California cotton. Entomophaga
Gonzalez, D., B. R. Patterson, T. F. Leigh, and L. T. Wilson. 1982. Mites: aprimary food source for two predators in San Joaquin Valley cotton. Calif. Agric. 36: 18- 20.
Gravena, S. and W. L. Sterling. 1983. Natural predation on the cotton leafworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 76:779-784.
Hagedorn, C., W. D. Gould, and T. R. Bardinelli. 1985. Characterization of the bacterial populations associated with the cotton rhizosplane. Proc. Beltwide Cot- ton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp.
Hartstack, A. W. and W. L. Sterling. 1986. Texas Cotton Fleahopper Model: Version 2-Basic. Tex. Agric. Exp. Stn. Misc. Publ. MP 1595. 68 pp.
Hartstack, A. W. and W. L. Sterling. 1988. The Texas Cotton Insect Model-TEXCIM User’s Guide ver. 2.3 Tex. Agric. Exp. Stn. Misc. Publ. MP-1646. 37 pp.
Hartstack, A. W., J. A. Witz, and R. L. Ridgway. 1975. Suggested applications of a dynamic Heliothis model (MOTHZV-1) in pest management decision making. Proc. Beltwide Cotton Prod, Res. Conf., National Cotton Council of America, Memphis, TN. pp. 118-122.
Henneberry, T. J. and T. E. Clayton. 1982. Pink bollworm: seasonal oviposition, egg predation, and square and boll infestations in relation to cotton plant devel- opment. Environ. Entomol. 11:663-666.
27: 3 1-43.
31-32.

References 185
Henneberry, T. J. and T. E. Clayton. 1985. Consumption of pink bollworm (Lepi- doptera: Gelechiidae) and tobacco budworm (Lepidoptera: Noctuidae) eggs by some predators commonly found in cotton fields. Environ. Entomol. 14:416-419.
Herrera Aranguena, J. M. 1965. Investigations of mirids of the genus Rhinacloa, important agents in the control of Heliothis virescens on cotton. Rev. Peru. En- tomol. 8:44-60.
Hinds, W. E. 1907. Some Factors in the Natural Control of the Mexican Cotton Boll Weevil. U.S. Dep. Agric. Bur. Entomol. Bull. 74. pp. 1-79.
Hornby, D. 1983. Suppressive soils. Annu. Rev. Phytopathol. 21:65-85. Howell, C. R. 1982. Effect of Gliocladium virens on Pythium ultimum, Rhizoctonia
soluni, and dampingoff of cotton seedlings. Phytopathology 72:496-498. Howell, C. R. and R. D. Stipanovic. 1979. Control of Rhizoctonia soluni on cotton
seedlings with Pseudomonas jluorescens and with an antibiotic produced by the bacterium. Phytopathology 69:480-482.
Howell, C. R. and R. D. Stipanovic. 1980. Suppression of Pythium ultimum induced dampingoff of cotton seedlings by Pseudomonas fluorescens and its antibiotic, pyoluteorin. Phytopathology 70:7 12-7 15.
Howell, M. L., L. S. Bird, K. M. El-Zik, and P. M. Thaxton. 1987. Identification of three bacteria associated with plant tissues of Tamcot CAMD-E. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN.
Huffaker, C. B. 1959. Biological control of weeds with insects. Annu. Rev. Entomol. 4:25 1-76.
Hunter, W. D. and W. D. Pierce. 1912. The Mexican Boll Weevil: A Summary of the Investigations of This Insect up to Dee. 31, 1911. U S . Dep. Agric. Bur. Entomol. Bull. 114. 188 pp.
Irwin, M. E., R. W. Gill, and D. Gonzalez. 1974. Field-cage studies of native egg predators of the pink bollworm in southern California cotton. J . Econ. Entomol.
Jackson, R. M. 1965. Antibiosis and fungistasis of soil microorganisms, in K. F. Baker and W. C. Snyder (eds.), Ecology of Soil-Borne Plant Pathogens. Uni- versity of California Press, Berkeley, CA. pp. 363-373.
Johnson, S. J., H. N. Pitre, J. E. Powell, and W. L. Sterling. 1986. Control of Heliothis spp. by conservation and importation of natural enemies, in S . J. John- son, E. G. King, and J. R. Bradley, Jr. (eds.), Theory and Tactics of Heliothis Population Management: Cultural and Biological Control. South. Coop. Ser.
