
AN IMMUNOCYTOCHEMICAL MAPPING OF METHIONINE-atarazanas.sci.uma.es/docs/articulos/16493783.pdf · AN...
17
AN IMMUNOCYTOCHEMICAL MAPPING OF METHIONINE- ENKEPHALIN-ARG 6 -GLY 7 -LEU 8 IN THE HUMAN BRAINSTEM R. COVEÑAS, a * F. MARTÍN, a P. SALINAS, b E. RIVADA, b V. SMITH, b L. A. AGUILAR, a Z. DÍAZ-CABIALE, c J. A. NARVÁEZ c AND G. TRAMU d a Institute of Neurosciences of Castilla y León, Laboratory of Neuro- anatomy of the Peptidergic Systems, Facultad de Medicina, Campus Unamuno, c/ Alfonso X El Sabio s/n, 37007 Salamanca, Spain b University of Málaga, School of Medicine, Department of Anatomy, 29080 Málaga, Spain c University of Málaga, School of Medicine, Department of Physiology, 29080 Málaga, Spain d University of Bordeaux I, Laboratory of Functional Neurocytochemis- try, C.N.R.S., 33405 Talence, France Abstract—Using an indirect immunoperoxidase technique, we studied the distribution of immunoreactive fibers and cell bodies containing methionine-enkephalin-Arg 6 -Gly 7 -Leu 8 in the adult human brainstem. Immunoreactive cell bodies were found in the reticular formation of the medulla oblongata (in which we observed the highest density of immunoreactive cell bodies) and the pons, the solitary nucleus, the hypoglos- sal nucleus, the medial and spinal vestibular nuclei, the lat- eral cuneate nucleus, the nucleus prepositus, the central gray of the pons and mesencephalon, the central and peri- central nuclei of the inferior colliculus, the superior collicu- lus, ventral to the superior olive and in the midline region of the pons and mesencephalon. The highest density of immu- noreactive fibers containing methionine-enkephalin-Arg 6 - Gly 7 -Leu 8 was found in the spinal trigeminal nucleus, the central gray and the reticular formation of the medulla oblon- gata, pons and mesencephalon, the solitary nucleus, the spinal vestibular nucleus, the dorsal accessory olivary nu- cleus, the raphe obscurus, the substantia nigra and in the interpeduncular nucleus. The widespread distribution of im- munoreactive structures containing methionine-enkephalin- Arg 6 -Gly 7 -Leu 8 in the human brainstem indicates that this neuropeptide might be involved in several physiological mechanisms, acting as a neurotransmitter and/or neuromodulator. © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: man, medulla oblongata, mesencephalon, pons, pro-enkephalin. Methionine-enkephalin-Arg 6 -Gly 7 -Leu 8 (Met 8 ) is a peptide derived from the precursor pro-enkephalin. It is known that Met 8 is exclusively produced from pro-enkephalin and not by the other two precursors (pro-opiomelanocortin and pro-dynorphin) of opioid peptides. Thus, the presence of Met 8 is a good marker for the pro-enkephalin system (Abe et al., 1987; Murakami et al., 1987). This peptide has been shown to act as a neurotransmitter or neuromodulator and has been implicated in pain, cardiovascular, and motor mechanisms (Akill et al., 1984; Basbaum and Fields, 1984; Pasternak, 1987; Douglas and Kitchan, 1992). Its distribu- tion has been studied using immunocytochemical and ra- dioimmunoassay techniques in the rat, cat and monkey central nervous systems (Pittius et al., 1984; Zamir et al., 1985; Fallon and Leslie, 1986; Ibuki et al., 1989; Murakami et al., 1987, 1989; Tanaka et al., 1993; Belda et al., 2003). Previous reports using a combination of radioimmunoas- say, gel filtration, and high-performance liquid chromatog- raphy have only demonstrated the presence of Met 8 in eight nuclei of the human brainstem (e.g. periaqueductal gray, substantia nigra) and in a general form in the dorsal and ventral medulla oblongata (Pittius et al., 1984; Iadarola et al., 1991). Thus, at the present very few data are avail- able on the distribution of Met 8 in the human brainstem (Pittius et al., 1984; Iadarola et al., 1991), and in this sense, to date the distribution of Met 8 -immunoreactive (Met 8 -ir) fibers and cell bodies in the human CNS has not been studied. In light of the above and in order to know a more complete distribution of Met 8 in the human brainstem, here we were prompted to study the distribution of cell bodies and fibers containing Met 8 in the whole human brainstem using an immunocytochemical technique. EXPERIMENTAL PROCEDURES Brainstem material was obtained from five adult human brains (School of Medicine, University of Málaga, Spain) with no previous history of neurological or psychiatric disease (two men, 81 (M-1) and 82 (M-2) years; three women, 69 (W-1), 84 (W-2) and 87 (W-3) years), who died from renal failure, anemia, or myocardial infarct. The experimental design, protocols, and procedures of this work were performed under the guidelines of the ethics and legal recommendations of Spanish and European law, as well as in accordance with the Declaration of Helsinki. This work has been approved by the research commission of the University of Málaga (Spain). All procedures were carried out with the adequate under- standing and written consent of the subjects. Autopsies were carried out within 24 – 48 h after death. The brains were removed and perfused ex situ via the carotid and vertebral arteries. Both arteries were connected to a pressure- driven pump and the brains were perfused at normal mean arterial pressure with 1000 ml of 0.15 M phosphate-buffered saline (PBS; pH 7.2), followed by 3000 ml of 4% paraformal- dehyde in the same buffer. After postfixation (30 days in the latter fixative solution) at 4 °C, the brains were kept in PBS at 4 °C and cryoprotected, at the same temperature, by immersion in increasing sucrose baths (10 –30%) until they sank. The brainstems were dissected out and, using a cryostat, 60 m- *Corresponding author. Tel: 34-923-294400x1856; fax: 34-923-294549. E-mail address: [email protected] (R. Coveñas). Abbreviations: ir, immunoreactive; Met 8 , methionine-enkephalin-Arg 6 - Gly 7 -Leu 8 ; PBS, phosphate-buffered saline. Neuroscience 128 (2004) 843– 859 0306-4522/04$30.000.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2004.07.009 843
Transcript of AN IMMUNOCYTOCHEMICAL MAPPING OF METHIONINE-atarazanas.sci.uma.es/docs/articulos/16493783.pdf · AN...

