
Advances in Pest Management in Legume Crops - CCS HAU, Hisar
228
Transcript of Advances in Pest Management in Legume Crops - CCS HAU, Hisar



Vice-Chancellor
CCS Haryana Agricultural University
HISAR- 125004 (Haryana) India
FOREWORD
Grain legumes have historically been cul�vated for their protein- and energy-rich dry seeds which are o�en called pulses. Legumes, par�cularly pigeonpea and chickpea, are major protein sources for poor communi�es in many parts of semi-arid tropical areas of Africa and Asia. They have long been recognized by farmers and scien�sts alike as builders and restorers of soil fer�lity. However, in Asia, the green revolu�on concentrated exclusively on increasing cereal produc�on and neglected all other food crops, including pulses that had previously played an important part in cereal cropping systems. Far less acreage was given over to pulses, which were relegated to marginal areas where soils were less produc�ve.
The produc�vity of legume crops, especially the pulses, has not increased markedly in 30 years. This is a serious concern in this �me of exponen�al human popula�on growth because legume crops provide essen�al dietary components that are not present in cereals. The demand for food is likely to increase about by 50 per cent by 2050. This target will have to be achieved in the backdrop of decrease in cul�vable land, increased water deficit, and increased uncertainty of clima�c condi�ons. With the
0 predic�on of 2- 4 rise in temperature in the next 100 years, condi�ons will become further challenging to grow crops successfully.
Food legumes are damaged by a large number of insect pests. Losses due to pod borers such as Helicoverpa armigera are incalculable on a global scale, although on chickpea and pigeonpea have been es�mated as exceeding US$ 600 million in India annually. Insec�cide use against pod borers is prevalent in developing countries but the control achieved is largely unsa�sfactory. Globally, lot of efforts are being made by the scien�sts to manage insect pests in legume crops. Sources of resistance to insects in grain legumes have been iden�fied, but these have not been used effec�vely in crop improvement. There is a need to place greater emphasis on u�liza�on of wild rela�ves of crops with different resistance mechanisms, gene�c engineering of plants for insect resistance, and iden�fica�on of molecular markers associated with resistance to insect pests.
Final Correction Rakesh 19.11.2013

Future research must focus on environmentally sound pest management strategies that are compa�ble with the needs and limita�ons of legume producing farmers. Cultural manipula�on of the crop and its environment, popula�on monitoring and pest forecas�ng, use of natural plant products and bio-pes�cides alone or in combina�on with synthe�c pes�cides, use of natural enemies, deployment of insect resistant varie�es derived through conven�onal breeding, or gene�c engineering, and ra�onal use of selec�ve chemicals can be exploited for pest management in food legumes.
I am happy that a compendium of lectures delivered during the advanced training course on “Advances in Pest Management in Legume Crops” is being published by the Centre of Advanced Faculty Training in the Department of Entomology. Needless to say that this course would benefit the par�cipants immensely. I hope that the knowledge so gained by the par�cipants would be helpful in further refinement of their research projects.
(K. S. KHOKHAR)

PREFACE
Legume crops are an important component of Indian agriculture. They are a major source of protein, vitamins, minerals and roughages, par�cularly for the vegetarian popula�on, apart from their miscellaneous uses in animal feed. A hallmark trait of legumes is their ability to develop root nodules and to fix nitrogen in symbiosis with compa�ble rhizobia. The produc�vity of legume crops, especially the pulses, has not increased markedly in 30 years. The most important produc�on problems (a�er uncontrollable abio�c factors like drought and flooding) are pests and diseases, especially in the tropics and subtropics. Therefore, with the objec�ve to update the theore�cal and prac�cal knowledge of the trainees with respect to important pests of legume crops and their management and to provide an opportunity for sharing the experiences of the par�cipa�ng scien�sts among themselves, the present training course on “Advances in pest management in legume crops” was organized from September 25 to October 15, 2013.
Important aspects covered during this course included current status of entomological research in legume crops; key pests of legume crops and their management; host plant resistance, management of insect vectors; sampling and monitoring techniques for pests of legume crops; pest damage assessment; ecological factors affec�ng popula�ons of important pests; transgenic protec�on of legume crops; use of bio-pes�cides; post harvest pest management; exploita�on of natural enemies; disease diagnosis and nutrients deficiency symptoms diagnosis in legume crops; etc. Most of the lectures were contributed by the specialists from CCS Haryana Agricultural University, Hisar. However, some of these were delivered by experts from different agricultural universi�es/ins�tutes such as Navsari Agricultural University, Navsari (Gujarat); Indian Agricultural Research Ins�tute, New Delhi; Agricultural Research Sta�on, Rajasthan Agricultural University, Durgapura (Jaipur); and Punjab Agricultural University, Ludhiana. Eleven par�cipants represen�ng seven Agricultural University from five states a�ended this course.
The financial assistance received from Indian Council of Agricultural Research (ICAR), New Delhi for organizing the training course, and help and coopera�on received from different resource persons, faculty and staff of Department of Entomology and other departments of the University who have been associated with this course are gratefully acknowledged.
I am indebted to worthy Vice-Chancellor, Prof. K. S. Khokhar, for the patronage, support and encouragement given by him to this training programme. I express deep sense of gra�tude to Dr R. K. Pannu, Dean, College of Agriculture, for his keen interest and extending all sorts of help and guidance. I owe my sincere thanks to Dr. S. S. Siwach, Director of Research, for his encouragement, coopera�on and help. Support from members of various commi�ees engaged with this programme and the �reless efforts made by the Course Coordinator, Dr. S. S. Sharma, are thankfully acknowledged. I hope this compendium will be of great help to students, researchers and teachers in understanding various basic and applied aspects of legume entomology.
Course Director
“ADVANCES IN PEST MANAGEMENT IN LEGUME CROPS” TH THFROM 25 SEPTEMBER TO 15 OCTOBER, 2013
LIST OF PARTICIPANTS
Rahul M. Wadaskar
Pulses Research Unit
Dr. PDKV, Akola (M.S.).
E-mail: [email protected]
Mob: 099229-34949
Mr. Balu Narayan Bhangare
AICRP on Tuber Crops
Dr. B.S.K.K.V.,
Dapoli, Ratnagiri (MS).
E-mail: [email protected]
Mob: 09272388436
Dr.Kamlesh Bali
Jr.Scien�st-cum-Ass�.Professor
(Entomology), SKUAST-Jammu, RARS,
Rajouri (J&K).
E-mail:
Mob: 094191-56289
Dr. F.K. Chaudhary
Subject Ma�er Specialist (Pl.Prot.)
KVK, S.D. A.U., Deesa
Dis�. Banaskantha, Gujarat.
E-mail: �[email protected]
Mob: 09273531557
Dr.Sunita Yadav
Ass�.Scien�st
Dep�.of Entomology
CCSHAU, Hisar
E-mail: [email protected]
Mob: 08930930874
Dr. Gajanan K. Lande
Ass�.Professor (Entomology), University
Dep�.of Entomology
Dr. PDKV, Akola (M.S.).
E-mail: [email protected]
Mob: 09421749429
Dr. U�am Kisan Kadam
AICRP on Citrus,
M.P.K.V., Rahuri (M.S.).
E-mail: [email protected]
Mob: 09273531557
Dr. Vidyasagar Kishanrao Biradar
AICRP on Linseed
College of Agriculture, Nagpur (M.S.).
(Under Dr. PDKV, Akola).
E-mail: [email protected]
Mob: 09273531557
Dr. Amandeep Kaur
DES (Entomology)
Pa�ala (Pb.)
PAU, FASS.
E-mail: [email protected].
Mob: 09815158866
Dr.J.R. Patel
Subject Ma�er Specialist (Pl.Prot.)
KVK, S.D. A.U., Khembrahma
Dis�. Sabarkantha, Gujarat.
E-mail: [email protected]
Mob: 09428197133
Dr. Pravin K. Rathod
Assistant Entomologist
AICRP on PHT,
Dr. PDKV, Akola (M.S.).
E-mail: [email protected]
Mob: 09423384974

CONTENTS
No. TITLE AND NAME
1. CURRENT STATUS OF ENTOMOLOGICAL RESEARCH IN LEGUME CROPS: AN OVERVIEW R. K. SAINI 1
2. ADVANCES IN USE OF MODERN KEYS FOR PEST IDENTIFICATION ROSHAN LAL 11
3. MANAGEMENT STRATEGIES FOR INSECT PESTS OF COWPEA AND CLUSTERBEAN S.P. SINGH 30
4 MANAGEMENT STRATEGIES FOR INSECT PESTS OF PEA AND FRENCH BEAN S. S. SHARMA 35
5. NOVEL INSECTICIDES FOR LEGUME PEST MANAGEMENT S.S. YADAV AND KRISHNA ROLANIA 50
6. ROLE OF CULTURAL METHODS IN SUPPRESSING INSECT- PESTS IN LEGUME CROPS K. K. DAHIYA 57
7. USE OF BIOPESTICIDES IN LEGUME PEST MANAGEMENT V.S. MALIK 65
8. MANAGEMENT STRATEGIES OF LEPIDOPTEROUS INSECT PESTS ESPECIALLY THROUGH NUCLEAR POLYHEDROSIS VIRUS H.D.KAUSHIK 72
9. MANAGEMENT STRATEGIES FOR HAIRY CATERPILLARS IN LEGUME CROPS OMBIR AND R.K. SAINI 79
10. MANAGEMENT OF WHITEGRUBS IN KHARIF LEGUME CROPS SWAROOP SINGH* AND VIJAY KUMAR AGRAWAL 86
11. PROSPECTS OF EXPLOITATION OF NATURAL ENEMIES ASSOCIATED WITH PESTS OF LEGUME CROPS PALA RAM 94
12. DETAILED ACCOUNT OF WHITEFLY, BEMISIA TABACI (GENNADIUS) A VECTOR OF GEMINIVIRUS IN LEGUME CROPS S.S. SHARMA 103

13. MANAGEMENT STRETAGIES FOR INSECT PESTS OF PIGEONPEA KRISHNA ROLANIA AND S. S. YADAV 106
14. TERMITE MANAGEMENT IN LEGUME CROPS S S YADAV, J.P.BHANOT AND KRISHNA ROLANIA 113
15. ROLE OF INSECT POLLINATORS IN PRODUCTION OF LEGUME CROPS S.K.SHARMA AND YOGESH KUMAR 119
16. ROLE OF POLLINATORS IN PRODUCTION OF LEGUMINOUS VEGETABLES YOGESH KUMAR AND S.K.SHARMA 126
17. HOST PLANT RESISTANCE TO INSECT PESTS IN CHICKPEA MUKESH K. DHILLON 135
18. HOST PLANT RESISTANCE TO INSECT PESTS IN PIGEONPEA MUKESH K. DHILLON 142
19. ECOLOGICAL FACTORS AFFECTING POPULATION OF IMPORTANT INSECT PESTS OF LEGUME CROPS G.S. YADAV 148
20. DISEASE DIAGNOSIS OF LEGUME CROPS AND THEIR MANAGEMENT S.K. KHIRBAT 153
21. NEMATODE INFESTATION AND ITS DIAGNOSES IN LEGUME CROPS K. R. DABUR* & I. J. PARUTHI 158
22. MITE PESTS OF LEGUMES AND THEIR MANAGEMENT* ABHISHEK SHUKLA AND S. S. MANDAPE 165
23. IDENTIFICATION OF MITES ASSOCIATED WITH LEGUME CROPS* ABHISHEK SHUKLA AND G.G. RADADIA 187
24. STRATEGIES TO UNDERSTAND INSECT RESISTANCE IN PLANTS THROUGH THE RICE-GALL MIDGE INTERACTION SURESH NAIR 192
25. MANAGEMENT OF INSECT VECTORS IN MUNGBEAN AND URDBEAN RAVINDER SINGH GURWARA 196
26. INSECT SAMPLING METHODS FOR IMPORTANT LEGUME CROPS R. K. SAINI 200

CURRENT STATUS OF ENTOMOLOGICAL RESEARCH IN LEGUME CROPS: AN OVERVIEW
R. K. SAINIDepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
Grain legumes are in the botanical family Fabaceae (formerly known as the
Leguminosae family) and are used for human food and animal feed. There are more
than forty species and many varie�es of grain legumes. Grain legumes have historically
been cul�vated for their protein- and energy-rich dry seeds which are o�en called
pulses. Legumes, par�cularly pigeonpea (Cajanus) and chickpea (Cicer) are major
protein sources for poor communi�es in many parts of semi-arid tropical areas of Africa
and Asia. They can be totally destroyed by insects and diseases. The produc�vity of
legume crops, especially the pulses, has not increased markedly in 30 years. This is a
serious ma�er in this �me of exponen�al human popula�on growth because legume
crops provide essen�al diet components that are not present in cereals.
Food legumes such as chickpea, pigeonpea, cowpea, field pea, len�l, faba bean,
blackgram, greengram, grasspea, and Phaseolus beans play an important role in the
daily diets of people worldwide. These crops are damaged by a large number of insect
pests, of which pod borers, Helicoverpa armigera and H. punc�gera; spo�ed pod borer,
Maruca vitrata; spiny pod borer, E�ella zinckenella; pod fly, Melanagromyza obtusa;
leaf miner, Liriomyza cicerina; stem fly, Ophiomyia phaseoli; pea and bean weevil,
Sitona spp.; aphids, Aphis craccivora, Aphis fabae, and Acyrthosiphon pisum; white fly,
Bemisia tabaci; defoliators, Spodoptera litura, S. exigua, and Amsacta spp.;
lea�oppers, Empoasca spp., thrips, Megaleurothrips dorsalis, and Caliothrips indicus;
blister beetles, Mylabris spp.; and the bruchids, Collasobruchus chinensis and Bruchus
pisorum cause extensive losses worldwide (Sharma et al. (2005).
Losses due to notorious borers such as Helicoverpa armigera are incalculable on
a global scale, although on chickpea and pigeonpea have been es�mated as exceeding
US$600million in India annually. Insec�cide use against pod borers is prevalent in
developing countries and largely ineffec�ve. Apart from the well-documented social
and environmental hazards of liberal use of pes�cides in the developing tropics, two
par�cular problems have emerged: (a) development of resistance and (b) destruc�on
of the associated natural enemy complex. Focussing control strategy on non-chemical
control methods has the greatest likelihood of providing benefits on mul�ple fronts.
1

ENTOMOLOGICAL RESEARCH IN LEGUME CROPS IN ASIA
In general, in East and South-East Asia legumes are most likely to be eaten as
green vegetables. Green gram and soybean are o�en processed in the manufacture of
noodles and fermenta�on products. In predominantly vegetarian South Asia, nearly all
thelegumes are eaten, but the pulses (dried seeds) are significant as supplementsto the
staple rice and wheat dishes. Groundnut and soybean are dominant sources of high
quality vegetable oil but are also eaten in many forms. Produc�on levels have not
approached the 1:9 pulse to cereal ra�o (1 1% pulse) considered normal for Asia for
many years. The produc�on of legume oil seeds (groundnut and soybean) presents only
a slightly brighter picture in terms of increasing produc�vity and the area sown. All
produc�on figures are not gloomy: soybean and groundnut produc�on are increasing
in India and Indonesia. The most important produc�on problem (a�er uncontrollable
abio�c factors like drought and flooding) is a cohort of insect pests. Its members
regularly reduce the yield of legume crops to levels at which it is not in the interest of
the farmers to harvest them. However, other components of the farming systems in
which these crops grow are also involved. For instance, in India and Thailand the heavy
applica�on of insec�cides to co�on has enhanced the injury caused by Helicoverpa spp.
and whiteflies to legume crops in neighboring fields.
The problem created by these pests is different and considerably more intense
than it was perhaps 10 years ago. Many insects are now out of control despite the liberal
applica�on of insec�cides. The best documented example within the current context is
that of Helicoverpa resistance to several insec�cide classes. Specialists from The
Philippines, Vietnam, Indonesia. Thailand, Sri Lanka and India have reported that
farmers apply insec�cides to legume crops at least once a week to kill Maruca, E�ella,
Helicoverpa and the large Heteroptera. The natural control process has been destroyed
and the target insects are highly resistant to pes�cides. Marketed produce must be
highly contaminated with pes�cides. Farmers can see no answer to the problem except
to apply more insec�cides-the insec�cide treadmill.
Entomological research in legume crops in India
Three All India Coordinated Projects, each for Chickpea, Pigeonpea and MULLaRP
(Mungbean, Urdbean, Len�l, Lathyrus, Rajmash and Pea), are involved in strategic and
applied research, and demonstra�on of technologies developed through the
coordinated efforts for increasing and sustaining produc�on of respec�ve crops in the
country. The headquarter of all these AICRPs is at Indian Ins�tute of Pulses Research,
Kanpur. These projects are operated through different centres located in various agro-
ecological zones of country. The research centres under AICRPs have been playing a
2

crucial role in improvement of respec�ve crops not only as tes�ng centres but also as
major contributors of technologies for tes�ng across the country.
All India Coordinated Research Project on Chickpea:
Systema�c research on Chickpea started with the establishment of the All India Coordinated Pulses Improvement Project (AICPIP) in 1967. The area under chickpea has increased from 6.45 million ha in 1992-93 to 8.56 million ha in 2009-10. Madhya Pradesh, U�ar Pradesh, Rajasthan, Maharashtra, Gujarat, Andhra Pradesh and Karnataka are the major chickpea producing states sharing over 95% area. The north Indian states have lost more than 29% chickpea area which has caused about 28% reduc�on in produc�on during 1975 to 1990. This reduc�on was because of increase in irriga�on facili�es and non-availability of input responsive varie�es of chickpea.
The All India Coordinated Research Project on MULLaRP under the auspices of Indian Council of Agricultural Research (ICAR) is responsible for carrying out applied research on mungbean, urdbean, len�l, lathyrus, rajmash and fieldpea in a coordinated manner at na�onal level.
AICRP on Arid Legumes, Jodhpur
Project coordina�ng unit with HQs at CAZRI, Jodhpur is coordina�ng research ac�vi�es pertaining to gene�c improvement, crop produc�on, plant protec�on, quality components, breeder seed produc�on and conduc�ng Front Line Demonstra�ons. These ac�vi�es relate to 4 mandatory crops of guar, cowpea, moth bean and horse gram. Aim is to increase produc�vity to guar and moth bean in Rajasthan and area in other states, extending area of guar in Anantpur, Chi�oor and Kodappa districts, horse gram in HP and WB regions. The major thrust is on:
Iden�fica�on of genotypes with in-built mul�-stress tolerance towards drought, heat and salt and field resistance against important diseases and storage pests.
Finger prin�ng of released varie�es and lifelines of arid legumes.
Breeding guar (clusterbean) for high galactomannan gum content (>35%), improve meal quality and high seed protein content (25-27%) with increased propor�on of amino acids and improved cooking quality of cowpea and moth bean grains.
Finding out an�-nutri�onal compounds in horse gram seeds and making it an alternate source of edible pulse food, par�cularly in tribal areas.
Legume pod borer, Maruca (testulalis) vitrata (Geyer) is one of the major constraints in increasing the produc�on and produc�vity of grain legumes in the tropics. Screening for resistance has been carried out using natural infesta�on, and
3

mul�- and no-choice tests under greenhouse/laboratory condi�ons. Informa�on is available on genotypic resistance to M. vitrata in cowpea, while such informa�on on pigeonpea and other legumes is limited. Stem and pod wall thickness, trichomes and podding habit are associated with resistance to Maruca. Several natural enemies have been recorded on M. vitrata. Cultural prac�ces such as intercropping, weeding, �me of plan�ng, and plan�ng density reduce its damage in cowpea. Several insec�cides have been found to be effec�ve for controlling this insect. There is a need to generate informa�on on insect-plant-environment interac�ons, screening techniques, mechanisms and diversity of resistance, gene�c transforma�on of host plants involving Bt genes, and use of natural enemies for integrated pest management in diverse agro-ecosystems.
Pigeonpea (Cajanus cajan) is an important crop in semi-arid tropical and subtropical farming systems, providing high quality vegetable protein, animal feed, and firewood. Insect pests feeding on flowers, pods, and seeds are the most important bio�c constraint affec�ng pigeonpea yields. Important pests are: flower- and pod-feeding Lepidoptera, pod-sucking Hemiptera, and seed-feeding Diptera and Hymenoptera. Pigeonpea pest management research has focused un�l recently on the iden�fica�on and development of resistant cul�vars and on chemical control. Future research must focus on environmentally sound pest management strategies that are compa�ble with the needs and limita�ons of pigeonpea farmers.
Entomological research in legume crops in Nepal
Na�onal Grain Legumes Research Program (NGLRP) in Nepal was established in 1985 with the main aim of the program is to develop and recommend suitable technologies on different grain legumes and increase produc�on and produc�vity at na�onal level. Research ac�vi�es at NGLRP are variety development, crop management (agronomical and integrated pest and disease management), outreach (tes�ng and up scaling of promising genotypes through RARS, ARS, NGOs, DADOs/ NGLRP command areas), source seed produc�on and dissemina�on of technology (training, field visit and fair). At present, research ac�vi�es on len�l, pigeonpea, blackgram, soybean, chickpea, cowpea, mungbean, Phaseolus bean, ricebean and grasspea are being conducted at NGLRP Rampur and at other tes�ng research sta�ons and farmers fields. NGLRP has been working in collabora�on with na�onal (Department of Agriculture, NGO, INGO, farmers groups, seed company etc) and interna�onal organiza�ons such as ICRISAT, India (pigeonpea/ deshi chickpea, groundnut); ICARDA, Syria (len�l, Kabuli chickpea, fababean, and grasspea), AVRDC, Taiwan (vegetable soybean, mungbean), CLIMA Australia, IITA, Nigeria (grain type soybean, cowpea), IIPR Kanpur India for germplasm exchange, funding, technical
4

support, human resource development. Grain legumes improved technologies have been developed with the collabora�on with various research and extension partners including valuable support of farmers. The major thrust areas for increasing pulse produc�on are: Germplasm collec�on and evalua�on, Varietal improvement program, Promising genotypes, Produc�on technology, Nutrient management, Water management, Seed produc�on, Crop protec�on.
Integrated Pest Management
Integrated management of grain legumes insect-pests: Prepara�on and use of HNPV, integrated insect management of grain legumes, monitoring of aphid species in cowpea, field experimenta�on to manage aphid and management of cow pea pest in storage condi�on.
Determining status of priority insect pests and review on entomological research in Nepal.
ENTOMOLOGICAL RESEARCH IN LEGUME CROPS IN AFRICA
African agriculture is largely tradi�onal—characterized by a large number of smallholdings of no more than one ha per household. Crop produc�on takes place under extremely variable agro-ecological condi�ons, with annual rainfall ranging from 250 to 750 mm in the Sahel in the northwest and in the semi-arid east and south, to 1500 to 4000 mm in the forest zones in the central west. Farmers o�en select well-adapted, stable crop varie�es, and cropping systems are such that two or more crops are grown in the same field at the same �me. These diverse tradi�onal systems enhance natural enemy abundance and generally keep pest numbers at low levels. Pest management prac�ce in tradi�onal agriculture is a built-in process in the overall crop produc�on system rather than a separate well-defined ac�vity. Increased popula�on pressure and the resul�ng demand for increased crop produc�on in Africa have necessitated agricultural expansion with the concomitant decline in the overall biodiversity.
Increases in plant material movement in turn facilitated the accidental introduc�on of foreign pests. At present about two dozen arthropod pests, both introduced and na�ve, are recognized as one of the major constraints to agricultural produc�on and produc�vity in Africa. Although yield losses of 0% to 100% have been observed on-sta�on, the economic significance of the majority of pests under farmers' produc�on condi�ons is not adequately understood. Economic and social constraints have kept pes�cide use in Africa the lowest among all the world regions. The majority of African farmers s�ll rely on indigenous pest management approaches to manage pest problems, although many government extension programs encourage the use of pes�cides.
5

The current pest management research ac�vi�es carried out by na�onal or
interna�onal agricultural research programs in Africa focus on classical biological
control and host plant resistance breeding. With the excep�on of classical biological
control of the cassava mealybug, research results have not been widely adopted. This
could be due to African farmers facing heterogeneous condi�ons, not needing fixed
prescrip�ons or one ideal variety but a number of op�ons and genotypes to choose
from. Indigenous pest management knowledge is site-specific and should be the basis
for developing integrated pest management (IPM) techniques. Farmers o�en lack the
biological and ecological informa�on necessary to develop be�er pest management
through experimenta�on. Formal research should be instrumental in providing the
input necessary to facilitate par�cipatory technology development such as that done
by Farmer Field Schools, an approach now emerging in different parts of Africa.
ENTOMOLOGICAL RESEARCH IN LEGUME CROPS IN USA
Pest Management Strategic Plan for Succulent Edible Legumes in the North Central
Region (USA)
The North Central Region produces about one half of the snap bean, pea, and
lima bean produc�on of the United States. Most of this produc�on resides within an
area from Michigan through northern Illinois, and southern Wisconsin and Minnesota.
Although some legumes are consumed as part of the fresh market crop the vast
majority is processed as canned or frozen produce.
Issues concerning insect pest management were:
Research is needed to address virus-aphid e�ology, epidemiology and
management models in snap beans. Viral diseases can be devasta�ng and producers
feel vulnerable to the possibility of major losses.
Plant breeders need to maintain efforts to iden�fy available germplasm for
resistance to diseases (e.g. white mold, root rot pathogens, viruses) and insects (e.g.
leaf hoppers, ear worm, corn borer, aphids) for edible legumes. Producers realize that
varietal improvement is an ongoing and important need.
An important part of pest management is being able to predict the
development, dispersal, and distribu�on of weeds, insects and diseases of edible
legumes. A regional pest mapping system for all major pests might significantly improve
their ability to control seasonal pests.
There are several products which are as yet unregistered for edible legumes
that would benefit growers. Cruiser, an insec�cide, should also be considered for fast
track registra�on in group 6 crops as a replacement for threatened OPs.
6

In northern Florida, many different insects a�ack leguminous vegetables.
Aphids damage terminals, whiteflies feed on sap and transmit bean golden mosaic
virus, and caterpillars, like bean leafroller, and beetles feed on leaves. Flower thrips
feed in blossoms and s�nk bugs, corn earworm, and leaffooted bugs damage seeds and
pods. Snap beans are becoming an important crop for Florida. Southern peas (a bean)
are common.
ENTOMOLOGICAL RESEARCH IN LEGUME CROPS IN AUSTRALIA
The Helicoverpa problem
Helicoverpa spp. represent a significant challenge to field crops in many parts of the world. In Australia, H. armigera and H. punc�gera are extremely well adapted to exploit the natural environment and managed agricultural systems. They remain the target for concerted management with pes�cides and concerted research efforts seeking to develop alterna�ves to pes�cides, which allow more sustainable integrated approaches for management. While the last decade has seen considerable advances in the development and adop�on of IPM systems in certain crops, further advances are needed. There is unlikely to be a simple solu�on to the control of such adaptable and mobile pests as Helicoverpa spp., but by taking an integrated approach to the Helicoverpa problem in Australia we may be able to achieve the goal of long term, sustainable pest management strategies with minimal environmental impacts.
The two examples of current research outlined here – Bt co�on and development of host plant resistance in chickpea – represent a small part of the current IPM focused research effort. Other areas of research ac�vity include the ecology and management of beneficial insects, the use of trap crops and refuge crops, popula�on structure and movement of Helicoverpa spp., resistance management and the gene�c basis of pes�cide resistance, “so�” non-disrup�ve pes�cides and behaviour modifying chemicals (a�ractants, repellants) for use in management systems.
IPM systems will, of necessity, be more complex than the pes�cide based systems of the past, and will require greater effort on the part of crop managers whether they be professional consultants or farmers themselves. Likewise there remains a need for ongoing research to develop a suite of management tac�cs and integrate them into IPM systems. For example, transgenic co�ons expressing insec�cidal proteins provide great scope to drama�cally reduce pes�cide dependence, but they must be supported with well-researched resistance management strategies and by a range of other pest management approaches.
Centre for Legumes in Mediterranean Agriculture (CLIMA)
The Centre for Legumes is a research alliance between the Department of Agriculture Western Australia,The University of Western Australia,CSIRO and Murdoch University,
7

formed to con�nue the research collabora�on begun under the Commonwealth Government's Coopera�ve Research Centre Program. CLIMA's research projects are funded largely by the Grains Research and Development Corpora�on (GRDC),Australian Centre for Interna�onal Agricultural Research (ACIAR), Rural Industries Research and Development Corpora�on (RIRDC), Grains Research Commi�ee of Western Australia (GRC-WA) and the increasingly ac�ve WA-based Council of Grain Growers Organisa�ons (COGGO). The following programmes/projects are being taken up by the Centre:
1. Germplasm Development for Grain Legumes
The Germplasm Development for Grain Legumes Sub-program con�nues to add value to Western Australian and na�onal grain legume breeding programs through the acquisi�on and screening of new germplasm and through the development and applica�on of new breeding technologies to our mandate species: lupin, chickpea, field pea, faba bean and len�l. Over the past two years interna�onal linkages enabled the Cenre to import in excess of 2000 new accessions of lupins and pulses. These accessions include lines that have resistance to major diseases that are a threat to our agricultural industries. In addi�on, our linkages with the GRDCcoordinated na�onal breeding programs ensure that relevant advanced breeding lines are evaluated locally with the goal of releasing the best adapted lines to Western Australian farmers.
2. Interna�onal collabora�on to develop interspecific hybrids between chickpea and its wild rela�ves
Wild rela�ves within the genus Cicer may be the answer to very low gene�c varia�on in domes�cated chickpea. A range of valuable agronomic traits have already been iden�fied within the eight wild annual species.These include resistance to diseases such as ascochyta blight, fusarium wilt, phytophora, botry�s grey mould; pests such as leaf miner, bruchid and cyst nematode; and improved tolerance to abio�c stresses like cold and drought. Two wild annual Cicer species, C. re�culatum and C. echinospermum, share the primary gene pool with chickpea (C. arie�num) and can be crossed using conven�onal crossing methods to produce fer�le hybrids.
3. Disease and Pest Management
The Disease and Pest Management Sub-program seeks to protect grain legume crops from the threat of disease and pests by:
improving understanding of the mechanisms of pathogenicity adopted by pests and disease-causing organisms;
iden�fying exo�c threats to exis�ng grain species;
improving understanding of the mechanisms of Lupin resistance to to pests and diseases;
8

facilita�ng the selec�on of resistant germplasm;
iden�fying and characterising germplasm that is resistant to pests and diseases; and
iden�fying genes that could be transferred to grain legumes that will confer resistance.
This sub-program works in close coordina�on with the Division of Entomology Plant Pathology Sec�on and CSIRO,the ACNFP at Murdoch University and the grain legume breeding programs at DAWA. It also con�nues to strengthen linkages with the interna�onal Medicago community as exemplified by the recent 1st Australian Medicago truncatula workshop at Ro�nest.
FUTURE
Poten�al role of transgenic approaches in the control of insect pests
Crops' incompa�bility makes conven�onal breeding approaches untenable in transferring available insect resistance from wild species into cul�vated ones. The alterna�ve recourse is to isolate and transfer alien resistance genes using gene�c transforma�on. Ar�ficial diet bioassays carried out on Maruca pod borer, pod sucking bugs and cowpea weevil indicate that these insects can be controlled by Bacillus thuringiensis crystal proteins, plant lec�ns, protease, alpha-amylase inhibitors, chi�nases and/or ribosome inac�va�ng proteins. The challenge now is to express the genes encoding these proteins in transgenic cowpea and hope that what happens in ar�ficial diets will, at least in some cases, be replicated in transgenics. Other candidate genes include enzymes encoding biochemical pathways in secondary metabolism. It can be an�cipated that useful informa�on emerging from current global genomics efforts in crop species, including model legumes, will have a bearing on cowpea improvement through gene�c engineering. What researchers need to do is develop a comprehensive pest resistance management strategy that incorporates transgenic approaches.
Sources of resistance to insects in grain legumes have been iden�fied, but these have not been used effec�vely in crop improvement. The transfer of resistance conferring genes from resistant material to regionally adapted lines has been constrained by several biological and technological factors, including a lack of sufficient informa�on about the chemical and physical nature and gene�c bases of insect resistance in plants; the need for breakthrough research and technology to overcome barriers to the development of cul�vars with mul�ple insect and disease resistance; and the requirement for new and improved technology to overcome barriers to inter-specific hybridiza�on so that resistance genes can be transferred from non-adapted to adap�ve background. Largely unexplored by pulse entomologists are the effects that different types and levels of plant resistance could have on the success or failure of
9

chemical and biological control methods. With new progress by interdisciplinary, mission-orient research team at ICARDA and ICRISAT, there is reason to be op�mis�c about the future development of insect-resistant grain legumes, especially chickpea for the developing countries. For example, entomologists and chemists have learnt much about the biochemical bases of resistance in Cicer to pod borer, Helicoverpa armigera, and the leafminer, Liriomyza cicerina and the factors governing host-selec�on behaviour of these major insect pests.
There is a need to place greater emphasis on gene�c engineering of plants for insect resistance, and iden�fica�on of molecular markers associated with resistance to insect pests. Cultural manipula�on of the crop and its environment, popula�on monitoring and pest forecas�ng, manipula�on of the crop environment to encourage the ac�vity of natural enemies, use of natural plant products and bio-pes�cides alone or in combina�on with synthe�c pes�cides, deployment of insect resistant varie�es derived through conven�onal breeding, wide hybridiza�on, or gene�c engineering, and ra�onal use of selec�ve chemicals can be exploited for pest management in food legumes.
Suggested Reading:
Belmain, S.R., Haggar, J., Holt, J. and Stevenson, P.C. (2013). Managing legume pests in sub-Saharan Africa: Challenges and prospects for improving food security and nutri�on through agroecological intensifica�on. Chatham Mari�me (United Kingdom): Natural Resources Ins�tute, University of Greenwich. 34p.
Clement, S. L., El-Din, N. E. D. S., Weigand, S. And Lateef, S. S. 1994. Research achievements in plant resistance to insect pests of cool season legumes. Euphy�ca 73: 41-50.
Fatokun, C. A., Tarawali, S. A., Singh, B. B., Kormawa, P. M. And Tamo, M. (editors). 2002. Challenges and opportuni�es for enhancing sustainable cowpea produc�on. Proceedings of the world cowpea conference III held at the Interna�onal Ins�tute for Tropical Agriculture (IITA), Ibadan, Nigeria, 4-8 September 2000. IITA, Ibadan, Nigeria.
Sharma, H C et al., (2005) Insect pest management in food legumes: The future strategies. In: Proceedings of the Fourth Interna�onal Food Legumes Research Conference, 18-22 Oct 2005, New Delhi, India.
Singh,S. R. and Emden, H. F. V. 1979. Insect pests of grain legumes. Annual Review of Entomology 24: 255-278.
Vadez, V., Berger, J. D., Warken�n, T., Asseng, S., Raanakumar, P., Rao, K. P. C., Gaur, P. M., Munier-Jolain, N., Larmure, A., Voicin A.S., Sharma, H. C. , Pande, S., Sharma, M., Krishnamurthy, L., and Zaman, M. A. 2011. Adapta�on of grain legumes to climate change: a review. Agronomy Sustainable Development. DOI 10.1007/s 13593-011-0020-6.
10

ADVANCES IN USE OF MODERN KEYS FOR PEST IDENTIFICATION
ROSHAN LALDepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
In science and biology, a “key” is a wri�en tool used for accurate iden�fica�on of such things as plants, animals, and rocks. In this guide, the key is used to iden�fy insect orders.
Keys work by a process of elimina�on, gradually narrowing down the number of possibili�es. It is important to understand that a key is much more trustworthy in proving that your insect is not A, than in proving that it is B. It might, for instance, actually be C, a species not men�oned in the key. A key does not prove anything posi�ve, it only suggests possibili�es. People used to be faddy about the sort of key they liked, but these days the arrangement that has the greatest acceptability for simplicity and directness is the dichotomous key, so-called because at each step it asks you to choose between two alterna�ves.
There are so many different kinds of insects (about 1 million) and they are divided into 30 different orders. Human beings are having probably familiar rela�onships with many common insects. These include the flies (house flies, mosquitoes, horse flies, and others) belong to the order Diptera, moths and bu�erflies are in the order Lepidoptera and beetles are in Coleoptera. Keys give the characteris�cs to accurately tell the difference between insect orders with a li�le prac�ce people can be able to recognize most insects without using a key.
However, people can iden�fy to an insect rapidly by comparing it to pictures. Pictures are a good tool, but by the using of a key provide accurate iden�fica�on.Immature stages and adult stages are o�en very different. S�ll there are is a few insect orders those are not included on the key. These orders are either extremely rare, or are not present in normal environmental condi�ons.
To use these keys there is need to know the basic anatomy of the body parts of the insects. Grasshopper as a universal dissec�ng insect can be use as a reference when the key asks you to look at certain body parts. Most of the insects have all of these body parts, but a few excep�ons (many insects don't have wings, for example, and most insects other than grasshoppers and crickets do not have ears).
External morphological keys are designed like “choose-your-own-adventure” books. As you read the key, you can choose which part of the key to read next based on what your insect looks like.
11

Insect makes the diverse bulk of species diversity on the earth. Millions of insect species exist and entomologists/zoologists have divided them into a manageable number of units called orders. The members of each insect order have arisen from their common ancestors which shares similar phenological characteris�cs and have certain biological a�ributes in common. All insect orders are not equal in species numbers; some of them having just a few hundred species while others have more than 100,000 species. The range of structural characteris�cs and biological features tends to be broader in the more species-rich orders.
Predic�ons of the biology, behavior and ecology of an insect can be made through knowing its order. An insect can be iden�fied in various ways. Comparing a specimen with a book of illustra�ons of iden�fied insects is one way andusing a printed key is another way. These computer key combines the advantages of these methods and adds a new dimension of simplicity and power to the process of iden�fica�on. The simple key is designed to iden�fy most common adult insects to order. The key has also been designed for use by advanced researchers, and others interested in entomology.
The use of gene�c markers represents a valuable addi�on to tradi�onal morphological methods of species diagnosis, an addi�on that has been deployed in a recent interac�ve trilingual iden�fica�on system to 99 species of pest Thripidae of the world (Moritz et al..2004). The polymerase chain reac�on (PCR) (Saiki et al., 1988) and PCR-based typing methods have previously provided ideal markers for iden�fica�on of difficult to recognize species (Stouthamer et al. 1999, Toda and Komazaki 2002, Borghuis et al. 2004). By its nature, PCR is an ideal tool for use in species diagnosis.It is technically simple, requiring only basic laboratory skills, minimal amounts of DNA, and once established, it is rapid, sensi�ve, and specific.
Pest iden�fica�on key to successful management:
A good pest management program starts with the early iden�fica�on. When pests are correctly iden�fied early, there are more op�ons.As crops emerge, our focus turns to protec�ng them from the threat of weeds, diseases and insects. The first step in an Integrated Pest Management is accurate iden�fica�on of the pest we are dealing with.To ensure an accurate iden�fica�on, samples must be prepared in a specific way. Here one sample type that can be submi�ed and how to submit them properly.
· Insect samples should be sent in a leak-proof vial of alcohol or vinegar.
· A sample of the damage caused to the plants should also be sent in along with the insect sample.
· If the insect is in larval stages, it should be lightly boiled while alive then placed in alcohol or it can be sent in s�ll alive.
· Moths and bu�erflies should be frozen for 30 minutes and then shipped in a vial with �ssue paper.
12

A�er complete packing samples should be send to the iden�fica�on specialists or
taxonomists for further iden�fica�on.
In biological control programs using parasitoids of the genus Trichogramma a
very important step is the iden�fica�on and the use of correct species to be released in
the field. The iden�fica�on of these wasps is difficult due to their size (0.25 mm in
length) and only specialists can reliably iden�fy specimens, using characteris�cs of the
male genitalia. In the past, Trichogramma iden�fica�on was based on body color and
seta�on as morphological parameters. Later, Nagarka� & Nagaraja (1971) found out
the taxonomic use of male genitalic characteris�cs. This leads to an improvement of the
iden�fica�on of these minute wasps. Using this technique, around 200 species have
been described (Pinto 1998). Due to the small size and few characters available, specific
iden�fica�on is s�ll difficult, par�cularly when sibling species need to be dis�nguished.
In addi�on, the specimen prepara�on for morphological iden�fica�on is �me
consuming and requires much training. At the species and intraspecific levels, the ITS1
and ITS2 regions have been o�en used as a taxonomic tool for insect iden�fica�on
(Campbell et al. 1993, Hoy 1994). The sequence and restric�on analysis of the ITS2
rDNA has been described in recent studies as a tool for Trichogramma iden�fica�on as
well (Kan et al. 1996, Pinto et al. 1997, Stouthamer et al. 1999). Using this technique, it
was possible to iden�fy seven Trichogramma species and to elaborate a molecular key
to these species recorded in Brazil.
The Interna�onal Network for the Barcoding of Invasive and Pest Species
(INBIPS) is an informal interna�onal network of researchers, research ins�tutes and
government agencies concerned with the recogni�on of invasive and pest species.
INBIPS's goal is to provide informa�on about applying DNA barcoding to invasive and
pest species iden�fica�on and to facilitate new barcoding ac�vi�es in this area.
Iden�fica�on Key to the Classes of Adult Arthropoda:
Insects represent one Class of animals within the Phylum Arthropoda. If we do
not immediately recognize an insect we may need to iden�fy some arthropods to first
determine if they are in fact insects before proceeding further. Biologists have adopted
the use of dichotomous keys to iden�fy organisms. A dichotomous key is constructed
with a series of couplets, which is a pair of phrases consis�ng of two mutually exclusive
statements. From the characters described in each couplet we can be able to choose
one of the statements as the one that best fits your specimen. Then follow the key to
the next couplet, make a decision, and con�nue un�l you iden�fy the unknown
specimen. By process of elimina�on we can arrive to at iden�fica�on. Compare the
results with pictures and other notes and in books to see if we have arrived at a likely
iden�fica�on. If we are sa�sfied with our result, proceed to the next key that we wish to
13

use and follow the same process. As we move from Class to Order to Family and perhaps
to Genus and Species we will no�ce that choices may become more difficult. This is due
to the details necessary to separate these categories. Since these keysare designed to
help us recognize insects, and to also recognize Arthropods that might be confused with
insects, we will start with an obvious and surefire couplet, 1. There are many insects
which do not appear to have wings or actually lack wings. However, many have easily
observable and func�onal wings which immediately iden�fy the creature as an insect.
1a. Wings present. —– go to 21b. Wings absent. —– go to 9
3a. Front and hind wings not similar in texture. Front wings parchment-like, shell-like or leathery or thickened. Hind wings more delicate or membranous. —– go to 4
3b. Front and hind wings similar in texture.------------ go to 7
2a. One pair of membranous wings. The insect may look like a fly or bee. (Lookcarefully for a second pair of wings hidden beneath the front pair.) —– Diptera (flies)
2b. Two pair of wings present. —— go to 3
4a. Chewing mouthparts. —– go to 5
4b. Piercing-sucking mouthparts. —– go to 6
5a. Leathery front wings, without veins, lay over body in a shell-like fashion, making a straight line between wings. Hind wings membranous. No cerc i ( ta i l - l ike appendages). —– Coleoptera (beetle)
5b. Front wings more or less parchment like with a network of veins. Hinds wings membranous, usually broad with folds like a fan. Hind leg enlarged for jumping —– Orthoptera(grasshoppers, crickets, katydids)
14

5c. Front wings more or less parchment like with a network of veins.Fla�ened bodies, thread-like antennae — Bla�aria (cockroach)
7a. Wings membranous —– go to 8
7b. Wings usually covered with powdery-like scales. Mouthparts in the form of a coiled up tube beneath the head. —– Lepidoptera(bu�erflies, moths)
7d. Wings covered with fine hairs, held roof-like over body atrest. Looks like moth with hairy wings. Long antenna eextended back over body. –Trichoptera (caddisfly)
8a. Piercing-sucking mouthparts. Beak-like mouthparts not very visible and appear to arise from the area between the front pair of legs. Wingsgenerally held roof-like over body when at rest. —– Homoptera (aphids, cicadas, lea�oppers, psyllids, tree hoppers, whiteflies)
5d. Front wings more or less parchment like with a network of veins. Front legs modified to catch and hold prey. -Mantodea(man�ds)
5e. Front wings short and leathery. Hind wings membranous. Elongated fla�ened body with dis�nct forceps-like pinchers (cerci). —– Dermaptera(earwigs)
6a. Front wings thickened at base but membranous and overlapping at �ps, so that the wings form a triangle pa�ern on the back. Beak-like mouthparts usually easily visible and appears to arise from front of head, ahead of eyes. —–Hemiptera (true bugs)
6b. Wings membranous folded tent-like at rest. Beak-like mouthparts not very visible and appear to arise from the area between the front pair of legs. —– Homoptera (lea�oppers, treehoppers)
7c. Wings very narrow and fringed with hair. Tiny insect (less than 1/8inch). Foot ending in balloon-like swelling. —– Thysanoptera (thrips)
15

8b. Front wing usually larger than hind wing.Wings may be connected. Chewing or chewing sucking mouthparts. Bee, wasp, hornet like. -Hymenoptera (bees, wasps, hornets, wingsants)
8c. Large wings in comparison to pe�te body. Wings usually with many veins and cross veins, held roof-like over body when at rest.Mouthparts chewing. No cerci (tail-like appendages). – Neuroptera (lacewing)
8d. Wings longer than body. Body whi�sh, looks like a fat ant without a slim waist.Straight and beaded antennae. —– Isoptera (winged termite)
8e. Tiny (less than 1/8 inch). Wings held roof-like over body when atrest. Veins prominent. Hair-like antennae as long or longer than body. Whi�sh body. —– Psocoptera (book lice)
8f. Insect dominated by very large eyes. Wings elongated, highly veined. Slender,needle-like body. Chewing mouth parts used to capture and consume prey inflight. —– Odonata (dragonflies and damselflies)
8g. Elongated wings fold flat over body when at rest. Long thread-like antennae. Two tail-like filaments. —– Plecoptera (stonefly)
8h. Front wings large and triangular shaped. Hind wings small and rounded. Held ver�cal over body. Two-very long tail-likefilaments. —– Ephemeroptera(Mayfly)
9a. Piercing-sucking mouthparts. —– go to 10
9b. Chewing mouthparts. —– go to 11
9c. Rasping-sucking mouthparts. Tiny (less than 1/8 inch), slender insects. Balloon-likeswelling on end of foot. O�en no�ced by scarring and deforma�on of leaves andflowers where it feeds. —– Thysanoptera(thrips)
16

10a. Bug-like with jointed beak. Mouthparts usually easily visible and appear to arise from front of head, in front of eyes. Hemiptera(true bugs)
10b. Bug-like with jointed beak. Mouth parts not very visible and appear to arise from the area betweenthe front pair of legs. Homoptera(aphids,cicadas. lea�oppers, etc.)
10c. Tiny, hard bodied, fla�ened sideways, covered with bristles that project backwards. Large hind legs designed for jumping, fast moving. Siphonaptera(fleas)
10d. Small, fla�ened parasites of animals and people. Forelegs with claw designed to grasphair or feathers. Head narrower than thorax. Anoplura(sucking lice,including head lice and body lice)
11a. Larva-like (caterpillar-like, grub-like, maggot-like). go to 12
11b. Bug-like with tail-like features (cerci,filaments). —– go to 15
11c. Ant-like. —– go to 16
11d. Dragon-like. —– go to 17
11e. Tiny, bug-like. —– go to 18
12a. Caterpillar-like: so� bodied, three pair of legs on thorax,fleshy leg-like prolegs on abdomen. —– go to 13
12c. Legless —– go to 14
12b. Grub-like: head capsule, three pair of legs on thorax, no legs on abdomen. typical of Coleoptera (beetle grub)
13a. Up to five pair of prolegs on abdomen. —– Lepidoptera(bu�erfly and moth caterpillar)
13b. Six or more pair of prolegs on abdomen. Hymenoptera(sawfly larva)
14a. Maggot-like: no legs, no head capsule, mouth-hooks. typical ofDiptera (fly maggot)
17

14b. Legless with enlarged head area. —– typical of Coleoptera (borer larva) and some Hymenoptera larva
15a. Elongated fla�ened body with dis�nct forceps-like pinchers (cerci). —– Dermaptera(earwigs)
15b. Hind legs designed for jumping. Antennae tread-like. one pairof tail-like cerci on most adults. Orthoptera(grasshoppers, crickets, katydids)
15c. Fla�ened body, long thread-like antennae. Bla�aria(cockroaches)
15d. Front legs designed for grasping and holding prey. Bodyelongated. Long, thread-like antennae. —– Mantodea(man�ds)
15e. Very elongated, s�ck-like insect. Phasmida, (walking s�ck)
15f. Two or three long, tail-like cerci. Fast moving. Small 1/4 to 1/2 inch. Zygentomaa (Silverfish, firebrats)
15g. Tiny (1-2 mm) so�-bodied insect. “Spring tail” o�en present, used to jump. —– Collembola(springtail)
16a. Three dis�nct body segments. Ant like with narrow ant waist. Elbowedantennae. —– Hymenoptera (ant)
16b. Looks like a fat ant without a narrow waist. Creamy white. Straight beaded antennae. —– Isoptera(termite)
17a. Generally dark colored with bright markings and spines. Lacking jaw as in17b. —– Coleoptera (lady beetle larva)
17b. Forward-projec�ng, curved pointed jaws designed to grasp prey, which they crush and suck out the insides of their prey.Neuroptera(antlion)
18

18a. Tiny parasite of birds. Fla�ened body. Head as wide or wider than thorax. Mallophaga(chewing or bi�ng lice)
18b. Tiny, less than 1/8 inch. Hair-like antennae as long or longer than body.Whi�sh body. —– Psocoptera(book lice)
Computer-based iden�fica�on keys:
Ingeborg et al. (2004) developed computer based key for thrips. Besides being a pest thrips itself, some species spread to spread viruses due to their feeding behaviour. Iden�fica�on of pest thrips and transmi�ed viruses as early as possible is important to implement the appropriate pest control measures. They present a computer-aided iden�fica�on key based on LucID 3.4 for pest thrips using typical character states visible under bright field or phase contrast microscope. More than 1000 computerised microscopic photomicrographs and fact sheets for each species are included in the iden�fica�on key. They also established a web-accessible data base for molecular iden�fica�on (ITS-RFLP) of thrips of any developmental stage (adult, prepupa/pupa, larva, egg). A�er DNA-extrac�on from insect or leaf material, PCR, diges�on with restric�on enzymes and gel electrophoresis, the resul�ng fragment size can be compared online with our php-data base to iden�fy the species. They are developing also in a next step a low density bio chip to iden�fy pest thrips of any developmental stage. The new biochip will iden�fy both thrips and tospoviruses transmi�ed by the insect simultaneously. The newly developed technique will be small, robust and cheap enough to be used both in lab and field. All three parts will be integrated into one computer-based iden�fica�on tool running on most computer pla�orms (MS Windows 98 or higher, Linux, Sun, MacOS X).
DNA BARCODING:
DNA barcoding is a technique that uses short DNA sequences from standardized regions of the genome as a species iden�fier. A ~ 650 base-pair region of a mitochondrial gene (COI) has been shown to be an effec�ve barcode region for many animals, (Hebert et al. 2003a; Hebert et al. 2004a, b; Hogg and Hebert 2004; Armstrong and Ball 2005; Ball et al. 2005; Barre� and Hebert 2005).
DNA BARCODING AND TAXONOMY:
DNA barcoding can also contribute to the categoriza�on of many invasive and pest groups by iden�fying gene�cally dis�nct individuals or popula�ons, flagging poten�ally new species (Hebert et al. 2004b) and unveiling cryp�c species complexes (Hebert et al. 2004a). For invasive and pest species, understanding the taxonomy and associa�ng other biological informa�on is cri�cal to preven�ng their establishment and could play a role in predic�ng the invasiveness of species.
19

Numerous thrips species cause economic damage to a range of agricultural crops in southern Africa. Much of this economic damage may be prevented by an accurate system for iden�fying pest thrips species. At present the inability to iden�fy thrips species leads both to poor control in the field and a decrease in exports due to the presence of species of quaran�ne importance. A molecular iden�fica�on tool, based on nucleo�de sequencing analysis of the mitochondrial cytochrome oxidase I (COI) gene, was developed. Nine thrips species were included in analyses: Frankliniella occidentalis, F. schultzei, Fulmekiola serrata, Haplothrips nigricornis, H. clarise�s, Heliothrips sylvanus, Scirtothrips auran�i, Synaptorthrips psoraleae and Thrips tabaci. Analysis of 163 individuals showed that each of the nine thrips species could be dis�nguished on the basis of their COI sequences. Informa�on generated was integrated with that produced previously for thrips species present in Europe and allowed a rapid, accurate and simple means of iden�fying the species of major economic importance present in southern Africa. The molecular key will provide much-needed informa�on on thrips iden�fica�on for South African pest management officers and quaran�ne purposes (Timm et al., 2008).
Effec�ve plant quaran�ne and biological control ini�a�ves require rapid and accurate iden�fica�on of exo�c and poten�ally invasive taxa that may cause high economic losses or environmental damage. The genus Scirtothrips Shull includes several species that are serious agriculturalpests, and, because of their minute size and cryp�c behavior, prone to undetected transport through interna�onal trade of plant material. Although assigning specimens to the genus Scirtothrips is straight forward using tradi�onal taxonomic methods, iden�fica�on of species is much more difficult and requires expert knowledge of the genus. Furthermore, the validity of some Scirtothrips species is ques�onable. Therefore, an easy, accurate, and highly reliable technique is desirable for Scirtothrips iden�fica�on. Here, we provide a simple molecular key based on the internal transcribed spacer regions 1 and 2 (ITS1 and ITS2) of nuclear ribosomal DNA. Individual specimens can be iden�fied by amplifica�on of their ITS1 and ITS2 regions with general primers and determining the size of the products by using standard agarose gel electrophoresis, followed in some instances by DNA diges�on with the restric�on enzymes SacII or PspOM I. The advantage of this iden�fica�on system is that non-specialists are able to quickly and cheaply iden�fy individual specimens. They iden�fied seven pest species with the molecular-based methods described. It is hoped that this system can be extended to other members of the genus as their ITS1and ITS2 sequences become available. We also provide molecular confirma�on for two new Scirtothrips species, one species from Honduras and one species from New Zealand (Paul et al., 2006).
Brunner et al. (2002) reported that treatments against pathogens or pests are o�en very specific and, as a fundamental first step, require the ability to iden�fy taxa
20

correctly and unambiguously. They used PCR amplifica�on techniques to successfully establish a molecular iden�fica�on key for economically important thrips species. A PCR amplified 433 bp long fragments of the mitochondrial COI coding gene was analyzed by automated direct sequencing and RFLP. Sequencing of 264 individual thrips represen�ng 10 named species detected 17 haplotypes. Varia�on within species was low, whereas among species varia�on was high resul�ng in an average sequence divergence of 18.6% and an average pairwise species differen�a�on (calculated as F -ST
value) of 0.9896. Two restric�on enzymes (AluI, Sau3AI) produced pa�erns that allowed unambiguous iden�fica�on of all thrips species. Sta�s�cal support for the quality of the key was given by (i) a highly significant permuta�on approach, assigning individual haplotypes to the correct species groups and (ii) a hierarchical NJ cluster analysis in which all conspecific individual sequences clustered together with maximal (100%) bootstrap support. This study has shown that the use of gene�c markers represents a valuable alterna�ve for situa�ons, such as epidemiological research, in which correct iden�fica�on with classical morphological methods is either very difficult and �me consuming or virtually impossible.
Reliable and rapid iden�fica�on of exo�c pest species is cri�cal to biosecurity. However, iden�fica�on of morphologically indis�nct specimens, such as immature life stages, that are frequently intercepted at borders is o�en impossible. Several DNA-based methods have been used for species iden�fica�on; however, a more universal and an�cipatory iden�fica�on system is needed. Shelley and Karen (2006) tested the ability of DNA "barcodes" to iden�fy species of tussock moths (Lymantriidae), a family containing several important pest species. They sequenced a 617 base pair fragment of the mitochondrial gene cytochrome c oxidase 1 for 20 lymantriid species. They used these, together with other Noctuoidea species sequences from GenBank and the Barcoding of Life Database to create a "profile" or reference sequence data set. Then they tested the ability of this profile to provide correct species iden�fica�ons for 93 addi�onal lymantriid specimens from a data set of mock unknowns. Of the unknowns, 100% were correctly iden�fied by the cytochrome c oxidase 1 profile. Mean interspecific sequence (Kimura 2-parameter) divergence was an order of magnitude greater (14%) than mean intraspecific divergence (<1%). Four species showed deeper gene�c divergences among popula�ons. They conclude that DNA barcodes provide a highly accurate means of iden�fying lymantriid species and show considerable promise as a universal approach to DNA-based iden�fica�on of pest insects.
Seunggwan et al. (2013) studied that on shiitake farms, mycophagous maggots can cause serious damage by preven�ng forma�on of the frui�ng body. Recently, these pests have significantly reduced shiitake produc�on in Korea. However, larvae and female adults cannot be iden�fied due to their lack of morphological characteris�cs. Therefore, farmers and applied entomologists are unable to determine which species is
21

the primary cause of the shiitake damage. They were collect mycophagous flies (colonized larvae) from damaged shiitake farms and subsequently iden�fied by matching iden�fied males with the cytochrome c oxidase subunit I (COI) sequences from the larvae. Divergences of the COI sequences among the species discriminated the clusters clearly, and the mycophagous pests were iden�fied as Camptomyia cor�calis and C. heterobia. Interes�ngly, these two species coexisted under the bark of shiitake oak bed logs.
The Collembola were collected from Igloolik, Cornwallis, and Somerset islands and included representa�ves from all major families found in the Arc�c. Members of 13 genera and 19 species were examined, including 4 species of the genus Folsomia and 3 species of the genus Hypogastrura. In all cases, species were successfully discriminated. Sequence divergences within species were generally less than 1%, whereas divergences between species were greater than 8% in all cases. Divergences among individuals of one species of Folsomia were much higher (up to 13%), but this likely represents the presence of an undescribed sibling species. Conclusion was that DNA barcoding is a powerful tool for iden�fying species of Collembola and should regularly be useful as a complement to tradi�onal, morphological taxonomy (Hogg and Hebert, 2004).
With almost 40000 species, the spiders provide important model systems for studies of sociality, ma�ng systems, and sexual dimorphism. However, work on this group is regularly constrained by difficul�es in species iden�fica�on. DNA-based iden�fica�on systems represent a promising approach to resolve this taxonomic impediment, but their efficacy has only been tested in a few groups. Demonstra�on was carried out and concluded that sequence diversity in a standard segment of the mitochondrial gene coding for cytochrome c oxidase I (COI) is highly effec�ve in discrimina�ng spider species. A COI profile containing 168 spider species and 35 other arachnid species correctly assigned 100% of subsequently analyzed specimens to the appropriate species. In addi�on, no overlap between mean nucleo�de divergences at the intra- and inter-specific levels was found. Results establish the poten�al of COI as a rapid and accurate iden�fica�on tool for biodiversity surveys of spiders (Barre� and Hebert, 2005).
Barr et al., (2006) reported that the predominantly Afrotropical fruit fly genus Cera��s contains many species of agricultural importance. Consequently, quaran�ne of Cera��s species is a major concern for governmental regulatory agencies. Although diagnos�c keys exist for iden�fica�on of all described Cera��s species, these tools are based on adult characters. Flies intercepted at ports of entry are usually immatures, and Cera��s species cannot be diagnosed based on larval morphology. To facilitate iden�fica�on of Cera��s pests at ports of entry, they explores the u�lity of DNA-based diagnos�c tools for a select group of Cera��s species and related tephri�ds, some of
22
which infest agriculturally important crops in Africa. The applica�on of the polymerase chain reac�on–restric�on fragment length polymorphism (PCR–RFLP) method to analyse three mitochondrial genes (12S ribosomal RNA, 16S ribosomal RNA, and NADH-dehydrogenase subunit 6) is sufficient to diagnose 25 species and two species clusters. PCR analysis of the internal transcribed spacer region 1 (ITS-1) is able to dis�nguish three of the five species le� unresolved by mitochondrial DNA analysis.
Americo et al. (2001) Using the ITS2 sequences of the ribosomal DNA together with some restric�on enzymes, a simple and precise molecular key to some Brazilian species of Trichogramma was created. This key is very easy to make and quick results can be obtained on the iden�fica�on of this minute parasitoid (0.25 mm). The five individuals used for DNA extrac�on was enough to obtain good PCR products for cloning and for sequencing. All the sequences was sent to the Genbank and have the accession numbers: AF282222-AF282239. Using the sizes of the PCR products with the enzymes EcoRl, Mse l, Pvu I and Nru I, all the species was dis�nguished in this work. The key made shows that the sequence size of the ITS2 region of the rDNA of some species of Trichogramma is useful and can be used as a tool to quickly iden�fy species. It is important to know that this key fits well for the popula�ons studied in this work. More samples are needed in order to construct a complete key in the future with all other Trichogramma species from Brazil. With a molecular key of all Brazilian species as well as all the sequences, it can be easily known if new species are occurring in Brazil just comparing the recorded sequences of each species.
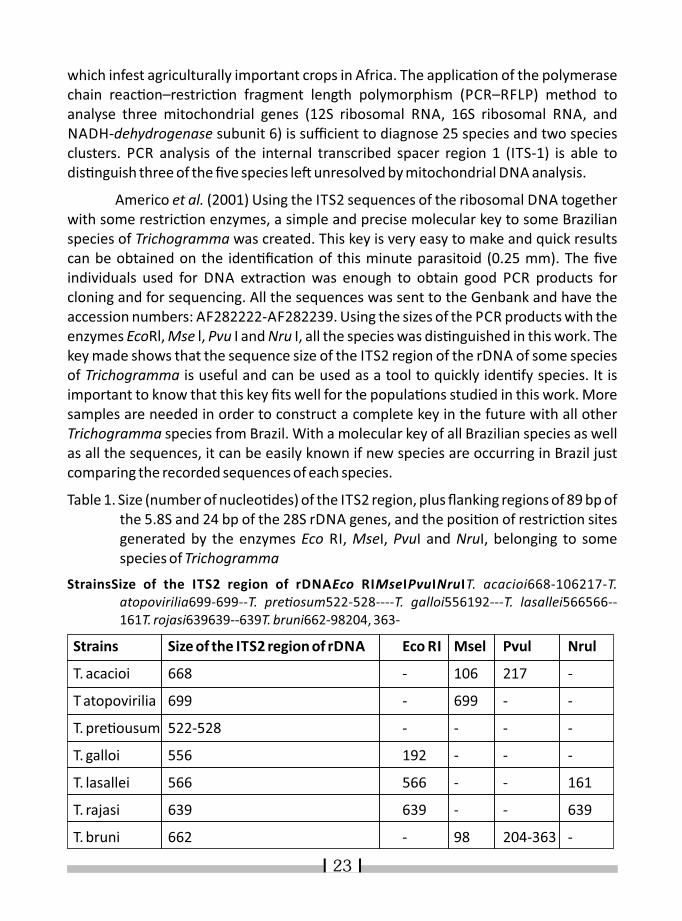
Table 1. Size (number of nucleo�des) of the ITS2 region, plus flanking regions of 89 bp of the 5.8S and 24 bp of the 28S rDNA genes, and the posi�on of restric�on sites generated by the enzymes Eco RI, MseI, PvuI and NruI, belonging to some species of Trichogramma
StrainsSize of the ITS2 region of rDNAEco RIMseIPvuINruIT. acacioi668-106217-T. atopovirilia699-699--T. pre�osum522-528----T. galloi556192---T. lasallei566566--161T. rojasi639639--639T. bruni662-98204, 363-
Strains Size of the ITS2 region of rDNA Eco RI Msel Pvul Nrul
T. acacioi 668 - 106 217 -
T atopovirilia 699 - 699 - -
T. pre�ousum 522-528 - - - -
T. galloi 556 192 - - -
T. lasallei 566 566 - - 161
T. rajasi 639 639 - - 639
T. bruni 662 - 98 204-363 -
23

Table 2. Molecular key to seven Brazilian species of Trichogramma based on the PCR
product plus flanking regions and the enzymes Eco RI, Msel, PvuI and NruI
1. PCR product bigger than 660 bp*........................................................................................... 21' PCR product smaller than 660 bp........................................................................................... 3
2. PCR product cut by the enzyme MseI..................................................................................... 42' PCR not cut by the enzyme MseI .........................................................................T. atopovirilia
3. PCR product bigger than 550 bp ............................................................................................ 53' PCR product between 522-528 bp ........................................................................T. pre�osum
4. PCR product cut once by the enzyme PvuI .................................................................T. acacioi4' PCR product cut twice by the enzyme PvuI ...................................................................T. bruni
5. PCR product cut by the enzyme Eco RI .........................................................................T. galloi5' PCR product not cut by the enzyme Eco RI............................................................................. 6
6. PCR product cut once by the enzyme NruI .................................................................T. lasallei6' PCR not cut by the enzyme NruI ...................................................................................T. rojasi* bp = base pairs
Farris et al. (2010) studied that the thrips Scirtothrips dorsalis Hood (Thysanoptera:
Thripidae) is an invasive pest that poses a significant economical threat to U.S.
agriculture and trade. DNA sequence data and polymerase chain reac�on (PCR) was
u�lized to develop a molecular diagnos�c marker for S. dorsalis. The DNA sequence
varia�on from the internal transcribed spacer 2 (ITS2) region of nuclear ribosomal DNA
(rDNA) was analyzed from various thrips species, including S. dorsalis. A primer set and
polymerase chain reac�on cycling parameters was designed for the amplifica�on of a
single marker fragment of S. dorsalis ITS2 rDNA. Specificity tests performed on ten
thrips species, efficacy tests performed on fi�een S. dorsalis popula�ons, and tests on
primer sensi�vity and robustness all demonstrated the diagnos�c u�lity of this marker.
This diagnos�c PCR assay provides a quick, simple, and reliable molecular technique to
be used in the iden�fica�on of S. dorsalis.
� Parasitoid detec�on and iden�fica�on is a necessary step in the
developmentand implementa�on of fruit fly biological control strategies employing
parasitoid augumen�ve release. In recent years, DNA-based methods have been used
to iden�fynatural enemies of pest species where morphological differen�a�on is
problema�c.Molecular techniques also offer a considerable advantage over tradi�onal
morphologicalmethods of fruit fly and parasitoid discrimina�on as well as within-host
parasitoididen�fica�on, which currently relies on dissec�on of immature parasitoids
from the host,or lengthy and labour-intensive rearing methods (Cheryl et al., 2012).
24

