BB14 Best of Botswana, Namibia, Zimbabwe & South Africa in 14 Days
Upload
giovanni-cirilloCategory
view
214download
0
Biotechnology Advances 30 (2012) 223–232
Contents lists available at ScienceDirect
Biotechnology Advances
j ourna l homepage: www.e lsev ie r.com/ locate /b iotechadv
BB14, a Nerve Growth Factor (NGF)-like peptide shown to be effective in reducingreactive astrogliosis and restoring synaptic homeostasis in a rat model of peripheralnerve injury
Giovanni Cirillo a, Anna Maria Colangelo b, Maria Rosaria Bianco a, Carlo Cavaliere a, Laura Zaccaro d,Paolo Sarmientos c, Lilia Alberghina b, Michele Papa a,⁎a Laboratory of Morphology of Neural Networks, Department of Medicina Pubblica Clinica e Preventiva, Second University of Napoli, 80138 Napoli, Italyb Laboratory of Neuroscience “R. Levi-Montalcini”, Department of Biotechnology and Bioscience, University of Milano-Bicocca, Milano, Italyc PRIMM Srl, Milano, Italyd Centro Interuniversitario di Ricerca sui Peptidi Bioattivi, Department of Biological Sciences, University of Napoli “Federico II” and Istituto di Biostrutture e Bioimmagini,Consiglio Nazionale delle Ricerche, 80138 Napoli, Italy
Abbreviations: CCI, Chronic Constriction Injury; CEAAC1, excitatory amino acid carrier 1; GAD65/67, GlGFAP, Glial Fibrillary Acidic Protein; GSH, glutathione;Growth Factor-like peptide; MCB, monochlorobimaneNGF, Nerve Growth Factor; SNI, Spared Nerve Injury; vGA⁎ Corresponding author at: Department of Medicina
Institute of Human Anatomy. Second University of NaTel./fax: +39 081 296636.
E-mail address: [email protected] (M. Papa).
0734-9750/$ – see front matter © 2011 Published by Edoi:10.1016/j.biotechadv.2011.05.008
a b s t r a c t
a r t i c l e i n f oAvailable online 18 May 2011
Keywords:Neuro-glial networkNerve injuryNGF-like peptideReactive astrocytosisSynaptic homeostasis
Peptidomimetics hold a great promise as therapeutic agents for neurodegenerative disorders. We previouslydescribed a Nerve Growth Factor (NGF)-like peptide, now named BB14, which was found to act as a strongTrkA agonist and to be effective in the sciatic nerve injury model of neuropathic pain. In this report we presentthe effects of BB14 in reducing reactive astrocytosis and reverting neuroplastic changes of theglutamate/GABAergic circuitry in the lumbar spinal cord following spared nerve injury (SNI) of the sciaticnerve. Immunohistochemical analysis of spinal cord sections revealed that SNI was associated with increasedmicroglial (Iba1) and astrocytic (GFAP) responses, indicative of reactive gliosis. These changes were paralleledby (i) decreased glial aminoacid transporters (GLT1 and GlyT1) and increased levels of (ii) neuronalglutamate transporter EAAC1, (iii) neuronal vesicular GABA transporter (vGAT) and (iv) the GABAergicneuron marker GAD65/67. A remarkable increase of the Glutamate/GABA ratio and the reduction ofglutathione (GSH) levels were also indicative of modifications of glial function in neuroprotection. All thesemolecular changes were found to be linked to an alteration of endogenous NGF metabolism, as demonstratedby decreased levels of mature NGF, increase of proNGF and increased activity of NGF-degrading methallo-proteinases (MMPs). Biochemical alterations and SNI-related neuropathic behavior, characterized byallodynia and hyperalgesia, were reversed by 7-days i.t. administration of the NGF-like peptide BB14, aswell as by increasing endogenous NGF levels by i.t. infusion of GM6001, a MMPs inhibitor. All together, whileconfirming the correlation between reactive astrogliosis and perturbation of synaptic circuitry in the SNImodel of peripheral nerve injury, these data strongly support the beneficial effect of BB14 in reducing reactiveastrogliosis and restoring synaptic homeostasis under pathological conditions linked to alteration of NGFavailability and signaling, thereby suggesting a potential role of BB14 as a therapeutic agent.
NS, Central Nervous System;utamate decarboxylase 65/67;i.t., intrathecal; BB14®, Nerve; MMPs, metalloproteinases;T, vesicular GABA transporter.Pubblica Clinica e Preventiva.ples. 80100 – Naples – Italy.
lsevier Inc.
© 2011 Published by Elsevier Inc.
1. Introduction
Recently, the drug discovery process has been greatly implemen-ted by new biotechnological approaches, such as in silico libraryscreening and molecular modeling, which allow to predict properties
and activities of new chemical entities. These tools proved to beparticularly useful for neurotrophin-based interventions in neurode-generative diseases, which currently lack of effective therapies.Neurotrophins (NGF, BDNF, NT-3 and NT-4/5) regulate severalfunctions in distinct classes of neurons through interaction with twotypes of receptors: the tyrosine kinase receptors (TrkA, TrkB andTrkC) and the common p75NTR receptor. While Trk activation isessential for neuronal survival, differentiation and synaptic function,the p75NTR signaling is more complex: depending on cellularenvironment, expression of co-receptors (Nykjaer et al., 2004) andligands availability (mature neurotrophins or their precursors, such asproNGF and proBDNF), the p75NTR receptor can transduce signals thatare either pro-survival, pro-differentiation, or pro-apoptotic (Chao etal., 2006). Functional mimicry of specific pharmacophores has

224 G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
allowed the development of small peptide or non-peptide moleculesfunctioning as ligands of specific neurotrophin receptors and/ormimicking selected components of the neurotrophin signaling thatmodulate specific ligand/receptors functions. Likewise, several mol-eculesmimicking specific neurotrophin signalings have been reportedto be a valuable tool to overcome limitations of native proteins aspharmacological agents (Longo et al., 1997; Maliartchouk et al., 2000;Massa et al., 2010; Peleshok and Saragovi, 2006; Yang et al., 2008;Zaccaro et al., 2005).
Our previous studies, based on crystallographic structures of theNGF/TrkA complex (McDonald and Chao, 1995; Wiesmann et al.,1999), as well as site-specific mutagenesis (Kullander et al, 1997;Rydén and Ibànez, 1997; Shih et al., 1994) and molecular modelingstudies, led to development of several new NGF-like molecules.Among them, the NGF-mimetic peptide, now named BB14, displayedTrkA agonist activity and, like the native NGF protein, was found to beeffective in reducing biochemical and structural modifications linkedto reactive astrogliosis in the chronic constriction injury (CCI) modelof peripheral neuropathy (Colangelo et al., 2008).
The astrocytic network actively influences neuronal and synapticproperties and plasticity by providing metabolic support to neuronsand participating to reuptake and release of aminoacid transmitters(Allen and Barres, 2009; Giaume et al., 2010; Hamilton and Attwell,2010; Henneberger et al., 2010; Ni and Parpura, 2009; Rouach et al.,2008). Phenotypic changes of reactive astrocytes (Pekny and Nilsson,2005) impair neuron-astrocyte cross-talk and synaptic efficacy due tochanges of glutamate clearance and extracellular levels of gliotrans-mitters, which modulate N-methyl-D-aspartate (NMDA) receptor-mediated synaptic transmission (Panatier et al. 2006). Thesealterations are believed to contribute to progression of neurodegen-erative diseases (Cirillo et al., 2010a; Giovannoni et al., 2007; Lobsigerand Cleveland, 2007).
