Auxin-binding protein located in the endoplasmic reticulum of maize ...
5
Proc. Nadl. Acad. Sci. USA Vol. 86, pp. 3564-3568, May 1989 Biochemistry Auxin-binding protein located in the endoplasmic reticulum of maize shoots: Molecular cloning and complete primary structure (cDNA sequence/N-glycosylation/plant hormone receptor/retention signal) NAOHIRO INOHARA, SHOJI SHIMOMURA*, TOSHIO FUKUI, AND MASAMITSU FUTAI Institute of Scientific and Industrial Research, Osaka University, Ibaraki, Osaka 567, Japan Communicated by Harry Beevers, February 3, 1989 ABSTRACT We previously purified an auxin-binding pro- tein (ABP) from the microsomal fraction of maize shoots (Zea mays L. cv. Golden Cross Bantam). In the present study cDNA clones derived from mRNAs encoding the ABP were isolated and sequenced. The nucleotide sequence of the 822-base-pair cDNA includes a 603-base-pair open reading frame. RNA blot hybridization analysis indicated a single mRNA species of =1.0 kilobase. The predicted precursor of ABP is composed of 201 amino acid residues and has a molecular weight of 21,976. The NH2-terminal sequence of 38 residues is hydrophobic and may be a signal peptide for translocation of the ABP across the membrane of the endoplasmic reticulum. The mature ABP, composed of 163 residues with a molecular weight of 18,352, contains a potential N-glycosylation site (Asn-Thr-Thr), and the COOH-terminal tetrapeptide (Lys-Asp-Glu-Leu) may be a signal for retention of the ABP in the lumen of the endoplasmic reticulum. The plant hormone auxin (indole-3-acetic acid) in higher plants functions in regulating normal growth and develop- ment, including cell elongation, division, and differentiation (1, 2). Rapid responses to auxin have been observed in organelles such as plasma membrane (3-6), Golgi apparatus (7), endoplasmic reticulum (ER) (8), and nucleus (2, 9, 10) as well as in the cell wall (11). The specificity of auxin is sup- posed to be due to the presence of a specific receptor(s) (12). However, it is unknown whether the diverse responses of different organelles to auxin are mediated by a single or multiple species of receptors. Auxin can enter cells because it is a membrane-permeant, lipophilic weak acid. In addition, specific transporters for auxin have been demonstrated in plasma membranes (13-16). Specific and saturable auxin-binding sites are also found in other organelles such as ER, tonoplast, and nucleus and in the cytosol (12), and these should be taken into consideration as possible candidates for auxin receptors in addition to those located in the plasma membranes. We previously purified an auxin-binding protein (ABP) from maize shoot membranes (17). This ABP was found in the ER (18), and its amount seemed to be correlated with auxin-induced cell elongation of the coleoptile and mesocotyl (18, 19). The specificity of the binding site for various natural and synthetic auxins was roughly parallel to that in the cell elongation response (20). Information on the structure of the ABP should help in understanding its roles in the action of auxin. Therefore, in this study we cloned a cDNA encoding the ABP. The primary structure deduced from the nucleotide sequence showed the characteristics of proteins localized in the lumen of the ER, consistent with the location of the ABP in maize shoots.t MATERIALS AND METHODS Construction and Screening of a cDNA Library. Poly(A)+ RNA used for cDNA synthesis was isolated from the poly- some fraction (21) of whole shoots of 3-day-old maize seedlings (Zea mays L. cv. Golden Cross Bantam, Fujita Seed, Osaka). cDNA was synthesized according to Gubler and Hoffman (22) and inserted into the EcoRI site of plasmid pUC18 by using EcoRI linkers (5'-GGAATTCC-3'). The cDNA library was screened either with 32P-labeled oligonu- cleotide probes with a final washing with 3 M tetramethyl- ammonium chloride/50 mM Tris-HCl, pH 8.0/2 mM EDTA/0.1% NaDodSO4 at 50'C (23) or with a nick-translated cDNA insert with a final washing with 0.1 x SSC (1 x is 0.15 M NaCl/0.015 M trisodium citrate, pH 7.0) containing 1% NaDodSO4 at 650C (24). Nucleotide sequences were deter- mined by the dideoxynucleotide chain-termination method (25). Oligonucleotide probes, which were constructed on the basis of the NH2-terminal sequence of the isolated ABP, were synthesized with an Applied Biosystems model 391A DNA synthesizer. The NH2-terminal sequence was determined with an Applied Biosystems model 470A gas-phase protein sequencer. In preparation of a cDNA probe, an ABP cDNA (pABP370 as described later) was digested with EcoRI and the fragment containing the cDNA insert was isolated by NaDodSO4/PAGE and then nick-translated (24). RNA Blot Hybridization. The mRNA encoding the ABP was identified by blot hybridization of glyoxal-denatured poly(A)+ RNA that had been subjected to electrophoresis in a 1% agarose gel (24). Hybrid-Selected Translation and Immunoprecipitation. Plasmid DNA (50 ,ug) carrying ABP cDNA of 822 base pairs (bp) (pABP420 as described later) was bound to a small piece of nylon filter, and 190 ,g of poly(A)+ RNA from maize shoots was hybridized at a concentration of 