
Analiza varijabilnosti: funkcionalna varijabilnost...
30
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović ----------------------------------------------------------------------------------------------------------- 1 Analiza varijabilnosti: funkcionalna varijabilnost, varijabilnost DNK Genetička varijabilnost (različitost) potiče iz dva osnovna procesa: mutacija i rekombinacija. Mutacije su izvor genetičkih promena, stvaranja sasvim novih oblika genskih alela. Rekombinacije su procesi tokom kojih dolazi do stvaranja novih kombinacija genskih alela. Drugim rečima, aleli se rekombinacijama preuređuju, čime se stvara ogroman broj mogućih genotipova. Preživljavanje bioloških vrsta obezbeĎeno je mehanizmima održavanja genetičkog integriteta i verodostojnog prenošenja genetičke informacije iz generacije u generaciju. Ipak, proces biološke evolucije nezamisliv je bez povremenog unošenja promena u sekvenci nukleotida ili drugih oblika genetskih rearanžmana. Osnovni izvor genetičkih novina u populacijama su mutacije i one obezbeĎuju nastanak „novog“ („sirovog“) materijala, pri čemu se kroz proces prirodne selekcije vrši favorizovanje onih mutacija koje su u skladu sa ekološkim uslovima u odreĎenom vremenskom periodu. Ukoliko ne bi postojale mutacije, svi geni bi egzistirali samo u jednom obliku, ne bi imali alternativne oblike – genske alele. Rekombinacije gena samo preuređuju genetičku varijabilnost do novih kombinacija, dok selekcija vodi ka favorizovanju najbolje prilagoĎenih kombinacija u odnosu na postojeće uslove u sredini. Obzirom da od genetiĉke varijabilnosti zavisi i fenotipska varijabilnost, jasno je da procesi mutacija i rekombinacija doprinose stvaranju velikog broja razliĉitih fenotipova. Kada su u populaciji prisutne fenotipski različite jedinke, prirodna selekcija ima šta da odabere, pri čemu će favorizovati jedinke sa većom šansom za preživljavanje i boljim reproduktivnim sposobnostima, odnosno boljim sposobnostima za prilagoĎavanje ekološkim uslovima (adaptibilnije jedinke). Ukoliko je za opstanak pod promenjenim uslovima sredine „zaslužna” neka mutacija, onda se putem prirodne selekcije favorizuju mutacije koje datim uslovima obezbeĎuju opstanak (npr. mutacije koje obezbeĎuju opstanak u zagaĎenoj životnoj sredini – čak i nakon ekoloških katastrofa, eksplozija nuklearnih elektrana). Procesi koji stoje u osnovi genetičke varijabilnosti omogućavaju da se kroz generacije menja genetička struktura populacije u skladu sa promenama uslova u životnoj sredini.
Transcript of Analiza varijabilnosti: funkcionalna varijabilnost...

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
1
Analiza varijabilnosti: funkcionalna varijabilnost,
varijabilnost DNK
Genetička varijabilnost (različitost) potiče iz dva osnovna procesa: mutacija i rekombinacija.
Mutacije su izvor genetičkih promena, stvaranja sasvim novih oblika genskih alela.
Rekombinacije su procesi tokom kojih dolazi do stvaranja novih kombinacija genskih
alela. Drugim rečima, aleli se rekombinacijama preuređuju, čime se stvara ogroman
broj mogućih genotipova.
Preživljavanje bioloških vrsta obezbeĎeno je mehanizmima održavanja genetičkog integriteta i
verodostojnog prenošenja genetičke informacije iz generacije u generaciju.
Ipak, proces biološke evolucije nezamisliv je bez povremenog unošenja promena u sekvenci
nukleotida ili drugih oblika genetskih rearanžmana.
Osnovni izvor genetičkih novina u populacijama su mutacije i one obezbeĎuju nastanak
„novog“ („sirovog“) materijala, pri čemu se kroz proces prirodne selekcije vrši favorizovanje
onih mutacija koje su u skladu sa ekološkim uslovima u odreĎenom vremenskom periodu.
Ukoliko ne bi postojale mutacije, svi geni bi egzistirali samo u jednom obliku, ne bi imali
alternativne oblike – genske alele.
Rekombinacije gena samo preuređuju genetičku varijabilnost do novih kombinacija, dok
selekcija vodi ka favorizovanju najbolje prilagoĎenih kombinacija u odnosu na postojeće
uslove u sredini.
Obzirom da od genetiĉke varijabilnosti zavisi i fenotipska varijabilnost, jasno je da procesi
mutacija i rekombinacija doprinose stvaranju velikog broja razliĉitih fenotipova. Kada su
u populaciji prisutne fenotipski različite jedinke, prirodna selekcija ima šta da odabere, pri
čemu će favorizovati jedinke sa većom šansom za preživljavanje i boljim reproduktivnim
sposobnostima, odnosno boljim sposobnostima za prilagoĎavanje ekološkim uslovima
(adaptibilnije jedinke). Ukoliko je za opstanak pod promenjenim uslovima sredine „zaslužna”
neka mutacija, onda se putem prirodne selekcije favorizuju mutacije koje datim uslovima
obezbeĎuju opstanak (npr. mutacije koje obezbeĎuju opstanak u zagaĎenoj životnoj sredini –
čak i nakon ekoloških katastrofa, eksplozija nuklearnih elektrana).
Procesi koji stoje u osnovi genetičke varijabilnosti omogućavaju da se kroz generacije menja
genetička struktura populacije u skladu sa promenama uslova u životnoj sredini.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
2
Sa druge strane, mutacije su često uzrok anomalija, ali posledice mutacija nisu uvek očigledne,
odnosno, fenotipski efekti mutacija se u mnogim slučajevima mogu detektovati samo
delikatnim biohemijskim ili genetičkim analizama, a samo u nekim slučajevima mutacije
dovode do velikih morfoloških promena i letalnih efekata.
Nažalost, u stočarstvu, selekcija je jako dugo obavljana na osnovu fenotipa i primenom metoda
selekcijskog ukrštanja. Ove procedure selekcije primenjivane su u velikom obimu bez
detaljnog znanja o broju gena koji su uključeni u ekspresiju datih osobina, kao i bez ikakvog
znanja o efektu tih gena na objektima selekcije i njihovom učešću u plejotropnom efektu ili
vezanom delovanju na druge osobine. Primena ovakvog pristupa pokazala se korisnom u
velikoj meri u poboljšanju prosečne količine mleka po laktaciji kod mnogih populacija mlečnih
krava. MeĎutim, pri korišćenju ovakvih odgajivačkih šema dešava se da najbolje rangirani
bikovi (odabrani isključivo na osnovu fenotipa), koji se koriste u programima veštačkog
osemenjavanja, budu nosioci mutiranih alela. Takvi bikovi značajno doprinose širenju
naslednih anomalija kod potomaka.
Korišćenjem molekularno-genetičkih analiza moguće je identifikovati mutacije koje su uzrok
anomalijama (koje se nekad manifestuju odmah po roĎenju, ali često tek kada stupe u proces
reprodukcije) i sprečiti dalekosežno širenje mutiranih alela.
Nakon identifikacije mutacije, dizajniraju se molekularni markeri za njihovu detekciju, čime je
omogućeno da se nosioci tih alela isključe iz daljeg programa selekcije.
Analiza genetiĉke varijabilnosti
Prouĉavanje fenotipskih razlika meĎu jedinkama jedne populacije predstavlja jedan od
najjednostavnijih naĉina analize genetiĉke varijabilnosti (raznovrsnosti). Pod fenotipskim
razlikama podrazumevamo npr. razlike u graĎi tela, visini i težini, pigmentaciji kože (dlake,
perja), krvnim grupama, ponašanju, dužini života, zdravstvenom stanju ali i razlike u tzv.
proizvodnim osobinama gajenih životinja - dnevni prirast, plodnost, kvalitet trupa i ukus
mesa, kvantitet trupa i prinos mesa, mlečnost, odnosno kvantitet i kvalitet mleka kod krava,
produkcija jaja kod živine, mesnatost kod svinja itd.
Fenotipska varijabilnost je veoma česta pojava u prirodi, ali najvaţnije pitanje je koliko je za
tu varijabilnost odgovorna nasledna osnova, a koliko faktori sredine (videti: Box 1).
Prethodno navedene proizvodne osobine, odnosno ekonomski znaĉajne osobine
(economically relevant traits-ERT) su kompleksne, odnosno pod kontrolom velikog broja
gena i pod uticajem faktora spoljašnje sredine. Kod goveda su to, na primer, osobine vezane
za rast, unos hrane, mlečnost, karakteristike trupa (mesnatost, količina masti) i reprodukciju.
Markeri koji su otkriveni za te osobine, vezani su za samo jedan gen od brojnih gena koji
kontrolišu kompleksnu osobinu. MeĊutim, ekspresija ţenjenog fenotipa (npr. povećana
mleĉnost, bolji kvalitet mleka i mesa) zavisi i od brojnih drugih “nemarkiranih” gena, ali
i brojnih faktora sredine. Zbog toga je pri donošenju odluke o selekciji uvek neophodno
uzeti u obzir oĉekivane razlike u potomstvu (Expected Progeny Differences-EPDs), čak i

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
3
kada postoje podaci dobijeni primenom markera. Selekcija goveda u kojoj se koriste
molekularni markeri je prihvaćena i preporuĉena kao dopuna tradicionalnim metodama
selekcije. Najveća korist molekularnih markera u selekciji je pre svega kod osobina koje
imaju jednostavan naĉin nasleĊivanja (boja krzna, genetiĉke anomalije), a zatim kod
kompleksnih osobina kao što su: kvalitet trupa i ukus mesa, kvantitet trupa i prinos
mesa, mleĉnost, odnosno kvantitet i kvalitet mleka, fertilnost i reproduktivna efikasnost,
sposobnost za materinstvo, unos hrane, ponašanje vezano za ishranu i prirast. Ipak, u
svetu postoje standardizovani testovi za k-kazein, beta-laktoglobulin i alfa S1 kazein na
osnovu kojih se može odrediti kvalitet mleka krave čiji je uzorak analiziran, ali i genotip za te
osobine, što je značajno za procenu količine i kvaliteta mleka koji se može očekivati kod
potomaka. Na primer, krave BB genotipa za k-kazein, proizvode veću koliĉinu mleka sa
većim sadrţajem proteina nego krave AB ili AA genotipa. Osim toga, mleko krava BB
genotipa je bolje za proizvodnju sira. Krave B genotipa za beta-laktoglobulin, glavnu
komponentu surutke, proizvode mleko sa većim sadrţajem masti i kazeina, komponenti
koje su poţeljne za proizvodnju sira. Sa druge strane krave AA genotipa daju veće
koliĉine mleka. Alfa S1 kazein takoĊe utiĉe na prinos mleka. Krave BB genotipa za alfa
S1 kazein daju više mleka nego krave reĊe prisutnog BC genotipa. Genetičke analize
navedenih parametara obavnjaju se iz korena dlake ili uzoraka krvi. Preciznost rezultata je
99%. Najznačajniji faktor koji je povezan sa praktično svim produktivnim, reproduktivnim, pa
i imunim karakteristikama goveda jeste hormon leptin, proizvod leptin gena („gena za
gojaznost”). Leptin sintetišu i luče uglavnom adipocite belog masnog tkiva. Leptin ima ulogu u
regulisanju apetita, unosa hrane, telesne težine, telesnog sastava i potrošnje energije. TakoĎe
utiče reprodukciju i neke funkcije imunog sistema. Koncentracija leptina u serumu povezana sa
depoima masti u trupu i ekonomski značajnim karakteristikama goveĎeg mesa. U cilju
ispitivanjem molekularnih mehanizama koji stoje u osnovi uticaja leptina, najpre je mapiran
gen za leptin na hromozomu 2, a zatim je utvrĎeno da u kodirajućem regionu leptin gena
postoji polimorfizam koji je značajan za koncentraciju leptina u serumu. Polimorfizam u
kodirajućem regionu leptin gena ima veliki uticaj na ekonomski značajne karakteristike
goveda: na unos hrane, na vrednosti trupa, odnosno utovljenost i kvalitet mesa, kvantitet i
kvalitet mleka. Osim toga, postoje dokazi da polimorfizam u regulatornom regionu
(promotoru) gena za leptin utiče na koncentraciju leptina u serumu, prirast, telesnu težinu,
pokazatelje vrednosti trupa, unos hrane i ponašanje vezano za ishranu. Predloženi SNP markeri
za navedene osobine su UASMS2 i UASMS3. Polimorfizam u promotoru leptin gena može
uticati i na prinos i sastav mleka. SNP markeri UASMS1 i UASMS3 u promotoru leptin gena
imaju signifikantan uticaj na prinos masti, ali za UASMS2 nije potvrĎena signifikantna
povezanost ni sa jednom osobinom vezanom za kvalitet mesa goveda. MeĎutim, definisani su
molekularni SNP markeri unutar leptin gena za sledeće ekonomski značajne osobine goveda.
