
BDNF and NT-3 increase excitatory input connectivity in ... · PDF fileBDNF and NT-3 increase...
13
SYNAPTIC MECHANISMS BDNF and NT-3 increase excitatory input connectivity in rat hippocampal cultures Shimshon Jacobi, 1, * Jordi Soriano, 1,2, * Menahem Segal 3 and Elisha Moses 1 1 Department of Physics of Complex Systems, Weizmann Institute of Science, Rehovot, 76100 Israel 2 Department ECM, Facultat de Fı ´sica, Universitat de Barcelona, Av. Diagonad 647, Barcelona, 08028 Spain 3 Department of Neurobiology, Weizmann Institute of Science, Rehovot, 76100 Israel Keywords: BDNF, network connectivity, NT-3, percolation, synaptic connectivity Abstract The neurotrophic factors brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) have been shown to promote excitatory and inhibitory synapse development. However, a quantitative analysis of their influence on connectivity has proven in general difficult to achieve. In this work we use a novel experimental approach based on percolation concepts that provides a quantification of the average number of connections per neuron. In combination with electrophysiological measurements, we characterize the changes in network connectivity induced by BDNF and NT-3 in rat hippocampal cultures. We show that, on the one hand, BDNF and NT-3 accelerate the maturation of connectivity in the network by about 17 h. On the other hand, BDNF and NT-3 increase the number of excitatory input connections by a factor of about two, but without modifying the number of inhibitory input connections. This scenario of a dominant effect on the excitation is supported by the analysis of spontaneous population bursts in cultures treated with either BDNF or NT-3, which show burst amplitudes that are insensitive to the blockade of inhibition. A leaky integrate-and-fire model reproduces the experimental results well. Introduction The neurotrophins constitute a family of peptides that are secreted and sensed by neurons, influencing their development and survival. Of the many known neurotrophins, brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) play critical roles in controlling neuronal growth and synaptic efficacy. In general, neurotrophins play diverse roles in development, and each may have dissimilar, even contradictory, effects in different areas of the brain (for a review, see Huang & Reichardt, 2001). To shed light on the influence of neurotrophins in neural circuits, we considered rat hippocampal cultures, a well-studied system in which both BDNF and NT-3 have been shown to drive development in excitatory neurons and synapses (Morfini et al., 1994; Bartrup et al., 1997; Baker et al., 1998; Vicario-Abejon et al., 1998; Bolton et al., 2000; Labelle & Leclerc, 2000). BDNF also plays an important, though complex, role in regulating inhibition (Segal & Fishbein, 2005). On the one hand, BDNF promotes inhibition by elongating the axon and dendrites in inhibitory neurons, and by increasing their synaptic density and strength (Murphy et al., 1998; Vicario-Abejon et al., 1998; Jin et al., 2003; Palizvan et al., 2004; Baldelli et al., 2005). On the other hand, acute exposure to BDNF reduces the strength of inhibitory synapses (Tanaka et al., 1997; Frerking et al., 1998; Mizoguchi et al., 2003). Given these intriguing, sometimes conflicting, observations, we chose to examine whether the effects of BDNF and NT-3 could be clearly delineated in terms of global network connectivity. Thus far, however, neurotrophic action on connectivity has been measured by local means, such as electrophysiological measurements of synaptic efficacy, and by estimation of the dendritic and axonal dimensions. How neurotrophins influence the global connectivity of the network has yet to be elucidated. Measuring connectivity in networks that contain a large number of neurons is a highly non-trivial task even in vitro, where neurons and their connections are relatively accessible. For this reason, we have recently developed a novel measurement technique based on graph and percolation theory (Breskin et al., 2006; Soriano et al., 2008), which enables the global, statistical measurement of the average number of input connections per neuron. Here, we apply this technique to quantify changes in connectivity induced by the neurotrophic agents BDNF and NT-3. Materials and methods Neural cultures Rat hippocampal neurons from 17- (E17) or 19- (E19) day-old embryos were used. All procedures were approved by the Weizmann Institute’s Animal Care and Use Committee. Following Segal & Manor (1992), dissection was carried out in ice-cold L-15 medium enriched with glucose (0.6%) and Gentamicin (0.4%; Sigma-Aldrich, St Louis, MO, USA). The hippocampus was dissociated by mechan- Correspondence: Dr E. Moses, as above. E-mail: [email protected] *S.J. and J.S. contributed equally to this study. Received 28 March 2009, revised 22 June 2009, accepted 8 July 2009 European Journal of Neuroscience, Vol. 30, pp. 998–1010, 2009 doi:10.1111/j.1460-9568.2009.06891.x ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd European Journal of Neuroscience
Transcript of BDNF and NT-3 increase excitatory input connectivity in ... · PDF fileBDNF and NT-3 increase...

SYNAPTIC MECHANISMS
BDNF and NT-3 increase excitatory input connectivity in rathippocampal cultures
Shimshon Jacobi,1,* Jordi Soriano,1,2,* Menahem Segal3 and Elisha Moses1
1Department of Physics of Complex Systems, Weizmann Institute of Science, Rehovot, 76100 Israel2Department ECM, Facultat de Fı́sica, Universitat de Barcelona, Av. Diagonad 647, Barcelona, 08028 Spain3Department of Neurobiology, Weizmann Institute of Science, Rehovot, 76100 Israel
Keywords: BDNF, network connectivity, NT-3, percolation, synaptic connectivity
Abstract
The neurotrophic factors brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) have been shown to promote excitatoryand inhibitory synapse development. However, a quantitative analysis of their influence on connectivity has proven in general difficultto achieve. In this work we use a novel experimental approach based on percolation concepts that provides a quantification of theaverage number of connections per neuron. In combination with electrophysiological measurements, we characterize the changes innetwork connectivity induced by BDNF and NT-3 in rat hippocampal cultures. We show that, on the one hand, BDNF and NT-3accelerate the maturation of connectivity in the network by about 17 h. On the other hand, BDNF and NT-3 increase the number ofexcitatory input connections by a factor of about two, but without modifying the number of inhibitory input connections. This scenarioof a dominant effect on the excitation is supported by the analysis of spontaneous population bursts in cultures treated with eitherBDNF or NT-3, which show burst amplitudes that are insensitive to the blockade of inhibition. A leaky integrate-and-fire modelreproduces the experimental results well.
Introduction
The neurotrophins constitute a family of peptides that are secreted andsensed by neurons, influencing their development and survival. Of themany known neurotrophins, brain-derived neurotrophic factor(BDNF) and neurotrophin-3 (NT-3) play critical roles in controllingneuronal growth and synaptic efficacy.In general, neurotrophins play diverse roles in development, and
each may have dissimilar, even contradictory, effects in different areasof the brain (for a review, see Huang & Reichardt, 2001). To shed lighton the influence of neurotrophins in neural circuits, we considered rathippocampal cultures, a well-studied system in which both BDNF andNT-3 have been shown to drive development in excitatory neurons andsynapses (Morfini et al., 1994; Bartrup et al., 1997; Baker et al., 1998;Vicario-Abejon et al., 1998; Bolton et al., 2000; Labelle & Leclerc,2000). BDNF also plays an important, though complex, role inregulating inhibition (Segal & Fishbein, 2005). On the one hand,BDNF promotes inhibition by elongating the axon and dendrites ininhibitory neurons, and by increasing their synaptic density andstrength (Murphy et al., 1998; Vicario-Abejon et al., 1998; Jin et al.,2003; Palizvan et al., 2004; Baldelli et al., 2005). On the other hand,acute exposure to BDNF reduces the strength of inhibitory synapses(Tanaka et al., 1997; Frerking et al., 1998; Mizoguchi et al., 2003).
Given these intriguing, sometimes conflicting, observations, we choseto examine whether the effects of BDNF and NT-3 could be clearlydelineated in terms of global network connectivity.Thus far, however, neurotrophic action on connectivity has been
measured by local means, such as electrophysiological measurementsof synaptic efficacy, and by estimation of the dendritic and axonaldimensions. How neurotrophins influence the global connectivity ofthe network has yet to be elucidated.Measuring connectivity in networks that contain a large number of
neurons is a highly non-trivial task even in vitro, where neurons andtheir connections are relatively accessible. For this reason, we haverecently developed a novel measurement technique based on graphand percolation theory (Breskin et al., 2006; Soriano et al., 2008),which enables the global, statistical measurement of the averagenumber of input connections per neuron. Here, we apply thistechnique to quantify changes in connectivity induced by theneurotrophic agents BDNF and NT-3.
Materials and methods
Neural cultures
Rat hippocampal neurons from 17- (E17) or 19- (E19) day-oldembryos were used. All procedures were approved by the WeizmannInstitute’s Animal Care and Use Committee. Following Segal &Manor (1992), dissection was carried out in ice-cold L-15 mediumenriched with glucose (0.6%) and Gentamicin (0.4%; Sigma-Aldrich,St Louis, MO, USA). The hippocampus was dissociated by mechan-
Correspondence: Dr E. Moses, as above.E-mail: [email protected]
*S.J. and J.S. contributed equally to this study.