QPAIS: A Web-based expert system for assisted iden�fica�on of quaran�ne stored insect pests:
� Stored insect pests can seriously depredated stored products causing worldwide economic losses. Pests enter countries travelling with transported goods. Inspec�on and Quaran�ne ac�vi�es are essen�al to prevent the invasion and spread of pest s. iden�fica�on of quaran�ne stored insect pests is an important component of the China's Inspec�on and Quaran�ne procedure, and it is necessary not only to iden�fy weather the species captured is an invasive species, but determine control procedures for stored insect pests. With the development of informa�on technologies, many expert systems that aid in the iden�fica�on of agricultural pests have been developed. Expert system for the iden�fica�on of quaran�ne stored insect pests are rare and are mainly developed for stand-alone PCs. Huang et al. (2009) describe the development of a web-based expert system for iden�fica�on of quaran�ne stored insect pests as part of
thChina 11 Five-year Na�onal Scien�fic and Technological Support Project. Based on user needs, txtual knowledge and images were gathered from literature and expert interviews. ASP.NET, C# and SQL language were used to program the system. Improvement of iden�fica�on efficiency and flexibility was achieved using a new inference method called characteris�c-select-based spa�al distance method. The expert system can assist iden�fying 150 species of quaran�ne stored insect pests and provide detailed informa�on for each species. The expert system has also been evaluated using two steps: system tes�ng and iden�fica�on tes�ng. With 85% rate of correct iden�fica�on and high efficiency, the system evalua�on shows that this expert system can be used in iden�fica�on work of quaran�ne stored insect pests.
� Jianigning et al. (2012) designed a new automa�c iden�fica�on system to iden�fy insect's specimen images at the order level. Several rela�ve features were designed according to the methods of digital image progressing, pa�ern recogni�on and the theory of taxonomy. Ar�ficial neural networks (ANNs) and a support vector machine (SVM) are used as pa�ern recogni�on methods for the iden�fica�on tests. During tests on nine common orders and sub-orders with an ar�ficial neural network, the system performed with good stability and accuracy reached 93%. Results from tests using the support vector machine further improved accuracy. They also did tests on eight- and nine-orders with different features and based on these results they compare the advantages and disadvantages of their system and provide some advice for future research on insect image recogni�on.
Automated Insect Iden�fica�on through Concatenated Histograms of Local Appearance Features:
� Larios et al. (2008) describes a computer vision approach to automated rapid-
throughput taxonomic iden�fica�on of stonefly larvae. The long-term objec�ve of this
research is to develop a cost-effec�ve method for environmental monitoring based on 25

automated iden�fica�on of indicator species. Recogni�on of stonefly larvae is
challenging because they are highly ar�culated, they exhibit a high degree of
intraspecies varia�on in size and color, and some species are difficult to dis�nguish
visually, despite prominent dorsal pa�erning. The stoneflies are imaged via an
apparatus that manipulates the specimens into the field of view of a microscope so that
images are obtained under highly repeatable condi�ons. The images are then classified
through a process that involves (a) iden�fica�on of regions of interest, (b)
representa�on of those regions as SIFT vectors (c) classifica�on of the SIFT vectors into
learned “features” to form a histogram of detected features, and (d) classifica�on of the
feature histogram via state-of-the-art ensemble classifica�on algorithms. The steps (a)
to (c) compose the concatenated feature histogram (CFH) method. They apply three
region detectors for part (a) above, including a newly developed principal curvature-
based region (PCBR) detector. This detector finds stable regions of high curvature via a
watershed segmenta�on algorithm. They compute a separate dic�onary of learned
features for each region detector, and then concatenate the histograms prior to the
final classifica�on step. They evaluate this classifica�on methodology on a task of
discrimina�ng among four stonefly taxa, two of which, Calineuria and Doroneuria, are
difficult even for experts to discriminate. The results show that the combina�on of all
three detectors gives four-class accuracy of 82% and three-class accuracy (pooling
Calineuria and Doro-neuria) of 95%. Each region detector makes a valuable
contribu�on. In par�cular, our new PCBR detector is able to discriminate Calineuria and
Doroneuria much be�er than the other detectors.
Prototype System of Insects Iden�fica�on based on Computer Vision:
� Zhang and Fareed (2013) presents the insects' image preprocessing, feature
extrac�on and target recogni�on. The objec�ve of this research was to design a new
type of insect mul�media databases. Implementa�on of three types of basic feature
extrac�on methods: color features of image, texture features and shape features,
mainly the color vector, Gabor-wavelet transform, Fourier, GLCM, etc was done and
then experimental results were analyzed. Authors suggested a new assisted GPS
feature which can improve the retrieval efficiency of large insect's dataset. Designed
and implemented a mul� feature asynchronous insect retrieval system, mul�-feature
fusion research retrieval features normaliza�on and similarity measure algorithm.
Implemented insect iden�fica�on sample tested in Mat lab and Visual C++
environment and the test results were analyzed. The proposed database will include
audio, video and special image (such as the infra red image). This research will add an
assistant feature of GPS applica�ons, and this feature will effec�vely enhance the
efficiency of retrieval in large dataset
26

Suggested Reading:
Americo, I., Ciociola, J. R., Roberto, A., Zucchi and Richard Stouthamer (2001).Molecular Key to Seven Brazilian Species of Trichogramma (Hymenoptera: Trichogramma�dae) Using Sequences of the ITS2 Region and Restric�on Analysis. Neotropical Entomology, 30(2): 259-262.
Armstrong, K.F. and Ball, S.L. (2005).“DNA barcodes for biosecurity: invasive species iden�fica�on,” Philosophical Transac�ons of the Royal Society London B 360:1813–1823.
Ball, S.L., Hebert, P.D.N., Burian, S.K. and Webb, J.M. (2005). “Biological iden�fica�ons of mayflies (Ephemeroptera) using DNA barcodes,” Journal of the North American Benthological Society 24: 508–524.
Barr, N.B., R.S. Copeland, M. De Meyer, D. Masiga, H.G. Kibogo, M.K. Billah, E. Osir, R.A. Wharton and B.A. McPheron. (2006). Molecular diagnos�cs of economically important Cera��s fruit fly species (Diptera: Tephri�dae) in Africa using PCR and RFLP analyses. Bulle�n of Entomological Research, 96(5): 505-521.
Barre�, R.D.H., and Hebert, P.D.N. (2005). “Iden�fying spiders through DNA barcodes,” Canadian J Zoology 83: 481–491.
Borghuis, A., J. D. Pinto, G. R. Platner, and R. Stouthamer, (2004). Par�al cytochrome oxidase II sequences dis�nguish the sibling species Trichograma minutum Riley and Trichogram aplatneri Nagarka�. Biol. Control 30: 90-94.
Brunner, P. C., C. Fleming, J. E. Frey (2002). A molecular iden�fica�on key for economically important thrips species (Thysanoptera: Thripidae) using direct sequencing and a PCR-RFLP-based approach. Agricultural and Forest Entomology, 4(2): 127–136:
Campbell, B.C., J. D. Steffen-Campbell and J. H. Werren. (1993). Phylogeny of the Nasonia species complex (Hymenoptera: Pteromalidae) inferred from an internal transcribed spacer (ITS2) and 28S rDNA sequences. Insect Mol. Biol. 2: 225-237.
Cheryl, Jenkins, Toni, A. Chapman, Jessica, L. Micallef and Olivia, L. (2012).Reynolds Molecular Techniques for the Detec�on and Differen�a�onof Host and Parasitoid Species and the Implica�ons forFruit Fly Management. Insects, 3: 763-788.
Farris R.E, Ruiz-Arce R, Ciomperlik, M., Vasquez, J. D. and Deleón R. (2010).Development of a ribosomal DNA ITS2 marker for the iden�fica�on of the thrips, Scirtothrips dorsalis. J. Insect Sci. 10(26): 15.
27

Hebert, P.D.N., Cywinska, A., Ball, S.L., and deWaard, J.R. (2003a).“Biological iden�fica�ons through DNA barcodes,” Proceedings of the Royal Society London B 270: 313–321.
Hebert, P.D.N., Penton, E.H., Burns, J.M., Janzen, D.H., and Hallwachs, W. (2004a). “Ten species in one: DNA barcoding reveals cryp�c species in the neotropical skipper bu�erfly Astraptesfulgerator,” Proceedings of the Na�onal Academy of Science 101: 14812–14817.
Hebert, P.D.N., Stoeckle, M.Y., Zemlak, T.S., and Francis, C.M. (2004b). “Iden�fica�on of birds through DNA barcodes,” Public Library of Science, Biology 2: e312.
Hogg, I.D. and Hebert, P.D.N. (2004). “Biological iden�fica�ons of springtails (Collembola: Hexapoda) from the Canadian arc�c, using mitochondrial DNA barcodes,” Canadian J. Zoology 82: 1–6.
Hoy, M. (1994). Insect molecular gene�cs.An introduc�on to principles and applica�ons. Academic Press, Inc. San Diego, California, 546.
Huang, H., Rajo�e, E.G., Li, Z., Chen, K. and Zhang, S. (2009). In IFIP Interna�onal federa�on for Informa�on Processing, Volume 293, Computer and Compu�ng Technologies in Agriculture II, volume I, eds. D. Li, Z. Chunjiang, (Boston: Springer ), pp. 701-714.
Ingeborg, Schuetz, Antje, Breitenstein and Gerald, Moritz (2004). Computer-based iden�fica�on key for pest thrips and tospoviruses by use of LucID 3.4, ITS-RFLP and low-density Bio Chip technology. J. Insect Sci., 12: 45-49.
Jiangning, W., Cong�an, L., Liqiang, Ji. And Aiping, L. (2012). A new automa�c iden�fica�on system of insect images at the order level. Knowledge-Based System, 33: 102-110.
Kan, F. J. P. M. van, I. M. M. S. Silva, M. Schilthuizen, J. D. Pinto and R. Stouthamer (1996). Use of DNA-based methods for the iden�fica�on of minute wasps of the genus Trichogramma. Proc. Exp. Appl. Entomol., N.E.V. 7: 233-237.
Larios, N., H. Deng, W. Zhang, M. Sarpola, J. Yuen, R. Paasch, A. Moldenke, D. A. Lytle, S. R. Correa, E. N. Mortensen, L. G. Shapiro, and T. G. Die�erich. (2008). Automated insect iden�fica�on through concatenated histograms of local appearance features: feature vector genera�on and region detec�on for deformable objects. Machine Vision and Applica�ons 9(2): 105-124.
Moritz, G., L. A. Mound, D. C. Morris, and A. Goldarazena.(2004). Pest thrips of the world, visual and molecular iden�fica�onof pest thrips. Interac�ve CD ROM distributedby Lucid, University of Queensland, Australia.
28

Nagarka�, S. and Nagaraja, H. (1971). Redescrip�on of some known species of Trichogramma, showing the importance of the male genitalia as a diagnos�c character. Bul. Entomol. Res. 61: 13-31.
Paul F. Rugman-jones, Mark S. Hoddle, Laurence A. Mound, and Richard Stouthamer (2006).Molecular Iden�fica�on Key for Pest Species of Scirtothrips (Thysanoptera: Thripidae). J. Econ. Entomol. 99(5): 1813-1819.
Pinto, J. D. (1998). The systema�cs of the North American species of Trichogramma (Hymenoptera: Trichogramma�dae). Mem. Entomol. Soc. Wash. Allen Press Inc., Washington D.C. 22, 287.
Pinto, J. D., R. Stouthamer and G. R. Platner (1997). A new cryp�c species of Trichogramma (Hymenoptera Trichogramma�dae) from the Mojave desert of California as determined by morphological, reproduc�ve and molecular data. Proc. Entomol. Soc. Wash. 99: 238-247.
Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi,G. T. Horn, K. B. Mullis, and H. A. Erlich (1988). Primer directed enzyma�c amplifica�on of DNA with a thermostable DNA polymerase. Science (Wash. DC) 239: 487-491.
Seunggwan Shin, Sunghoon Jung, Heungsik Lee, Seunghwan Lee (2013). Molecular iden�fica�on of dipteran pests (Diptera: Sciaroidea) from shiitake mushroom. Molecular Ecology Resources, 13(2): 200-209.
Shelley L Ball, Karen F Armstrong (2006). DNA barcodes for insect pest iden�fica�on: a test case with tussock moths (Lepidoptera: Lymantriidae). Canadian J. Forest Res., 36(2): 337-350.
Stouthamer, R., J. Hu, F.J.P.M. van Kan, G. R. Platner, and J. D. Pinto (1999). The u�lity of internally transcribedspacer 2 DNA sequences of the nuclear ribosomal genefor dis�nguishing sibling species of Trichogramma. Biocontrol 43: 421-440.
29

MANAGEMENT STRATEGIES FOR INSECT PESTS OF COWPEA AND CLUSTERBEAN
S.P. SINGHDepartment of Entomology,
CCS Haryana Agricultural University, Hisar – 125 004, India.
Cowpea (Vigna unguiculata L. Walp.) and clusterbean (Cyamopsis tetragonolaba L. Taub) are major arid legume crops covering about 70 per cent area of arid crops in India during kharif season. Both cowpea and clusterbean are grown for grain, green pods and fodder purposes. Insect pests are major constraints in reducing the produc�on and produc�vity of these crops. Integrated pest management (IPM) approach has been accepted worldwide for achieving sustainability in agriculture. This type of approach has numerous advantages like pes�cides free produces, safety to environment and beneficial for natural enemies and low input based crop produc�on programme especially for arid legume crops. Therefore, IPM is an eco-friendly approach for pest management by using all the available methods and techniques such as cultural including host plant resistance, biological, mechanical and chemical.
A number of insect pests inflict moderate to severe quan�ta�ve and qualita�ve losses to these crops. About 55 insect pests a�ack both the crops among which lea�opper, Empoasca kerri Pruthi, aphid, Aphis craccivora Koch and whitefly, Bemisia tabaci Gennadius are most serious pests adversely affec�ng the vegeta�ve growth and yield of the crop. The pod borer, Helicoverpa armigera (Hubner) feeds on the developing grains inside the pods, thus reducing grain yield significantly. Pest wise informa�on on important pests of cowpea and clusterbean crops is described here under.
a) Aphid, Aphis craccivora Koch: Aphids are locally known as chepa, mahun or tela. They are black in colour with green �nge, small, viviparous insects, measuring about one tenth to one-eight of an inch in length. Colonies of aphids are found on the stems, leaves, and pods of cowpea and guar plants. Nymphs and adults suck sap from underside of the leaves, top shoots and stems, as a result of which the plants become discoloured and weak. Infesta�on in the early stage causes stun�ng of the plants as well as reducing the vigour. The female may produce 8-30 young ones in a life span of 10-12 days. The nymphs transform into adults in 5-8 days a�er passing through four moults. The apterous female starts producing broods within 24 hours of a�aining that stage. The pest breeds throughout the year. The aphids secrete honeydew on which sooty moulds develop. Females increase at an enormous rate and spread over a big area in a very short �me. The pest is most ac�ve in early stages of the crop and damages young plants. This pest also a�acks sorghum and other vegetable crops. The aphids appear on
30

the cowpea and guar from July to September. There are several genera�ons during the rainy season. These insects generally thrive best in damp weather and when moist wind blows. The economic threshold is 10 per cent infested plants. The avoidable losses due to this pest in cowpea and cluster bean seed crops are about 20 per cent.
Management approaches: Early sowing of cowpea and clusterbean crops escapes the aphids incidence. Grow improved / tolerant cowpea and clusterbean varie�es, CS 88, GC 3 and HG 365, HG 563, respec�vely. Intercropping with non-leguminous crops such as sorghum and maize reduces the aphids incidence in cowpea and clusterbean crops. Removal of aphid infested tops of the plants, at early stage of aphid infesta�on. The fog, frost, rain, severe heat and cold are the main natural mortality factors of aphids. The predators viz., Coccinellids, syrphids and lace wing and some entomogenous fungi viz., Entomophthora and Cephalosporium contribu�ng a lot in checking the aphid popula�on. If 10 per cent plants infested, crop may be protected from aphids by spraying 0.05 per cent malathion.
b) Lea�opper, Empoasca kerri Pruthi : Lea�opper, commonly known as jassid is a
small size about 12 mm long, delicate and yellowish green insect. Both nymphs and
adults suck sap from the leaves, which in severe cases of a�ack turn yellow to reddish
brown. The a�acked leaves later curl up, become distorted and fall down. The nymphs
and adults prefer shady areas and generally remain on the lower surface of the leaves. A
female hopper lays 30 to 50 yellowish eggs singly on the underside of the leaves,
embedding them into the leaf veins. The nymphs emerge from the eggs in 4 to 10 days,
which are wedge shaped and are very ac�ve. Nymphal period lasts for 10 to 20 days.
These are light green and fast moving in zig-zag manner. The adult stage lasts from 5 to 7
days. The insect passes through 8 to 10 genera�ons in a year. The pest appears on these
crops during the rainy season and a�acks through out the growth stage. Besides
cowpea and guar, it also a�acks berseem, lucerne, soybean, potato and tomato crops.
Economic threshold for lea�opper is 2 nymphs per leaf based on 30 leaves or 20 per
cent fully developed leaves start curling. The avoidable losses due to this pest in cowpea
and clusterbean seed crops are about 30 per cent.
Management approaches: Early sowing of cowpea and clusterbean crops (up to first
week of June) escapes lea�opper incidence. Grow resistant cowpea and clusterbean
varie�es, CS 88, GC 8927 and HG 75, HG 884, respec�vely. Removal and destroy the
weed as they provide food and shelter to the pest. Use balanced fer�lizers based on soil
test. Excessive use of nitrogenous fer�lizers a�racts more pests. If nymphal popula�on
exceeds more than one per leaf, spray the crop with 0.05 per cent malathion.
c) Whitefly, Bemisia tabaci (Gennadius): The female lays eggs singly on the underside
of the leaves, averaging 120 eggs per female. The eggs are stalked and light yellow at
31

first, turn brown later on. They hatch in 3-5 days. Both nymphs and adults feed on cell
sap from under surface of the leaves and ul�mately the crop yield is reduced
considerably. The quality of fodder as well as grains is also reported to deteriorate.
Winged adults are 1.0 to 1.5 mm long and their yellowish bodies are slightly dusted with
white waxy powder. They have two pairs of pure white wings. The nymphs grow into
three stages to form the pupae with in 9-14 days. In 2-8 days, the pupae change into
adults. The life cycle is completed in 15-120 days and 10 genera�ons completed in a
year. Moreover, white fly acts as vector of number virus diseases. They also excrete
honeydew interfering with the photosynthesis, on which black mould fungi grow. This
insect appears more on cowpea and guar crops in dry and hot weather condi�ons.
Economic threshold for white fly is 6 adults per leaf or 20 nymphs per leaf. The
avoidable losses due to this pest in cowpea and clusterbean seed crops are about 30 per
cent (Singh, 2003).
Management approaches: Grow improved / tolerant varie�es of cowpea and
clusterbean, CS 88, HC 98-46 and HG 75, HG 94, respec�vely. Use s�cky traps or yellow
sheet coated with grease helps mass trapping white fly adults. Removal and destroy the
weeds as they provide food and shelter to the pest. The parasitoids, Encarsia formosa
and Eretmocerus mundus contribu�ng a lot in checking the white fly popula�on. Spray
the crop with 0.03 per cent oxydemeton methyl or dimethoate (Singh, 2004).
d) Pod Borer, Helicoverpa armigera (Hubner): It is a serious polyphagous pest. The female lays spherical yellowish eggs singly on tender parts of the plants. A single female may lay as many as 740 eggs in four days. The incuba�on period is 2-4 days. The young larva feeds on the foliage for some �me and later damages the flower buds, pods and feed on the developing grains and can reduce the seed yield up to 60 per cent. A single larva may destroy 30-40 pods before it reaches maturity. Characteris�cally, while feeding, the head will be thrust inside leaving rest of the body out side. Cannibalism among larvae is common. The full-grown larvae are greenish with dark broken gray lines along the side of the body measuring about 40 mm long. They come out of the pods and pupate in an earthen cell. The pupa is dark brown and has a sharp spine at the posterior end. The pupal period lasts 8-15 days in ac�ve season, which may be prolonged in winter. The moth is large and brown with a V-shaped speck and dull black border on the hind wings. There may be as many as 8 genera�ons in a year. The pest appears on these crops during the rainy season and a�acks through out the growth stages of the crop. Besides cowpea and guar, it also a�acks berseem, lucerne and tomato crops. Economic threshold for pod borer is 0.5 larva per plant (10 larvae per 20 plants) or 5 per cent pods infested. The avoidable losses due to this pest in cowpea and clusterbean seed crops are about 25 per cent.
32

Management approaches: Grow early maturing, short dura�on and tolerant varie�es of cowpea and clusterbean, CS 88, GC 13 and HG 365, HG 563, respec�vely. Removal and destroy the crop stubbles/remnants as they serve as ini�al source of infesta�on. Removal and destroy the weed as they provide food and shelter to the pest. Use balanced fer�lizers based on soil test. Excessive use of nitrogenous fer�lizers a�racts more pests. Use pheromone and light traps to monitor and collect male moth popula�on. Release egg parasitoid, Trichogramma chilonis @ 3 lakh adults/ha and larval parasitoid, Campole�s chloridae Uchida to manage this pest in field condi�ons. Spray the crop with 0.07 per cent carbaryl to control this pest effec�vely and second spray can be done 10 days a�er, if pest persists. Neem based insec�cide like nimbecidine is also effec�ve, which acts as an�oviposi�on (repellant) and an�feedant against this pest.
Integrated pest management module in cowpea and clusterbean crops: Grow early maturing, short dura�on varie�es of cowpea and clusterbean during June so as to avoid the heavy incidence of insect pests. Destroying the stubbles, weeds and other alternate hosts by deep ploughing the fields a�er harvest may reduce the poten�als of pest carryover. Use balanced fer�lizers based on soil test. Excessive use of nitrogenous fer�lizers a�racts more pests. Release of parasitoids to manage different pests at proper �me. Use resistant / tolerant cowpea and clusterbean varie�es, par�cularly CS 88 and HG 75, respec�vely which having mul�ple resistance against majority of the pests.
Arora, R.K. and Kashyap, R.K. 2003. Insect pests. Pages: 149-169. In Guar in India. (Eds. D.Kumar and N.B. Singh) Scien�fic Publishers (India), P.O. Box 91, Jodhpur, Rajasthan, India.
Faleiro. J.R., Singh, K.M., and Singh, R.N. 1986. Pest complex and succession of insect pests in cowpea Vigna unguiculata (L.) Walp. Indian J. Ent. 48 (1): 54-61.
Singh, S.P. 2000. Insect pest management in forage crops. Proc. Advanced Training Course on Recent Advances in Integrated Pest Management. 1-21, December, 2000, Department of Entomology, CCS Haryana Agricultural University, Hisar, India.pp. 283-292.
Singh, S.P. 2003. Insect – pests management in cowpea. Indian Farming, 72(11): 23-26.
Singh, S.P. 2003. Insect pest management in cowpea and clusterbean crops. Pages 448-452 in Advances in Arid Legumes Research. (Eds. A. Henry, D. Kumar, N.B. Singh), Indian Arid Legumes Society, CAZRI, Jodhpur, Rajasthan, India.
Singh, S. P. 2004. Pest management strategies in clusterbean. Pages: 112-120 in Guar (Eds. J.V. Singh and B.S. Dahiya), Forage Research Society, CCS HAU, Hisar, India.
Singh, S.P. and Chhillar, B.S. 2010. Insect-pest management in legume forage crops.
33

pages: 241-262 in Forage Legume (Eds. Jai Vir Singh, B. S. Chhillar, B.D. Yadav and U.N. Joshi), Scien�fic Publishers (India), Jodhpur, Rajasthan.
Singh, S.P. and Sangwan, R.S. 2000. Rela�ve suscep�bility of some cowpea genotypes to lea�opper and whitefly. Haryana J. Hort. Sci., 29(3&4): 261-263.
Verma, A.N., Singh, S.P. and Gandhi, S.K. l992. Role of plant protec�on in forage crop produc�vity. Pages: 169-177 In Strategy for Forage Produc�on and Improvement. (Eds: R.S. Paroda, Arora, S.K., Faroda, A.S.). Indian Society of Forage Research, H.A.U., Hisar, India.
34

MANAGEMENT STRATEGIES FOR INSECT PESTS OF PEA AND FRENCH BEAN
S. S. SHARMADepartment of Entomology
CCS Haryana Agricultural University Hisar - 12001, India
Insect pests of pea Peas (Pisum sa�vum L)
Pea is a cool season important vegetable crop. It has sweet taste and starchy in nature. It contains sugar, starch and health protec�ve phytonutrients as polyphenol and colemestrol. It is cul�vated in Syria, Turkey, Jordan, Egypt, Georgia, Pakistan and India. It is a�acked by a variety of insects but the major insect pests have been described below:
White grubs: Holotrichia longipennis, Brahmina coriacea, Maladera furcicauda and
Anomala sp. F: Scarabidae O: Coleoptera
These species of white grubs are found in different parts of Himachal Pradesh. They are polyphagous pests. The white grubs completely cut the roots of the plants and the damaged plants show varying degree of yellowing, browning and wil�ng. They can be pulled out easily. The patchy areas can be seen in the infested fields.
Loss: In Sangla Valley of Kinnaur (H.P.) white grubs cause 20-25 per cent plant mortality in off season crop in the month of June-July. The damage remains most serious in fields located in the vicinity of apple orchards.
Management: Timely sowing reduces the damage of white grubs and delay of sowing increases the incidence of L. s�gma white grubs. 2) Early ploughing of the field exposes the grubs to predators like birds and sun. 3) Apply chlorpyriphos 20 EC @ 800 g/ha. 4) Mix the insec�cide in sand and broadcast in the field prior to harves�ng so as to mix it thoroughly in the soil. 5) Treat the pea seeds with quinalphos 25 EC in chlorpyriphos 20 EC @ 4 ml/kg seed.
Leaf miner: Chromatomyia hor�cloa (Gourea) F: Agromyzidae O: Diptera
Host plants: Cruciferous plants, pea, linseed, potato, leguminous crops, cucurbits, tomato and le�uce etc.
Iden�fica�on and life cycle:
Adult (Fly) : The adult is a �ny black fly with transparent two wings. It has grayish-black mesonotum and yellowish frons. These flies become ac�ve from December to April or May. The adults emerge in the beginning of December and a�er ma�ng it starts egg laying singly in leaf �ssues.
35

Egg: The eggs are deposited in leaf �ssue. The egg period is 2-3 days
Larvae (maggots): The maggots are legless creamy in colour and legless. The larval (maggot) period is 5 days. Just a�er hatching the maggot makes mine, in the leaf in zigzag fashion.
Pupa: Pupa�on takes place in the mines or galleries made in the leaf. Pupil period lasts for 6 days.
The insect completes one genera�on in 13-14 days and pass through several genera�ons during the period of its ac�vity.
Nature of damage: The larvae (maggots) feed between the lower and upper epidermis of the leaf by making zig-zag tunnels/mines. The infested leaves show prominent whi�sh shiny tunnels/ streaks against the green background. When the damaged leaves are held against bright light, the minute slender larvae can be seen feeding within the tunnels/mines. The large numbers of tunnels/mines made by the larvae interfere with photosynthesis and proper growth of plants.
Management: Grow leafless varie�es. Spray one litre of dimethoate 30EC in 250 litres of water per ha and repeat spray at 15-day interval. A wai�ng period of 20 days should be observed for picking of pods.
Pea aphid, (Acyrthosiphon pisum Harris) F: Aphididae O: Homoptera
Host plant: Field peas, alfalfa, clover, len�l, faba beans, vetches, sweet clover, sweet pea, trefoil and dry beans.
Iden�fica�on and life cycle:-
Egg: � Winged male and female mate and lay eggs on leaves and stem.
Nymph: Nymphs are wingless, green, small but resembled to adults.
Adult: They are so� bodied insects, slow moving, light to dark green coloured, pear shaped and about 3 mm in length. It survives 5-50 days.
Nature of damage: Both the nymphs and adults suck the cell sap from the lower side of the leaves. The damaged leaves turn yellow. The plant remains stunted resul�ng the lesser number of pods per plant.
ET: 3-4 aphids/stem �p (Bommarco, 1991) or 2-3 aphids/8 inch plant �p or 9-12 aphid / sweep.
Management: Clip off the twigs having aphid colonies and destroy them. Chemical control same as prescribed for leaf miner. Early maturing cul�vars escape from its a�ack parasitoid like Aphidius ervi and Beauveria fungus in warm weather reduce its
36

popula�on. Save the predators like lady beetles, larvae of syrphid flies, flower bugs and damsel bugs etc. Spray malathion 50 EC @ 400-500 ml/acre.
Blue bu�erfly, Polyomatus boe�cus (Linnaeus) F: Lycaenidae O: Lepidoptera.
Host plants: Pigeonpea, Lucerne, Sturt's Desert pea, Ra�le-Box, English Broom, Hyacinth bean, Sweet Pea. Trefoil, Lupines, Siratro, Garden peas and Broad beans etc.
Iden�fica�on and life cycle:
Adult: The bu�erflies have a wingspan of about 3 cms. It is small blue colured bu�erfly.
The tops of the wings of the males are blue, whereas those of the females are blue with wide dark brown edges. Underneath, they both have a brown and white pa�ern. The adults are dimorphic: the males and females being different. They both have a li�le tail on each hind wing, with a pair of small black eye-spots beside each tail. The pair of eye-spots and tails (fake antennae) are useful for confusing predators about which end of the animal is which. The species is found all over the world. (Ground 2010.)
Egg: The eggs are small, ini�ally pale green, later turning bluish white, hemispherical, strongly fla�ened top and bo�om, slightly depressed on top. The short blunt spines are present on the side, but absent on the top of the egg. The eggs period is about seven days. They are laid singly on the food plant, usually on flower buds, but some�mes on the open flowers and delicate green parts.
Larva: The full fed larva looks like a maggot and 5- 7mm in length. It is flat, sluggish and green in colure and becomes pink or purple if they are ea�ng red or purple flower buds, or green or yellowish green if they are on yellow flowers or green pods. The larval period is about 14 days.
Pupa : The full grown up larva find a suitable loca�on, some �mes it crawls to the base of the food plant, or simply drop off or lower themselves by silk from the food plant to the ground and find a suitable place for pupa�on. This can be among leaf and other debris within or around the base of the plant, beneath nearby rocks, or they can pupate in holes in the ground or in soil. In sandy country they will
37

actually force themselves into the loose sand to pupate if there is not enough debris about. They will also pupate within flowers on the food plant, so that when the flower eventually dies, shrivels, and falls to the ground, the pupa falls hidden within it. They also some�mes pupate inside the consumed seedpods if there is enough room.
The pupa is short cylindrical, mostly smooth with a few short bristles near the head, rounded anteriorly and posteriorly, about 10 mm long, polymorphic in various shades of brown, pink brown or green brown. The pupa�on period is variable, even within the same popula�on. It can be as short as 10 days in summer, extending to a month in winter. Pupal period varies from 10-30 days depending upon temperature.
Nature of damage: Caterpillars of L. boe�cus larvae remain inside the pod and feed on the internal contents/ green grains of the pod. The larval entry hole is not visible. The full-fed larva comes out of the pod for pupa�on a�er making an exit hole.
Management: Same as men�oned for Helicoverpa armigera
Fruit borer: Helicoverpa (=Heliothis) armigera (Hubner) F: Noctuidae O: Lepidoptera
Host plants: It is a polyphagous insect pest. Various Leguminous crops, co�on, tomato, berseem, wheat many ornamental plants as rose, poppy and many cul�vated and wild fruit trees like citrus etc.
Iden�fica�on and life cycle: -
Adult : Adults are yellowish brown, stoutly built moths with dark brown black circular spots on fore wings. Total life cycle is completed in 4-6 weeks.
Egg: Moth lays 1200-1600 creamy white eggs singly on twigs, leaves and flowers. Incuba�on period is 2-4 days.
Larva (Caterpillar): The caterpillar is the only damaging stage. It is green in colour with small hairs on its body. White grey broken longitudinal stripes are present on lateral sides of the larvae. The larval dura�on is 15-20 days.
Pupa: Pupa is brown in colour and pupa�on usually takes place in soil. Pupal period is 10-14 days.
Nature of damage: The young larvae feed on tender leaves and advanced stage larvae feed on flower buds and pods. Larva cuts circular hole in the pod through which it thrusts its head inside the pod and feeds the green grains inside the pod. The larva remains outside protruding its head inside the pod while feeding on green seeds. Some�mes whole larva goes inside the pod and all the grains are consumed.
38

Economic Threshold (E.T.): One larva per square meter area or one larva or one egg or one damaged fruit per plant.
Management
Cultural control: 1. Deep ploughing before transplan�ng helps to hi1l the pupae hiding in the soil. 2. Grow pest resistant varie�es like leafless varie�es. 3. Early transplanted crop escapes from the damage of this pest. Mechanical control: 1. Pick up the caterpillars and kill them. 2. Provide perching places for birds in the field so that they may pick up the larvae from the crop. Use of traps: 1 Use pheromone and light traps to monitor and collect male moth popula�on. Biological control:1 Grow cowpea or pulses on the bunds to build up natural enemy fauna.2 Release Trichogramma chilonis parasitoids twice a�er mid March at 2-5 day interval @ 500000 parasi�zed per hectare. Spray biopes�cides like Bacillus thuringiensis (B.t.) formula�ons viz., Biolep, Bioasp, Delfin, Dipel (@ 400 per acre) or NPV prepara�ons @ 300 LE per acre. Botanical insec�cides: Spray nimbicidine @ 2.5 litre per hectare 12 days a�er release of Trichogramma wasps. Chemical Control: Spray 150 ml cypermethrin 10 EC or 60 ml cypermethrin 25 EC or 200 ml decamethrin 2.8 EC or 75ml fenvalerate 20 EC or 500 g carbaryl 50 WP per acre in 200-250 litre water if needed, repeat the spray a�er 12 days.
Note: The above-men�oned insec�cides should be used alternately. Ten days interval should be given between spray applica�ons and fruit picking.
Pea stem fly, Ophiomyia phaseoli (Tryon) F: Agromyzidae O: Diptera
Host plants: It is also a polyphagous pest and feeds on almost all types of beans, gram, pigeonpea and pea. It is widely distributed in India, Srilanka, Philippines and China.
Adult fly : The adult flies are metallic black, 2.0-2.5 mm long with a wing expanse of 5 mm and are ac�ve in summer. The female flies live for 8-22 days and the males for 11 days. The pest completes 8-9 genera�ons 1n a year.
Eggs: The female lays 14-64 elongate, oval and white eggs into the leaf �ssue. The incuba�on period is 2-4 days.
Larva (Maggot): Ini�ally the maggot is white in colour but later on it becomes yellowish. These are small in size being less than 1.0 mm in length. The larval periods is 6-7 days and pass through three instars.
Pupa: The pupa is barrel-shaped and brown in colour. Pupa�on takes place in the gallery made by the larva within the stem itself. The pupal period varies from 5-19 days depending upon the temperature.
Nature of damage: The larvae are leaf and stem miners. The maggots just a�er emergence make mines in the leaf and goes towards the main stem. A�er entering the
39

main it feeds on the pith of the stem and goes down wards. The adults also cause damage by puncturing the leaves. The damaged plant turn yellow and the bearing capacity of the plant are reduced. The damage is more serious on seedlings.
Management: 1. Remove and destroy all the affected branches during the ini�al stages of a�ack. 2. Early sown crop can escape from the damage of the pest. 3. Spray the crop three �mes with 750 ml of methyl oxydemeton 25EC in 250 litres of water per ha. The first applica�on should be just a�er germina�on and the other two at an interval of 2 weeks each.
Pod borers: E�ella zinckenella (Treitschke) F: Pyralidae O: Lepidoptera
Host plants: It is serious pest of green pea and len�ls in northern India and also a�acks other pulses in various parts of the country.
Iden�fica�on and life cycle: -
Adult (Moth): The adult is a purple brown moth having grayish brown forewings with a wing expanse of 25 mm. The moths emerge in February/March and are nocturnal in habit.
Egg: The eggs are small, 0.5 to 0.7 mm long, ellip�cal and glistening white in colour. A female generally lays 50 to 180 eggs singly or in clusters usually at the junc�on of calyx and pod or occasionally on young pods. The egg period is 5 days.
Larva (Caterpillar): The �ny caterpillar is reddish pink dorsally and pale green ventrally. The full grown larva is rosy with a purplish �nge. The larval stage is completed in 10-27 days.
Pupa: Pupa�on takes place in soil which lasts for 10-15 days.
Nature of damage: The �ny greenish caterpillars bore into the pods and eat away the young grains; generally one caterpillar is found in one pod. The larva makes a large exit hole and comes out of the pod for pupa�on.
Management : 1. Grow resistant varie�es, leafless varie�es should be preferred. 2. Collect and burn the infested leaves. Spray 400 ml dimethoate 30 EC or 500 ml oxy-demeton methyl 25 EC or the insec�cide recommended for other pod borers will take care this pest also.
Insect pests of field bean (Dolichos lablab)
Field bean or French is cul�vated throughout the world. In India also it is very popular
and widely cul�vated in the southern states of the country. It is a rich source of protein
and fiber. This crop is damaged by a large number of insect pests due to which its
produc�on is badly affected. The descrip�ons of the major insect pests and their
management have been men�oned as follows.
40

Lablab bugs, Coptosoma cribraria (Fabricius.) F: Plataspidae O:
Heteroptera
Riptortus pedestris (Fabricius) F: Alydidae O: Heteroptera
Nature of damage: Among the sucking pests, lablab bug, Coptosoma cribraria
(Fabricius.) and Riptortus pedestris (Fabricius) occurred commonly and found in large
numbers throughout the cropping period (Govindan, 1974). The bugs were found
congrega�ng on tender vines and sucking sap resul�ng in fading of vines and shoots
(Ayyar 1963)
Aphid, Aphis craccivora Koch F: Aphididae O: Homoptera
Host plants: It is a serious pest of leguminous crop kidney bean,
French beans, cowpea, alfalfa, len�l, gram, fenugreek and some
flowering plant also.
Nymph: The nymphs and adults are black in colour.
Nature of damage: The aphid nymphs and adults suck the sap from tender shoots,
inflorescence and pods resul�ng in drying up of tender shoot and premature fall of
flower buds, flowers and tender pods.
3. Whitefly, Bemisia tabaci (Gennadius) F: Aleyrodidae O: Homoptera
Host plant: It is a polyphagous insect pest damage cooton.vegetable crop, pulse crops,
flowering plants, fruit trees, many herbs, shrubs and weed plants.
Adult: Adult whiteflies are almost 1-3 mm in body length and
possess seven segmented antennae and three pairs of legs with a
simple vena�on in fore-wing. A structure known as 'Vasiform
orifice' is unique in white fly. This vasiform orifice is present in all
larval (Nymphal) stages and adult. Two pairs of wings fully mobile
with white powdery wax all over the body, three pairs of legs and
fore wings with simple vena�on. Reproduce sexually and
occasionally parthenogene�cally.
Eggs: Pyriform or ovoid with creamy to brown in colour, pedicel
inserted in leaf �ssue and egg stands on pedicel. The eggs are laid
on lower side of leaf. A female lays from 48 to 394 eggs on the
lower surface of leaf. . Op�mum temperature for hatching is 028.5 C.
Crawler: The first instar nymph has legs and antennae. This is mobile in nature and called as called crawler. It crawls for a short distance to locate a suitable feeding site in
41

search of miner vein. They se�le at one in few hrs and insert their mouth parts into phloem �ssue and extract the sap and remain sessile un�l they reach the adult stage.
Nymph: A�er first moult the remaining three larval instars are nd rd
sessile and yellowish in colour. 2 and 3 Instars have oval or elongate oval body, may be circular or heart shaped. Body depressed, ellip�cal, pale greenish yellow and antennae are
rdpresent. In 3 instars antennae directed towards the median line and become hook shaped.
Pupa: Body convex, colour deep yellow, eyes well developed have red eyes, legs as stump like suckers and 7 pairs of dorsal bristles. The final whitefly larval stage is usually termed as a 'puparium'. The adult fly emerges out by rupturing of lines of weakness which are termed as 'transverse and longitudinal' moul�ng sutures. The vacated puparium is o�en termed as pupal case.
Female: Abdomen bigger and broader and dis�nctly yellow.
Male: Abdomen narrow and tapering posterior and creamy yellow in colour.
Nature of damage: Whitefly pierces stylet in plant �ssue and suck sap from phloem �ssue. Plant becomes yellow week by excessive drainage of sap. They produce large amount of sugar excreta (Honey dew) on which black sooty mould develops which interfere the photosynthesis. Honey dew a�racts ants which disturb the parasitoids and predators. The leaves are deforming by sucking the sap.
Pod borer, Helicoverpa armigera (Hübner) F: Noctuidae Order: Lepidoptera
Host plants: It is widely distributed in Africa, Asia, Europe and Oceania. The larvae feed on over 120 plant species of cul�vated and wild plants. Some of its host crop plants include alfalfa, beans, cabbage, carna�on, cauliflower, corn, chrysanthemum, geranium, peas, pepper, pink, small grains, squash, tomato and zucchini.
Adult (Moth): The moth is stout and light brown in colour with 35-40 mm wingspan and 12-20 mm long. Forewings are grey to brown with a network of fine, wavy, dark lines; brown band near the edge of forewing; hind wings buff with a dark band near the edge, which contains a pale sec�on; undersides of wings buff with dark bands near the edge; forewing has a dark, comma-like mark underside. Flights occur at dusk and night
Egg: The spherical egg is 0.5 mm in diameter, white to brown in colour and have a costate surface. The incuba�on period is three days. Female moth lays several hundred eggs eggs singly or in small clusters, on various parts of their host plants, most commonly on the top third of the plant and growing points.
42

Larva: Larva is up to 40 mm long; body color varies from green, yellow, pink and red-brown to almost black; a broad cream stripe running over spiracles along each side. Several pale, fine, wavy, broken lines run along the body dorsally; a single dark stripe running along the middle of the back; head pale brown with several spots. The larval period is 13 to 22 days. Larva passes through six instars. Three dark stripes extend along the dorsal side and one yellow light stripe is situated under the spiracles on the lateral side. The ventral parts of the larvae are pale. They are rather aggressive, some�mes carnivorous. The elder larvae when come in contact cannibalize with each other. If disturbed, they fall from the plant and curl up on the ground
Pupa: The pupa is pale to dark brown in and 15-20 mm in length. The pupae develop inside a silken cocoon in soil at a depth of 4–10 cen�meters. Pupal period is 10 to 15 days. The whole life cycle can be completed in just over a month.
Nature of damage: Larvae feed on various plant parts including leaves, growing �ps, buds, flowers, seeds, and fruits, causing extensive damage (Carter 1984). Larvae burrow into fruits of tomato, ears of corn and seed pods of pulses. They pupate in the cocoon underground, 5-10 cm deep. They overwinter as pupae. Two to three genera�ons develop per year in a year.
Management: Same as prescribed for other pod borers.
Bean Pod Borer, Maruca testulalis (Geyer) Maruca vitrata F: Pyralidae O: Lepidoptera
Host plants: It is a serious insect pest of leguminous crops like pigeon pea, cowpea, mung bean and soybean. Its common names include the Maruca pod borer, Bean pod borer, Soybean pod borer, mung bean, moth bean, and the legume pod borer.
Adult : The adult is creamy white to brown having medium brown wings and long legs. The longevity of male and female was 5-8 days and 6-11 days, respec�vely. Ma�ng period with an average 1-3 days. Total life span from egg to death of adult for male and female was 35.10 ± 2.42 and 39.60 ± 3.68 days, respec�vely or whole life cycle is about 20–57 days. (Sureja et al. 2010) R. S. Ochieng (1981) Okeyo-owuor and Ochieng, (1981), Ramasubramanian and Babu (1988) (Visakantaiah and Babu1980)
Eggs: The eggs are translucent and extremely difficult to see against the background of the oviposi�on site. Freshly laid eggs are pale yellowish white in color but turn white at the �me of hatching. Incuba�on period is 2-4 days. The female moth lays eggs either singly or in overlapping groups on the flower buds and flowers surface in large numbers. The female moth was 6 and 194 eggs in its life.
Larva: The larva passes through five instars. The total larval period is 12-15days.
43

Pupa: Pre-pupal and pupal period last for 1-3 days and 6-8 days, respec�vely.
Nature of damage: The early instar larva feeds on flower peduncles and young plant stems while the older larva feeds on flower-buds, flowers and young pods.
Loss: It can cause losses of 20-80% on the harvests of cowpeas.
Chemical control: one or two sprays of a protec�ve mixture of cypermethrin and dimethoate at different stages in the crop phenology, and full protec�on with the same mixture (Jackai, 1995).
The plume moth, Exelas�s atomosa Walshingham F: Pterophoridae O: Lepidoptera:
It is found in Cape Verde, Ethiopia, Kenya, Madagascar, South Africa, Swaziland, Tanzania, India, Nepal and Iran.
Host Plant: Leguminous and cucubitaceous crops.
Adult: Adult is a small moth with yellowish brown wings. The forewings are cut into two plumes and hind wings into three. The mean longevity of the adult is 6.59±0.38 days. The female lays about 93 to 101eggs in her life.
Egg: Egg is dirty in colour. It hatches in 2 to 4 days.
Larva: The lava is greenish-brown and fringed with short hairs and spines. There were five larval instars and the larval period is 22-24 days.
Pupa: Pupa�on takes place on pod surface or entrance of hole or in the burrow of infested pods and the pupal period is 7-9 days.
The life cycle of E. atomosa is 40 to 42 days (Subharani and Singh 2008).
Nature of damage: It is a serious pest of pigeon pea. The larva enters into the pod and feeds on developing grains. It also damages the flowers, buds and pods to drop.
White plume moth (Bo�le Gourd Plume Moth), Sphenarches caffer (Zeller)
F: Pterophoridae O:Lepidoptera
Host plants: The larvae feed on the leaves, flowers and fruits of various plants, including sweet potato. It is found India, Malaysia, Mauri�us, the Seychelles and South Africa.
Adult: It is slender shaped moth with lobed wings and fringed with scales
Egg: Eggs are laid singly on buds and leaves.
Larva: The larva is small, yellowish green, cylindrical with short spines all over the body.
Pupa: It is greenish brown in colour.
44

Nature of damage: Larva feeds on the leaves by making small holes.
Management: Collect and destroy larvae and pupae. Spray malathion 50 EC @ 500 ml or dimethoate 30 EC 500 ml or methyl demeton 25 EC @ 500 ml/ ha (agritech.tnau.ac.in)
Limabean pod borer, gold-banded moth, pea pod borer, legume pod moth, E�ella zinkenella (Treitschke) F: Pyralidae O: Lepidoptera
Host plants: It feeds on more than 80 species of cul�vated and wild legumes including soybeans, pea, len�l, lupine, vetch, white (silver-chain) and yellow (Siberian pea shrub) acacia, clover, alfalfa, sainfoin, vetchling, astragals, Pigeon pea, cowpea, soyabean limabean, mung bean, other species of Fabaceae and Catha edulis (Celastraceae). It is found in Southern & Eastern Europe, Africa and Asia, North America and Australia. Russia, North Caucasus, south Siberia, Bal�c States, Byelorussia, Ukraine, Moldavia, Transcaucasia, Kazakhstan, Middle Asia. Western Europe, north Africa, Asia Minor, India, China, Korea, Japan, South-East Asia, Australia and America.
Adult (Moth): The moth is quite dis�nc�ve, with a bright creamy-yellow costal streak and a yellowish and chestnut fascia at one third. Body length is 8-11 mm; wing span 19-27 mm and the wings are longer than abdomen which is folded as a roof. Forewing is yellow- or brown-grayish with characteris�c light stripe along fore edge, with orange spot on basal third, and with dark fringe. Hind wings are light gray, with dark vena�on and dark double line near fringe; the fringe is long and light in color. Top of abdomen is with a tu� of golden-yellow hairs. Moths are ac�ve during deep twilights and at night. Flight of adults begins in May-June.
Egg: A female lays maximum 600 eggs one by one in clusters on fully mature pods or on ovaries with dried corolla or on green fruits. Egg lasts for 4-21 days.
Larva: The young larvae are green, but become pinkish- red as they get older with the body length ranging from 15-22 mm. The larval period is 19-40 days. Larva overwinters within a cocoon in soil at a depth of 2-5 cm.
Pupa: Pupa is brilliant, brown, fine punctured, to 7-10 mm in length; cocoon is thick, white, and usually covered with soil par�cles. The pupal period is 12-18 days. It pupates in spring. Pupa�on takes place in the soil.
Life span of adult is 20 days, average fecundity is about 100-300, eggs.
Nature of damage: The larva feeds on maturing seeds a�er entering the pods. Fecal pellets are seen inside the damaged pods. It lives inside the fruit and eats away the seeds. The larva easily passes from one bean to another. Damaged seeds lose their commodity value and become unsuitable for sowing. Sowings of grain legumes located
45

near forest shelter belts composed of true or false acacia (serving as reserva�ons for the pest) are especially strongly damaged. Larvae are generally found infes�ng maturing and dried pods. Popula�ons of this pest build up by the end of the season, when the temperature is high.
Management: E�ella zinckenella pheromone trap are used to lure and kill the adults and reduce breeding. Eco-friendly! Cost-Effec�ve! Safe! sowing of grain legumes at early-op�mal periods; harvest executed at the proper �me; 20-25 cm deep under-winter plowing; spa�al isola�on from planta�ons of true and false acacia; insec�cide treatments soon before larva penetra�on into beans; resistant varie�es. Forecas�ng is achieved by means of sex pheromone traps.
Chemical Control: Apply chemical insec�cide only if the pest popula�on crosses ET. Spraying of monocrotophos 36SL 0.04% (1 ml of 36 SL /litre of water) or chlorpyriphos 20 EC @ 3.5 ml/litre of water at 600-1000 litre of spray material per ha with hand sprayer.
Blue bu�erfly, Lampides (Polyomatous) boe�cus (Linnaeus) : as men�oned under pea .
Field bean pod borer: Adisura atkinsoni,
Adult (Moth): Adult Moths are yellowish brown in colour. Forewings are yellow with 'V' shaped specks and hind wings have pale brown markings.
Eggs: The single female lays about 413 eggs. The incuba�on period is 3-5 days.
Larva: The larva is green in colour having brown lateral marking. It has humped anal segment. Old larva shows lateral brown strips and yellow to green color. The head is brown to black. The larva passes through six instars. Larva period is 17-21days.
Pupa: The pre-pupal dura�on is 2.15 ± 0.16 days, whereas pupal period is 13.15 ± 0.27 days.
Nature of damage: The larva feeds on flower buds, opened flowers, tender and mature pods. It bores inside the pod and feeds on the seeds within.
Economic Threshold (ET): 10% affected plants.
Management : Put 50 bird perches per ha. Mechanical collec�on of grown up larva. Deep summer ploughing in 2-3 years to eliminate quiescent pupa. Early sowing, short dura�on varie�es. Avoid closer plant spacing. Grow tall sorghum as comparison crop to serve as biological bird perches. Install pheromone traps at a distance of 50 m @ 5 traps/ha for each insect pest. Se�ng of light traps (1 light trap/5 acre) to kill moth popula�on.
46

Biological Control: Sequen�al applica�on of NSKE-HaNPV-Bt is effec�ve against insect pests. HaNPV is effec�ve against H. armigera larvae, but ineffec�ve to other pod borers Padmanaban and Arora (2002). Release Trichogramma chlionis at weekly intervals @1.5 lakh/ha/ week for four �mes. Conserve green lacewing, predatory s�nk bugs, spider, and ants. Applica�on of NPV 250 LE /ha with teepol 0.1% and Jaggery 0.5% thrice at 10 – 15 days interval commencing from flowering stage or spray Bt @ 600 g/h.
Chemical Control: Spray triazophos 0.05% , quinalphos 25 EC @ 1000 ml/ha, or 1000g carbaryl 50WP/ha.
Suggested Reading
AgroAtlas - Pests - E�ella zinckenella Tr. - Limabean Pod Borer, Pea ... h�p://www.agroatlas.ru/en/content/pests/E�ella_zinckenella/
E�ella Zinckenella Pheromone Trap - Made-in-China.com – h�p://www.made-in-china.com/showroom/�enjoy/product-detailPMUnfKdEHorC/China-E�ella-Zin.
R. Taghizadeh, A. A.Talebi Y. Fathipour and J. Khalghani 2012. Effect of ten soybean cul�vars on development and reproduc�on of lima bean pod borer,E�ella zinckenella (Lepidoptera: Pyralidae) under laboratory condi�ons. Appl. Ent. Phytopath.Vol. 79 (2) : 15.
AgroAtlas - Pests - E�ella zinckenella Tr. - Limabean Pod Borer, Pea ... h�p://www.agroatlas.ru/en/content/pests/E�ella_zinckenella/
h�p://agritech.tnau.ac.in/crop_protec�on/crop_prot_crop_insect-veg_cucurbit.html
Subharani S., Singh T.K. 2008. Biological Studies of Plume Moth, Exelas�s atomosa Wals. on Cajanus cajan (L.) Millsp. Annals of Plant Protec�on Sciences. 16(2):367- 369.
Louis E.N. Jackai The legume pod borer Maruca testulalis, and its principal host plant, Vigna unguiculata (L.) Walp. — use of selec�ve insec�cide sprays as an aid in the iden�fica�on of useful levels of resistance. Crop Protec�on 14(4): 299-306.
S. Rekha & C. P. Mallapur 2007. Abundance and Seasonability of Sucking Pests of Dolichos Bean. Karnataka J. Agric. Sci.,20(2): (397-398).
A. M. Byrappa, N. G. Kumar and M. Divya (2012). Impact of biopes�cides applica�on on pod borer complex in organically grown fieldbean ecosystem. J. Biopest. 5(2): 148-160.
Byre gowda , M.2006. Uses and chemical composi�on of Dolichos bean green pod. A report to the University of Agricultural Sciences, Bangalore, India,8-10PP
Kogan, M. and Herzog, D. C.1980.Sampling methods in soybean Entomology. Springes-Verlog Inc., New York, 82-126 PP. Dhamdhere, S. G. and Khaire, V. M. 1986.Field evalua�on of different doses of NPV
47

Heliothis armigera (Hub.). Current Research Report, Mahatma Phule Krishi Vidyapeeta, Rahuri, Maharashtra, India, 221-226PP.
Govindan, R. 1974. Insects of the field bean, Lablab nigervar. Lignosus Medikus with special reference to the biology and ecology of the pod borer, Adisura atkinsoni Moore (Leipdoptera : Noctuidae). M. Sc. (Agri.) Thesis, University of Agricultural Sciences, Bangalore, India.
Jayaraj, S., Rabindra, R. J. and Santharam, G.1987, Control of Heliothis armigera (Hub.) on chickpea and lablab beans by nuclear polyhedrosis virus. Indian Journal of Agricultural Sciences, 57: 738-741.
Katagihallimath, S. S. and Siddappaji, C.1962.Observa�ons on the incidence of lepidopteron pod borers of Dolichos lablab and the results of preliminary insec�cidal trails to control them. Second All India Congress of Zoology, 59PP.
Mallikarjunappa, S. 1989.Field bean pod borer complex with reference to germplasm screening, life table, ac�on threshold, crop loss and chemical control of Adisura atkinsoni. Ph.D Thesis, University of Agricultural Sciences, Bangalore, India.
Mallikarjuna, J. 2009, Studies on pod borers of Dolichos bean, Lablab purpureus L. Sweet) and their management. M.Sc (Agri.) Thesis, University of Agricultural Sciences, Bangalore,India.
Mallikarjuna, J., Ashok kumar, C. T. and Rashmi, M. A. 2009. Field evalua�on of indigenous materials and newer insec�cide molecules against pod borers of dolichos bean. Karnataka Journal of Agricultural Science, 22(3): 617.
Pawar, V. M. and Thombre, U. T. 1992 Prospects of baculovirus in integrated pest management of pulses. In: Emerging Trends in Biocontrol of Phytophagous Insects. (Ananthakrishnan, T. N.Ed.) Oxford and IBH Publishing Company Private limited, New Delhi, 253-258 P.
Padmanaban, N. and Arora, R. 2002. Field evalua�on of na�ve nucleopolyhedrovirus for the management of tomato fruit borer, Helicoverpa armigera (Hübner). Pes�cide Research Journal,14: 113-119.
Rekha, S. 2005. Status and management of pod borer complex in Dolichos bean, Lablab. purpureus L. M.Sc. (Agri.) Thesis, University of Agricultural Sciences, Dharwad, India.
Thejaswi, L.2007.Studies on popula�on dynamics of pest complex of field bean (Lablab purpureus L.) with special reference to pod borers. M.Sc. (Agri.) thesis,University of Agricultural Sciences,India.
Surelivelu, T., Chellaih, S. and Balasubramanaian, G., 1978, Efficacy of certain insec�cides and Bacillus thuringiensis in the control of lablab pod borers. Madras Agricultural Journal. 65(2): 120-121.
48

Mishra, A., Yadava, D. A., Patel, R. C. and Pawar, B. S. 1984.Field evalua�on of nuclear polyhedrosis virus against Heliothis armigera(Hub.) (Lepidoptera: Noctuidae) in Gujarat. Indian Journal of Plant Protec�on,12: 31-32.
B V Sureja, B G Pachani and A V Khanpara 2010. Biology of Spo�ed Pod Borer, Maruca testulalis (Geyer) on Cowpea. Research Journal of Agricultural Sciences 1(4):477-478.
Ramasubramanian G V and Babu S P C. 1988. Effect of host plants on some biological aspects ofspo�ed pod borer (Maruca testulalis) (Lepidoptera:Pyralidae). Indian Journal of AgriculturalSciences. 58:618-620.
Sardana H R and Verma S. 1986. Preliminary studies on the prevalence of insect pests and their natural enemies on cowpea in rela�on to weather factors at Delhi. Indian Journal of Entomology. 48(4):448-458.
Visakantaiah, M and Babu J C S. 1980.Bionomics of tur webworm, Maruca testulalis Geyer (Lepidoptera:Pyralidae). Mysore Journal of Agricultural Sciences14:529-532.
Okeyo-owuor J B and Ochieng R S. 1981. Studies on the legume pod-borer, Maruca testulalis Geyer. I. Life cycle and behaviour. Insect Science and its Applica�on. 1(3):262-268.
Saji, K. 2013. Lampides boe�cus Linnaeus, 1767 - Pea Blue. In K. Kunte, S. Kalesh & U. Kodandaramaiah (eds.). Bu�erflies of India, v. 2.00. Indian Founda�on for Bu�erflies. h�p://www.ifoundbu�erflies.org/sp/596/Lampides-boe�cus
49

NOVEL INSECTICIDES FOR LEGUME PEST MANAGEMENT
S.S. YADAV AND KRISHNA ROLANIADepartment of Entomology
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
India is the largest producer and consumer of pulses in the world. Major pulses
grown in India include chickpea or bengal gram (Cicer arie�num), pigeonpea or red
gram (Cajanus cajan), len�l (Lens culinaris), urdbean or black gram (Vigna mungo),
mungbean or green gram (Vigna radiata), lablab bean (Lablab purpureus), moth bean
(Vigna aconi�folia), horse gram (Dolichos uniflorus), pea (Pisum sa�vum var. arvense),
grass pea or khesari (Lathyrus sa�vus), cowpea (Vigna unguiculata), and broad bean or
faba bean (Vicia faba). More popular among these are chickpea, pigeonpea,
mungbean, urdbean and len�l. These crops are damaged by a large number of insect
pests, of which pod borers, Helicoverpa armigera and H. punc�gera; spo�ed pod borer,
Maruca vitrata; spiny pod borer, E�ella zinckenella; pod fly, Melanagromyza obtusa;
leaf miner, Liriomyza cicerina; stem fly, Ophiomyia phaseoli; pea and bean weevil,
Sitona spp.; aphids, Aphis craccivora, Aphis fabae, and Acyrthosiphon pisum; whitefly,
Bemisia tabaci; defoliators, Spodoptera litura, S. exigua, and Amsacta spp.;
lea�oppers, Empoasca spp., thrips, Megaleurothrips dorsalis, and Caliothrips indicus;
blister beetles, Mylabris spp.; and the bruchids, Collasobruchus chinensis and Bruchus
pisorum cause extensive losses worldwide. "The injudicious use of pes�cides has
resulted in many problems eg. destruc�on of non target organisms, development of
insect resistance to insec�cides, outbreak of secondary pests, pes�cide poisoning and
presence of pes�cide residues in food and feed. During the past four decades, efforts
have been made to reduce the risk to human exposure to pes�cides, especially
insec�cides. Accordingly, many conven�onal pes�cides have been replaced by newer
insec�cides which are more selec�ve than conven�onal insec�cides. Hence safer and
fit well in IPM programmes. To overcome these problems, number of newer
insec�cides with unique mode of ac�on have been registered during late 1990s and
early 2000s for insect pest control in agriculture (Table 1). These insec�cides play an
important role in IPM of legume crops with proven bio-efficacy, high selec�vity and low
mammalian toxicity, that render them a�rac�ve replacement for organochlorines,
organophosphates, carbamates and pyrethroid insec�cides.
Neonico�noids
The neonico�noid group of insec�cides (viz. imidacloprid, acetamiprid,
50

Thiamethoxam, thiachloprid, clothianidin and dinotefuran) are highly ac�ve
insec�cides for foliar applica�on, soil and seed treatment. They have combining
systemic proper�es with rela�vely low applica�on rates and act agonis�cally at the pest
synap�c nico�nic acetylcholine receptors in the central nervous system of insects and
also have par�cularly strong ac�vity against sucking insects such as aphids, jassids,
whiteflies and thrips. They show good compa�bility with all relevant crops and
phytotoxicity is also found to be nil. They are notable for their rela�vely low toxicity to
most animals other than insects due to its specificity for novel type of receptor which is
found more o�en in insect nervous systems than that of animals and used for control all
target insect pest.
Phenyl pyrazoles (Fipronil)
Fipronil is broad spectrum neurotoxin that works as a GABA agonist. It has
contact ac�vity on both chewing and sucking insects and controls Coleoptera,
Lepidoptera, Diptera, Homoptera, Isoptera and Thysanoptera. It has systemic ac�vity,
with long residual and show excellent poten�al for control of most sucking and chewing
insects, thrips, beetles, Termites and grasshoppers. Fipronil is effec�ve at low field
applica�on rates against insects that are resistant to other chemical agents such as
pyrethroids, organophosphates and carbamates.
Oxadiazines (Indoxacarb)
Indoxacarb is the first commercialized insec�cide of the oxadizine group. It is a
propes�cide with unique mode of ac�on. It inhibits the flow of sodium ions into nerve
cell in insects that cause paralysis and death. It enters into insect body in two ways like
through inges�on of treated foliage and also penetrates through insect cu�cle. The
major use is against chewing insects such as lepidopterous larvae, beetles on
Pigeonpea, soybean and other legume crops (pod borer complex).
Pyrroles (Chlorfenapyr)
The pyrroles are compounds that act as metabolic toxins and work uncoupling
oxida�ve phosphoryla�on in the mitochondria, a key cellular process. They have
translaminar ac�vity and are toxic both by contact and inges�on to chewing & sucking
arthropods.
Pyridine azomethines (Pymetrozine)
Pymetrozine, a novel pyridine azomethine insec�cide, is highly specific against
sucking insect pests. The mode of ac�on of pymetrozine in insects has not been
precisely determined biochemically but it may involve effects on neuro-regula�on or
nerve-muscle interac�on. Physiologically, it appears to act by preven�ng these insects
51

from inser�ng their stylus into the plant �ssue. The compound is powerful toxicant
against aphids. Pymetrozine has systemic and translaminar ac�vi�es and can be used as
drench or foliar applica�on.
Tetronic acids deriva�ve (Spiromesifen)
Spiromesifen is a novel insec�cide/acaricide belonging to the new chemical
class of spirocyclic phenyl-subs�tuted tetronic acids, recently introduced by Bayer Crop
Science. Spiromesifen is especially ac�ve against whiteflies and tetranychid spider mite
species as Tetramychus spp. a�er foliar applica�on. Its mode of ac�on is by inhibi�on of
lipid biosynthesis that affects the egg and immature stages of these pests.
Pyridine carboxamid (Flonicamid)
Flonicamid pyridine carboxamide novel compound with a unique mode of
ac�on has contact ac�vity and is upwardly systemic. Flonicamid effec�vely manages
popula�on of aphids, thrips, leaf hoppers and other sucking insects of legume crops. It
has no nega�ve impact on beneficial insects or natural enemies. Flonicamid acts as an
an�feedant and a behavioural modifier on aphids within 30 minutes of exposure
resul�ng in insect mortality from starva�on within 5 to 7 days a�er exposure. It has
systemic and translaminar ac�vity and gives long term control. Flonicamid is a systemic
insec�cide that immediately suppresses the feeding of sucking insects. Its mode of
ac�on appears to be unique and can help in pest resistance management.
Tetramic acid deriva�ve (Spirotetramat)
Spirotetramat, a novel insec�cide belonging to chemical class of Ketoenois is a
tetramic acid deriva�ve. It is said to interfere with lipid biosynthesis, leading to death of
juvenile within 2-10 days a�er applica�on. It is systemic in ac�on, xylem and phloem
mobile, allowing acropetal and basipetal transloca�on in the plant. A�er foliar
applica�on, spirotetramat penetrates through leaf cu�cle and is translocated as
spirotetramat-enol via xylem and phloem, upto growing shoots and down to roots. This
full ambimobility or two way systemicity ensures the control of hidden and soil living
sucking pests a�er foliar applica�on and protects new shoots.
Diamides (Flubendiamide, Chlorantraniliprole)
Flubendiamide belongs to phthalic acid diamide group developed by Bayer Crop
Science, Germany in collabora�on with Nihon Nohyaku Co. Ltd., Tokyo, Japan. In
contrast to other insec�cide classes targe�ng the insect nervous system,
flubendiamide act at receptor in insect muscles causing an immediate cessa�on of
feeding and thus avoids crop damage. It is well suited for the control of a broad range of
lepidopteran pests. The unique mode of ac�on makes the compound well suited as a
52

tool in insect resistance management programmes (Pigeonpea pod borers).
Chlorantraniliprole belongs to new chemical class, the anthranilic diamides, and
has a novel mode of ac�on as an ac�vator of insect ryanodine receptors, causing rapid
muscle disfunc�on and paralysis. The insec�cide has proven to be effec�ve against
lepidopteran pests as well as against selected species in the order Coleoptera, Diptera
and Hemiptera.
Coragen SC is an anthranilamide discovered and currently being
commercialized by Dupont crop protec�on, acts primarily via inges�on on whitefly
nymphs. Although adult whiteflies are not directly controlled, feeding behaviour and
reproduc�on are affected. Field efficacy tests conducted around the world indicate that
in addi�on to its outstanding efficacy on lepidopterian pests. Rynaxypyr can be a
valuable tool for the management of key whitefly species, providing inhibi�on of virus
transmission and overall crop protec�on.
Dichloropropenyl ethers
Pyridalyl is the only member of this class. Pyridalyl gives very good control of
various lepidopterous and thysanopterous pests, without phyto toxicity. It controls
popula�on of Helicoverpa armigera and Spodoptera litura as well as against
Thysanopteran insects. It also produces unique insec�cidal symptoms, so it may have
different mode of ac�on from exis�ng insec�cides. Pyridalyl is also less harmful than
exis�ng insec�cides to various beneficial arthropods, so it should provide an important
tool in Integrated Pest Management (IPM) and insec�cidal management programmes
for the control of lepidopterous and thysanopterous pests.
Bacterial fermenta�on products
Spinosyns (Spinosad and Spinetorum)
Spinosyns are class of fermenta�on derived macro cyclic lactone
bioinsec�cides, produced from Sccharopolyspora spinosa. Spinosad is derived from
mixture of spinosyn A and B and spinetoram is derived from the mixtue of spinosyn J
and L. They are par�cularly effec�ve against lepidopterous larvae, dipterous leaf miners
and thrips provide both improved efficacy and an expanded spectrum while
maintaining the excep�onal environmental and toxicological profile already
established for spinosyn chemistry. Both spinosad and spinetoram furnish an excellent
combina�on of ac�vity and residual control while maintaining the low mammalian
toxicity and short environmental persistence. It acts by depolarizing insect neurons
involving acetylcholine and GABA receptors. No cross resistance exhibited with other
insect control agents.
53

