
Biochemistry and molecular biology oi galactoside transport and
10
Lait (1993) 73, 87-96 © Elsevier/INRA Metabolim - Regulation Biochemistry and molecular biology oi galactoside transport and metabolism in lactic acid bacteria B Poolman Department of Microbiology, University of Groningen, Kerklaan 30 9NN 9751 Haren, The Netherlands Summary - Until a few years ago the description of the pathways of transport and metabolism of galactosides in lactic acid bacteria was mainly phenomenological. Different transport and enzymatic activities had been detected but the individual components (proteins, enzymes) were largely un- known and the genes had not yet been isolated. In this paper 1will address the different transport mechanisms and metabolic pathways of galactosides in factic acid bacteria, and indicate the melee- ular level at which these studies have evolved. The genes encoding galactosidetransport ATPases ion Iinked galactoside transporters and phosphoenolpyruvate:galactoside phosphotransferase sys- tems (PTS) have been isolated and characterized from a number of lactic acid bacteria. In addition, the genes encoding subsequent steps of lactose (galactose) metabolism, ie those encoding the Le- loir and Tagatose 6 phosphate pathway enzymes, have been characterized at a genetic and to some extent at a biochemical javel. The genes encoding the transport proteins and metabolic en- zymes are often clustered in a single operon or regulon which allows co-ordinate expression of the polypeptides. Transport of galactosides has most been thoroughly studied in Streptococcus thermo- phi/us where the transport gene encodes a chimeric protein (LacS) that is composed of a carrier do- main homofogous to various ion Iinked carbohydrate transporters and a hydrophilic domain hornolo- gous to the liA phosphoryl transfer protein of various PTSs. LacS has been extracted from membrane vesicles and functionally reconstituted into proteoliposomes, and the energetics and ki- netic mechanism of transport have been analyzed. lacS mediates galactoside H+ symport and ga- lactoside exchange (eg lactose/galactose exchange) of which the latter reaction is highly favoured under physiological conditions. Details of the galactoside translocation pathway have been estab- Iished by studying mutant LacS proteins in which the galactoside movement is uncoupled from the H+ transport whereas the exchange reaction takes place normally. The data offer a rationale for the observed excretion of galactose by Gal- and Galt strains of S thermophilus under conditions of growth in the presence of excess lactose. sugar transport ATPase / ion-Iinked transport / exchange / PTS / galactoside metabolism Résumé - Biochimie et biologie moléculaire du transport et du métabolisme des galacto- sides chez les bactéries lactiques. 1/ y a quelques années encore, la description des voies de transport et du métabolisme des galactosides chez les bactéries lactiques était principalement phé- noménologique. Des systèmes de transport et des activités enzymatiques différentes avaient été dé- tectés, mais leurs composants individuels (protéines, enzymes) étaient pour la plupart inconnus et 87
Transcript of Biochemistry and molecular biology oi galactoside transport and

Lait (1993) 73, 87-96© Elsevier/INRA
Metabolim - Regulation
Biochemistry and molecular biologyoi galactoside transport and metabolism
in lactic acid bacteria
B Poolman
Department of Microbiology, University of Groningen, Kerklaan 30 9NN 9751 Haren,The Netherlands
Summary - Until a few years ago the description of the pathways of transport and metabolism ofgalactosides in lactic acid bacteria was mainly phenomenological. Different transport and enzymaticactivities had been detected but the individual components (proteins, enzymes) were largely un-known and the genes had not yet been isolated. In this paper 1will address the different transportmechanisms and metabolic pathways of galactosides in factic acid bacteria, and indicate the melee-ular level at which these studies have evolved. The genes encoding galactosidetransport ATPasesion Iinked galactoside transporters and phosphoenolpyruvate:galactoside phosphotransferase sys-tems (PTS) have been isolated and characterized from a number of lactic acid bacteria. In addition,the genes encoding subsequent steps of lactose (galactose) metabolism, ie those encoding the Le-loir and Tagatose 6 phosphate pathway enzymes, have been characterized at a genetic and tosome extent at a biochemical javel. The genes encoding the transport proteins and metabolic en-zymes are often clustered in a single operon or regulon which allows co-ordinate expression of thepolypeptides. Transport of galactosides has most been thoroughly studied in Streptococcus thermo-phi/us where the transport gene encodes a chimeric protein (LacS) that is composed of a carrier do-main homofogous to various ion Iinked carbohydrate transporters and a hydrophilic domain hornolo-gous to the liA phosphoryl transfer protein of various PTSs. LacS has been extracted frommembrane vesicles and functionally reconstituted into proteoliposomes, and the energetics and ki-netic mechanism of transport have been analyzed. lacS mediates galactoside H+ symport and ga-lactoside exchange (eg lactose/galactose exchange) of which the latter reaction is highly favouredunder physiological conditions. Details of the galactoside translocation pathway have been estab-Iished by studying mutant LacS proteins in which the galactoside movement is uncoupled from theH+ transport whereas the exchange reaction takes place normally. The data offer a rationale for theobserved excretion of galactose by Gal- and Galt strains of S thermophilus under conditions ofgrowth in the presence of excess lactose.
