
AGING AS A PROCESS OF COMPLEXITY LOSS · advances in the fields of nonlinear dynamics, chaos...
24
1 AGING AS A PROCESS OF COMPLEXITY LOSS Lewis A. Lipsitz, MD In: Complex Systems Science in Biomedicine Deisboeck, Kresh, and Kepler, editors. Kluwer Academic Publishers, 2003 From: The Hebrew Rehabilitation Center for Aged, Beth Israel Deaconess Medical Center, and Harvard Medical School, Boston MA Address correspondence to: Lewis A. Lipsitz, MD Hebrew Rehabilitation Center for Aged 1200 Centre Street, Boston, MA 02131 Phone: 617-363-8318, Fax: 617-363-8929 E-mail: [email protected] Supported by Grants AG04390 and AG08812 from the National Institute on Aging, Bethesda, MD. Dr. Lipsitz holds the Irving and Edyth S. Usen and Family Chair in Geriatric Medicine at the Hebrew Rehabilitation Center for Aged.
Transcript of AGING AS A PROCESS OF COMPLEXITY LOSS · advances in the fields of nonlinear dynamics, chaos...

1
AGING AS A PROCESS OF COMPLEXITY LOSS
Lewis A. Lipsitz, MD
In: Complex Systems Science in Biomedicine
Deisboeck, Kresh, and Kepler, editors.
Kluwer Academic Publishers, 2003
From: The Hebrew Rehabilitation Center for Aged,
Beth Israel Deaconess Medical Center,
and Harvard Medical School, Boston MA
Address correspondence to:
Lewis A. Lipsitz, MD
Hebrew Rehabilitation Center for Aged
1200 Centre Street, Boston, MA 02131
Phone: 617-363-8318, Fax: 617-363-8929
E-mail: [email protected]
Supported by Grants AG04390 and AG08812 from the National Institute on Aging, Bethesda,
MD. Dr. Lipsitz holds the Irving and Edyth S. Usen and Family Chair in Geriatric Medicine at
the Hebrew Rehabilitation Center for Aged.

2
INTRODUCTION
Classical research in the field of aging has been largely reductionistic, defining the
process of aging as a linear decline in many organs and physiologic systems until functional
disability results (Figure 1) (1). However, the process of aging is a nonlinear, multi-dimensional
process that is associated not only with changes in individual systems, but probably more
importantly, alterations in the connections and interactions between system components. The
complex mechanisms by which these components interact to enable an organism to perform a
variety of functions necessary for survival, is the subject of physiology.
Healthy physiologic processes require the integration of complex networks of control
systems and feedback loops that operate on multiple scales in space and time (2). For example,
physiologic systems exist at molecular, subcellular, cellular, organ, and systemic levels of
organization. Continuous interplay between the electrical, chemical, and mechanical components
of these systems ensures that information is constantly exchanged, even as the organism rests.
These dynamic processes give rise to a highly adaptive, resilient organism that is prepared to
respond to internal and external perturbations.
Recognition of the dynamic nature of regulatory processes challenges the concept of
homeostasis, which is taught by physiologists as a function of all healthy cells, tissues, and
organs to maintain static or steady-state conditions in their internal environment (3). However,
with the introduction of techniques that can acquire continuous data from physiologic processes
such as heart rate, blood pressure, nerve activity, or hormonal secretion, it became apparent that
these systems are in constant flux, even under so-called steady-state conditions. Dr. Eugene
Yates introduced the term homeodynamics to convey the fact that the high level of bodily control

