
Anatomical distribution of NPY-like immunoreactivity in the domestic chick brain (Gallus domesticus)
16
Anatomical Distribution of NPY-Like Immunoreactivity in the Domestic Chick Brain (Gallus domesticus) VINCENZO ESPOSITO,* GAETANO VINCENZO PELAGALLI, PAOLO DE GIROLAMO, AND GIULIANA GARGIULO Dipartimento di Strutture, Funzioni e Tecnologie Biologiche, Universita’ di Napoli Federico II, 80137 Napoli, Italia ABSTRACT Neuropeptide Y-immunoreactive (NPY-ir) fibers and neurons in the brain of the domestic chick (Gallus domesticus) were described using an immunohistochemical technique. NPY-ir neurons were seen in the lobus parolfactorius; hyperstria- tum, neostriatum, paleostriatum, and archistriatum; hippocampal and parahippocampal areas; dorsolateral corticoid area; piriform cortex; two thalamic areas contiguous to the n. rotundus; n. dorsolateralis anterior thalami, pars lateralis, and pars magnocellularis; n. periventricularis hypothalami; n. paraventricularis magnocellularis; regio lateralis hypothalami; n. infundibuli; inner zone of the median eminence; dorsal and lateral portions of the n. opticus basalis; n. raphes; and n. reticularis paramedianus. NPY-ir fibers were seen throughout the entire chick brain, but were more abundant in the hypothalamus where they formed networks and pathways. They were also observed in some circumventricular organs. The anatomical data of the present study regarding the distribution of NPY ir in the chick brain, together with the physiological findings of other studies, suggest that NPY plays a key role in the regulation of the neuroendocrine, vegetative, and sensory systems of birds by acting as a neuromodulator and/or neurotransmitter. Anat Rec 263:186 –201, 2001. © 2001 Wiley-Liss, Inc. Key words: neuropeptide Y (NPY); avian brain; immunohistochemistry; domestic chick; Gallus do- mesticus; neuromodulators; neurotransmitters Abbreviations: AA, archistriatum anterior; AId, archistri- atum intermedium, pars dorsalis; AIv, archistriatum interme- dium, pars ventralis; AL, ansa lenticularis; AM, nucleus anterior medialis hypothalami; Ap, archistriatum posterior; APH, area parahippocampalis; BCS, brachium colliculi superioris; BO, bul- bus olfactorius; CA, commissura anterior; Cb, cerebellum; CHCS, tractus cortico-habenularis et cortico-septalis; CDL, area corti- coidea dorsolateralis; CO, chiasma opticum; CP, commissura pos- terior; CPa, commissura pallii; CPi, cortex piriformis; CTz, corpus trapezoideum; DLAl, nucleus dorsolateralis anterior thalami, pars lateralis; DLAmc, nucleus dorsolateralis anterior thalami, pars magnocellularis; E, ectostriatum; FLM, fasciculus longitu- dinalis medialis; FPL, fasciculus prosencephali lateralis; FV, fu- niculus ventralis; GCt, substantia grisea centralis; GLv, nucleus geniculatus lateralis, pars ventralis; GLdp, nucleus geniculatus lateralis, pars dorsalis principalis; HA, hyperstriatum accesso- rium; HD, hyperstriatum dorsale; HIS, hyperstriatum inter- calatum supremum; HM, nucleus habenularis medialis; Hp, hippocampus; HV, hyperstriatum ventrale; ICo, nucleus inter- collicularis; IH, nucleus inferioris hypothalami; IN, nucleus in- fundibuli; INP, nucleus intrapeduncularis; LA, nucleus lateralis anterior thalami; LHy, regio lateralis hypothalami; LPO, lobus parolfactorius; ME, median eminence; MM, nucleus mamillaris medialis; MPO, nucleus magnocellularis preopticus; (d), pars dor- salis; (m), pars medialis; (v), pars ventralis; N, neostriatum; NF, neostriatum frontale; NC, neostriatum cau dale; NH, neurohy- pophysis; NI, neostriatum intermedium; NIII, nervus oculomoto- rius; NIV, nervus trochlearis; NV, nervus trigeminus; NVI, ner- vus abducens; NVIIIv, nervus octavus, pars vestibularis; nBOR, nucleus opticus basalis; nBORd, nucleus opticus basalis, pars dorsalis; nBORl, nucleus opticus basalis, pars lateralis; nCPa, nucleus commissura pallii; NIX, nucleus nervi glossopharyngei; OM, tractus occipitomesencephalicus; OVLT, organum vasculo- sum lamina terminalis; P, glandula pinealis; PA, paleostriatum augmentatum; PHN, nucleus periventricularis hypothalami; POM, nucleus preopticus medialis; POP, nucleus preopticus periventricularis; PP, paleostriatum primitivum; PVN, nucleus paraventricularis magnocellularis; PVO, organum paraventricu- lare; PVT, paleostriatum ventrale; QF, tractus quintofrontalis; R, nucleus raphes; ROT, nucleus rotundus; RpaM, nucleus reticu- laris paramedianus; RPO, nucleus reticularis pontis oralis; SCNm, nucleus suprachiasmaticus, pars medialis; SCO, organum subcommissurale; SGFS, stratum griseum et fibrosum superfi- ciale; SL, nucleus septalis lateralis; SM, nucleus septalis medialis; Sme, stria medullaris; SO, stratum opticum; SOv, nucleus supraop- ticus, pars ventralis; SSO, organum subseptale; TeO, optic tectum; TIO, tractus isthmo-opticus; TPO, area temporo-parieto-occipitalis; TrO, tractus opticus; TSM, tractus septomesencephalicus; TT, trac- tus tectothalamicus; 3V, third ventricle; VeM, nucleus vestibularis medialis; VL, ventriculus lateralis; VLT, nucleus ventrolateralis thalami; VMN, nucleus ventromedialis hypothalami; VT, ventricu- lus tecti mesencephali. *Correspondence to: Dr. Vincenzo Esposito, Dipartimento di Strutture, Funzioni e Tecnologie Biologiche, Universita’ di Napoli Federico II, Via Veterinaria, 1-80137 Napoli, Italia. E-mail [email protected] Received 19 June 2000; Accepted 7 February 2001 Published online 00 Month 2001 THE ANATOMICAL RECORD 263:186 –201 (2001) © 2001 WILEY-LISS, INC.
-
Upload
vincenzo-esposito -
Category
Documents
-
view
215 -
download
1
Transcript of Anatomical distribution of NPY-like immunoreactivity in the domestic chick brain (Gallus domesticus)

