
AVALIAÇÃO DO EFEITO DO LASER EM λ= 830nm NA EXPRESSÃO ... · EXPRESSÃO GÊNICA DE CÉLULAS...
66
AUTARQUIA ASSOCIADA À UNIVERSIDADE DE SÃO PAULO AVALIAÇÃO DO EFEITO DO LASER EM λ = 830nm NA EXPRESSÃO GÊNICA DE CÉLULAS MESENQUIMAIS INDIFERENCIADAS DE POLPA DENTAL EM CULTURA VISANDO TERAPIA CELULAR VINICIUS MARCHIORI SILVA Dissertação apresentada como parte dos requisitos para obtenção do Grau de Mestre Profissional em Lasers em Odontologia. Orientadora: Profª. Drª. Martha Simões Ribeiro Co-orientadora: Profª. Drª. Luciane Hiramatsu Azevedo SÃO PAULO 2009
Transcript of AVALIAÇÃO DO EFEITO DO LASER EM λ= 830nm NA EXPRESSÃO ... · EXPRESSÃO GÊNICA DE CÉLULAS...

AUTARQUIA ASSOCIADA À UNIVERSIDADE DE SÃO PAULO
AVALIAÇÃO DO EFEITO DO LASER EM λ= 830nm NA
EXPRESSÃO GÊNICA DE CÉLULAS MESENQUIMAIS INDIFERENCIADAS DE POLPA DENTAL EM CULTURA
VISANDO TERAPIA CELULAR
VINICIUS MARCHIORI SILVA
Dissertação apresentada como parte dos requisitos para obtenção do Grau de Mestre Profissional em Lasers em Odontologia.
Orientadora: Profª. Drª. Martha Simões Ribeiro Co-orientadora: Profª. Drª. Luciane Hiramatsu Azevedo
SÃO PAULO
2009
2
Mestrado Profissionalizante Lasers em
Odontologia

3
AUTARQUIA ASSOCIADA À UNIVERSIDADE DE SÃO PAULO
AVALIAÇÃO DO EFEITO DO LASER EM λ= 830nm NA
EXPRESSÃO GÊNICA DE CÉLULAS MESENQUIMAIS INDIFERENCIADAS DE POLPA DENTAL EM CULTURA
VISANDO TERAPIA CELULAR
VINICIUS MARCHIORI SILVA
Dissertação apresentada como parte dos requisitos para obtenção do Grau de Mestre Profissional em Lasers em Odontologia.
Orientadora: Profª. Drª. Martha Simões Ribeiro Co-orientadora: Profª. Drª. Luciane Hiramatsu Azevedo
SÃO PAULO
2009

4
Dedico este trabalho à futura geração em nome do meu filho Gabriel
Raggio Marchiori, que todo esforço seja feito para melhorar seu
caminho, espero que este trabalho sirva de inspiração pra você e pra
alguns que vierem depois de mim.

5
AGRADECIMENTOS
À minha orientadora Professora Doutora Martha Simões
Ribeiro pela paciência, pelos conhecimentos compartilhados,
pelas ótimas idéias e perfeita sintonia de trabalho, espero
poder continuar trabalhando com você.
À minha co-orientadora, Professora Doutora Luciana
Hiramatsu Azevedo pela dedicação e pelas respostas rápidas
sempre que foi solicitada, além do empenho clínico excepcional.
A todos os professores do curso de mestrado Lasers em
Odontologia, com destaque especial aos professores Nilson Dias
Vieira Junior e Carlos de Paula Eduardo pela força e coragem de
manter um curso tão especial envolvendo três instituições de
renome: FOUSP, LELO e IPEN.
Aos funcionários ligados ao curso, especialmente a Andrea
Malavazzi do fundo do meu coração pelo empenho em fazer
nossa vida acadêmica funcionar nestes anos.
Aos colegas que compartilharam a sala de aula deste
curso comigo e ao amigo Rodrigo que compartilhou a clínica, os
textos, as fotos e bons momentos.
Ao Professor Doutor Sang Won Han, do CINTERGEN da
UNIFESP pelas orientações, pelos equipamentos gentilmente
cedidos e pelos conhecimentos compartilhados nos últimos
anos.
A Minha grande amiga, pois é assim que chamamos
alguém que está sempre presente de forma importante em
nossa vida, Bianca Lisboa, que muito colaborou para que este
trabalho e outros pudessem acontecer.
A minha grande amiga e professora Flávia Helena da
Silva, companheira de laboratório e de muitas conversas
ótimas, sem você eu estaria na estaca zero, tenho certeza que

6
nosso futuro reserva ótimas oportunidades para trabalharmos
juntos.
Aos amigos Dr. Henrique Celebrone, Dra. Angélica Luppi e
sua equipe por dar suporte fundamental durante todo o período
da pesquisa.
Ao professor Doutor Niels Olsen Câmara, que acreditou em
meu trabalho e ajudou cedendo tempo, recursos e muito
conhecimento valioso.
A minha amiga Patricia Semedo, que em meio a toda
dificuldade para cuidar da sua própria pesquisa ainda arrumou
tempo pra cuidar da minha e me ensinar caminhos para ser feliz
no mundo científico.
Agradeço aos amigos Raphael Pacheco e professora
Doutora Rejane Daniele Reginato da morfologia da UNIFESP
pela grande ajuda no ano passado e nos trabalhos atuais,
espero trabalhar muito mais com vocês.
Aos amigos Gil, Eduardo, Priscila e Pri, Rô, Leonardo, Xuxu
por me aguentarem perguntando e pedindo coisas dentro do
laboratório, ajudarem com cálculos, cederem tempo, material e
até por fazerem procedimentos para mim sem perder o bom
humor.
A minha paciente Ligia Shizuko Sumi Vieira do ICB da USP
por tão gentilmente me arrumar dispase no início do meu
trabalho, sem você eu teria perdido muito tempo.
A minha sócia, amiga e mãe do meu filho, Andrea Raggio
por garantir o funcionamento do consultório enquanto eu estive
fora.
A minha cunhada Professora Doutora Daniela Raggio que
me deu muitas orientações como ótima acadêmica que é.
Meus pais, Elena Marchiori e Antonio Carlos Bueno que me
deram suporte de todas as formas que existem e mais algumas
que eles inventaram.

7
Minha irmã Bianca e meu cunhado Eduardo Claro que me
receberam em sua casa e em seus corações para que eu
pudesse levar meu trabalho para a Europa.
Ao meu irmão Emerson que sempre me deu socorro
linguístico quando precisei me virar em outros idiomas.
Ao amigo Fernando Henrique Costa e Silva que moveu um
mundo pra me ajudar quando eu não conseguia me levantar
sozinho, me trouxe luz e me levou às águas.
Ao amigo Professor Doutor Marcelo Abla, graças a você
isso tudo vai ter um sentido, nossa caminhada está apenas
começando apesar de já ter 20 anos de parcerias diversas e
amizade intensa.
A amiga Rana Rached: veterinária, revisora, chefe de
cozinha, enóloga e companheira de dança, além de mulher do
meu amigo Marcelo Abla, amo vocês.
A mio amico Carmine Gentile, sono grato per l’appoggio in
2008 a Ginevra, Svizzera i per iChat!
A minhas amigas que acreditaram que era possível fazer
uma especialização e um mestrado ao mesmo tempo,
pesquisar envolvendo as duas áreas e ainda me deram suporte
emocional durante o percurso: Renata Chinaite e Cristiane
Henrique Soares excelentes fonoaudiólogas.

8
EFEITOS DO LASER EM λ= 830nm NA EXPRESSÃO GÊNICA DE
CÉLULAS MESENQUIMAIS INDIFERENCIADAS DE POLPA
DENTAL EM CULTURA VISANDO TERAPIA CELULAR
VINICIUS MARCHIORI SILVA
Resumo
Este estudo visa investigar a possível relação entre a irradiação
laser λ= 830 nm e a expressão gênica dos genes ligados à
diferenciação de células mesenquimais indiferenciadas em células de
linhagem osteoblástica. Dentes extraídos de pacientes por motivos
clínicos foram levados ao ambiente estéril em laboratório BL-2 e
abertos para acesso às câmaras pulpares. O tecido pulpar foi então
digerido para que a separação das células mesenquimais ocorresse.
Amostras iniciais coletadas das células recém obtidas foram analisadas
por citometria de fluxo e foi comprovada a presença de marcadores de
superfície correspondentes aos das células indiferenciadas,
caracterizando-se desta forma a população estudada como sendo de
células mesenquimais indiferenciadas, ou células tronco de adultos.
Após obtenção por digestão fracionada, foi feito o plaqueamento e
essas células foram mantidas em culturas, que foram expandidas e
divididas em dois grupos: GL, grupo que recebeu irradiação laser e
GC, que não recebeu irradiação laser e foi utilizado como controle.
Diferenças nas expressões dos genes ligados à diferenciação
celular foram detectadas por análise molecular tipo RT-PCR e PCR e
nossos resultados indicam que a diferenciação celular foi mais rápida
nas células que receberam irradiação laser.

9
EFFECTS OF LASER AT λ= 830nm ON THE GENIC EXPRESSION
OF MESENCHIMAL STEAM CELLS FROM DENTAL PULP IN
CULTURE AIMING CELLULAR THERAPY
VINICIUS MARCHIORI SILVA
Abstract
This study aims to investigate the possible relation between 830
nm laser radiation and the genic expression of the genes linked to
differentiation of the mesenchymal steam cells (MSC) to osteoblastic
lineage. Tooth extracted from patients for clinical purposes were taken
to the sterile ambience at biosecurity level 2 and opened to access to
dental pulp chamber. The pulp tissue was them digested to separate
the MSC. Samples collected from the cell population freshly extracted
were analyzed by flow citometry and the presence of markers
corresponding to MSC was confirmed, characterizing in this way the
population studied as MSC. After obtaining the cells by digestion, those
were put into culture, expanded and divided into two groups: GL,
group that received laser radiation and the other called GC, that didn’t
received laser radiation and it was used as a control. Differences
between the expression of the genes linked to cellular differentiation
were detected by molecular analysis RT-PCR and PCR. Our results
indicate that differentiation occurs more rapidly in the cells that
received laser radiation.

10
SUMÁRIO
1. INTRODUÇÃO
2. REVISÃO DE LITERATURA
2.1 CÉLULAS E LASER: APLICABILIDADE NA MEDICINA
REGENERATIVA
2.2 - BIOLOGIA MOLECULAR DA CÉLULA
2.2.1 DNA, CÓDIGO DA VIDA.
2.2.2 TODAS AS CÉLULAS TRANSCREVEM PORÇÕES DE
SUAS INFORMAÇÕES HEREDITÁRIAS EM UMA MESMA
FORMA INTERMEDIÁRIA, O RNA.
2.2.3 TRADUÇÃO DOS RNAS EM PROTEÍNAS
2.3 USO DE CÉLULAS EM REGENERAÇÃO TECIDUAL.
2.4 TECIDO ÓSSEO E REGENERAÇÃO
2.5 SINALIZAÇÃO CELULAR ESPECÍFICA: REGULAÇÃO DA
EXPRESSÃO GÊNICA.
3.OBJETIVOS.
11
13
13
18
18
23
27
35
37
38 42
4.MATERIAL E MÉTODOS 43
5. RESULTADOS E DISCUSSÃO. 50
8.CONCLUSÕES
9.REFERÊNCIAS BIBLIOGRÁFICAS
61
62

11
1. Introdução
A utilização de células tronco de adultos em regeneração tecidual
e na construção de órgãos artificiais vem ganhando espaço e está
presente atualmente como uma realidade possível.
Construção de órgãos artificiais, como por exemplo válvulas
cardíacas recobertas de células, membranas com cobertura celular
para reconstrução óssea, pele artificial, ou mesmo um spray rico em
células para reparo de descontinuidades de derme causados por
queimaduras ou feridas diversas, já são realidade e envolvem o uso de
células coletadas, na maioria das vezes, do próprio paciente. Os
tecidos coletados são levados para o laboratório, as células de
interesse são isoladas e colocadas em cultura. Normalmente, são
células mesenquimais indiferenciadas, que após sofrerem expansão, ou
seja, aumento na quantidade de células da cultura, são diferenciadas
por estímulos dados pelos meios de cultura modificados de forma que
estas células são levadas a cumprir uma função específica como
fibroblastos, osteoblastos, cardiomiócitos, entre outros.
As fases de obtenção, isolamento e expansão celulares seguem
tempos ditados pelo metabolismo celular e por algumas inovações
tecnológicas, como os meios de cultura aperfeiçoados, ou culturas em
perfusão ou microgravidade. É bem conhecida a capacidade dos lasers
em acelerar o reparo de tecidos in vivo e aumentar a proliferação
celular em cultura, mas essa inovação tecnológica ainda não entrou
nos protocolos de expansão de culturas celulares e esta introdução
poderá ocorrer caso comprove-se eficiente para esta utilização e caso
os parâmetros de irradiação sejam definidos para os diversos tipos
celulares. É importante também o conhecimento de como o laser pode
atuar na expressão dos genes envolvidos na diferenciação celular. Se
alguma modificação na expressão ocorre, é possível que as culturas

12
celulares se tornem diferenciadas mais rapidamente para o tipo celular
desejado, ou então que se mantenham indiferenciadas, o que permite
expansão maior, uma vez que células indiferenciadas tem maior
capacidade de diferenciação do que células diferenciadas. Outra
possibilidade é a expansão das culturas diferenciadas, mas somente
uma análise dos genes envolvidos poderá dizer em que fase de
diferenciação as células se encontram e se algo muda com o uso do
laser.
Este trabalho, portanto, tem como objetivo investigar os efeitos
doa radiação laser emitindo em λ= 830 nm na expressão gênica dos
genes ligados à diferenciação de células mesenquimais indiferenciadas
em células de linhagem osteoblástica.