3
required to survive depends on a dynamic interplay of multiple regulatory mechanisms rather
than constancy in the internal environment (4).
Although it is often difficult to separate the effects of aging from those of disease and
lifestyle changes such as reduced physical activity, healthy aging in the absence of such
confounding factors appears to have a profound impact on physiologic processes. Because of the
progressive age-related degeneration of various tissues and organs, and the interruption of
communication pathways between them, complex physiologic networks break down, become
disconnected, and lose some of their capacity to adapt to stress.
There is considerable redundancy in many of the biologic and physiologic systems in
higher organisms; for example, humans have far more muscle mass, neuronal circuitry, renal
nephrons, and hormonal stores than are needed to survive. This creates a physiologic reserve that
allows most individuals to compensate effectively for age-related changes. Because the network
structure of physiologic systems also enables alternate pathways to be used to achieve the same
functions, physiologic changes that result from aging alone usually do not have much impact on
everyday life. However, these changes may become manifest at times of increased demand,
when the body is subjected to high levels of physiologic stress. For this reason, elderly
individuals are particularly vulnerable to adverse events such as falls, confusion, and
incontinence when exposed to environmental, pharmacologic, or emotional stresses.
MEASURES OF COMPLEXITY LOSS
With the development of monitoring devices that can measure the output of regulatory
processes on a moment-to-moment basis, it has become apparent that the dynamics of many
systems lose complexity with advancing age (5). The continuous heart rate time series of a

4
healthy young subject and healthy elderly subject shown in Figure 2 provides a good example.
The average heart rate over the 8-minute period of recording is 64.7 beats per minute in the
young subject and 64.5 in the older subject; nearly identical. Furthermore, the standard deviation
of the heart rate is also nearly identical; 3.9 in the young subject and 3.8 in the elderly subject.
However, it is apparent that the dynamics of the heart rate time series are strikingly different.
Until recently, scientists have lacked the tools to describe these dynamics. However, new
advances in the fields of nonlinear dynamics, chaos theory, and complex systems have provided
new ways to quantify the aging process and understand its mechanisms. One particularly useful
concept that can be used to quantify the complexity of various anatomic structures or physiologic
processes is the concept of fractals.
The classic definition of a fractal, first described by Mandelbrot (6), is a geometric object
with “self-similarly” over multiple measurement scales. For example, many anatomic structures
demonstrate self-similarity in their structures and have fractal properties. The branches upon
branches upon branches of bronchi in the respiratory tree (7) or smaller and smaller vessels in the
circulatory system (8) look similar whether they are measured in microns, millimeters,
centimeters, or meters. In fact, the smaller the measuring device, the larger the length of a fractal
object. This is a property known as “power law scaling”. The output of dynamic physiologic
processes such as heart rate, which are measured over time rather than space, also have fractal
properties (9). Their oscillations appear self-similar when observed over seconds, minutes, hours,
or days. Furthermore, they demonstrate power-law scaling in the sense that the smaller the
frequency of oscillation of these signals, the larger their amplitude or “power”.
The heart rate time series demonstrates complex irregularity that has been described as a
fractal process because it looks similar whether it is plotted over days, hours, or minutes. In fact,

5
the heart rate time series has “1/f” or power law scaling, in that the amplitude of oscillations (A)
is proportional to the inverse of oscillation frequency, according to the formula: A ≅ 1/fβ. The
exponent β can be derived from the slope of the log – log transformation of the Fourier power
spectrum. A fractal process (most complex) has a slope of 1, while a loss of complexity occurs as
the slope approaches 0 (white noise) or 2 (Brownian noise). The presence of 1/f fractal scaling
indicates that there are long-range (power-law) correlations in the data.
Unfortunately, the computation of the power spectrum using Fourier analysis requires
stationary data, which most physiologic signals are not. Another particularly useful technique
that minimizes the effect of non-stationarities in the data is “detrended fluctuation analysis”
(DFA), which has been well validated in a number of dynamic systems (10). The DFA algorithm
is a two-point correlation method that computes the slope of the line relating the amplitude of
fluctuations to the scale of measurement, after detrending the data. The root-mean-square
fluctuation of the integrated and detrended data are measured in observation windows of
different sizes then plotted against the size of the window on a log-log scale. The slope of the
regression line which relates log-fluctuation to log-window size quantifies the complexity of the
data (1=fractal, 0.5=random, 1.5=random-walk).
Other indicators of complexity loss in physiologic systems include an increase in
periodicity (e.g., the tremor of Parkinson’s disease), increased randomness (e.g., atrial fibrillation
of the heart), and loss of long-range correlations (e.g., gait dynamics).
It is important to recognize that complexity and variability are not the same. For
example, a high amplitude sine wave signal is quite variable, but not at all complex.
Alternatively, an irregular low amplitude signal such as the heart rate of a healthy young subject
shown in Figure 2 can be quite complex but much less variable. Similarly, irregularity and