We previously reported that glial changes following peripheralnerve injury were associated with increased sprouting of primaryafferent nociceptive fibers (C and A-δ fibers) entering the spinal cord(Colangelo et al., 2008) and down-regulation of glial aminoacidtransporters (Cavaliere et al., 2007; Cirillo et al., 2011), suggesting astrict correlation between neuro-glial plasticity changes and periph-eral sensitization (Giaume et al., 2010; Scholz and Woolf, 2007; Todd,2010).
Superficial laminae of the dorsal horns of spinal cord represent anodal point for modulation and integration of peripheral sensorystimuli through complex networks involving glutamate receptors andlocal inhibitory GABAergic interneurons (Meisner et al., 2010). In thespared nerve injury (SNI) model of peripheral nerve lesion, werecently found, in agreement with previous reports (Polgár and Todd,2008), that nerve injury was able to perturb spinal synaptic circuitryby producing profound morpho-functional changes affecting neuro-glial interaction and the glutamatergic/GABAergic system (Cirillo etal., 2011).
Based on our previous findings regarding the ability of intrathe-cal (i.t.) administration of NGF or the NGF-like peptide BB14 torestrain reactive gliosis (Cirillo et al., 2010b, 2011; Colangelo et al.,2008), we evaluated the effect of this molecule in restoring spinalsynaptic homeostasis by modulating the expression of glial (GLT1and GlyT1) and neuronal excitatory aminoacid transporters(EAAC1). To further characterize the role of BB14 in reverting theeffects of decreased NGF availability, we also compared the efficacyof BB14 with the effect of increasing endogenous NGF by inhibitingits proteolytic degradation by the MMPs system (Bruno and Cuello,2006). We show that both i.t. BB14 supply and GM6001-mediatedinhibition of MMPs restored neuronal circuitry and metabolichomeostasis, thus providing a strong evidence of the efficacy ofthis NGF-mimetic molecule in targeting pathological conditionslinked to alteration of NGF availability/signaling and mechanisms ofreactive astrogliosis.
2. Materials and methods
2.1. Animals
Adult (250–300 g; Charles River, Calco, Italy) male SpragueDawley rats (n=40) were used. Rats were maintained on a 12/12 hlight/dark cycle and allowed free access to food and water. Eachanimal was housed under specific pathogen-free conditions in iron-sheet cages with solid floors covered with 4–6 cm of sawdust. Cageswith thin-plate floors were avoided on the assumption that theywould exacerbate the discomfort arising from the affected hind paw(Bai et al., 1999). All surgerier and experimental procedures wereperformed during the light cycle and were approved by the AnimalEthics Committee of the Second University of Naples. Animal care wasin compliance with Italian (D.L. 116/92) and EC (O.J. of E.C. L358/118/12/86) regulations on the care of laboratory animals. All effortswere made to reduce animal numbers.
2.2. SNI model
Spared sciatic nerve injury was made according to the method ofDecosterd and Woolf (2000). Briefly, each rat was anaesthetized withchlorydrate tiletamine (30 mg/kg) during surgery. The skin on thelateral surface of the thigh was incised and a section was madedirectly through the biceps femoris muscle exposing the sciatic nerveand its three terminal branches: the sural, common peroneal andtibial nerves. The SNI procedure involved the axotomy and ligation ofthe tibial and common peroneal nerves leaving the sural nerve intact.For the sham-operated control group (CTR), nerves were exposed butnot truncated. Great care was taken to avoid any contact with orstretching of the intact sural nerve. Muscle and skin were closed intwo layers.
2.3. Drug delivery
To reduce the bias of discomfort caused by lumbar spinal catheter,the chronic intrathecal lumbar spinal catheter was positioned duringthe SNI surgery, according to the method described previously(Decosterd and Woolf, 2000). Briefly, a small opening was madethrough the laminas of the lumbar tract of the spine and a catheter[polyethylene (PE) 10 tubing attached to PE 60 tubing for connectionto an osmotic pump] was inserted into the subarachnoid space anddirected to the lumbar enlargement of the spinal cord. After anchoringthe catheter across the careful apposition of a glass ionomer lutingcement triple pack (Ketac Cem radiopaque; 3 M ESPE, Seefeld,Germany), the wound was irrigated with saline and closed in twolayers with 3–0 silk (fascial plane) and surgical skin staples. Onrecovery from surgery, lower body paralysis was induced byintrathecal lidocaine (2%) injection to confirm proper catheterlocalization. Each rat was placed on a table, and the gait and postureof the affected hind paw were carefully observed for 2 min. Onlyanimals exhibiting appropriate, transient paralysis to lidocaine, aswell as lack of motor deficits, were used for treatments [BB14, orGM6001, or artificial CSF (ACSF) infusion; n=10/each group] andbehavioural testing. Three-days neuropathic rats were anesthetizedby intraperitoneal chlorydrate tiletamine (30 mg/kg), the freeextremity of the catheter was connected to an osmotic minipump,and the pump was implanted subcutaneously. Osmotic pumpsattached to intrathecal lumbar spinal catheters were filled with theNGF-like peptide BB14 (37,5 μg/μl) in ACSF containing rat serumalbumin (1 mg/ml; Sigma, Italy), or GM6001 (Calbiochem, Germany)(180 μg/μl, corresponding to 100 mg/kg body weight), or vehicle only(ACSF). The osmotic pumps were model 2001 Alzet (Cupertino, CA)pumps, which pumped at a rate of 1 μl/h for 7 d. This rate produced anintrathecal infusion dose of 37,5 μg/h of BB14, and 180 μg/h ofGM6001.

225G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
2.4. Behavioural testing
Only animals exhibiting no motor deficits were used forbehavioural tests. Animals were habituated to the testing environ-ment daily for at least 2 days before baseline testing. The experi-mental groups that underwent SNI lesion were behaviorally testedon day 0 (before surgery and i.t. lumbar spinal catheter positioning),day 3 (3 days after SNI), and day 10 (7 days after the implant of thepump). On day 10 all animals were sacrificed. Mechanical allodyniawas assessed by using the von Frey filaments (Ugo Basile, Comerio,Italy) according to the method described by Chaplan et al. (1994).Animals were allowed to habituate for 30 min before testing.Filaments were applied in either ascending or descending strengthas necessary to determine the filament closest to the threshold ofresponse. The time of response to a progressive force applied to hindpaw limb (30 g in 20 sec) was evaluated six times using the injuredhind limb, with an interval of 5 min between stimulations. Thethreshold was the lowest force that evoked a consistent, brisk,withdrawal response. Thermal nociceptive thresholds were mea-sured using a device based on the design by Hargreaves et al. (1988).Animals were allowed to habituate for 30 min before testing. Paw-withdrawal latency in response to radiant heat (infra-red) wasmeasured using the plantar test apparatus (Ugo Basile, Comerio,Italy). The heat source was positioned under the plantar surface ofthe affected hind paw and activated at a setting of 7.0. The digitaltimer connected to the heat source automatically recorded theresponse latency of paw withdrawal to the nearest 0.1 sec. Theintensity of the infra-red light beam was chosen to give baselinelatencies of 15 sec. in control rats. A cut-off time of 20 sec wasimposed to prevent tissue damage. The injured hind limb was testedtwice at each time point, with an interval of 5 min betweenstimulations. All testing was performed blind.
2.5. Tissue preparation
Rats were deeply anesthetized by intraperitoneal injection ofchloral hydrate (300 mg/kg body weight) and perfused transcardiallywith saline solution (Tris HCl 0.1 M/ EDTA 10 mM) followed by 4%paraformaldehyde/0.1% glutaraldehyde in 0.01 M phosphate-buffer(PBS), pH 7.4 at 4 °C. For light microscopy, spinal cords were removedand post-fixed for two hr in the same fixative, then soaked in 30%sucrose/PBS and frozen in chilled isopentane on dry ice. Serial sections(25 μm thickness) were cut with a slide microtome and collected incold PBS for immunohistochemistry.