1 mg/ml (26). The hybridized RNA was eluted from the filter and translated in a rabbit reticulocyte lysate (Amersham) in the presence of [35S]methionine. The products were incubated with an anti- ABP antiserum (18), and the immunocomplexes were pre- cipitated with Pansorbin (fixed Staphylococcus aureus cells, Calbiochem) (27) and analyzed by NaDodSO4/PAGE in a 12.5% polyacrylamide gel (28). Treatment of ABP with Endoglycosidase H. The purified ABP (5 ,ug) was denatured by boiling for 1 min and incubated for 16 hr at 37°C in 0.2 ml of 50 mM sodium citrate, pH 5.5/0.01% NaDodSO4/0.5 mM phenylmethylsulfonyl fluoride/50 mM 2-mercaptoethanol in the presence or ab- sence of 0.01 unit of endoglycosidase H from Streptomyces Abbreviations: ABP, auxin-binding protein; ER, endoplasmic retic- ulum. *To whom reprint requests should be addressed. tThe sequence reported in this paper is being deposited in the EMBL/GenBank data base (accession no. J04550). 3564 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of Auxin-binding protein located in the endoplasmic reticulum of maize ...

Proc. Nadl. Acad. Sci. USAVol. 86, pp. 3564-3568, May 1989Biochemistry
Auxin-binding protein located in the endoplasmic reticulum ofmaize shoots: Molecular cloning and complete primary structure
(cDNA sequence/N-glycosylation/plant hormone receptor/retention signal)
NAOHIRO INOHARA, SHOJI SHIMOMURA*, TOSHIO FUKUI, AND MASAMITSU FUTAI
Institute of Scientific and Industrial Research, Osaka University, Ibaraki, Osaka 567, Japan
Communicated by Harry Beevers, February 3, 1989
ABSTRACT We previously purified an auxin-binding pro-tein (ABP) from the microsomal fraction of maize shoots (Zeamays L. cv. Golden Cross Bantam). In the present study cDNAclones derived from mRNAs encoding the ABP were isolatedand sequenced. The nucleotide sequence of the 822-base-paircDNA includes a 603-base-pair open reading frame. RNA blothybridization analysis indicated a single mRNA species of =1.0kilobase. The predicted precursor of ABP is composed of 201amino acid residues and has a molecular weight of 21,976. TheNH2-terminal sequence of 38 residues is hydrophobic and maybe a signal peptide for translocation of the ABP across themembrane of the endoplasmic reticulum. The mature ABP,composed of 163 residues with a molecular weight of 18,352,contains a potential N-glycosylation site (Asn-Thr-Thr), andthe COOH-terminal tetrapeptide (Lys-Asp-Glu-Leu) may be asignal for retention of the ABP in the lumen of the endoplasmicreticulum.
The plant hormone auxin (indole-3-acetic acid) in higherplants functions in regulating normal growth and develop-ment, including cell elongation, division, and differentiation(1, 2). Rapid responses to auxin have been observed inorganelles such as plasma membrane (3-6), Golgi apparatus(7), endoplasmic reticulum (ER) (8), and nucleus (2, 9, 10) aswell as in the cell wall (11). The specificity of auxin is sup-posed to be due to the presence of a specific receptor(s) (12).However, it is unknown whether the diverse responses ofdifferent organelles to auxin are mediated by a single ormultiple species of receptors.Auxin can enter cells because it is a membrane-permeant,
lipophilic weak acid. In addition, specific transporters forauxin have been demonstrated in plasma membranes (13-16).Specific and saturable auxin-binding sites are also found inother organelles such as ER, tonoplast, and nucleus and inthe cytosol (12), and these should be taken into considerationas possible candidates for auxin receptors in addition to thoselocated in the plasma membranes.We previously purified an auxin-binding protein (ABP)
from maize shoot membranes (17). This ABP was found in theER (18), and its amount seemed to be correlated withauxin-induced cell elongation of the coleoptile and mesocotyl(18, 19). The specificity of the binding site for various naturaland synthetic auxins was roughly parallel to that in the cellelongation response (20). Information on the structure of theABP should help in understanding its roles in the action ofauxin. Therefore, in this study we cloned a cDNA encodingthe ABP. The primary structure deduced from the nucleotidesequence showed the characteristics of proteins localized inthe lumen of the ER, consistent with the location of the ABPin maize shoots.t
MATERIALS AND METHODSConstruction and Screening of a cDNA Library. Poly(A)+
RNA used for cDNA synthesis was isolated from the poly-some fraction (21) of whole shoots of 3-day-old maizeseedlings (Zea mays L. cv. Golden Cross Bantam, FujitaSeed, Osaka). cDNA was synthesized according to Gublerand Hoffman (22) and inserted into the EcoRI site of plasmidpUC18 by using EcoRI linkers (5'-GGAATTCC-3'). ThecDNA library was screened either with 32P-labeled oligonu-cleotide probes with a final washing with 3 M tetramethyl-ammonium chloride/50 mM Tris-HCl, pH 8.0/2 mMEDTA/0.1% NaDodSO4 at 50'C (23) or with a nick-translatedcDNA insert with a final washing with 0.1 x SSC (1x is 0.15M NaCl/0.015 M trisodium citrate, pH 7.0) containing 1%NaDodSO4 at 650C (24). Nucleotide sequences were deter-mined by the dideoxynucleotide chain-termination method(25).