To su dva SNP markera (E2JW, E2FB) značajna su za prinos i rasporeĎenost masti i za prinos
posnog mesa i posredno za mekoću mesa. Sa druge strane, najnovija istraživanja otkrila su da
SNP markeri UASMS2 i R25C imaju značajan uticaj na parametre porasta, telesne težine i
leĎne slanine. Navedeni markeri omogućavaju identifikaciju goveda koja imaju željenu
genetsku predisponiranost u odnosu na navedene osobine
Savremene analize su pokazale da je mnogo veći stepen genetiĉke varijabilnosti nego što
bi se oĉekivalo na osnovu analize fenotipske varijabilnosti.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
4
Metode analize genetiĉke varijabilnosti
kvalitativnih osobina:
MORFOMETRIJSKE METODE - analize morfoloških karakteristika, koje je
moguće koristiti ukoliko se zna priroda genetičke determinacije posmatrane osobine;
CITOGENETIĈKE METODE – analize kariotipa (hromozoma), kojima se
mogu otkriti promene u broju ili strukturi hromozoma kod jedinki ispitivane populacije;
Morfološki i hromozomski markeri obiĉno pokazuju nizak nivo polimorfizma i stoga
nisu posebno korisni kao genetiĉki markeri.
BIOHEMIJSKE METODE - analize proteina, jer zbog kolinearnosti izmeĎu gena i
proteina, promene nukleotidne sekvence DNK (tj. gena) mogu da dovedu do izmene
primarne strukture kodiranog proteina.
Iako su proteinski polimorfizmi bili prvi markeri za genetiĉka istraţivanja (koji su se
veoma dugo i obimno koristili u stočarstvu), analize proteina se danas smatraju
prevaziĊenom metodom zbog veoma niske rezolucije, odnosno zbog velikih ograniĉenja
u ispitivanju genetiĉke varijabilnosti.
Naime, analizom proteina mogu se detektovati samo neki genetiĉki polimorfizmi i
nivo polimorfizma koji se detektuje je najĉešće nizak.
OdreĎivanje amino-kiselinske sekvence proteina je veoma zahtevno (vremenski i
finansijski), tako da se najĉešće analize proteina obavljaju gel-elektroforezom, koja se
zasniva na tome da proteini imaju razliĉitu elektropokretljivost (na gelu u rastvoru
slabog elektrolita) ukoliko se razlikuju po sekvenci aminokiselina (drugim rečima,
elektroforetska separacija proteina zasniva se na razlikama u električnom naboju ili
razlikama u molekulskim masama kod različitih proteina).
MeĎutim, ĉak i analiza amino-kiselinske sekvence ne moţe otkriti svaku promenu u
genomu, jer postoje mutacije koje NE DOVODE do promene amino-kiselinske
sekvence usled izroĊenosti genetskog koda (tzv. tihe mutacije).
Sa druge strane, elektroforetska analiza proteina ima još manju rezoluciju, jer ne moţe
otkriti ĉak ni svaku promenu u sekvenci amino-kiselina, jer se neke zamene amino-
kiselina ne odraţavaju na elektroforetsku pokretljivost proteina (ukoliko se jedna
amino-kiselina u proteinu zameni nekom hemijski bliskom amino-kiselinom).
Konaĉno, navedeni markeri odraţavaju varijabilnost u sekvencama koje ih kodiraju, a
koje predstavljaju manje od 10% ukupnog genoma.
MOLEKULARNE METODE – predstavljaju metode izbora za analizu genetiĉke
varijabilnosti, obzirom da se njima otkrivaju razlike u samom molekulu DNK (tzv.
DNK polimorfizmi koji podrazumevaju svaku razliku u nukleotidnoj sekvenci
(unutar gena i/ili nekodirajućih regiona DNK). Markeri kojima se detektuju
razlike na nivou DNK nazivaju se molekularni ili DNK markeri.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
5
Molekularni markeri, sposobni da detektuju genetiĉke varijacije na nivou sekvenci DNK,
ne samo da su prevazišli ograniĉenja prethodno korišćenih metoda (morfometrijske,
citogenetiĉke, biohemijske), nego poseduju i jedinstvena genetiĉka svojstva koja ih ĉine
mnogo korisnijim od ostalih genetiĉkih markera. Oni su brojni i rasporeĎeni svuda po
čitavom genomu. NasleĎuju se po tipičnom Mendelovom načinu nasleĎivanja koji se obično
ispoljava u ko-dominantnom obliku i često su multialelni tako da se u proseku heterozigotnost
ostvaruje u više od 70%. Na njih ne utiču faktori spoljašnje sredine i generalno nemaju
plejotropni efekat na lokuse za kvantitativne osobine (quantitative trait loci-QTL). Obzirom da
genska ekspresija nije preduslov, korišćenjem molekularnih markera moţe se vizuelizovati
praktiĉno celokupan genom ukljuĉujući nekodirajuće regione.
Osim znatno veće preciznosti, molekularni markeri nude i brojne metodološke prednosti:
- mogućnost korišćenja bilo kog uzorka koji sadrži DNK životinje (ne samo krv, nego i
dlake, briseve bukalne ili vaginalne sluznice, mleko, sperma, arhivirani preparati);
- mogućnost retrospektivne analize u slučajevima kada životinje više nisu dostupne (iz
sačuvanih uzoraka tkiva ili sperme),
- uzorkovanje za DNK analize je neinvanzivno
- uzorci DNK se mogu jako dugo čuvati u laboratoriji bez nekih posebnih uslova i lako
se mogu transportovati izmeĎu laboratorija,
- analiza DNK može se obaviti u veoma ranima fazama života jedinke, odnosno čak i na
stupnju embriona (prenatalna dijagnostika), nezavisno od pola,
- kada se jednom prenese na čvrstu podlogu, kao što su filter membrane, može se više
puta koristiti za hibridizaciju sa različitim probama, a mogu se koristiti heterologe
probe i in vitro-sintetizovane oligonukleotidne probe,
- metode bazirane na PCR metodologiji mogu se automatizovati.
Postoje brojne mogućnosti primene molekularnih markera u programima poboljšanja
stoke kako u konvencionalnim, tako i u transgenim strategijama gajenja. U programima
poboljšanja putem konvencionalnih strategija gajenja, primena molekularnih markera može biti
kratkoročna ili neposredna (za odreĊivanje roditeljstva, procenu genetiĉke distance,
determinaciju tipa blizanaĉke zigotije i frimartinizma, odreĊivanja pola embriona pre
implantacije, kao i za identifikaciju nosioca bolesti), ali i dugoročna (u postupcima
mapiranja gena, kao i za selekciju u kojoj se koriste markeri, za koju se koristi termin:
marker-assisted selection-MAS). Kod transgenog gajenja, molekularni markeri se mogu
koristiti kao referentne tačke za identifikaciju, izolaciju i kloniranje relevantnih gena,
manipulaciju tim genima i identifikaciju životinja koje nose transgene. Ovde je značajno
napomenuti značaj molekularnih markera u identifikaciji genetički modifikovanih životinja
(GMO) i proizvoda od GMO životinja. Osim toga, progres u razvoju molekularnih markera
ukazuje na mogućnost njihove primene za genetičko poboljšanje gajenih vrsta životinja.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
6
Najznaĉajnije molekularne metode koje se koriste za analizu genetiĉke i
funkcionalne varijabilnosti
Postupak analize genetičke i funkcionalne varijabilnosti putem molekularnih markera
počinje izolacijom (ekstrakcijom) i multiplikacijom (kloniranjem) DNK ili RNK iz
bioloških uzoraka.
Izolacija DNK iz ćelije podrazumeva oslobaĎanje DNK iz jedra (nuklearna DNK) ili
organela (mitohondrijalna). Pri tome se koriste hemikalije kojima se razbijaju i liziraju: ćelijska
membrana (animalnih ćelija), ćelijski zid (kod biljaka i gljiva) i membrane organela
(mitohondrija i plastida). http://learn.genetics.utah.edu/content/labs/extraction/
Nakon izolacije DNK (ili RNK) iz ćelije, sledeći korak je amplifikacija (kloniranje) željenog
(ciljanog) gena ili fragmenta DNK, što se može obaviti in vivo i in vitro.
U slučaju in vivo kloniranja željenog (ciljnog) fragmenta DNK, najpre se restrikcionim
enzimima (videti Box 2) „iseĉe” ciljani deo DNK, a zatim taj deo ugradi u DNK vektora
(plazmida ili virusa), odnosno stvori se rekombinantna DNK. Da bi ona mogla da se
umnoţava, neophodno je da se vektor ubaci u ćeliju domaćina (najčešće u bakteriju
Escherichia coli). DNK vektora je u mnogim slučajevima cirkularna i dvolančana. Pri svakoj
replikaciji DNK vektora, istovremeno se umnoţava i ugraĊeno parĉe DNK, odnosno
replikuje se ĉitava rekombinantna DNK. Na ovaj način se još od 1982. godine proizvodi
humani insulin u ćelijama bakterija – ubacivanjem dela humane DNK koji sadrži gen za insulin
u vektor, a zatim u bakterije čijim umnožavanjem se obezbeĎuje proizvodnja sintetskog
humanog insulina. To je, inače, prvi lek dobijem primenom metoda genetičkog inžinjeringa.
In vitro kloniranje željenog (ciljnog) fragmenta DNK obavlja se putem reakcije lančane
polimerizacije, odnosno putem PCR amplifikacije – (Polymerase Chain Reaction – PCR)
u aparatu koji ima mogućnost brze promene temperaturnih uslova. Tvorac ove tehnike Kary
Mullis je za taj svoj izum dobio Nobelovu nagradu 1993. godine.
Ključna komponenta u PCR procesu jeste DNA polimeraza izolovana iz bakterije Thermus
aquaticus, koja je rezistentna na visoke temperature (živi i replicira se pri teperaturama do
95°C). Ta termostabilna polimeraza nazvana je Taq polimeraza (po bakteriji domaćinu) i
koristi se za replikaciju in vitro čime se omogućava geometrijski porast broja kopija ciljane
DNK.
Za PCR amplifikaciju potrebne su sledeće komponente:
izolovana DNK čiji odreĎeni fragment želimo da amplifikujemo,
prajmeri (oligonukleotidne sekvence) koji se dizajniraju tako da budu komplementarni
sekvencama koji okružuju ciljani region DNK (npr. gen) koji želimo da umnožimo,
Taq polimeraza,
slobodni nukleotidi, odnosno gradivne jedinice za sintezu novih lanaca DNK, u vidu
mešavine deoksiribonukleozid trifosfata (dNTP): adeninskih (dATP), timinskih (dTTP),
guaninskih (dGTP) i citozinskih (dCTP),
Magnezijumovi joni (Mg2 +
),
PCR pufer,
sterilna dejonizovana voda.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
7
Proces PCR amplifikacije obuhvata sledeće korake:
1. Inicijalna denaturacija DNK u trajanju od dva do četiri minuta (zavisno od udela GC
parova).
2. Denaturacija DNK – rasplitanje i razdvajanje lanaca DNK. Obavlja se na 94-96°C u
trajanju od 30 sec do nekoliko minuta (zavisno od udela GC parova),
3. Hibridizacija para prajmera sa komplementarnim sekvencama koji okružuju ciljani
region DNK. Obavlja se na temperaturi od 45-65°C u trajanju od 30 sekundi do
nekoliko minuta,
4. Elongacija prajmera (ekstenzija), odnosno sinteza novih DNK lanaca počev od
prajmera, tako što se Taq polimeraza vezuje za mesta hibridizacije prajmera i katalizuje
ugraĎivanje novih nukleotida komplementarih inicijalnim sekvencama. Ovaj proces se
obavlja na 72°C i traje od 45 sec do 1 minuta.
Koraci od 2. do 4. ponavljaju se tokom 25 do 40 ciklusa, kako bi se obezbedilo
umnožavanje dovoljnog broja kopija ciljnog fragmenta DNK. Umnoţeni ciljni fragmenti
DNK nazivaju se amplikoni ili PCR produkti. U svakom ciklusu količina amplikona se
duplicira (u prvom ciklusu nastaju 2 aplikona, u drugom 4, u trećem 8, u četvrtom 16, u petom
32, u šestom 64 ...) tako da na kraju PCR procesa nastaje od milion do bilion amplikona.