13
2. Revisão de literatura
2.1 Células e laser: aplicabilidade na medicina
regenerativa
Há muitos anos, a ciência vem tentando reabilitar pacientes com
perdas ósseas de etiologias diversas e muitos progressos ocorreram
desde as primeiras tentativas de regeneração feitas na década de
1960, com materiais que nada mais faziam do que separar defeitos
ósseos e suas células reparadoras da influência de outros tipos
celulares (Boyne, 1964).
Sem dúvida os maiores avanços vieram com o avanço
tecnológico, e técnicas de reabilitação óssea surgiram nas décadas
posteriores (Buser et al., 1994; Lynch et al., 1999) bem como o uso do
laser como adjuvante no reparo ósseo (Freitas et al., 2000)
A interação de estímulos físicos e seu efeito sobre as células
foram alvo de estudos e atualmente têm ganhado maior atenção
devido à possibilidade de uso destas células em terapias baseadas em
engenharia de tecidos (Tsai et al., 2007; Burger & Klein-Nulend,
1998; Mai et al. 2007; zheng et al., 2007) e o efeito do laser de baixa
potência vem sendo citado como efetivo na aceleração cicatricial e
reparação de tecidos duros do corpo humano bem como no controle da
sintomatologia dolorosa crônica e reparo de tecidos moles (Hawkins et
al., 2006). A fototerapia laser pode, além de estimular o crescimento
celular em tecidos ósseos, promover regeneração celular e
remodelação vascular além de promover maior atividade enzimática,
aumentar a formação de produtos pelas células e alterar potencial de
ação em células nervosas (Ribeiro e Zezell, 2004).
Em odontologia, o laser tem várias aplicações e sua efetividade
em acelerar o reparo do tecido duro auxilia tratamentos relacionados
ao osso em diversas especialidades, tais como: ortodontia, cirurgia,

14
implantodontia (Blay, 2001) e outras onde o potencial bioestimulatório
possa ser usado (Saito & Shimizu, 1997; Kawasaki & Shimizu, 2000).
Estudos in vivo mostram maior rapidez na recuperação de fraturas
ósseas (Trelles & Mayayo, 1987) e aumento da reparação óssea em
tratamentos ortopédicos (Saito & Shimizu, 1997).
O efeito do laser de baixa potência também foi analisado in vitro
(Ozawa et al., 1998) e foi observado aumento da proliferação celular e
da atividade de fosfatase alcalina bem como formação de nódulos
ósseos nas placas de cultura. Outro estudo (Fujihara, 2005)
comprovou aumento na proliferação e adesão celular em osteoblastos
em cultura estimulados por laser
A proliferação celular de células mesenquimais indiferenciadas,
conhecidas como células tronco adultas, também aumenta com a
irradiação das culturas com laser em baixa intensidade conforme
descreve Tuby e colaboradores (2007), estes autores demonstraram
este fato e creditam à irradiação laser a indução de síntese de
proteínas regulatórias celulares devido à ativação de genes do ciclo
celular responsáveis pelo início da mitose como o MAPK (mitogen
activated protein kinase) e ERK (extracellular signal regulated kinase).
Estas reações são dependentes de transporte de energia e reações de
oxi-redução que ocorrem nas células (Voet et al. 2002).
Segundo Alberts e colaboradores (2004), reações de oxidação e
redução são constantes nas células e fazem parte do processo de
transformação de energia em função dentro destas. O NAD
(nicotinamida adenina dinucleotídeo) e FAD (flavina adenina
dinucleotídeo) são moléculas carreadoras de elétrons de grande
importância na célula e participam de processos ligados à energia e
seu uso nas mesmas, da mesma forma é o ATP (adenosina tri-fosfato).
Estas moléculas estão envolvidas diretamente no funcionamento do
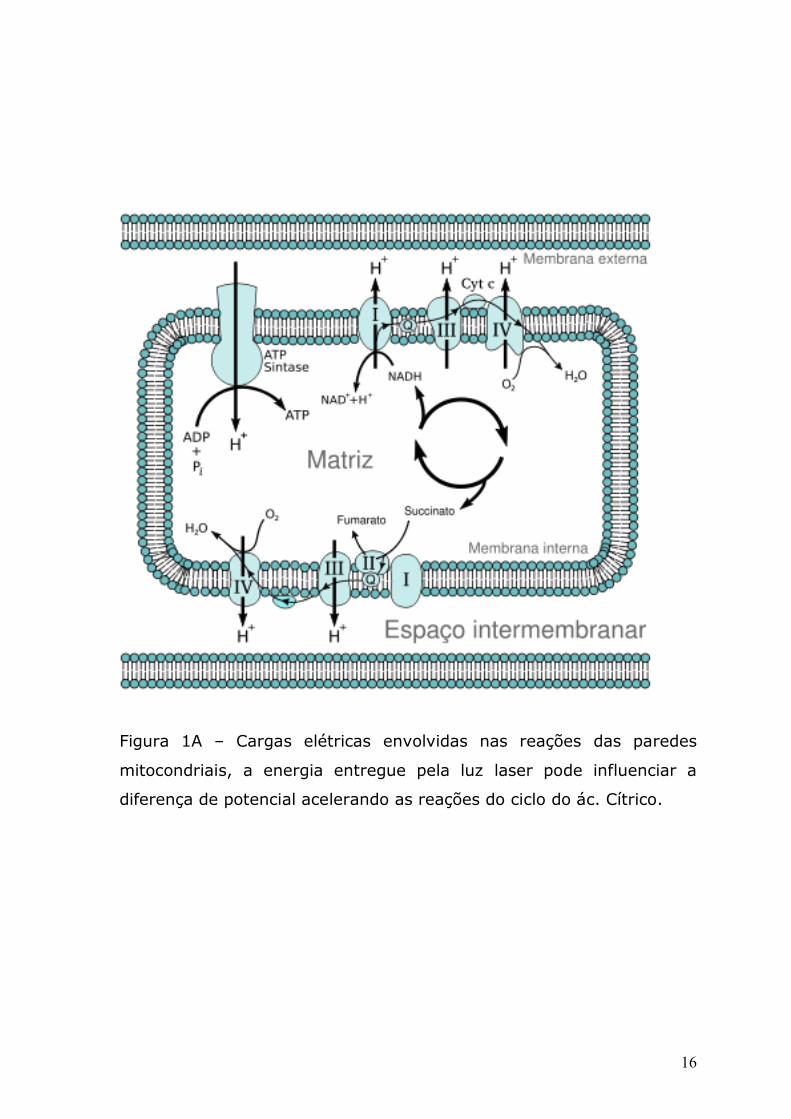
ciclo do ácido cítrico ou ciclo de Krebs que ocorre nas mitocôndrias
(figura 1 A). As reações bioquímicas encontradas neste sistema
acontecem devido ao potencial redox das moléculas envolvidas e

15
quanto maior o potencial padrão de redução, maior a afinidade por
elétrons, ou seja, existe a tendência da forma oxidada de um par redox
aceitar elétrons e tornar-se reduzida. Baseado nesta constatação
bioquímica, algumas explicações surgiram para que o laser tenha efeito
sobre tal sistema. A dualidade onda-partícula da luz leva alguns
autores a explicar sua influência sobre as reações de oxi-redução
ocorridas nas mitocôndrias pela transferência de energia para esta
organela.
Segundo Wilden e Karthein (2000), o modelo clássico de
transferência de energia mitocondrial é dado pela bioquímica do
transporte de elétrons nesta organela e a função do elétron no sistema
é operar a fosforilação oxidativa neste local produzindo ATP. A
fosforilação oxidativa somente pode ocorrer na conexão entre um
doador de elétron e um aceitator do mesmo e os mais importantes
doadores são o NADH e o FADH2. O NADH doa seu elétron a proteínas
de membrana ligadas à cadeia respiratória a partir de colisões
randômicas na membrana mitocondrial interna e o FADH por estar na
parede mitocondrial entre as membranas externa e interna da mesma
doa seu elétron diretamente à cadeia respiratória. Para que as reações
envolvendo estes parceiros ocorram com maior eficiência ou
velocidade, a energia deve estimular pelo menos um deles de forma a
mudar a distribuição de cargas por pelo menos um instante, ou seja, a
presença de radiação em freqüência específica, comprimento de onda
adequado e intensidade ideal pode ser fator decisivo para que as
reações tomem lugar ou não (figura 1 B).
Os estudos in vitro corroboram com os resultados dos estudos in
vivo, entretanto, as células envolvidas nos trabalhos pesquisados não
eram visadas para uso em terapia celular com pacientes, mas apenas
para pesquisa sobre a influência da irradiação laser nas células in vitro,
mas será que a irradiação laser poderia ser útil no caso dessas culturas
serem usadas em terapia celular?
16
Figura 1A – Cargas elétricas envolvidas nas reações das paredes
mitocondriais, a energia entregue pela luz laser pode influenciar a
diferença de potencial acelerando as reações do ciclo do ác. Cítrico.

17
Figura 1B Esquema de Tiina Karu: a presença de radiação em
freqüência específica, comprimento de onda adequado e intensidade
ideal pode ser fator decisivo para que as reações tomem lugar ou não

