
The Autonomic Nervous System. Visceral sensory Visceral motor &

Arteriolar Function in Visceral Adipose Tissue Is Impairedin Human Obesity
Melissa G. Farb, Lisa Ganley-Leal, Melanie Mott, YanMei Liang, Bahadir Ercan,Michael E. Widlansky, Sherman J. Bigornia, Antonino J. Fiscale, Caroline M. Apovian, Brian Carmine,
Donald T. Hess, Joseph A. Vita, Noyan Gokce
Objective—The purpose of this study was to characterize the relationship between adipose tissue phenotype anddepot-specific microvascular function in fat.
Methods and Results—In 30 obese subjects (age 42�11 years, body mass index 46�11 kg/m2) undergoing bariatricsurgery, we intraoperatively collected visceral and subcutaneous adipose tissue and characterized depot-specific adiposephenotypes. We assessed vasomotor function of the adipose microvasculature using videomicroscopy of small arterioles(75–250 �m) isolated from different fat compartments. Endothelium-dependent, acetylcholine-mediated vasodilation wasseverely impaired in visceral arterioles, compared to the subcutaneous depot (P�0.001 by ANOVA). Nonendotheliumdependent responses to papaverine and nitroprusside were similar. Endothelial nitric oxide synthase inhibition withN�-nitro-L-arginine methyl ester reduced subcutaneous vasodilation but had no effect on severely blunted visceral arteriolarresponses. Visceral fat exhibited greater expression of proinflammatory, oxidative stress-related, hypoxia-induced, andproangiogenic genes; increased activated macrophage populations; and had a higher capacity for cytokine production ex vivo.
Conclusion—Our findings provide clinical evidence that the visceral microenvironment may be intrinsically toxic toarterial health providing a potential mechanism by which visceral adiposity burden is linked to atherosclerotic vasculardisease. Our findings also support the evolving concept that both adipose tissue quality and quantity may play significantroles in shaping cardiovascular phenotypes in human obesity. (Arterioscler Thromb Vasc Biol. 2012;32:467-473.)
Key Words: endothelium � vasodilation � adiposity � arteries � inflammation
The global obesity epidemic is expanding at an alarmingrate producing a major health care burden.1,2 Obesity is
associated with endothelial dysfunction and early atheroscle-rosis,3,4 and recent data highlight obesity as a major cause ofpremature death from ischemic heart disease and stroke.5
Clinical studies link the accumulation of intraabdominalvisceral fat and central adiposity to coronary heart disease,metabolic syndrome, and type 2 diabetes risk.6–8 As such,there is mounting evidence that intrinsic properties of visceraladipose tissue may be deleterious to the cardiovascular systemowing to its high capacity for toxic lipolysis and increasedproduction of adipokines and inflammatory cytokines.7,9–11 Werecently demonstrated that adipose tissue inflammation is linkedto whole body metabolic dysregulation and systemic vasculardysfunction, providing evidence that qualitative features of fatmay shape clinical phenotypes.12,13
While experimental studies infer a pathogenic connectionbetween visceral fat and cardiovascular risk, mechanisms arelargely unknown. We hypothesized that if adipose tissue is aregulator of vascular function and the visceral milieu more
pathogenic, depot-specific differences would be manifest inarterioles intrinsic to different adipose domains. The goal of thepresent study was to determine whether arterioles isolated fromvisceral compared to subcutaneous (sc) regions exhibit a morediseased profile manifest by endothelial vasomotor dysfunction,and to determine whether differences in inflammatory charac-teristics exist between the 2 different compartments.
MethodsStudy SubjectsObese men and women (body mass index �30 kg/m2, age �18)were enrolled in the Boston Medical Center Bariatric SurgeryProgram. Subjects with unstable medical conditions such as activecoronary syndromes, congestive heart failure, systemic infection,acute illness, malignancy, or pregnancy were excluded. The studywas approved by Boston Medical Center Institutional Review Boardand all subjects gave written informed consent.
Anthropometric and Metabolic MeasuresClinical characteristics including blood pressure, heart rate, height,weight, body mass index, and waist circumference were recorded.
Received on: July 28, 2011; final version accepted on: November 7, 2011.From the Department of Medicine and Whitaker Cardiovascular Institute (M.G.F., M.M., B.E., S.J.B., J.A.V., N.G.), Department of Medicine (L.G.-L.,
Y.M.L.), Section on Infectious Diseases, Boston University School of Medicine, Boston, MA; Departments of Medicine and Pharmacology (M.E.W.),Division of Cardiovascular Medicine, Medical College of Wisconsin, Milwaukee, WI; Department of General Surgery (A.J.F., B.C., D.T.H.), Departmentof Medicine (C.M.A.), Section of Endocrinology, Diabetes and Nutrition, Boston University School of Medicine, Boston, MA.
Correspondence to Noyan Gokce, Boston Medical Center, 88 East Newton St, D-8, Cardiology, Boston, MA 02118. E-mail [email protected]© 2011 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.111.235846
467
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Biochemical analyses including lipids, glucose, insulin, homeostasismodel assessment of insulin resistance, glycosylated hemoglobin,and high-sensitivity C-reactive protein were quantified from bloodsamples collected in a fasting state.
Adipose Tissue Collection and Vessel PreparationSubcutaneous and visceral adipose tissue biopsies were collectedintraoperatively during planned bariatric surgery. Subcutaneousadipose tissue was harvested from the lower abdominal wall andvisceral tissue secured from the greater omentum, respectively.Specimens were placed immediately in cold HEPES buffer solution,pH 7.51 (American Bioanalytical, Natick, MA). Small adiposearterioles (75–250 �m internal diameter) were carefully removed ofsurrounding fat and connective tissue and suspended in an organchamber containing Krebs solution, cannulated securely with glassmicropipettes.14 The organ chamber was then mounted onto a stagewith inverted microscope (magnification �200) and video cameramonitor (model VIA-100; Boeckler Instruments, Tuscon, AZ) forvascular diameter measurements using videomicroscopy. Arterioleswere pressure equilibrated and continuously perfused with Krebsbuffer aerated with a gas mixture of 5% O2, 21% CO2, and 74% N2.
Assessment of Adipose Arteriolar FunctionThe internal arterial diameter of each vessel was initially measuredat a steady state followed by administration of endothelin-1 (ET-1,2�10�6 M, Sigma-Aldrich, St. Louis, MO) to preconstrict vessels to50% to 70% of internal diameter. Endothelial nitric oxide synthase-dependent vasodilation was assessed by measuring arteriolar changein diameter to increasing doses of receptor mediated NO-agonistacetylcholine (Ach, 10�10 to 10�5 M, Sigma-Aldrich) in the pres-ence or absence of endothelial nitric oxide synthase inhibitorN�-nitro-L-arginine methyl ester (L-NAME, 10�4 M, Sigma-Al-drich). Endothelium-independent vasodilation was determined usingpapaverine (Pap, 2�10�4 M, Sigma-Aldrich) and sodium nitroprus-side (SNP, 10�10 to 10�4 M, Sigma-Aldrich). Pharmacologicalagents were added to the external bathing solution of the organchamber, and the indicated concentration represents the final cham-ber molar concentration.
