
Autonomic Nervous... · Web viewسلول های عصبی در این هسته دارای پی...
53
The Autonomic Nervous System and the Hypothalamus Susan Iversen Leslie Iversen Clifford B. Saper WHEN WE ARE FRIGHTENED our heart races, our breathing becomes rapid and shallow, our mouth becomes dry, our muscles tense, our palms become sweaty, and we may want to run. These bodily changes are mediated by the autonomic nervous system, which controls heart muscle, smooth muscle, and exocrine glands. The autonomic nervous system is distinct from the somatic nervous system, which controls skeletal muscle. As we shall learn in the next chapter, even though the neural control of emotion involves several regions, including the amygdala and the limbic association areas of the cerebral cortex, they all work through the hypothalamus to control the autonomic nervous system. The hypothalamus coordinates behavioral response to insure bodily homeostasis, the constancy of the internal environment. The hypothalamus, in turn, acts on three major systems: the autonomic nervous system, the endocrine system, and an ill-defined neural system concerned with motivation. In this chapter we shall first examine the autonomic nervous system and then go on to consider the hypothalamus. In the next two chapters, we shall examine emotion and motivation, behavioral states that depend greatly on autonomic and hypothalamic mechanisms. P.961 وس ملا ا وت پ هیدکار و و خ ی صب ع م ت س سی# ون س ور ی ا# ان ور س# ون س ور ی ی ا سل ل ورد ف ی کلر پ ی. سا ب ف5 ک، # ان5 م ا ر5 م لات 5 ض ع ود،< 5 س ی م> ک< 5 س خ ا5 م# ان5 ود ، ده< 5 س ی م ق5 م ع م ک ع و ی ر5 س س ف یL ت، ب ل ق ا5 ادهP ر پ ده ر ت< 5 س خ ا و5 ه م5 ی ک م ا5 گ ن ه له 5 ض ع ه5 ار ، ک5 ودک خ ی ب5 ص ع م ت5 س سی ط5 وس ی# 5 دن 5 ت رات ی 5 ی غ ت# ن ی ود. ا< 5 س را 5 ج ه ا 5 د ب 5 ی ه وا خ ب ت5 س ا# ن مک م ا5 د و م وت< 5 س ی م رق5 ع ا5 م ت5 س د ه5 ، ک> ک ی ت وما5 س ی ب5 ص ع م ت5 س سی ا ار ر 5 ج م ار5 ودک خ ی ب5 ص ع م ت5 س سی طه .5 واس ا 5 رل ت یt 5 ی ک ی ج ار 5 حه خ< 5 ش ر مید د 5 ، و غ اف5 ص لات 5 ض ع، ب ل ق ل م ا< 5 س ات5 اس س خ ا ی ار ب5 ص ع رل یt 5 ی کر5 گ ی ا ب5 خ د ، 5 رت گی ی م اد 5 د ت5 غ تل5 ض ف در د 5 ات ا ت5 ه م5 ور ک ط ن ا5 م ه د. 5 ی ک ی م رل یt 5 ی ک ی را کلب5 س ا لات 5 ض ع رل یt 5 ی ک رای 5 پ ار5 وس ک ملا ا 5 وت پ هی ق 5 ی ر ط را ار ا5 ه ن ه ا5 م ه، خ م ر< 5 ش ق ار> ک 5 ی می ی ل اط 5 ی ت ار ق ط ا 5 ی ملا ودا5 گ ی م ه ا5 ل م ج ، ار 5 ف5 ل ت خ م ق ط ا 5 ی م. 5 ی ل خ ط دا 5 ی خ م# ودن 5 ی ب ب ا< 5 ، ت# دن 5 ت ار ی س و م ه م ی و< س# ن¢ مئ ط م ا اری ، ت ت ق ر خ ش ا ت ضاتL ی خ م وس ملا ا وت پ هی. ت سر ا ودکا خ ی صب ع م ت س سی ود، در 5 خ ه 5 وب ی ه 5 وس ، ب ملا ا 5 وت پ هی ه : ر ی5 گ ت ا ا 5 ه ت5 ط در ران ف 5 ی ر ع ت ی ب و 5 خ ه ی ب صب ع م ت س سی و ر پ ر# د درون د م غ ت س سی ی ، صب ع ودکار خ م ت س سی# ن ت ق ر5 گ ر‹ 5 ظ ن ه در 5 ب# ن ا س ار پ ار و5 ودک خ ی ب5 ص ع م ت5 س سی ی5 سرر پ د 5 ات دا ت 5 ی ت ا ا5 ل م5 ض ف# ن ی. در ا د 5 ی ک ی م ل5 م ع ی ل5 ص م ا ت5 س سی ه5 س
Transcript of Autonomic Nervous... · Web viewسلول های عصبی در این هسته دارای پی...

The Autonomic Nervous System and the HypothalamusSusan IversenLeslie IversenClifford B. Saper
WHEN WE ARE FRIGHTENED our heart races, our breathing becomes rapid and shallow, our mouth becomes dry, our muscles tense, our palms become sweaty, and we may want to run. These bodily changes are mediated by the autonomic nervous system, which controls heart muscle, smooth muscle, and exocrine glands. The autonomic nervous system is distinct from the somatic nervous system, which controls skeletal muscle. As we shall learn in the next chapter, even though the neural control of emotion involves several regions, including the amygdala and the limbic association areas of the cerebral cortex, they all work through the hypothalamus to control the autonomic nervous system. The hypothalamus coordinates behavioral response to insure bodily homeostasis, the constancy of the internal environment. The hypothalamus, in turn, acts on three major systems: the autonomic nervous system, the endocrine system, and an ill-defined neural system concerned with motivation. In this chapter we shall first examine the autonomic nervous system and then go on to consider the hypothalamus. In the next two chapters, we shall examine emotion and motivation, behavioral states that depend greatly on autonomic and hypothalamic mechanisms.P.961
هیپوتاالموس و خودکار عصبی سیستمایورسون سوزان
ایورسون لسلیساپر. کلیفورد بی
می خشک ما دهان ، شود می عمق کم و سریع تنفس قلب، نژادها زده وحشت ما که هنگامی . این شود اجرا به بخواهید است ممکن ما و شوند می عرق ما دست کف ، زمان ما عضالت شود،
مترشحه غدد و ، صاف عضالت ، قلب عضله که ، خودکار عصبی سیستم توسط بدن تغییراتعضالت . که ، سوماتیک عصبی سیستم از مجزا خودکار عصبی سیستم واسطه با کنترل خارجی
. عصبی کنترل اگر حتی ، گیرند می یاد بعد فصل در باید ما که همانطور کند می کنترل را اسکلتیهمه ، مخ قشر از لیمبیک ارتباط مناطق و آمیگداال جمله از ، مختلف مناطق شامل احساسات از
. هیپوتاالموس است خودکار عصبی سیستم کنترل برای کار هیپوتاالموس طریق از را آنها . سیستم داخلی محیط بودن ثابت بدن، هموستاز شویم مطمئن تا ، رفتاری پاسخ مختصات
: انگیزه با رابطه در تعریف خوبی به عصبی سیستم و ریز درون غدد سیستم ، عصبی خودکار . بررسی باید ابتدا ما فصل این در کند می عمل اصلی سیستم سه در خود، نوبه به ، هیپوتاالموس
باید . ما ، بعدی فصل دو در هیپوتاالموس گرفتن نظر در به آن از پس و خودکار عصبی سیستمو خودکار مکانیسم در دارد بستگی زیادی حد تا که رفتاری حاالت ، انگیزه و احساسات
. کند بررسی را هیپوتاالموسP.961
The Autonomic Nervous System Is a Visceral and Largely Involuntary Sensory and Motor System
In contrast to the somatic sensory and motor systems, which we considered in Parts IV and V of this book, the autonomic nervous system is a visceral sensory and motor system. Virtually all visceral reflexes are mediated by local circuits in the brain stem or spinal cord. Although these reflexes are regulated by a network of central autonomic control nuclei in the brain stem, hypothalamus, and forebrain, these visceral reflexes are not under voluntary control, nor do they impinge on consciousness, with few exceptions. The autonomic nervous system is thus also referred to as the involuntary motor system, in contrast to the voluntary (somatic) motor system.
غیرارادی عمدتا و احشایی حرکتی و حسی سیستم یک که است خودکار عصبی سیستم

قطعات در را آن ما که سیستم، حرکتی و حسی جسمی به مقابل عصبی Vو IVدر سیستم شده، گرفته نظر در کتاب این از . نخاع یا و مغز ساقه در محلی مدارات توسط احشایی های واکنش تمام تقریبا است احشایی حرکتی و حسی سیستم خودکار
می. تنظیم مغز و هیپوتاالموس، مغز، ساقه در مرکزی خودکار کنترل های هسته از ای شبکه توسط رفلکس این چه اگر است . در که است خودکار عصبی سیستم استثنا چند با تجاوز، آگاهی در آنها نه و نیست، ارادی کنترل تحت احشایی رفلکس این شود،
. ) موتور ) سیستم سوماتیک داوطلبانه مقابل در گویند، می ارادی غیر سیستم موتور عنوان به نیز نتیجه
The autonomic nervous system has three major divisions: sympathetic, parasympathetic, and enteric. The sympathetic and parasympathetic divisions innervate cardiac muscle, smooth muscle, and glandular tissues and mediate a variety of visceral reflexes. These two divisions include the sensory neurons associated with spinal and cranial nerves, the preganglionic and postganglionic motor neurons, and the central nervous system circuitry that connects with and modulates the sensory and motor neurons. The enteric division has greater autonomy than the other two divisions and comprises a largely self-contained system, with only minimal connections to the rest of the nervous system. It consists of sensory and motor neurons in the gastrointestinal tract that mediate digestive reflexes.
. : عضله کردن پی دارای پاراسمپاتیک و سمپاتیک بخش روده و پاراسمپاتیک، دلسوز، عمده بخش سه دارای خودکار عصبی سیستم . با ارتباط در حسی نورونهای از عبارتند بخش دو این احشایی رفلکس انواع واسطه و غددی های بافت و صاف، عضالت قلب،
حرکتی نورونهای مغزی، و نخاعی با postganglionicو preganglionicاعصاب که است مرکزی عصبی سیستم مدارات و ، . حد تا سیستم یک شامل و دیگر بخش دو از بیشتر استقالل است روده تقسیم حرکتی و حسی نورونهای مدوله و شده متصل
. است گوارش دستگاه در حرکتی و حسی عصبی های سلول این عصبی سیستم بقیه به حداقل به اتصال با تنها شامل، خود زیادی. است شده تشکیل گوارشی های رفلکس میانجیگری که
The American physiologist Walter B. Cannon first proposed that the sympathetic and parasympathetic divisions have distinctly different functions. He argued that the parasympathetic nervous system is responsible for rest and digest, maintaining basal heart rate, respiration, and metabolism under normal conditions. The sympathetic nervous system, on the other hand, governs the emergency reaction, or fight-or-flight reaction. In an emergency the body needs to respond to sudden changes in the external or internal environment, be it emotional stress, combat, athletic competition, severe change in temperature, or blood loss. For a person to respond effectively, the sympathetic nervous system increases output to the heart and other viscera, the peripheral vasculature and sweat glands, and the piloerector and certain ocular muscles. An animal whose sympathetic nervous system has been experimentally eliminated can only survive if sheltered, kept warm, and not exposed to stress or emotional stimuli. Such an animal cannot, however, carry out strenuous work or fend for itself; it cannot mobilize blood sugar from the liver quickly and does not react to cold with normal vasoconstriction or elevation of body heat.
والتر آمریکایی کرد. . Bفیزیولوژیست استدالل او متفاوت توابع پاراسمپاتیک و سمپاتیک بخش که کرد پیشنهاد بار اولین برای کاننشرایط در بدن ساز و سوخت و تنفس قلب، پایه نرخ حفظ ، است هضم و استراحت برای مسئول پاراسمپاتیک عصبی سیستم که
اضطراری . . مواقع در گریز و جنگ واکنش یا و ، اضطراری های واکنش بر حاکم ، دیگر سوی از ، سمپاتیک عصبی سیستم عادیرقابت ، با مبارزه عاطفی، استرس را آن شود می داخلی، یا و خارجی های محیط در ناگهانی تغییرات به پاسخگویی به نیاز بدن که
. و قلب به سمپاتیک عصبی سیستم ، پاسخگویی در فرد یک برای است خون دادن دست از یا دما، در شدید تغییرات ورزشی، هایو عرق، غدد و محیطی احشا،عروق و امعا که . piloerectorسایر حیوانی خروجی دهد می افزایش را چشمی عضالت از برخی و
یا و استرس به و گرم، شود می نگهداری ، پناه اگر بمانند زنده توانند می تنها حذف تجربی است شده عصبی سیستم دلسوز . قند تواند می را آن ، خود از دفاع یا و شدید کار انجام ، حال این با ، توانند نمی حیوانات چنین ندارند قرار ، احساسی های محرک
. دهند نمی نشان واکنش بدن حرارت رفتن باال یا طبیعی عروق انقباض با سرما به و سرعت به کبد از نه بسیج را خون

Figure 49-1 Anatomical organization of the somatic and autonomic motor pathways.A. In the somatic motor system, effector motor neurons in the central nervous system project directly to skeletal muscles.B. In the autonomic motor system, the effector motor neurons are located in ganglia outside the central nervous system and are controlled bypreganglionic central neurons.
.1-49شکل خودمختار حرکتی مسیرهای و جسمی از تشریحی سازمانA. اسکلتی. عضالت به مستقیم طور به مرکزی عصبی سیستم پروژه در موثر حرکتی نورونهای حرکتی، جسمی سیستم درB .های سلول توسط و دارند قرار مرکزی اعصاب سیستم از خارج در نخاع، در حرکتی نورونهای موثر خودکار، موتور سیستم در
مرکزی .preganglionicعصبی شود می کنترل
The relationship between the sympathetic and parasympathetic pathways is not as simple and as independent as suggested by Cannon, however. Both divisions are tonically active and operate in conjunction with each other and with the somatic motor system to regulate most behavior, be it normal or emergency. Although several visceral functions are controlled predominantly by one or the other division, and although both the sympathetic and parasympathetic divisions often exert opposing effects on innervated target tissues, it is the balance of activity between the two that helps maintain an internal stable environment in the face of changing external conditions.
. عارضه از بخش دو هر حال این با پیشنهاد، کانن توسط که مستقل و ساده بسیار است، پاراسمپاتیک و سمپاتیک مسیر بین رابطهعادی را آن شود می کنند، می عمل رفتار ترین تنظیم برای حرکتی جسمی سیستم با و یکدیگر با ارتباط در و هستند فعال مانند ها . بخش دو هر چه اگر و شود می کنترل دیگر های بخش یا یک توسط عمدتا احشایی توابع چندین چه اگر است اضطراری یا و
هدف بافت بر متضاد اثرات اغلب پاراسمپاتیک و کمک innervatedسمپاتیک که است دو این بین های فعالیت از تعادل آن اعمال،. خارجی شرایط تغییر است صورت در ثبات با داخلی محیط حفظ تا کند می
The idea of a stable internal environment in the face of changing external conditions was first proposed in the nineteenth century by the French physiologist Claude Bernard. This idea was developed further by Cannon, who put forward the concept of homeostasis as the complex

P.962فرانسوی فیزیولوژیست توسط نوزدهم قرن در بار اولین برای خارجی شرایط تغییر مقابل در ثبات با داخلی محیط یک از ایده این
. شد داده توسعه بیشتر پیچیده عنوان به هموستاز مفهوم جلو به را خود که کانن، توسط ایده این شد پیشنهاد برنارد کلودP.962physiological mechanisms that maintain the internal milieu. In his classic book The Wisdom of the Body published in 1932, Cannon introduced the concept of negative feedback regulation as a key homeostatic mechanism and outlined much of our current understanding of the functions of the autonomic nervous system.
. سال در بدن از عقل خود کالسیک کتاب در است داخلی محیط حفظ که است فیزیولوژیکی های شده، 1932مکانیسم منتشرسیستم عملکرد از ما فعلی درک از خیلی و متعادل کلیدی مکانیسم یک عنوان به منفی بازخورد مقررات مفهوم معرفی کانن
. است شده خالصه خودکار عصبی
Figure 49-2 Anatomical organization of the sympathetic preganglionic and postganglionic axons. (Adapted from Loewy and Spyer 1990.)
آکسون 2-49شکل از تشریحی از. ) postganglionic و preganglionic سازمان اقتباس .(Spyer 1990 و Loewy دلسوز
If a state remains steady, it does so because any change is automatically met by increased effectiveness of the factor or factors that resist the change. Consider, for example, thirst when the body lacks water; the discharge of adrenaline, which liberates sugar from the liver when the concentration of sugar in the blood falls below a critical point; and increased breathing, which reduces carbonic acid when the blood tends to shift toward acidity.
که است عواملی یا عامل بخشی اثر افزایش خودکار طور به تغییر گونه هر خاطر به را کار این ماند، می باقی پایدار حالت یک اگر . آزاد که آدرنالین، از ترشح آب، فاقد بدن که زمانی تشنگی مثال، عنوان به بگیرید، نظر در کرد مالقات را تغییر برابر در مقاومت
وقتی که کربن اکسید دی دهد می کاهش را تنفس، افزایش و بحرانی نقطه یک از کمتر خون در قند غلظت که زمانی کبد از قنداسیدیته سمت به تمایل .خون
Cannon further proposed that the autonomic nervous system, under the control of the hypothalamus, is an important part of this feedback regulation. The hypothalamus regulates many of the neural circuits that mediate the peripheral components of