sugar transport ATPase / ion-Iinked transport / exchange / PTS / galactoside metabolism
Résumé - Biochimie et biologie moléculaire du transport et du métabolisme des galacto-sides chez les bactéries lactiques. 1/ y a quelques années encore, la description des voies detransport et du métabolisme des galactosides chez les bactéries lactiques était principalement phé-noménologique. Des systèmes de transport et des activités enzymatiques différentes avaient été dé-tectés, mais leurs composants individuels (protéines, enzymes) étaient pour la plupart inconnus et
87

88 B Poolman
les gènes n'avaient pas été isolés. Dans cet article, je parlerai des différents mécanismes de trans-port et des voies métaboliques des galactosides des bactéries lactiques, et j'indiquerai le niveau mo-léculaire auquel ces études ont abouti. Les gènes codant pour les A TPases du transport des galacto-sides, les transporteurs ions-dépendants et les systèmes phosphoénolpyruvate:galactosidephosphotransférase (PTS) de nombreuses bactéries lactiques ont été isolés et caractérisés. De plus,les gènes codant pour les étapes suivantes du métabolisme du lactose (galactose), c'est-à-dire ceuxcodant pour les enzymes des voies de Leloir et du tagatose-6-phosphate, ont été caractérisés au ni-veau génétique et, dans une certaine mesure, au niveau biochimique. Les gènes codant pour les pro-téines de transport et les enzymes du métabolisme sont souvent groupés en un opéron ou régulonunique, ce qui permet leur expression coordonnée. Le transport des galactosides a été étudié demanière approfondie chez Streptococcus thermophilus où les gènes de transport codent pour uneprotéine-chimère (LacS) constituée d'un domaine carrier homologue à des transporteurs de glucidesions-dépendants variés, et un domaine hydrophile homologue à la protéine phosphotransférase liAde différents systèmes PTS. LacS a été extraite de vésicules membranaires et fonctionnel/ement re-constituée dans des protéoliposomes, et les mécanismes énergétiques et cinétiques du transport ontété analysés. LacS intervient dans le symport galactoside H+ et dans l'échange des galactosides(c'est-à-dire l'échange lactose/galactose), ce demier étant hautement favorisé dans les conditionsphysiologiques. Les détails de la voie de translocation des galactosides ont été établis en étudiant lesprotéines mutantes LacS dans lesquel/es le mouvement des galactosides est découplé du transportdes ions H+ alors que la réaction d'échange se déroule normalement. Les données offrent une expli-cation pour l'excrétion de galactose observée dans les souches GaJ+ et eer de S thermophilus dansdes conditions de croissance en présence d'un excès de lactose.
ATPase du transport des glucides / transport ions-dépendant / échange / PTS / métabolismedes galactosides
INTRODUCTION
The initial event in the metabolism of mono-and disaccharides involves the transloca-tion of these molecules across the cyto-plasmic membrane. The systems by whichthe carbohydrate molecules are transport-ed can be subdivided into 3 classes thatdiffer in their mechanism of energycoupling: i) in case of primary transport thetranslocation of a sugar is driven by thehydrolysis of an energy-rich chemical bond(eg ATP; fig 1); ii) for secondary transportthe driving force for the translocation of asugar molecuJe is supplied by the sugarconcentration gradient, and, if anothermolecuJeis co- or counter transported withthe carbohydrate, the (electro-) chemicalgradient of this coupling molecuJe,ie.accu-mulation is achieved by the downhill move-ment of another molecule (fig 2); Hi) forgroup translocation transport the translo-cation of the sugar molecule across the cy-
toplasmic membrane is coupied to thechemical modification of the molecule, ietransport concomitant with phosphorylationof the substrate by the PTS (fig 3). Thesedifferent mechanisms of transport havebeen observed for a wide variety of sugarsbut only those that mediate lactose (galac-toside) transport in lactic acid bacteria willbe described.