18
2.2 - Biologia Molecular da célula
2.2.1 DNA, código da vida.
Todas as células replicam e mantém sua informação hereditária
seguindo um padrão por isso podemos dizer que o mecanismo da vida
está contido na estrutura dupla da molécula de DNA (Alberts e cols.,
2004).
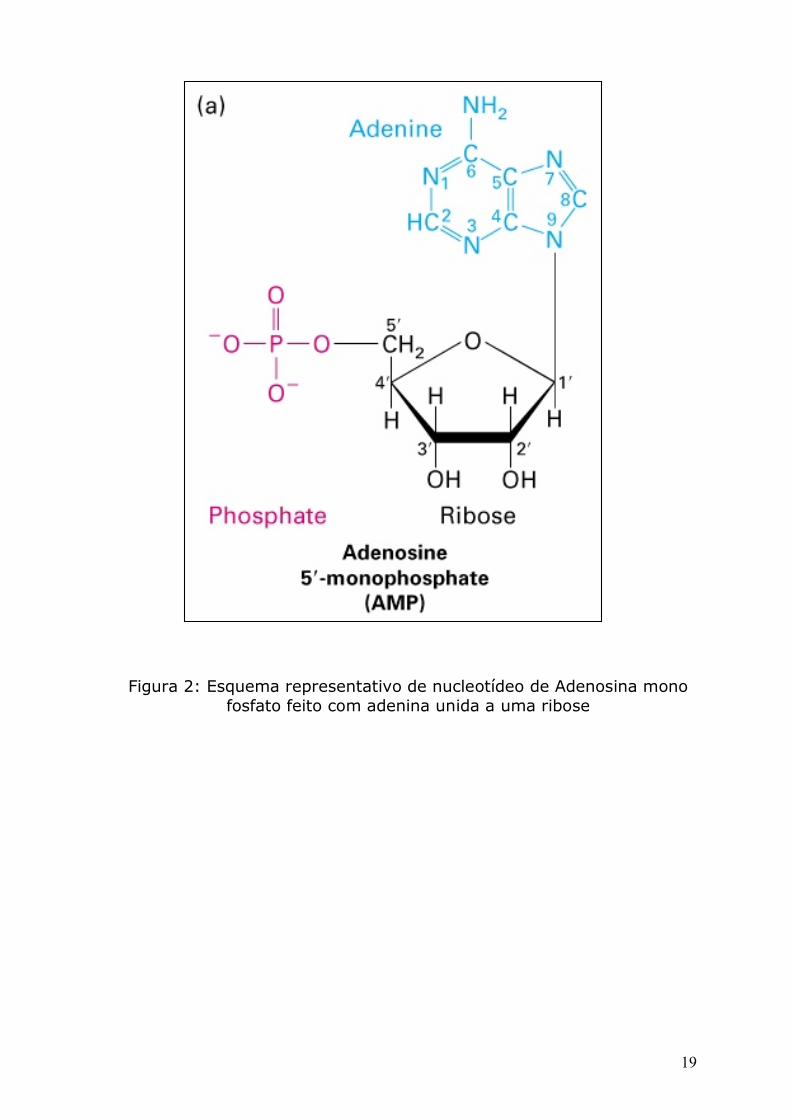
Cada monômero, chamado nucleotídeo, presente em uma cadeia
simples de DNA consiste em duas partes: um açúcar (desoxiribose)
com um grupo fosfato ligado a ele e uma base que pode ser: Adenina
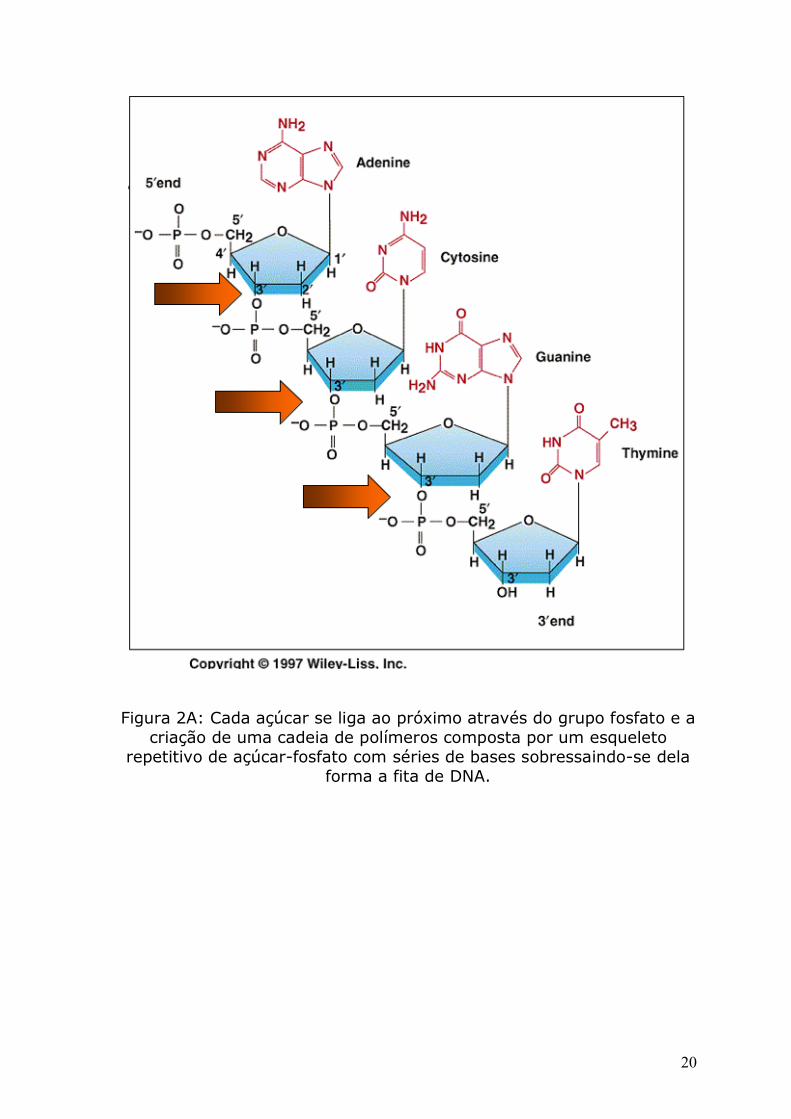
(A), Timina (T), Citosina (C) ou Guanina (G) (figura 2). Cada açúcar
se liga ao próximo através do grupo fosfato e a criação de uma cadeia
de polímeros composta por um esqueleto repetitivo de açúcar-fosfato
com séries de bases sobressaindo-se dela forma a fita de DNA (figura
2A). Nas células esta fita não é sintetizada isoladamente como uma fita
livre mas sim formada por complemento a um molde padrão de fita
DNA pré-existente de forma que há uma regra definida pelas
estruturas das bases nitrogenadas de forma que: “A” liga-se com “T” e
“C” liga-se com “G” (figura 2 B), por este pareamento de bases os
monômeros são adicionados conforme o molde original e a fita dupla é
formada por 2 sequências exatamente complementares de As, Ts, Cs e
Gs, essas duas fitas torcidas entre si formam a estrutura da dupla
hélice (figura 2C). As ligações entre os pares de bases das duas fitas é
feita por pontes de Hidrogênio e é mais fraca que a ligação entre
açúcar-fosfato de forma que é possível a separação dessas fitas sem
causar dano ao esqueleto do DNA (figura 2B e 2C). A partir da
separação das fitas a sequência pode ser copiada em pontos
específicos que são os genes, conforme explicado a seguir.
19
Figura 2: Esquema representativo de nucleotídeo de Adenosina mono fosfato feito com adenina unida a uma ribose
20
Figura 2A: Cada açúcar se liga ao próximo através do grupo fosfato e a criação de uma cadeia de polímeros composta por um esqueleto
repetitivo de açúcar-fosfato com séries de bases sobressaindo-se dela forma a fita de DNA.
21
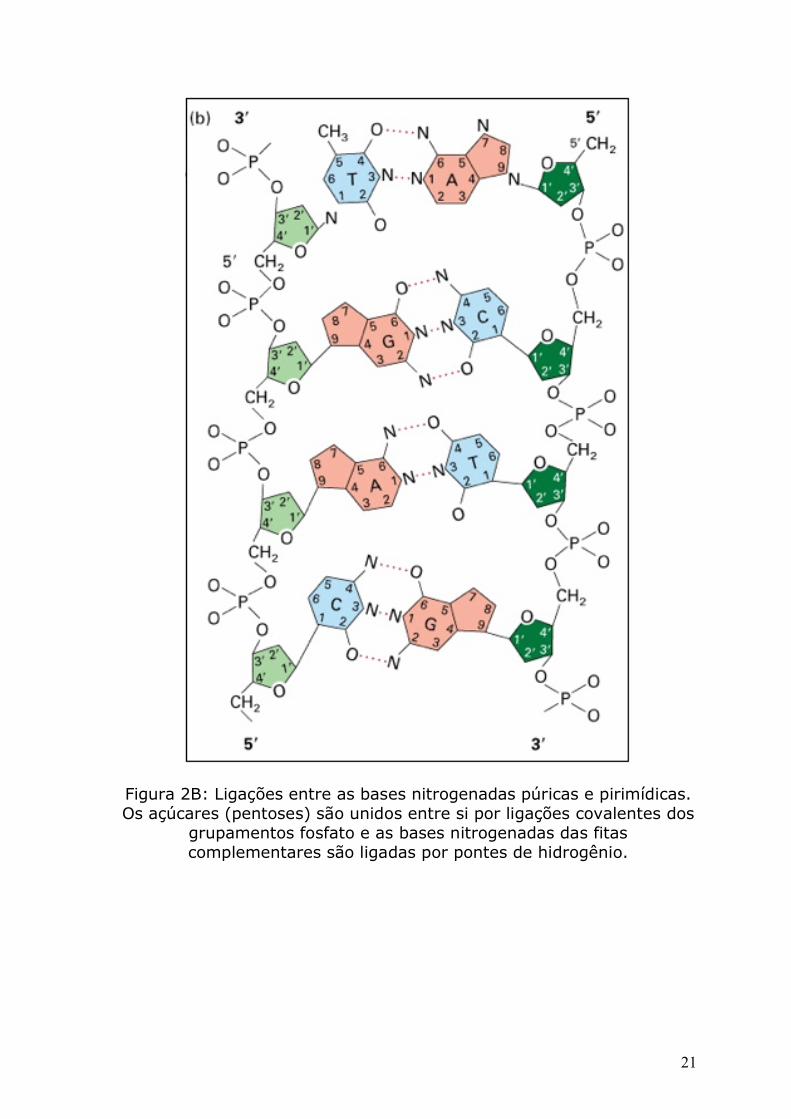
Figura 2B: Ligações entre as bases nitrogenadas púricas e pirimídicas. Os açúcares (pentoses) são unidos entre si por ligações covalentes dos
grupamentos fosfato e as bases nitrogenadas das fitas complementares são ligadas por pontes de hidrogênio.

22
Figura 2c: Dupla fita de DNA. As ligações entre as bases nitrogenadas dentro da mesma fita são feitas por ligação tipo covalente entre os grupamentos fosfato, esta ligação é mais forte que as ligações feitas
por pontes de hidrogênio e que mantém as fitas unidas (em vermelho), por isso é possível a separação das duas fitas para leitura do DNA sem que cada fita sofra rompimento na estrutura principal, isto garante a
integridade durante a transcrição.

23
2.2.2 Todas as células transcrevem porções de suas
informações hereditárias em uma mesma forma intermediária,
o RNA.
O DNA não faz somente cópias de si mesmo mas é capaz de
expressar as informações que contém e guiar a síntese de outras
moléculas na célula, isso ocorre pela formação de outras classes de
polímeros: RNAs e proteínas.
O processo sempre se inicia pela polimerização (a partir de
moldes do DNA) de uma molécula de RNA chamada de transcrição ou
transcrito primário, na qual sequências de DNA (genes) são usados
como moldes para guiar a síntese de moléculas menores de polímeros
de ácido ribonucléico, os RNA mensageiros (mRNA). Depois, em um
processo mais complexo, chamado tradução, muitas dessas moléculas
de RNA direcionam a síntese de polímeros pertencentes a classes
químicas radicalmente diferentes, as proteínas.
No RNA o esqueleto molecular é formado por um açúcar
ligeiramente diferente daquele do DNA com ribose em vez de
desoxirribose (figura 3) e uma das quatro bases também é
ligeiramente diferente: Uracila (U) no lugar de Timina (T).
Durante a transcrição os monômeros de RNA são alinhados e
selecionados de acordo com a fita molde de DNA pela enzima RNA
polimerase e assim a fita simples formada representa uma parte da
informação genética da célula mas em um “alfabeto” ligeiramente
diferente. Um mesmo fragmento de DNA (gene) pode ser lido repetidas
vezes e produzir uma série de RNAs, que são os intermediários entre a
informação genética contida no núcleo e a proteína final e são
chamados RNAs mensageiros, desta forma enquanto o arquivo de
informação genética da célula na forma de DNA é fixo, os RNAs
transcritos são produzidos em massa e descartados (figura 4).
As moléculas de RNA possuem estruturas distintas que também
podem conferir-lhes outras características químicas especializadas.