Mec�ns
The mec�n family are complex macrocyclic lactones with (Abamec�n,
Emamec�n benzoate) or without (Milbemec�n) conjuga�on with sugar residues.
Abamec�n is a mixture of two naturally occurring avermec�ns that are derived
by fermenta�on from the soil micro-organism Streptomyces avermi�s. Its primary
ac�vity against mites and limited range of insects.
Emamec�n benzoate is a “Semisynthe�c” epiamino deriva�ve of abamec�n. It
shows much improved ac�vity against lepidopterous pests. Emamec�n affects the
nervous system of arthropods by increasing chloride ion flux at the neuromuscular
junc�on, resul�ng in cessa�on of feeding and irreversible paralysis. It has translaminar
ac�vity, providing a rela�vely prolonged residual ac�vity. Surface residues of
emamec�n benzoate decompose rapidly in sunlight, resul�ng in rela�vely low toxicity
to beneficial insects. Hence, this compound is considered important component in IPM
programmes for controlling some agricultural pests.
Milbemec�n is primarily a mi�cide act as allosteric ac�vators of glutamate and
GABA-mediated neurotransmission causes rapid and prolonged paralysis and
ul�mately death.
Benzoylphenyl urea (Novaluron, Buprofezin, Diflubenzuron, Lufenuron)
They are chi�n synthesis inhibitors, affect on moul�ng, act by inges�on and
contact, causes abnormal endocu�cular deposi�on and abor�ve moul�ng due to
reduc�on of biochemical process. Among the Benzoyl phenyl urea (BPU's) group,
Novaluron, Diflubenzuron and Lufenron are most important in controlling lepidopteran
and coleopteran larvae which infest the legume crops. They act on insects by inhibi�ng
chi�n forma�on, thereby causing abnormal endocu�cular deposi�on and abor�ve
moul�ng and also cease feeding. They affect on the larval stage of insect which
synthesize chi�n during their moul�ng process. Hence, the adult of beneficial species,
predators and parasitoids are seldom affected. For this reason, they are considered as
important component as biora�onal pes�cides in IPM programmes.
Novaluron: Novaluron is an insec�cide of benzoylphenyl urea class of insect-growth
regulators (IGR)/ It is a rela�vely new CSI (Chi�n synthesis inhibitor), that inhibits the
chi�n forma�on on larvae of various insects (Lepidoptera, Coleoptera, Homoptera and
Diptera). It has a potent insec�cidal ac�vity against several important foliage feeding
insect pests. As it is a powerful suppressor of lepidopteran larvae such as Spodoptera
litura and Helicoverpa armigra (by inges�on) and whitefly, Bemisia tabaci nymphs (by
contact). Novaluron is more ac�ve in suppressing developing stages of leaf miners,
Liniomyza sp.
54

Table 1. Novel Insec�cides
No. of Class of the insec�cides Name of the insec�cidesclass1 Neonico�noids Imidacloprid, Acetamiprid, Thiamethoxam, Thiachloprid, Clothianidin and Dinotefuran2 Phenyl pyrazoles Fipronil3 Oxadiazines Indoxacarb4 Pyrroles Chlorfenapyr5 Pyridine Azomethines Pymetrozine6 Tetronic Acid Deriva�ve Spiromesifen, Spirodiclofen7 Tetramic Acid Deriva�ve Spirotetramat8 Pyridine carboxamid Flonicamid9 Diamides Flubendiamide, Chlorantraniliprole10 Dichloropropenyl ethers Pyridalyl11 Bacterial Fermenta�on Products Spinosyns, Avermec�ns12 Benzoyl phenyl Urea Novaluron, Buprofezin, Diflubenzuron, Lufenuron
Table 2. Novel Insec�cides recommended for legume pest management
Spinosad 45.0% SC Pigeonpea: Pod borerThiamethoxam 30% FS Soybean: StemflyThiacloprid 21.7% SC Soybean: Girdle beetleSpinosad 45.0% SC Pigeonpea: Pod borerNovaluron 10% EC Chick pea: Pod borerLufenuron 5.4% EC Pigeonpea: Pod borer, podfly Black gram: Pod borer Indoxacarb 15.8% EC Pigeonpea: Pod borer complex Soybean: Tobacco caterpillar, green semilooper, stem flyIndoxacarb 14.5% SC Pigeonpea: Pod borer complex.Imidacloprid 17.8% SL Sucking pestsImidacloprid 48% FS Soybean: JassidsFlubendiamide 39.35% M/M SC Pigeonpea, black gram : Pod borer complexEmamec�n Benzoate 5% SG Chick pea: Pod borerChlorantraniliprole 18.5% SC Pigeonpea, black gram, Chick pea: Pod borer complex. Soybean: Green Semilooper, stem fly, girdle beetle.
55

Suggested Reading:
Gopali, J. B., Sharma, O.P., Yelshe�y, S. and Rachappa, V. 2013. Effect of insec�cides and biora�onals against pod bug (Clavigralla gibbosa) in pigeonpea. Indian Journal of Agricultural Sciences 83 (5): 582–5.
h�p://cibrc.nic.in/
h�p://www.irac-online.org/teams/crop-protec�on/
Ishaaya, I. 1990. Benzoylphenyl ureas and other selec�ve control agents—Mechanism and applica�on. In Pes�cides and alterna�ves, ed. J. E. Casida, 365–76. Amsterdam: Elsevier.
Ishaaya, I., and Horowitz, A. R. 1998. Insec�cides with novel modes of ac�on: An overview. In Insec�cides with novel modes of ac�on: Mechanism and applica�on, ed. I. Ishaaya and D. Degheele, 1–24. Berlin: Springer.
Ishaaya, I., et al. 2002b. Novaluron (Rimon), a novel IGR—Mechanism, selec�vity and importance in IPM programs. Med. Landbouww Rijksuniv. Gent. 67:617–26.
Ishaaya, I., S. Kontsedalov, and A. R. Horowitz. 2002. Emamec�n, a novel insec�cide for controlling field crop pests. Pest. Manage. Sci. 58:1091–95.
Memon, N.A. and Memon, A.A. 2005. Efficacy of different insec�cides against len�l pod borer (Helicoverpa spp.). Research Journal of Agriculture and Biological Sciences 1(1): 94-97.
56

ROLE OF CULTURAL METHODS IN SUPPRESSING INSECT- PESTS IN LEGUME CROPS
K. K. DAHIYADepartment of Entomology, CCS Haryana Agricultural University,
Hisar-125 004
Surprisingly simple modifica�ons of a pest's environment or habitat o�en prove to be effec�ve methods of pest control. As a group, these tac�cs are usually known as cultural control prac�ces. Low cost is the advantage of cultural control tac�c. S�ll a wide variety of insect pests are there which cannot be suppressed by cultural methods alone. Cultural prac�ces are simple modifica�ons or adapta�ons of regular farm opera�ons. The extra cost of their incorpora�on into pest management system is minimal in most cases. These are o�en the only control op�ons ecologically feasible for low value crops. These opera�ons produce no or negligible harmful ecological consequences and their primary aims are to prevent and reduce the pest out breaks and are effec�ve means of pest control. These do not result in total elimina�on of the pest thus allowing the conserva�on of beneficial insects. These prac�ces make cropping systems less friendly to the establishment and prolifera�on of pest popula�ons. They are designed for their posi�ve effects on farm ecology and pest management. The implementa�on of cultural control tac�cs require through knowledge of pest ecology and its interac�on with the cropping system. Area wide development of cultural prac�ces is essen�al for the effec�veness of the prac�ce in pest management system. These prac�ces are o�en pest, crop and region specific. Care should be emphasized in transferring tac�cs to region with markedly different agro-ecological condi�ons. To overcome insect-pest problems in legume crops, some of the modern cultural methods are developed to strengthen pest management and enhance crop produc�vity.
Chickpea, Cicer arie�num L.
Chickpea (Cicer arie�num L.) is the third most important legume crop in the world. In India, it is generally grown under rainfed condi�ons in Haryana, Rajasthan, U�ar Pradesh, Madhya Pradesh and Maharashtra. Nearly 60 insect species are known to feed on chickpea. The pod borer, H. armigera and the aphid, A. craccivora are the major pests of chickpea in the Indian Subcon�nent. The black aphid, A. craccivora is important as a vector of the chickpea stunt disease, while C. chinensis is the most dominant species in storage.
Intercropping/Mixed cropping:
Intercropping or strip-cropping with marigold, sunflower, linseed, mustard and coriander can minimize the extent of damage to the main crop. Intercropping of crops
57

viz., linseed, wheat, barley, safflower, coriander and mustard with gram directly affected the pod borer incidence. Intercropping of chickpea with safflower in 6:1 ra�o reduced pod damage and increased chickpea yield (Gowda et. al., 2004). The sole crop of chickpea a�racted more Helicoverpa compared to intercrops with wheat, barley, linseed, mustard and safflower. Parasi�sm was more near the rows of coriander. Strip-cropping also increases the efficiency of chemical control (Pimberi and Srivastava, 1989).
Crop rota�on:
Rota�ons do not help to manage these polyphagous and very mobile insects, although it has been noted that some crops (e.g. lucerne) are more a�rac�ve to the moths, and suscep�ble crops should not be planted too close to the main crop. Habitat diversifica�on to enhance pest control has been a�empted in Australia.
Trap crop:
Trap crops are managed in the same way as commercial crops, but destroyed by cul�va�on before larvae begin to pupate. The trap crops reduce the size of the local H. armigera popula�on before it can infest summer crops and start to increase in size. As a result, the overall H. armigera pressure on summer crops is reduced, resul�ng in greater opportunity for the implementa�on of so� control op�ons, reduced insec�cide use and greater natural enemy ac�vity. More eggs were laid on field pea as compared to vetch and faba bean. A par�al life table study of the fate of eggs oviposited on the field pea and chickpea suggested that large propor�ons of the eggs laid on the field pea suffered mortality due to their dislodging from the plants a�er oviposi�on. Plan�ng field pea as a trap crop in chickpea under commercial condi�ons confirmed the high level of a�rac�on of this crop to the moths ready for oviposi�on (Grundy et al., 2004).
Timely sowing of the crop:
Timely sown crop generally escape the infesta�on of major insect-pests. Delay
in plan�ng may cause increase in damage (Srivastava and Singh, 1979). Popula�on
fluctuated on variable dates of sowing (Hossaina et al. 2008). In Northern India
(Haryana and Punjab), late maturing crops suffering more damage while in Southern
part of India (ICRISAT) pod borer incidence was more in early maturing cul�vars of
chickpea (Dahiya, 1998). Sequen�al/staggered sowing extends the �me of availability
of chickpea as a host and permits the popula�on to develop to the addi�onal
genera�ons. This generally leads to the outbreak of the pod borer in north Indian
condi�ons. Cultural prac�ces such as removing leguminous weeds, trap cropping with
Crotalaria spp., intercropping monocots like sorghum, maize, pearl millet or finger
millet, mung and urd beans reduced pod damage in main crop (Ganapathy, 2010). The
black aphid, A. craccivora causes substan�al damage to chickpea in North India.
58

Varie�es with low trichome density or devoid of trichomes are highly suscep�ble to
aphid damage. The aphid incidence is greater under drought condi�ons. There are
three popula�on peaks on chickpea at Hisar, Haryana, India (Sithanantham et al.,
1984). Early sowing leads to early canopy closure, which also helps reduce virus spread
in chickpea.
Presence of weeds:
The presence of diverse vegeta�on within or near the main field may add
essen�al resources for predators or parasites and so enable them to find all their
requirements near the pest popula�ons. Such resources include food, cover or
alternate prey. Conversely, weeds may also adversely affect the orienta�on of
predators and parasite to their prey. The weeds may even directly contribute to pest
mul�plica�on by providing preferred surface for oviposi�on. Vicia sa�va is a common
weed associated with chickpea in northern India. Removal of the weed at a �me when
maximum eggs are laid substan�ally reduces the incidence of pod borer, H. armigera
(Rathore and Nwanze, 1993).
Pigeonpea, Cajanus cajan (L.) Millspaugh
Pigeonpea, Cajanus cajan is a major grain legume crop of the tropics and sub-
tropics (Nene and Sheila, 1990). However, the yield level of this crop is not very
encouraging, as the crop has been reported to be a�acked by several pest species. Its
produc�vity is far below the poten�al yield. The constraint for the low yield is the heavy
infesta�on of an array of pest complex. Pod borers are the key impediments for the low
produc�vity in India. The borers together damage 57% pods and 35% seeds incurring
yield loss of 28% (Sahoo, 1998). Among these, spo�ed pod borer, Maruca vitrata Geyer,
Gram pod borer, Helicoverpa armigera Hubner, blue bu�erfly, Lampides boe�cus (L),
plume moth, Exelas�s atomosa Walshingham, pod bug, Clavigralla gibbosa Spinola,
pod fly, Melanagromyza obtusa Malloch and pod wasp, Tanaos�gmodes cajaninae
Lasalle are considered important in causing economic losses to the farmers.
Sowing Time:
Early and mid-late maturing varie�es of pigeonpea (Cajanus cajan) were more
infested by H. armigera. Early maturing crop escape pod borer damage as the variety
completes podding stage and pods become harder prior to the ini�a�on of infesta�on
(Sachan and Lal, 1997). Incidence of pod fly and plume moth was more in mid late and
late maturing varie�es as compared to early ones (Yadav et al. 1989). Short dura�on
pigeon pea lines, ICPL 88037 and MPG 679 that recorded low damage (10 to 25%)
showed excellent recovery from damage evaluated for recovery resistance to M.
testulalis.
59

Intercropping:
The incidence of Helicoverpa was low in pigeonpea intercropped with soybean. Due to increased ac�vity of natural enemies like Chrysoperla, Chilomenes sexmaculatus, Orius spp. and spiders in sorghum, pigeonpea intercropped fields, resulted in the reduced pod damage and higher yields. It may also be interpreted that due to increased ac�vity of predatory birds like balckdrongo, house sparrow, Indian myna and green bee eater in sorghum + pigeonpea intercropped plots where sorghum acts as a live perch for birds to pray on H. armigera larvae from greater distance and height. Changes in crop canopy brought out by intercropping with blackgram, greengram, cowpea, sorghum and pearl millet delayed the incidence of pod borers in pigeonpea (Singh and Singh, 1978). Ganapathy (2010) reported that removing of leguminous weeds, trap cropping with Crotalaria spp., intercropping monocots like sorghum, maize, pearl millet or finger millet, mung and urd beans reduced pod damage in pigeon crop.
Summer ploughing:
Summer ploughing and mixing of sorghum seeds (300gm/ha) as bird perches increased the average produc�on of pigeonpea and control the pod borer up to 85 per cent and gain the addi�onal net returns over the farmers prac�ces (Sandeep et al. 2013).
Cowpea, Vigna unguiculata L. Walp.
Cowpea (Vigna unguiculata) is an important grain legume of the tropics. It is an inexpensive source of vegetable protein, and a hardy crop well adapted to rela�vely dry environments. More than 100 field pests of cowpea can be found in most of the crop produc�on agro-ecologies in world, but for of these – aphids (Aphis craccivora Koch), flower thrips (Megalurothrips sjostedi Trybom), the legume pod borer (Maruca vitrata Fab. Syn. Maruca testulalis Gayer) and the pod sucking bugs- are commonly encountered and are of economic importance.
Sowing �me:
The phenology of most crop species affords flexibility in choosing a crop plan�ng date. Plan�ng date may be manipulated to reduce insect densi�es and damage in a variety of crops. Pod borer infesta�on increases on the late sown crop. Grain yield also decreases in late planted crops. Simultaneously plan�ngs of maize and cowpea increase pod borer infesta�on in cowpea (Ezueh and Taylor, 1984), whereas sowing cowpea 12 weeks a�er maize reduces the pod borer damage.
Intercropping:
Intercropping offers remedial control, but the crop combina�on must consider the pest profile, cowpea/sorghum intercrop being effec�ve against aphids and thrips, and cowpea/green gram against legume pod borers and pod sucking bugs. Selected
60

combina�ons of agronomic, cultural control measures (IPM), especially when combined with early plan�ng, offer be�er management op�ons than the use of sole treatments.
Trap cropping:
The trap cropping and a plan�ng date modifica�on reduced plant bug, L. hesperus abundance in the crop plots and consequently reduce damage. An IPM strategy for L. hesperus in black eyed cowpea should incorporate a well-�med plan�ng date and well-chosen cowpea variety. Trap cropping may work to a�ract L. hesperus from the cowpeas (Bensen and Temple, 2008).
Urdbean/black gram, Vigna mungo L.
Over 64 species of insect-pests have been found associated with black gram. Of these, Caliothrips indicus, Megalurothrips distalis (Karny), leaf hopper, Empoasca kerri (Pruthi), whitefly, Bemisia tabaci (Gennadius), hairy caterpillars, Spilosima obliqua (Walk.) and Amsacta moorei (Butl.), tobacco caterpillar, Spodoptera litura (Fab.), pod borers, Helicoverpa armigera (Hub.) and Maruca vitrata (Geyer) are the common pests. Late sowing and dry spell condi�ons results an outbreak of Maruca testulalis which resulted in almost 100 per cent loss of flower buds and pods (Giraddi et al., 2000). Increasing in the phosphorus fer�lizer level during crop period resul�ng in the decreases of the pest infesta�on and the combina�on of 40 kg phosphorus with 15 kg nitrogen/ha gave the pest tolerance and be�er crop yield (Kumar et al. 2000). Through intercropping of black gram with non-host and taller crops has been reported to decrease the incidence of hoppers and whiteflies. Sowing of mixed biodiversity crops enhance the ac�vity of predators and parasitoids. Reduc�on of pests complexes and increased the biological agents was observed when black gram crop was intercropped with sorghum or pigeonpea crop (Dhuri et al., 1986). Intercropping of black gram and pigeonpea, black gram and maize, black gram and castor, castor and pigeonpea, castor and maize, sesamum and pigeonpea, pure crops of black gram and castor and sesamum resulted maximum S. obliqua abundance on pure crops, while intercropping of these crops with pigeonpea resulted lowest pest popula�on and the rate of parasi�sm by Trichogramma chilonis was rela�vely high in pure crops of castor followed by black gram, while it adversely affected with the intercropping of black gram and pigeonpea (Yadav et al., 1992).
Mungbean, Vigna radiata L.
The mungbean crop is a�acked by a number of insect pests. The most serious
pest problems include the whitefly (Bemisia tabaci), bean thrips (Megalurothrips
distalis), gram pod borer (Helicoverpa armigera) and legume pod borer (Maruca
vitrata). Pod borer infesta�on can be minimized by avoiding successive plan�ngs of
summer legumes. Good agronomic prac�ces and soil moisture are crucial as large,
61

luxurious growing plants suffer less defolia�on and have less risk of terminal damage.
Vigorously growing plants with adequate available moisture are be�er able to replace
damaged leaves and compensate for flower and pod damage. The effect so of
intercropping green gram with maize, ricebean, soybean, black gram or groundnut on
infesta�on of Monolepta signata, A. craccivora, Nizara viridula were inves�gated.
Intercropping with maize reduced popula�ons of pests on greengram than
intercropping with leguminous crops (Das and Du�a, 1996). Intercropping of maize and
sorghum along the periphery significantly reduced the whitefly incidence and the pod
damage caused by M. vitrata and L. boe�cus in greengram. Popula�on of the pest -1increased at higher seed rate of 37.5-40 kg ha compared with low seed rate (25-30 kg
-1ha ). Narrow row-to-row spacing of 20 cm a�racted more thrips compared with wider
spacing of 30 cm. Lesser fer�lizer and more irriga�ons were effec�ve in lowering thrips
popula�on. Mired bugs are also of key pests of mungbens as they feeds on buds and
flowers causing them to abort. Shortening of mungbean flowering period reduces the
risk of mirid damage. Flowering periods can be shortened by plan�ng on a full moisture
profile and by watering crops just before budding.
Kidney Beans/Common beans, Phaseolus vulgaris
Pod borer damage in a mono crop is greater than the maize-cowpea-sorghum
inter-mixed crops. Pod borer incidence was significantly lower in intercropped and
higher plant popula�ons than in pure stands, and in a lower plant popula�on of
common bean, Phaseolus vulgaris (Karel, 1993). Flower and pod damage was
significantly lower in an intercrop combina�on of one third bean-two thirds maize, so
intercropping maize with bean was considered useful as a cultural method for
controlling pod borers in common bean.
Groundnut, Arachis hyposia
In India, early plan�ng has been found to reduce white grub damage in
groundnut. Closer spacing in groundnut lowered the incidence of thrips, jassids and leaf
miner, and also increased parasi�sm in the la�er. Deep summer ploughing is also
helpful in minimizing the infesta�on of white grub a�acking groundnut, pupae of hairy
caterpillars a�acking groundnut and destruc�on of alternate hosts is also helpful in
reducing red hairy caterpillar in groundnut crop. Intercropping of groundnut with pearl
millet reduced the incidence of thrips, jassids and leaf minor whereas, the same with
sunflower and castor increased the incidence of thrips, and jassid, respec�vely. When
pearl millet was grown as an intercrop in groundnut, the parasi�c ac�vity of Goniozu sp.
was considerably enhanced. The pollen grains of the millet were preferably used as
food by the adult parasitoids. Rota�on of groundnut with non-leguminous crops is
recommended as a method for minimizing the damage by leaf miner.
62

Lucerne (Medicago sa�va L.)
Lucerne, Medicago sa�va suffers great loss in yield due to variety of insect-pests a�ack during crop period. Among vast array of insect pests, leaf hoppers (Empoasca sp.), aphids (Therioaphis maculate and Aphis craccivora) and lucerne weevil (Hyper avariabilis) are the important ones. Clima�c condi�ons in a locality greatly influence the incidence of insect-pests. In an a�empt to find the op�mum �me of sowing for lucerne to manage insect pests to achieve a maximum yield of green fodder, sowing in early November, yielded the maximum, related to the least damaging intensity of lea�oppers, lucerne weevil and aphids, the op�mal �me for vegeta�ve growth and the temperature and humidity. Other sowings gave a poor yield. The popula�on of aphid, leaf hopper and lucerne weevil was managed by improved cultural prac�ces, viz., last suscep�ble variety and op�mum fer�lizer combina�on when crop was sown at the �me of the last week of October and increased the green fodder produc�on also (Shri Ram and Gupta, 1990). A reduc�on in the incidence of a number of insect pests including Empoasca sp., Therioaphis trifolii f. maculate and Aphis craccivora on lucerne following increasing applica�on of K either singly or in combina�on with P has been reported by Dhaliwal and Arora, (1993).
Suggested reading:
Bensen, T.A. and Temple, S.R. (2008). Trap cropping, plan�ng date, and cowpea variety as poten�al elements of an integrated pest management strategy for Lygus hesperus in blackeyed cowpea. Crop Protec�on, 27: 1343– 1353.
Das, R and Du�a, S.K. (1996). Effect of intercropping on infesta�on of insect-pests of green gram. J. Agric. Sci. 9: 220.
Dhaliwal, G.S. and Arora, R. (1993). Changing status of insect pests and their management strategies. In: K. S. Gill, G.S. Dhaliwal and B.S. Hansra (eds). Changing Scenario of Indian Agriculture. Commonwealth Publishers, New Delhi, India, pp. 98-145.
Dhuri, A. V., Singh, K. M. and Singh, R. N. (1986). Effect of intercropping on popula�on dynamics of insect pests of black gram.Indian J. Ent., 48: 329.
Ganapathy, N. (2010). Spo�ed Pod Borer, Maruca vitrata Geyer in Legumes: Ecology and Management. Madras Agric. J., 97(7-9): 199-211.
Giraddi, R.S., Amarnath, K., Chandrashekhra, K.B. and Pa�l, R.S. (2000). Late sowing and dry spell cause pest outbreak in kharif pulses. Insect Envt., 6: 124.
Gowda, D. K. S, Dhanaraj, Halle and Sharanbashappa (2004). Evalua�on of different IPM modules and intercropping systems for the management of pod borer in chickpea. Karnataka J. Agric. Sci., 17: 586.
Karel, A. K. (1993). Effects of intercropping with maize on the incidence and damage caused by pod borers of common beans. Environ. Entomol. 22: 1076-1083.
63

Pimberi, M. P. and Srivastava, C. P. (1989). Vegeta�on management and biological control of Helicoverpa armigera in chickpea. Int. Chickpea Newsl. 21: 16.
Sachan, J. N. and Lal, S. S. (1997). Integrated Pest management of pod borer complex in chickpea and pigeonpea in India. In: Recent Advances in Pulse Research, Asthana AN and Ali, M. (eds). pp. 349.
Sandeep, K., S. K. Kannaujia, Ajit Verma and Vinay Kumar Singh (2013). Study of the IPM module and net return in pigeonpea. Plant Archives, 13(1): 171-172.
Sharma, H.C., (1998). Bionomics, host plant resistance and management of legume pod borer, Maruca vitrata- a review. Crop Prot., 17: 373-386.
Singh, R. N. and Singh, K. M. (1978). Influence of intercropping on succession and popula�on buildup of insect pests of early varie�es of redgram. Indian J. Ent., 40: 361-375.
Sithanantham, S., Sethi, S.C. and Benitwal, S.P.S. (1984). A preliminary study of incidence of Aphis craccivora in chickpea at Hisar, India. Int. Chickpea Newsl. 10, 19–20.
Yadav, C. P., Lal, S. S., Dias, C. A. R. and Nigam, R. (1989). Host evasion-a prospec�ve approach for suppressing Heliothis armigera. Int Pigeonpea Newsl. 2: 62.
64

USE OF BIOPESTICIDES IN LEGUME PEST MANAGEMENT
V.S. MALIKDepartment of Entomology,
CCS Haryana Agricultural University, Hisar
Food legumes such as chickpea (Cicer arie�num L.), pigeonpea [Cajanus cajan (L.) Millsp.], cowpea (Vigna unguiculata Walp.), field pea (Pisum sa�vum L.), len�l (Lens culinaris Medik.), greengram [Vigna radiata (L.) Wilczek], blackgram [Vigna mungo (L.) Hepper], French bean (Phaseolus vulgaris L.), faba bean (Vicia faba L.), and grass pea (Lathyrus sa�vus L.) are the principle source of dietary protein among vegetarians and are an integral part of daily diet in several forms worldwide. They are low in fat and are excellent sources of protein, dietary fiber and a variety of micronutrients and phytochemicals. They are an important component of cropping systems to maintain soil health because of their ability to fix atmospheric nitrogen, extract water and nutrients from the deeper layers of the soil and add organic ma�er into the soil through leaf drop. However, food legumes are mainly grown under rain fed condi�ons and the produc�vity levels are low, mainly because of severe losses due to insect pests and diseases.
Grain legumes being a rich source of proteins, are damaged by a large number of insect pests, both under field condi�ons and in storage. These crops are damaged by a large number of insect pests, of which pod borers, Helicoverpa armigera and H. punc�gera; spo�ed pod borer, Maruca vitrata; spiny pod borer, E�ella zinckenella; pod fly, Melanagromyza obtusa; leaf miner, Liriomyza cicerina; stem fly, Ophiomyia phaseoli; pea and bean weevil, Sitona spp.; aphids, Aphis craccivora, Aphis fabae, and Acyrthosiphon pisum; white fly, Bemisia tabaci; defoliators, Spodoptera litura, S. exigua, and Amsacta spp.; lea�oppers, Empoasca spp., thrips, Megaleurothrips dorsalis, and Caliothrips indicus; blister beetles, Mylabris spp.; and the bruchids, Callasobruchus chinensis and Bruchus pisorum cause extensive losses worldwide. Insect pests in India cause an average of 30 per cent loss in pulses valued at $815 million, which at �mes can be 100 per cent. Being the rain fed, crops the use of costly agrochemicals for the management of these insect pests is not economically feasible for the resource constrained farmers. The problems associated with the excessive use of insec�cides such as resistance development, impacts on the human health and environment necessitates search for the cheaper and safer alterna�ves for the insect pest management in these crops. Biopes�cides are naturally occurring substances that control pests (biochemical pes�cides), microorganisms that control pests (microbial pes�cides), and pes�cidal substances produced by plants. The biopes�cides when used alone or in combina�on with the conven�onal insec�cides will help not only in reducing
65

the cost of pest management but also efficient management of the resistant popula�on of insect pests. Some of the commonly used biopes�cides and their role in the pest management in important legume crops are presented in this chapter.
Microbial biopes�cides:
Microbial pes�cides contain a microorganism (bacterium, fungus, virus, protozoan or algae) as the ac�ve ingredient. Microbial pes�cides can control different kinds of pests, although each separate ac�ve ingredient is rela�vely specific for its target pests.
Bacteria: Bacterial bio-pes�cides are probably the most widely used and cheaper than the other methods of pest bioregula�on. Insects can be infected with many species of bacteria but those belonging to the genus Bacillus are most widely used as pes�cides. One of the Bacillus species, Bacillus thuringiensis, has developed many molecular mechanisms to produce pes�cidal toxins; most of toxins are coded for by several cry genes. Since its discovery in 1901 as a microbial insec�cide, Bacillus thuringiensis has been widely used to control insect pests important in agriculture, forestry and medicine. Its principle characteris�c is the synthesis, during sporula�on, of a crystalline inclusion containing proteins known as
endotoxins or Cry proteins, which have insec�cidal proper�es. To date, over one hundred B. thuringiensis based bioinsec�cides have been developed, which are mostly used against lepidopteran, dipteran and coleopteran larvae. In addi�on, the genes that code for the insec�cidal crystal proteins have been successfully transferred into different crops plants, which have led to significant economic benefits. Spraying Bacillus thuringiensis (Bt) formula�ons in the evening results in be�er control than spraying at other �mes of the day in control of H.armigera in chickpea.
Laboratory efficacy of biopes�cides alone and in combina�on with insec�cides revealed that the treatment of endosulfan 0.07 per cent recorded significantly highest larval mortality (96.58%) and it was at par with Bt @ 1.0 kg/ha + endosulfan 0.035 per cent which recorded 95.60 per cent mortality of H. armigera larvae in pigeon pea (Khanapara and Kapadia, 2011). For the management of H.armigera infes�ng chickpea crop different combina�ons of Bacillus thuringiensis and HaNPV along with reduced concentra�ons of endosulfan were reported to be effec�ve (Bhalkare et al., 2007). Bt. Kurstaki (1liter/ha) with endosulfan (0.035%) give be�er yield in the field. Bt was also found effec�ve when combined with entomopathogenic fungi such as Metarhizium and Beauveria, resul�ng in be�er grain yield (Kale and Men, 2008). Singh and Ali (2005) tested the efficacies of Nuclear polyhedrosis virus, Ha NPV 250 LE/ha, Ha NPV 350 LE/ha, Ha NPV 450 LE/ha, B. thuringiensis- Bt 1 %(Biolep), neem seed kernel extract 5% and endosulfan 0.07 % against H. armigera in chick pea. Maximum larval mortality was recorded in endosulfan 0.07% followed by Bt formula�on @ 1 per cent and Ha NPV @ 450 LE/ha.
66

Baculoviruses: First well-documented introduc�on of baculovirus into the environment which resulted in effec�ve suppression of a pest occurred accidentally before the World War II. In the past, the applica�on of baculoviruses for the protec�on of agricultural annual crops, fruit orchards and forests has not matched their poten�al. The number of registered pes�cides based on baculovirus, though slowly, increases steadily. At present, it exceeds fi�y formula�ons, some of them being the same baculovirus prepara�ons distributed under different trade names in different countries. NPVs and GVs are used as pes�cides but the group based on nucleopolyhedrosis viruses is much larger. The first viral insec�cide Elcar™ was introduced by Sandoz Inc. in 1975. Elcar™ was a prepara�on of Heliothis zea NPV which is rela�vely broad range baculovirus and infects many species belonging to genera Helicoverpa and Heliothis. HzNPV is a product of choice for biocontrol of H. armigera . Countries with large areas of such crops like co�on, pigeonpea, tomato, pepper and maize, e.g. India and China, introduced special programs for the reduc�on of this pest by biological means.
12Applica�ons of samba isolate of NPV virus alone @ 1.5×10 OBs/ha in chickpea resulted in significant reduc�on in the pest damage and increase in the marketable yield. Among the Ha NPV isolates, CBE I (Coimbatore) and NEG (Negamum) applied at
121.5 × 10 POB/ha in chickpea with an adjuvant, crude sugar, significantly reduced the H. armigera larval popula�on and increased the yield under field condi�ons. In chickpea grain yields of 980, 983, and 973.3 kg/ha and cost/benefit ra�os of 1:1.36, 1:1.48 and 1:0.87, respec�vely, in CBE I, NEG and endosulfan treated plots were obtained. The isolate RAJ (Rajasthan) recorded the lowest yield comparable to that of the untreated control (Jeyarani et al. 2010).
Entomopathogenic fungi: Entomopathogenic fungi are important natural regulators of insect popula�ons and have poten�al as mycoinsec�cide agents against diverse insect pests in agriculture. These fungi infect their hosts by penetra�ng through the cu�cle, gaining access to the hemolymph, producing toxins and grow by u�lizing nutrients present in the haemocoel to avoid insect immune responses. Entomopathogenic fungi may be applied in the form of conidia or mycelium which sporulates a�er applica�on. The use of fungal entomopathogens as alterna�ve to insec�cide or combined applica�on of insec�cide with fungal entomopathogens could be very useful for insec�cide resistant management. The fungi such as Beauveria bassiana, Nomuraea rileyi, Metarhizium anisopliae and Ver�cilium lecani were show their effec�veness in controlling many insect pests that infest the legume crops.
6 The entomopathogenic fungus Nomuraea rileyi (@ 10 spores per ml) resulted in 90 to 100 per cent larval mortality of H.armigera, while Beauveria bassiana (@ 2.68 X
7 10 spores per ml) resulted in 6 per cent damage by H.armigera on chickpea compared to 16.3 per cent damage in untreated plots (Saxena and Ahmad,1997). The
67

bioinoculants such as Beauveria bassiana and Pseudomonas fluorescens talc formula�ons were tested against whitefly Bemisia tabaci, leaf hopper Empoasca kerri, legume pod borer Maruca vitrata and flea beetle, Madurasia obscurella in Urd bean. The P. fluorescens seed treated plots had lower whitefly popula�on compared to B.bassiana treated plots and untreated check. The foliar applica�on of B.bassiana showed effec�ve results on the pod borer complex. The damage by M. vitrata, H. armigera, L. boe�cus was low in the harvested pods as against the untreated control (Soundararajan and Chitra, 2011). Sreeekanth and Sheshamahalakshmi (2010) observed per cent inflorescence damage due to legume pod borer in pigeonpea was lowest in spinosad 45% SC @ 73 g a.i/ha (4.74%), followed by Bacillus thuringiensis-1 @ 1.5 kg/ha (10.52%) and Beauveria bassiana SC formula�on @ 300mg/Lt (14.15%) with 80.9, 57.6 and 42.9 per cent reduc�on over control, respec�vely as against control (24.79%).
12The treatment with M.anisopliae (isolate M-34412) applied @ 5×10 conidia/ha showed the remarkable reduc�on in the pod damage caused by the H. armigera resul�ng in higher grain yield by 10.84 % and 6.06 % over conven�onal prac�ce in pigeonpea and chickpea, respec�vely. The per cent mortality of pod borer with Metarhizium M-34412 and also two other isolates namely M-34311 and M-81123, ranged from 65-72 per cent and this was comparable to the performance of the chemical insec�cide endosulfan (74%) in chickpea.(Kulkarni et al., 2008)
Plant based biopes�cides: Use of botanicals is now emerging as one of the important means to be used in protec�on of crop produce and the environment from pes�cidal pollu�on, which is a global problem. Pes�cidal plants contain a mixture of ac�ve substances that may have many advantages in terms of efficacy and preven�ng the development of insec�cide resistance when compared to synthe�c insec�cides. Hundreds of na�ve plant species have been evaluated against range of insect pests on various crops. Botanical insec�cides act as a synergis�c component in several IPM strategies. Neem tops the list of 2,400 plant species that are reported to have pes�cidal proper�es and is regarded as the most reliable source of eco-friendly biopes�cidal property. Neem products are effec�ve against more than 350 species of arthropods, 12 species of nematodes, 15 species of fungi, three viruses, two species of snails and one crustacean species.
Neem Seed Kernel Extract (NSKE) was found most effec�ve in reducing the larval popula�on of H. armigera in chickpea and pod damage. Vegetable oils, neem oil and karanj oil provide effec�ve protec�on against bruchid damage in pulses (Reddy et al., 1994). Karanj oil, and leaf and seed extracts act as oviposi�on deterrents (Kumar and Singh, 2002). Neem and custard apple extracts and neem and karanj (Pongamia) oil based formula�ons have also been recommended for the management of H. armigera. Neem Seed Kernel Extract (5%) + DDVP @ 0.5 ml/ liter of water were found next best treatment for the management of spo�ed pod borer in pigeon pea a�er Profenofos + DDVP@ 0.5 ml/ liter.
68

Three biopes�cide prepara�ons (crude neem kernel seed extract (5%), neem oil (3000 ppm) and Pongamia oil were tested against pigeon pea pod fly recorded higher grain yield in comparison to control. Among these biopes�cides crude neem seed extract outperformed other biopes�cides in terms of per cent increase in the grain yield (Sharma et al., 2011). Spraying of NSKE (5%) resulted in good control of pulse beetle under field condi�ons. The mean per cent pod damage was significantly different between the treatments. NSKE (5%) recorded the lowest pod damage (4.64 %) when compared to malathion (5.96%) and spinosad (6.30%).
Biochemical Pes�cides:
Sex pheromones: Insect sex pheromones are biochemical pes�cides and have long been used as monitoring and mass-trapping tools in IPM strategies. Several sex pheromone lures including insects like pod borer, tobacco cutworm, and legume pod borer are commercially available. The pheromone traps are installed for monitoring the tobacco cut worm, gram pod borer and legume pod borers is an essen�al component of the integrated pest management modules against legume insect pests.
Plant vola�les: Certain secondary metabolites in plants act as deterrents for generalist feeders, and a�ractants for specialist feeders. For instance, glucosinolates and their metabolites act as a�ractants and s�mulants for specialist feeders. The plant vola�les can be effec�vely used for the manipula�on of pest popula�on if the role of the plant vola�les is specifically established. Applica�on of plant vola�le, cis jasmonate enhanced the parasi�sa�on percentage of bean aphid by Aphidus ervi. Similarly plant vola�les are sprayed on the plants to increase the parasi�sa�on by the egg parasi�ods such as Trichogramma.
Integrated pest management modules containing biopes�cides as major components:
Several biointensive modules containing biopes�cides were tested against the insect pests affec�ng the legume crops. Some of the bio intensive modules are scored best in controlling the insect pests compared farmers prac�ces that solely depend on the use of conven�onal insec�cides. The IPM module mainly comprised of seed treatment (Trichoderma + vitavax), pheromone traps for monitoring of Helicoverpa, spray of HaNPV followed by NSKE 5% and spray of endosulfan as last resort resulted in lower damage by pod bores and pod fly in pigeon pea and chick pea. Sequen�al applica�on of NSKE-HaNPV-Bt was effec�ve against insect pests of organically grown field pea. HaNPV was effec�ve against H. armigera larvae, but ineffec�ve to other pod borers. Among biopes�cides treated plots, sequen�al applica�on of NSKE-HaNPV-Bt recorded higher grain yield in field bean (Byrappa et al., 2010). Pigeonpea + sorghum intercropping with biointensive IPM module consis�ng of applica�on of Neem seed extract (5%), HaNPV 250 LE/ha and Bacillus thuringiensis (Bt) @ 1 kg/ha followed the pes�cide based IPM for the integrated pest management of pigeonpea pests.
69

Adop�on of IPM module in blackgram consis�ng of the components viz., seed treatment, erec�on of bird perches, installa�on of pheromone traps against Spodoptera litura and Helicoverpa armigera, use of bio-ra�onal pes�cides viz., Neem Seed Kernel Extract, Sl NPV/Ha NPV, mechanical management and need based applica�on of insec�cides registered reduced pod borer damage (4.2-10.8 %), and pod bug damage (2.7-5.8 %) as against the farmers' prac�ce viz.,dus�ng with lindane 1.3 D @ 25 kg/ha at peak flowering, which registered higher pod borer damage (5.8-13.8 %), and pod bug damage (3.8-7.8 %). The grain yield (528-852 kg/ha.) and Benefit: Cost (1.37-2.21) were also higher in IPM plots compared to farmer's prac�ce (Gajendran et al., 2006)
Conclusion:
Biopes�cides when used in alone or in combina�on with biochemical insec�cides proved to be as effec�ve as the conven�onal insec�cides. Although biopes�cides increasingly are being used as alterna�ve pest management strategies, several constraints such as developing stable formula�ons, standardizing appropriate delivery methods, lack of registra�on procedures, etc. are associated with their introduc�on and promo�on in most of the developing world. Improving stability would enhance the performance of biopes�cides under field condi�ons. In the case of microbial pes�cides, the formula�ons should maintain the viability of the spores. For example, the insolubility and poor stability of the ac�ve cons�tuent azadirach�n in water have limited the use of neem as a safe and effec�ve insec�cide for systemic applica�on. There is need to develop the stable formula�ons of microbial insec�cides and documenta�on and scien�fic valida�on of the plant based biopes�cides is need of the hour. The higher cost and lack of �mely availability of the some of the biopes�cides led to failure of much poten�al bio intensive integrated pest management.
Suggested Reading:
Bhalkare, S.K., Supare, N.R., Nimbalkar, S.K. and Sarode, S.V. (2007). Bio intensive m a n a g e m e n t o f p o d b o r e r , H e l i c o v e r p a a r m i g e r a (Hubner)(Lepidoptera:Noctuidae) in chickpea crop. J. Biol. Control., 21:17-24.
Byrappa, A.M., Kumar, N.G., and Divya, M. (2012). Impact of biopes�cides applica�on on pod borer complex in organically grown field bean ecosystem. J Biopest., 5(2): 148-160.
Gajendran, G., Chandrasekaran, M. and Jebaraj, S. (2006). Evalua�on of integrated pest management module against major pests of blackgram in rainfed ecosystem. Legume Res., 29 (1): 53 – 56.
Jeyarani, S., Sathiah, N. and Karuppuchamy, P. (2010). Field efficacy of Helicoverpa armigera Nucleopolyhedrovirus Isolates against H. armigera (Hubner) (Lepidoptera: Noctuidae) on co�on and chickpea. Plant Protect. Sci,. 46(3): 116–122.
70

Kale, S.N. and Men, U.B. (2008). Efficacy of microbial insec�cides and their combina�ons against Helicoverpa armigera (Hubner) in chickpea. J. Biol. Control., 22:205-208.
Khanapara, A.V. and Kapadia, M.N. (2011) . Efficacy of Bio-pes�cides alone and in combina�on with insec�cides against Helicoverpa armigera on pigeonpea. Res. J. Agri. Sci., 2(2): 340-343.
Kulkarni, S.A., Ghormade, V., Kulkarni, G., Kapoor, M., Chavan, S.B., Rajendran, A., Pa�l, S.K. , Shouche, Y. and Deshpande, M.V.(2008). Comparison of Metarhizium isolates for biocontrol of Helicoverpa armigera (Lepidoptera: Noctuidae) in chickpea. Biocontrol Sci. Technol.18: 809-828.
Kumar, M. and Singh. R. (2002). Poten�al of Pongamia glabra Vent. as an insec�cide of plant origin. Biol. Agri. Hort., 20: 29-50.
Reddy, V.S. Babu , T. R. Hussaini, S. H, and Reddy, B. M. (1994) Effect of edible and non-edible ol1s on the development of pulse beetle, Callasobruchus chinensis L and on viability of mungbean seeds. Pest Manage. Eco. Zool, 2: 15-17
Saxena, H. and Ahmad, R. (1997). FIeld evalua�on of Beauveria bassiana (Balsamo) Vuillemm against Heilcoverpa armigera (Hubner) infes�ng chickpea. J. Biol. Control,, 11: 93-96.
Sharma, O. P., Bhosle, B. B., Kamble, K. R., Bhede, B. V. and Seeras, N. R.(2011). Management of pigeonpea pod borers with special reference to pod fly (Melanagromyza obtusa). Indian J. Agri.Sci., 81 (6): 539–43.
Singh, R. and Ali, S. (2005) Efficacy of biopes�cides on the management of Helicoverpa armigera (Hubner) in chickpea. Ann. Pl. Protec. Sci., 13: 94-96.
Soundararajan, R.P. and Chitra, N. (2011). Effect of bioinoculants on sucking pests and pod borer complex in urdbean. J Biopest., 4 (1): 7 – 11.
Sreekanth, M. and Seshamahalakshmi, M. (2012). Studies on rela�ve toxicity of biopes�cides to Helicoverpa armigera (Hubner) and Maruca vitrata (Geyer) on pigeonpea (Cajanus cajan L.). J Biopest., 5(2): 191-195.
71

MANAGEMENT STRATEGIES OF LEPIDOPTEROUS INSECT PESTS ESPECIALLY THROUGH NUCLEAR POLYHEDROSIS VIRUS
H.D.KAUSHIKDepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
There are various insects feed upon from seedling stage of the pulse crops but they cause most of the economic damage to flowers and pods. More than 200 insect species have been recorded in India and a few of them cause severe damage at pod forma�on stage leading to significant reduc�on in the yield. Of these, pulse pod borer, Helicoverpa armigera(Hubner); hairy caterpillars, Spilarc�a obliqua Walker and Amsacta moorei (Butler); cutworm, Agro�s ypsilon (Huf.) and Agro�s flammatra Schiffer-Mueller; semilooper, Autographa nigrisigna (Walker),tur pod bug, Clavigralla gibbosa Spinola; tur pod fly, Melanagromyza obtusa Malloch; blister beetle, Mylabris spp.; bean black aphid, Aphis craccivora Koch, whitefly, Bemisia tabaci Gen. and termites, Ondontotermes obesus (Rambur) and Microtermes obesi Holm are of economic importance (Raheja et al., 1991). Very less interest has been shown towards the plant protec�on measures to check their a�ack where crop returns are not assured under dry land condi�ons. Number of management strategies based on use of insec�cides have been recommended and �me and again followed over a large area resul�ng into various problems. Much emphasis has always been made on almost total reliance on chemical control instead of natural and biocontrol. The applica�on of specific pes�cides to control directly the few pest species which surpass the economic damage level in a given crop has been the main principle of integrated pest management. To implement this, our inten�on is to leave the ecosystem as undisturbed as possible in order to protect and take full advantage of natural enemies in the intact fauna. Most of the chemical insec�cides are broad spectrum and do not fit properly into those strategies. It is obvious that in integrated control programs, a single us of broad spectrum pes�cide can annihilate the effect of all other selec�ve control measures. Therefore, lack of selec�ve insec�cides was become a major problem in these programmes. Agriculture without pest control measures is not economically feasible, either for producer or consumer. There are many methods of biological control which can give relief in these situa�ons. Fortunately enough, nature itself has provided highly specific insec�cides as insect viruses.
Biological control in totality will never replace chemical insec�cides though it maintains the hazards free ecosystem. The majority of bio-control agents are compa�ble with other conven�onal methods of control to sustain the agro ecosystem which are cost effec�ve and self sustaining too. Besides parasitoids and predators,
72

several entomopathogens such as viruses like nuclear polyhedrosis virus (NPV) and granulosis virus (GV), fungi like Beauveria, Ver�cillium, Metarhizium, bacteria like Bacillus thuringiensis (Bt) are also known to cause diseases in insects thereby destroying them. Among the pathogens, nuclear polyhedrosis virus (NPV) found promising and effec�ve in checking the lepidoplerous pests of pulses. There viruses can produce either lethal infec�ons or epizoo�es or chronic diseases in pest popula�on. Because of narrow host range (host specific), Baculoviruses group have been studied extensively as effec�ve entomopathogen. Polyhedral occlusion bodies having vision as ac�ve cons�tuents are ingested/infected per os to create infec�on in the insect haemocoel.
Insect viruses: Viruses pathogenic for insects do not form a taxonomic en�ty. We can find them in such different virus families as Iridoviridae, Parvoviridae, Poxviridae, Reoviridae, and Baculoviridae. Most of these families also have representa�ves which infect not only insects but also vertebrates or even plants. There is one excep�on to this: the members of the family of the Baculoviridae, the baculoviruses, which infect arthropods only. Un�l today no baculovirus has ever been isolated from a non-arthropod host. They have been found in crustacea and mites, but mostly in insects. This absence from organisms other then arthropods is strong indirect evidence that baculoviruses are safe for man and the environment. This is one of the main reasons why it is mostly these viruses that are being used in pest control. About 523 species of insects from 52 families and 8 orders were recorded as hosts of NPVs. Of the 523 species, 455 species are in the order Lepidoptera and 107 species in the family Noctuidae, which contains many agricultural pests of economic importance. The order Hymenoptera has 31 species listed as hosts of NPVs, 19 of which are in the family Diprionidae and are pests of forest and shade trees. The order Diptera has 27 species as hosts of NPVs, 20 of them are in the family culicidae. Most NPVs are pathogenic to only a few closely related species of insects. Most NPVs loose 50% or more of their original ac�vity within a day or two when sprayed on plants in the field by UV rays and plant foliage factors. However, NPVs can persist for years in the soil which acts as a reservoir of virus and provides inoculum to ini�ate new infec�ons each year. These NPVs can be augmented in the field for effec�ve pest suppression.
Poten�al in pest control: Baculoviruses are characterized by double stranded circular DNA which is included in rod shaped capsids. They are formed mostly in the nucleus of the host cells. In common with many insect viruses from other virus families, the virions of most baculoviruses are contained within proteinaceous par�cles called occlusion bodies, which o�en have a polyhedral shape. The thick layers of protein provide protec�on against adverse physical and chemical factors in the environment, allow survival outside the host and enable the viruses to kill their host rapidly without jeopardizing their own existence. It is obvious that the good protec�on of the virus
73

par�cles by the occlusion bodies also is of a great advantage for the use of these viruses as biological insec�cides. Since they are so resistant, in general, they can be used like chemical pes�cides.
Baculoviruses have several proper�es which make them ideally suited for use in integrated plant protec�on programmes. Their most important a�ribute in this regard is their extremely high host specificity. This means, when used in plant protec�on against a given pest species, only the target pest is affected and all the beneficial or even neutral arthropod species in the same ecosystem are le� unharmed. Therefore, the whole poten�al of the natural antagonists of the pest, present in an intact ecosystem, can be exploited. Many secondary pests are kept below economic damage levels and the necessity for addi�onal plant protec�on measures is greatly reduced. As a consequence, a treadmill situa�on as o�en results from the use of broad spectrum pes�cides can be avoided. At the same �me, selec�vity means that baculoviruses are harmless for man, environmentally and ecologically safe, and do not present residue problems.In contrast to chemicals, viruses have the ability to mul�ply in their host. A�er dissemina�on for pest control, they may persist or even spread in the popula�on of the target pest. They can even ini�ate real epizoo�cs which may keep the pest at a low level for several years and make further control measures unnecessary. So far we have no indica�on that resistance in the pest popula�on will become a major problem, as it is for many chemical pes�cides. In nature, insect popula�ons have been exposed to viruses for thousands of years without ever becoming resistant. From the more than 1100 insect viruses known, about 60% belong to the baculoviruses. There are es�mates that baculoviruses can be used against nearly 30% of all the major pests of food and fibre crops. In Central America, by replacing chemical insec�cides with insect viruses, pes�cide consump�on could be reduced by nearly 80%. This vast poten�al for pest control so far has hardly been exploited.
Helicoverpa armigera: Research over the last three decades on the NPV of H. armigera (HaNPV) has been focused on the natural occurrence of the virus, virus-host rela�onship, mass produc�on and formula�on, field tes�ng and safety evalua�on against several non-target species. The gram pod borer, armigera is an important and a serious pest of pulses in India and damages both chickpea and pigeonpea. On an average 30% crop loss is reported. Due to absence of true resistant varie�es in both the crops, resor�ng to chemicals had been the only alterna�ve Among various alterna�ves to chemicals microbial control with nuclear polyhedrosis virus (NPV) is favoured for the management of H. armigera on many crops including chickpea (Jayaraj et al., 1985). It is mainly due to its host specificity, safety to ecosystem and having no effect on non-target species so that all can work together.The following review traces some of the research and developmental ac�vi�es, which have led to the commercial scale produc�on and registra�on of HaNPV as a microbial pes�cide in India. The occurrence of the virus was
74

first reported by Patel et al. (1968) in Gujarat. Rabindra and Subramaniam (1974) described a very virulent NPV which was singly occluded. Sudhakar and Mathavan (1999) reported a mul�ply occluded NPV from H. armigera. Natural occurrence of the virus was also reported from Maharashtra by Chundurwar and Pawar (1991). Being an obligate pathogen, the virus can be mass produced only in H. armigera larvae. Sathiah (2001) manipula�ng the inoculum dose, temperature of incuba�on and stage of larvae
10could obtain a high yield of 1.2x10 polyhedral occlusion bodies (POB)/larva. Earlier
9reports have indicated lower yields of 2x10 POB/larva. Obtaining high yield/larva is cri�cal to economic produc�on of the virus. A successful technique of mass produc�on of H. armigera for in vivo produc�on of the virus has also been developed (Sathiah, 2001). Comparing different semisynthe�c diets for H. armigera mass produc�on for in vivo NPV produc�on Srinivasan et al. (1998) found that the chickpea flour based diet was significantly be�er than cowpea or french bean based diets. However, french bean flour subs�tuted at 75% was the most cost-effec�ve diet for NPV produc�on.
Adjuvants enhancing the efficacy of the virus: Several adjuvants with phago-s�mulant and possible UV protectant property were found to increase the efficacy of the virus. Dhandapani et al. (1994) incorporated NPV into different bait formula�ons containing various combina�ons of maize, soybean or chickpea flour, crude sugar, co�on seed kernel and extracts of larvae of H. armigera. Results of bioassays revealed that baits containing chickpea or maize flour along with H. armigera larval extracts were the most effec�ve. Adjuvants may also protect the virus from inac�va�on by the leaf environment. These results indicate the role of adjuvants in increasing the persistence of the virus on the crop plants.Host plant surface environment was shown to reduce the ac�vity of the virus enormously. Co�on followed by chickpea leaf environment was found to be the most detrimental to the virus. (Rabindra et al., 1994). The LC was 50
lowest on lablab followed by sunflower but highest on co�on. More or less a similar trend was observed with LT . Among the different plant parts of co�on, the leaf 50
environment was the most detrimental. This detrimental effect, however, could be offset by the addi�on of an adjuvant mix consis�ng of co�on seed kernel extract 10% + crude sugar 10%. The compa�bility of NPV with host plant resistance was studied by Rabindra et al. (1992). The mortality caused by NPV in H. armigera larvae was much higher on a suscep�ble chickpea-Shoba than on a resistant variety ICC506. It is well known that the performance of NPV can be increased when sprayed in the evening hours through enhanced persistence. But not many field experiments have been conducted with HaNPV. Pokharkar and Chaudhary (1999) found that applica�on of the virus in the evening was significantly be�er than applica�on in the morning hours in reducing the larval popula�on and increasing the yield of chickpea. NPV would have to be integrated with insec�cides like endosulfan for the management of H. armigera on crops like pigeon pea and co�on, which are infested with other lepidopterous pests and in such an event, the NPV would be expected to enhance the suscep�bility to
75

pes�cides. This phenomenon would make NPV a powerful component of IPM and an important tool in insec�cide resistance management (IPM).
Field efficacy of HaNPV: A large number of field experiments on the efficacy of NPV against H. armigera on chickpea in different states of India have been conducted (Table 6). On chickpea the efficacy of HaNPV (Helicoverpa armigera NPV) has been demonstrated in Gujarat and Hyderabad where high mortality of Helicoverpa larvae is obtained. Around 47% increase in grain yield has been indicated over untreated
10controls. In TamilNadu HaNPV at the rate of 1.5x10 polyhedral occlusion bodies (POB)/ha sprayed three to four �mes in the evening hours at an interval of 7-10 days when the pest is in the very early stages, effec�vely controlled Helicoverpa on pigeonpea, (Santharam & Balasubramaniam,1981) chickpea, (Rabindra & Jayaraj,1987) lab lab bean (Jayaraj et al.,1987) and blackgram (Mahadevan & Kumaraswamy,1980). Applica�on of HaNPV alongwith 0.035% endosulfan has been found to give be�er control of Helicoverpa on chickpea (Rabindra & Jayaraj, 1987) and pigeonpea (Santharam & Balasubramaniam, 1981). There was effec�ve control with 250 LE of HaNPV/ha or 125 LE HaNPV/ha+endosulfan 0.035% against Helicoverpa on chickpea in Hyderabad (Anonymous, 1993). Two spraying of endosulfan 0.07% or HaNPV @ 500 LE/ha alternated with endosulfan significantly reduced the pod damage in pigeonpea and increased grain yield (Snap & Deshmukh, 1988). The results reveal
12that the virus at 1.5x10 POB/ha applied 4-5 �mes when the larval popula�on is in the early states gave effec�ve control of the pest and increased the grain yield. Ultra low
12volume applica�on of the virus at 1.5x10 POB/ha along with 20 per cent crude sugar gave control equivalent to that of endosulfan (Rabindra and Jayaraj, 1988). The virus at
12a concentra�on of 1.5x10 POB/ha was found to control the popula�on of H. armigera and increase the yield of pigeonpea. The virus has been used in combina�on with other components of pest management like B.t., chemical insec�cides, azadirach�n, neem seed kernel extract, Chrysoperla carnea and bird perches (such integra�on would be necessary when mixed popula�ons of different instars par�cularly of grown up larvae of H. armigera or different species of pests occur along with H. armigera on crops like pigeonpea or co�on.Various spray equipment have been used for applying the virus. Controlled droplet applica�on of the virus with 20% crude sugar as an adjuvant was found to give very good control of H. armigera on chickpea (Rabindra and Jayaraj, 1988). Applica�on of the virus with the CDA sprayer was also found effec�ve against H. armigera on pigeonpea (Muthiah and Rabindra, 1991). These reports indicate that the virus may be applied with any of the spray equipment. Among the several alterna�ve methods for management of H. armigera, the nuclear polyhedrosis virus (NPV) is promising, whereas Trichoderma sp. have shown promising results against chickpea wilt. The biocontrol package against H. armigera and wilt disease under field condi�ons were evaluated and lowest H. armigera larval popula�on (0.71 larvae/plant) was recorded in chemical control, which was at a par with biocontrol package (0.91
76

larvae/plant), and both the treatments were significantly be�er than control. Lowest per cent pod damage (3.85%) was recorded in chemical control followed by biocontrol treatment (5.08%) and unsprayed control (8.61%). The yields from biocontrol package (13.45 q/ha) and chemical control (15.37 q/ha) were significantly higher than unsprayed control (10.7 q/ha).There was no disease incidence in all treatments in both 2008 and 2009 (Joshi and Virk, 2011).
Spodoptera litura: The NPV of S. litura was reported by Ramakrishnan and Tiwari (1969). An epizoo�c occurrence of the virus was recorded by Dhandapani et al. (1982) on S. litura larvae migra�ng to Sesbania bispinosa.The rela�ve persistence of SlNPV was studied on soybean, cabbage, co�on, castor bean, groundnut, mulberry, potato, straw berry, sunflower and tomato plants by Kulkarni and Hugar (1999). Results of bioassays showed that three days a�er spray, persistence was highest on soybean and groundnut but lowest on co�on.
Field efficacy: Control of S.litura has been studied on banana, blackgram, castor, cauliflower, co�on and tobacco using the NPV alone or in combina�on with low concentra�ons of insec�cides. Reduc�on in plant damage was assessed in some experiments. The leaf damage index (05 scale) in co�on was reduced from 3.84 in
12 control to 2.03 following the applica�on of NPV at 1.5x10 POB/ha (Jayaraj et al., 1981).
9Spilosoma obliqua: Applica�on of SoNPV @ 10 POB/ml recorded 0.1 larvae/plant as against 2.10 in control and the virus persisted on the foliage of cowpea for a period of 5 days with 53.3% of its original ac�vity and the LC for 4, 12 and 16 day old larvae were 50
4 6 62.6 x 10 , 1.1 x 10 and 3.16 x 10 POB/ml (Chaudhari, 1997).
Limita�ons: It cannot be denied that the wider use of insect viruses in pest control is confronted with some reluctance. Insect viruses have also some features which are nega�ve, par�cularly with regard to their economic use in plant protec�on. As profitable and desirable specificity is from an ecological viewpoint, it causes problems with regard to the economics of the commercializa�on of viral pes�cides. In most countries, insect viruses have to be officially registered for use as insec�cides and are subjected to the same regula�ons as chemical pes�cides. Therefore, the expenses for their commercializa�on are in the same order of magnitude as for conven�onal insec�cides. But, due to their selec�vity, their market size and their sales poten�al is very limited - an aspect which is not very a�rac�ve for a poten�al producer, especially not for big companies which have the necessary experience for handling registra�on, but which need a big market to make produc�on profitable. As a consequence, microbial pes�cides, if they ever make it to the market, find it difficult to compete in price with chemical pes�cides. Furthermore, the use of selec�ve pes�cides in the framework of integrated pest management programmes requires good knowledge of
77

the biology of the pest species and their antagonists in the crop, and is therefore not as simple as the use of broad spectrum chemicals. Since insect viruses do not act on contact and are less persistent than most chemical pes�cides, much more a�en�on has to be given to the correct �ming and applica�on of the sprays. All this makes the use of selec�ve pes�cides more cumbersome and more costly for the farmer. So, whereas the main advantage of using a selec�ve insec�cide is on an environmental and social level, there is usually hardly any immediate economic benefit for the farmer as the direct user of the product. The basic constraints for a wider use of viral pes�cides arise therefore from the fact that the nega�ve and the posi�ve aspects of the use of selec�ve pes�cides do not afflict the same group of people.
Future prospects: Recent advantages in gene�c engineering have raised hopes that this novel technique would be capable of improving some of the nega�ve pes�cide quali�es of insect viruses. It has been suggested for instance to create viruses which a) exhibit a broader host range, b) produce a toxin for faster kill, c) show increased virulence, or d) have a be�er environmental stability than natural occurring parent viruses. Since, as we have seen above, the-main hindrance for wider use of insect viruses is not so much lack of efficacy as economical constraints; this would not solve the problem anyhow.To summarize, only if we accept that the preserva�on of nature has its price and if we are willing to pay it, then, and only then, will viral pes�cides be as widely esteemed and used in the future as they should be with regard to our environment.
Suggested Reading
Chaudhari, S. 1997. Indian J.Entomol.., 59(1): 5961.
Joshi, N. and Virk, J. S.2011 Archives Phytopath Plt. Protect., 44|(18)|: 1815–1821.
Kulkarni, G. G. and Hugar, P. S. 1999. Haryana Agril. Univ. J. Res., 29(3-4): 95-99.
Muthiah, C. and Rabindra, R. J. 1991. Indian J. Agricul. Sci., 61(6): 449-452.
Pokharkar, D.S. and Chaudhary, S. D. 1999. J. Maharashtra Agril. Univ., 24(2): 147-151.
Rabindra, R. J. and Balasubramanian, M. 1980. Current Sci., 49: 279.
Rabindra, R. J. and Jayaraj, S. 1988. Trop. Pest Mgmt., 34, 441-444.
Rabindra, R. J. and Jayaraj, S. 1990. J. Biol. Contr., 4(1): 31-34.
Rabindra, R. J. and Jayaraj, S. 1995. J.Biol. Contr., 9: 34-36.
Rabindra, R. J., Sathiah, N. and Jayaraj, S. 1992. Crop Protect., 11(4): 320-322.
Rabindra, R. J., Muthuswami, M. and Jayaraj, S. 1994. J. Appl. Ent., 118: 4-5, 453-460.
78