6
complexity are not the same. Traditional entropy-based algorithms such as Approximate
Entropy (11), that are used to quantify the regularity of a time series, indicate greater irregularity,
which has been interpreted as greater complexity, for certain uncorrelated random signals
associated with pathologic processes. However, as highlighted by the recent work of Costa et.
al. (12), these algorithms fail to account for the multiple time scales over which healthy
physiologic processes operate. By calculating the entropy of heart rate dynamics over multiple
time scales (multiscale entropy analysis), these investigators were able to distinguish healthy
from pathological (e.g., atrial fibrillation and congestive heart failure) dynamics, and showed
consistently higher multiscale entropy values for dynamic processes with long-range (1/f-type)
correlations, compared to those with uncorrelated random noise. Therefore, this measure appears
useful for quantifying the complexity of a time series and distinguishing it from irregularity due
to uncorrelated noise.
EXAMPLES OF COMPLEXITY LOSS WITH AGING
There are numerous examples of the progressive loss of complexity in the fractal
architecture of anatomic structures and the dynamics of physiologic processes with advancing
age. As shown in Table 1, structures such as neurons, bone trabeculae, kidney glomeruli, and
elastic fibers or dermal papillae beneath the skin all lose structural complexity with aging (5).
The loss of connectivity in bone trabeculae, which is characteristic of osteoporosis, is shown in
figure 3. Not only anatomic structures, but also physiologic processes lose complexity with
advancing age. These include heart rate (13), blood pressure (13), and respiratory dynamics (14),
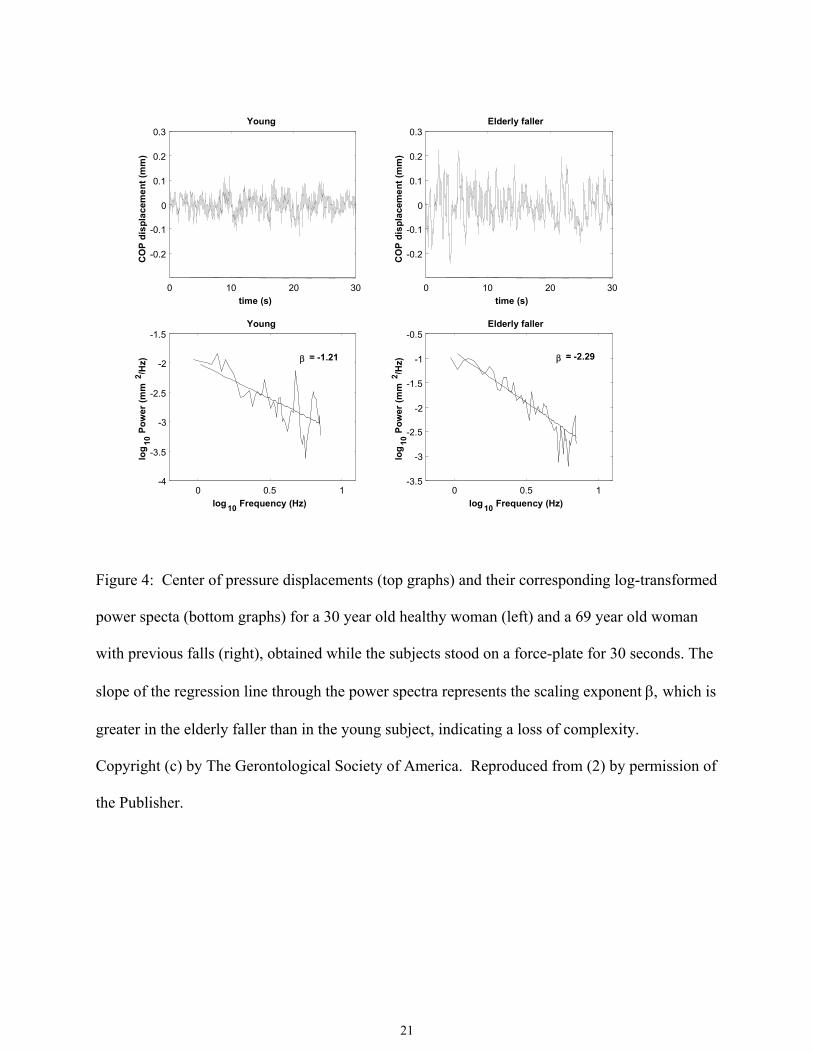
center of pressure trajectories when measured on a balance platform (Figure 4), and gait
dynamics (15)(Table 2). Moreover, complexity loss has been shown to be associated with a