Depending on the transport mechanismthe following step in lactose metabolism in-volves hydrolysis of the free sugar by B-galactosidase (LacZ), yielding glucose andgalactose (fig 2), or hydrolysis of lactose 6-phosphate by a phospho-B-galactosidase(LacG), yielding glucose and galactose 6-phosphate (fig 3). The 2 major pathways ofglucose degradation in lactic acid bacteriaare the glycolytic (or Embden-Meyerhof-Parnas) and the phosphoketolase path-ways (KandJer, 1983). The glycolyticpathway is operative in Lactococcus spp,homofermentative lactobacilli (eg Lactoba-
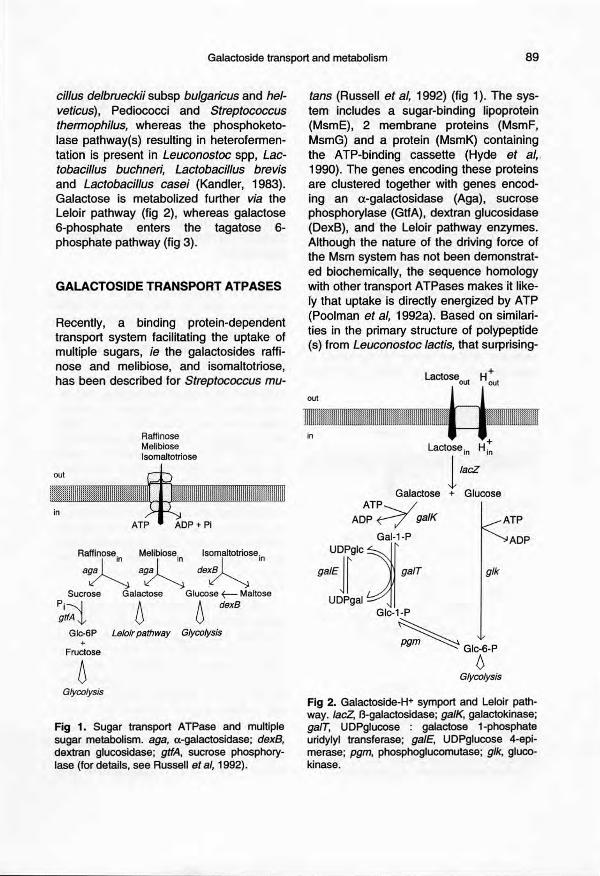
Galactoside transport and metabolism 89
cillus delbrueckii subsp bulgaricus and hel-veticus), Pediococci and Streptococcusthermophilus, whereas the phosphoketo-lase pathway(s) resulting in heterofermen-tation is present in Leuconostoc spp, Lac-tobacillus buchneri, Lactobacillus brevisand Lactobacillus casei (Kandler, 1983).Galactose is metabolized further via theLeloir pathway (fig 2), whereas galactose6-phosphate enters the tagatose 6-phosphate pathway (fig 3).
GALACTOSIDE TRANSPORT ATPASES
Recently, a binding protein-dependenttransport system facilitating the uptake ofmultiple sugars, ie the galactosides raffi-nose and melibiose, and isomaltotriose,has been described for Streptococcus mu-
RaffinoseMelibioseIsomaltotriose
ATP ADP + Pi
Raffinose Melibiose Isomaltotriose
a~a~ de~in
Sucrase Galactose Glucose f-- MaltosePi~ A dexBgtfA -i ()
Glc-6P Leloir pathway Glycolysis6
+Fructose
6Glycolysis
Fig 1. Sugar transport ATPase and multiplesugar metabolism. aga, a-galactosidase; dexB,dextran glucosidase; gtfA, sucrose phosphory-lase (for details, see Russell et al, 1992).
tans (Russell et al, 1992) (fig 1). The sys-tem includes a sugar-binding lipoprotein(MsmE), 2 membrane proteins (MsmF,MsmG) and a protein (MsmK) containingthe ATP-binding cassette (Hyde et al"1990). The genes encoding these proteinsare clustered together with genes encod-ing an a-galactosidase (Aga), sucrosephosphorylase (GtfA), dextran glucosidase(DexB), and the Leloir pathway enzymes.Although the nature of the driving force ofthe Msm system has not been demonstrat-ed biochemically, the sequence homologywith other transport ATPases makes it Iike-Iy that uptake is directly energized by ATP(Poolman et al, 1992a). Based on similari-ties in the primary structure of polypeptide(s) from Leuconostoc tectis, that surprising-
Lactose H+ln in
l-+ GlucoseGalactose
ATP-JADP~ galK
Gal·1·P
UDP9IC)gaiE Jf galT
UDPgalGlc-1-P
~GIC-6-P
6Glycolysis
ATP
ADP
glk
Fig 2. Galactoside-H+ symport and Leloir path-way. laeZ, B-galactosidase; galK, galactokinase;galT, UDPglucose : galactose 1-phosphateuridylyl transferase; gaIE, UDPglucose 4-epi-merase; pgm, phosphoglucomutase; glk, gluco-kinase.

90
Iy complement an Escherichia coli lacYmutation, and components of binding pro-tein dependent transporters (David, 1992)it is tempting to speculate that a lactosetransport ATPase is also present in this or-ganism. The number of ATP molecules hy-drolyzed per solute taken up by the trans-port ATPases is most Iikely 1-2 (Poolmanet al, 1992a) which makes these transport-ers energetically expensive as comparedto the ion-linked transporters, exchangesystems and PTS (see below). Generallythe binding protein dependent transportersbind their substrates with very high affinitywhich allows efficient transport in thenano- and submicromolar range of con-centrations.