emotional states: changes in heart rate, blood pressure, temperature, and water and food intake. It also controls the pituitary gland and thereby regulates the endocrine system.
. در تغییر است بازخورد مقررات این از مهمی بخش هیپوتاالموس، کنترل تحت خودکار، عصبی سیستم که کرد پیشنهاد بیشتر کانن : اجزای واسطه که است عصبی مدارهای از بسیاری هیپوتاالموس غذا مصرف و آب و حرارت، درجه خون، فشار قلب، ضربان
. ریز درون غدد سیستم تنظیم نتیجه در و هیپوفیز غده کنترل این همچنین کند می تنظیم هیجانی حاالت از .محیطی
Each of the Three Divisions of the Autonomic Nervous System Has a Distinctive Anatomical Organization
The Motor Neurons of the Autonomic Nervous System Lie Outside the Central Nervous System
In the somatic motor system the motor neurons are part of the central nervous system: They are located in the spinal cord and brain stem and project directly to skeletal muscle. In contrast, the motor neurons of the sympathetic and parasympathetic motor systems are located outside the spinal cord in the autonomic ganglia. The autonomic motor neurons (also known as postganglionic neurons) are activated by the axons of central neurons (the preganglionic neurons) whose cell bodies are located in the spinal cord or brain stem, much as are the somatic motor neurons. Thus, in the visceral motor system a synapse (in the autonomic ganglion) is interposed between the efferent neuron in the central nervous system and the peripheral target (Figure 49-1).
متمایز تشریحی سازمان یک دارای خودکار عصبی سیستم از بخش سه این از یک هر
مرکزی عصبی سیستم از خارج در سیستم دروغ عصبی اتونوم های نورون موتور
: مستقیم طور به پروژه و مغز ساقه و نخاع در آنها مرکزی عصبی سیستم از بخشی حرکتی نورونهای حرکتی جسمی سیستم در . نخاعی طناب از خارج در پاراسمپاتیک و سمپاتیک موتور سیستم از حرکتی نورونهای مقابل، در دارد قرار اسکلتی های ماهیچه در
( . عصبی های سلول عنوان به همچنین اتونوم حرکتی نورونهای است شده واقع اتونوم گانگلیون می postganglionic در شناختهنورون( ) مرکزی عصبی های سلول های آکسون توسط قرار ( preganglionic شود مغز ساقه یا نخاعی طناب در ها سلول بدن که
) ( . اتونوم گانگلیون در سیناپس یک احشایی موتور سیستم در بنابراین، است سوماتیک حرکتی نورونهای اندازه فعال، است گرفتهشکل ) محیطی هدف به و مرکزی اعصاب سیستم در وابران نورون .interposed( 1-49بین
The sympathetic and parasympathetic nervous systems have clearly defined sensory components that provide input to the central nervous system and play an important role in autonomic reflexes. In addition, some sensory fibers that project to the spinal cord also send a branch to autonomic ganglia, thus forming reflex circuits that control some visceral autonomic functions.
واکنش در مهمی نقش و مرکزی عصبی سیستم به ورودی فراهم که حسی وضوح به پاراسمپاتیک و سمپاتیک عصبی های سیستم . گانگلیون به شاخه یک همچنین پروژه نخاع به که حسی های رشته از برخی این، بر عالوه است شده تعریف بازی خودکار های
احشایی خودکار توابع از بعضی کنترل که است رفلکس مدارات گیری شکل نتیجه در ارسال، .اتونوم
The innervation of target tissues by autonomic nerves also differs markedly from that of skeletal muscle by somatic motor nerves. Unlike skeletal muscle, which has specialized postsynaptic regions (the end-plates; see Chapter 14), target cells of the autonomic nerve fibers have no specialized postsynaptic sites. Nor do the postganglionic nerve endings have presynaptic specializations such as the active zones of somatic motor neurons. Instead, the nerve endings have several swellings (varicosities) where vesicles containing transmitter substances accumulate (see Chapter 15).
innervation حرکتی اعصاب توسط اسکلتی عضالت از آن از توجهی قابل طور به نیز اتونوم اعصاب توسط هدف بافت از . مناطق متخصص که اسکلتی، عضله خالف بر است متفاوت فصل )postsynaptic سوماتیک بشقاب، ( 14پایان های سلول ، ببینید را
تخصصی های سایت هیچ اتونوم عصبی های رشته از عصب . postsynaptic هدف انتهای از نه های postganglionic و تخصص دارای . تورم چندین عصب انتهای عوض، در سوماتیک حرکتی نورونهای فعال مناطق جمله از سیناپس کیسه ( varicosities )پیش آن در که
فصل ) جمع فرستنده مواد حاوی ببینید 15های (.را
Synaptic transmission therefore occurs at multiple sites along the highly branched axon terminals of autonomic nerves. The neurotransmitter may diffuse for distances

P.963as great as several hundred nanometers to reach its targets. In contrast to the point-to-point contacts made in the somatic motor system, neurons in the autonomic motor system exert a more diffuse control over target tissues, so that a relatively
small number of highly branched motor fibers can regulate the function of large masses of smooth muscle or glandular tissue.
. انتقال دهد می رخ اتونوم اعصاب از شاخه بسیار آکسون های پایانه طول در مختلف های سایت در سیناپسی انتقال بنابراینمنتشر های مسافت برای است ممکن عصبی دهنده
P.963
. در شده ساخته تماس نقطه به نقطه به مقابل در است آن اهداف به دستیابی برای نانومتر صد چند عنوان به بزرگ عنوان بهکه طوری به هدف، بافت در بیشتری منتشر کنترل اعمال خودکار موتور سیستم در عصبی های سلول حرکتی، جسمی سیستم
. غده بافت یا و صاف عضله تنظیم بزرگ توده عملکرد تواند می منشعب بسیار موتور الیاف از کمی نسبتا تعداد
Sympathetic Pathways Convey Thoracolumbar Outputs to Ganglia Alongside the Spinal Cord
Preganglionic sympathetic neurons form a column in the intermediolateral horn of the spinal cord extending from the first thoracic spinal segment to rostral lumbar segments. The axons of these neurons leave the spinal cord in the ventral root and initially run together in the spinal nerve. They then separate from the somatic motor axons and project (in small bundles called white myelinated rami) to the ganglia of the sympathetic chains, which lie along each side of the spinal cord (Figure 49-2).
نخاع کنار در عقده به خروجی کمری و سینهای مهرههای انتقال سمپاتیک مسیرهای
سمپاتیک عصبی های شاخ Preganglionic سلول در ستون از intermediolateral یک که نخاع از . نورون این های آکسون دهند می تشکیل را دار نوک سطوح به پشتی فقرات ستون بخش اولین
. های آکسون از را آنها سپس نخاعی عصب در اجرا هم با ابتدا در و شکمی ریشه در نخاع ترک ) ( های عقده به میلین سفید رامی نام به کوچک افزاری نرم بسته در پروژه و حرکتی جسمی
شکل ) فقرات ستون از طرف هر طول در که سمپاتیک، جدا( 2-49زنجیره .دروغ
Axons of preganglionic neurons exit the spinal cord at the level at which their cell bodies are located, but they may innervate sympathetic ganglia situated either more rostrally or more caudally by traveling in the sympathetic nerve trunk that connects the ganglia (Figure 49-2). Most of the preganglionic axons are relatively slow-conducting, small-diameter myelinated fibers. Each preganglionic fiber forms synapses with many postganglionic neurons in different ganglia. Overall, the ratio of preganglionic fibers to postganglionic fibers in the sympathetic nervous system is about 1:10. This divergence permits coordinated activity in sympathetic neurons at several different spinal levels.
عصبی های سلول های بدن preganglionicآکسون آن در که سطح در نخاعی طناب از خروجعصب تنه در سفر با سمپاتیک گانگلیون است ممکن آنها اما دارد، قرار آن در را خود همراه
( شکل گانگلیون که است یا( 2-49سمپاتیک شده واقع یا rostrallyمتصل .caudallyبیشتر پی ترآکسون از . preganglionicبسیاری فیبر هر میلین الیاف قطر کوچک انجام، آهسته نسبتا
preganglionic عصبی های سلول از بسیاری با سیناپس های postganglionicتشکیل گره درالیاف. نسبت کلی، طور به عصبی postganglionicبه preganglionicمختلف سیستم در الیاف
مورد در . 1:10سمپاتیک سمپاتیک عصبی های سلول در هماهنگ فعالیت اجازه واگرایی این است. فقرات ستون مختلف سطوح چند در
The axons of postganglionic neurons are largely unmyelinated and exit the ganglia in the gray unmyelinated rami. The postganglionic cells that innervate structures in the head are located in the superior cervical ganglion, which is a rostral extension of the sympathetic chain. The axons of these cells travel along branches of the carotid arteries to their targets in the head. The postganglionic fibers innervating the rest of the body travel in spinal nerves to their targets; in an average spinal nerve about 8% of the fibers are sympathetic postganglionic axons. Some neurons of the cervical and upper thoracic ganglia innervate cranial blood vessels, sweat glands, and hair follicles; others innervate the glands and visceral organs of the head and chest, including the lacrimal and salivary glands, heart, lungs, and blood vessels. Neurons in the lower thoracic and lumbar paravertebral ganglia innervate peripheral blood vessels, sweat glands, and pilomotor smooth muscle (Figure 49-3).

عصبی های سلول های زیادی postganglionic آکسون حد در unmyelinated تا هسته از خروج و. unmyelinated رامی های سلول است در postganglionic خاکستری سر در ها سازه پی دارای که
شده واقع سمپاتیک زنجیره از دار نوک فرمت یک که است، برتر رحم گردن گانگلیوندر. خود اهداف به کاروتید شریان های شاخه امتداد در سفر ها سلول این از آکسون است
الیاف. یک postganglionic innervating سر در خود، اهداف به نخاعی اعصاب در بدن سفر بقیهمتوسط طور به نخاعی آکسون 8حدود عصب الیاف از ٪ postganglionic . از برخی هستند دلسوز
غدد خون، مغزی عروق کردن پی دارای باال رحم دهانه و سینه های عقده از عصبی های سلولسینه، قفسه و سر از احشایی های اندام و غدد کردن پی دارای برخی و مو های فولیکول و عرق
. پایین سینه قفسه در عصبی های سلول خونی رگهای و ها ریه قلب، بزاقی، غدد و اشکی جمله ازو عرق، غدد خون، محیطی عروق کردن پی دارای های عقده فقرات ستون کمری و pilomotor تر
شکل ) صاف (.3-49عضله
Some preganglionic fibers pass through the sympathetic ganglia and branches of the splanchnic nerves to synapse on neurons of the prevertebral ganglia, which include the coeliac ganglion and the superior and inferior mesenteric ganglia (Figure 49-3). Neurons in these ganglia innervate the gastrointestinal system and the accessory gastrointestinal organs, including the pancreas and liver, and also provide sympathetic innervation of the kidneys, bladder, and genitalia. Another group of preganglionic axons runs in the thoracic splanchnic nerve into the abdomen and innervates the adrenal medulla, which is an endocrine gland, secreting both epinephrine and norepinephrine into circulation. The cells of the adrenal medulla are developmentally and functionally related to postganglionic sympathetic neurons.
فیبرهای از دو preganglionic برخی تماس محل به تصویب احشاء به وابسته اعصاب های شاخه و سمپاتیک گانگلیون طریق ازهسته های نرون در شکل ) prevertebral عصب مزانتریک های عقده تر پست و برتر و سلیاک گانگلیون شامل که سلول(. 49-3،
و کبد، و پانکراس جمله از جانبی، لوازم و گوارش دستگاه های اندام و گوارش دستگاه کردن پی دارای هسته این در عصبی هایارائه . innervation همچنین آکسون از دیگری گروه تناسلی دستگاه و مثانه ها، کلیه از در preganglionic دلسوز شود می اجرا
را نفرین نوراپی و نفرین اپی دو هر ترشح ریز، درون غده یک که آدرنال، تحریک و شکم به سینه قفسه احشاء به وابسته اعصاب . به عملکرد و تکامل حال در آدرنال سلولهای خون گردش است postganglionic به مرتبط سمپاتیک عصبی های .سلول
Parasympathetic Pathways Convey Outputs From the Brain Stem Nuclei and Sacral Spinal Cord to Widely Dispersed Ganglia
The central, preganglionic cells of the parasympathetic nervous system are located in several brain stem nuclei and in segments S2-S4 of the sacral spinal cord (Figure 49-3). The axons of these cells are quite long because parasympathetic ganglia lie close to or are actually embedded in visceral target organs. In contrast, sympathetic ganglia are located at some distance from their targets.
پراکنده ای گسترده طور به عقده نخاع خاجی و هسته مغز ساقه از خروجی انتقال پاراسمپاتیک مسیرهای
های سلول بخش preganglionic مرکزی، در و مغز ساقه های هسته چند در پاراسمپاتیک عصبی شکل ) S2-S4 سیستم خاجی نخاع49-3 . اندامهای( در واقع در یا و نزدیکی در پاراسمپاتیک های عقده خاطر به طوالنی بسیار ها سلول این از آکسون است شده واقع
. است گرفته قرار خود اهداف از دور راه از برخی در سمپاتیک گانگلیون مقابل، در است شده تعبیه احشایی .هدف
The preganglionic parasympathetic nuclei in the brain stem include the Edinger-Westphal nucleus (associated with cranial nerve III), the superior and inferior salivary nuclei (associated with cranial nerves VII and IX, respectively), and the dorsal vagal nucleus and the nucleus ambiguus (both associated with cranial nerve X). Preganglionic axons exiting the brain stem through cranial nerves III, VII, and IX and project to postganglionic neurons in the ciliary, pterygopalatine, submandibular, and otic ganglia (Figure 49-3). Parasympathetic preganglionic fibers from the dorsal vagal nucleus project via nerve X to postganglionic neurons embedded in thoracic and abdominal targets—the stomach, liver, gall bladder, pancreas, and upper intestinal tract (Figure 49-3). Neurons of the ventrolateral nucleus ambiguus provide the principal parasympathetic innervation of the cardiac ganglia, which innervate the heart, esophagus, and respiratory airways.
پاراسمپاتیک هسته preganglionic هسته شامل مغز ساقه کرانیال ) -Edinger در اعصاب با ارتباط در برتر )III وست های هسته ، جمجمه ) اعصاب با ارتباط در بزاقی تر پست ( IX و VII و و پشتی واگ عصب هسته و ، ترتیب به ، ambiguus ( در دو هر هسته
با (. X ارتباط آکسون جمجمه جمجمه Preganglionic عصب اعصاب طریق از مغز ساقه از III خروج ، VII ، IX به پروژه و وpostganglionic ،مژگانی در عصبی های ) pterygopalatineسلول شکل سمعی های عقده و فکی تحت الیاف(. 49-3،

preganglionic طریق از پشتی واگ هسته پروژه از به X پاراسمپاتیک در postganglionic عصب شده جاسازی های نورونشکل ) فوقانی گوارش دستگاه و لوزالمعده صفرا، کبد، معده، هدف، شکم و سینه هسته(. 3-49قفسه های ventrolateral نرون
ambiguus ارائه innervation تنفسی هوایی های راه و مری، قلب، کردن پی دارای که قلبی، های عقده از اصلی .پاراسمپاتیک
In the sacral spinal cord the parasympathetic preganglionic neurons occupy the intermediolateral column. Axons of spinal parasympathetic neurons leave the spinal cord through the ventral roots and project in the pelvic nerve to the pelvic ganglion plexus. Pelvic ganglion neurons innervate the descending colon, bladder, and external genitalia (Figure 49-3).
نورون خاجی نخاع ستون preganglionic در اشغال را پاراسمپاتیک . intermediolateral پاراسمپاتیک عصبی های سلول های آکسون . دارای لگنی عقده نورونهای نخاع ترک را لگن گانگلیونی های شبکه به لگن عصب در پروژه و شکمی های ریشه طریق از نخاعی
شکل ) خارجی تناسلی دستگاه و مثانه نزولی، کولون کردن (.3-49پی
Figure 49-3 Sympathetic and parasympathetic divisions of the autonomic nervous system. Sympathetic preganglionic neurons are clustered in ganglia in the sympathetic chain alongside the spinal cord extending from the first thoracic spinal segment to upper lumbar segments. Parasympathetic preganglionic neurons are located within the brain stem and in segments S2-S4 of the spinal cord. The major targets of autonomic control are shown here.
P.964

. 3-49شکل نورون خودکار عصبی سیستم پاراسمپاتیک و سمپاتیک سمپاتیک preganglionic بخش زنجیره در های گره در سمپاتیک . نورون گرفتند قرار فوقانی سطوح به پشتی فقرات ستون بخش اولین از که نخاعی طناب کنار در preganglionic در پاراسمپاتیک
بخش در و مغز ساقه . S2-S4 داخل است شده داده نشان اینجا در خودکار کنترل اصلی اهداف است شده واقع .نخاع
P.964
The sympathetic nervous system innervates tissues throughout the body, but the parasympathetic distribution is more restricted. There is also less divergence, with an average ratio of preganglionic to postganglionic fibers of about 1:3; in some tissues the numbers may be nearly equal.
. با کمتر، اختالف همچنین است تر محدود پاراسمپاتیک توزیع اما است، بدن سراسر در بافت تحریک را سمپاتیک عصبی سیستممتوسط الیاف preganglionic نسبت حدود postganglionic به است 1:03در برابر تقریبا است ممکن اعداد بافت برخی در .و
The Enteric Nervous System Is Largely Autonomous
مستقل زیادی حد تا ای روده عصبی سیستم
The enteric nervous system controls the function of the gastrointestinal tract, pancreas, and gallbladder. It contains local sensory neurons and interneurons as well as motor neurons and is responsive to alterations in the tension of gut walls and changes in the chemical environment in the gut. The enteric motor neurons control smooth muscle of the gut, local blood vessels, and secretion by the mucosa. The human enteric nervous system has 80-100 million neurons, approximately as many as are found in the spinal cord.
. و محلی حسی نورونهای شامل این صفرا کیسه و پانکراس گوارش، دستگاه عملکرد کنترل ای روده عصبی interneuronsسیستم. نورونهای است روده در شیمیایی های محیط در تغییرات و روده دیواره تنش در تغییرات به پاسخگو و حرکتی نورونهای همچنین و
. انسان روده عصبی سیستم کنترل مخاط از ترشح و محلی، خونی های رگ روده، از روده صاف عضالت 100.000.000-80حرکتی. شود می یافت نخاع در که اندازه همان به تقریبا نورون،
Two major plexuses of nerve cell bodies and fibers extend continuously along the entire length of the gastrointestinal tract (Figure 49-4). These are the myenteric (Auerbach's) plexus, between the outer longitudinal and inner circular smooth muscle layers, and the submucous (Meissner's) plexus between the circular muscle layer and the mucosa. In general, the submucous plexus is concerned
P.965With control of the secretory functions of the gut, while the myenteric plexus controls gut motility. The two plexuses are interconnected, and they contain motor neurons that innervate both smooth muscle and secretory cells in the mucosa, as well as sensory neurons that respond to stretch, tonicity, and specific chemical signals.
شکل ) گوارش دستگاه کل طول امتداد در مداوم طور به الیاف و عصبی های سلول بدن اصلی شبکه . 4-49دو، این( دهد گسترشmyenteric ) ( و صاف، عضله دایره درون و طولی بیرونی های الیه بین ، آورباخ ( submucous شبکه ( الیه بین در مایسنر شبکه
. شبکه کلی، طور به است مخاط و حلقوی است submucous عضالنی نگران
P.965
شبکه کنترل که حالی در روده، ترشحی توابع از کنترل . myenteric با شامل و هستند پیوسته هم به شبکه دو، این روده تحرککشش، به پاسخ در که است حسی نورونهای همچنین و مخاط، در ترشحی های سلول و صاف عضله دو هر که حرکتی نورونهای
کردن پی دارای خاص شیمیایی های سیگنال و .صدا