2.6. Antibodies
The following antibodies were used for immunodetection:mouse antibodies directed against Glial Fibrillary Acidic Protein(GFAP) (1:400; Sigma-Aldrich Milano, Italy); rabbit antibodies toionized calcium binding adaptor molecule 1 (Iba1) (1:500; WakoChemicals, VA, USA); guinea pig antibodies to glutamate transporter(GLT1) (1:200; Chemicon Inc Temecula, CA, USA); goat antibodies toglycine transporter 1 (GlyT1) (1:1000; Chemicon Inc Temecula, CA,USA); guinea pig antibodies raised against vesicular glutamatetransporter 1 (vGLUT1) (1:5000; Chemicon Inc Temecula, CA, USA);mouse antibodies to vesicular GABA transporter (vGAT) (1:500;Synaptic Systems, Gottingen, Germany); rabbit antibodies againstGlutamic Acid Decarboxylase 65/67 (GAD65/67) (1:1000; Sigma-Aldrich, Milano, Italy); goat antibodies to neuronal glutamatetransporter EAAC1 (1:4000; Chemicon Inc Temecula, CA, USA);rabbit antibodies against NGF (1:250; Chemicon Temecula, CA,USA); rabbit antibodies against proNGF (1:250; Sigma-Aldrich,Milano, Italy).
2.7. Spinal cord immunohistochemistry
Spinal cord sections were blocked in 10% normal serum in 0.01 MPBS/0.25% Triton-X100 for 1 h at room temperature (RT). Eachprimary antibody (GFAP, Iba1, GLT1, GlyT1, EAAC1, NGF, proNGF)was diluted in 0.01 M PBS containing 10% normal serum and 0.25%Triton. Following incubation for 48 hr at 4 °C, sections were washedseveral times in PBS and incubated with the appropriate biotinylatedsecondary antibody (1:200; Vector Labs Inc., Burlingame, CA, USA) for90 min at RT, washed in PBS and processed with the Vectastainavidin-biotin peroxidase kit (Vector Labs Inc., Burlingame, CA, USA)for 90 min at RT. Sections were then washed in 0.05 M Tris-HCl andreacted with 3.3-diaminobenzidine tetrahydrochloride (DAB; Sigma,0.5 mg/ml in Tris-HCl) and 0.01% hydrogen peroxide. Sections weremounted on chrome-alume gelatine coated slides, dehydrated andcoverslipped. Immunofluorescence staining was performed as de-scribed previously (Papa et al., 2003). Sections were incubated withthe primary antibody (vGLUT1, vGAT and GAD65/67) for 48 hr at 4 °C.Following incubation with primary antibodies, sections were incu-bated with the appropriate secondary antibody (Alexa Fluor 488 anti-guinea pig IgG, Alexa Fluor 488 anti-mouse IgG, Alexa Fluor 546 anti-rabbit IgG and Alexa Fluor 488 anti-rabbit IgG (1:200; Invitrogen,Carlsbad, CA, USA) for 2 hr. Sections were mounted and coverslippedwith Vectashield (Vector Laboratories).
2.8. In situ zymography for MMPs activity
In situ gelatinolytic activity was performed on frozen lumbar spinalcord sections (25 μm thick) using a commercial kit (EnzCheckGelatinase Assay Kit; Molecular Probes, Invitrogen, Carlsbad, CA,USA). Sections were incubated with DQ gelatin conjugate, thefluorogenic substrate, at 37 °C overnight and, after washing, fixed in4% paraformaldehyde in PBS. Cleavage of DQ gelatin by proteasesresulted in a green fluorescent product and was analyzed by confocalmicroscopy.
2.9. GSH assay
After washing with PBS, frozen lumbar spinal cord sections wereincubated overnight in ACSF containing 40 μM of monochlorobimane(MCB), a thiol reactive reagent, and then transferred to fresh ACSF for30 min. Slides were fixed in 4% paraformaldehyde and then analyzedby confocal microscopy.
2.10. Confocal microscopy
vGLUT, vGAT and GAD65/67 expression levels and proteasesactivity were analyzed by using a laser scanning microscope Zeiss(Oberkochen, Germany) LSM 510 Meta. Confocal images of dorsalhorns of lumbar spinal cords were acquired. Images were captured ata resolution of 512x512 pixels. The appropriate argon laser fluores-cence for visualization of the vGLUT1 and vGAT was used with anexcitation wavelength of 488 nm and emission filter bandpass 505–530 nm. The HeNe laser fluorescence for the GAD65/67 signal with anexcitation wavelength of 546 nm and emission filter long-pass560 nm was used. GSH levels were analyzed with a UV laser(351 nm, 364 nm).
2.11. HPLC analysis of aminoacids
Aminoacids levels were analyzed by RP-HPLC analysis using anAgilent 1200 Series Liquid Chromatograph, equipped with a binarypump delivery system (G1312B), robotic autosampler (G1317B),column thermostat (G1316A) and multi-wavelength detector(G1315B). Briefly, tissues samples were diluted in borate buffer(0.15 M, pH=10.2), followed by addition of the derivatization

226 G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
solution [o-Phthalaldehyde (OPA) (10 mg/ml), β-mercaptoethanol(10 mg/mL)] and diluent solution (mobile phase A: 1.5% v/v H3PO4).After derivatization, the mixture (20 μl) was injected on a reverse-phase Jupiter 5 μm C18 300 Å (250 mm x 4.6 mm) column at 40 °Cand derivatives absorption detected at 338 nm. Separation wasobtained at a flow rate of 1 mL/min with a gradient of the mobilephase A [Na2HPO4 (10 mM)/Na2B4O7.10 H2O (10 mM)] and phase B[methanol:acetonitrile:water (9:9:2, v:v:v)]. Spinal cord sampleswere automatically derivatized by the robotic autosampler andanalyzed blindly by using amino acid standard samples [glutamicacid, glutamine, glycine and gamma-aminobutyric acid (GABA),0.25 mM each]. Method reproducibility was assessed by analyzingall samples for five consecutive times. Aminoacid concentrations wereexpressed as the mean of the peak areas, and the standard error of themean (SEM) and coefficient of variation (% CV) were calculated.Results were reported as the ratio between the percentage of the areasversus the total area of the investigated amino acids.
2.12. Measurements and Statistical analysis
Slides were imaged with a Zeiss Axioskope 2 light microscopeequipped with a high-resolution digital camera (C4742-95, Hama-matsu Photonics, Italia). Measurements of markers in the dorsal hornof spinal cords were performed by using a computer assisted imageanalysis system (MCID 7.0; Imaging Res. Inc, Canada). For glialmarkers the morphometric approach was preferred because of theperfect visualization of single positive elements. Therefore, values ofGFAP and Iba1, markers for astrocytes and microglia respectively,were expressed as proportional areas: number of positive elementsrelative to the scanned area. The densitometric values of GLT1, GlyT1,EAAC1, NGF and proNGF were expressed as the total target measuredarea relative to the scanned area. For confocal images analysis, thedensitometric method by MCID was used for vGLUT1, vGAT,GAD65/67 and GSH, while the morphometric approach was appliedfor in situ zymography analyses. Averages were obtained from fiverandomly selected spinal cord sections for each animal, andcomparisons were made between treatments (BB14, and GM6001)versus control groups (ACSF and CTR). Data were exported andconverted to frequency distribution histograms by using the Sigma-Plot 10.0 program (SPSS Erkrath Germany).
Fig. 1. Evaluation of glial markers in the dorsal horn of spinal cord. Sections of the dorsal hotreated for 7 days with BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF (vehicle),separate experiments with five independent samples (n=10 for each treated groups; n=10test). Scale bar: 50 μm, objective 40x.