AE
RVJa
aUb
2c
2d
t
AwbtfwcsegcltnGcgscimAnmnr
Kp
MdMbpM
*�EAG
Neuroscience 128 (2004) 843–859
0d
N IMMUNOCYTOCHEMICAL MAPPING OF METHIONINE-NKEPHALIN-ARG6-GLY7-LEU8 IN THE HUMAN BRAINSTEM
eshmPtdc1ePsregaea(s(b
cwau
B(ha(iwraa(s
bvdasdl4i
. COVEÑAS,a* F. MARTÍN,a P. SALINAS,b E. RIVADA,b
. SMITH,b L. A. AGUILAR,a Z. DÍAZ-CABIALE,c
. A. NARVÁEZc AND G. TRAMUd
Institute of Neurosciences of Castilla y León, Laboratory of Neuro-natomy of the Peptidergic Systems, Facultad de Medicina, Campusnamuno, c/ Alfonso X El Sabio s/n, 37007 Salamanca, Spain
University of Málaga, School of Medicine, Department of Anatomy,9080 Málaga, Spain
University of Málaga, School of Medicine, Department of Physiology,9080 Málaga, Spain
University of Bordeaux I, Laboratory of Functional Neurocytochemis-ry, C.N.R.S., 33405 Talence, France
bstract—Using an indirect immunoperoxidase technique,e studied the distribution of immunoreactive fibers and cellodies containing methionine-enkephalin-Arg6-Gly7-Leu8 in
he adult human brainstem. Immunoreactive cell bodies wereound in the reticular formation of the medulla oblongata (inhich we observed the highest density of immunoreactiveell bodies) and the pons, the solitary nucleus, the hypoglos-al nucleus, the medial and spinal vestibular nuclei, the lat-ral cuneate nucleus, the nucleus prepositus, the centralray of the pons and mesencephalon, the central and peri-entral nuclei of the inferior colliculus, the superior collicu-us, ventral to the superior olive and in the midline region ofhe pons and mesencephalon. The highest density of immu-oreactive fibers containing methionine-enkephalin-Arg6-ly7-Leu8 was found in the spinal trigeminal nucleus, theentral gray and the reticular formation of the medulla oblon-ata, pons and mesencephalon, the solitary nucleus, thepinal vestibular nucleus, the dorsal accessory olivary nu-leus, the raphe obscurus, the substantia nigra and in the
nterpeduncular nucleus. The widespread distribution of im-unoreactive structures containing methionine-enkephalin-rg6-Gly7-Leu8 in the human brainstem indicates that thiseuropeptide might be involved in several physiologicalechanisms, acting as a neurotransmitter and/or
euromodulator. © 2004 IBRO. Published by Elsevier Ltd. Allights reserved.
ey words: man, medulla oblongata, mesencephalon, pons,ro-enkephalin.
ethionine-enkephalin-Arg6-Gly7-Leu8 (Met8) is a peptideerived from the precursor pro-enkephalin. It is known thatet8 is exclusively produced from pro-enkephalin and noty the other two precursors (pro-opiomelanocortin andro-dynorphin) of opioid peptides. Thus, the presence ofet8 is a good marker for the pro-enkephalin system (Abe
Corresponding author. Tel: �34-923-294400x1856; fax:34-923-294549.-mail address: [email protected] (R. Coveñas).bbreviations: ir, immunoreactive; Met8, methionine-enkephalin-Arg6-
7 8
bly -Leu ; PBS, phosphate-buffered saline.306-4522/04$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.07.009
843
t al., 1987; Murakami et al., 1987). This peptide has beenhown to act as a neurotransmitter or neuromodulator andas been implicated in pain, cardiovascular, and motorechanisms (Akill et al., 1984; Basbaum and Fields, 1984;asternak, 1987; Douglas and Kitchan, 1992). Its distribu-
ion has been studied using immunocytochemical and ra-ioimmunoassay techniques in the rat, cat and monkeyentral nervous systems (Pittius et al., 1984; Zamir et al.,985; Fallon and Leslie, 1986; Ibuki et al., 1989; Murakamit al., 1987, 1989; Tanaka et al., 1993; Belda et al., 2003).revious reports using a combination of radioimmunoas-ay, gel filtration, and high-performance liquid chromatog-aphy have only demonstrated the presence of Met8 inight nuclei of the human brainstem (e.g. periaqueductalray, substantia nigra) and in a general form in the dorsalnd ventral medulla oblongata (Pittius et al., 1984; Iadarolat al., 1991). Thus, at the present very few data are avail-ble on the distribution of Met8 in the human brainstemPittius et al., 1984; Iadarola et al., 1991), and in thisense, to date the distribution of Met8-immunoreactiveMet8-ir) fibers and cell bodies in the human CNS has noteen studied.
In light of the above and in order to know a moreomplete distribution of Met8 in the human brainstem, heree were prompted to study the distribution of cell bodiesnd fibers containing Met8 in the whole human brainstemsing an immunocytochemical technique.
EXPERIMENTAL PROCEDURES
rainstem material was obtained from five adult human brainsSchool of Medicine, University of Málaga, Spain) with no previousistory of neurological or psychiatric disease (two men, 81 (M-1)nd 82 (M-2) years; three women, 69 (W-1), 84 (W-2) and 87W-3) years), who died from renal failure, anemia, or myocardialnfarct. The experimental design, protocols, and procedures of thisork were performed under the guidelines of the ethics and legal
ecommendations of Spanish and European law, as well as inccordance with the Declaration of Helsinki. This work has beenpproved by the research commission of the University of MálagaSpain). All procedures were carried out with the adequate under-tanding and written consent of the subjects.
Autopsies were carried out within 24 – 48 h after death. Therains were removed and perfused ex situ via the carotid andertebral arteries. Both arteries were connected to a pressure-riven pump and the brains were perfused at normal meanrterial pressure with 1000 ml of 0.15 M phosphate-bufferedaline (PBS; pH 7.2), followed by 3000 ml of 4% paraformal-ehyde in the same buffer. After postfixation (30 days in the
atter fixative solution) at 4 °C, the brains were kept in PBS at°C and cryoprotected, at the same temperature, by immersion
n increasing sucrose baths (10 –30%) until they sank. The
rainstems were dissected out and, using a cryostat, 60 �m-ved.

topsi
I
If(TfTs4(wthPdstnc
S
Pg(wBcit
t
ahAaiwec
MMhtw
sprrpwhtc(
Csbagn
G
A
FHaa
R. Coveñas et al. / Neuroscience 128 (2004) 843–859844
hick frontal sections of the mesencephalon, pons and medullablongata were cut and collected in PBS, kept at 4 °C, androcessed for immunostaining. In general, six out of sevenections were used for immunocytochemistry, and the remain-
ng section was stained for the Nissl technique.
mmunocytochemistry
n order to avoid possible interference by endogenous peroxidase,ree-floating sections were treated with distilled water containing NH3
20%), H2O2 (30%) and NaOH (1%) for 20 min (Guntern et al., 1989).hen, the sections were washed for 20 min in PBS and preincubated
or 30 min in PBS containing 1% normal horse serum and 0.3%riton X-100. The sections were then washed for 30 min in the latterolution containing casein (0.5%) and were incubated overnight at°C in PBS containing Triton X-100 (0.3%) and normal horse serum
1%), as well as anti-Met8 antiserum (diluted 1/30,000). The sectionsere then washed in PBS (30 min) and incubated for 60 min at room
emperature with biotinylated antirabbit immuno-� globulin (made inorse) (Vector, USA) diluted 1/200 in PBS. After a 30 min wash withBS, the sections were incubated for 1 h with avidin–biotin–peroxi-ase complex (Vectastain; diluted 1/100). Finally, after washing theections in PBS (30 min) and Tris–HCl buffer (pH 7.6; 10 min), theissue-bound peroxidase was developed by H2O2 using 3,3= diami-obenzidine as chromogen. The sections were rinsed with PBS andoverslipped with PBS/Glycerol (1/1).
pecificity of the antiserum
olyclonal anti-Met8 antibody was raised in rabbits against immuno-ens prepared by coupling the whole synthetic Met8 peptideBachem, Switzerland) to a carrier protein (human serum albumin)ith glutaraldehyde, as previously reported (Marcos et al., 1999;elda et al., 2003). Rabbit antiserum was preabsorbed with thearrier protein and the couplin agent in order to prevent non-specific
mmunoreactivity due to the anti-carrier antibodies. This preabsorp-ion was carried out before the immunocytochemical applications.
To determine the specificity of the immunostaining, several his-
ig. 1. Distribution of Met8-ir fibers and cell bodies in frontal planesaines (1987). Cell bodies containing the neuropeptide are representre represented by dotted lines (single fibers or low density), continbbreviations for nomenclature.
ologic controls were carried out. The primary antibody was pre- w
bsorbed with synthetic Met8 (1.11�10�4 M) and after the immuno-istochemical protocol no immunostaining appeared (Fig. 17A–C).lso, sections were incubated omitting the synthetic Met8 antibodynd no labeling was detected. In addition, no significant reduction in
mmunoreactivity was observed upon incubating the first antibodyith an excess (10�7 M) of synthetic methionine-enkephalin, leucine-nkephalin, methionine-enkephalin-Arg6-Phe7, �-, �-, and �-melano-yte-stimulating hormone, dynorphin A, or �-neo-endorphin.
appingapping was carried out according to Haines’ (1987) atlas of theuman brain, and the same atlas was used for the terminology ofhe brainstem nuclei. In addition, the atlas of Paxinos et al. (1990)as consulted.
The number of immunostained cell bodies appearing in eachection from each person was counted: a high density of Met8-irerikarya was considered when we found more than 20 profiles/egion/section; a moderate density when we found 10–20 profiles/egion/section, and a low density when we found fewer than 10rofiles/region/section. The density of the immunopositive fibersas graded under microscopic observation into four categories:igh, moderate, low and single. This was accomplished by viewinghe sections under bright-field illumination at a constant magnifi-ation with reference to photographs of defined series of densitiessee Marcos et al., 1998).
Finally, photomicrographs were obtained with an 8-bitoolSNAP digital camera attached to a Zeiss Axiophot micro-cope. To improve the visualization of the results, only therightness and contrast of the images were adjusted, withoutny further manipulation of the photographs. Adobe Photo-raph 6.0 was used to view the images and alter their bright-ess and contrast.
RESULTS
eneral considerations
s shown in Figs. 1– 6, Met8 structures were found to be
man medulla oblongata according to the atlas of the human brain ofsed circles (high density), and closed squares (low density). Ir fiberses (moderate density) and crossed lines (high density). See list of
of the hued by clouous lin
idely distributed throughout the human brainstem. Our