Figure 49-4 The locations of the mucosal, submucous, and myenteric plexuses between the layers of intestinal wall are shown in three dimensions (A) and in cross-section (B). (Adapted from Furness and Costa 1980.)
مخاط، 4-49شکل از شبکه submucousمکان و، ، myenteric بعد سه در ها روده دیواره از هایی الیه مقطع ( A )بین در نشان( B )و ( . کوستا و فرنس از اقتباس است شده .1980داده
The enteric nervous system is relatively independent of the central nervous system. Although it does have both sympathetic and parasympathetic inputs, these are relatively sparse in relation to the large numbers of enteric neurons. Parasympathetic preganglionic fibers project to enteric ganglia in the stomach, colon, and rectum through the vagus, pelvic, and splanchnic nerves. The sympathetic fibers originate primarily in paravertebral ganglia, although some originate in the prevertebral ganglia, and project mainly to the myenteric and submucous plexuses.
. و سمپاتیک ورودی نوع دو هر دارای که چند هر است مرکزی عصبی سیستم از مستقل نسبتا ای روده عصبی سیستم . الیاف پروژه پاراسمپاتیک است نادر نسبتا روده عصبی های سلول از زیادی تعداد با ارتباط در این به preganglionicپاراسمپاتیک،
. در سرچشمه سمپاتیک الیاف احشاء به وابسته اعصاب و لگن، واگ، طریق از رکتوم و بزرگ روده معده، در ای روده های عقدهگانگلیون در سرچشمه از برخی چند هر فقرات، ستون های گره در اول شبکه prevertebralدرجه به، عمدتا پروژه و ،myenteric و
submucous.
Disruption of enteric connections to the central nervous system results in little or no impairment in function of the small and large bowels; the esophagus and stomach, however, appear to be more dependent on sympathetic and parasympathetic innervation for normal function. The innervation of parts of the gastrointestinal system by the sympathetic and parasympathetic systems may be a way that the other divisions of the autonomic nervous system can override the local nervous control of gut function.
با معده، و مری بزرگ، و کوچک روده عملکرد در هیچ یا و کم اختالل در مرکزی اعصاب سیستم نتایج به روده ارتباطات در اختاللبه وابسته بیشتر رسد می نظر به حال، .innervationاین طبیعی عملکرد برای پاراسمپاتیک و های innervationسمپاتیک قسمت از
تواند می خودکار عصبی سیستم دیگر های بخش که نحوی به است ممکن پاراسمپاتیک و سمپاتیک سیستم توسط گوارش دستگاه. بگذارند پا زیر روده عملکرد از محلی عصبی کنترل
Sensory Inputs Produce a Wide Range of Visceral ReflexesTo maintain homeostasis the autonomic nervous system responds to many different types of sensory inputs. Some of these are somatosensory. For example, a noxious stimulus activates sympathetic neurons that regulate local vasoconstriction (necessary to reduce bleeding when the skin is broken). At the same time, the stimulus activates nociceptive afferents in the spinothalamic tract with axon collaterals to an area in the rostral ventrolateral medulla that coordinates reflexes. These inputs cause widespread sympathetic activation that increases blood pressure and heart rate to protect arterial perfusion pressure and prepares the individual for vigorous defense.
رفلکس احشایی از ای گسترده طیف تولید حسی ورودی
. این از برخی حسی های ورودی مختلف انواع از بسیاری به پاسخ در خودکار عصبی سیستم تعادل حفظ می somatosensoryبرای ( کاهش. منظور به الزم محلی عروق انقباض تنظیم سمپاتیک عصبی های سلول فعال مضر محرک یک مثال، عنوان به باشد
.) دستگاه در درد آوران اعصاب فعال محرک زمان، همان در است شکسته پوست که زمانی Collateralsبا spinothalamicخونریزیمدوال در منطقه به . ventrolateralآکسون است گسترده سمپاتیک شدن فعال باعث ورودی این رفلکس هماهنگی به که دار نوک
. شدید دفاع برای فرد آماده و دهد می افزایش را شریانی خون پرفیوژن فشار از محافظت برای قلب ضربان و خون فشار کهHomeostasis also requires important information about the internal state of the body. Much of this information from the thoracic and abdominal cavities reaches the brain via the vagus nerve. The glossopharyngeal nerve also conveys visceral sensory information from the head and neck. Both of these

nerves and the facial nerve relay special visceral sensory information about taste (a visceral chemosensory function) from the oral cavity. All of these visceral sensory afferents synapse in a topographic fashion in the nucleus of the solitary tract. Taste information is represented most anteriorly; gastrointestinal information, in an intermediate
P.966position; cardiovascular inputs, caudomedially; and respiratory inputs, in the caudolateral part of the nucleus.
. شکم و سینه قفسه حفره از را اطالعات این از بسیاری است بدن داخلی های دولت مورد در مهم اطالعات به نیاز نیز هموستاز . عصب واگ عصب طریق از مغز رسد . glossopharyngealمی از دو هر گردن و سر از احشایی حسی اطالعات انتقال وسیله نیز
( تابع یک مزه و طعم مورد در حسی احشایی ویژه صورت اطالعات عصبی های رله و اعصاب ( chemosensoryاین از احشایی . . اطالعات انفرادی دستگاه از هسته در توپوگرافی مد در عصب دو تماس محل احشایی حسی آوران اعصاب این همه دهان حفره
واسط حد در گوارش، دستگاه اطالعات داده، نشان قدام ترین طعم
P.966
عروقی،. و قلبی های ورودی بخش caudomediallyموقعیت در تنفسی، های ورودی .caudolateralو هسته ازBox 49-1 First Isolation of a Chemical Transmitter
The existence of chemical messengers was first postulated by John Langley and Henry Dale and their students on the basis of their pharmacological studies dating from the beginning of the century. However, convincing evidence for a neurotransmitter was not provided until 1920, when Otto Loewi, in a simple but decisive experiment, examined the autonomic innervation of two isolated, beating frog hearts. In his own words:
The night before Easter Sunday of that year I awoke, turned on the light, and jotted down a few notes on a tiny slip of paper. Then I fell asleep again. It occurred to me at six o'clock in the morning that during the night I had written down something most important, but I was unable to decipher the scrawl. The next night, at three o'clock, the idea returned. It was the design of an experiment to determine whether or not the hypothesis of chemical transmission that I had uttered seventeen years ago was correct. I got up immediately, went to the laboratory, and performed a simple experiment on a frog heart according to the nocturnal design. I have to describe briefly this experiment since its results became the foundation of the theory of chemical transmission of the nervous impulse.
شیمی 1-49جعبه فرستنده یک جداسازی نخست
مطالعات اساس بر را خود آموزان دانش و دیل هنری و النگلی جان توسط بار اولین برای شیمیایی های رسان پیام وجودعصبی . دهنده انتقال یک برای ای کننده قانع شواهد حال، این با است شده طرح قرن آغاز به آن قدمت که را خود فارماکولوژیک
سال تا اتو 1920شد که زمانی ،Loewi قرار بررسی مورد قاطع، اما ساده آزمایش یک در ،innervation ، شده جدا دو از اتونوم: . او خود گفته به است نشده ارائه قورباغه قلب شتم و ضرب
و نور، در تبدیل ، شد بیدار خواب از من سال آن از یکشنبه پاک عید از قبل لغزش jottedشب یک روی بر یادداشت چند کردن . . شب طول در که است صبح شش ساعت در من به را آن برد خوابش دوباره من سپس کاغذ نوشته Iکوچک مهم چیزی کردن
. برای . آزمایش یک از طرح این بازگشت ایده سه، ساعت در ، بعد شب خطی خط کشف به قادر من اما ، است داده رخ بود شده . به بالفاصله، ، شدم بلند من بود درست آمده زبان به پیش سال هفده من که است شیمیایی انتقال فرضیه نه یا آیا اینکه تعیین . از آزمایش این مختصر، طور به من شود می انجام شبانه طراحی به توجه با قورباغه قلب در ساده آزمایش یک و رفت آزمایشگاه
شد . عصبی ضربه از شیمیایی مواد انتقال تئوری اساس و پایه آن نتایجThe hearts of two frogs were isolated, the first with its nerves, the second without. Both hearts were attached to Straub cannulas filled with a little Ringer solution. The vagus nerve of the first heart was stimulated for a few minutes. Then the Ringer solution that had been in the first heart during the stimulation of the vagus was transferred to the second heart. It slowed and its beat diminished just as if its vagus had been stimulated. Similarly, when the accelerator nerve was stimulated and the Ringer from this period transferred, the second heart speeded up and its beat increased. These results unequivocally proved that the nerves do not influence the heart directly but liberate from their terminals specific chemical substances which, in their turn, cause the well-known modifications of the function of the heart characteristic of the stimulation of its nerves.
Loewi called this substance Vagusstoff (vagus substance). Soon after, Vagusstoff was identified chemically as acetylcholine.
کمی . رینگر محلول با شده پر اشتراوب کانول به دل دو هر بدون دوم آن، اعصاب با بار اولین برای شد جدا ، قورباغه دو قلب . . در بار اولین برای قلب در که رینگر محلول سپس بود شده تحریک دقیقه چند برای بار اولین برای قلب واگ عصب شد متصل . آن واگ که همانطور درست ، است یافته کاهش آن شتم و ضرب و کند این شد منتقل دوم قلب به است بوده واگ تحریک طول
. و شتاب دوم قلب ، و بود شده تحریک انتقال دوره این از دهندهرینگر شتاب اعصاب که وقتی مشابه، طور به بود شده تحریک . نیست، قرار تاثیر تحت مستقیم طور به را قلب اعصاب که کرد ثابت صراحت به نتایج این است یافته افزایش آن شتم و ضرباعصاب تحریک قلب مشخصه تابع از شده شناخته تغییرات باعث ، خود نوبه به ، که خاص شیمیایی مواد خود های پایانه از آزاد بلکه
است.
Loewi ماده این نام ، ) ( . Vagusstoffبه آن از پس بالفاصله واگ .Vagusstoffماده است شده شناخته کولین استیل شیمیاییThe nucleus of the solitary tract distributes visceral sensory information within the brain along three main pathways. Some neurons in the nucleus of the solitary tract directly innervate preganglionic neurons in the medulla and spinal cord, triggering direct autonomic reflexes. For example, there are direct inputs from the nucleus of the solitary tract to vagal motor neurons controlling esophageal and gastric motility, which are important for ingesting food. Also, projections from the nucleus of the solitary tract to the spinal cord are involved in respiratory reflex responses to lung inflation.
. دستگاه از هسته در نرونها برخی اصلی مسیر سه طول در مغز در احشایی حسی اطالعات توزیع انفرادی دستگاه از هستهعصبی های سلول کردن پی دارای مستقیم طور به . preganglionicانفرادی به اتونوم مستقیم رفلکس تحریک نخاع، و مغز در

غذا خوردن برای که معده، و مری کنترل واگ عصب حرکتی نورونهای به انفرادی دستگاه از هسته از مستقیم ورودی مثال، عنوان. . است درگیر ریه تورم به تنفسی رفلکس پاسخ در نخاع به انفرادی دستگاه از هسته از بینی پیش همچنین، دارد وجود هستند مهم
Other neurons in the nucleus project to the lateral medullary reticular formation, where they engage populations of premotor neurons that organize more complex, patterned autonomic reflexes. For example, groups of neurons in the rostral ventrolateral medulla control blood pressure by regulating both blood flow to different vascular beds and vagal tone in the heart to modulate heart rate. Other groups of neurons control complex responses such as vomiting and respiratory rhythm (a somatic motor response that has an important autonomic component and that depends critically on visceral sensory information).
عصبی های سلول جمعیت درگیر آنها که جایی جانبی، مدوال رتیکوالر تشکیل به هسته پروژه در دیگر که premotorنورون . در عصبی های سلول گروه مثال، عنوان به الگو خودکار های رفلکس تر، پیچیده مغز ventrolateralسازماندهی خون فشار کنترل
. های سلول از دیگر گروه قلب ضربان بم و زیر به قلب در واگ تن و مختلف عروق بستر به خون جریان دو هر تنظیم با منقاریاست ) وابسته و اتونوم مهم جزء یک دارای که است حرکتی جسمی پاسخ تنفسی ریتم و استفراغ مانند پیچیده های واکنش عصبی
. ) کنید کنترل را احشایی حسی اطالعاتThe third main projection from the nucleus of the solitary tract provides visceral sensory input to a network of cell groups that extend from the pons and midbrain up through the hypothalamus, amygdala, and cerebral cortex. This network coordinates autonomic responses and integrates them into ongoing patterns of behavior. These will be described in more detail after we consider more elementary autonomic reflexes.
گسترش را پونز از که سلولی های گروه از ای شبکه به احشایی حسی ورودی انفرادی دستگاه از هسته از اصلی طرح سومین . را آنها ادغام و هماهنگ خودکار های پاسخ شبکه این کند می فراهم میانی مغز مغز قشر و آمیگدال هیپوتاالموس، طریق از و داده. . تر ابتدایی اتونوم های رفلکس نظر در ما از بعد شده داده شرح بیشتر جزئیات با شد خواهد این رفتار انجام حال در الگوهای در
Discrete Autonomic Reflexes Produce Both Slow and Rapid Visceral ResponsesThe usual role of the autonomic nervous system is to control a variety of visceral and ocular reflexes. Some of these reflexes are relatively fast, for example, adjustment of pupil size in response to light. Others, such as glandular secretion or gastrointestinal responses to food, are slow. Some bodily functions are under the dual control of the autonomic and somatic motor systems.
احشایی سریع و آهسته دو هر پاسخ با رفلکس خودکار گسسته
. به سریع، نسبتا واکنش این از برخی چشم و احشایی رفلکس انواع کنترل برای است خودکار عصبی سیستم از معمول نقش . آهسته غذا، به گوارش دستگاه های پاسخ یا و غددی ترشح مانند دیگران، است نور به پاسخ در مردمک اندازه تنظیم مثال، عنوان
. . باشد می جسمی و خودکار موتور سیستم از دوگانه کنترل تحت بدن عملکردهای از بعضی باشد می
Ocular ReflexesThe autonomic nervous system controls two movements of the eye: opening the pupils and focusing the lens. Pupil size determines the amount of light impinging on the retina. Sympathetic fibers from the superior cervical ganglion innervate the muscles of the iris that dilate the pupil, while parasympathetic fibers innervate circular muscle fibers of the iris that constrict the pupil. Ordinarily, the parasympathetic and sympathetic controls
P.967are balanced to achieve the appropriate pupillary opening, although fine-tuning of pupil size may be largely under parasympathetic control. Under conditions of excitement or alarm there is a shift in this balance, inhibiting pupillary constriction and increasing tone in the pupillodilator muscle of the iris. Focusing of the lens is regulated almost entirely by parasympathetic control of ciliary muscles, whereas Muller's muscle, which retracts the eyelids, is under sympathetic control.
چشمی رفلکس
. : روی بر نور برخورد میزان تعیین مردمک اندازه لنز تمرکز و آموزان دانش کردن باز چشم جنبش دو کنترل خودکار عصبی سیستمالیاف. که حالی در مردمک، شدن گشاد که عنبیه عضالت کردن پی دارای رحم گردن برتر گانگلیون از سمپاتیک الیاف شبکیه
. سمپاتیک و پاراسمپاتیک کنترل معمول، مردمک انقباض که عنبیه حلقوی عضالنی الیاف پی پاراسمپاتیک
P.967
حد تا است ممکن مردمک اندازه تنظیم ریز چند هر است، مناسب چشم مردمک کردن باز به رسیدن برای شده کننده متعادل و ها . در تن افزایش و مردمک انقباض مهار تعادل، این در تغییر است هشدار زنگ یا و هیجان شرایط در پاراسمپاتیک کنترل تحت زیادی
. pupillodilatorعضله تنظیم، مژگانی عضالت پاراسمپاتیک کنترل توسط کامل طور به تقریبا است لنز از تمرکز دارد وجود عنبیه از. دلسوز کنترل تحت پلک، جمع که مولر، عضله که حالی در

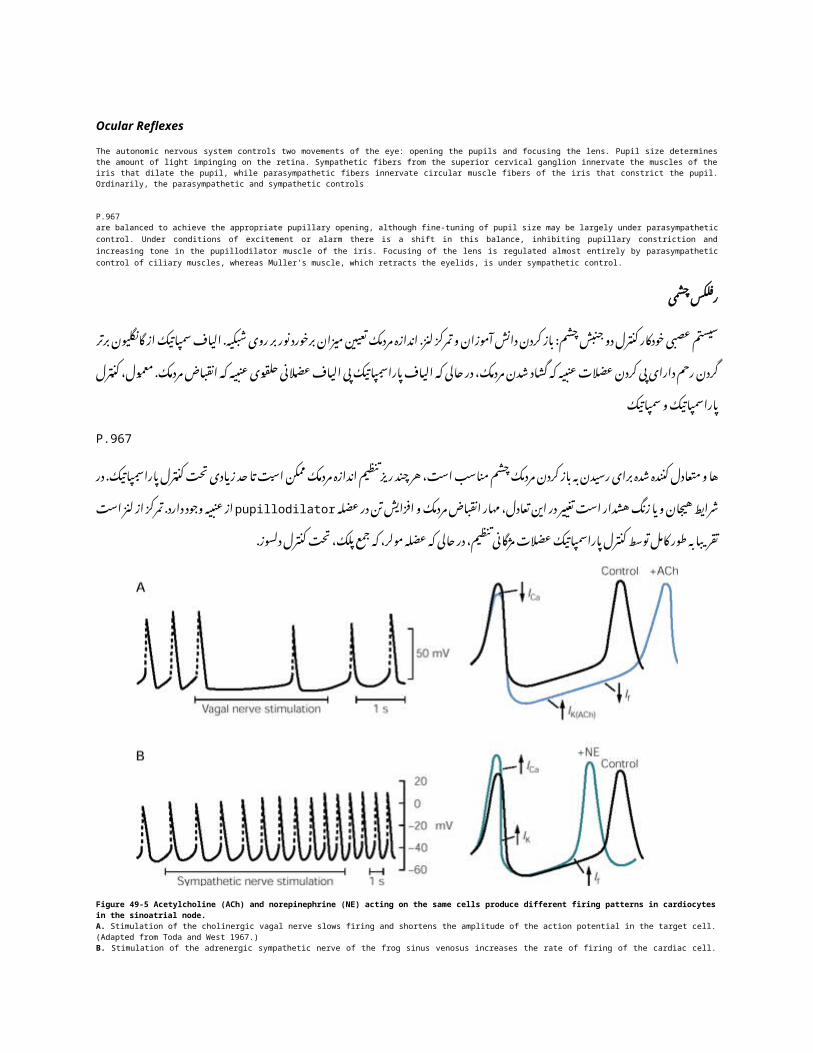
Figure 49-5 Acetylcholine (ACh) and norepinephrine (NE) acting on the same cells produce different firing patterns in cardiocytes in the sinoatrial node.A. Stimulation of the cholinergic vagal nerve slows firing and shortens the amplitude of the action potential in the target cell. (Adapted from Toda and West 1967.)B. Stimulation of the adrenergic sympathetic nerve of the frog sinus venosus increases the rate of firing of the cardiac cell. (Adapted from Hutter and Trautwein 1956.)
کولین )5-49شکل نفرین( )ACHاستیل نوراپی در( NEو مختلف شلیک الگوهای تولید همان های سلول به در cardiocytesاقدام. سینوسی گره
A ( . و. تودا از اقتباس هدف سلول در عمل پتانسیل دامنه شدن کوتاه باعث و شلیک کند می کند را کولینرژیک واگ عصب تحریک.(1967غرب
B ( . از. اقتباس دهد می افزایش را قلبی های سلول از شلیک نرخ قورباغه سیاهرگی سینوس از آدرنرژیک سمپاتیک عصب تحریکHutter وTrautwein 1956).
Cardiovascular Reflexes
Arterial blood pressure is determined by the rate of output of blood from the heart and the resistance to blood flow through the blood vessels. The sympathetic system increases heart rate and strength of contraction; the parasympathetic slows the heart. Sympathetic stimulation increases blood pressure by increasing cardiac output and peripheral resistance (by constricting small arterioles). Parasympathetic stimulation has a smaller effect on peripheral resistance, although some vasodilatory responses occur, as in blushing. Parasympathetic vasodilation may involve unconventional chemical messengers such as nitric oxide. Under resting conditions almost all systemic arterioles are constricted to approximately half maximal diameter by ongoing sympathetic tonic activity. A decrease in sympathetic output leads to vasodilation; an increase, to further constriction. Without ongoing tonic activity of the sympathetic system, sympathetic output could only increase and thus control only constriction.
عروق و قلب واکنش
خونی عروق در را خون جریان برابر در مقاومت و قلب از خون خروجی میزان توسط خون فشار . . قلب کند می کند را پاراسمپاتیک انقباض قدرت و قلب ضربان سمپاتیک سیستم شود می تعیین
مقاومت. و قلبی ده برون افزایش با دهد می افزایش را خون فشار سمپاتیک تحریک است . ) ( مقاومت بر کمتری اثر پاراسمپاتیک تحریک کوچک شریانی محدودیت اساس بر محیطی
. خون تاخیر سرخ در دهد، می رخ عروق کنندگی گشاد های پاسخ از برخی چند هر ، محیطی