Anatomical Distribution of NPY-LikeImmunoreactivity in the DomesticChick Brain (Gallus domesticus)
VINCENZO ESPOSITO,* GAETANO VINCENZO PELAGALLI,PAOLO DE GIROLAMO, AND GIULIANA GARGIULO
Dipartimento di Strutture, Funzioni e Tecnologie Biologiche, Universita’ di NapoliFederico II, 80137 Napoli, Italia
ABSTRACTNeuropeptide Y-immunoreactive (NPY-ir) fibers and neurons in the brain of the domestic chick (Gallus domesticus)
were described using an immunohistochemical technique. NPY-ir neurons were seen in the lobus parolfactorius; hyperstria-tum, neostriatum, paleostriatum, and archistriatum; hippocampal and parahippocampal areas; dorsolateral corticoid area;piriform cortex; two thalamic areas contiguous to the n. rotundus; n. dorsolateralis anterior thalami, pars lateralis, and parsmagnocellularis; n. periventricularis hypothalami; n. paraventricularis magnocellularis; regio lateralis hypothalami; n.infundibuli; inner zone of the median eminence; dorsal and lateral portions of the n. opticus basalis; n. raphes; and n.reticularis paramedianus. NPY-ir fibers were seen throughout the entire chick brain, but were more abundant in thehypothalamus where they formed networks and pathways. They were also observed in some circumventricular organs. Theanatomical data of the present study regarding the distribution of NPY ir in the chick brain, together with the physiologicalfindings of other studies, suggest that NPY plays a key role in the regulation of the neuroendocrine, vegetative, and sensorysystems of birds by acting as a neuromodulator and/or neurotransmitter. Anat Rec 263:186–201, 2001.© 2001 Wiley-Liss, Inc.
Key words: neuropeptide Y (NPY); avian brain; immunohistochemistry; domestic chick; Gallus do-mesticus; neuromodulators; neurotransmitters
Abbreviations: AA, archistriatum anterior; AId, archistri-atum intermedium, pars dorsalis; AIv, archistriatum interme-dium, pars ventralis; AL, ansa lenticularis; AM, nucleus anteriormedialis hypothalami; Ap, archistriatum posterior; APH, areaparahippocampalis; BCS, brachium colliculi superioris; BO, bul-bus olfactorius; CA, commissura anterior; Cb, cerebellum; CHCS,tractus cortico-habenularis et cortico-septalis; CDL, area corti-coidea dorsolateralis; CO, chiasma opticum; CP, commissura pos-terior; CPa, commissura pallii; CPi, cortex piriformis; CTz, corpustrapezoideum; DLAl, nucleus dorsolateralis anterior thalami,pars lateralis; DLAmc, nucleus dorsolateralis anterior thalami,pars magnocellularis; E, ectostriatum; FLM, fasciculus longitu-dinalis medialis; FPL, fasciculus prosencephali lateralis; FV, fu-niculus ventralis; GCt, substantia grisea centralis; GLv, nucleusgeniculatus lateralis, pars ventralis; GLdp, nucleus geniculatuslateralis, pars dorsalis principalis; HA, hyperstriatum accesso-rium; HD, hyperstriatum dorsale; HIS, hyperstriatum inter-calatum supremum; HM, nucleus habenularis medialis; Hp,hippocampus; HV, hyperstriatum ventrale; ICo, nucleus inter-collicularis; IH, nucleus inferioris hypothalami; IN, nucleus in-fundibuli; INP, nucleus intrapeduncularis; LA, nucleus lateralisanterior thalami; LHy, regio lateralis hypothalami; LPO, lobusparolfactorius; ME, median eminence; MM, nucleus mamillarismedialis; MPO, nucleus magnocellularis preopticus; (d), pars dor-salis; (m), pars medialis; (v), pars ventralis; N, neostriatum; NF,neostriatum frontale; NC, neostriatum cau dale; NH, neurohy-pophysis; NI, neostriatum intermedium; NIII, nervus oculomoto-rius; NIV, nervus trochlearis; NV, nervus trigeminus; NVI, ner-vus abducens; NVIIIv, nervus octavus, pars vestibularis; nBOR,nucleus opticus basalis; nBORd, nucleus opticus basalis, pars
dorsalis; nBORl, nucleus opticus basalis, pars lateralis; nCPa,nucleus commissura pallii; NIX, nucleus nervi glossopharyngei;OM, tractus occipitomesencephalicus; OVLT, organum vasculo-sum lamina terminalis; P, glandula pinealis; PA, paleostriatumaugmentatum; PHN, nucleus periventricularis hypothalami;POM, nucleus preopticus medialis; POP, nucleus preopticusperiventricularis; PP, paleostriatum primitivum; PVN, nucleusparaventricularis magnocellularis; PVO, organum paraventricu-lare; PVT, paleostriatum ventrale; QF, tractus quintofrontalis; R,nucleus raphes; ROT, nucleus rotundus; RpaM, nucleus reticu-laris paramedianus; RPO, nucleus reticularis pontis oralis;SCNm, nucleus suprachiasmaticus, pars medialis; SCO, organumsubcommissurale; SGFS, stratum griseum et fibrosum superfi-ciale; SL, nucleus septalis lateralis; SM, nucleus septalis medialis;Sme, stria medullaris; SO, stratum opticum; SOv, nucleus supraop-ticus, pars ventralis; SSO, organum subseptale; TeO, optic tectum;TIO, tractus isthmo-opticus; TPO, area temporo-parieto-occipitalis;TrO, tractus opticus; TSM, tractus septomesencephalicus; TT, trac-tus tectothalamicus; 3V, third ventricle; VeM, nucleus vestibularismedialis; VL, ventriculus lateralis; VLT, nucleus ventrolateralisthalami; VMN, nucleus ventromedialis hypothalami; VT, ventricu-lus tecti mesencephali.
*Correspondence to: Dr. Vincenzo Esposito, Dipartimento diStrutture, Funzioni e Tecnologie Biologiche, Universita’ di NapoliFederico II, Via Veterinaria, 1-80137 Napoli, Italia.E-mail [email protected]
Received 19 June 2000; Accepted 7 February 2001Published online 00 Month 2001
THE ANATOMICAL RECORD 263:186–201 (2001)
© 2001 WILEY-LISS, INC.