Received 28 March 2009, revised 22 June 2009, accepted 8 July 2009
European Journal of Neuroscience, Vol. 30, pp. 998–1010, 2009 doi:10.1111/j.1460-9568.2009.06891.x
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience

ical trituration, and the neurons plated on 13-mm coverslips (Mari-enfeld, Germany), with a final density in the range of 500–800 neurons ⁄ mm2 (Fig. 1A). Coverslips were prepared 1 day earlierby 20 min sonication in ethanol, flaming and overnight immersion inpoly-l-lysine solution (0.007% in borate buffer). Prior to plating,coverslips were washed twice with DDW and once with platingmedium, consisting of MEM+3G [Eagle’s minimum essential medium(MEM) with Earle’s salts (Sigma-Aldrich) with Glutamax (Gibco-Invitrogen, Carlsbad, CA, USA), 0.6% glucose and 0.4% Gentamicin(Sigma-Aldrich)] supplemented with 5% foetal calf serum and 5%horse serum (HS) (both from Gibco-Invitrogen). On day in vitro (DIV)4, the medium was switched to MEM supplemented with 10% HS,20 mg ⁄ mL 5¢-fluoro-2¢-deoxyuridine and 50 mg ⁄ mL uridine (all fromSigma-Aldrich) to limit glial growth. Finally, on DIV 7, the mediumwas changed to MEM supplemented with 10% HS. The latter mediumwas refreshed every 2 days by replacing 1 ⁄ 3 of the well’s volume.
Neurotrophin treatment
Mature forms of recombinant BDNF and NT-3 (20 ng ⁄ mL; AlomoneLabs, Jerusalem, Israel) were chronically applied, starting within
6–12 h after plating, and refreshed every 2 days. We observed that theresults for mature cultures (DIV 12-16) were neither sensitive to theexact embryonic age of the plated neurons (E17 or E19), nor to theprecise day (DIV 0-4) of initial neurotrophin treatment (data notshown).It is important to note that BDNF was not present during the time of
measurement, nor during fluorescent dye loading. Hence, the culturewas not exposed to exogenous BDNF for at least 1 h prior to theexperiment.We also carried out control experiments with BDNF scavengers to
verify that changes in connectivity were solely induced by theneurotrophins. We used the BDNF scavenger protein TrkB-Igg1 (BDBiosciences) at concentrations in the range 0.5–1.5 lg ⁄ mL. Higherconcentrations resulted in cell death. The scavenger was applied ontothe BDNF-treated cultures at the same time as the neurotrophin, andwas refreshed every 2 days.
Pharmacology
We studied the response of the neural network to partial blocking ofthe a-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)-
Fig. 1. Effect of brain-derived neurotrophic factor (BDNF) on synaptic strength. (A) Phase micrograph image of an E19 rat hippocampal culture at DIV 14. (B)Sample of whole-cell voltage-clamp recordings for BDNF-treated and controls cultures. Cells were held at -40 mV to record spontaneously generated both excitatorypostsynaptic currents (EPSCs; marked by ticks below traces) and inhibitory postsynaptic currents (IPSCs; ticks above traces). (C) Cumulative amplitude distributionof solitary EPSC and IPSC events, for E19 cultures at DIV 14 treated with BDNF (average over n = 6 cells from four cultures) compared with controls (average overn = 6 cells from three cultures). Error bars indicate standard errors. (D) Measurements of neuronal excitability and input resistance for BDNF-treated cultures (n = 8cells from four cultures) and controls (n = 7 cells and four cultures). Main plot: number of current spikes recorded while the membrane potential was held for 250 msbetween )70 and )10 mV. Inset: current-voltage measurements for the same cells. The input resistance was obtained as linear fits between )70 and )40 mV,providing 173 ± 21 MX for BDNF-treated cultures and 249 ± 4 MX for controls.
BDNF and NT-3 input rat hippocampal cultures 999
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

glutamate receptors in excitatory neurons, and used 6-cyano-2,3-dihydroxy-7-nitro-quinoxaline acid (CNQX; Sigma-Aldrich) asAMPA antagonist. CNQX reduces synaptic efficacy by blockingAMPA receptors, and we used the fraction of active receptors as ameasure of the efficacy, given by c = 1 ⁄ (1 + [CNQX] ⁄ Kd) (Sorianoet al., 2008), where Kd = 300 nm is the CNQX dissociation constant(Honore et al., 1988). N-methyl-d-aspartate (NMDA) receptors inexcitatory neurons were completely blocked with 20 lm 2-amino-5-phosphonovaleric acid (APV) (Sigma-Aldrich) to monitor theresponse of the network due solely to CNQX.We considered two types of networks, one containing both
excitatory and inhibitory neurons (labelled GEI), and one containingonly excitatory neurons (GE). For the latter c-aminobutyric acid(GABA)ergic synapses were blocked with 40 lm bicuculline metho-chloride (Tocris Bioscience, Bristol, UK). In our cultures, theinhibitory action of GABAergic synapses is present starting at DIV7-8 (Breskin et al., 2006; Soriano et al., 2008). Because we studiedmature cultures (DIV 12-16) we did not consider GABAergicsynapses to have an excitatory role.For experiments in which inhibition was partially blocked, we
considered the fraction of active receptors as a measure of the strengthof inhibitory synapses 1 ⁄ (1+[BIC] ⁄ Kd) with Kd = 3 lm as thedissociation constant for bicuculline (Zukin et al., 1974; Ault &Nadler, 1982; Shirasaki et al., 1991).
Electrophysiology
To determine the change in the synaptic strength due to the action ofthe neurotrophins, we measured synaptic currents by means ofa whole-cell patch-clamp technique (Korkotian & Segal, 2001).Cultures were perfused with the same recording solution as the oneused for fluorescence imaging. Cells were patch-clamped with a1.5-mm outer diameter micropipette linked to an Axoclamp 2Bamplifier (Axon Instruments, Foster City, CA, USA). The patchpipette contained (in mm): K-gluconate, 140; NaCl, 2; HEPES, 10;EGTA, 0.2; Na-GTP, 0.3; Mg-ATP, 2; phosphocreatine, 10 (pH 7.4),with a resistance in the range 6–12 MX. Voltage-clamping at )40 mVenabled the recording of both inward-going excitatory and outward-going inhibitory postsynaptic currents (EPSC and IPSC, respectively)that resulted from spontaneous network activity, as shown in Fig. 1B.We observed that up to 85% of the detected postsynaptic currents
(PSCs) occurred in population bursts. In such cases, PSC events fromnumerous pre-synaptic cells piled up, thereby masking the amplitudeof single events. Because we were interested in determining thestrength of single synapses, we eliminated the piled-up events byrestricting the analysis to events that occurred at least dt = 20 ms apartfrom each other, and at least 500 ms after action potentials. We finallycomputed the PSC cumulative amplitude distributions and comparedBDNF-treated cultures with controls. We verified that the distributionswere insensitive to the value of dt.We also studied the effect of BDNF on neural excitability. To limit
population bursts we used a modified recording medium with elevatedlevels of Mg2+ (4 mm) and Ca2+(2 mm). EPSC amplitudes wererecorded in cells clamped to )60 mV over 20 s. Excitability wasdetermined by holding the membrane at )70 mV and stepping themembrane voltage up at 10 mV steps for 250 ms to differentdepolarized potentials. The number of action currents generated wasthen counted. Finally, the input resistance of the cell was determinedby measuring the current that was required to hold the cell at differentpreset voltages. We studied four voltages, between )70 and )40 mVin steps of 10 mV, and obtained the value of the resistance as a linearfit of the current-voltage values.
Calcium fluorescence imaging
Neural activity was monitored using fluorescence calcium imaging,with the following procedure: cultures were first incubated for 60 minin the recording solution (that included, in mm: NaCl, 128; KCl, 4;CaCl2, 1; MgCl2, 1; glucose, 10; HEPES, 10; the pH was titrated to7.4 and osmolarity to 320 mOsm with sucrose) in the presence of2 lg ⁄ mL of the cell-permeant Fluo4-AM (Molecular Probes), acalcium-sensitive dye. Cultures were then placed in fresh recordingsolution and imaged on an Axiovert 135TV inverted microscope(Zeiss, Germany). The analysis of the fluorescence signal is describedbelow.
Measurement of neural activity
Variations in calcium fluorescence were measured in single neuronsusing the microscope with an online analysis program, as described inBreskin et al. (2006) and Soriano et al. (2008). Spontaneous activitywas recorded for 10–15 min at every experimental condition in orderto obtain sufficient statistics from several tens of bursts.Evoked neural activity was obtained by applying a global electrical
stimulation across two parallel platinum wires, separated by about15 mm, and located at the bottom of the measurement chamber. Thecoverslip that contained the culture was centred between the twoplatinum wires. A bipolar current pulse with amplitude in the range1–30 mA and duration of 20 ms was applied across the wires, with aresulting electrical field of 0.5–20 V ⁄ cm. The inter-stimulus intervalwas set in the range of 2–4 min to ensure sufficient resting time for theneurons, thus preventing memory effects or increased sensitivity to theelectric excitation.To accurately measure the response of the neurons upon stimu-
lation, the activity of each neuron was recorded over intervals of�30 s surrounding the onset of stimulation. A neuron was regarded asresponding to the stimulation if its fluorescence signal, integrated over1 s following stimulation, differed from the baseline by at least fourstandard deviations, as exemplified in Fig. 2A. Although we typicallyobserved several hundred neurons in the field of view of themicroscope, we found that it was sufficient to monitor only about80 of them, which were selected based on a visual assessment of theirfluorescent signal. The error introduced by monitoring only a fractionof the accessible neurons was less than 10%, which is about theoverall systematic error associated with the graph theoretical, statis-tical analysis (Soriano et al., 2008) that is described in the nextsection.It was previously shown that the fluorescence amplitude is linearly
related to the rate of spikes fired. This relation works well for a smallnumber of spikes that occur within a measurement window smallerthan 200 ms (Canepari & Mammano, 1999; Murayama et al., 2005).These conditions are fulfilled in our cultures (based on MEArecordings, data not shown; see Eytan & Marom, 2006; Eckmannet al., 2008). Therefore, we use the fluorescence amplitude to estimatethe maximal firing rate of the burst.