rsdast2iti
odiyoomupM
4AAAAAAACCCCCCCCCCDDDDDDDEFFFFFFgGGHIIIIIIIIIIJLLLLLm
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 845
esults indicate that Met8 is present in the human brain-tem, at least until 48 h after death. In general, both theistribution of the ir structures (fibers and cell bodies),nd their density, were fairly similar in the five brain-tems studied. Thus, we did not find great differences inhe human brainstems, which were perfused ex situ from4 h to 48 h after death, nor in the density of the
mmunoreactivity nor in the distribution of the ir struc-ures. In general, no sex or age differences were foundn the distribution and density of the Met8-ir structures
Abbreviation
area postremabdNr abducens nervebdNu abducens nucleusccNu accessory nucleusCSp anterior corticospinal tractLS anterolateral systemMV anterior medullary velumrNu arcuate nucleusA cerebral aqueductbl cerebellumC crus cerebrieGy central grayoBul corticobulbar fibersocNr cochlear nerveortNig corticonigral fibersSNu chief sensory nucleus (caudal end)Sp corticospinal fibersTT central tegmental tractAO dorsal accessory olivary nucleusCNu dorsal cochlear nucleusLF dorsal longitudinal fasciculusMNu dorsal motor nucleus of the vagusNu dentate nucleusSCT dorsal spinocerebellar tractTTr dorsal trigeminothalamic tractAF external arcuate fibersac, G facial nerve, internal genuacNr facial nerveacNu facial nucleusCu cuneate fasciculusGr gracile fasciculusPon frontopontine fibers
gelatinosa part of the spinal trigeminal nucleusIF glossopharyngeal fibers (intramedullary)INr glossopharyngeal nerveyNu hypoglossal nucleus
AF internal arcuate fibersC, Br inferior colliculus, brachiumC, CNu inferior colliculus, central nucleusC, Com inferior colliculus, commissureC, LZ inferior colliculus, lateral zoneCP inferior cerebellar peduncleC, PNu inferior colliculus, pericentral nucleusPFo interpeduncular fossaPNu interpeduncular nucleusSNu inferior salivatory nucleusRB juxtarestiform bodyCNu lateral cuneate nucleusL lateral lemniscusL, Nu lateral lemniscus, nucleusoCer locus ceruleusRNu lateral reticular nucleus
magnocellular part of the spinal trigeminal nucleus
bserved in the human brainstems studied. Thus, theistribution and density of Met8-ir fibers and cell bodies
n the brainstem of the youngest person studied (69ears) were in general similar to those observed in thelder persons (81, 82, 84 and 87 years). However, webserved some minor differences among the speci-ens. Thus, ir cell bodies located in the medial vestib-lar nucleus, the hypoglossal nucleus and in the nucleusrepositus were respectively only found in the brains-1 (81 years; perfused 24 h after death), W-2 (84
the figures
O medial accessory olivary nucleusP middle cerebellar pedunclesNu mesencephalic nucleussTr mesencephalic tract
medial lemniscusF medial longitudinal fasciculusN medial vestibular nucleusSt nigrostriatal fibersAm nucleus ambiguusCu cuneate nucleusGr gracile nucleusPp Nucleus prepositusbF olivocerebellar fiberson occipitopontine fiberslNig pallidonigral fibersNu pontobulbar nucleusbF pontocerebellar fibers
principal (inferior) olivary nucleusnNu pontine nucleion parietopontine fibers
pyramidrestiform body
tF reticular formationtSp reticulospinal tractSp rubrospinal tract
superior colliculusP superior cerebellar peduncleP, Dec superior cerebellar peduncle, decussation
stria medullares of fourth ventriclesubstantia nigrasuperior olive
lNu solitary nucleuslTr solitary tractTec spinotectal tractTh spinothalamic tractTNu spinal trigeminal nucleusTT spinal trigeminal tractVN spinal (inferior) vestibular nucleusNu superior salivatory nucleusN superior vestibular nucleuscSp tectospinal tracton temporopontine fiberspB trapezoid bodypNu trapezoid nucleus
MoNu trigeminal motor nucleus (caudal part)Nr trigeminal nerveNr trochlear nerveNu trochlear nucleusNu ventral cochlear nucleussSp vestibulospinal tractCT ventral spinocerebellar tractTr ventral trigeminothalamic tract
s used in
MAMCMeMeMLMLMVNigNuNuNuNuOCOPPaPBPCPOPoPPPyRBReReRuSCSCSCSMSNSOSoSoSpSpSpSpSpSSSVTeTPTraTraTriTriTroTroVCVeVSVT

yp
(e
dtpbo
FHs(
FCdS
R. Coveñas et al. / Neuroscience 128 (2004) 843–859846
ears; perfused 37 h after death) and W-1 (69 years;erfused 40 h after death).
In all the cases studied, in the three brainstem regionsmedulla oblongata, pons and mesencephalon), the high-st density of ir structures was generally observed in their
ig. 2. Distribution of Met8-ir fibers and cell bodies in frontal planes oaines (1987). Cell bodies containing the neuropeptide are representequares (low density). Ir fibers are represented by dotted lines (singlehigh density). See list of abbreviations for nomenclature.
ig. 3. Distribution of Met8-ir fibers and cell bodies in frontal planes oell bodies containing the neuropeptide are represented by closed cirensity). Ir fibers are represented by dotted lines (single fibers or low
ee list of abbreviations for nomenclature.orsal parts. In addition, the clusters of cell bodies con-aining Met8 were almost always observed in the dorsalart of the human brainstem. The highest density of ir cellodies was found in the reticular formation of the medullablongata.
an medulla oblongata, according to the atlas of the human brain ofed circles (high density), closed triangles (moderate density) or closed
low density), continuous lines (moderate density) and crossed lines
an pons, according to the atlas of the human brain of Haines (1987).density), closed triangles (moderate density) or closed squares (low
continuous lines (moderate density) and crossed lines (high density).
f the humd by closfibers or
f the humcles (highdensity),