Adipose Tissue Gene ExpressionImmediately following biopsy collection, adipose tissue sampleswere stored in RNAlater (Sigma Aldrich) solution at �80°C. TotalRNAs were isolated from homogenized whole adipose tissues usingthe miRNeasy kit (Qiagen, Germantown, MD). Quantitative real-time–PCR reactions were performed using a high throughput instru-ment (BioMark, Fluidigm, San Francisco, CA) by using GeneExpression Assays (Applied Biosystems, Foster City, CA) andDynamicArray chips (Fluidigm, San Francisco, CA). We addition-ally separated endothelial cells from nonvascular fractions in adiposetissue as previously described.15,16 Briefly, fat samples were digestedin a collagenase I solution (300 U/mL in phosphate-buffered salineand 2% bovine serum albumin; Worthington Biochemical Corp,Lakewood, NJ), treated with erythrocyte-lysing buffer (R&D sys-tems, Minneapolis, MN), and endothelial cells isolated using CD31MACS MicroBead Kit (Miltenyi Biotec Auburn, CA). Cells werelysed with Qiazol lysis solution and kept at �80°C until RNAisolation. Total RNA isolation for vascular endothelial cells wasperformed as described above.
HistologyBlood vessels isolated from fat were fixed in 10% buffered formalinfor analysis by the Pathology Department of Boston Medical Center.Paraffin embedded arterioles were stained with hematoxylin andeosin and also characterized by immunohistochemistry with endo-thelial cell-specific CD31 (BioGenex, Freemont, CA). Microvesselswere stained for macrophages using a cell-specific target to CD68(Ventana Medical Systems, Tucson, AZ). Microvascular morphol-ogy was further assessed by elastica-van Gieson staining (VentanaMedical Systems). Media to lumen ratio was quantified using anestablished NIH Image J program as previously described.17
Flow Cytometry and Leukocyte IsolationResident macrophages were isolated from fat as previously de-scribed.18 In brief, tissues were gently homogenized and filteredthrough 70 �m cell strainers (BD Falcon, Bedford, MA) for singlecell suspension. Macrophages were isolated by dual density gradient(Histopaque-1077 and Histopaque-1119, Sigma Aldrich) to accountfor changes in cellular density of tissue. Recovered cells wereassessed by flow cytometry to measure macrophage surface recep-tors CD14, CD209, and CD206 (BD Pharmingen, San Diego, CA)and TLR4 (eBioscience, San Diego, CA).
In a subset of specimens, isolated leukocytes were harvested andallowed to adhere to cell culture plates for 4 hours. Adherent cellscomprised of 85% to 95% CD14� macrophages, cultured overnight�1 �g/mL of E.coli lipopolysaccharide (Alexis Corporation,Lausen, Switzerland), and supernatants assessed for secreted inter-leukin (IL)-6 and IL-8 concentrations using standard ELISA (R&DSystems, Minneapolis, MN).
Statistical AnalysisGroup differences in clinical characteristics, gene expression, cyto-kine production, flow-cytometry measures, and baseline vascularparameters were examined by Mann-Whitney–Wilcoxon and Chisquare tests for continuous and categorical variables, respectively.Repeated-measures ANOVA was used to compare depot-specificvascular dose-response curves to pharmacological probes. Associa-tions between vascular dilator responses (defined by dose-responsearea under the curve), adipose tissue gene expression, and clinicaldata were examined using Spearman’s rank correlation analyses.Statistical significance was defined as P�0.05. All data are ex-pressed as mean�SD, unless otherwise indicated. All data wereanalyzed using SPSS for Windows, version 13.1.
ResultsWe harvested a total of 40 adipose arterioles from sc (n�17) andvisceral (n�23) fat depots in 30 patients. Clinical characteristicsof subjects that provided adipose samples during bariatricsurgery are displayed in Table 1. There were no significantdifferences in clinical parameters or medication use listedbetween subjects that provided sc versus visceral arterioles.
Depot-Specific Adipose Arteriolar ResponsesMean resting internal diameter of arterioles isolated from scfat was 147�57 �m and in the visceral 171�50 �m(P�0.20). Additionally, the media/lumen ratio was not dif-ferent between microvessels isolated from the sc (2.1) com-pared to visceral adipose depot (2.0, P�0.88). The degree ofmicrovascular preconstriction to ET-1 was also similar inboth groups (57�12% and 56�12% of basal diameter in scand visceral, respectively, P�0.87). Endothelium-dependentvasodilation was then assessed with dose-response relationshipto increasing Ach concentrations (10�10–10�5 M). As shown inFigure 1, Ach-mediated vasodilation was significantly impairedin visceral compared to sc adipose arterioles (P�0.001 byANOVA), even exhibiting paradoxical vasoconstriction in someindividuals by �5%. In contrast, responses to papaverine (Pap)as an endothelium-independent agonist were similar in bothgroups, indicating intact vascular smooth muscle function. In asubset of 10 patients that provided paired samples simulta-neously from both visceral and sc depots, we again observed aprofound decrement in endothelium-dependent vasorelaxationof visceral arterioles, similar in magnitude to results for thegroup as a whole (P�0.001, Supplemental Figure I, availableonline at http://atvb.ahajournals.org). These latter findings areparticularly important because vessels for both depots originated
468 Arterioscler Thromb Vasc Biol February 2012
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

from the same person, removing any potential confoundingeffect of unmeasured systemic parameters between individuals.Stratification of subjects based on presence or absence ofdiabetes mellitus (Supplemental Figure II) or hypertension (datanot shown) did not alter the results.
Vessel viability was confirmed by preserved responses toET-1 (preconstriction) and papaverine (maximal vasodila-tion). We further excluded the possibility of endothelialdenudation or anatomic damage to blood vessels by histolog-ical confirmation of structural integrity. Immunohistochem-istry targeted to CD31 (Supplemental Figure IIIA), and H&Estaining (Supplemental Figure IIIB) demonstrated intact nu-cleated vascular endothelial and smooth muscle layers inadipose vessels (Supplemental Figure III). Additionally,elastica-van Gieson and CD68 stains showed no evidence ofintimal thickening, atherosclerotic changes, or vascular mac-rophage infiltration (Supplemental Figure IV).
To determine the role of nitric oxide bioaction in endothe-lium-dependent responses of adipose microvessels, in asubset of individuals, we examined vascular dilation toAch�L-NAME (10�4 M). As shown in Figure 2, L-NAMEsignificantly reduced Ach-mediated relaxation by 40% in scvessels (n�6, P�0.005), whereas no significant effect wasobserved on visceral arterioles that already exhibited pro-
found endothelial dysfunction (n�6, P�0.36). Additionally,we examined vasodilator responses to increasing doses ofsodium nitroprusside, which demonstrated no significantinterdepot differences in vasodilation (n�6, P�0.5; Supplemen-tal Figure V), further supporting the notion that depot-specificvasodilator defects are related to endothelial dysfunction.