MANAGEMENT STRATEGIES FOR HAIRY CATERPILLARS IN LEGUME CROPS
OMBIR AND R.K. SAINI Department of Entomology,
CCS Haryana Agricultural University, Hisar
Different species of genus Amsacta Family Arc�idae) a�acking legume crops are: Amsacta moorei Butler, Amsacta albistriga Walker, Amsacta lineola Fabricius, and Amsacta flavicosta Hampson).Another species damaging legume crops is Spilosoma (= Diacrisa) obliqua Walker (Arc�idae : Lepidoptera). Family Arc�idae includes small to moderate sized moths commonly called as �ger moths or woolly bears, o�en of bright colouring. The white colour may be a form of warning colouring. The body is robust and hairy or well scaled. The round sculptured eggs are commonly laid in clusters. Larvae are commonly hairy, with tu�s of long hair or dense uniform clothing. All the known larvae are herbivorous; pupa�on takes place on the soil in a cocoon of silk and hair. Hiberna�on is passed as a pupa in the cocoon in a sheltered place or in the soil. The majority of the moths do not emerge un�l the first heavy fall of rain. There are excep�ons to this rule, some moths emerging in the dry hot weather. The number of broods vary from 1-8. Several species are injurious owing to their very great mul�plica�on under favourable circumstances and to their omnivorous habits. They are the common “hairy caterpillars” which are well known pest in plains. The larvae curl into compact mass when disturbed and fall of the plants and feign death and are called woolly bears. They are parasi�zed by parasi�c Hymenoptera and Tachinidae.
1. Red hairy caterpillar, Amsacta moorei Butler (Arc�idae: Lepidoptera)
Distribu�on: A moorei is typically a pest of arid sandy zones where rainfall is scanty and irriga�on facili�es are poor and soil remains rela�vely less disturbed over longer periods. This species is predominant in the Indian sub-con�nent though it has been reported from Senegal and China also. It is widely distributed in the northern India, par�cularly in the states of Rajasthan, Gujarat, Haryana and Punjab.
Host range and nature of damage: Vigna acconi�folius, Vigna mungo, Cyamopsis tetragonoloba, Vigna radiata, Crotalaria juncea, Arachis hypogaea, Citrullus lanatus, Pennisetum typhoides, Ricinus communis, Cajanus cajan, Sesamum indicum, Vigna unguiculata, Glycine max. The host range of A. moorei is very wide. It feeds on virtually all vegeta�on (cul�vated or wild) growing in or around the fields during rainy (end June to September) season. Among the cul�vated hosts, a number of plants a�acked belong to the families Leguminoseae and Gramineae. However, plants of some other families, namely, Malvaceae, Compositae, Cucurbitaceae, Pedaliaceae, Euphorbiaceae, etc. are also frequently a�acked. Some of the common weed hosts like puncture vine (Tribulus
79

terrestris) and carpet weed (Trianthema portulacastrum) are equally good in suppor�ng the development and survival of this pest.
Early symptoms of larval damage are characterized by the presence of leaves completely devoid of chlorophyll leaving behind the network of veins intact. Minute pellets of larval excreta and/or o�en early instars feeding gregariously in large numbers on the undersurface of leaves are also visible. The early instar larval damage is generally no�ced in pockets in the field. The late instars feed on leaves, stems, growing points and whole plant (in seedling stage) causing extensive damage. Some�mes, resowing of the crop may be necessitated if seedlings are damaged heavily.
Morphology
Egg: Eggs are spherical, about 7 mm in size, creamish yellow to yellow which change to orange red and finally dark brown to blackish brown in colour before hatching. They are laid in clusters.
Larva: Newly hatched larvae are about 1.25 to 1.5 mm long, light brown in colour and possess long grayish brown hair; mature larvae reach a length of about 44 mm. The head is reddish brown and the body carries tu�s of long hair arising from the fleshy tubercles. Mathur (1966) and Chaudhary et al. (1971) have categorized larvae on the basis of colour pa�ern e.g. larvae having whole body with reddish brown hair or black hair or with a mixture of these or anterior and/or posterior ends with black hair and the remaining part with reddish brown hair. However, about 50 per cent larvae have reddish brown hair on the whole body.
Pupa: Pupae are 17-20 mm long, 7-8 mm broad and dark reddish brown to blackish brown in colour. Their shape is peculiar being quite broader in the middle and tapering towards both ends. Pupae are found enclosed in cocoons formed with the help of silken threads and shed larval hair.
Adult: The adults are strongly built moths with white wings having black spots. Forewings are with scarlet costa. Anterior margin of thorax and abdomen is scarlet. There are black bands and dots on abdomen. Hind wings have black elongated spots along the posterior outer margin. Most adults have above colour pa�ern but a few have the scarlet colour replaced by yellow. The females are larger (20-22 mm) than the males (18-20 mm). Wing expanse of females is 35-50 mm while that of males 36-44 mm. A. moorei resembles A. albistriga in appearance and habits and cross-pairing between the two species has been observed.
Biology and ecology: Adult emergence starts with first monsoon showers during end of June. The moths invariably emerge during night and early hours of the day and mate within 10 to 36 hours a�er emergence. Moths are sluggish and have a rather clumsy flight. The females, in par�cular, which may be filled with several hundred eggs, fly very
80

li�le are easily differen�ated from males by their larger size, tubular body with a large slit like gonopore and by being more sluggish. The copula�on generally starts a�er 8.00 p.m. and copulatory period lasts for 6.5 to 9.5 hours. Moths almost invariably mate once and start laying eggs about half an hour to four hours a�er end of copula�on. Eggs are laid in irregular batches of 40 to 600 eggs. Females do not require any food for laying fer�le eggs and oviposi�on con�nues for 2 to 5 days. Maximum eggs are laid generally on the second day of start of oviposi�on and about 90 per cent are deposited by the third day. A female lays 500 to 1900 eggs during its life span.
The newly hatched larvae feed on egg chorion and are able to survive without food for 13 to 15 hours. The late instars begin to spread in the field and feed solitarily on the foliage causing heavy defolia�on. Damage by larvae differs from the crop to crop. The larvae pass through 5 to 7 instars and become full-grown in 15-25 days. They stop feeding a few hours to a few (1-3) days before pupa�on but keep on moving quite fast probably in search of a suitable site for pupa�on. In years of outbreak, army of caterpillars moving from one to the other field is a common sight.
Most larvae pupate in the soil at varying depth. However, a few pupate on the soil surface mainly under plant debris. About 65 per cent larvae pupate deeper than 10 cm and some reach upto 28 cm or more deeper. A�er reaching the required depth, the larva sheds its hair and prepares a cocoon with the help of shed hair and silken threads and pupates inside. Pupal period is quite variable being 10-20 days in the mul�vol�ne race and 9 to over 10 months in the univol�ne race.
Moth emergence is o�en irregular and is governed by the rainfall pa�ern. In years of good and frequent/ intermi�ent rains, bulk (>80%) of the moths emerge within a short period of about 7 to 10 days. On the contrary, when rains are sca�ered, moth emergence is spread over several weeks. Under field condi�ons moths start emerging when, a�er the rains, the r.h. exceeds 70 per cent coupled with the lowering of mean
otemperature to around 30 C. From July to September, there are 2 to 3 genera�ons under laboratory condi�ons as well as in the field, though during most years the build-up of larval popula�on from second or third genera�on moths is not sufficient enough to cause any significant damage to the crops.
The moths have strong a�rac�on for ar�ficial light. About 50 per cent of the moth popula�on in a night is a�racted within the first two hours (8.00 to 10.00 p.m.) a�er dusk.
Natural enemies
Parasitoids: Trichogramma basiliense and Telenomus sp. near (Telenomus proditor Nixon) a�ack eggs; Apanteles obliquae; Apanteles bosei; Exorista xanthaspis; Exorista fallax; Plaxorista laxa (Palexorista laxa); Atherigona orientalis and Meteorus sp., a�ack larvae.
81

Nematodes: Mermis indica, a�acks larvae.
Viruses: NPV and Entomopoxvirus (Am EPV), a�ack larvae.
Fungi: Aspergillus fumigatus, a�acks larvae.
Most reports on natural enemies of A. moorei are simply records mainly from India. Preliminary field trials with Trichogramma brasiliense as egg parasitoid show some promise while larval parasitoids, mainly Exorista spp., have low parasi�sa�on in nature.
Management: Some cultural methods like deep ploughing a�er harvest, destruc�on of weeds growing on bunds, fence lines and on nearby uncul�vated land may help in reducing some pest damage but have limited scope given the deep pupa�ng habit of the diapausing larvae and availability of large tracts of uncul�vated land in the desert areas. Marching bands of caterpillars can be destroyed by digging trenches around the fields and burying them. Addi�onally, collec�on and destruc�on of egg masses and young larvae in the gregarious phase of the pest have been recommended. Light traps have been found useful in controlling A. moorei. Since majority of the moths a�racted to light are males, these are effec�ve par�ally only. Not much effort has been made on biological control of this pest. However, some bio-agents like Trichogramma and NPV seem to hold some promise.
Chemical control : The pest can be effec�vely controlled with the help of insec�cides. However, because of the spreading nature of the grown-up larvae from one to the other field, co-opera�ve approach in the form of �mely control campaigns would be more useful. Recent insec�cides reported to be effec�ve against this pest are quinalphos (0.05%), monocrotophos (0.05%), triazophos (0.06%) and chlorpyriphos (0.05%).
2. Amsacta albistriga Walker : It is a serious pest in southern India.
Distribu�on: Distributed throughout the plains of southern India, par�cularly in Karnataka, Andhra Pradesh and Tamil Nadu.
Host Range: Groundnut, pulses, cholam, cumber, castor, co�on, pearlmillet, sorghum, pigeonpea, etc. Feeds on almost all low growing plants and crops.
Biology: Adults emerge immediately a�er the south-west monsoon showers in May to July. Each female lays 600-700 eggs indiscriminately on any substrate, from which the larvae emerge in about four days. The larvae become full grown in 40-50 days, feeding on almost every green plant in their way. The larvae are blackish-brown covered with long blackish hairs. Pupa is squat, red brown and pupa�on is in soil at a depth of 10-20 cm.
Natural enemies
Telenomous manolus Nixon (Scelionidae : Hymenoptera) on eggs; Apanteles
colemani Vier and Apanteles creatono� Vier (Braconidae : Hymenoptera) on young
82

larvae; Sturmia incospicuella Bar. and Exorista civiloides Bar. (Tachinidae: Diptera) on
mature larvae; and Megaselia scalaris Liv. (Phoridae: Diptera) on pupae.
3. Amsacta lineola Fabricius
It has been reported as a serious pest of kharif crops around Udaipur
(Rajasthan).
Host plants: Maize, cowpea, lucerne, Bermuda grass, cauliflower, co�on, groundnut,
orange, le�uce, etc.
4. Amsacta flavicosta Hampson: Reported from Nigeria on cowpea.
The management of other spp. of Amsacta is similar to that of A. moorei.
5. Spilosoma (= Diacrisa) obliqua Walker (Arc�idae : Lepidoptera).
It is commonly called as Bihar hairy caterpillar or soybean hairy caterpillar or
jute hairy caterpillar.
Distribu�on : It is a sporadic polyphagous pest in the Oriental region. In India, it
damages the crops in Bihar, U.P., M.P., Punjab and Haryana.
Host plants : It has been recorded to feed on a large number of host plants, especially on
oilseed crops. Its host plants include sesame, groundnut, mustard, sunflower, castor,
linseed, co�on, mungbean, mothbean, black gram, lablab, cowpea, soybean, tomato,
cabbage, cauliflower, carrot, sweet potato, potato, cluster bean, pigeon pea, rice,
sannhemp, lucerne, Lantana, ragi, banana, Abu�lon indicum, etc. The young larvae
feed gregariously on chlorophyll of leaves. The order larvae disperse and skeletonize
leaves and feed on stems and growing points also.
Biology and ecology : The pest breeds from March to April and from July to November.
It passes the ho�est part of summer (May-June), and winter (December-February) in
the pupal stage amidst plant debris. Moths emerge in March. A female may lay 400-
1200 light green, spherical eggs in clusters on the underside of leaves. The eggs hatch in
8-13 days and the �ny caterpillars during the first two stages feed gregariously but
a�erwards disperse. There are 7 larval instars over a period of 4-8 weeks. The ground
colour of the larva in black and yellow and it is profusely covered with long black or
black and white hair. Full grown larva measures 40-45 mm. When full grown, the larva
spins a loose silken cocoon in which pupa�on takes place in plant debris or soil. Pupal
stage lasts for 1-2 weeks. The pest passes through 3 or 4 broods. Build-up of popula�on
of this pest on sunflower at Delhi was ascribed to unseasonal rains during February,
followed by gradual rise in temperature, fairly high humidity and longer sunshine
dura�on, and peak larval popula�on (i.e. upto 1350 caterpillars/plant) was recorded in
the third week of March (Sethi et. al., 1979).
83

Egg parasitoid : Teleomous molorchus (causing 50 to 83.3% parasi�za�on at
Hyderabad), Apanteles ter, A. vitripennis;
Larval parasitoids : Apanteles ter and A. vitripennis.
Insec�cides : The insec�cides reported to be effec�ve against this pest are
monocrotophos (0.02%), quinalphos (0.05-0.075%), endosulfan (0.05-0.15%),
methomyl (0.03%), dichlorvos (0.05%), chlorpyriphos (0.05%) and carbaryl (0.1%).
Diflubenzuron has also been reported to have adverse effect on its development.
Prophylac�c sprays against the migra�ng caterpillars did not prove fully effec�ve.
Azadirach�n showed some an�feedant ac�vity against the younger larvae but had li�le
insec�cidal effect when applied topically on the larvae.
Integrated pest management
i) Use of light trap to monitor the pests ac�vity.
ii) Collec�on and destruc�on of egg masses and hand picking and destroying the leaves containing young larvae in the gregarious phase.
iii) Use of effec�ve insec�cides as men�oned above.
Suggested Reading:
Atwal, A.S. and Dhaliwal, G.S. 1997. Agricultural Pests of South Asia and their
Management. Kalyani Publishers, New Delhi-Ludhiana. 487 pp.
Chaudhary, J.P.; Sidhu, A.S. and Ramzan, M. 1971. Studies on the biology of the red hairy
caterpillar, Amsacta moorei Butler (Arc�idae: Lepidoptera). Haryana
Agricultural University. J. Res. 1(4): 24-28.
Kulshrestha, S.P.; Diwakar, M.C. 1987. Some observa�ons on the incidence of red hairy
caterpillar during kharif 1986 in Gujarat. Plant Prot. Bulle�n, India, 39(3): 36-37.
Patel, C.C.; Patel, N.C.; Patel, J.R.; Patel, A.M.; Broad, P.K. and Jyani, D.B. 1995. Natural
control of Gujarat hairy caterpillar, Amsacta moorei Butler with bio and
microbial agents in middle-Gujarat. Gujarat agric. Univ. Res. J. 21(1): 54-58.
Ramaswamy, K.A. and Kuppuswamy, S. 1973. Some observa�ons on the biology of the red
hairy caterpillar (Amsacta spp.) on groundnut. Madras Agril. J. 60(7): 637-639.
Saini, R.K. 1993. Development and survival of red hairy caterpillar, Amsacta moorei Butler
on some cul�vated plants and weeds in Haryana. J. Insect Sci. 6(1): 64-68.
Saini, R.K.; Pala Ram and Chaudhary, S.D. 1997. Some behavioural aspects of Amsacta
moorei Butler with par�cular reference to pupa�on and light a�rac�on. J.
Insect Sci. 10(1): 12-15.
84

Saini, R.K. and Ram, P. 2000. Emergence of Amsacta moorei moths as influenced by ambient weather parameters. Indian J. Agril.Sci. 70(11): 765-768.
Joshi, R.C., Rao, P.K. and Rao, B.H.K. 1983. Occurrence of Telenomus (Acholeus) molorchus Nixon as an egg parasite of Bihar hairy caterpillar. Indian J. Ent. 45(4): 499.
Kalra, V.K. 1984. Record of two new species of Apanteles parasi�zing Spilosoma obliqua (Wkr.) Indian J. Pl. Prot. 12(2) : 155.
Sinha, P.K., Singh, R.P., Yadav, R.P. and Kumar, K. 1984. Rela�ve efficacy of some insec�cides for the control of Bihar hairy caterpillar, Diacrisia obliqua Walker infes�ng sweet potato. Indian J. Ent. 46(3): 331-335.
85

MANAGEMENT OF WHITEGRUBS IN KHARIF LEGUME CROPS
SWAROOP SINGH* AND VIJAY KUMAR AGRAWAL***Network Coordinator, **Professor
Division of Entomology,Swami Keshwanand Rajasthan Agricultural University
Rajasthan Agricultural Research Ins�tute,Durgapura, Jaipur
India is reportedly the largest pulse growing country in the world both in terms of area as well as produc�on covering 43.30 per cent of land area under pulses with 33.15 per cent produc�on. In another report, it has been described that India is the largest producer and consumer of pulses in the world accoun�ng for 33 per cent of world's area and 22 per cent of world's produc�on of pulses. Green gram is one of the most widely cul�vated pulse crops a�er chickpea and pigeonpea. The major producing states in India are Andhra Pradesh, Orissa, Maharashtra, Madhya Pradesh and Rajasthan accoun�ng for about 70 per cent of total produc�on.
An es�mated 200 insect pests that belong to 48 families in Coleoptera, Diptera, Hemiptera, Hymenoptera, Isoptera, Lepidoptera, Orthoptera, Thysanoptera, and 7 mites of the order Acarina are known to infest green gram and black gram. Scrab beetles (Coleoptera: Scarabaeids) commonly called as white grub are one of the major limi�ng factor in agricultural crop produc�on (Sharma, 2002). The damage caused by the grubs under rainfed condi�on, which account for about 80% of cul�vated land, may vary on an average from 10%-30% but some�mes complete crop failure is also observed. Due to intensive cul�va�on and destruc�on of forests, white grubs have a�ained the status of pest of na�onal importance during the last three decades. The grubs, feed on the roots of many agricultural crops, weed hosts, forest trees, ornamental and hor�cultural crops where as the adults feed on foliage, flower, pollen and fruits of the crops. There is a complex of white grub species prevalent in the agricultural areas of different parts of the country. Existence of as many as about 80 species belonging to sub families Rutelinae, Melolonthinae, Dynas�nae, Cetoniinae, Aphodiinae, Euchirinae and Scarabaeinae has been documented .The C-shaped larvae, grubs are pale yellow or white. Adult beetles are nocturnal, although the flower chafers (Cetoniinae) and many leaf chafers (Rutelinae) are ac�ve during the day. The grubs mostly live underground or under debris, so are not exposed to sunlight. Whitegrubs are root feeders and their beetle feeds on the leaves of host plants. The major species of whitegrubs namely Holotrichia consanguinea, H. serrata Fab., H .fissa Br., Leucopholis lepidophora Bl. (Melolonthidae), Anomola sp. (Rutelinae) are cosmopolitan in distribu�on. The fauna of Indian sub region is very rich and diverse, but it is yet to be fully explored. Availability
86

of abundant roots and adequate moisture for a longer �me in sugarcane, groundnut and pea crop tend to increase the white grub build up markedly.
Descrip�on of Holotrichia consanguinea:
Holotrichia consanguinea is the most serious scarab pest in India. It finds loose, sandy well drained soil to be quite suitable for its survival and mul�plica�on. It is a dominant whitegrub species in the states of Rajasthan, Gujarat, Haryana, Punjab, U.P. and Bihar. It feeds on all kharif crops but the damage is more evident on plants having tap roots and so in many parts of the country, it is the main constraint in groundnut cul�va�on. The beetles of Holotrichia consanguinea emerge from the soil during dusk a�er good pre monsoon or monsoon rain in mid May or later. Any rain received prior to mid-May does not ini�ate emergence of beetles as they are not sexually mature. If there is good pre-monsoon rain, two peaks of beetle emergence are observed, Once ac�ve, the beetles follow the daily rhythm of emergence, congrega�on on host trees at dusk, and return to soil at dawn. The beetles are polyphagous, and may feed on the foliage of a wide variety of host trees and bushes found in the close vicinity. However, they do exercise some preference for certain hosts like jujube (ber), Prosopis cineraria (khejri) neem, cluster fig (gular), jambolana (Jamun) and drums�ck (sainjana). In some situa�ons, large number of beetles se�les on a non-host tree, only for ma�ng and just a�er, shi� for feeding to nearby preferred hosts. The host preference depends on the combina�on of hosts available in a par�cular locality. The beetles usually a�er emergence, do not fly long distances in the evening but try to avail the short twilight hour for ma�ng and selec�ng a suitable host as they are unable to see clearly in dark. It is the female which first comes out of soil, se�les on a host and secretes a pheromone which a�racts the male for ma�ng. If a female fails to mate in first 10 to 15 minutes of se�ng on the host tree, she tries ma�ng next evening. The process of congrega�on of beetles on host con�nues every evening for a period of about three weeks to ensure ma�ng of all females. The mated females return to soil in early morning and start egg laying within 2 to 3 days of ma�ng. However, some females, depending upon the condi�on of maturity of their ovarioles, may lay eggs upto 10 days or more a�er ma�ng. The female prefer light sandy soil for egg-laying. The longevity of the emerged beetles varies from 35 to 49 days, during the period daily host visits are made for feeding. On an average 20 eggs are laid by mated females in instalments, star�ng from third day a�er ma�ng. The eggs are laid at the depth of 6 to 10 cm inside the soil. Freshly laid eggs are white, ellip�c, about 3.22 mm in length and 1.95mm in width. Prior to hatching the eggs become brownish black and globular, and measure about 3.27 mm in length and 2.73 mm in width. The incuba�on period under laboratory condi�ons ranges from 7 to 13 days.
Life cycle of whitegrub:
The newly hatched grubs are creamy white in colour and may feed on organic ma�er for some �me �ll they come in contact with living roots. On an average, the first
87

instar grub measures 15.1 mm in length; while, the head length and width are 4 mm and 3 mm, respec�vely. The average dura�on of first instar grub is 16 days. The second instar grub measures 34.5 mm in length; the head length and width being 6 mm and 5 mm, respec�vely. The average dura�on of second instar grub is 32 days. The third instar grub measures 40.7 mm in length; the head length and width being 8 mm and 7 mm, respec�vely. The average dura�on of third instar is 49.5 days. Total grub dura�on ranges from 82 to 113 days. For feeding, the grub makes chamber by compressing the surrounding soil par�cles and then eats the rootlets exposed into the chamber; therea�er it moves ver�cally, a li�le, to eat more of the same root. A�er this, the grubs move horizontally making chambers and feeding on the exposed toots. The grubs con�nue ac�ve feeding from July to mid - October. All the grubs pupate by the beginning of November. Before pupa�on the grubs enter deep into the soil to the depth of 40 to 70 cm or more in search of suitable moisture Zone. The average pupal length and width is 27.3 mm and 14.2 mm, respec�vely. A short prepupal period is spent before entering into pupa�on, during which, the grub prepares and earthen chamber for pupa�on. The average pupal period is 14.2 days. The freshly emerged beetle is white in colour but with the lapse of �me it becomes dull brown with light brown abdomen and dark brown legs. The average beetle length and width is 21.3 mm and 11.8 mm, respec�vely. The beetles remain in the soil in inac�ve state upto middle of May at a depth of about one meter. The average dura�on of one life cycle is 122 days and there is only one genera�on in a year.
Damage in legume crops at different centres:
(i) Damage in moongbean at Durgapura (Jaipur): Moongbean has tape root system; it is highly damaged by whitegrub. The second instar grubs of Holotrichia consanguinea species cut the root of moongbean. The damaged plant show varying degree of yellowing than wilt and ul�mately die. Such plant can be easily pulled out. The damage ranges from 20-80 per cent in the presence of one grub/m2. In endemic areas incidence of whitegrub is 80-100 per cent in moongbean.
(i) Pea in Himachal Pardesh: pea is a leading off season vegetable of the state. There are certain ecological niches providing environmental condi�ons congenial for growing peas during Kharif in higher hills. In Sangla valley of Kinnaur, whitegrubs cause 20-25 per cent plant mortality in offseason crop in the month of June-July. The major species which were collected from different locali�es in the valley were Holotrichia longipennis, Brahmina coriacea, Melolontha furcicauda and Anomala sp. The damage was most serious in fields located in the vicinity of apple orchards. There was patchy growth in the infested fields and the damaged plants showed varying degree of yellowing, browning and wil�ng (Fig. 12 ). The popula�on of whitegrubs was very high and 4-5 grubs were found feeding on a single plant. The roots were totally pruned and the infested plants can be pulled out very easily. The pea crop fetches premium price
88

during off season, hence white grub damage incurs heavy losses to farmers in that area.
1.� Treatment of pea seeds with quinalphos 25 EC or chlorpyriphos 20 EC @ 4 ml/kg seed is recommended for the control of H. longipennis.
2.� The required quan�ty of the insec�cides should be thoroughly mixed with the seed in a polythene bag by adding small quan�ty of water (20-30 ml) and the seed should be shade dried before sowing.
(ii) Damage in pulses at Jorhat (Assam) : Jorhat centre has reported following species on different pulses
White grub species� � Crop� � � Infesta�on
(i) Apogonia sp. � Green gram� � 28-32% � Black gram� � 25-30% � French bean� � 10-15%
(ii) A. aerea Blanchard � Green gram� � 27-33% � Black gram� � 25-30% � French bean� � 13-15%
(iii) Adoretus sp. � Green gram� � 22-26% � Black gram� � 15-20%
(iv) A. aerial Arrow � Green gram� � 25-30%
Management
Although whitegrubs can be a problem every year, the most serious damage occurs in regular three year cycles. The greatest damage to crops occurs the year a�er the appearance of the adults. During the years of heavy May beetle infesta�on, deep-rooted legumes, such as alfalfa or clovers, should be planted. If corn or small grains are present, every effort should be made to keep the field free of grass and weed growth, as this will reduce the number of eggs laid. The year following heavy flights of May beetles, plan�ng corn or potatoes should be avoided in fields that were previously under sod or grass. Late spring or early autumn plowing destroys many larvae, pupae, and adults in the soil and also exposes the insects to predators, such as birds and skunks. For this cultural prac�ce to be effec�ve, ploughing must occur before the grubs migrate below the plough depth. No-�llage or reduced �llage crop management enhances grub popula�ons. Natural enemies that control these white grubs include parasi�c wasps and flies in the genera Tiphia and Myzinum (Hymenoptera: Tiphiidae), and Pelecinus polyturator Drury (Hymenoptera: Pelecinidae), and the fly, Pyrgota undata (Diptera: Pyrgo�dae).
Control measures
89

As with other soil dwelling pests, control with insec�cides is difficult. Best results occur when treatments are applied to young grubs, generally in August. Larger grubs are more difficult to control and may require more than one treatment. Management of soil moisture will improve results. Crop rota�on, however, is the most effec�ve cultural control method. Deep-rooted legumes, like alfalfa, is excellent crop with which to rotate corn or small grains, especially following years of unusually heavy May beetle flights.
Management through beetle control
1. The preferred hosts of beetles are sprayed with imidacloprid 17.8 SL at 1.5 ml/L water during day �me with the onset of monsoon. No �me gap is given between the onset of rain and spraying and efforts be made to spray all the preferred hosts within 3-4 days of the first emergence of the beetles.
2. As an alterna�ve to spraying mechanical collec�on of beetles be done during the night by jerking the host trees, collec�ng the fallen beetles and killing them in kerosenized water. Where host trees are not available nearly, cut branches of host trees like neem may be planted in the evening to a�ract the beetles for collec�on.
3. Light trap be set up in endemic areas to collect the beetles with the onset of monsoon in June July coinciding with emergence of beetle and the trapped beetles be destroyed.
4. Pheromone traps can also be very effec�ve management tool to trap the adults if the pheromone of that species is available. The adults of Holotrichia consanguinea emerge from soil with first good shower (premonsoon or monsoon) and congregate on preferred hosts. These beetles can be killed conveniently by spraying above insec�cide during day �me just a�er the first shower of the season. In any case the spraying opera�on should be completed within three days of the showers with the objec�ve to kill the adults before they are able to lay eggs in the soil. This technique is very effec�ve, but at some places, where the popula�on of host trees per unit area is very high, spraying of all the host trees within a very short period is a problem, consuming more �me, labour and insec�cide and also pollutes the environment. To over come these problems, Durgapura centre of All India Network Project on White grubs and Other Soil Arthropods has generated whitegrub management technology using pheromone. The pheromone (methoxy benzene), isolated from the abdominal glands of calling female adults of H. consanguinea have a unique property of ac�ng as aggrega�ng pheromone, a�rac�ng both the sexes from a distance of 15 meters. Use of this pheromone has made beetle control opera�on easier, economical and less hazardous as only one tree within a radius of 15 meters is required to be sprayed with insec�cide and loaded with "Pheromone Dispensers". For the purpose, a single tree from a group of host trees within a radius
90

of 15 meters could be selected and sprayed with imidacloprid 17.8 SL (0.02%) during day �me. "Pheromone Dispensers" be placed (3-4 dispensers/tree) on this host tree in the evening, con�nuously for three evenings at the �me of beetle emergence. No �me gap should be given between onset of rain, spraying and pheromone loading. This should be done for 3-4 days a�er first emergence of beetles. The beetles which are a�racted by pheromone, se�le and feed on the sprayed host trees and get killed soon by contact as well as stomach ac�on of the insec�cide and thus, not able to return to soil next morning for egg laying. By pheromone loading, number of trees to be sprayed is dras�cally reduced, which make the beetle control opera�on more effec�ve, economical and less �me consuming.
How a farmer can prepare the "Pheromone Dispenser" himself:
Materials: 1. Wire: 30-40 cm long , 2. Sponge: 10 cm2 , 3. Pheromone: 3 ml , 4. Stone: About 50 g
Methods: Tie the sponge piece with one end and stone with another end of the wire. Pour the pheromone on the sponge drop by drop and throw it on the tree (3-4/tree/night).
Management through grub control
· Whenever beetle control is not possible and crops like maize is to be protected pre-sowing soil applica�on of phorate 10G of quinalophos 5 G both at the rate of 25kg/ha .
· In standing crops Imidacloprid 17.8 SL @ 300 ml/ha be applied with irriga�on.
· In March –April deep ploughing of fallow lands should be undertaken with a view to expose the immature state i.e. grubs and pupae of this pest for preda�on by the avian predators' viz. crow, mynahs. Besides grubs and pupae exposed during the ploughing should be collected and destroyed.
· Farmers should apply only well decomposed farmyard manure in fields. The partly decomposed FYM provides congenial condi�ons to survive the newly hatched grubs.
· Sowing of tolerant crops viz. amaranthus, buckwheat, maize, and horse gram in the severely infested areas is recommended which are less damaged by the grubs when compared with millets, vegetables etc.
· Deep ploughing of fallow lands in august- September to expose the third instar grubs for predatory birds is quite effec�ve in reducing the popula�on.
Cultural prac�ces to control grubs:
1. In endemic areas summer fodder crops provide suitable place for survival of large
91

number of grubs. These fields are ploughed from end of July to middle of august, a�er taking the last cu�ng to expose the grubs to predatory birds. If predatory birds are absent a man with a bucket in hand may follow the plough and pick up the exposed grubs. Two cross ploughing may expose about 70% of grubs present in the field.
2. Early (premonsoon, irriga�on) sowing of crops should be done to allow roots to get established so that a certain level of grub popula�on may be tolerated by the crops without any no�ceable damage. This will be useful in areas where grub popula�on is quite low and should be adopted as an adhoc strategy. If this is prac�ced over the years it will increase the popula�on of the pest.
3. Intercropping of pearl millet with pulses significantly reduce the white grub damage on pearl millet crops.
Biological control measure of whitegrub:
There are no any known effec�ve biological agents which can bring down the white grub popula�on to a non pest level in a short �me. However, the impact of biological agents was found effec�ve when congenital condi�ons prevail. Large number of predators, Parasitoids and entomogemous microbes have been recorded.
1. An insect predator, the common crock beetle was observed to prey upon the beetles of H. longipennis during night hours. The emergence of predatory beetles coincides with the emergence of scarab beetles in June – July. In lab. Condi�ons, the preda�on efficiency of predator was observed to be 1-6 beetles/24hr.
2. The spo�ed owlet and grey jungle fowl were observed to prey upon the beetles se�led on various host plants during evening hours. Bio-ra�onal control: The bio-ra�onal control measures are recently developed and adopted to control the white grubs. Bio-ra�onal controls used in combina�on may impart a synergis�c mortality on grub popula�ons. This may occur if the two agents a�ack different sub-popula�ons (i.e., grubs feeding at different depths) or if one agent (i.e. sublethal dose of merit) predisposes an insect to another agent.
Summary:
Whitegrubs are the pests of na�onal Importance and are a serious constraint to almost all the kharif crops grown under rainfed condi�ons. Both the grubs and adults are polyphagus and univol�ne in nature. No single control measure is effec�ve for their management hence is the only op�on for their effec�ve management in the hilly zone.
Life Cycle of Whitegrub (one genera�on in a year)
92

Farmers' preparing sponge
Spraying of Selected WhitegrubHost Tree
Large Number of Beetles Attractedon Pheromone Loaded Trees
Preparation of sponge for loading on host trees
93

PROSPECTS OF EXPLOITATION OF NATURAL ENEMIES ASSOCIATED WITH PESTS OF LEGUME CROPS
PALA RAMDepartment of Entomology,
CCS Haryana Agricultural University,Hisar-125 004 (Haryana)
The yield of legume crops especially the pulses has not increased considerably over the several years. Among various factors insect pests are also responsible for reduc�on in yields to levels at which harves�ng is not economic. Key pests have also developed resistance to insec�cides and have become uncontrollable. So there is a need to think about long term solu�ons by emphasising on some important components of pest management especially biological control. The present review discusses about the role of natural enemies in the management of important pests in major legume crops like chickpea, groundnut, pigeonpea and soybean.
Chickpea:The egg parasitoids, Trichogramma spp. and Telenomus spp. destroy large numbers of eggs of H. armigera and H. punc�gera, but their ac�vity levels are too low in chickpea and pigeonpea because of trichome exudates. The acidic exudates of the plant have been reported to be responsible for the deterrence and failure of egg parasitoids. A number of larval parasitoids have also been reported to a�ack various stages of H. armigera larvae. Hymenopterans parasitoids generally a�ack early instar larvae and the dipterans choose late instars. Of the large number of larval parasitoids recorded on H. armigera in chickpea ichneumonid, Campole�s chlorideae has been found to be most poten�al and effec�ve parasitoid causing 20-80% parasiza�on. Srinivas (1989) recorded 43.9% parasi�sm of neonates during the first fortnight of December but only 12% during last week of January, however, Eriborus sp., another ichneumonid, a poten�al larval parasitoid of pod borer on chickpea resulted in 9% parasi�sm during early December and 43.8% during late January. Parasi�sa�on by C. chlorideae was high on crop sown in October than in November (Teggalli, 1995). In India, Carcelia illota (Curran), and to a lesser extent, Goniophthalmus halli Mesnil, and Palexorista laxa (Curran) parasi�ze up to 54% of H. armigera larvae on chickpea (A.B.S. King, unpublished). There are a few reliable es�mates of pre-pupal and pupal mortality of H. armigera, which may be as high as 80% (King, 1994). Six species of parasitoids have been recorded from field-collected Helicoverpa pupae (Fi�, 1989). As far as bruchids, Callosobruchus species in stored grain legumes are concerned the specialized egg parasitoid, Uscana lariophaga Steffan, U. mukerjii (Mani) offer some perspec�ves due to their strong impact on Callosobruchus maculatus popula�on in stored cowpea. The larval-pupal parasitoid, Dinarmus vagabundus Timberlake was found to have good poten�al for controlling stored grain pest, Callosobruchus maculatus F. infes�ng various pulses in storage (Dongret et al., 2002).
94

There is voluminous informa�on on parasi�sm, and to a lesser extent on preda�on of insect pests of different food legumes. The most common predators of insect pests of food legumes are Chrysopa spp., Chrysoperla spp., Nabis spp., Geocoris spp., Orius spp., Polistes spp., and species belonging to Pentatomidae, Reduviidae, Coccinellidae, Carabidae, Formicidae and Araneida (Zalucki et al., 1986; van den Berg et al., 1988; Romeis and Shanower, 1996; Sharma, 2001). Among predators, Chrysoperla zastrowi arabica (=Chrysoperla carnea) when released @1,00,000 second instar grubs per ha was found quite effec�ve (Rabindra et al., 1994) but the economic considera�ons in its mass produc�on and release disproved its poten�al and efficiency as a biocontrol agent. Among other different predatory insects of H. armigera recorded in chickpea crop adults of Calosoma indicum Hope and Polistes herbraens Fab. were found promising (Singh et al., 1990). However, there are no reports of large scale use or these predators in chickpea ecosystem. Among birds, common mynah, Acridotheres tris�s (L.), house sparrow, Passer domes�cus and parakeet, Psi�acula krameri (Scopli) are the common predatory birds in chickpea crop (Chhabra, 1992). Exploita�on of biocontrol agents in the integrated management of legume crop pests is confined to use of microbial pathogens. Odak et al. (1982) evaluated commercial formula�ons of Bacillus thuringiensis (Bt) against H. armigera and reported that Thuricide and Bactospine were highly pathogenic both in the laboratory and pot experiments causing 70-100% and 20-65% larval mortality, respec�vely. Nuclear polyhedrosis virus has emerged as the most poten�al component in the suppression of H. armigera. Effec�veness of HaNPV at varying dosages (125-500LE/ha) has been reported and addi�on of adjuvants and UV protectants in solu�on further increased the effec�veness of HaNPV. Use of this bioagent in India has been on increase. However, its availability in �me and space and cost seems to be the major constraints at grower's level. Establishment of mass mul�plica�on units from the field collected host larvae in a co-opera�ve approach should resolve the constraints. Nematodes also parasi�ze H. armigera larvae, especially during the rainy season. These nematodes grow to 10 cm or more. They coil up inside H. armigera larvae, and kill them before pupa�on by consuming and disrup�ng the internal organs of the host. Steinernema glaseri @4000 infec�ve juveniles (IJs)/20ml dis�lled water caused highest mortality (24.6%) of pod borer on chickpea a�er 6 days of spraying followed by 3000 IJs/20ml dis�lled water (11.9%) (Patel and Vyas, 1995). Fungus, Beauveria bassiana (Balsamo) Vuillemin has been reported pathogenic to H. armigera. Field inves�ga�ons indicated that the fungus controls H. armigera effec�vely resul�ng in lower pod damage and higher yield. Pod damage of 6.8 and 8.7% and grain yield of 2.38 and 2.19q/ha was recorded in chickpea
7 7plots treated with spore concentra�on of 2.82x10 and 2.82x10 spores/ml, respec�vely, in contrast to that in untreated control which registered 16.3% pod damage and 1.84q/ha grain yield (Saxena and Ahmad, 1997).
95

Groundnut:Egg parasitoid, Trichogramma chilonis Ishii (=Trichogramma evanescens minutum Riley) had been found to parasi�se the eggs of Amsacta albistriga Walker and Diacrisia obliqua Walker (Krishnamurthi and Usman, 1954). For the control of Spodoptera litura, T. chilonis appears to offer effec�ve control under inunda�ve releases (Khan et al., 2013). Telenomus manolus Nixon parasi�sed A. albistriga eggs in Tamil Nadu with 94.3% parasi�sm (Sundaramurthy et al., 1976 and Gunathilagaraj and Babu, 1987). In Tamil Nadu, Trichogramma spp. was found parasi�zing 10.3% of A. modicella eggs in groundnut (Muthaiah and Kareem, 2000). Hymenopterous parasitoids a�acking leaf miner larvae were the most important group of natural enemies. Nine primary and eight secondary parasitoids emerged from host larvae, and killed up to 50% of the leaf miner larvae sampled (Shanower et al., 1992). Braconid, Avga nixoni and eulophid Stenomesioides ashmeadi Subba Rao and Sharma were recorded for the first �me on Stomopteryx subsecivella (Subba Rao and Sharma, 1966). The eulophid, Stenomesius japonicus Ashmead and the chalcid, Brachymeria wi�ei Schmitz were found parasi�zing for the first �me on A. modicella in Tamil Nadu (Muthaiah, 1991). Braconid, Asympiesiella indica, Bracon spp., Chelonus spp., Apanteles spp. and the bethylid Goniozus spp. were also found to parasi�se the leaf miner in Tamil Nadu. The parasi�sa�on by B. gelechiae on A. modicella ranged from 5.2-33.3% at Bangalore (Rajagopal et al., 1988). The parasitoid Goniozus spp., Brachymeria spp., S. japonicus, B. gelechiae, Chelonus curvimaculatus Cam. and Apanteles spp. parasi�sed S. subsecivella during 3rd week of August and per cent parasi�sa�on reached its peak in mid September (Khan and Raodeo, 1978). B. gelechiae and Brachymeria spp. parasi�sed 7.7-24.0% larvae (Bakhe�a, 1974). Among ten larval parasitoids recorded on A. modicella in Tamil Nadu, Goniozus indicus Ashmead and B. wi�ei parasi�zed 16.7 and 20.0% larvae, respec�vely (Muthaiah and Kareem, 2000). Carcelia illota Curran was found parasi�zing the pupae of H. armigera in groundnut in Gujarat (Koshiya and Patel, 1987). Pupal parasitoid Chelonus spp. was found parasi�zing 26.0% of A. modicella pupae in Tamil Nadu (Muthaiah and Kareem, 2000). Several predators were also reported to be associated with pest complex of groundnut. In Gujarat Aphis craccivora Koch popula�on was suppressed with in a week by predatory coccinellids, Menochilus sexmaculata F. and C. septempunctata L. (Patel et al., 1976). Brumoides suturalis F., M. sexmaculata and C. septempunctata were found feeding on groundnut aphid at Ludhiana (Bakhe�a and Sidhu, 1977). On an average M. sexrnaculata consumed 474 aphids during 6 days of nymphal development and 16-49 days of adult life (Patel and Vyas, 1984). Syrphids, Ischiodon scutellaris F., Xanthogramma scutellaris, and chrysopid, Chrysoperla carnea Stephens were also found to predate upon A. craccivora (Patel et al., 1976, and Patel and Vyas, 1984). Coccinellids, B. suturalis, C. septempunctata, M. sexmaculata and Hippodamia variegata Goeze were also found to feed on jassid nymphs and adults (Singh et al., 1991). C. septempunctata, Orius sp. and M. sexmaculata were found to control thrips
96

and the predatory mite, Balaus�um sp. was recorded as a predator of lea�oppers and thrips (Sherasiya and Butani, 1999). Rhinicoris marginatus F. reduced 92.73 and 94.91% popula�on of H. armigera and S. litura, respec�vely, under field condi�ons (Sahayaraj, 1999).
Wide array of microbial agents have been found suppressing various pests in the groundnut ecosystem, which include bacteria, fungi, nematodes, viruses and others. Bacillus thuringiensis was effec�ve in bringing down popula�on of A. modicella to the tune of 5.6-36.4% at Bangalore (Rajagopal et al., 1988). Spore suspensions of Bacillus popillae var holotrichiae obtained from diseased grubs of H. consanguinea was highly virulent and controlled grubs up to 72.2% under field condi�ons when applied at 5
2 billion spores/m (Vyas et al., 1991). Metarrhizium anisopliae Metchnikoff was also found infec�ng H. consanguinea (Kalra and Kulshrestha, 1961), S. litura (Siddaramaiah et al., 1986) and A. modicella (Rajagopal et al., 1988). Infec�on of Beauveria bassiana (Balsamo) Vuillemin on A. modicella ranged from 11.6-30.8% at Bangalore (Rajagopal et al., 1988). Beauveria brongniar�i Saccardo was reported to infect the grubs of H. consanguinea (Vyas et al., 1990). D. obliqua was found infected with Entomophthora grylli Fres. (Thontadarya et al., 1973), while the fungus, Aspergillus spp. was found infec�ng Amsacta moorei Butler and Ver�cillium lecanii Zimmerman infec�ng H. consanguinea (Gour and Dabi, 1988). Nomuraea rileyi (Farlow) Samson brought 36.9% mortality of S. litura larvae in Andhra Pradesh (Sridha and Prasad, 1996). Nuclear polyhedrosis virus (NPV) was recorded infec�ng A. moorei, A. albistriga, H. armigera, S. litura and S. obliqua. Grubs of H. consanguinea harboured entomopox virus (EPV) along with Bacillus popillae (Narayanan and Tandi, 2001). A protozoan, Nosema sp. was found infec�ng 13.8 per cent larvae on S. litura in Andhra Pradesh (Sridhar and Prasad, 1996). Among four nematodes Steinernema carpocapsae Weiser (strain DD 136), Heterorrhabdi�s bacteriophora (strain Burliar), Heterorhabdi�s sp. (strain Chekkanuari) and Heterorrhabdi�s sp. (strain Melur) tested against fourth instar larvae of red hairy caterpillar, A. albistriga, S. carpocapsae was the most effec�ve in killing the larvae in Tamil Nadu. Steinernema glasseri Steiner and S. carpocapsae were found infec�ng the grubs of H. consanguinea (Shan� and Sivakumar, 1991). Shanower et al. (1992) reported that pathogens killed up to 30% of the leaf miner larvae. The combined effects of all mortality agents killed up to 95% of the leaf miner larvae per sample period.
Pigeonpea: Extensive studies on host plant a�ributes have shown that in both pigeon pea and chickpea, trichogramma�d egg parasitoids are not likely to perform sa�sfactorily for control of H. armigera. The egg parasitoids occurring in or used for biocontrol in legume crop ecosystem in India are mainly Scelionids (Telenomus sp.) and Trichogramma�ds (Trichogramma, 14 species; Trichogrammatoidea, 7 species), besides the braconoid egg-larval parasitoid (Chelonus, 6 species) (Khan et al., 2013).
97

Lingappa et al. (2001) published a comprehensive list of 83 parasitoids of H. armigera in India. Parasi�sm of H. armigera eggs by Trichogramma chilonis on pigeonpea was reported very low ranging from 0.1- 5.2% (Bhatnagar et al., 1982; Ram et al., 1998). Parasitoids belonging to genus Trichogramma do not prefer to parasi�se Helicoverpa eggs laid on chickpea and pigeonpea. When T. chilonis was released at flowering stage in pod borer infested pigeonpea fields, it failed to parasi�se the eggs of pod borers. Romeis et al. (1997) based on their field experiments concluded that T. chilonis releases were ineffec�ve on pigeonpea. Bisane et al. (2008) found that ichneumonids, Eriborus argenteopilosus, Cample�s chlorideae and braconid, Bracon sp. resulted in about two-third of the total mortality of early instar H. armigera larvae on pigeonpea. They also found that tachinid parasitoids which were found to be associated with the late larval instar and pupae resulted in their 17.8% and 8.3-26.3% popula�on reduc�on, respec�vely. In India, Carcelia illota (Curran), and to a lesser extent, Goniophthalmus halli Mesnil, and Palexorista laxa (Curran) parasi�ze up to 22% of H. armigera larvae on pigeonpea (Bhatnagar et al., 1983). However, dipterous parasitoids appear to be of lesser importance as these kill H. armigera in the prepupal or pupal stages a�er substan�al damage has been caused by the caterpillars to pods (Bhatnagar and Davies, 1980). In general, hymenopteran a�acked small larvae and the dipteran preferred large ones. An exo�c dipterous parasitoid, Eucelatoria brayani introduced from USA, was successfully reared on the pod borer in the laboratory and field establishment has been reported (Pawar et al., 1981). Pod fly, an important pod feeder on pigeonpea was parasi�sed by four parasitoids, namely, Euderus sp., Ormyrus sp., Eurytoma and An�strophoplex sp. Their cumula�ve extent of parasi�za�on varied from 4.1-49.8% and these parasites had a significant nega�ve correla�on with the host larval popula�on (Das and Katyar, 1998). Gryon clavigrallae Mineo, an egg parasitoid of Clavigralla scutellaris Spinola and C. gibbosa (Westwood) parasi�sed 58 and 40% eggs, respec�vely (Romeis et al., 2000). Clavigralla scutellaris laid significantly larger egg clusters than C. gibbosa (19.9 versus 10.5 eggs per cluster). A total of 29 species of insects and spiders have been recorded to predate on egg and larvae of H. armigera. Chrysoperla carnea Stephens a general predator, though reported in pigeonpea ecosystem (Srinivas and Jayaraj, 1989), has been found ineffec�ve and inunda�ve releases were unlikely to be economic (King el al., 1982; Hegde and Lingappa, 1998). Laboratory studies at ICRISAT revealed that a spider, Clubiona sp. can consume 59.3 eggs or 3.2 neonate larvae of the pod borer. These spiders were also reported to trap the adult pods files (Reed and Lateef, 1990). Among birds, black drongo, Dicrurus adsimilus (Bechstein), house sparrow (Passer domes�cus L.) blue jays (Coracias bengalensis L.), ca�le egret (Bulbulcus ibis (Boddaot), rosy pastor (Sturmus roseus L.) and mynah (Acridotheres ginginianus Lathrim) have been recorded feeding on Helicoverpa armigera and other lepidopterous insects on chickpea and pigeonpea crops (Yadava, 1991; Lingappa and Gopali, 1994). It was also found that the ac�vity of
98

predatory birds was found to be high when some grain crops like sorghum, are grown as intercrop in pigeonpea (Venugopal Rao, 1993; Kashyap, 1998). Wooden log or branch bifurcated at the top provided with an earthen container filled with water mixed with cooked rice a�racts the birds (Kashyap, 1998).
Microbial pathogens of H. armigera namely, nuclear polyhedrosis virus (virus), Bacillus thuringiensis (bacterium), Beauveria bassiana, B. brongniar�i, Metarhizium anisopliae, Nomuraea rileyi (fungi) Vairimorpha (protozoa) and Steinernema fel�ae (nematode) have been documented on legume crops. However, NPV has been reported to be more poten�al agent and therefore, has been used in field experiments either alone or in combina�on with other biological or chemical agents. Three to four sprays of HaNPV @ 250 larval equivalent (LE) per ha in the evening hours at an interval of 7-10 days during the early stages of the pest effec�vely controlled it on pigeonpea (Santharam et al., 1981). Santharam et al. (1994) no�ced equal amount of reduc�on in the borer infesta�on by B. thuringiensis subsp. kurstaki alone and in combina�on with HaNPV and the predator Chrysoperla carnea in two successive field experiments under irrigated situa�on. Bt formula�ons Biobit @1.0 and 1.5 kg/ha (Shankar et al., 1992) and Halt @1.0 kg/ha (Bakhe�a et al., 1998) controlled pod borers and increased grain yield. Of the four fungal pathogens recorded to cause disease to the lepidopteran borer in pigeonpea, li�le a�en�on is being given to exploit the poten�al of Nomuraea rileyi on this pest. Environmental humidity plays key role in the epizoo�cs of N. rileyi. Higher rates of fungal infec�on in H. armigera were found on pigeonpea (37%) compared to Phaseolus vulgaris (28.2%) and tomato (28.5%) in Karnataka (Gopalkrishnan and Narayanan, 1989).
Soybean: Helicoverpa armigera, leaf miner Aproaerema modicella, girdle beetle, Obereopsis brevis, tobacco caterpillar, Spodoptera species, green semilooper, Chryrodecxis acuta, lea�opper, Empoasca kerri and whiteflies, Bemisia tabaci are the major insect-pests of soybean. Shanower et al. (1992) reported nine primary and eight secondary parasitoids emerged from larvae of Aproaerema modicella, and killed up to 50% of the leaf miner larvae sampled. They reported that the combined effects of all mortality agents including pathogens killed up to 95% of the leaf miner larvae per sample period. However, use of insec�cides in sprayed plots reduced the efficacy of parasitoids. Two species of ladybird beetles, Coccinella septempunctata and Menochillus sexmaculata and two predatory spiders, lynx spider and golden predatory spider were found preda�ng mainly on lea�oppers and whiteflies (Netam et al., 2013). A predatory pentatomid bug, Eocanthecona furcellata was also reported feeding on lepidopterous larvae. Gupta (2002) reported natural occurrence of the entomopathogenic fungus, Nomuraea rileyi on soybean green semilooper, Chrysodeixis acuta, in Rajasthan. The influence of pathogens of this species is reported for the first �me. Shanower et al. (1992) reported that pathogens killed up to 30% of the
99

leaf miner, A. modicella larvae. The combined effects of all mortality agents killed up to 95% of the leaf miner larvae per sample period. However, use of insec�cides in sprayed plots reduced the efficacy of parasitoids.
Conserva�on and augmenta�on of natural enemies: Efficacy of natural enemies can be enhanced in legume crops by making specific changes in the cul�va�on prac�ces. Parasi�sm of pod borer larvae on chickpea rows near coriander was high (5.3-6.8%) than without coriander (0.7-1.0%). High ac�vity of larval parasitoids was a�ributed to the a�rac�on of the parasitoids to flowers of coriander (Pimbert and Srivastava, 1989). Close plan�ng (20x10cm) and mulching with rice straw caused significant reduc�on in groundnut leaf miner incidence and increase in percentage parasi�sm (Logiswaran and Mohanasundaram, 1985). A reduc�on in pest incidence and increase in coccinellids, Coccinella sp. and M. sexmaculata as well as Chelonus spp was observed when groundnut intercropped with pearl millet compared to groundnut monoculture (Kennedy et al., 1990). The intercropping of groundnut with red gram, green gram and soybean is favourable for build up of spider popula�on by serving as ideal niches in turn reduced the pest popula�on (Singh et al., 1991). Intercropping groundnut with Pennisetum glacum enhanced parasi�sm by Goniozus spp. on leaf miner, A. modicella and gave highest net return of 4.7% (Baskaran et al., 1993). Popula�on of coccinellids was higher on groundnut when intercropped with maize, green gram and soybean whereas Chrysopa spp. popula�on was higher when intercropped with maize and soybean intercrops (Sherasiya and Butani, 1999). Intercrops like groundnut, sorghum, setaria, maize, etc. were found to increase the popula�on of natural enemies of pigeonpea pod borer (Venugopal Rao, 1993). Bhatnagar and Davies (1980) however, concluded that within field diversity provided by the intercropping systems may not always contribute to increased preda�on and parasi�sm of insect pest on the principal crops. However, intercrop like co�on in pigeonpea favoured high levels of dipteran parasitoids on both co�on and pigeonpea and parasitoids also got transferred with the pest from co�on to pigeonpea but these dipterans are of li�le benefit as they kill larvae in prepupal or pupal stage.Duffield (1993) observed that adults of Trichogramma were more abundant on pigeonpea, when it flowered a�er sorghum than it flowered simultaneously with sorghum. He concluded that the failure of Trichogramma to parasi�se eggs of H. armigera on pigeonpea was not due to the failure of the adult parasitoids to enter pigeonpea fields but due to other reasons. Romeis et al. (1999) reported that on pigeonpea, H. armigera oviposited more than 74.8% of its eggs on calyxes and pods. Parasi�sm levels in host eggs collected from different plant structures varied significantly with 3.6, 0.3, and 40.7% of eggs on calyxes, pods, and leaves parasi�zed. Earlier studies have shown that calyxes and pods possess long glandular and non-glandular trichomes, and are covered by s�cky trichome exudates which inhibit parasitoid searching behaviour. According to Duffield (1994), oviposi�on of H.
100

armigera was concentrated on the early podding stages of both flushes of pigeonpea. Parasi�sm level on sorghum increased rapidly as egg density increased and reached a peak of 74.6%. Parasi�sm on pigeonpea was concentrated on pod borer eggs laid on the 1st flush of flowers and reached a maximum of 69.2%. However, these high levels of parasi�sm on pigeonpea coincided with the period of parasitoid ac�vity on sorghum. The levels of parasi�sm then declined rapidly and only very low levels were detected on the 2nd flush of flowering on pigeonpea to the extent of 7.8% and it was 34.4% on sorghum. The pa�ern of parasi�sm suggested that transfer of parasitoid occurred from sorghum to pigeonpea but the rapid decline of parasi�sm on pigeonpea indicated that parasitoid popula�ons could not be sustained on pigeonpea, once the influence from sorghum had stopped. Thus the possible method of encouraging the transfer of parasitoids to short dura�on pigeonpea is by producing a more con�nuous cropping environment. When pigeonpea was intercropped with cowpea, soybean, setaria, sorghum and bajra, the coccinellied beetle popula�on was high on pigeonpea upto 90-100 days under intercropping systems as compared to sole crop of pigeonpea. However, this difference was not maintained a�er the harvest of intercrop. Egg and larval parasi�sm levels on pigeonpea were very low (1.4 and 0.2%) as compared to that on sorghum (30.6 and 20.8%) (Hegde and Lingappa, 1995).
Conclusions: Legume crops ecosystem provides a rich diversity of natural enemies but their exploita�on is highly limited. The future research thrust aspects include focused survey to capture na�ve biodiversity of poten�al natural enemies in representa�ve agro-clima�c regions, improving their mass produc�on and release systems and iden�fying ecology-based favourable components for integra�on. Manipula�on of crop ecosystem through crop mixing for conserva�on and build up insect pest dependent arthropods to prey on pest species needs to be explored with more inves�ga�ons. Understanding on the interac�on between biological components in the ecosystem at various tropic levels appears to be the key for successful explora�on of the bioagents as management tools. Insec�vorous birds, especially black drongo, find greater scope as an ecologically suppor�ve approach.
Suggested reading:
Bisane, K. D., Borkar, S. L., Lande, G. K., & Bhalkare, S. K. 2008. Parasi�za�on of Helicoverpa armigera (Hubner) on pigeonpea. Journal of Biological Control, 22(2), 455-461.
Giraddi, R.S., R.A. Balikai, L.K. Naik, S. Yelshe�y and H.N. Sa�gi. 1997. Bioefficacy of NPV compared to insec�cides in the management of Helicoverpa armigera (Hubner). Karnataka J. Agric. Sci., 10: 1197-1199.
Khan, M. A., Tiwari, S., Agnihotri, M., and Pandey, S. 2013. Egg Parasitoids in Legume Crops Ecosystem. In Biological Control of Insect Pests Using Egg Parasitoids (pp. 373-388). Springer India.
101

Lingappa, S., and Hegde, R. 2001. Exploita�on of biocontrol poten�al in the management of insect pests of pulse crops. In R. K. Upadhyay, K. G. Mukerji, B. P. Chamola (Eds.) Biocontrol Poten�al and its Exploita�on in Sustainable Agriculture, Springer, USA, pp. 321-344.
Reed, W., Lateef, S.S., Sithanantham,S. and C.S. Pawar, 1989. Pigeonpea and Chickpea Insect Iden�fica�on Handbook. ICRISAT Informat ion Bulle�n no. 26, pp. 120.
Ridgeway, R L , E G Kmg and J L Carillo 1977 Augmenta�on of natural enemies for control of plant pests m the Western Hemisphere p 379-416 In R L Ridgeway, and S B Vinson (ed ) Biological Control by Augmenta�on of Natural Enemies, Plenum Press, New York, USA.
Romeis, J., Shanower, T. G. and Madhuri, K. 2000. Biology and field performance of Gryon clavigrallae (Hymenoptera: Scelionidae), an egg parasitoid of Clavigralla spp. (Hemiptera: Coreidae) in India. Bulle�n of Entomological Research, 90, pp 253-263. doi:10.1017/S0007485300000377.
Romels, J and T G Shanower 1996 Arthropod natural enennes of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) in India. Biocontrol Science and Technology, 6:481-508.
Saxena, H and R Ahmad 1997, FIeld evalua�on of Beauvena bassiana (Balsamo) Vuillemm against Heilcoverpa armigera (Hubner) infes�ng chickpea. Journal of Biological Control, 1:1 93-96
Shanower, T. G., Wightman, J. A., Gu�errez, A. P., and Rao, G. R. 1992. Larval parasitoids and pathogens of the groundnut leaf miner, Aproaerema modicella (Lep.: Gelechiidae), in India. Entomophaga, 37(3), 419-427.
Sharma, H. C. 2005. Insect pest management in food legumes: The future strategies. M. C. Kharkwal (ed.) Proceedings of the Fourth Interna�onal Food Legumes Research Conference (IFLRC-IV), October 18-22, 2005, New Delhi, India, 522-544.
Soroka J. and J. Otani. 2011. Arthropods of Legume Forage Crops. In Arthropods of Canadian Grasslands (Volume 2): Inhabitants of a Changing Landscape. Edited by K. D. Floate. Biological Survey of Canada. pp. 239-264.
102

DETAILED ACCOUNT OF WHITEFLY, BEMISIA TABACI (GENNADIUS)A VECTOR OF GEMINIVIRUS IN LEGUME CROPS
S.S. SHARMADepartment of Entomology,
CCS Haryana Agricultural University,Hisar-125 004 (Haryana)
Whiteflies belong to order Hemiptera, super family Aleyrodoidea, Sub Order: Sternorrhyncha Family Aleyrodidae are sap sucking insects whose adults bear a remarkable superficial resemblance to �ny moths. In 1758 Linnaeus described European cabbage whitefly (Aleyrodes proletella) ini�ally as a moth. But in 1795 laterielle recognized it as a hemipterous insect. 1450 species whiteflies have been described.
Whitefly name was derived from the presence of secreted powdery wax preened over the body and wings by the adults in almost all the species. Adult whiteflies are almost 1-3 mm in body length and possess seven segmented antennae and a fore-wing vena�on reduced to simple or one branched major vein (R + R ) with R variably s 1
developed. A structure known as 'Vasiform orifice' is unique to aleyrodids and comprises the anus, a 'lingula' which ejects excreta and an 'operculum' which par�ally or wholly covers the orifice. This vasiform orifice is present in all larval (Nymphal) stages and adult.
Whiteflies of both sexes posses a feeding rostrum and are four winged and fully mobile. Reproduc�on is usually sexual, occasionally parthenogene�c. Eggs are always laid on the plant surface. The first instars whitefly larvae are mobile and walk a short distance to locate a suitable feeding site. Once the first moult has taken place, however the remaining three larval instars are sessile and individuals are unable to relocate themselves if feeding condi�ons deteriorate. The final whitefly larval stage is usually termed as a 'puparium' a name which reflects the extreme morphological difference between this stage and the winged adult, which emerges out by rupturing of lines of weakness which are termed as 'transverse and longitudinal' moul�ng sutures. The vacated puparium is o�en termed as pupal case.
Bemisia tabaci transmits geminiviruses to cucurbitaceae, Leguminosae, Euphorbiacea, Malvaceae and Solanaceae and serious pest of both open air and protected cropping system.
History
Bemisia tabaci O. Hemiptera Sub order Sternorrhyncha ; Super Family: Aleyrodoidea Family. Aleyrodidae
103

Host Plants : Total hosts are more than 600 plant species world wide which includes field crops (open air crops), Protected crops, Herbs, Shrubs, Trees, Vegetable, frui�ng trees and flowering plant (ornamental), medicinal plants.
B.tabaci over winters on a variety of cul�vated and wild vegeta�on such as vegetables and cheeseweed and move to spring hosts as potato and cul�vated sunflower. The wild and cul�vated hosts grow in close proximity to one another.
Eggs: Pyriform or ovoid with creamy to brown in colour ,pedicel inserted in leaf �ssue and egg stands on pedicel. The eggs are laid on lower side of leaf. Eggs laid per female
0vary from 48 to 394. Op�mum temperature for hatching is 28.5 C.
Crawlers : (Ist instars nymphs) have func�onal walking legs and antennae. They walk on the leaf surface in search of miner vein. They se�le at one in few hrs and insert their mouth parts into phloem �ssue and extract the sap and remain sessile un�l they reach the adult stage.
nd rdNymphs : (2 and 3 Instars) They have oval or elongate oval body, may be circular or heart shaped. Body depressed, ellip�cal, pale greenish yellow and antennae are present.
rdIn 3 instars antennae directed towards the median line and become hook shaped.
Pupa: Body convex, colour deep yellow, eyes well developed have red eyes, legs as stump like suckers and 7 pairs of dorsal bristles.
Adult: Two pairs of wings fully mobile with white powdery wax all over the body and wings about 1-3 mm body length possessing seven segmented antennae, three pairs of legs and fore wings with simple vena�on. Presence of vasiform orifice. Reproduce sexually and occasionally parthenogene�cally.
Female : Abdomen bigger and broader and dis�nctly yellow.Male : Abdomen narrow and tapering posterioly and creamy yellow in colour.Nature of damage :
1. Whitefly pierces stylet in plant �ssue and suck sap from phloem �ssue. Plant becomes yellow week by excessive drainage of sap.
2. They produce large amount of sugar excreta (Honey dew) (Black sooty mould grows which interfere the photosynthesis).
3. Honey dew a�racts ants which disturb the parasitoids and predators.
4. Produce wooly wax secre�ons.
5. Deform the leavesby sucking the sap.
6. Act as vector of Gemini virus (70 viruses) especially in cucurbits, leguminous, Malvaceae solenaceae & Euphobiaceae families.
Behaviour: Adult whitefly mate during day �me and deposit eggs single or groups of 3-10 eggs on the lower surface of leaf crawlers, nymphs and pupae remain on lower
104

surface of the lower leaves. The adults can be seen feeding on lower surface upper fresh leaves during morning and evening hours and in the lower surface of middle canopy leaves during day �me.
1. Rise in temperature and decrease in RH (Sept.) hastens the life cycle. Drop in temp and increase RH (Dec.) prolongs the life dura�on.
0 02. Eggs don't hatch above 36 C and very less below 10 C.0
3. Op�mum temp. for incuba�on is 23-30 C04. Op�mum temp. for development : 20-31 C
5. Op�mum RH 70%6. Total genera�ons at op�mum condi�ons = 16-17.
Dispersal and migra�on: The migra�on of whiteflies is wing added. The greatest distance covered by co�on whiteflies 7 Km and can go over 20 �. above the crop. The flight starts as soon as there is sufficient light available to s�mulate lake off and goes up to midday. The migrant take maximum advantage of the low wind speed to gain height and take them away. The whiteflies can take off and ignore plant cues in the field. Short range migra�on takes place regularly. Flight occurs during the morning and midday hours.
Natural enemies: Total 200 parasitoids of all species of whiteflies. Encarsia spp.(19 spp) parasitoids on B. tabaci.
Mortality key factors: Among the developmental stages highest mortality remains that of egg and crawler stage and then pupal stage. The nymphs and adults compara�vely stronger than these. The mortality factors are climate and the parasitoids. The mortality due to parasi�sm remains moderate. The higher
ndparasi�sm and less survival per cent on co�on in 2 genera�on but higher st
parasi�za�on and higher survival per cent on brinjal in 1 genera�on indicate that parasi�sm is not a decisive mortality factor.
1. Eggs are most resistant to Bakian giving 3.14% mortality.
2. Methanolic extract are more effec�ve than aqueous in giving the nymphal mortality.
3. Pupae are more resistant as body is protected by fat bodies.
4. Bakian seed extract (1:5 w/v) and Neem seed aqueous extract (0.2-2%) act as repellent and an�feedant for adults, thus reducing the eggs on co�on leaves.
105

