Agenda 11/13 Makeup Test Check Ch. 12 Notes Finish cell cycle, regulation and cancer Homework –...
59
Agenda 11/13 • Makeup Test • Check Ch. 12 Notes • Finish cell cycle, regulation and cancer Homework – Print AP Lab 7 – Mitosis and Meiosis (on my website) – read first 4 pages – will do part one tomorrow Ch. 13 notes and concept checks due Friday, online assignment will be due end of break Me – copy old lab modeling, pop beads ready TA’s – upload Powerpoints
-
Upload
asher-byrd -
Category
Documents
-
view
214 -
download
0
Transcript of Agenda 11/13 Makeup Test Check Ch. 12 Notes Finish cell cycle, regulation and cancer Homework –...

Agenda 11/13
• Makeup Test• Check Ch. 12 Notes• Finish cell cycle, regulation and cancer
Homework –Print AP Lab 7 – Mitosis and Meiosis (on my
website) – read first 4 pages – will do part one tomorrow
Ch. 13 notes and concept checks due Friday, online assignment will be due end of break
Me – copy old lab modeling, pop beads ready
TA’s – upload Powerpoints

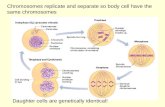
• The mitotic (M) phase of the cell cycle alternates with the much longer interphase.– The M phase includes mitosis and cytokinesis.– Interphase accounts
for 90% of the cell cycle.
1. The Cell Cycle
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.4
Let’s watch an animation – online lesson 4.1 – Activity 2

Figure 12.5-3
ChromosomesChromosomal
DNA molecules
Centromere
Chromosomearm
Chromosome duplication(including DNA replication)and condensation
Sisterchromatids
Separation of sisterchromatids intotwo chromosomes
1
2
3
•The centromere is the narrow “waist” of the duplicated chromosome, where the two chromatids are most closely attached

Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.5 left

Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.5 right

• Assembly of the spindle microtubules starts in the centrosome.– The centrosome (microtubule-organizing
center) of animals has a pair of centrioles at the center, but the function of the centrioles is somewhat undefined.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.6a

• When a chromosome’s kinetochore is “captured” by microtubules, the chromosome moves toward the pole from which those microtubules come.
• When microtubules attach to the other pole, this movement stops and a tug-of-war ensues.
• Eventually, the chromosome settles midway between the two poles of the cell, the metaphase plate.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• One hypothesis for the movement of chromosomes in anaphase is that motor proteins at the kinetochore “walk” the attached chromosome along the microtubule toward the opposite pole.– The excess microtubule sections
depolymerize.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.7a

• Nonkinetichore microtubules are responsible for lengthening the cell along the axis defined by the poles.– These microtubules interdigitate across the
metaphase plate.– During anaphase motor proteins push
microtubules from opposite sides away from each other.
– At the same time, the addition of new tubulin monomers extends their length.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• On the cytoplasmic side of the cleavage furrow a contractile ring of actin microfilaments and the motor protein myosin form.
• Contraction of the ring pinches the cell in two.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.8a

• Cytokinesis in plants, which have cell walls, involves a completely different mechanism.
• During telophase, vesicles from the Golgi coalesce at the metaphase plate, forming a cell plate.– The plate enlarges until its
membranes fuse with the plasma membrane at the perimeter, with the contents of the vesicles forming new wall material in between.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.8b

© 2011 Pearson Education, Inc.
Animation: Cytokinesis Right-click slide / select ”Play”

© 2011 Pearson Education, Inc.
Animation: Animal Mitosis Right-click slide / select ”Play”

© 2011 Pearson Education, Inc.
Animation: Sea Urchin (Time Lapse) Right-click slide / select ”Play”

Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.9

• Prokaryotes reproduce by binary fission, not mitosis.
• Possible intermediate evolutionary steps are seen in the division of two types of unicellular algae.
4. Mitosis in eukaryotes may have evolved
from binary fission in bacteria