Dm
wcdvp88w1tt
MdFrrsdDcn
otnft(t(B2b
ndpIi((Hfir
aM
FCbn
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 847
istribution of Met8-ir structures in the humanedulla oblongata
Met8-ir cell bodies. A high density of ir cell bodiesas found along the reticular formation, except in itsaudalmost part (Figs. 1B, 2A, B, 7A, B). A moderateensity was visualized in the caudal part of the medialestibular nucleus (Figs. 2A, 7E, F) and in the rostralarts of the spinal (inferior) vestibular nucleus (Figs. 2B,A, B) and the medial longitudinal fasciculus (Figs. 2B,C, D), while a low density of cell bodies containing Met8
as observed in the solitary nucleus (caudal part) (Fig.B), the rostral part of the hypoglossal nucleus (Fig. 2A),he nucleus prepositus (Fig. 2B) and the rostral part ofhe lateral cuneate nucleus (Fig. 2A).
Met8-ir fibers. A high density of ir fibers containinget8 was observed in the central gray (Fig. 1A, B), theorsal motor nucleus of the vagus (Figs. 1A, B, 2A, 9D,), the solitary nucleus (Figs. 1B, 2A, B, 3A, 9A, B), theeticular formation (Figs. 1B, 2A, B, 7B, 9A, C), theaphe obscurus (Fig. 10A, B) and in the dorsal acces-ory olivary nucleus (Figs. 2A, B, 8E, F). A moderateensity was found in the gracile nucleus (Fig. 1B, 10C,), the lateral cuneate nucleus (Figs. 1B, 2A), the nu-leus ambiguus (Figs. 1B, 2A, B), the lateral reticular
ig. 4. Distribution of Met8-ir fibers and cell bodies in frontal planes oell bodies containing the neuropeptide are represented by closed circy dotted lines (single fibers or low density), continuous lines (modomenclature.
ucleus (Figs. 1B, 2A, 10F, 11A), the medial accessory r
livary nucleus (Figs. 2A, B, 11B), the nucleus preposi-us (Fig. 2B) and in the dorsal and ventral cochlearuclei (Figs. 2B, 11C–F). A low density of ir fibers was
ound in the rostral midline (Fig. 2B), the medial longi-udinal fasciculus (Figs. 1B, 2B), the tectospinal tractFigs. 1B, 2B), the lateral vestibular nucleus, the solitaryract (Figs. 2A, B, 3A, 9A, B), the medial lemniscusFigs. 1B, 2A, B), the anterolateral system (Figs. 1B, 2A,, 10F, 11A) and in the spinal trigeminal tract (Figs. 1B,A, B). Finally, single fibers were found in the restiformody (Figs. 1B, 2A, B) and the red nucleus.
Moreover, the gelatinosa part of the spinal trigeminalucleus (pars caudalis; Figs. 1A, 12C, D) showed a highensity of ir fibers, but a moderate density was found inars interpolaris and pars oralis (Figs. 1B, 2A, B, 12A, B).n addition, a moderate density of Met8-ir fibers was foundn the magnocellular part of the spinal trigeminal nucleuspars caudalis; Fig. 1A). In the spinal vestibular nucleusFigs. 2A, B, 8A, B), we observed a low density of ir fibers.owever, in its ventral region, we found a high density of irbers at caudal level (Fig. 2A) and a moderate density atostral level (Fig. 2B).
In the hypoglossal nucleus (Figs. 1A, B, 2A, 9D, E),t caudal level a moderate density of fibers containinget8 was visualized (Fig. 1A, B) and a low density at
an pons, according to the atlas of the human brain of Haines (1987).density), closed triangles (moderate density). Ir fibers are representedsity) and crossed lines (high density). See list of abbreviations for
f the humles (higherate den
ostral level (Fig. 2A). In the cuneate nucleus (Fig. 1A, B,
F(o
F(o
R. Coveñas et al. / Neuroscience 128 (2004) 843–859848
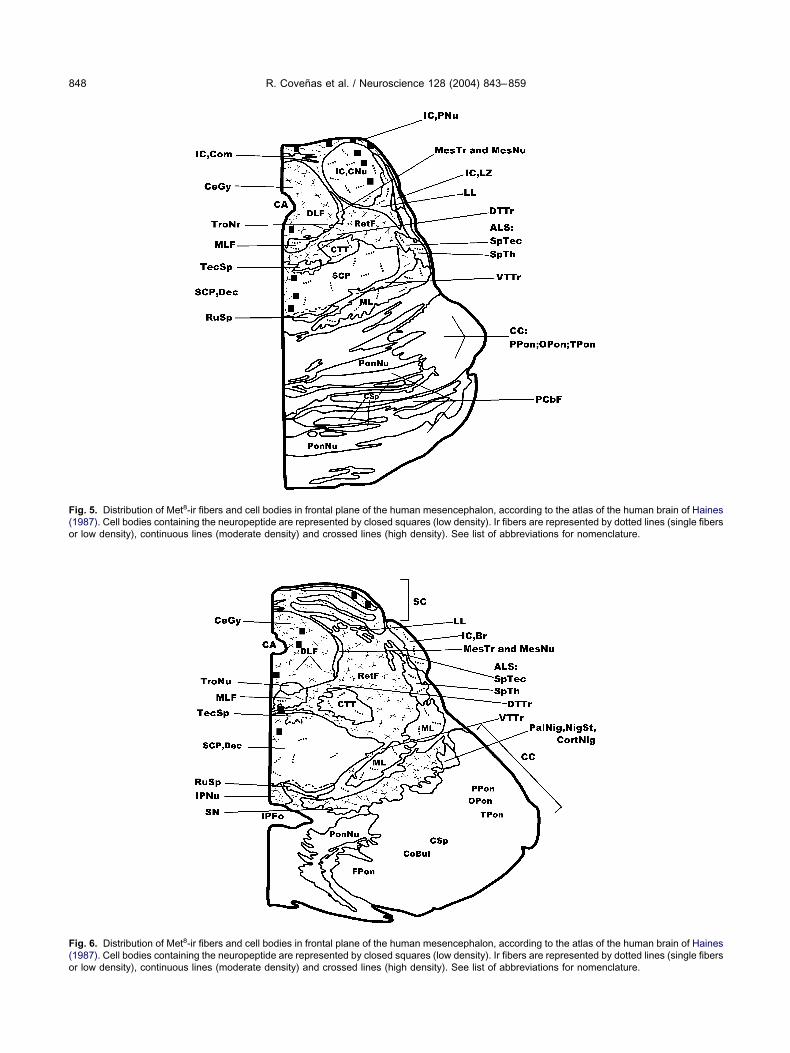
ig. 6. Distribution of Met8-ir fibers and cell bodies in frontal plane of the human mesencephalon, according to the atlas of the human brain of Haines1987). Cell bodies containing the neuropeptide are represented by closed squares (low density). Ir fibers are represented by dotted lines (single fibers
ig. 5. Distribution of Met8-ir fibers and cell bodies in frontal plane of the human mesencephalon, according to the atlas of the human brain of Haines1987). Cell bodies containing the neuropeptide are represented by closed squares (low density). Ir fibers are represented by dotted lines (single fibersr low density), continuous lines (moderate density) and crossed lines (high density). See list of abbreviations for nomenclature.
r low density), continuous lines (moderate density) and crossed lines (high density). See list of abbreviations for nomenclature.