7
variety of diseases and adverse outcomes (Tables 3 and 4). On the basis of these observations,
we have hypothesized that the age-related loss of complexity in physiologic systems, results in
an impaired ability to adapt to stress and the ultimate development of disease and disability (2).
In response to a given stress or perturbation, physiologic systems mount a specific
adaptive response that restores the organism to a new dynamic equilibrium. Therefore, the
dynamics observed during resting and stimulated conditions are often quite different. For
example, when glucose is ingested, insulin is secreted in pulsatile fashion in order to promote
glucose metabolism. When an individual stands up, blood pools in the lower extremities, and
blood pressure suddenly falls. In response, the sympathetic nervous system secretes
norepinephrine to restore the blood pressure to its resting state. Thus, the complex dynamics
observed during resting conditions differ from the more focused singular response that occurs
during stress. The dynamics of this adaptive response have been referred to as “reactive tuning”
(2). In the field of nonequilibrium statistical mechanics, the relationship between the correlation
properties of the fluctuations of a system and its relaxation to equilibrium has be described by the
fluctuation-dissipation theorem (16). If the complex interactions of physiologic systems during
rest enable an organism to mount a focused adaptive response during a perturbation, the loss of
complexity in resting dynamics may indicate an impaired ability to adapt to stress and a
predisposition to functional decline.
MECHANISMS OF PHYSIOLOGIC COMPLEXITY
A variety of mechanisms probably underlie the complexity of physiologic systems,
including neuronal networks in the nervous system, biochemical pathways in metabolic control
systems, signaling pathways within and between cells, genetic switches, and transcription control

8
elements. Two experiments highlight the importance of the autonomic nervous system in
generating the complexity of heart rate dynamics. When autonomic nervous system influences
on the heart are eliminated through the administration of the muscarinic receptor blocker
atropine and the beta-receptor blocker propranalol, the complex dynamics observed under
control conditions are lost (Figure 5).
In another study, baby pigs were shown to develop increasing heart rate complexity as
they matured from 8 to 33 days after birth (17) (Figure 6). During this period of time, the heart
becomes innervated by sympathetic nerves from the right stellate ganglion. When the right
stellate ganglion is denervated at birth, heart rate complexity does not develop. Thus, during
healthy development, complexity appears to emerge in physiologic systems such as heart rate,
and with senescence, system complexity is lost.
LOSS OF COMPLEXITY AS A PATHWAY TO FRAILTY IN OLD AGE
These considerations have led to the theory that the loss of complexity is the physiologic
basis of frailty in old age (2). This is illustrated in Figure 7. During youth, a multitude of
physiologic systems interact to produce a highly complex output signal (such as heart rate) that is
associated with a high level of physical function. With progressive aging, many of the
physiologic inputs and their connectivity are lost, resulting in a less complex output signal. This
loss of complexity is associated with a decline in functional ability. Finally, late in life,
physiologic inputs are diminished to the extent that the output becomes periodic or random, and
the individual crosses a frailty threshold. At this point they lose their ability to adapt to stress and
relatively minor perturbations such as new medications, a viral illness, or emotional trauma may
result in serious disability and death.