Data from all quantitative analyses were analyzed by one-wayANOVA, using all pairwise Holm-Sidak method for multiple compar-isons (*p≤0,01; **p≤0.001). All data shown are presented as themean±SEM. Individual images of control and treated rats wereassembled and then the same adjustments were made for brightness,contrast and sharpness using Adobe Photoshop (Adobe Systems, SanJose, CA).
3. Results
3.1. The NGF-like peptide BB14 reduces reactive gliosis induced by SNI ofperipheral nerves
We have previously reported the activity of the peptide BB14 inrestoring neuro-glial rearrangement following CCI of the rat sciaticnerve to an extent similar to that of NGF (Colangelo et al., 2008). Tofurther investigate mechanisms underlying the activity of this NGFmimetic molecule, we have used in this report the SNI model ofperipheral nerve injury. As previously indicated (Cirillo et al., 2010b,2011), peripheral nerve injury correlated with increased expression ofglial markers indicative of reactive astrogliosis. Indeed, immunohisto-chemical analysis of glial markers in the dorsal horns of lumbar spinalcord sections revealed the presence ofmarked gliosis 3 days after SNI, asdemonstrated by the strong increase of GFAP staining (18.02±0.73)(Fig. 1A-B) and Iba1 levels (4.62±0.23) (Fig. 1C-D) in the ACSF-treatedanimals, as compared to the CTR group (GFAP: 9.42±0.34; Iba1: 1.58±0.27) (p≤0.001). GFAP and Iba1 levels were reduced by i.t. treatmentwith BB14 (37,5 μg/μl/hr) for 7 days to 12.73±0.46 and 3.03±0.31,respectively (p≤0.001, BB14 vs ACSF). These data, by establishing therole of BB14 in reducing reactive astrocytosis andmicroglial recruitmentto an extent similar to that elicited by NGF (Cirillo et al., 2011), clearlysupport the notion that BB14 can modulate reactive gliosis, a processthat is involved in several neurodegenerative diseases (Lobsiger andCleveland, 2007).
3.2. NGF and proNGF content in the lumbar spinal cord
To better understand the role played by NGF in reducing glialactivation, we analyzed NGF levels in the dorsal horns of lumbar spinalcord of SNI-injured animals. Moreover, since the decrease of endoge-nous NGF might be due to increased MMPs activity (Fig. 2) (Bruno and
rns of lumbar spinal cords were prepared from sham-operated (CTR) and SNI animalsand immunostained for Iba1 (A-B) and GFAP (C-D). Data are the mean±SEM of twofor CTR group); (**p≤0.001, ACSF vs BB14, ACSF vs GM6001; ANOVA and Holm-Sidak

Fig. 2. Schematic overview of NGF degradation by the tPA/plasminogen/plasmin/MMPdegrading system. Plasmin, resulting from tPA/plasminogen activation, is required forproNGF processing to mature NGF. On the other hand, plasmin promotes the cleavageof proMMP to its mature form (MMP), which rapidly degrades NGF protein. MMPs-dependent degradation of NGF is inhibited by GM6001, a generic MMPs inhibitor.
227G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
Cuello, 2006), we examined the effect of increasing endogenous NGF byinhibiting the plasminogen/plasmin/MMPs degrading system byGM6001 (Fig. 2). Immunohistochemical analyses revealed that endog-enous NGF levels were reduced in the ACSF group (25.54±3.01) and
Fig. 3. Immunohistochemistry for endogenous NGF (A-B) and proNGF (C-D) expression in thfrom sham-operated (CTR) and SNI animals treated for 7 days with BB14 (37.5 μg/μl/hr) or G(C-D) antibodies. (E-F) In situ zymography for MMPs proteolytic activity. In situ zymographhorns of lumbar spinal cords prepared from sham-operated (CTR) and SNI animals treated foexpressed as the mean±SEM of two separate experiments with five independent samples
BB14-treated rats (22.32±3.23) as compared to CTR animals (40.02±2.75) (Fig. 3A-B). The reduction of NGF content in SNI-injured animalswas counterbalanced by a strong increase of proNGF levels, as indicatedby the immunostaining for proNGF in ACSF rats (41.14±3.13),as compared to CTR animals (23.05±3.52) (Fig. 3C-D). Interestingly, a7-days i.t. treatment with BB14 also restored proNGF to the basal levels(20.31±2.56) (Fig. 3C-D) (p≤0.001, BB14 vs ACSF). As expected,modification of NGF/proNGF levels following SNI injury was paralleledby a strong induction of the proteolytic activity due to MMPs (Fig. 2)(Cirillo et al., 2010b; Cuello et al., 2007). Indeed, in situ zymography oflumbar spinal cord sections revealed a strong increase of thegelatinolytic activity in the ACSF group (0.16±0.04), as comparedto CTR animals (0.11±0.004) (p≤0.001) (Fig. 3E-F). Treatment withBB14 was able to reduce MMPs activity (0.12±0.005) to CTR values(Fig. 3E-F) (p≤0.001, BB14 vs ACSF).
To further dissect mechanisms underlying BB14 activity, wepharmacologically blocked the proteolytic activity of the MMPssystem to increase endogenous NGF concentrations. Indeed, i.t.administration of GM6001, a generic MMPs inhibitor, restored NGFlevels in SNI animals, as indicated by the densitometric value of NGFstaining (37.42±2.02) (p≤0.001, GM6001 versus ACSF) (Fig. 3A-B).The increase of endogenous NGF levels was paralleled by a reductionof proNGF expression after a 7-days i.t. administration of GM6001(24.23±2.43), as compared to ACSF-treated animals (p≤0.001)(Fig. 3C-D). The efficacy of GM6001 on NGF protein content throughMMPs inhibition was also confirmed by in situ zymography. Confocalmicroscopy of spinal cord sections showed that the proteolyticactivity was fully abolished in SNI-rats treated with GM6001 (0.10±0.03), compared to ACSF-treated rats (0.16±0.04) (Fig. 3E-F)(p≤0.001).
Restoring the NGF content by i.t. treatment of SNI animals withGM6001 had the same effect on glial markers as that induced by i.t.administration of BB14. In fact, i.t. infusion of GM6001 to SNI animals
e lumbar spinal cord. Sections of the dorsal horns of lumbar spinal cords were preparedM6001 (180 μg/μl/hr) or ACSF (vehicle), and immunostained with NGF (A-B) or proNGFy for proteolytic activity was performed as described in M&M on sections of the dorsalr 7 days with BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF (vehicle). Data are(**p≤0.001, ACSF vs GM6001; ANOVA and Holm-Sidak test). Scale bar: 50 μm.
228 G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
for 7 days reduced both GFAP (11.35±0.43) (Fig. 1A-B) and Iba1(2.91±0.32) staining (Fig. 1C-D) (p≤0.001, GM6001 vs ACSF).