1atsp
D
w37
wcudto
i(
FMboMbdC
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 849
0C, E) a low density was found at caudal level (Fig. 1A)nd a moderate density at rostral level (Fig. 1B). Finally,
he medial vestibular nucleus (Figs. 2A, B, 7E, F)howed a moderate density, but its anteroventrolateralart showed a low density of ir fibers (Fig. 2B).
istribution of Met8-ir structures in the human pons
Met8-ir cell bodies. A high density of ir cell bodiesas caudally located ventral to the superior olive (Figs.A, 12E, F), in the reticular formation (laterally; Figs. 3B,
ig. 7. Met8-ir cell bodies and fibers in the human medulla oblongata aL. Thick dashed lines indicate the midline. This photograph was takear�500 �m. (B) High-power image of the area delimited in Fig. 7Ablongata. Scale bar�100 �m. (C) Low-power image showing the CeG, medial. Scale bar�500 �m. (D) High-power image of the area dar�100 �m. (E) Low-power image of the region around the SolNu aelimited into the MVN is shown at higher magnification in Fig. 7F. Phluster of ir cell bodies in the MVN. Scale bar�100 �m.
C, D, 13A, B) and in the central gray (rostral; Fig. 4), 4
hereas a moderate density was found in the midline (ataudal level; Figs. 3A, 13C–F) and rostrally in the retic-lar formation located close to the midline (Fig. 4). A lowensity was visualized in the caudal midline (Fig. 3A, B),
he central gray (Figs. 3 B, 14A), and in the rostral partf the medial longitudinal fasciculus (Fig. 3A).
Met8-ir fibers. A high density of ir fibers was observedn the nucleus prepositus (Fig. 3A), the reticular formationFigs. 3A, B, 4, 7C, D, 13A, B) and the central gray (Figs. 3B,
. (A) A low magnification of the RetF of the medulla oblongata and therain M-1 (see Experimental Procedures). D, dorsal; M, medial. Scaleof ir perikarya and Met8-ir fibers located in the RetF of the medullaRetF of the pons (RetF). Photograph taken from brain W-1. D, dorsal;
in Fig. 7C. Ir cell bodies and fibers in the RetF of the pons. ScaleFor a nomenclature of the nuclei, see list of abbreviations. The areataken from brain M-1. D, dorsal; M, medial. Scale bar�500 �m. (F)
nd ponsn from b
. Clustery and theelimitednd SolTr.otograph
, 14A). A low density was seen in the abducens nucleus
(t4(mp
Dm
t
nca
sratw
FoPAs8aW
R. Coveñas et al. / Neuroscience 128 (2004) 843–859850
Fig. 3A), the facial nucleus (Fig. 3A), the spinal trigeminalract (Fig. 3A), the medial longitudinal fasciculus (Figs. 3A, B,), the tectospinal tract (Fig. 3A), the central tegmental tractFigs. 3A, B, 4), the anterolateral system (Figs. 3A, B, 4), theedial lemniscus (Figs. 3A, B, 4) and the superior cerebellareduncle (Fig. 4).
istribution of Met8-ir structures in the humanesencephalon
Met8-ir cell bodies. A low density of cell bodies con-8
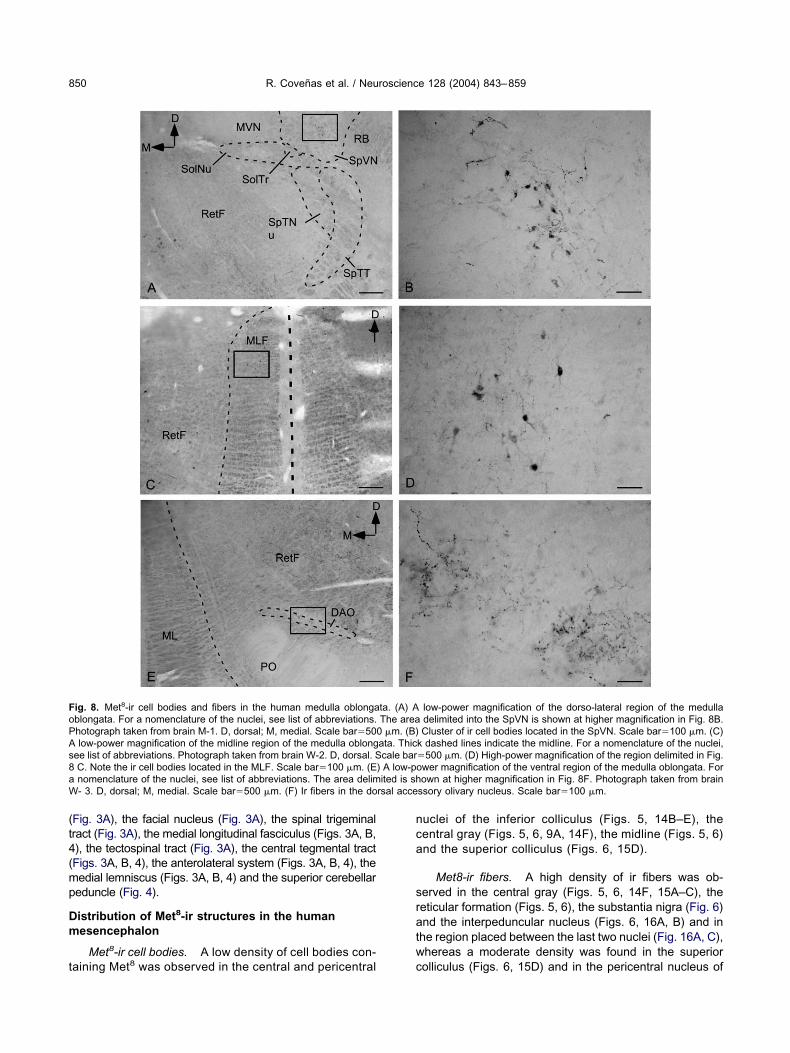
ig. 8. Met8-ir cell bodies and fibers in the human medulla oblongablongata. For a nomenclature of the nuclei, see list of abbreviations.hotograph taken from brain M-1. D, dorsal; M, medial. Scale bar�50low-power magnification of the midline region of the medulla oblong
ee list of abbreviations. Photograph taken from brain W-2. D, dorsal. SC. Note the ir cell bodies located in the MLF. Scale bar�100 �m. (Enomenclature of the nuclei, see list of abbreviations. The area delim- 3. D, dorsal; M, medial. Scale bar�500 �m. (F) Ir fibers in the do
aining Met was observed in the central and pericentral c
uclei of the inferior colliculus (Figs. 5, 14B–E), theentral gray (Figs. 5, 6, 9A, 14F), the midline (Figs. 5, 6)nd the superior colliculus (Figs. 6, 15D).
Met8-ir fibers. A high density of ir fibers was ob-erved in the central gray (Figs. 5, 6, 14F, 15A–C), theeticular formation (Figs. 5, 6), the substantia nigra (Fig. 6)nd the interpeduncular nucleus (Figs. 6, 16A, B) and inhe region placed between the last two nuclei (Fig. 16A, C),hereas a moderate density was found in the superior
low-power magnification of the dorso-lateral region of the medulladelimited into the SpVN is shown at higher magnification in Fig. 8B.Cluster of ir cell bodies located in the SpVN. Scale bar�100 �m. (C)
k dashed lines indicate the midline. For a nomenclature of the nuclei,�500 �m. (D) High-power magnification of the region delimited in Fig.ower magnification of the ventral region of the medulla oblongata. Forown at higher magnification in Fig. 8F. Photograph taken from brain
ssory olivary nucleus. Scale bar�100 �m.
ta. (A) AThe area
0 �m. (B)ata. Thiccale bar
) A low-pited is sh
rsal acce
olliculus (Figs. 6, 15D) and in the pericentral nucleus of

tMifattis(
Cb
TMw
o
FtdmdSS
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 851
he inferior colliculus (Figs. 5, 14B–D). A low density ofet8-ir fibers was located in the central nucleus of the
nferior colliculus (Figs. 5, 14E), the medial longitudinalasciculus (Figs. 5, 6), the lateral lemniscus (Fig. 5), thenterolateral system (Figs. 5, 6), the central tegmentalract (Figs. 5, 6), the superior cerebellar peduncle (Fig. 5),he medial lemniscus (Figs. 5, 6) and the brachium of thenferior colliculus (Fig. 6). Finally, single fibers were ob-erved in the oculomotor nucleus and in the red nucleus
ig. 9. Met8-ir fibers in the human medulla oblongata. (A) A low maghe nuclei, see list of abbreviations. Photograph taken from brain W-1elimited in Fig. 9A including the SolNu and the SolTr. Note the ir fibagnification of the region delimited in the corner of Fig. 9A showingorso-medial region of the medulla oblongata. For a nomenclature of thcale bar�500 �m. (E) High-power image of the region delimited in tcale bar�50 �m. (F) High-power magnification of the region delimite
not shown in Figs. 1–6). i
DISCUSSION
omparison of the distribution of Met8 in the humanrainstem
his is the first detailed study showing the distribution ofet8-ir fibers and cell bodies in the human brainstemithout neurological disease.
In comparison with two previous studies carried outn several nuclei (eight in total) of the human brainstem,
of the dorsal region of the medulla oblongata. For a nomenclature ofal; L, lateral. Scale bar�500 �m. (B) High-power image of the areaws) located in the solitary tract. Scale bar: 100 �m. (C) High-powerlaced in the RetF. Scale bar�100 �m. (D) A low magnification of thesee list of abbreviations. Photograph taken from brain M-2. D, dorsal.l rectangle of Fig. 9D. Note the ir fibers containing Met8 in the HyNu.DMNu of Fig. 9D. Ir fibers located in the DMNu. Scale bar�50 �m.
nification. D, dorsers (arro
ir fibers pe nuclei,
he ventrad in the
n which a combination of radioimmunoassay, gel filtra-

