Introduction to Carbohydrates UNIT II: Intermediary Metabolism.
Upload
buck-mcbrideCategory
view
258download
0
Basic Concepts of Regulation ofIntermediary Metabolism
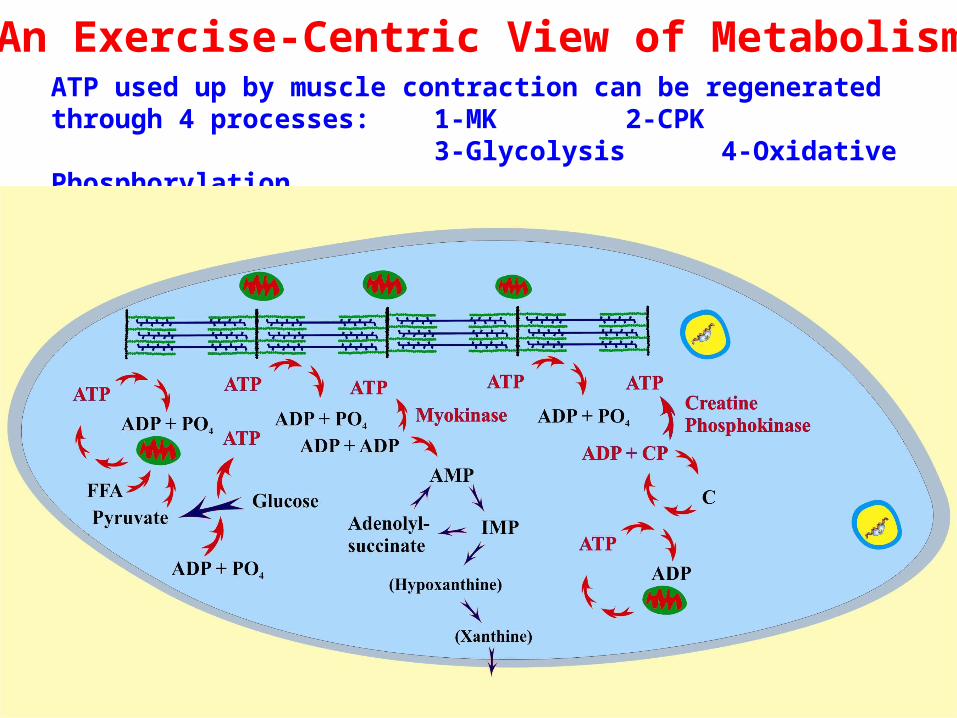
ATP used up by muscle contraction can be regenerated through 4 processes: 1-MK 2-CPK
3-Glycolysis 4-Oxidative Phosphorylation
An Exercise-Centric View of Metabolism

The Myokinase reaction: ADP + ADP → ATP + AMP
is impossible to sustain at maximal rates because AMP cannot beregenerated to ADP and ATP; it can, however, be used to generatefumarate through the purine nucleotide cycle (costs NRG):
AMP + H2O → IMP + NH4+
IMP + Aspartate + GTP → AMP + Fumarate + GDP + Pi
The Creatine Phosphokinase reaction: ADP + CP ↔ ATP + C
is impossible to sustain at maximum activity because of relatively limited supplies of CP in the cell and CP can’t be regenerated very quickly if the vast majority of ATP is being used up for the contractile