9
INTERVENTIONS TO RESTORE COMPLEXITY IN PHYSIOLOGIC SYSTEMS
Fortunately, a number of novel interventions may be able to restore healthy dynamics in
elderly individuals and enhance their ability to adapt to a variety of external stimuli. Some
single interventions that have multi-system effects have already shown to improve functional
ability in older individuals. These include exercise (18), which can improve physical and mental
function, and medications such as beta-blockers, which reverse many of the physiologic
consequences of congestive heart failure. Other multifactorial interventions that address the
multiple systems that are impaired in individuals with syndromes such as falls (19) or delirium
(20) have also proven useful in clinical trials. Recently, Dr. Collins and colleagues have
demonstrated that low levels of mechanical or electrical noise can be used to enhance
somatosensation and thus improve postural control in healthy elderly subjects and patients with
diabetic neuropathy or stroke (21). New dynamic drug delivery systems provide more
physiologic drug administration and thereby enhance physiologic effects. For example, insulin
given in oscillatory fashion has a greater effect on glucose metabolism that when given
continuously (22). Similarly, when parathyroid hormone is give intermittently rather than
continuously, it increases bone mass in osteoporotic patients (23). Finally, external dynamic
control techniques such as pacing procedures to terminate cardiac arrhythmias have proven
useful in cardiac patients (24).
CONCLUSION
In conclusion, aging results in a number of changes in anatomic structures and
physiologic control processes that result in a reduction in system complexity and a loss of ability

10
to adapt to common stresses in the external and internal environment. This loss of complexity
may be the physiologic basis of frailty. New interventions aimed at restoring complex dynamics
may be able to enhance physiologic adaptation and prevent the onset of disease and disability.
REFERENCES
1. Shock NW GR, Andres R, Arenberg D, Costa PT, Lakatta EG, Tobin JD, ed.
Normal Human Aging: The Baltimore Longitudinal Study of Aging. Vol. NIH Publication
No. 84-2450. Baltimore: U.S. Department of Health and Human Services; 1984.
2. Lipsitz L. The dynamics of stability: the physiologic basis of functional health and
frailty. J. Gerontol. Biol. Sciences. 2002;57A(3):B115-B125.
3. Guyton AC. Textbook of Medical Physiology. Eighth ed Philadelphia: W,B. Saunders
Company; 1991.
4. Yates FE BL. Loss of integration and resiliency with age: a dissipative destruction. In:
EJ M, ed. Handbook of physiology. New York: Oxford University Press; 1995.
5. Lipsitz LA GA. Loss of 'complexity' and aging. Potential applications of fractals and
chaos theory to senescence. JAMA. 1992;267(13):1806-1809.
6. Mandelbrot B. The Fractal Geometry of Nature New York, NY: WH Freeman and Co.;
1983.
7. McNamee JE. Fractal perspectives in pulmonary physiology. J. Appl. Physiol.
1991;71(1):1-8.
8. Goldberger AL R, DR, West BJ. Chaos and fractals in human physiology. Scientific
American. 1990;262:43-49.

11
9. Goldberger A. Non-linear dynamics for clinicians: chaos therory, fractals, and
complexity at the bedside. Lancet. 1996;347:1312-1314.
10. Peng C-K HS, Stanley HE, Goldberger AL. Quantification of scaling exponents and
crossover phenomena in nonstationary heart-beat time series. Chaos. 1995;5:82-87.
11. Pincus S, Goldberger AL. Physiological time-series analysis: what does regularity
quantify? Am J Physiol. 1994;266:H1643-H1656.
12. Costa M GA, Peng C-K. Multiscale entropy analysis of complex physiologic time
series. Physical Review Letters. 2002;89(6):068102.
13. Kaplan DT FM, Pincus SM, Ryan SM, Lipsitz LA. Aging and complexity of
cardiovascular dynamics. Biophysical J. 1991;59:945-949.
14. Peng C-K MJ, Liu Y, Lee C, Hausdorff JM, Stanley HE, Goldberger AL, Lipsitz,
LA. Quantifying fractal dynamics of human respiration: age and gender effects. Ann.
Biomed. Eng. 2002;30(5): 683-692.
15. Hausdorff JM MS, Firtion R, et al. Altered fractal dynamics of gait: reduced stride-
interval correlations with aging and Huntington's disease. J Appl Physiol. 1997;82:262-
269.
16. Lauk M CC, Pavlik AE, Collins JJ. Human balance out equilibrium: nonequilibrium
statistical mechanics in posture control. Phys Rev Let. 1998;80(2):413-416.
17. Lipsitz LA PS, Morin RJ, Tong S, Eberle LP, Gootman PM. Preliminary evidence for
the evolution in complexity of heart rate dynamics during autonomic maturation in
neonatal swine. J Auto Nerv Syst. 1997;65:1-9.