Neuropeptide Y (NPY) is an amidated 36-amino-acidpeptide that was first isolated from, and characterized for,porcine brain (Tatemoto, 1982; Tatemoto et al., 1982). Itbelongs to a family of structurally related peptides, someof which are found in all vertebrates (peptide YY), othersonly in tetrapods (pancreatic polypeptide), and still othersonly in certain fish species (peptide Y and peptide methi-onine-tyrosine) (Conlon et al., 1991, 1994; Larhammar etal., 1993, 1995, 1998; Larhammar, 1996). The molecularsequence of NPY shows a remarkable degree of conserva-tion across all studied mammalian species, the chicken,frog, goldfish, and the ray. In particular, it is 92% homol-ogous between cartilaginous fish and mammals, and itdiffers by only one amino acid in the chicken compared tothe rat or human (Chartrel et al., 1991; Blomqvist et al.,1992; Larhammar et al., 1992, 1993; Larhammar, 1996).In addition, the molecular forms of certain receptors thatare selectively activated by NPY are also highly conservedin vertebrates (Hoyle, 1999).
In mammals, NPY is distributed throughout the centraland peripheral nervous systems where it is involved inseveral different biological processes (for review seeColmers and Wahlestedt, 1993; Heilig and Widerlov,1995). The distribution of NPY has also been studied inthe brain of avian species; however, the majority of thesestudies are limited to specific areas of the avian brain, andonly one deals with the whole brain of an avian, theJapanese quail (Aste et al., 1991).
Physiological studies of NPY in avian species suggestthat NPY is involved in the regulation of feeding behav-iour (Kuenzel et al., 1987; Lee et al., 1994; Richardson etal., 1995), insulin secretion (Kuenzel and McMurtry,1988), sexual maturation (Fraley and Kuenzel, 1993;Walsh and Kuenzel, 1997), and GnRH release (Contijochet al., 1993; Barker-Gibb et al., 1995).
The present study provides data regarding the distribu-tion of NPY immunoreactive neuronal structures through-out the whole brain of the chick (Gallus domesticus),which can serve as a basis to better understand the phys-iological functions of NPY in avian species, and, as aconsequence, the general rule underlying the distributionof NPY.
MATERIALS AND METHODSTen brains of 2-week-old male chicks (Gallus domesti-
cus) were obtained and processed as previously described(Esposito et al., 1993), in order to have 5–7-mm sagittaland coronal paraffin sections.
For immunohistochemistry, deparaffinized and rehy-drated sections were initially washed in 0.01 M phos-phate-buffered saline (PBS; pH 7.4), subsequently incu-bated in 3% H2O2 for 20 min at room temperature (RT) toinhibit endogenous peroxidase activity, rinsed for 10 minin PBS, and then incubated in normal goat serum (Dako,Denmark) diluted 1:5 in PBS for 30 min at RT to reducebackground staining. The sections were then exposed tothe specific antiserum (rabbit anti-synthetic porcine NPYantiserum, IHC 7172, Peninsula Lab) at a dilution of1:2,000 in PBS for 24 hr at 4°C. This antiserum is knownto be highly specific for mammalian NPY and does notcross-react with PYY, PP, VIP, Amylin, prepro NPY 68–97, insulin, and somatostatin. Furthermore, it has beenrecently used successfully for the detection of NPY immu-noreactivity in the avian central nervous system (Medinaet al., 1998). After several rinses in PBS, the sections were
incubated for 30 min at RT with anti-rabbit IgG (Dako)diluted 1:50 in PBS, washed in PBS, and subsequentlytreated for 30 min at RT with the peroxidase-antiperoxi-dase (PAP) complex (Dako) diluted 1:100 in PBS. All in-cubations were performed in moist chambers. The perox-idase reaction was revealed with a fresh solutioncontaining 10 mg of 3,39 diaminobenzidine in 15 ml of 0.5M Tris-buffer, pH 7.6, and 1.5 ml of H2O2 at 0.03%. Someof the sections were counterstained with Cresyl violet. Foranatomical identification of the nervous structures, somedeparaffinized and rehydrated sections were stained withLuxol fast blue/Cresyl violet. All sections were dehydratedand coverslipped, and then observed and photographedusing a Leitz Aristoplan.
ControlsThe specificity of the immunoreactivity was tested by
successively substituting either the NPY antiserum, theanti-rabbit IgG, or the PAP complex with a buffer, inrepeated trials. Furthermore, the primary antibody wasalso replaced by normal rabbit serum (Dako).
The cross-reactivity of the primary antibody was testedby incubation of sections with antibody that had beenpreabsorbed with an excessive amount of homologous an-tigen (synthetic NPY, Peninsula Lab.; up to 50 mg/ml inthe final dilution).
None of the negative controls performed yielded specificimmunostaining. Moreover, the incubations of NPY anti-serum preabsorbed with its homologous antigen showedno immunoreactivity.
RESULTSImmunoreactive cells and fibers are described for the
coronal sections that progress from the rostral to the cau-dal brain, and for the sagittal sections. Their distributionsin the brain are shown in Figure 1. The nomenclatureused in the present study is based on the sterotaxis atlasof the chicken (Kuenzel and Masson, 1988).
TelencephalonThe olfactory bulb did not display NPY immunoreactiv-
ity. The lobus parolfactorius (LPO) showed few NPY-irneurons located mainly in the most rostral and ventralparts, and few immunoreactive fibers (Figs. 1B, R, 2A). Inthe hyperstriatum and neostriatum, few labeled fiberswere observed overall; small bipolar and triangular im-munostained neurons were observed in the peripheralarea (Figs. 1A–G, 2B, D). The paleostriatum showed im-munoreactive perikarya in an area lateral to the tractusseptomesencephalicus (TSM) and ventral to the fasciculusprosencephali lateralis (FPL), and a few immunostainedneurons in the paleostriatum ventrale (PVT) (Figs. 1C, D,2E). The hippocampus (Hp), the parahippocampal area(APH), the dorsolateral corticoid area (CDL), and the piri-form cortex (CPi) contained numerous NPY-immunoposi-tive neurons that were either fusiform or triangular inshape. NPY-ir fibers were rarely present (Figs. 1E–L,O–R, 2C). The archistriatum intermedium, pars ventralis(AIv) showed a small number of labeled cells (Fig. 1F–H).The septum contained NPY-ir fibers that were primarilylocated in the lateral septal nucleus (SL), from where theyran ventrally to join those of the pallial commissure (CPa).In addition, the nucleus of the pallial commissure (nCPa)also contained NPY-ir fibers (Figs. 1C–F, Q, 3A,B). The
187NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN

corticohabenular and corticoseptal tract (CHCS) had im-munoreactive fibers, which ran caudally to the ventrolat-eral region, and then gradually disappeared (Figs. 1G,3C).
DiencephalonIn the thalamus, an extensive NPY-ir cell population
was observed (Figs. 1F–H, O–P, 4, 5). The majority of thecells were bipolar; the others were either oval or round.Their overall distribution, based on a 3-dimensional re-construction of the coronal and sagittal sections, was di-vided into two general areas contiguous to the nucleusrotundus (ROT): (1) along anterior lateral and anterior
medial projections that proceeded away from the ventralanterior border of the ROT, and terminated, respectively,along the dorsal border of the nucleus lateralis anteriorthalami (LA), and along the dorsal borders of both thenucleus ventrolateralis thalami (VLT) and nucleusgeniculatus lateralis, pars ventralis (GLv) (Figs. 1F–G,4A); and (2) around the antero-lateral, lateral, medial,ventral, and dorsal borders of the ROT (Figs. 1H, P, 4B).Regarding the lateral, medial, and ventral borders of theROT, the anterior coronal sections showed cell clusters(Fig. 5A), whereas the posterior coronal sections showedonly a single strata of cells. The cells near the antero-lateral border of the ROT seemed to belong to the nucleus
Fig. 1. Schematic drawings of 12 coronal rostro-caudal (A–N) and 4parasagittal latero-medial (O–R) sections of the chick brain showing arepresentation of the immunohistochemical distribution of neuropeptideY (NPY). On the right side of each section, NPY-ir cell bodies are
represented by dots and NPY ir-fibers are represented by little triangles.On the left side are shown the locations of neuronal structures identifiedin luxol fast blue-cresyl violet-stained sections. For abbreviations, seepage 186.
188 ESPOSITO ET AL.

dorsolateralis anterior thalami, pars magnocellularis(DLAmc) (Figs. 1F, 5B), and to the nucleus dorsolateralisanterior thalami, pars lateralis (DLAl) (Figs. 1H, O, 5C).
NPY-ir fibers intermingled with all cells of areas (1) and(2). In addition, fibers were observed in the dorsal andperiventricular zones of the thalamus; the fibers of thelatter zone extended into the hypothalamic periventricu-lar region (Figs. 1G–I, 3C).
In the hypothalamus, the majority of the NPY-ir cellswere observed along the walls of the third ventricle (3V)
inside the nucleus periventricularis hypothalami (PHN)(Fig. 1E–H, 6). Additional cells were seen in the nucleusparaventricularis magnocellularis (PVN) (Figs. 1G, H, R,6C) and a few, small cells were present in the regio late-ralis hypothalami (LHy). In the posterior hypothalamicregion, NPY-ir cells were localized in the most caudalportion of the nucleus infundibuli (IN) (Figs. 1I, R, 7A),and in the zona interna of the median eminence (ME)(Figs. 1I, R, 7B). These latter cells were fusiform with theiraxes frequently running parallel to the ependymal layer.
Fig. 1. (continued)
189NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN

Immunopositive fibers were observed throughout theentire hypothalamus. They were densely distributed alongthe 3V walls, and formed networks in the nuclei of su-praopticus, pars ventralis (SOv), magnocellularis preopti-
cus (MPO), preopticus periventricularis (POP), suprachi-asmaticus, pars medialis (SCNm), PVN, and IN (Figs.1C–I, R, 8A). Basket-like NPY-ir fiber terminals sur-rounded the immunonegative neurons of the PVN (Figs.
Fig. 1. (continued)
190 ESPOSITO ET AL.

Fig. 2. Light micrographic localization of NPY-ir within the chicktelencephalon. PAP method. A: Immunoreactive cells and fibers in theLPO, parasagittal section, 3250. Left side is rostral. B: Medial portion ofthe HA in coronal section showing ir neurons and fibers, 3400. C:Parasagittal section of the hippocampal complex showing ir cells and
fibers. 3250. Left side is rostral. D: Rostral portion of the Hp showing irneurons and fibers, coronal section 3 400. E: Immunoreactive neuronsand fibers in the paleostriatal region located under the FPL, coronalsection 3400. Left side is lateral. For abbreviations, see page 186.
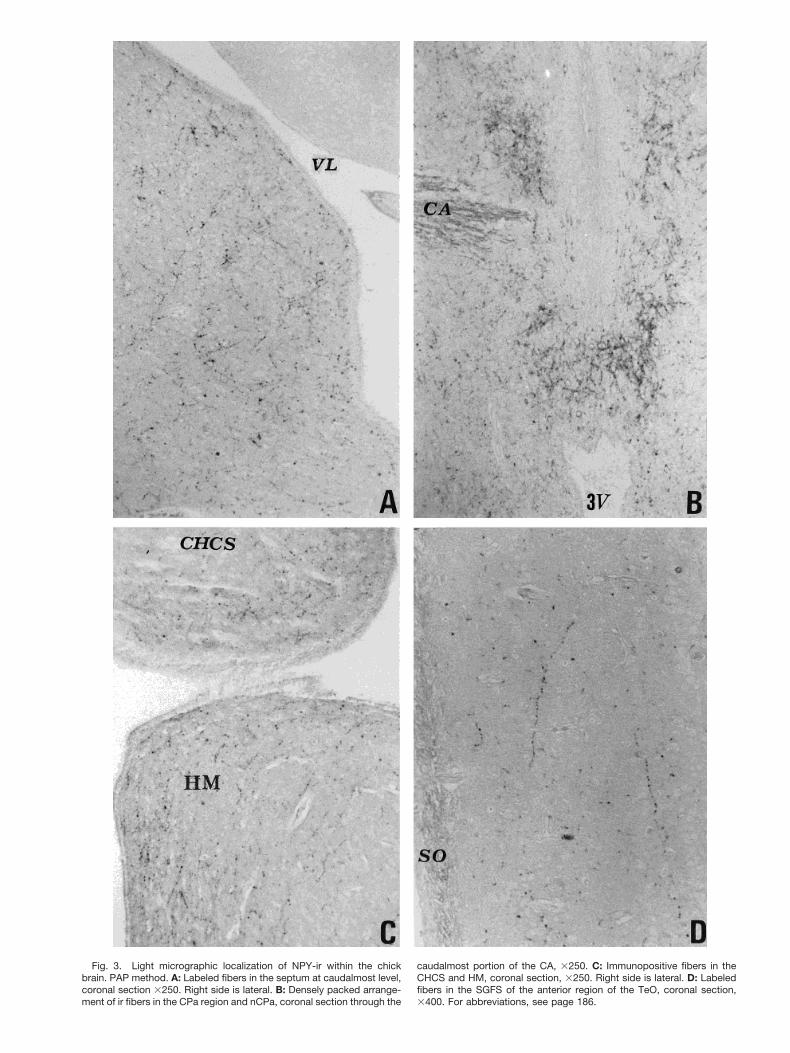
Fig. 3. Light micrographic localization of NPY-ir within the chickbrain. PAP method. A: Labeled fibers in the septum at caudalmost level,coronal section 3250. Right side is lateral. B: Densely packed arrange-ment of ir fibers in the CPa region and nCPa, coronal section through the
caudalmost portion of the CA, 3250. C: Immunopositive fibers in theCHCS and HM, coronal section, 3250. Right side is lateral. D: Labeledfibers in the SGFS of the anterior region of the TeO, coronal section,3400. For abbreviations, see page 186.