demands. It is however, more active than MK because it has a lower Km: ~0.02 mM vs. ~0.120mM; [ADP] rest ~ .08, Ex ~0.15) therefore CPK is always active – indicating an important role in shuttling regulating ATP/ADP/Pi while MK is active only at higher [ADP]

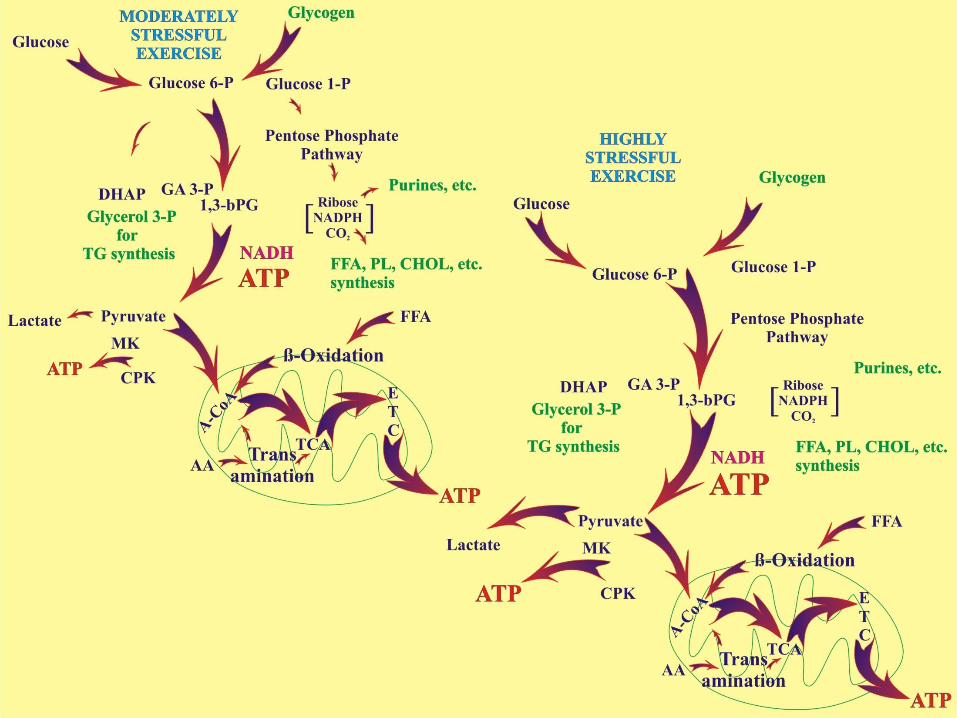
Glycolysis (+ PDH), β-Oxidation, & several transamination reactions lead to the production of acetyl CoA which transfers the acetate group to oxaloacetate in the mitochondria for further breakdown to CO2 and H2O.
Glycolysis is the metabolic pathway through which glucose 6-phosphate is “broken down” to pyruvate (or lactate) in the cytosol.
Note that there are other metabolic pathways for “glucose” metabolism: storage as glycogen, glycerol synthesis for Triglyceride synthesis, pentose-phosphate pathway for synthesis of a variety of other compounds (including ribose and NADPH).
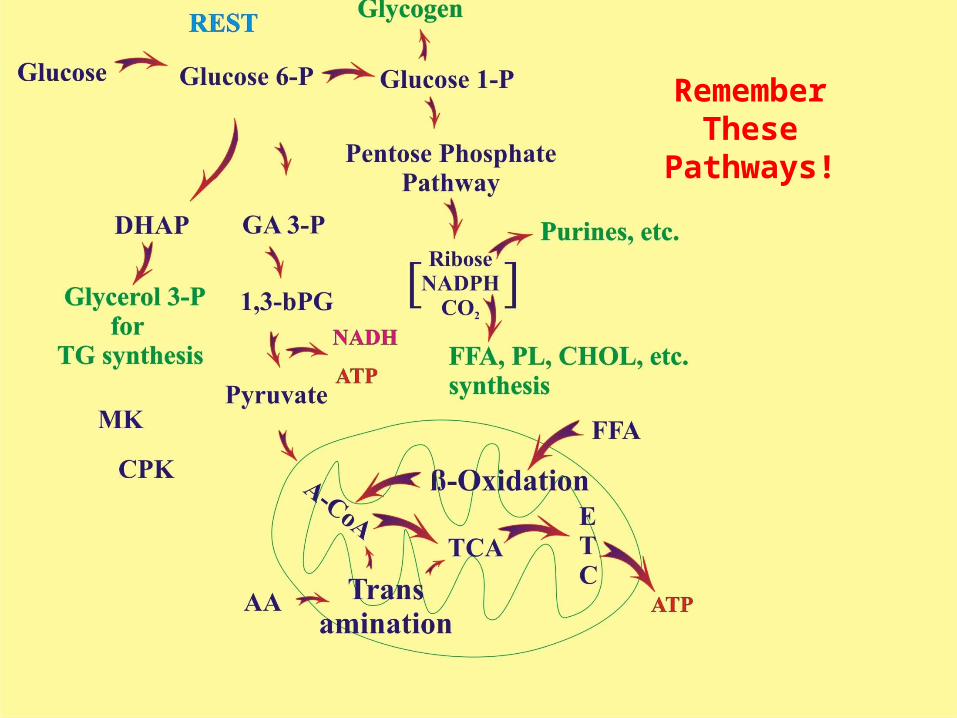
Remember These Pathways!