Downstream of the putative lactosetransport ATPase genes of Lc lsctis, 2'translationally coupied genes (IacL and
Lactose
liBin HPr-P-( liA
EI~ rllA-pPEP-( HPr Lac-6P
rEI-pPyruvate l,acG
Gal-6P + Glucose
laeAS II glkkTP
Tag-6P G1c-6P ADPlaeC ATP~ 1~ ATP
ADP ~ t '~r
TDP FDp~ADPlaeD Lz DHAP)-l
G3P
6Glycolysis
Fig 3. Lactose-PTS and tagatose 6-phosphatepathway. lacG, phospho-B-galactosidase;lacAS, galactose 6-phosphate isomerase; lace,tagatose 6-phosphate kinase; lacO, tagatose1,6-diphosphate aldolase; glk, glucokinase.
B Poolman
lacM) have been found that encode a func-tional B-galactosidase (David et al, 1992).The deduced amino acid sequences ofLacL and LacM are homologous to theamino- and carboxy-terminal parts, respec-tively, of the B-galactosidases (LacZ) of Sthermophilus, Lb bulgaricus and E coli.
ION-LiNKED GALACTOSIDETRANSPORTERS AND GALACTOSIDEEXCHANGE
lon-linked galactoside transport has firstbeen described for the atypical L lactis7962 where methyl-B-D-galactopyranoside(TMG) accumulation has been shown tobe coupied to the downhill movement of aproton (Kashket and Wilson, 1973). Thesystem is highly specific for galactose,TMG and other galactose analogs, but ex-hibits poor affinity for lactose. A similarsystem has been described for Lc lactisML3 (Thompson, 1980). Lactose transport-ers driven by the electrochemical protongradient have been described for S ther-mophilus and Lb bulgaricus (Poolman etal, 1989; unpublished results). These lac-tose transporters turn out to be specific notonly for lactose (~-galactoside) but al50 formelibiose (a-galactoside), galactose (mono-saccharide) and to a lesser extent raffi-nose (trisaccharide) (Poolman et al,1992b).
The genes encoding the lactose (galac-toside) transport proteins (LacS) of S ther-mophilus and Lb bulgaricus have beenc1oned, characterized and functionally ex-pressed in E coli (Poolman et al, 1989,1992b). Both proteins are composed of anamino-terminal polytopic membrane pro-tein domain (carrier domain) and a car-boxy-terminal hydrophilic domain. The car-rier domain is homologous to the melibiosecarrier protein (MeIS) of E coli, the xylosecarrier protein (XyIP) of Lactobacillus pen-

Galactoside transport and metabolism 91
tosus, and some other carrier proteins thatmost Iikely couple the uphill movement ofsugars to the downhill movement of a cat-ion (proton, sodium or lithium). The hydro-philic domain of LacS is homologous tovarious liA protein(s) or protein domains ofPTS, previously indicated as enzyme III orenzyme 1I1-likedomain. The chimeric na-ture of LacS makes this transporter uniqueand suggests that it shares features ofboth ion-Iinked transporters and PTS.
Studies on the kinetic mechanism ofLacS have indicated that a full transloca-tion cycle involves stoichiometric trans-membrane movement of galactosides andprotons (fig 4). On the other hand, with sat-urating concentrations of galactosides (eglactose and galactose) on either side of themembrane, release of substrate (and pro-ton) is immediately followed by (re)bindingof substrate (and proton) rather than by re-orientation of the 'empty' binding site(s)(Foucaud and Poolman, 1992). Underthese conditions the carrier protein per-forms an exchange reaction with no netproton translocation. Since S thermophilusand Lb bulgaricus strains excrete galac-tose into the medium in amounts stoichio-metric with the uptake of lactose, the sug-gestion has been made that underphysiological conditions lactose is taken upin exchange for a galactose molecule(Poolman et al, 1989; Poolman, 1990). Evi-dence in favour of this notion has been ob-tained from kinetic studies of galactosidetransport (Foucaud and Poolman, 1992)and comparison of in vivo transport andmetabolism rates (Poolman and Foucaud,unpublished). The recent suggestion thatthe galactoside transporter of S thermophi-lus is a strict lactose/galactose antiporter(Hutkins and Ponne, 1991) is not correct.Although the lactose/galactose exchangereaction may be favoured under many con-ditions, the exchange mode simply reflectspartial steps, forward and backward reac-tions with no net proton translocation, of a
complete translocation cycle involving sug-ar and proton uptake on one side and re-lease on the other side of the membrane,and reorientation of loaded and unloadedsubstrate binding sites (Foucaud and Pool-man, 1992; see below). Attempts are beingmade to refine the kinetic scheme for LacSfacilitated transport by using mutants[LacS(H379D) and LacS(E379Q)] that areconditionally uncoupled in the galactoside-H+symport reaction.