5. Elongacija preostalih produkata (na 72 °C, u trajanju od dva do čeiti minuta).
http://learn.genetics.utah.edu/content/labs/pcr/
Po završetku PCR amplifikacije, neophodno je izvršiti vizuelizaciju PCR produkata.
Dobijeni amplikoni razdvajaju se elektroforezom na gelu zbog toga što fragmenti DNK
razliĉite duţine imaju razliĉitu elektropokretljivost, odnosno putuju razliĉitom brzinom
(na gelu u rastvoru slabog elektrolita) pod uticajem elektriĉnog polja. Zbog toga za isto
vreme kraći fragmenti (koji se kreću brže) preĎu duži put u odnosu na duže fragmente. Nakon
završene elektroforeze gel se boji etidijum bromidom (koji se interkalira izmeĎu lanaca DNK)
koji fluorescira pod UV svetlošću, te se gel postavlja na UV transiluminator (vizuelizacija PCR
produkata), što omogućava očitavanje rezultata. http://learn.genetics.utah.edu/content/labs/gel/
Postoje razliĉite modifikacije PCR metode:
Duplex PCR – je varijanta PCR metode u kojoj se, umesto jednog, koriste dva para
prajmera, pri ĉemu oni prepoznaju razliĉite fragmente DNK i omogućavaju
diferencijalnu dijagnostiku) (Primer: razlikovanje vrsta Nosema apis i N. ceranae putem
duplex PCR uz primenu species-specifiĉnih prajmera).
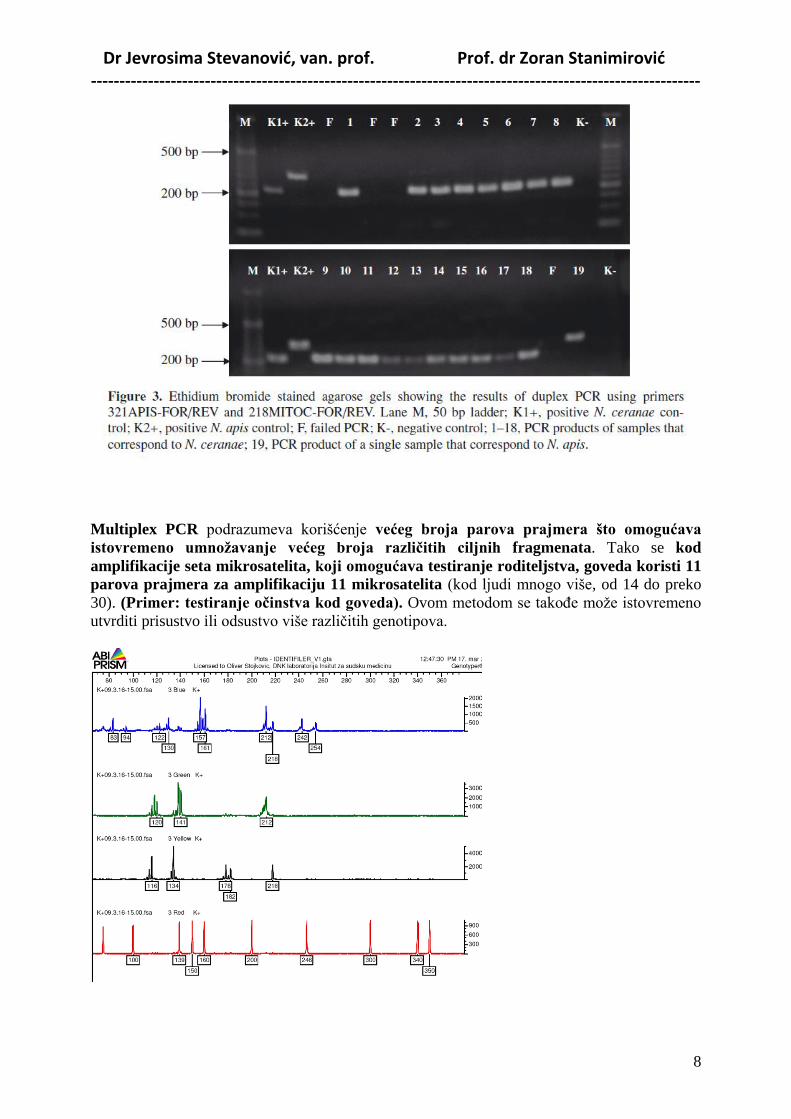
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
8
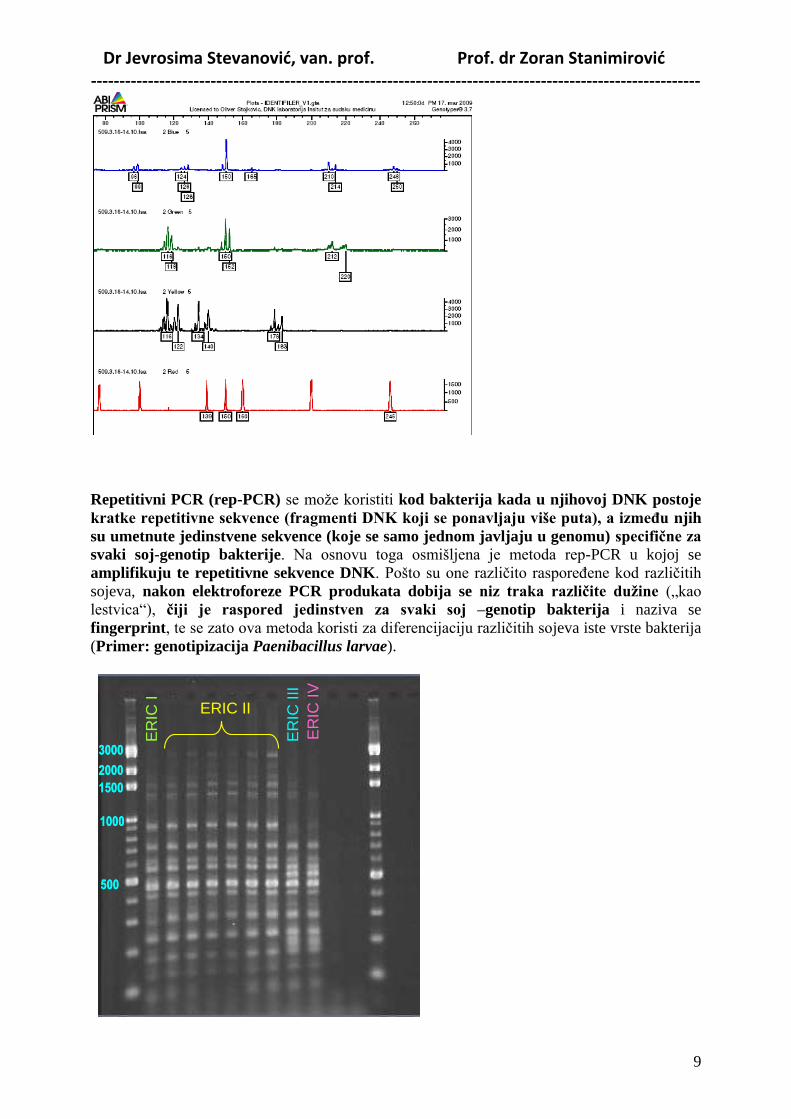
Multiplex PCR podrazumeva korišćenje većeg broja parova prajmera što omogućava
istovremeno umnoţavanje većeg broja razliĉitih ciljnih fragmenata. Tako se kod
amplifikacije seta mikrosatelita, koji omogućava testiranje roditeljstva, goveda koristi 11
parova prajmera za amplifikaciju 11 mikrosatelita (kod ljudi mnogo više, od 14 do preko
30). (Primer: testiranje oĉinstva kod goveda). Ovom metodom se takoĎe može istovremeno
utvrditi prisustvo ili odsustvo više različitih genotipova.
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
9
Repetitivni PCR (rep-PCR) se može koristiti kod bakterija kada u njihovoj DNK postoje
kratke repetitivne sekvence (fragmenti DNK koji se ponavljaju više puta), a izmeĊu njih
su umetnute jedinstvene sekvence (koje se samo jednom javljaju u genomu) specifiĉne za
svaki soj-genotip bakterije. Na osnovu toga osmišljena je metoda rep-PCR u kojoj se
amplifikuju te repetitivne sekvence DNK. Pošto su one različito rasporeĎene kod različitih
sojeva, nakon elektroforeze PCR produkata dobija se niz traka razliĉite duţine („kao
lestvica“), ĉiji je raspored jedinstven za svaki soj –genotip bakterija i naziva se
fingerprint, te se zato ova metoda koristi za diferencijaciju različitih sojeva iste vrste bakterija
(Primer: genotipizacija Paenibacillus larvae).
ER
IC I
ER
IC III
ER
IC I
V
ERIC II
500
1000
1500
3000
2000
ER
IC I
ER
IC III
ER
IC I
V
ERIC II
500
1000
1500
3000
2000
500
1000
1500
3000
2000

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
10
Real-time PCR - Real-time PCR se razlikuje od tradicionalne PCR metode po tome što se
uzorci obeleţe fluorescentnim markerom, a real-time PCR aparat ima detektor
fluorescencije i zahvaljujući tome Real-time PCR omogućava kvantifikaciju umnoţenog
segmenta DNK, a time i zastupljenost u uzorku, kao i detekciju PCR produkta i praćenje
kinetike reakcije od samog početka reakcije, odnosno u svakom momentu dok traje reakcija.
Ovo su velike prednosti u odnosu na konvencionalni PCR koji je samo kvalitativna metoda
bez mogućnosti kvantifikacije PCR produkta i kod koga je postupak mnogo duži jer je
potrebno da se ceo postupak završi, pa tek onda očitaju rezultati (najpre je neophodno sačekati
da se kompletno završi amplifikacija, pa obaviti elektroforezu PCR produkata, obojiti gel
etidijum bromidom i vizuelizovati PCR produkte na transiluminatoru).
Zahvaljujući opisanim karakteristikama real-time PCR pruža mogućnost primene u proceni
efikasnosti terapije lekovima, u merenju stepena oštećenja DNK, u kvantifikaciji genske
ekspresije, detekciji patogena, genotipovanju, analizi SNPs, kontroli kvaliteta i validaciji
testova.
Sekvencioniranje – odreĊivanje redosleda nukleotida. Bazira se na dideoksi metodi u
kojoj se pri in vitro sintezi DNK pored normalnih gradivnih jedinica (dNTP), dodaju i
dideoksinukleotidi (ddNTP) koji u pentoznom šećeru nemaju OH grupe na pozicijama 2' i 3',
te kada se oni ugrade u polinukleotidni lanac na tom mestu se dalja sinteza zaustavlja (jer nema
slobodne OH grupe na poziciji 3' za koju bi sledeći nukleotid trebao da se veže). Zato se ova
metoda naziva i „metoda prekida lanca“. Dideoksinukleotidi su obeleženi fluorescentnim
markerom koji je različite boje za svaki od četiri tipa dideoksinukleotida. Svaki ddNTP
fluorescira različitom bojom kada ga osvetli laserski zrak: adeninski (ddATP) zeleno,
guaninski (ddGTP) žuto, citozinski (ddCTP) plavo, timinski (ddTTP) crveno, što se automatski
zabeleži putem skenera (npr. svaki put kada se regustruje crvena boja – automatski skener
registruje da taj nukleotid nosi timin). Obeležavanje ddNTP različitim fluorescentnim bojama
omogućava da se koristi samo jedna reakciona smeša i razdvajanje (čitanje rezultata) obavlja
na jednom gelu. Čitav proces se odvija u sekvencioneru, aparatu koji sve radi automatski od
unošenja uzoraka do čitanja rezultata sa gela.
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/D/DNAsequencing.html

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
11
Molekularni (DNK) markeri
za procenu genetiĉkog diverziteta
1. NUKLEARNI DNK MARKERI
Brojni markeri su danas dostupni za detekciju polimorfizama nuklearne DNK. U prouĉavanju
genetiĉkog diverziteta i za potrebe genetiĉke karakterizacije ţivotinja, najčešće korišćeni
markeri su mikrosateliti.
MIKROSATELITI su kratke jedinice DNK (tzv. ponovci) koje se više puta ponavljaju u
tandemu - (Short Tandem Repeat - STR loci). Veličina jedinice (ponovka) je od 2 do 6
baznih parova. Najčešći ponovak je CA. Mikrosateliti su hipervarijabilni, često imaju desetine
alela po lokusu, pri čemu se različiti aleli razlikuju po broju ponovaka. RasporeĎeni su duž
čitavog genoma. Na primer, odreĎeni mikrosatelit može obuhvatati 10 ponovljenih jedinica CA
(CACACACACACACACACACA). Tokom evolucije, neka mutacija na DNK može dovesti
do toga da neke jedinke imaju alel od 8 jedinica CA, dok druge mogu imati alel od 6 ili 12 tih
jedinica na istom lokusu.