24
Sendo fita simples, seu esqueleto é flexível, podendo dobrar-se para
permitir que uma parte da molécula forme ligações fracas com outra
parte desta mesma molécula. Isso acontece quando os fragmentos da
mesma sequência são localmente complementares: um segmento
...GGGG... por exemplo tenderá a se associar a outro ...CCCC... Esses
tipos de associações iternas fazem com que uma cadeia de RNA se
dobre em uma forma específica, dirigida por sua própria sequência
(figura 5). A forma da molécula de RNA por sua vez pode habilitá-la a
reconhecer outras moléculas, ligando-se a elas seletivamente e até
catalizar mudanças químicas nas moléculas às quais se ligam.
Figura 3: No RNA o esqueleto molecular é formado por um açúcar ligeiramente diferente daquele do DNA com ribose em vez de desoxirribose e uma das quatro bases também é ligeiramente
diferente: Uracila (U) no lugar de Timina (T).
25
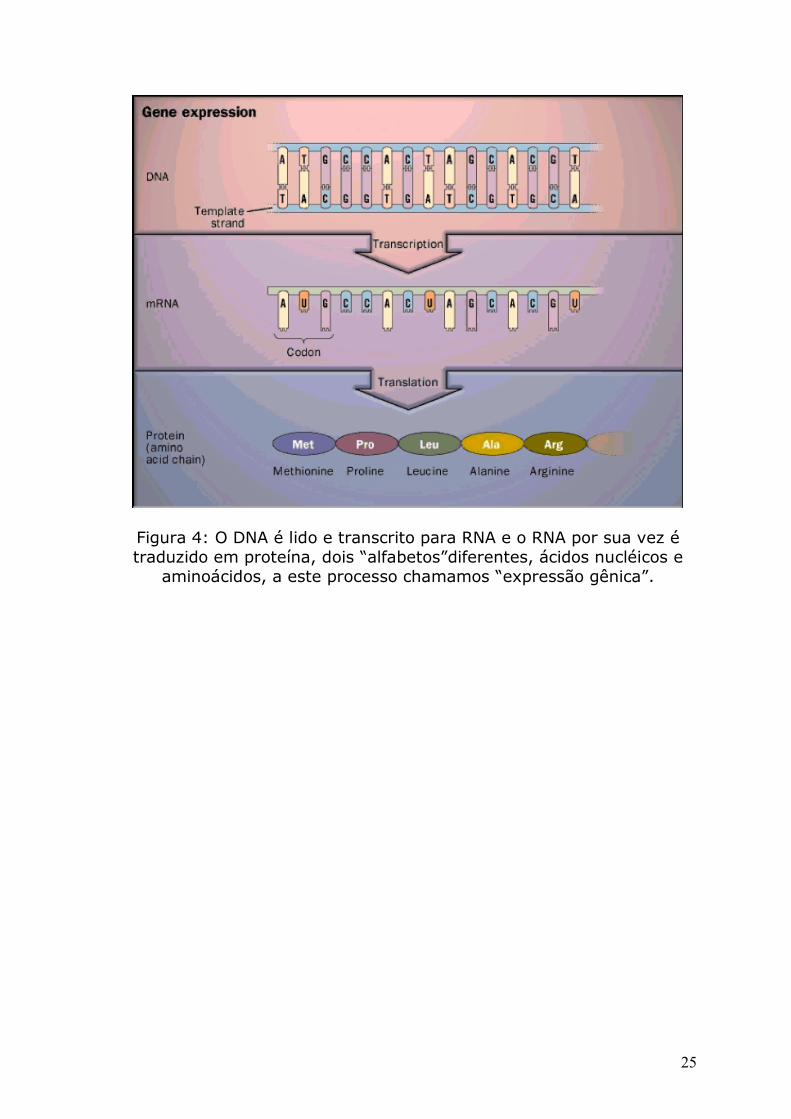
Figura 4: O DNA é lido e transcrito para RNA e o RNA por sua vez é traduzido em proteína, dois “alfabetos”diferentes, ácidos nucléicos e
aminoácidos, a este processo chamamos “expressão gênica”.
26
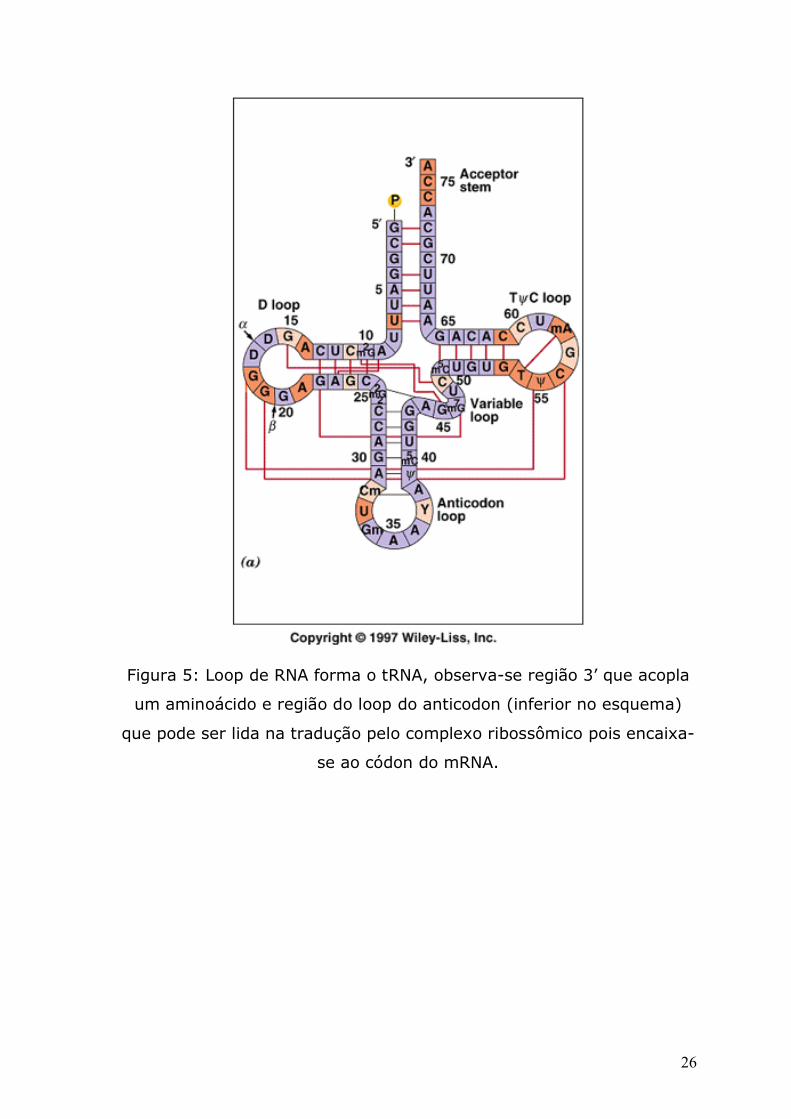
Figura 5: Loop de RNA forma o tRNA, observa-se região 3’ que acopla
um aminoácido e região do loop do anticodon (inferior no esquema)
que pode ser lida na tradução pelo complexo ribossômico pois encaixa-
se ao códon do mRNA.

27
2.2.3 Tradução dos RNAs em proteínas.
Todas as células traduzem RNAs em proteínas de uma mesma
maneira e como regra as moléculas de RNA são muito grandes e
contém as especificações para milhares de proteínas. Os segmentos de
sequências inteiras de DNA são, portanto, transcritos em moléculas de
mRNA separadas, com cada segmento codificando uma proteína
diferente. Um gene é definido como um segmento de DNA
correspondentes a uma única proteína ou a uma única molécula de
RNA catalítico ou estrutural para aqueles genes que produzem RNA
mas não proteínas uma vez que RNAs também podem ter funções por
si só que não a confecção de proteínas.
As moléculas de proteínas, como as moléculas de DNA e RNA,
são cadeias poliméricas longas não ramificadas, formadas por
sequências de blocos construtores monoméricos retirados de um
repertório padrão encontrado em todas as células vivas. Assim como o
DNA e RNA, elas carregam informações em uma forma de sequências
lineares de símbolos como um “alfabeto”. Existem muitas moléculas
diferentes de proteínas em cada célula e descontando-se a água a
massa celular é dada na sua maior parte por elas.
Os monômeros de uma proteína são os aminoácidos e são
completamente diferentes dos monômeros de RNAs e DNAs por isso
damos o nome de tradução à leitura dos mRNAs para confecção das
proteínas, afinal é outro tipo e “alfabeto” que foi utilizado! Ao invés de
4 monômeros foram usados 20 e cada aminoácido é constituído da
mesma estrutura básica, por meio da qual pode-se ligar de um modo-
padrão a qualquer outro aminoácido e ligadas a este esqueleto ou
estrutura básica existem cadeias laterais que atribuem a cada
aminoácido propriedades químicas distintas (figura 4). Cada molécula

28
de proteína, ou polipeptídeo, formada pela união de aminoácidos em
determinada sequência particular adquire conformações específicas,
formando uma estrutura tridimensional de alta especificidade e com
sítios reativos em sua superfície que se unem a outras moléculas e
atuam como enzimas das reações celulares, catalizando-as por
formação e quebra de ligações covalentes. As proteínas tem funções
diversas tais como: estruturais, sensoriais, geradoras de movimentos,
sinalização celular e desta forma colocam em ação as informações
genéticas das células em um ciclo de retroalimentação de forma que
DNAs formam RNAs que formam por sua vez proteínas e estas entram
nos processos celulares incluindo aquelas onde novas moléculas de
DNA são produzidas.
Para a produção dos polímeros de aminoácidos temos quatro
letras de nucleotídeos que se combinam para formar 20 de
aminoácidos, isso é um processo complexo (figura 6A e 6B).
A informação contida nos RNAs é lida em grupos de três
nucleotídeos de forma que para cada triplete de nucleotídeos, chamado
códon, temos um aminoácido correspondente ou seja: codificado na
proteína que irá se formar. Como existem 64 códons possíveis (4 x 4 x
4) mas apenas 20 aminoácidos sabe-se que muitos códons
correspondem a um mesmo aminoácido (figura 7). Os códons dos
RNAs são lidos por pequenas moléculas de outros RNAs, os RNAs
transportadores (tRNAs) de forma que cada tRNA tem em uma
extremidade um aminoácido acoplado e na base (que nada mais é do
que um loop do próprio tRNA) expõe parte da sua sequência de forma
que três letras ou bases nitrogenadas que estão livres para combinar
com as três do mRNA na região do códon e por isso essa região do
tRNA é chamada anticódon e é essa porção que o habilita ao
reconhecimento, por pareamento das bases, desta forma o aminoácido
acoplado a esta molécula de tRNA é levado à sequência e ligado à
cadeia de proteína crescente, esta reação ocorre no ribossomo, que é
uma estrutura ou organela celular formada por outras duas moléculas

29
de RNA (o RNA ribossômico ou rRNA) e mais de 50 proteínas
conjugadas a eles (figura 8). Esta superestrutura une-se à molécula de
mRNA e vai acoplando a ela, de acordo com os códons encontrados, as
moléculas de tRNA correspondentes de forma a ir unindo os
aminoácidos que estas carregam para formar os polipeptídeos (figura
9).
Os marcadores de superfície são moléculas protéicas expressas
na membrana celular, produzidas a partir de genes celulares e ajudam
a identificar uma população de células (Voet D, et al., 2002).
O gene é uma seqüência específica de nucleotídeos do DNA
celular que é lido e transcrito para RNA no núcleo celular e este é
traduzido para uma seqüência de aminoácidos nos ribossomos do
citoplasma da célula formando molécula de proteína que é então
dobrada (folded) e endereçada às organelas para exercer funções
diversas, para ser secretada, ou ainda à membrana celular (figura 10 a
e 10b) para comunicação com o meio extracelular e com outras células
em situações específicas (Alberts et al. 2004), ao processo de
transcrição e tradução do gene damos o nome de expressão gênica e
sua regulação foi explicada adiante.
No caso das proteínas de superfície, ou seja, aquelas que foram
endereçadas e encontram-se fixas à membrana plasmática, é possível
detectar sua presença com o uso de anticorpos que se ligam a estas
proteínas e que podemos ler com ajuda do citômetro de fluxo conforme
descrito na metodologia e assim comprovar qual é a célula em questão
ou qual seu grau de diferenciação dentro de uma linhagem pois células
em estágios diferentes de diferenciação apresentarão proteínas
diferentes e específicas em sua superfície (Voet et al., 2002). Outra
possibilidade para estudar quais proteínas estão sendo produzidas
pelas células de uma amostra em determinado momento e assim
comprovar seu grau de diferenciação ou sua interação com o meio é
detectar diretamente do citoplasma qual ou quais os RNAs que a célula
está produzindo e que vão dar origem a determinadas proteínas, como

30
por exemplo o do gene da STRO-1 (stromelisina 1) que é expresso por
células mesenquimais indiferenciadas que se comprometem com a
linhagem osteoblástica mas que ainda permanecem com suas
características de célula indiferenciada conforme descrito no ítem 1.3
abaixo, para esta detecção é usado o método de RT-PCR também
descrito a seguir.
Figura 6A: Para a produção dos polímeros de aminoácidos temos quatro letras de nucleotídeos que se combinam para formar 20 de aminoácidos. Nesta representação descobrimos qual o aminoácido corresponde a determinado códon partindo de uma letra na região
central e seguindo para fora.