3.3. Reactive gliosis-induced changes of spinal synaptic homeostasis arereverted by i.t. BB14 administration
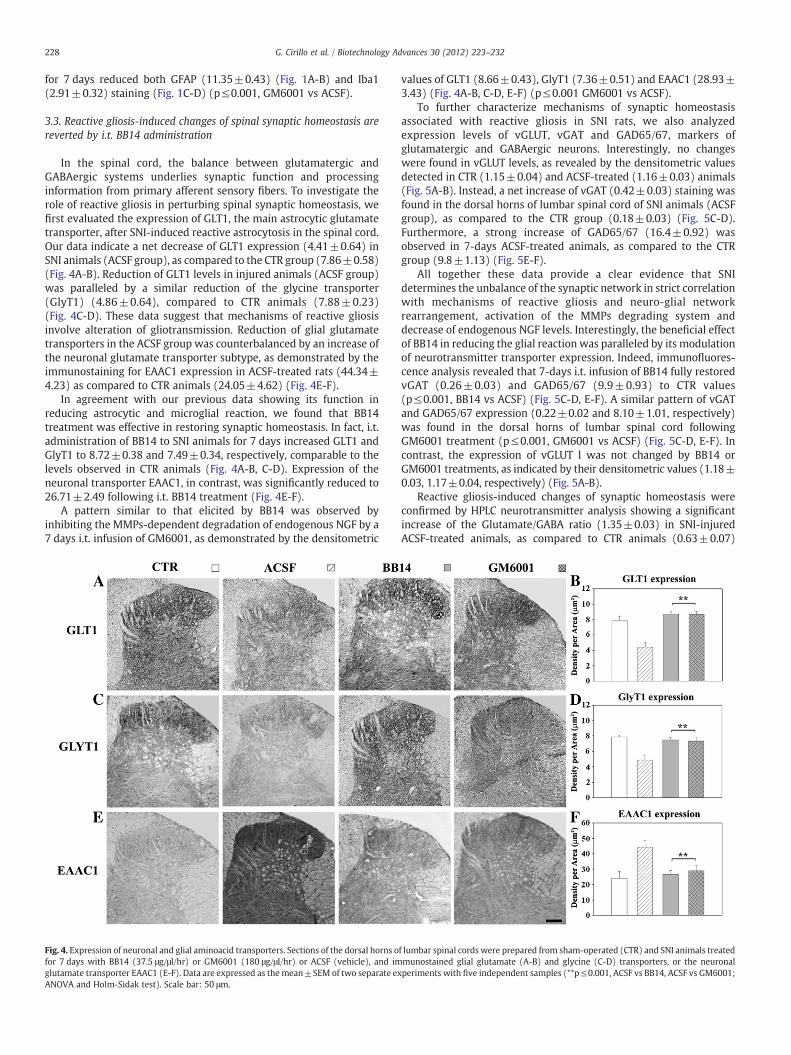
In the spinal cord, the balance between glutamatergic andGABAergic systems underlies synaptic function and processinginformation from primary afferent sensory fibers. To investigate therole of reactive gliosis in perturbing spinal synaptic homeostasis, wefirst evaluated the expression of GLT1, the main astrocytic glutamatetransporter, after SNI-induced reactive astrocytosis in the spinal cord.Our data indicate a net decrease of GLT1 expression (4.41±0.64) inSNI animals (ACSF group), as compared to the CTR group (7.86±0.58)(Fig. 4A-B). Reduction of GLT1 levels in injured animals (ACSF group)was paralleled by a similar reduction of the glycine transporter(GlyT1) (4.86±0.64), compared to CTR animals (7.88±0.23)(Fig. 4C-D). These data suggest that mechanisms of reactive gliosisinvolve alteration of gliotransmission. Reduction of glial glutamatetransporters in the ACSF group was counterbalanced by an increase ofthe neuronal glutamate transporter subtype, as demonstrated by theimmunostaining for EAAC1 expression in ACSF-treated rats (44.34±4.23) as compared to CTR animals (24.05±4.62) (Fig. 4E-F).
In agreement with our previous data showing its function inreducing astrocytic and microglial reaction, we found that BB14treatment was effective in restoring synaptic homeostasis. In fact, i.t.administration of BB14 to SNI animals for 7 days increased GLT1 andGlyT1 to 8.72±0.38 and 7.49±0.34, respectively, comparable to thelevels observed in CTR animals (Fig. 4A-B, C-D). Expression of theneuronal transporter EAAC1, in contrast, was significantly reduced to26.71±2.49 following i.t. BB14 treatment (Fig. 4E-F).
A pattern similar to that elicited by BB14 was observed byinhibiting the MMPs-dependent degradation of endogenous NGF by a7 days i.t. infusion of GM6001, as demonstrated by the densitometric
Fig. 4. Expression of neuronal and glial aminoacid transporters. Sections of the dorsal horns ofor 7 days with BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF (vehicle), and imglutamate transporter EAAC1 (E-F). Data are expressed as the mean±SEM of two separate exANOVA and Holm-Sidak test). Scale bar: 50 μm.
values of GLT1 (8.66±0.43), GlyT1 (7.36±0.51) and EAAC1 (28.93±3.43) (Fig. 4A-B, C-D, E-F) (p≤0.001 GM6001 vs ACSF).
To further characterize mechanisms of synaptic homeostasisassociated with reactive gliosis in SNI rats, we also analyzedexpression levels of vGLUT, vGAT and GAD65/67, markers ofglutamatergic and GABAergic neurons. Interestingly, no changeswere found in vGLUT levels, as revealed by the densitometric valuesdetected in CTR (1.15±0.04) and ACSF-treated (1.16±0.03) animals(Fig. 5A-B). Instead, a net increase of vGAT (0.42±0.03) staining wasfound in the dorsal horns of lumbar spinal cord of SNI animals (ACSFgroup), as compared to the CTR group (0.18±0.03) (Fig. 5C-D).Furthermore, a strong increase of GAD65/67 (16.4±0.92) wasobserved in 7-days ACSF-treated animals, as compared to the CTRgroup (9.8±1.13) (Fig. 5E-F).
All together these data provide a clear evidence that SNIdetermines the unbalance of the synaptic network in strict correlationwith mechanisms of reactive gliosis and neuro-glial networkrearrangement, activation of the MMPs degrading system anddecrease of endogenous NGF levels. Interestingly, the beneficial effectof BB14 in reducing the glial reaction was paralleled by its modulationof neurotransmitter transporter expression. Indeed, immunofluores-cence analysis revealed that 7-days i.t. infusion of BB14 fully restoredvGAT (0.26±0.03) and GAD65/67 (9.9±0.93) to CTR values(p≤0.001, BB14 vs ACSF) (Fig. 5C-D, E-F). A similar pattern of vGATand GAD65/67 expression (0.22±0.02 and 8.10±1.01, respectively)was found in the dorsal horns of lumbar spinal cord followingGM6001 treatment (p≤0.001, GM6001 vs ACSF) (Fig. 5C-D, E-F). Incontrast, the expression of vGLUT l was not changed by BB14 orGM6001 treatments, as indicated by their densitometric values (1.18±0.03, 1.17±0.04, respectively) (Fig. 5A-B).
Reactive gliosis-induced changes of synaptic homeostasis wereconfirmed by HPLC neurotransmitter analysis showing a significantincrease of the Glutamate/GABA ratio (1.35±0.03) in SNI-injuredACSF-treated animals, as compared to CTR animals (0.63±0.07)
f lumbar spinal cords were prepared from sham-operated (CTR) and SNI animals treatedmunostained glial glutamate (A-B) and glycine (C-D) transporters, or the neuronalperiments with five independent samples (**p≤0.001, ACSF vs BB14, ACSF vs GM6001;

Fig. 5. Expression of vGLUT, vGAT and GAD65/67 in dorsal horn of lumbar spinal cord. Confocal images of dorsal horns of the lumbar spinal cord immunostained for vGLUT (A-B),vGAT (C-D) and GAD65/67 (E-F). Sections of the dorsal horns of lumbar spinal cords were prepared from sham-operated (CTR) and SNI animals treated for 7 days with BB14(37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF (vehicle). Following immunostaining, sections were analyzed by confocal microscopy. Data are expressed as the mean±SEM oftwo separate experiments with five independent samples (**p≤0.001, ACSF vs BB14, ACSF vs GM6001; ANOVA and Holm-Sidak test). Scale bar: 10 μm.