Pošto su mikrosateliti relativno kratke sekvence, lako se amplifikuju putem PCR metode iz
prethodno izolovane DNK. Vizuelizacija polimorfizama nakon amplifikacije obavlja se na
gelu, a dostupnost automatizovanih sekvencera omogućava detaljnu i preciznu analizu velikog
broja uzoraka u isto vreme.
Zbog visoke stope mutacije i kodominantne prirode, mikrosateliti su najadekvatniji markeri za:
- proučavanje diverziteta (procenu unutar-rasnog i meĎurasnog genetičkog diverziteta,
kao i procenu mešanja meĎu rasama čak i kada su one blisko srodne);
- analize roditeljstva (najčešće očinstva), kontrolu pedigrea, odnosno individualnu
identifikaciju;
- mapiranje lokusa za kvantitativne osobine (Quantitative Trait Loci – QTL).
Prosečan broj alela (number of alleles – MNA) po populaciji, uočena (zabeležena) i očekivana
heterozigotnost (Ho i He) su parametri koji se najčešće koriste za procenu unutar-rasnog
diverziteta. Najjednostavniji parametri za procenu genetičke različitosti izmeĎu rasa su
genetička diferencijacija ili fiksacioni indeksi. Postoji nekoliko načina procene, ali najčešće se
koristi FST koji predstavlja meru stepena genetičke diferencijacije subpopulacija putem
proračuna standardnih varijansi alelskih frekvenci meĎu populacijama.
Organizacija za hranu i poljoprivredu pri Ujedinjenim nacijama (Food and Agriculture
Organization of the United Nations - FAO) zvaniĉno preporuĉuje panele mikrosatelita
koje treba koristiti za procenu diverziteta glavnih vrsta domaćih ţivotinja, na osnovu
odluke i odobrenja MeĎunarodnog društva za animalnu genetiku (International Society for
Animal Genetics-ISAG) http://www.fao.org/dad-is/
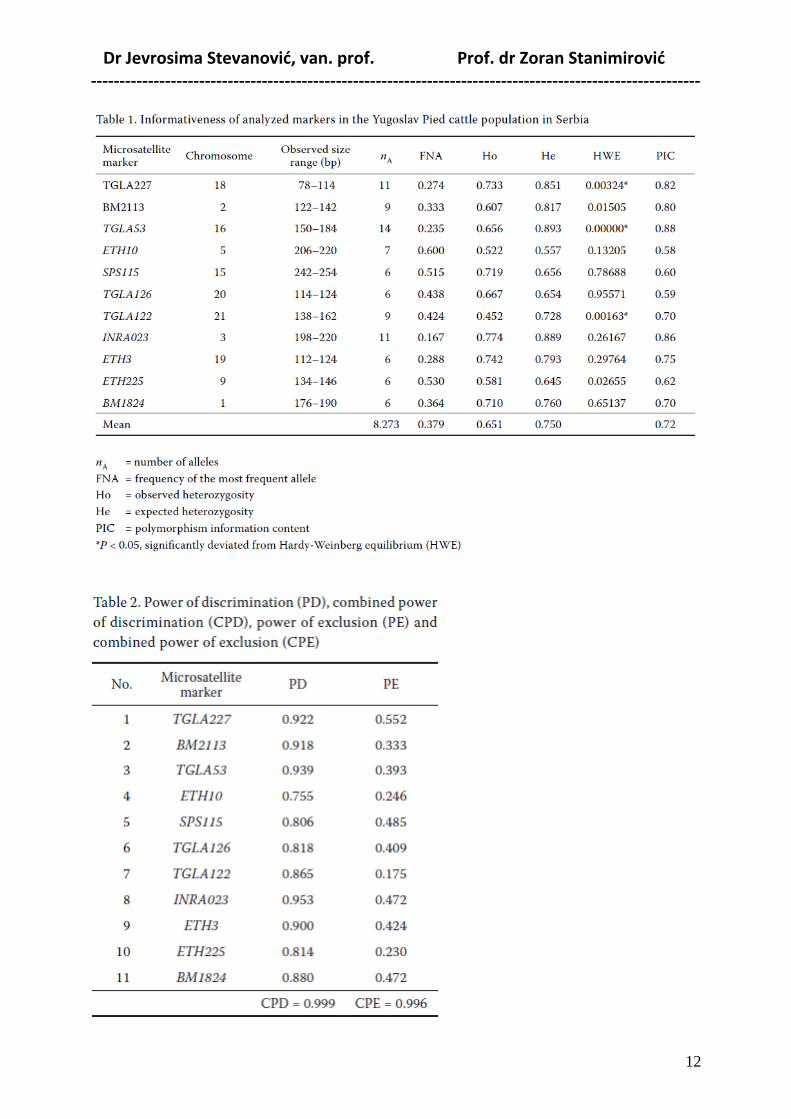
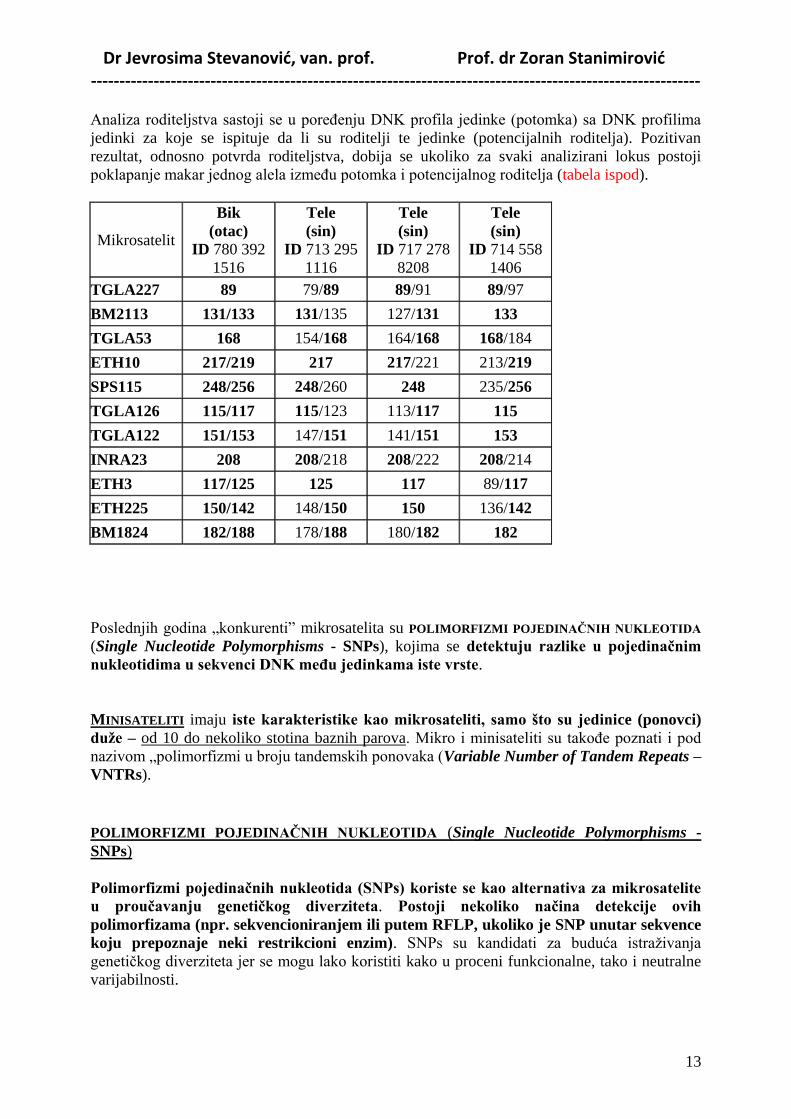
U tabelama ispod su rezultati ispitivanja informativnosti ISAG markera i mogućnosti njihovog
korišćenja u verifikaciji roditeljstva YU šarenog govečeta u tipu simentalca.
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
12
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
13
Analiza roditeljstva sastoji se u poreĎenju DNK profila jedinke (potomka) sa DNK profilima
jedinki za koje se ispituje da li su roditelji te jedinke (potencijalnih roditelja). Pozitivan
rezultat, odnosno potvrda roditeljstva, dobija se ukoliko za svaki analizirani lokus postoji
poklapanje makar jednog alela izmeĎu potomka i potencijalnog roditelja (tabela ispod).
Mikrosatelit
Bik
(otac)
ID 780 392
1516
Tele
(sin)
ID 713 295
1116
Tele
(sin)
ID 717 278
8208
Tele
(sin)
ID 714 558
1406
TGLA227 89 79/89 89/91 89/97
BM2113 131/133 131/135 127/131 133
TGLA53 168 154/168 164/168 168/184
ETH10 217/219 217 217/221 213/219
SPS115 248/256 248/260 248 235/256
TGLA126 115/117 115/123 113/117 115
TGLA122 151/153 147/151 141/151 153
INRA23 208 208/218 208/222 208/214
ETH3 117/125 125 117 89/117
ETH225 150/142 148/150 150 136/142
BM1824 182/188 178/188 180/182 182
Poslednjih godina „konkurenti” mikrosatelita su POLIMORFIZMI POJEDINAĈNIH NUKLEOTIDA
(Single Nucleotide Polymorphisms - SNPs), kojima se detektuju razlike u pojedinaĉnim
nukleotidima u sekvenci DNK meĊu jedinkama iste vrste.
MINISATELITI imaju iste karakteristike kao mikrosateliti, samo što su jedinice (ponovci)
duţe – od 10 do nekoliko stotina baznih parova. Mikro i minisateliti su takoĎe poznati i pod
nazivom „polimorfizmi u broju tandemskih ponovaka (Variable Number of Tandem Repeats –
VNTRs).
POLIMORFIZMI POJEDINAĈNIH NUKLEOTIDA (Single Nucleotide Polymorphisms -
SNPs)
Polimorfizmi pojedinaĉnih nukleotida (SNPs) koriste se kao alternativa za mikrosatelite
u prouĉavanju genetiĉkog diverziteta. Postoji nekoliko naĉina detekcije ovih
polimorfizama (npr. sekvencioniranjem ili putem RFLP, ukoliko je SNP unutar sekvence
koju prepoznaje neki restrikcioni enzim). SNPs su kandidati za buduća istraživanja
genetičkog diverziteta jer se mogu lako koristiti kako u proceni funkcionalne, tako i neutralne
varijabilnosti.
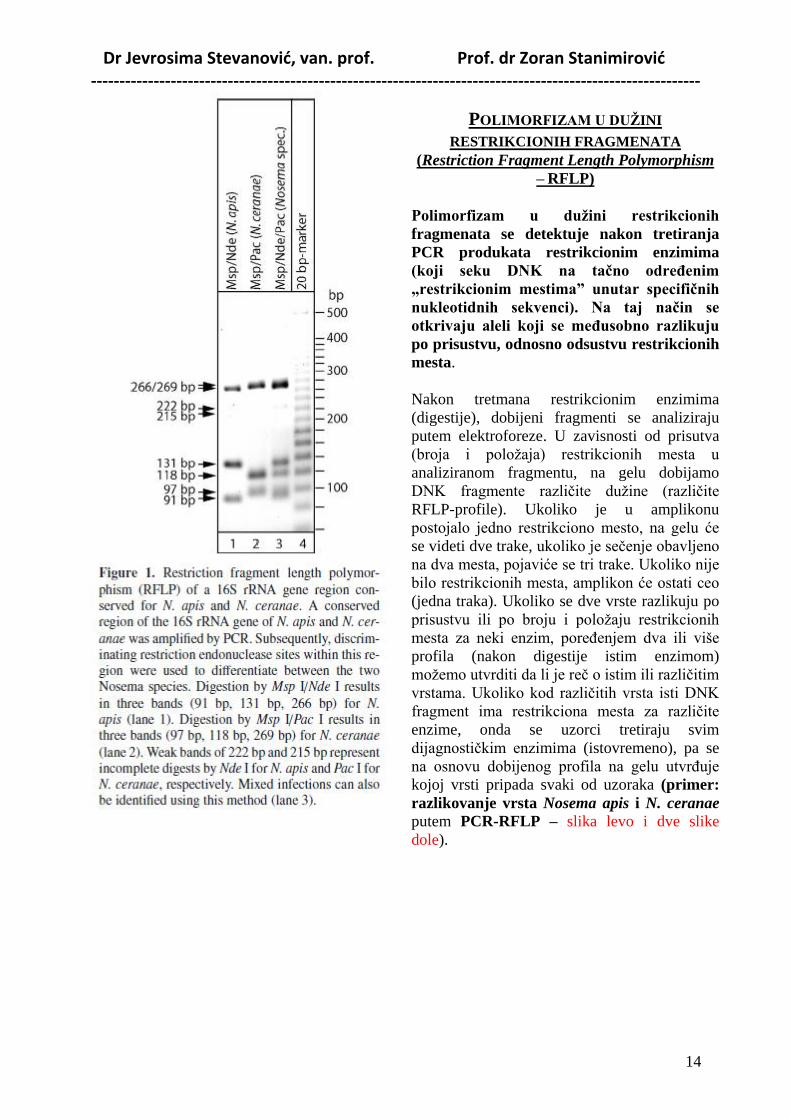
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
14
POLIMORFIZAM U DUŢINI
RESTRIKCIONIH FRAGMENATA
(Restriction Fragment Length Polymorphism
– RFLP)
Polimorfizam u duţini restrikcionih
fragmenata se detektuje nakon tretiranja
PCR produkata restrikcionim enzimima
(koji seku DNK na taĉno odreĊenim
„restrikcionim mestima” unutar specifiĉnih
nukleotidnih sekvenci). Na taj naĉin se
otkrivaju aleli koji se meĊusobno razlikuju
po prisustvu, odnosno odsustvu restrikcionih
mesta.