Oligonucleotide probes, which were constructed on thebasis ofthe NH2-terminal sequence ofthe isolated ABP, weresynthesized with an Applied Biosystems model 391A DNAsynthesizer. The NH2-terminal sequence was determinedwith an Applied Biosystems model 470A gas-phase proteinsequencer. In preparation of a cDNA probe, an ABP cDNA(pABP370 as described later) was digested with EcoRI andthe fragment containing the cDNA insert was isolated byNaDodSO4/PAGE and then nick-translated (24).RNA Blot Hybridization. The mRNA encoding the ABP
was identified by blot hybridization of glyoxal-denaturedpoly(A)+ RNA that had been subjected to electrophoresis ina 1% agarose gel (24).
Hybrid-Selected Translation and Immunoprecipitation.Plasmid DNA (50 ,ug) carrying ABP cDNA of 822 base pairs(bp) (pABP420 as described later) was bound to a small pieceof nylon filter, and 190 ,g of poly(A)+ RNA from maizeshoots was hybridized at a concentration of 1 mg/ml (26). Thehybridized RNA was eluted from the filter and translated ina rabbit reticulocyte lysate (Amersham) in the presence of[35S]methionine. The products were incubated with an anti-ABP antiserum (18), and the immunocomplexes were pre-cipitated with Pansorbin (fixed Staphylococcus aureus cells,Calbiochem) (27) and analyzed by NaDodSO4/PAGE in a12.5% polyacrylamide gel (28).Treatment of ABP with Endoglycosidase H. The purified
ABP (5 ,ug) was denatured by boiling for 1 min and incubatedfor 16 hr at 37°C in 0.2 ml of 50 mM sodium citrate, pH5.5/0.01% NaDodSO4/0.5 mM phenylmethylsulfonylfluoride/50 mM 2-mercaptoethanol in the presence or ab-sence of 0.01 unit of endoglycosidase H from Streptomyces
Abbreviations: ABP, auxin-binding protein; ER, endoplasmic retic-ulum.*To whom reprint requests should be addressed.tThe sequence reported in this paper is being deposited in theEMBL/GenBank data base (accession no. J04550).
3564
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 86 (1989) 3565
1 2 3
AS
kb
-2.3
FIG. 1. RNA blot hybridization of ABP mRNA. Lane 1, 2 ,ug ofpoly(A)+ RNA from maize shoots; lane 2, 18 ,ug of poly(A)+ RNA;lane 3, DNA fragments of the ABP cDNA clone, obtained bydigestion with restriction endonucleases.
plicatus (Boehringer Mannheim). Glycoproteins were visu-alized by the periodic acid/Schiff(PAS) staining method (29).
RESULTSNH2-Terminal Sequence of ABP. The NH2-terminal amino
acid sequence of the ABP isolated from maize shoots wasdetermined to be Ser-Xaa-Val-Arg-Asp-Asn-Ser-Leu-Val-Arg-Asp-Ile-Ser-Gln-Met-Pro-Gln-Ser-Ser-Tyr-Gly-Ile-Glu-Gly-Leu-Ser-His-Ile-Thr-Val-Ala-. No phenylthio-hydantoin derivative was identified in the second cycle ofEdman degradation, suggesting that the residue (Xaa) atposition 2 is either a cysteine or a modified amino acidresidue.