31
Figura 6: Detalhe da figura anterior: observe que a combinação AUG é a única que leva à METionina, este aminoácido sempre significa início
de transcrição dos mRNAs nos ribossomos.
32
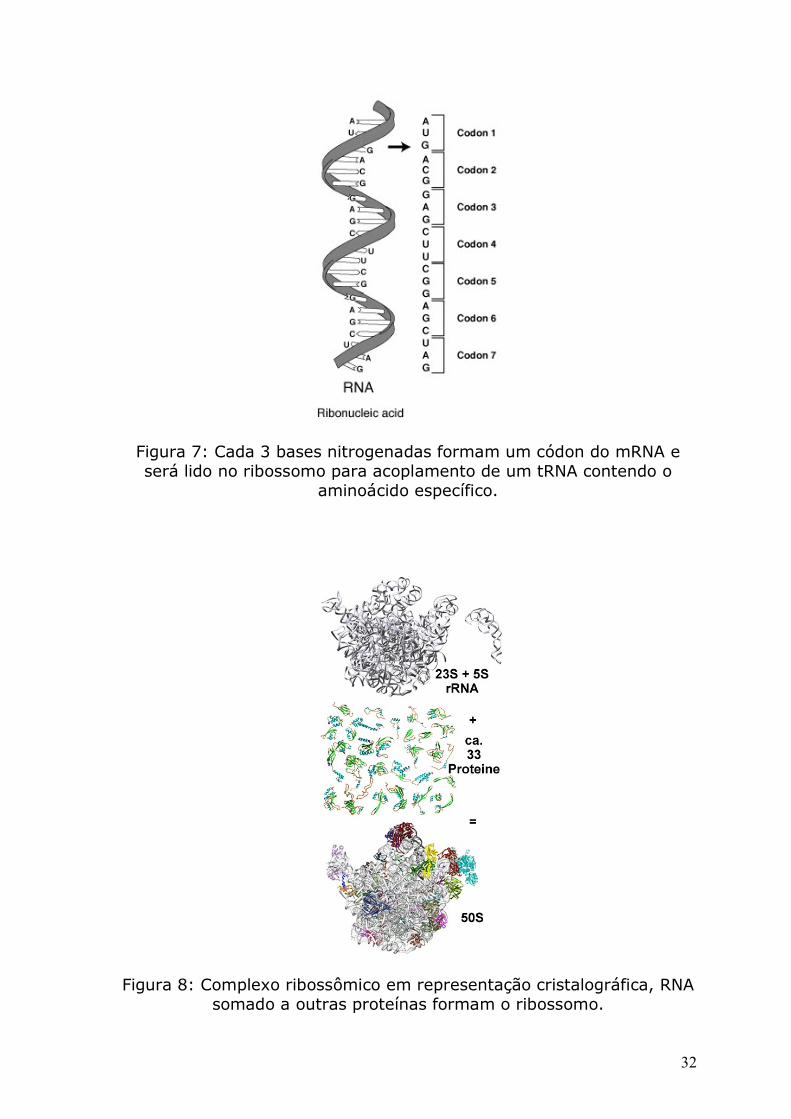
Figura 7: Cada 3 bases nitrogenadas formam um códon do mRNA e será lido no ribossomo para acoplamento de um tRNA contendo o
aminoácido específico.
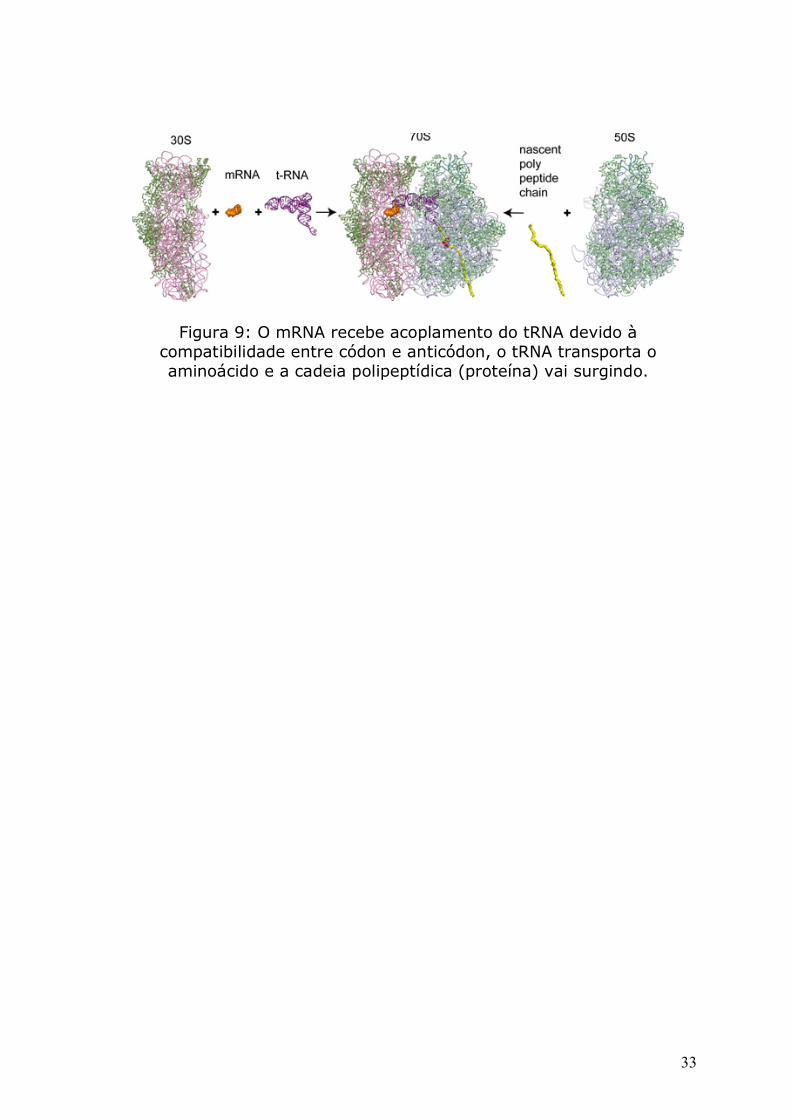
Figura 8: Complexo ribossômico em representação cristalográfica, RNA somado a outras proteínas formam o ribossomo.
33
Figura 9: O mRNA recebe acoplamento do tRNA devido à compatibilidade entre códon e anticódon, o tRNA transporta o aminoácido e a cadeia polipeptídica (proteína) vai surgindo.

34

35
2.3 Uso de células em regeneração tecidual.
O uso de células tronco embrionárias gera questões éticas (Hug,
2006), mas a medicina regenerativa as emprega na tentativa de
reparar tecidos lesionados ou repor perdas teciduais devido à
característica destas células de proliferar em cultura indefinidamente e
seu potencial de diferenciar-se em outros tipos celulares (Gepstein,
2002). São classificadas em três grandes grupos: multipotentes,
pluripotentes e totipotentes.
Totipotentes são células com capacidade de gerar o organismo
completo (Klimankaya et al., 2006), como exemplo temos o ovócito
fecundado e células do blastômero. Pluripotentes são as células com
capacidade de divisão indefinida e que podem se diferenciar em células
dos três folhetos embrionários (Prelle et al., 2002) como as células do
interior do blastocisto, núcleo do embrião e produzem fator de
transcrição Oct-4, que as mantém indiferenciadas e com capacidade de
divisão celular indefinida. Nestes dois casos os questionamentos éticos
surgem, pois é necessário o uso de um embrião para obtenção das
células.
As células multipotentes podem se transformar em vários tipos
celulares e podem ser obtidas de tecidos de indivíduos, como por
exemplo do sangue, de onde obtemos as células tronco
hematopoiéticas (Kaufman et al., 1999). Outra alternativa é a
obtenção de células tronco mesenquimais (MSC) ou células
mesenquimais indiferenciadas, encontradas em diversos tecidos no
decorrer da embriogênese e em adultos (Kassem et al., 2004).
Quanto à utilização das células mencionadas acima em terapia
celular, podemos encontrar na literatura a sua diferenciação em
linhagem condrogênica, osteogênica, adipogênica e miogênica,
sugerindo que é possível a reconstrução de tecidos a partir destes tipos

36
celulares. (Zuk. A et al., 2001). Pode-se, por exemplo, obter células de
tecido adiposo por lipoaspiração (Gimble, 2003) ou polpa dental
(Gronthos, 2000) e tentar diferenciar estas células sob condições
osteogênicas de forma que estas células passam a expressar genes e
proteínas típicas ao fenótipo descrito para osteoblastos, como por
exemplo fosfatase alcalina (ALP), colágeno tipo 1, osteopontina,
osteonectina, osteocalcina, sialoproteína óssea, receptores da proteína
morfogenética de osso (BMP) tipo 2 e 4 e receptor de parathormônio
(Gronthos, 2000; Halvorsen et al., 2001; Zuk et al., 2002).
Encontramos nestas células os marcadores de superfície que
identificam potencial de multidiferenciação (CD105, CD166 e STRO-1)
(Gronthos, 1999), o que as torna candidatas a terapias celulares
diversas inclusive em terapia de regeneração óssea conforme a
presente proposta.
A obtenção de outros tipos celulares chamados de células
mesenquimais indiferenciadas, tais como as hematopoiéticas (HSC)
também foram descritas de forma que é possível obter estes tipos
celulares da medula óssea, do sangue ou do cordão umbilical (Dzierzak
et al., 1998; Kaufman et al., 1999). Estas células são responsáveis
pela constante reposição da linhagem celular hematopoiética e tem
capacidade de proliferação similar a das células embrionárias e estudos
comprovam que estas células têm capacidade de se diferenciar em
outros tipos celulares como neurônios (Mezey et al., 2000), mioblastos
(Bittner et al., 1999) e hepatócitos (Lagasse et al., 2000).
Na área da terapia celular, um substrato para a cultura que seja
absorvível pelo corpo humano pode compor, com as células
osteoblásticas cultivadas sobre ele, um enxerto que pode ser devolvido
ao paciente do qual as células se originaram, desde que sob
determinadas circunstâncias (Sittinger et al., 1996 e Marchiori-Silva et
al., 2008). Experimentos de outros pesquisadores (Perka et al., 1999 e
Schimming & Schmelzeinsen, 2004) comprovam esta possibilidade
terapêutica, mas ainda existe uma lacuna no estudo da ação do laser

37
neste tipo de procedimento que comprove algum benefício e que
indique sua utilidade como parte do protocolo nesta terapia durante a
fase laboratorial de cultivo celular.
2.4 Tecido ósseo e regeneração.
A constituição do tecido ósseo dá-se por uma combinação de
matriz majoritariamente composta por hidroxiapatita e colágeno tipo 1
e componentes celulares. Os componentes celulares são: osteoblastos,
osteócito, esteoclastos, células mesenquimais indiferenciadas.
Os osteoblastos são responsáveis pela deposição da matriz e pela
mineralização. Após depositarem matriz ao redor de si mesmos,
mantêm pequenos canalículos alimentadores e se tornam reféns do
seu próprio material secretado, modificam sua estrutura e passam a
constituir os osteócitos. Os osteoclastos são células de tamanho
grande, multinucleadas e responsáveis pela remodelação constante
sofrida pelo tecido ósseo. Os osteoclastos fazem a reabsorção da
matriz óssea e, acompanhados dos osteoblastos em formação
denominada por Daniel Buser (1994) de cone osteoclástico, o osso vai
sendo aos poucos reabsorvido e refeito em uma troca constante de
íons que mantém o equilíbrio dos minerais envolvidos no corpo do
indivíduo.
As células mesenquimais indiferenciadas no tecido ósseo são
células com capacidade de se diferenciar para promover a reposição de
osteoblastos quando necessário (Gartner e Hiatt, 2001).
Todos os componentes do tecido ósseo se organizam de forma
que a estrutura macroscópica tridimensional possa suportar cargas e
exercer a função de suporte, proteção de órgãos internos, locomoção e
ainda servir como reservatórios de minerais. A medula óssea exerce
também a importante função de fornecer células do sistema
hematopoiético a partir de células precursoras lá presentes (Satija et
al. 2007, Freitas e Dalmau, 2006).

38
Todo este tecido é regulado por estímulos mecânicos, fatores
hormonais, fatores de crescimento e citocinas (Gartner e Hiatt, 2001)
de forma que todo o tecido ósseo sofre remodelação de tempos em
tempos e calcula-se que a cada quatro anos todo o esqueleto adulto
seja renovado (Parfitt, 2002).
O desenvolvimento de processos e técnicas para produzir tecido
ósseo in vitro tem avançado nos últimos anos (Goessler et al., 2006;
Sanmartin et al., 2006; Salgado et al., 2007; Perka et al., 2000) o que
possibilitará no futuro a reposição de perdas ósseas por causas
variadas. Esta área da ciência avançou com técnicas de cultura celular
avançadas, cuidadosamente feitas com intenção de produzir formação
de tecido autólogo in vitro (Sittinger et al., 1996 e 2004; Risbud e
Sittinger 2002).
2.5 Sinalização celular específica: regulação da expressão
gênica.
Toda alteração da expressão gênica da célula está relacionada a
estímulos recebidos pela célula para que determinado gene seja
transcrito e traduzido e desta forma uma resposta é dada a tal
estímulo.
Os estímulos indutores ou inibidores da expressão gênica podem
ser recebidos das células vizinhas, da matriz extra-celular (Alford e
Hankenson, 2006) ou de fonte externa de energia tal como uma fonte
de luz laser (Carvalho et al., 2006) ou um campo eletromagnético (Tsai
et al., 2007) e desta forma são chamados parácrinos ou podem ser de
origem da própria célula e desta forma serem denominados autócrinos
(Alberts, 2004), neste caso substâncias sinalizadoras produzidas pela
própria célula vão atuar em cascatas moleculares nela mesma e
produzir efeito inibitório ou estimulatório.
A transcrição de um gene não depende somente da enzima RNA
polimerase como descrito anteriormente, mais do que isso, o primeiro

39
passo para que um gene seja expresso, ou seja, transcrito, é a ligação
de uma proteína indutora ou “fator de transcrição” à sequência
promotora do gene (promotor). Estes fatores de transcrição se ligam a
sequências específicas do promotor, principalmente na sequência
TATAAA, também conhecida como “TATA box”. Esta sequência é
comum à maioria dos promotores de genes de células eucarióticas.
Assim, o fator de transcrição, que é uma proteína, possui sítios de
reconhecimento desta sequência. A ligação do fator de transcrição ao
“TATA box” propicia a ligação da enzima RNA polimerase ao sítio de
iniciação de transcrição do gene e a síntese do RNA começa (Schor et
al., 2003).
O sítio de iniciação de transcrição (ou domínio do fator de
transcrição) “runt” presente no DNA é um componente integral das
cascatas de sinalização mediadas pela TGF-β e pelas BMPs (Bone
Morphogenetic Proteins) e é ativado a partir da presença destas duas
proteínas nos seus respectivos receptores de membrana.
Quando TGF-β ou BMPs se unem aos seus respectivos receptores
estes se ativam como serinas kinases ou seja, o domínio extra-celular
que contém um sítio de ligação para a molécula sinalizadora ao receber
tal molécula muda de conformação e ativa por fosforilação os resíduos
serina do domínio intra-celular que a partir desta modificação
funcionará como sítio de ligação de proteínas intra-celulares. No caso
das TGF-β as proteínas intra-celulares ativadas pela ligação com o sítio
intra-celular dos receptores são as Smad2 e Smad3 e no caso das
BMPs as Smad1, Smad5 e Smad8 (Miyazono, 1999). Essas proteínas
reguladas por receptores Smad, R-Smad, se associam com a Smad-4
(comum a todas, Co-Smad) e entra no núcleo, lá o complexo R-
Smad/Co-Smad se une ao fator de transcrição para regular a
transcrição do gene, este fator de transcrição é chamado RUNX, assim
chamado por ser homólogo ao gene runt presente na Drosophila ou
seja: “runt related” (Ito e Miyazono, 2003)