Adipose Tissue CharacterizationAs systemic clinical parameters failed to explain differencesin depot-specific vascular responses, we sought to character-ize disparities between fat depots focusing on inflammatoryactivation. As shown in Table 2, using RT-PCR we demon-strated significantly higher expression of proinflammatoryand oxidative stress-related genes in visceral compared to thesc depot. Among the genes measured, the greatest differencewas observed for IL-6, a cytokine upregulated in obesity and
Table 1. Study Population Characteristics
Clinical Parameter n�30
Age (yrs) 42�11
Female (%) 79%
BMI (kg/m2) 46�11
Waist circumference (cm) 140�20
Weight (kg) 129�36
Insulin (mU/ml) 12�7
Glucose (mg/dl) 107�38
HOMA 3�2
Triglycerides (mg/dl) 123�61
Total cholesterol (mg/dl) 190�39
HDL-C (mg/dl) 47�10
LDL-C (mg/dl) 120�35
hs-CRP (mg/dl) 8.8�9.7
HbA1c (%) 6.0�0.9
Duration of adulthood obesity (yrs) 23�11
Diabetes (%) 44%
Hypertension (%) 44%
Hypercholesterolemia (%) 22%
ARB/ACE-inhibitor use (%) 27%
DPP-4 inhibitor use (%) 3%
Insulin use (%) 3%
Metformin use (%) 40%
Statin use (%) 10%
Thiazolidinedione use (%) 3%
BMI indicates body mass index; HOMA, homeostasis model assessment ofinsulin resistance; hs-CRP, high-sensitivity C-reactive protein; HbA1c, glyco-sylated hemoglobin; ARB/ACE, angiotensin receptor blocker/angiotensin con-verting enzyme; DPP-4, dipeptidyl peptidase-4.
Figure 1. Adipose tissues arteriolar responses. Endothelium-dependent, acetylcholine (Ach)-mediated vasodilation wasseverely impaired in visceral compared to sc adipose tissuearterioles (P�0.001 by ANOVA), while responses to papaverine(PAP) were similar (n�40). Data presented as mean � SEM. ETindicates endothelin.
Figure 2. Effect of N�-nitro-L-arginine methyl ester (L-NAME) onadipose vasomotor responses. A, L-NAME (10-4 M) reducedvasodilator responses in subcutaneous microvessels indicatingpartial dependence on nitric oxide bioaction (n�6, P�0.005 byANOVA). B, In contrast, L-NAME had no effect on severelyblunted visceral arteriolar responses (n�6, P�0.36 by ANOVA).Data are presented as mean�SEM. ET indicates endothelin;PAP, papaverine; Ach, acetylcholine.
Farb et al Visceral Adiposity and Vascular Function 469
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

strongly implicated in the pathogenesis of diabetes and acutecoronary syndromes.19,20 Proangiogenic and hypoxia-relatedgenes were also upregulated in visceral fat in line withrecent data demonstrating depot-specific differences invascularity and angiogenic potential.21 Results were direc-tionally similar for subjects that provided paired sc andvisceral fat, despite the smaller sample size (SupplementalTable I). In addition, gene expression analysis specificallyperformed in isolated endothelial cells for select cytokinesyielded similar results (Supplemental Figure VI). Ach-mediated vasodilatation (AUC) correlated negatively withthe following adipose tissue genes: IL-6, NF-�B, MYD88,CCL5, TGF-�, CD8, HIF1-�, and VEGF (P�0.05 for all).
Infiltrating macrophages are thought to be the primarysource of proinflammatory cytokine production in the adipose
tissue.22 The notion that macrophages exist in proinflamma-tory (M1) and reparative (M2) states that may be relevant todisease mechanisms is an evolving concept.23 We identified asubset of activated adipose tissue macrophages expressingproinflammatory cell surface toll-like receptor 4 suggestiveof M1 phenotype. As shown in Figure 3, using flow cytom-etry we demonstrated higher population of toll-like receptor 4macrophages in visceral versus sc fat (58�20% versus43�20%, P�0.001, n�19). The same relationship was evi-dent for macrophages expressing 2 surface markers tradition-ally associated with M2 populations: CD209 (31�18% ver-sus 21�17%, P�0.01, n�14) and CD206 (27�21%versus18�11% P�0.01, n�17).
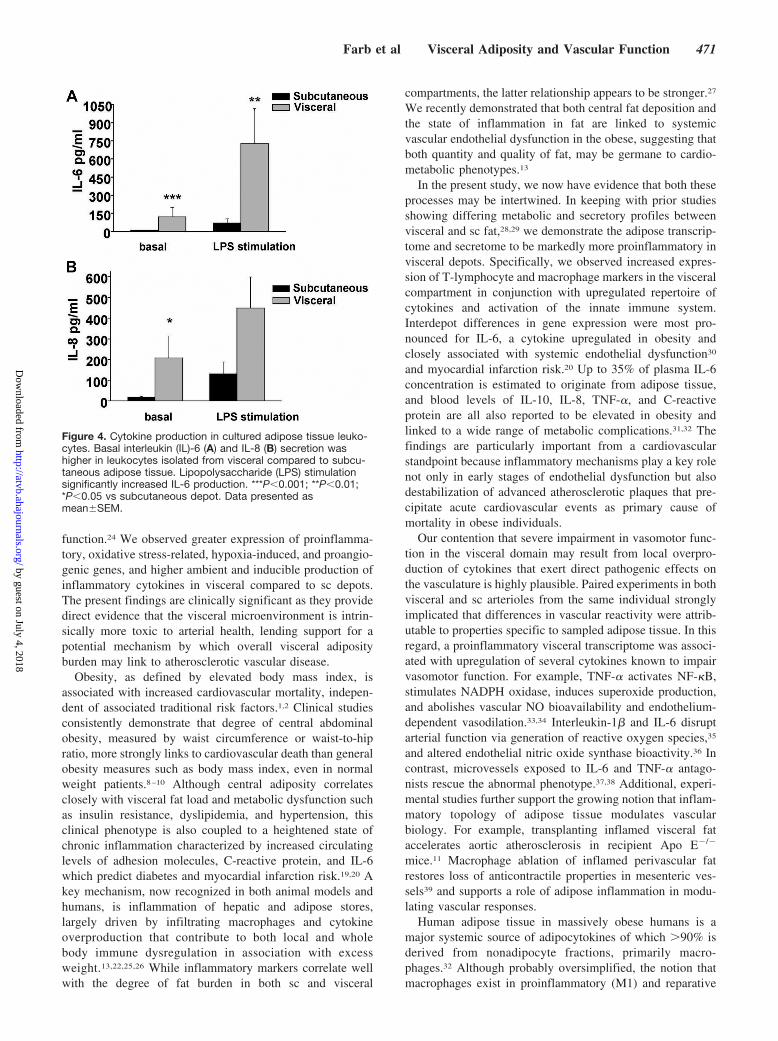
To further characterize functional aspects of adipose im-mune cells, leukocytes primarily comprising of macrophageswere isolated from different depots and cultured in thepresence or absence of lipopolysaccharide. As shown inFigure 4, leukocytes recovered from visceral tissue producedhigher basal levels of IL-6 (P�0.001) and IL-8 (P�0.05)compared to sc fat. Further, lipopolysaccharide-stimulatedproduction of IL-6 was higher in visceral fat (P�0.01). Thefindings suggest that immune cells within visceral domainsexpress higher basal and stimulated levels of cytokines thatcontribute to regional differences in proinflammatory profiles.