. در شود می نیتریک اکسید مانند متعارف غیر شیمیایی های رسان ممکن پاراسمپاتیک شاهرگدر های فعالیت حداکثر از نیمی حدود قطر به سیستمیک شریانی تمام تقریبا استراحت شرایط
. ، شاهرگ خون تاخیر به منجر سمپاتیک خروجی در کاهش منقبض دلسوز تونیک انجام حال . ، سمپاتیک سیستم مقوی انجام حال در های فعالیت بدون است بیشتر انقباض به ، افزایش
انقباض . فقط کنترل نتیجه در و دهد افزایش تواند می تنها دلسوز خروجیSympathetic vasoconstrictor tone results from continuous firing of mainly adrenergic neurons in the rostral ventrolateral medulla, which innervate sympathetic vasoconstrictor preganglionic neurons. Activation of pressure-sensitive (baroreceptor) neurons that innervate the aortic arch and the carotid sinus signal an increase in blood pressure to the nucleus of the solitary tract. Neurons of this nucleus excite interneurons in the caudal ventrolateral medulla, which in turn both inhibit the tonic vasomotor neurons and excite vagal cardiomotor neurons. The result, the baroreceptor reflex, is a fall in both arterial blood pressure and heart rate.
آدرنرژیک عمده طور به عصبی های سلول از مداوم شلیک از ناشی سمپاتیک عروق کننده تنگ تنمنقاری مدوال نورون ventrolateralدر دلسوز عروق کننده تنگ که ،preganglionic . سازی فعال پی
) سینوس ) و آئورت قوس کردن پی دارای که فشار به حساس عصبی های سلول بارورسپتورها از . تحریک به هسته این های نورون انفرادی دستگاه از هسته به خون فشار افزایش سیگنال کاروتید
interneurons مدوال وازوموتور ventrolateralدر نورون تونیک دو هر خود نوبه به که دم، به وابستهعصبی های سلول تحریک و مهار . cardiomotorرا در سقوط بارورسپتورها، رفلکس نتیجه، در واگ
. است قلب ضربان و خون فشار دو هرThe actions of norepinephrine and acetylcholine (ACh) on the heart are worth considering in detail as examples of the complex cellular regulatory systems involved in autonomic control. Norepinephrine acts on cardiac muscle to stimulate heart rate and force of contraction. It increases the force of contraction by acting on b-adrenergic receptors that activate the cyclic adenosine monophosphate (cAMP) second-messenger system, which in turn increases the long-lasting (L-type) Ca2+ channel current in the muscle (Chapter 14). Activation of the b-adrenergic receptors also decreases the threshold for firing the cardiac pacemaker cells in the sinoatrial node, thereby increasing heart rate. These effects of norepinephrine can be potently reinforced by circulating epinephrine released from the adrenal medulla.
( کولین استیل و نفرین نوراپی عنوان( ACHاقدامات به جزئیات در ارزش به توجه با قلب در . نفرین نوراپی باشد می خودکار کنترل در دخیل پیچیده سلولی نظارتی های سیستم از هایی نمونه . نیروی این شود می انقباض قدرت و قلب ضربان تحریک برای کند می عمل قلب عضله در
( حلقوی فسفات منو آدنوزین منوفسفات که آدرنرژیک بتا های گیرنده بر اثر با (CAMPانقباض( دهد می افزایش خود نوبه به که رسول، دوم ( Lسیستم + CA2نوع در مدت طوالنی کانال
فصل ) باعث( . 14عضالت نیز آدرنرژیک بتا های گیرنده از سازی فعال دهد می افزایش را فعالافزایش موجب که سینوسی، گره در قلبی ساز ضربان های سلول شلیک برای آستانه کاهش
. در آدرنال از شده آزاد نفرین اپی های طور به توان می را نفرین اپی نور اثرات این قلب ضربان. است شده تقویت خون گردش
ACh is released from parasympathetic nerve terminals, as first shown by Otto Loewi in his classic experiment proving the existence of chemical neurotransmitters (Box 49-1). ACh slows the heart by acting on muscarinic receptors in the cardiocytes of the sinoatrial and atrioventricular nodes of cardiac muscle, thus increasing a resting K+ conductance in these cells. The
P.968increase in K+ conductance hyperpolarizes sinoatrial cells, thus slowing conductance through the atrioventricular node. Hyperpolarization of the sinoatrial cells appears to involve direct gating of a K+ channel by a G protein activated by the muscarinic receptor. ACh also decreases heart rate by increasing the threshold for firing the pacemaker cells in a manner opposite to that of norepinephrine, thereby slowing the heart rate (Figure 49-5). ACh also reduces the force of contraction by decreasing intracellular cAMP, thus reducing the L-type Ca2+ current.

ACH اتو توسط بار اولین عنوان به شد، منتشر پاراسمپاتیک عصبی های پایانه وجود Loewiاز اثبات به خود معروف آزمایش درجعبه ) شیمیایی عصبی های دهنده . 1-49انتقال است( شده داده موسکارینی ACHنشان های گیرنده بر اثر با قلب کند می کند را
استراحت cardiocytesدر افزایش نتیجه در است، قلب عضله دهلیزی و سینوسی گره .Kاز ها + سلول این در هدایت
P.968. hyperpolarizesهدایت + Kافزایش دهلیزی گره طریق از هدایت کاهش نتیجه در سینوسی، های Hyperpolarizationسلول
کانال یک از مستقیم راهگاهی شامل رسد می نظر به و نخورده دست سلولهای پروتئین + Kاز یک توسط Gتوسط شده فعال . موسکارینی های مقابل ACHگیرنده در را ساز ضربان های سلول شلیک برای آستانه افزایش با قلب ضربان یابد می کاهش نیز
شکل ) قلب ضربان شدن کند نتیجه در نفرین، اپی نور به که ای کاهش ACH(. 5-49شیوه با انقباض نیروی کاهش cAMPباعثکاهش نتیجه در سلولی، .CA2 Lداخل است + جاری نوع از
Glandular ReflexesNasal, lacrimal, and many gastrointestinal glands are strongly stimulated by parasympathetic inputs. The enteric glands most strongly stimulated by the parasympathetic system are in the upper alimentary tract, particularly in the mouth and stomach. Glandular secretion in lower parts of the alimentary tract is mostly under the autonomous control of the enteric nervous system. Salivary glands respond to both parasympathetic and sympathetic stimulation with secretion. Sympathetic stimulation elicits viscous secretion with a high amylase content, and parasympathetic stimulation elicits a more copious, watery saliva.
Sympathetic activity generally reduces glandular secretion because it causes vasoconstriction, whereas parasympathetic stimulation increases local blood flow, promoting secretion. Sweat glands are an exception to this rule, as sympathetic stimulation increases sweating. Most of the sympathetic fibers are cholinergic rather than adrenergic, but in humans many sympathetic fibers to sweat glands are under {a}-adrenergic control.
غده رفلکس
. توسط شدت به اغلب روده غدد شده تحریک پاراسمپاتیک ورودی شدت به گوارش دستگاه غدد از بسیاری و ، اشکی ، بینی . پایین های قسمت در غده ترشح است معده و دهان در خصوص به هستند، فوقانی گوارش دستگاه در تحریک پاراسمپاتیک سیستم
و . پاراسمپاتیک دو هر تحریک به ترشح با بزاقی غدد است روده عصبی سیستم از مستقل کنترل تحت عمدتا گوارش دستگاه از تر . بزاق ، تر موجبفراوان پاراسمپاتیک تحریک و باال، آمیالز محتوای با چسبناک ترشح موجب سمپاتیک تحریک دهند پاسخ سمپاتیک
آبکی .
افزایش پاراسمپاتیک تحریک که حالی در ، عروق انقباض باعث چون غددی ترشح دهد می کاهش کلی طور به سمپاتیک فعالیتالیاف . . از بسیاری کردن عرق سمپاتیک تحریک افزایش با ، قانون این در استثنا یک عرق غدد ترشح ترویج محلی، خون جریان
. باشد }{ می آدرنرژیک کنترل تحت عرق غدد به سمپاتیک الیاف از بسیاری انسان در اما هستند، آدرنرژیک نه و کولینرژیک سمپاتیکGastrointestinal ReflexesGastrointestinal function is controlled by many autonomic reflexes. Some depend on input from the parasympathetic or sympathetic nervous systems (eg, control of gastric acid secretion in the stomach), while others are mainly under local control of the enteric nervous system. For example, intestinal peristalsis —the wave of muscle contractions along the length of the intestine that propels intestinal contents toward the anus—is controlled entirely by the enteric nervous system.
As food enters the intestine it pushes the intestinal wall outward, thus stretching sensory neurons in the wall. When sufficiently stretched, these neurons activate interneurons and motor neurons in the myenteric plexus to move the food forward. Peristalsis starts with the activation of excitatory motor neurons whose fibers project orally, causing the circular muscle at the oral end of the intestinal distention to contract. At the same time, reflex activation of inhibitory motor neurons, whose fibers project anally, relaxes the circular smooth muscle at the anal end of the distention. The waves of contraction and relaxation of the intestinal wall propel the food through the intestines. During peristalsis, parasympathetic nerves excite enteric neurons through nicotinic receptors and contracts smooth muscle through muscarinic receptors. Nitric oxide is thought to mediate smooth muscle relaxation in peristalsis.
روده و معده رفلکس . سیستم از ورودی در از برخی شود می کنترل خودکار های رفلکس از بسیاری توسط که است گوارش دستگاه عملکرد
) عمده ) طور به دیگران که حالی در دارد، بستگی معده در معده اسید ترشح کنترل مثال، عنوان به عصبی سمپاتیک و پاراسمپاتیک - . که است روده طول در عضالنی انقباضات از موج دودی حرکت روده مثال، عنوان به روده عصبی سیستم از محلی کنترل تحت
. شود می کنترل ای روده عصبی سیستم توسط کامل طور به مقعد، سمت به روده محتویات سوق . اندازه به که هنگامی دیوار در حسی نورونهای کشش نتیجه در بیرون، به روده دیواره دهد می هل را آن روده وارد غذا عنوان به
فعال را ها نورون این کشیده، شبکه interneuronsکافی در حرکتی نورونهای . myentericو دودی حرکت جلو به غذا حرکت بهدهان پایان در حلقوی عضالنی باعث شفاهی صورت به پروژه الیاف که تحریکی حرکتی نورونهای شدن فعال با شود می شروع
. پروژه الیاف که بازدارنده، حرکتی نورونهای از رفلکس شدن فعال زمان، همان در قرارداد به روده شدن anallyاتساع شل ، . . حرکت طول در ها روده طریق از غذا حرکت روده دیواره شدن شل و انقباض امواج اتساع مقعد پایان در دایره صاف عضالت
موسکارینی های گیرنده طریق از و نیکوتین های گیرنده طریق از روده عصبی های سلول تحریک پاراسمپاتیک اعصاب دودی،. . دودی حرکت در صاف عضالت شدن شل واسطه به نیتریک اکسید که شود می تصور صاف عضله قرارداد

Urogenital Reflexes
The control of bladder emptying is unusual because it involves both involuntary autonomic reflexes and some voluntary control. The excitatory input to the bladder wall that causes contraction and promotes emptying is parasympathetic. Activation of parasympathetic postganglionic neurons in the pelvic ganglion plexus near to and within the bladder wall contracts the bladder's smooth muscle. These neurons are quiet when the bladder begins to fill but are activated reflexly by visceral afferents when the bladder is distended.
The sympathetic nervous system relaxes the bladder smooth muscle. Axons of preganglionic sympathetic neurons project from the thoracic and upper lumbar spinal cord to the inferior mesenteric ganglion. From there, postganglionic fibers travel to the bladder in the hypogastric nerve. When the sympathetic system is activated by low-frequency firing in sensory afferents that respond to tension in the bladder wall, the parasympathetic neurons in the pelvic ganglion are inhibited, relaxing bladder smooth muscle and exciting the internal sphincter muscle. Thus, during bladder filling the sympathetic system promotes relaxation of the bladder wall directly while maintaining closure of the internal sphincter.
Somatic motor neurons in the ventral horn of the sacral spinal cord innervate striated muscle fibers in the external urethral sphincter, causing it to contract. These motor neurons are stimulated by visceral afferents that are activated when the bladder is partially full. As the bladder fills, spinal sensory afferents relay this information to a region in the pons that coordinates micturition. This pontine area, sometimes called Barrington's nucleus after the British neurophysiologist who first described it, also receives important descending inputs from the forebrain concerning behavioral cues for emptying the bladder. Descending pathways from Barrington's nucleus cause coordinated inhibition of sympathetic and somatic systems, relaxing both sphincters. The onset of urinary flow through the urethra causes reflex contraction
P.969of the bladder that is under parasympathetic control.
تناسلی ادراری رفلکس . ورودی ارادی کنترل از برخی و ارادی غیر اتونومیک رفلکس دو هر شامل را آن که چرا است معمول غیر مثانه تخلیه کنترل . عصبی های سلول از سازی فعال است پاراسمپاتیک تخلیه ترویج و انقباض باعث که است مثانه جدار به تحریکی
postganglionic . سلول این است مثانه صاف عضله منقبض مثانه جدار در و به نزدیک لگن گانگلیونی های شبکه در پاراسمپاتیکاما کردن پر به شروع مثانه که هستند آرام عصبی است .reflexlyهای متسع مثانه که زمانی فعال احشایی آوران اعصاب توسط
سمپاتیک . عصبی های سلول های آکسون مثانه صاف عضالت شدن شل سمپاتیک، عصبی و preganglionicسیستم ای سینه از . الیاف آنجا، از باشد می پروژه تحتانی مزانتریک گانگلیونی به نخاعی طناب فوقانی در postganglionicکمری مثانه به سفر
در . تنش به که است حسی آوران اعصاب در پایین فرکانس با شلیک توسط سمپاتیک سیستم که هنگامی شکم زیر در واقع عصبانگیز هیجان و صاف عضله مثانه بخش آرامش ، مهار لگن گانگلیون در پاراسمپاتیک عصبی های سلول فعال، پاسخ مثانه جدارحالی . در مستقیم طور به ترویج را مثانه جدار از آرامش سمپاتیک سیستم کردن پر مثانه طول در بنابراین، داخلی اسفنکتر عضله
داخلی . اسفنکتر شدن بسته حفظ کهبه را آن باعث و خارجی، مجرای اسفنکتر در دار خط عضالنی فیبرهای پی خاجی نخاع قدامی شاخ در سوماتیک حرکتی نورونهای
. شده. تحریک است پر نیمه مثانه که زمانی و شوند می فعال که است احشایی آوران اعصاب توسط حرکتی نورونهای این قرارداد . ، پونز منطقه این ادرار مختصات که پونز در منطقه یک به را اطالعات این رله نخاعی حسی آوران اعصاب ، پر مثانه که همانطور
نزولی مهم های ورودی همچنین شود، می نامیده توصیف را آن بار اولین که انگلیسی اعصاب از پس برینگتون هسته اوقات گاهی . در برینگتون هماهنگ مهار علت هسته از مسیر نزولی کند می دریافت مثانه تخلیه برای رفتاری های نشانه مورد در مغز جلو از
رفلکسی . انقباض باعث مثانه خروجی مجرای طریق از ادرار جریان شروع اسفنگتر دو هر استراحت ، جسمی و سمپاتیک سیستمP.969
پاراسمپاتیک . کنترل تحت که است مثانه
Figure 49-6 Both acetylcholine (ACh) and a luteinizing hormone-releasing hormone (LHRH)-like peptide are released by presynaptic cells at synapses in the sympathetic chain ganglia in the bullfrog. The two transmitters produce different types of postsynaptic potentials in different postganglionic neurons because of their actions on different receptors. (Adapted from Jan and Jan 1983.)A. In one type of postganglionic neuron a single presynaptic stimulus evokes a fast excitatory postsynaptic potential (fast EPSP) at a nicotinic ACh receptor. Repetitive stimulation evokes a slow inhibitory postsynaptic potential (slow IPSP) at a muscarinic ACh receptor and a slow EPSP at a peptidergic receptor.B. In another class of postganglionic neurons a single presynaptic stimulus also evokes a fast EPSP at the nicotinic ACh receptor but repetitive stimulation leads to a slow EPSP at the muscarinic ACh receptor. This class of neurons also evokes the slow peptidergic EPSP, but only in response to stimulation of the preganglionic fibers shown in A. The peptide diffuses from these terminals to distant receptors.
) 6-49شکل کولین استیل دو ) ACHهر ) زرد جسم هورمون کننده آزاد هورمون ( LHRHو پیش های سلول در پپتید مانند . پتانسیل از مختلف انواع تولید فرستنده دو شد منتشر امریکایی بزرگ غوک در سمپاتیک زنجیره هسته در سیناپس در سیناپس