Percolation approach and giant component
The input connectivity of the neurons in culture was measured usinga recent experimental approach that allows the determination ofseveral important statistical properties in living neural networks(Breskin et al., 2006; Soriano et al., 2008). Conceptually, thisapproach is based on the relationship between the average number ofinput connections per neuron, k, and the number of active inputsrequired for a neuron to fire, m. In the framework of percolation,
1000 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

neuronal activity propagates across the culture only if the averageconnectivity k is higher than the input threshold m (Soriano et al.,2008). Therefore, for k � m the neurons are either isolated orforming disconnected clusters, while for k > m the neurons form asingle connected cluster that spans the entire network. This network-spanning cluster, termed giant component (g), enables the spontane-ous synchronous bursting of the whole network, and forms the basisfor the analysis of our experimental results.
The value of g was experimentally determined by measuring thefraction of neurons / that responded together in response to anexternal excitation at voltage V. We identified the size of the giantcomponent as the largest fraction of such responding neurons. Thebehaviour of the response curves /(V ) depends on the connectivity ofthe network (Breskin et al., 2006; Soriano et al., 2008). In a fully
disconnected network, neurons cannot be ignited by their neighboursand therefore they respond only to the external excitation. Hence, thecurve /(V ) increases smoothly, and g = 0. At the other extreme, in afully connected network, a small number of responding neuronssuffices to light up the entire network, /(V ) jumps abruptly at athreshold voltage and g = 1. For intermediate connectivity strengthsbetween neurons, the appearance of a characteristic jump in the /(V )curve signals the existence of a giant component; the size of the jumpconstitutes our measure of g.The behaviour of the giant component was studied using two
different methodologies, as described below. In both approaches,batches of identically prepared E17 hippocampal cultures wereseparated into three groups, according to their treatment: control;BDNF-treated; and NT-3-treated cultures.
Fig. 2. Effect of neurotrophins on network input connectivity. (A) Examples of calcium fluorescence signals for five neurons upon electrical stimulation, comparingBDNF-treated cultures with controls, and for two concentrations of CNQX. The dashed lines and the vertical arrows indicate the excitation time. Arrow heads markthe responding neurons. (B) Size of the giant component g as a function of the concentration of CNQX for GE cultures at DIV 12-15. BDNF-treated cultures (circles,average over n = 6 experiments) require higher concentrations of CNQX for the network to disintegrate compared with controls (open squares, n = 5).(C) Normalized input connectivity, given by 1 + [CNQX]critical ⁄ Kd, where [CNQX]critical is the concentration of CNQX at which g = 0, for GEI and GE networks,comparing neurotrophin-treated cultures (n = 5 for BDNF and n = 6 for NT-3) with controls (n = 7). The value obtained for GE networks is directly related to theaverage number of excitatory input connections in the network, and is higher for neurotrophin-treated cultures than for controls, with P-values of 2 · 10)5 (BDNFtreatment) and 10)4 (NT-3). (D) Relative amount of inhibition in the network. Control cultures have about 70% excitation and 30% inhibition, values thatsignificantly change to about 80% and 20% for neurotrophin-treated cultures, with P-values of 3 · 10)3 (BDNF) and 2 · 10)3 (NT-3). Error bars in (B), (C) and(D) indicate standard errors.
BDNF and NT-3 input rat hippocampal cultures 1001
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

In the first approach, the behaviour of the response curves /(V) andthe size of the giant component g was studied as a function of theculture age in vitro. This methodology allowed us to follow thedevelopment of the network in each of the three different treatments.No synaptic blockers were applied to the culture, so that theemergence and growth of a giant component in the network wasdirectly associated to the formation of new connections. The firstmeasurement was carried out at DIV 2, and repeated in 4–8-h intervalswith another culture from the same treatment and batch. Themeasurements were extended along 2–3 days or until a value ofg = 1 was attained, allowing the identification of the time that a fullyconnected network emerges.In the second approach, we studied the disintegration of the
network as a function of the AMPA receptor antagonist CNQX. Weconsidered mature cultures (DIV 12–16) only. For each culture andtreatment we measured 6–8 response curves /(V) in increasingconcentrations of CNQX, and analysed the curves to obtain the sizeof the giant component g. This gave us g as a function of theconcentration of CNQX, and we could determine the criticalconcentration of antagonist at which g = 0, as exemplified inFig. 2B. The value of this critical concentration is special as itcharacterizes the ‘percolation’ point that separates a connectednetwork from a disconnected one. This critical point is used tocompare the effect of the different treatments on neural connectivity,and to derive the average number of input connections in the culture(see next section).
Determining the number of input connections
In the framework of percolation the critical concentration of CNQX atwhich the network disintegrates is related to the average number ofconnections per neuron. For simplicity, we initially consider networkswith only excitatory inputs (GE networks), which is achieved by fullblocking of the GABAergic synapses with 40 lm bicuculline. Therole of inhibition is treated later.Following Soriano et al. (2008), the percolation approach considers
the simplest model of spiking neurons, in which a neuron has toexceed the membrane firing threshold VT in order to fire and propagatethe activation. Each input onto a neuron has an average contribution gsto the total voltage, and therefore m0 = VT ⁄ gs inputs are initiallyrequired for a neuron to fire. The average synaptic strengthgs decreases as CNQX is applied to the culture. We assume thatgs decreases proportionally to the fraction of active AMPA receptors c,given by c = 1 ⁄ (1 + [CNQX] ⁄ Kd). Therefore, the number of inputconnections necessary for a neuron to fire gradually increases fromm = m0 to m = m0 ⁄ c as CNQX is administered.At a critical concentration [CNQX]E the network disintegrates and
g = 0. In the framework of our percolation model, the critical inputthreshold m at this concentration is approximately equal to the averagenumber of input connections kE (Soriano et al., 2008), and thus
kE ¼m0
c¼ m0ð1þ ½CNQX�E=kdÞ:
This is the mathematical expression that we use to relate the criticalconcentration of CNQX with the average number of excitatory inputconnections. Note, however, that because VT = 15–30 mV and gs =1–2 mV, the value of m0 typically varies in the range 7–30. Directmeasurements of m0 have proven difficult, but reported values fallwithin this range (Otmakhov et al., 1993; Deuchars & Thomson,1996; Bolton et al., 2000; Feinerman et al., 2005). Hence, we takem0 = 15, which fixes kE with an error of a factor of 2.
Our analysis is based on the assumption that the number of inputconnections of every neuron is drawn from the same probabilitydistribution. As the axons are a few hundred microns long, reachingneurons that are not directly in the field of view, neural densityfluctuations are smoothed and our assumption seems justified.However, we carried out control experiments to verify that fluctuationsin neural density did not modify the results within our experimentalerror (data not shown).
Measuring the effect of neurotrophins on the number of inputconnections
Treatment of neural cultures with neurotrophins may modify both theaverage number of connections kE and their synaptic strength gs. Welabel the modified quantities by k
0E and g¢s, respectively, and the
corresponding critical concentration by [CNQX]¢E.The membrane firing threshold VT is insensitive to neurotrophin
treatment (Bolton et al., 2000). Hence, m¢0 = m0gs ⁄ g¢s, and theaverage number of connections in treated cultures is given by
k0E ¼ m00 1þ ½CNQX�0E=Kd
� �¼ m0gs=g0sð1þ ½CNQX�
0E=KdÞ:
It is clear from this equation that the critical concentration[CNQX]¢E is sensitive to changes in both k
0E and g¢s. Thus, to
accurately determine the value of k0E from the measured critical
concentration [CNQX]¢E, we need to quantify the change in thesynaptic strength g¢s due to neurotrophin treatment by using anindependent measurement. This measurement is provided through therecording of synaptic input currents by means of the whole-cell patch-clamp technique.