Isolation of ABP cDNA Clones. A mixture of two sets of17-base oligonucleotides, 5'-GGCATYTGRCTDATRTC-3'and 5'-GGCATYTGNGADATRTC-3' (where Y is C or T; Ris A or G; D is A, G, or T; and N is A, C, G, or T), wassynthesized and used as a probe for screening the cDNAlibrary. These oligonucleotides are complementary to the 72possible coding sequences for amino acid residues 11-16 ofthe ABP. A clone carrying a cDNA insert of 454 bp(pABP370) was isolated from 100,000 transformants. As thisinsert was shorter than the predicted ABP mRNA [--1.0kilobase (kb); Fig. 1], we used it as a probe for screening thelibrary to isolate the corresponding full-length cDNA. Eightclones carrying cDNA inserts of 388-822 bp were obtainedand sequenced.
Nucleotide Sequence of ABP cDNA. Fig. 2 shows thenucleotide sequence of the cDNA clone (pABP420) contain-ing the longest insert (822 bp). The sequence at positions 191-207 was complementary to a sequence of the oligonucleotideprobes. This clone also contained a poly(A) tract with twopotential polyadenylylation signals (AATAAA; ref. 31) 22and 72 bases upstream of the tract. Slight differences in thesequence were observed in the overlapping regions of thenine clones (Fig. 2). The nucleotides at positions 82, 83, 325,468, and 469 were T, C, A, G, and T, respectively, in twoclones (including pABP420), while in other clones thesenucleotides were T, C, A, A, and T (three clones), T, G, G,A, and C (one clone), or G, G, A, A, and T (two clones). Thechanges of the nucleotides at positions 83 (C -> G) and 468(G-> A) resulted in changes of the codons leading to differentamino acid residues (Pro 26-> Ala and Ser103 -+ Asn; Fig. 2).The changes at nucleotides 82 and 325 did not alter theencoded amino acids. One cDNA clone (pABP370) lackedthe sequence at positions 281-298. The sequence surroundingthe boundary of this deleted sequence had similar character-istics to an exon/intron boundary sequence (32). The de-duced amino acid sequence of this clone lacked 6 residues asa consequence of the deletion, but the deletion did not alterthe coding frame. The reason for the heterogeneity in thecDNA clones is unknown. A possibility is that there is a
ATCTCATTCCACTCCGACATTCACGTGCAGCTGTCGGGAGCAGGCA ATG GCG CCG GAT CTA AGC GAA CTC 70Met Ala Pro Asp Leu Ser Glu Leu -31
G GGCC GCCC GCG GCT CCA GCC CGT GGC GCC TAC CTC GCC GGC GTC GGT GTC GCG GTC CTC CTC GCT GCC TCC TTC CTC CCA GTA GCC GAG TCG 160Ala Ala Ala Ala Pro Ala Arg Gly Ala Tyr Leu Ala Gly Val Gly Val Ala Val Leu Leu Ala Ala Ser Phe Leu Pro Val Ala Glu Ser -1
Ala