40
RUNX-2 é componente importante para a maturação de
osteoblastos pois induz células mesenquimais indiferenciadas à
diferenciação em osteoblastos sendo chave reguladora para
osteocalcina e fosfatase alcalina além de conduzir o sinal dado pela
BMP para regular os genes responsáveis pela osteogênese. Segundo
Gronthos (1999) o Runx-2 se une ao DNA na região promotora do
gene da osteocalcina, regulando assim a sua expressão. Não foram
aboradadas aqui as outras formas do gêne: RUNX-1 e RUNX-3 por
serem respectivamente ligados à hematopoiese/angiogênese e à
supressão de tumores, características que fogem ao propósito deste
trabalho mas merecem serem citadas (Ito e Miyazono, 2003).
Segundo Gronthos (1999) os osteoblastos podem ser
caracterizados histologicamente pela alta expressão de fosfatase
alcalina e bioquimicamente pela síntese de certas proteínas de matriz
extra-celular específicas tais como: colágeno tipo I, osteocalcina,
sialoproteína óssea, osteonectina e osteopontina. entretanto esta
heterogeneidade de fenótipos acontece de acordo com a fase
proliferativa e funcional da população celular. Juntos estes marcadores
são úteis em determinar comprometimento osteogênico mas não
podem ser associados de forma indubitável com células em discreto
estágio de diferenciação, além disso as precursoras menos
diferenciadas continuam sem uma identificação pela ausência de
marcadores confiáveis para tais células.
O grupo de Gronthos, liderado por Simmons em 1991, já havia
identificado um marcador de superfície para células precursoras de
medula óssea chamado STRO-1 que segundo estes autores estaria
associado e serviria para caracterização de células fibroblásticas
formadoras de colônias, ou seja, células precursoras indiferenciadas
que sob diferentes condições de cultura se diferenciam em diferentes
tipos celulares como osteoblastos, adipócitos e células musculares.
O STRO-1 está presente em células precursoras de osteoblastos
e quando as células expressam a fosfatase alcalina é possível observar

41
mineralização in vitro (Gronthos 1999) o que demonstra diferenciação
para osteoblasto, segundo estes autores a diferenciação leva a uma
redução na expressão do STRO-1 e aumento na expressão da ALP
(fosfatase alcalina). Estudo mais recente (D’Aquino 2007) revela que o
gene da STRO-1 está associado a células com potencial osteogênico e
no caso de células da polpa dental este gene estaria presente após o
cultivo destas células em meio osteogênico por pelo menos 20 dias no
entanto o gene RUNX-2 somente estaria presente após 45 dias em
cultura osteogênica uma vez que este, segundo os autores, seria um
gene expresso apenas em osteoblastos maduros. Neste mesmo
trabalho é possível entender como a população de células da polpa é
heterogênea pois os autores encontraram uma grande porcentagem de
células STRO-1+ e cerca de 30% de STRO-1 – e afirmam que esta
presença de células STRO-1 – é características das células precursoras
endoteliais e por isso esta população de células tem capacidade de se
diferenciar em tecido ósseo vascularizado. Ainda neste trabalho ficou
constatado que nenhuma molécula precursora de dentina foi
encontrada nessa população, ao contrário do que se imagina uma vez
que o tecido de onde as células saíram é contido na câmara pulpar de
terceiros molares.

42
3. Objetivos
• Desenvolver protocolo de obtenção de células
mesenquimais indiferenciadas a partir de explantes e de
cultivo dessas células da polpa dental;
• Investigar a ação do laser de λ=830nm sobre as células
isoladas dos explantes;
• Avaliar a expressão dos genes: STRO-1, Runx-2, ALP, OC,
OCT-4 e NANOG sob ação da luz laser neste comprimento
de onda.
• Caracterizar fenotipicamente as linhagens obtidas

43
4. MATERIAL E MÉTODOS 4.1 Meios de cultura 4.1.1 Meio de cultura Basal α-MEM (Minimum Essential Medium, Gibco) suplementado com
5mM de bicarbonato de sódio (Merck), 10% de soro fetal bovino
(Gibco); 100 unidades de penicilina/mL; 0,23mg de streptomicina/mL
(Gibco).
4.1.2 Meio de cultura osteogênico
Para indução de diferenciação de células mesenquimais em
linhagem osteogênica foi usado α-MEM (Minimum Essential Medium,
Sigma) enriquecido com FBS (soro fetal bovino) 10%; 10mM de β-
glicerofosfato; 10nM de dexametasona e 50 µM de ácido ascórbico
(vitamina C).
4.2 Linhagens celulares: células-tronco humanas isoladas de
polpa dental.
Terceiros molares extraídos de pacientes com indicação para tal
procedimento (figuras 11 e 12) na clínica do CETAO
(www.cetao.com.br) e doados pelos pacientes sob termo de doação e
consentimento foram encaminhados ao laboratório em meio de cultura
basal para extração da polpa dental. A extração foi feita em condições
assépticas por corte nos dentes e fratura do remanescente de forma a
evitar contaminação do tecido por broca. No fluxo laminar em
laboratório BL-2 (Biosecurity Level 2) no CINTERGEN (Centro
Interdisciplinar de Terapia Gênica, Escola Paulista de Medicina) foi
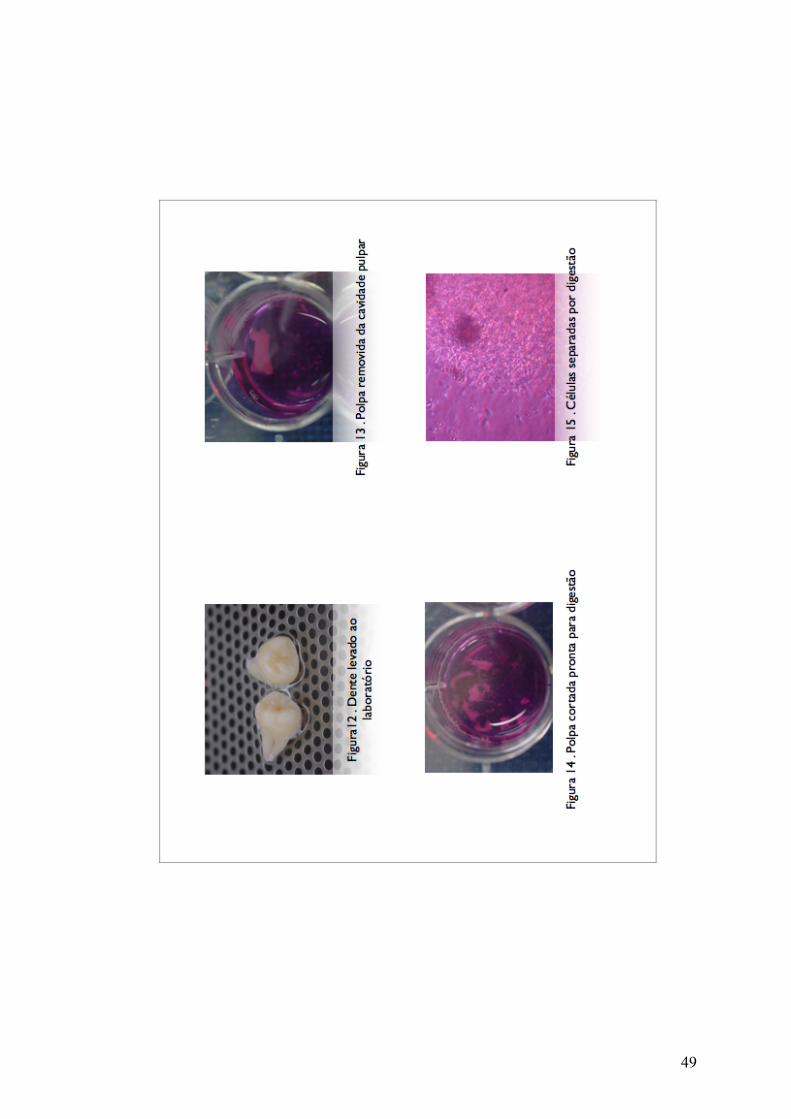
retirado o tecido da câmara pulpar (figura 13) e em seguida este foi

44
digerido (figura 14) por uma mistura de colagenase tipo I 3mg/mL e
dispase 4mg/mL a 37°C por 1 hora, filtrado com filtros Falcon 70µm
(BD Biosciences), centrifugados a 1200rpm por 2 minutos. O pellet
formado foi ressuspendido em meio basal com FBS (Gibco) 10%,
penicilina/streptomicina 1% e as células obtidas foram plaqueadas em
frascos de cultura (figura 15) e incubadas a 37°C com 5% CO2 por 4
dias antes da primeira troca de meio para que aconteça adesão celular,
conforme descrito por Gronthos et al. (2000) e aperfeiçoado por nossa
equipe (Marchiori-Silva et al., 2008) com a metodologia de seccionar
parcialmente os dentes durante o procedimento clínico e fraturá-lo em
ambiente estéril para remover a polpa.
Após a primeira troca de meio de cultura as trocas de meio
passaram a ser feitas duas vezes por semana e as passagens, ou seja,
as trocas de placa, ocorreram no décimo e no vigésimo dias quando as
culturas estavam em subconfluência.
Uma amostra inicial foi coletada para a citometria de fluxo e para
o RT-PCR inicial.
4.3 Citometria de fluxo
A análise das moléculas de superfície das células-tronco
mesenquimais isoladas de polpa dental humana foram realizadas por
citometria de fluxo em amostra de células retiradas de polpas recém
extraídas. Os resultados foram plotados na forma de histograma.
As células mesenquimais indiferenciadas de polpa dental são
caracterizadas por citometria de fluxo através da análise da presença
(+) de moléculas da superfície celular: CD117 (+), CD81 (+), CD105
(+), conforme encontrado na literatura (Lindros et al., 2008).
3.3.1 Marcação celular com anticorpos primários e secundário.

45
As células-tronco da polpa foram removidas das placas de cultura
pela utilização de tripsina 10% e contadas através da câmara de
Neubauer. Aproximadamente 5.105 células foram incubadas por 30
min, a 4ºC com o anticorpo para marcação de moléculas de superfície
CD105 (BD), CD81 (BD), e CD 117 (BD) na diluição de 1:10. Após o
período, as células foram lavadas duas vezes com PBS e fixadas (1%
paraformaldeído, 0,1% azida sódica e 0,5% SFB). As células foram
analisadas em FACSCalibur flow cytometer (BD Biosciences).
4.4 Extração de RNA e RT-PCR
O RNA total foi isolado de 1 x 106 células usando o reagente
TRIZol (Gibco). O RNA foi precipitado na fase aquosa com álcool
isopropílico. O precipitado resultante foi ressuspendido em água DEPC
(di-etil pirocarbonato). A concentração do RNA foi determinada pela
leitura da absorbância em 260/280nm (nanodrop, Thermo Scientific).
O cDNA foi sintetizado a partir de 1µg de RNA total, oligo dt e a enzima
transcriptase reversa SuperScript III (Invitrogen). O cDNA resultante
foi diluído a uma concentração de 20µg /µl e utilizado nas reações de
PCR (Polymerase Chain Reaction) utilizando-se o reagente Mastermix
(Qiagen) e os primers (fermentas) para: OCT-4, sinalizador de
indiferenciação forward: GAGAACAATGAGAACCTTCAGGA e reverse:
TTCTGGCGCCGGTTACAGAACCA; Nanog, sinalizador de indiferenciação
forward: TGCCTCACACGGAGACTGTC e reverse:
TGCTATTCTTCGGCCAGTTG; GAPDH, house keeping genes forward:
AGCCACATCGCTCAGACACC e reverse: GTACTCAGCGGCCAGCATCG;
STRO-1, que sinaliza capacidade das células mesenquimais
indiferenciadas para diferenciar em linhagem osteoblástica forward:
CGCAAATCCCTCAGGAAGCTTGAA e reverse:
AAACCTAGGGTGTGGATGCCTCTT; RUNX-2, sinalizador de
diferenciação para linhagem osteoblástica forward: CGC CCC TCC CTG
AAC TCT e reverse: TGC CTG CCT GGG ATC TGT A; Osteocalcina (OC)