Nakon tretmana restrikcionim enzimima
(digestije), dobijeni fragmenti se analiziraju
putem elektroforeze. U zavisnosti od prisutva
(broja i položaja) restrikcionih mesta u
analiziranom fragmentu, na gelu dobijamo
DNK fragmente različite dužine (različite
RFLP-profile). Ukoliko je u amplikonu
postojalo jedno restrikciono mesto, na gelu će
se videti dve trake, ukoliko je sečenje obavljeno
na dva mesta, pojaviće se tri trake. Ukoliko nije
bilo restrikcionih mesta, amplikon će ostati ceo
(jedna traka). Ukoliko se dve vrste razlikuju po
prisustvu ili po broju i položaju restrikcionih
mesta za neki enzim, poreĎenjem dva ili više
profila (nakon digestije istim enzimom)
možemo utvrditi da li je reč o istim ili različitim
vrstama. Ukoliko kod različitih vrsta isti DNK
fragment ima restrikciona mesta za različite
enzime, onda se uzorci tretiraju svim
dijagnostičkim enzimima (istovremeno), pa se
na osnovu dobijenog profila na gelu utvrĎuje
kojoj vrsti pripada svaki od uzoraka (primer:
razlikovanje vrsta Nosema apis i N. ceranae
putem PCR-RFLP – slika levo i dve slike
dole).
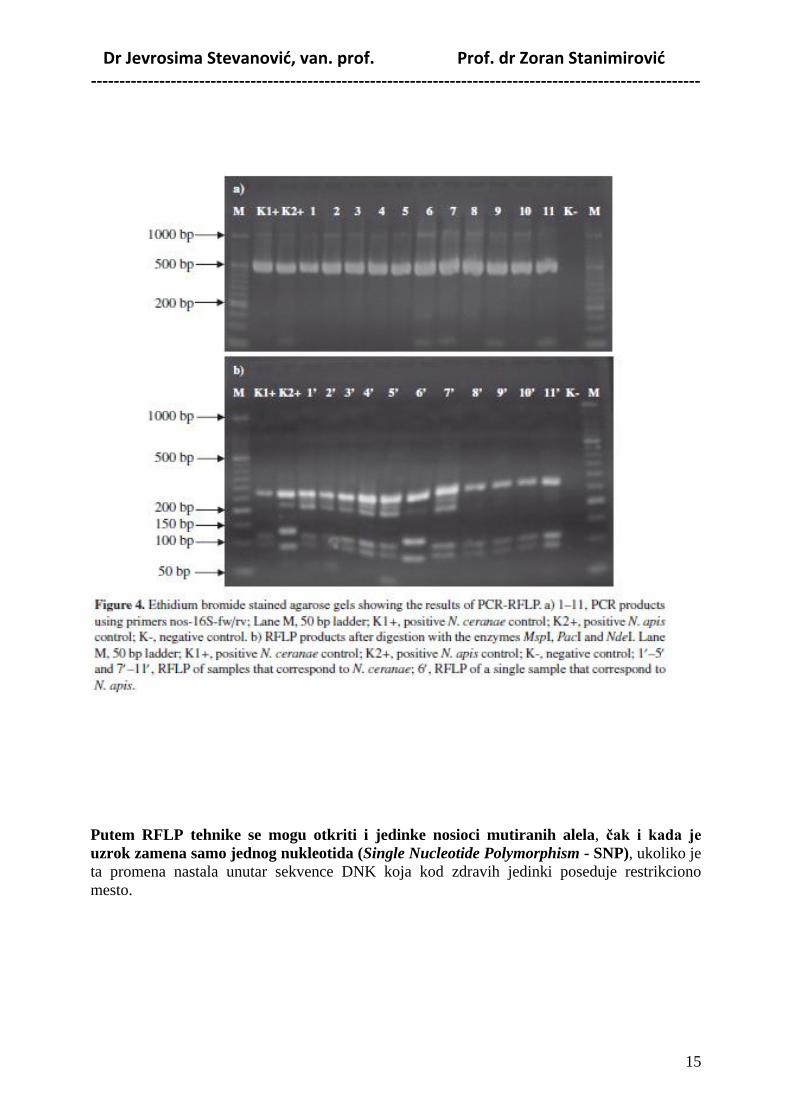
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
15
Putem RFLP tehnike se mogu otkriti i jedinke nosioci mutiranih alela, ĉak i kada je
uzrok zamena samo jednog nukleotida (Single Nucleotide Polymorphism - SNP), ukoliko je
ta promena nastala unutar sekvence DNK koja kod zdravih jedinki poseduje restrikciono
mesto.
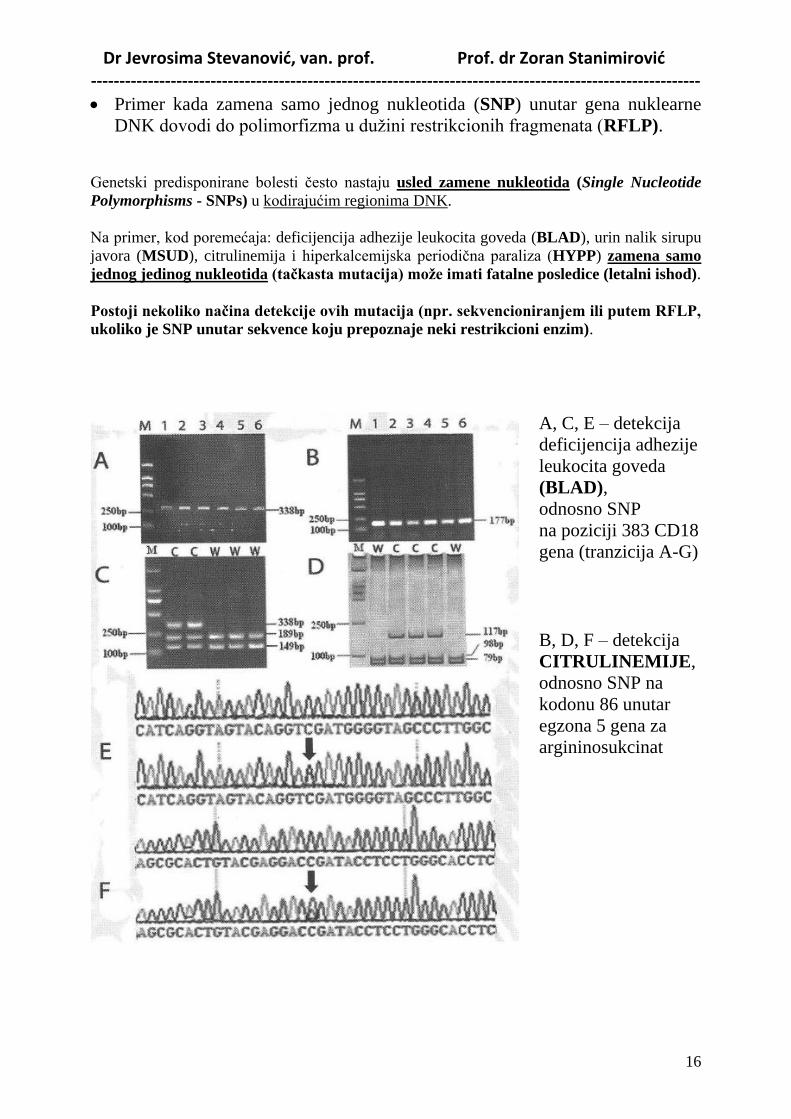
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
16
Primer kada zamena samo jednog nukleotida (SNP) unutar gena nuklearne
DNK dovodi do polimorfizma u dužini restrikcionih fragmenata (RFLP).
Genetski predisponirane bolesti često nastaju usled zamene nukleotida (Single Nucleotide
Polymorphisms - SNPs) u kodirajućim regionima DNK.
Na primer, kod poremećaja: deficijencija adhezije leukocita goveda (BLAD), urin nalik sirupu
javora (MSUD), citrulinemija i hiperkalcemijska periodična paraliza (HYPP) zamena samo
jednog jedinog nukleotida (taĉkasta mutacija) moţe imati fatalne posledice (letalni ishod).
Postoji nekoliko naĉina detekcije ovih mutacija (npr. sekvencioniranjem ili putem RFLP,
ukoliko je SNP unutar sekvence koju prepoznaje neki restrikcioni enzim).
A, C, E – detekcija
deficijencija adhezije
leukocita goveda
(BLAD),
odnosno SNP
na poziciji 383 CD18
gena (tranzicija A-G)
B, D, F – detekcija
CITRULINEMIJE,
odnosno SNP na
kodonu 86 unutar
egzona 5 gena za
argininosukcinat
sintazu (tranzicija C–
T).

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
17
Kod oboljenja pod nazivom „anemija srpastih ćelija“, zdrave jedinke imaju makar na jednom
hromozomu normalni alel za glutaminsku kiselinu na šestom mestu beta lanca hemoglobina
(normalni betaA lanac), dok su obolele jedinke homozigoti za mutirani alel i sintetišu valin
(imaju izmenjeni betaS lanac). Mutaciju je izazvala zamena samo jednog A nukleotida sa T
nukleotidom, jer je time promenjeno značenje šifre GAG (koja kodira glutaminsku kiselinu) u
GTG koja kodira valin), ali istovremeno je nestalo restrikciono mesto koje postoji samo kod
normalnog alela.
Kada je DNK fragment zdrave jedinke (koja nosi normalni alel (betaA
) na makar jednom
hromozomu), tretira restrikcionim enzimom, nakon elektroforeze se će dobiti obavezno kraća
traka (na slici obeležena kao beta-A), jer je u normalnom alelu prisutno restrikciono mesto koje
seče odeĎeni restrikcioni enzim (videti crvene strelice desno od slike gela). MeĎutim, kod
obolelih jedinki na oba hromozoma je mutirani alel (betaS) i u njemu ne postoji restrikciono
mesto, te primenjeni restrikcioni enzim ne nailazi na mesto sečenja i tretirani DNK fragmenti
tih jedinki ostaju neiseceni, pa se na gelu vidi samo duža traka (na slici obeležena kao beta-S).
Rodoslovno stablo pokazuje situaciju u porodici gde su heterozigotni roditelji (nosioci jednog
betaA i jednog beta
S alela) imali prvog sina bolesnog (homozigota za beta
S alel), drugo dete je
bila heterozigotna ćerka i treće dete normalna homozigotna ćerka (homozigot za betaA alel).
Opisana metoda omogućava prenatalnu dijagnostiku iz uzorka amnionske tečnosti ili
horionskih čupica u slučajevima kada su oba roditelja heterozigotna jer je šansa 25% da će
dobiti obolelo dete (homozigot za betaS alel), sa teškom, često smrtonosnom, anemijom
srpastih ćelija.