MANAGEMENT STRETAGIES FOR INSECT PESTS OF PIGEONPEA
KRISHNA ROLANIA AND S. S. YADAVDepartment of Entomology
CCS HAU, Hisar
Pigeonpea (Cajanus cajan (L.) Millsp.) is the second important pulse crop in
India. It accounts for about 11.8 percent of total pulse area and 17 percent of total pulse
produc�on of the country. Pigeonpea grows well in tropical and subtropical
environments. Pigeonpea is a�acked by 250 insect species belonging to 8 orders and 61
families with 12 major insect pests (Chhillar, B. S. and Singh Ram, 2007). The insect-
pests infes�ng pigeonpea are lepidopterans, dipterans, coleopterans and
hymenopterans bugs which some�me cause considerable economic losses to
pigeonpea. Farmers in many areas apply insec�cides in an a�empt to manage these
pests. This chapter provides descrip�ons of the most common species, their biology,
distribu�on, and damage symptoms. Possible modes of control are also included with
an emphasis on integrated pest management and reduced reliance on insec�cides.
GRAM CATERPILLAR, Helicoverpa armigera (Hubner) (Noctuidae: Lepidoptera)
Amongst medium and late pigeonpea gram caterpillar is a predomenent borer
of southern states of India. H. armigera, a notorious polyphagous pest with wide
distribu�on. Varia�on in adult and larval phases is observed due to its polyphagous
nature. Damage ranges from 46 to 67 per cent on redgram due to this pest. Helicoverpa
readily adjusts with any newly introduced variety. It is observed throughout the year on
one or the other crops viz., peas, tomato, co�on, maize, tobacco, safflower, groundnut,
chillies etc.
The young caterpillars feed on the tender foliage and as they grow they bore
into the pods and destroy the seeds, while feeding it thrusts its head inside the pod
leaving the rest of its body outside. Make large round on each locule. The newly
hatched larvae are sluggish and white green in colour, become ac�ve a�er 3-4 hours.
The full grown larvae are 3.5-4 mm in length with pale green in colour. The larval period
extends from weeks to two months or more depending upon temperature and quality
of food (Sithanantham 1987). Single larvae may destroy 30-40 pods before it reaches
maturity. Full grown caterpillar is cylindrical 40 – 48 mm in length with variable colour,
dark green or reddish brown or brownish and marked with a white broken lines and a
prominent white line along lower part of sides. The emergence and ac�ve period of
moth observed during evening hours and night.
106

Management : It is a major pest on redgram and gram. Hence IPM is important. Among cultural prac�ces deep summer ploughing should be done to expose pupae in soil and follow crop rota�on with less favourable crops like jowar, gingelly, blackgram, horsegram, dry paddy (in redgram). Manipula�on of sowing dates and crop dura�on i.e sowing in mid june shows less pod damage as compared to late sown upto mid july. Incidence of H.armigera was reduced when pigeonpea intercropped with cowpea, groundnut, mungbean, urdbean and fingermillet. (Patnaik et al.1989). As a guard crop 4 rows of sorghum can be planted all around redgram crop. Tolerant varie�es like ICPL–332 and LRG – 41 and varie�es with recupera�ng ability like LRG – 30 should be selected. Avoid cul�va�on of varie�es of pigeonpea with different maturity groups in the same area. Clipping off the terminal twig to remove oviposi�onal niches and raising of rabi redgram to avoid pest.
Pheromone traps @ 10/ha and light traps during August –September and November – December were helpful to monitor the pest popula�on. For effec�ve control need basis spray should be followed and avoid indiscriminate use of insec�cides, synthe�c pyrethroids and mixtures. Spray of NSKE 5 % or neem based insec�cides when eggs and early instar larvae are no�ced · Among microbial insec�cides: spray NPV @ 200 LE/ha and B.t formula�on 400g or 400 ml/ac thrice at weekly interval. For chemical control of H.armigera the economic threshold level is 8-10 eggs or 3-5 small larvae per plant at flowering stage of crop should be considered as signal to start the control. Spray chlorpyriphos 2.5 ml/l or quinalphos 2 ml/l or acephate 1.5 g/l at flowering and frui�ng using 750 –1000 l of spray fluid with High Volume sprayer. In severe incidence, indoxacarb 1 ml/l or spinosad 0.3 ml/l were effec�ve and adopt community approach.
REDGRAM PLUME MOTH, Exelas�s atomosa (Walschingham) Spenarches caffer
(Pterophoridae: Lepidoptera)
It is a specific pest of redgram in India, AP. Assam, Bihar, Maharashtra and Tamilnadu. Minute, single eggs are laid on flower buds and pods. Tiny caterpillar scrapes the pod surface and cuts a hole and thrusts the head into it and feeds on seed by remaining outside. The caterpillars bore into green pods and feed on the developing seeds which are more or less completely devoured or eaten away. Also feeds on flower buds. This pest is usually found at flowering and known to cause heavy damage to redgram. A�ack by this pest can cause severe bud, flower and pod drop. The larva never enters inside the pod and feeds remaining outside the pod. The damaging symptoms are small hole on seeds, dropping of flower buds and flowers in severe cases, completely eaten and devoured seeds. The damage is more in determinate varie�es as compared to indeterminate ones ( Reddy et al., 2001)The full grown caterpillar is about 12 mm long greenish brown, and are fringed with short hairs and spines all over the
107

body. The pest completes about 6 genera�ons November to April (Lal and Ka� 1998).
Management : Pigeonpea cul�vars PDA-92-3E, PDA-89-2E, GAUT 85035 AND BDN-2 are reported to posses resistance against this pest (Kalariya et al.1998). Early sown varie�es exhibit less damage from plume moth as compared to late sown variety. In ini�al stage caterpillars can be collected by shaking shoots and destroy them. The incidence of plume moth can be reduced by using combina�on of NPV, Bacillus thuringiensis subsp. Kurstaki and predator chrysoperla carrnea (Santharam et al.,1994). Foliar sprays should commence at 50% flowering with quinalphos 2 ml/l or carbaryl 3 g/l. Spray of fenvalerate 20EC (0.02%) and methomyl 24L (0.1%) are the effec�ve against this pest ( Singh et al. 2001).
REDGRAM POD FLY, Malanagromyza obtuse (Malloch), (Agromyzidae: Diptera)
It is a major pest of redgram, soybean and cowpea. A�ack is more in north and central India and Karnataka. The grain damage caused by this insect pest varies from 20 to 80%. Small metallic black fly thrusts its minute eggs into the �ssues of the tender pod and flower buds. Fly pierces pericarp with oviposi�or and lay eggs which are seen like needles projec�ng inwards from the pods. Tiny maggots burrow into pods and feed on young seeds. In affected pods, no visual symptoms are observed regarding its entrance. Ini�ally larva bores into epidermis without rupturing the seed coat. In the second and third instar stages, the larva bores into cotyledons and in most instances one seed is sufficient for the maggot to complete its development. The final instar larva leaves the seed and prior to pupa�on, windows the pods and pupates either in the pod cavity or in the pod wall �ssue.The damaged seeds are unfit for consump�on because par�ally damaged seeds easily comes under bacterial and fungal a�ack (Khedar.O.P, 2008). Diagnos�c symptoms are: Discoloura�on of the infested pods visible in green podded varie�es. At the later stage of infesta�on, the holes about 1mm in diameter covered with a thin membrane readily seen on the infested pod. Exit holes visible a�er the adult emergence. The pest infesta�on can be iden�fied only a�er adult emergence of first genera�on. Maggot is creamy white in colour. Adults emerge by cu�ng the thin spot already made by maggot.
Management : In endemic areas early sowing should be done to avoid the pest incidence. The popula�on of redgram pod fly can be reduced by proper sanita�on through removal of alternate host like Rhycosia minima, Atylosia spp. etc. Removal of affected pods of first brood during winter also helpful to minimize the pest popula�on. Mul�plica�on rate of pigeonpea pod fly is low during winter month, therefore early maturing crop (February- March) has low damage than late maturing (April-May). Mixed or intercropping of pigeonpea with other crops helps to reduce the damage of pod fly as compared to sole crop.(Singh and Singh,1978). Destruc�on of precocious plants (early flowering plants) helps in reduc�on of popula�on build up of pest. Use of
108

resistant varie�es like PDA-89-2E, PDA-88-2E provides 60-70% crop protec�on in comparison to local checks. Use of parasite like Euderus agromyzae, or E.Agromyzae for pre-pupal stag and Euderus lividus for pupa are beneficial to minimize the pest popula�on. Foliar sprays with monocrotophos 1.5 ml/l or dimethoate 2 ml/l are effec�ve against this pest.
SPOTTED POD BORER, Maruca vitrata (Geyer) (Pyralidae: Lepidoptera)
This pest is found throughout South India on pulses and beans. The larvae web the leaves and inflorescence, and feed inside on flowers, flower buds, and pods. This typical feeding habit protects the larvae from natural enemies and other adverse factors, including insec�cides. The flower bud stage is preferred most for oviposi�on, and it is at this stage that the young larvae cause substan�al damage, and reduce the crop poten�al for flowering and fruit se�ng. The young larvae bore into the flower buds, and cause flower shedding by destroying the young flower parts enclosed in the sepals. The successful establishment of this pest at the flower bud stage is significant in rela�on to subsequent damage, reduc�on in grain yield, and efficiency of control. At this stage the damage is largely internal and there is li�le or no sign of damage externally. Usually more than one larva is present in each flower. The larval movement is facilitated by the silken threads, which are used as bridges between flowers. A�er ini�al dispersal, larval development is completed on several flowers /pods. The larvae move from one flower to another as they are consumed, and a larva may consume 4 -6 flowers before larval development is completed. Third to fi�h-instar larvae were capable of boring into the pods. Adults of legume / spo�ed pod borer are 13 mm in length with a wing span of 29 mm. The forewings are dark brown while hind wings are light brown in colour. Mass excreta can be seen at the entrance of larval burrow.
Management: Affected pods should be removed. Use of resistant varie�es MPG-537-M 1-2-1B and MPG 664-M1-2-M13 also helpful to reduce the pest indidence. For chemical control 10 Larvae / 100 flowers are considered as a tenta�ve threshold for legume pod borer. Foliar spray from flower bud ini�a�on with combina�on of chlorpyriphos 2.5 ml/l or quinalphos 2 ml/l or novaluron 0.75 ml/l or spinosad 0.75 ml/l or lamda cyhalothrin 1 ml/l in combina�on with dichlorvos 1 ml /l at wekly intervals is effec�ve.
BEAN APHIDS, Aphis craccivora Koch (Aphididae: Hemiptera)
Both nymphs and adults suck sap from tender parts,i.e., top soots, flowers, buds, developing pods and usually from the underside of the leaves. The leaves, shoots and flowers deform and do not develop properly. The insect secretes honey dew, due to which black mould developes on infested plant which interferes in photosynthesis. This pest is act as a vector of viral disease. In len�l it transmits tobacco streak virus and cucumber mosaic virus. The adults are small sized insects about 2mm long, pear shaped green or greenish black in colour. Males are rare. The most common mode of
109

reproduc�on is vivipary and parthenogenesis. The colour of newly laid nymph is translucent except the reddish-brown compound eye. The apterous female (wingless) is shining dark brown and greenish black when alate (winged) with transparent wings.
Management: By keeping the crop free from moisture stress and maintaing wider spacing pest popula�on can be minimize. Predators Coccinella septumpuncata L., C.transversalis (Fabr), C.nigritus, Vernia sp. and the parasites Trioxys indicus and Praon sp. are known to control aphid. Spraying with tobaccoo decoc�on (1 kg tobacco boiled in 10 lit of water of ½ hour and make up to 30 lit + 100 g soap) and systemic insec�cides like dimethoate 2 ml/l or phosphamidon 2 ml/l or malathion 2 ml/l are effec�ve.
TUR POD BUG/REDGRAM POD BUG, Cavigralla gibbosa Spinola, (Coreidae: Hemiptera)
Hundreds of nymphs and adults suck sap from the shoots and pods. Shoots fade, pods shrivel and seeds with dark patch loose germina�on capacity due to the feeding of bugs. The adult bugs are greenish brown in colour, having a spined pronotum and femur, swollen at the typical end. The bugs are about 20 mm long. The young nymph is reddish and show prominent lateral spines on the prothoracic and abdominal segments.
Management: The pest popula�on can be minimize by collec�on of bugs and their destruc�on by dipping into kerosinized water. Dus�ng or spraying with carbaryl 10D @ 10 kg/ac or foliar spray with dimethoate 2 ml/l or monocrotophos 1.5 ml /l are effec�ve.
GREEN PLANT BUG/STINK BUG, Nezara viridula L. (Pentatomidae: Hemiptera)
It is a polyphagous pest. Nymphs and adults suck sap from tender shoots and developing pods in large numbers, due to that shoots fade. The seed become shriveled with dark patches. They are usually green in colour, but may occasionally be a reddish brown. The “s�nk bug” is so named because of the strong odor emi�ed from scent gland when disturbed. Adults ac�vate during April to June and with first rains they mate and oviposit.
Management: Mechanical collec�on of nymphs and adults and destroying them. Foliar spray with carbaryl 3 g/l is found effec�ve.
BLISTER BEETLE, Mylabris pustulata Thunb. (Meloidae: Coleopteran)
It is a polyphagous pest a�acking various pulses, malvaceous and cucurbitaceous crops. Pigeonpea is a preferred host of blister beetle than cowpea and urdbean in term of foral damage. Adult beetles feed on different floral parts including petals, anthers, s�gma and ovary. Some�mes, ovary is completely eaten away resul�ng no pod forma�on. The larvae do not feed on plant. The beetle is medium-sized and 12.5-25.0 mm long. Each black elytron has one orange red spot anteriorly, one wide
110

orange red band in the middle and second wide orange band posteriorly. They are bright metallic-blue, green, black and yellow or brown in colour.
Management: Manual picking and killing in early morning hours is benificial. As a natural enemy long-eared Indian hedgehog, Hemiechinus auritus (Grey), feed on blister beetle and minimize the pest popula�on. Dus�ng of malathion (5%) or Carbaryl (10%) @ 25 kg/ha and spray of dimethoate 0.03% is found effec�ve to manage the pest.
GREEN POD BORING CATERPILLAR OR LENTIL POD BORER, E�ella zinckenella Treitschke (Pyralidae: Lepidoptera)
It occurs on redgram, horsegram and other pulses and green manure crop like sunhemp. The full -grown larvae are rosy and feed on floral parts, newly formed pods and seeds in developing pods. Faucal pellets inside damaged pods and small round holes on redgram pods plugged with excreta can be no�ced. The moths are grey with a wing expanse of 25 mm. The fore wings have dark marginal lines and are interspersed with ochreous scales. Hind wings are light grey.
Management : The incidence can be reduced by growing resistance genotype like: ICEAP00073 of pigeonpea showed less damage to this pest (Ritchie et al,2000). Spray the crop with 2.25 kg Sevin 50 WP (Carbaryl) in 250 litres of water per hectare at flower ini�a�on stage. The soil applica�on of aldicarb 1.5 kg per hectare before sowing of the crop which gave 27-29 per cent reduc�on in the infesta�on of pest.
BLUE BUTTERFLIES, Catochrysops cnejus, Lampides boe�cus, Catochrysops strabo
(Lycaenidae: Lepidoptera)
The larva bores into the buds, flowers and green pods just within couple of hours. The eggs are laid on flower buds. A�er hatching the �ny caterpillars enter into unopened flower bud and feed inside. A�erwards they may a�ack another flower or enter a pod and feed on the developing seeds. The holes can be seen even a�er maturing of the pods. The female is bigger than the male. The colour of wings is violet metallic blue to dusky blue.
Management : Use of resistant verie�es ICEAP 00073 to reduce the pest poula�on. Foliar spray with carbaryl (sevin) 1.75 kg of 50 WP and parasi�sa�on with Neotypus intermedius and Brachymeria lasus is effec�ve.
RED GRAM MITE, Aceria cajani (Eriophyidae: Acarina)
It infests underside of tender leaves, causing yellowing of leaves and suppression of flowering and frui�ng. It transmits pigeonpea sterility mosaic virus. A single eriophyiid mite is sufficient to transmit disease. Disease can be iden�fied from a distance as patches of bushy, pale green plants without flowers or pods. Leaves are small, show a light and dark green mosaic pa�ern.
111

Management: Destruc�on of perennial and volunteer pigeonpea and the ratooned growth of harvested plants which provide reservoir of mite vectors and pathogens. Tolerant varie�es like ICPL 87119, ICPL 227, Jagruthi, Bahar should be sown to reduce the pest incidence and foliar spray with dicofol 3 ml/l or we�able sulphur 3g/l in early stages of plant growth is effec�ve against mite vector.
LEAFHOPPER, Empoasca kerri (Cicadellidae: Hemiptera)
Small greenish yellow nymphs and adults suck sap from leaves resul�ng in severe case, the leaves turn brown, dry and bri�le, a condi�on called “hopper burn”. A�acked leaflets become cup shaped and yellow at edges. Heavy a�ack results in the leaflets turning red-brown with subsequent defolia�on and stun�ng. The adults are greenish- yellow, acquiring a reddish �nge in winter. They are a�racted to light at night �me.
Management: Intercropping of urd bean or cowpea with pigeonpea delayed appearance of lea�opper on this crop. Chrysopa symbela, Coccinella septumpunctata and Camponotus sp.preda�ng upon the pest and reduce the pest popula�on. Spraying with thiamethoxam 0.4g/l or imidacloprid 0.4 ml/l have been found effec�ve.
Suggested Reading:
Lal,S.S. and Ka�,G.(1998). IPM of pod borer complex infes�ng pigeonpea. In IPM system in agriculture. Vol.IV.Pulses (Eds R.K.Upadhyey,K.G.Mukerji and R.L.Rajak), pp.79-128.New Delhi: Aditya books Pvt.Ltd.
Ritchie,J.M., Polaszek,A.,Abeyasekera,S.,Minja,E.M. and Mviha,P. (2000). Insect pest incidence in seed pods of pigeonpea genotypes in on-farm trial in Southern Malawi. Interna�onal Chickpea and Pigeonpea Newsle�er 7, 50-52.
Singh,R.N. and Singh,K.M. (1978). Influence of intercropping on succession and popula�on build up of insect pests in early variety of red gram, Cajanus cajan (L.) Millsp. Indian Journal Of Entomology 40, 361-375.
112

TERMITE MANAGEMENT IN LEGUME CROPS
S S YADAV, J.P.BHANOT AND KRISHNA ROLANIA Department of Entomology,
CCS Haryana Agricultural University,Hisar-125 004 (Haryana)
Termites are a group of insects belonging to class Isoptera consis�ng of 2,500 species of which 300 are considered as pests (Pearce, 1997). Termites are one of the most damaging pests in the tropics and can cause considerable problems in agriculture, forestry and housing. They feed on dead organic material such as crop residues, mulches and soil organic ma�er (humus). However when this type of food is not available they will eat live plant material including crops such as groundnuts, millets and maize. Termites can a�ack plants at any stage of development from the seed to the mature plant. They are social insects that live together as a colony in a nest. Colony members belong to one of three interdependent groups with specialized form and func�on known as castes. The three basic castes present in the colony are workers, soldiers and reproduc�ve forms. Workers and soldiers are wingless, sterile and blind. Workers construct the dis�nc�ve shelter tubes and collect food to feed the young and other members of the colony. The primary func�on of the soldiers is to defend the colony, usually against ants, which are their main enemies. The reproduc�ve caste is usually referred to as the king and the queen. They are responsible for the produc�on of fer�lized eggs for the colony and of specialized chemicals (hormones) important for managing the inhabitants of the colony. Mature colonies produce winged reproduc�ve forms or alates at certain �mes of the year. A�er the dispersal flight, the alates a�empt to find mates and formed new colonies. U�lizing cellulose as their food source and living in colonized form, termites are the most successful and long lived of all the social insects. They depend en�rely on wood, either living or dead, or the woody �ssue of plants, intact or par�ally decayed and also in the form of humus and dried animal dung.
Different methods of termite management1. Cultural methoda. Adding organic material to the soil
Termites prefer to eat dead plant material. Their a�acks are thought to be related to soils with low organic ma�er content. This is because such soils do not contain enough food for termites to live and they resort to feeding on living plant material. Adding compost or well-ro�ed manure to the soil and growing green manures helps to increase the organic ma�er in the soil. Where possible, green manure crops can be ploughed into the soil. Moisture plus organic ma�er a�ract the termites and prevent them from a�acking the target crop. Castor press cake can be incorporated into
113

the soil in order to control the termite infesta�on. It is suggested to add it into the opened furrow before sowing the crop.
b. Date of sowing
By growing crop when the pest is not present or least abundant, damage can be checked upto an extent. Gram crop, when sown on five different dates at three week interval (mid October to mid January), resulted in more termite damage in the crop sown during mid October or early November than in the later dates (Bhanot et al., 1994; Sharma, 1986).
c. Cul�va�ng on ridges
Ridges are usually made along contours and the soil is finally shaped into a ridge form. In these opera�ons, termite colonies are destroyed or exposed to predators such as birds.
d. Crop rota�on
Plan�ng the same crop on the same land year a�er year reduces soil fer�lity and structure. Crops growing in such condi�ons will be weaker and suscep�ble to termites. Crop rota�on can play an important role in reducing termite a�ack. This can prevent pest and disease buildup and also help the soil to recover nutrients. In some parts of India, farmers cul�vate castor (Ricinus communis) crop in severely termites infested field and doing so they found that the termite infesta�on has substan�ally been decreased in the next cropping season (HDRA Report, 2001).
e. Irriga�on
Irriga�on is also used to minimize the effect of the termites. When the farm is regularly irrigated, the ac�vi�es or damages of termites are reduced. Termite damage was lesser in surface irrigated than in drip irrigated groundnut crop (Jayan� et al., 1993). It can be concluded that termite damage is nega�vely correlated with irriga�on levels.
f. Timing of harves�ng
Crops are more seriously damaged towards harvest than earlier period of the season. Therefore prompt harvest is recommended.
g. Plant density/seed rate
Seed rate influences the spacing and this may affect the rela�ve rate of growth of a plant and its popula�on per unit area as well as the incidence of insect- pests over them. Close spacing increases the effec�veness of natural enemies and results in greater control of pests popula�on (NAS, 1969; Sharma, 1986).
h. Mulching
114

Termite caused more damage to drying groundnut pods in case of sunhemp mulch, while Celosia argentea mulch afforded intermediate protec�on (Gold et al., 1980). They also observed lesser termite damage in pods placed on the top of mulch than mixed in them.
i. Type of soil/crop varie�es
Termite damage is more in light soils under condi�ons of low-inputs and high temperature (Verma and Kashyap, 1980). No termite damage in black soils of northern Gujarat was observed (Jhaveri, 1971).
j. Other prac�ces
i) Remove diseased or damaged plants and weeds taking care not to damage the roots of healthy plants.
ii) Use only well ro�en manure, otherwise termite incidence is aggravated.
iii) Subterranean termites can be controlled by destroying queen by digging it out from termitarium.
iv) Harvest quickly and remove and burn plant residues which form source of further infesta�on.
2. Mechanical control
Digging of shallow trenches in the fields reduced the termite incidence. Removal of plant debris and other crop residues resulted in reduced termite damage.
3. Biological control
Biological control measures against termites are generally difficult because of their social nature and secure enclosed environments that protect them against most antagonists. Entomopathogenic fungi, nematodes and bacteria are some of the biological control agents of termite control.
a. Use of bio-pes�cides
Botanical pes�cides possess an array of proper�es including insec�cidal ac�vity, repellency to pests, an�feedency, insect growth regula�on, toxicity to agricultural pests. Plant parts and plant extracts can be used effec�vely. These can be removed from the plant and used as a natural insec�cide by grinding up the relevant parts, placing in boiling water, s�rring and leaving to soak. The mixture is then sprayed onto the infested crop. Powdered fruit, leaves, bark and seeds of Melia azedirach and their methanol extracts were effec�ve against Odontotermes formosanus and Coptotermes formosanus (Lin and Wang, 1988). Feeding for 7 days resulted in termite mortali�es of 100, 90, 85 and 65 per cent for fruits, leaves, bark and seeds, respec�vely.
115

b. Fungi
Entomogenous fungi Beauveria bassiana and Metarihizium anisopliae have been widely used for the control of termites. Ac�ve workers were killed within 12 days when exposed to sporula�ng fungus for 24 hours. B. bassiana isolate (Baptala) was the most
4effec�ve fungal pathogen when applied at 7.98xl0 conidia/ml (Khan et al., 1993 and Delate et al., 1995). Whereas Chai (1995) observed that Metarihizium anisopliae was the most effec�ve @ 6x 10 spores/ml. In Australia, M. anisopliae has been applied to kill the colonies of mound and tree nes�ng termites (Milner and Steples, 1996). The live conidia are repellent to the termites and can be used as a barrier treatment. Bioblast is an important commercial available formula�on of M. anisopliae (Quarles, 1999).
c. Nematodes
An entomogenous nematode, Steinernema fel�ae was tested as a biocontrol agent against a sub-terranean termite (Re�culitermes �bialis). However, a large number of nematodes were required to get effec�ve kill of termites under laboratory
7 2condi�ons (Epsky and Capinera, 1988). In field, nematodes were applied @ 1x10 /m , which were effec�ve only for 2 to 4 weeks. But Khan et al. (1993) got almost 100 per cent mortality of termites with in a week under laboratory condi�ons.
d. Viruses
Al Fazairy and Ahlam (1987) showed that a nuclear polyhedrosis virus isolated from the co�on leaf worm, Spodoptera li�oralis could infest termites.
e. Encouraging predators
Termites have many predators because they provide a source of protein. Insects that eat termites include spiders, beetles, flies, wasps and especially ants. Other predators including frogs, rep�les, birds and mammals such as bats and monkeys. Encouraging this kind of wildlife will help to reduce the number of termites.
4. Chemical control
Before the advent of organic synthe�c insec�cides, the termite control in crops was largely done by adop�ng suitable cultural prac�ces. Later on inorganic insec�cides viz., arsenicals and mercuric compounds were used, which posed hazards to human beings and ca�les. Systema�c studies on termite control were, however, taken up only a�er the World War -II. Insec�cides have been applied as presowing soil applica�on in fields and with irriga�on water for termite control in various crops. Of late, seed treatment with insec�cides has been found not only cheaper and ecofriendly but also more effec�ve than above conven�onal methods. Therefore, it is becoming more popular. Seed treatment with insec�cides in gram (Verma et al., 1978; Bhanot et al., 1995) earlier with aldrin and at present with chlorpyriphos, formothion and monocrotophos has been perfected and recommended.
116

Thiamethoxam (Actara) 25 WG was evaluated @ 0.03 and 0.10 ppm against three species of termites: Trinernitermes trinervius, Odontotermes smeathmani and Amitermea evuncifer (pests of principal tropical food crops like (maize, rice, sorghum, sugarcane). The effec�ve dose for these termites was 0.03 ppm, which resulted in 100% mortality within 2-8 days-depending upon the species studied (Delgrade and Rouland-Lefevre, 2002).
5. Monitoring of termite popula�on
Termite popula�on in the fields can be monitored with the help of suscep�ble wooden stakes, suscep�ble maize stalks and by the use of soil augurs. Termite popula�on in field has been found posi�vely correlated with termite damage. Bhanot et al. (1984) studied the popula�on build up of termites in barley field using kikar (Acacia arabica) wood stakes.
Conclusion
Termites are the most troublesome pest of agricultural crops. Although, chemical control is an effec�ve measure of protec�on by termites but their excessive use is harmful for our environment and the results are not sustainable and also costlier for farmers. Preven�on is the best feasible and effec�ve op�on to termite control. Field especially during crop produc�on period must be kept in good hygienic condi�on by removal of dead plant material like wood, stalks and any trash. The extracts of many plants were found to be effec�ve against termites. Although some botanicals were not as effec�ve as chemical but they are nontoxic and safe for the environment, biodegradable and renewable source. The use of botanicals and biocontrol agents are a promising alterna�ve to chemical control.
Suggested Reading
Al-Fazairy, A. and Ahlam, A. 1987. Infec�on of termites by Spodoptera li�oralis nuclear polyhedrosis virus. Insect Science & its Applica�on. 9: 37-39.
Bhanot, J.P.; Sharma, A.K.; Batra, G.R. and Verma, A.N. 1994. Influence of different levels of irriga�on and fer�lizer on termite damage and yield of gram crop raised from aldrin treated and untreated seed. J. Insect. Sci. 7: 115-116.
Bhanot, J.P.; Verma, A.N. and Kashyap, R.K. 1984. Popula�on dynamics of termites in barley fields and correla�on between termite popula�on and termite damage. Z. ang. Ent. 98(3) : 234-239.
Delgrade, S. and Rouland-Lefevre, C. 2002. Evalua�on of the effects of thiamethoxam on three species of African termite (Isoptera : Termi�dae) crop pests. J. econ. Ent. 95(3): 531-536.
Epsky, N.D. and Capinera, I.L. 1988. Efficacy of the entomogenous nematode, Steinernema felitae against a subterranean termite, Re�clilitermes �bialis (Isoptera : Rhinotermi�dae). J. econ. Ent. 81(5): 1313-1317.
117

H D R A r e p o r t , 2 0 0 1 . T e r m i t e C o n t r o l : A c c e s s e d o n l i n e a t h�p://www.gardenorganic.org.uk
Jayan�, M.; Singh, K.M. and Singh, R.N. 1993. Differen�al influence of drip and surface irriga�on on pest complex of groundnut. Indian J.Ent. 55(2) : 124- 13 1.
Khan, H.K.; Jayaraj, S. and Gopalan, M. 1993. Muscardine fungi for the biological control of agroforestry termite, Odontotermes obesus (Rambur). Insect. Sci. & its Applica�on 14(4): 529-535.
Lin, T.S. and Wang, C.L. 1988. The an�-termite proper�es of extracts from Melia azedarach Linn. Bulle�n of the Taiwan Forestry Research Ins�tute 3 (4) : 255-261.
Pearce, M.J., 1997. Termites – Biology and pest management. CAB Interna�onal, Wallingford, Oxon, UK, pp. 172.
Sharma, A.K. 1986. Effect of crop management prac�ces on termite incidence in gram crop. M.Sc. Thesis, Haryana Agri. Univ. Hisar, pp. 42.
Verma, A.N. 1980. Effect of number of irriga�ons on termite damage in wheat crop. Haryana agric. Univ. J. Res. 10(4) : 564-565.
Verma, A.N. and Kashyap, R.K. 1980. Termites - their damage and control in field crops. Memoir No.8, New Delhi. Entomological Society of India, pp. 53.
118

ROLE OF INSECT POLLINATORS IN PRODUCTION OF LEGUME CROPS
S.K.SHARMA AND YOGESH KUMARDepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
Many entomophilous crops require cross pollina�on for their seed/fruit set. Cross-pollina�on by bees is the most effec�ve and cheapest method of increasing yield in most of these crops (McGregore, 1976; Free, 1993). India presents diverse clima�c and geographical condi�ons, therefore, wide variety of crops are grown here. Many of these are entomophilous and their pollina�on status is needed to be known. Legume plants are vital to world agriculture as they help to improve livestock and the soil. Large amounts of legume seed are required annually for plan�ng, especially when intervals between crop rota�ons become shorter. Improved varie�es with higher yields are always difficult to obtain so that good pollina�on of exis�ng varie�es is essen�al. Poor weather, inadequate irriga�on, insect pests, diseases and inefficient harves�ng have caused the lower yields. However, inadequate pollina�on is the most common cause albeit insect pests frequently inflict dras�c reduc�ons.The pollina�on requirements for legume crops vary according to whether they are self-fer�le or self-sterile. For self-pollina�ng varie�es bees are of li�le value for peas, soybeans, peanuts, snap beans and subterranean clover, however there are certain legume such as pigeonpea in which bees play vital role in pollina�on and seed produc�on. In this ar�cle the role of insect pollinators in pigeonpea and berseem are discussed.
Pigeonpea (Cajanus cajan (L.) Millsp.) is a member of the family Leguminosae. It has vernacular names like arhar, tur and red gram. It is an important pulse crop in the semi arid tropics. Pulses are important sources of protein, which perhaps help to meet the most important nutri�onal need of people in the developing countries. Pigeonpea forms the main source of protein in vegetarian diet of people. It contains 23.00-28.60% protein which is rich in amino acids like cys�ne, tryptophan and methionine. It also contains 52.50 to 59.90% starch by weight (Jambunathan and Singh, 1982).Pigeonpea forms a large many branched shrub of about 1 to 3m in different agro-ecological zones of India (Williams, 1983). The yellow or orange papilionaceous flowers develop into green or brown, hairy, 4-7 seeded long beaked pods. Although flowers are o�en cross-pollinated, natural cross-pollina�on ranging from 0.1-48.0% has been reported by various workers (Howard et al., 1919). Variable amounts of cross-pollina�on are associated with differences in locality, cul�var, and �me of year and pollinator popula�ons. S�gma is recep�ve upto 12.00 h and pollen germina�on only 24.00 h a�er dehiscence, thus crea�ng a chance for foreign pollen to germinate. The foreign pollen
119

tube grows much faster than self (Onim et al., 1979a). Flowers provide subsistence for pollinators as these produce pollen and nectar. Insects collect these food substances and during the process, they transfer pollen from one flower to the other. Cross pollina�on provides chances of gene recombina�on and hybrid seed produc�on; thus exploi�ng the heterosis and hybrid vigour in crop seed (Kevan and Baker, 1983). Honeybees, megachilids and wasps are known for their ac�ve involvement in cross pollina�on and improving quan�ty and quality of pigeonpea yield.
The problem with honeybees is that they visit pigeonpea more o�en for nectar than for pollen. When they are searching for nectar they learn to avoid the tripping apparatus, so that only about one percent of the visits results in tripping. When they are looking specifically for pollen they can be very efficient, however. For high yields to occur it is o�en necessary to have more bees on the field than are required to make a honey crop, which is costly for the beekeeper? Problems that occur with wild bees are that they almost always go to pigeonpea primarily for pollen. Many species are very rapid in their movements and trip at about twice the rate of pollen-collec�ng honeybees. But they are not abundant enough on most fields to do a thorough pollina�on, and their reliability is low because their numbers fluctuate from season to season. It is desirable for farmers to cooperate with neighbors in their region especially in the acquisi�on of beekeeping services, which o�en require remunera�on because of low honey yields.
Various workers conducted different trials to establish the role of insect pollinators in pigeonpea are discussed below:
Case studies -1: Pigeonpea flowers are o�en cross-pollinated; natural cross-pollina�on ranging from 0.1-48.0% has been reported by various workers (Howard et al., 1919). Variable amounts of cross-pollina�on are associated with differences in locality, cul�var, and �me of year and pollinator popula�ons. S�gma is recep�ve upto 12.00 h and pollen germina�on only 24.00 h a�er dehiscence, thus crea�ng a chance for foreign pollen to germinate. The foreign pollen tube grows much faster than self (Onim et al., 1979a). Flowers provide subsistence for pollinators as these produce pollen and nectar. Insects collect these food substances and during the process, they transfer pollen from one flower to the other. Cross pollina�on provides chances of gene recombina�on and hybrid seed produc�on; thus exploi�ng the heterosis and hybrid vigour in crop seed (Kevan and Baker, 1983). Honeybees, megachilids and wasps are known for their ac�ve involvement in cross pollina�on and improving quan�ty and quality of pigeonpea yield.
Case studies-2: Studies conducted at Hisar reaveled that 24 insect species belonging to Hymenoptera (17), Lepidoptera (2) and diptera (5) have been reported to visit pigeonpea blossoms. Among them, Megachile spp. was the most dominant cons�tu�ng 42.66 per cent of total insect visitors (Salunke, 2003). Pollina�on studies
120
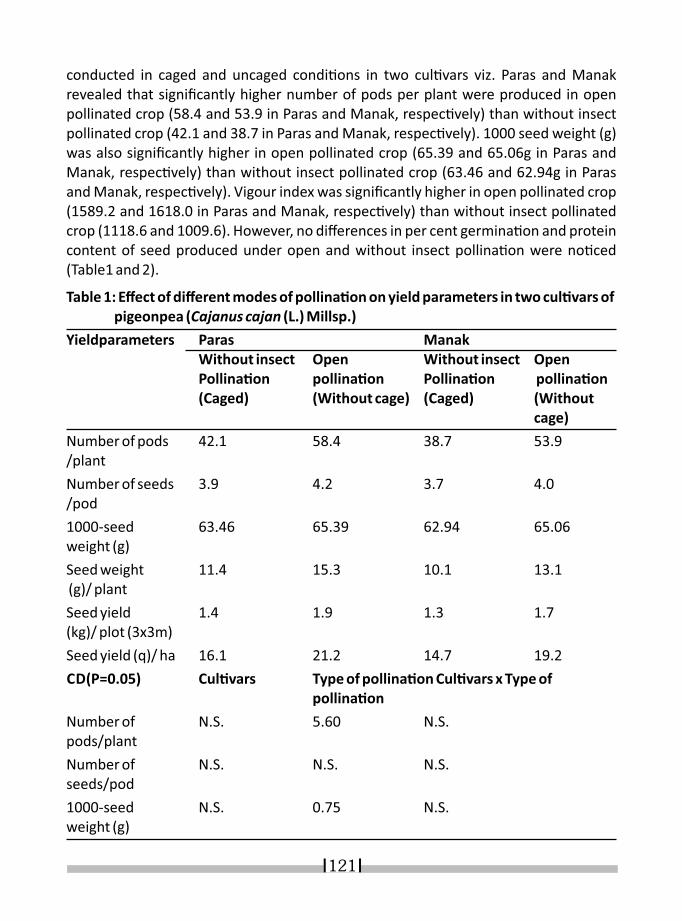
conducted in caged and uncaged condi�ons in two cul�vars viz. Paras and Manak revealed that significantly higher number of pods per plant were produced in open pollinated crop (58.4 and 53.9 in Paras and Manak, respec�vely) than without insect pollinated crop (42.1 and 38.7 in Paras and Manak, respec�vely). 1000 seed weight (g) was also significantly higher in open pollinated crop (65.39 and 65.06g in Paras and Manak, respec�vely) than without insect pollinated crop (63.46 and 62.94g in Paras and Manak, respec�vely). Vigour index was significantly higher in open pollinated crop (1589.2 and 1618.0 in Paras and Manak, respec�vely) than without insect pollinated crop (1118.6 and 1009.6). However, no differences in per cent germina�on and protein content of seed produced under open and without insect pollina�on were no�ced (Table1 and 2).
Table 1: Effect of different modes of pollina�on on yield parameters in two cul�vars of pigeonpea (Cajanus cajan (L.) Millsp.)
Yieldparameters Paras Manak Without insect Open Without insect Open Pollina�on pollina�on Pollina�on pollina�on (Caged) (Without cage) (Caged) (Without cage)
Number of pods 42.1 58.4 38.7 53.9/plant
Number of seeds 3.9 4.2 3.7 4.0/pod
1000-seed 63.46 65.39 62.94 65.06weight (g)
Seed weight 11.4 15.3 10.1 13.1 (g)/ plant
Seed yield 1.4 1.9 1.3 1.7(kg)/ plot (3x3m)
Seed yield (q)/ ha 16.1 21.2 14.7 19.2
CD(P=0.05) Cul�vars Type of pollina�on Cul�vars x Type of pollina�on
Number of N.S. 5.60 N.S.pods/plant
Number of N.S. N.S. N.S.seeds/pod
1000-seed N.S. 0.75 N.S.weight (g)
121
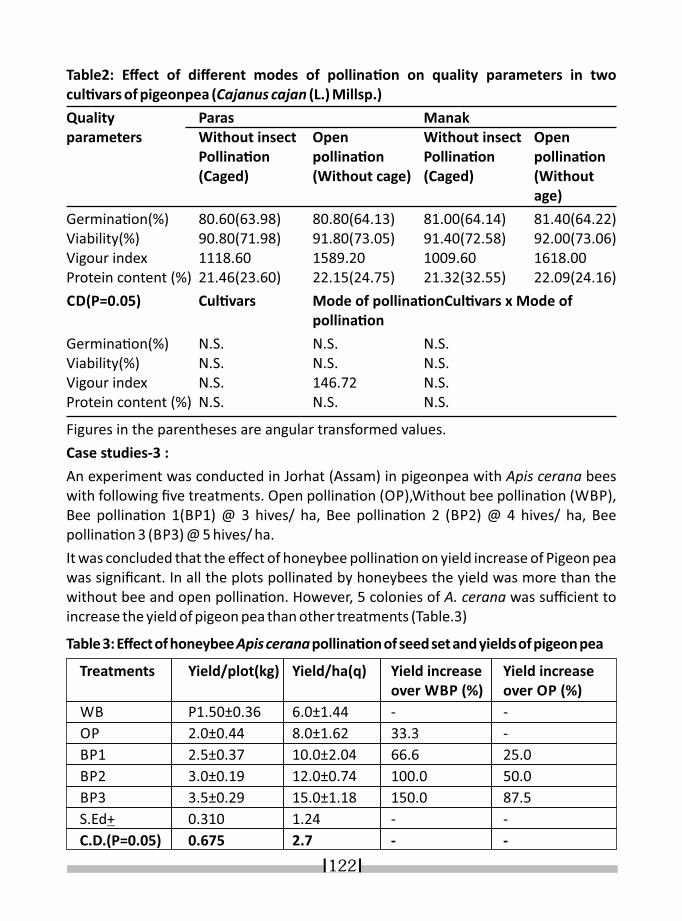
Table2: Effect of different modes of pollina�on on quality parameters in two �cul�vars of pigeonpea (Cajanus cajan (L.) Millsp.)
Quality Paras Manakparameters Without insect Open Without insect Open Pollina�on pollina�on Pollina�on pollina�on (Caged) (Without cage) (Caged) (Without age)
Germina�on(%) 80.60(63.98) 80.80(64.13) 81.00(64.14) 81.40(64.22)Viability(%) 90.80(71.98) 91.80(73.05) 91.40(72.58) 92.00(73.06)Vigour index 1118.60 1589.20 1009.60 1618.00Protein content (%) 21.46(23.60) 22.15(24.75) 21.32(32.55) 22.09(24.16)
CD(P=0.05) Cul�vars Mode of pollina�onCul�vars x Mode of pollina�on
Germina�on(%) N.S. N.S. N.S.Viability(%) N.S. N.S. N.S.Vigour index N.S. 146.72 N.S.Protein content (%) N.S. N.S. N.S.
Figures in the parentheses are angular transformed values.
Case studies-3 :
An experiment was conducted in Jorhat (Assam) in pigeonpea with Apis cerana bees with following five treatments. Open pollina�on (OP),Without bee pollina�on (WBP), Bee pollina�on 1(BP1) @ 3 hives/ ha, Bee pollina�on 2 (BP2) @ 4 hives/ ha, Bee pollina�on 3 (BP3) @ 5 hives/ ha.
It was concluded that the effect of honeybee pollina�on on yield increase of Pigeon pea was significant. In all the plots pollinated by honeybees the yield was more than the without bee and open pollina�on. However, 5 colonies of A. cerana was sufficient to increase the yield of pigeon pea than other treatments (Table.3)
Table 3: Effect of honeybee Apis cerana pollina�on of seed set and yields of pigeon pea
Treatments Yield/plot(kg) Yield/ha(q) Yield increase Yield increase over WBP (%) over OP (%)
WB P1.50±0.36 6.0±1.44 - -
OP 2.0±0.44 8.0±1.62 33.3 -
BP1 2.5±0.37 10.0±2.04 66.6 25.0
BP2 3.0±0.19 12.0±0.74 100.0 50.0
BP3 3.5±0.29 15.0±1.18 150.0 87.5
S.Ed+ 0.310 1.24 - -
C.D.(P=0.05) 0.675 2.7 - -
122

Berseem/ Egyp�an clover, Trifolium alexandrinum L.: Berseem flowers are highly a�rac�ve to bees, which visit it avidly for nectar and pollen. It sets seed abundantly, more than 70 per head, if pollina�ng insects are present (Narayanan et al. 1961).
Pollina�on Requirements: The pollina�on of berseem is quite well established. Chowdhury et al. (1966) stated that it is self-compa�ble, but tripping is essen�al for seed set. They stated that wind was an important pollina�ng agent but presented no data to support this statement. Shamel (1905) reported that bees are absolutely necessary for pollina�on. This has been verified with caged and open plots by Hassanein (1953), La�f (1956), and Narayanan et al. (1961). The reports leave li�le doubt that insect pollina�on is absolutely necessary for profitable seed produc�on. Narayanan et al. (1961) obtained from 19.58 to 70.54 seeds per head of open pollinated berseem plants but only 0.27 to 0.64 seed per head where insects were excluded by 16-mesh wire gauze cages.
Pollinators: Honey bees are the primary pollinators of berseem. They collect both nectar and pollen (Narayanan et al. 1961).
Pollina�on Recomenda�ons and Prac�ces: There are no recommenda�ons on the use of insect pollinators on berseem. Considering its flowering characteris�cs, the absolute necessity of bees in its pollina�on, and the �me of year it blooms, the equivalent of two to four bees per square yard should be sufficient to set a maximum crop of seed.
Trifolium hybridum L.: Alsike clover has been an important summer legume in north central and Northeastern United States; however, it is being replaced by alfalfa. Alsike clover is a perennial but usually grows as a biennial, and in some situa�ons it behaves as an annual. A floret will produce two to three seeds. Flower heads with 100 seeds indicate a good seed crop. The flowers are quite a�rac�ve to bees, especially honey bees, for the nectar and pollen. Pelle� (1923) stated that some beekeepers es�mated that alsike clover might produce 500 pounds of honey per acre in good seasons. This seems abnormally high but indicates the importance beekeepers a�ach to this crop as a source of honey.
Pollina�on Requirements: The florets are largely self-incompa�ble so they must receive pollen from another plant to produce seed (Pieters and Hollowell 1937). In Ohio, Dunham (1939) showed that three cul�vars set only 0.4 to 5.5 seeds per head when selfed, 3.4 seeds per head when wind pollinated, and 120 to 125 seeds per head in a cage with bees. Open plots had 2.6 to 90.4 seeds per head. He calculated that the seed yielding capacity of a field with 1,000 heads per square yard, averaging 50 seeds per head, would be 350 pounds per acre, 90 seeds per head would produce 625 lb/acre and with 120 seeds per head the yield would be 825 lb/acre. In Oregon, Scullen (1956) reported 5.1 seeds per head where bees were excluded but 69.2 seeds per head where bees had access to them. (About 500 lb/acre, according to Dunham's (1939) method of
123

calcula�on.) Oregon averages 300 to 415 lb/acre, but occasional phenomenal yields of 1,000 lb/acre are obtained. The overall United States average is about 140 lb/acre (Wheeler and Hill 1957). Evidently, seed produc�on in most areas could be significantly increased with adequate pollina�on.
Pollinators: Few detailed studies have been made on the insect pollinators of alsike clover. Megee and Kelty (1932) concluded that the honey bee was an effec�ve pollinator on alsike clover in Michigan. Dunham (1957) studied alsike clover seed se�ng for a number of years in an area of Ohio where intensive farming was prac�ced. He found that na�ve bees set only 1.5 to 3 percent of the seeds. Valle (1960) reported that in Finland the honey bee was a much more important pollinator of alsike clover than bumble bees. According to Pankiw and Ellio� (1959), the honey bee is the primary pollinator of alsike clover in western Canada. They found that fields with higher opula�ons of pollinators matured earlier as well as produced more seed. Harrison et al. (1945) concluded that honey bees were essen�al to alsike clover in Michigan. Tucker et al. (1958) reported that honey bees comprised 93 to 99 percent of the pollina�ng insects on alsike clover in Minnesota. Smith (1960) stated that honey bees represent 83 percent of the total pollinator popula�on on alsike fields in southern Ontario. Pankiw and Ellio� (1959) stated that honey bees are excellent pollinators of alsike clover and that they visited the florets at the rate of 18.7 per minute, as compared to 20.0 per minute for leafcu�er bees, and 28.6 per minute for bumble bees. Fischer (1954) reported that honey bees will leave alsike for sweetclover. Wahlin (1962) bserved that when widespread cul�va�on of oil plants occurred in the red and alsike clover seed growing area of Sweden, the bees visited these crops, which resulted in a reduced clover seed harvest. oldawav et al. (1957) in three observa�ons over 2 years obtained yields of 15, 20, and 20 pounds of seed per acre without insect pollina�on; 102, 207, and 368 lb/acre with insect pollina�on, no harmful insect control and no fer�lizer added; and 685, 691, and 808 pounds of seed per acre with pollina�on, harmful insect control, and the addi�on of phosphorus and potash fer�lizers. They recommended at least two colonies per acre, the colonies placed at the edge of the field. Studies have established that the honey bee is the primary insect pollinator of alsike clover and its ac�vity accounts for the bulk of the seed produced. Evidently, seed produc�on of this crop can be significantly increased over current commercial averages if honey bees are present in adequate numbers.
Pollina�on Recommenda�ons and Prac�ces: Pankiw and Ellio� (1959) recommended one colony of honey bees per acre of alsike clover for fields of 50 acres or more but up to three colonies per acre for smaller fields. They obtained about 375 pounds of seed per acre with a bee popula�on of about threequarters of a bee per square yard. Smith (1960) recommended two to three colonies per acre with the colonies placed in or close to the field. Holdaway et al. (1957) recommended at least two colonies per acre. Tucker
124

et al. (1958) concluded that seed yields were increased about 260 lb/acre for each 2colony per acre. They calculated that one bee per 3 yd set 175 pounds of seed per acre,
but one bee per square yard set 800 pounds of seed per acre. The number of colonies per acre necessary to provide the one bee per square yard was not indicated.
Suggested Reading:
Jambunathan, R. and Singh, U. 1982. Grain quality of pigeonpea. Proc. Symp. On Increasing Pulse Produc�on in India-Constraints and Opportuni�es. Oxford and IBH Publ., New Delhi. Pp.389-395.
Kevan, P.G. and Baker, H.G. 1983. Insects as flower visitor and pollinators. Ann. Rev. Entomol. 28: 407-453.
Onim, F.J.M., Eijna�en, C.L.M. and Pathak, R.S. 1979a. Some factors which encourage out-crossing in pigeonpea. Proc. Fourth Int. Symp. Polln., Maryland, pp. 219-225.
Rathi, A. 1993. Field study of bee pollinator-pes�cide interac�on on pigeonpea (Cajanus cajan (L.) Millsp.). Ph.D. Thesis CCS Haryana Agricultural University, Hisar.
Williams, I.H. 1983. The pollina�on of pigeonpea (Cajanus cajan (L.) Millsp.) in India. Second Int. Conf. Apic Torp., New Delhi. pp. 661-666.
125

ROLE OF POLLINATORS IN PRODUCTION OF LEGUMINOUS VEGETABLES
YOGESH KUMAR AND S.K.SHARMADepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
Legume plants are vital to as they help to improve human health, livestock and
the soil. Large amounts of legume seed are required annually for plan�ng, especially
when intervals between crop rota�ons become shorter. Improved varie�es with higher
yields are always difficult to obtain so that good pollina�on of exis�ng varie�es is
essen�al. Poor weather, inadequate irriga�on, insect pests, diseases and inefficient
harves�ng have caused the lower yields. However, inadequate pollina�on is the most
common cause dras�c reduc�ons.
The pollina�on requirements for legume crops vary according to whether they
are self-fer�le or self-sterile. For self-pollina�ng varie�es bees are of li�le value for
peas, soybeans, peanuts, snap beans and subterranean clover. But bees have some
value for lespedeza, lima beans and vetches. In the not self-pollina�ng varie�es some
outside agent like bees is required. Legume crops can be very a�rac�ve to pollinators,
especially Hymenoptera, for both their pollen and nectar. They are capable of yielding
good honey crops .Numerous species of plants in different genera of the family
Leguminosae are called clover. They have certain characteris�cs in common. The leaf is
normally composed of three leaflets. The papilionaceous (bu�erfly-like) or pealike
floret of the flower head consists of a large dorsal standard petal, two lateral wing
petals, and two lower keel petals. It usually has 10 stamens, one of which is free and the
other nine united, which form a tube enclosing the long ovary. The flower trips exposing
the s�gma, which returns to its original posi�on a�er pressure on the petals is removed.
Many species are good sources of honey and pollen for bees. A typical legume flower
comprises a pa�ern consis�ng of sepals, and five petals, including one standard, two
wings, and two keels, stamens and carpels. In addi�on, extra floral organs, such as
s�pule extra-floral nectaries may also play a role in pollinator a�rac�on (Westerkamp
and Weber 1999). Soybean floral nectaries are small, highly developed,and surround
the carpel base. These are heteromorphic structures that produce fluids and/or
vola�les that can a�ract insects (Horner et al. 2003; ).The soybean flower also has well-
defined visible and ultraviolet nectar guides, a tongue channel, tongue guides, and a
nectary ridge that contains stomates that exude vola�les (Erickson 1979; Erickson and
Garment 1979; Horner et al. 2003). Honeybees can be a�racted to soybean flowers and
recognize them by their colour, shape, and vola�les
126

There are special problems with some important legume varie�es. In alfalfa, for example, the blossoms must be tripped in order for pollina�on to occur. To result in the required cross-pollina�on bees must carry out the tripping. In Western North America honeybees, alkali bees, leafcu�er bees and bumblebees are of greatest importance. Although honeybees are usually abundant, they are rather inefficient as pollinators when compared with alkali bees. Their large numbers tends to offset this deficiency. Among the alkali bees, Nomia melanderia is very common. Leafcu�er bees (Megachile spp.) are usually present but in low numbers. Bumblebees (Bombus spp.) occur locally in moderate abundance. The problem with honeybees is that they visit alfalfa more o�en for nectar than for pollen. When they are searching for nectar they learn to avoid the tripping apparatus, so that only about one percent of the visits results in tripping. When they are looking specifically for pollen they can be very efficient, however. For high yields to occur it is o�en necessary to have more bees on the field than are required to make a honey crop, which is costly for the beekeeper? Problems that occur with wild bees is that they almost always go to alfalfa primarily for pollen. Many species are very rapid in their movements and trip at about twice the rate of pollen-collec�ng honeybees. But they are not abundant enough on most fields to do a thorough pollina�on, and their reliability is low because their numbers fluctuate from season to season. It is desirable for farmers to cooperate with neighbors in their region especially in the acquisi�on of beekeeping services, which o�en require remunera�on because of low honey yields.
With the excep�on of the sweet clovers that are a�rac�ve to wasps and bees, bees are the only reliable pollinators for commercial legume crops. Many hundreds of species may be involved. And although they are usually highly efficient, their numbers are rarely high enough to do sa�sfactory pollina�on. Wild bees are best for such crops as red clover and alfalfa. There are a few species of bumblebee that re detrimental to pollina�on of vetch and red clover because they cut into the bases of the corolla tubes. This can ruin red clover seed produc�on in some areas. At the global level, ecosystems are under threat; both produc�vity and geographic distribu�onof crop species will be affected. This, in turn, will affect insect pollinators and plant pollina�on. For example, air pollu�on can lead to reduc�ons in the concentra�on of vola�le compounds that insects detect as they forage for nectar and pollen (McFrederick et al. 2008). Faba bean plants growing under elevated CO2 condi�ons showed an increased floral display and may be more a�rac�ve to pollinators, but pollen flow may not necessarily be improved (Osborne et al. 1997). In the par�cular case of legumes, plant breeders could help preserve and enhance bee fauna by providing suitable floral resources within the crops themselves. In parallel, legume breeding for sustainable agriculture is linked to the development of non-food services such as environmental services (Helenius and Stoddard 2007). Legume improvement demands a shi� in emphasis in breeding programmes from yield alone towards addi�onal emphasis on environmental func�on.
127

Legumes are visited by a great number of social, eusocial, and solitary bees, which can be commercial, feral, or wild (Free 1993; Delaplane and Mayer 2000). Foraging places and nes�ng sites for solitary and social bees are some of the ecological services provided by legumes in a sustainable agriculture. The provision of floral resources within the crop for suppor�ng beneficial insect pollinator popula�ons could be a promising strategy to enhance environmental func�on of legumes. Consequently, we face a situa�on where the development of pollinator-friendly cul�vars is needed. Such a situa�on may require a re-thinking of crop breeding strategies and objec�ves. The role of bee pollinators as agents of pollina�on and of hybridisa�on needs to be considered. Pollinators help increase seed set and self-pollina�on, but more importantly enhance cross-pollina�on (Richards 2001). Pollinators as agents of cross-pollina�on can contribute to breeding strategies. Crops with suitable floral a�rac�veness and rewards can be used to foster pollinator conserva�on, which in turn may lead to improved seed set and/or improved crop yields as well as increased resistance to bio�c and abio�c stresses. There are good breeding reasons as well as ecological reasons for mproved targe�ng of crop pollinator rela�onships (CPR) (Westerkamp and Go�sberger 2000; Aizen et al. 2008). Op�mising CPR would be a key to the establishment of breeding strategies that increase yield and its stability by using social and solitary bees, thereby facilita�ng the development of ''pollinator-friendly'' cul�vars. The beauty of this breeding strategy is that farmers might get addi�onal income from conserva�on payments,and simultaneously from conserving biodiversity(Morandin and Winston 2006). And the farmer would s�ll con�nue to get high yield and yield stability.
Cowpea , Vigna spp.
Pollina�on Requirements: Robbins (1931) stated that the cowpea is capable of self-fer�liza�on, and this is probably the common occurrence, although the flowers are o�en visited by honey bees or bumblebees, a�racted chicfly by the extrafloral nectar.
Pollinators: Purseglove (1968) stated that a heavy insect is required to depress the wings of the flower and expose the stamens and s�gma. The pollen is s�cky and heavy, indica�ng that the plant is not wind-pollinated (Mackie 1946). Cross-pollina�on seems to be associated with areas where bumble bees are numerous. Mackie and Smith (1935) stated that bumble bees are the primary pollinators. The actual value of frequent visita�on by pollina�ng insects to blossoms of cowpeas has not been determined; however, the study by Warnock and Hagedorn (1954) would indicate that such ac�vity is beneficial in increasing the number of pod set, the number of seeds per pod, or both.
Pollinators recorded are given below: European honeybee (Apis mellifera Lin.), Asia�c honeybee (Apis cerana Fab.), Rock bee (Apis dorsata Fab.), Carpenter bee (Xylocopa
128

sp.), Riceskipper (Pelopidas mathias (F.), Cowpea borer (Lampides boe�cus L.), Bumble bee (Bombus spp.), Oriental wasp (Vespa orientalis (Lin.), Tabanidfly (Tabanus spp), Syrphid fly (Eristalis sp.), Housefly (Musca domes�ca Lin.), Fruit fly (Bactrocera sp.), Castor bu�erfly (Ergolis merione Cram.),Cabbage bu�erfly (Pieris brassicae Lin., P. Canidia Lin.), Golden wasp (Vespa magnifica (Smith), Cyntomid fly (Cyntomis passalis (F.), Mustard sawfly (Athalia lugens proxima (Klug.)
Pigeonpea Cajanus cajan (L.) Millsp. Krauss (1932) and Wilsie and Takahashi (1934) considered the pigeon pea to be a normally self-pollinated species. However, a high degree of cross- pollina�on, ranging from 5 to 40 percent, has been observed (Wilsie and Takahashi 1934, Purseglove 1968*, Ma�a and Dave 1931, Krauss 1927, Shaw 1932, Abrams 1967).
Pollinators recorded are as under: European honeybee (Apis mellifera Lin.), Asia�c honeybee (Apis cerana Fab.), Rock bee (Apis dorsata Fab.), Cowpea borer (Lampides boe�cus L.), Bumble bee (Bombus spp.), Tabanid fly (Tabanus spp), Syrphid fly (Syrphus sp.), Housefly (Musca domes�ca Lin.), Rice skipper (Pelopidas mathias (F.), Castor bu�erfly (Ergolis merione Cram.), Cabbage bu�erfly (Pieris brassicae Lin., P. canidia Lin.), Golden wasp (Vespa magnifica (Smith), Mustard sawfly (Athalia lugens proxima (Klug.)
Broad bean,Vicia faba L.: Rowlands (1960) recorded an average of 24 percent set in the open but only 7.3 percent set in the glass house He concluded that some plants are highly self-sterile and some cross be�er than others. Free (1970) stated that the amount and concentra�on of field bean floral nectar was rela�vely low, so the flowers are not a�rac�ve to nectar collec�ng insects. In discussing the field bean inflorescence, nectar secre�on, and bee visita�on, Free (1970*) stated that bee visitors to extra-floral nectaries were more numerous at midday, whereas those visi�ng the flowers were most numerous from 2 to 4 p.m., when pollen was available. He concluded that pollen was more a�rac�ve to bees than the floral nectar.
Pollina�on Requirements: The pollina�on of broad bean and field bean was studied by Free (1966) who concluded that insect pollina�on greatly increased produc�on of broad beans but had li�le effect on produc�on of field beans. Later, Free (1970) concluded that the pollina�on requirements of broad bean and field bean were similar although inadequate pollina�on limited seed produc�on in broad beans more so than in field beans. Darwin (1889) showed that 17 broad bean plants covered with a net to exclude pollinators produced only 40 seeds, whereas 17 exposed plants produced 135 seeds. Free (1966) showed that plants caged with bees produced about twice as many seeds as similar plants caged without bees. .
Pollinators: Bond and Hawkins (1967) and Free (1962) have studied the behavior of bees on field bean. Free (1962) concluded that the ac�vity of the honey bees may vary. They may visit only the extrafloral nectaries, they may visit only the holes cut in the
129

bases of the corollas by bumble bees, or they may visit the corolla for nectar and pollen. Only in the la�er case are the bees of value to the beans as a pollina�ng agent. Bond and Hawkins (1967) placed two colonies by an acre of field bean, and the bees collected sizeable amounts of pollen. However, these workers concluded that bumble bees were primarily responsible for the cross-pollina�on that occurred. Wafa and Ibrahim (1960) concluded that the carpenter bee, Xylocopa aestuans (L.), was the most important wild pollina�ng insect in the Gaza region. Free (1959) concluded that be�er visita�on to the bean flowers resulted when the honey bee colonies were moved to the crop a�er it started to flower. Wa�s and Marshall (1961) showed about 26 percent increase in seed produc�on due to the presence of bees. They also showed the value of bees in se�ng the seed early. In their plots caged with bees and also in their open plots, they found an average of nine, nine, and two pods, respec�vely on the bo�om, mid- and top por�on of the plant, but in the no-bee cage three, five, and four pods were in the same areas.Free (1970) concluded that only insects with long tongues could reach the nectar in the bean flower and that honey bees and short-tongued bumble bees that enter the flower probably obtain only pollen most of the �me. However, some bumble bee species with short tongues, male carpenter bees (Xylocopa aestuans), and certain ants (Cataglyphis bicolor (F.)) bite holes in the base of the corolla and "rob" the nectar without contribu�ng to pollina�on. Honey bees do not make holes, but they will rob nectar from holes made by other insects.
Pollina�on Recommenda�ons and Prac�ces: Allen and Scriven (1957) concluded that one colony of honey bees per acre should give adequate coverage of field beans
Mung bean/Green gram, Phaseolus spp: The inflorescence is an axillary raceme, with 10 to 25 pale-yellow flowers, 1 1/2 to 2 cm long, and clustered at the top. Pollen is shed the a�ernoon before the flower opens the following morning. The flower fades the same a�ernoon. Only about half of the flowers (64 percent) open to permit possible cross-pollina�on.
Pollina�on Requirements: Purseglove stated, "The flowers are fully self-fer�le when bagged and almost en�rely selfpollinated." However, van Rheenen (1964) stated that when he alternated varie�es in the row, he obtained 2.8 to 3 percent crossing.
Phaseolus vulgaris L.: The bean flower is of the typical legume shape; usually whi�sh, but may be �nged to deep violet, purple, or red; and is l/2 to 3/4 inch long. The keel is prolonged in a spirally twisted beak. The style follows the spirals of the keel. There are the usual 10 stamens, the upper one free while the other nine are united into a tube enclosing the long ovary and part of the style. The blooms are loosely sca�ered along a 2- to 3-inch rachis (stem). Weinstein (1926) reported that many pollen grains germinate in the anther, but whether such pollen plays a part in fer�liza�on is unknown. Knuth (1908) stated that although the anthers surround the style and the pollen is released
130

before the flower opens, the pollen does not get onto the s�gma before tripping occurs. When pressure, for example, the weight of a large bee, is applied to the wing petals, the style, with pollen adhering to the brush of hairs on it, projects from the �p of the keel. When pressure is released, the style retracts into the keel, some�mes leaving some pollen exposed. When the s�gma is first exposed, it is free of pollen, and may touch a pollen-coated bee during its brief period of exposure. If this happens, cross-pollina�on can occur. As the s�gma retracts into the keel, it comes in contact with the pollen released by the anthers and selfing may also occur.
Pollina�on Requirements: The flowers of most bean cul�vars are capable of self-pollina�on although cross-pollina�on can and does occur to varying degrees, depending upon the cul�var and the pollinator popula�on (Hawthorn and Pollard 1954, Mackie and Smith 1935, Barrons 1939). Much of the research on bean pollina�on has been concerned only with the impact of the cross pollina�on on varietal seed contamina�on, not on quan�ty or quality of fruit set. Rutger and Beckham (1970) stated that P. vualgaris is primarily a selfing species, but studies of controlled insect pollina�on are needed. Free (1966) concluded that the pollina�on requirements of different species of beans could not be predicted so they must be studied individually. Thorough studies on the possible benefits of maximum pollinator ac�vity on the various types and cul�vars would appear to be highly valuable.Knuth (1908) stated that the anthers surround the style and shed their pollen upon it, but the s�gma is never dusted un�l the keel is depressed. Then the style, with its pollen-covered brush, springs out of the opening. In case the s�gma is touched by the bee, coated with pollen from other plants, before being contacted by its own pollen, cross pollina�on results. However, Taylor (1919) stated that the flowers are en�rely self-fer�le as is abundantly proved by their produc�veness when grown in greenhouses where bees are not present. Yet, he stated that they can be cross-fer�lized by bees, although the percentage of crossing is not great. Mackie and Smith (1935) and Barrons (1939) indicated that cross- pollina�on may amount to more than 8 percent. The pollinator popula�on associated with the crossing was never measured.Jones and Rosa (1928) stated that because the s�gma and the anthers are enclosed within the keel, insects reach them only with difficulty; however, visita�on occurs and crossing results. They considered the- amount negligible. They also stated that selfing occurs freely when flowers are enclosed. Kristofferson (1921) reported 0 to 1.4 percent crossing, and Emerson (1904), 0 to 10 percent crossing. Free (1966) working with 'Processor' cv. of P. vulgaris concluded that "honey bees are unlikely to have much effect on the yield." His data, however, showed that the plants in the cages with bees produced 21 percent more seed weight, with 6 percent more pod weight than plants from which bees were excluded. Beans are visited only sparingly by honey bees, and beekeepers do not consider this crop of significance as a source of nectar or pollen.
131