postsynaptic مختلف های نورون ژان . ) postganglionicدر و ژان از اقتباس مختلف های گیرنده در را خود اقدامات دلیل به1983) .
A .نورون نوع یک بالقوه postganglionicدر تداعی سیناپس پیش واحد محرک سریع ) postsynapticیک سریع EPSPتحریکیگیرنده( یک . ACHدر مهار آهسته بالقوه تداعی تکراری تحریک ) postsynapticنیکوتین ( IPSPآهسته های گیرنده در
گیرنده EPSPو ACHموسکارینی یک در .peptidergicآهستهB .عصبی های سلول از دیگری دسته تداعی postganglionicدر نیز سیناپس پیش واحد محرک گیرنده EPSPیک در ACHسریع
به منجر تکراری تحریک اما گیرنده EPSPنیکوتین در . ACHکند کند تداعی نیز عصبی های سلول از دسته این موسکارینیpeptidergic EPSP الیاف از تحریک به پاسخ در تنها اما ،preganglionic در شده داده پایانه. Aنشان این از شده منتشر پپتید
. دور های گیرنده بهIn patients with spinal cord injuries at the cervical or thoracic levels, the spinal reflex control of micturition remains intact, but the connections with the pons are severed. As a result, micturition cannot be voluntarily controlled. When it does occur as a spinal reflex resulting from bladder overfilling, urination is incomplete. As a result, urinary tract infections are common, and it may be necessary to empty the bladder mechanically by catheterization
Sexual reflexes are organized in a pattern that is analogous to those controlling bladder function. Erectile tissue is controlled largely by the parasympathetic nervous system, involving neurons that produce nitrous oxide as their main mediator. Glandular secretion is also parasympathetically mediated. Ejaculation in males is caused by sympathetic control of the seminal vesicles and vas deferens, and emission involves control of striated muscles in the pelvic floor as well. Supraspinal inputs play an important role in producing the coordinated pattern of sexual response, although some simple sexual reflexes can be activated even after spinal transection (eg, penile erection can be elicited by local sensory stimuli).
می باقی نخورده دست ادرار از نخاعی رفلکس کنترل سینه، قفسه یا و رحم گردن سطح در نخاعی های آسیب به مبتال بیماران در . . می رخ کند را آن که هنگامی شود می کنترل داوطلبانه طور به توان نمی را ادرار نتیجه، در هستند جدا پونز با ارتباط اما ماند،
از ناشی نخاعی رفلکس یک عنوان به . overfillingدهد شایع ادراری دستگاه های عفونت نتیجه، در است ناقص کردن ادرار مثانه،سوند توسط مکانیکی مثانه کردن خالی برای باشد الزم است ممکن و است،
. که است شونده سیخ بافت است یافته سازمان مثانه عمل کنترل که کسانی با مقایسه در که است الگوی یک در جنسی رفلکسعنوان به نیتروژن اکسید تولید برای که است عصبی های سلول شامل که کنترل، پاراسمپاتیک عصبی سیستم توسط زیادی حد تا
. نیز غده ترشح آنها اصلی . parasympatheticallyمیانجی سمینال وزیکول از دلسوز کنترل با که است مردان در انزال واسطه . در مهمی نقش نخاعی، فوق های ورودی هست نیز لگن کف در مخطط عضالت کنترل شامل انتشار و شود، می ایجاد دفران و
نخاع قطع از پس حتی توان می را ساده جنسی های واکنش از برخی چند هر بازی، می جنسی های پاسخ از هماهنگ الگوی تولید.) مشخص ) محلی حسی های محرک با توان می را تناسلی آلت نعوظ مثال، عنوان به فعال
Autonomic Neurons Use a Variety of Chemical Transmitters
Autonomic ganglion cells receive and integrate inputs from both the central nervous system (through preganglionic nerve terminals) and the periphery (through branches of sensory nerves that terminate in the ganglia). Most of the sensory fibers are nonmyelinated and may release neuropeptides, such as substance P and calcitonin gene-related peptide (CGRP), onto ganglion cells. Preganglionic fibers primarily use ACh and norepinephrine as transmitters.
شیمی انتقال انواع از استفاده با عصبی های سلول اتونومعصبی ) های پایانه طریق از مرکزی عصبی سیستم دو هر از ورودی خودگردان گانگلیونی های حاشیه( preganglionicسلول و
. ) حسی) های رشته از بسیاری دارد اختیار در و دریافت را خاتمه گانگلیون در که حسی اعصاب های شاخه طریق ازnonmyelinated نوروپپتید است ممکن و مانند neuropeptideاست کلسیتونین )Pآزاد، ژن با مرتبط پپتید و بر( CGRPمواد ،
. الیاف گانگلیونی های سلول از Preganglionicروی استفاده اول درجه .ACHدر فرستنده عنوان به را نفرین نوراپی و
Ganglionic Transmission Involves Both Fast and Slow Synaptic Potentials
Preganglionic activity induces both brief and prolonged responses from postganglionic neurons. ACh released from preganglionic terminals evokes fast excitatory postsynaptic potentials (EPSPs) mediated by nicotinic ACh receptors. The fast EPSP is often large enough to generate an action potential in the
postganglionic neuron, and it is thus regarded as the principal synaptic pathway for ganglionic transmission in both the sympathetic and parasympathetic systems.
آهسته و سریع دو هر سیناپسی پتانسیل شامل انتقال گانگلیونی
عصبی Preganglionicفعالیت های سلول از مدت طوالنی و کوتاه پاسخ دو . postganglionicهر پایانه ACHباعث از شده آزادتحریک preganglionicهای سریع پتانسیل نیکوتین postsynaptic (EPSPS)تداعی رسپتورهای واسطه EPSPسریع. ACHبه
نورون در عمل پتانسیل یک تولید برای بزرگ کافی اندازه به های postganglionicاغلب مسیر عنوان به بنابراین و است ، . گرفته نظر در پاراسمپاتیک و سمپاتیک سیستم دو هر در گانگلیونی انتقال برای اصلی سیناپسی
ACh also evokes slow EPSPs and inhibitory postsynaptic potentials (IPSPs) in postganglionic neurons. These slow potentials can modulate the excitability of these cells. They have been most often studied in sympathetic ganglia but are also known to occur in some parasympathetic ganglia. Slow EPSPs or IPSPs are mediated by muscarinic ACh receptors (Figure 49-6). The slow excitatory potential results when Na+ and Ca2+ channels open and M-type K+ channels close. The M-type channels are normally active at the resting membrane potential, so their closure leads to membrane depolarization (Chapter 13). The slow inhibitory potential results from the opening of K+ channels, allowing K+ ions to flow out of the nerve terminals, resulting in hyperpolarization.
ACH تداعی پتانسیل EPSPSنیز و نورون( IPSPsمهاری )postsynapticکند می. postganglionicدر آهسته های پتانسیل این . همچنین اما گرفته قرار مطالعه مورد سمپاتیک های گره در اغلب و اند شده آنها بم و زیر ها سلول این از پذیری تحریک تواند
. دهد می رخ پاراسمپاتیک های عقده از برخی به شود می یا EPSPSشناخته و موسکارینی IPSPsآهسته های گیرنده ACHتوسط: + 6-49شکل) . و( سدیم های کانال که زمانی تحریکی پتانسیل شماره از نتایج آهسته واسطه و + CA2با نوع Mکانال + Kباز از
. های کانال است به- Mنزدیک منجر آنها شدن بسته از پس هستند، فعال معمول طور به استراحت حال در غشا پتانسیل در نوعفصل ) دپالریزاسیون : 13غشاء های(. کانال شدن باز از مهاری پتانسیل شماره از نتایج های + Kآهسته یون دهد می اجازه ،K + به
نتیجه در و عصبی، های پایانه از .hyperpolarizationجریان
The fast cholinergic EPSP reaches a maximum within 10-20 ms; the slow cholinergic synaptic potentials take up to half a second to reach their maximum and last for a second or more (Figure 49-6). Even slower synaptic potentials, lasting up to a minute, are evoked by neuropeptides, a variety of which are present in the terminals of preganglionic neurons and sensory nerve endings. The actions of one peptide have been studied in detail and reveal important features of peptidergic transmission.
کولینرژیک در EPSPسریع حداکثر رسد برای 20-10می دوم نیمه به تا را کند کولینرژیک سیناپسی های پتانسیل از ثانیه، میلی ( شکل بیشتر یا ثانیه یک برای را خود آخرین و حداکثر به دقیقه،(. 6-49رسیدن یک مدت به پتانسیل سیناپسی تر آهسته حتی
نوروپپتید عصبی neuropeptideتوسط های سلول های پایانه در حاضر حال در که انواع ،preganglionic حسی عصب انتهای و . انتقال مهم های ویژگی از دهد می نشان و گرفته قرار بررسی مورد مفصل طور به پپتید یک اقدامات برانگیخته می
peptidergic.
In some, but not all, preganglionic nerve terminals in bullfrog sympathetic ganglia, ACh is colocalized with a luteinizing hormone-releasing hormone (LHRH)-like peptide. High-frequency stimulation of the preganglionic nerves causes the peptide to be released, evoking a slow, long-lasting EPSP in all postganglionic neurons (Figure 49-6), even those not directly innervated by the peptidergic fibers. The peptide must diffuse over considerable distances to influence distant receptive neurons. The slow peptidergic EPSP, like the slow cholinergic excitatory potential, also results from the closure of M-type channels and the opening of Na+ and Ca2+ channels. The peptidergic excitatory potential alters the excitability of autonomic ganglion cells for long periods after intense activation of preganglionic inputs. No mammalian equivalent of the actions of the LHRH-like peptide in amphibians has yet been identified, but the neuropeptide substance P released from sensory afferent terminals in mammals evokes a similar slow, long-lasting EPSP.
عصبی های پایانه ، همه نه اما ، برخی ، preganglionicدر سمپاتیک گانگلیون امریکایی بزرگ غوک آزاد ACHدر هورمون بازرد ) جسم هورمون پپتید ( LHRHکننده اعصاب . colocalizedمانند از باال فرکانس با شود preganglionicتحریک می باعث
فراخوانیکند، ، شده منتشر شود می پپتید عصبی EPSPکه های سلول در مدت حتی ( 6-49شکل ) postganglionicطوالنی ، الیاف توسط مستقیم طور به که . peptidergicinnervatedآنهایی نفوذ برای از بیش منتشر توجهی قابل فاصله باید پپتید نیست
. آهسته دور پذیرای عصبی های سلول شدن peptidergic EPSPدر بسته از همچنین ، کولینرژیک تحریک پتانسیل آهسته مانند ، های و- + Mکانال سدیم از باز و : . CA2نوع تحریکی + پتانسیل شماره از نتایج سلول peptidergicکانال پذیری تحریک تغییر
ورودی از شدید سازی فعال از پس طوالنی مدت برای خودکار گانگلیونی . preganglionicهای از پستانداران معادل بدونمانند پپتید اما LHRHاقدامات ، است شده شناسایی هنوز دوزیستان در Pدر حسی آوران های پایانه از شده آزاد نوروپپتید مواد
مدت طوالنی ، آهسته مشابه تداعی .EPSPپستانداران
Norepinephrine and Acetylcholine Are the Predominant Transmitters in the Autonomic Nervous System
Most postganglionic sympathetic neurons release norepinephrine, which acts on a variety of different adrenergic receptors. There are five major types of adrenergic receptors, and these are the target for several medically important drugs (Table 49-1).

خودکار عصبی سیستم در غالب انتقال به کولین استیل و نفرین نوراپی
سمپاتیک عصبی های سلول می postganglionicبیشتر عمل آدرنرژیک مختلف های گیرنده انواع در که نفرین، نوراپی آزادسازیجدول. ) مهم پزشکی دارو چند برای هدف این و دارد، وجود آدرنرژیک های گیرنده از عمده نوع پنج (.1-49کند
ATP and Adenosine Have Potent Extracellular Actions
Adenosine triphosphate (ATP) is an important cotransmitter with norepinephrine in many postganglionic sympathetic neurons. By acting on ATP-gated ion channels (P2 purinergic receptors), it is responsible for some of the fast responses seen in target tissues (Table 49-1). The proportion of ATP to norepinephrine varies considerably in different sympathetic nerves. The ATP component is relatively minor in nerves to blood vessels in the rat tail and rabbit ear, while the responses of guinea pig submucosal arterioles to sympathetic stimulation appear to be mediated solely by ATP.
The nucleotide adenosine is formed from the hydrolysis of ATP and is recognized by P1 purinergic receptors (Table 49-1) located both pre- and postjunctionally. It is thought to play a modulatory role in autonomic transmission, particularly in the sympathetic system. Adenosine may dampen sympathetic function after intense sympathetic activation by activating receptors on sympathetic nerve endings that inhibit further norepinephrine and ATP release. Adenosine also has inhibitory actions in cardiac and smooth muscle that tend to oppose the excitatory actions of norepinephrine.
ATP قوی سلولی خارج عملیات آیا آدنوزین و
( فسفات تری سمپاتیک ATP) cotransmitterآدنوزین عصبی سلولهای از بسیاری در نفرین نوراپی با postganglionicمهمیونی . های کانال در اقدام با های- ) ATPاست گیرنده می ( purinergic P2دردار دیده سریع های پاسخ از برخی مسئول را آن ،
جدول ) هدف بافت در نسبت ( . 1-49شود متفاوت ATPاست توجهی قابل بطور مختلف سمپاتیک اعصاب در نفرین نوراپی بهجزء. گینه ATPاست پاسخ که حالی در ، است جزئی نسبتا خرگوش گوش الله و موش دم در خونی های رگ به اعصاب در
توسط تنها رسد می نظر به سمپاتیک تحریک به خوک زیرمخاطی شود .ATPشریانی واسطه
هیدرولیز از که است نوکلئوتید های ATPآدنوزین گیرنده توسط و است شده دو ( 1-49جدول ) purinergic P1تشکیل هر واقعو . postjunctionallyقبل سیستم در خصوص به ، بازی خودکار انتقال در تعدیلی نقش این که میشود تصور است شده شناخته
سمپاتیک . عصب انتهای در گیرنده کردن فعال با شدید سمپاتیک شدن فعال از پس دلسوز عملکرد است ممکن آدنوزین سمپاتیکآزادی و بیشتر نفرین نوراپی مانع . ATPکه تمایل که صاف و قلب عضله در بازدارنده اقدامات همچنین آدنوزین کنید مرطوب کمی
نفرین . نوراپی از تحریکی اقدامات با مخالفت به
Many Different Neuropeptides Are Present in Autonomic Neurons
Neuropeptides are colocalized with norepinephrine and ACh in autonomic neurons. Cholinergic preganglionic neurons in the spinal cord and brain stem and their terminals in autonomic ganglia may contain enkephalins, neurotensin, somatostatin, or substance P. Noradrenergic postganglionic sympathetic neurons may also express a variety of neuropeptides. Neuropeptide Y is present in as many as 90% of the cells and modulates sympathetic transmission. In tissues in which the nerve endings are distant from their targets (more than 60 nm, as for the rabbit ear artery),
P.971
P.972neuropeptide Y potentiates both the purinergic and adrenergic components of the tissue response, probably by acting postsynaptically. In contrast, in tissues with dense sympathetic innervation and where the target is closer (20 nm, such as the vas deferens), neuropeptide Y acts presynaptically to inhibit release of ATP and norepinephrine, thus dampening the tissue response. The peptides galanin and dynorphin are often found with neuropeptide Y in sympathetic neurons, which can contain several neuropeptides. Cholinergic postganglionic sympathetic neurons commonly contain CGRP and vasoactive intestinal polypeptide (VIP) (Figure 49-7).
از نورون NEUROPEPTIDESبسیاری اتونوم در حاضر حال در است متفاوت می
NEUROPEPTIDES اتونوم عصبی های سلول در آه و نفرین نوراپی با در preganglionicنورون . colocalizedمی کولینرژیک ، انکفالین است ممکن اتونوم گانگلیون در را خود های پایانه و فقرات ستون ساقه مغز و ناف ، neurotensinبند
ماده یا ، سمپاتیک. Pسوماتوستاتین عصبی های سلول نوروپپتید postganglionicنورآدرنرژیک انواع است ممکن همچنین حاویneuropeptide . نوروپپتید از Yبیان بسیاری عنوان به حاضر حال است . 90در بافت در دلسوز انتقال تعدیل و است سلول از ٪
از ) بیش باشد می خود اهداف از عصبدور انتهای از آن در ( 60که ، خرگوش گوش عروق برای عنوان به ، نانومترP.971P.972
دو Yنوروپپتید هر اقدام purinergicباعث با احتماال بافت، پاسخ آدرنرژیک اجزای . postsynapticallyو بافت در مقابل، دربا است ) innervationهای تر نزدیک هدف به که جایی و متراکم ( 20دلسوز نوروپپتید ، است دفران مجرای مانند ، Yنانومتر
کند می شدن presynapticallyعمل آزاد . ATPمهار بافت پاسخ نیرو، تقلیل ترتیب این به ، نفرین نوراپی و galaninو پپتیدهاdynorphin نوروپپتید با نوروپپتید Yاغلب چندین تواند می که ، سمپاتیک عصبی های سلول .neuropeptideدر یافت حاوی
سمپاتیک عصبی های شامل postganglionicسلول معموال روده CGRPکولینرژیک پپتیدی پلی شکل )vasoactive ( VIP )و49-7. )
Table 49-1 Pharmacology of the Autonomic Nervous System
Receptor category Functional roles1 Drugs that act selectively at these receptors
Medical use

Norepinephrine
Adrenergic a1 Contractile effects of NE on smooth muscle, especially blood vessels, urogenital, and sphincter muscles
Prazosin (antagonist) Hypertension
Adrenergic a2 Presynaptic control (inhibitory) of release of NE, ATP, and ACh from nerve terminals
Yohimbine (antagonist) Delay ejaculation
Adrenergic b1 Stimulatory effects of NE and circulating epinephrine on heart
Atenolol (antagonist) Hypertension
Adrenergic b2 Relaxant effects of NE on smooth muscle in gastrointestinal tract, urinogenital system, and airways
Salbutamol (agonist) Bronchodilator for asthma
Adrenergic b3 Stimulate release of free fatty acids from adipose tissue
None Potential in obesity
Acetylcholine
Cholinergic-nicotinic (ganglionic type) Fast excitation of postganglionic neurons in autonomic ganglia
Hexamethonium (antagonist) Hypertension (formerly)
Cholinergic-muscarinic M1 Inhibit ACh and NE release from autonomic nerve terminals
Pirenzepine (antagonist) Anti-ulcerogenic
Cholinergic-muscarinic M2 Effects of ACh on heart and smooth muscle Atropine (nonselective antagonist) Mydriatic
Cholinergic-muscarinic M3 ACh-induced secretion from glandular tissues (eg, salivary gland)
Atropine (nonselective antagonist) Reduced drooling in Parkinson disease
Others
Purinergic P1 (Four subtypes) Modulatory effects of adenosine on autonomic effector tissues
Theophylline (antagonist) Bronchodilator
Purinergic P2 (Two subtypes) Fast and slow responses to ATP in smooth muscle
Few drugs; suramin is P2Y antagonist None
Nitric oxide (NO) Relaxant effects on smooth muscle, especially blood vessels
Glyceryl trinitrate and nitroprusside (generate NO)
Coronary vasodilators for angina
1ACh = acetylcholine; ATP = adenosine triphosphate; NE = norepinephrine.
Figure 49-7 A variety of neuropeptides coexist with norepinephrine and acetylcholine in neurons of the sympathetic ganglia, as shown here for the cat and the guinea pig. The sympathetic preganglionic nuclei extend from T1 to L3.MCG = WW middle cervical ganglion; WSCG = superior cervical ganglion; SG = stellate ganglion. (Adapted from Elfvin et al. 1993.)
نوروپپتید 7-49شکل سمپاتیک، neuropeptide انواع های عقده از عصبی سلولهای در کولین استیل و نفرین نوراپی با همزیستی . هسته است شده داده نشان اینجا در هندی خوکچه و گربه برای عنوان از preganglionic به L3.MCG = WW به T1 دلسوز