The exchange reaction has advantagesover galactoside-H+ symport for the uptakeof lactose and excretion of galactose. First,although uptake of lactose is stimulated bythe transmembrane electrochemical protongradient (àp), efflux is inhibited and netexit will only occur if the galactose concen-tration gradient (LlJlgatlF)exceeds the Llp.
out 1 in
C-, 5 ,C- 1
H1~tLiC~H~'flrCLT2 2 1 +
Li'" J-7 Hi
HLC" 3 >, CLHI1
membrane
4
Fig 4. Kinetic scheme for LacS- mediated galac-toside-Hr symport and galactoside exchange. C,carrier protein; L, ligand (galactoside); H+, pro-ton. In the model ordered binding and release ofsubstrate and proton (substrate first, proton last(bold type letters), and proton first, galactosidelast) on the inner surface of the membrane areindicated. The order of binding and release onthe outer surface of the membrane are not spec-ified. The negative charge carried by the unload-ed carried and binary carried-Iigand complex isindicated (for details, see Foucaud and Pool-man, 1992).

92
Instead the linkage of lactose uptake togalactose excretion via an exchange reac-tion assures tight coupling to the metabo-lism, which minimizes (excessive) galac-tose accumulation and possible productinhibition (Poolman, 1990). Second, effluxis significantly rate-Iimited by the releaseof the proton at the outer surface of themembrane, and, as a consequence, ex-tremely slow below pH 7; similarly, influx isinhibited by a low intracellular pH whereasexchange is less drastically affected by theabsolute H+ concentrations (Foucaud andPoolman, 1992). In fact, exchange pro-ceeds maximally between pH 6-7 and withrates at least one order of magnitude fast-er than the corresponding galactoside-H+symport.
Carboxy-terminal deletion mutants ofLacS have been constructed which lackpart or the entire liA domain (approximate-Iy 170 amino acids) (Pool man and Nieu-wenhuis, unpublished observations). Stud-ies with LacS(~ 166) and other mutantsindicate that the basic properties of trans-port are retained in these mutants. In thepresence of PEP, and purified enzyme 1and HPr, the general phosphoryl transferproteins of the PTS, the liA domain ofLacS can be phosphorylated at a specifichistidine residue (His-552) (Poolman et al,1992b). Currently, the kinetic mechanismof 'phosphorylated' LacS is compared withthat of the unphosphorylated protein bothin vitro and in vivo.
Galactoside (TMG) transport has alsobeen studied in the heterofermentative lac-tobacilli Lb brevis and Lb buchneri (Roma-no et al, 1987). In the presence of argi-nine as exogenous energy source TMG isaccumulated by these cells most likely viaa galactoside-H+ symport mechanism. Ad-dition of glucose results in a rapid efflux ofTMG, and it has been suggested thatglu-cose converts the ion-linked transporterinto an uniport mechanism (Romano et al,1987). Since addition of glucose also elic-
B Poolman
its the phosphorylation of the energycoupling proteins of the PTS, it is conceiv-able that phosphorylation of a domain simi-lar to liA in LacS mediates the apparentconversion in transport mechanism.
The lactose transport genes (laCS) of Sthermophilus and Lb bulgaricus are orga-nized in an operon that also contains theB-galactosidase gene (/acZ) (Poolman etal, 1989; Schmidt et al, 1989). Down-stream of the lac operon of Lb bulgaricus,an open reading frame (ORF) has beenfound that shows similarity to various re-pressor proteins, including Lacl, GaIR, andPurR (Yoast et al, 1990). It has been sug-gested that this ORF does not encode afunctional repressor protein which wouldexplain the constitutive expression of thelac genes in Lb bulgaricus. In contrast, ex-pression of the lac genes in S thermophi-lus is inducible but the regulatory protein(s)have not yet been identified. Induction ofthe lac (and ga~ genes is effected by ga-lactose and galactose analogs, but not bylactose, whereas glucose represses (Pool-man et al, 1990; Foucaud and Poolman,unpublished observations). Sequencesstrongly resembling those of catabolite-repressed genes of Gram-positive bacteriahave been detected in the lac promoter re-gion of S thermophilus, suggesting that ex-pression of the lac genes is also under glu-cose (catabolite) repression control.
PEP:GALACTOSIDE PHOSPHOTRANS-FERASE SYSTEMS (PTS)
The genes encoding IISC (or lactose spe-cific enzyme Il) and liA (previously, factorIIILac or enzyme III-lac) components of thelactose PTS as weil as the phospho-B-galactosidase of L lactis and Lb casei havebeen cloned and sequenced (Boizet et al,1988; Porter and Chassy, 1988; Alpert andChassy, 1988, 1990; de Vos et al, 1990).