229G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
(Fig. 6). In agreement with the immunohistochemical data, theincrease of Glutamate/GABA ratio was significantly reduced by i.t.BB14 or GM6001 to 0.95±0.04 and 0.90±0.02, respectively(p≤0.001), strongly supporting the beneficial role of the NGF-likepeptide BB14 in maintaining the synaptic network followingperipheral nerve injury through modulation of glial Glutamate/GABAtransporters and neurotransmitters levels at the synaptic cleft.
3.4. BB14 preserves the function of the astrocytic neuroprotective GSHsystem
Glutamate and glycine uptake through GLT1 and GLYT1 isimportant for both normal gliotransmission and neuroprotection. Infact, besides clearing glutamate from synaptic cleft, thus protectingneurons from glutamate excitotoxicity, astrocytes are an essential
Fig. 6. HPLC analysis of Glutamate/GABA ratio. Amino acid levels were measured byHPLC in the dorsal horns of lumbar spinal cords dissected from sham-operated (CTR)and SNI animals treated for 7 days with BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr)or ACSF (vehicle). The Glutamate/GABA ratio was calculated as described in M&M. Dataare expressed as the mean±SEM of two separate experiments with five independentsamples (**p≤0.001, ACSF vs BB14, ACSF vs GM6001; ANOVA and Holm-Sidak test).
source of GSH for neurons and provide themselves the aminoacidiccomponents (glutamate, glycine and cysteine) for glutathione (GSH)synthesis (Shih et al., 2006). To examine the role of reactive gliosis inreducing the main neuronal antioxidant defense, we analyzed GSHlevels by confocal microscopy following MCB staining. In agreementwith the observed down regulation of GLT1 and GlyT1, we found astrong reduction of GSH levels in ACSF-treated rats (2.63±0.45), ascompared to the CTR group (6.78±0.36) (Fig. 7A-B). Consistent withthe role of BB14 in restoring GLT1 and GlyT1 expression, i.t. BB14treatment increased GSH levels to 7.69±0.34 (p≤0.001) (Fig. 7A-B).I.t. treatment with GM6001 also increased GSH levels to 8.22±0.04,thus establishing a strong correlation between decreased GSH andendogenous NGF levels.
These data confirm the relevance of astrocytes in maintenance ofneuronal homeostasis and clearly indicate a role of BB14 inneuroprotection, a mechanisms that appears to involve modulationof glial function.
3.5. I.t. administration of BB14 reduces the neuropathic pain behavior inSNI rats
I.t. delivery of NGF or the NGF-like peptide was found to restorethe neuro-glial network in the CCI model of neuropathic pain (Cirilloet al., 2010b, 2011; Colangelo et al., 2008). To further investigate therelevance of neuro-glial alterations in neuropathic pain, SNI animalswere tested for neuropathic pain behavior on days 0, 3, and 10 afterSNI surgery by analyzing mechanical and thermal sensitivity, aspreviously described (Cirillo et al., 2010b; Colangelo et al., 2008). Themean baseline of normal mechanical threshold before SNI (day 0) was26.95±0.52 g across the different experimental groups. In CTRanimals this value was unmodified on day-3 and day-10 showing amechanical threshold of 27.12±0.32 g and 27.11±0.42 g, respec-tively (Fig. 8A). SNI lesion determined the appearance of a typicalneuropathic behavior (ACSF group), as indicated by a significantreduction of mechanical nociceptive threshold (10.19±0.46) on day-3 after surgery, indicative of an allodynic state. The Hargreaves test

Fig. 7. GSH levels in the dorsal horns of lumbar spinal cords. GSH levels were measured by the MCB assay as described in M&M in the dorsal horns of lumbar spinal cords dissectedfrom sham-operated (CTR) and SNI animals treated for 7 days with BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF (vehicle). Data are expressed as the mean±SEM of twoseparate experiments with five independent samples (**p≤0.001, ACSF vs BB14, ACSF vs GM6001; ANOVA and Holm-Sidak test). Scale bar: 50 μm.
230 G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
also showed a strong reduction of the reaction time to the thermalstimulus 3 days after SNI with a very short time-response to infraredstimulation (6.18±0.90 sec), as compared to the basal values(16.23±0.72 sec) of normal thermal sensitivity (Fig. 8B), indicatingthe onset of a hyperalgesic state. Hargreaves test recordings in theCTR group were almost unmodified on day-3 (16.21±0.62) andday-10 (16.24±0.44) after SNI surgery. The hyperalgesic behaviorwas still evident in the ACSF-treated animals and no improvementwas observed on day-10, after 7 days of vehicle infusion (6.28±0.57). A 7-days i.t. administration of BB14, instead, significantlyrestored both mechanical (22.24±0.73) and thermal (12.46±0.49)sensitivity in SNI-operated rats, as compared to ACSF treated animals
Fig. 8. Antinociceptive effect of i.t. administration of BB14 peptide, GM6001. SNI- andsham-operated rats were tested for responses to the von Frey (A) and the Plantar test(B) for baseline sensitivity (day 0), three days after SNI (day 3) and after seven days ofi.t. infusion (day 10) of BB14 (37.5 μg/μl/hr) or GM6001 (180 μg/μl/hr) or ACSF(vehicle). Data are expressed as the mean±SEM of two separate experiments with fiveindependent samples. (**p≤0.001, BB14, and GM6001 vs ACSF; ANOVA and Holm-Sidak test).
(13.15±0.46 and 6.28±0.57, respectively) (p≤0.001, BB14 vs ACSF)(Fig. 8A-B).
Based on relevance of the tPA/plasminogen/MMP system in theproteolytic degradation of NGF and generation of the histochemicaland biochemical alterations described above, we have also analyzedthe effect of increasing endogenous NGF by i.t. administration ofGM6001. Indeed, we found that i.t. administration of GM6001 for7 days to SNI animals restored mechanical sensitivity to 26.45±0.70(Fig. 8A) and reduced thermal sensitivity to 12.55±0.70 (Fig. 8B)(p≤0.001, GM6001 vs ACSF). These data strongly suggest thatmechanisms underlying chronic pain may involve a decrease of NGFavailability.
4. Discussion
Development of neurotrophin-based therapeutics is regarded as apromising strategy to target neurodegenerative diseases in whichneuroprotection is compromised. The therapeutic potential of NGFhas been established by a large number of preclinical and clinicalstudies in peripheral neuropathies and Alzheimer's disease (Apfel,2002; Tuszynski et al., 2005). However, its pharmacological applica-tions are hindered by its poor pharmacokinetics and bioavailability atthe desired targets (Poduslo and Curran, 1996). Our previousmolecular modeling studies led to the synthesis of the NGF-likepeptide BB14, that behaved as a strong TrkA agonist and, like thenative NGF protein, was found to be effective in reducing hallmarks ofneuropathic pain following CCI (Colangelo et al., 2008). In this report,we further refined the functional characterization of BB14 byestablishing its efficacy in targeting NGF-dependent maintenance ofneuronal homeostasis, notably, its efficacy in reducing reactive gliosisand synaptic modifications following peripheral nerve injury.
We previously reported that reactive astrocytosis following CCIdetermined increased sprouting of primary afferent nociceptive C-and Aδ- fibers (Colangelo et al., 2008), morphological alterations ofnerve myelination and dorsal root ganglia architecture (Cirillo et al.,2010b), down-regulation of glial aminoacid transporters (Cavaliere etal., 2007; Cirillo et al., 2011) and changes of neurotrophin receptorsexpression (Cirillo et al., 2010b). This new morphological andmolecular structure establishes a strict correlation between neuro-glial plasticity changes and peripheral sensitization, and produces inthe spinal cord an adaptive plasticity that facilitates neuropathic paintransmission (Cirillo et al., 2011; Wang et al., 2009).