REGULATION OF GLYCOLYSIS
The rate limiting enzyme of glycolysis: PFK, is the slowest one of the pathway. It is regulated predominantly by ATP, citrate, AMP, H+ and fructose 2,6 Bis-P; with the 2,6 BP being the major regulator in liver and a relatively minor regulator in muscle.
ATP, H+, and citrate greatly inhibit it, promoting storage of glucose as glycogen whenever ATP levels are high. When ATP levels decrease (slightly) as a result of extreme rates of ATP use, such as with maximum muscle contraction, the inhibition of PFK is attenuated and rates of pyruvate and ATP production increase.
When AMP levels increase from increasing MK activity due to increasing production of ADP during exercise rates of glycolysis are greatly speeded up.
Maximally stimulated rates of glycolytic enzyme activity can exceed the maximal capacity of mitochondrial enzymes by about 40x.

Regulation of TCA & ETC
PDH activity allosterically regulated by: NAD+, Co-A, Ca++, and insulin (not really), enhancing activity, and by Acetyl CoA, NADH + H+, and ATP, inhibiting avtivity
Citrate synthase is inhibited by ATP
α-ketoglutarate dehydrogenase activity is inhibited by NADH + H+ and Succinyl CoA and both α-ketoglutarate dehydrogenase and isocitrate dehydrogenase are activated by Ca++
Cytochrome oxidase activity is enhanced by ADP and thyroid hormone (T2), inhibited by ATP, while the ATP synthase and the ETC is activated by Ca++.
-note that just about all of the regulated enzymes in mitochondria can be activated by Ca++ (they are actually more sensitive to calcium than the other allosteric regulators) ensuring increased ATP supply immediately as it is needed while there are multiple inhibitors to prevent unnecessary electron transfer (kinase-decrease/phosphatase-increase).

Recall from the ETC slide that some of the hydrogens that make it to the intermembrane space can leak out when the membrane gets too hot and some leak back into the matrix through the mitochondrial permeability transition which is activated by calcium coming in through the calcium uniporter.
Also note that electron carriers can autooxidize directly to oxygen, creating oxygen radicals (Co-Q is the major site of autooxidation) with as much as 5% of resting oxygen use due to this phenomenon.
All of these processes represent a significant amount of electron transfer to oxygen without concomitant ATP synthesis and all become much more active when exercising hard, creating interesting complications when trying to interpret oxygen consumption and its association with athletic performance.
When accounting for ATP synthesis on the basis of the actual proton cost, you will get ~ 2.5 ATP for each NADH + H+ and ~ 1.5 ATP for each FADH2.
Because of hydrogen leaks & oxygen-radical chemistry, actual yields of ATP from the electron donors are actually less than the proton cost-based ~ 2.5 ATP & ~ 1.5 ATP; a yield that will diminish at high temperature and high calcium conditions (think exercise here).

Recall that H+ and citrate inhibit glycolysis and that it can only be maximally stimulated a lot by AMP (citrate leaks out of the mitochondria when you make lots of it!). This ensures that rates of glycolysis will more or less match rates of oxidative phosphorylation at all rates of ATP demand, at least until rates of glycolysis speed up due to increasing muscle contraction.
We must remember that both mitochondria and LDH enzymes co-exist in cells and therefor they “compete” for the pyruvate that is produced by glycolysis. Because the muscle form of LDH maintains [lactate] >> {pyruvate] some of the pyruvate produced through glycolysis will always be converted to lactate while some is picked up by the mitochondria. Losing lactate from the cell would be awkward because then it wouldn’t be made into pyruvate by LDH so it is transported out of the cell only at relatively high [lactate]. Thus lactate diffuses out of muscle cells at high rates of lactate production (i.e. during exercise) and is an indicator that the LDH is out-competing the mitochondria for pyruvate - a situation that can change when more mitochondria are synthesized.
