
As histonas se ligam ao DNA graças à interação de seus ...º período Faciplac/Biologia...
23
Transcript of As histonas se ligam ao DNA graças à interação de seus ...º período Faciplac/Biologia...


* As histonas se ligam ao DNA graças à interação de seus radicais amino com os radicais fosfato do DNA. Nem todos os radicais fosfato estão neutralizados pelas histonas, o que confere à cromatina um caráter ácido (afinidade por corantes básicos).

Cada cromossomo contém um único duplex de DNA, compactado em uma fibra que se estende continuamente ao longo de todo o cromossomo. Assim, no que diz respeito à cromatina interfásica e ao cromossomo mitótico, podemos explicar a compactação de uma molécula de DNA única e extremamente longa, em uma forma na qual pode ser transcrita e replicada e também ser ciclicamente mais ou menos comprimida.A disposição da cromatina dentro do núcleo e o seu grau de condensação variam de um tipo celular para outro (são característicos de cada tipo celular). Além disso, a mesma célula pode apresentar a cromatina com vários graus de condensação, de acordo com o estágio funcional e com o estado de diferenciação em que se encontra.

Os genes localizados na eucromatina podem se expressar ou não, dependendo do tipo celular e de suas necessidades metabólicas. Existem pelo menos duas formas de eucromatina: cerca de 10% na forma de cromatina ativa, que é menos condensada, e o restante na forma de cromatina inativa, que é mais condensada. A heterocromatina constitutiva está sempre condensada. Consiste, na maior parte, de DNA repetitivo e é encontrada nos centrômeros e ao redor deles e em algumas outras regiões. Ela passa pelo ciclo celular com poucas mudanças no seu grau de condensação. Forma uma série de massas discretas, mas, freqüentemente, as diversas regiões de cromatina agregam-se em um cromocentro densamente corado.A heterocromatina facultativa pode existir tanto na forma geneticamente ativa (descondensada) quanto na forma inativa (condensada), como no caso da inativação do cromossomo X em mamíferos.A mesma fibra cromatínica pode apresentar regiões eucromáticas contínuas com regiões heterocromáticas. Assim, o material genético é organizado de modo que diferentes estados de compactação sejam mantidos lado a lado, possibilitando a ocorrência de alterações cíclicas no nível de compactação da cromatina entre a intérfase e a divisão, e entre as diferentes fases da vida da célula.

Os cromossomos normais têm um só centrômero, o qual é visto ao miscroscópio como uma constrição primária, a região em que as cromátides-irmãs estão unidas. O centrômero é essencial para a segregação durante a divisão celular. Funções:-ponto de ligação das fibras de microtúbulos durante a divisão celular;-ponto de reunião de cromátides-irmãs;-motor mecano-químico responsável pelo movimento dos cromossomos para os pólos da célula e conclusão da divisão celular.Durante a prófase tardia, um par de cinetócoros forma-se em cada centrômero, cada um ligado a uma das cromátides-irmãs. Múltiplos microtúbulos ligam-se a cada cinetócoro, ligando o centrômero de um cromossomo aos dois pólos do fuso. Os cinetócoros desempenham um papel fundamental nesse processo, controlando a reunião e a separação dos microtúbulos acoplados e, por meio da presença de moléculas motoras, dirigindo o movimento cromossômico.Os telômeros são estruturas especializadas, constituídas por DNA repetitivo (repetições em tandem ou agrupadas) e proteína, que cobrem as extremidades dos cromossomos eucarióticos. Funções prováveis: -manter a integridade estrutural do cromossomo: se um telômero for perdido, a extremidade cromossômica resultante fica instável, tendendo a fundir-se com as extremidades de outros cromossomos quebrados ou envolver-se em eventos de recombinação ou ser degradada.-garantir a replicação completa das extremidades codificadoras dos cromossomos: durante a replicação do DNA, a síntese da fita atrasada é descontínua e requer a presença de algum DNA à frente para um primerde RNA.-auxiliar o estabelecimento da arquitetura tridimencional do núcleo e/ou do pareamento cromossômico: as extremidades dos cromossomos parecem estar ligadas à membrana nuclear, sugerindo que os telômeros auxiliam no posicionamento dos cromossomos.-auxiliar no reconhecimento entre cromossomos homólogos para pareamento na meiose.Origens de replicação: necessárias para que ocorra a replicação do DNA na fase S da intérfase. São seqüências de DNA cis-ativas, situadas próximas aos pontos onde a síntese de DNA é iniciada, que controlam o início da replicação. Nos eucariotos existem várias origens da replicação (replicadores) autônomas (ARS – autonomously replicating sequences) ao longo de uma única molécula (para cada origem, duas forquilhas de replicação bidirecionais).