Galactoside transport and metabolism
The genes are contained in an operonstructure which has best been studied for Llactis. The lactose operon of L lactis alsocomprises the tagatose 6-phosphate path-way genes (van Rooyen et al, 1991; seebelow). The expression of the lac genes ofL lactis and Lb casei is induced duringgrowth on lactose and galactose. A repres-sor protein (LacR) acting at the level oftranscription mediates the expression ofthe lac genes in L lactis (van Rooyen et al,1991; 1992); tagatose 6-phosphate mostlikely functions as the effector molecule (in-ducer). LaeR belongs to the family of DNA-binding proteins that includes the E coli re-pressors DeoR, FucR and GutR, and LaeRof Staphylococcus aureus.
Sugar transport mediated by the PTS in-volves phosphoryl transfer from PEP viathe general cytoplasmic components en-zyme 1 and HPr to the sugar specifie com-ponent(s) IIABC. The phosphoenzyme in-termediates of enzyme l, HPr and liA of ailPTS studied to date carry the phosphorylgroup on a histidine residue (see Pas andRobillard, 1988). In addition to the histi-dine, HPr in Gram-positive bacteria carriesa specifie serine residue that is phosphory-lated by an ATP dependent protein kinase(Reizer et al, 1985). This phosphorylationsite does not play a role in the phosphoryltransfer per se but modulates the PTS ac-tivity. Biochemically it has been demon-strated that phosphorylation of liB-mannitolof E coli is at a specifie cysteine residue(Pas and Robillard, 1988). This phosphory-lation site actually transfers the phosphorylgroup to the sugar that is translocated byIIC. By replacing each of the histidine andcysteine residues of IIBC of the lactosePTS of Lb casei evidence has been ob-tained that also a single cysteine (Cys-384) mediates the final phosphoryl transferto lactose (AIpert and Chassy, 1990).
ln additionto catalyzing galactoside up-take concomitant with phosphorylation ofthe sugar, IIBC of the lactose PTS is also
93
believed to catalyze efflux (expulsion) of in-tracellularly formed free galactosides(Reizer et al, 1985; Thompson, 1987). Thisexpulsion of galactosides is phenomeno-logically similar to that observed in the het-erofermentative lactobacilli where a secon-dary transport mechanism has beenimplicated in the efflux of free galactosides(see above, ion-Iinked galactoside trans-porters and galactoside exchange). For de-tails on galactoside efflux in lactic acid(and other Gram-positive) bacteria thereader is referred to Reizer (1989).
LELOIR PATHWAY
The enzymes of the Leloir pathway of ga-lactose metabolism include galactokinase(GaIK) which phosphorylates galactose atthe C-1 position, UDPglucose:galactose-1-phosphate uridylyltransferase (GaIT) whichtransfers the UDP group from UDPglucoseto galactose-1-phosphate, UDPglucose 4-epimerase (GaiE) which catalyzes the epi-merization of UDPgalactose to UDPglu-cose (fig 2). UDPglucose synthetase(GaIU, not shown) and phosphoglucomu-tase (Pgm) are also needed for galactosemetabolism but the corresponding genesdo not form part of the gal operon. In E colithese enzymes are synthesized constitu-tively (Adhya, 1987). The reversible reac-tions of the Leloir pathway allow cells tomake UDPgalactose in the absence of ex-ogenous galactose which can be used forthe synthesis of complex (extracellular)polysaccharides.
The genes encoding GalT and GaiE ofS thermophilus (Poolman et al, 1990; un-published observations), and GalK andGalT of Lactobacillus helveticus (Molletand Pilloud, 1991) have been isolated andcharacterized. The primary structures ofGaiE from S thermophilus, and GalK fromLb helveticus show significant similarity to

94
the corresponding polypeptides of otherpro- and eukaryotic organisms. The de-duced amino acid sequences of GalT fromLb helveticus and S thermophilus indicatethat, so far, this enzyme is unique for lacticacid bacteria (Mollet and Pilloud, 1991;Poolman, unpublished observations). In-terestingly, a gene encoding an aldose1-epimerase (mutarotase) (GaIM) is asso-ciated with the gal and lac gene cluster ofS thermophilus and Lb helveticus. GalMcatalyzes the anomeric interconversion ofD-glucose and other aldoses, a process(mutarotation) that also occurs spontane-ously (Poolman et al, 1990). The role ofGalM in galactoside metabolism in the lac-tic acid bacteria is unclear, but its pres-ence suggests that one or more steps inthe carbohydrate metabolism requires ei-ther an (X- or B-sugar anomer.