Foraging and pollina�on ac�vi�es of Xylocopa olivacea (Hymenoptera, Apidae) on Phaseolus vulgaris flowers .Twenty-four insect species visit P. vulgaris flowers. X. olivacea was the most frequent visitor and they intensely and exclusively foraged nectar. The foraging speed was 9.94 flowers/min. The foraging ac�vity of X. olivacea resulted in a significant increment in frui�ng rate by 48.43 and 78.18%, the number of seeds/pod by 19.38 and 18.58% and the normal seeds/pod by 15.67 and 38.25%, respec�vely in 2009 and 2010. Researchers have shown no set of seeds from undisturbed bean flowers, whereas, flowers manipulated by wing petals have a great number of seeds (Darwin, 1858). Bumblebees were indicated to be the most effec�ve pollinator of beans when visi�ng blossoms to collect pollen and nectar in South Africa (Palmer, 1967).Genus Bombus was associated with pollina�on ac�vity in common beans in North America (McGregor, 1976). P. vulgaris flowers were reported to produce fewer seeds per pod in the absence of efficient pollinators in the United States of America (Ibarra-Perez et al., 1999).
Recent research conducted in Kenya has revealed Apis mellifera to be the most abundant insect species visi�ng P. vulgaris flowers, followed by X. calens (currently Xylocopa olivacea) and X. inconstans (Kasina et al., 2009). Xylocopa olivacea was the main floral visitor of P. vulgaris during the observa�on period. Elsewhere, bumblebees in South Africa (Palmer, 1967), A. mellifera in Western Kenya (Kasina et al., 2009) have respec�vely been reported as the main floral visitor of this crop. The peak ac�vity of X. olivacea on P. vulgaris flowers was located between 10.00 and 13.00 h, which correlated with the highest availability period of nectar on P. vulgaris flowers. However, this decreased ac�vity a�er 16.00 to17.00 h could be related to decreased temperature in the experimental field. Although, foragers preferred warm or sunny days for good floral ac�vity (Kasper et al., 2008), the enhanced temperature posi�vely influenced the insect ac�vity on foraged flowers. Similarly, rainfall has been documented as an environmental factor that can disrupt the floral insect ac�vity (McGregor, 1976). The abundance of X. olivacea foragers on 1000 flowers and the posi�ve and highly significant correla�on between the number of P. vulgaris flowers at bloom, as well as, the number of X. olivacea visits indicates the a�rac�veness of P. vulgaris nectar with respect to this bee. In fact, weather during bloom was demonstrated to affect the abundance and foraging of pollinator insects (Bramel et al., 2004, Julianna and Rufus, 2010). Among the 24 insect species visi�ng P. vulgaris flowers, X. olivacea was the most abundant (28.7%), followed by Coryna sp. (17.92%), Chalicodoma rufipes (15.24%) and Cera�na sp. (7.65%).
During the collec�on of nectar on each flower, X.olivacea foragers regularly come into contact with the s�gma. They were also able to carry pollen with their hairs, legs and mouth accessories from a flower of one plant to s�gma of another flower of the same plant (geitonogamy), to the same flower (autogamy) or to that of another
132

plant (xenogamy). The contribu�on of X. olivacea to P. vulgaris produc�on through its pollina�on efficiency was significantly higher than that of all insects on the exposed flowers. The weight of X. olivacea played a posi�ve role during nectar collec�on. X. olivacea shook flowers facilita�ng the libera�on of pollen by anthers for the op�mal occupa�on of the s�gma. Our results confirmed those of Mensah and Kudom (2011) who revealed that the development of fruits from L. aegyp�aca flowers that have received a single visit of X. olivacea produced a mean weight of 428.7 g that was 1.5 �mes heavier than fruits from flowers visited by A. mellifera (286.76 g). This phenomenon was also reported by Vanderborght and Rasmont (1987) for X. bariwal, an efficient P. coccineus pollinator.Higher produc�vity of pods and seeds in unlimited visits when compared with bagged flowers showed that insect visits were effec�ve in increasing cross-pollina�on It is concluded that P. vulgaris black seed outlets is a highly nectariferous bee plant that obtained benefits from the pollina�on by insects among which X. olivacea is of great importance. The comparison of pods and seeds set of unprotected flowers with that of flowers visited exclusively by X. olivacea underscores the value of this bee in increasing pods and seed yields as well as seed quality. The installa�on of X. olivacea nests at the proximity of P. vulgaris small black seed fields should be recommended for the increase of pods and seed yields of this valuable crop.
Pollinators: Mommers (1971 ) studied the pollina�on of beans grown under glass and concluded that bees do not influence development of P. vulgaris.Diwan and Salvi (1965) stated that beans in India are eagerly visited by Apis dorsata, A. florea, and Trigona spp., but the flowers are generally ignored by A. cerana. Those who have studied the problem agree that bumble bees and honey bees are the most frequent visitors to bean flowers in the United States. Darwin (1858) and Palmer (1967) indicated that bees are of benefit to beans, but the kind of bean they referred to is not clear. The data indicate that, if pollina�ng insects are of benefit, the bumble bee is the best pollinator. Honey bees are apparently next in importance. They visit the blossom for both pollen and nectar. The exact effect on the flower or value to the different cul�vars of P. vulgaris has not been well established.
Dependence of crops on bee pollina�on.The bagged and un-bagged flowers were used to assess the effects of bee pollina�on on crop yield. The bagged flowers were assumed to have had spontaneous self-pollina�on (or non-bee pollina�on) while there was a strong possibility that the un-bagged flowers were visited by bees. The number of seeds per pod of the legumes (pulses) obtained from bagged and un-bagged flowers were found to be significantly different.
However, this difference was rela�vely lower for beans (25%) and green grams (27%) compared to the other legumes, while cowpeas showed the highest difference (43%). The results show that the mean number of seeds was lower than the maximum a�ainable in the un-bagged flowers, implying that bee pollina�on of the legumes in the study area was not sufficient.
133

Soybean ,Glycine max (L.) Merr.: Soybean flowers a�ract rela�vely few bees (Blickenstaff and Huggans 1962); however, bees do visit the flowers for both pollen and nectar. The anthers dehisce before the flower opens so that the s�gma is in contact with and recep�ve to the pollen on the anthers. Pollina�on and fer�liza�on is usually accomplished before the flower opens. Beekeepers in some areas frequently report honey crops from soybeans ( Hambleton 1936, Jaycox 1970, Pelle� 1947). Beekeepers in other areas report that bees never visit the flowers, or if they do no honey is obtained.
Pollina�on Requirements: The soybean is considered to be self-fer�le and not benefited by insect pollina�on (Morse and Car�er 1937, Rubis 1970).There is no experimental evidence to support them. Tests with plants caged to exclude bees have shown no decrease in produc�on over exposed plots (Woodhouse and Taylor 1913, Milum 1940, Piper and Morse 1910). Culter (1934) obtained an es�mated 5 percent cross-pollina�on in open plots surrounded by 36-inch cheesecloth fences with an apiary nearby.
Pollinators:Woodworth (1932) indicated that thrips and honey bees might be responsible for some transfer of pollen; however, Caviness (1970) showed that thrips are uneffec�ve as pollinators of soybeans, but that honey bees were repsonsible for 7.7 percent crossing.
Suggested Reading:
Kevan, P.G. and Baker, H.G. 1983. Insects as flower visitor and pollinators. Ann. Rev. Entomol. 28: 407-453.
Pedersen, M. W., Jones, L. G., and Rogers, T. H. 1961. Producing seed of legumes. U.S. Dept. Agr. Yearbook 1961: 171-181.
134

HOST PLANT RESISTANCE TO INSECT PESTS IN CHICKPEA
MUKESH K. DHILLONDivision of Entomology, Indian Agricultural Research Ins�tute (IARI),
New Delhi 110012, India.
Chickpea (Cicer arie�num L.) is the world's third most important food legume, grown in 42 countries in South Asia, North and Central America, the Mediterranean Region, West Asia, and North and East Africa; and is the premier pulse crop in the Indian subcon�nent. In recent years, it has become an important crop in Australia, Canada, and the USA. It is valued for its nutri�ve seeds with high protein content (25.3 to 28.9%). Chickpea seeds are consumed fresh as a green vegetable, fried, roasted, and boiled as a snack food. The grain a�er dehulling is largely consumed as split seeds as 'dhal', or the split seeds are ground as flour, which is used to make bread, snacks, and sweets. The straw is used as feed for livestock. It is grown on about 10.38 million ha with a produc�on of 8.57 million tonnes worldwide (FAO, 2004). India is the largest producer as well as consumer of chickpea. In India, chickpea is grown on about 6.67 million ha with a produc�on of 5.3 million tonnes (Majumder, 2009). Chickpea yields are low (400
-1to 600 kg ha ), because of several bio�c and abio�c constraints. Among the bio�c stresses, insect pests are one of the main yield reducing factors. Nearly 60 insect species are known to feed on chickpea, of which pod borer, Helicoverpa armigera (Hubner); leaf caterpillar, Spodoptera exigua Hubner; leaf miner, Liriomyza cicerina (Rondani); aphids, Aphis craccivora Koch; black cutworm, Agro�s ipsilon (Hufnagel); and termites, Microtermes obesi (Holmgren) and Odontotermes spp. are one of the most important insect pests of chickpea in different agro-ecological regions of India. Insect pests in India cause an average of 30% loss in pulses valued at $815 million, which at �mes can be 100% (Dhaliwal and Arora, 1994). Helicoverpa armigera – the single largest yield reducing factor in food legumes, causes an es�mated loss of $328 million in chickpea alone (ICRISAT, 1992). The development of chickpea cul�vars resistant or tolerant to insects has a major poten�al for use in integrated pest management (IPM), par�cularly under subsistence farming condi�ons in developing countries.
Host plant resistance (HPR) assumes a central role, and is the backbone of pest management in order to increase produc�on and produc�vity of grain legumes, par�cularly the chickpea to meet the increasing demand of pulses in the vegetarian diets. In spite of the importance of HPR as an integral part of IPM, breeding for resistance to insects has not been as successful as breeding for disease resistance because of the rela�ve ease with which insect control is achieved through insec�cide use, slow progress in developing insect-resistant cul�vars as a result of the difficul�es
135

involved in ensuring adequate insect infesta�on for resistance screening, slow transfer of insect resistance traits due to complex and polygenic inheritance, and lack of due importance and place to breeding for insect resistance in na�onal agricultural research system. With the development of insect resistance to insec�cides, adverse effects of insec�cides on natural enemies, and public awareness of environment conserva�on, there has been a renewed interest in the development of crop cul�vars with resistance to insect pests. However, the levels of insect resistance in most of the recently released varie�es are inadequate, and therefore recent years have observed a paradigm shi� in advocacy and deployment of techniques to diversify the bases of resistance through gene pyramiding from cul�vated germplasm and the closely related wild rela�ves for developing insect-resistant chickpea genotypes. The ar�cle overviews the recent research developments on different components of host plant resistance to insect pests in chickpea over the last two decades.
Mechanisms of resistance: Knowledge of the mechanisms, nature, and inheritance of resistance is cri�cal for developing germplasm with durable and stable resistance to insects. Except H. armigera, there has not been any ac�ve research on mechanisms and breeding for resistance to insect pests in chickpea in India. All the three mechanisms viz., an�xenosis, an�biosis and tolerance have been reported against H. armigera in chickpea.
An�xenosis. Oviposi�on non-preference is one of the components of resistance to H. armigera in chickpea, for example fewer eggs were recorded on ICC 506 than on ICCC 37 and Annigeri (Cowgill and Lateef, 1996). Lateef (1985) recorded 38 eggs per 5 plants in ICC 506 compared to 64 eggs per plant on Annigeri among the early flowering genotypes. Similarly, 57 and 77 eggs per 5 plants were recorded in ICC 10619 and ICC 3137, respec�vely, among the medium-maturity genotypes. Among the late-flowering genotypes; there were 36 eggs on ICC 7320-11-1, 53 on ICC 5264-E9, and 57 on ICC 8835. Thus, oviposi�on preference is one of the important components of resistance to pod borer in chickpea.
An�biosis. An�biosis is expressed in terms of larval mortality, decreased larval and pupal weights, prolonged larval and pupal development, failure to pupate, and reduced fecundity, and egg viability. Life table analysis indicated that there was considerable varia�on for net reproduc�ve ra�o (142.1 to 268.6), mean genera�on �me (39.1 to 45.2 days), intrinsic rate of daily increase (0.12 to 0.14), finite rate of daily increase (1.13 to 1.15), and weekly mul�plica�on rate (2.57 to 3.02) on different genotypes (Cowgill and Lateef, 1996). Based on weekly mul�plica�on rate, NDG 90-27, BG 1027, and BG 267 showed greater an�biosis to the pod borer than P 256. Net reproduc�ve rate was greater on BG 1027 than on other genotypes. Pupae of H. armigera reared on ICC 506 and ICCV 7 weigh less than those reared on ICCC 37. Larvae reared on leaves or pods of ICCV 7 weighed significantly lower than those reared on ICCC 37. There are
136

considerable differences in numbers of H. armigera larvae on different genotypes. Lateef (1985) recorded 58 larvae per 5 plants on ICC 506 compared to 103 larvae on Annigeri; 99 versus 202 on ICC 10619 and ICC 3137, respec�vely; and 112 versus 147 on ICC 7320-11-1 and ICC 8835, respec�vely. Ollela et al. (2002) studied the feeding preference of the third-instar larvae of H. armigera on different plant parts of chickpea. In no-choice feeding tests, H 92-67, H 91-47 showed less leaf and flower damage than H 86-18, H 89-96, and HK 89-131. Pods of H 92-67, H 91-47, and L 550 were also less preferred than that of H 86-18. In mul�-choice tests; H 92-67, H 91-47, and C 235 were less preferred than the other genotypes tested.
Tolerance. The levels of resistance to H. armigera in the germplasm accessions are low to moderate. This has necessitated the need for selec�ng genotypes with greater ability to tolerate or recover from the pod borer damage.
Physico-chemical traits associated with resistance to insects: The physico-chemical components play an important role in host plant resistance to insect pests. Several morphological traits such as pod shape, pod wall thickness, and crop dura�on influence H. armigera damage in chickpea. Varie�es with larger leaflets are preferred by the leaf miner than those with small leaflets. The glandular trichomes in dicotyledons produce s�cky exudates that may contain chemicals ac�ng as phagos�mulants, an�feedants, and/or repellents, and disrupt development and survival of insects. Chickpea varie�es with low trichome density or those devoid of trichomes, and low acid exudates were found highly suscep�ble to aphid damage (Sharma and Crouch, 2004). Secondary plant substances and poor nutri�onal quality of the host plant are important components of resistance to insects. An�biosis to insects in general is due to secondary plant metabolites, and these chemicals inhibit feeding by the insects. Chickpea exudates have malate and oxalate as the main components, and there are characteris�c differences among genotypes depending on diurnal cycles and growth stage. High acidity in the leaf exudates of chickpea is associated with resistance to H. armigera. Malic acid acts as an�feedant to the H. armigera larvae, and the varie�es with highest amounts of malic acid in chickpea results in resistance to pod borer (Sharma, 2009). Oxalic acid inhibits the growth of H. armigera larvae when incorporated into ar�ficial diet, while malic acid shows no growth inhibi�on. Oxalic acid content in chickpea leaves has also been found to be associated with resistance to leaf miner. The chickpea flavonoids judaicin 7-O-glucoside, 2 methoxy judaicin, judaicin, and maakiain present in wild rela�ves of chickpea (Cicer bijugum and C. judaicum) have shown an�feedant ac�vity towards the larvae of H. armigera (Simmonds and Stevenson, 2001). The other important class of an�-nutri�onal factors present in chickpea is protease inhibitors, which have also shown varia�on in H. armigera gut protease inhibitory ac�vity in developing seeds of wild and cul�vated chickpea genotypes.
Iden�fica�on and u�liza�on of insect resistance sources: Several thousands of
137

germplasm collec�ons have been evaluated for resistance against insect species in chickpea, and several germplasm accessions (ICC 506EB, ICC 10667, ICC 10619, ICC 4935, ICC 10243, ICCV 95992, and ICC 10817) have been iden�fied with resistance to H. armigera. Several new sources of insect resistance have also been iden�fied and supplemented to the exis�ng resistance sources against legume pod borer, H. armigera in chickpea. The insect resistance breeding efforts in chickpea have developed many Helicoverpa-resistant varie�es such as C 235, PantG 114, Anupam, JG 74, ICCV 10, Dulia, Pusa 261, Vijay, Vishal, ICCV 7, ICCV 10 and ICCL 86103, and released for cul�va�on in India (Sharma et al., 2005). There is no published informa�on on resistance of chickpea varie�es to S. exigua. However, several progenies from interspecific hybridiza�on involving cul�vated chickpea and a wild rela�ve (C. re�culatum) have exhibited high levels of resistance to S. exigua. Two accessions of C. cuneatum (ILWC 40 and ILWC 187) and 10 accessions of C. judaicum with high levels of resistance, while 18 lines of C. judaicum, and four lines of C. pinna�fidum and C. re�culatum have been iden�fied with resistance to leaf miner in chickpea (Singh and Weigand, 1994). The germplasm lines viz., ILC 3800, ILC 5901, and ILC 7738 have been inden�fied and registered as sources of resistance to Liriomyza cicerina. Seven of these lines (FLIP 2005-1C, FLIP 2005-2C, FLIP 2005-3C, FLIP 2005-4C, FLIP 2005-5C, FLIP 2005-6C, and FLIP 2005-7C) have good agronomic background, seed size, and plant type (Malhotra et al., 2007), and have been distributed to na�onal programs for evalua�on under local condi�ons.
Gene�cs and inheritance of resistance to insect pests: Informa�on on gene�cs and inheritance of resistance to insect pests is important for crop improvement, which indicates the degree of ease or difficulty involved in incorpora�ng resistance genes into the improved cul�vars. In desi chickpea, highly significant variances have been observed for pod borer, H. armigera damage, sugges�ng the involvement of addi�ve type of gene ac�on, but there was preponderance of non-addi�ve type of gene ac�on in the kabuli types. From line x tester studies, it was concluded that resistance to H. armigera is controlled by mul�ple genes. In most studies, gene ac�on was found to be predominantly addi�ve, although non-addi�ve type of gene ac�on was also observed (Gowda et al., 2005).
Search of insect resistance genes from wild rela�ves of crops : Wild species of crops are important sources of genes for resistance to insect pests. Genotypes showing high levels of resistance to insects can be used in wide-hybridiza�on to increase the levels and diversify the basis of resistance to the target insect pests. Last two decades have observed a paradigm shi� in iden�fica�on and deployment of wild species of chickpea (Dhillon and Sharma, 2012). Wild chickpea species such as C. bijugum, C. re�culatum, C. judaicum, C. pinna�fidum, C. microphyllum, and C. cuneatum have shown high levels of resistance to H. armigera (Sharma et al., 2007). Accessions belonging to C. bijugum, C.
138

pinna�fidum, and C. echinospermum have also shown resistance to the bruchid, Callosobruchus chinensis L. (Singh et al., 1998).
Deployment of molecular markers for host plant resistance to insects: Considerable progress has been made in the recent past in iden�fying genomic regions and genes associated with resistance traits in several crops to different insect pests. Several types of molecular markers have been used to evaluate DNA polymorphism, and for developing gene�c linkage maps of different crops, to iden�fy quan�ta�ve trait loci (QTLs) associated with resistance to insects. Mapping popula�ons from interspecific crosses are o�en used for gene�c linkage studies due to high level of detectable polymorphism, but linkage maps derived from such crosses may have limited relevance in crop breeding programs due to different recombina�on pa�erns. However, markers developed from such maps may be valuable tools for introgression of genes of interest from the wild rela�ves into the cul�gen. Furthermore, marker assisted selec�on (MAS) has shown the poten�al to drama�cally speed up the process by reducing the number of genera�ons and the size of the popula�ons required to iden�fy individuals with appropriate combina�on of genes, with minimal amount of linkage drag from the wild rela�ves.
Many studies have used interspecific mapping popula�ons for developing gene�c linkage maps of chickpea. The preliminary linkage map was based on interspecific crosses of C. arie�num × C. re�culatum and C. echinospermum, and intraspecific crosses of C. re�culatum (Gaur and Slinkard, 1990a,b). Winter et al. (1999) developed the first genomic map of chickpea based on 90 RILs derived from a cross of C. re�culatum (PI 489777) and the cul�vated chickpea, ICC 4958, using 120 STMS markers. This map was then augmented using 118 STMS, 96 DAFs, 70 AFLP, 37 ISSR, 17 RAPD, 2 SCAR, 3 cDNA, and 8 isozyme markers screened across 130 RIL from the same cross (Winter et al., 2000). Mapping complex traits such as resistance to pod borer, H. armigera in chickpea is only just beginning, and the efforts are underway to evaluate inter-specific mapping popula�ons to iden�fy QTLs linked to various components of resistance to H. armigera in chickpea (Sharma et al., 2005b, Sharma, 2009).A mapping popula�on of 126 F RILs of ICCV 2 x JG 62, has been evaluated for resistance to H. 13
armigera. The overall resistance score (1 = <10 leaf area and/or pods damaged, and 9 = >80% leaf area and/or pods damaged) varied from 1.7 to 6.0 in the RIL popula�on compared to 1.7 in the resistant check, ICC 506EB, and 5.0 in the suscep�ble check, ICCV 96029. There were 4 to 31 larvae per 10 plants in the mapping popula�on compared to 10 larvae on ICC 506EB and 18 on ICCV 96029. These results indicated that there is considerable varia�on in this mapping popula�on for suscep�bility to H. armigera. Another RIL mapping popula�on from the cross Vijay (suscep�ble) × ICC 506EB (resistant) has been evaluated for resistance to H. armigera. Efforts are also underway to evaluate interspecific mapping popula�ons based on the crosses ICC 3137
139

(C. arie�num) x IG 72933 (C. re�culatum) and ICC 3137 × IG 72953 (C. re�culatum) for resistance to pod borer to iden�fy QTLs linked to various components of resistance to H. armigera.
Suggested Reading
Cowgill, S.E. and Lateef, S.S. 1996. Iden�fica�on of an�bio�c and an�xeno�c resistance to Helicoverpa armigera (Lepidoptera: Noctuidae) in chickpea. Journal of Economic Entomology 89: 224–229.
Dhaliwal, G.S. and Arora, R. 1994. Trends in Agricultural Insect Pest Management. Commonwealth Publishers, New Delhi, India.
Dhillon, M.K. and Sharma, H.C. 2012. Paradigm shi�s in research on host plant resistance to insect pests. Indian Journal of Plant Protec�on 40(1): 1-11.
Food and Agriculture Organiza�on (FAO). 2004. Produc�on Sta�s�cs. Food and Agriculture Organiza�on, Rome, Italy.
Gaur, P.M. and Slinkard, A.E. 1990a. Inheritance and linkage of isozyme coding genes in chickpea. Journal of Heredity 81: 455-461.
Gaur, P.M. and Slinkard, A.E. 1990b. Gene�c control and linkage rela�ons of addi�onal isozyme markers in chickpea. Theore�cal and Applied Gene�cs 80: 648-656.
Gowda, C.L.L., Ramesh, S. Chandra, S. and Upadhyaya, H.D. 2005. Gene�c basis of pod borer (Helicoverpa armigera) resistance and grain yield in desi and Kabuli chickpea (Cicer arie�num L.) under unprotected condi�ons. Euphy�ca 145: 199-214.
ICRISAT. 1992. The Medium Term Plan. Interna�onal Crops Research Ins�tute for the Semi-Arid Tropics (ICRISAT), Patancheru 502 324, Andhra Pradesh, India.
Lateef, S.S. 1985. Gram pod borer (Heliothis armigera (Hub) resistance in chickpea. Agriculture, Ecosystem, and Environment 14: 95–102.
Majumder, N.D. 2009. All India coordinated research project on chickpea: a profile. Indian Ins�tute of Pulses Research, Kanpur, U�ar Pradesh, India.
Malhotra, R.S., Singh, M. and Erskine, W. 2007. Genotype × environment interac�on and iden�fica�on of dual-season cul�vars in chickpea. Euphy�ca 158: 119–127.
Ollela, G.S., Saini, R.K. and Dahiya, B. 2002. Evalua�on of some promising chickpea genotypes for their feeding preference by the third-instar larvae of Helicoverpa armigera (Hubner). Indian Journal of Pulses Research 15: 83-85.
Sharma, H.C., Gowda, C.L.L., Stevenson, P.C., Ridsdill-Smith, T.J., Clement, S.L., Ranga Rao, G.V., Romies, J., Miles, M. and Bouhssini, M. 2007. Host plant resistance and insect pest management in chickpea. In: Chickpea Breeding and Management
140

(Yadav, S.S., Redden, R.J., Chen, W. and Sharma, B., eds). CAB Interna�onal, Oxfordshire, UK, pp. 520-537.
Sharma, H.C. 2009. Gene�c engineering of natural enemies for integrated pest management. In: Biotechnological Approaches for Pest Management and Ecological Sustainability. CRC Press, Tailor & Francis Group, Boca Raton, FL, USA, pp. 293-315.
Sharma, H.C., Ahmad, R., Ujagir, R., Yadav, R.P., Singh, R. and Ridsdill-Smith, T.J. 2005a. Host plant resistance to co�on bollworm/legume pod borer, Helicoverpa armigera. In: Strategies for Heliothis/Helicoverpa Management: Emerging Trends and Strategies for Future Research (Sharma, H.C., ed.). Oxford and IBH, New Delhi, India, pp 167-208.
Sharma, H.C., Gaur, P.M. and Hoisington, D.A. 2005b. Physico-chemical and molecular markers for host plant resistance to Helicoverpa armigera. In: Recent Advances in Helicoverpa Management (Saxena, H., Rai, A.B., Ahmad, R. and Gupta, S., eds.). Indian Society of Pulses Research and Development, Indian Ins�tute for Pulses Research, Kanpur, U�ar Pradesh, India, pp. 84-121.
Sharma, H.C., Sharma, K.K, and Crouch, J.H. 2004. Gene�c transforma�on of crops for insect resistance: poten�al and limita�ons. Cri�cal Reviews in Plant Sciences 23: 47-72.
Simmonds, M.S.J. and Stevenson, P.C. 2001. Effects of isoflavonoids from Cicer on larvae of Helicoverpa armigera. Journal of Chemical Ecology 27: 965-977.
Singh, K.B., and Weigand, S. 1996. Registra�on of three leafminer-resistant chickpea lines:ILC 3800, ILC 5901 and ILC 7738. Crop Science 36: 472.
Singh, K.B., Ocampo, B. and Robertson, L.D. 1998. Diversity for abio�c and bio�c stress resistance in the wild annual Cicer species. Gene�c Resources and Crop Evolu�on 45: 9-17.
Winter, P., Benko-Iseppon, A.M., Hu�el, B., Ratnaparkhe, M., Tullu, A., Sonnante, G., Pfaff, T., Tekeoglu, M., Santra, D., Sant, V.J., Rajesh, P.N., Kahl, G. and Muehlbauer, F.J. 2000. A linkage map of chickpea (Cicer arie�num L.) genome based on recombinant inbred lines from a C. arie�num x C. re�culatum cross: localiza�on of resistance genes for fusarium wilt races 4 and 5. Theore�cal and Applied Gene�cs 101: 1155-1163.
Winter, P., Pfaff, T., Udupa, S.M., Hu�el, B., Sharma, P.C., Sahi, S., Arreguin-Espinoza, R., Weigand, F., Muehlbauer, F.J. and Kahl, G. 1999. Characteriza�on and mapping of sequence-tagged microsatellite sites in the chickpea (Cicer arie�num L.). Genome 262: 90-101.
141

HOST PLANT RESISTANCE TO INSECT PESTS IN PIGEONPEA
MUKESH K. DHILLONDivision of Entomology, Indian Agricultural Research Ins�tute (IARI),
New Delhi 110012, India.
Introduc�on
Pigeonpea [Cajanus cajan (L.) Millsp.] is grown in about 50 countries in Asia, Africa, and the Caribbean for various uses including food, fodder, firewood, and building material. India accounts for 85% of the world pigeonpea produc�on, and more than 4.0 million ha are under pigeonpea cul�va�on. Despite appreciable technological advances, major breakthrough in pigeonpea produc�vity is s�ll elusive, and the gap between poten�al and actual yields of pigeonpea is one-third or even less of the poten�al yields. While low inputs and sub-op�mal crop management accounts for a significant share of this gap, the losses due to insect pests result in harvested yields much lower than that is possible with resistant/tolerant varie�es. Although, more than 250 insect species have been recorded feeding on pigeonpea, however, only a few of them only cause significant and consistent damage (Sachan et al., 1994). Of these, the legume pod borers, Helicoverpa armigera (Hubner) and Maruca vitrata (Geyer), pod fly, Melanagromyza obtusa Malloch., pod bug, Clavigralla tomentosicollis Stal., and blister beetle, Mylabris spp. are one of the most important constraints to increase the produc�on and produc�vity of pigeonpea in India. To meet the future demands for pulses, strategies need to be developed for solving the pressing problem of management of these insect pests involving iden�fica�on of stable sources of resistance and introgression of the resistance genes into high yielding cul�vars using conven�onal and biotechnological approaches.
Screening of cul�vated germplasm in pigeonpea has revealed low to moderate levels of resistance to insect pests, however, several accessions of wild rela�ves have shown high levels of resistance due to diverse mechanisms of resistance to insect pests (Sharma et al., 2008, 2009). Development and release of pigeonpea cul�vars with tolerance/resistance can play a major role in minimizing the extent of losses due to insect pests without addi�onal monetary investment by the farmers. Also, there has been very limited success in deployment of molecular tools for resistance to insect pests in pigeonpea. Therefore, it is important to develop improved cul�vars of pigeonpea with stable resistance to insect pests through introgression of genes from wild rela�ves into insect-resistant tolerant/resistant cul�vars to make host plant resistance an effec�ve weapon for the management of insect pests.
142

Physico-chemical traits associated with resistance to insects: Mechanisms and inheritance of resistance is cri�cal for developing genotypes with durable and stable resistance to insect pests in pigeonpea. All the three mechanisms viz., an�xenosis, an�biosis and tolerance have been reported against insect pests in pigeonpea. Pigeonpea genotypes with determinate growth habit, clustered pods, and dense plant canopy are more suscep�ble to pod borers, H. armigera and M. vitrata than genotypes with non-clustered pods, while the genotypes with smaller pods, pod wall �ghtly fi�ng to the seeds, and a deep constric�on between the seeds are less suscep�ble to H. armigera (Sharma, 2009). Several morphological features such as pod wall thickness, differences in the structure of pod �ssue and the presence of different types of trichomes on the pod surface have been reported to confer resistance to H. armigera. Shanower et al. (1997) observed five types of trichomes viz., Type A, Type B, Type C, Type D and Type E on pods of Cajanus species and reported their importance in mechanism of resistance against H. armigera. The phagos�mulant / an�feedant ac�vity of glandular trichomal secre�ons towards H. armigera larvae has been reported (Green et al., 2003). Wild rela�ves of pigeonpea such as C. scarabaeoides and C. acu�folius with non-glandular trichomes are not preferred by H. armigera females for egg laying. Trichomes act as a defense mechanism of plants to insects, although in some instances, they also provide a suitable substrate for egg laying, a�achment, or interfere with the ac�vity of natural enemies. Glandular trichomes in pigeonpea are linked to suscep�bility to H. armigera, and influence the feeding by larvae of spo�ed pod borer, M. vitrata.
Pigeonpea is known to contain some an�-nutri�onal factors such as proteinase inhibitors, oligosaccharides, phenols, isoflavonoids, tannins and phy�c acid. The late maturing cul�vars of pigeonpea resistant to pod borer damage have higher content of polyphenols and lower amino acids, sugars and proteins compared to the suscep�ble medium and early maturity varie�es. Several chemicals on the pod surface of pigeonpea, that are absent from the pods of wild rela�ves, influence host plant selec�on for feeding by the larvae of H. armigera. Acetone extracts of pods of pigeonpea and its wild rela�ve, C. platycarpus have a significant feeding s�mulant effect on H. armigera larvae, whereas the extracts from C. scarabaeoides pods have no such effect (Shanower et al., 1997). Querce�n, quercetrin, and guerce�n-3-methyl ether (present in pod surface exudates) play an important role in food selec�on behavior of H. armigera larvae, while S�lbene - a phytoalexin, occurs at high concentra�ons in pigeonpea cul�vars with resistance to H. armigera (Green et al., 2002, 2003).
Iden�fica�on and u�liza�on of insect resistance sources: Resistance to pod borers has been reported by several scien�sts, but the available sources of resistance have not been ac�vely exploited in breeding programs. The levels of resistance to pod borers are
143

low to moderate, but are quite effec�ve when deployed in combina�on with synthe�c pes�cides or natural plant products. More than 14,000 pigeonpea accessions have been screened for resistance to different insect pests at ICRISAT and in the na�onal programs (Lateef and Pimbert, 1990; Shanower et al., 1998; Sharma, 2009). Based on extensive tes�ng of pigeonpea lines several pigeonpea genotypes such as PPE 45-2, BDN 2, ICPL 4, Bori, T 21; ICPL 269 and ICPL 88039 - early maturity; ICPL 332, ICPL 84060, LRG 41, and ICPL 187-1 - medium maturity;, and ICP 7035 medium-long maturity and vegetable type, have been found to be resistant/tolerant to H. armigera (Sharma, 2009; Srivastava and Joshi, 2011). Of these, ICPL 88039 has been extensively tested in the IndoGange�c plains in North India and has been found to be suitable for rice-wheat cropping system. ICPL 332WR has been found to be promising in Andhra Pradesh, while ICP 7035 is preferred by the farmers as a vegetable type. Pigeonpea genotypes GP 75, GP 118, GP 233, and GP 253 represen�ng different plant types and maturity groups were found to be resistant to M. obtusa, sugges�ng that resistance to pod fly is not linked to growth type and maturity period of the genotype (Moudgal et al., 2008). LRG 41 has been developed by ANGARU, and has been released for cul�va�on in Andhra Pradesh. The pigeonpea genotypes viz., ICP 10531E1, ICP 7941E1, ICP 7946E1, ICP 71765, PDA 88-2E and PDA 89-2E have also been reviewed to be resistant to pod fly, M. obtusa (Srivastava and Joshi, 2011).
Gene�cs and inheritance of resistance to insect pests : Combining ability studies have indicated the preponderance of non-addi�ve type of gene ac�on for resistance to H. armigera and M. vitrata (Lal et al., 1989). Verulkar et al. (1997) indicated the involvement of a single dominant gene in an�xenosis mechanism of resistance in C. scarabaeoides to H. armigera and M. obtusa. Non-glandular trichomes, which are associated with resistance to H. armigera in C. scarabaeoides, are inherited as a dominant trait (Rupakala et al., 2005).
Search of insect resistance genes from wild rela�ves of crops: Wild species of pigeonpea are important sources of genes for resistance to insect pests. Genotypes showing high levels of resistance to insects can be used in wide-hybridiza�on to increase the levels and diversify the basis of resistance to the target insect pests. Last two decades have observed a paradigm shi� in iden�fica�on and deployment of wild species of pigeonpea (Dhillon and Sharma, 2012). Cul�vars with mul�ple-resistances to insects will be in greater demand in future because of the concerns associated with environmental pollu�on due to synthe�c pes�cides, and the changes in rela�ve importance and severity of damage due to climate change. However, lack of precision in evalua�ng germplasm accessions and breeding lines for resistance to insect pests has resulted in missing many poten�ally good sources of resistance as severity of of damage is quite o�en variable across seasons and loca�ons. Wild species of Cajanus have been iden�fied as poten�ally valuable source of germplasm for improving the levels of
144

resistance in pigeonpea against insect pests (Pundir and Singh, 1987; Sharma et al., 2001). Wild rela�ves of pigeonpea such as C. scarabaeoides, C. acu�folius, C. platycarpus, and C. sericeus have been found to be highly resistant to H. armigera and M. obtusa, which can be easily crossed with the cul�vated pigeonpea (Sharma et al., 2005; Sharma, 2009). The wild accessions of pigeonpea ICPW 141, ICPW 278, and ICPW 280 (C. scarabaeoides), ICPW 214 (C. bracteata), ICPW 14 (C. albicans), and ICPW 202 (F. stricta) have also been reported to have resistance to pod fly (Sharma et al., 2003). With the use of interspecific hybridiza�on, it would be possible to transfer genes with diverse mechanisms of resistance to insect pests from the wild rela�ves to the cul�gen.
Deployment of molecular markers for host plant resistance to insects: A few studies have been conducted to inves�gate polymorphism and its wild rela�ves in pigeonpea (Sharma et al., 2005). Low levels of gene�c diversity were observed in the cul�vated pigeonpea using DArTs and oly 64 markers were found polymorphic among the cul�vated pigeonpea (Yang et al., 2006). The DArT markers also revealed gene�c rela�onships among the accessions of different species consistent with the available informa�on and systema�c classifica�on. Panguluri et al. (2006) used AFLP markers to detect polymorphism in cul�vated pigeonpea and two of its wild rela�ves C. volubilis and Rhynchosia bracteata. The two wild species shared only 7.15% of the bands with the pigeonpea, whereas 86.71% common bands were observed among the pigeonpea cul�vars. Similarly, 62.08% bands were polymorphic between C. volubilis and pigeonpea in comparison to 63.33% of polymorphic bands between R. bracteata and pigeonpea, and 13.28% polymorphic bands among pigeonpea cul�vars. Some efforts have also been made to derive interspecific deriva�ves from the secondary and ter�ary gene pools, however, there have not been any defini�ve efforts to iden�fy QTLs associated and marker-aided selec�on for resistance to insect pests in pigeonpea (Sharma, 2009). A mapping popula�on based on C. cajan x C. scarabaeoides has also been developed, and is under evalua�on for resistance to H. armigera to iden�fy QTLs linked for resistance to pod borer in pigeonpea.
Suggested Reading
Dhillon, M.K. and Sharma, H.C. 2012. Paradigm shi�s in research on host plant resistance to insect pests. Indian Journal of Plant Protec�on 40(1): 1-11.
Green, P.W.C., Stevenson, P.C., Simmonds, M.S.J. and Sharma, H.C. 2002. Can larvae of the pod-borer, Helicoverpa armigera (Lepidoptera: Noctuidae), select between wild and cul�vated pigeonpea [Cajanus sp. (Fabaceae)]?, Bulle�n of Entomological Research 92: 45-51.
Green, P.W.C., Stevenson, P.C., Simmonds, M.S.J. and Sharma, H.C. 2003. Phenolic compounds on the pod-surface of pigeonpea, Cajanus cajan, mediate feeding behavior of Helicoverpa armigera larvae. Journal of Chemical Ecology 29: 811-821.
145

Lal, S.S., Singh, I.P. and Jeet, V. 1989. Heterosis and combining ability for resistance to pod fly and lepidopteran borers in pigeonpea (Cajanus cajan). Indian Journal of Agricultural Sciences 69(11): 786–788.
Lateef, S.S. and Pimbert, M.P. 1990. The search for host plant resistance of Helicoverpa armigera in chickpea and pigeonpea at ICRISAT. Pages 14-18 in Proceedings of the Consulta�ve Group Mee�ng on the Host Selec�on Behavior of Helicoverpa armigera, 5-7 March 1990. Interna�onal Crops Research Ins�tute for the Semi-Arid Tropics, Patancheru, Andhra Pradesh, India.
Moudgal, R.K., Lakra, R.K., Dahiya, B. and Dhillon, M.K. 2008. Physico-chemical traits of Cajanus cajan (L.) Millsp. pod wall affec�ng Melanagromyza obtusa (Malloch) damage. Euphy�ca 161(3): 429-436.
Pundir, R.P.S. and Singh, R.B. 1987. Possibility of gene�c improvement pigeonpea (Cajanus cajan) u�lizing wild gene sources. Euphy�ca 36: 33-37.
Rupakala, A., Rao, D., Reddy, L.J., Upadhyaya, H.D. and Sharma, H.C. 2005. Inheritance of trichomes and resistance to pod borer (Helicoverpa armigera) and their associa�on in interspecific crosses between cul�vated pigeonpea (Cajanus cajan) and its wild rela�ve C. scarabaeoides. Euphy�ca 145: 247-257.
Sachan, J.N., Yadava, C.P., Ahmad, R. and Ka�, G. 1994. Insect pest management in pulse crops. In: Trends in Agricultural Insect Pest Management (Dhaliwal G.S. and Arora, R., eds.). Commonwealth Publishers, New Delhi, pp 308-344.
Shanower, T.G., Lal, S.S. and Bhagwat, V.R. 1998. Biology and management of Melanagromyza obtusa (Malloch) (Diptera: Agromyzidae). Crop Protec�on 17(3): 249-263.
Shanower, T.G., Yoshida, M. and Peter, A.J. 1997. Survival, growth, fecundity and behavior of Helicoverpa armigera (Lepidoptera: Noctuidae) on pigeonpea and two wild Cajanus species. Journal of Economic Entomology 90: 837-841.
Sharma, H.C. 2009. Biotechnological approaches for pest management and ecological sustainability. CRC Press, Taylor and Francis Group LLC. Pp. 526.
Sharma, H.C., Ahmad, R., Ujagir, R., Yadav, R.P., Singh, R. and Ridsdill-Smith, T.J. 2005. Host plant resistance to co�on bollworm/legume pod borer, Helicoverpa armigera. In: Heliothis / Helicoverpa Management: Emerging Trends and Strategies for Future Research (Sharma, H.C., ed.). Oxford and IBH, New Delhi, India, pp. 167-208.
Sharma, H.C., Green, P.W.C., Stevenson, P.C. and Simmonds, M.S.J. 2001. What makes it tasty for the pest? Iden�fica�on of Helicoverpa armigera (Hübner) feeding s�mulants and loca�on of their produc�on on the pod-surface of pigeonpea [Cajanus cajan (L.) Millsp.]. Compe��ve Research Facility Project R7029 C, Final Technical Report, Department for Interna�onal Development, London, UK.
146

Sharma, H.C., Pampapathy, G. and Reddy, L.J. 2003. Wild rela�ves of pigeonpea as a source of resistance to the pod fly (Melanagromyza obtusa Malloch) and pod wasp (Tanaos�gmodes cajaninae La Salle). Gene�c Resources and Crop Evolu�on 50: 817-824.
Srivastava, C.P. and Joshi, N. 2011. Insect pest management in pigeonpea in Indian scenario: A cri�cal review. Indian Journal of Entomology 73(1): 63-75.
Verulkar, S.B., Singh, D.P. and Bha�acharya, A.K. 1997. Inheritance of resistance to pod fly and pod borer in the interspecific cross of pigeonpea. Theore�cal and Applied Gene�cs 95: 506–508.
Yang, S., Pang, W., Harper, J., Carling, J., Wenzl, P., Hu�ner, E., Zong, X. and Kilian, A. 2006. Low level of gene�c diversity in cul�vated pigeonpea compared to its wild rela�ves is revealed by diversity arrays technology (DArT). Theore�cal and Applied Gene�cs 113: 585-595.
Panguluri, S., Janaiah, K., Govil, J., Kumar, P. and Sharma, P. 2006. AFLP fingerprin�ng in pigeonpea (Cajanus cajan (L.) Millsp.) and its wild rela�ves. Gene�c Resources and Crop Evolu�on 53: 523-531.
147

ECOLOGICAL FACTORS AFFECTING POPULATION OF IMPORTANT INSECT PESTS OF LEGUME CROPS
G.S. YADAVDepartment of Entomology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
Chickpea, pigeonpea, urdbean, moongbean, mothbean, cowpea, len�l and pea are the commonly grown pulses. Of the several insect pests associated with these crops, Helicoverpa armigera (Hubner), Melanagromyza obtusa (Malloch), Spilarc�a obliqua Walker, Amsacta moorei (Butler), Ophiomya phaseoli (Tryon), E�ella zinckenella (Treitschke) and Lampides boe�cus (Linnaeus) are of much importance. There is approximately 15 to 20 per cent loss in produce amoun�ng to 2 to 2.5 million tonnes, due to the pest complex. Of the various insect pests, Helicoverpa. armigera and M. obtusa are the most damaging, hence much studied. However, rela�ve importance of these insect pests differs from region to region as influenced by weather condi�ons, soil type etc. Chickpea is generally grown under rainfed condi�ons but some part of the area is under irrigated and late sown condi�ons. In Haryana, pod borer appears in
ndthreshold level a�er 2 week of March and by that �me the chickpea is almost at maturity. The per cent pod damage inflicted by pod borer varies from 0-17 per cent at farmer's field in contrast to heavy damage incurred in irrigated crops with an average of 30-40 per cent in chickpea.
Possible ecological management of insect pests: Farmers generally pay less a�en�on with respect to pest control in pulses as compared to other crops. Pulses are grown on marginal lands and under low-input condi�ons. The survey conducted at Kanpur and also at Haryana indicated that 5 per cent or even less farmers go for plant protec�on. Therefore, self perpetua�ng and low cost technology is desired. So management of the pest species should be based on ac�ons favouring mankind and nature in pest popula�on and basic ecological studies on the concerned pests are essen�al.Kaushik and Naresh (1984) studied popula�on dynamics, distribu�on pa�ern and damage of H. armigera on chickpea. The larval popula�on was low �ll 8 March, slightly increased by
215 March and increased rapidly in subsequent week, a�aining peak (19.02 larvae/m ) in st 2
the 1 week of April. The popula�on declined to 12.12 l/m in the 2nd week of April, when the crop was at maturity and some of the larvae had pupated. Frequent rainfall during the crop season ini�ated moth emergence, ma�ng, oviposi�on and larval hatch.
thPopula�on increased from 16 March onwards because of favourable weather and nd
good growth of the crop. The average minimum temperature from 2 March onwards orose above 10 C when the crop was at the pod stage with developing grains. There was
highly significant correla�on between the minimum temperatures and larval popula�on.
148

� Yadava et al. (1991) studied popula�on abundance of pod borer complex on chickpea at Kanpur for 7 years. It was recorded that pod borer complex consis�ng of H. armigera and Autographa nigrisigna (Walker) on chickpea cv. K850 varied during different stages of crop growth. Larval popula�on of H. armigera and A. nigrisigna occurring in a ra�o of 62.5 and 34.3 per cent, respec�vely. Both H. armigera and A. nigrisigna appeared from standard week 47 (middle of November). Abundance of H. armigera was high (83.3%) during standard week of 47, declined therea�er, reaching the lowest level (21.7%) by standard week 4 (end of Jan.) and increasing again. By standard week 16 (middle of April) there were only larvae of H. armigera. Meaning thereby that H. armigera was abundant during 47-51 and 10-17 standard weeks. Abundance of A. nigrisigna was just reverse to that of H. armigera. With the growth of chickpea plants, abundance of A. nigrisigna started to increase from 14.5 per cent in standard week 47 to 74 per cent by standard week 4 under Haryana condi�ons. Chauhan and Dahiya (1991) reported maximum incidence of H. armigera on 'L144',
ndL550, H75-35 and HMS30 during 2 fortnight of March as recorded in Punjab. Regarding the influence of abio�c factors, correla�on coefficient (R) was posi�vely significant with maximum and minimum temperatures and nega�vely significant with rela�ve humidity in H. armigera. The R values for maximum and minimum temperature showed nega�ve rela�onship and posi�ve with rela�ve humidity against A. nigrisigna.
2The coefficient of determina�on (R ) revealed that all the 3 factors i.e. maximum, minimum and rela�ve humidity contributed 55, 49 and 50 per cent, respec�vely of total larval varia�on of H. armigera and 49, 36 and 43 per cent in A. nigrisigna.
� Studies of popula�on ecology and its parasites on pulses were conducted by Bilapate et al. (1988). Campole�s chlorideae Uchida, Carcelia sp., Eribours argenteopilosus Cameron and Goniophthalmus halli Mesnil were iden�fied as mortality factors for Heliothis on arhar. Parasi�za�on of early instar (I-III) larvae owing to C. chlorideae was 1.38 per cent. Carcelia sp. parasi�zed the late instar larvae (IV-VI)
st nd rdand the parasi�za�on was 1.95, 1.08 and 2.89 per cent in the 1 , 2 and 3 genera�on, st nd rdrespec�vely. The parasi�za�on of G. halli in 1 , 2 and 3 genera�on was 5.40, 3.52 and
7.88 per cent, respec�vely. The parasite C. chlorideae and E. argenteopilosus were key mortality factors for H. armigera on gram. The pest completed 3 genera�ons on gram.
stC. chlorideae existed only in the 1 genera�on of Heliothis on gram and the parasi�za�on was to the extent of 14.73 per cent. E. argenteopilosus parasi�zed the
st nd rdlarvae to the extent of 1.53, 16.41 and 10.44 per cent during 1 , 2 and 3 genera�ons of the pest. Mortality due to NPV was 6.86, 1.98 and 24.52 per cent, respec�vely. The
stnega�ve trend index value (1=0.23) in 1 genera�on indicated that mortality factors opera�ng during this genera�on were effec�ve in reducing the popula�on of the pest
nd nd(N1) in 2 genera�on while the posi�ve trend index value (1=1.33) in 2 genera�on
rdrevealed the increase in popula�on in the 3 genera�on. Larval popula�on was at its st
peak during 1 genera�on was slowly decreased therea�er, on arhar. Mortality of
149

larvae and pupae because of bio�c factor was compara�vely more in the last genera�on as compared to that of earlier two genera�ons.
� Yadav et al. (1982) reported that the parasite C. chlorideae began its ac�vity ndtowards the 2 week of September on tomato and this synchronized with ac�vity of H.
armigera on tomato. Op�mum parasi�sm was recorded from December to February. The parasi�za�on of H. armigera by C. chloridae was to the extent of 34.5 per cent on gram (Gangrade, 1964). Studies conducted at Hisar indicated the ac�vity of C. chlorideae on early instars of H. armigera during January – February but the parasi�za�on was of low levels. Sahoo and Behera (2001) reported a posi�ve correla�on between temperature and larval popula�on build up of Maruca vitrata on UPAS-120 var. of pigeonpea. RH had a nega�ve effect on the popula�on of the pod borers except Maruca. Rainfall showed a non-significant posi�ve effec�ve on the larval incidence on these pests.
Shan�bala and co-workers (2004) reported a varying response of blue bu�erfly, Lampides boe�cus (L.) on pigeonpea by the abio�c factors of environment in different cropping seasons. The peak popula�on was a�ained during first week of January, 2000 (10.86 and 13.61%) on flowers and buds, respec�vely. Yadav and Associates (2011) studied seasonal ac�vity of podfly, Melanagromyza obtusa (Malloch.) on pigeonpea and observed the first appearance of maggots in the first week of October (90-100 days
th stold crop) and peaked upto 47 week i.e. 1 week of November. The maggot popula�on o
started building up when the maximum temperature dropped below 32 C and a�ained peak when declined.�
Pest monitoring through light traps and pheromone traps: Keeping a vigil on the popula�on fluctua�ons through monitoring of the pest is one of the important components of prac�cal pest management.
� Light traps: Use of light traps is the primary method of monitoring adults of H. armigera.. The studies revealed that female moths were ac�ve during pre-mid night period while males were more ac�ve during post mid-night period. Maximum catches were recorded between 2.0-5.0 h.
� Pheromone traps: Use of synthe�c pheromone traps made monitoring easy. The pheromone used was a mixture of (Z)-II Hexadecinal and (Z)-9 Hexadecinal in a 97:3 ra�os. Eight years studies on synthe�c pheromones at different loca�ons revealed that in general peak catches were recorded around mid-April at most northern loca�ons. The catches during these peaks o�en averaged over 100 moths per trap/night. Local pest popula�ons were considered to have bred mainly on chickpea. At most of these loca�ons, a small peak in catches was recorded around the late October. These moths were thought to have bred on short dura�on pigeonpea, co�on and wild hosts. The cold winter limits H. armigera ac�vity at northern loca�ons from November to February.
150

oThe night temperature drops below 10 C quite o�en which is detrimental to larval development. Vaishampayan and Veda (1980) reported that a minimum daily
o otemperature of 10 to 14 C was op�mum for the ac�vity of H. armigera and that a
otemperature below 8 C sharply suppressed larval ac�vity. In loca�ons like Solan, Hisar, Ludhiana and Pantnagar popula�on development is prolonged by the low temperatures prevailing during the winter. In most of the southern loca�ons like Coimbatore, the pheromone trap catches were generally lower than those recorded at northern loca�ons and without well defined peaks.
nd� In Punjab, there are two periods of peak ac�vity i.e. during 2 half of March and st1 half of April and another during October, coinciding with reproduc�ve growth stage
of chickpea and pigeonpea, respec�vely. Under Hisar condi�ons, trap catches of pod borer were maximum during March-April and again in October-November indica�ng the peak ac�vity period corresponding to reproduc�ve stage of chickpea and pigeonpea. The lowest ac�vity was observed during December-January. Moths catches were low upto the end of January. From beginning of February, increasing trend of moth emergence was observed. Maximum of 71.21 and 58.07 moths/ trap/ night were
th th th thtrapped between 11 March to 18 March and 18 March to 24 March, respec�vely. Increase in moth emergence was followed by increase in larval popula�on in the field. Adult popula�on started declining gradually in the month of April and remained very
stlow �ll 1 week of October. There was abrupt increase in the number of adults emerged nd rd
in the 2 week of October and were maximum in the 3 week of October. A�er November, there was decline. So moth catches were good indicator of seasonal ac�vity of pod borer as the damage to pigeonpea crop occurred during October-November. Generally meanagement is required at pod ini�a�on stage in Haryana, 7-10 days following increase in trap catches.
Economic threshold and economic injury levels of pod borer
Chickpea Economic injury level: 4 l/sq.m. (Odak and Thakur, 1975) Economic threshold level: 0.524 l/metre row length (mrl) per 10 plants (Singh, 1980) One l/mrl (Chaudhary & Sharma, 1982) One l /5 plants (DPR, Kanpur) 1.75 l/mrl (Sharma, 1985)
Pigeonpea 10.36 per cent bored pods in pigeonpea variety ICPL 270 is conomic injury level One l /20 plants (Venugopal et al., 1990) One l/ 1-2 plants or 5-10 per cent pod damage (Sachan et al. 1990)
Ecological methods for other insect-pests: Collec�on and destruc�on of egg masses and young larvae of hairy caterpillars. Early sowing of moongbean in the month of July
151

results in lesser insect pests on moongbean. Yellow mosaic resistant variety of moongbean, 'Asha' (MH83-20) in Haryana and ML267, ML5 and ML613 in Punjab are recommended for cul�va�on. In pea early sowing is avoided to reduce stem fly a�ack. A leafless variety of pea, 'Aparna' (HFP4) with low leaf miner incidence is recommended.
Suggested Reading:
Alamzeb, Kha�ak, S.U., Abid, F. and Khan, A.U. 2002. Evalau�on of pheromone traps at different heights for management of chickpea pod borer (H. armigera). Proc. Pakistan Cong. Zool. 22: 79-83.
Bilapate, G.G.; Mokat, R.B.; Lovekar, R.C. and D.N. Bagade, 1988. Popula�on Ecology of Heliothis armigera (Hub) and its parasites on Pulses. J. Maharashtrra agric. Univ. 13(3): 299-302.
Chauhan,R. 2007. Popula�on dynamics and ecological management of insect-pests of pulse crops. Lecture in compendium “Applica�on of ecological methods for management of crop pests” by J.P. Bhanot, R.K. Saini and G.S. Yadav. pp. 149-156.
Chauhan, R.; Dahiya, B. and K.S. Khokhar, 1991. Weeding Vicia sa�va L. to help check pod borer, Helicoverpa armigera Hb. damage in chickpea, Cicer arie�num L. ACIAR Food Legume Newsle�er No.15.
Sahoo, B.K. and Behera, U.K. 2001. Influence of abio�c factors on the incidence of pigeonpea pod borer in coastal half of Orissa. Environ. & Ecology 19(4): 882-84.
Shan�bala, T.; Subharani, S. and Singh, T.K. 2004. Seasonal incidence of Lampidas boc�cus (Linn.) and influence of weather factors on its abundance on pigeonpea, Cajanus cajan (L.) Millsp. in Manipur. Indian J. Ent. 66(3): 198-201.
Yadav, S.K.; Ahuja, D.B. and Dhandapani, A. 2011. Seasonal ac�vity of pod fly, Melanagromyza obtusa (Malloch.) (Diptera: Agromyzidae) and effect of abio�c factors on its incidence in pigeonpea. Indian J. Ent. 73(2): 162-165.
152

DISEASE DIAGNOSIS OF LEGUME CROPS AND THEIR MANAGEMENT
S.K. KHIRBATDepartment of Entomology,
CCS Haryana Agricultural University,Hisar-125 004 (Haryana)
A.� Chickpea diseases
a)� Wilt : Pathogen,�Fusarium oxysporum f sp. ciceri
Symptom: Appeared 3-5 weeks a�er sowing and later on February on wards. Drooping of the pe�oles and rachis and ul�mately plant dries up. Internal xylem discoloura�on. External root ro�ng absent. More in western part of Haryana
Disease cycle: Soil and seed-borne in nature. Found in disease debris also. 30°C and dry condi�ons favourable for disease
b)� Root rot:� Two types of root rot
Dry root rot Pathogen:� Rhizotonia bata�cola� �
Symptom: Yellowing and drying of plant. External root rot of most of roots
Tap root bri�le and can be pulled easily from ground. Minute sclero�a or sparse grey mycelium in pith cavity of roots and in bark
Wet Root rot
Symptom: Mostly in seedling stage in soil with higher moisture content. Affected seedling gradually turn yellow and pe�oles as well as leaflets droops. District dark brown lesion appears just above the collar regions on stem associated with pinkish mycelia growth. Do not produce sclero�a on host plants
Management (Wilt and Root rot): Sow the crop in month of November. Collect the debris and burn it. Deep sowing during summer month. Seed treatment with vitavax (1 g) + Trichoderma viridae (4 g). For this purpose, a paste of Bioderma and vitavax in equal amount of water (5 ml) and then treat the seed. Crop rota�on for three years. Grow resistant varie�es like HC-1, HC-3, HK-1 and HK-5
Stem rot, Pathogen:� Sclero�nia sclero�orum
Symptom: Appeared during February-March under moist condi�ons where plant canopy was more. White mycelium could be seen around the whole stem. Later plants
153

become straw coloured and die. Disease debris play important role for primary spread
Management: Collect the debris and burn it. Remove the plants where disease appeared and allow to pass the air. Plough up the field during summer months
Stunt Virus disease transmi�ed by an Aphis crassivora
Symptom: The leaves of black gram become reddish brown in colour. The leaves of kabuli gram become yellow in colour. Internal phloem discoloura�on. Transmi�ed by an aphid
Management: Rogue out the infected plants. Spray with Metasystox @ 200 ml/200 L water/acre
Ascochyta blight, Seed as well as soil-borne in nature
Symptom: Affected all the above ground plant parts. Light brown spot appeared on leaves, stem and pods. Pycnidia arranged in concentric rings on leaves and pods. The lesion elongated on the stem and girdled in severe case
Management: Collect plant debris and burn it. Use diseased free seed. Grow resistant variety like C235 and Gaurav. Treat seed with Bavis�n 2.5 g/kg seed before sowing
Botry�s grey mould, Pathogen: Botry�s cinerea
Symptoms: Grey or dark brown lesion on leaflet branches and pods. Growing �p and flowers are suscep�ble to infec�on. Yellowing, shreading and death of infected flowers
Disease cycle: Disease debris. Externally and internally seed-borne
Management: Seed treatment with Bavis�n 2.5 g/kg. Spray with Dithane M-45 @ 400/200 L water/acre
Diseases of Pea
Powdry mildew, Pathogen:� Erysiphe polygoni
Symptoms: Occurred severely at flowering stage. White patches on leaves, stem and pods. In severe cases flour could be seen on all plant parts. Disease cycle:� Disease debris
Management: Spray with Karathene 250 ml in 250 water/Acre or Spray with selfex 500ml in 250 water/Acre
Wilt, Pathogen:� Fusarium oxysporum f. sp. pisi
Symptoms: Dwarfing, yellowing of foliage and ul�mately drying of plants. Internal
154

discoloura�on of xylem vessel. Disease cycle:�Seed borne, soil born and disease debris
Root rot of Pea, Pathogen:� F. solani f. sp. pisi
Symptoms: Yellowing and wil�ng of plant. Brown lesion along the top root and side roots. The symptom extend and spread up to 1-2”above the soil. Root shrivelled and death of plant
Disease cycle:� Seed borne, soil borne and plant debris. Chlaymydospore (5 yr in soil in absence of pea)
Management of wilt and root rot: Treat the seed with Bavis�n 2.5 g/kg. Crop rota�on – 3 yea�
Diseases of Len�l
Wilt , Pathogen:� Fusarium oxysporum f. sp. len�s
Symptom: Appeared in patches at seedling and adult stage. Drooping of the leaves and seedling dried up. Dull green colour followed by wil�ng at adult stage. Internal discoloura�on of xylem �ssue
Disease cycle:� Seed borne, soil borne and diseased debris
Management: Seed treatment Benomyl 3 g/kg seed. Late sowing. Crop rota�on – 4.5 years
Stem rot, Pathogen:� Sclero�nia sclero�orum
Symptom: Ro�ng of stem �ssue above the collar region. Yellowing and drooping of leaves above the infec�on point. Rolling and presence of white mycelium growth on stem por�on
Management: Destruc�on of debris. Spray with Dithane M-45 @ 400 g/200 L water/acre
Disease of mungbean and urdbean
Cercospora leaf spot, Pathogen:� Cercospora canescens
Symptoms: Spots appeared on leaves, stalk and pods.
The upper surface turned light yellow
Spots dark
Rhizoctonia leaf spot, Pathogen:� R. bata�cola
Symptoms: The disease caused seed rot seedling mortality and dry root rot of tap root.
The seed killed before emergence. Two or three weeks old seedlings infected at collar region and produce stem canker. Light yellow spot appeared on margin then dark
155

brown and progress towards mid-rib. Shreading of leaves
Disease cycle:� Seed as well as soil borne in nature
Management: Spray with Zineb @ 800 g/300 L water/acre. Bud spray 45 days a�er sowing
Bacterial leaf spot, Pathogen: Xanthomonas campestris pv vignaeradiata
Symptoms: Water soaked spot on under surface of leaves. Spots dark brown, raised which do not exceed 1 mm size. Covered all the leaf surface
Disease cycle:� Seed borne in nature
Management:�Spray with Streptocycline 25 g/100 L water
Yellow mosaic, Pathogen:� Vector – Whitefly
Symptom: Yellow specks appeared on the leaves. Green and yellow patches on leaves. The infected plants bears less pods and machine alter. Pods become yellow in colour. Transmi�ed by whitefly
Management: Rogue out the infected plants. Grow resistant variety – Sa�ya, Basan�. Spray crop with Metasystox 200 ml/200 L water/acre
Leaf crinkle of urdbean, Pathogen:� Virus disease – seed-borne
Symptoms: Appeared 3-4 weeks a�er sowing. Leaflets become crinkled and curl downward. The pe�ole of third trifoliate become smaller and closely touch the other leaves. Beard flowers 8-10 days a�er normal plants. Seed-borne and sap transmi�ed.
Management: Rogue out the infected plants. Grow resistant variety Pant U-19 and Pant U-26
Diseases of Clusture bean
Bacterial blight, Pathogen:� Xanthomonas cyamopsidis
Symptom: Spots interveinal and water soaked in appearance. Spot mixed together result blight phase. Invaded the vascular �ssue and cause flaccidity of affected por�on. The spot developed from the margin appear V-shaped and progressed to mid-rib. Infec�on advanced systemically through pe�oles into the stem and produced longitudinal streaks.
Disease cycle:� Internally seed-borne in nature
Management: Dissolve 5 g Streptocycline 5 L water and dip 5 kg seed for 40 minutes. Spray with Streptocycline (30 g) + Blitox (400 g) /200 L water
Alternaria leaf spot, Pathogen:� Alternaria spp.
156

Symptom: Round to irregular dark brown spot appeared in leaf surface. Concentric rings could be seen around the spots. In severe infec�on leaf defoliate
Disease cycle:� Seed borne and debris
Management: Collect the debris and burn it. Spray with Mancozeb @ 0.2%
Diseases of Pigeonpea
Wilt, Pathogen: Fusarium udum
Symptom: Interveinal claring of the leaves and banding on stem. Chlorosis of foliage and ul�mately drying. Browning of xylem vessel
Disease cycle:� Soil borne
Management:�Use of Trichoderma harzianum @ 4 g/kg seed. Soil solarisa�on
Sterility mosaic, Pathogen:� Pigeonpea sterility mosaic virus
Symptoms: Pale green plants without flowers and pods.
Excessive vegeta�ve growth. Light and pale green patches ini�ally appeared in young leaves.
Management: Rogue out the infected plants. Spray with Rogor 200ml in 200 ml water/Acre
Phytophthora blight, Pathogen: Phytophthora cajani
Symptoms: Water soaked lesion. Swelling of the stem. Main problem in standing water.
Girdling of the stem. Disease cycle:� Soil borne
Management: Do not allow the water to stand in the field. Deep ploughing during summer months
157