As histonas H3 e H4 apresentam seqüências idênticas em organismos tão distintos quanto a ervilha e o boi, sugerindo que elas desempenham funções idênticas em todos os eucariontes. Os tipos H2A e H2B possuem também seqüências idênticas, com algumas variações espécie-específicas.Em alguns tecidos, a H1 é substituída por histonas especiais. Por exemplo, em eritrócitos nucleados de aves, a histona H5 é encontrada em substituição à H1.

A porção enovelada da molécula das histonas contém alta percentagem de aminoácidos hidrofóbicos, e sua associação com o DNA deve-se à interações hidrofóbicas. Pode-se separar, por processos químicos, a molécula de DNA das moléculas de histonas. Mas quando o DNA e as histonas são colocados juntos em condições favoráveis, ocorre novamente a formação espontânea de nucleossomos.

O nucleossomo é uma partícula de forma cilíndrica achatada, com 10 nm de diâmetro e 6 nm de altura. Cada nucleossomo é constituído por 200 pb de DNA associados a um octâmero de histonas e a uma molécula de histona H1. O octâmero é formado por duas moléculas de cada uma das histonas H2A, H2B, H3 e H4. A molécula de H1 se associa externamente ao DNA que envolve o octâmero. Cada nucleossomo é formado por um centro ou cerne, constituído pelo octâmero de histonas H2A, H2B, H3 e H4, em torno do qual se enrola um segmento de DNA de aproximadamente 146 pb. Conectando um centro de nucleossomo a outro, encontra-se um segmento de DNA não associado a proteínas com 15 até 100 pb, chamado DNA de ligação.

Dois tipos de fibras cromatínicas são encontradas no núcleo interfásico: a fibra de 10 nm de diâmetro ou nucleofilamento e a fibra de 30 nm ou solenóide.A fibra de 10 nm constitui o primeiro nível de compactação da cromatina e é formada pela associação de nucleossomos adjacentes. A organização dessa fibra depende da interação entre as histonas H1 de nucleossomos vizinhos. A histona H1 de um nucleossomo liga-se através de sua extremidade amino-terminal, à extremidade carboxi-terminal da H1 do nucleossomo adjacente, em um arranjo “cabeça-cauda”. Com essa organização, o DNA de ligação não é mais observado na fibra de 10 nm.A fibra de 30 nm constitui o segundo nível de organização da cromatina e é formada pelo enovelamento da fibra de 10 nm em uma estrutura helicoidal. Cada volta da espiral contém 6 nucleossomos organizados radialmente, ficando a histona H1 localizada no interior da fibra. A histona H1, juntamente com íons Mg2+ em concentração adequada, tem papel preponderante na formação e estabilização dessa fibra.Durante a intérfase, a cromatina que contém os genes ativamente transcritos é formada, em sua maioria, por fibras de 30 nm, enquanto cerca de 10% estão na forma de fibras de 10 nm, permitindo o acesso às enzimas envolvidas na transcrição.
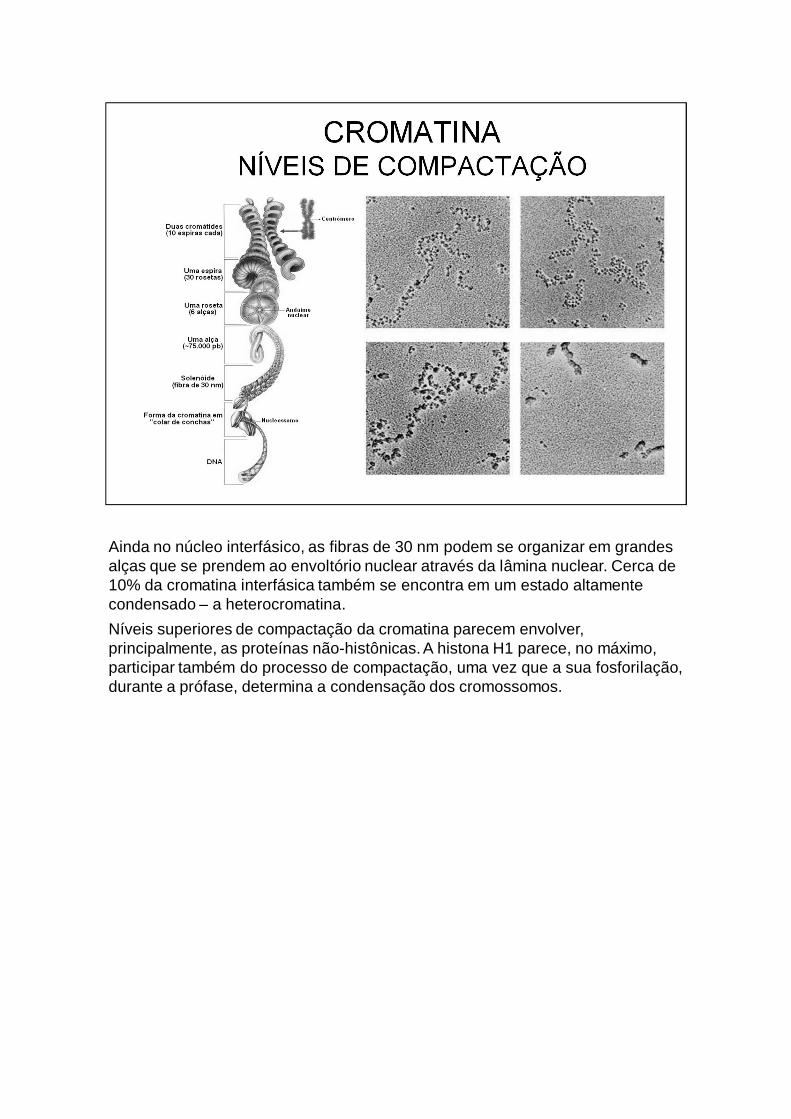
Ainda no núcleo interfásico, as fibras de 30 nm podem se organizar em grandes alças que se prendem ao envoltório nuclear através da lâmina nuclear. Cerca de 10% da cromatina interfásica também se encontra em um estado altamente condensado – a heterocromatina.Níveis superiores de compactação da cromatina parecem envolver, principalmente, as proteínas não-histônicas. A histona H1 parece, no máximo, participar também do processo de compactação, uma vez que a sua fosforilação, durante a prófase, determina a condensação dos cromossomos.