Oxygen consumption will continue to increase at higher workloads (up to the your maximum ability to remove electrons from “food”) while increasing inefficiencies in oxidative phosphorylation occur at increasing temperature-, ROS-, and calcium-loads. Thus your maximum capacity to produce ATP through oxidative (aerobic) pathways; “aerobic max” (to make ATP), is reached at a workload somewhat similar to that producing the appearance of lactate in the blood (lactate threshold, OBLA). This approximates to that point where increasing inefficiencies in coupling O2 consumption to ATP synthesis (due to increasing work-loads) match the increasing rate of flow of electrons from substrates to O2 through the metabolic pathways to produce no net gain in “aerobically” produced ATP.
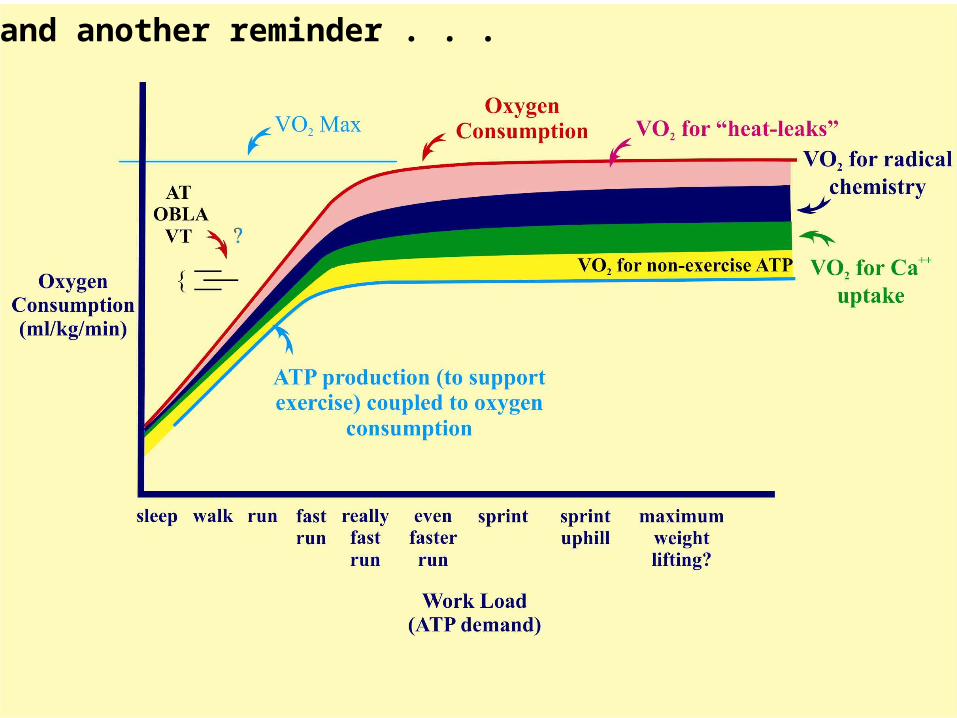
… and another reminder . . .