Determining the effect of neurotrophins on inhibitoryconnectivity
To estimate the amount of inhibition in the network, we studied thecritical concentration of CNQX at which the giant componentdisintegrates, but for the case in which inhibition was left active(GEI network). We describe inhibition using a highly simplified modelof ‘subtractive’ inhibition. If kI is the average number of inhibitoryinputs onto a neuron, then the net input connectivity is given bykEI ¼ kE � kI. This relation assumes that the average EPSC and IPSCamplitudes are practically equivalent (Shadlen & Newsome, 1998;Varela et al., 1999), so that excitatory and inhibitory synapses have asimilar strength. It also assumes that the role of inhibition in neuralconnectivity is to effectively reduce the amount of excitatoryconnections. Hence, we consider GABAergic synapses to be onlyinhibitory, and we disregard other differences between excitatory andinhibitory inputs such as the synaptic kinetics, density and distributionacross the neurons, that may lead to more complex effects such asshunting inhibition.With these assumptions, kEI for control cultures is given by
kEI ¼ m0ð1þ ½CNQX�EI=KdÞ;
while for neurotrophin-treated cultures it is given by
k0EI ¼ m0ð1þ ½CNQX�0EI=KdÞ ¼ m0
gsg0sð1þ ½CNQX�0EI=KdÞ
Here, [CNQX]EI and [CNQX]¢EI are the critical concentrations atwhich g = 0 for control and neurotrophin-treated cultures, respec-tively, in the GEI network.
1002 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

The excitatory input fraction is given by the ratio between thenumber of excitatory inputs and the total number of inputs, which isobtained from the measured values of [CNQX]EI and [CNQX]E by
kEkE þ kI
¼ 2� kEIkE
� ��1¼ 2� 1þ ½CNQX�EI=Kd
1þ ½CNQX�E=Kd
� ��1
This ratio is independent of the synaptic strength between neurons,and is therefore particularly suitable for the assessment of the ‘relative’change in the amount of inhibition due to the action of neurotrophins.
Results
Effect of BDNF on synaptic strength
The synaptic strength between neurons may be altered by theneurotrophin treatment, therefore affecting our analysis of theconnectivity of the network, as described in Materials and methods.To quantify the influence of neurotrophins on the synaptic strength gs,we compared the response of mature (DIV 12–16) BDNF-treatedcultures and controls in single-cell patch-clamp recordings.
For both BDNF-treated cultures and controls we recorded PSCsthat are generated spontaneously. The cells were voltage-clamped at)40 mV to simultaneously measure both inward EPSCs and outwardIPSCs.
The results are shown in Fig. 1B. For both control and BDNF-treated cultures we measured PSC amplitudes from four differentcultures, with a total of six cells. EPSC measurements for controlsprovided an average current of 20 ± 4 pA, which increased to25 ± 3 pA for BDNF-treated cultures. To probe the statisticalsignificance of these values, we computed the cumulative probabilitydistributions (Fig. 1C) and analysed them using the Kolmogorov–Smirnov test. We observed that treated cells have EPSC amplitudesthat are, on average, (24 ± 4)% higher than control amplitudes. Thisincrease is significant (P-value of 10)5) and in agreement with the�30% increase observed by others (Bolton et al., 2000).
IPSC amplitude measurements (Fig. 1C) for the same cells providedan average current of 25 ± 8 pA for controls (four of the six cellsshowed IPSCs only) and 27 ± 9 pA for BDNF-treated cultures (all sixcells showed IPSCs). The measured amplitudes for the two treatmentswere not statistically different, within an estimated error of the order of30% in our measurements of the IPSC.
We conclude from these measurements that BDNF-treated culturesincrease the postsynaptic excitatory but not inhibitory currents by upto 25% compared with controls.
Effect of BDNF on neurons’ excitability and input resistance
To verify that BDNF did not influence the excitability of the neurons,we carried out additional electrophysiological measurements to studyneuronal excitability and input resistance. We used a recordingsolution with elevated concentrations of Mg2+ and Ca2+ to reducepopulation bursts.
We quantified the excitability of the neurons by determining thethreshold potential that resulted in a significant increase in spikegeneration. As shown in Fig. 1D, we explored voltages between)70 mV and )10 mV, and measured similar numbers of spikes forboth control and BDNF-treated cultures. The threshold potential wasalso similar, and between )40 and )30 mV.
The inset of Fig. 1D shows the results of the membrane inputresistance. For control cultures (n = 8 cells) we measured249 ± 4 MX, which decreased to 173 ± 21 MX for BDNF-treated
cultures (n = 8 cells, P-value = 2 · 10)4). This reduced membraneresistance upon BDNF treatment is in agreement with previousobservations (Bolton et al., 2000), and may be associated withan increase of the soma area (Bartrup et al., 1997), among otherinterpretations (Sherwood & Lo, 1999).The reduced input resistance for BDNF-treated cultures has an
interesting implication. Following Bolton et al. (2000) we assume thatthe postsynaptic potentials (PSP) can be quantified as the synapticcurrent times the input resistance. Hence, the measured increase insynaptic currents combined with the decrease in the input resistanceleads to an effective balance of the two contributions, so that the PSPsare practically the same for BDNF-treated cultures and controls. Ourmeasurements provide a ratio of 0.86 ± 0.13 between the PSPs oftreated and untreated cultures. Because this ratio is the result of acomplex balance of the two factors and is nevertheless so close tounity, it is convenient in the analysis of the connectivity to assume thatthe synaptic strength between neurons does not increase upontreatment. If the synaptic strength does differ from unity (which ispractically within the experimental error), then it would change thevalue of the number of inputs needed to excite the cell, m = VT ⁄ gs,by the same amount. Using g¢s = gs, we are effectively measuring alower bound on the increase of the number of input connections. Atmost, this would introduce an error of underestimation in the increasein m by up to 14%.
Effect of neurotrophins on the number of input connections
Long-term treatment with neurotrophins may increase the connectivityof the neural network. To prove that this is the case and providea quantitative estimation, we used the percolation approach (seeMaterials and methods) to determine the critical CNQX concentrationat which the giant component g of the network disintegrates.We studied mature cultures at DIV 12–16, and compared BDNF-and NT-3-treated cultures with controls.Figure 2A shows examples of the response of five neurons to an
electric stimulation, for both BDNF and control cultures, at twoextreme concentrations of CNQX. It shows that a larger fraction ofneurons remains connected for BDNF-treated cultures compared withcontrols. A more systematic analysis is provided through thedependence of the largest fraction of connected neurons g on theCNQX concentration. As shown in Fig. 2B for GE networks(inhibition blocked), BDNF-treated cultures required a substantiallyhigher concentration of CNQX, by a factor 2, to break off the network,which indicates a substantial increase in the connectivity of thenetwork for BDNF-treated cultures.We extended the study of the dependence of g on [CNQX]E to
NT-3-treated cultures, and compared GE and GEI networks. The resultsare shown in Fig. 2C. For clarity, we give the results in terms of thenormalized input connectivity, defined as (1 + [CNQX]E ⁄ Kd).The normalized input connectivity provides, in units of m0,
the number of excitatory inputs kE for GE networks. Figure 2C showsthat BDNF- and NT-3-treated cultures have kE values about 12m0 and10m0, respectively, which are much higher than the value of 6m0 forcontrols. Hence, with m0 = 15 for control cultures, and assumingm0¢ = m0 = 15 for both BDNF and NT-3, we find that the averagenumber of connections per neuron kE increases from kE = 95 ± 8 forcontrols (n = 7 cultures) to 178 ± 7 for BDNF (n = 5) and to 147 ± 3for NT-3 (n = 5). The increase is significant, with P-values of2 · 10)5 and 10)4, respectively.Figure 2C also shows the values of the normalized input
connectivity for GEI networks. They are related to the net number of
BDNF and NT-3 input rat hippocampal cultures 1003
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

input connections kEI, and are lower than the values for kE due toinhibition. The relative difference between kEI and kE provides anestimation of the fraction of inhibitory connections in the network.With kI ¼ kE � kEI (see Materials and methods), we observed that thetotal number of inhibitory input connections kI shows no statisticallysignificant change due to treatment. For m0 = 15, we obtained kEIto be 56 ± 5 for controls (n = 7 cultures) and 132 ± 5 for BDNF-treated cultures (n = 7), resulting in a kI of 41 ± 5 for controls, and46 ± 9 for BDNF-treated cultures. For NT-3-treated cultures (n = 7),we obtained kEI = 105 ± 4 and kI= 42 ± 4.The substantial increment in the number of excitatory input
connections for treated cultures, which is not followed by a propor-tional increase in the inhibitory connectivity, leads to an elevatedexcitatory fraction of input connections kE=ðkE þ kIÞ (see Materialsand methods). The excitatory fraction increased from (69 ± 1)% forcontrol (n = 6 cultures in each condition) to (80 ± 3)% for BDNF-treated (n = 5) and (79 ± 2)% for NT-3-treated cultures (n = 5), asshown in Fig. 2D. The increases are statistically significant comparedwith controls, with P-values of 2 · 10)2 and 3 · 10)3, respectively.‘Shunting’ inhibition, in which the effect of inhibitory synapses is
multiplicative rather than additive, is a potential complication that mustbe addressed. However, an average neuron will spike a number of timesduring the population bursts, typically 3–5 spikes per neuron. In thiscase, the membrane potential continuously varies between the restingpotential and the threshold potential, remaining most of the time awayfrom the reversal potential. This causes shunting inhibition to have anoverall effective subtractive effect on the synaptic input (Koch, 1999),as we have assumed throughout this work, rather than a divisive one.Within the experimental resolution of our measurement, which is anaverage over the contributions of the various channels and receptors, weare unable to observe the possible effects of shunting inhibition.