As histonas são suscetíveis a uma grande variedade de modificações pós-traducionais, tais como acetilação, fosforilação, metilação e ubiquitinação. A maioria dessas modificações acontece no domínio N-terminal das histonas, que é rico nos aminoácidos básicos lisina e arginina (mas também podem ocorrer nos domínios globulares). Enzimas como a acetiltransferase, as quinases e as metiltransferases, que depositam marcadores químicos nas histonas (acetil, fosfato e metil, respectivamente), são reguladores importantes da atividade gênica, da dinâmica dos cromossomos, da regulação do ciclo celular e da organização do genoma.


Em 1993, Alan Wolffe mostrou que acetilases/desacetilases formam complexos com fatores de transcrição que ligam/desligam os genes.Em 1998 Adrian Bird et al. mostraram que as desacetilases podem funcionar em conjunto com metilases: se a desacetilase for inibida, a metilação não inativa os genes.

In vivo, os resíduos de lisina podem ser mono, di ou trimetilados, enquanto os de arginina, mono ou bimetilados.

Formas ubiquitiladas de histonas H2A e H2B foram associadasespecificamente com genes ativos, tornando a ubiquitilação de histona umdos primeiros marcadores de cromatina transcricionalmente activo a serreconhecidoA ubiquitilatição da histona H2B, mediada pela enzima ubiquitina-conjugante (Ubc) Rad6, é implicada na repressão transcricional do geneargininosuccinato sintase (ARG1) e manutenção do silenciamentotelomérico em Saccharomyces cerevisiae. A ubiquitilação de da histonaH2B (uH2B) é necessária para a metilação da histona H3 nos resíduos delisina 4 (K4) e 79 (K79). A metilação histona H3, por sua vez, é necessáriapara o silenciamento do gene-telomérico.
Figura: A histona H2B é reconhecida pela Rad6 e suas proteínasauxiliares e é ubiquitilada na sua lisina 123. Esta ubiquitinação servecomo um sinal de reconhecimento direto ou indireto para COMPASS, quecatalisa a metilação da quarta lisina da histona H3, resultando nosilenciamento transcriptinal de genes localizados perto do telômero.COMPASS: a metilação de lisina 4 na cauda amino-terminal da histonaH3, mediada por um complexo multiproteico chamdo compass, énecessária para silenciar a expressão de genes localizados perto dostelômeros cromossômicos e dentro do rDNA.