DiscussionIn the present study of severely obese subjects, we show thathuman arterioles isolated from visceral omental fat exhibitseverely impaired endothelium-dependent vasodilation com-pared to sc adipose microvessels. The degree of vascularimpairment in the visceral compartment was profound withsome arterial segments exhibiting paradoxical vasoconstric-tion suggesting a significant defect in endothelial vasodilator
Table 2. Gene Expression in Visceral Compared toSubcutaneous Adipose Tissue
Gene
Fold Differencein Visceral
Compared toSubcutaneous Fat P Value
Immune cell markers CD3 3.89 �0.001*
CD4 4.67 0.008*
CD8 7.54 �0.001*
CD68 1.70 0.032*
CD163 2.95 0.025*
FOXP3 1.26 0.073
Inflammation andoxidative stress
Adiponectin 1.90 0.095
CCL2 1.03 0.929
CCL5 6.32 �0.001*
CCR2 4.27 0.004*
FSTL1 3.42 0.009*
IFN-� 2.20 0.831
ICAM-1 1.54 0.055
IL-1� 4.23 0.077
IL-6 10.05 �0.001*
IL-10 3.93 0.059
MYD88 2.53 0.002*
NF-�B 2.06 0.001*
NOX1 1.85 0.044*
NOX2 4.41 �0.001*
NOX4 1.55 0.186
TGF-� 5.58 0.001*
TLR-4 2.80 0.003*
TNF-� 4.77 0.008*
VCAM-1 1.54 0.180
eNOS 2.55 0.149
iNOS 5.50 0.05
Angiogenic andhypoxia-relatedgenes
ANGPT2 4.55 0.004*
HIF1-� 3.05 0.004*
VEGF 3.24 �0.001*
ICAM-1 indicates intercellular adhesion molecule-1; IL, interleukin; TLR-4,toll-like receptor 4; NOX, NADPH oxidase; eNOS, endothelial nitric oxidesynthase; iNOS, inducible nitric oxide synthase; HIF1, hypoxia-inducible factor 1.
*P�0.05.
Figure 3. Depot-specific macrophage polarization in adiposetissue. Adipose flow cytometry identified higher populations of(A) TLR4� M1 polarized macrophages in visceral vs subcutane-ous adipose tissue. Similarly, macrophages expressing markerstraditionally associated with M2 phenotypes (B) CD209 and (C)CD206 were also significantly higher in visceral fat. ***P�0.001;**P�0.01 compared to subcutaneous depot. Data presented asmean�SEM.
470 Arterioscler Thromb Vasc Biol February 2012
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from
function.24 We observed greater expression of proinflamma-tory, oxidative stress-related, hypoxia-induced, and proangio-genic genes, and higher ambient and inducible production ofinflammatory cytokines in visceral compared to sc depots.The present findings are clinically significant as they providedirect evidence that the visceral microenvironment is intrin-sically more toxic to arterial health, lending support for apotential mechanism by which overall visceral adiposityburden may link to atherosclerotic vascular disease.
Obesity, as defined by elevated body mass index, isassociated with increased cardiovascular mortality, indepen-dent of associated traditional risk factors.1,2 Clinical studiesconsistently demonstrate that degree of central abdominalobesity, measured by waist circumference or waist-to-hipratio, more strongly links to cardiovascular death than generalobesity measures such as body mass index, even in normalweight patients.8–10 Although central adiposity correlatesclosely with visceral fat load and metabolic dysfunction suchas insulin resistance, dyslipidemia, and hypertension, thisclinical phenotype is also coupled to a heightened state ofchronic inflammation characterized by increased circulatinglevels of adhesion molecules, C-reactive protein, and IL-6which predict diabetes and myocardial infarction risk.19,20 Akey mechanism, now recognized in both animal models andhumans, is inflammation of hepatic and adipose stores,largely driven by infiltrating macrophages and cytokineoverproduction that contribute to both local and wholebody immune dysregulation in association with excessweight.13,22,25,26 While inflammatory markers correlate wellwith the degree of fat burden in both sc and visceral
compartments, the latter relationship appears to be stronger.27
We recently demonstrated that both central fat deposition andthe state of inflammation in fat are linked to systemicvascular endothelial dysfunction in the obese, suggesting thatboth quantity and quality of fat, may be germane to cardio-metabolic phenotypes.13
In the present study, we now have evidence that both theseprocesses may be intertwined. In keeping with prior studiesshowing differing metabolic and secretory profiles betweenvisceral and sc fat,28,29 we demonstrate the adipose transcrip-tome and secretome to be markedly more proinflammatory invisceral depots. Specifically, we observed increased expres-sion of T-lymphocyte and macrophage markers in the visceralcompartment in conjunction with upregulated repertoire ofcytokines and activation of the innate immune system.Interdepot differences in gene expression were most pro-nounced for IL-6, a cytokine upregulated in obesity andclosely associated with systemic endothelial dysfunction30
and myocardial infarction risk.20 Up to 35% of plasma IL-6concentration is estimated to originate from adipose tissue,and blood levels of IL-10, IL-8, TNF-�, and C-reactiveprotein are all also reported to be elevated in obesity andlinked to a wide range of metabolic complications.31,32 Thefindings are particularly important from a cardiovascularstandpoint because inflammatory mechanisms play a key rolenot only in early stages of endothelial dysfunction but alsodestabilization of advanced atherosclerotic plaques that pre-cipitate acute cardiovascular events as primary cause ofmortality in obese individuals.