12
18. Schuit AJ VAL, Verheij TC,Rijneke RD, Maan AC, Swenne CA, Schouten EG.
Exercise training and heart rate variability in older people. Med Sci Sport Exer.
1999;31(6):816-821.
19. Tinetti ME BD, McAvay MS, Claus EB, Garrett M, Koch ML, Trainor K, Horwitz
RI. A multifactorial intervention to reduce the risk of falling among elderly people living
in the community. NEJM. 1994;331(13):821-827.
20. Inouye SK BS, Charpentier PA, Leo-Summers L, Acampora D, Holford TR, Cooney
LM. A multicomponent intervention to prevent delirium in hospitalized older patients.
NEJM. 1999;340(9):669-676.
21. Liu W CJ, Montero-Odasso M, Bean J, Kerrigan C, Lipsitz LA. Noise-enhanced
vibrotactile sensitivity in older adults, patients with stroke, and patients with diabetic
neuropathy. Arch. Phys. Med. Rehab. 2001;83:171-176.
22. Sturis J SA, Leproult R, Plonsky KS, Van Cauter E. 24-hour glucose profiles during
continuous or oscillatory insulin infusion. Demonstration of the functional significance
of ultradian insulin oscillations. J Clin Invest. 1995;95:1464-1471.
23. Neer RM AC, Zanchetta JR, Prince R, Gaich GA, Reginster JY, Hodsman AB,
Eriksen EF, Ish-Shalom S, Genant HK, Wang O, Mitlak BH. Effect of parathyroid
hormone (1-34) on fractures and bone mineral density in postmenopausal women in
osteoporosis. New Eng J Med. 2001;344(10):1434-1441.
24. Christini DJ SK, Markowitz SM, et al. Nonlinear-dynamical arrhythmia control in
humans. Proc Natl Acad Sci USA. 2001;98:5827-5832.

13
25. Jelles B vB, Slaets JPJ, Hekster REM, Jonkman EJ, Stam CJ. Decrease on non-linear
structure in the EEG of Alzheimer patients compared to healthy controls. Clin
Neurophysiol. 1999;110:1159-1167.
26. Mishma M H, Itoh H, Nakano Y, Sakai H, Nishimura K, Oku Y, Chin K, Ohi M,
Nakamura T, Bates JHT, Alencar AM, Suki B. Complexity of terminal airspace
geometry assessed by lung computed tomography in normal subjects and patients with
chronic obstructive pulmonary disease. Proc Natl Acad Sci. 1999;96:8829-8834.
27. Makikallio TH KJ, Jordaens L, Tulppo MP, Wood N, Golosarsky B, Peng C-K,
goldberger AL, Huikuri HV. Heart rate dynamics before spontaneous onset of
ventricular fibrillation in patients with healed myocardial infarcts. Heart rate
dynamics before spontaneous onset of ventricular fibrillation in patients with healed
myocrdial infarcts. Am J Cardiol. 1999;83:880-884.
28. Vikman S MT, Yl-Mayry S, Pikkujamsa S, Koivisto A-M, Reinkainen P, Airaksinen
KEJ, Huikuri HV. Altered complexity and correlation properties of R-R interval
dynamic before the spontaneous onset of paroxysmal atrial fibrillation. Circulation.
1999;100:2079-2084.
29. Huikuri HV MT, Peng C-K, Goldberger AL, Hintze U, Moller M. Fractal correlation
properties of R-R interval dynamics and mortality in patients with depressed left
ventricular function after an acute myocardial infarction. Circulation. 2000;101:47-53.
30. Velanovich V. Fractal analysis of mammographic lesions: a feasibility study quantifying
the difference between benigh and malignant masses. Am J Med Sci. 1996;311:211-214.