1F, R, 8B). The zona interna and the zona externa of theME also contained labelled fibers (Figs. 1I–L, R, 7B).
The brain parasagittal sections exhibited two NPY-irfiber pathways: (1) in the ventral hypothalamus, from thepreoptic area just above the optic chiasm (CO) to the ME;and (2) in the dorsal hypothalmus, from the PVN region tothe tubero-infundibular region (Figs. 1R, 9). The coronalsections also showed a fiber pathway running from the INtoward the median eminence (Fig. 1I).
BrainstemThe brainstem coronal sections exhibited three NPY-ir
cell groups. The first group was localized dorsally to thenucleus opticus basalis (nBOR) and consisted of polygonalor fusiform neurons (Figs. 1M, Q, 10A, B). The cells of thesecond and third groups were located in the caudal pons;they extended from the nervus abducens (NVI) to thenucleus nervi glossopharyngei (nIX), and were containedin, respectively, the nucleus reticularis paramedianus(RPaM) and the nucleus raphes (R) (Figs. 1N, Q, 10C). Thecells of these latter two groups were either polygonal oraffusolate in shape.
The NPY-ir fibers located in the whole brainstem werelocalized in the ventral portion, and were less denselypacked than those of the other brain regions (Fig. 1N,Q,R).Fibers were also located in the nBOR (Fig. 1M, Q, 10A), inthe stratum griseum et fibrosum superficiale (SGFS) ofthe optic tectum (TeO) (Figs. 1I–M, 3D), and in the areabelow the ventriculus tecti mesencephali (VT), i.e., wherethe nucleus intercollicularis (ICo) is located (Fig. 1L, M).
Circumventricular OrgansSome circumventricular organs showed NPY immuno-
reactivity. Specifically, NPY-ir fibers were located in theorganum vasculosum laminae terminalis (OVLT), in theorganum subcommissurale (SCO), and near the organumsubseptale (SSO) (Fig. 8C–E).
DISCUSSIONThe present immunohistochemical study shows, for the
first time, the distribution of NPY immunoreactivitywithin the whole domestic chick brain. Our data will bediscussed both in relation to the only other data regardingNPY distribution in the whole brain of an avian species,i.e., the quail, and in relation to data regarding NPYdistribution in circumscribed brain regions in other avianspecies.
Results of the present study showed NPY-ir perikaryain the chick LPO. This accords with observations ofNPY-ir in pigeon forebrain (Anderson and Reiner, 1990),but not in quail (Aste et al., 1991). The LPO of the domes-tic chick plays a key role in the storage of informationconcerning the passive avoidance response (Gilbert et al.,1991; Rose, 1991). Thus, it is possible that NPY is involvedin this function.
In accord with data reported for the quail (Aste et al.,1991) and the pigeon (Anderson and Reiner, 1990), ourobservations found NPY-ir cells in the local circuits of thecortex- and striatal-equivalent parts of the avian telen-cephalon. In addition, the hippocampal NPY-ir perikaryaobserved in our study resembled the multi-angular/ovoidlocal circuit neurons described in chick and pigeon hip-pocampus (Tombol et al., 2000).
Our description of the thalamic NPY-ir cell complex issimilar to descriptions reported in other studies of chicken
Fig. 4. Light micrographic localization of NPY-ir within the chickthalamus. PAP method. A: Labeled neurons in the area surrounded bythe TSM, AL, LA, and VLT, coronal section, 3125. Left side is lateral. B:Parasagittal section showing the arrangement of the ir neurons aroundthe ROT, 3100. Left side is rostral. For abbreviations, see page 186.
193NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN

Fig. 5. Light micrographic localization of NPY-ir within the chick thalamus. PAP method. A: Immunopo-sitive cell cluster ventral to the ROT, coronal section, 3250. Left side is lateral. B: Labeled neurons and fibersin the DLAl, coronal section, 3250. Right side is lateral. C: Some ir neurons dorsally bordering the FPL,coronal section, 3250. Right side is lateral. For abbreviations, see page 186.

Fig. 6. Light micrographic localization of NPY-ir within the chickhypothalamus. PAP method. A: Labeled neurons and fibers in the ros-tralmost portion of the PHN, coronal section, 3250. B: Labeled neuronsand fibers in the medial portion of the PHN, coronal section, 3250. The
major axis of the cells is prevalently parallel to the ventricle surface. C:Labeled neurons and fibers in the PVN, coronal section, 3300. Forabbreviations, see page 186.
195NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN

(Kuenzel and McMurtry, 1988; Walsh and Kuenzel, 1997)and quail (Aste et al., 1991), with the exception that theselatter studies did not observe immunoreactive cells in theDLAl and DLAmc. Our observations also agree with thosemade for the pigeon thalamus regarding an area of NPYcells between the GLv and the principal optic nucleus ofthe dorsolateral thalamus that encircles the rostral pole ofROT and LA. These cells (called the nucleus of the mar-
ginal optic tract) projected into the optic tectum, and,hence, might be the avian equivalent of the mammalianintergeniculate leaflet (Gunturkun and Karten, 1991).The NPY-positive somata in the pigeon intergeniculatearea and the few neurons in the dorsolateral margin of theGLv and in DLAmc have been shown to represent themain, and perhaps exclusive, source of NPY-ir fiberswithin the pigeon nBOR (Hamassaki and Britto, 1990a).This intergeniculate area is also the source of NPY-irfibers that occur in the suprachiasmatic nucleus and inmany other retinorecipient and visually related areas inthe pigeon brain (Hamassaki and Britto, 1990b). Based onthese findings, and on our results that NPY-ir fibers andterminals are located in the optic tectum, in the SCNmand nBOR, and that NPY-ir cells are located in the tha-lamic NPY cell complex, it may be hypothesized that NPYplays a role in the functional organization of the visualsystem of the chick brain. In addition, the localization ofvasotocin-ir fibers and neurons in the area surroundingthe ROT of the chick brain (Sugita, 1994) suggests anavenue of future research regarding the interaction be-tween NPY and vasotocin.
The distributions of NPY-ir cells and fibers in the hypo-thalamus were similar to those observed by Kuenzel andMcMurtry (1988) in chick. Our findings, however, showeda more extensive distribution of cells in the periventricu-lar region, and three separate fiber pathways. In the quail,these cells and fibers were observed only in the n. preop-ticus periventricularis and in the n. preopticus medialis,and not in the PHN, PVN, and IN (Aste et al., 1991).
Previous physiological studies of avian species (Kuenzelet al., 1987; Kuenzel and McMurtry, 1988; Boswell et al.,1999) and our study of NPY distribution in the paraven-tricular region of chick hypothalamus indicate that NPY isinvolved in the control of feeding behaviour in birds. Thisagrees with results obtained in similar studies of mam-mals (Gehlert, 1999; Hokfelt et al., 1998; Palmiter et al.,1998; Yokosuka et al., 1999).
In both quail (Aste et al., 1991) and chick (the presentstudy), NPY-ir fibers and terminals are present in thepreoptic and anterior hypothalamic areas. Since theseareas contain both warm and cold sensitive neurons (Satoand Simon, 1988), NPY may play a role in thermoregula-tion. Our observations of NPY-ir neurons in the IN andME agree with those made by Walsh and Kuenzel (1997).These neurons may influence the early stages of sexualmaturation in the chick by interacting with the terminalfield of GnRH neurons in the ME. In fact, it has beensuggested that the influence of NPY on LH secretion in theewe (Barker-Gibb et al., 1995) and in hens (Contijoch etal., 1993) is probably exerted by way of the presynapticaction of NPY on GnRH terminals in the ME.
A further fact of interest is that several hypothalamicareas of chick brain containing NPY-ir cells also containedGnRH (Kuenzel and Blahser, 1991). Ultrastructural stud-ies are needed to clarify these anatomical correlations.
Our study has shown that there are NPY-ir terminals inthe vicinity of the capillaries of the hypothalamo-hypophy-sial portal system, and that there is a fiber pathway run-ning from the periventricular region to the ME. Based onthese observations, it may be hypothesized that theNPY-ir perikarya located in the periventricular region(PHN, PVN) project into the ME, and that NPY is releasedinto the portal circulation, thereby directly regulating an-terior pituitary functions. This hypothesis is supported by
Fig. 7. Light micrographic localization of NPY-ir within the chickhypothalamus. PAP method. A: Arrangement of ir neurons and fibers inthe IN, coronal section, 3250. B: Innervation of the internal (i.z.) andexternal zone (e.z.) of the ME. Note the palisade arrangement of thefibers in the e.z. of ME (arrowheads). Some bipolar cells with their majoraxis parallel to the ependymal layer in the i.z. of ME (arrows), coronalsection, 3300. For abbreviations, see page 186.
196 ESPOSITO ET AL.

Fig. 8. Light micrographic localization of NPY-ir within the chickbrain. PAP method. A: A dense ir fiber arrangement in the dorsal (d),medial (m), and ventral (v) portions of the MPO, coronal section, 3250.B: Basket-like terminals around the immunonegative cells of the PVN
(arrows), coronal section, 3250. C: Fibers in the SCO (arrows), coronalsection, 3250. D: Fibers (arrows) near the SSO (arrowhead), coronalsection, 3250. E: Fibers in the OVLT (arrows), coronal section, 3250.For abbreviations, see page 186.
197NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN

the presence of NPY-binding sites both in cultured adeno-hypophysial cells and in the pituitary of chicken (Merck-aert and Vandesande, 1990, 1996). Previous studies con-ducted by us showed that the periventricular region ofchick hypothalamus contained several non-NPY peptides(Esposito et al., 1992, 1993, 1997). The colocalization andinteraction of these peptides with NPY still need to bedetermined.
In the quail nBOR, a network of NPY-containing fibershas been reported (Aste et al., 1991). In chick, we havefound NPY-ir perikarya in dorsal and lateral portions ofthe nBOR, and NPY-ir fibers in the nBOR proper. In birds,the nBOR is a nucleus of the accessory optic system thatresponds to whole-field visual motion, which is used foroptokinetic eye movements (Wallman et al., 1981). TheNPY present in this nucleus may modulate this function.
The mesencephalic Ico, which is the counterpart of themammalian peduncolopontine nucleus and is known toplay a central role in mediating vocal behavior in birds(Phillips et al., 1972; De Lanerolle and Andrew, 1974) wasfound in the present study to contain NPY-ir fibers. Inaddition, the preoptic and hypothalamic regions of severalavian species have been observed to contain peptide-irperikarya projecting towards the ICo (Deviche and Gun-turkun, 1992). Our observations, which require eventualconfirmation by tracing studies, suggest that a pathwaycontaining NPY originates in the hypothalamus and ter-minates in the ICo. Thus, NPY may also be involved in thecontrol of chick vocalizations.
In our study, the chick rhombencephalon has shownNPY ir-cells only in the caudal pons, i.e., in the RpaM andR. In the quail brain, however, two NPY-ir cell popula-tions have been localized: one in the isthmic region, andthe other in the caudal pons where it borders the medulla.The significance of the localization of these cell popula-tions still needs to be determined. Our results, however,confirm the finding that there are fewer NPY-ir cell groupsin the rhombencephalon of non-mammalian vertebratespecies than in the rhombencephalon of mammals (Me-dina et al., 1992; Vecino et al., 1994).
In conclusion, the morphological data of the presentstudy regarding the NPY-ir in the chick brain show that:(1) the pattern of NPY-ir distribution in the whole brain is,for the most part, similar in chick and quail (Aste et al.,1991); (2) the NPY-containing cells and fibers are widelydistributed throughout the avian brain, and play diversefunctional roles; and (3) NPY in birds, as in other verte-brates (Andersen et al., 1992; Medina et al., 1992; Heiligand Widerlov, 1995; Chiba, 1997; Castro et al., 1999), mayplay a key role in the regulation of the neuroendocrine,vegetative, and sensory systems by acting as a neuro-modulator and/or neurotransmitter.
Fig. 9. Light micrographic localization of NPY-ir within the chick hypo-thalamus. PAP method. A: Innervation of PVN region, parasagittal sec-tion 3100. Note the numerous basket-like terminals surrounding thePVN cells (arrows). B: A fiber pathway (arrows) connecting the PVNregion (left) and IN region (right), parasagittal section, 3100. C: A rostro-caudally (left-right) fiber pathway (arrows) running above the CO, para-sagittal section, 3100. In all photos the left side is rostral. For abbrevi-ations, see page 186.
198 ESPOSITO ET AL.