46
forward: CTG ACA AAG CCT TCA TGT CCA A e reverse: GCG CCG GAG
TCT GTT CAC TA; Fosfatase Alcalina (ALP) forward: TTG TGC GAG AGA
AAG GAG A e reverse: GTT TCA GGG CAT TTT TCA AGG T que
identificam atividade osteoblástica (Gronthos et al., 1999).
4.5 Protocolo de irradiação e cultura celular.
O tecido pulpar sofreu digestão em dispase e colagenase
conforme descrito acima, a seguir as células foram filtradas e
centrifugadas em seguida foram plaqueadas em meio basal.
Um equipamento emissor de laser diodo de GaAlAs λ=830nm,
DMC equipamentos, foi usado para a irradiação laser. Foi feita de
forma a obter-se a densidade de energia de 3J/cm2 (Fujihara, 2001)
seguindo para isto a seguinte fórmula: D=PxT/A onde teremos como
potência 80mW determinados no equipamento. A irradiação foi feita
sob a placa de cultura de 6cm com espaçador fornecido pelo fabricante
para garantir que a incidência do feixe laser coincidisse com os
parâmetros mostrados no painel do equipamento e de forma contínua,
em movimento espiral do centro para as bordas da placa e no sentido
inverso, até que se esgotasse o tempo de irradiação pré-determinado
pelo painel.
No plaqueamento as células foram divididas em dois grupos, o
grupo C (GC)- grupo controle que não recebeu irradiação laser e o
grupo L-(GL), que recebeu irradiações conforme protocolo descrito a
seguir. Ambos os grupos passaram pelas mesmas trocas de meio de
cultura e passagens.
O grupo LASER foi irradiado segundo o seguinte protocolo:
Primeiro dia sem laser – isolamento das células e plaqueamento.
Segundo dia sem laser
Terceiro dia sem laser

47
Quarto dia 3J/cm2 – primeira troca do meio de cultura
Quinto dia
Sexto dia 3J/cm2 – troca do meio de cultura
Sétimo dia
Oitavo dia 3J/cm2 – troca do meio de cultura
Nono dia
Décimo dia 3J/cm2 – Primeira passagem e substituição do meio
de cultura basal por meio de cultura osteogênico.
Décimo primeiro dia
Décimo segundo dia 3J/cm2 – troca do meio de cultura
Décimo terceiro dia
Décimo quarto dia 3J/cm2 – troca do meio de cultura
Décimo quinto dia
Décimo sexto dia 3J/cm2 – troca do meio de cultura
Décimo sétimo dia
Décimo oitavo dia 3J/cm2 – troca do meio de cultura
Décimo nono dia
Vigésimo dia 3J/cm2 – Segunda passagem
Vigésimo primeiro dia
Vigésimo segundo dia – 3J/cm2 coleta de RNA para RT-PCR
48
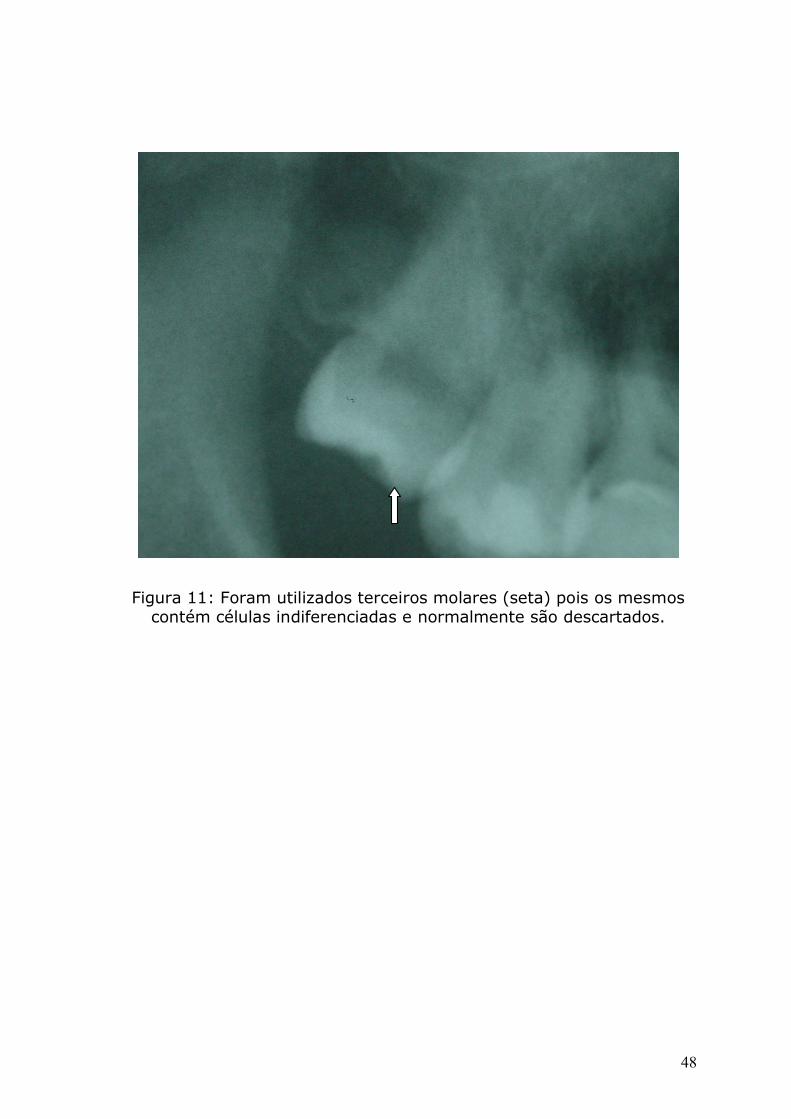
Figura 11: Foram utilizados terceiros molares (seta) pois os mesmos
contém células indiferenciadas e normalmente são descartados.
49

50
5. Resultados e discussão
Células de polpa dental foram isoladas pelos métodos descritos
acima. As células isoladas das polpas dentais de pacientes foram
caracterizadas com uso de citômetro de fluxo conforme descrição da
técnica informada acima e foi constatado que a população de células
obtidas corresponde ao esperado, ou seja células mesenquimais
indiferenciadas de polpa dental (DPMSC).
A presença dos marcadores de superfície celular CD117, CD 81 e
CD 105 confirmaram que a população de células obtidas pela
metodologia descrita pode ser considerada como uma população de
células mesenquimais indiferenciadas, ou seja, células tronco de
adultos (figura 16). A população celular apresentou negatividade para
os marcadores CD45, CD14 e CD34 (figura 17) comprovando também
serem o tipo celular buscado. A parte da população positiva para o
marcador de superfície CD34 representa células tronco
hematopoiéticas dentro desta cultura, esta presença já era esperada
uma vez que o tecido hematopoiético e vasos estão presentes na polpa
dental entretanto as células hematopoiéticas não de desenvolvem em
culturas deste tipo pois são células que não crescem aderidas mas sim
em suspensão, desta forma é esperado que esta contaminação
desapareça no tipo de cultura celular em placas que foi utilizado aqui,
da mesma forma células endoteliais já diferenciadas não se mantém
por muito tempo no meio de cultura utilizado e também tendem a
desaparecer, o meio de cultura α-MEM é adequado às células
indiferenciadas por ser um meio pobre em nutrientes inclusive
facilitando a não diferenciação num primeiro momento mas as
endoteliais não se mantém bem com este meio de cultura.
As células obtidas foram então cultivadas (figura 18) em placas
de cultura estática em meio basal α-MEM não osteogênico inicialmente
e osteogênico após a primeira passagem até o final e dividida em dois

51
grupos: grupo A, que recebeu irradiação laser conforme descrito
(figura 19 e 21) e grupo B, que não recebeu irradiação laser (figura 20
e 22).
Amostras do RNA das células foram obtidas logo após seu
isolamento da polpa bem como depois da irradiação laser no grupo A e
depois da segunda passagem do grupo B e foram analisadas por RT-
PCR para determinar se os RNAs presentes nas células, nesses tempos,
correspondiam àqueles esperados.
Foram encontrados RNAs nas células isoladas correspondentes à
expressão dos genes Oct-4 e Nanog logo após a obtenção destas dos
dentes extraídos (figura 23), a presença de mRNAs correspondentes a
estes genes comprovaram que estas células são indiferenciadas, como
as descritas por Prelle em 2002, as células ainda indiferenciadas tem
maior capacidade de proliferação e também capacidade de diferenciar-
se em alguns tipos celulares e a presença destes genes sugere
portanto que este grupo celular pode ser expandido em cultura e assim
pode ser utilizado em terapias celulares no que se refere à quantidade
de células que pode ser obtida a partir do explante, no entanto
técnicas de cultura avançadas são necessárias para que o número de
células aumente sem que ocorra diferenciação para tipos celulares
indesejados como por exemplo em caso de utilização em regeneração
óssea ocorrer diferenciação para fibroblastos. Células cultivadas em
placas como foi o caso do presente estudo tendem a se diferenciar com
mais facilidade pois o contato com as células vizinhas por si só já é um
estímulo e esta diferenciação pode ocorrer para tipos celulares
indesejados pois não é uma diferenciação ocorrida por estímulo
específico do meio de cultura osteogênico por exemplo mas sim por
estímulo do contato entre as células, além disso a cultura pára de
expandir quando as placas estão em confluência e há pouco espaço
para que as células se desenvolvam, conforme observamos ao
microscópio as placas irradiadas com laser rapidamente chegavam à
subconfluência (figuras 19 e 21).

52
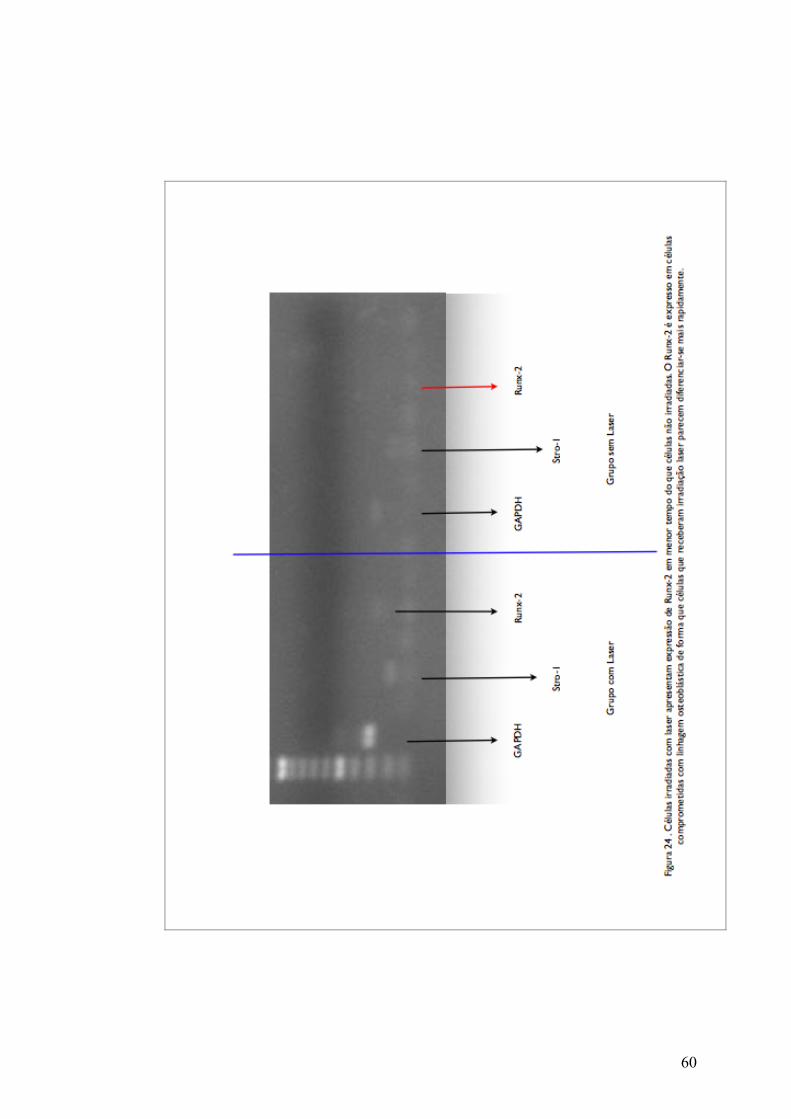
Observamos também que o gene STRO-1 estava ativo em fase
inicial (figura 24), segundo Gronthos este gene caracteriza células em
estágio inicial de diferenciação mas ainda precursoras de tipos
celulares variados pois este autor constatou a diferenciação destas
para outros tipos celulares além da linhagem osteogênica, de qualquer
forma a presença deste gene indica uma capacidade osteogênica,
existe uma dificuldade grande na literatura em se determinar
diferenças entre os tipos celulares de uma mesma linhagem em estágio
de diferenciação diferentes mas muito próximos, ou seja, células que
estão ainda num estágio muito inicial de diferenciação apresentam
poucas diferenças em relação umas às outras, mesmo que entre elas
os estágios de diferenciação não sejam os mesmos. Também segundo
estes autores o gene da Stro-1 vai aos poucos sendo menos evidente
ao longo da diferenciação e o gene Runx-2 passa ser expresso ao longo
da diferenciação osteoblástica, no presente trabalho detectamos por
PCR a presença do gene Runx-2 nas células irradiadas com o laser
antes que esta expressão ocorresse nas células não irradiadas (figura
24) e isso nos permite afirmar que o estágio de diferenciação das
células irradiadas para linhagem osteoblástica aconteceu antes.
Podemos assim ter duas conclusões possíveis, a primeira é que não
necessariamente o laser causa diferenciação mas como os processos
ligados ao metabolismo celular são acelerados a diferenciação que
ocorreria normalmente acaba acontecendo em menos tempo portanto
a expressão dos genes aconteceu antes de uma forma indireta e não
pela sua ativação por parte do estímulo da luz laser ou então a outra
conclusão que é a de que o laser pode causar sim diferenciação pela
ativação direta dos genes responsáveis e neste caso será necessária
uma investigação mais específica da relação entre a irradiação e a
expressão de cada gene com por exemplo medindo-se a proliferação
da cultura em relação à ativação dos genes. Ainda que a expressão dos
genes possa ser alterada pela irradiação com o laser temos que levar
em conta também a observação nossa e de outros autores de que o