Figure 12.13
(a) Bacteria
(b) Dinoflagellates
(d) Most eukaryotes
Intact nuclearenvelope
Chromosomes
Microtubules
Intact nuclearenvelope
Kinetochoremicrotubule
Kinetochoremicrotubule
Fragments ofnuclear envelope
Bacterialchromosome
(c) Diatoms andsome yeasts

• The distinct events of the cell cycle are directed by a distinct cell cycle control system.– These molecules trigger and coordinate key
events in the cell cycle. – The control cycle has
a built-in clock, but it is also regulated by external adjustments and internal controls.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.13

• For many cells, the G1 checkpoint seems to be the most important
• If a cell receives a go-ahead signal at the G1 checkpoint, it will usually complete the S, G2, and M phases and divide
• If the cell does not receive the go-ahead signal, it will exit the cycle, switching into a nondividing state called the G0 phase
© 2011 Pearson Education, Inc.

Figure 12.16
G1 checkpoint
G1 G1
G0
(a) Cell receives a go-ahead signal.
(b) Cell does not receive a go-ahead signal.

• Cyclin levels rise sharply throughout interphase, then fall abruptly during mitosis.
• Peaks in the activity of one cyclin-Cdk complex, MPF, correspond to peaks in cyclin concentration.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.14a

(b) Molecular mechanisms that help regulate the cell cycle
Cdk
Degradedcyclin
Cyclin isdegraded
MPF
G2checkpoint
Cdk
Cyclin
M
S
G 1G 2
Figure 12.17b

The M phase checkpoint • Ensures all chromosomes properly attached to
spindle at metaphase plate before anaphase– This ensures that daughter cells do not end up with
missing or extra chromosomes.
• A protein signal to delay anaphase originates at kinetochores that have not yet attached to spindle microtubules.– This keeps the anaphase-promoting complex (APC) in
an inactive state.– When all kinetochores are attached, the APC activates,
triggering breakdown of cyclin and inactivation of proteins uniting sister chromatids together.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• All necessary nutrients must be present for cell division and usually growth factor also.– For example, platelet-derived growth factors
(PDGF), produced by platelet blood cells, bind to tyrosine-kinase receptors of fibroblasts, a type of connective tissue cell.
– This triggers a signal-transduction pathway that leads to cell division.
• In a living organism, platelets release PDGF in the vicinity of an injury.
• The resulting proliferation of fibroblasts help heal the wound.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• The role of PDGF is easily seen in cell culture.– Fibroblasts in culture will only divide in the
presence of medium that also contains PDGF.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.15

• Growth factors appear to be a key in density-dependent inhibition of cell division.– Cultured cells normally
divide until they form a single layer on the inner surface of the culture container.
– If a gap is created, the cells will grow to fill the gap.
– At high densities, the amount of growth factors and nutrients is insuffi-cient to allow continued cell growth.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.16a

• Most animal cells also exhibit anchorage dependence for cell division.– To divide they must be anchored to a
substratum, typically the extracellular matrix of a tissue.
• Cancer cells are free of both density-dependent inhibition and anchorage dependence.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.16b

• Cancer cells divide excessively and invade other tissues because they are free of the body’s control mechanisms. – Cancer cells do not stop dividing when growth
factors are depleted either because they manufacture their own, have an abnormality in the signaling pathway, or have a problem in the cell cycle control system.
3. Cancer cells have escaped from cell cycle controls
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• Cancer cells may divide indefinitely if they have a continual supply of nutrients.– In contrast, nearly all mammalian cells divide
20 to 50 times under culture conditions before they stop, age, and die.
– Cancer cells may be “immortal”.• Cells (HeLa) from a tumor removed from a woman
(Henrietta Lacks) in 1951 are still reproducing in culture.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• The abnormal behavior of cancer cells begins when a single cell in a tissue undergoes a transformation that converts it from a normal cell to a cancer cell.– Normally, the immune system recognizes and destroys
transformed cells.– However, cells that evade destruction proliferate to
form a tumor, a mass of abnormal cells.
• If the abnormal cells remain at the originating site, the lump is called a benign tumor. – Most do not cause serious problems and can be
removed by surgery.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