NEMATODE INFESTATION AND ITS DIAGNOSES IN LEGUME CROPS
K. R. DABUR* & I. J. PARUTHIDepartment of Nematology,
CCS Haryana Agricultural University, Hisar-125 004 (Haryana)
Leguminous pulses occupy an important posi�on in human dietary and are very good source of vegetable proteins varying between 17-43 per cent and they supplement to cereal based diet (Swaminathan, 1974). With the help of their root nodules, pulses fix atmospheric nitrogen and contribute substan�ally to the soil fer�lity. Pulse crops have been the backbone of Indian Agriculture, enabling the land to emliorate and maintain its fer�lity. Generally, the pulse crops are included in a crop rota�on to maintain the soil fer�lity which affects the crop produc�vity. Some pulses are considered to be excellent forages and grain concentrates in the feed for the large ca�le popula�on and some of them serve as green manure crop, which add the required humus and major plant nutrients to the soil. Pulses in India are being cul�vated in 22.5 million hectares with an average of 11.2 million tones of produc�on per year. At present the na�onal average produc�on of pulses is 6-8 q/ha which is quite low in comparison to that of other countries of the world. With a steep increase in the human popula�on, per capita availability of pulses has declined, resul�ng in under-nutri�on and malnutri�on problems in the country. Therefore, it becomes impera�ve to substan�ally increase the pulse produc�on to a level at which protein requirements of our people could be met out. Unfortunately, growing pulse crops suffer several constraints, of which pests and diseases have their leading role. Plant parasi�c nematodes are invariably found in the soil around plant root, which may act as a limi�ng factor in the crop produc�on. The economic damage becomes higher when the nematodes interact with other micro-organisms. In India a large number of phytophagous nematodes have been reported around the roots of various pulse crops (table1). A breakthrough in the produc�vity of pulse crops can be achieved by adop�ng suitable measures of management prac�ces through advanced scien�fic technologies. Among pests and diseases of pulse crops, the nematodes have remained neglected or over seen since long, however, in recent �mes Meloidogyne spp. Heterodera spp. Pratylenchus spp. Hoplolaimus spp. Helicotylenchus spp. Telotylenchus spp. and Rotylenchus reniformis have been found potent parasites causing poten�al damage to the pulses, resul�ng in economic losses in yield. These damages are reflected in the form of yellowing, stun�ng, poor plant growth and reduc�on in yield. The ill effects of nematodes have been variable on different important pulse crops of India viz. gram, black gram, moth, black gram, len�l, field pea, cowpea, guar, french bean, soyabean, limabean, pigeonpea, etc.
158

Root-knot nematode (Meloidogyne spp.): Root-knot nematodes are considered as most important pests of pulse crops and are rated as number one nematode pest because of its widespread occurrence, endoparasi�c nature and wide host range. Out of 12 species of this nematode reported from India, two namely, M. incognita and M. javanica are the most frequent species , infes�ng almost on all the pulse crops.
Hosts: These are polyphagous nematodes with wide host range. They are known to invade almost all the pulse crops like chickpea, cowpea, mungbean, blackgram, pea, len�l and soyabean.
Biology: Very few reports are available on the biology of root-knot nematode on pulse crops. The nematode is reported to complete its lifecycle in 3-4 weeks during kharif season but this dura�on is increased in rabi season due to low temperature prevailing in that dura�on. In general nematode completes several genera�ons on a suitable host during a crop season.
Nature of damage: The root-knot disease of pulses is characterized by the presence of simple or mul�ple knots on the root system, especially on root-�ps and can be easily seen with naked eyes. Size of knots varies depending on crop, age of the plant and nematode species involved. Generally, M. javanica exhibits smaller knots than M. incognita while the la�er produces profuse kno�ng on the en�re root system. Patches of poorly growing plant stand may be the first indica�on of nematode infesta�on in a crop. Stunted growth, pale green foliage, reduc�on in branches, wil�ng, poor pod forma�on, poorer yields and fewer rhizobium nodules, are common symptoms associated with root-knot disease of pulses. Heavy galling on the roots results in the development of wil�ng symptoms in the field. Seed produced on such plants may be deformed, under developed and under size. Plants infected at an early stage further carry the infec�on and the severity of the symptoms aggravate with age of crop. Symptoms of nematode infesta�on are more severe in light textured soils having low organic ma�er. Fields infested with root-knot nematodes in preceding cropping season exhibit poor germina�on of the crop grown in succeeding season. Under severe infec�on condi�ons they may succumb to root-knot and wilt disease complex. Popula�ons of M. javanica may alter Fusarium wilt resistance in chickpea, pigeonpea and len�l.
Pathogenecity: Pathogenecity of this nematode in various pulse crops has been recorded by various workers. Inoculum level of 1j2/g soil has been reported pathogenic in case of chickpea (Dhangar & Gupta, 1983), len�l (Mishra & Gaur, 1980), mungbean (Hussani & Seshadri, 1975).
Losses: Losses caused by root-knot nematode in pulses is well recognized in all the pulse growing areas of the country (table 2). Gaur (1973) reported 8-82 % reduc�on in mungbean depending upon the level of infesta�on of M. incognita in the soil.
159

Pigeonpea cyst nematode (Heterodera cajani): Pigeonpea cyst nematode was reported for the first �me from Delhi by Koshy (1967) on Cajanus cajan. This species is known to occur on various pulse crops in different states of India like Andhra Pradesh, Bihar, Delhi, Haryana, Punjab, Rajasthan, U�ar Pradesh and Tamil Nadu.
Hosts: The nematode has wide host range and parasi�zes many pulse crops like pigeonpea, cowpea, uradbean, mungbean, mothbean, etc.
Biology: It is a sedentary endoparasi�c nematode and has been reported to complete one genera�on in 16 days at 84° F. Under cooler condi�ons temperature with temperature range of 10-25 °c the nematode completes one genera�on in about 45 days. On pigeonpea, this nematode has been reported to complete 8-9 genera�ons per cropping season.
Nature of damage: Delay in flowering and pod forma�on has been reported in pigeonpea, infested with cyst nematode, Heterodera cajani. The roots of such plants bear pearly white cyst. The plants exhibit yellow foliage, extensive necrosis and early senescence. Patches of stunted and pale looking plants with few flowers are the visual symptoms when pigeonpea crop is infested with cyst nematode, H. cajani.
Interac�ons with other micro organisms: This nematode causes more damage to the crops in the presence of several soil borne fungi. Edward and Singh (1979) recorded more damage to pigeonpea due to H. cajani in associa�on with Fusarium udum , a soil borne fungus.
Losses: The informa�on on losses due to H. cajani in various pulse crops is very limited (table 2). Saxena and Reddy (1987) reported losses due to H. cajani as 30.1 % in terms of grain yield. But the loss due to this nematode in associa�on with Fusarium has been reported to be as high as 93.6 % in pigeonpea crop by Hassan (1984).
Reniform nematode (Rotylenchulus reniformis): Linford and Oliveira (1940) reported R. reniformis for the first �me, on cowpea grown in pineapple field at Hawaai in U.S.A. and is rated as an important pest of pulse crops in India also. This is a sedentary semi-endoparasi�c nematode.
Hosts: Pigeonpea, chickpea, cowpea, beans and blackgram are the main host crops of this nematode.
Biology: This nematode has been reported to complete one life cycle in 15-25 days in pulse crops (Leinford and Oliveira, 1940; Siva Kumar and Sashadri, 1971). The nematode has two races, namely race A and race B (Dasgupta and Sashadri, 1971). Race A parasi�zes cowpea, castor and co�on crops, whereas race B parasi�zes cowpea only. Young females of the nematodes are seen a�ached to the roots of infected plants.
Nature of damage: Symptom produced by reniform nematode, R. reniformis, is
160

malforma�on of root system in pigeonpea which is also called 'dirty root disease'. It is due to the fact that when the roots of infected plants are washed with water, the soil adhered to the gela�nous matrix is not completely washed off. Affected plants appear light green in colour and generally appear in the field as patches of stunted plants. Symptoms of nematode infesta�on are discernible as patches of plants with uneven growth even in early stage of the crops viz. chickpea, mungbean, urdbean and len�l, however at maturity of the crop it is not easy to dis�nguish between nematode infested and healthy plants as the symptoms are mixed up with senescence of the crop. Early senescence, some�mes even at flowering stage, appears in patches in field pea and common bean. The presences of female nematodes on rootlets may be observed on en�re root system a�er a gentle wash. This is seen at flowering and at maturity stages and is the only remarkable symptom of reniform nematode infec�on.
Interac�on with other organisms: This nematode in associa�on with root-knot nematode (M. incognita) has been reported to cause more damage in blackgram than the individual nematodes, Mishra and Gaur (1981).
Root lesion (Pratylenchus spp.) : This group of plant parasi�c nematodes feeds on most of the pulse crops. Many species of this nematode are found associated with pulse crops but P. thoronei is the most important and potent species of this group invading the pulse crops.
Biology: This is a migratory endoparasi�c nematode. All the stages of the nematode are capable of penetra�ng the roots. A�er entering, the nematodes feed on cor�cal cells and cause necrosis. The female nematode lays eggs inside the roots or in soil. Complete lifecycle of this nematode requires 30-92 days depending upon temperature and host crop. Pathogenic level of this nematode in chickpea has been reported to be 4000 nematodes/kg soil.
Nature of damage: Generally, forma�on of necro�c lesions on the roots is the characteris�c symptoms of Pratylenchus spp. infesta�on in chickpea. In case of heavy infesta�on the necro�c spots coalesce, leading to the development of necrosis in the en�re root system. In early stage of the crop, infesta�on is manifested by appearance of patches of stunted plants and pale green foliage. The stun�ng becomes more pronounced as cropping season advances. No characteris�c symptoms are evident on above ground parts of infested chickpea plants. As compared to root-knot nematodes, the damage caused by lesion nematodes is less impressive. The plant show stun�ng, chlorosis and lack of vigor. On the roots, infec�on is characterized by dark brown to black lesions. The above ground symptoms are similar to those of other nematode infesta�on in plants and resemble with those of malnutri�on.
Other nematodes: Besides above men�oned nematodes, several other nematodes like lance nematode (Hoplolaimus spp.), spiral nematode (Helicotylenchus spp.), stunt
161
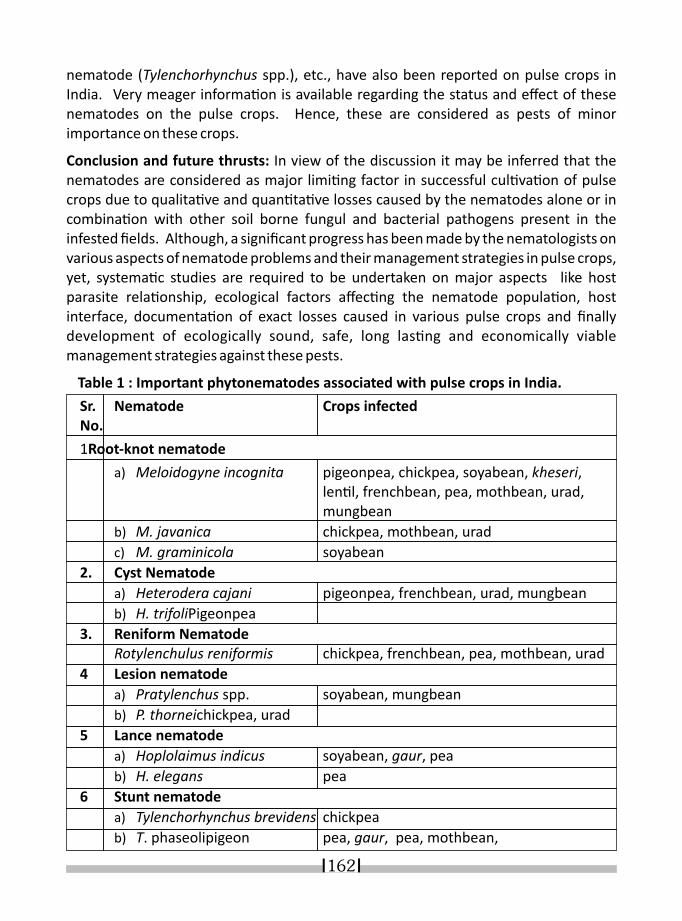
nematode (Tylenchorhynchus spp.), etc., have also been reported on pulse crops in India. Very meager informa�on is available regarding the status and effect of these nematodes on the pulse crops. Hence, these are considered as pests of minor importance on these crops.
Conclusion and future thrusts: In view of the discussion it may be inferred that the nematodes are considered as major limi�ng factor in successful cul�va�on of pulse crops due to qualita�ve and quan�ta�ve losses caused by the nematodes alone or in combina�on with other soil borne fungul and bacterial pathogens present in the infested fields. Although, a significant progress has been made by the nematologists on various aspects of nematode problems and their management strategies in pulse crops, yet, systema�c studies are required to be undertaken on major aspects like host parasite rela�onship, ecological factors affec�ng the nematode popula�on, host interface, documenta�on of exact losses caused in various pulse crops and finally development of ecologically sound, safe, long las�ng and economically viable management strategies against these pests.
Table 1 : Important phytonematodes associated with pulse crops in India.
Sr. Nematode Crops infectedNo.
1Root-knot nematode
a) Meloidogyne incognita pigeonpea, chickpea, soyabean, kheseri, len�l, frenchbean, pea, mothbean, urad, mungbean
b) M. javanica chickpea, mothbean, urad
c) M. graminicola soyabean
2. Cyst Nematode
a) Heterodera cajani pigeonpea, frenchbean, urad, mungbean
b) H. trifoliPigeonpea
3. Reniform Nematode Rotylenchulus reniformis chickpea, frenchbean, pea, mothbean, urad
4 Lesion nematode
a) Pratylenchus spp. soyabean, mungbean
b) P. thorneichickpea, urad
5 Lance nematode
a) Hoplolaimus indicus soyabean, gaur, pea
b) H. elegans pea
6 Stunt nematode
a) Tylenchorhynchus brevidens chickpea
b) T. phaseolipigeon pea, gaur, pea, mothbean,
162
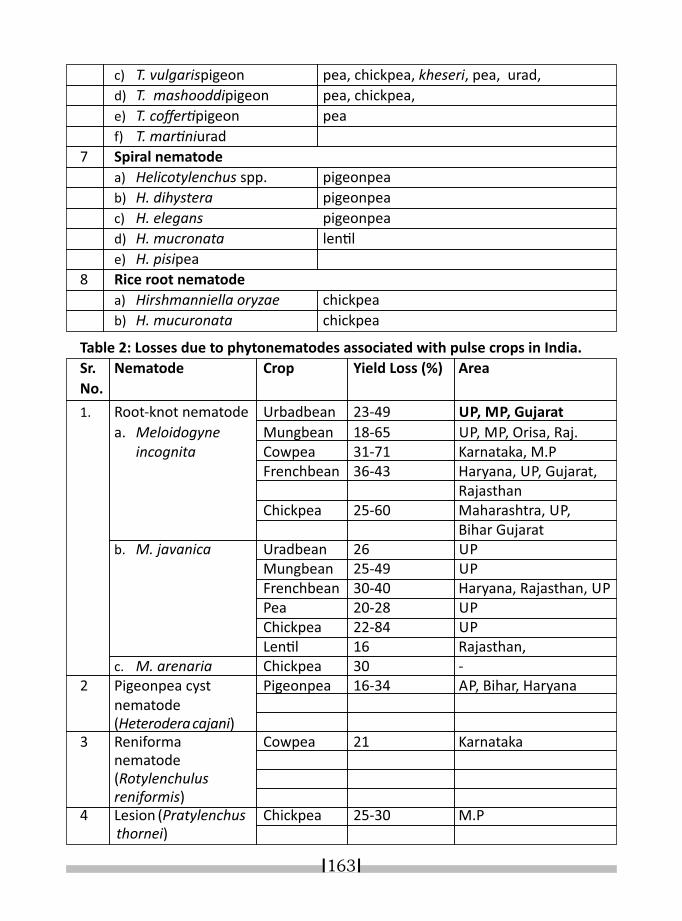
c) T. vulgarispigeon pea, chickpea, kheseri, pea, urad,
d) T. mashooddipigeon pea, chickpea,
e) T. coffer�pigeon pea
f) T. mar�niurad
7 Spiral nematode
a) Helicotylenchus spp. pigeonpea
b) H. dihystera pigeonpea
c) H. elegans pigeonpea
d) H. mucronata len�l
e) H. pisipea
8 Rice root nematode
a) Hirshmanniella oryzae chickpea
b) H. mucuronata chickpea
Table 2: Losses due to phytonematodes associated with pulse crops in India.
Sr. Nematode Crop Yield Loss (%) Area
No.
1. Root-knot nematode Urbadbean 23-49 UP, MP, Gujarat
a. Meloidogyne Mungbean 18-65 UP, MP, Orisa, Raj. incognita Cowpea 31-71 Karnataka, M.P Frenchbean 36-43 Haryana, UP, Gujarat, Rajasthan
Chickpea 25-60 Maharashtra, UP, Bihar Gujarat b. M. javanica Uradbean 26 UP
Mungbean 25-49 UP Frenchbean 30-40 Haryana, Rajasthan, UP Pea 20-28 UP Chickpea 22-84 UP Len�l 16 Rajasthan,
c. M. arenaria Chickpea 30 -2 Pigeonpea cyst Pigeonpea 16-34 AP, Bihar, Haryana nematode (Heterodera cajani)3 Reniforma Cowpea 21 Karnataka nematode (Rotylenchulus reniformis)4 Lesion (Pratylenchus Chickpea 25-30 M.P thornei)
163
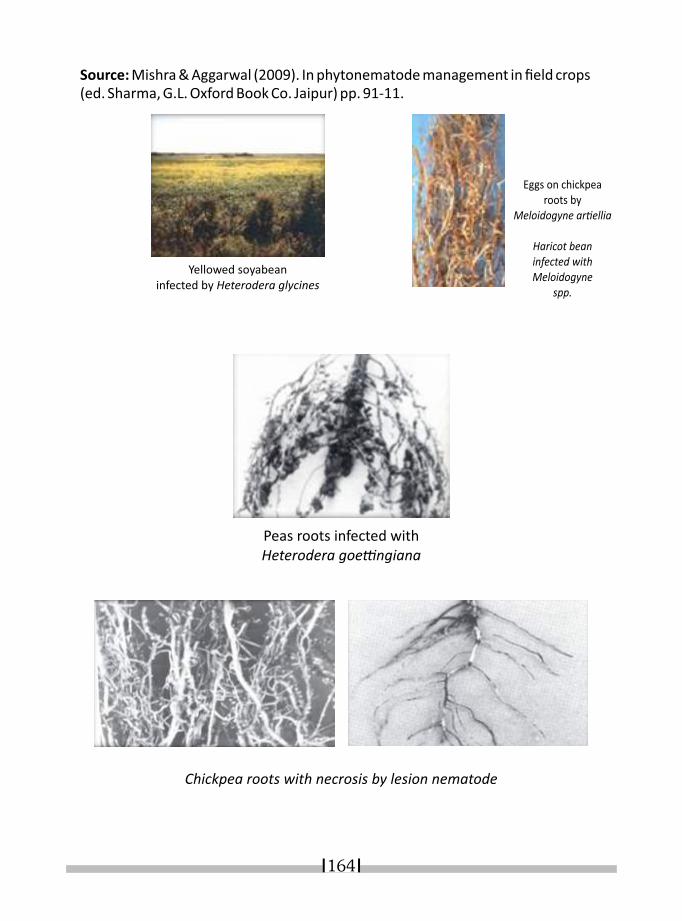
Source: Mishra & Aggarwal (2009). In phytonematode management in field crops(ed. Sharma, G.L. Oxford Book Co. Jaipur) pp. 91-11.
Yellowed soyabean infected by Heterodera glycines
Eggs on chickpea roots by
Meloidogyne ar�ellia
Haricot bean infected with Meloidogyne
spp.
Peas roots infected with Heterodera goe�ngiana
Chickpea roots with necrosis by lesion nematode
164

MITE PESTS OF LEGUMES AND THEIR MANAGEMENT*ABHISHEK SHUKLA AND S. S. MANDAPE
*All India Network Project on Agricultural AcarologyDepartment of Entomology, N.M. College of AgricultureNavsari Agricultural University, Navsari-396 450, Gujarat
Acarology dealing with mites and �cks were unknown in India un�l the middle of the last century which has gained tremendous momentum during the later decades
thof the 20 century. Agricultural importance of mites as pests of hor�cultural and agricultural crops are being appreciated all over the world and have caused great concern to both farmers and agricultural scien�sts with regard to their management. The main contribu�ng factor for this has been the wide spread use of non-selec�ve pes�cides mainly of chlorinated hydrocarbons and organophosphorous types as they were innocuous before the use of such chemicals. Mites occupy diverse habitat and can be found in any conceivable habitat, though because of their small size they are not easily seen, unlike many insects, mite damage to plants is o�en slow, and in the ini�al stages damage symptoms are o�en confused with those due to nutrient deficiencies. The crop losses due to mite damage are as much as 50-80 % in mango, 30-40% in litchi, 20-25% in paddy, 20-30% in sugarcane, 15-30% in tea and 25-30% in ber. Besides, total failure of crop due to mite infesta�on is not uncommon. However, many mites are beneficial to man in that they prey on undesirable arthropods. Other aid in the breakdown of forest li�er and implement nutrient recycling.
DISTINGUISHING FEATURES OF MITES FROM RELATED FORMS
Mites are closely related to the insects. For a common man, mites, �cks insects and spiders like alike but mites can easily be differen�ated from them by following characters:
From Insects: Mites lack wings, mandibles and antennae and normally possess eight, instead of six legs in case of insects in adult stage. The body consists of two divisions, anterior “gnathosoma” (bearing oral opening and mouth parts) and posterior “idiosoma” (bearing anal and genital openings and legs) unlike insects who have three divisions head, thorax and abdomen. Apart from smallness of size, their most conspicuous character is a reduc�on in segmenta�on (inconspicuous or absent) which shows characteris�cs of the typical arthropod.
From spider: The mites and �cks are closest rela�ve of the spider. The abdomen in former group is broadly joined to gnathosoma forming a single piece and lacks the waist (pedicel) which is characteris�c of spiders who have two dis�nct body regions, anterior (cephalothorax) bearing mouth appendages and four pairs of legs and posterior
165

(abdomen) devoid of any appendages and both the regions are separated by constric�on (waist).
From �cks: The �cks, although very similar to the mites in morphology, differ from later in possessing hypostome with recurved teeth. Haller's organ on tarsus of leg I, presence of s�gmata posteriolateral to the coxae of leg IV and leathery texture of body. Besides, the mites are plant parasi�c and animal parasi�c both while �cks are exclusively animal parasi�c (blood suckers).
Senior Acarologist; [email protected]
ACARINES OF ECONOMIC IMPORTANCE IN AGRICULTURE1. Plant feeding (Phytophagous) mites: Mites mainly causing economic damage
to food crops are spider mites (Family Tetranychidae), false spider mites (Family Tenuipalpidae), gall mites or bud mites or blister mites (Family Eriophyidae) and yellow or broad mites (Family Tarsonemidae). Among phytophagous mites, eriophyids are to a great extent host specific. The mites of other families men�oned as above are, however, not host specific. Some of the salient features of these groups are:Spider mites (Family Tetranychidae): These mites are exclusively plant feeders, so� bodied, variously coloured, colony forming and normally inhabit the under surface of the leaves. Many species except Bryobia and Petrobia spin webs like spiders to cover their colonies hence the name. Larva 6-lagged, while nymph and adult 8-legged, size is 0.5 to 0.6 mm. (Plate-1)
MORPHOLOGICAL DIFFERENCE BETWEEN MITES AND INSECTMORPHOLOGICAL DIFFERENCE BETWEEN MITES AND INSECTMORPHOLOGICAL DIFFERENCE BETWEEN MITES AND INSECT
Body divided into two part Adult stage has four pairs of leg Do not have antennae Do not have wings In place of mandible it possess pair of
chelicerae and padipalp
Body divided into three part Adult stage has three pairs of leg Have one pair of antennae Have to pairs of wing Have one pair of mandible
Mite Insect
Mite Insect
1. Gnathosoma
2. Ideosoma
1. Head
2. Thorax
3. Abdomen
166

False spider mites (Family Tenuipalpidae): They look like spider mites but unlike those they do not spin webs. These mites are flat, pear shaped, mostly bright coloured, slow moving and normally found on the under surface of leaves, twigs, fruits etc. some inhabit inside the galls and feed on barks, possesses 4 pairs of legs. Size is generally 0.3 to 0.35 mm. (Plate-2)Yellow or broad mite (Family Tarsonemidae): They are �ny, glossy or yellow mites, fast moving found on under surface of leaves as well as in associa�on of fungus. Males o�en carry female deutonymphs and possess 4 pairs of legs. Size is generally 0.25 to 0.30 mm. (Plate-3)Gall mite (Family Eriophyidae): They are �ny, elongated, worm like and have only two pairs of legs. Apart from leaves, they also live within buds, may form blister and most commonly form gall, hence the name. Size is generally 0.2 to 0.25 mm. (Plate-4)
2. Beneficial mites: Unlike the injurious mites, there are beneficial mites, too which are of significant importance in achieving biological control of mites. Predatory mites mainly belong to the family Phytoseiidae, S�gmaeidae, Cunaxidae, Bdellidae, Cheyle�dae, Erythraeidae and Tydeidae, etc. Of these, the mites of the family phytoseiidae are of great promise in effec�vely regula�ng mite pests if judiciously u�lized, while the rest are of rela�vely less importance.
GENERAL BIOLOGY OF AGRICULTURAL MITES
Reproduc�on in Acarina is normally through Arrhenotoky (produc�on of males from unfer�lized eggs) and in some cases through Thelytoky (produc�on of females from unfer�lized eggs i.e. Bryobia). The primi�ve life cycle consists of 6-legged larva hatching from eggs followed by 8-legged two nymphal stages (spider mites and false spider mites) or three nymphal stages (many predatory mites) or no nymphal stage (many tarsonemid mites) and finally reach adult stage, also provided with 8-legs.
A common red spider mite (Tetranychidae)
A common false spider mite (Tenupalpidae)
A common broad / yellow mite
(Trarsonemidae)
A common erineum mite (Eriophyidae)
Phytophagous Mites causing economic damage
to crops plants are belong to 4 families
ACARINES OF ECONOMIC IMPORTANCE IN AGRICULTURE
167

However, in Eriophyidae, two pairs of legs are present throughout the developmental stages, including the adults.
ECONOMIC IMPORTANCE OF AGRICULTURAL MITES
Some of the mites associated with plants are quite injurious as they cause damage to their hosts either directly or indirectly. On the contrary there are good number of species which act as predators of phytophagous mites and some small insect pests as aphids, coccids etc. and thereby help in doing natural control of pests.
Direct damage:
i) Phytophagous mites penetrate the plant �ssues by their needle like (styli form)
chelicerae and suck the exuding sap. During this process, they damage leaf
�ssue, destroy chlorophyll resul�ng in change of photosynthesis and
transpira�on rates. Plants o�en remain covered with dense webs (in case of
spider mite infesta�on) where dust par�cles adhere to and retard normal
physiological ac�vi�es of the plant. All these s�mulate the appearance of
common mite feeding symptoms as light yellow spots, s�ppling, bronzing,
defolia�on, retarda�on of growth, curling, crinkling, bri�leness of leaves,
reduc�on in size, quality and quan�ty of fruits and flowers and all these affect
the yield of the crop. Normally spider mites and false mites show these
symptoms.
LOSSES CAUSES BY VEGETABLES MITES
Vegetable crops Losses ReferenceBrinjal Okra Chilli All vegetables
16.16% (due to T. ur�cae)23-25% (due to T.macfarlanei)61.23% (due to P.latus)15-20% (due to all mites)
Anonymous,(2007)
Female adult 4 pairs of
legs
Egg
Deitpmu,[j4 pairs of
legs
Larva 3 pairs of
legs
Protonymph 4 pairs of
legs
Two spo�ed mite life cycle
8-12 days at 30-32⁰C
168

ii) While feeding, some eriophyids and tenupalpids inject toxins through saliva
which causes malforma�ons and deformi�e (citrus, guava), gall forma�on
(pongamia) erineum (brinjal, co�on, jasmine, litchi), witche's broom effect
(mango, chrysanthemum), blisters (sugarcane), russe�ng (citrus) etc.
Tarsonemides cause curling and twis�ng of leaves may be because of toxin
injec�on.
Indirect damage:
i) Among the plant associated mites, eriophyids like Aceria cajani cause pigonpea
sterility mosaic disease, fig mosaic disease by Aceria ficus, streak mosaic
disease by Aceria tulipae, sugarcane streak mosaic virus disease by Aceria
sacchariand tetranychid mite, Tetranychus ludeni transmites dolichos ena�on
mosaic virus.
ii) Some mites are also known to cause fungal diseases (they act as vector of these
diseases or spread the spores from one place to another).
Seasonal occurrence: As a rule, mites are numerous during summer months, mul�ply
quickly and pass through several genera�ons in a very short period of �mes like aphids
and can build up a very high popula�on. Most of the phytophagous mites and predatory
mites occur in the field throughout the year in tropical climate except, of course, in rainy
season when the popula�on declines considerably due to washing away of the leaf
popula�on and also during severe winter months when the egg laying ceases due to
dropping down of temperature below the development threshold. Temperature,
humidity and light are important factors influencing dynamism of mites.
Dispersal and spread: The dispersal and spread of infesta�on in mites accomplished
through various mechanisms as:
I. Transporta�on of hosts in case of mites infes�ng ornamental plants and fruit trees.
II. The mite-dispersal from one field to another takes place through crawling.
III. Mites are o�en carried by different organisms from one field to another by insects, birds and even man himself.
IV. Mites are blown away by wind.
V. In some tetranychids, the dispersal in a very peculiar way called “Balooning” in which they hang themselves from leaves (when popula�on pressure is very high) with the help of fine silken threads (ac�ng as balloon or as parachute) spun by themselves and are carried away to a distant places through a gentle wind current.
169

MAIN REASONS FOR THE PROBLEM OF MITES IN AGRICULTURE
1. Elimina�on of natural enemies due to random and indiscriminate use of chlorinated hydrocarbons and synthe�c pyrethroids for general pest control.
2. Use of improves agricultural prac�ces, high yielding varie�es and higher dose of nitrogenous fer�lizers make the plants more suscep�ble to mite a�ack and also induce mite mul�plica�on.
3. Development of resistance and cross resistance in mite with the use of sublethal doses of chemicals.
4. Some pes�cides like DDT, diazinone, carbaryl, synthe�c pyrethroids cause sudden pest upsurge due to increased reproduc�ve poten�ality of mites as post-spray effect leading to physiological inducement in the pests as well as biochemical changes in plants as affected by these chemicals.
5. Plan�ng an area with a single crop (monoculture) provides extensive food supplies for mite pest and consequent increase in popula�on.
Mite Pests of Legumes (Pulses):- Pulses are important source of dietary protein and have unique property of maintaining and restoring soil fer�lity through biological nitrogen fixa�on as well as conserving and improving physical proper�es of soil by virtue of their deep root system and leaf fall. India grows pulse crops in 23 million ha producing 14.4 million tones of pulse grains (Anan. 2009). Common pulse grains of India are bengal gram, pigeonpea, green gram, black gram, len�l, field pea, etc. The mite species which infest, various pulse crops in India are as below:
Pigeonpea (Cajanas cajan)
Aceria cajani Channa Basavanna
This is popularly called Pigeonpea sterility mosaic (PPSM) mite. This mite act as a vector of PPSM disease which occurs more to the young plants. The damage is so serious that the pods are not produced, though the vegeta�ve growth of the plant is normal and looks greenish and healthy. The infesta�on of the mite and transmi�ng the virus make the flowers sterile, shed off and no pod forma�on takes place. If the plants are infested with virus at the later stage, the least damage is caused and can give yield up to 50-75%.
Pigeonpea Sterility Mosaic Disease
Pigeonpea is cul�vated in tropics and subtropics and semi arid areas of the world. This crop is severely a�acked by a sterility mosaic disease in India like U�ar Pradesh, Bihar, Punjab, Gujarat, Maharashtra, Tamil Nadu, Andhra Pradesh, Karnataka and Cha�shgarh (Srinivasa et al., 2004) and beside India, it also occurs in Bangladesh, Nepal, Sri Lanka, Myanmar. This disease is incriminated to a mite, Aceria cajani.
170

Economic loss:- Infesta�on at early stages within 45 DAS causes complete
sterility causing total yield loss. The yield reduc�on is related to percentage of infec�on
of plants whereas when it affects some of the branches, it causes par�al sterility and
that is not correlated with percentage of infesta�on. Dras�c reduc�on in pod length,
pod width, number of grains per pod and 1000 grain weight, shoot and root, nodule
weight, nodule number, etc are caused due to this disease. The ratoon crops and
perennial crops are more seriously a�acked and losses in those cases are huge. The
es�mated yield loss may be more than two lakh tones in India (Kannaiyen et al., 1984,
Srinivasa et al., 2004).
Symptoms:- The sterility mosaic virus which is transmi�ed by Aceria cajani
causes the sterility mosaic disease. Infested plants show sterility (the flower drop off, no
pod forma�on while mosaic symptoms appear on leaves). Such infested plants are
bushy, pale green with leaf size reduced showing mo�ling. The secondary and ter�ary
branches from the leaf axils appear and no pod forma�on takes place. The reproduc�ve
structures show complete cessa�on. Severe reduc�on in internodal length, shortening
of branches and filliformed leaves are the other symptoms. The symptoms can be
categorized under three types:
a) Complete sterility: - Leaflets show severe mosaic, no flower and pod forma�on
take place and this is caused when infesta�on takes place within 45 days a�er
sowing.
b) Par�al sterility:- The leaflets show mild mosaic symptoms. The infected branches produce no flowers and pods. This occurs when infesta�on takes place beyond 45 days a�er sowing.
c) Ring spot:- In this case the infected leaflets show green island surrounded by chloro�c halo. However, with the maturity, those symptoms may disappear. This type of symptoms are common in some of the sterility tolerant cul�vars e.g. ICP-2376 (Reddy et al., 1998). However, due to this the flowering may not be affected.
Causal organisms: - According to Capoor (1950, 1952), a virus is involved for causing this disease. The viral natural of pathogen is hypothised because of transmissibility through gra�ing, associated eriophyid mites and mechanical means. Several workers ruled out the theory that the sterility mosaic disease has no rela�on with mite toxaemia/fungi/bacteria/mycoplasma/nematode, etc.
Distribu�on pa�ern of mite: - This mite preferred basal part of leaflet at both top and middle canopies. Thus, basal ¼ por�on of leaflet recorded significantly more number of mites followed by other successive por�ons of leaflet in the same order. The basal half
171

of leaflet which harbored nearly 60-65% of mite popula�on was suggested for sampling as it gave more reliable es�mate of mite vector popula�on. Further, it was also no�ced that vectors mite popula�ons were concentrated towards the pe�ole and younger leaves in sterility mosaic infected pigeonpea plant.
Transmission:- The transmission of the disease is most probably by gra� transmission method and it is neither seed born as reported by Nene and Rathi (1983) and Ghanekar et al., (1992) nor by sap transmission using leaf juices from the infected plants. Mites from infested plants successfully transmi�ed PPSMV to pigeonpea. Stapling of mosaic infected leaflets carrying Aceria cajani on to 2-weeks old healthy pigeonpea seedlings resulted in successful transmission of sterility mosaic and mosaic symptoms were no�ced from 1-2 weeks a�er stapling, the extent of disease transmission with a single stapling on suscep�ble varie�es like Maru� and TTB7 ranged from 18-46% and 32-66%, respec�vely (Srinivasa et al., 2004).
Vector: - The transmission of PPSM y Acria cajani from diseased to healthy plants was reported by Seth (1962) and later was confirmed by Nene (1972). Further Nane and Reddy (1976) confirmed that a single mite is capable of transmi�ng this disease.
The life cycle of this mite is as below:-
Eggs: - 30-40 µm, parallel sided, milky white, rounded at the ends, loosely a�ached to filamentous trichomes, hatch within 4-5 days.
Nymphs: - I stage nymph –dura�on unknown, II stage nymph-48 hours to less than 5 days.
Adult: - II stage nymph moults to adult. Adult is 200-250 µm long.
Total life cycle:- About 2 weeks.
Adult male: - smaller than female, deposits stalked spermatophores which remain
firmly a�ached to leaf with an expanded base.
Adult female: - Have parallel ribbed flap covering the genitalia.
Pre-oviposi�on Period:- 24 hours
Oviposi�on: - 1-3 eggs are laid per day.
Dispersal of vector:- According to Naine et al., (1976) and the dispersal of vector is
mostly by air current. The disease spreads in the downwind direc�on up to 2 km from
the source of inoculums while the spread in the upwind direc�on is very limited.
However, the incidence and spread of disease depend upon vector popula�on in the
field (Reddy et al., 1989).
Popula�on dynamics:- The vector mite remains in field throughout the cropping period
and popula�on gradually increases from 45-60 days a�er sowing and the peak is
172

a�ained during 120-150 days old crop. (Between October-November). The increase in
vector popula�on was associated with simultaneous increase in the extent of disease.
They reported popula�on increase from 45-60 days reaching peak in mid October to
mid November (Prabhuswamy et al., 1995).
Survival of Aceria cajani on Ratoon crops:- Narayanswamy (2004) studied survival of
this mite on ratoon crop and Atylosia scarabaeoides and reported that the survival on
the la�er crop was from January-October in various propor�on ranging from 10-53
mites/trifoliate leaf and lowest was in February, though it survived on Atylosia
scarbaeoides throughout the year and that may act as poten�al source for spread of
vector in absence of other possible sources like stubbles, ra�ons, etc.
Spa�al distribu�on:- Srinivasa et al. (2004) studied the distribu�on pa�ern of Aceria
cajani on pigeonpea and reported that this mite preferred basal por�on of leaflet both
at top and middle canopies which recorded significantly more number of mites
followed by other successive por�ons of leaflets in the same order. The basal half of
leaflet harboured nearly 60-65% of mite popula�on on a leaflet and for that it is
suggested that the area for sampling as it gives more reliable es�mate.
Control of sterility mosaic disease/mite vector:-
1) Applica�on of phorate 10 g @1 kg/a.i./ha at sowing + foliar spray with
monocrotophos on 30 days from sowing (Arjunan et al., 1989).
2) Seed treatment with aldicarb 10 G protected crop from sterility mosaic (Rathi
1983).
3) Dicifol (0.05%) between 30 and 45 days a�er sowing (Patel et al., 1990).
4) We�ablle sulphur @ 2.5 g/l of water 60 and 90 days a�er sowing (Natarajan and Beevi, 1992) and Singh (1999).
5) Seed dressing with carbosulfan @ 15 g/ha or soil applica�on of phorate @1.5 kg a.i./ha, followed by foliar spray with dicofol at 30 and 45 days a�er sowing (De et al., 1996).
6) Reddy et al., (1993) suggested following measures for control of vectors
i) Grow resistant varie�es (Bahar/ICPL-151, Hyd-3C)
ii) Breaking pigeonpea cycle by crop rota�on, changing sowing date.
iii) Destroying infected plants during crop season and voluntary diseased plants during off season.
iv) Spraying acaricide once during first two months of age.
7) Chinniah and Mohanasundaram (1995) reported mortality of A. cajani with
173

different chemicals giving mortality as dicofol (91.62%), NSKE (90.95%), ethion (88.65%), monocrotophos (88.31%), we�able sulphur (87.77%), neem oil emulsion (87.73%), neem cake extract (86.41%), phosalone (86.41%), dithane M (64.19%). All the neem based formula�ons were very effec�ve.
Predators:- Transeius teranychivorous can keep the popula�on of eriophyid mite at low level.
Eotetranychus broodryki Meyer
� This mite is popularly called Pigeonpea green mite. The adults are green white, colonise on lower surface of leaves near midrib. The infesta�on is more acute on the young leaves at the �ps where due to feeding brownish discoloura�on appears. This a�ack is more during December-January and affects the pod yield. According to Patel et al., (1999) popula�on of Eotetranychus broodryki started building from September and a�ained peak in January I fortnight in kharif shown pigeonpea. Popula�on was higher on lower leaves. Sunshine had significant posi�ve correla�on with mite popula�on. Dicofol 0.04% was significantly superior to monocrotophos (0.05%), methyl-o-demeton (0.025%), ethion (0.1%), dimethoate (0.03%) and deltamethrin (0.05%), which were at par with each other. Neemark (0.5%) was superior to NSKE (5%).
Schizotetranychus cajani Gupta
� This mite also infests pigeonpea in different parts of India (Satpathi et al.,1992). Chillar et al., (2007) studied biology of S. cajani on red gram. Life cycle was completed in 7.87 days in August and 43.18 days in February. Incuba�on, larval, nymphal period varied from 2-22, 1-9 and 1-13 days, respec�vely. Popula�on was maximum in December- February, heavy rainfall in July-August dras�cally reduced popula�on. Agistemus sp and Amblyseius sp were predators. Karuppuchamy et al., (1990) studied biology, varietal screening and control of S. cajani on red gram. The varie�es T-21, 4758, Khargaon-2, 148 were equally suscep�ble. P-1141 was least suscep�ble in West Bengal (Gupta et al., 1985).
Tetranychus ur�cae Koch.
� According to Kalisekar and Thakur (2007) this mite assumes as major pest of pigeonpea in Meghalaya. The pest appears in august and remains in field upto September. The affected leaves progressively loose turgidity at points of feeding. Finally, such leaves dry up. As many as 12 mites per leaf may cause the onset of hypersensi�vity symptoms a�er 3 days of feeding. The studies of Latrou et al., (1995) showed that mite density of 1-32 per 1,5 cm sq of leaf area on bean plant for a feeding period of 1-5 days produce the symptoms. The ac�vity of oxida�ve enzymes, peroxide and polyphenol oxidase increased in leaves of injured plants as a result of mite feeding which led to drying of leaves and leaf dropping.
174

Green gram/ Black gram/Bengal gram/ Rajmah/French bean and Khesari (Lathyras sa�vum)/ Masur/Indian Bean
The mite associated with these pulses are quite common and the details of these mite pests ia given below-
Tetranychus ur�cae Koch.
This mite is commonly known as red spider mite. It is a polyphagous pest and found on wide range of host plants throughout the world. It colonize on the under surface of leaves and envelops the plant heavily with dense webs.
Damage:- This mite feeds on undersurface of leaves causing appearance of yellow spots on leaves near the basal part. Gradually with increasing of feeding the spots increase, which coalesce to form reddish brown patches. The infested leaves gradually dry up and defoliate.
Seasonal occurrence:- The high temperature is more preferred by this mite during which the mite problem is more acute.
Biology:- The incuba�on period is 3-4 days. The larvae are sluggish, period lasts for 3-5 days. The protonymphal and deutonymphal periods are of 2-3 days and 2-3 days, respec�vely. The life cycle is completed in 10-15 days, depending upon the season. The number of genera�ons may be as high as 30 per year.
Management:- Docofol (0.05%) will provide effec�ve control. Fluvanilate 25 EC at 75 g.ai/ha gave temporary reduc�on of mite popula�on but a�erwards it increased. Trizophos (Hostathion) 40 EC (at 0.1%), dicofol were also found to be very effec�ve. A combina�on of monocrotophos (40 EC) and dicofol (20 EC) ar 0.05% was found to be best for reducing mite popula�on.
Figure 1. Two-Spo�ed Spider Mite Life Cycle 8-40 Days
Eggs
Larva
Adult
Nymph1st stage
Nymph2nd stage
175

Tetranychus ludeni Zacher
According to Gupta (1985) T. ludeni infests over 70 hosts of which beans, brinjal, cucurbitaceous plants, etc., are the important ones. Outside India it has been recorded to feed on Hibiscus, bean, eggplant, potato, co�on, alfalfa, castor, etc. The life cycle of this mite is a typical one, the stages being egg, larva, protonymph, deutonymphs and adult. The three immature stages viz., larva, protonymph and deutonymphs were each followed by short period of quiescence called nymphochrysalis, deutochrysalis and teleochrysalis, respec�vely. Malaviya (1994) recorded three phytophagous mites viz., T. ludeni, P.latus and uniden�fied eriophyids mite and predatory A. (E) alstoniae (Phytoseiidae) as well as predatory predatory thrips, S. indicus (Thripidae) on Indian
thbean. The incidence of T. ludeni commenced from 15 STW (6.13 mite/trifoliate) and con�nued �ll crop maturity. The crop remained uninfested from 50st STW (first week of
thDecember) to 14 STW (first week of April). Major ac�vity of the pest was no�ced
ndduring warmer months (May-June) with a peak during 22 STW (May) (39.89 mites /trifoliate). Similar, trends were also observed in case of egg incidence on the crop. Among the three canopy level of the plant, the mite (mobile stage) preferred the lower canopy leaves (24.45 mites/trifoliate leaf) than middle and top canopy. Similar trend was also observed in the incidence of eggs of spider mite on different canopy level. The-life studied suggested that the net reproduc�ve rate (Ro) was 32.079 female/female/genera�on in a genera�on �me of 19.81 days. The innate capacity for increase, finite rate of increase, weekly mul�plica�on of popula�on and hypothe�cal F 2
females were 0.1750 female/female/day,1.19. 1.38 and 1029.06, respec�vely. On reaching stable age-distribu�on, the contribu�on of various stage age distribu�on, the contribu�on of various stage viz., egg, larva, protonymph, deutonymphs and adult were to the tune of 7-.16, 13.21, 5.23, 5.03 and 6.36 per cent, respec�vely. Further in field evalua�on of chemical pes�cides including botanical pes�cides, dicofol, monocrotophos and methyl-o-dematon were highly effec�ve against T. ludeni. The plant product neemark was comparable in its effec�veness with quinalphos, ethion and methyl-o-dematon.Channabasavanna and Virakamath (1989) obtained best result in the field by spraying dicofol (0.02 to 0.01 per cent) and sulphur (0.05 to 0.01 per cent) against T. ludeni infes�ng Indian bean. Sangeetha and Ramani (2011) studied the post embryonic development of the spider mite, Tetranychus ludeni Zacher infes�ng velvet
obean (Mucuna deeringiana) at 30 + 2 C and 70 + 5% rela�ve humidity. The dura�ons of pre-oviposi�on period, oviposi�on period and post-oviposi�on period were recorded to be 0.5 + 0, 11.5 + 0.38 and 0.5 + 0 days respec�vely. Fecundity recorded was 83.6 + 3.4 eggs and longevity, 12.6 + 0.37 days. The total dura�on of sexual and parthenogene�c reproduc�on respec�vely were 10.04 + 0.13 days and 9.04 + 0.13 days. The parthenogene�c development required compara�vely shorter dura�on than sexual ones. Sex ra�o (male: female) was 2: 10.
176

Polyphagotarsonemus latus (Banks)
This is also known as yellow mite. It is a serious pest of chilli. All the life stages will be found on undersurface of young apical leaves and tender shoots. The a�ack of this mite on young apical leaves, causes retarding of apical growth of developing plants. The affected leaves become silvery white and ul�mately dry up and drop down prematurely. Consequent upon it, the growth is retarded resul�ng in reduced fibres. The infested apical leaves curve downwards, become boat-shaped. In some cases, the infested leaves become darker than normal leaves. The internodes become shortened and fibre quality is affected. The infesta�on dras�cally affects plant height. The extent of loss may be as much as 36-42%. The mite suck sap from the ventral surface of leaves as a result, the leaves do not become of normal size, turn copperish brown and drop down prematurely.th mite popula�on was less on fresh leaf than on crinkled leaf. The ver�cal vegeta�ve growth is arrested.
Average longevity of female and male is 13.44 days and 12.0 days, respec�vely. The intrinsic rate of increase is rm=0.359, with a mean genera�on �me of 10.34 days and net reproduc�ve rate Ro= 41.0. a single genera�on is completed in 4-5days during summer, 7-10 days in winter. Average number of eggs laid per female per day is 3.6, laid on leaves or fruit surface. Males o�en carry the pharate deutonymphs to help in dispersal of the colony.
Propergite, dicofol, carbophenothion, avamec�n in their recommended doses can be used. Dicofol 3 ml/l and difenthiuron (0.5 g/l) were found effec�ve even 7 days a�er spraying and that registered mortality of 84.78% and 79.15%, respec�vely. Galecron registered 90.8% mortality (Gupta et al., 1976).
Groundnut
Tetranychus ur�cae Koch.
� This mite a�acks groundnut in many groundnut growing areas of India. This mite heavily envelops the plants by web where dust par�cles adhere affec�ng normal physiological ac�vi�es of plants. The heavily a�acked plants look seriously unhealthy and may die.
Ravi and Pu�aswamy (1995) conducted histological and biochemical studies on three varie�es of groundnut genotypes Robut 33-1, TMV-2, ICGV-87160. Direct damage can be caused to hypodermis and sponge chlorenchyma and even in the palisade parenchyma. Robut 33-1, is a tolerant genotype having more leaflet thickness followed by moderately tolerant genotype TMV-2 and suscep�ble genotype ICGV-87160. Thickness in leaflet in tolerant genotype did not alter much a�er mite feeding. Reduc�on of thickness of leaflets was much in suscep�ble genotypes. Histochemical observa�ons showed change in number and size in starch grains and protein and RNA
177

contents in the mesophyll �ssues of three genotypes due to feeding of T. ur�cae. Suscep�ble genotypes had higher reduc�on in starch grains than moderately tolerant and tolerant genotypes indica�ng that size and number of starch grains impart resistance against T. ur�cae not much difference in protein and RNA contents was seen among the three genotypes.
Varietal preference:- Kateshia et al., (1999) performed varietal screening of 10 varie�es of groundnut. The variety TG-26 had least number of mites (9-12 mites/15 leaves), followed by GGT-4, ICGS-37, GG-3, TG-26 and ICGS-37 which were at par. The varie�es J-11, Kaderi-3, JL-24, ICGS-44 and GG-20 were moderately suscep�ble, variety GG-2 was most suscep�ble.
Tetranychus macfarlanei Baker and Pritchard
This mite has also been reported on groundnut and its feeding makes the leaves yellowish and then brownish. Subsequently, such leaves dry up and pod forma�on is affected.
Tetranychus hypogea Gupta
This mite was described from specimens collected on groundnut from West Bengal. All the infested leaves become heavily covered with webs and all those leaves turn yellow. The infested plants look sickly and growth is adversely affected (Gupta, 1976, 1985).
The following insect predators also recorded from these legumes
Insect Predator Wollastoniella sp. (Hemiptera: Anthocoridae) Oligota oviform (Coleoptera: Staphylinidae) Stethorus pauperculus (Coleoptera:Coccinellidae) Sceliothrips sexmaculatus (Thysanoptera:Thripidae)
The following predatory mites are poten�al natural enemies under field condi�ons
Insect Predator Amblyseius sp (Family-Phytoseiidae), Amblyseius finlandicus, Amblyseius alstonia (Family Phytoseiidae)
178
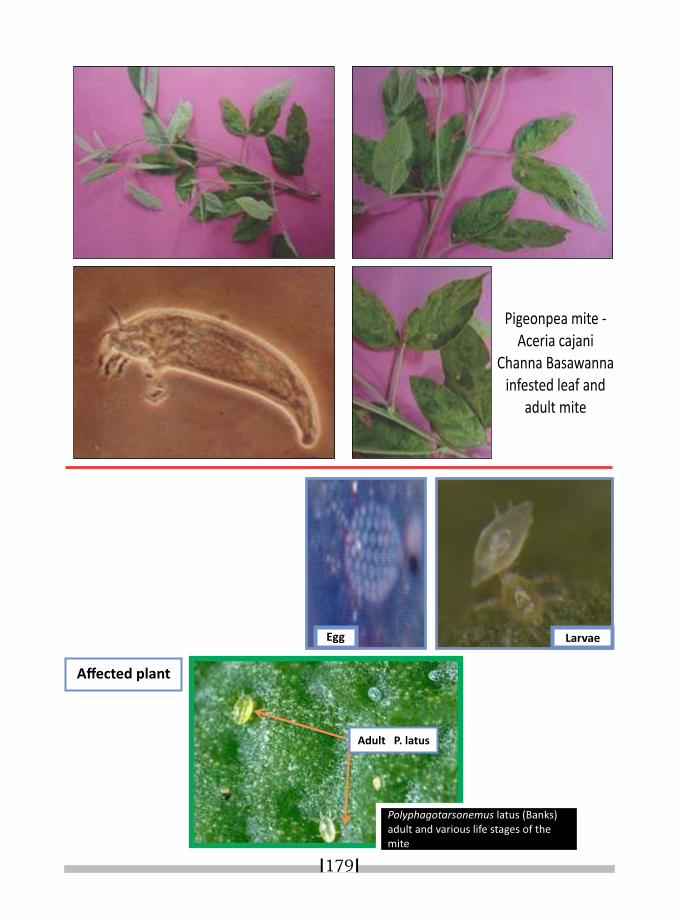
Pigeonpea mite - Aceria cajani
Channa Basawanna infested leaf and
adult mite
Polyphagotarsonemus latus (Banks)adult and various life stages of the mite
LarvaeEgg
Affected plant
Adult P. latus
179

Affected leaf
Ballooning
French bean (Rajma) Severely affected by the r e d s p i d e r m i t e Tetranychus urticae (Koch) The mite is active round the year on french bean crop
Ballooning
180

Pigeonpea, Schizotetranychus cajani Gupta
Groundnut -Teranychus urticae (Koch)
181

Photo courtesy - H.S. Gohani
Egg Grub
Pupa
190
Adult
Mite destroyer ladybird beetle, stethorus pauperculus Weise(Coccinellidae : Coleoptera) feed upon T. urticae and O. indicus (F: Tetranychidae). It is noticed in crop like sroghum, Surgarane, Baru Grass, Okra, Brinjal and marigold. The peak population was recorded in October.
182

DeutonymphProtonymph
LarvaEggs
Adult female
P h y t o s e i i d p r e d a t o r Amblyseius longispennis ( E v a n s ) a p o t e n t i a l predator of Tetranychus urticae.
With 1:10 and 2:20 raties, a l l t h e p r e y s w a s consumed with in three day. Generally it prefers eggs stage of T. urticae.
183
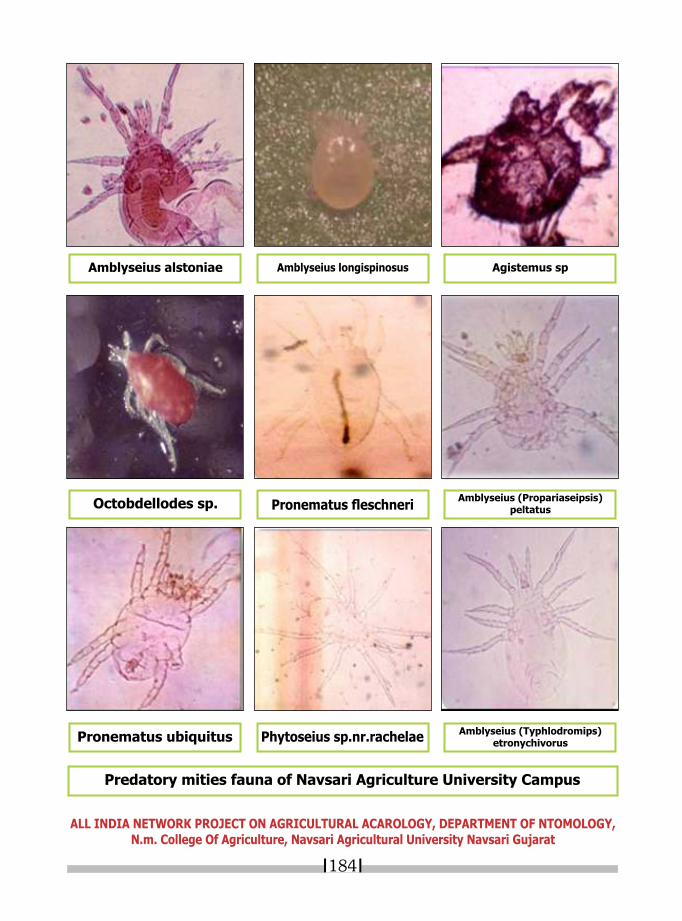
Predatory mities fauna of Navsari Agriculture University Campus
Amblyseius alstoniae Amblyseius longispinosus Agistemus sp
Octobdellodes sp. Pronematus eschneriAmblyseius (Propariaseipsis)
peltatus
Pronematus ubiquitus Phytoseius sp.nr.rachelae Amblyseius (Typhlodromips)
etronychivorus
ALL INDIA NETWORK PROJECT ON AGRICULTURAL ACAROLOGY, DEPARTMENT OF NTOMOLOGY,N.m. College Of Agriculture, Navsari Agricultural University Navsari Gujarat
184

Suggested Reading:
Anan. 2009. Annual Report of AINP on Agricultural Acarology (April 2007- March 2011). Dir. Of Research, BCKV, Kalyani, 1-80.
Arjunan, G., Swamiyappan, R. and Reddy, M.V. 1989. Studies on red gram sterility mosaic disease and its vector. Abst. Nat. Sem. Red gram, February 1989, IRS, Gulbarga, Karnataka.
Capoor, S.P. 1950. Report on pigeonpea sterility with symptoms and transmission. Proc.Ind. Sci. Congr., 22: 270-274.
Capoor, S.P. 1952. Observa�on on sterility disease of pigeonpea in Bombay.Ind. J. Agric. Sci.22: 270-274.
Channa Basavanna , G.P. and Viraktamath, C.A. 1989. (Eds.) Progress in Acarology Vol.-2. Oxford and IBH Pub., New Delhi.
Chillar, B.S., Gula�, R. and Bhatanagar, P. 2007. Agricultural Acarology. Daya Pub. House, Delhi. 1-355
Chinnaiah, C. and Mohanasundaram, M. 1995. Evalua�on of the efficacy of neem products and certain pes�cides against Aceria cajani on pigeonpea. Abst. G.B. Pant Univ. of Agric. And Tech., Pantnagar Nat. Symp. Acarology, Bangalore.
De, R. K., Dhar, V. and Rathore, Y.S. 1996. Control of mosaic in pigeonpea with insec�cides and acaricides. Indian J. Pulses Res., 9 (1): 87-89.
Ghanekar, A.M., Shiela, V,K., Beniwal, S.P.S., Reddy, M.V. and Nane, Y.L. 1992. Sterility mosaic of pigeonpea. In: Plant disease of Interna�onal Importance. Vol.- 1. (Disease of cereals and pulses). Pren�ce Hall Inc. Englewood Cliffs. New Jersey, USA: 415-428.
Gupta, S. K. 1985. Handbook-Plant mites of India, Zoological Survey of India, Calcu�a:10520.
Gupta, S. K., Dhooria, M.S. and Sidhu, A.S. 1976. Seasonal abundance of Tetranychus telarus (Linn.) on castor in the Punjab. Oilseeds Jour.6: 16-18.
Gupta, S. K., Dhooria, M.S. and Sandhu, G.S. 1973. Varietal suscep�bility of wheat to brown wheta mite, Petrobia lateens (Muller). Sci. & Cilt., 39: 312-314.
*Kalisekar and Thakur. 2007. J. acarol.17: 7-8.*Kateshia et al., (1999). J. Acarol., 14:47-48.
*Karuppnchamy et al., 1990. Legume Research, 13: 183-187.
Kannaiyan,J., Nene, Y.L.,Reddy, M.V., Ryan, J.G. and Raju, T.N. (1984). Prevalence of pigeonpea disease and associated crop losses in Asia, Africa and in the America,. Tropical pest Management, 30 (1): 62-71.
185

Malaviya, M.D. (1994). Incidence of different mite species on Indian bean with
emphasis to biology and control of Tetranychus ludeni (Zacher)
(Acari:Tetranychidae). M.Sc. (Agri.) thesis, GAU, S.K. Nagar, Gujarat, 210 pp.
Nane, Y.L. 1972. A survey of viral diseases of pulse crops in u�ar Pradesh.Res. Bull.,4:
G.B. Pant Univ. of Agric. And Tech., Pantnagar, u�ar Pradesh:1-191.Nane, Y.L. and Rathi, Y.P.S. 1972. Disease of (sterility mosaic and yellow mosaic) viral
disease of pulse crops in U�ar Pradesh. Final Tech. Rept. G.B. Pant Univ. of Agric. And Tech., Pantnagar: 151-162.
Nane, Y.L. and Redd, M.V. 1976. A new technique to screen pigeonpea for resistance to sterility mosaic. Tropical grain Legume Bull., 5:23.
Narayanaswamy, C.A. 2004. Studies on Aceria cajani Channabasavanna (Acari: Eriophyidae) mite vector of pigeonpea sterility mosaic disease with special reference to varietal screening and its interplant distribu�on. M.Sc. Agri. Thesis, UAS, Bangalore:1-90.
Natarajan, E. and Beevi, D. 1992. Control of Acaria cajani (Eriophyidae: Acarina) on Cajanas cajani (L) Millsp. Internat. Pigeonpea Newsl.,16 : 22-24.
Patel, J.R., Patel, C.B., Patel, M.B., Desai, H.R., Patel, J. and Rai, A.B. 1990. Some ecological studies on eriophyids mite, Acaria cajani infes�ng pigeonpea in south Gujarat, Abst. V Nat. Symp. Acarology, Bangalore: 29-30.
Prabhuswamy, H.P., Srinivasa, N., and Gupta, S.K. 1995. Popula�on fluctua�on of eriophyids mite Aceria cajani vector of pigeonpea sterility mosaic disease. Abst. V Nat. Symp. Acar. ,Bangalore:29.
Rathi, Y.P.S. 1983. Studies on sterility mosaic disease of pigeonpea (Cajanas cajan (L) Millsp.). FinalTech. Report G.B. Pant Univ. of Agri. And Tech., Pantnagar, U�ar Pradesh, India.
Ravi, K.C. and Pu�aswamy, 1995. Reac�on of some groundnut genotypes to Tetranychus ur�cae. Abst. V Nat. Symp. Acarology:38.
Reddy, M. V., Raju, T.N. and Lenne, J.M. 1998. Disease of pigeonpea. In: The pathology of food and pasture legumes (eds. D.J. Allen and J.M. Lenned). CAB Interna�onal ICRISAT:517-518.
Reddy, M.V., Nane, Y.L., Raju, T.N., Sheila, V.K., Kanaiyan, J. and Beniwal, S.P.S. 1989. Evalua�on of pigeonpea lines for resistance to wilt, sterility mosaic and phytopthora blight. Interna�onal Pigeonpea Newsle�er, 10: 22.
Reddy, M.V., Sheila, V.K. and Nane, Y.L. 1993. Cajanas scara baeoides an alternate host of pigeonpea sterility mosaic pathogen and its vector Aceria cajani. Internat. Pigeonpea Newsl., : 18-24.
Satpathi et al., 1992. Annals of Ento. 10:51-53.Seth, M.L. 1962. Transmission of pigeonpea sterility by a eriophyids mite. Indian
Phytopathology, 15:225-227. *only review seen.
186

IDENTIFICATION OF MITES ASSOCIATED WITH LEGUME CROPS
* ABHISHEK SHUKLA AND G.G. RADADIAAll India Network Project on Agricultural Acarology
Department of Entomology, N.M. College of AgricultureNavsari Agricultural University, Navsari-396 450, Gujarat
Family- Tetranychidae (Spider mites)
Introduc�on
Spider mites belong to the family Tetranychidae of the order Pros�gmata. They are so named because many members of this family produce silk webbing on host plants. Spider mites are without doubt the most important mites a�acking plants. Most spider mite species are polyphagous. They occur on virtually every major food crop and ornamental plant. They include some of the most important pests on economic plants. The Tetranychidae is a large family of worldwide distribu�on. Some 1,200 species of spider mites belonging to over 70 genera are known in the world and more are yet to be discovered, especially in the Southern Hemisphere. The family consists of two subfamilies: Bryobinae and Tetranychinae. Most pest species belong to the Tetranychinae.
Morphological characters
Spider mites are so�-bodied, medium-sized mites (about 400 mm for an average adult female). They are o�en red, green, orange or yellow in colour when alive. The gnathosoma has a capsule-like structure known as the stylophore, which is formed by the fusion of the cheliceral body. The movable digits of the chelicerae are very long, o�en whip-like and recurved proximally. They are well suited for piercing. A pair of s�gmata is located near the base of the chelicerae, where the peritremes arise. The palps are five-segmented. The palpal tarsus and �bia (with a distal claw-like seta known as the �bial claw) o�en form a thumb-claw complex. The tarsus o�en has an enlarged distal eupathidium (spinneret) in the Tetranychinae and this is used to spin webbing in many species. The size and shape of the spinneret is of taxonomic significance. The idiosoma is o�en covered with a striate cu�cle. The pa�ern of the stria�on and the shape/density of lobes distributed on the striae are useful diagnos�c characters. There are three or four pairs of normal setae in two rows (v1-2, sc1-2) and two pairs of eyes on the dorsal propodosoma. On the opisthosomal dorsum, there are five rows of setae: c, d, e, f and h. The number, loca�on, length and structure of dorsal setae are of taxonomic significance. Female genital pores are transverse and are bordered anteriorly by a genital flap and laterally by characteris�c cu�cular folds. The structures of the paired lateral claws and the medial empodium are of taxonomic importance. The claws may be
187

claw-like or pad-like with tenent hairs, and so is the empodium. Claws may bear dorsal or ventral hairs. The tarsi of legs I and II bear duplex setae (a long solenidion and a short normal tac�le seta with their bases joined together. The number of duplex setae and their posi�ons are of taxonomic significance. Wedge-shaped males are smaller than ovoid females and have a tapering opisthosoma. Males have a protrudable aedeagus, the shape of which is very important in species iden�fica�on.
Tarsonemid Mites
Tarsonemid mites belong to the family Tarsonemidae of the order Pros�gmata. No common names have been proposed for the family. Many tarsonemid species are fungivores, algivores and herbivores, and others are predators of other mites, parasites of insects and possibly symbionts of insects. Some phytophagous tarsonemids are pests of agricultural crops and are important in greenhouses. The Tarsonemidae is a large family of worldwide distribu�on. More than 500 species belonging to some 40 genera are known in the world and many others are yet to be discovered. The family consists of three subfamilies: Pseudotarsonemoidinae, Acarapinae and Tarsoneminae. Most of the described species are distributed in the Tarsoneminae, which includes two large genera Tarsonemus (over 270 species) and Steneotarsonemus (over 70 species).Most plant feeding species belong to a few genera in the Tarsoneminae, except Polyphagotarsonemus, which is a member of the Pseudotarsonemoidinae.
Morphological characters
Tarsonemid mites are small in size (100 to 300 µm). Most species are semitranslucent, pale or whi�sh, but the actual colour is affected by the food ingested. Some phytophagous species assume a greenish appearance when they feed on green leaves. Tarsonemids are dimorphic, with males differing markedly from females in both size and structure. Adult females are generally ovoid, but many species of Steneotarsonemus are elongate. Males are smaller than females, characterized by the subterminal genital capsule and strong leg IV with a single claw. The gnathosoma is capsulate. The movable digit of the chelicerae is a fine stylet. The reduced palps are closely appressed. Adult females have s�gmata but no peritremes; the opening of the s�gmata is on the anterior lateral dorsum of the propodosoma. Females have a pair of capitate trichobothria and two pairs of simple setae on dorsal propodosoma, whereas males have three or four pairs of simple setae. The dorsal surface of the opisthosoma is covered with a series of plates. The structure, length and loca�on of dorsal setae on the plates are used in classifica�on. Coxal apodemes are strongly developed. Leg IV of females are three-segmented and slender, termina�ng in a characteris�c long seta. Leg IV of males are generally four-segmented and strong, termina�ng in a claw (in some species the tarsus and �bia of leg IV are fused. Legs II and III usually terminate in a pair of claws and a membranous empodium.
188