53
laser aumenta a proliferação celular, como exposto anteriormente a
proliferação ocorre em momento anterior à diferenciação e após a
diferenciação as células proliferam menos por isso a hipótese de que o
laser pode acelerar os processos naturais nos parece mais realista uma
vez que caso o laser levasse à ativação dos genes ligados à
diferenciação antes que ocorresse a proliferação destas células na
cultura poderíamos observar menos proliferação pois células
diferenciadas proliferam menos, acreditamos que a diferenciação
acontece antes porque todos os processos relacionados às culturas
celulares aconteceram antes também e assim a expressão dos genes
estudados ocorreu antes no grupo irradiado mas de forma indireta, ou
seja, pelo aumento da velocidade do processo como um todo. Podemos
ainda levantar a hipótese de que o laser ative os genes estudados e
ainda assim mantenha as células em proliferação mas isso seria como
dizer que a célula já diferenciada poderia continuar proliferando o que
é incompatível tanto com a literatura quanto com a experiência clínica
pois células diferenciadas em proliferação estariam em um mecanismo
muito similar ao de um tecido tumoral e isso não tem sido associado ao
uso do laser.
Ambos os grupos testados não apresentaram mRNAs
correspondentes aos genes da osteocalcina e da fosfatase alcalina, esta
constatação pode ser explicada devido ao pouco tempo de cultura
utilizado neste trabalho, segundo Gronthos (2007) a osteocalcina e
principalmente a fosfatase alcalina são produzidas por osteoblastos já
maduros e a diferenciação dos tipos celulares estudados segundo a
literatura ocorreria próxima aos sessenta dias de cultura celular de
forma que mesmo acelerando a cultura a observação da produção
destas moléculas não seria possível antes de trinta dias, é possível que
a osteocalcina esteja presente pouco antes da fosfatase alcalina mas
mesmo esta primeira é um marcador de célula osteoblástica madura e
não em estágios iniciais de diferenciação ou então em diferenciação
não completa.

54
As células irradiadas com laser nos parâmetros determinados
para este trabalho apresentaram proliferação mais organizada em
relação às células que não receberam laser.

55

56

57

58

59
60

61
6. CONCLUSÕES É possível obter células mesenquimais indiferenciadas de polpa dental pela técnica proposta, caracterizá-las e cultivá-las para expandir sua quantidade. A irradiação laser λ=830nm promove mudanças no metabolismo celular que influenciam na velocidade da expressão dos genes aqui estudados. Foi possível demonstrar que as células indiferenciadas de polpa dental podem iniciar diferenciação para linhagem osteoblástica de forma mais rápida com o uso da luz laser em λ=830nm. Para a completa diferenciação das células estudadas para osteoblastos será necessário mais tempo de cultura. Mais estudos serão necessários para determinar a exata influência do laser na expressão gênica celular.

62
Referências Bibliográficas
1. Alberts B. et al. Biologia Molecular da Célula. 1463pp, 4 ed. Porto Alegre: Artmed 2004.
2. Alford A. I.; Hankenson K. D. Matricellular proteins: Extracellular modulators of bone development, remodeling and regeneration. Bone 38: 749-757, 2006.
3. Bittner et al. Recruitment of bone-marrow-derived cells by skeletal and cardiac muscle in adult dystrophic mdx mice. Anatomy and Embryology 199: 391-396, 1999.
4. Blay, A. Efeitos da radiação laser em baixa intensidade no mecanismo de osseointegração de implantes: estudo “in vitro”. Dissertação apresentada, IPEN-FOUSP, 2001.
5. Boyne P.J. Regeneration of alveolar bone beneath cellulose acetate filter implants. J Dent Res, 43:827, 1964.
6. Burger E.H.; Klein-Nulend J. Microgavity and bone cell mechanosensitivity. Bone, vol. 22 n°5 suplemento:127s-130s, 1998.
7. Buser D. et al. Guided Bone Regeneration. 270pp, Quintessence pub. 1 ed. 1994.
8. Carvalho P. T. C. et al. Effect of 650 nm low-power laser on bone morphogenetic protein in bone defects induced in rat femors. Acta Cirúrgica Brasileira, 21:63-68, 2006.
9. D’Aquino R et al. Human post natal dental pulp cells co-differentiate into osteoblasts and endotheliocytes: a pivotal synergy leading to adult bone tissue formation. Cell Death and Differentiation, 14:1162-1171, 2007.
10. Dzierzak E; et al. Qualitative and quantitative aspects of hematopoietic cell development in the mammalian embryo. Immunol Today, 19: 228-236, 1998.
11. Freitas CS., Dalmau SR. Multiple sources of non-embryonic multipotent stem cells: processed lipoaspirates and dermis as promissing alternatives to bone –marrow-derived cell therapies. Cell Tissue Res 325(3): 403-411, 2006.
12. Freitas I.G.F.; et al. Laser effects on osteogenesis. Applied surface Science, 154-155:548-554, 2000.
13. Fujihara N A Estudo da adesão, proliferação e síntese de proteínas por osteoblastos cultivados e submetidos à ação do laser de baixa potência. Tese de mestrado em patologia bucal, USP, 2002.
14. Gartner LP; Hiat JL Color textbook of histology. 2nd ed. Philadelphia, London, New York, St. Louis, Sydney and Toronto: WB Saunders Co., 2001.

63
15. Gepstein L. Derivation and potential applications of human embryonic stem cells. Circulation research, 91: 866-876, 2002.
16. Gimble J., Guilak F. Adipose derived adult stem cells: isolation, characterization and differentiation potential. Cytotherapy 5: 362-369, 2003.
17. Goessler et al. Perspectives of gene therapy in stem cell tissue engineering. Cells Tissues Organs 183(4): 169-179, 2006.
18. Gronthos S. et al. Differential cell surface expression of the STRO-1 and Alkaline Phosphatase Antigens on discrete develomental stages in primary culture of human bone cells. J Bone Miner Res 14: 47-56, 1999.
19. Gronthos S.; et al. Postnatal human dental pulp stem cells (DPSCs). Proc Nati Acad Sci, 97(25):13625-13630, 2000.
20. Halvorsen YD et al. Extracellular matrix mineralization and osteoblast gene expression by human adipose tissue derived stromal cells. Tissue Eng 7: 729-741, 2001b.
21. Hawkins D.; Abrahamse H. Effect of multiple exposures of low-level laser therapy on the cellular responses of wounded human skin fibroblasts. Photomedicine and Laser Surgery, 24, 6:705-714, 2006.
22. Hug K. Therapeutic perspectives of human embryonic stem cell research versus the moral status of a human embryo – does one have to be compromissed for other? Medicina (kaunas). 42(2):107-114, 2006.
23. Ito Y; Miyazono K RUNX transcription factors as key targets of TGF-β superfamily signaling. Current Opinion in Genetics and Development 13: 43-47, 2003.
24. Janssen F.; te al. A perfusion bioreactor system capable of producing clinically relevant volumes of tissue-engineered bone: in vivo proof of concept. Biomaterials, 27: 315-323, 2006.
25. Karu T. The Science of Low-Power Laser Therapy. 299pp, Overseas Publishers Association, New Delhi,1998.
26. Kassem M; et al. Mesenchymal stem cells: cell biology and potencial use in therapy. Basic Clin Pharmacol Toxicol 95: 209-214, 2004.
27. Kaufmann D S, et al. Directed differentiation of human embryonic stem cells into hematopoietic colony forming cells. Blood, 94: (supplement part 1) 34a, 1999.
28. Klimankaya I. et al. Human embryonic stem cells lines derived from single blastomeres. Nature, 2006: 1-5.
29. Lagasse E et al. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat. Med, 6: 1229-1234, 2000.

64
30. Lynch S.E. et al. Tissue Engineering. 286pp, Quintessence pub Co, Illinois, 1999.
31. Mai J., et al. A microfabricated platform probing cytoskeleton dynamics using multidirectional topografical cues. Biomed Microdevices, 9:523-531, 2007.
32. Marchiori-Silva V. et al. Polyhydroxybutirate (PHB) from sugar cane as biomaterial for bone tissue engineering using Mesenchymal Stem Cells derived from human dental pulp. The Int’l Journal of Artificial Organs 31(7): 600-601, 2008.
33. Mezey E et al. Turning blood into brain: cells bearing neuronal antigens generated in vivo from bone marrow. Science 290: 1779-1782, 2000.
34. Miyazono K Signal Transduction by Bone Morphogenetic Protein Receptors: Functional Roles of Smad Proteins. Bone 25(1): 91-93, 1999
35. Nozaki T et al. Differentiation of rat dental pulp-derived cells into an osteoblastic lineage. Oral Science International 2(2):118-125, 2005.
36. Ozawa Y et al. Low energi laser irradiation stimulates bone nodule formation at early stages of cell culture in rat calvarial cells. Bone 22(4): 347-354, 1998.
37. Parfitt AM. Targeted and nontargeted bone remodeling: relatioinship to basic multicellular unit origination and progression. Bone 30: 5-7, 2002.
38. Perka, C. et al. Segmental bone repair by tissue-engineered periosteal cell transplants with bioresorbable fleece and fibrin scaffolds in rabbits. Biomaterials 21: 1145-1153, 2000.
39. Prelle K; et al. Pluripotent stem cells – model of embryonic development, tool for gene targeting and basis of cell therapy. Anat Histol Embryol 31(3): 169-186, 2002.
40. RIBEIRO, M. S. ; ZEZELL, Denise Maria . Laser de Baixa Intensidade. In: Carlos de Paula Eduardo; Norbert Gutknecht. (Org.). A Odontologia e o Laser. Atuação do laser na especialidade odontológica. 1 ed. São Paulo: Quintessence Editora Ltda., 2004, v. 1, p. 217-240.
41. Risbud M. e Sittinger M. Tissue engineering: advances in in vitro cartilage generation. Trends in Biotechnology 20:8, 2002.
42. Saito S.; Shimizu N. Stimulatory effects of low-power laser irradiation on bone regeneration in midpalatal suture during expansion in the rat. Am J Orthod Dentofacial Orthop 111(5):525-532, 1997.
43. Salgado AJ, Coutinho OP., Reis, RL., Davies, JE. In vivo response to starch-based scaffolds designed for bone

65
tissue engineering applications. J Biomed Mater Res 80A: 983-989, 2007.
44. Sanmartin A. et al. Stem cells in cell transplantation. Stem Cells Dev 15(6): 963-966, 2006.
45. Satija NK. et al. Mesenchymal stem cells: molecular targets for tissue engineering. Stem Cells Dev. 16: 7-23, 2007.
46. Shimming R; Schmelzeisen R. Tissue-engineered bone for maxillary sinus augmentation. J Oral Maxillofac Surg 62: 724-729, 2004.
47. Schor N. et al. Bases moleculares da Biologia, da Genética e da Farmacologia. 382pp, ed. Atheneu: São Paulo, 2003.
48. Simmons P J; Torok-Storb B Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody, STRO-1. Blood 78:55-62, 1991.
49. Sittinger M. et al. Tissue engineering and autologous transplant formation: practical approaches with resorbable biomaterials and new cell culture techniques. Biomaterials 17: 237-242, 1996.
50. Sittinger M. et al. Current strategies for cell delivery in cartilage and bone regeneration. Current Opinion in Biotechnology 15:411-418, 2004
51. 52. Trelles M; Mayayo E Bone fracture consolidates faster
with low-power laser. Lasers Surg Med, 7(1): 36-45, 1987. 53. Tsai, M.; et al. Pulsed electromagnetic fields affect
osteoblast proliferation and differrentiation in bone tissue engineering. Bioelectromagnetics 2007. Disponivel em http://www.interscience.wiley.com
54. Tuby H et al. Low-Level Laser Irradiation (LLLI) Promotes proliferation of mesenchymal and cardiac stem cells in culture. Lasers in Surgery and Medicine 39: 373-378, 2007.
55. Voet, D et al. Fundamentos de Bioquímica 930pp, 1a reimpressão: Artmed 2002.
56. Wilden L.; Karthein R. Import of radiation phenomena of electrons and therapeutiy low-level laser in regard to the mitochondrial energy transfer. Journal of clinical Laser Medicine and Surgery, 16, 3: 159-165, 2000.
57. Zheng Q.; et al. Could the effect of modeled microgravity on osteogenic differentiation of human menchymal stem cells be reversed by regulation of signaling pathways? Biol. Chem., 388:755-763, 2007.
58. Zuk PA. et al. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng 7: 211-228, 2001.

66


