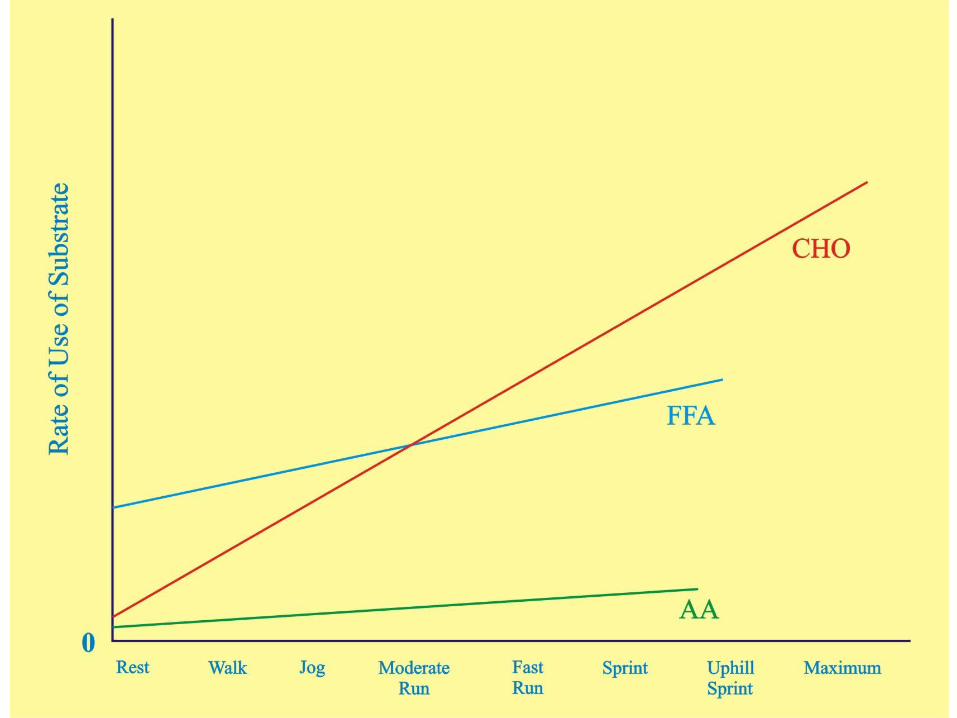
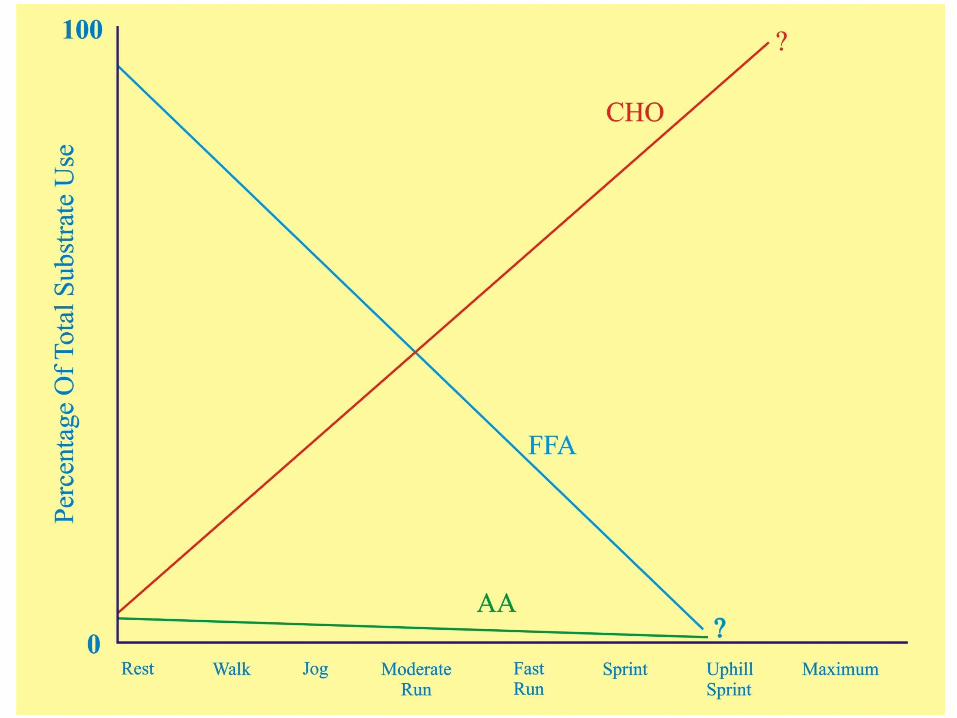
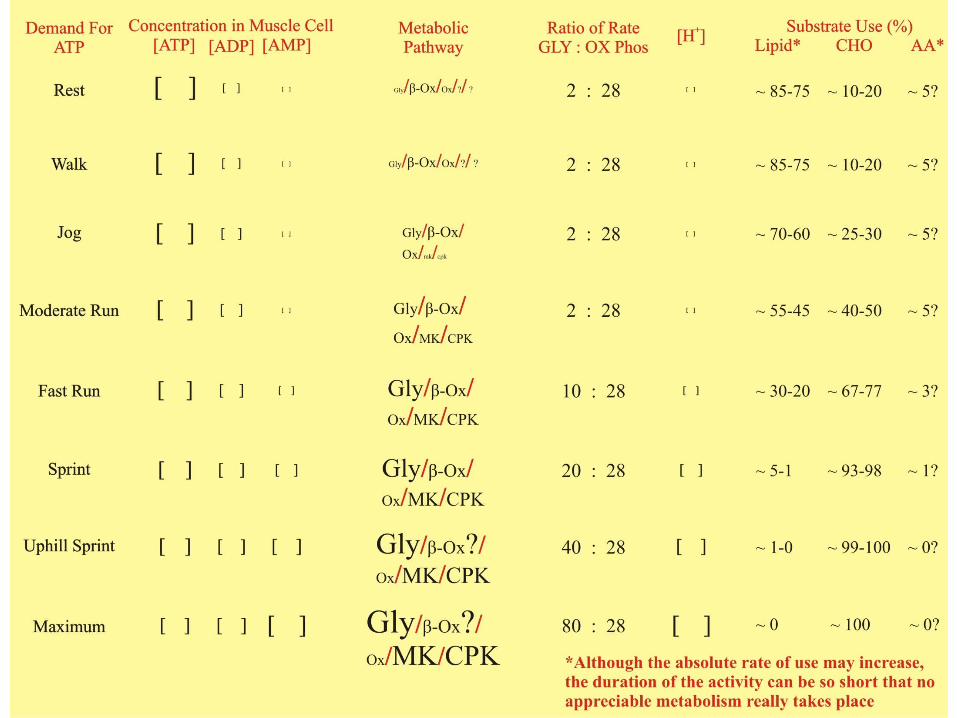
Abbreviated Summary of RegulationWhen you put all this regulation of metabolism stuff together you can see why the relative use of carbohydrates and lipids changes so dramatically as the intensity of exercise changes . . .
To make it easier, you think about metabolic pathways rather then individual enzymes and focus only on a couple of the molecules that regulate the pathways (ATP, ADP, AMP – ok, that was a few):
There are 6 basic “pathways” 4 of which can regenerate ATP and 3 of which supply Acetyl CoA to the TCA cycle of the Oxidative Phosphorylation (Ox) pathway and/or electrons for the Electron Transport Chain of the Ox pathway:
Pathway Abbr. Products RegulatorsGlycolytic Pathway Gly (ATP + Acetyl CoA + e-) ATP↓ AMP↑ Ca++ ↑Oxidative Phosphorylation Ox (ATP/H2O) ATP ↓ ADP ↑ Ca++ ↑
TCA (e-, ie.: NADH/FADH)ETC (ATP/H2O)
Myokinase MK (ATP) ADP ↑Creatinephosphokinase CPK (ATP) ADP ↑β-oxidation β-Ox (Acetyl CoA + e-) “ADP” ↑ Ca++ ↑Transamination Tam (Acetyl CoA) “ADP” ↑ Ca++ ↑
A table of substrate use based on “Regulators” follows:
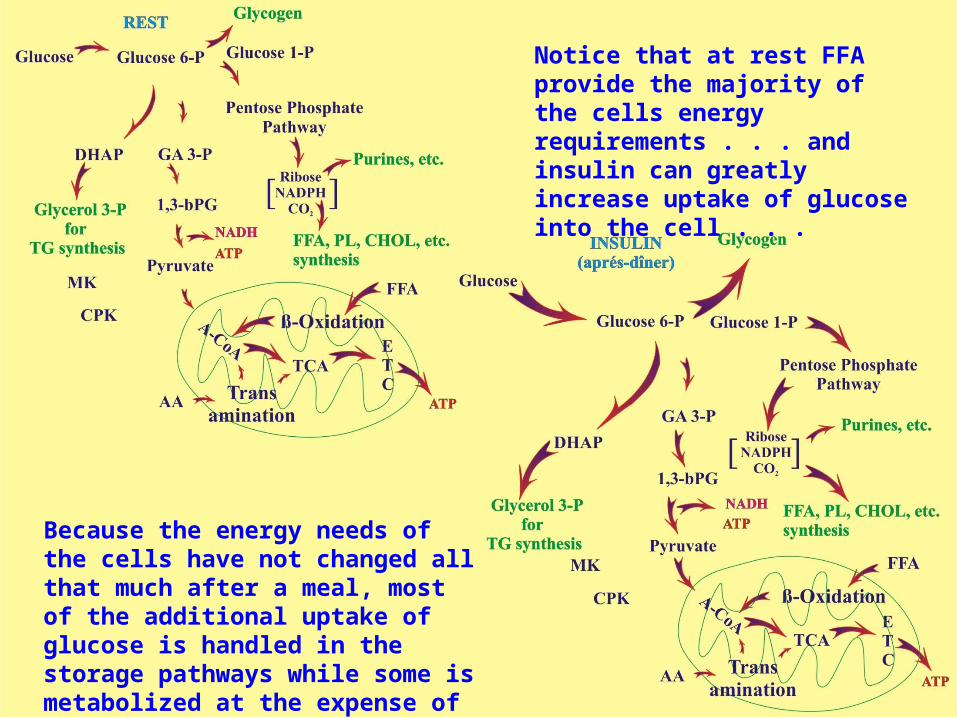
Because the energy needs of the cells have not changed all that much after a meal, most of the additional uptake of glucose is handled in the storage pathways while some is metabolized at the expense of FFA