Network development
The study of the disintegration of the network with CNQX thusprovided information on the average connectivity of the network. Wehave seen how mature cultures treated with neurotrophins substan-tially increase their number of input connections. In addition,neurotrophins play a pivotal role in the formation and developmentof connections synapses (Bartrup et al., 1997; Vicario-Abejon et al.,1998), and therefore we have studied how the formation of a giantcomponent in the network during development is influenced by theneurotrophins treatment. We previously studied developmental aspectsin neural cultures derived from different embryonic stages (E17, E19and postnatal pups), and observed that the appearance of the giantcomponent is timed to coincide with full term of the pregnancy, so thatcultures from more mature neurons formed a network earlier (Sorianoet al., 2008).Here we followed the development of the network in E17 cultures,
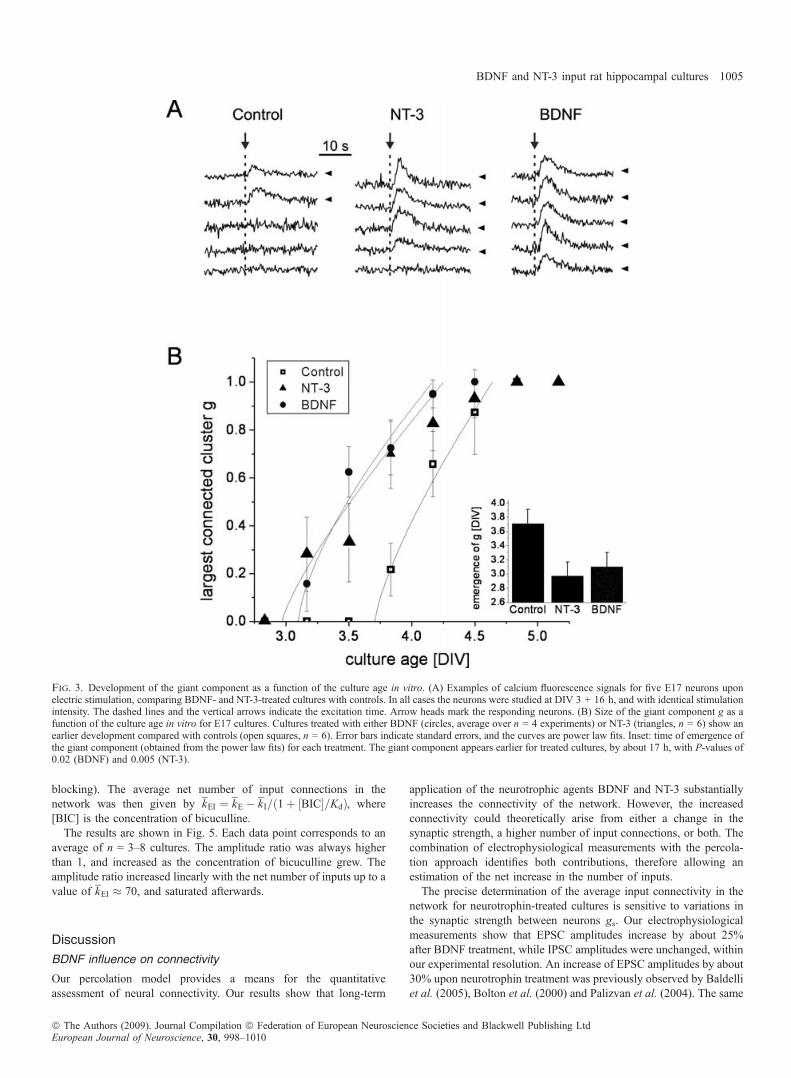
and compared BDNF- and NT-3-treated cultures with controls. Weobserved three major distinctive features. First, cultures treated withneurotrophins responded for the first time to electric excitation earlierthan controls (data not shown). Second, neurons in treated culturesshowed a stronger response to the excitation, as exemplified inFig. 3A. And third, the fraction of neurons that fired together in treatedcultures was significantly higher than in controls of the same age.A detailed study of the evolution of the size of the giant component
as a function of time is shown in Fig. 3B. Neurotrophin-treatedcultures showed electric response by DIV 2.5, while for controls theresponse was not observed until DIV 3. The emergence of the giantcomponent for treated cultures was observed at about DIV 3, while for
controls g appeared about 17 h later. In all cases, however, the giantcomponent grew with a similar rate, reaching g = 1 in about 1 day.The inset of Fig. 3B compares the time of emergence of the giant
component for the three culture types. For controls, g appeared at DIV3.7 ± 0.2 (n = 6 experiments), which decreased to DIV 3.1 ± 0.2 forBDNF-treated cultures (n = 4) and to DIV 3.0 ± 0.2 for NT-3-treatedcultures (n = 6). The decrease was statistically significant, withP-values of 0.02 and 0.005 for BDNF and NT-3.
Neurotrophic effects on spontaneous population bursts
Neural cultures exhibit spontaneous activity in the form of populationbursts, an observation that was previously used as a measure of neuralconnectivity (Bartrup et al., 1997; Vicario-Abejon et al., 1998;Feinerman et al., 2005, 2007; Jacobi & Moses, 2007; Soriano et al.,2008). Here, we studied the influence of the neurotrophin treatment onspontaneous activity.Spontaneous bursting in mature cultures (DIV 12–16) was
measured for both GEI and GE networks. For the latter, inhibitionwas completely blocked with application of 40 lm bicuculline. Weused the peak fluorescence amplitude as a measure of the neuron firingrate in the burst. Examples of network bursting activity are shown inFig. 4A for controls (top), NT-3- (centre) and BDNF (bottom)-treatedcultures. For controls we observed a strong increase in burstingamplitude after blocking of inhibition. However, both BDNF- andNT-3-treated cultures exhibited a very small variation in burstingamplitude between GEI and GE networks.Figure 4B shows an analysis of the burst amplitude increase
following inhibition compared with the baseline amplitude. While forcontrols the amplitude increased by about 180% (n = 9 cultures),treated cultures experienced no statistically significant variation, withan increase of about 20% (n = 8 cultures treated with BDNF andn = 5 cultures treated with NT-3). The difference between the controlsand the treated cultures is statistically significant, with P < 0.001(BDNF) and P < 0.005 (NT-3). Similarly, the analysis of the averageinter-burst interval (IBI) with and without inhibition, shown inFig. 4C, indicates that the IBI almost tripled for control cultures,increasing from 45 ± 9 s to 138 ± 33 s (n = 9 cultures, a statisticallysignificant increase with P = 0.03). In neurotrophin-treated culturesthe IBI had a statistically non-significant increase, from 41 ± 15 s to60 ± 19 s with NT-3 treatment (n = 5), and from 47 ± 9 s to 55 ± 8 swith BDNF treatment (n = 8).Despite the statistically insignificant changes in amplitude and
frequency, we did observe that the coefficient of variation (CV) of theburst amplitude, which is defined as the standard deviation of thesignal divided by its mean, decreased in treated cultures with theblocking of inhibition. The CV changed from 0.24 ± 0.03 to0.11 ± 0.03 for controls (n = 9 cultures), and similarly from0.20 ± 0.05 to 0.12 ± 0.03 for BDNF-treated (n = 8), and from0.23 ± 0.05 to 0.13 ± 0.05 for NT-3-treated (n = 4) cultures.
Influence of partial blocking of inhibition on the amplitudeof spontaneous bursting
To understand how the net number of inputs affects spontaneousbursts, we studied how the bursting amplitude changed when theamount of inhibition in the network was gradually reduced. Weconsidered only untreated control cultures, and measured the ratio ofthe average bursting amplitude between a network with graduallyhigher disinhibition (from GEI towards GE) and the normal GEI
network. We used bicuculline to reduce the inhibitory synapseefficacy, and explored concentrations between 375 nm and 40 lm (full
1004 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010
blocking). The average net number of input connections in thenetwork was then given by kEI ¼ kE � kI=ð1þ ½BIC�=KdÞ, where[BIC] is the concentration of bicuculline.
The results are shown in Fig. 5. Each data point corresponds to anaverage of n = 3–8 cultures. The amplitude ratio was always higherthan 1, and increased as the concentration of bicuculline grew. Theamplitude ratio increased linearly with the net number of inputs up to avalue of kEI � 70, and saturated afterwards.