14
31. Ho KKL MG, Peng C-K, Mietus JE, LarsonMG, Levy D, Goldberg AL. Predicting
survival in heart failure case and control subjects by use of fully automated method.
Circulation. 1997;96:842-848.
32. Huikuri HV MT, Airaksinen J, Seppanen T, Puukka P, Raiha J, Sourander LB.
Power-law relationship of heart rate variability as a predictor of mortality in the elderly.
Circulation. 1998;97:2031-2036.
33. Makikallio TH HH, Hintze U, Videbaek J, Mitrani RD, Castellano A, Myerburg RJ,
Moeller M. Fractal analysis and time- and frequency-domain measures of heart rate
variability as predictors of mortality in patients with heart failure. Am J Cardiol.
2001;87:178-182.
34. Colantonio A KS, Ostfeld AM, Berkman LF. Psychosocial predictors of stroke
outcomes in an elderly population. J Gerontol. 1993;49(5):S261-S268.
35. Fratiglioni L WH-X, Ericsson K, Maytan M, Winblad B. Influence of social network
on occurrence of dementia: a community-based longitudinal study. The Lancet.
2000;355:1315-1319.

15
Table 1: Decreased complexity of anatomic structures with aging.
Structure Measure Age Effect Neurons Dendritic arbor Loss of branches and neural
connections Bone Trabecular meshwork Trabecular loss and
disconnection Kidney Glomerular capillary tuft Degeneration and loss of
capillaries Subepidermis Elastic fibers and dermal
papillae Loss and collapse of subepidermal structure
Table 2: Decreased complexity of physiologic systems with aging.
System Measure Age Effect Heart rate dynamics 1/f slope, DFA, & approx.
entropy of interbeat intervals Decreased fractal scaling, more regular dynamics
Blood pressure dynamics 1/f slope, DFA, & approx. entropy of BP fluctuations
Decreased fractal scaling, more regular dynamics
Respiratory dynamics DFA of interbreath intervals
Loss of long range correlations in elderly males
Postural control 1/f slope of center-of-pressure trajectories
Decreased fractal scaling
Gait dynamics 1/f slope, DFA Decreased fractal scaling

16
Table 3: Decreased Complexity in Disease
Disease
(ref) Measure
Alzheimer’s Disease (25)
EEG correlation dimension is reduced
Emphysema (26)
Reduced 1/f slope of size distribution of terminal airspace clusters
Ventric. Fibrillation (27)
Reduced HR fractal scaling (1/f slope*) and long-range correlations (DFA**)
Atrial Fibrillation (28)
Increased RR regularity (ApEn+) and decreased long-range correlations (DFA).
CHF and CAD (10, 29)
Reduced HR fractal scaling (1/f) and long-range correlations (DFA).
Aging and Hunting-ton’s Disease (15)
Loss of stride interval long-range correlations (DFA)
Breast Cancer (30)
Fractal Dimension of mammographic mass.
* 1/f = the slope of the log-transformed power spectrum (see text). **DFA = detrended
fluctuation analysis, a technique that quantifies the fractal-like correlation properties of time-
series data (see text).
+ ApEn = Approximate entropy, a measure of regularity in time series data (11). ApEn
quantifies the (logarithmic) likelihood that a series of data points that are a certain distance apart
for a given number of observations remain within the same distance on next incremental
comparisons.
Copyright (c) by The Gerontological Society of America. Reproduced from (2) by
permission of the Publisher.