tnMrTttsstsf
s(utMt
ti(t
FDRs(dW
R. Coveñas et al. / Neuroscience 128 (2004) 843–859852
ion and high-performance liquid chromatography tech-iques were applied in order to detect the presence ofet8 (Pittius et al., 1984; Iadarola et al., 1991), our
esults are in general in agreement with these studies.hus, using these techniques and immunocytochemistry
he presence of Met8 was detected in the central gray ofhe midbrain, the inferior and superior colliculi, the sub-tantia nigra and in the red nucleus of the human brain-tem. Pittius et al. (1984) found the highest concentra-ions of Met8 in the human periaqueductal gray, sub-tantia nigra and in other non-brainstem nuclei. In the
ig. 10. Met8-ir fibers in the human medulla oblongata.. (A) Ir fibers in, dorsal. Scale bar�100 �m. (B) A higher magnification of the regionegions delimited in the NuGr and NuCu are respectively observed aee list of abbreviations. Photograph taken from brain W-3. D, dorsal. SE) Presence of Met8-ir fibers in the NuCu. Scale bar�50 �m. (F) A loelimited can be observed at higher magnification in Fig. 11A. For a no-2. D, dorsal. L, lateral. Scale bar�500 �m.
ormer two nuclei, we also observed a high density of ir o
tructures containing Met8. By contrast, Pittius et al.,1984) reported the presence of Met8 in the locus cer-leus, the inferior olivary nucleus and in the area pos-
rema of the human brainstem, in which we failed to findet8-ir structures using an immunocytochemical
echnique.These differences could be due to the sensitivity of
he techniques used (radioimmunoassay, gel filtration,mmunocytochemistry, etc.). Moreover, Pittius et al.1984) show a higher concentration in the dorsal than inhe ventral medulla oblongata. This is in agreement with
. Photograph taken from brain M-1. Dashed lines indicate the midline.in Fig. 10A. Scale bar�50 �m. (C) A low magnification of the DMNu.agnification in Figs. 10D and 10E. For a nomenclature of the nuclei,500 �m. (D) Ir fibers located in the gracile nucleus. Scale bar�50 �m.cation of the ventro-lateral part of the medulla oblongata. The regionre of the nuclei, see list of abbreviations. Photograph taken from brain
the RaNudelimitedt higher mcale bar�w magnifimenclatu
ur results, since we also observed a higher density of ir

svvbIts
Cb
Od
odtglcfitM1d
Fctmbt
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 853
tructures containing Met8 in the dorsal than in theentral medulla oblongata. In sum, our results show aery more widespread distribution of Met8 in the humanrainstem than previously reported (Pittius et al., 1984;adarola et al., 1991), since this is the first study in whichhe presence of Met8 in a large number of human brain-tem nuclei and tracts has been demonstrated.
omparison of the distribution of Met8 in the primaterainstem
ur results are also in general in agreement with those
ig. 11. Met8-ir fibers in the human medulla oblongata. (A) Ir fibersontaining Met8 in the MAO. Photograph taken from brain W-3. D, dorsaken from brain W-3. D, dorsal; L, lateral. Scale bar�200 �m. (D) Higagnification of the ventro-lateral region of the medulla oblongata. Forrain M-1. D, dorsal; L, lateral. Scale bar�500 �m. (F) A high magnifihe ventral cochlear nucleus. Scale bar�50 �m.
escribed in the monkey brainstem for the distribution m
n the Met8-ir fibers (Ibuki et al., 1989). However, someifferences are apparent. Thus, in the human, but not in
he monkey, Met8-ir fibers have been found in the hypo-lossal nucleus, the nucleus ambiguus, the medial and
ateral vestibular nuclei and in the abducens nucleus. Byontrast, in the monkey but not in the human, Met8-irbers have been observed in the locus ceruleus and inhe trapezoid body. In comparison with the distribution ofet8-ir cell bodies in the monkey brainstem (Ibuki et al.,989), our results for the human brainstem showedifferences. In this sense, whereas in both human and
8
located in the ALS and in the LRNu. Scale bar�100 �m. (B) Fibersdial. Scale bar�100 �m. (C) Ir fibers located in the DCNu. Photographmage of the region delimited in Fig. 11C. Scale bar�50 �m. (E) A lowclature of the nuclei, see list of abbreviations. Photograph taken fromthe region delimited in Fig. 11E (VCNu). Note the ir fibers located in
(arrows)al; M, meh-power ia nomencation of
onkey cell bodies containing Met are found in the

rtcnomslcMmg
ttbbwticaon
FTPS(Wb
R. Coveñas et al. / Neuroscience 128 (2004) 843–859854
eticular formation of the medulla oblongata and pons,he solitary nucleus, the nucleus prepositus and in theentral gray of the pons and midbrain, in other brainstemuclei the presence of Met8-ir cell bodies is found in onlyne of the two species. Thus, in humans but not in theonkey, Met8-ir perikarya are present in the hypoglos-
al nucleus, the lateral cuneate nucleus, the medialongitudinal fasciculus and in the inferior and superiorolliculi. However, in the monkey, but not in humans,et8-ir cell bodies are located in the central gray of theedulla oblongata, the dorsal motor nucleus of the va-
ig. 12. Met8-ir fibers and cell bodies in the human medulla oblongatahe area delimited (SpTNu) can be observed at higher magnificatihotograph taken from brain M-2. D, dorsal; M, medial. Scale bar�50cale bar�50 �m. (C) A low magnification of the SpNu. Photograph tak
D) High-power image of the region delimited in Fig. 11C. Scale bar�1-3. D, dorsal; M, medial. Scale bar�200 �m. (F) High-power mag
ar�100 �m.
us, the spinal trigeminal nucleus, the reticular forma- a
ion of the midbrain, the interpeduncular nucleus and inhe trapezoid body. It therefore appears that the distri-ution of cell bodies containing Met8 in the monkeyrainstem shows significant differences in comparisonith the distribution of perikarya containing the octapep-
ide in the human brainstem. The differences observedn both species concerning the distribution of cell bodiesontaining Met8 could be due to technical consider-tions; for example the antisera used, the administrationf colchicine, and/or species differences. It should beoted that such a discrepancy could be due to the
. (A) A low magnification of the lateral region of the medulla oblongata.. 12B. For a nomenclature of the nuclei, see list of abbreviations.) Ir fibers (arrows) in the rostral part of the spinal trigeminal nucleus.rain M-2. D, dorsal; g, gelatinosa part; M, medial. Scale bar�500 �m.) Ir cell bodies located ventral to the SO. Photograph taken from brainof the region delimited in Fig. 12E. Note the ir cell bodies. Scale
and ponson in Fig0 �m. (Ben from b
00 �m. (Enification
dministration of colchicine, since in the monkey the