O grupo fosfato é fortemente negativo, então sua adição induz forças nacadeia protéica que podem levar a uma radical alteração em suaconformação. Desse modo, uma proteína pode expor os aminoácidosantes escondidos em seu centro e mudar muito suas características. Porexemplo, uma proteína apolar e hidrofóbica pode se tornar polar ehidrofílica.Figura de cima: após a estimulação pelo fator de crescimento, a via daMAP quinase é ativada, resultando na fosforilação da histona H3 em doisresíduos específicos de serina (S10 e S28). Estes eventos de fosforilaçãocorrelacionam com a ativação da transcrição de genes.MAP quinase (Mitogen Activated Protein Kinases): Proteínas-quinaseativadas por mitógenos. É uma subfamília de proteínas-quinaseespecíficas de serina/treonina que respondem a estímulos extracelulares(mitógenos) e regulam várias atividades celulares, como expressãogênica, mitose, diferenciação, sobrevivência celular e apoptose (mortecelular).Figura de baixo: a fosforilação de H3 promove também a acetilação damesma histona e estas modificações funcionam em conjunto para ativar aexpressão do gene.A desfosforilação é realizada pelas fosfatases.

SAH: S-adenosil-homocisteína, produzida por desmetilação da SAM.A natureza das desmetilases é desconhecida.
A metilação consiste em uma modificação covalente do DNA na qual umgrupamento metil (CH3) é transferido da S-adenosilmetionina para ocarbono 5 de uma citosina (5-Mec ou 5-metil-citosina) que geralmenteprecede uma guanina (dinucleotídeo CpG), pela ação de uma família deenzimas que recebe o nome de DNA metiltransferases (DNMT).

As DNA metiltransferases estão divididas em duas classes derepresentantes: aquelas envolvidas na metilação de fitas hemimetiladasdo DNA (fitas de DNA em processo de replicação), conhecidas comometilases de manutenção como a DNMT1; e outro grupo, responsávelpela maioria dos processos de metilação de novo, que ocorre em sítioscom nenhum tipo de indicação de metilação, ou seja, sem a presençade metilação prévia, como as DNMT2, DNMT3A e DNMT3B. Osdoadores de radical metil são obtidos pela dieta e são principalmente ametionina, seguido do folato, colina e vitamina B12.
A desmetilação pode ocorrer na ausência de DNMT1 com rodadascontínuas de replicação do DNA (desmetilação passiva), bem comoativamente (sem a replicação do DNA). O processo denominado dedesmetilação ativa envolve as desmetilases e parece ser necessário paraativar genes específicos ou apagar a marca epigenética durante odesenvolvimento ou em respostas a perturbações ambientais. Adesmetilação ainda pode ser passiva quando não há envolvimento dedesmetilases e ocorre quando a manutenção pelas metiltransferases éinativada durante o ciclo celular. Assim, o nível e o padrão de 5-Mec sãodeterminados por ambos os processos de metilação e desmetilação,e as enzimas envolvidas nesses processos devem estar altamentereguladas.

Os dinucleotídeos CpG aparecem esparsos pelos genomas eucariotos ouagrupados em regiões definidas como ilhas CpG. Essas ilhas sãofrequentes em regiões promotoras de certos genes, incluindo geneshousekeeping. A transcrição gênica pode ser fortemente inibida pelaadição de radical metil. A presença de um “capuz” metil sobre umacitosina que precede a uma guanina pode inibir a ligação de fatores detranscrição a essas regiões. A não ligação de fatores de transcrição aosseus sítios específicos resulta na ausência de transcrição gênica. Aocontrário, a desmetilação leva ao aumento da transcrição gênica. Assim,CpGs metilados estão associados com DNA silenciados (transposons,genes imprintados), enquanto CpGs não-metilados estão associados agenes ativos.
Definindo: “ilhas CpG” são regiões do DNA com mais de 200 pares debases (pb), contendo aproximadamente 50% de bases C e G e com umapresença esperada de proximadamente 60% de dinucleotídeos CG.
Em células embrionárias indiferenciadas, a porcentagem de metilação emCpA, CpT ou CpC é alta e isso poderia estar relacionado com apluripotência dessas células.




