Fig. 10. Light micrographic localization of NPY-ir within the chick brainstem. PAP method. A: Immuno-reactive neurons and fibers in the nBOR, parasagittal section, 3160. Left side is rostral. B: Magnification ofthe neurons in the nBOR, parasagittal section, 3400. C: Labeled neurons within the RPaM, coronal section,3250. Right side is lateral. For abbreviations, see page 186.

ACKNOWLEDGMENTSThe authors thank Miss S. Alı for technical assistance
and Mr. A. Calamo for the photographic art work.
LITERATURE CITEDAndersen AC, Tonon M-C, Pelletier G, Conlon JM, Fasolo A, Vaudry
H. 1992. Neuropeptides in the amphibian brain. Int Rev Cytol138:89–210.
Anderson KD, Reiner A. 1990. Distribution and relative abundance ofneurons in the pigeon forebrain containing somatostatin, neuropep-tide Y, or both. J Comp Neurol 299:261–282.
Aste N, Viglietti-Panzica C, Fasolo A, Andreone C, Vaudry H, Pelle-tier G, Panzica GC. 1991. Localization of neuropeptide Y-immuno-reactive cells and fibers in the brain of the Japanese quail. CellTissue Res 265:219–230.
Barker-Gibb ML, Scott CJ, Boublik JH, Clarke IJ. 1995. The role ofneuropeptide Y (NPY) in the control of LH secretion in the ewe withrespect to season, NPY receptor subtype and the site of action in thehypothalamus. J Endocrinol 147:565–579.
Blomqvist AG, Soderberg C, Lundall I, Milner R, Larhammar D. 1992.Strong evolutional conservation of neuropeptide Y: sequences ofchicken, goldfish, and Torpedo marmorata DNA clones. Proc NatlAcad Sci USA 89:2350–2354.
Boswell T, Dunn IC, Corr SA. 1999. Hypothalamic neuropeptide YmRNA is increased after feed restriction in growing broilers. Poul-try Sci 78:1203–1207.
Castro A, Becerra M, Manso MJ, Anadon R. 1999. Development ofimmunoreactivity to neuropeptide Y in the brain of brown trout(Salmo trutta fario). J Comp Neurol 414:13–32.
Chartrel N, Conlon JM, Danger J-M, Fournier A, Tonon M-C, VaudryH. 1991. Characterization of melanotropin-release-inhibiting factor(melanostatin) from frog brain: homology with human neuropeptideY. Proc Natl Acad Sci USA 88:3862–3866.
Chiba A. 1997. Distribution of neuropeptide Y-like immunoreactivityin the brain of bichir, Polypterus senegalus, with special regard tothe terminal nerve. Cell Tissue Res 289:275–284.
Colmers WF, Whalestedt C. 1993. The biology of neuropeptide Y andrelated peptide. Totawa, NY: Humana Press.
Conlon JM, Bjornholm B, Jorgensen FS, Youson JH, Schwartz TW.1991. Primary structure and conformational analysis of peptidemethionine-tyrosine, a peptide related to neuropeptide Y and pep-tide YY isolated from the lamprey intestine. Eur J Biochem 199:293–298.
Conlon JM, Balasubramaniam A, Sower SA. 1994. Purification andsteroidogenic activity of a neuropeptide Y-related peptide from thebrain of the sea lamprey. Reg Peptides 50:167–175.
Contijoch AM, Malamed S, McDonald JF, Advis J-P. 1993. Neuropep-tide Y regulation of LHRH release in the median eminence: immu-nocytochemical and physiological evidence in hens. Neuroendocri-nology 57:135–145.
De Lanerolle N, Andrew RJ. 1974. Midbrain structures controllingvocalization in the domestic chick. Brain Behav Evol 10:354–376.
Deviche P, Gunturkun O. 1992. Peptides for calling? An immunohis-tochemical study of the avian n. intercollicularis. Brain Res 569:93–99.
Esposito V, de Girolamo P, Gargiulo G. 1992. Vasoactive intestinalpolypeptide (VIP)-, somatostatin (SOM)-, methionine-enkephalin(m-ENK)- and b-endorphin (b-END)-like immunoreactivity in thehypothalamus of Gallus domesticus. Neuropeptides 22:22 (abstr).
Esposito V, de Girolamo P, Gargiulo G. 1993. Immunoreactivity tovasoactive intestinal polypeptide (VIP) in the hypothalamus of thedomestic fowl Gallus domesticus. Neuropeptides 25:83–90.
Esposito V, de Girolamo P, Gargiulo G. 1997. Neurotensin-like immu-noreactivity in the brain of the chicken, Gallus domesticus. J Anat191:537–546.
Fraley GS, Kuenzel WJ. 1993. Precocious puberty in chicks (Gallusdomesticus) induced by central injection of neuropeptide Y. Life Sci52:1649–1656.
Gehlert DR. 1999. Role of hypothalamic neuropeptide Y in feedingand obesity. Neuropeptides 33:329–338.
Gilbert DB, Patterson TA, Rose SPR. 1991. Dissociation of brain sitesnecessary for registration and storage of memory for a one-trialpassive avoidance task in the chick. Behav Neurosci 105:553–561.
Gunturkun O, Karten HJ. 1991. An immunocytochemical analysis ofthe lateral geniculate complex in the pigeon (Columba livia).J Comp Neurol 314:721–749.
Hamassaki DE, Britto LRG. 1990a. Thalamic origin of neuropeptide Yinnervation of the accessory optic nucleus of the pigeon (Columbalivia). Visual Neurosci 5:249–259.
Hamassaki DE, Britto LRG. 1990b. Efferent projection of the pigeonintergeniculate leaflet containing neuropeptide Y. Braz J Med BiolRes 23:1051–1055.
Heilig M, Widerlov E. 1995. Neurobiology and clinical aspects ofneuropeptide Y. Crit Rev Neurobiol 9:115–136.
Hokfelt T, Broberger C, Zhang X, Diez M, Kopp J, Xu Z.-Q, Landry M,Bao L, Schelling M, Koistinaho J, De Armond SJ, Prusiner S, GongJ, Walsh JH. 1998. Neuropeptide Y: some viewpoints on a multi-faceted peptide in the normal and diseased nervous system. BrainRes Rev 26:154–166.
Hoyle CH. 1999. Neuropeptide families and their receptors: evolution-ary perspectives. Brain Res 848:1–25.
Kuenzel WJ, Blahser S. 1991. The distribution of gonadotrophin-releasing hormone (GnRH) neurons and fibers throughout the chickbrain (Gallus domesticus). Cell Tissue Res 264:481–495.
Kuenzel WJ, Masson M. 1988. A stereotaxic atlas of the brain of thechick (Gallus domesticus). Baltimore: Johns Hopkins UniversityPress.
Kuenzel WJ, McMurtry J. 1988. Neuropeptide Y: brain localizationand central effects on plasma insulin levels in chicks. Physiol Behav44:669–678.
Kuenzel WJ, Douglass LW, Davison BA. 1987. Robust feeding follow-ing central administration of neuropeptide Y or peptide YY inchicks, Gallus domesticus. Peptides 8:823–828.
Larhammar D. 1996. Evolution of neuropeptide Y, peptide YY, andpancreatic polypeptide. Regul Peptides 62:1–11.
Larhammar D, Soderberg C, Blomqvist AG. 1992. The neurobiology ofneuropeptide Y and related peptides. In: Wahlestedt C, ColmersWF, editors. Contemporary neuroscience series. Clifton NJ: Hu-mana Press, p 65–73.
Larhammar D, Soderberg C, Blomqvist AG. 1993. Evolution of theneuropeptide Y family of peptides. In: Colmers WF, Wahlestedt C,editors. The biology of neuropeptide Y and related peptides. NewJersey: Humana Press, p 1–41.
Larhammar D, Soderberg C, Blomqvist AG, Lundell I, Brodin L. 1995.Evolution of the evolution neuropeptide Y-peptide YY family. Neth-erlands J Zool 45:15–17.
Larhammar D, Soderberg C, Lundell I. 1998. Evolution of the neu-ropeptide Y family and its receptors. Ann NY Acad Sci 839:35–40.
Lee MC, Schiffman SS, Pappas TN. 1994. Role of neuropeptides in theregulation of feeding behaviour: a review of cholecystokinin, bomb-esin, neuropeptide Y and galanin. Neurosci Behav Rev 18:313–323.
Medina L, Marti E, Artero C, Fasolo A, Puelles L. 1992. Distributionof neuropeptide Y-like immunoreactivity in the brain of the lizardGallotia galloti. J Comp Neurol 319:387–405.
Medina M, Reperant J, Miceli D, Bertrand C, Bennis M. 1998. Animmunohistochemical study of putative neuromodulators andtransmitters in the centrifugal visual system of the quail (Coturnixjaponica). J Chem Neuroanat 72:75–79.
Merckaert J, Vandesande F. 1990. Study of a direct effect of neuropep-tide Y (NPY) on the pituitary of the chicken (Gallus domesticus).15th Conference of European Comparative Endocrinologists, Leu-ven, Belgium. p 148 (abstr).
Merckaert J, Vandesande F. 1996. Autoradiographic localization ofreceptors for neuropeptide Y (NPY) in the brain of broiler andleghorn chickens (Gallus domesticus). J Chem Neuroanat 12:123–134.
Palmiter RD, Erickson JC, Hollopeter G, Baraban SC, Schwartz MW.1998. Life without neuropeptide Y. Rec Progr Horm Res 53:163–1999.
Phillips RE, Youngren OM, Peek FW. 1972. Repetitive vocalizationsevoked by local electrical stimulation of avian brains. I. Awakechickens (Gallus gallus). Anim Behav 20:689–705.
200 ESPOSITO ET AL.