Our contention that severe impairment in vasomotor func-tion in the visceral domain may result from local overpro-duction of cytokines that exert direct pathogenic effects onthe vasculature is highly plausible. Paired experiments in bothvisceral and sc arterioles from the same individual stronglyimplicated that differences in vascular reactivity were attrib-utable to properties specific to sampled adipose tissue. In thisregard, a proinflammatory visceral transcriptome was associ-ated with upregulation of several cytokines known to impairvasomotor function. For example, TNF-� activates NF-�B,stimulates NADPH oxidase, induces superoxide production,and abolishes vascular NO bioavailability and endothelium-dependent vasodilation.33,34 Interleukin-1� and IL-6 disruptarterial function via generation of reactive oxygen species,35
and altered endothelial nitric oxide synthase bioactivity.36 Incontrast, microvessels exposed to IL-6 and TNF-� antago-nists rescue the abnormal phenotype.37,38 Additional, experi-mental studies further support the growing notion that inflam-matory topology of adipose tissue modulates vascularbiology. For example, transplanting inflamed visceral fataccelerates aortic atherosclerosis in recipient Apo E�/�
mice.11 Macrophage ablation of inflamed perivascular fatrestores loss of anticontractile properties in mesenteric ves-sels39 and supports a role of adipose inflammation in modu-lating vascular responses.
Human adipose tissue in massively obese humans is amajor systemic source of adipocytokines of which �90% isderived from nonadipocyte fractions, primarily macro-phages.32 Although probably oversimplified, the notion thatmacrophages exist in proinflammatory (M1) and reparative
Figure 4. Cytokine production in cultured adipose tissue leuko-cytes. Basal interleukin (IL)-6 (A) and IL-8 (B) secretion washigher in leukocytes isolated from visceral compared to subcu-taneous adipose tissue. Lipopolysaccharide (LPS) stimulationsignificantly increased IL-6 production. ***P�0.001; **P�0.01;*P�0.05 vs subcutaneous depot. Data presented asmean�SEM.
Farb et al Visceral Adiposity and Vascular Function 471
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

(M2) states that may be relevant to disease mechanisms is anevolving concept.40 Although macrophages within the adi-pose tissue of obese mice exhibit M1 characteristics,41 im-mune dysregulation in human fat appears more complex andlacks a distinct phenotype at this time.42,43 We identified ahigher percentage of macrophages in visceral fat expressingcell surface TLR4, known to exhibit proinflammatory M1characteristics in atherosclerotic plaques.44 Similarly, weobserved parallel upregulation of macrophages expressingCD206 or CD209, traditionally associated with M2 subpopu-lations43 with no significant depot-specific difference inM1/M2 ratio. We speculate that adipose remodeling may notonly be species-specific but also relate to obesity durationthat have generally been short-term in animal studies, incontrast to our human cohort afflicted with an average of 20years of obesity. Understanding mechanisms involved inresolution of adipose tissue inflammation may reveal noveltherapeutic targets.45
What sets off the inflammatory cascade in adipose tissue isof great interest and likely to provide treatment clues forseveral obesity-related diseases that have inflammatorymechanistic basis including atherosclerosis, diabetes, andcancer. Experimental data suggest that adipose overexpan-sion, particularly in visceral fat, may suffer from capillaryrarefaction and tissue hypoperfusion that locally triggers avicious cycle of ischemia, hypoxia, necrosis, and inflamma-tion within the adipose milieu.46 As such, our observation ofsignificantly greater hypoxia-related and proangiogenic im-print in visceral fat prompts speculation that tissue-specificfunctional defects in vascularity and perfusion may haveprofound effects on the adipose microenvironment and theorganism as a whole, and leaves room for additional studies.
Our present study has several limitations. First, we studiedindividuals with advanced class III–IV obesity referred forbariatric surgery, which provided clinical feasibility to accessmultiple adipose depots simultaneously. We recognize thatstudying extreme weight categories may not be representativeof the general population with milder degrees of obesity.However, recent data show that even modest visceral fat gainin normal weight subjects promotes vascular endothelialdysfunction, suggesting a biological connection.47 Second,most participants in the study were women, reflecting thefemale predominance of our bariatric service and clinicalpractice nationally.48 Third, we speculated that TLR4 expres-sion signified M1 polarization, although the functional char-acterization of this specific macrophage phenotype in humanfat remains unproven. Fourth, medications could not beentirely washed out owing to medical treatment indications,and depot-specific effects in treated patients cannot be com-pletely excluded. Lastly, we attributed the cytokine sourcefrom cultured leukocytes to primarily macrophages based onprior experimental data, but we recognize that adipocytes andother immune cell populations including B and T lympho-cytes, neutrophils, and mast cells likely contribute to chemo-kine and cytokine release in adipose tissue.32,49
In conclusion, we provide direct human evidence that thevisceral microenvironment is intrinsically toxic to arterialhealth lending support for a potential mechanism by whichoverall visceral adiposity burden may link to atherosclerotic
vascular disease. Our data also support the growing paradigmthat qualitative features of adipose tissue may play a role inshaping disease phenotypes in obese individuals.
AcknowledgmentsWe thank Jingli Wang for her assistance with technical aspects of theadipose tissue microvessel videomicroscopy methodology.
Sources of FundingDr Gokce is supported by National Institutes of Health (NIH) grantsR01 HL084213 and P01 HL081587. Dr Vita is supported by NIHgrants HL083269, HL083801, HL081587, and HL75795. Dr Wid-lansky is supported by NIH grant K23 HL089326 and AHAGrant-in-Aid 10GRNT3880044.
DisclosuresDr Apovian has served on the advisory boards for Allergan, Amylin,Orexigen, Merck, Johnson and Johnson, Arena, and Sanofi-Aventis,and has received research funding from Lilly, Amylin, Pfizer,Sanofi-Aventis, Orexigen, MetaProteomics, and the Dr Robert C.and Veronica Atkins Foundation. There are no conflicts of interestwith the current article.
References1. Berrington de GA, Hartge P, Cerhan JR, Flint AJ, Hannan L, MacInnis
RJ, Moore SC, Tobias GS, nton-Culver H, Freeman LB, Beeson WL,Clipp SL, English DR, Folsom AR, Freedman DM, Giles G, HakanssonN, Henderson KD, Hoffman-Bolton J, Hoppin JA, Koenig KL, Lee IM,Linet MS, Park Y, Pocobelli G, Schatzkin A, Sesso HD, Weiderpass E,Willcox BJ, Wolk A, Zeleniuch-Jacquotte A, Willett WC, Thun MJ.Body-mass index and mortality among 1.46 million white adults. N EnglJ Med. 2010;363:2211–2219.
2. Whitlock G, Lewington S, Sherliker P, Clarke R, Emberson J, Halsey J,Qizilbash N, Collins R, Peto R. Body-mass index and cause-specificmortality in 900 000 adults: collaborative analyses of 57 prospectivestudies. Lancet. 2009;373:1083–1096.
3. Arkin JM, Alsdorf R, Bigornia S, Palmisano J, Beal R, Istfan N, Hess D,Apovian CM, Gokce N. Relation of cumulative weight burden to vascularendothelial dysfunction in obesity. Am J Cardiol. 2008;101:98–101.
4. McGill HC Jr, McMahan CA, Herderick EE, Zieske AW, Malcom GT,Tracy RE, Strong JP. Obesity accelerates the progression of coronaryatherosclerosis in young men. Circulation. 2002;105:2712–2718.