Notice that at rest FFA provide the majority of the cells energy requirements . . . and insulin can greatly increase uptake of glucose into the cell . . .
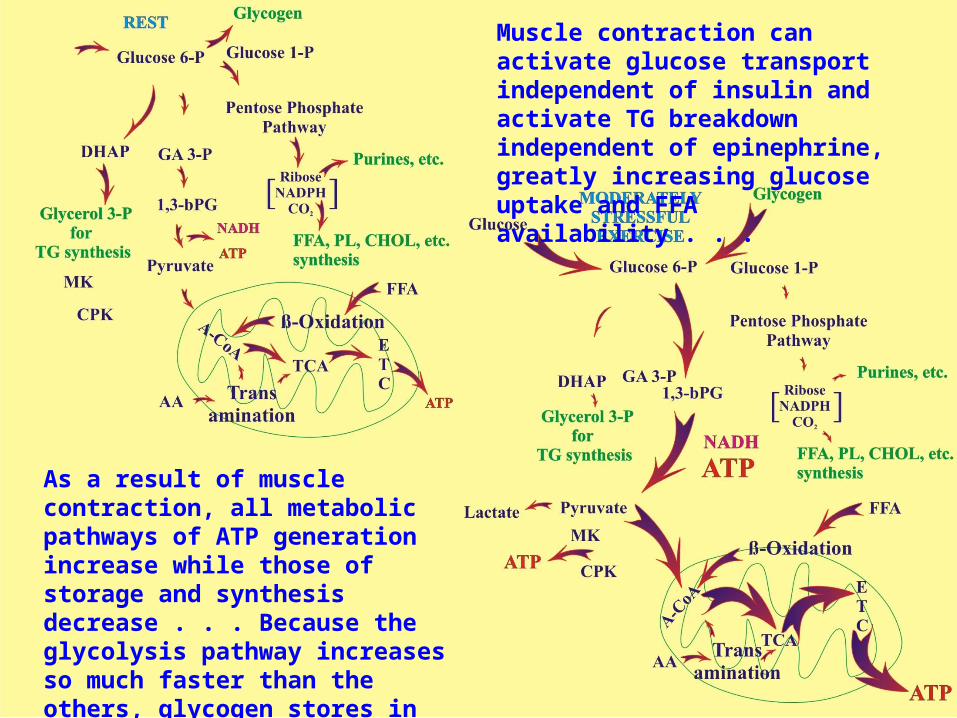
Muscle contraction can activate glucose transport independent of insulin and activate TG breakdown independent of epinephrine, greatly increasing glucose uptake and FFA availability . . .
As a result of muscle contraction, all metabolic pathways of ATP generation increase while those of storage and synthesis decrease . . . Because the glycolysis pathway increases so much faster than the others, glycogen stores in the muscle will decrease very quickly compared to TG stores . . .