• In a malignant tumor, the cells leave the original site to impair the functions of one or more organs.– This is what you think of as “cancer”– These cancer cells often lose attachment to
nearby cells, are carried by the blood and lymph system to other tissues, and start more tumors in a event called metastasis.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Fig. 12.17

• Treatments for metastasizing cancers include high-energy radiation and chemotherapy with toxic drugs.– These treatments target actively dividing cells.
• Researchers are beginning to understand how a normal cell is transformed into a cancer cell.– The causes are diverse.– However, cellular transformation always
involves the alteration of genes that influence the cell cycle control system.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings

Apoptosis
• Cell suicide for the good of the body –
• Cell signaling pathways to turn on enzymes to digest the cell and implode

Lab 3
• Lab bench
• http://www.phschool.com/science/biology_place/labbench/lab3/intro.html

Agenda 11/14
• Do background and getting started questions for Lab 7• Then do modeling mitosis while looking at onion root tip
slides
Homework – • Read part 4 and 5 of lab and do prelab for
meiosis/Sordaria Ch. 13 notes and concept checks due Friday, online
assignment will be due end of break
Me – get access to genetics lab, get Paul online, finalize BioRad PO,

Getting Started Question #1

Figure 12.UN05

Agenda 11/15• Go over modeling mitosis questions
• Meiosis Bioflix video
http://media.pearsoncmg.com/bc/bc_0media_bio/bioflix/bioflix.htm?9apmeiosis
• Meiosis slides• Meiosis simulation
Homework – Ch. 13 notes and concept checks due tomorrow, online assignment will be due end of
break – 64 minutes
• Review manual – read Ecology section, highlight or add to your chapter notes as needed, and do practice questions on separate paper due Tuesday back
• Skim and summarize Ecology article – due Monday back – I will email link

• In humans, each somatic cell (all cells other than sperm or ovum) has 46 chromosomes.– Each chromosome can be distinguished by its size,
position of the centromere, and by pattern of staining with certain dyes.
• A karyotype display of the 46 chromosomes shows 23 pairs of chromosomes, each pair with the same length, centromere position, and staining pattern.
• These homologous chromosome pairs carry genes that control the same inherited characters.
1. Fertilization and meiosis alternate in sexual life cycles

• Karyotypes, ordered displays of an individual’s chromosomes, are often prepared with lymphocytes.
Fig. 13.3

• Fertilization restores the diploid condition by combining two haploid sets of chromosomes.
• Fertilization and meiosis alternate in sexual life cycles.
Fig. 13.4

3 Types of Life Cycles

• Meiosis reduces chromosome number by copying the chromosomes once, but dividing twice.
• The first division, meiosis I, separates homologous chromosomes.
• The second, meiosis II, separates sister chromatids.
Fig. 13.6

Figure 13.8
MEIOSIS I: Separates homologous chromosomes
Prophase I Metaphase I Anaphase I Telophase I andCytokinesis
Centrosome(with centriole pair)
Sisterchromatids
Chiasmata
Spindle
Homologouschromosomes
Fragmentsof nuclearenvelope
Duplicated homologouschromosomes (red and blue)pair and exchange segments;2n 6 in this example.
Centromere(with kinetochore)
Metaphaseplate
Microtubuleattached tokinetochore
Chromosomes line upby homologous pairs.
Sister chromatidsremain attached
Homologouschromosomesseparate
Each pair of homologous chromosomes separates.
Cleavagefurrow
Two haploid cellsform; each chromosomestill consists of twosister chromatids.
MEIOSIS I: Separates sister chromatids
Prophase II Metaphase II Anaphase IITelophase II and
Cytokinesis
Sister chromatidsseparate
Haploid daughtercells forming
During another round of cell division, the sister chromatids finally separate;four haploid daughter cells result, containing unduplicated chromosomes.