In the spinal cord, nociceptive processing relies on modulation andintegration of a complex network involving glutamate transmission andlocal inhibitory GABAergic interneurons (Bai et al., 1999; Yasaka et al.,2010). Indeed, we have found that the onset of reactive gliosis followingSNI (increase of Iba1 and GFAP) (Fig. 1) was paralleled by remarkablechanges of glial and neuronal neurotransmitter transporters, as shownby the reduction of glial glutamate and glycine transporters, andincrease of the neuronal glutamate transporter EAAC1 (Fig. 4) andvesicular GABA transporter vGAT (Fig. 5). The increase of theGlutamate/GABA ratio (Fig. 6), counterbalanced by an evident sprouting

231G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
of the GABAergic neurons (increase of GAD65/67 staining) (Fig. 5), alsoaccounted for a remarkable unbalance of the Glutamatergic/GABAergiccompartments. Under these conditions, the decrease of glutamate andglycineuptake in SNI animals (Fig. 4)mightbe responsible for the failureof the twomain neuroprotective functions of astrocytes in the tripartitesynapse: clearing excess of glutamate from synaptic clefts and providingneuronswithGSH, themajor antioxidant system. Indeed, the substantialreduction of GSH levels in the ACSF-SNI animals (Fig. 7) was consistentwith the reduced transport of glutamate and glycine, two aminoacidicconstituents of GSH. Notably, all these molecular changes weresubstantially reduced by the i.t. administration of BB14 (Figs. 1 and4–7). In addition, restoring synaptic homeostasis by continuous i.t.BB14 treatment also reduced mechanical and thermal sensitivity,that are the hallmarks of neuropathic behavior following nerve injury(Fig. 8).
The relevance of the NGF-like activity of BB14 in revertingmorpho-molecular changes of reactive astrocytes and modificationof synaptic homeostasis was confirmed by the observation thatendogenous NGF levels were reduced following SNI, together with astrong increase of proNGF levels (Fig. 3). These findings are inagreement with previous evidence of altered NGF/proNGF ratiofollowing CNS lesions (Harrington et al., 2004). Regulation ofproNGF/NGF conversion occurs in the extracellular space throughthe tPA/plasminogen/plasmin/MMPs system (Bruno and Cuello,2006). Restoring endogenous NGF levels by i.t. injection of theMMPs generic inhibitor GM6001 (Figs. 2 and 3) reduced reactivegliosis (Fig. 1), normalized the expression of neuronal and glialglutamate and glycine transporters (Fig. 4) and the GABAergicmarkers (vGAT and GAD65/67) (Fig. 5), and reduced neuropathicbehavior (Fig. 8) to an extent similar to that achieved by continuousi.t. BB14 infusion. Interestingly, the effect of both BB14 and GM6001treatments in modulating neuronal, glial and vesicular transporterswas paralleled by a concomitant adjustment of both synaptichomeostasis (reduction of the Glutamate/GABA ratio) (Fig. 5 and 6)and the neuroprotective function of astrocytes (GSH production)(Fig. 7).
These findings strongly suggest that the beneficial effect of BB14 isextended well beyond the peripheral context to invest the spinalsynaptic system responsible for pain processing and integration.Notably, i.t. BB14 supply also reduced MMPs activity and proNGFlevels and re-established the balance between mature and precursorforms of NGF (Fig. 3), clearly suggesting a strict correlation betweenglial function, synaptic homeostasis, alteration of MMPs activity andregulation of endogenous NGF levels. This is a novel and attractivefunction for BB14, given that i) mechanisms of reactive gliosis,ii) alteration of NGF metabolism and increase of proNGF, andiii) proNGF/p75-mediated neuron death, all have been described inCNS injury models and suggested to participate to neurodegenerativediseases (Domeniconi et al., 2007; Harrington et al., 2004). Thus it isconceivable that, besides its efficacy in diseases that are specificallycaused by NGF-dependent neurons, BB14 might be a useful thera-peutic tool for a wide range of pathologies that are associated withreactive astrogliosis.
5. Conclusions
In conclusion, our findings confirm the importance of glialactivation in pain processing based on: i) reduction of glial glutamatetransporters and perturbation of synaptic circuitry homeostasis;ii) reduction of glutamate and glycine uptake, thus affecting glialfunction in GSH production and neuroprotection against excitotoxi-city; iii) increased activity of the MMPs degrading system, thusreducing endogenous NGF and its neuroprotective function. On theother hand, our data strongly support the relevance of the peptideBB14 in modulating glial function to an extent similar to that elicitedby the native NGF protein, and its role in: i) maintaining the synaptic
circuitry through modulation of glutamatergic and GABAergiccomponents; ii) restoration of the balance between mature andprecursor forms of NGF; iii) neuroprotection through control of glialfunction in maintaining synaptic glutamate levels and providingneurons with GSH. In conclusion, our data support the relevance ofBB14 peptide as a new useful therapeutic agent for a wide range ofneurodegenerative pathologies characterized by reactive astrogliosis.
Conflict of interest statement
We declare no conflicts of interest.
Acknowledgements
This work was supported by grants from Regione Campania (L.R.N.5 Bando 2003 to M.P.), the Italian Minister of Research andUniversity (PRIN2007 to M.P. and to A.M.C.), Regione Campania(Prog. Spec art 12 E.F. 2000 to M.P.), the CNR (Neurobiotecnologie2003 to M.P.), FIRB-ITALBIONET to L.A., PRIMM srl, Blueprint Biotechand Associazione Levi-Montalcini (fellowships to MRB).
References
Allen NJ, Barres BA. Neuroscience: Glia - more than just brain glue. Nature 2009;457:675–7.
Apfel SC. Nerve growth factor for the treatment of diabetic neuropathy: what wentwrong, what went right, and what does the future hold? Int Rev Neurobiol2002;50:393–413.
Bai YH, Takemitsu M, Atsuta Y, Matsuno T. Peripheral mononeuropathy induced byloose ligation of the sciatic nerve in the rat: behavioral, electrophysiological andhistopathologic studies. Exp Anim 1999;48:87–94.
Bruno MA, Cuello AC. Activity-dependent release of precursor nerve growth factor,conversion to mature nerve growth factor, and its degradation by a proteasecascade. Proc Natl Acad Sci USA 2006;103:6735–40.
Cavaliere C, Cirillo G, Bianco MR, Rossi F, De Novellis V, Maione S, et al. Gliosis altersexpression and uptake of spinal glial amino acid transporters in a mouseneuropathic pain model. Neuron Glia Biol 2007;3:141–53.
Chao MV, Rajagopal R, Lee FS. Neurotrophin signalling in health and disease. Clin Sci2006;2:167–73.
Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactileallodynia in the rat paw. J Neurosci Methods 1994;53:55–63.
Cirillo G, Maggio N, Bianco MR, Vollono C, Sellitti S, Papa M. Discriminative behavioralassessment unveils remarkable reactive astrocytosis and early molecular correlatesin basal ganglia of 3-nitropropionic acid subchronic treated rats. Neurochem Int2010a;56:152–60.
Cirillo G, Cavaliere C, Bianco MR, De Simone A, Colangelo AM, Sellitti S, et al. IntrathecalNGF Administration Reduces Reactive Astrocytosis and Changes NeurotrophinReceptors Expression Pattern in a Rat Model of Neuropathic Pain. Cell MolNeurobiol 2010b;30:51–62.
Cirillo G, Bianco MR, Colangelo AM, Cavaliere C, De Luca D, Zaccaro L, et al. Reactiveastrocytosis-induced perturbation of synaptic homeostasis is restored by NerveGrowth Factor. Neurobiol Disease 2011;41:630–9.
Colangelo AM, Bianco MR, Vitagliano L, Cavaliere C, Cirillo G, De Gioia L, et al. A newnerve growth factor-mimetic peptide active on neuropathic pain in rats. J Neurosci2008;28:2698–709.