TCC TGC GTG CGA GAT AAC TCA TTG GTG AGA GAC ATA AGC CAA ATG CCG CAA AGC AGC TAT GGG ATT GAA GGA TTG TCA CAT ATA ACA GTT 250Ser Cys Val Arg Asp Asn Ser Leu Val Arg Asp Ile Ser Gln Met Pro Gln Ser Ser Tyr Gly Ile Glu Gly Leu Ser His Ile Thr Val 30
CGCT GGT GCG CTC AAT CAT GGG ATG AAG GAG GTG GM GTG TGG CTT CAG ACA ATA AGT CCA GGT CAA AGG ACG CCA ATC CAC AGG CAT TCC 340Ala Gly Ala Leu Asn His Gly Met Lys Glu Val Glu Val Trp Leu Gln Thr Ile Ser Pro Gly Gln Arg Thr Pro Ile His Arg His Ser 60
TGT GAA GAA GTT TTC ACT GTC CTC AAA GGG AAG GGT ACG CTC TTG ATG GGA TCA AGC TCA CTA AAG TAC CCA GGG CAG CCA CAG GAA ATT 430Cys Glu Glu Val Phe Thr Val Leu Lys Gly Lys Gly Thr Leu Leu Met Gly Ser Ser Ser Leu Lys Tyr Pro Gly Gln Pro Gln Glu Ile 90
ACCCT TTC TTT CAG AAT ACC ACA TTT TCA ATC CCT GTA AGT GAT CCA CAC CAG GTT TGG AAT TCT GAC GAG CAC GAA GAT TTG CAA GTT CTT 520Pro Phe Phe Gln Asn Thr Thr Phe Ser Ile Pro Val Ser Asp Pro His Gln Val Trp Asn Ser Asp Glu His Glu Asp Leu Gln Val Leu 120
Asn
GTG ATC ATA TCG AGA CCG CCT GCT AAG ATA TTT TTA TAT GAT GAT TGG AGC ATG CCT CAT ACA GCC GCG GTA CTG AAA TTC CCC TTC GTC 610Val Ile Ile Ser Arg Pro Pro Ala Lys Ile Phe Leu Tyr Asp Asp Trp Ser Met Pro His Thr Ala Ala Vai Leu Lys Phe Pro Phe Vai 150
TGG GAT GAG GAC TGC TTC GAA GCA GCA AAA GAGAA CTC TAG GTCACAAGTGTTTCCTGCAATTTATCTGCTTCATCCATGATCCTGCTGGTGCTGGACTACTAC 715Trp Asp Giu Asp Cys Phe G1U Ala Ala Lys Asp Giu Leu 163
AATTCTCAGCACTAGTTGTAATAAAGCCAGTGCGCTTTTCATGTATAATTCTGTA.TTGTGGCTCGCGAAAATAAAATTTGGCAACGGTTTATGAACTAAAAAAA 822
FIG. 2. Nucleotide sequence and deduced amino acid sequence of the ABP cDNA in the cDNA clone (pABP420) carrying the longest insert.The different nucleotides and amino acids at various positions in nine cDNA clones are shown. Numbering of the amino acid sequence startsat the NH2-terminal residue of the ABP isolated from maize shoots. The amino acid sequence determined by protein sequencing is underlined.Two polyadenylylation signals and a poly(A) tract are also underlined. Asterisks indicate the potential N-glycosylation site. The consensussequence necessary for retention in the ER lumen (30) is marked by double underlining.
Biochemistry: Inohara et al.
3566 Biochemistry: Inohara et al.
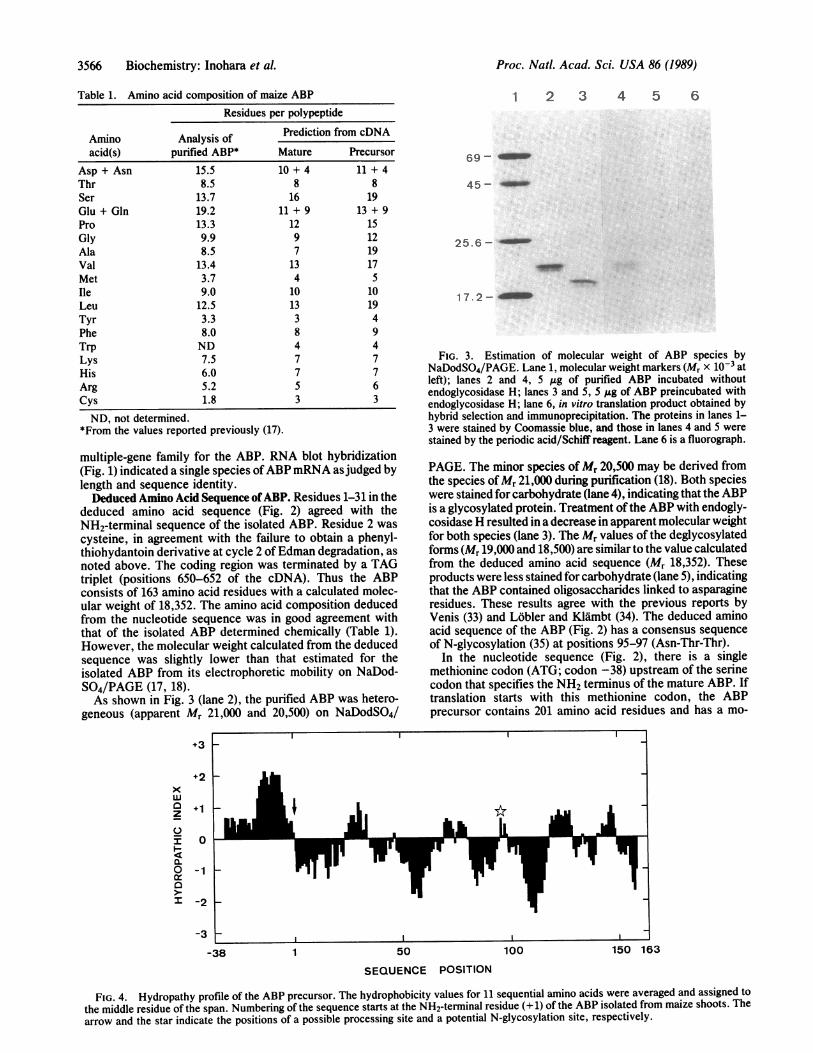
Table 1. Amino acid composition of maize ABP
Residues per polypeptide
Amino Analysis of Prediction from cDNAacid(s) purified ABP* Mature Precursor
Asp + Asn 15.5 10 + 4 11 + 4Thr 8.5 8 8Ser 13.7 16 19Glu + Gln 19.2 11 + 9 13 + 9Pro 13.3 12 15Gly 9.9 9 12Ala 8.5 7 19Val 13.4 13 17Met 3.7 4 5Ile 9.0 10 10Leu 12.5 13 19Tyr 3.3 3 4Phe 8.0 8 9Trp ND 4 4Lys 7.5 7 7His 6.0 7 7Arg 5.2 5 6Cys 1.8 3 3
ND, not determined.*From the values reported previously (17).