Discussion
BDNF influence on connectivity
Our percolation model provides a means for the quantitativeassessment of neural connectivity. Our results show that long-term
application of the neurotrophic agents BDNF and NT-3 substantiallyincreases the connectivity of the network. However, the increasedconnectivity could theoretically arise from either a change in thesynaptic strength, a higher number of input connections, or both. Thecombination of electrophysiological measurements with the percola-tion approach identifies both contributions, therefore allowing anestimation of the net increase in the number of inputs.The precise determination of the average input connectivity in the
network for neurotrophin-treated cultures is sensitive to variations inthe synaptic strength between neurons gs. Our electrophysiologicalmeasurements show that EPSC amplitudes increase by about 25%after BDNF treatment, while IPSC amplitudes were unchanged, withinour experimental resolution. An increase of EPSC amplitudes by about30% upon neurotrophin treatment was previously observed by Baldelliet al. (2005), Bolton et al. (2000) and Palizvan et al. (2004). The same
Fig. 3. Development of the giant component as a function of the culture age in vitro. (A) Examples of calcium fluorescence signals for five E17 neurons uponelectric stimulation, comparing BDNF- and NT-3-treated cultures with controls. In all cases the neurons were studied at DIV 3 + 16 h, and with identical stimulationintensity. The dashed lines and the vertical arrows indicate the excitation time. Arrow heads mark the responding neurons. (B) Size of the giant component g as afunction of the culture age in vitro for E17 cultures. Cultures treated with either BDNF (circles, average over n = 4 experiments) or NT-3 (triangles, n = 6) show anearlier development compared with controls (open squares, n = 6). Error bars indicate standard errors, and the curves are power law fits. Inset: time of emergence ofthe giant component (obtained from the power law fits) for each treatment. The giant component appears earlier for treated cultures, by about 17 h, with P-values of0.02 (BDNF) and 0.005 (NT-3).
BDNF and NT-3 input rat hippocampal cultures 1005
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

groups also reported that evoked IPSC amplitudes increase by up to60%, which we did not observe. The different observations may resultfrom the difference between spontaneous and evoked IPSCs.In our cultures, the spontaneous PSC amplitudes increase is
balanced by the decrease of input resistance, leading to PSPamplitudes that do not change much with the BDNF treatment, andtherefore we can assume for simplicity that g¢s = gs.The application of the percolation approach to the analysis of
excitation showed that neuotrophin treatment increased the number ofexcitatory inputs by 80%. This substantial increase gives a quantitativeassessment of results from previous studies on the densification ofexcitatory input synapses induced by BDNF (Singh et al., 2006). Italso relates to previous reports regarding the role of BDNF in synapticdevelopment and maturation (reviewed in Cohen-Cory, 2002; Vicario-
Abejon et al., 2002), as well as the effect of BDNF on the extension ofdendrites and axons (Cohen-Cory & Fraser, 1995; Labelle & Leclerc,2000).In addition to an increase in the average network’s connectivity, our
study also identified a faster development of the network, by about17 h, for cultures treated with either BDNF or NT-3. In comparison,Bartrup et al. measured that the fraction of neurons receiving synapticinputs appeared earlier, by about a day, in BDNF-treated cultures(Bartrup et al., 1997). Vicario-Abejon et al. showed a rescue ofsynaptic development, as well as spontaneous activity, in E16 culturesfollowing application of either BDNF or NT-3 (Vicario-Abejon et al.,1998).The influence of neurotrophin treatment on inhibition is more
complex. The inhibitory contribution is obtained from the relative shift
Fig. 4. Effects of BDNF and NT-3 on spontaneous activity. (A) Examples of spontaneous activity measurements for GE (grey) and GEI (black) networks, comparingcontrol cultures (top) with NT-3- (centre) and BDNF (bottom)-treated cultures. Each signal is an average over the synchronized activity of 80 neurons. (B) Ratiobetween the average bursting amplitude after blocking of inhibition (GE network) and before (GEI). Control cultures (average over n = 8 experiments) show anincrease in amplitude by a factor 3, while BDNF- (n = 8) and NT-3- (n = 5) treated cultures maintain a ratio close to 1. The difference between the ratio for treatedand control cultures is significant, with P-values of 0.001 for BDNF and 0.01 for NT-3 treatments. (C) Comparison of the change in the IBIs before and afterblocking of inhibition, and for the same set of cultures. Control cultures exhibited a significant increase of the inter-burst duration (P-value = 0.03) that was notobserved in neurotrophin-treated cultures. Error bars indicate standard errors.
1006 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

between the curves for GEI and GE networks. Our results show thattreated cultures do not exhibit a statistically significant change in thenumber of inhibitory input connections compared with controls.Previous morphological studies reported an increase in inhibitorysynapse density. Elmariah et al. studied the relation between GABAreceptors and density. They described a two- to threefold increase inreceptor cluster density following exposure to BDNF, without changeof the receptor cluster size (Elmariah et al., 2004, 2005). A similarstudy by Marty et al. showed that BDNF treatment increased thedensity of the GAD65-immunoreactive puncta in CA1 hippocampalslice cultures by 40–60% (Marty et al., 2000). On the other hand,Singh et al. have shown that local BDNF expression in BDNF-deficient cultures reduces the number of inhibitory inputs to post-synaptic neurons, while increasing the corresponding number ofexcitatory inputs (Singh et al., 2006).
Our results are therefore in conflict, albeit indirect, with some ofthese previous observations. While we observe that the number ofinhibitory inputs increases by about 10%, we do not see the expectedeffects of a �50% increase in inhibitory apparatus reported by others.As our measurement of the number of inhibitory inputs is indirect, itsmeasurement error is relatively large, and can reach 25%. What wecan observe very reliably and with small errors is that the relativeweight of inhibition, or the ratio of inhibitory ⁄ excitatory inputs,decreases significantly. This is mainly due to the large increase in thenumber of the excitatory input connections.
Spontaneous activity and the excitatory-inhibitory balance
Our measurements of neurotrophic action on spontaneous burstingamplitudes provided additional insight into the role of inhibition innetwork’s connectivity. Interestingly, while for control cultures the
spontaneous bursting amplitude increased by 180% after the blockingof inhibition, the increase was minor for treated cultures, by only 20%.This remarkable contrast in amplitude variation between treated and
control cultures is an important result, and supports the observationthat inhibition is not significantly affected by the neurotrophins.Indeed, treated cultures behave as though they were permanentlydisinhibited.To shed light on the role of inhibition in the observed burst dynamics,
we consider a simplified computational model of a single leakyintegrate and fire (LIF) neuron with multiple excitatory and inhibitoryinputs, which represents a typical neuron in the bursting network(Koch, 1999). As key ingredients, we introduce a synaptic depressionand synaptic delay. The system is described as a single postsynapticneuron, which accepts kE excitatory and kI inhibitory inputs, each firingat the same rate r(t). The values of kE and kI are obtained from ourmeasurements, and we assume as before m0 = VT ⁄ gs = 15. For controlcultures we take kE = 90 and kI = 40, while for BDNF-treated cultureswe consider kE = 144 and kI = 36. We also set kI = 0 for thedisinhibited case for both control and treated neurons.The LIF model is a simple yet realistic description for the dynamics
of the neuron (Koch, 1999). The temporal profile of the EPSC (IPSC)is described by an exponential growth followed by an exponentialdecay with two different time constants. Its input parameters arestandard, and we take them from the literature: the exponential growthrate is srise = 2 ms and the decay rate sE = 6 ms (sI = 20 ms for IPSC;Segal & Barker, 1984; Wilcox et al., 1994; Bolton et al., 2000; Magee& Cook, 2000). The membrane time-constant is sm = 15 ms (Brownet al., 1981; Segal, 1983) and the absolute refraction period is 3 ms(Koch, 1999).We assume a synaptic delay of 4 ms to shift the response of the
postsynaptic neuron for both EPSP and IPSP (Segal, 1983; Barker &Harrison, 1988; Medina et al., 1995; Bi & Poo, 1999; Kouznetsovaet al., 2002). This delay is a necessary ingredient that creates a timescale for recruitment of activity, which is of the order of a few tens ofms, as experimentally observed (Eckmann et al., 2008). The synapticdepression is calculated as in Abbott et al. (1997) and Varela et al.(1997) using their values for all parameters.The total input onto the LIF neuron is given by the single PSC times
the number of inputs. We include a low background noise to initiatethe burst, consisting of an average spiking rate of 10)2 spikes persecond and input. While the time for burst initiation decreases with theinput noise amplitude, the maximal firing in the burst is insensitive toit as it only depends on the balance between excitatory and inhibitoryinputs on the network.The dynamics of the LIF neuron are simulated by calculating the
postsynaptic output at a given time step. The output is then used as thepresynaptic firing rate of each of the synaptic inputs in the next timestep, until the entire temporal profile of the burst is constructed.Figure 6 shows the temporal profiles of the simulated control and
treated neurons. Interestingly, while for the control neuron themaximal firing rates increase with disinhibition by a factor of 2.8,there is only a small increase by a factor of 1.2 for the BDNF-treatedneuron. Similarly, the total number of spikes in the burst increasesby a factor of 2.6 for controls, and by 1.2 for the BDNF-treatedneurons.The LIF model with synaptic depression and delay reproduces the
experimental results well. It also indicates that the key ingredient forthe different burst dynamics between treated and untreated cultures isthe strong difference in the net number of input connections,kEI ¼ kE � kI. For control cultures kEI increases from kEI � 55 tokE � 95 after disinhibition (75% increase), while for treated culturesthe change is from kEI � 130 to kE � 180 (40% increase).
Fig. 5. Effect of partial disinhibition on spontaneous bursting amplitude incontrol cultures. Ratio between the average burst amplitude for the networkwith partial blocking of inhibition and the standard GEI network, as a functionof the bicuculline concentration. Bicuculline was administered in logarithmicscale, in the range 0.38–40 lm, and the corresponding number of inputconnections was obtained as kE � kI=ð1þ ½BIC�=KdÞ, with [BIC] the concen-tration of bicuculline and Kd = 3 lm its dissociation constant. Each data pointis an average over n = 3–8 cultures. The relative increase in the burst amplitudegrows with the number of input connections, and reaches saturation for about70 net input connections. Error bars indicate standard errors.