The majority of 'wild type' strains ofS thermophilus (and Lb bulgaricus) cannotmetabolize galactose, and- in case ofS thermophilus the Gal- phenotype hasbeen attributed to a defect in the inductionmechanism for galactokinase (Thomasand Crow, 1984; Hutkins et al, 1985). Un-der the selective pressure of the chemo-stat Gal+ variants of S thermophilus couldbe isolated indicating that galK is presentin the organisms (Thomas and Crow,1984). The level of galactokinase wasmany times greater in the Galt variantthan in the parent strain. By contrast wehave been unable to detect activity of anyof the Gal enzymes in Lb bulgaricus(strains B131 and B032; Poolman, unpub-lished observations). Moreover, the regionupstream of the lac operon of Lb bulgari-eus B131 does not code for any of the Galenzymes as does the corresponding re-gion in S thermophilus. This suggests thatthese Lb bulgaricus strains are not onlyphenotypically but also genotypically Galnegative.
ln the section on ion-linked galactosidetransporters and galactoside exchange it
B Poolman
has been indicated that the lactose/galactose exchange reaction is favouredover lactose uptake and galactose excre-tion via separate steps. Since GaIK+strains of S thermophilus selected in lac-tose-limited chemostats are very unstableand lose their ability to utilize galactoserapidly upon transfer to media containingan excess of lactose (Thomas and Crow,1984), it has been suggested that the Gal-phenotype of most strains may be a fa-vourable trait due to the more rapid uptake(and metabolism) of lactose in the absenceof galactokinase (Poolman, 1990). Onlywhen lactose becomes limiting, which willnot easily happen in the 'milk environment',the organism may switch to the Gal+ phe-notype.
TAGATOSE 6-PHOSPHATE PATHWAY
ln the tagatose 6-phosphate pathway ga-lactose 6-phosphate, obtained from the hy-drolysis of lactose 6-phosphate or the cou-pied transport and phosphorylation ofgalactose mediated by PTS (Thompson,1980), is isomerized to tagatose 6-phosphate by galactose 6-phosphate iso-merase (LacAB) (fig 3). Subsequently, ga-lactose 6-phosphate is phosphorylated atthe expense of ATP by tagatose 6-phosphate kinase' (LacC), whereafter thetagatose 1,6-diphosphate is cleaved to di-hydroxyaceton phosphate and glyceralde-hyde 3-phosphate by an aldolase (LacD)(fig 3).
The genes encoding LacAB, LacC andLacD have been isolated and character-ized genetically (van Rooyen et al, 1991),and they form part of the operon that alsocontains the lactose specifie componentsof the PTS (LacE and LacF) and phospho-B-galactosidase (LacG) in L lactis (de Voset al, 1990). Expression of both lacA andlacB is required for galactose 6-phosphate

Galactoside transport and metabolisrn 95
isomerase activity suggesting a multimericstate of the enzyme. The tagatose 1,6-diphosphate aldolase enzyme of L lactishas been purified and characterized bio-chemically (Crow and Thomas, 1982).
ACKNOWLEDGMENT
The author's research has been made possibleby a fellowship from the Royal NetherlandsAcademy of Arts and Sciences.
REFERENCES
Adhya S (1987) The galactose operon. In: Es-cherichia coli and Salmonella typhimurium:Cellular and Molecular Biology (NeidhardtFC, ed) Am Soc Microbiol, Washington, OC
Alpert CA, Chassy BM (1988) Molecular cloningand nucleotide sequence of the factor IlIlacgene of Lactobacillus cesei. Gene 62, 277-288
Alpert CA, Chassy BM (1990) Molecular cloningand DNA sequence of lacE, the gene encod-ing the lactose-specifie enzyme Il of thephosphotransferase system of. Lactobacilluscasei. J Biol Chem 265, 22561-22568
_Boizet B, Villeval D, Sios P, Novel M, Novel G,Mercenier A (1988) Isolation and structuralanalysis of the phospho-13-galactosidasegene from Streptococcus lactis Z268. Gene62,249-261
Crow VL, Thomas TD (1982) D-Tagatose 1,6-diphosphate aldolase from lactic streptococ-ci: purification, properties, and use in meas-uring intracellular tagatose 1,6-diphosphate.J Bacterio/151 , 600-608
David S (1992) Genetics of mesophilic citratefermenting lactic acid bacteria. PhD thesis,Agric Univ Wageningen, The Netherlands
David S, Stevens H, van Riel M, Simons G, deVos WM (1992) Leuconostoc lactis 13-galactosidase is encoded by Iwo overlappinggenes. J Bacterio/174, 4475-4481
De Vos WM, Boerrigter l, van Rooijen RJ,Reiche B, Hengstenberg W (1990) Charac-terization of the lactose-specific enzymes of
the phosphotransferase system in Lactococ-cus lactis. J Biol Chem 265, 22554-22560