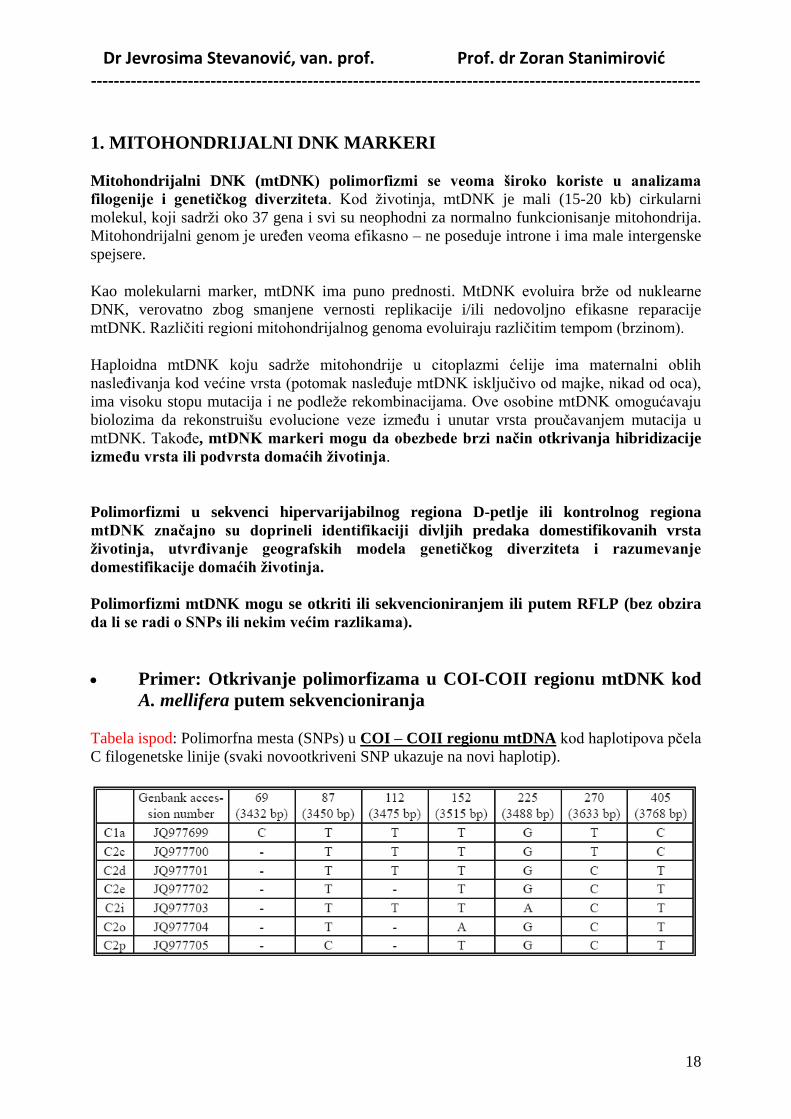
Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
18
1. MITOHONDRIJALNI DNK MARKERI
Mitohondrijalni DNK (mtDNK) polimorfizmi se veoma široko koriste u analizama
filogenije i genetiĉkog diverziteta. Kod životinja, mtDNK je mali (15-20 kb) cirkularni
molekul, koji sadrži oko 37 gena i svi su neophodni za normalno funkcionisanje mitohondrija.
Mitohondrijalni genom je ureĎen veoma efikasno – ne poseduje introne i ima male intergenske
spejsere.
Kao molekularni marker, mtDNK ima puno prednosti. MtDNK evoluira brže od nuklearne
DNK, verovatno zbog smanjene vernosti replikacije i/ili nedovoljno efikasne reparacije
mtDNK. Različiti regioni mitohondrijalnog genoma evoluiraju različitim tempom (brzinom).
Haploidna mtDNK koju sadrže mitohondrije u citoplazmi ćelije ima maternalni oblih
nasleĎivanja kod većine vrsta (potomak nasleĎuje mtDNK isključivo od majke, nikad od oca),
ima visoku stopu mutacija i ne podleže rekombinacijama. Ove osobine mtDNK omogućavaju
biolozima da rekonstruišu evolucione veze izmeĎu i unutar vrsta proučavanjem mutacija u
mtDNK. TakoĎe, mtDNK markeri mogu da obezbede brzi naĉin otkrivanja hibridizacije
izmeĊu vrsta ili podvrsta domaćih ţivotinja.
Polimorfizmi u sekvenci hipervarijabilnog regiona D-petlje ili kontrolnog regiona
mtDNK znaĉajno su doprineli identifikaciji divljih predaka domestifikovanih vrsta
ţivotinja, utvrĊivanje geografskih modela genetiĉkog diverziteta i razumevanje
domestifikacije domaćih ţivotinja.
Polimorfizmi mtDNK mogu se otkriti ili sekvencioniranjem ili putem RFLP (bez obzira
da li se radi o SNPs ili nekim većim razlikama).
Primer: Otkrivanje polimorfizama u COI-COII regionu mtDNK kod
A. mellifera putem sekvencioniranja
Tabela ispod: Polimorfna mesta (SNPs) u COI – COII regionu mtDNA kod haplotipova pčela
C filogenetske linije (svaki novootkriveni SNP ukazuje na novi haplotip).

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
19
Sedam SNPs definiše sedam haplotipova pčela A. mellifera u Srbiji: C1a, C2c, C2d, C2e, C2i
(ranije otkrivenih u drugim zemljama) i dva prvi put otkrivena haplotipa, C2o (u
banatskom regionu) i C2p (u sjeniĉko-pešterskom regionu) - utvrĎenih na osnovu dva
novootkrivena polimorfna mesta (jedne transverzije i jedna tranzicije).
Table 1. Broj analiziranih društava (n), učestalost haplotipova i diverzitet (D) u društvima
pčela A. mellifera iz Srbije i referentnim populacijama
Sekvenca COI-COII regiona mtDNK A. mellifera otkriven u cilju
otkrivanja polimorfizama – slika ispod (SNPs su obeleženi brojevima od 1 do 8).

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
20
PoreĎenje sekvence COI-COII regiona mtDNA dva haplotipa A. mellifera – slika ispod
(SNPs su obeleženi brojevima od 1 do 8)
1
1 GTATTTTTAAACTTTTATTAAAATTTCCC-ACTTAATTCATATTAATTTAAAAATAAATT 59
2 GTATTTTTAAACTTTTATTAAAATTTCCC-ACTTAATTCATATTAATTTAAAAATAAATT 59
***************************** ******************************
2 3
1 AATAACAATTTTTAATAAAATAAATAATTAATTTTATTTTTATATTGAATTTTAAATTCA 119
2 AATAACAATTTTTAATAAAATAAATAATTAATTTTATTTTTATATTGAATTTTAAATTCA 119
************************************************************
1 ATCTTAAAGATTTAATCTTTTTATTAAAATTAATAAATTAATATAAAATAAAACAAAATA 179
2 ATCTTAAAGATTTAATCTTTTTATTAAAATTAATAAATTAATATAAAATAAAACAAAATA 179
************************************************************
4*5* 6
1 TAACAGAATATATTTATTAAAATTTAATTTATTAAAATTTCCACATGATTCATATTTATA 239
2 TAACAGAATATATTTATAATAATTTAATTTATTAAAATTTCCACATGATTCATATTTATA 229
***************** * ****************************** *********
1 TTTCAAGAATCAAATTCATATTATGCTGATAATTTAATTTCATTTCATAATATAGTTATA 299
2 TTTCAAGAATCAAATTCATATTATGCTGATAATTTAATTTCATTTCATAATATAGTTATA 399
************************************************************
7*
1 ATAATTATTATTATAATTTCAACATTAACTGTATATATTATTTTAGATTTATTTATAAAC 359
2 ATAATTATTATTATAATTTCAACATTAACTGTATATATTATTTTAGATTT-TTTATAAAC 358
************************************************** *********
8
1 AAATTTTCAAATTTATTTTTATTAAAAAATCATAATATTGAAATTATTTGAACAATTATT 419
2 AAATTTTCAAATTTATTTTTATTAAAAAATCATAATATTGAAATTATTTGAACAATTATT 418
***** ******************************************************
Primer: polimorfizam u COI genskom segmentu mtDNK kod A. mellifera
koji se moţe detektovati putem RFLP uz korišćenje restrikcionih enzima
NcoI i StyI, ĉime je omogućeno razlikovanje podvrsta A. mellifera koje
dele isti (C2D) haplotip (tabela ispod).

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
21
Box 1.
Razlike u fenotipskoj ekspresiji genotipova – knjiga 28. str
Razlike u fenotipskoj ekspresiji genotipova postoje zbog interakcije gena sa sredinom (pod
sredinom se podrazumeva kako unutrašnje okruženje u kome se ćelije nalaze, tako i uticaji
spoljašnje sredine u kojoj se organizam razvija). Znači, sredina može tokom individualnog
razvoja da utiče na stepen ispoljavanja gena. Zbog toga odreĊeni genotip ne mora uvek da
se ispolji kod svih jedinki koje su njegovi nosioci.
Učestalost ispoljavanja datog gena u populaciji naziva se penetrabilnost. Penetrabilnost
zavisi kako od genotipa (prisustvo epistaze ili nekog drugog tipa interakcije), tako i od
sredine. Ukoliko se kod klasičnog dominanto-recesivnog tipa naspeĎivanja recesivni fenotip
ispolji kod svih recesivnih homozigota, a dominantan kod svih dominantnih homozigota i
heterozigota, kažemo da je penetrabilnost potpuna (100%) – npr. kod osobina graška koje je
Mendel ispitivao i kod ABO sistema krvnih grupa. MeĎutim, ukoliko manje od 100% jedinki
ispolji očekivani fenotip, penetrabilnost je nepotpuna. Kod čoveka gen za brahidaktiliju
(kratki prsti) ima penetrabilnost od 50-80%. Smatra se da i mnogi geni koji dovode do
predispozicije za nastanak malignih bolesti ispoljavaju nepotpunu penetrabilnost.
Pored razlika u ispoljavanju gena na nivou populacije, geni mogu imaju različit stepen
ispoljenosti kod jedinke. Ekpresivnost predstavlja stepen do kojeg se penetrabilan gen ili
genotip fenotipski ispoljava kod jedinke. Kao i penetrabilnost, ekspresivnost zavisi kako od
sredine, tako i od genotipa (tj. genskih interakcija). Kod nasleĎivanja prošaranosti dlake
nekih rasa domaćeg psa postoji razlika u ekspresivnosti Sp genskog alela, te se uočava
gradacija od desetak različitih fenotipova.
Konačno, neki geni pokazuju nepotpunu penetrabilnost i varijabilnu ekspresivnost, što
umnogome otežava interpretaciju rezultata različitih ukrštanja, kao i analize rodoslovnih
stabala.
Pod uticajem sredine se ponekad javljaju nenasledne promene fenotipa (fenokopije) koje su
jako slične ili identične fenotipovima nastalim usled mutacije gena (iako imaju normalnu
genetičku konstituciju, jedinke sa fenokopijom ispoljavaju mutantan fenotip). Fenokopije
mogu da nastanu pod uticajem razliĉitih hemijskih agenasa tokom ranog razvića.
primer: Katarakta, gluvoća i malformacije srca mogu da budu posledica recesivno
homozigotnog stanja odreĎenih gena, ali mogu da se jave i kod osoba normalnog fenotipa
ukoliko su njihove majke bile inficirane virusom rubeole tokom prvih nekoliko nedelja
trudnoće.
primer: Nemogućnost razvoja dugih kostiju ekstremiteta može biti posledica retke
dominantne mutacije sa varijabilnom ekspresivnošću, ali može biti izazvana i primenom
sedativa talidomida tokom graviditeta.
Brojni su uticaji sredine koji menjaju vreme ili stepen ispoljavanja gena. Na stepen
ispoljenosti gena utiču: hemijski agensi, hormonski status (različito ispoljavanje kod mužjaka i

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
22
ženki), period života u kome se gen ispoljava, različiti fizički faktori – npr. temperatura utiče
na obojenost dlake kod kunića i sijamskih mačaka).
Relativan doprinos genetičke konstitucije i činilaca sredine na razviće odreĎenog fenotipa
moţe dosta da varira zavisno od broja gena uključenih u determinaciju date osobine, kao i
od toga u kojoj je meri ispoljavanje osobine pod uticajem spoljašnje sredine.
Na visinu, pored većeg broja gena, utiču i brojni spoljašnji faktori (ishrana, fizička aktivnost,
zdravstvena zaštita). Zato je za poslednjih 100 godina u razvijenim zemljama povećana
prosečna visina ljudi zbog bolje ishrane, sporta i brige o zdravlju. U ovom slučaju, geni daju
potencijal za razviće fenotipa u odreĎenim granicama. Raspon mogućih fenotipova koji
mogu nastati od jednog istog genotipa pod različitim uslovima sredine naziva se norma
reakcije. Za neke genotipove norma reakcije je mala i oni će dati gotovo iste fenotipove u
veoma različitim uslovima sredine. Nasuprot tome, neki genotipovi imaju veliku normu
reakcije, tako da isti genotip daje niz različitih fenotipova u različitim uslovima sredine.
Pored norme reakcije, treba razlikovati fenotipsku plastiĉnost kao stepen ispoljavanja osobine
pod dejstvom faktora spoljašnje sredine.
Plejotropnost gena
Poligeno nasleĊivanje

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
23
Box 2.