گسترش رحم گردن وسط رحم، = WSCG. گانگلیون گردن برتر . ) = SGگانگلیون از اقتباس ستاره همکاران Elfvin گانگلیون و1993.(
In postganglionic parasympathetic neurons that express VIP together with ACh, the peptide may contribute to the response of the target tissue because of its powerful vasodilator effects. For example, ACh triggers salivary gland secretion while VIP is responsible for the local increase in blood flow, which is important to the secretory response. Some of the complex modulatory functions that neuropeptides perform are illustrated in Figure 49-8.
پاراسمپاتیک عصبی های سلول بیان postganglionicدر با VIPکه دلیل ACHهمراه به هدف بافت پاسخ به است ممکن پپتید ، . مثال، عنوان به کند می کمک آن قدرتمند عروق گشادکننده که ACHاثرات حالی در بزاقی غدد ترشح افزایش VIPباعث مسئول
. نوروپپتید که است ای پیچیده تعدیلی توابع از برخی است مهم ترشحی های پاسخ به که خون، جریان در neuropeptideمحلیشکل در .8-49انجام است شده داده نشان
A Central Autonomic Network Coordinates Autonomic Function
Autonomic functions ultimately must be coordinated with one another and the ongoing behavioral needs of the individual. This coordination is carried out by a highly interconnected set of structures in the brain stem and forebrain that form a central autonomic network.
A key component of this network is the nucleus of the solitary tract. The nucleus receives visceral input from cranial nerves VII, IX, and X and then uses this information to modulate autonomic function in two ways (Figure 49-9).
خودکار مختصات مرکزی شبکه اتونومیک عملکرد
. مجموعه توسط که است هماهنگی این هماهنگ فرد از رفتاری انجام حال در نیازهای به و یکدیگر با باید نهایت در اتونوم عملکرد. است شده انجام مرکزی اتونوم شبکه یک تشکیل که است مغز و مغز ساقه در ساختار از پیوسته هم به بسیار ای
. جمجمه اعصاب از احشایی ورودی دریافت هسته است انفرادی دستگاه از هسته شبکه این از کلیدی جزء VIIیک ، IX وX سپس وشکل ) روش دو به خودکار تابع بم و زیر به اطالعات از استفاده (.9-49با
First, the nucleus of the solitary tract projects to neurons forming brain stem and spinal circuits that control simple autonomic responses. For example, visceral
P.973afferents relayed through the nucleus of the solitary tract directly regulate vagal motor control of the stomach and heart rate. Other outputs from the nucleus of the solitary tract innervate neurons in the ventrolateral medullary reticular formation and control blood pressure by regulating the blood flow in different vascular beds (Figure 49-10).
Second, the nucleus of the solitary tract acts to integrate autonomic function with more complex endocrine and behavioral responses, a process in which the hypothalamus, which we will consider below, plays an important role.
ساده پاسخ که است نخاعی مدارهای و مغز ساقه تشکیل عصبی های سلول به انفرادی های دستگاه های پروژه از هسته نخست، . احشایی مثال، عنوان به کنترل خودکار
P.973 . خروجی دیگر قلب و معده نرخ از واگ موتور کنترل تنظیم مستقیم طور به انفرادی دستگاه از هسته طریق از رله آوران اعصاب
خون فشار کنترل و گیری شکل مشبک مدوال در دستگاه کردن پی دارای نورون انفرادی از هسته تنظیم ventrolateralاز باشکل ) مختلف عروق بستر در خون (.10-49جریان
یک تر، پیچیده های پاسخ رفتاری و ریز درون غدد با خودکار عملکرد از استفاده به کند می عمل انفرادی دستگاه از هسته دوم،. کند می ایفا را مهمی نقش گرفت، خواهد نظر زیر را آن ما که هیپوتاالموس، آن در که است فرایند
The visceral sensory outflow from the nucleus of the solitary tract is relayed to the forebrain by the parabrachial nucleus, which is important for the behavioral responses to taste and other visceral sensations. Lesions of the parabrachial nucleus prevent conditioned behavioral responses resulting from gustatory cues. The parabrachial nucleus surrounds the superior cerebellar peduncle in the upper pons and provides inputs to the hypothalamus, the periaqueductal gray matter, the amygdaloid complex, the visceral sensory thalamus, and the cortex. In turn, the parabrachial nucleus receives descending connections from these regions.
The periaqueductal gray matter surrounds the cerebral aqueduct in the midbrain. It receives inputs from the nucleus of the solitary tract, the parabrachial nucleus, and the hypothalamus and projects to the medullary reticular formation, where it produces behaviorally coordinated patterns of autonomic response. For example, during a “fight-or-flight” response, the periaqueductal gray matter redirects blood flow from internal organs to the hind limbs to support running behavior.
هسته توسط انفرادی دستگاه از هسته از احشایی حسی خروج احشایی parabrachialاز حس و طعم به رفتاری پاسخ برای که ، . هسته ضایعات میرساند مغز جلو به است جلوگیری parabrachialمهم چشایی های نشانه از ناشی مشروط رفتاری های پاسخ
. هسته کند ، parabrachialمی هیپوتاالموس به کند می فراهم ورودی و فوقانی پل در برتر مخچه ساقه است کرده احاطه راخاکستری پیچیده periaqueductalماده ،amygdaloid . هسته خود، نوبه به قشر و ، احشایی حسی تاالموس ،parabrachial
مناطق . این از اتصاالت نزولی دریافت

خاکستری . periaqueductalماده هسته ، انفرادی دستگاه از هسته از این میانی مغز در مغزی قنات کرده احاطه راparabrachial پاسخ از هماهنگ رفتاری الگوهای تولید به را آن که جایی ، مشبک مغز گیری شکل به پروژه و هیپوتاالموس و ،
خاکستری . " " ماده پاسخ، گریز و جنگ یک طول در مثال، عنوان به ورودی دریافت جریان periaqueductalخودکار مسیر تغییراجرا . حال در رفتار از حمایت برای عقبی اندام به داخلی اندامهای از خون
The amygdaloid complex plays a key role in regulation of the autonomic components of conditioned behavioral responses. Inputs to the amygdala from areas of the cortex and the thalamus concerned with behavior enter the lateral and basal nuclei, whereas the central nucleus receives inputs from the central autonomic system. Complex internal circuits allow the amygdala to associate autonomic responses with specific behaviors. For example, as we shall learn in the chapter on emotion (Chapter 50), when a rat learns that an auditory cue is followed by an electric shock, the auditory cue itself eventually produces an elevation of heart rate and behavioral freezing previously associated with the shock. Lesions of the central nucleus of the amygdala, which projects to the hypothalamus and the medulla, prevent this response.
مجموعه . amygdaloidاین آمیگدال به ورودی کند می بازی شرطی رفتاری های پاسخ از خودکار قطعات تنظیم در کلیدی نقشورودی مرکزی هسته که حالی در قاعده، و جانبی های هسته کردن وارد با رفتار با رابطه در تاالموس و کورتکس مناطق از
. رفتارهای با خودکار های پاسخ ارتباط به آمیگدال دهد می اجازه مجتمع داخلی مدارات مرکزی اتونوم سیستم از شده دریافتفصل. ) احساسات در فصل این در باید ما که حالی در مثال، عنوان به که( 50خاص گیرد می یاد موش یک که هنگامی یادگیری ،
با قبال که رفتاری انجماد و قلب ضربان ارتفاع تولید نهایت در را خود شنوایی نشانه بعد، الکتریکی شوک توسط شنوایی نشانه. . پاسخ این از جلوگیری مغز، و هیپوتاالموس به پروژه که آمیگدال، مرکزی هسته ضایعات شوک وجود
Visceral sensory areas of the thalamus and the visceral sensory cortex both receive visceral sensory afferents directly from the parabrachial nucleus. In primates, the taste component of the nucleus of the solitary tract also projects to the thalamus, thus providing an even more direct relay of taste information to consciousness. The visceral sensory thalamic areas are located in a small-celled nucleus adjacent to the ventroposterior (somatic sensory) complex, the ventroposterior parvocellular nucleus. This thalamic nucleus relays taste and other visceral
P.974sensations (hunger pangs, abdominal fullness, breath-holding sensations) to the anterior insular cortex, where there is a topographic map of the internal organ systems. Taste is located most anteriorly in the insular cortex, with gastrointestinal, then cardiorespiratory sensations, more posteriorly.
هسته از مستقیم طور به احشایی حسی آوران اعصاب دو هر احشایی حسی کورتکس و تاالموس از احشایی حسی مناطقparabrachial . نتیجه در و نموده پروژه تاالموس به نیز انفرادی دستگاه از هسته طعم جزء پستانداران، در کرد خواهید دریافت
. به مجاور سلولی کوچک هسته یک در احشایی حسی تاالموس مناطق آگاهی به مزه و طعم اطالعات از بیشتر حتی مستقیم رلهventroposterior ) هسته ) پیچیده، حسی . parvocellular ventroposteriorجسمی تاالموس هسته رله این است شده واقع
احشایی و طعم
P.974 ) از ) توپوگرافی نقشه یک آن در که قدامی، منزوی قشر به برگزاری نفس احساس شکم، پری احساس گرسنگی، چنگال احساس
. قلبی احساس آن از پس گوارش، دستگاه با است، شده واقع منزوی قشر در قدام به که است طعم دارد وجود بدن داخلی اعضاء. است خلف تنفسی،

Figure 49-8 Complex modulatory functions of neuropeptides. Sensory neurons in the wall of the colon that excite sympathetic ganglion cells in the inferior mesenteric ganglion (IMG) use several neuropeptides along with acetylcholine (ACh) as transmitters. The neurons that contain cholecystokinin and the vasoactive intestinal polypeptide (VIP) are mechanosensory. Other mechanosensory fibers originate in spinal ganglion cells and contain substance P (SP). They provide excitatory synapses to the sympathetic ganglion cells. Cholinergic fibers originating in the preganglionic spinal cord nuclei contain either enkephalins (ENK) or neurotensin (NT). The cholinergic neurons form an excitatory input to the ganglion cells. The enkephalin pathway inhibits release of ACh and SP, whereas the NT pathway facilitates release of SP and gives rise to an excitatory potential in some IMG neurons. Excitatory neurotransmitters and peptides are indicated in orange; inhibitory in gray. NA = noradrenaline; NE = norepinephrine; NPY = neuropeptide Y; SOM = somatostatin. (Adapted from Elfvin et al. 1993.)
نوروپپتید 8-49شکل مجتمع تعدیلی . neuropeptideتوابع روده دیواره در حسی های نورونتحتانی ) مزانتریک گانگلیونی در دلسوز گانگلیونی های سلول تحریک که از ( IMGبزرگ استفاده
نوروپپتید ) neuropeptideچند کولین استیل با . ACHهمراه ) های سلول فرستنده عنوان بهحاوی که روده cholecystokininعصبی پپتیدی پلی vasoactiveو ( VIP )
mechanosensory . الیاف دیگر باشد نخاع mechanosensoryمی گانگلیونی های سلول درشامل و الیاف ( . . SPماده ) Pمنشا دلسوز گانگلیونی های سلول به ارائه تحریکی سیناپس آنها
نخاعی طناب هسته در آن منشاء انکفالین ) preganglionicکولینرژیک دو هر یا ( ENKشامل وneurotensin ( NT ) . گانگلیونیتشکیل های سلول به تحریکی ورودی یک کولینرژیک های نورون
. مسیر دهند انتشار enkephalinمی مسیر SPو ACHمهار که حالی در ،NT انتشار SPتسهیلعصبی های سلول از برخی در تحریکی پتانسیل یک به دهد می افزایش . IMGو عصبی ناقالن
خاکستری . . رنگ به مهاری داد نشان پرتقال در پپتیدها و نوراپی = NEنورآدرنالین = . NAتحریکاز = . ) SOM؛ Yنوروپپتید = NPYنفرین . اقتباس همکاران Elfvinسوماتوستاتین (.1993و

Figure 49-9 Pathways that distribute visceral sensory information in the brain. Visceral afferent information (solid line) enters the brain through the nucleus of the solitary tract. It is distributed to preganglionic neurons, to an area in the ventrolateral medulla that coordinates autonomic and respiratory reflexes, and via an ascending pathway to the forebrain. Less direct inputs (dotted line) relayed from the parabrachial nucleus bring visceral sensory information to the hypothalamus, the amygdala, the septum (not shown), the cortex, and the periaqueductal gray matter.
( 9-49شکل ( . از هسته طریق از مغز وارد جامد خط احشایی آوران اطالعات مغز در احشایی حسی اطالعات توزیع که مسیرهای . عصبی های سلول به که است این انفرادی مدوال preganglionicدستگاه در منطقه به شده، به ventrolateralتوزیع که
) ( . از رله خال خال خط مستقیم کمتر ورودی مغز جلو به صعودی مسیر یک طریق از و تنفسی، و اتونوم های رفلکس هماهنگی( parabrachialهسته ( ماده و قشر، ، است نشده داده نشان سپتوم آمیگدال، هیپوتاالموس، به را احشایی حسی اطالعات
.periaqueductalخاکستری
The visceral sensory cortex interacts with a portion of the anterior tip of the cingulate cortex, called the infralimbic area, which is a visceral motor region. Electrical or chemical stimulation here can cause gastric contractions or changes in blood pressure. Both the anterior insular and infralimbic areas project to the amygdala, hypothalamus, periaqueductal gray matter, parabrachial nucleus, nucleus of the solitary tract, and medullary reticular formation. Lesions of the visceral sensory cortex cause loss of conscious appreciation of visceral sensation such as taste. The visceral motor cortex is part of a region of cingulate cortex in which injury causes abulia, a condition in which patients fail to show emotional reactions to external stimuli.
منطقه نام به کمربندی، قشر قدامی نوک از بخشی با تعامل احشایی حسی .infralimbicقشر است احشایی حرکتی منطقه ، . و منزوی مناطق از دو هر شود خون فشار در تغییر یا و معده انقباضات توانید می اینجا در شیمیایی یا الکتریکی تحریک
infralimbic خاکستری ماده هیپوتاالموس، آمیگداال، به پروژه هسته periaqueductalقدامی ،parabrachial دستگاه از هسته ، . و طعم مانند احشایی حس آگاه قدردانی دادن دست از علت قشر حسی احشایی ضایعات مدوالری رتیکوالر تشکیل و انفرادی،در. که است بیماری یک اراده، ضعف باعث آسیب آن در که است کمربندی قشر از منطقه یک از بخشی احشایی حرکتی قشر مزه
. است خارجی های محرک به عاطفی العمل عکس دهد می نشان به قادر افراد آن

The Hypothalamus Integrates Autonomic and Endocrine Functions With Behavior
The hypothalamus plays a particularly important role in regulating the autonomic nervous system and was once referred to as the “head ganglion” of the autonomic nervous system. But recent studies of hypothalamic function have led to a somewhat different view. Whereas early studies found that electrical stimulation or lesions in the hypothalamus can profoundly affect autonomic function, more recent investigations have demonstrated that many of these effects are due to involvement of descending and ascending pathways of the cerebral cortex or the basal forebrain passing through the hypothalamus. Modern studies indicate that the hypothalamus functions to integrate autonomic response and endocrine function with behavior, especially behavior concerned with the basic homeostatic requirements of everyday life.
رفتار با توابع ریز درون غدد و خودکار قدرتمندترین هیپوتاالموس
" عصبی " سیستم از سر عقده عنوان به بار یک و کند می بازی خودکار عصبی سیستم تنظیم در مهمی ویژه به نقش هیپوتاالموس . . مطالعات که حالی در است شده منجر متفاوت حدودی تا نظر به هیپوتاالموس عملکرد از اخیر مطالعات اما شد ارجاع خودکار
اخیر تحقیقات اتونومیک، عملکرد بر تواند می شدت به هیپوتاالموس در ضایعات یا الکتریکی تحریک که است داده نشان اولیههیپوتاالموس از عبور پایه مغز جلو یا مغز قشر از مسیر صعودی و نزولی دخالت دلیل به اثرات این از بسیاری که اند داده نشان
. به رفتار، با ریز درون غدد عملکرد و خودکار پاسخ ادغام به هیپوتاالموس عملکرد که دهد می نشان اخیر مطالعات باشد می. است روزمره زندگی از فرمول پایه الزامات با رابطه در رفتار خصوص
It controls blood pressure and electrolyte composition by a set of regulatory mechanisms that range from control of drinking and salt appetite to the maintenance of blood osmolality and vasomotor tone.
It regulates body temperature by means of activities ranging from control of metabolic thermogenesis to behaviors such as seeking a warmer or cooler environment.
P.975
It controls energy metabolism by regulating feeding, digestion, and metabolic rate.
It regulates reproduction through hormonal control of mating, pregnancy, and lactation.
It controls emergency responses to stress, including physical and immunological responses to stress by regulating blood flow to muscle and other tissues and the secretion of adrenal stress hormones.
کنترل• از که است قانونی های مکانیسم از ای مجموعه توسط الکترولیت ترکیب و خون فشار. کند می کنترل محدوده وازوموتور تن و خون اسمواللیته حفظ به اشتها نمک و آشامیدنی آب
رفتارهای• به بدن ساز و سوخت حرارت تولید از کنترل از مختلف های فعالیت از استفاده با این. بدن حرارت درجه تنظیم را سرد و گرم محیط یک جستجوی مانند
P.975
. ساز• و سوخت و هضم تغذیه، تنظیم با کنترل را انرژی متابولیسم این
. کند• می تنظیم شیردهی و بارداری گیری، جفت از هورمونی کنترل طریق از مثل تولید این
تنظیم• با استرس به ایمنی و فیزیکی های واکنش جمله از استرس، به اضطراری های پاسخ. کند می کنترل این آدرنال استرس های هورمون ترشح و ها بافت سایر و عضالت به خون جریان