Cuello AC, Bruno MA, Bell KF. NGF-cholinergic dependency in brain aging, MCI andAlzheimer's disease. Curr Alzheimer Res 2007;4:351–8.
Decosterd I, Woolf CJ. Spared nerve injury: an animal model of persistent peripheralneuropathic pain. Pain 2000;87:149–58.
Domeniconi M, Hempstead BL, Chao MV. Pro-NGF secreted by astrocytes promotesmotor neuron cell death. Mol Cell Neurosci 2007;34:271–9.
Giaume C, Koulakoff A, Roux L, Holcman D, Rouach N. Astroglial networks: a stepfurther in neuroglial and gliovascular interactions. Nat Rev Neurosci 2010;11:87–99.
Giovannoni R, Maggio N, Rosaria Bianco M, Cavaliere C, Cirillo G, Lavitrano M, et al.Reactive astrocytosis and glial glutamate transporter clustering are early changes ina spinocerebellar ataxia type 1 transgenic mouse model. Neuron Glia Biol 2007;3:335–51.
Hamilton NB, Attwell D. Do astrocytes really exocytose neurotransmitters? Nat RevNeurosci 2010;11:227–38.
Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method formeasuring thermal nociception in cutaneous hyperalgesia. Pain 1988;32:77–88.
Harrington AW, Leiner B, Blechschmitt C, Arevalo JC, Lee R, Mörl K, et al. SecretedproNGF is a pathophysiological death-inducing ligand after adult CNS injury. ProcNatl Acad Sci USA 2004;101:6226–30.
Henneberger C, Papouin T, Oliet SH, Rusakov DA. Long-term potentiation depends onrelease of D-serine from astrocytes. Nature 2010;463:232–6.

232 G. Cirillo et al. / Biotechnology Advances 30 (2012) 223–232
Kullander K, Kaplan D, Ebendal T. Two restricted sites on the surface of the NerveGrowth Factor molecule independently determine specific trkA receptor bindingand activation. J Biol Chem 1997;272:9300–7.
Lobsiger CS, Cleveland DW. Glial cells as intrinsic components of non-cell-autonomousneurodegenerative disease. Nat Neurosci 2007;10:1355–60.
Longo FM, Manthorpe M, Xie YM, Varon S. Synthetic NGF peptide derivatives preventneuronal death via a p75 receptor-dependent mechanism. J Neurosci Res 1997;48:1-17.
Maliartchouk S, Feng Y, Ivanisevic L, Debeir T, Cuello AC, Burgess K, et al. A designedpeptidomimetic agonistic ligand of TrkA nerve growth factor receptors. MolPharmacol 2000;57:385–91.
Massa SM, Yang T, Xie Y, Shi J, Bilgen M, Joyce JN, et al. Small molecule BDNF mimeticsactivate TrkB signaling and prevent neuronal degeneration in rodents. J Clin Invest2010;120:1774–85.
McDonald NQ, Chao MV. Structural determinants of neurotrophin action. J Biol Chem1995;270:19669–72.
Meisner JG, Marsh AD, Marsh DR. Loss of GABAergic interneurons in Laminae I-III of thespinal cord dorsal horn contributes to reduced GABAergic tone and neuropathicpain following spinal cord injury. J Neurotrauma 2010;27:729–37.
Ni Y, Parpura V. Dual regulation of Ca2+−dependent glutamate release fromastrocytes: vesicular glutamate transporters and cytosolic glutamate levels. Glia2009;57:1296–305.
Nykjaer A, Lee R, Teng KK, Jansen P, Madsen P, Nielsen MS, et al. Sortilin is essential forproNGF-induced neuronal cell death. Nature 2004;427:843–8.
Panatier A, Theodosis DT, Mothet JP, Touquet B, Pollegioni L, Poulain DA, et al. Glia derivedD-serine controls NMDA receptor activity and synaptic memory. Cell 2006;125:775–84.
PapaM, Boscia F, CanitanoA, Castaldo P, Sellitti S, Annunziato L, et al. Expression pattern ofthe ether-a-gogo-related (ERG)K+channel-encodinggenes ERG1, ERG2, and ERG3 inthe adult rat central nervous system. J Comp Neurol 2003;466:119–35.
Pekny M, Nilsson M. Astrocyte activation and reactive gliosis. Glia 2005;50:427–34.Peleshok J, Saragovi HU. Functional mimetics of neurotrophins and their receptors.
Biochem Soc Trans 2006;34:612–7.Poduslo JF, Curran GL. Permeability at the blood-brain and blood-nerve barriers of the
neurotrophic factors: NGF, CNTF, NT-3, BDNF. Mol Brain Res 1996;36:280–6.
Polgár E, Todd AJ. Tactile allodynia can occur in the spared nerve injury model in the ratwithout selective loss of GABA or GABA(A) receptors from synapses in laminae I-IIof the ipsilateral spinal dorsal horn. Neuroscience 2008;156:193–202.
Rouach N, Koulakoff A, Abudara V, Willecke K, Giaume C. Astroglial metabolic networkssustain hippocampal synaptic transmission. Science 2008;322:1551–5.
Rydén M, Ibànez CF. A second determinant of binding to the p75 neurotrophin receptorrevealed by Alanine-Scanning mutagenesis of a conserved loop in Nerve GrowthFactor. J Biol Chem 1997;272:33085–91.
Scholz J, Woolf CJ. The neuropathic pain triad: neurons, immune cells and glia. NatNeurosci 2007;10:361–8.
Shih A, Laramee GR, Schmelzer CH, Burton LE, Winslow JW. Mutagenesis identifiesamino-terminal residues of nerve growth factor necessary for Trk receptor bindingand biological activity. J Biol Chem 1994;269:27679–86.
Shih AY, Erb H, Sun X, Toda S, Kalivas PW, Murphy TH. Cystine/glutamate exchangemodulates glutathione supply for neuroprotection from oxidative stress and cellproliferation. J Neurosci 2006;26:10514–23.
Todd AJ. Neuronal circuitry for pain processing in the dorsal horn. Nat Rev Neurosci2010;12:823–36.
Tuszynski MH, Thal L, PayM, Salmon DP UHS, Bakay R, Patel P, Blesch A, Vahlsing HL, HoG, Tong G, Potkin SG, Fallon J, Hansen L, Mufson EJ, Kordower JH, Gall C, Conner J. Aphase 1 clinical trial of nerve growth factor gene therapy for Alzheimer disease. NatMed 2005;11:551–5.
WangW,WangW,Mei X, Huang J,Wei Y,Wang Y, et al. Crosstalk between spinal astrocytesand neurons in nerve injury-induced neuropathic pain. PLoS One 2009;4:e6973.
Wiesmann C, Ultsch MH, Bass SH, de Vos AM. Crystal structure of nerve growth factor incomplex with the ligand-binding domain of the TrkA receptor. Nature 1999;401:184–8.
Yang T, Knowles JK, Lu Q, Zhang H, Arancio O, Moore LA, et al. Small molecule, non-peptide p75 ligands inhibit Abeta-induced neurodegeneration and synapticimpairment. PLoS One 2008;3:e3604.
Yasaka T, Tiong SY, Hughes DI, Riddell JS, Todd AJ. Populations of inhibitory andexcitatory interneurons in lamina II of the adult rat spinal dorsal horn revealed by acombined electrophysiological and anatomical approach. Pain 2010;2:475–88.
Zaccaro MC, Lee HB, Pattarawarapan M, Xia Z, Caron A, L'Heureux PJ, et al. Selectivesmall molecule peptidomimetic ligands of TrkC and TrkA receptors afford discreteor complete neurotrophic activities. Chem Biol 2005;12:1015–28.