Jones, D. and W. L. Sterling. 1979. Manipulation of red imported fire ants in a trap crop for boll weevil suppression, Environ. Entomol. 8: 1073- 1077.
King, C. J., C. Hope, and E. D. Eaton. 1934. Some microbiological activities affected in manurial control of cotton root rot. J . Agric. Res. 49: 1093-1 107.
King, E. G., R. J. Coleman, J. R. Phillips, and W. A. Dickerson. 1985. Heliothis spp. and selected natural enemy populations in cotton: a comparison of three insect control programs in Arkansas (1981-1982) and North Carolina. Southwest. Entomol. Suppl. 8:71-98.
p. 35.
671193-196.
Bull. 316. pp. 132-154.

186 Biological Control of Pest Populations
Leigh, T. F. and D. Gonzalez. 1976. Field cage evaluation of predators for control of Lygus hesperus Knight on cotton. Environ. Entomol. 5:948-952.
Lingren, P. D., R. L. Ridgway, and S. L. Jones. 1968. Consumption by several common arthropod predators of eggs and larvae of two Heliothis species that attack cotton. Ann. Entomol. SOC. Am. 61518-522.
Lopez, J. D., Jr., R. L. Ridgway, and R. E. Pinnell. 1976. Comparative efficacy of four insect predators of the bollworm and tobacco budworm. Environ. Entomol.
Mankau, R. 1980. Biological control of nematode pests by natural enemies. Annu. Rev. Phytopathol. 18:415-440.
Mankau, R. 1981. Microbial control of nematodes, in B. M. Zuckerman, W. F. Mai, and R. A. Rhode (eds.), Plant Parasitic Nematodes. Vol. 111. Academic Press, Inc., New York, pp. 475-494.
McDaniel, S. G. and W. L. Sterling. 1979. Predator determination and efficiency on Heliothis virescens eggs in cotton using 32 p. Environ. Entomol. 8: 1083-1087.
McDaniel, S. G. and W. L. Sterling. 1982. Predation of Heliothis virescens (F.) eggs on cotton in east Texas. Environ. Entomol. 11:60-66.
McDaniel, S. G . , W. L. Sterling, and D. A. Dean. 1981. Predators of tobacco bud- worm larvae in Texas cotton. Southwest. Entomol. 6: 102- 108.
Miller, J. H., H. M. Kempen, K. M. El-Zik, D. W. Cudney, B. P. Fischer, and P. Keeley. 1977. Weed Control in Cotton. Univ. Calif. Div. Agric. Sci. Leafl. 2991.
Morris, R. F. 1965. Contemporaneous mortality factors in population dynamics. Can. Entomol. 97:1173-1184.
Mussett, K. S. , J . H. Young, R. G. Price, and R. D. Momson. 1979. Predatory arthropods and their relationship to fleahoppers on Heliothis-resistant cotton va- rieties in southwestern Oklahoma. Southwest. Entomol. 4:35-39.
National Academy of Sciences. 1975. Pest Control: An Assessment of Alternative Technologies. Vol. 3. Cotton Pest Control. National Academy of Sciences, Wash- ington, DC. 139 pp.
Orphanides, G. M., D. Gonzalez, and B. R. Bartlett. 1971. Identification and eval- uation of pink bollworm predators in southern California. J . Econ. Entomol.
Orr, C. C. 1981. Nothanguina phyllobiu, a nematode biocontrol of silverleaf night- shade. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of Amer- ica, Memphis, TN. pp. 36-37.
Papavizas, G. C. and R. D. Lumsden. 1980. Biological control of soilborne fungal propagules. Annu. Rev. Phytopathol. 18:389-413.
Perkins, P. V. and T. F. Watson. 1972. Nabis alternatus as a predator of Lygus hesperus. Ann. Entomol. SOC. Am. 65:625-629.
Reinhard, H. J. 1926. The Cotton Fleahopper. Tex. Agric. Exp. Stn. Bull. B-339.
Ridgway, R. L. and S. L. Jones. 1968. Field-cage releases of Chrysopa carnea for suppression of populations of the bollworm and the tobacco budworm on cotton. J . Econ. Entomol. 61:892-898.
5:1160-1164.
19 PP.
64 142 1-424.
32 PP.

References 187
Ridgway, R. L. and P. D. Lingren. 1972. Predaceous and parasitic arthropods as regulators of Heliothis populations, in Distribution Abundance and Control of Heliothis Species in Cotton and Other Host Plants. South. Coop. Ser. Bull, 169.