Foucaud C, Poolman B (1992) Lactose trans-port system of Streptococcus thermophilus :reconstitution of the protein and characteriza-tion of the kinetic mechanism of transport. JBiol Chem 267,22087-22094
Hutkins RW, Ponne C (1991) Lactose uptakadriven by galactose efflux in Streptococcusthermophilus: evidence for a galactose-lactose antiporter. Appl Environ Microbio/57,941-944
Hutkins R, Morris HA, McKay LL (1985) Galac-tokinase activity in Streptococcus thermophi-lus. Appl Environ Microbio/50. 777-780
Hyde SC, Emsley P, Hartshorn MJ, MimmackMM, Gileadi U, Pearce SR, Gallagher MP,Gill DR, Hubbard RE, Hubbard RK, HigginsCF (1990) Structural model of ATP-bindingprotein associated with cystic fibrosis, multi-drug resistance and bacterial transport. Na-ture 346, 362-365
Kandler 0 (1983) Carbohydrate metabolism inlactic acid bacteria. Antonie van Leeuwen-hoek 49, 209-224
Kashket ER, Wilson TH (1973) Proton-coupledaccumulation of galactoside in Streptococcuslactis 7962. Proc Nat! Acad Sci USA 70,2866-2869
Mollet B, Pilloud N (1991) Galactose utilizationin Lactobacillus helveticus : isolation andcharacterization of the galactokinase (galK)and galactose-1-phosphate uridyl transferase(galT) genes. J Bacterio/173, 4464-4473
Pas HH, Robillard GT (1988) S-Phospho-cysteine and phosphohistidine are intermedi-ates in the phosphoenolpyruvate-dependentmannitol transport catalyzed by Escherichiacoli EIIMtI.Biochemistry 27, 5835-5839
Poolman B (1990) Precursor/product antiport inbacteria. Mol Microbio/4,1629-1636
Poolman B, Royer TJ, Mainzer SE, Schmidt BF(1989) Lactose transport system of Strepto-coccus thermophilus: a hybrid protein withhomology to the melibiose carrier and en-zyme III of phosphoenol-pyruvate-dependentphosphotransferase systems. J Bacteriol171, 244-253
Poolman B, Royer TJ, Mainzer SE, Schmidt BF(1990) Carbohydrate utilization in Streptococ-cus thermophilus: characterization of the

96
genes for aldose 1-epimerase (mutarotase)and UDPglucose 4-epimerase. J Bacteriol172,4037-4047
Poolman B, Molenaar D, Konings WN (1992a)Diversity of transport mechanisms in bacteria.ln: Handbook of Biomembranes (ShinitzkyM,ed) Balaban Publ Rehovot, Israel, vol 2
Poolman B, Modderman R, Reizer J (1992b)Lactose transport sytem of Streptococcusthermophilus: the role of histidine residues. JBiol Chem 267,9150-9157
Porter EV, Chassy BM (1988) Nucleotide se-quence of the l3-o-phosphogalactosidegalac-tohydrolase gene of Lactobacillus casei :comparison to analogues pbg genes of otherGram-positive organisms. Gene 62, 263-276
Reizer J (1989) Regulation of sugar uptake andefflux in Gram-positive bacteria. FEMS Mi-crobiol Rev 63, 149-156
Reizer J, Deutscher J, Sutrina S, Thompson J,Saier MH .(1985) Sugar accumulation inGram-positive bacteria: exclusion and expul-sion mechanisms. Trends Biochem Sei 10,32-35
Romano AH, Brino G, Peterkofsky A, Reizer J(1987) Regulation of J3-galactosidetransportand accumulation in heterofermentative lac-tic acid bacteria. J Bacterio/169, 5589-5596
Russell RRB, Aduse-Opoku J, Sutcliffe IC, TaoL, Ferretti JJ (1992) A binding protein-dependent transport system in Streptococ-eus mutans responsible for multiple sugarmetabolism. J Biol Chem 267,4631-4637
B Poolman
Schmidt BF, Adams LM, Requadt C, Power S,Mainzer SE (1989) Expression and nucleo-tide sequence of the Lactobacillus bulgaricusl3-galactosidase gene cloned in Escherichiacoli. J Bacterio/171 , 625-635
Thomas TD, Crow VL (1984) Selection of galac-tose-fermenting Streptococcus thermophilusin lactose-Iimited chemostat cultures. ApplEnviron Microbio/48, 186-191
Thompson J (1980) Galactose transport sys-tems in Streptococcus lactis. J Bacterio/144,683-691
Thompson J (1987) Sugar transport in the lacticacid bacteria. In : Sugar Transport and Me-tabolism in Gram-Positive Bacteria (Reizer J,Peterkofsky A, eds) Ellis Horwood Ltd, Chi-chester, UK
Van Rooijen RJ, Schalkwijk S, de Vos WM(1991) Molecular c1oning, characterization,and nucleotide sequence of the tagatose 6-phosphate pathway gene cluster of the lac-tose operon of Lactococcus lactis. J BiolChem266,7176-7181
Van Rooijen RJ, Gasson MJ, de Vos WM (1992)Characterization of the Lactococcus lactislactose operon promoter: contribution off1anking sequences and LacR repressor topromoter activity. J Bacterio/174, 2273-2280
Voast S, Schmidt BF, Royer TJ, Poolman B(1990) Characterization of the 3' end of thelactose operon from Lactobacillus delbrueckiisubsp bulgaricus. FEMS Microbiol Rev 87,18


