Figure 49-10 Pathways that control autonomic responses. Direct outputs to autonomic preganglionic neurons (solid line) arise from the paraventricular and lateral hypothalamus, the parabrachial nucleus, the nucleus of the solitary tract, certain monoamine groups such as the A5 noradrenergic neurons (not shown), serotonergic raphe neurons (not shown), and adrenergic neurons in the ventrolateral medulla. Less direct outputs from the cerebral cortex, amygdala, and periaquaductal gray matter (dotted lines) are relayed into the cell groups with direct input to the preganglionic inputs. Nearly all of the cell groups illustrated in these drawings are also connected with one another, forming a central autonomic network.
. 10-49شکل عصبی های سلول به مستقیم خروجی کنترل خودکار های پاسخ که ( preganglionic مسیرهای از ) جامد خط خودکارparaventricular هسته هیپوتاالموس، جانبی مانند parabrachial و خاص مونوآمین های گروه انفرادی، دستگاه از هسته ،
نورآدرنرژیک های ( (A5 نورون ( ) سلول و ، آیند می بوجود است نشده داده نشان سروتونرژیک رافه نورون ، است نشده داده نشانمدوال در آدرنرژیک عصبی ماده . ventrolateral های و آمیگدال، مغز، قشر از کمتر مستقیم های periaquaductal خروجی
) ( ورودی به مستقیم ورودی با همراه گروه به نقطه خطوط سلولی. preganglionic خاکستری های گروه تمام تقریبا میرساندمرکزی اتونوم شبکه یک تشکیل اند، متصل دیگر یک با نیز نقشه این در شده داده .نشان

The hypothalamus regulates these basic life processes by recourse to three main mechanisms. First, the hypothalamus has access to sensory information from virtually the entire body. It receives direct inputs from the visceral sensory system and the olfactory system, as well as the retina. The visual inputs are used by the suprachiasmatic nucleus to synchronize the internal clock mechanism to the day-night cycle in the external world (Chapter 3). Visceral somatosensory inputs carrying information about pain are relayed to the hypothalamus from the spinal and trigeminal dorsal horn (Chapters 23 and 24). In addition, the hypothalamus has internal sensory neurons that are responsive to changes in local temperature, osmolality, glucose, and sodium, to name a few examples. Finally, circulating hormones such as angiotensin II and leptin enter the hypothalamus at specialized zones along the margins of the third ventricle called circumventricular organs, where they interact directly with hypothalamic neurons.
. اول، کند می تنظیم اصلی مکانیسم سه به توسل با زندگی اساسی فرآیندهای این هیپوتاالموس . از مستقیم ورودی این است بدن کل تقریبا از است حسی اطالعات به دسترسی هیپوتاالموس
. های ورودی کند می دریافت چشم شبکیه همچنین و ، بویایی سیستم و احشایی حسی های سیستمهسته توسط مکانیزم suprachiasmatic تصویری سازی همزمان برای را شود می استفاده
( فصل خارجی جهان در شب روز، در چرخه به داخلی . 3ساعت ) somatosensory ورودی ( فصل قلو سه عصب و نخاع خلفی شاخ از درد مورد در اطالعات حمل ( 24و 23احشایی به
. به پاسخ در که حسی داخلی های نورون است هیپوتاالموس این، بر عالوه میرساند هیپوتاالموس . نهایت، در نمونه چند نام به هستند، سدیم و قند ، اسمواللیته محلی، حرارت درجه تغییرات
آنژیوتانسین مانند خون گردش در اندام II هورمون نام به سوم بطن از حاشیه امتداد در لپتین وcircumventricular وارد مستقیم طور هیپوتاالموس عصبی های سلول با آنها که جایی ،
تخصصی مناطق در .هیپوتاالموسSecond, the hypothalamus compares sensory information with biological set points. It compares, for example, local temperature in the preoptic area to the set point of 37°C and, if the hypothalamus is warm, activates mechanisms for heat dissipation. There are set points for a wide variety of physiological processes, including blood sugar, sodium, osmolality, and hormone levels.
. به مقایسه، این است بیولوژیکی ای مجموعه نقاط با مقایسه حسی اطالعات هیپوتاالموس دوم،منطقه در محلی دمای مثال، از preopticعنوان ای مجموعه نقطه ° 37به C اگر و ،
. طیف برای ای مجموعه نقاط حرارت اتالف برای مکانیزم شدن فعال است، گرم هیپوتاالموسها هورمون سطح و اسمواللیته سدیم، خون، قند جمله از فیزیولوژیکی، فرایندهای از ای گسترده
. دارد وجود
Finally, when the hypothalamus detects a deviation from a set point, it adjusts an array of autonomic, endocrine, and behavioral responses to restore homeostasis. If the body is too warm, the hypothalamus shifts blood flow from deep to cutaneous vascular beds and increases sweating, to increase heat loss through the skin. It increases vasopressin secretion, to conserve water for sweating. Meanwhile, the hypothalamus activates coordinated behaviors, such as seeking to change the local ambient temperature or seeking a cooler environment.
آرایه تنظیم را آن تشخیص، ای مجموعه نظر نقطه از انحراف هیپوتاالموس که زمانی نهایت، در . گرم، خیلی بدن اگر تعادل بازگرداندن برای رفتاری های واکنش و ریز درون غدد خودکار، از ای
از افزایش تعریق، افزایش و پوستی عروق بستر به عمق از را خون جریان تغییر هیپوتاالموس . برای آب حفظ برای ادراری، ضد هورمون ترشح افزایش این پوست طریق از گرما دادن دست
. حرارت درجه تغییر دنبال به جمله از هماهنگ، رفتار فعال هیپوتاالموس حال، همین در کردن عرق. است تر خنک محیط یک دنبال به یا و محلی محیط


Figure 49-11 The structure of the hypothalamus.A. Frontal view of the hypothalamus (section along the plane shown in part B).B. A medial view shows most of the main nuclei. The hypothalamus is often divided analytically into three areas in a rostocaudal direction: the preoptic area, the tuberal level, and the posterior level.
.11-49شکل هیپوتاالموس ساختاربخش. ) Aمشاهده در شده داده نشان صفحه امتداد در ها بخش هیپوتاالموس از (.Bنما
B . منطقه. دهد می نشان را داخلی نمایی اصلی های هسته از سطح preopticبسیاری ،tuberal : هیپوتاالموس خلفی سطح و ، جهت یک در قسمت سه به را تحلیلی اغلب که .rostocaudalاست است شده تقسیم
All of these processes must be precisely coordinated. For example, adjustments in blood flow in different vascular beds are important for such diverse activities as thermoregulation, digestion, response to emergency, and sexual intercourse. In order to do this, the hypothalamus contains an array of specialized cell groups with different functional roles.
P.976
P.977 . های فعالیت چنین برای مختلف عروق بستر در خون جریان تنظیم مثال، عنوان به است شده هماهنگ دقت به باید فرآیند این تمام
. هیپوتاالموس کار، این انجام منظور به است مهم جنسی رابطه و اورژانس به پاسخ هضم، حرارت، تنظیم عنوان به گوناگون. کاربردی مختلف های نقش با تخصصی های گروه از ای مجموعه شامل
P.976
P.977
The Hypothalamus Contains Specialized Groups of Neurons Clustered in Nuclei
Although the hypothalamus is very small, occupying only about 4 grams of the total 1400 grams of adult human brain weight, it is packed with a complex array of cell groups and fiber pathways (Figure 49-11). The hypothalamus can be divided into three regions: anterior, middle, and posterior. The most anterior part of the hypothalamus, overlying the optic chiasm, is the preoptic area. The preoptic nuclei, which include the circadian pacemaker (suprachiasmatic nucleus), are mainly concerned with integration of different kinds of sensory information needed to judge deviation from physiological set point. The preoptic area controls blood pressure and composition; cycles of activity, body temperature, and many hormones; and reproductive activity.
هسته در خوشه نورون تخصصی های گروه شامل هیپوتاالموس
حدود در تنها اشغال است، کوچک بسیار هیپوتاالموس چه کل 4اگر از مجموعه 1400گرم یک با را آن انسان، مغز بالغ وزن گرمشکل ) فیبر مسیرهای و های گروه از ای : 11-49پیچیده . سه( به توان می را هیپوتاالموس خلفی و میانی قدامی، شده بندی بسته
. ی اندازه از بیش هیپوتاالموس، از بیشتر قدامی بخش است شده تقسیم منطقه chiasmمنطقه . preopticنوری، هسته استpreoptic ( هسته روزی شبانه ساز ضربان شامل که ،suprachiasmatic )حسی اطالعات مختلف انواع ادغام با عمده طور به ،
. منطقه و؛ باروری های فعالیت است کرده نگران فیزیولوژیکی مجموعه نقطه از انحراف قضاوت برای نیاز فشار preopticمورد. ها هورمون از بسیاری و بدن، حرارت درجه فعالیت، چرخه کنترل، ترکیب و خون
The middle third of the hypothalamus, overlying the pituitary stalk, contains the dorsomedial, ventromedial, paraventricular, supraoptic, and arcuate nuclei. The paraventricular nucleus includes both magnocellular and parvocellular neuroendocrine components controlling the posterior and anterior pituitary gland. In addition, it contains neurons that innervate both the parasympathetic and sympathetic preganglionic neurons in the medulla and the spinal cord, thus playing a major role also in regulating autonomic responses. The arcuate and periventricular nuclei, along the wall of the third ventricle, like the paraventricular nucleus contain parvocellular neuroendocrine neurons, whereas the supraoptic nucleus contains additional magnocellular neuroendocrine cells. The ventromedial and dorsomedial nuclei project mainly locally within the hypothalamus and to the periaqueductal gray matter, to regulate complex integrative functions such as control of growth, feeding, maturation, and reproduction.
Finally, the posterior third of the hypothalamus includes the mammillary body and the overlying posterior hypothalamic area. In addition to the mammillary nuclei, whose function remains enigmatic, this region includes the tuberomammillary nucleus, a histaminergic cell group that is important in regulating wakefulness and arousal.

The major nuclei of the hypothalamus are located for the most part in the medial part of the hypothalamus, sandwiched between two major fiber systems. A massive longitudinal fiber pathway, the medial forebrain bundle, runs through the lateral hypothalamus. The medial forebrain bundle connects the hypothalamus with the brain stem below, and with the basal forebrain, amygdala, and cerebral cortex above. Large neurons scattered among the fibers of the medial forebrain bundle provide long-ranging hypothalamic outputs that reach from the cerebral cortex to the sacral spinal cord. They are involved in organizing behaviors as well as autonomic responses.
شامل ، هیپوفیز ساقه پوشاننده ، هیپوتاالموس میانی سوم dorsomedialیک ، ventromedial ، paraventricular ، supraoptic . هسته قوسی های هسته و ،paraventricular دو هر غدد magnocellularشامل و اعصاب اجزای و
parvocellular . نورون دو هر که است عصبی های سلول حاوی را آن ، این بر عالوه قدامی هیپوفیز غده و خلفی کنترلpreganglionic های پاسخ تنظیم در نیز بازی مهمی نقش نتیجه در ، کردن پی دارای نخاع و مغز در سمپاتیک و پاراسمپاتیک
. هسته مانند ، سوم بطن از دیوار امتداد در ، بطن اطراف های هسته و کمانی های paraventricularخودکار سلول حاویعصبی غددی هسته parvocellularعصبی که حالی در ،supraoptic غدد و اعصاب های سلول magnocellularشامل
پروژه. هسته ventromedialاضافی خاکستری dorsomedialو ماده به و هیپوتاالموس در محلی صورت به عمدتاperiaqueductal. مثل تولید و بلوغ تغذیه، ، رشد کنترل مانند یکپارچه پیچیده توابع تنظیم برای ،
بدن شامل هیپوتاالموس خلفی سوم نهایت، . mammillaryدر هسته بر عالوه پوشاننده خلفی هیپوتاالموس منطقه وmammillary هسته شامل منطقه این است، مانده باقی مبهم تابع که ،tuberomammillary گیرنده های سلول گروه یک ،
که . است مهم برانگیختگی و بیداری تنظیم در هیستامینی هایشده واقع فیبر عمده سیستم دو بین ساندویچ ، هیپوتاالموس میانی قسمت در ها قسمت بیشتر در ها هیپوتاالموس اصلی هسته
افزاری. . نرم بسته جانبی هیپوتاالموس طریق از شود می اجرا ، میانی مغز افزاری نرم بسته ، طولی فیبر گسترده مسیر است . میان در پراکنده بزرگ های نورون کنید مخ قشر و آمیگدال ، بازال مغز جلو با و ، زیر به مغز ساقه متصل هیپوتاالموس میانی مغز
. در آنها برسد خاجی نخاع به مغز قشر از که هیپوتاالموس خروجی تا گرفته طوالنی فراهم داخلی مغز افزاری نرم بسته از الیافدرگیر . خودکار های پاسخ همچنین و رفتار سازماندهی

Figure 49-12 The hypothalamus controls the pituitary gland both directly and indirectly through hormone-releasing neurons. Peptidergic neurons (5) release oxytocin or vasopressin into the general circulation through the posterior pituitary. Two general types of neurons are involved in regulation of the anterior pituitary. Peptidergic neurons (3, 4) synthesize and release hormones into the hypophyseal-portal circulation. The second type of neuron is the link between the peptidergic neurons and the rest of the brain. These neurons, some of which are monoaminergic, are believed to form synapses with peptidergic neurons either on the cell body (1) or on the axon terminal (2).
.12-49شکل آزاد هورمون عصبی های سلول طریق از مستقیم غیر و مستقیم طور به دو هر هیپوفیز غده کنترل هیپوتاالموس. Peptidergic (5)نورون از کلی نوع دو خلفی هیپوفیز طریق از کلی طور به خون گردش به را وازوپرسین و توسین اکسی آزاد
. نورون درگیر قدامی هیپوفیز تنظیم در ها Peptidergic (3نورون ، 4) . - نوع پورتال هیپوفیز خون گردش به هورمون انتشار و تولیدعصبی های سلول بین ارتباط نورون از . peptidergicدوم آنها از برخی که عصبی، های سلول این است مغز استراحت و
monoaminergic نورون با سیناپس تشکیل به اعتقاد ،peptidergic( سلولی جسم در آکسون( )1یا ترمینال در یا (.2و
A second, smaller fiber system is located medial to the major hypothalamic nuclei, in the wall of the third
P.978ventricle. This periventricular fiber system contains longitudinal fibers that link the hypothalamus to the periaqueductal gray matter in the midbrain. This pathway is thought to be important in activating simple, stereotyped behavioral patterns, such as posturing during sexual behavior. The periventricular system also conveys the axons of the parvocellular neuroendocrine neurons located in the periventricular region, and including the paraventricular and arcuate nuclei, to the median eminence, for control of the anterior pituitary gland. They are met in the median eminence by the axons from the magnocellular neurons, which continue down the pituitary stalk to the posterior pituitary gland.
سوم دیوار در هیپوتاالموس، اصلی هسته به گرفته قرار داخلی است کوچکتر فیبر سیستم دوم،
P.978خاکستری. ماده به هیپوتاالموس که طولی الیاف شامل بطن اطراف فیبر سیستم این بطن
periaqueductal . شدن، فعال در مهم مسیر این شود می تصور دارند پیوند میانی مغز در . منتقل نیز بطن اطراف سیستم جنسی رفتار در گیری موضع مانند ساده، کلیشه رفتاری الگوهای
عصبی غددی عصبی های سلول از از parvocellularآکسون و بطن، اطراف منطقه در واقعمتوسط paraventricularجمله برتری به هسته، کمانی . و آنها قدامی هیپوفیز غده کنترل برای ،
متوسط مقام نورون در از آکسون غده magnocellularتوسط به هیپوفیز ساقه ادامه که ، . کرد مالقات خلفی هیپوفیز

Figure 49-13 The paraventricular nucleus of the hypothalamus is a microcosm of the autonomic and endocrine control systems. Two distinct populations of magnocellular neuroendocrine neurons produce oxytocin or vasopressin, which are secreted into the bloodstream in the posterior pituitary gland. Parvocellular neuroendocrine neurons in the medial paraventricular nucleus (medial parvocellular neuroendocrine neurons) contain hypothalamic releasing hormones, such as corticotropin-releasing hormone and release-inhibiting hormones, such as dopamine and somatostatin. Their axons project to the median eminence, where they release their hormones into the hypophysial portal circulation to control the anterior pituitary gland. Dorsal, ventral, and lateral (not shown) parvocellular neurons project to the preganglionic cell groups in the medulla and the spinal cord, as well as to other autonomic control nuclei in the brain stem. They use mainly oxytocin and vasopressin as neuromodulators. However, this is a completely distinct population from the magnocellular oxytocin and vasopressin neurons, as few if any cells send axons to both the posterior pituitary and the brain stem.
دو . paraventricularهسته 13-49شکل است ریز درون غدد کنترل و خودکار های سیستم از کوچک جهان یک هیپوتاالموس ازعصبی غددی عصبی های سلول از متمایز غده magnocellularجمعیت در خون جریان به که ، وازوپرسین و توسین اکسی تولید
. عصبی غددی عصبی های سلول شود می ترشح خلفی هسته Parvocellularهیپوفیز ) paraventricularدر داخلی داخلیعصبی غددی عصبی های ( parvocellularسلول کننده آزاد هورمون مانند ، هیپوتاالموس کننده آزاد هورمون حاوی
متوسط . برتری به پروژه آنها آکسون سوماتوستاتین و دوپامین مانند ، هورمون مهارکننده انتشار و آنها کورتیکوتروپین که جایی ، پورتال خون گردش به را خود های . hypophysialهورمون به را جانبی و شکمی پشتی، قدامی هیپوفیز غده کنترل برای آزاد
های است ) (preganglionicگروه نشده داده نشان مغز ساقه در خودکار کنترل های هسته دیگر به همچنین و ، نخاع و مغز درنورون . parvocellularپروژه عنوان به وازوپرسین و توسین اکسی عمده طور به استفاده . neuromodulatorsآنها این با
وازوپرسین و توسین اکسی از متمایز کامل طور به جمعیت این هر magnocellularحال، اگر است چند عنوان به ها، نورونمغز . ساقه و خلفی هیپوفیز غده دو هر به ارسال آکسون سلول
The Hypothalamus Controls the Endocrine System
The hypothalamus controls the endocrine system directly, by secreting neuroendocrine products into the general
P.979circulation from the posterior pituitary gland, and indirectly, by secreting regulatory hormones into the local portal circulation, which drains into the blood vessels of the anterior pituitary (Figure 49-12). These regulatory hormones control the synthesis and release of anterior pituitary hormones into the general circulation. The highly fenestrated (perforated) capillaries of the posterior pituitary and median eminence of the hypothalamus facilitate the entry of hormones into the general circulation or the portal plexus. Direct and indirect control form the basis of our modern understanding of hypothalamic control of endocrine activity.
ریز درون غدد سیستم کنترل هیپوتاالموس
کلی طور به غدد و اعصاب های فرآورده ترشح با مستقیم، طور به کنترل را ریز درون غدد سیستم هیپوتاالموس