Ridgway, R. L. and R. K. Morrison. 1985. Worldwide perspective on practical uti- lization of Trichogramma with special reference to control of Heliothis on cotton. Southwest. Entomol. Suppl. 8:190-198.
Ridgway, R. L. and S. B. Vinson. 1976. Biological Control by Augmentation of Natural Enemies. Plenum Press. New York. 480 pp.
Ridgway, R. L., R. K. Morrison, and R. E. Kinzer. 1973. Programmed releases of parasites and predators for control of Heliothis spp. on cotton. Proc. Beltwide Cotton Res. Conf., National Cotton Council of America, Memphis, TN. pp. 92- 94.
Roach, S. H., J. W. Smith, S. B. Vinson, H. M. Graham, and J. A. Harding. 1979. Sampling predators and parasites of Heliothis species on crops and native host plants, in W. L. Sterling (ed.), Economic Thresholds and Sampling of Heliothis Species on Cotton, Corn, Soybeans and Other Host Plants. South. Coop. Ser.
Roncadori, R. W. and R. S. Hussey. 1980. The role of vesicular-buscular mycor- rhizae in rootknot nematode infected cotton. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 26-27.
Royama, T. 1981. Evaluation of mortality factors in insect life table analysis. Ecol. Monogr. 5 1 :495-505.
Shipton, P. J. 1977. Nonoculture and soilborne plant pathogens. Annu. Rev. Phy- topathol. 15:387-407.
Simmonds, F. J . 1948. Some difficulties in determining by means of field samples the true value of parasitic control. Bull. Entomol. Res. 39:435-440.
Smith, R. F. and R. van den Bosch. 1967. Integrated control, in W. W. Kilgore and R. L. Doutt (eds.), Pest Control: Biological, Physical, and Selected Chemical Methods. Academic Press, Inc., New York. pp. 295-340.
Smith, G. S., R. W. Roncadori, and R. S. Hussey. 1986. Interaction of endomy- corrhizal fungi, superphosphate, and Meloidogyne incognita on cotton in micro- plot and field studies. J . Nematol. 18:208-216.
Sterling, W. L. 1978. Fortuitous biological suppression of the boll weevil by the red imported fire ant. Environ. Entomol. 7564-568.
Sterling, W. L. 1983. Predaceous insects and spiders, in Bohmfalk et al. (eds.), Identification, Biology and Sampling of Cotton Insects. Tex. Agric. Ext. Serv.
Sterling, W. L. 1984. Action and Inaction Levels in Pest Management. Tex. Agric. Exp. Stn. Bull. B-1480. 20 pp.
Sterling, W. L., D. A. Dean, D. A. Fillman, and D. Jones. 1984. Naturally-occurring biological control of the boll weevil (Col.: Curculionidae). Entomophaga
Stimac, J. L. and R. J. O’Neil. 1985. Integrating influences of natural enemies into models of croplpest systems, in M. A. Hoy and D. C. Herzog (eds.), Biological Control in Agricultural IPM Systems. Academic Press, Inc., Orlando, FL.
pp. 48-56.
Bull. 231. pp. 133-145.
Bull. 933. pp. 25-31.
29: 1-9.

188 Biological Control of Pest Populations
Stinner, R. E. 1977. Efficacy of inundative releases. Annu. Rev. Entomol. 22:515- 531.
Sturm, M. M. and W. L. Sterling. 1986. Assessment of boll weevil mortality factors within flower buds of cotton. Bull. Entomol. Soc. Am. 32:239-247.
Tamaki, G., J. U. McGuire, and J. E. Turner. 1974. Predator power and efficacy: a model to evaluate their impact. Environ. Entomol. 3:625-630.
TeBeest, D. 0. 1981. Biological control of weeds in cotton with fungal plant path- ogens. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of Amer- ica, Memphis, TN. 36 pp.
Templeton, G. E. 1982. Biological herbicides: discovery, development, deployment. Weed Sei. 30:430-433.
Templeton, G. E., D. 0. TeBeest, and R. J. Smith, Jr. 1979. Biological weed control with mycoherbicides. Annu. Rev. Phytopathol. 17:301-310.
Trichilo, P. 1986. Influence of the host plant on the interaction of spider mites with their natural enemies in a cotton agroecosystem. Ph.D. dissertation. University of California, Davis, CA.