Restrikcioni enzimi (endonukleaze) prepoznaju tzv. specifične nukleotidne sekvence (tzv.
palindromske sekvence) i seku ih na tačno odreĎenim „restrikcionim mestima”. Naime, za
svaki restrikcioni enzim postoji specifična sekvenca koju samo on prepoznaje i seče je na
specifičan način. (Mesta sečenja nekih restrikcionih enzima prikazana su na slici dole).
Za potrebe sekvencioniranja DNK, neophodno je prethodno iseći DNK u manje fragmente i za
tu svrhu se koriste restrikcioni enzimi.
Neki restrikcioni enzimi (kao npr. HaeIII i AluI) seku DNK ravno tako da nastaju fragmenti sa
ravnim krajevima. Ako se specifična sekvenca koju prepoznaje i seče npr. enzim HaeIII nalazi
na 11 mesta duž cirkularne DNK virusa, nastaće 11 fragmenata razliĉitih po duţini i
sekvencama nukleotida. Nastali fragmenti se mogu odvojiti jedan od drugog
elektroforezom, a zatim odrediti redosled nukleotida u njima postupkom
sekvencioniranja .
MeĎutim, mnogi restrikcioni enzimi (BamHI, HindIII, Eco RI) seku DNK neravno tako da
nastaju fragmenti sa “lepljivim” jednolanĉanim krajevima. Ukoliko se DNK poreklom iz
dva različita izvora tretiraju istim restrikcionim enzimom, nastaće fragmenti sa istim lepljivim
krajevima, te ukoliko se dobijeni fragmenti pomešaju, dolazi do njihovog povezivanja,
odnosno spajanja njihovih lepljivih krajeva po principu komplementarnosti baza.
Konačno, dodaje se DNK ligaza koja spaja mesta prekida kovalentnim vezama (formira
kovalentne veze unutar svakog lanca i kao rezultat nastaje “rekombinantna” DNK (rDNA).
Stvaranje rekombinante DNK je revolucionarno otkriće, ne samo zbog toga što je usavršilo
genetička istraživanja, nego i zbog velike mogućnosti primene u biotehnologiji i medicini.
Humani insulin (za dijabetiĉare), humani faktor VIII (za hemofiliĉare) i drugi preparati
danas se komercijalno proizvode zahvaljujuću otkriću načina stvaranja rekombinantne DNK.
http://learn.genetics.utah.edu/content/labs/extraction/

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
24
REFERENCE ISTRAŢIVAĈA SA KATEDRE ZA BIOLOGIJU U KOJIMA SU KORIŠĆENE
MOLEKULARNE METODE BAZIRANE NA PCR TEHNOLOGIJI:
1. Kozmus P, Stevanovic Jevrosima, Stanimirovic Z, Stojic V, Kulisic Z, Meglic V
(2007) Analysis of mitochondrial DNA in honey bees (Apis mellifera) from Serbia.
Acta Veterinaria 57 (5-6) 465-476.
2. Forsgren E, Stevanovic Jevrosima, Fries I (2008) Variability in germination and in
temperature and storage resistance among Paenibacillus larvae genotype. Veterinary
Microbiology 129 (3-4) 342-349.
3. Stevanovic Jevrosima, Stanimirovic Z, Dimitrijevic V, Stojic V, Fratric N, Lazarevic M
(2009) Microsatellite DNA polymorphism and its usefulness for pedigree verification in
Simmental cattle from Serbia. Acta Veterinaria 59 (5-6) 621-631.
4. Stevanovic Jevrosima, Stanimirovic Z, Dimitrijevic V, Maletic M (2010) Evaluation of
11 microsatellite loci for their use in paternity testing in the Yugoslav Pied cattle (YU
Simmental cattle). Czech Journal of Animal Science 55 (6) 221-226.
5. Stevanovic Jevrosima, Stanimirovic Z, Radakovic Milena, Kovacevic R Sanja (2010)
Biogeographic study of the honey bee (Apis mellifera L.) from Serbia, Bosnia and
Herzegovina and Republic of Macedonia based on mitochondrial DNA analyses. Russian
Journal of Genetics 46 (5) 603-609.
6. Stevanovic Jevrosima, Stanimirovic Z, Genersch Elke, Kovacevic R Sanja,
Ljubenkovic J, Radakovic Milena, Aleksic Nevenka (2011) Dominance of Nosema
ceranae in honey bees in the Balkan countries in the absence of symptoms of colony
collapse disorder. Apidologie 41 (1) 49-58.
7. Vucicevic M, Stevanov-Pavlovic Marija, Stevanovic Jevrosima, Bosnjak Jasna, Gajic
B, Aleksic Nevenka, Stanimirovic Z (2012) Sex determination in 58 bird species and
evaluation of CHD gene as a universal molecular marker in bird sexing, Zoo Biology 32
(3) 269-276.
8. Muñoz Irene, Stevanovic Jevrosima, Stanimirovic Z, De la Rúa Pilar (2012) Genetic
variation of Apis mellifera from Serbia inferred from mitochondrial analysis. Journal of
Apicultural Science 56 (1) 59-69.
9. Stevanov–Pavlovic Marija, Vucicevic Milos, Bosnjak Jasna, Stevanovic Jevrosima,
Dimitrijevic Vladimir, Resanovic Radmila, Stanimirovic Zoran (2013) Molecular sex
determination of 20 bird species protected in the Republic of Serbia. Acta Veterinaria,
63 (1) 45-51.
10. Vucicevic Milos, Stevanov-Pavlovic Marija, Stevanovic Jevrosima, Bosnjak Jasna,
Gajic Bojan, Aleksic Nevenka, Stanimirovic Zoran (2013) Sex determination in 58 bird
species and evaluation of CHD gene as a universal molecular marker in bird sexing.
Zoo Biology 32 (3) 269-276.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
25
11. Gajic Bojan, Radulovic Zeljko, Stevanovic Jevrosima, Kulisic Zoran, Vucicevic Milos,
Simeunovic Predrag, Stanimirovic Zoran (2013) Variability of the honey bee mite
Varroa destructor in Serbia based on mtDNA analysis. Experimental and Applied
Acarology 61 (1) 97-105.
12. Stevanovic Jevrosima, Simeunovic Predrag, Gajic Bojan, Lakic Nada, Radovic Dejan,
Fries Ingemar, Stanimirovic Zoran (2013) Characteristics of Nosema ceranae infection
in Serbian honey bee colonies. Apidologie 44 (5) 522-536.
13. Bosnjak Jasna, Stevanov-Pavlovic Marija, Vucicevic Milos, Stevanovic Jevrosima,
Simeunovic Predrag, Resanovic Radmila, Stanimirovic Zoran (2013) Feasibility of
non-invasive molecular method for sexing of parrots. Pakistan Journal of Zoology 45
(3) 715-720.
14. Dimitrijevic Vladimir, Stevanovic Jevrosima, Savic Mila, Petrujkic Branko,
Simeunovic Predrag, Milosevic Ivan, Stanimirovic Zoran (2013) Validation of 10
microsatellite loci for their use in parentage verification and individual identification in
the Yugoslavian Shepherd Dog – Sharplanina, Annals of Animal Science 13 (4) 715-
722.
15. Simeunovic Predrag, Stevanovic Jevrosima, Vidanovic Dejan, Nisavic Jakov, Radovic
Dejan, Stanisic Ljubodrag, Stanimirovic Zoran (2014) A survey of deformed wing virus
and acute bee paralysis virus in honey bee colonies from Serbia using real-time RT-
PCR. Acta Veterinaria 64 (1) 81-92.
16. Glavinic Uros, Stevanovic Jevrosima, Gajic Bojan, Simeunovic Predrag, Đuric
Spomenka, Vejnovic Branislav, Stanimirovic Zoran (2014) Nosema ceranae DNA in
honey bee haemolymph and honey bee mite Varroa destructor. Acta Veterinaria 64 (3)
349-357.
17. Gajic Bojan, Bogunovic Danica, Stevanovic Jevrosima, Kulišić Zoran, Simeunovic
Predrag, Stanimirovic Zoran (2014) Canine and feline thelaziosis caused by Thelazia
callipaeda in Serbia. Acta Veterinaria 64 (4) 447-455.
18. Simeunovic Predrag, Stevanovic Jevrosima, Cirkovic Dragan, Radojicic Sonja, Lakic
Nada, Stanisic Ljubodrag, Stanimirovic Zoran (2014) Nosema ceranae and queen age
influence the reproduction and productivity of the honey bee colony. Journal of
Apicultural Research 53 (5) 545-554.
19. Davitkov Darko, Vucicevic Milos, Stevanovic Jevrosima, Krstic Vanja, Tomanovic
Snezana, Glavinic Uros, Stanimirovic Zoran (2015) Clinical babesiosis and molecular
identification of Babesia canis and Babesia gibsoni infections in dogs from Serbia.
Acta Veterinaria Hungarica 63 (2) 199–208.
20. Stevanov-Pavlović Marija, Dimitrijević Vladimir, Marić Saša, Radović Dejan,
Stevanović Jevrosima, Stanimirović Zoran (2015) Applicability assessment of a
standardized microsatellite marker set in endangered Busha cattle. Slovenian Veterinary
Research 52 (3) 133-139.
21. Özvegy József, Marinković Darko, Vučićević Miloš, Gajić Bojan, Stevanović
Jevrosima, Krnjaić Dejan, Aleksic-Kovacevic Sanja (2015) Cytological and molecular

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
26
identification of Haemogregarina stepanowi in blood samples of European pond turtle
(Emys orbicularis) from quarantine at Belgrade zoo. Acta Veterinaria 65 (4) 525-533.
22. Davitkov Darko, Vucicevic Milos, Stevanovic Jevrosima, Krstic Vanja, Slijepcevic
Dajana, Glavinic Uros, Stanimirovic Zoran (2016) Molecular detection and prevalence
of Theileria equi and Babesia caballi in horses of central Balkan. Acta Parasitologica
61 (2) 337-342.
23. Vucicevic Milos, Slijepcevic Dajana, Davitkov Darko, Avdalovic Vladimir, Aleksic
Kovacevic Sanja, Stevanovic Jevrosima, Stanimirovic Zoran (2016) First report of
Polycystic kidney disease occurrence in Persian cats in Serbia. Veterinaria Italiana 52
(1) 51-56.
24. Stevanovic Jevrosima, Schwarz Ryan S, Vejnovic Branislav, Evans Jay D, Irwin
Rebecca E, Glavinic Uros, Stanimirovic Zoran (2016) Species-specific diagnostics of
Apis mellifera trypanosomatids: a nine-year survey (2007-2015) for trypanosomatids
and microsporidians in Serbian honey bees, Journal of Invertebrate Pathology, DOI:
10.1016/j.jip.2016.07.001
25. Radakovic Milena, Davitkov Darko, Borozan Sunčica, Stojanovic Srdjan, Stevanovic
Jevrosima, Krstic Vanja, Stanimirovic Zoran (2016) Oxidative stress and DNA damage
in horses naturally infected with Theileria equi. The Veterinary Journal 217, 112–118.
26. Stanisic Ljubodrag, Dimitrijevic Vladimir, Simeunovic Predrag, Glavinic Uros,
Jovanovic Biljana, Stevanovic Jevrosima, Stanimirovic Zoran (2017) Assessment of 17
microsatellite loci for their use in parentage verification and individual identification in
the balkan donkey breed. Genetika-Belgrade 49 (1) DOI: 10.2298/GENSR1601009E
27. Vucicevic Milos, Vucicevic Ivana, Davitkov Darko, Davitkov Dajana, Stevanovic
Jevrosima, Resanovic Radmila, Stanimirovic Zoran (2017) Detection and analysis of
new psittacine beak and feather disease virus (PBFDv) nucleotide sequences. Journal of
the Hellenic Veterinary Medical Society, in press.
28. Davitkov Dajana, Davitkov Darko, Vucicevic Milos, Stanisic Ljubodrag, Radakovic
Milena, Glavinic Uros, Stanimirovic Zoran (2017) A molecular and hematological
study of Theileria equi in Balkan donkeys. Acta Veterinaria Hungarica 65 (2), in press.
RADOVI SA SKUPOVA
1. Stevanović Jevrosima, Maletić M, Stanimirović M, Stanimirović Z (2008) Molekularno
genetičke metode u ekologiji i menadžmentu divljači. Zbornik radova X Savetovanja iz
kliničke patologije i terapije životinja Clinica Veterinaria2005, Sept 1-5, pp 108-109,
Kragujevac, Srbija.