Richardson RD, Boswell T, Raffety BD, Seeley RJ, Wingfield JC,Woods SC. 1995. NPY increases food intake in white-crownedsparrows: effect in short and long photoperiods. Am J Physiol 268:R1418–R1422.
Rose SPR. 1991. What the chick can tell us about the process andstructure of memory. In: Squire LR, Weinberger N, Lynch G, Mc-Gaugh JL, editors. Memory: organization and locus of change. NewYork: Oxford University Press. p 392–412.
Sato H, Simon E. 1988. Thermal characterization and transmitteranalysis of single units in the preoptic and anterior hypothalamusof conscious ducks. Pflug Arch 411:34–41.
Sugita S. 1994. Vasotocin fibers and neurons in the brain of the domesticfowl (Gallus gallus domesticus). Acta Anat Nippon 69:22–33.
Tatemoto K. 1982. Neuropeptide Y: complete amino acid sequence ofthe brain peptide. Proc Natl Acad Sci USA 79:5485–5489.
Tatemoto K, Carlquist M, Mutt V. 1982. Neuropeptide Y: a novelbrain peptide with structural similarities to peptide YY and pan-creatic polypeptide. Nature 296:659–660.
Tombol T, Davies DC, Nemeth A, Sebesteny T, Alpar A. 2000. Acomparative Golgi study of chicken (Gallus domesticus) and homingpigeon (Columba livia) hippocampus. Anat Embryol 201:85–101.
Vecino E, Perez M-TR, Ekstrom P. 1994. In situ hybridization ofneuropeptide Y (NPY) mRNA in the goldfish brain. NeuroReport6:127–131.
Wallman J, McKenna OC, Burns S, Velez J, Weinstein B. 1981.Relation of accessory optic system and pretectum to optokineticresponses in chickens. In: Fuchs A, Becker W, editors. Progress inoculomotor research. New York: Elsevier-North Holland Pub. Co. p435–442.
Walsh KM, Kuenzel W. 1997. Effect of sulfamethazine on sexualprecocity and neuropeptide Y neurons within the tuberoinfundibu-lar region of the chick brain. Brain Res Bull 44:707–713.
Yokosuka M, Kalra PS, Kalra SP. 1999. Inhibition of neuropeptide Y(NPY)-induced feeding and c-Fos response in magnocellular para-ventricular nucleus by a NPY receptor antagonist: a site of NPYaction. Endocrinology 140:4494–4500.
201NPY-IMMUNOREACTIVITY IN THE CHICK BRAIN


