dasaMtd
Cm
IM
bbMpbmTplnp
FmSHcb
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 855
rug was administered into the ventricle in order toccumulate the neuropeptide in cell bodies of the brain-tem (Ibuki et al., 1989). Finally, in agreement with thebove, this paper shows for the first time the presence ofet8-ir structures in the primate hypoglossal nucleus,
he medial, spinal and lateral vestibular nuclei, the ab-ucens nucleus and in the facial nucleus.
omparison of the distribution of Met8 in theammalian non-primate brainstem
n comparison with the distribution of ir fibers containing8
ig. 13. Met8-ir cell bodies in the human pons. (A) A low magnificationagnification in Fig. 13B. Photograph taken from brain M-1. D, dorsal;cale bar�100 �m. (C) Ir cell bodies located in the midline. Photoigh-power magnification of the region delimited in Fig. 13C. Scale baran be observed at higher magnification in Fig. 13F. Dashed linear�500 �m. (F) Cluster of ir cell bodies located in the midline. Scale
et found in the brainstem of the cat (Cruz and Bas- i
aum 1985; Belda et al., 2003), our results in the humanrainstem point to a more widespread distribution ofet8-ir fibers. On comparing the distribution of Met8-irerikarya in the brainstem, it seems that in the humanrainstem the distribution of Met8-ir cell bodies is slightlyore widespread than in the cat (Belda et al., 2003).hus, for example, in humans, but not in the cat, Met8-irerikarya were found in the hypoglossal nucleus, the
ateral cuneate nucleus, the medial and spinal vestibularuclei, the reticular formation, the central gray of theons, and in the pericentral and central nuclei of the
rsal region of the pons. The area delimited can be observed at higherl. Scale bar�200 �m. (B) Cluster of ir cell bodies located in the RetF.en from brain W-2. D, dorsal; M, medial. Scale bar�500 �m. (D). (E) A low magnification of the midline of the pons. The area delimited
the midline. Photograph taken from brain M-2. D, dorsal. Scale0 �m.
of the doM, mediagraph tak�100 �ms indicate
bar�10
nferior colliculus. In both species, the methodology and

ttvoabggdc
sLPoohct
FdotFbmM
R. Coveñas et al. / Neuroscience 128 (2004) 843–859856
he primary antiserum used was the same, except that inhe feline colchicine was administered into the fourthentricle (Belda et al., 2003). Despite the administrationf the drug in the cat, as mentioned, Met8-ir cell bodiesre slightly more widespread distributed in the humanrainstem than in the same region of the cat. This sug-ests that the discrepancies found in both species re-arding the distribution of Met8-ir perikarya would beue to species differences and not to methodological
ig. 14. Met8-ir cell bodies and fibers in the human pons and meseorsal; M, medial. Photograph taken from brain W-2. Scale bar�10f the nuclei, see list of abbreviations. Photograph taken from braihe region delimited in Fig. 14B. The areas delimited in the dorsaligs. 14D and 14E. Scale bar�200 �m. (D) Ir cell bodies (arroar�50 �m. (E) Cell bodies (arrows) in the central nucleus ofesencephalic central gray. The area delimited is observed at high, medial. Scale bar�500 �m.
onsiderations. e
Finally, on comparing the distribution of Met8-irtructures in the human and rat brainstems (Fallon andeslie 1986; Harlan et al., 1987; Murakami et al., 1987;alkovits 1988), it seems that in general the distributionf Met8-ir fibers is quite similar. However, the distributionf Met8-ir perikarya is more widespread in the rat than inumans. This difference could also be due to technicalonsiderations, since in the rat colchicine was adminis-ered in the ventricle (Fallon and Leslie 1986; Murakami
n. (A) Ir fibers and cell bodies in the pons central gray (CeGy). D,) A low magnification of the inferior colliculus. For a nomenclature, dorsal; M, medial. Scale bar�500 �m. (C) High-power image ofral rectangles are respectively observed at higher magnification infibers in the pericentral nucleus of the inferior colliculus. Scale
ior colliculus. Scale bar�50 �m. (F) A low magnification of theification in Fig. 15A. Photograph taken from brain W-3. D, dorsal;
ncephalo0 �m. (Bn W-1. Dand ventws) andthe inferer magn
t al., 1987), and in this rodent not only immunocyto-

c1iabc
P
Rcmemttdibo
Ph
Thinp
star1osbaha
dasnMvdaCst
cs
Fb(c le bar�5
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 857
hemical techniques were applied (Fallon and Leslie986; Murakami et al., 1987; Palkovits 1988) but also an
n situ hybridization (Harlan et al., 1987) technique waspplied in the brainstem. By contrast, here in the humanrainstem only immunocytochemical techniques werearried out.
ossible Met8-containing pathways in human
egarding the results found here, we have no data indi-ating whether the Met8-ir perikarya observed in the hu-an brainstem are local and/or projecting neurons. How-ver, it can be suggested, for example, that the dorsalotor nucleus of the vagus, the spinal trigeminal nucleus,
he dorsal accessory olivary nucleus, the reticular forma-ion of the midbrain, the substantia nigra and the interpe-uncular nucleus would receive Met8-ir afferences, since
n these nuclei a high density of ir fibers containing Met8,ut no cell bodies containing the neuropeptide, werebserved.
ossible physiological functions of Met8 in theuman brainstem
he widespread distribution of Met8-ir structures in theuman brainstem suggests that this peptide might be
nvolved in several physiological actions, acting as aeurotransmitter and/or neuromodulator. Some of these
ig. 15. Met8-ir cell bodies and fibers in the human mesencephalonar�100 �m. (B) Low-power image of the mesencephalic central graC) High-power image of the region delimited in Fig. 15B. Note the iolliculus. Photograph taken from brain M-1. D, dorsal; L, lateral. Sca
hysiological actions are well documented. For in- f
tance, there is substantial evidence that opioid pep-ides in brainstem nuclei, including the solitary nucleus,re involved in respiratory, cardiovascular, and gastricegulation (Palkovits and Eskay, 1987; Cheng et al.,996; Yuan and Foss, 1999), and also on the modulationf taste responses (Cheng-Shu et al., 2003). In thisense, it has been described that the respiratory distur-ances observed in Rett syndrome are associated withdecreased expression of Met8 in the parabraquial,
ypoglossal, dorsal vagal and solitary nuclei (Saito etl., 2001).
Furthermore, an analgesic role for Met8 has beenescribed in the spinal trigeminal nucleus (Langemark etl., 1995), and also in patients suffering infantilepasms, Met8 was almost completely vanished in thisucleus (Hayashi et al., 2000). Finally, the presence ofet8 in the substantia nigra suggests its possible in-
olvement on motor function. Moreover, it has beenemonstrated a loss of Met8 in this area in Parkinson’snd Huntington’s diseases (Iadarola et al., 1991; deeballos and López-Lozano, 1999). Thus, the data pre-ented in this paper support the involvement of Met8 inhese physiological functions in humans
We hope that our study on the distribution of fibers andell bodies containing Met8 in the human brainstem willerve to compare the distribution of Met8-ir structures
l body (arrow) and fibers containing Met8 in the central gray. Scaleraph taken from brain W-. D, dorsal; M, medial. Scale bar�200 �m.cale bar�50 �m. (D) Ir cell body (arrow) and fibers in the superior
0 �m.
. (A) Cely. Photogr fibers. S
ound in normal human brainstems with those observed in

bAiiw
ps
Fmedt(g
FtchbafiabS
R. Coveñas et al. / Neuroscience 128 (2004) 843–859858
rainstems showing a given pathology (e.g. Parkinson orlzheimer disease), as well as for studying the physiolog-
cal actions of Met8 in the human brainstem. In this sense,t has been demonstrated that the concentration of Met8
ig. 16. Met8-ir fibers in the human mesencephalon. (A) Low-poweragnification of the ventro-medial mesencephalon. Observe the pres-nce of ir fibers in the IPNu. Photograph taken from brain M-2. D,orsal; M, medial. Scale bar�200 �m. (B) High-power magnification ofhe area delimited in the left rectangle of Fig. 16A. Scale bar�50 �m.C) High-power magnification of the area delimited in the right rectan-le of Fig. 16A. Scale bar�50 �m.
as significantly elevated in the substantia nigra of schizo- 1
hrenic patients in comparison with those found in theame brainstem region of normal patients (Iadarola et al.,
ig. 17. Specificity of the Met8 antiserum. (A) Met8-ir fibers (arrows) inhe dorsal medulla oblongata in a section to which the immunocyto-hemistry method was applied. The delimited area is observed atigher magnification in Fig. 17C. D, dorsal; M, medial. Scalear�200 �m. (B) No immunoreactivity appeared in the same region insection adjacent to that shown in Fig. 17A when preabsorption of the
rst antiserum (anti-Met8) with synthetic Met8 was carried out prior topplying the immunocytochemical technique. Arrowheads indicate alood vessel. Scale bar�200 �m. (C) Ir fibers in the midline region.cale bar�100 �m.
991).