5. Roger VL, Go AS, Lloyd-Jones DM, Adams RJ, Berry JD, Brown TM,Carnethon MR, Dai S, de SG, Ford ES, Fox CS, Fullerton HJ, GillespieC, Greenlund KJ, Hailpern SM, Heit JA, Ho PM, Howard VJ, KisselaBM, Kittner SJ, Lackland DT, Lichtman JH, Lisabeth LD, Makuc DM,Marcus GM, Marelli A, Matchar DB, McDermott MM, Meigs JB, MoyCS, Mozaffarian D, Mussolino ME, Nichol G, Paynter NP, RosamondWD, Sorlie PD, Stafford RS, Turan TN, Turner MB, Wong ND, Wylie-Rosett J. Heart disease and stroke statistics–2011 update: a report fromthe American Heart Association. Circulation. 2011;123:e18–e209.
6. Van Gaal LF, Mertens IL, De Block CE. Mechanisms linking obesitywith cardiovascular disease. Nature. 2006;444:875–880.
7. Despres JP, Lemieux I. Abdominal obesity and metabolic syndrome.Nature. 2006;444:881–887.
8. Coutinho T, Goel K, Correa de SD, Kragelund C, Kanaya AM, Zeller M,Park JS, Kober L, Torp-Pedersen C, Cottin Y, Lorgis L, Lee SH, Kim YJ,Thomas R, Roger VL, Somers VK, Lopez-Jimenez F. Central obesity andsurvival in subjects with coronary artery disease a systematic review ofthe literature and collaborative analysis with individual subject data. J AmColl Cardiol. 2011;57:1877–1886.
9. Canoy D, Boekholdt SM, Wareham N, Luben R, Welch A, Bingham S,Buchan I, Day N, Khaw KT. Body fat distribution and risk of coronaryheart disease in men and women in the European Prospective Investi-gation Into Cancer and Nutrition in Norfolk cohort: a population-basedprospective study. Circulation. 2007;116:2933–2943.
10. Lakka HM, Lakka TA, Tuomilehto J, Salonen JT. Abdominal obesity isassociated with increased risk of acute coronary events in men. EurHeart J. 2002;23:706–713.
472 Arterioscler Thromb Vasc Biol February 2012
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

11. Ohman MK, Shen Y, Obimba CI, Wright AP, Warnock M, Lawrence DA,Eitzman DT. Visceral adipose tissue inflammation accelerates atherosclerosisin apolipoprotein E-deficient mice. Circulation. 2008;117:798–805.
12. Apovian CM, Bigornia S, Mott M, Meyers MR, Ulloor J, Gagua M,McDonnell M, Hess D, Joseph L, Gokce N. Adipose macrophage infiltrationis associated with insulin resistance and vascular endothelial dysfunction inobese subjects. Arterioscler Thromb Vasc Biol. 2008;28:1654–1659.
13. Farb MG, Bigornia S, Mott M, Tanriverdi K, Morin KM, Freedman JE,Joseph L, Hess DT, Apovian CM, Vita JA, Gokce N. Reduced adiposetissue inflammation represents an intermediate cardiometabolic pheno-type in obesity. J Am Coll Cardiol. 2011;58:232–237.
14. Phillips SA, Hatoum OA, Gutterman DD. The mechanism of flow-induced dilation in human adipose arterioles involves hydrogen peroxideduring CAD. Am J Physiol Heart Circ Physiol. 2007;292:H93–H100.
15. Curat CA, Miranville A, Sengenes C, Diehl M, Tonus C, Busse R,Bouloumie A. From blood monocytes to adipose tissue-resident macro-phages: induction of diapedesis by human mature adipocytes. Diabetes.2004;53:1285–1292.
16. Peinado JR, Jimenez-Gomez Y, Pulido MR, Ortega-Bellido M, az-LopezC, Padillo FJ, Lopez-Miranda J, Vazquez-Martinez R, Malagon MM. Thestromal-vascular fraction of adipose tissue contributes to major dif-ferences between subcutaneous and visceral fat depots. Proteomics. 2010;10:3356–3366.
17. Kalk P, Godes M, Relle K, Rothkegel C, Hucke A, Stasch JP, Hocher B.NO-independent activation of soluble guanylate cyclase prevents diseaseprogression in rats with 5/6 nephrectomy. Br J Pharmacol. 2006;148:853–859.
18. Noronha AM, Liang Y, Hetzel JT, Hasturk H, Kantarci A, Stucchi A,Zhang Y, Nikolajczyk BS, Farraye FA, Ganley-Leal LM. HyperactivatedB cells in human inflammatory bowel disease. J Leukoc Biol. 2009;86:1007–1016.
19. Pradhan AD, Manson JE, Rifai N, Buring JE, Ridker PM. C-reactiveprotein, interleukin 6, and risk of developing type 2 diabetes mellitus.JAMA. 2001;286:327–334.
20. Ridker PM, Rifai N, Stampfer MJ, Hennekens CH. Plasma concentrationof interleukin-6 and the risk of future myocardial infarction amongapparently healthy men. Circulation. 2000;101:1767–1772.
21. Gealekman O, Guseva N, Hartigan C, Apotheker S, Gorgoglione M,Gurav K, Tran KV, Straubhaar J, Nicoloro S, Czech MP, Thompson M,Perugini RA, Corvera S. Depot-specific differences and insufficient sub-cutaneous adipose tissue angiogenesis in human obesity. Circulation.2011;123:186–194.
22. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A,Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays acrucial role in the development of obesity-related insulin resistance.J Clin Invest. 2003;112:1821–1830.
23. Mantovani A, Garlanda C, Locati M. Macrophage diversity and polar-ization in atherosclerosis: a question of balance. Arterioscler ThrombVasc Biol. 2009;29:1419–1423.
24. Ludmer PL, Selwyn AP, Shook TL, Wayne RR, Mudge GH, AlexanderRW, Ganz P. Paradoxical vasoconstriction induced by acetylcholine inatherosclerotic coronary arteries. N Engl J Med. 1986;315:1046–1051.
25. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, FerranteAW Jr. Obesity is associated with macrophage accumulation in adiposetissue. J Clin Invest. 2003;112:1796–1808.
26. Wellen KE, Hotamisligil GS. Inflammation, stress, and diabetes. J ClinInvest. 2005;115:1111–1119.
27. Pou KM, Massaro JM, Hoffmann U, Vasan RS, Maurovich-Horvat P,Larson MG, Keaney JF Jr, Meigs JB, Lipinska I, Kathiresan S, MurabitoJM, O’Donnell CJ, Benjamin EJ, Fox CS. Visceral and subcutaneousadipose tissue volumes are cross-sectionally related to markers of inflam-mation and oxidative stress: the Framingham Heart Study. Circulation.2007;116:1234–1241.