Tsai, A. H. Y. and L. S. Bird. 1975. Microbiology of host-pathogen interactions for resistance to seedling disease and multiadversity resistance in cotton. Proc. Belt- wide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis,
van den Bosch, R. 1978. The Pesticide Conspiracy. Doubleday & Company, Inc., New York. 226 pp.
van den Bosch, R. and K. S. Hagen. 1966. Predaceous and Parasitic Arthropods in California Cotton Fields. Calif. Agric. Exp. Stn. Bull. 820. 32 pp.
van den Bosch, R., T. F. Leigh, D. Gonzalez, and R. E. Stinner. 1969. Cage studies on predators of the bollworm in cotton. J . Econ. Entomol. 62:1486-1489.
Van Gundy, S. D. 1972. Nonchemical control of nematodes and root infecting fungi, in Pest Control Strategies for the Future, National Academy of Sciences, Wash- ington, DC. 317 pp.
Walker, H. L. 1980. Spurred anoda (Anoda cristata (L.) Schlecht) bicontrol with a plant pathogen. Proc. South. Weed Sci. Soc. 33:65.
Wheeler, A. G., Jr. 1976. Lygus bugs as facultative predators, in D. R. Scott and L. E. O'Keeffee (eds.), Lygus Bug: Host Plant Interactions. Proc. Workshop 15th Int. Congr. Entomol. University of Idaho Press, Moscow, ID. pp. 28-33.
Whitcomb, W. H. 1967. Field studies on predators of the second-instar bollworm, Heliothis zea (Boddie) (Lepidoptera: Noctuidae). J . Ga. Entomol. Soc. 2: 113- 118.
Whitcomb, W. H. and K. Bell. 1964. Predaceous Insects, Spiders and Mites of Arkansas Cotton Fields. Ark. Agric. Exp. Stn. Bull. 690. 84 pp.
Wiggins, E. A. and E. A. Curl. 1979. Interactions of collembola and microflora of cotton rhizosphere. Phytopathology 69:244-249.
Wilson, L. T. 1985. Estimating the abundance and impact of arthropod natural ene- mies in IPM systems, in M. A. Hoy and D. C. Herzog (eds.), Biological Control in Agricultural IPM Systems. Academic Press, Inc., Orlando, FL. pp. 303-322.
Wilson, L. T. 1986. Developing economic thresholds in cotton, in R. Frisbie and P.
I
TN. pp. 39-45.

References 189
L. Adkisson (eds.), CZPM, Integrated Pest Management on Major Agricultural Systems. Tex. Agric. Exp. Stn. MP 1616. pp. 308-344.
Wilson, L. T. and A. L. Bishop. 1982. Responses of Deltapine 16 cotton, Gossypium hirsutum L. to simulated attack by known populations of Heliothis larvae (Lep- idoptera: Noctuidae) in a field experiment in Queensland, Australia. Prot. Ecol.
Wilson, L. T. and A. P. Gutierrez. 1980. Within-plant distribution of predators on cotton: comments on sampling and predator efficiencies. Hilgardia 48:3-11.
Wilson, L. T., A. P. Gutierrez, and T. F. Leigh. 1980. Within-plant distribution of the immatures of Heliothis zea (Boddie) on cotton. Hilgardia 48: 12-23.
Wilson, L. T., D. Gonzalez, T. F. Leigh, V. Maggi, C. Foristiere, and P. Goodell. 1983. The within-plant distribution of spider mites (Acari: Tetranychidae) on cot- ton: a developing implementable monitoring program. Environ. Entomol. 12: 128- 134.
Wilson, L. T., D. Gonzalez, and R. Plant. 1985. Predicting sampling frequency and economic status of spider mites on cotton. Proc. Beltwide Cotton Prod. Res. Conf., National Cotton Council of America, Memphis, TN. pp. 168-170.
Windels, C. E. and S. E. Lindow. 1985. Biological Control on the Phylloplane. The American Phytopathological Society, St. Paul, MN. 169 pp.
Yin, S . Y., D. C. Keng, K. Y. Yang, and D. Cheu. 1957. A further study on the biological control of verticillium wilt of cotton. Acta Phytopathol. Sin. 3:55-61.
Yin, S . Y., J. K. Chang, and P. C. Xun. 1965. Studies in mechanisms of antagonistic fertilizer "5406." IV. The distribution of the antagonist in soil and its influence on the rhizosphere. Acta Microbiol. Sin. 11:259-288.
4~371-380.


