P.979محلی، های پورتال خون گردش به را کننده تنظیم های هورمون ترشح با مستقیم، غیر طور به و خلفی، هیپوفیز غده از خون گردش
شکل ) قدامی هیپوفیز از خونی های رگ داخل به تخلیه های(. 12-49که هورمون آزادسازی و سنتز کنترل کننده تنظیم هورمون این ) ( . میانی برجستگی و خلفی هیپوفیز غده از ها مویرگ سوراخ سوراخدار بسیار عمومی خون گردش به را قدامی هیپوفیز
. دهند می تشکیل را مستقیم غیر و مستقیم کنترل پورتال شبکه یا عمومی خون گردش به را ها هورمون ورود تسهیل هیپوتاالموسریز درون غدد فعالیت هیپوتاالموس کنترل از ما مدرن درک .اساس
Magnocellular Neurons Secrete Oxytocin and Vasopressin Directly From the Posterior Pituitary
Large neurons in the paraventricular and supraoptic nuclei, constituting the magnocellular region of the hypothalamus, project to the posterior pituitary gland (neurohypophysis). Some of the magnocellular neuroendocrine neurons in the paraventricular and supraoptic nuclei release the neurohypophyseal hormone oxytocin, while others release vasopressin into the general circulation by way of the posterior pituitary (Figure 49-13). These peptides circulate to target organs of the body that control water balance and milk release.Oxytocin and vasopressin are peptides that contain nine amino acid residues (Table 49-2). Like other peptide hormones, they are cleaved from larger prohormones (see Chapter 15). The prohormones are synthesized in the cell body and cleaved within transport vesicles as they travel down the axons. The peptide neurophysin is a cleavage product of the processing of vasopressin and oxytocin and is released along with the hormone in the posterior pituitary. The neurophysin formed in neurons that release vasopressin differs somewhat from that produced in neurons that release oxytocin.
Magnocellular هیپوفیز خلفی از مستقیم طور به وازوپرسین و توسین اکسی ترشح عصبی های سلول
در بزرگ های هسته paraventricular نورون منطقه supraoptic و تشکیل ، magnocellular غده به پروژه ، هیپوتاالموس ازخلفی عصبی )neurohypophysis É. É ( هیپوفیز غددی عصبی های سلول از و paraventricular در magnocellular برخی
هورمون supraoptic هسته توسین اکسی طور neurohypophyseal آزاد به خون گردش به را وازوپرسین دیگران که حالی در ، شکل ) خلفی هیپوفیز طریق از . 13-49کلی آزادی ( و آب تعادل که بدن، های اندام دادن قرار هدف به گردش پپتیدهای این کند آزاد را
کنترل .شیر ( جدول آمینه اسید های مانده باقی نه شامل که پپتیدهای وازوپرسین و توسین . 2-49اکسی ) هورمونهای سایر مانند باشد می
از را آنها ، فصل ) prohormones پپتیدی شکاف ببینید 15بزرگتر در prohormones). را شکاف و شده سنتز سلولی جسم درآکسون کردن سفر عنوان به آنها نقل و حمل های کیسه اکسی neurophysin . درون و وازوپرسین پردازش برش محصول پپتید
شد منتشر خلفی هیپوفیز غده در هورمون با همراه و است آزاد neurophysin. توسین وازوپرسین که است عصبی های سلول دراست متفاوت ترشح توسین اکسی که است عصبی های سلول در شده تولید آن از حدودی تا شده .تشکیل
Parvocellular Neurons Secrete Peptides That Regulate Release of Anterior Pituitary Hormones
Geoffrey Harris proposed in the 1950s that the anterior pituitary gland is regulated indirectly by the hypothalamus. He demonstrated that the hypophysial portal veins, which carry blood from the hypothalamus to the anterior pituitary gland, convey important signals that control anterior pituitary secretion. In the 1970s the structure of a series of peptide hormones that carry these signals was elucidated. These hormones fall into two classes: releasing hormones and release-inhibiting hormones (Table 49-3). Of all the anterior pituitary hormones, only prolactin is under predominantly inhibitory control. Hence transection of the pituitary stalk causes insufficiency of adrenal cortex, thyroid, gonadal, and growth hormones, but increased prolactin secretion.
Systematic electrical recordings have not been made from neurons that secrete releasing hormones. However, they are believed to fire in bursts because of the pulsatile nature of secretion of the anterior pituitary hormones, which show periodic surges throughout the day. Episodic firing may be particularly effective for causing hormone release and may limit receptor inactivation.
Parvocellular ها هورمون هیپوفیز قدامی انتشار تنظیم پپتیدهای ترشح عصبی های سلول
Table 49-2 Hormones of the Posterior Pituitary GlandName Structure Function
Vasopressin H-Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-Gly-NH2 S-S
Vasoconstriction, water resorption by the kidney
Oxytocin H-Cys-Tyr-Ile-Glu-Asn-Cys-Pro-Leu-Gly-NH2 S-S
Uterine contraction and milk ejection

در هریس . 1950sجفری او شود می تنظیم هیپوتاالموس توسط مستقیم غیر طور به است قدامی هیپوفیز غده که کرد پیشنهادپورتال وریدهای که داد است hypophysial نشان مهم های سیگنال انتقال ، قدامی هیپوفیز غده به هیپوتاالموس از خون حمل که ،
. در قدامی هیپوفیز ترشح کنترل . 1970sکه این شد روشن سیگنال این حمل که پپتیدی های هورمون از سری یک از ساختار ( : جدول هورمون مهارکننده رها و آزاد هورمونها شوند می تقسیم دسته دو به ها . 3-49هورمون ) هیپوفیز های هورمون همه از
. ، تیروئید ، کلیوی فوق غده قشر نارسایی باعث هیپوفیز ساقه قطع رو این از است مهار عمدتا کنترل تحت پروالکتین تنها ، قدامیپروالکتین ترشح افزایش اما ، رشد های هورمون و ، جنسی .غدد
. به آنها ، حال این با است شده ساخته آزاد های هورمون ترشح که است عصبی های سلول از اند شده مند نظام الکتریکی ضبططول در دورهای افزایشهای موج دهد می نشان که ، قدامی هیپوفیز های هورمون ترشح ضربانی طبیعت دلیل به انفجار در آتش
می . محدود را گیرنده سازی فعال غیر است ممکن و موثر هورمون ترشح ایجاد برای خصوص به است ممکن ای دوره شلیک روز .کند
The neurons that make releasing hormones are found mainly along the wall of the third ventricle. The gonadotropin-releasing hormone (GnRH) neurons tend to be located most anteriorly, along the basal part of the third ventricle. Neurons that make somatostatin, corticotropin-releasing hormone (CRH), and dopamine are located more dorsally and are found in the medial part of the paraventricular nucleus. Neurons that make growth hormone-releasing hormone (GRH), thyrotropin-releasing hormone (TRH), GnRH, and dopamine are found in the arcuate nucleus, an expansion of the periventricular gray matter that overlies the median eminence, in the floor of the third ventricle (see Figure 49-10). The median eminence contains a plexus of fine capillary loops. These are fenestrated capillaries, and the terminals of the neurons that contain releasing hormones end on these loops. The blood then flows from the median eminence into a secondary (portal) venous system, which carries it to the anterior pituitary gland (See Figure 49-11).
آزادکننده . هورمون داشت بر در سوم بطن از دیوار امتداد در عمده طور به هورمون شدن آزاد باعث که است عصبی های سلولکه . )GNRH É ( گنادوتروپین است عصبی های سلول سوم بطن قاعده امتداد در ، جلو بیشتر دارد قرار به تمایل عصبی های سلول
هورمون کورتیکوتروپین کننده آزاد ، )CRH É ( سوماتوستاتین هسته داخلی قسمت در و دارد قرار تر پشتی هستند دوپامین و ، paraventricular . رشد هورمون کننده آزاد هورمون که است عصبی های سلول داشت بر تیروئید )GRH É ( در محرک هورمون ،
آزاد گذاری )TRH É ( هورمون تخمک محرک ، GnRH که خاکستری بطن اطراف به توجه ،گسترش کمانی هسته در دوپامین و ، overlies متوسط افتخاری ) از شکل بطن سوم طبقه در که ،49-10 . ) متوسط که افتخاری های از حلقه از ای شبکه شامل
. . یابد پایان حلقه این در آزاد های هورمون حاوی که ها نورون های ترمینال و ، باشد می سوراخدار مویرگ این است خوب مویرگیمتوسط مقام از آن از پس شکل ) ( ) خون حمل قدامی هیپوفیز غده به را آن که ، ثانویه وریدی سیستم پورتال یک به -49جریان
11. ( P.980
An Overall View
Table 49-3 Hypothalamic Substances That Release or Inhibit the Release of Anterior Pituitary Hormones
Hypothalamic substance Anterior pituitary hormone
Releasing
Thyrotropin-releasing hormone (TRH) Thyrotropin, prolactin
Corticotropin-releasing hormone (CRH) Adrenocorticotropin, b-lipotropin
Gonadotropin-releasing hormone (GnRH) Luteinizing hormone (LH), follicle-stimulating hormone (FSH)
Growth hormone-releasing hormone (GHRH or GRH)
Growth hormone (GH)
Prolactin-releasing factor (PRF) Prolactin
Melanocyte-stimulating hormone-releasing factor (MRF)
Melanocyte-stimulating hormone (MSH), b-endorphin
Inhibiting
Prolactin release-inhibiting hormone (PIH), dopamine
Prolactin
Growth hormone release-inhibiting hormone (GIH or GHRIH; somatostatin)
Growth hormone, thyrotropin
Melanocyte-stimulating hormone release-inhibiting factor (MIF)
Melanocyte-stimulating hormone (MSH)

The three divisions of the autonomic nervous system comprise an integrated motor system that acts in parallel with the somatic motor system and is responsible for homeostasis. Esential to the functioning of the motor outflow are the visceral sensory afferents that are relayed from the nucleus of the solitary tract through a network of central autonomic control nuclei. The hypothalamus integrates somatic, visceral, and behavioral information from all of these sources, thus coordinating autonomic and endocrine outflow with behavioral state.
Several features of the autonomic nervous system permit rapid integrated responses to changes in the environment. The activity of effector organs is finely controlled by coordinated and balanced excitatory and inhibitory inputs from tonically active postganglionic neurons. Moreover, the sympathetic system is greatly divergent, permitting the entire body to respond to extreme conditions
P.980
کلی مشخصات
حرکتی جسمی سیستم با موازی صورت به که است شده ادغام حرکتی سیستم یک شامل خودکار عصبی سیستم بخش سه از . دستگاه از هسته از که است احشایی حسی آوران اعصاب موتور خروجی عملکرد به همزمان است تعادل مسئول و کند می عمل . اطالعات و احشایی جسمانی، ادغام هیپوتاالموس باشد می مخابره مرکزی خودکار کنترل های هسته از ای شبکه طریق از انفرادی
رفتاری دولت با ریز درون غدد و خودکار خروج هماهنگی نتیجه در منابع، این همه از .رفتاری
. موثر بدن اعضای فعالیت است زیست محیط در تغییرات به سریع یکپارچه های پاسخ اجازه خودکار عصبی سیستم از ویژگی چندنورون از مهار های ورودی و متعادل و هماهنگ تحریکی های ریز . postganglionic است شود می کنترل فعال مانند ها عارضه از
دشوار شرایط به پاسخ برای بدن کل اجازه متفاوت، زیادی حد تا است سمپاتیک سیستم این، بر عالوه
In addition to the small molecule neurotransmitters— ACh and norepinephrine—a wide variety of peptides are thought to be released by autonomic neurons either onto postganglionic cells or their targets. Many of these peptides act to alter the efficacy of cholinergic or adrenergic transmission. The autonomic nervous system uses a rich variety of chemical mediators, several of which may commonly coexist in single autonomic neurons. The release of different combinations of chemical mediators from autonomic neurons may represent a means of “chemical coding” of information transfer in the different branches of the autonomic nervous system, although we are still only beginning to learn how to read the code
As we shall also see in the following two chapters, the autonomic nervous system is a remarkably adaptable system of homeostatic control. It can function locally through branches of primary sensory fibers that terminate in autonomic ganglia, or intrinsically through the entire nervous system on the functions of the digestive tract. Control centers in the brain stem are involved in several autonomic reflexes. While the hypothalamus integrates behavioral and emotional responses arising from the forebrain with ongoing metabolic need to produce highly coordinated autonomic control and behavior.
، عصبی های دهنده انتقال کوچک مولکول بر توسط ACHعالوه شود می تصور پپتیدهای از ای گسترده طیف ، نفرین نوراپی وهای سلول روی بر یا اتونوم عصبی پپتید . postganglionic سلولهای این از بسیاری شود می عرضه بازار در را خود اهداف یا و
. های واسطه از غنی انواع از استفاده با خودکار عصبی سیستم آدرنرژیک انتقال یا و کولینرژیک اثر تغییر برای کنند می عملهای . واسطه از مختلف ترکیبات انتشار باشند داشته وجود اتونوم تک عصبی های سلول در است ممکن معموال که چند شیمیایی،
" " مختلف های شاخه در اطالعات انتقال شیمیایی نویسی برنامه از استفاده است ممکن اتونوم عصبی های سلول از شیمیاییکد خواندن به ی نحوه یادگیری به شروع فقط هم هنوز ما چه اگر ، نشان خودکار عصبی سیستم
کنترل از سازگار ای مالحظه قابل سیستم یک خودکار عصبی سیستم کنید، می مشاهده زیر فصل دو در باید نیز ما که همانطورسیستم . طریق از ذاتا یا ، فسخ اتونوم گانگلیون در که اولیه حسی الیاف از شاخه طریق از را محلی تواند می این است متعادل
. . که حالی در دارد نقش اتونوم های رفلکس چند در مغز ساقه در کنترل مراکز کنند می عمل گوارش دستگاه عملکرد در کل عصبیبسیار خودکار کنترل تولید برای انجام حال در ساز و سوخت نیاز با مغز جلو از ناشی عاطفی و رفتاری های پاسخ هیپوتاالموس
ادغام رفتار و .هماهنگ
Selected Readings
Bacq ZM. 1974. Chemical Transmission of Nerve Impulses: A Historical Sketch. New York: Pergamon.
P.981
Burnstock G. l972. Purinergic nerves. Pharmacol Rev 24:509–581.

Burnstock G, Hoyle CHV (eds). 1995. The Autonomic Nervous System. Vol. 1, Autonomic Neuroeffector Mechanisms. London: Harwood Academic.
Cannon WB. 1932. The Wisdom of the Body. New York: Norton.
Costa M, Brookes SJ. 1994. The enteric nervous system. Am J Gastroenterol 89 (Suppl):S129–137.
Furness JB, Bornstein JC, Murphy R, Pompolo S. 1992. Roles of peptides in transmission in the enteric nervous system. Trends Neurosci 15:66–71.
Langley JN. 1921. The Autonomic Nervous System. Cambridge: Heffer & Sons.
Milner P, Burnstock G. 1995. Neurotransmitters in the autonomic nervous system. In: AD Korczyn (ed). Handbook of Autonomic Nervous System Dysfunction, pp. 5-32. New York: Marcel Dekker.
برگزیدهای
Bacq ZM . عصبی. 1974سال پالسهای شیمیایی : A: انتقال . پرگامون نیویورک طرح .تاریخی
P.981
Burnstock G. l972 .اعصاب Purinergic. Pharmacol 581-24:509برگرد.
Burnstock G هویل ، CHV (EDS) . 1995سال . . جلد. خودکار عصبی خودکار 1سیستم ، Neuroeffector : هاروود. لندن ساز.علمی
: 1932سال . WB کانن . نورتون. نیویورک بدن از .حکمت
M کوستا ، SJ .سال . 1994بروکس هستم. روده عصبی .S129-137): نمودار (J Gastroenterol 89 سیستم
JB فرنس ، Bornstein JC مورفی ، R ، Pompolo S. 1992 . . روند روده عصبی سیستم در انتقال در پپتید Neurosci نقش15:66-71.
. . JN. 1921 النگلی کمبریج خودکار عصبی پسران Heffer: سیستم .و
P میلنر ، Burnstock G. 1995 . . در خودکار عصبی سیستم در عصبی های دهنده خودکار(. (AD Korczyn: انتقال هندبوک ادصص سیستم، عملکرد اختالل : 32-5عصبی یکسان. .نیویورک
References
Brown DA, Adams PR. 1980. Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. Nature 283:673–676.
Elfvin LG, Lindh B, Hokfelt T. 1993. The chemical neuroanatomy of sympathetic ganglia. Annu Rev Neurosci 16:471–507.
Fredholm BB, Abbracchio MP, Burnstock G, Daly JW, Harden TK, Jacobson KA, Leff P, Williams M. 1994. Nomenclature and classification of purinoceptors. Pharmacol Rev 46:143–156.
Furness JB, Costa M. 1980. Types of nerves in the enteric nervous system. Neuroscience 5:1–20.
Gershon M. 1998. The Second Brain. New York: Harper Collins.
Guillemin R. 1978. Control of adenohypophysial functions by peptides of the central nervous system. Harvey Lect 71:71–131.
Hugdahl K. 1995. Classical conditioning and implicit learning: The right hemisphere hypothesis. In: RJ Davidson, K Hugdahl (eds). Brain Asymmetry, pp. 235-268. Cambridge, MA: MIT Press.
Hugdahl K, Berardi A, Thompson WL, Kosslyn SM, Macy R, Baker DP, Alpert NM, LeDoux JE. 1995. Brain mechanisms in human classical conditioning: a PET blood flow study. Neuroreport 6:1723–1728.
Hutter OF, Trautwein W. 1956. Vagal and sympathetic effects on the pacemaker fibers in the sinus venosus of the heart. J Gen Physiol 39:715–733.
Jan LY, Jan YN. 1983. A LHRH-like peptidergic neurotransmitter capable of action at a distance in autonomic ganglia. Trends Neurosci 6:320–325.

Loewy AD, Spyer KM (eds). 1990. Central Regulation of Autonomic Function. New York: Oxford Univ. Press.
Randall WC (ed). 1984. Nervous Control of Cardiovascular Function. New York: Oxford Univ. Press.
Reuter H, Scholz H. 1977. The regulation of the calcium conductance of cardiac muscle by adrenaline. J Physiol (Lond) 264:49–62.
Silverman AJ, Zimmerman EA. 1983. Magnocellular neurosecretory system. Annu Rev Neurosci 6:357–380.
Swanson LW, Sawchenko PE. 1983. Hypothalamic integration: organization of the paraventricular and supraoptic nuclei. Annu Rev Neurosci 6:269–324.
Toda N, West T. 1967. Interactions of K, Na and vagal stimulation in the S-A node of the rabbit. Am J Physiol 22:416–423.
Tranel D, Damasio AR. 1985. Knowledge without awareness: an autonomic index of facial recognition by prosopagnosics. Science 228:1453–1454.
Yuan SY, Bornstein JC, Furness JB. 1995. Pharmacological evidence that nitric oxide may be a retrograde messenger in the enteric nervous system. Br J Pharmacol 114:428–432.


