2. Muñoz Irene, Dall’Olio R, Stevanovic Jevrosima, Stanimirovic Z, de la Rúa Pilar
(2009) Diversidad genetica y estructura poblacional de Apis mellifera en Europa
Oriental, Book of abstracts, Secundo Congreso de la Sociedad Espanola de Biologia
Evolutiva, Nov 29-Dec 02, pp. 78, Valencia, Spain.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
27
3. Stevanović Jevrosima, Stanimirović Z, Radaković Milena, Maletić M, Đelić Н (2009)
Molekularno genetička identifikacija roditeljstva u analizi pedigrea životinja. Zbornik
radova XI Savetovanja iz kliničke patologije i terapije životinja Clinica Veterinaria2009,
Jun 19-21, pp 47-49. Subotica, Srbija.
4. Stevanović Jevrosima, Stanimirović Z, Radaković M (2009) Species identification of
Nosema microsporidian pathogen in samples of Apis mellifera from Serbia using PCR-
RFLP method, Book of Abstracts, IV Congress of the Serbian Genetic Society, June 1-
5, pp 171, Tara, Serbia.
5. Stevanović Jevrosima, Stanimirović Z, Đelić N, Radaković M (2009) Parentage
verification and sex determination in cattle using molecular markers, Book of Abstracts,
IV Congress of the Serbian Genetic Society, June 1-5, pp 156, Tara, Serbia.
6. Stevanović Jevrosima, Stanimirović Z, Radaković M (2009) Investigations of genetic
diversity of Paenibacillus larvae from Serbia using rep-PCR fingerprint technique,
Book of Abstracts, IV Congress of the Serbian Genetic Society, June 1-5, pp 141, Tara,
Serbia.
7. Stevanović Jevrosima, Stanimirović Z, Radaković Milena, Đelić N (2009) UtvrĎivanje
roditeljstva i pola goveda iz uzoraka biološkog materijala primenom molekulskih
markera, Zbornik predavanja sa XXX Seminara za inovacije znanja veterinara, Feb 13,
pp 47-53, Beograd, Srbija.
8. Muñoz Irene, Stevanovic Jevrosima, Stanimirovic Z, de la Rúa Pilar (2010) Molecular
analysis discriminated among Serbian ecotypes of Apis mellifera carnica. Proceedings
of the 4th European Conference of Apidology EurBee 2010. Sept 7-9, pp 132, Metu-
Ankara, Turkey.
9. Maletić M, Stevanović Jevrosima, Stanimirović Z (2010) Polimorfizam laktoferin i
β4defensin gena i njihov značaj u otpornosti krava na mastitis. Zbornik radova XI
Savetovanja iz kliničke patologije i terapije životinja Clinica Veterinaria2010, Jun 18-20,
pp 55-56, Subotica, Srbija.
10. Vučićević M, Stevanov-Pavlović Marija, Bošnjak Jasna, Stevanović Jevrosima,
Stanimirović Z (2010) Determinacija pola ptica primenom molekularnih markera.
Zbornik radova XI Savetovanja iz kliničke patologije i terapije životinja Clinica
Veterinaria2010, Jun 18-20, pp 53-54, Subotica, Srbija.
11. Stevanović Jevrosima, Stanimirović Z (2010) Specijska identifikacija mikrosporidija
Nosema apis/N. ceranae korišćenjem molekularnih metoda dupleks PCR i PCR-RFLP.
Zbornik radova XI Savetovanja iz kliničke patologije i terapije životinja Clinica
Veterinaria2010, Jun 18-20, pp 45-47, Subotica, Srbija.
12. Gajić Bojan, Radulović Željko, Stevanovic Jevrosima, Kulišić Zoran, Stanimirović
Zoran (2011) Preliminarni rezultati molekularno-genetičkih analiza varijabilnosti
mtDNK Varroa destructor u pčelinjim zajednicama na teritoriji Srbije. Zbornik
plenarnih referata i rezimea, Simpozijum entomologa Srbije 2011, Sep 21-25, p 69.
Donji Milanovac, Srbija.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
28
13. Stanimirović Zoran, Stevanović Jevrosima (2012) Primena molekularno-genetičkih
analiza u veterinarskoj medicini. Zbornik predavanja sa XXXIII Seminara za inovacije
znanja veterinara, Feb 24, pp 17-33, Beograd, Srbija.
14. Vučićević Miloš, Stevanović Jevrosima, Simeunović Predrag, Vučićević Ivana, Đelić
Ninoslav, Stanimirović Zoran, Stojić Velibor (2012) Analysis of the CHD gene for sex
determination of protected bird species, Proceedings of the International symposium on
hunting »Мodern aspects of sustainable management of game population«, June 22–24,
pp. 83-86, Zemun-Belgrade, Serbia.
15. Vučićević Miloš, Stevanović Jevrosima, Vučićević Ivana, Pantelić Aleksandar, Đelić
Ninoslav, Resanović Radmila, Stanimirović Zoran (2012) Sex determination in game
birds management, Proceedings of the International symposium on hunting »Мodern
aspects of sustainable management of game population«, June 22–24, pp. 91-94,
Zemun-Belgrade, Serbia.
16. Vučićević Miloš, Stevanović Jevrosima, Stanimirović Zoran (2012) Provera uspešnosti
izolacije DNK iz različitih tkiva ptica, Zbornik radova XIV Regionalnog savetovanja iz
kliničke patologije i terapije životinja Clinica Veterinaria2012, Jun 14-16, pp. 132-133,
Subotica, Srbija.
17. Čobanović Nikola, Vučićević Miloš, Stevanović Jevrosima, Stanimirović Zoran (2012)
Uzorkovanje tkiva i izolacija DNK za molekularna ispitivanja kod gmizavaca, Zbornik
radova XIV Regionalnog savetovanja iz kliničke patologije i terapije životinja Clinica
Veterinaria2012, Jun 14-16, pp.130-131, Subotica, Srbija.
18. Gajić Bojan, Radulović Željko, Stevanović Jevrosima, Kulišić Zoran, Simeunović
Predrag, Stanimirović Zoran (2013) Varijabilnost mtDNK ektoparzita Varroa
destructor u Srbiji: detekcija novih haplotipova. Plenarni referati i rezimei, Simpozijum
entomologa Srbije sa meĎunarodnim učešćem 2013, 18-22. septembar 2013. pp. 31.
Tara, Srbija.
19. Stevanović Jevrosima, Munoz Irene, De la Rua Pilar, Gajić Bojan, Stanimirović Zoran
(2013) Genetički diverzitet medonosne pčele Apis mellifera u Srbiji na osnovu analize
mtDNK. Plenarni referati i rezimei, Simpozijum entomologa Srbije sa meĎunarodnim
učešćem 2013, 18-22. septembar 2013. pp. 33. Tara, Srbija.
20. Vučićević Miloš, Simeunović Predrag, Davitkov Darko, Stanišić Ljubodrag, Stevanović
Jevrosima, Stanimirović Zoran (2014) Analize DNK i RNK u funkciji detekcije i
genotipizacije patogena životinja i njihov značaj u veterinarskoj praksi. Zbornik
predavanja XVI regionalnog savetovanja iz kliničke patologije i terapije životinja
Clinica Veterinari 2014, Jun 23-25, pp. 152-159, Kopaonik, Srbija.
21. Gajić Bojan, Bogunović Danica, Stevanović Jevrosima, Kulišić Zoran, Simeunović
Predrag, Glavinić Uroš, Stanimirović Zoran (2014) Molecular identification of Thelazia
callipaeda in dogs and cats from Serbia. Book of Abstracts, V Congress of the Serbian
Genetics Society, Sept 28-Oct 2, p. 74, Kladovo, Serbia.
22. Gajic Bojan, Stevanovic Jevrosima, Radulovic Zeljko, Glavinic Uros, Kulisic Zoran,
Stanimirovic Zoran (2014) Haplotype determination of Varroa destructor mites in

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
29
Serbia using ARMS and RFLP methods, Book of Abstracts, Sixth European
Conference of Apidology (EURBEE6). Sept 9-11, pp. 105, Murcia, Spain.
23. Stanimirovic Zoran, Simeunovic Predrag, Stevanovic Jevrosima, Vidanovic Dejan,
Glavinic Uros (2014) Occurrence and distribution of deformed wing virus, acute bee
paralysis virus, sacbrood virus and chronic bee paralysis virus in honey bees (Apis
mellifera) in Serbia – a Real-time RT-PCR based survey, Book of abstracts, Sixth
European Conference of Apidology (EURBEE6). Sept 9-11, pp. 47, Murcia, Spain.
24. Davitkov Darko, Vučićević Miloš, Stevanović Jevrosima, Krstić Vanja, Gajić Bojan,
Glavinić Uroš, Stanimirović Zoran (2014) Značaj PCR-RFLP metode u preciznoj,
specijskoj identifikaciji uzročnika babezioze pasa (plenarni rad). Zbornik radova i
kratkih sadržaja, 25. Savetovanje veterinara Srbije, Sept 11-14, pp. 299-303, Zlatibor,
Srbija.
25. Davitkov Darko, Vučićević Milos, Stevanović Jevrosima, Krstić Vanja, Tomanović
Snežana, Glavinić Uroš, Stanimirović Zoran (2014) Molecular characterization of
Babesia canis and B. gibsoni from naturally infected dogs from Serbia. Book of
Abstracts, V Congress of the Serbian Genetics Society, Sept 28-Oct 2, p. 219,
Kladovo, Serbia.
26. Stevanov-Pavlović Marija, Dimitrijević Vladimir, Marić Saša, Stevanović Jevrosima,
Stanimirović Zoran (2014) Verification of a standardized microsatellite marker set for
paternity testing in endangered Busha cattle. Book of Abstracts, V Congress of the
Serbian Genetics Society, Sept 28-Oct 2, p. 195, Kladovo, Serbia.
27. Radaković Milena, Djelić Ninoslav, Stevanović Jevrosima, Soković Marina, Glavinić
Uroš, Van Griensven L.J.L.D., Stanimirović Zoran (2014) Evaluation of antigenotoxic
potential of Agaricus blazei extract against thymol in the Comet assay. Book of
Abstracts, V Congress of the Serbian Genetics Society, Sept 28-Oct 2, p. 156,
Kladovo, Serbia.
28. Vučićević Milos, Slijepčevic Dajana, Davitkov Darko, Stevanović Jevrosima, Krstić
Vanja, Ilić Vojislav, Stanimirović Zoran (2014) Frequency of Polycystic Kidney
Disease (PKD) in population of Persian cats from Serbia and comparison between
ultrasound diagnosis and genetic testing. Book of Abstracts, V Congress of the
Serbian Genetics Society, Sept 28-Oct 2, p. 76, Kladovo, Serbia.
29. Maletić Milan, Vakanjac Slobodanka, Djelić Ninoslav, Lakić Nada, Stevanović
Jevrosima, Glavinić Uroš, Stanimirović Zoran (2014) Association of lactoferrin gene
polymorphism with mammary gland health and production characteristics of Holstein-
Friesian cows. Book of Abstracts, V Congress of the Serbian Genetics Society, Sept 28-
Oct 2, p. 75, Kladovo, Serbia.
30. Stanimirović Zoran, Simeunović Predrag, Vučićević Miloš, Davitkov Darko, Stanišić
Ljubodrag, Maletić Milan, Gajić Bojan, Glavinić Uroš, Stevanović Jevrosima (2015)
Primena molekularno-genetičkih analiza u forenzici i dijagnostici kod domaćih
životinja i divljači. Zbornik predavanja sa XXXVI Seminara za inovacije znanja
veterinara, Feb 20, pp 137-146, Beograd, Srbija.

Dr Jevrosima Stevanović, van. prof. Prof. dr Zoran Stanimirović -----------------------------------------------------------------------------------------------------------
30
31. Stevanovic Jevrosima, Schwarz S. Ryan, Vejnovic Branislav, Evans D. Jay, Irwin E.
Rebecca, Glavinic Uros, Stanimirovic Zoran (2016) The earliest record and first report
of Lotmaria passim in Serbian honey bees. Proceedings of the Seventh European
Conference of Apidology (EURBEE7). Sept 7-9, pp. 110-111, Cluj-Napoca, Romania.
32. Stanimirović Zoran, Simeunović Predrag, Stanišić Ljubodrag, Maletić Milan,
Stevanović Jevrosima (2016) Determinacija pola kod domaćih životinja, plenarni
referat, Zbornik predavanja, 7. Naučni Simpozijum Reprodukcija domaćih životinja, 6-
9. Okt, str. 105-116, Divčibare, Srbija.
SVA NAŠA GENOMSKA ISTRAŢIVANJA PUBLIKOVANA U NAUĈNIM
RADOVIMA, KAO I SVE NOVE SEKVENCE DEPONOVANE U GENSKOJ BAZI
nalaze se na internet stranici: http://katedre.vet.bg.ac.rs/~biolog/index.php?option=com_content&view=article&id=4&Itemid=6


