AD(
A
A
B
B
C
C
C
d
D
F
G
H
H
H
I
I
L
M
M
M
M
P
P
P
P
P
S
T
Y
Z
R. Coveñas et al. / Neuroscience 128 (2004) 843–859 859
cknowledgments—This work has been supported by the.G.I.C.Y.T. (PB99/0160) and Ministerio de Ciencia y Tecnología
BFI2001-1905), Spain.
REFERENCES
be J, Okamura H, Makino S, Yanaihara N, Ibata Y (1987) Theimmunocytochemical distribution of met-enkephalin-Arg6-Gly7-Leu8 immunoreactivity in the rat diencephalon. Brain Res Bull19:735–742.
kill H, Watson SJ, Young E, Lewis ME, Khachaturian H, Walker JM(1984) Endogenous opioids: biology and function. Annu Rev Neu-rosci 7:223–255.
asbaum AI, Fields HL (1984) Endogenous pain control systems:brainstem spinal pathways and endorphin circuitry. Annu Rev Neu-rosci 7:309–338.
elda M, Coveñas R, Narváez JA, Aguirre JA, Gonzalez-Barón S(2003) An immunocytochemical mapping of methionine-enkepha-lin-Arg6-Gly7-Leu8 in the cat brainstem. Anat Embryol206:399–408.
heng PY, Liu-Chen LY, Chen C, Pickel VM (1996) Immunolabellingof mu opioid receptors in the rat nucleus of the solitary tract:extrasynaptic plasmalemmal localization and association with leu5-enkephalin. J Comp Neurol 371:522–536.
heng-Shu L, Davis BJ, Smith DV (2003) Opioid modulation of tasteresponses in the nucleus of the solitary tract. Brain Res 965:21–34.
ruz L, Basbaum AI (1985) Multiple opioid peptides and the modula-tion of pain: immunohistochemical analysis of dynorphin and en-kephalin in the trigeminal nucleus caudalis and spinal cord of thecat. J Comp Neurol 240:331–348.
e Ceballos ML, López-Lozano JJ (1999) Subgroups of parkinsonianpatients differentiated by peptidergic immunostaining of caudatenucleus biopsies. Peptides 20:249–257.
ouglas H, Kitchan I (1992) Mechanisms involved in the cardiovascu-lar responses to opioid products of proenkephalin in the anaesthe-tised rat. Gen Pharmacol 23:269–277.
allon JH, Leslie FM (1986) Distribution of dynorphin and enkephalinpeptides in the rat brain. J Comp Neurol 249:293–336.
untern R, Vellet PG, Bouras C, Constantinidis J (1989) An improvedimmunohistostaining procedure for peptides in human brain. Ex-perientia 45:159–161.
aines DE (1987) Neuroanatomy: an atlas of structures, sections andsystems. Baltimore: Urban & Schwarzenberg.
arlan RE, Shivers BD, Romano GJ, Howells RD, Pfaff DW (1987)Localization of preproenkephalin mRNA in the rat brain and spinalcord by in situ hybridization. J Comp Neurol 258:159–184.
ayashi M, Itoh M, Araki S, Kumada S, Tanuma N, Kohji T, KohyamaJ, Iwakawa Y, Satoh J, Morimatsu Y (2000) Immunohistochemicalanalysis of brainstem lesions in infantile spasms. Neuropathology20:297–303.
adarola MJ, Ofri D, Kleinman JE (1991) Enkephalin, dynorphin andsubstance P in postmortem substantia nigra from normal and
schizophrenic patients. Life Sci 48:1919–1930.buki T, Okamura H, Miyazaki M, Yanaihara N, Zimmerman EA, IbataY (1989) Comparative distribution of three opioid systems in thelower brainstem of the monkey (Maccaca fuscata). J Comp Neurol279:445–456.
angemark M, Bach FW, Ekman R, Olessen J (1995) Increasedcerebrospinal fluid Met-enkephalin immunoreactivity in patienswith chronic tension-type headache. Pain 63:103–107.
arcos P, Coveñas R, Narváez JA, Aguirre JA, Tramu G, González-Barón S (1998) Neuropeptides in the cat amygdala. Brain Res Bull45:261–268.
arcos P, Coveñas R, Narváez JA, Díaz-Cabiale Z, Aguirre JA, TramuG, González-Barón S (1999) Immunohistochemical mapping ofenkephalins, NPY, CGRP and GRP in the cat amygdala. Peptides20:635–644.
urakami S, Okamura H, Pelletier G, Ibata Y (1989) Differentialcolocalization of neuropeptide Y and methionine-enkephalin-Arg6-Gly7-Leu8-like immunoreactivity in catecholaminergic neu-rons in the rat brain stem. J Comp Neurol 281:532–544.
urakami S, Okamura H, Yanaihara C, Yanaihara N, Ibata Y (1987)Immunocytochemical distribution of met-enkephalin-Arg6-Gly7-Leu8 in the rat lower brainstem. J Comp Neurol 261:193–208.
alkovits M (1988) Neuropeptides in the brain. In: Frontiers in neu-roendocrinology, Vol. 10 (Martini L, Ganong WF, eds), pp 1–44.New York: Raven Press.
alkovits M, Eskay RL (1987) Distribution and possible origin of beta-endorphin and ACTH in discrete brainstem nuclei of the rats.Neuropeptides 9:123–137.
asternak GW (1987) Opiod receptors. In: Psychopharmacology: thethird generation of progress (Meltzer Y, eds), pp 281–288. NewYork: Raven Press.
axinos G, Törk I, Halliday G, Mehler WR (1990) Human homologs tobrainstem nuclei identified in other animals as revealed by acetyl-cholinesterase activity. In: The human nervous system (Paxinos G,eds), pp 149–202. San Diego: Academic Press.
ittius CW, Seizinger BR, Pasi A, Mehraein P, Herz A (1984) Distri-bution and characterization of opioid peptides derived from proen-kephalin A in human and rat central nervous system. Brain Res304:127–136.
aito Y, Ito M, Ozawa Y, Matsuishi T, Hamano K, Takashima S(2001) Reduced expression of neuropeptides can be related torespiratory disturbances in Rett syndrome. Brain Dev23:S122–S126.
anaka M, Okamura H, Yanaihara N, Tanaka Y, Ibata Y (1993)Differential expression of serotonin and methionine-enkephalin-Arg6-Gly7-Leu8 in neurons of the rat brain stem. Brain Res Bull30:561–570.
uan CS, Foss JF (1999) Gastric effects of methylnaltrexone on mu,kappa, and delta opioid agonists induced brainstem unitary re-sponses. Neuropharmacology 38:425–432.
amir N, Palkovits M, Brownstein M (1985) Distribution of immunore-active met-enkephalin-Arg6-Gly7-Leu8 and leu-enkephalin in dis-
crete region of the rat brain. Brain Res 326:1–8.(Accepted 12 July 2004)



![Dietary supplementation with free methionine or methionine … · 2019. 6. 27. · with MHA or DL-methionine in heat stress-exposed broilers [23, 24]. In this study, we hypothesize](https://static.fdocuments.net/doc/165x107/60e337666b3f9a31a45a96d1/dietary-supplementation-with-free-methionine-or-methionine-2019-6-27-with-mha.jpg)