multiple-gene family for the ABP. RNA blot hybridization(Fig. 1) indicated a single species ofABP mRNA asjudged bylength and sequence identity.Deduced Amino Acid Sequence ofABP. Residues 1-31 in the
deduced amino acid sequence (Fig. 2) agreed with theNH2-terminal sequence of the isolated ABP. Residue 2 wascysteine, in agreement with the failure to obtain a phenyl-thiohydantoin derivative at cycle 2 ofEdman degradation, asnoted above. The coding region was terminated by a TAGtriplet (positions 650-652 of the cDNA). Thus the ABPconsists of 163 amino acid residues with a calculated molec-ular weight of 18,352. The amino acid composition deducedfrom the nucleotide sequence was in good agreement withthat of the isolated ABP determined chemically (Table 1).However, the molecular weight calculated from the deducedsequence was slightly lower than that estimated for theisolated ABP from its electrophoretic mobility on NaDod-S04/PAGE (17, 18).As shown in Fig. 3 (lane 2), the purified ABP was hetero-
geneous (apparent Mr 21,000 and 20,500) on NaDodSO4/
1 2 3 4 5 6
69
45--
25.6--
17.2-
FIG. 3. Estimation of molecular weight of ABP species byNaDodSO4/PAGE. Lane 1, molecular weight markers (Mr x 10-3 atleft); lanes 2 and 4, 5 ,ug of purified ABP incubated withoutendoglycosidase H; lanes 3 and 5, 5 ug of ABP preincubated withendoglycosidase H; lane 6, in vitro translation product obtained byhybrid selection and immunoprecipitation. The proteins in lanes 1-3 were stained by Coomassie blue, and those in lanes 4 and 5 werestained by the periodic acid/Schiff reagent. Lane 6 is a fluorograph.
PAGE. The minor species ofMr 20,500 may be derived fromthe species ofMr 21,000 during purification (18). Both specieswere stained for carbohydrate (lane 4), indicating that the ABPis a glycosylated protein. Treatment of the ABP with endogly-cosidase H resulted in a decrease in apparent molecular weightfor both species (lane 3). The Mr values of the deglycosylatedforms (Mr 19,000 and 18,500) are similar to the value calculatedfrom the deduced amino acid sequence (Mr 18,352). Theseproducts were less stained for carbohydrate (lane 5), indicatingthat the ABP contained oligosaccharides linked to asparagineresidues. These results agree with the previous reports byVenis (33) and Lobler and Klambt (34). The deduced aminoacid sequence of the ABP (Fig. 2) has a consensus sequenceof N-glycosylation (35) at positions 95-97 (Asn-Thr-Thr).
In the nucleotide sequence (Fig. 2), there is a singlemethionine codon (ATG; codon -38) upstream of the serinecodon that specifies the NH2 terminus of the mature ABP. Iftranslation starts with this methionine codon, the ABPprecursor contains 201 amino acid residues and has a mo-
+3
xw0zC-
t-0Ix
+1
0
-1
-2
-3 8
-38 1 50 100 150 163
SEQUENCE POSITION
FIG. 4. Hydropathy profile of the ABP precursor. The hydrophobicity values for 11 sequential amino acids were averaged and assigned tothe middle residue of the span. Numbering of the sequence starts at the NH2-terminal residue (+ 1) of the ABP isolated from maize shoots. Thearrow and the star indicate the positions of a possible processing site and a potential N-glycosylation site, respectively.
Proc. Natl. Acad. Sci. USA 86 (1989)

Proc. Natl. Acad. Sci. USA 86 (1989) 3567
lecular weight of 21,976, consistent with the observed value(Mr 22,000) of the in vitro translation product isolated byhybrid selection and immunoprecipitation (Fig. 3, lane 6).There is a similarity between the consensus sequence sur-rounding the initiator ATG codon in plant genes (31) and thecorresponding sequence of the ABP cDNA. In particular, 10nucleotides surrounding the first ATG codon of the ABP areexactly the same as those of maize alcohol dehydrogenase 1(31).Hydropathy Analysis of the Deduced Amino Acid Sequence.