BDNF and NT-3 input rat hippocampal cultures 1007
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

Further evidence for the importance of the net number of inputs inburst dynamics was provided by our experiments with gradualblocking of inhibition (Fig. 5), which showed that the burstingamplitude reached saturation at a value of kEI � 70. Hence, wehypothesize that, once a threshold value of the net number inputs isreached, the refractory period limits any further increase of the firingrate and the bursting amplitudes saturate. Indeed, we note thatundrugged BDNF-treated cultures have a relatively high number ofnet inputs kEI � 130, and therefore further increase of connections bythe blocking of inhibition does not substantially modify the burstamplitude.Interestingly, this saturated behaviour of undrugged BDNF-treated
cultures does not affect the decrease in burst amplitude variabilityshown in Fig. 4A, bottom. This variability is reduced by disinhibitionin the same way as it does the control cultures. Hence, the inhibitorynetwork still influences variability in burst initiation even withneurotrophin treatment, with a subsequent variation in the maximalfiring rate of the burst, as shown in Eckmann et al. (2008) and Jacobi &Moses (2007).
Implications on the homeostatic regulation framework
It was previously suggested that BDNF plays an important role in thescaling of synaptic strength for the maintenance of a constant aver-age neuronal firing rate. Specifically, it was reported that long-termapplication of BDNF inhibits strengthening of excitatory synapsesin excitatory neurons that occurs after activity blockade, and leadsto strengthening of the inhibitory network (Rutherford et al., 1998).As BDNF secretion is activity dependent, it was suggested that thisnegative feedback helps in the regulation of the neural activity level(Turrigiano & Nelson, 2000).Our findings may indicate a combined effect of BDNF and the
spontaneous activity that occurs over long time scales. We found that
the single synapse strength is maintained or increases only slightly,which is in agreement with the synaptic scaling framework. Thenumber of input connections, however, increases due to the contin-uous existence of BDNF in the culture medium. This interpretation issupported by the work of Singh et al., where localized BDNFexpression in BDNF-deficient cultures increased excitatory input andreduced inhibitory input in postsynaptic neurons, consequentlyaltering the excitatory-inhibitory balance, as was observed in ourcultures (Singh et al., 2006).
Conclusions
In summary, using a bath excitation protocol coupled with concepts ofpercolation theory, we measured the number of input connections inrat hippocampal cultures, and showed that chronic exposure to BDNFand NT-3 increases the excitatory connectivity by about a factor of 2.The inhibitory connectivity, however, appeared to be insensitive to thetreatment. We associate the increased excitatory connectivity to anincrease in the number of input connections, and not to thestrengthening of the synaptic efficacy between neurons. The strongincrease in the excitatory connectivity, which does not occur for theinhibitory one, is reflected in the spontaneous bursting dynamics. As aresult, burst amplitudes in treated cultures are practically insensitive tothe blocking of inhibition.In native tissue, some of the single neuron characteristics that we
observe may be similar. In particular, relative changes in neuronproperties before and after BDNF application may carry overnaturally. The relative increase of the number of input connectionsthat is induced by BDNF is probably a result of the single neuronresponse to BDNF, given that its environment is not very affected bythe BDNF. Therefore, the excitatory connectivity in native tissues mayincrease without being counterbalanced by an increase in inhibition,resulting in an increasing excitatory ⁄ inhibitory ratio.
Fig. 6. Numerical simulation of the burst time course for control and BDNF cultures. Temporal profiles of population bursts in a model of a LIF neuron withsynaptic depression and delay. The parameters of the model were based on the experimentally measured connectivity. Four cases were simulated: BDNF-treatedcultures (grey) and controls (black), both with either inhibition (solid lines) and without it (dotted lines). The maximal firing rate after disinhibition increased by afactor of 2.8 for controls, but only by a factor of 1.2 for BDNF-treated cultures.
1008 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

In addition, our finding that BDNF perturbs the excitatory ⁄ inhib-itory balance of synaptic inputs is supported by the observation bySingh et al. (2006) that excess synaptic activity induced by BDNFcauses deviation from the homeostatic regulation of the synapticstrength.
In our study, we used cultures of dissociated rat hippocampalneurons. Although the differences between cultures and native tissuesuch as the mammalian brain are substantial, our results suggest thatBDNF in organotypic networks is crucial to regulate excitatoryconnectivity and neural activity. Our percolation approach can beapplied to more complex topologies such as patterned networks orbrain slices. It would certainly be interesting to explore how theoverall connectivity, the balance with inhibition and the neuralactivity are modified by the combination of both neurotrophicagents and the network’s topology. In this direction, we haverecently started to study the influence of neurotrophins on one-dimensional neural cultures, and focused on the interplay betweenthe changes in connectivity reported here and the velocity of neuralactivity fronts.
Acknowledgements
We thank A. Rotem and O. Feinerman for advice, and V. Greenberger fortechnical assistance. This work was supported by the Minerva Foundation(Munich, Germany) and by the Israeli Science Foundation, Grant 993 ⁄ 05.
Abbreviations
AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; BDNF, brain-derived neurotrophic factor; CNQX, 6-cyano-2,3-dihydroxy-7-nitro-quinoxa-line acid; CV, coefficient of variation; DIV, days in vitro; EPSC, excitatorypostsynaptic current; GABA, c-aminobutyric acid; HS, horse serum; IBI, inter-burst interval; IPSC, inhibitory postsynaptic current; LIF, leaky integrate andfire; MEM, minimum essential medium; NT-3, neurotrophin-3; PSC, postsyn-aptic current; PSP, postsynaptic potential.
References
Abbott, L.F., Varela, J.A., Sen, K. & Nelson, S.B. (1997) Synaptic depressionand cortical gain control. Science, 275, 220–224.
Ault, B. & Nadler, J.V. (1982) Baclofen selectively inhibits transmission atsynapses made by axons of CA3 pyramidal cells in the hippocampal slice.J. Pharmacol. Exp. Ther., 223, 291–297.
Baker, R.E., Dijkhuizen, P.A., Van Pelt, J. & Verhaagen, J. (1998) Growth ofpyramidal, but not non-pyramidal, dendrites in long-term organotypicexplants of neonatal rat neocortex chronically exposed to neurotrophin-3.Eur. J. Neurosci., 10, 1037–1044.
Baldelli, P., Hernandez-Guijo, J.M., Carabelli, V. & Carbone, E. (2005) Brain-derived neurotrophic factor enhances GABA release probability andnonuniform distribution of N- and P ⁄ Q-type channels on release sites ofhippocampal inhibitory synapses. J. Neurosci., 25, 3358–3368.
Barker, J.L. & Harrison, N.L. (1988) Outward rectification of inhibitorypostsynaptic currents in cultured rat hippocampal neurones. J. Physiol., 403,41–55.
Bartrup, J.T., Moorman, J.M. & Newberry, N.R. (1997) BDNF enhancesneuronal growth and synaptic activity in hippocampal cell cultures.Neuroreport, 8, 3791–3794.
Bi, G.-Q. & Poo, M.-M. (1999) Distributed synaptic modification in neuralnetworks induced by patterned stimulation. Nature, 401, 792–796.
Bolton, M.M., Pittman, A.J. & Lo, D.C. (2000) Brain-derived neurotrophicfactor differentially regulates excitatory and inhibitory synaptic transmissionin hippocampal cultures. J. Neurosci., 20, 3221–3232.
Breskin, I., Soriano, J., Moses, E. & Tlusty, T. (2006) Percolation in livingneural networks. Phys. Rev. Lett., 97, 188102.
Brown, T.H., Fricke, R.A. & Perkel, D.H. (1981) Passive electrical constants inthree classes of hippocampal neurons. J. Neurophysiol., 46, 812–827.
Canepari, M. & Mammano, F. (1999) Imaging neuronal calcium fluorescence athigh spatio-temporal resolution. J. Neurosci. Methods, 87, 1–11.
Cohen-Cory, S. (2002) The developing synapse: construction and modulationof synaptic structures and circuits. Science, 298, 770–776.
Cohen-Cory, S. & Fraser, S.E. (1995) Effects of brain-derived neurotrophicfactor on optic axon branching and remodelling in vivo. Nature, 378, 192–196.
Deuchars, J. & Thomson, A.M. (1996) CA1 pyramid-pyramid connections inrat hippocampus in vitro: dual intracellular recordings with biocytin filling.Neuroscience, 74, 1009–1018.
Eckmann, J.P., Jacobi, S., Marom, S., Moses, E. & Zbinden, C. (2008) Leaderneurons in population bursts of 2D living neural networks. New J. Phys., 10,015011.
Elmariah, S.B., Crumling, M.A., Parsons, T.D. & Balice-Gordon, R.J. (2004)Postsynaptic TrkB-mediated signaling modulates excitatory and inhibitoryneurotransmitter receptor clustering at hippocampal synapses. J. Neurosci.,24, 2380–2393.