Figure 13.8a
Prophase I Metaphase I Anaphase I Telophase I andCytokinesis
Centrosome(with centriole pair)
Sisterchromatids
Chiasmata
Spindle
Homologouschromosomes
Fragmentsof nuclearenvelope
Duplicated homologouschromosomes (red and blue)pair and exchange segments;2n 6 in this example.
Centromere(with kinetochore)
Metaphaseplate
Microtubuleattached tokinetochore
Chromosomes line upby homologous pairs.
Sister chromatidsremain attached
Homologouschromosomesseparate
Each pair of homologous chromosomes separates.
Cleavagefurrow
Two haploid cells form; each chromosomestill consists of two sister chromatids.

Figure 13.8b
Prophase II Metaphase II Anaphase II Telophase II andCytokinesis
Sister chromatidsseparate
Haploid daughtercells forming
During another round of cell division, the sister chromatids finally separate;four haploid daughter cells result, containing unduplicated chromosomes.

• Three events, unique to meiosis, occur during the first division cycle.
1. During prophase I, homologous chromosomes pair up in a process called synapsis.– A protein zipper, the synaptonemal complex, holds
homologous chromosomes together tightly.– Later in prophase I, the joined homologous
chromosomes are visible as a tetrad.– At X-shaped regions called chiasmata, sections of
nonsister chromatids are exchanged.– Chiasmata is the physical manifestation of crossing
over, a form of genetic rearrangement.

2. At metaphase I homologous pairs of chromosomes, not individual chromosomes are aligned along the metaphase plate.
• In humans, you would see 23 tetrads.
3. At anaphase I, it is homologous chromosomes, not sister chromatids, that separate and are carried to opposite poles of the cell.– Sister chromatids remain attached at the
centromere until anaphase II.
• The processes during the second meiotic division are virtually identical to those of mitosis.

• Sister chromatid cohesion allows sister chromatids of a single chromosome to stay together through meiosis I
• Protein complexes called cohesins are responsible for this cohesion
• In mitosis, cohesins are cleaved at the end of metaphase
• In meiosis, cohesins are cleaved along the chromosome arms in anaphase I (separation of homologs) and at the centromeres in anaphase II (separation of sister chromatids)
© 2011 Pearson Education, Inc.

• Mitosis produces two identical daughter cells, but meiosis produces 4 very different cells.
Fig. 13.8

Fig. 13.8

Three mechanisms of genetic variation
• 1) Independent Assortment
•For humans with n = 23, there are 223 or about 8 million possible combinations of chromosomes.

Figure 13.11-5Prophase Iof meiosis
Nonsister chromatidsheld togetherduring synapsis
Pair of homologs
Chiasma
Centromere
TEM
Anaphase I
Anaphase II
Daughtercells
Recombinant chromosomes
2) Crossing over
• produces recombinant chromosomes which combine genes inherited from each parent.

• 3) Fertilization – The random nature of fertilization adds to the
genetic variation arising from meiosis.– Any sperm can fuse with any egg.
• An ovum is one of approximately 8 million possible chromosome combinations (actually 223).
• The successful sperm represents one of 8 million different possibilities (actually 223).
– The resulting zygote is composed of 1 in 70 trillion (223 x 223) possible combinations of chromosomes.
– Crossing over adds even more variation to this.
Sex and mutations are two sources of the continual generation of new genetic variability.

© 2011 Pearson Education, Inc.
Animation: Genetic Variation Right-click slide / select “Play”

The Evolutionary Significance of Genetic Variation Within Populations
• Natural selection results in the accumulation of genetic variations favored by the environment
• Sexual reproduction contributes to the genetic variation in a population, which originates from mutations
© 2011 Pearson Education, Inc.

Figure 13.UN03

Agenda 11/16Watch Lab Bench Sordaria part
• http://www.phschool.com/science/biology_place/labbench/lab3/intro.html
• Check and go over prelab • Do Sordaria part 5• Go over analysis questions
Homework - • Online assignment Ch. 13 due Monday back 8am• Review manual – read Ecology section, highlight or add to your chapter
notes as needed, and do practice questions on separate paper due Tuesday back
• Skim and summarize Ecology article on Socotra (National Geographic) – I’ll email link – due Monday back