28. Alvehus M, Buren J, Sjostrom M, Goedecke J, Olsson T. The humanvisceral fat depot has a unique inflammatory profile. Obesity (SilverSpring). 2010;18:879–883.
29. Samaras K, Botelho NK, Chisholm DJ, Lord RV. Subcutaneous andvisceral adipose tissue gene expression of serum adipokines that predicttype 2 diabetes. Obesity (Silver Spring). 2010;18:884–889.
30. Esteve E, Castro A, Lopez-Bermejo A, Vendrell J, Ricart W,Fernandez-Real JM. Serum interleukin-6 correlates with endothelial dys-function in healthy men independently of insulin sensitivity. DiabetesCare. 2007;30:939–945.
31. Eder K, Baffy N, Falus A, Fulop AK. The major inflammatory mediatorinterleukin-6 and obesity. Inflamm Res. 2009;58:727–736.
32. Fain JN, Madan AK, Hiler ML, Cheema P, Bahouth SW. Comparison ofthe release of adipokines by adipose tissue, adipose tissue matrix, andadipocytes from visceral and subcutaneous abdominal adipose tissues ofobese humans. Endocrinology. 2004;145:2273–2282.
33. Gao X, Belmadani S, Picchi A, Xu X, Potter BJ, Tewari-Singh N, Capo-bianco S, Chilian WM, Zhang C. Tumor necrosis factor-alpha induces en-dothelial dysfunction in Lepr(db) mice. Circulation. 2007;115:245–254.
34. Wimalasundera R, Fexby S, Regan L, Thom SA, Hughes AD. Effect oftumour necrosis factor-alpha and interleukin 1beta on endothelium-dependent relaxation in rat mesenteric resistance arteries in vitro. Br JPharmacol. 2003;138:1285–1294.
35. Vila E, Salaices M. Cytokines and vascular reactivity in resistancearteries. Am J Physiol Heart Circ Physiol. 2005;288:H1016–H1021.
36. Hung MJ, Cherng WJ, Hung MY, Wu HT, Pang JH. Interleukin-6 inhibitsendothelial nitric oxide synthase activation and increases endothelialnitric oxide synthase binding to stabilized caveolin-1 in human vascularendothelial cells. J Hypertens. 2010;28:940–951.
37. Greenstein AS, Khavandi K, Withers SB, Sonoyama K, Clancy O,Jeziorska M, Laing I, Yates AP, Pemberton PW, Malik RA, HeagertyAM. Local inflammation and hypoxia abolish the protective anticon-tractile properties of perivascular fat in obese patients. Circulation. 2009;119:1661–1670.
38. Virdis A, Santini F, Colucci R, Duranti E, Salvetti G, Rugani I, SegnaniC, Anselmino M, Bernardini N, Blandizzi C, Salvetti A, Pinchera A,Taddei S. Vascular generation of tumor necrosis factor-alpha reducesnitric oxide availability in small arteries from visceral fat of obesepatients. J Am Coll Cardiol. 2011;58:238–247.
39. Withers SB, gabiti-Rosei C, Livingstone DM, Little MC, Aslam R, MalikRA, Heagerty AM. Macrophage activation is responsible for loss ofanticontractile function in inflamed perivascular fat. Arterioscler ThrombVasc Biol. 2011;31:908–913.
40. Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat RevImmunol. 2005;5:953–964.
41. Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switchin adipose tissue macrophage polarization. J Clin Invest. 2007;117:175–184.
42. ron-Wisnewsky J, Tordjman J, Poitou C, Darakhshan F, Hugol D, BasdevantA, Aissat A, Guerre-Millo M, Clement K. Human adipose tissue macro-phages: m1 and m2 cell surface markers in subcutaneous and omental depotsand after weight loss. J Clin Endocrinol Metab. 2009;94:4619–4623.
43. Bourlier V, Zakaroff-Girard A, Miranville A, De BS, Maumus M,Sengenes C, Galitzky J, Lafontan M, Karpe F, Frayn KN, Bouloumie A.Remodeling phenotype of human subcutaneous adipose tissue macro-phages. Circulation. 2008;117:806–815.
44. Cole JE, Georgiou E, Monaco C. The expression and functions of toll-likereceptors in atherosclerosis. Mediators Inflamm. 2010;2010:393946.
45. Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. AnnuRev Immunol. 2011;29:415–445.
46. Pasarica M, Sereda OR, Redman LM, Albarado DC, Hymel DT, Roan LE,Rood JC, Burk DH, Smith SR. Reduced adipose tissue oxygenation in humanobesity: evidence for rarefaction, macrophage chemotaxis, and inflammationwithout an angiogenic response. Diabetes. 2009;58:718–725.
47. Romero-Corral A, Sert-Kuniyoshi FH, Sierra-Johnson J, Orban M, Gami A,Davison D, Singh P, Pusalavidyasagar S, Huyber C, Votruba S, Lopez-Jimenez F, Jensen MD, Somers VK. Modest visceral fat gain causes endo-thelial dysfunction in healthy humans. J Am Coll Cardiol. 2010;56:662–666.
48. Flum DR, Belle SH, King WC, Wahed AS, Berk P, Chapman W, PoriesW, Courcoulas A, McCloskey C, Mitchell J, Patterson E, Pomp A, StatenMA, Yanovski SZ, Thirlby R, Wolfe B. Perioperative safety in thelongitudinal assessment of bariatric surgery. N Engl J Med. 2009;361:445–454.
49. Hotamisligil GS. Inflammation and metabolic disorders. Nature. 2006;444:860–867.
Farb et al Visceral Adiposity and Vascular Function 473
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Donald T. Hess, Joseph A. Vita and Noyan GokceWidlansky, Sherman J. Bigornia, Antonino J. Fiscale, Caroline M. Apovian, Brian Carmine, Melissa G. Farb, Lisa Ganley-Leal, Melanie Mott, YanMei Liang, Bahadir Ercan, Michael E.
Arteriolar Function in Visceral Adipose Tissue Is Impaired in Human Obesity
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2011 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.111.2358462011;
2012;32:467-473; originally published online November 17,Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/32/2/467World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2011/11/17/ATVBAHA.111.235846.DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 4, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Supplement Material
Arteriolar function in visceral adipose tissue is impaired in human obesity
Melissa G. Farb PhD, Lisa Ganley-Leal PhD, Melanie Mott, YanMei Liang, Bahadir Ercan PhD,
Michael E. Widlansky MD, Sherman J. Bigornia MA, Antonino J. Fiscale, Caroline M. Apovian
MD, Brian Carmine MD, Donald T. Hess MD, Joseph A. Vita MD, Noyan Gokce MD
Supplemental Figure I Supplemental Figure II Supplemental Figure III Supplemental Figure IV Supplemental Figure V Supplemental Figure VI Supplemental Table I

Supplemental Figure I: Adipose tissues arteriolar responses, paired data. In a subset of
individuals (n=10) that provided paired samples simultaneously from both visceral and
subcutaneous depots, endothelium-dependent vasorelaxation of visceral arterioles was
severely impaired, similar in magnitude to results for the group as a whole (p<0.001 by
ANOVA). Data presented as mean ± SEM.