Elmariah, S.B., Oh, E.J., Hughes, E.G. & Balice-Gordon, R.J. (2005)Astrocytes regulate inhibitory synapse formation via Trk-mediated modula-tion of postsynaptic GABAA receptors. J. Neurosci., 25, 3638–3650.
Eytan, D. & Marom, S. (2006) Dynamics and effective topology underlyingsynchronization in networks of cortical neurons. J. Neurosci., 26, 8465–8476.
Feinerman, O., Segal, M. & Moses, E. (2005) Signal propagation alongunidimensional neuronal networks. J. Neurophysiol., 94, 3406–3416.
Feinerman, O., Segal, M. & Moses, E. (2007) Identification and dynamicsof spontaneous burst initiation zones in unidimensional neuronal cultures.J. Neurophysiol., 97, 2937–2948.
Frerking, M., Malenka, R.C. & Nicoll, R.A. (1998) Brain-derived neurotrophicfactor (BDNF) modulates inhibitory, but not excitatory, transmission in theCA1 region of the hippocampus. J. Neurophysiol., 80, 3383–3386.
Honore, T., Davies, S.N., Drejer, J., Fletcher, E.J., Jacobsen, P., Lodge, D. &Nielsen, F.E. (1988) Quinoxalinediones: potent competitive non-NMDAglutamate receptor antagonists. Science, 241, 701–703.
Huang, E.J. & Reichardt, L.F. (2001) Neurotrophins: roles in neuronaldevelopment and function. Annu. Rev. Neurosci., 24, 677–736.
Jacobi, S. & Moses, E. (2007) Variability and corresponding amplitude-velocityrelation of activity propagating in one-dimensional neural cultures.J. Neurophysiol., 97, 3597–3606.
Jin, X., Hu, H., Mathers, P.H. & Agmon, A. (2003) Brain-derived neurotrophicfactor mediates activity-dependent dendritic growth in nonpyramidalneocortical interneurons in developing organotypic cultures. J. Neurosci.,23, 5662–5673.
Koch, C. (1999) Biophysics of Computation. Oxford University Press, NewYork.
Korkotian, E. & Segal, M. (2001) Spike-associated fast contraction of dendriticspines in cultured hippocampal neurons. Neuron, 30, 751–758.
Kouznetsova, M., Kelley, B., Shen, M. & Thayer, S.A. (2002) Desensitizationof cannabinoid-mediated presynaptic inhibition of neurotransmissionbetween rat hippocampal neurons in culture. Mol. Pharmacol., 61, 477–485.
Labelle, C. & Leclerc, N. (2000) Exogenous BDNF, NT-3 and NT-4differentially regulate neurite outgrowth in cultured hippocampal neurons.Brain Res. Dev. Brain Res., 123, 1–11.
Magee, J.C. & Cook, E.P. (2000) Somatic EPSP amplitude is independent ofsynapse location in hippocampal pyramidal neurons. Nat. Neurosci., 3, 895–903.
Marty, S., Wehrle, R. & Sotelo, C. (2000) Neuronal activity and brain-derivedneurotrophic factor regulate the density of inhibitory synapses in organotypicslice cultures of postnatal hippocampus. J. Neurosci., 20, 8087–8095.
Medina, I., Filippova, N., Charton, G., Rougeole, S., Ben-Ari, Y., Khrestch-atisky, M. & Bregestovski, P. (1995) Calcium-dependent inactivation ofheteromeric NMDA receptor-channels expressed in human embryonickidney cells. J. Physiol., 482, 3.
Mizoguchi, Y., Ishibashi, H. & Nabekura, J. (2003) The action of BDNF onGABA(A) currents changes from potentiating to suppressing duringmaturation of rat hippocampal CA1 pyramidal neurons. J. Physiol., 548,703–709.
Morfini, G., DiTella, M.C., Feiguin, F., Carri, N. & Caceres, A. (1994)Neurotrophin-3 enhances neurite outgrowth in cultured hippocampal pyra-midal neurons. J. Neurosci. Res., 39, 219–232.
Murayama, M., Miyazaki, K., Kudo, Y., Miyakawa, H. & Inoue, M. (2005)Optical monitoring of progressive synchronization in dentate granule cellsduring population burst activities. Eur. J. Neurosci., 21, 3349–3360.
Murphy, D.D., Cole, N.B. & Segal, M. (1998) Brain-derived neurotrophicfactor mediates estradiol-induced dendritic spine formation in hippocampalneurons. Proc. Natl. Acad. Sci. U S A, 95, 11412–11417.
BDNF and NT-3 input rat hippocampal cultures 1009
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010

Otmakhov, N., Shirke, A.M. & Malinow, R. (1993) Measuring the impact ofprobabilistic transmission on neuronal output. Neuron, 10, 1101–1111.
Palizvan, M.R., Sohya, K., Kohara, K., Maruyama, A., Yasuda, H., Kimura, F.& Tsumoto, T. (2004) Brain-derived neurotrophic factor increases inhibitorysynapses, revealed in solitary neurons cultured from rat visual cortex.Neuroscience, 126, 955–966.
Rutherford, L.C., Nelson, S.B. & Turrigiano, G.G. (1998) BDNF has oppositeeffects on the quantal amplitude of pyramidal neuron and interneuronexcitatory synapses. Neuron, 21, 521–530.
Segal, M. (1983) Rat hippocampal neurons in culture: responses to electricaland chemical stimuli. J. Neurophysiol., 50, 1249–1264.
Segal, M. & Barker, J.L. (1984) Rat hippocampal neurons in culture: voltage-clamp analysis of inhibitory synaptic connections. J. Neurophysiol., 52, 469–487.
Segal, M. & Fishbein, I. (2005) Multiple actions of Brain-Derived Neuro-trophic Factor on Central Neurons. In Binder, D.K. & Scharfman, H.E. (Eds)Growth Factors and Epilepsy. Nova Science Publishers, Hauppauge, NewYork, pp. 71–82.
Segal, M. & Manor, D. (1992) Confocal microscopic imaging of [Ca2+]iin cultured rat hippocampal neurons following exposure to N-methyl-D-aspartate. J. Physiol., 448, 655–676.
Shadlen, M.N. & Newsome, W.T. (1998) The variable discharge of corticalneurons: implications for connectivity, computation, and information coding.J. Neurosci., 18, 3870–3896.
Sherwood, N.T. & Lo, D.C. (1999) Long-term enhancement of central synap-tic transmission by chronic brain-derived neurotrophic factor treatment.J. Neurosci., 19, 7025–7036.
Shirasaki, T., Klee, M.R., Nakaye, T. & Akaike, N. (1991) Differentialblockade of bicuculline and strychnine on GABA- and glycine-inducedresponses in dissociated rat hippocampal pyramidal cells. Brain Res., 561,77–83.
Singh, B., Henneberger, C., Betances, D., Arevalo, M.A., Rodriguez-Tebar, A.,Meier, J.C. & Grantyn, R. (2006) Altered balance of glutamatergic ⁄ GAB-Aergic synaptic input and associated changes in dendrite morphology afterBDNF expression in BDNF-deficient hippocampal neurons. J. Neurosci., 26,7189–7200.
Soriano, J., Rodrıguez Martınez, M., Tlusty, T. & Moses, E. (2008)Development of input connections in neural cultures. Proc. Natl. Acad.Sci. U S A, 105, 13758–13763.
Tanaka, T., Saito, H. & Matsuki, N. (1997) Inhibition of GABAA synapticresponses by brain-derived neurotrophic factor (BDNF) in rat hippocampus.J. Neurosci., 17, 2959–2966.
Turrigiano, G.G. & Nelson, S.B. (2000) Hebb and homeostasis in neuronalplasticity. Curr. Opin. Neurobiol., 10, 358–364.
Varela, J.A., Sen, K., Gibson, J., Fost, J., Abbott, L.F. & Nelson, S.B. (1997)A quantitative description of short-term plasticity at excitatory synapses inlayer 2 ⁄ 3 of rat primary visual cortex. J. Neurosci., 17, 7926–7940.
Varela, J.A., S., S., Turrigiano, G.G. & Nelson, S.B. (1999) DifferentialDepression at Excitatory and Inhibitory Synapses in Visual Cortex.J. Neurosci., 19, 4293–4304.
Vicario-Abejon, C., Collin, C., McKay, R.D. & Segal, M. (1998) Neurotrophinsinduce formation of functional excitatory and inhibitory synapses betweencultured hippocampal neurons. J. Neurosci., 18, 7256–7271.
Vicario-Abejon, C., Owens, D., McKay, R. & Segal, M. (2002) Role ofneurotrophins in central synapse formation and stabilization. Nat. Rev.Neurosci., 3, 965–974.
Wilcox, K.S., Buchhalter, J. & Dichter, M.A. (1994) Properties of inhibitoryand excitatory synapses between hippocampal neurons in very low densitycultures. Synapse, 18, 128–151.
Zukin, S.R., Young, A.B. & Snyder, S.H. (1974) Gamma-aminobutyric acidbinding to receptor sites in the rat central nervous system. Proc. Natl. Acad.Sci. U S A, 71, 4802–4807.
1010 S. Jacobi et al.
ª The Authors (2009). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 30, 998–1010


