17
Table 4: Long-term Consequences of Complexity Loss
Study Authors/ref.
Subjects Complex System
Complex Measure
Adverse Outcome
Ho, et al. Circ.1997 (31)
52 CHF pts.+ 52 matched controls
RR interval (2o ECG)
DFA Mortality- 1.9 yrs.
Huikuri, et al. Circ.1998 (32)
347 random elders >65 yrs.
Heart rate (24oECG)
1/f slope Mortality- 10 yrs
Makikallio et al., AJC 1999) (27)
Case-control, post-MI +/- VF
RR interval (24oECG)
DFA 1/f slope
Ventricular Fibrillation
Huikuri, et al. Circ. 2000 (29)
446 MI pts w/ decreased LV fxn
RR interval (24oECG)
DFA Mortality- 1.9 yrs.
Makikallio, et al. AJC 2001 (33)
499 CHF pts. w/ EF< 35%
RR interval (24oECG)
DFA Mortality- 1.8 yrs.
Colantonio, et al. J. Geron. 1993 (34)
87 elderly stroke survivors
Psycho-social fxn.
Social net-work index
Fx’l loss, NH adm.
Fratiglioni, et al. Lancet 2000 (35)
Popn. of 1203 elders > 75 yrs.
Psycho-social Fxn.
Social network
Dementia
Copyright (c) by The Gerontological Society of America. Reproduced from (2) by permission of
the Publisher.

18
Figure 1: Age-related decrements in physiologic performance. From Shock N, et. al.,
Baltimore Longitudinal Study on Aging (1).

19
Figure 2: Continuous heart rate time series over 8 minutes for a healthy young subject (top
graph) and a healthy elderly subject (bottom graph). Note the similar average heart rate and
standard deviation of heart rate, but different dynamics as quantified by Approximate Entropy.
From (5) with permission from the American Medical Association.

20
Normal Bone Osteoporotic Bone
Figure 3: Loss of Complexity in Bone
21
0 10 20 30
-0.2
-0.1
0
0.1
0.2
0.3Young
time (s)
CO
P di
spla
cem
ent (
mm
)
0 10 20 30
-0.2
-0.1
0
0.1
0.2
0.3Elderly faller
time (s)
CO
P di
spla
cem
ent (
mm
)
0 0.5 1-4
-3.5
-3
-2.5
-2
-1.5
log 10 Frequency (Hz)
log
10Po
wer
(mm
2 /Hz)
Young
0 0.5 1-3.5
-3
-2.5
-2
-1.5
-1
-0.5
log 10 Frequency (Hz)
log
10Po
wer
(mm
2 /Hz)
Elderly faller
β = -1.21 β = -2.29
Figure 4: Center of pressure displacements (top graphs) and their corresponding log-transformed
power specta (bottom graphs) for a 30 year old healthy woman (left) and a 69 year old woman
with previous falls (right), obtained while the subjects stood on a force-plate for 30 seconds. The
slope of the regression line through the power spectra represents the scaling exponent β, which is
greater in the elderly faller than in the young subject, indicating a loss of complexity.
Copyright (c) by The Gerontological Society of America. Reproduced from (2) by permission of
the Publisher.

22
Figure 5: The effect of autonomic blockade with atropine and propanolol on RR interval
dynamics in a healthy human subject.
SALINE
time, sec0 200 400 600 800 1000120014001600
RR
inte
rval
, sec
0.6
0.9
1.2
ATROPINE/PROPRANOLOL
time, sec0 200 400 600 800 1000120014001600
RR
inte
rval
, sec
0.6
0.9
1.2

23
Figure 6: Interbeat interval (RR interval) time series over 200 seconds in 5 piglets as they
mature from 8 to 33 days of life. Note the increasing amplitude (standard deviation) and
irregularity (approximate entropy, ApEn) of the interbeat intervals with advancing postnatal age.
From (17) with permission of Elsevier Science.

24
The Physiologic Basis of Frailty
Figure 7. The physiologic basis of frailty. Multiple interacting physiologic inputs (top) produce
highly irregular, complex dynamics (middle), that impart a high level of functionality (bottom)
on an organism. As the inputs and their connections degrade, the output signal becomes more
regular and less complex, resulting in functional decline. Ultimately, with continued loss of
physiologic complexity, function may fall to the critical level below which an organism can no
longer adapt to stress (the frailty threshold).
Copyright (c) by The Gerontological Society of America. Reproduced from (2) by permission of
the Publisher.
Inpu
ts
Out
put S
igna
l Fu
nctio
nal L
evel
Frailty Threshold
Age


