Fig. 4 shows the result of analysis of the hydropathy of theABP precursor by the procedure of Kyte and Doolittle (36).The possible processing site between residue -1 and 1 islocated at a break point between regions of low and highhydrophobicity. N-glycosylation at residue 95 may producea long sequence exhibiting a high degree of hydrophilicity.The mature protein region is relatively hydrophilic andexhibits no hydrophobic sequence long enough to span themembrane (36). In contrast, the extended sequence of 38amino acid residues upstream of the NH2 terminus of themature protein is relatively hydrophobic.
DISCUSSIONIn this study we have cloned a cDNA encoding a maize ABP.RNA blot hybridization indicated a single mRNA species of-1.0 kb. The 822-bp cDNA clone may, therefore, containnearly the full length of the sequence for ABP mRNA exceptfor the sequence corresponding to the poly(A) tail of 150-200nucleotides, The predicted precursor for the ABP is composedof 201 amino acid residues. The NH2-terminal 38 residues maybe a signal peptide for membrane transport, since this se-quence contains a stretch of 20 hydrophobic residues (posi-tions -22 to -3) surrounded by two charged residues, char-acteristics of the hydrophobic core of a signal peptide (37).The ABP was found in the ER-rich fraction on sucrose
density gradient centrifugation of tissue homogenates (18),and its distribution was distinguishable from those of markerenzymes of plasma membranes, tonoplasts, dictyosomes,mitochondria, and proplastids (38-40). The ABP was solu-bilized without using detergent (41), suggesting that it is notan integral membrane protein. Hydropathy analysis of theamino acid sequence (Fig. 4) is consistent with this possibil-ity. As N-glycosylation is carried out solely on the luminalside of the ER (35), the ABP is proposed to be a proteinlocated in the ER lumen.The proteins located in the ER lumen of animal cells
possess a common sequence (Lys-Asp-Glu-Leu) at theirCOOH termini (30). Maize ABP contains the same sequenceat its COOH terminus (Fig. 2). This sequence is thought to bea signal for retaining soluble proteins in the ER lumen (30,42). The ABP sequence has an Asp/Glu cluster (Asp-Glu-Asp-Xaa-Xaa-Glu; Fig. 2) near this signal. Animal pro-teins in the ER lumen also possess an Asp/Glu cluster in theirCOOH-terminal region (30). This negatively charged regionmay function to expose the COOH-terminal signal on theprotein surface. The mechanism by which ER proteins aresorted in plant cells is unknown. However, the presentresults suggest that the same sorting system as in animal cellsmay operate in plant cells. A similar sequence for ERretention has also been postulated for yeast (43).
Lobler and Klambt (44) reported that anti-ABP antibodies,purified by affinity chromatography using ABP-linked Seph-arose, inhibited auxin-mediated coleoptile elongation. Thisresult suggests that the ABP is located and functions in thecell wall or on the plasma membrane facing the cell wall; thisis inconsistent with our conclusion concerning the cellularlocalization. A possibility is that the ABP found in the ER isa precursor that will be secreted into the external face of theplasma membrane. The ABP of Mr 21,000 was converted to
the Mr 20,500 protein during isolation (18). Since this de-crease in molecular weight could be due to removal of theCOOH-terminal ER-retention signal, the above possibilitycannot be ruled out.The ABP shows no significant homology with any of the
proteins in the National Biomedical Research Foundation'sprotein data bank,t including two auxin-regulated gene prod-ucts (Aux28 and Aux22) from soybean (45). The presentfindings provide a basis for comparative studies on proteinspossibly related to auxin functions and for studies on themechanism of auxin action.
tProtein Identification Resource (1988) Protein Sequence Database(Natl. Biomed. Res. Found., Washington, DC), Release 16.0.
We thank Professor Hiroyuki Kagamiyama and Dr. SeikiKuramitsu (Osaka Medical College) for NH2-terminal sequenceanalysis of maize ABP. We also thank Dr. Takato Noumi (OsakaUniversity) for expert advice on cDNA cloning and sequencing andMr. Wei Liu for technical assistance. This study was supported inpart by a Grant-in-Aid for Scientific Research from the Ministry ofEducation, Science, and Culture of Japan and by a Special Coordi-nation Fund for Promotion of Science and Technology from theScience and Technology Agency of the Japanese Government.
1. Evans, M. L. (1985) CRC Crit. Rev. Plant Sci. 2, 317-365.2. Guilfoyle, T. J. (1986) CRC Crit. Rev. Plant Sci. 4, 247-276.3. Gabathuler, R. & Cleland, R. E. (1985) Plant Physiol. 79, 1080-
1085.4. Brightman, A. O., Barr, R., Crane, F. L. & Morrd, D. J. (1988)
Plant Physiol. 86, 1264-1269.5. Morr6,D. J., Gripshover, B., Monroe, A.&Morrd, J. T. (1984)
J. Biol. Chem. 259, 15364-15368.6. Ettlinger, C. & Lehle, L. (1988) Nature (London) 331, 176-178.7. Ray, P. M. (1985) Plant Physiol. 78, 466-472.8. Buckhout, T. J., Young, K. A., Low, P. S. & Morrd, D. J.