020406080
100120
% v
esse
l dila
tion
Subcutaneous Visceral
Ach LOG (M)
ET-1 PAP -10 -9 -8 -7 -6 -5
*p<0.001

Supplemental Figure II: Adipose tissue arteriolar responses in non-diabetic (n=17)
versus diabetic subjects (n=13). Visceral microvascular dilation was severely impaired
compared to subcutaneous vasorelaxation irrespective of clinical diabetes status (p<0.001
by ANOVA). Data presented as mean ± SEM. DM=diabetes mellitus
Ach LOG (M)
ET-1 PAP-10 -9 -8 -7 -6 -5
020406080
100120
% v
esse
l dila
tion
Subcutaneous non-DM Subcutaneous DM Visceral non-DM Visceral DM
*p<0.001

Supplemental Figure III: Adipose arteriolar histology. Representative cross-sectional histology of
arteriolar staining for (A) endothelium-specific CD31 (B) H&E, demonstrating architecturally intact
nucleated endothelium (indicated by arrows) surrounded by smooth muscle layers demonstrating
endothelial integrity during experimental conditions.
A. B. A. B.

Supplemental Figure IV: Representative cross-sectional histology of adipose arterioles
stained for (A) elastica-van Gieson, and (B) CD68, demonstrating no evidence of
atherosclerotic changes or vascular macrophage infiltration, respectively.
A. B.

Supplemental Figure V: Adipose tissues arteriolar responses to sodium nitroprusside (SNP). No
difference in endothelium-independent, SNP-mediated vasodilation was observed between visceral as
compared to subcutaneous adipose arterioles. (n=6, p=0.5 by ANOVA). Data presented as mean ±
SEM.
SNP LOG (M)
ET-1 PAP -10 -8 -7 -4-6 -5-9
020406080
100120
% v
esse
l dila
tion
Subcutaneous Visceral
p=NS

Supplemental Figure VI: Vascular endothelial cell populations isolated from visceral fat exhibited
higher expression of inflammatory cytokines as compared to the subcutaneous depot (n=6). Data are
presented as fold difference in visceral compared to subcutaneous expression ± SEM. * p<0.05.
CCL5 IL6 TLR40
2
4
6
Rel
ativ
e G
ene
Expr
essi
on
Subcutaneous Visceral*
**
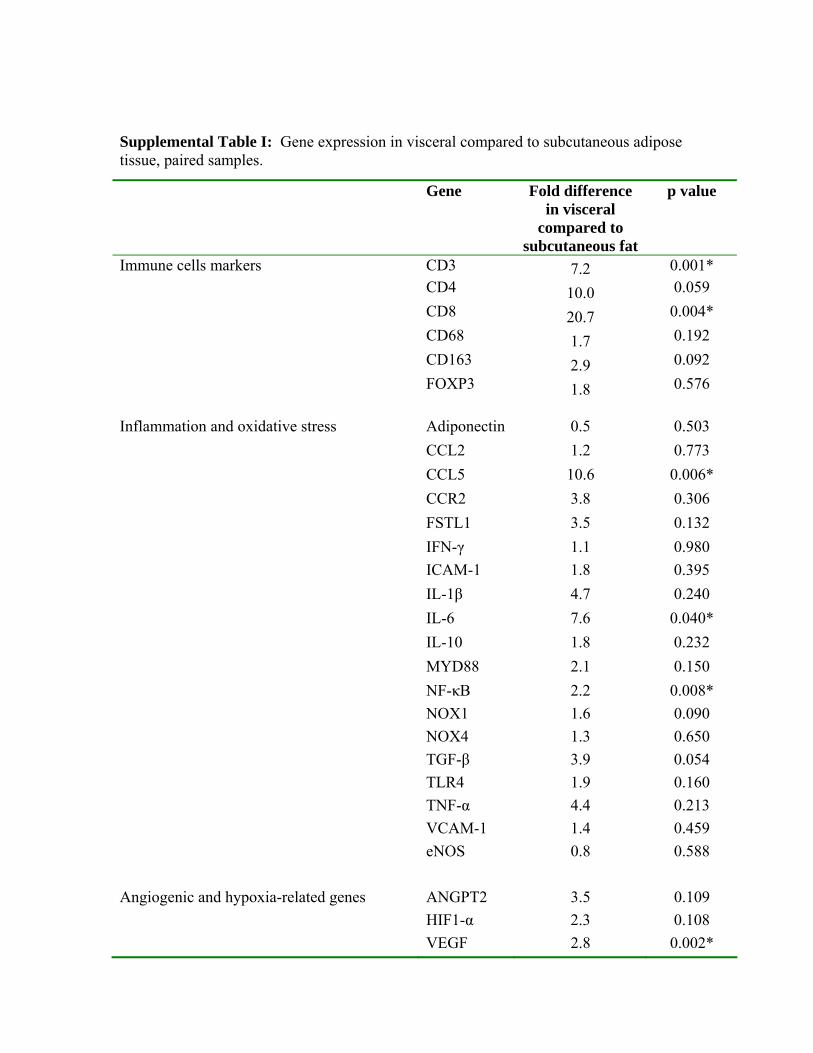
Supplemental Table I: Gene expression in visceral compared to subcutaneous adipose tissue, paired samples.
Gene Fold difference in visceral
compared to subcutaneous fat
p value
Immune cells markers CD3 7.2 0.001* CD4 10.0 0.059 CD8 20.7 0.004* CD68 1.7 0.192 CD163 2.9 0.092 FOXP3 1.8 0.576 Inflammation and oxidative stress Adiponectin 0.5 0.503 CCL2 1.2 0.773 CCL5 10.6 0.006* CCR2 3.8 0.306 FSTL1 3.5 0.132 IFN-γ 1.1 0.980 ICAM-1 1.8 0.395 IL-1β 4.7 0.240 IL-6 7.6 0.040* IL-10 1.8 0.232 MYD88 2.1 0.150 NF-κB 2.2 0.008* NOX1 1.6 0.090 NOX4 1.3 0.650 TGF-β 3.9 0.054 TLR4 1.9 0.160 TNF-α 4.4 0.213 VCAM-1 1.4 0.459 eNOS 0.8 0.588 Angiogenic and hypoxia-related genes ANGPT2 3.5 0.109 HIF1-α 2.3 0.108 VEGF 2.8 0.002*
















![Adipokines and the role of visceral adipose tissue in infl ......adipokines show correlations with the activity of a variety of autoimmune as well as infectious diseases [13,47]. Given](https://static.fdocuments.net/doc/165x107/5f704ebef0a62560e40f3170/adipokines-and-the-role-of-visceral-adipose-tissue-in-infl-adipokines-show.jpg)