(1981) Plant Physiol. 68, 512-515.9. Key, J. L., Kroner, P., Walker, J., Hong, J. C., Ulrich, T. H.,
Ainley, W. M., Gantt, J. S. & Nagao, R. T. (1986) Phil. Trans.R. Soc. London Ser. B. 314, 427-440.
10. Theologis, A. (1986) Annu. Rev. Plant Physiol. 37, 407-438.11. Kutschera, U. & Briggs, W. R. (1987) Proc. Natl. Acad. Sci.
USA 84, 2747-2751.12. Rubery, P. H. (1981) Annu. Rev. Plant Physiol. 32, 569-596.13. Hertel, R., Lomax, T. L. & Briggs, W. R. (1983) Planta 157,
193-201.14. Lomax, T. L., Mehlhorn, R. J. & Briggs, W. R. (1985) Proc.
Natl. Acad. Sci. USA 82, 6541-6545.15. Benning, C. (1986) Planta 169, 228-237.16. Jacobs, M. & Gilbert, S. F. (1983) Science 220, 1297-1300.17. Shimomura, S., Sotobayashi, T., Futai, M. & Fukui, T. (1986)
J. Biochem. 9, 1513-1524.18. Shimomura, S., Inohara, N., Fukui, T. & Futai, M. (1988)
Planta 175, 558-566.19. Walton, J. D. & Ray, P. M. (1981) Plant Physiol. 68, 1334-
1338.20. Ray, P. M., Dohrmann, U. & Hertel, R. (1977) Plant Physiol.
60, 585-591.21. Christoffersen, R. E. & Laties, G. G. (1982) Proc. Natl. Acad.
Sci. USA 79, 4060-4063.22. Gubler, U. & Hoffman, B. J. (1983) Gene 25, 263-269.23. Wood, W. I., Gitschier, J., Lasky, L. A. & Lawn, R. M. (1985)
Proc. NatI. Acad. Sci. USA 82, 1585-1588.24. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular
Cloning: A Laboratory Manual (Cold Spring Harbor Lab., ColdSpring Harbor, NY).
25. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl.Acad. Sci. USA 74, 5463-5467.
26. Jagus, R. (1987) Methods Enzymol. 152, 567-572.27. Goding, J. W. (1984) in Molecular and Chemical Character-
ization ofMembrane Receptors, eds. Venter, J. C. & Harrison,L. C. (Liss, New York), pp. 31-60.
28. Laemmli, U. K. (1970) Nature (London) 227, 680-685.
Biochemistry: Inohara et al.

3568 Biochemistry: Inohara et al.
29. Zacharius, R. M., Zell, T. E., Morrison, J. H. & Woodlock,J. J. (1969) Anal. Biochem. 30, 148-152.
30. Munro, S. & Pelham, H. R. B. (1987) Cell 48, 899-907.31. Heidecker, G. & Messing, J. (1986) Annu. Rev. Plant Physiol.
37, 439-466.32. Lewin, B. (1980) Cell 22, 324-326.33. Venis, M. A. (1987) NATOAdv. Stud. Inst. Ser., Ser. H 10, 27-39.34. Lobler, M. & Klambt, D. (1987) NATO Adv. Stud. Inst. Ser.,
Ser. H 10, 41-49.35. Kornfeld, R. & Kornfeld, S. (1985) Annu. Rev. Biochem. 54,
631-664.36. Kyte, J. & Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132.
Proc. Nail. Acad. Sci. USA 86 (1989)
37. Van Heijne, G. (1985) J. Mol. Biol. 184, 99-105.38. Dohrmann, U., Hertel, R. & Kowalik, H. (1978) Planta 140,97-
106.39. Jacobs, M. & Hertel, R. (1978) Planta 142, 1-10.40. Ray, P. M. (1977) Plant Physiol. 59, 594-599.41. Venis, M. A. (1977) Nature (London) 266, 268-269.42. Pelham, H. R. B. (1988) EMBO J. 7, 913-918.43. Pelham, H. R. B., Hardwick, K. G. & Lewis, M. J. (1988)
EMBO J. 7, 1757-1762.44. Lobler, M. & Klambt, D. (1985) J. Biol. Chem. 260, 9854-9859.45. Ainley, W. M., Walker, J. C., Nagao, R. T. & Key, J. L.
(1988) J. Biol. Chem. 263, 10658-10666.