Family Tenupalpidae (False Spider Mites)
The false spider mites, as the Tenuipalpidae are commonly known, also belong to the superfamily Tetranychoidea in the order Pros�gmata, as does the spider mite family Tetranychidae. They are not true spider mites because they do not produce silk webbing on plants. They are also known as flat mites because most species are dorsoventrally fla�ened. False spider mites are phytophagous. They are slow-moving and are usually found on the lower surface of the leaves near the midrib or veins. Some species feed on the bark. Others live in flower heads, under leaf sheaths or in galls. Only a small number of species belonging to a few genera have become pests of economic plants and they are most commonly found on tropical fruit crops and ornamental plants. The family Tenuipalpidae has some 800 described species belonging to over 25 genera, but many others are yet to be discovered and described. The family consists of three subfamilies, with most species belonging to the Brevipalpinae and Tenuipalpinae. Tenuipalpus and Brevipalpus are the two largest genera and also the economically most important ones.
Morphological characters
False spider mites are small mites. Most species range between 200 µm and 400 µmin body length andmany are orange to red in colourwhen alive. False spider mites have an elongate stylophore fused by cheliceral bases. Like spider mites, the cheliceral movable digits are long, styliform and recurved proximally. As the name suggests, the palps are simple and o�en reduced in size. They lack the very strong �bial claw in spider mites, and are comprised of one to five palpal segments. The adults usually have a fla�ened idiosoma which is ornamented dorsally with ridges and re�cula�on. There is an obvious sejugal furrow separa�ng the propodosoma and hysterosoma. There are two pairs of eyes and three pairs of setae on the dorsal propodosoma, which o�en has a bifid anteromedian lobe. The legs are short and wrinkled and some gall-forming genera have only three pairs. The legs have five free segments and terminate in paired claws and a pad-like empodium. Both claws and the empodium have tenent hairs
Family Eriophididae (Eriophyoid Mites, Gall mites)
Eriophyoid mites belong to the superfamily Eriophyoidea of the order Pros�gmata. These worm-like or fusiform mites are too small to be seen by the naked eye, but they cause many forms of plant abnormali�es such as galls, leaf blisters and rusts, which are very obvious and indica�ve of the presence of these mites. Most species are monophagous and many species are limited to plant species within a single genus, with few excep�ons. Most species cause li�le harm to their host plants. However, some rust mites and gall mites are important pests on economic plants. The Eriophyoidea is a large superfamily of worldwide distribu�on. Over 3,000 species belonging to over 250 genera are known in the world and ten of thousands of more
189

species are yet to be discovered. The superfamily consists of three families: Eriophyidae, Phytop�dae and Dip�lomiopidae. About three-quarters of the described species of the superfamily belong to the Eriophyidae.
Morphological characters
Eriophyoid mites are �ny annulate mites ranging in size from 90-350 µm. They are worm-like or fusiform and are o�en pale yellowish to brownish in colour when alive. The gnathosoma is modified as a median rostrum enclosed by indis�nc�vely segmented lateral palpi. The movable digits of the chelicerae are whip-like and lie in a dorsal groove of the palpal bases; they are very long in the Dip�lomiopidae and rela�vely short in the Eriophyidae and Phytop�dae. There are no eyes on the prodorsum. S�gmata and peritremes are also absent. The propodosoma o�en bears a dorsal shield with up to five setae. The dorsal shield may have a median line, a pair of admedian lines and a pair of submedian lines. The subcapicular setae, when present, may each arise from a tubercle. The hysterosoma has a maximum of nine pairs of setae. The genital pore in the female is transverse and located behind coxae II, with an anteriorly hinged flap. There are only two pairs of legs on the propodosoma. Both pairs of legs terminate in a feather-like empodium and are without true claws.
BENEFICIAL MITES
Phytoseiid mites
Phytoseiid mites belong to the family Phytoseiidae of the order Mesos�gmata. Phytoseiid mites are predators of spider mites and other small mites and insects on plants. Some species also feed on nematodes, fungal spores, pollen and exudates from plants, but rarely plant �ssue. Several members of this family are of great importance in the biological control of spider mites and thrips in greenhouse crop produc�on. The Phytoseiidae is a large family of worldwide distribu�on. More than 1,600 species belonging to over 70 genera are known in the world. The family consists of three subfamilies: Amblyseiinae, Phytoseiinae and Typhlodrominae. Effec�ve biocontrol agents occur in all three subfamilies, although most commercially available species that are commonly used in greenhouses belong to the genera Neoseiulus and Phytoseiulus in the Amblyseiinae.
Morphological characters
Phytoseiids are medium-sized mites (mostly 250-400 mm) with rela�vely long legs. Many species are pale, yellowish, orange or dark brown. The idiosoma is covered with a single en�re shield (rarely two shields) with no more than 24 pairs of setae (dorsal setae J1, J3 and J4 are absent). The s�gmata open ventrolaterally at the level between the coxae of leg III and leg IV. The sternal shield of the female has three pairs of setae. A pair of metasternal shields is small, each bearing one seta. The female genital
190

shield is truncated posteriorly. The male genital opening is on the anterior margin of the sternal shield. The male chelicera bears a spermatodactyl, the shape of which has diagnos�c value. Leg I terminates in well-developed ambulacra. Tibia I has five dorsal and two or three ventral setae. The distal leg segments o�en have one elongate or differently-shaped macroseta.
Brevipalpus phoenicisTetranychus urticae
Polyphagotarsonemus latus. Aceria cajani
Neaseiulus longispinosus a potential Predator of phytophagous mite
191

STRATEGIES TO UNDERSTAND INSECT RESISTANCE IN PLANTS THROUGH THE RICE-GALL MIDGE INTERACTION
SURESH NAIRInterna�onal Centre for Gene�c Engineering and Biotechnology (ICGEB)
Aruna Asaf Ali Marg New Delhi 110 067, India
The rice gall midge (Orseolia oryzae Wood-Mason; Diptera) is a major pest of rice and is responsible for severe yield loss. Host plant resistance is an effec�ve, cost-efficient and environment-friendly way to control many pests. However, the challenge is the constant development of new gall midge biotypes that overcome the deployed resistance genes in rice hosts. To make the deployed resistance genes durable, there is an urgent need to understand: (a) how new biotypes emerge, and, (b) how host resistance genes impart resistance against the pest.
In an effort to understand plant-insect interac�on in general and the rice-gall midge interac�on in par�cular, we are studying secreted salivary gland proteins (SSGPs) of the rice gall midge, as we believe it has a major role to play in the interac�on between the pest and its host, the rice plant. Towards this direc�on we have, for the first �me, successfully PCR amplified genes coding for SSGPs from the rice gall midge with the help of a cDNA library prepared from mRNA isolated from the salivary gland-enriched �ssue. We have successfully cloned cDNAs encoding different rice gall midge SSGPs. To study this interac�on, such an approach has never been a�empted before and the successful isola�on of full-length SSGPs will lead to a be�er understanding of the molecular basis that governs rice-gall midge interac�on. We have also determined the genomic organiza�on of some of these genes. Real-�me quan�ta�ve PCR analyses have provided us, for the first �me, a glimpse into the expression pa�ern of the respec�ve genes, coding for various SSGPs, in the various stages of the life cycle of this pest. We have also observed differen�al expression of these genes depending on whether the gall midge is feeding on a compa�ble or incompa�ble rice host. For example, �ssue-specific analysis of the expression of gamma subunit of oligosaccharyl transferase (OoOST) revealed an increase (> sevenfold) in the transcript levels in the salivary glands of maggots in suscep�ble plants when compared with the transcript level in the salivary glands of maggots feeding on resistant hosts. Using quan�ta�ve PCR, performed on different developmental stages of the maggots in two suscep�ble and two resistant hosts, we observed similar expression pa�erns (i.e. overexpression in the compa�ble interac�on). These results indicate the involvement of OoOST in maggot survival and establishment in the suscep�ble host. Our studies have also revealed that, during the interac�on, these genes have higher expression in the salivary glands than in the mid-gut and fat bodies �ssues. In this study, we have also studied other SSGPs including lipase.
192

We have overexpressed some of the gall midge SSGPs, using an E. coli-based heterologous expression system, and purified these to homogeneity. Rice hosts when injected with recombinant insect SSGPs showed �ssue necrosis -- typical of a hypersensi�ve reac�on observed in resistant hosts a�er a gall midge a�ack. We have also shown that another SSGP, Orseolia oryzae nucleoside diphosphate kinase (OoNDPK), is overexpressed in larvae feeding on suscep�ble hosts. In addi�on, heterologously expressed OoNDPK is capable of causing coleop�le cell elonga�on in rice hosts. Thus, it is for the first �me that it has being demonstrated that an insect pest protein is capable of influencing the physiology of the host in such a manner. Furthermore, isola�on of complete genes encoding the other SSGPs will provide an alternate and complementary approach to the isola�on of gall midge resistance genes from the rice host. Besides, informa�on on the interac�on between the host and its insect pest will allow this knowledge to be put to use to device be�er strategies to breed new rice varie�es with durable resistance to gall midge.
We have also carried out pyrosequencing-based transcriptome analysis of the Asian rice gall midge and our results revealed differen�al response of the gall midge depending on whether it is in a compa�ble or incompa�ble interac�on with its host. All the transcripts iden�fied from this study have been assigned Gene Ontology (GO) terms separately (Biological proper�es; Cellular components; Molecular func�on). Transcripts involved in Pseudouridine synthesis were observed only in maggots from resistant hosts. Also, transcripts of protein phosphoryla�on and proteolysis were rela�vely abundant in these maggots. Further, this set had abundance of genes involved in protein serine-threonine kinase and serine type endopep�dase ac�vity. When the maggots' a�empt to infest the host fails, it leads to events of proteolysis finally resul�ng in maggot mortality. In contrast, compara�vely higher number of transcripts involved in transla�on, regula�on of transcrip�on and electron transport chain were observed in maggots feeding on the suscep�ble rice variety. Also, transcripts encoding structural cons�tuents of ribosome, nucleic acid binding, ribosome, ribonucleoprotein complex and cytoplasm were more represented in this set. Presence of these transcripts suggests that in compa�ble interac�on the maggots successfully orchestrate the plant metabolic machinery and subvert the plant cell's natural func�ons for their own ends. However, this will need to be validated through quan�ta�ve real �me PCR. Future studies directed towards func�onal valida�on of differen�ally expressed genes of target pathways would decipher the mechanism of insect virulence. Also, the molecular markers generated from the study will enable development of diagnos�c tools for biotype differen�a�on and popula�on variability analysis. Furthermore, these studies will provide major molecular resource for both plant biologists and entomologists to understand plant-insect interac�on, which in turn, will help to develop be�er-integrated pest management strategies.
193

In addi�on, using suppressive subtrac�on hybridiza�on (SSH) cDNA libraries and microarray analysis, we show differen�al response of genes from gall midge-resistant and -suscep�ble rice varie�es to gall midge a�ack. Our results suggested involvement of at least three genes viz., NBS-LRR, Phenylalanine ammonia lyase (two ESTs each) and
(+)OsPR10a in HR type gall midge resistance in Suraksha. The study also suggested suscep�bility-related gene expression changes in rice variety TN1. These involved upregula�on of a number of genes of primary metabolism and two specific genes i.e. TCTP and NAC transcrip�on factor. Furthermore, the ini�al molecular clues into the rice-gall midge interac�on and access to the relevant transcriptome results can provide a pla�orm for gene discovery and help elucidate the resistance mechanism in Suraksha and suscep�bility mechanism in TN1. In future, we also propose to validate the role of
(+)SA-mediated defense pathway in HR type rice-gall midge resistance using a biochemical and proteomics approach. Analysis of the differen�ally expressed genes in the two interac�ons also revealed a marked similarity between rice-gall midge and plant-pathogen interac�on reitera�ng the overlap of global plant response to pest or pathogen a�ack. For iden�fying candidate resistance genes underlying insect resistance, our results further open up the line of thought that pathogen resistance genes as valid candidate genes for insect resistance. There is good poten�al that such genes, highlighted through our compara�ve transcriptome studies, could be tested as valid candidate resistance genes through transgenic approaches.
In another study, analysis of rice SSH cDNA libraries revealed up-regula�on of genes related to primary metabolism, nutrient reloca�on, cell organiza�on and DNA synthesis. Concomitantly, defense, secondary metabolism and signaling genes were suppressed. Further, real-�me PCR valida�on of a selected set of 20 genes, in three suscep�ble rice varie�es (TN1, Kavya and Suraksha) during the interac�on with the respec�ve virulent gall midge biotypes, also revealed varia�on in gene expression in Kavya as compared to TN1 and Suraksha. These studies showed that virulent insects induced the plants to step up metabolism and transport nutrients to their feeding site and suppressed defense responses. But Kavya rice mounted an elevated defense response during early hours of virulent gall midge infesta�on, which was over-powered later, resul�ng in host plant suscep�bility.
Currently, we are sequencing the mitogenome of the rice gall midge to get a be�er understanding of the popula�on structure of the rice gall midge and its various biotypes and also the evolu�onary pa�ern of this insect. Detailed gene�c informa�on on this important rice pest may help in the development of methods for its control, preven�on and biotype monitoring. Besides, certain regions of the mitogenome could be used to develop PCR-based diagnos�c markers for iden�fying gall midge biotypes. This will immensely help in integrated pest management for it can serve as an early warning system for the occurrence/arrival of specific gall midge biotypes in specific rice growing areas. And, this informa�on can also be used to thwart the pest a�ack by the �mely
194

deployment of rice varie�es carrying appropriate gall midge resistance gene(s). Furthermore, these markers can be used to con�nuously monitor the gall midge popula�on in a par�cular area and any change in the popula�on, as revealed by the markers, can be immediately used as a warning to use or deploy alternate resistance genes based on the occurrence of new biotypes. Addi�onally, we will also be studying the role of miRNAs in this interac�on. Based on leads obtained from the SSH studies we are carrying out metabolomics-based studies to understand the changes, in primary and secondary metabolites, taking place within the host, post gall midge infesta�on/challenge, in an effort to get a be�er understanding of gall midge-rice interac�on in par�cular and insect-plant interac�on in general.
Suggested Reading
Harris, M. O., Stuart, J. J., Mohan, M., Nair, S., Lamb, R. J. and Rohfritsch, O. (2003) Grasses and gall midges: Plant defense and insect adapta�on. Annual Review of Entomology 48: 549-577.
Rawat, N., Himabindu, K., Neeraja, C. N., Nair, S. and Bentur, J. S. (2013) Suppressive subtrac�on hybridiza�on reveals that rice gall midge a�ack elicits plant-pathogen-like responses in rice. Plant Physiology and Biochemistry 63 : 122-130.
Rawat, N., Neeraja, C. N., Nair, S. and Bentur, J. S. (2012) Differen�al gene expression in gall midge suscep�ble rice genotypes revealed by suppressive subtrac�on hybridiza�on (SSH) cDNA libraries and microarray analysis. Rice 5: 8 (DOI: 10.1186/1939-8433-5-8).
Rawat, N., Neeraja, C. N., Sundaram, R. M., Nair, S. and Bentur, J. S. (2012) A novel mechanism of gall midge resistance in the rice variety Kavya revealed by microarray analysis. Func�onal and Integra�ve Genomics 12: 249–264.
Sinha D.K., Bentur J.S., Nair S. (2011) Compa�ble interac�on with its rice host leads to enhanced expression of the gamma subunit of oligosaccharyl transferase in the Asian rice gall midge, Orseolia oryzae. Insect Molecular Biology 20: 567-575.
Sinha D.K., Lakshmi M., Anuradha G., Rahman S.J., Siddiq E.A., Bentur J.S., Nair S. (2011) Serine proteases-like genes in the Asian rice gall midge show differen�al expression in compa�ble and incompa�ble interac�ons with rice. Interna�onal Journal of Molecular Sciences 12: 2842-2852.
Sinha, D. K., Atray, I., Bentur, J. S. and Nair, S. (2012) Expression of Orseolia oryzae nucleoside diphosphate kinase (OoNDPK) is enhanced in rice gall midge feeding on suscep�ble rice hosts and its over-expression leads to salt tolerance in Escherichia coli. Insect Molecular Biology 21: 593–603.
Sinha, D. K., Nagaraju, J., Tomar, A., Bentur, J. S. and Nair, S. (2012) Pyrosequencing-based transcriptome analysis of the Asian rice gall midge reveals differen�al response during compa�ble and incompa�ble interac�on. Interna�onal Journal of Molecular Sciences 13: 13079-13103.
195

MANAGEMENT OF INSECT VECTORS IN MUNGBEAN AND URDBEAN
RAVINDER SINGH GURWARADepartment of Entomology
PAU, Ludhiana
Introduc�on
Mungbean (Vigna radiata (L.) Wilczek) and urdbean (Vigna mungo (L.) Hepper) are the most important pulse crops in India. Besides India, these are widely cul�vated throughout Southern Asia like Pakistan, Sri Lanka, Bangladesh, Thailand, Laos, Vietnam, Indonasia, China and Taiwan. In India, these crops are extensively grown in U�ar Pradesh, Madhya Pradesh, Rajasthan, Maharashtra, Orissa, Karnataka, Andhra Pradesh, Gujrat, Bihar, Haryana and Delhi during Kharif and Zaid season. These crops suffer from a large number of bio�c and abio�c stresses. Among bio�c stresses, insect pests are most important yield reducing factor. The average loss due to insect pests in mungbean and urdbean crops were es�mated to be 34.7 and 28.7% in different states of India (Asthana et al 1997). Lal and Ahmad (2002) reported nearly 60 insect species on mungbean and urdbean. Among these, 34 insects were serious pests on one or more of these pulse crops. Insect vectors, carrier of various viral diseases play an important role in transmission of pathogen from diseased to healthy plant . Vectors may be insects (whitefly, aphids, lea�oppers, mealy bugs, psyllids, membracids, beetles, thrips and flies), mites, nematodes and fungi (Honda and Iwaki 1991).
Types of virus – vector rela�onships
Virus can be divided into three groups depending upon various types of virus vector rela�onship. These are :
1. � Non-persistent type (Stylet borne) : Acquired and inoculated by vectors during short epidermal probes and pathogens are retained by vector only for few minutes/hours (e.g. Aphid born pathogens).
2. � Semi-persistent type (Circula�ve) : Acquired and inoculated by vectors during prolonged probes and circulate in the body. Latent period of 1-2 days in vector. Virus remains in vector throughout the life (e.g. whitefly born pathogens).
3.� Persistent type (Propaga�ve) : Circula�ve type of pathogens but these pathogens require slightly longer latent period (1-4 weeks) in vector. These pathogens mul�ply in vector (e.g. Lea�opper-born pathogens).
In mungbean and urdbean important vectors which transmit various viral diseases like mungbean yellow mosaic , leaf crinkle, mosaic mo�le and leaf curl are whitefly, aphids, mites and thrips.
196

WHITEFLY Bemisia tabaci (Gennadius)
It occurs throughout the northern and western regions of India. It is a serious pest of mung and mash. It also feeds on new crops like ricebean, soybean, arhar, co�on, cabbage, cauliflower, rapeseed and mustard, melon, potato, brinjal, okra, ornamental plants and some weeds, etc.
Nature of damage: It is a sucking pest and generally causes damage by sucking the sap from leaves and tender parts of the host plants. In case of grain legumes like mung, mash and soybean, it damages the plants in three ways: (i) sucking of cell sap from leaves, thus lowering the vitality of the plants. Under high popula�on pressure of the pest, the plant becomes very weak, giving a sickly look and may die. (ii) The insect excretes honey dew, on which growth of sooty mould takes place which result in blackening of leaves, resul�ng in decrease in photosynthesis. In severe a�acks there is total blackening of the crop, drying of leaves and total failure of the crop. (iii) The whitefly is a vector for a number of viral diseases especially mungbean yellow mosaic virus (MYMV).
Biology: The insect breeds throughout the year and all the developmental stages are no�ced. During winter season generally the adults are predominant. The female lays eggs singly on lower side of the leaves, averaging 120 eggs per female. The eggs are light yellow first, turning brown later on. The eggs hatch in 3-5, 5-17 and 33 days in April-September, October- November and December- January, respec�vely. The nymphs soon a�er emergence fix their mouth parts into the plant �ssues and feed on the cell sap. They grow into three stages to form the pupae within a period of 9-14 and 17-81 days in April –September and October-March, respec�vely. The pupal period generally lasts for 2-8 days. The total life cycle is completed in 14-122 days and there are 11-12 genera�ons of the insect in a year.
BLACK APHID Aphis craccivora Koch
It is found all over the world and has large host range including cul�vated crops, fruit trees, vegetables and ornamental plants. Among the legumes it damages beans, mungbean, urdbean, chickpea, clover, len�l, peas, cowpea, etc. (Hariri 1981; Lal 1992).
Nature and Extent of Damage
A. craccivora damages the plant by sucking the sap from tender parts, i.e., top shoots, flowers, buds, developing pods and usually from the underside of the leaves. Due to its feeding, there is heavy drain of vital plant sap and the deple�on of assimilates. As a result the rate of transpira�on is increased (van Emden 1973). The leaves, shoots and flowers deform and do not develop properly. The pods become shrivelled and deformed. The insect secretes honey dew, due to which black mould develops on infested plants, which interferes in photosynthesis. Under favourable condi�ons the damage caused is very high and there is substan�al reduc�on in yield (Thakur et al. 1984; Lal 1992; Ahmad and Lal 1998). Aphis. craccivora is also a potent vector of some viral diseases. In len�l it transmits tobacco streak virus and cucumber mosaic virus. The insect prefers dry weather condi�ons, therefore, the infesta�on is high when winter rainfall fails or is low (Lal 1992). The pest is vector of pea leaf roll virus that causes chickpea stunt disease. This disease is very damaging in some areas of India, West Asia and Northern Africa. Besides this, aphid is vector of alfalfa mosaic, bean common mosaic, urdbean leaf crinkle and bean yellow mosaic disease (Nene and Reddy 1976).
197

Biology and Ecology
The biology, host preferences and bionomics of A. craccivora have been studied extensively
(Dorges et al. 1966; Bakhe�a and Sidhu 1977). Detailed biology of the pest was studied by
Srikanth and Lakkundi (1988) on mungbean and urdbean at Bangalore, India. Fecundity and
development vary to a great extent with host plant, soil fer�lity, soil moisture and temperature.
Drought helps in mass mul�plica�on and spread of aphids over large areas. The adults are
small-sized insects, about 2 mm long, pear-shaped, green or greenish-black in colour. They have
black bodies with prominent cauda and brown to yellow apendages. Males are rare. The
common mode of reproduc�on is vivipary and parthenogenesis. A single gravid adult aphid is
observed reproducing 17431 numbers in 15 days. The apterous (wingless) females are shining
dark brown or black while the alate (winged) forms are greenish-black with transparent wings.
The colour of newly laid nymphs is translucent except the reddish-brown compound eye. A�er
a couple of minutes the transparency of body gradually disappears and changes to light yellow.
A�er about half an hour the nymphs start moving and within an hour start feeding. There are
generally four nymphal instars and the dura�on of each instar is 1-3 days. The apterous female
starts laying the young ones within 24 hours of a�aining that stage. A female can reproduce up
to a maximum of 12 days. Breeding occurs almost throughout the year on one crop or the other
and both alate and apterous forms are present (Chhabra et al. 1986; Ahmad and Lal 1998). A life
cycle may take 6 to 8 days and an adult can lay 2 or 3 nymphs per day.
BEAN MITE Polyphagotarsonemus latus (Banks)
The bean mite has assumed a pest status on a number of crops and fruit trees throughout India.
The pest has been recorded on mungbean, urdbean, common bean, horse gram, cowpea,
potato, co�on, sesame, citrus, hollyhock, marigold, tea, weeds and ornamental plants (Gupta
and Dhooria 1973; Dhooria 1984; David 1991; Singh 1996).
Nature of damage: Both the adults and nymphs suck the cell-sap from the leaves. In case of
serious infesta�on, it causes downward curling of leaves and necrosis of young leaves and
flowers. Ini�ally the infested leaves appear pale to light brown. Severely infested leaves first
turn light brown to brown and then to dark reddish brown. Beniwal and Chaudhary (1979)
recorded bean mite as a vector of viral disease mosaic mo�le in mungbean and urdbean
Biology: Adult mites are hyaline in colour, oval in shape and have four pairs of legs. The females
are larger than the males and are broader at abdominal end. The eggs are laid on the ventral
surface of the new leaves in the depressions and are firmly s�ck to the plant substrate. The eggs
are oval, elongate, covered with whi�sh longitudinal rows of tubercles with flat base and are
transparent shiny. Dhooria (1984) reported that Egg stage lasts for 1-3 days. Hatched out larvae
have 3 pairs of legs and resemble the adults. The larvae are pear-shaped, milky-white with a
dorsal white stripe in the middle region. Larval period ranges from 3 to11 days. Under op�mal
condi�ons a complete life cycle can be completed in about one week and 20-30 genera�ons
may actually be raised under tropical and subtropical regions. Various clima�c factors such as
temperature, rela�ve humidity and precipita�on affect field popula�ons of mite. Mite
popula�on is low during winter due to a combina�on of low temperature and heavy rains
(Gerson 1992).
198

BEAN THRIPS Frankliniella schultzei
It is distributed in Japan, Korea and throughout India. The important host plants include mungbean, urdbean, pigeonpea, cowpea, beans, peas, sunhemp, groundnut, Medicago sa�va, Dolichos lablab, etc. (Kooner et al. 1983; Chakraborty and Du�a 1999; Irulandi and Balasubramanian 2000).
Nature and Extent of Damage
Thrips can a�ack green beans from the moment the seedlings emerge from the ground through to flowering,but it is the damage some do during flowering that has the greatest impact. The feeding apparatus of thrips is unique amongst insects. Thrips have only one mandible, which they use to puncture the plant surface before inser�ng their stylets. Using their stylets thrips suck the sap from the damaged cells, inducing a range of symptoms. Silvering is the most common symptom. It is readily seen on leaf �ssue and occurs as a result of the cell contents being removed. Although of concern, leaf silvering does not cause significant yield reduc�on. Significant reduc�on in saleable yield does occur when thrips damage green bean pods. Scarring of the pods causes twis�ng and curling to varying degrees as well as mild rough patches on the skin. Kooner et al. (1983) reported that the thrips are found in flowers and due to their damage the flowers shed before opening. Infested inflorescence becomes abnormal, showing symptoms of flower drop. Even if any such flower opens and pod is formed, that is abnormal, the grains in the matured pods are shrivelled and small in size. Morphology of such plants is en�rely different from that of the healthy ones. Regenera�on of vegeta�ve growth of damaged plants occurs on axillary buds, the new leaves remain diminu�ve and consequently the plants a�ain a bushy growth. The crop looks dark green in colour. In case of severe incidence there is 100% loss in grain yield. Chhabra and Kooner (1985a) es�mated yield losses up to 54.3% and deteriora�on in grain quality by this pest. Thrips are known vectors of tospoviruses such as tomato spo�ed wilt virus (TSWV) causing leaf curl in mungbean and urdbean.(Amin et al 1985)
Biology and Ecology
Thrips are very small, dark brown insects. The body of the insect is quite shining. Adult thrips are small, cylindrical or cigar-shaped insects up to 2 mm long and range in colour from pale yellow to black. They have two pairs of narrow wings that are fringed with long hairs and rest along the length of their back. The young nymphs are similar in shape to the adults but smaller and wingless, and usually pale yellow to almost white. The life cycle of thrips includes an egg, two nymphal stages that ac�vely feed, followed by two non-feeding stages, a pre-pupal and pupal stage, which generally takes place in the soil or leaf li�er. The emerging adults are generally winged, but depending on the sex and species, some have short wings and others are wingless. Most thrips insert their eggs into plant �ssue. The length of the life cycle depends on the temperature and the quality of the food source. In warm condi�ons, around 30°C, the life cycle can be completed in as li�le as 10–12 days. At 20°C the life cycle takes around 19 days (Persley and Sharman 2007).
Important diseases transmi�ed by vectors and their symptoms :
Mungbean yellow mosaic
Among various viral diseases transmi�ed by insect vectors, mungbean yellow mosaic caused by mungbean yellow mosaic virus, a member of germinivirus group transmi�ed through whitefly (Bemisia tabaci Gen.), is a very severe disease. Yield losses due to this disease vary from 5 to 100
199

per cent depending upon disease severity, suscep�bility of cul�vars and popula�on of whitefly (Nene, 1972, Singh, 1980). The infec�on not only dras�cally reduces yield but also severely impairs the grain size and quality. The affected plant showed typical symptoms of mild sca�ered yellow spots on young leaves. The next trifoliate leaf emerging from the growing apex showed irregular alterna�ng yellow and green patches. The leaves showed slight puckering with reduc�on in size. The size of yellowing areas increased further resul�ng with complete yellow of apical leaves. The infected plants usually mature late and bear very few flowers and pods. Chenulu et al 1984 observed some facts on whitefly transmission of MYMV that (i) the virus may be transmi�ed by a single whitefly but infec�vity is greater with a group of 10 to 15 whiteflies. (ii) the virus may be acquired by the whitefly from infected mungbean or urdbean plant within ten minute feeding period but transmission will be higher with one to two hour feeding period, (iii) following its acquisi�on by the whitefly, the virus can be transmi�ed a�er 2 to 4 hours incuba�on period, (iv) the virus can be transmi�ed to the mungbean plant within 30-minute inocula�on feeding period, although the percentage of transmission will be increased if the feeding period is extended to one hour,(v) whiteflies retain infec�vity for 3 to 10 days, (vi) transovarial transmission does not occur, (vii) virus acquired during the hyphal stage by feeding on infected plants may be transmi�ed and (viii) whitefly adults can acquire the virus from a mungbean plant 1 to 3 days before the mungbean plants exhibits symptoms. Varia�ons in acquisi�on and inocula�on �me among varie�es may be due to the concentra�on of the virus in the host �ssue, occurrence of virus inhibitors in the host or host palatability for the virus.
Leaf crinkle
The disease was first no�ced on urdbean in the States of U�ar Pradesh and Delhi in 1966 and in 1967 in Tarai region of U�ar Pradesh. It has also been observed on mungbean. It has also been reported from states of Haryana, Himachal Pradesh, Andhra Pradesh and Madhya Pradesh. Leaf crinkle virus reduces the plant height, root length, nodula�on, pods per plant, length of pod, seeds per pod and seed weight. Reddy and Krishnaiah (1983) observed yield losses of 41.1 to 69.3% in different cul�vars of urdbean. Symptoms of the disease appear as extreme crinkling, curling, puckering and rugosity of leaves, stun�ng of plants and malforma�on of floral organs. Enlargement in size followed by crinkled surfaces of lamina are the characteris�c symptoms on affected trifoliates. Pollen fer�lity and pod forma�on is severely reduced on infected plants which decreased the yield dras�cally. � The ULCV is transmi�ed through sap inocula�on, gra�ing, insect vectors and seeds. It has been reported to be seed transmi�ed to an extent of 18.4% in urdbean and upto 15% in mungbean (Beniwal et al 1980). Many workers have reported its transmission by various insects viz. aphids (Aphis craccivora, A. gossypii, Acrythosiphon pisum, Myzus persicae), whitefly (Bemisia tabaci), Epilachnua beetle (Henosepilachna dodecas�gma (Bharathan and Beniwal, 1984). ULCV infected seeds when stored for one year lost their intensity by 50% at room temperature. Incuba�on period of the disease varied from 36 days in August to only 24 days in June-July, indica�ng that warmer and humid condi�ons favour the transmission of the virus (Brar and Rataul, 1986).
Mosaic Mo�le
The mosaic mo�le of urdbean and mungbean is quite common in India. The disease also occurs in Iran and Indonesia where it is described under the name of mungbean mosaic virus (MMV). The virus infects urdbean as well as mungbean and occurs in several South-East Asian countries including Philippines and Thailand (Tsuchizaki et al 1986).
200

The disease is reported to cause as high as 71.5% yield loss in some suscep�ble varie�es of urdbean showing symptoms at first trifoliate leaf stage. The reduc�on in yield is contributed mainly by reduced number of pods per plant and seed weight. The inflorescence is converted into leafy structures with severe infec�on resul�ng in a complete seed loss. Urdbean is more suscep�ble than mungbean. The disease first appears in the form of irregular light-green patches alterna�ng with normal green areas. The size of the leaf gets reduced and the margins show upward rolling. Later on, these young leaves show puckering and blistering and the normal green areas are seen on both sides of the larger veins. The leaves become rough and bri�le. Affected plants are stunted and o�en display excessive branching. In severe infec�on, complete inflorescence is changed into a leafy structure. The virus is transmi�ed both by spa and seed. It is seed transmi�ed in urdbean and mungbean to an extent of 3-12% and 8-32%, respec�vely. Singh and Nene (1978) also reported its transmission by aphis, Aphis craccivora and A. gossypii. It is transmi�ed by tarsonemid mite, Polyphagotarsonemus latus which is the first record of transmission of a plant virus by this mite (Beniwal and Chaubey, 1979).
Leaf Curl
In India, the leaf curl of mungbean and urdbean was first observed in 1968 from Pantnagar in U�ar Pradesh. The disease has also been reported from Andhra Pradesh and Delhi. Among the viral diseases recorded on mungbean and urdbean, leaf curl is a poten�al killer of the plants but meager informa�on is available on this disease. Majority of the plants showing symptoms within 5 weeks of sowing died due to top necrosis. Plants which escape the death remain stunted and fail to flower. Even late infec�on reduced number of pods which either produced smaller or larger seeds or no seeds at all. Seed produced by diseased plants are unfit both for sowing as well as consump�on purposes. Thus, every plant affected by the disease at any stage of crop growth becomes economically useless and extent of such affected plants may go as high as 25 and 42% in selec�ons of urdbean and mungbean, respec�vely. The ini�al symptoms appear on the youngest leaves as chlorosis around some lateral veins and their branches near the margin. The leaves show downward curling of margin. Some leaves may show twis�ng. The diseased plants show reddish-brown discolorua�on on the under surface which also extends to the pe�ole. Plants showing symptoms within 15 weeks a�er sowing invariably remain stunted and majority of these die due to top necrosis within one or two weeks. The virus is transmi�ed by sap, gra�ing and the thrip, Frankliniella schultzei (Amin et al 1985).
Management of vectors
Various methods are used for the control of the vectors. Some concern the elimina�on of the vector, but many others, the preven�on of infec�on by changing cultural prac�ces or areas of crop produc�on. Methods have to be applied successively or at the same �me and even then it may be difficult to completely stop the spread of virus. Great progress has been made in insect control by the development of several potent insec�cides. Although they provide good control in many cases but good control in all the vectors and all the crops is many �mes not possible. In such cases a number of methods of disease control are required which must be applied in combina�on with one another. So that maximum control of the vector and the disease is achieved through a proper strategy.
201

Management for whitefly
Cultural control
Ravindrababu (1987) reported that maize, sorghum and pearl millet when grown as barrier
crops for mung bean, significantly reduced the popula�on of B. tabaci and incidence of MYMV
and resulted in higher yield. Intercropping rice bean with sorghum had a suppressing effect on
the incidence of whitefly on rice bean (Satyanarayana et al.1997). Manipula�on in date of
sowing may help in minimizing the whitefly popula�on and thus reducing the disease incidence
but it may greatly differ from region to region. The incidence of MYMV is reduced in plots with
inter-varietal intercropping of mungbean and urdbean (Das Gupta and Chowdhury, 1985).
Biological control
Nene (1973) reported nuclear polyhedrosis virus (NPV) and a fungal pathogen, Poecilomyces
fairnosus, as biocontrol agents of adult whitefly. Lal (1987) reported Prospaltella flava and
Geocaris tricolor parasi�sing nymphal, pupal and immature stages. He further reported Brumus
sp. as a predator of immature stages of the pest.
Host plant resistance�
Many genotypes of mung bean and urd bean were screened for resistance to whitefly and
jassids by Kooner (1998), Kooner and Cheema (2007a, b), where genotypes ML 508, ML 537,
ML 1265, ML 1229, ML 1260, ML 1256 and ML 1191 of mungbean and were iden�fied as
resistant. Chhabra et al. (1981) while studying the mechanism of resistance in mungbean and
blackgram against whitefly inves�gated that bio-chemicals like phenols, amino acids and non-
reducing sugars were responsible for resistance to the insect. Kooner et al. (1994) studied the
phytochemical basis of resistance in urd bean and concluded that high contents of phenols, free
amino acids, total sugars and low content of non-reducing sugars governed the resistance.
Chemical control
Chhabra et al. (1993) reported that whitefly can be controlled by spraying the mung bean, urd
bean, soybean and rice bean with any of the following insec�cides using 250 litres of water per
hectare with manually operated sprayer: 950 ml of Malathion 50 EC (malathion), 625 ml of
Rogor 30 EC (dimethoate) or Anthio 25 EC (formothion) or Metasystox 25 EC (oxydemeton -1methyl), and 225 ml of Dimecron 85 WSC (phosphamidon). Disulfoton at the rate of 1.5 kg ha
as basal applica�on at sowing gave significant reduc�on of popula�on of whitefly (Chaudhary
et al. 1981). Combina�on of phorate l0 G and monocrotophos (Vadodaria and Vyas 1987),
endosulfan, monocrotophos, chlorpyriphos and dimethoate (Yein 1983) were most effec�ve
for the control of the pest. Cheema et al. (2009) reported that pest can be controlled by spraying
the crop with 1.25 litre triazophos 40 EC or 100 g thiamethoxam 25 WG /ha by using 250 litre
water. Shirale and Bidgire (2009) reported that spraying triazophos 40 EC @ 800 ml/ha twice,
recorded less number of whiteflies at three, seven and ten days a�er spraying. The treatment B.
bassiana @ 1 l/ha followed by Triazophos 40 EC @ 800 ml/ha was at par with treatment
triazophos 40 EC @ 800 ml/ha followed by B. bassiana @ 1 l/ha at three days a�er spraying.
Purwar and Yadav (2006) observed that Dimlin (diflubenzuron) 25 WP and triazophos 40 EC
were effec�ve to combat whitefly popula�on and to manage yellow mosaic in soybean.
202

Management for aphid
Cultural control
Aphids mul�ply and spread very fast under dry condi�ons. In endemic area, efforts should be made to keep the crop free from moisture stress. Wider spacing, i.e., 60 cm x 20 cm is more favourable to aphids on chickpea than narrow spacing, i.e. 30 cm x 10 cm. The ac�vity of aphid as a vector was slow down on paraffin treated plots which inhibited virus transmission (Kiss et al 2003). Proper weeding and destruc�on of weed hosts from the field area help to check the aphid popula�on and hence the diseases like leaf crinkle and mosaic mo�le (Singh et al 1997).
Biological control
Fi�een predators (Coccinella septumpuncata Linn, C. transversalis (Fabr), C. nigritus, Menochiles sexmaculatus (Fabr), Brumus suturalis (Fabr), Harmonia arcuata, Vernia sp., Ischiodan javana (Wied), I. scutellaris (Fabr), Xanthogramma scutellare (F.), Chrysopa camea Steph, Cyclosa insulana (Costa), Luecangs celebesiana (W), Theridion sp. and Leucopis sp. and three parasites (Trioxys indicus, Praon sp. and Diaerec�ella rapaecur�s) have been recorded as natural enemies on A. craccivora (Ahmad and Lal 1998; Yadava and Lal 1998).
Host plant resistance
Some genotypes resistant to A. craccivora have been iden�fied in mung bean, urd bean and cowpea but limited work seems to be undertaken in chickpea. Cul�vars H 75-35 and 2184 of chickpea support minimum aphid popula�on (Lal et al. 1989). Mustaque (1977) tested performance of 12 chickpea cul�vars and observed some of these as less suscep�ble. Chandra et al. (1992) reported that the varia�on in resistance to aphids infesta�on was correlated mainly with the colour of foliage of the genotypes; it has been observed that A. craccivora is quite a�racted towards yellow colour.
Chemical control
A large number of insec�cides [Acephate (0.02%), bromophos (0.05%), carbaryl (0.01%), cypermethrin (0.004%), oxydemeton methyl (0.05%), dimethoate (0.03%), endosulfan (0.07%), fenvalerate (0.01%), lindane (0.1%), malathion (0.05%), menazon (0.05%), methomyl (0.03%), phosdrin (0.02%), primicarb (0.03%), primiphos-methyl (0.03%), thiometon (0.05%)] have been evaluated for the management of A. craccivora on pulse crops and most of them have been found quite effec�ve to control this aphid (Singh and Sircar 1980; Thakur et al. 1984). Chhabra et al. (1993b) reported that the aphid can be controlled by spraying 625 ml of Malathion 50 EC (malathion) or 375 ml of Rogor 30 EC (dimethoate) or 375 ml of Metasystox 25 EC (oxydemeton methyl) per hectare.
Management for mite
Cultural control
Proper weeding and destruc�on of weed hosts from the field area help to check the vector popula�on and hence the mosaic mo�le disease. The �me of sowing may be adjusted to coincide with low vector popula�on (Singh et al 1997).
Biological control
About 11 natural enemies have been reported to predate on mite. Amblyseius laryoensis, Typhlodromus annectens, T. porresi, Euseius victoriensis, T. rickeri and T. s�pulatus are some of the major phytoseid mites which help to control mite in different parts of the world on different hosts.
203

Chemical control
The mite can be controlled by spraying 375 ml of Rogor 30 EC (dimethoate) in 200-250 liters of water per hectare (Annonymous 2010).
Management for thrips
Cultural control
Intercropping of mungbean with co�on minimizes incidence of thrips as compared to sole crop.
Biological control
Satyanarayan et al. (1995) reported Brumoides suturalis, Loccinella septumpunctata, Menochilus sexmaculatus (Coccinellids), Chrysopa scelestes, Polistes hebraeus, Geocoris tricolor, Acantholepis sp. (spiders) to be the primary natural enemies of M. distalis.
Host plant resistance
Malik (1990) observed that summer mung bean genotypes SML 77, UPM 82-4 and Pusa 107 were resistant to M. distalis under natural as well as screen house condi�ons. A number of mung bean and urd bean genotypes have been found resistant to the thrips (Chhabra and Kooner 1988, 1992, 1993). Urdbean varie�es, N 212 and Khargone 3 and mungbean selec�ons, P 15, P 56, P 101, P 135-2, P 382, P 386, P 435-1, P 435-2 and P 435-3 were found free from infec�on under natural condi�ons. Shanmugasundram (1988) reported variety Pant mung 3 to be moderately resistant to leaf curl virus. It was further inves�gated that low content of amino acids, total phenols, total minerals, total sugars, non-reducing sugars, calcium, potassium and high content of total carbohydrates were responsible in contribu�ng resistance in SML 99 and SML 100 (Chhabra et al. 1994).
Chemical control
The bean thrips/flower thrips can be effec�vely controlled by spraying the crop at bud ini�a�on stage with 250 ml of Rogor 30 EC (dimethoate) or Malathion 50 EC (malathion) or 300 ml of Metasystox 25 EC (oxydemeton methyl) or 90 ml of Dimecron 85 WSC (phosphamidon) in 200-250 litres of water per hectare (Kooner et al. 1983; Chhabra and Kooner 1985b, 1986). Damage caused by thrips gets reduced considerably with the applica�on of a combina�on of aldicarb at
-11 kg ha and monocrotophos 0.04% (Chander and Singh 1989). Gupta and Singh (1992) reported that one applica�on of granular formula�ons of phorate, mephasulfan, disulfoton,
-1carbofuran and aldicarb at 2.0 kg ha were effec�ve in controlling this pest. Irulandi and Balasubramanian (2000) reported neem seed kernal extract at 5% and neem oil at 2% to be as effec�ve as monocrotophos 0.05%.
Conclusions
· Minimize the inoculums ( virus source) and colonizing vectors by using cultural prac�ces.
· Follow the integrated pest management strategies for the management of insect vectors.
· Tac�cs for integra�on are resistant plants, trap crops, barrier crops, s�cky traps and insec�cide sprays.
204

Suggested Reading
Ahmad, R. and Lal, S.S. (1998). Insect pests of len�l and their management. In IPM System in Agriculture. Vol. IV. Pulses (Eds R.K. Upadhyay, K.G. Mukerji and R.L. Rajak), pp. 413-426. New Delhi : Aditya Books.
Annonymous (2010) Package of prac�ces for Kharif crops, pp 58- 63, Punjab Agricultural University, Ludhiana.
Asthana, A.N., Lal, S.S. and Dhar, V. (1997). Current problems in pulse crops and future needs. In Na�onal Seminar on Plant Protec�on towards Sustainability, pp. L-3. Hyderabad : Plant Protec�on Associa�on of India.
Bakhe�a, D.R.C. and Sidhu, A.S. (1977). Biology and seasonal ac�vity of the groundnut aphid, Aphis craccivora Koch. Journal of Research Punjab Agricultural University 14: 299-303.
Beniwal, S.P.S., Chaubey, S.N. and Bharathan, N. (1980). Presence of urdbean leaf crinkle virus bin seeds of mungbean germplasm. Indian Phytopath. 33:360-361.
Beniwal,S.P.S., and chaubey, S.N. (1979). Transmission of urdbean strain of bean common mosaic virus (UB-BCMV) by a tarsonemid mite, polyphagotarsoneemus latus. Indian phytopath. 32:163.
Bharathan, N. and beniwal, S.P.s. (1984). Transmission characteris�cs of urdbean leaf crinkle virus by the Epilachna beetle, henosepilachna dodecas�gma. Indian Phytopath. 37 : 660-664.
Brar,J.S. and Ratual, H.S. (1986). Some field characteris�cs of leaf crinkle virus of urdbean, Vigna mungo (L.) Hepper. Indain jour. Virology 2 : 49-56.
Chakraborty, A. and Du�a, S.K. (1999). Popula�on buildup of thrips, Megalurothrips distalis (Karny) in pea crop. Journal of Agricultural Science Society of North East India, 12: 48-51
Chander, S and Singh, Y. (1989). Control of thrips, Megalurothrips distalis and pod borer viz. Euchrysops cnejus Fab. and Maruca testulalis Gayer infes�ng greengram, Vigna radiata (L.) Wilczek. Indian Journal of Entomology 51: 131-135.
Chandra, S., Ahmad, R. and Asthana, A.N. (1992). Field tolerance to aphid in len�l. LENS Newsle�er 19: 45.
Chaudhary, J.P., Singh, A.K. and Yadav, L.S. (1981). Effect of disulfoton granules on pest complex, crop growth and yield of green-gram. Indian Journal of Entomology 43: 369-372.
Cheema, H.K., Taggar G.K., Singh R. and Kooner B.S. (2009). Evalua�on of insec�des against B. tabaci (Gennadius) on urdbean [Vigna mungo (L.) Hepper]. J. Insect Sci 22(4): 388-392
Chenulu, V.V. (1984). Plant Virology in India-past, Present and Future. Indian Phytopath. 37:1-20.
Chhabra, K.S. and Kooner, B.S. (1985). Losses in summer mungbean Punjab. Indian Journal of Entomology 47: 103-105
Chhabra, K.S. and Kooner, B.S. (1986). Compara�ve efficacy of different insec�cides in controlling thrips, Megalurothrips distalis (Karny) on summer mungbean, Vigna radiata (L.) Wilczek. Legume Research 9: 39-42.
205

Chhabra, K.S. and Kooner, B.S. (1988). Varietal resistance in mungbean (Vigna radiata (L.)
Wilczek) to thrips, Megalurothrips disatalis (Karny). The Tropical Vegetable Informa�on
Service News 3: 10-11.
Chhabra, K.S. and Kooner, B.S. (1992). Sources of resistance in mungbean against major insect-
pests and yellow mosaic virus. Legume Research 14: 175-184
Chhabra, K.S. and Kooner, B.S. (1993). Screening of mungbean germplasm against insect pests
and yellow mosaic virus. Indian Journal of Pulses Research 6: 69-75.
Chhabra, K.S., Kooner, B.S. and Cheema, H.S. (1986). Incidence of Aphis craccivora Koch. on
grain legumes in the Punjab. In Aphidology in India, Proceedings of the Na�onal
Symposium (Ed B.K. Agarwala.), pp. 23-28. Agartala : ASI.
Chhabra, K.S., Kooner, B.S., Saxena, A.K. and Sharma, A.K. (1981). Effect of biochemical
components on the incidence of insect pest complex and yellow mosaic virus in
mungbean. Crop Improvement 8: 56-59.
Chhabra, K.S., Kooner, B.S., Saxena, A.K. and Sharma, A.K. (1994). Role of allomones in varietal
resistance in summer mungbean against thrips, Megalurothrips distalis (Karny). In
Proceeding of Interna�onal Symposium on Pulses Research, p. 161. New Delhi.
Chhabra, K.S., Lal, S., Kooner, B.S. and Verma, M.M. (1993). Insect Pests of Pulses – Iden�fica�on
and Control Manual. Ludhiana : Punjab Agricultural University, and Kanpur :
Directorate of Pulses Research.\
Das gupta,B. and Chowdhury, A.K. (1985). Use of intervarietal intercropping to minimize the
yellow mosaic virus of urd and mungbeans. Indian Jour Plant Pathol. 3 : 100-101.
David, P.M. (1991). Resurgence of yellow mite, Polyphagotarsonemus latus (Acari :
Tarsonemidae) on chilli following applica�on of insec�cides. Madras Agricultural
Journal 7:, 20-25
Dhooria, M.S. (1984). Preliminary observa�on on the biology and host range of the tarsonemid
mite, Polyphagotarsonemus latus (Acari : Tarsonemidae). Acaricide Newsle�er 4: 7-9.
Dorges, S.K., Dalaya, V.P. and Kabi, T.D. (1966). Studies on bionomics and control of groundnut
aphid, Aphis craccivora Koch. Labdev Journal of Science and Technology 4 :165-167
Gerson, U. (1992). Biology and control of the broad mite, Polyphagotarsonemus latus (Banks)
(Acari : Tarsonemidae ). Experimental and Applied Acaricide 13: 163-178
Gupta, P.K. and Singh, J. (1992). Effec�ve control of thrips by soil applica�on of granular
insec�des in greengram. Indian Journal of Entomology 54: 465-470.
Gupta, S.K. and Dhooria, M.S. (1973). Occurrence of Hemitatorsonemus latus (Banks) on
blackgram and its control in Punjab. Science and Culture 37: 484-485
Hariri, G. (1981). Insect and other pests. In Len�ls (Eds G. Webs and G. Haw�n), pp. 173-189.
London : Commonwealth Agricultural Bureau.
Honda Y and Iwaki M (1991) studies on plant virus disease and their control in Japan. In
Integrated control of plant viral disease. pp 13-14
206

Irulandi, S. and Balasubramanian, G. (2000). Efficacy of insec�cides against pod borer, Lampides boe�cus L. (Lycaenidae : Lepidoptera) on green gram. Insect Environment 6: 108-109.
Kiss E F, Huszka T, Ockro I and Vidal S (2003) Pepper pathogen viruses and their control by resistance breeding and light summer oils. Bulle�n OILB-SROP 26:201-10.
Kooner, B. S. and Cheema, H. K.(2007a). Screening of mungbean germplasm against whitefly, Bemisia tabaci Genn. and MYMV. Journal of Food Legumes 20: 100-102.
Kooner, B. S. and Cheema, H. K.(2007b). Screening of mungbean germplasm against whitefly, Bemisia tabaci and MYMV. Acta Hor�culturae 752: 307-310.
Kooner, B.S. (1998). Iden�fica�on of sources of resistance to whitefly, jassids and MYMV in kharif mungbean. In Proceedings of Na�onal Symposium on Management of Bio�c & Abio�c Stresses in Pulse Crops, p. 40. Kanpur : Indian Society of Pulses Research and Development.
Kooner, B.S., Chhabra, K.S., Sekhon, H.S., Dhingra, K.K. and Cheema, H.S. (1983). A new deformity in summer mungbean, Vigna radiata (L.) Wilczek. Pulse Newsle�er 3: 23-26.
Kooner, B.S., Chhabra, K.S., Sharma, A.K. and Saxena, A.K. (1994). Phytochemical resistance in urdbean genotypes against major insect-pests. In Proceedings of Interna�onal Symposium on Pulses Research, pp. 162. Kanpur : Indian Society of Pulses Research and Development.
Lal, S.S. (1987). Insect pests of mung, urd, cowpea and pea and their management. In Plant Protec�on in Field Crops (Eds M. Veerabhadra Rao and S. Sithanantham), pp. 185-201. Hyderabad: Plant Protec�on Associa�on of India
Lal, S.S. (1992). Insect pest of len�l and their management : A review. Agricultural Review. 15: 225-232.
Lal, S.S. and Ahmad, R. (2002). Integrated insect pest management present status and future strategies in pulses. In Pulses for SustainableAgriculture and Nutri�onal Security (Eds M. Ali, S.K. Chaturvedi and S.N. Gurha), pp. 101-110. Kanpur : IIPR
Lal, S.S., Yadava, C.P., Dias, C.A.R. and Nigam, R. (1989). Effect of plan�ng density and chickpea cul�vars on the infesta�on of black aphid, Aphis craccivora Koch. Madras Agricultural Journal 76: 461-462.
Malik, S.P.S. (1990). Compara�ve resistance of summer mungbean genotype to the thrips, Megalurothrips distalis (Karny.). M.Sc. thesis, Punjab Agricultural University, Ludhiana.
Mushtaque, A. (1977). Preliminary studies on the infesta�on of Aphis craccivora Koch to chickpea and len�l. Journal of Agricultural Research 15 : 31-35.
Nene, Y.L. (1972). A survey of viral diseases of pulse crops in U�ar Pradesh. Final Technical Report of PL 480 Adhoc Project, pp. 88-108.
Nene, Y.L. (1973). Control of Bemisia tabaci Genn. : A vector of several plant viruses. Indian Journal of Agricultural Sciences 43: 433-436
Purwar J P and yadav S R (2006).Evalua�on of biora�onal insec�cides against whitefly (Bemisia tabaci Genn.): a vector of soybean yellow mosaic. Journal Experimental Zoology 9(2): 499-503
207

Ravindrababu, R. (1987). Studies on mungbean yellow mosaic virus disease. Ph.D. thesis,
Tamil Nadu Agricultural University, Coimbatore
Nene, Y.L. and Reddy, M.V. (1976). Preliminary informa�on on chickpea stunt. Tropical Grain
Legume Bulle�n 5: 31-32
Persley, D and Sharman, M ( 2007). Thrips and Tospovirus: A Management Guide, CRC for
Tropical Plant Protec�on, Brisbane, Department of Primary Industries and Fisheries,
Queensland.
Reddy,P.C. and Krishnaiah, K. (1983). Assessment of field losses due to leaf crinkle virus on
blackgram. Indian J.Mycol. Plant Pythol. 13: 19
Satyanarayana, J., Singh, K.M. and Singh, R.N. (1995). Insect pest succession in rice bean.
Bulle�n of Entomology 36: 78-83.
Satyanarayana, J., Singh, K.M., Singh, R.N. and Singh, S. (1997). Influence of intercropping and
insec�cide. Incidence of major pests in ricebean, Vigna umbellata. Indian Journal of
Entomology 59: 316-325.
Shanmugasundaram, S.(1988). A catalog of mungbean cul�vaters released around the world.
Asian vegetable research and development centre, Shanhua, Taiwan.
Shirale D. and Bidgire U. (2009). Integra�on of bioagents and synthe�c insec�cide in the
management of whitefly and its effect on yield in soybean. Karnataka Journal of
Agricultural Science 22(3): 631-632.
Singh, D.P. (1980). Inheritance of resistance to yellow mosaic irus in blackgram (Vigna mungo
(L.) Hepper). Theore�cal and applied gene�cs. 57: 233-356.
Singh, D.S. and Sircar, P. (1980). Rela�ve toxicity of pyrethroids to Lipaphis erysimi Kalt and
Aphis craccivora K. Indian Journal of Entomology 42: 597-605.
Singh, M. (1996). Studies on the biology of leaf curl mite Polyphagotarsonemus latus (Banks)
(Acari : Tarsonemidae) on chilli (Capsicum annum). M.Sc. thesis, Punjab Agricultural
Univerity, Ludhiana.
Singh, R A, Gurha S N and Ghosh A (1997) Diseases of mungbean and urdbean and their
management. In: Diseases of Field Crops and their Management (Eds T S Thind), pp.
179-204, Daya Publishing House, Delhi.
Singh, R.N and Nene, Y.L. (1978). Further studies on the mosaic mo�le disease of urdbean.
Indian Phytopath. 31:159-162.
Srikanth, J. and Lakkundi, N.H. (1998). Instar period, fecundity and longevity of cowpea aphid,
Aphis craccivora Koch. on seven leguminous hosts. Journal of Aphidology 2: 18-19.
Thakur, B.S., Verma, R., P�tunda, A. and Rawat, R.R. (1984) Chemical control of aphid, Aphis
craccivora Koch on len�l. Indian Journal of Entomology 46: 103-105.
Tsuchaizaki, T., Iwaki, M., Thongameearkom ,P., Sarindu, N. and Deema, N.(1996). Bean
common mosaic virus isolated from mungbean (vigna radiate) in Thailand. Technical
bulle�n of the tropical Agriculture research centre, No. 21:184-188.
208

Vadodaria, M.P. and Vyas, H.N. (1987). Control of whitefly, Bemisia tabaci (Gennadius) and its impact on yellow mosaic virus (YMV) in greengram, (Vigna radiata (L.) Wilczek) and the grain yield. Indian Journal of Agriultural Research 21: 21-26.
Van Emden, H.F. (1973). Aphid host plant rela�onship : some recent studies. In Perspec�ves in Aphid Biology (Ed A.D. Lowe), pp. 54-64. Auckland Entomology society of New Zealand.
Yadava, C.P. and Lal, S.S. (1998). Major insect pests of chickpea and their management. In IPM System in Agriculture. Vol. IV. Pulses (Eds R.K. Upadhyay, K.G. Mukerji and R.L. Rajak), pp. 197-231. New Delhi : Aditya Books.
Yein, B.R. (1983). Efficacy of certain insec�cides against B. tabaci (Genn.) and Pagria signata (Motsch) on blackgram. Journal of Research Assam Agricultural University 4: 45-48.
209

INSECT SAMPLING METHODS FOR IMPORTANT LEGUME CROPS
R. K. SAINIDepartment of Entomology,
CCS Haryana Agricultural University,Hisar-125004
Regular monitoring of the crop for the presence of insects pests and/or damage is necessary in order to make �mely decisions on control, especially when it is important to be targe�ng small and possibly insec�cide resistant larvae.
Sampling and detec�on Tools and Techniques
The success of any sampling programme depends on the effec�veness and efficiency of the detec�on and sampling tools and techniques.
Depending on the objec�ves, tools include traps, nets, microscopes, and molecular test kits. Techniques include knockdown, ne�ng, trapping, and visual inspec�on. Looking for characteris�c damage symptoms or signs of pest presence (such as faeces or frui�ng bodies) can also be useful, depending on the pest.
Visual Sampling: Examining a specified number of plants, plant parts, or infested areas for pests or their damage. Another method is to count the number of individuals that can be seen during a predetermined �me interval. These �med searches are used to monitor fruit tree leafroller, tussock moth larvae counted during a fixed amount of �me (e.g. larvae per hour of search).
Knockdown Techniques: These techniques sample arthropod pests that are easily dislodged from their habitat. It involves physically removing pests and beneficial from a por�on of a plant onto a tray, cloth, white painted can or bucket, or other collec�on device, then coun�ng them. Alterna�vely, for a quicker but less accurate es�mate, presence-absence sampling can be adopted. The drop cloth is another common sampling tool, especially for collec�ng lepidopterous caterpillars, leaf beetles, s�nk bugs, etc. A sheet, roll of plas�c or canvas, or tray is placed along a 30-45 cm sec�on of the plant row. Shaking the plants dislodges insects. The drop cloth method is highly efficient and low in cost but can be �me consuming.
Suc�on Techniques: Vacuum removal of arthropod pests using suc�on equipment, such as the D-Vac, modified leaf blowers, or any other suc�on devices dislodges pest species that are somewhat mobile. Vacuum devices are more effec�ve than bea�ng sheets for detec�ng lygus bugs and other mobile pests that are found on crop canopy, but not appropriate for so� bodied insects which may get crushed.
Ne�ng Techniques: Ne�ng techniques are easy to use and are low cost. A standard sweep net consists of a cone-shaped cloth bag 60 cm deep fi�ed into a wire loop 38 cm
210

in diameter and a�ached to a handle 65 cm long. Sweeps may be taken singularly or consecu�vely. A�er a series of sweeps, counts of each species are based on an average of all samples.
Trapping Techniques: Traps are used for mobile insects. They a�ract insects either through visual or chemosensory a�ractants or randomly catch them. But traps provoke a response by using the pest's food source as s�mulus. E.g. bait traps used to monitor dried fruit beetles; pheromone traps for disrup�on of ma�ng. Placement is usually cri�cal to op�mize their trapping efficiency. This generally means placement at the top of the canopy. Many pheromone traps drop dras�cally in efficiency or sensi�vity when placed only a few inches to a foot above the canopy. Light traps can be used to monitor the emergence or migra�on dates of various pests. One disadvantage of light traps is that they a�ract so many insect species that considerable �me and skill is required to sort the samples. S�cky traps are used to monitor adult whiteflies, thrips, aphids, and other pests and beneficial insect parasites. The ones used in most crops are bright yellow cards, each 7.5 by 12.5 cm or larger, covered with clear s�cky material. Certain wavelengths of yellow (about 550 to 600 nm) are more a�rac�ve to many insects than other wavelengths. Light blue s�cky traps capture more western flower thrips. To catch the most insects, hang the traps ver�cally. Placement of traps depends on the crop and the pest. A pi�all trap is a trapping pit for small animals, such as insects, amphibians and rep�les. A pi�all trap is an essen�al tool for catching and studying ground-dwelling insects, par�cularly springtails and ground beetles. Animals that enter a pi�all trap are unable to escape, and may either be killed by the trap or remain unharmed. It consists of a plas�c cup with a funnel, a hole is dug in the ground, the trap is sunk into the soil and the mouth is level with soil surface. Many ground dwelling spiders as well other arthropods fall into the trap and are enable to escape.
Defining the Sampling Universe and Sampling Unit
Coun�ng every insect or spider mite or measuring the length of every internode in a field is not feasible. However, a well planned sampling programme can provide the accuracy needed to evaluate a pest infesta�on.
Sample: A simple is a set of measurements taken from part of a popula�on or a sub set of the physical features of an environment. The degree to which the sample represents the whole field or managed system will determine the accuracy of the sampling programme.
Sampling Universe: Determining the boundaries of the area for which a specific pest management decision is to be made. For example, the area that will be covered in a single sampling programme. The sampling universe can consist of a single habitat, such as a whole field of corn; several smaller areas of a field if the area can be managed independently of one another; or even certain plants or plant pests if spot-trea�ng is feasible. For example, if in an orchard, there are different blocks, having different
211

varie�es, plan�ng �mes, or environmental condi�ons, it is best to sample them independently from one another.
Sampling Unit: The plant part or en�ty sampled in the sampling universe is the sampling unit. It may be a leaf, a bud, the area of plant covered with one sweep net sample, a square foot of soil, or the soil contained in a soil core sample. The choice of the sampling unit should take into considera�on the size, abundance, and distribu�on pa�ern and mobility of the pest organism, as well as balancing the cost of sampling against the accuracy of the informa�on obtained.
The sampling unit should be large enough to adequately assess the popula�on of the target pest, taking into account the pest's biology, but small enough to economically permit an adequate number of samples in a reasonable amount of �me. In general, choosing a small-sized sampling unit and collec�ng a large number of samples provides be�er informa�on than using a larger sampling unit and fewer samples.
SAMPLING METHODS FOR CHICKPEA PESTS
Pod borer, Helicoverpa armigera
A hand lens, microscope or USB microscope is cri�cal for examining small larvae. Small H. armigera larvae (3 rd instar) have a saddle on the fourth segment. In larger (5 th and 6 th instar) larvae, hair colour on the segment immediately behind the head are white. There is the presence of a pale patch in the hindwing of H. armigera. Sampling with a beat sheet is best prac�ce for monitoring helicoverpa in chickpeas.
Using a beatsheet
Check crops regularly (at least once a week) with a beatsheet from flowering through to harvest. In addi�on to larval counts; visual observa�on of the crop growth stage, progress of lowering/podding,and the presence of eggs, diseased larvae (NPV) and moths all provide useful
Informa�on for decision making. Each �me you inspect, check at least 5 x 1 row metre sec�ons at a number of sites in the field. Start sampling at least 50 m into the field, and include samples
from well into the field to enable a representa�ve average field popula�on to be calculated. Place the beat sheet with one edge at the base of a row. Spread the sheet out across the inter - row space and up against the base of the next row. Using a 1m long s�ck (dowel, heavy conduit), shake the row vigorously 10 �mes to dislodge larvae from the plants; size and
count larvae on the sheet.
Monitoring eggs and very small larvae - is it useful?
Eggs - There are no egg thresholds in chickpeas. Relying solely on egg counts for control
212

decisions in chickpea is unreliable and inconsistent. This is largely due to the difficulty in accurately detec�ng eggs on the chickpea plant. Egg survival through to larvae can also be highly variable so making decisions based on egg numbers is less accurate than those based on larval density. Use egg counts as an indica�on of an egg lay event.
Very small larvae - The number of very small larvae, hatched from the egg in the previous 24 hours, are difficult to es�mate in the field. Since very small larvae do not cause economic damage and have high mortality rates, they are only useful as indicators of an egg lay and poten�al ac�vity of larger larvae in a week or two.
Keep records
Keeping records should be a rou�ne part of insect checking. Successive records of crop inspec�ons will show whether pest numbers are increasing or decreasing, and help in deciding whether a spray is necessary. Insect checking records should include as a minimum:
· Date and �me of day· Crop growth stage and suscep�bility· Insects present (pest and beneficial)· Number of insects present (pest and beneficial) and their stage of development· Type of checking method used and number of samples taken per field· Control recommenda�on if any· Post spray counts
Higher numbers of helicoverpa can be tolerated during flowering than podding
From vegeta�ve to early flowering stages, high popula�ons of helicoverpa have no impact on yield or quality. In rare situa�ons, control may be warranted during the vegeta�ve and flowering stages if pest pressure is extreme.
Adjus�ng es�mates of larval density to include natural mortality of larvae
This natural mortality (most likely as a result of dislodgement from the plant, disease, cannibalism) is important because it means that there is a propor�on of the popula�on that will not survive to cause damage to the crop, even if they are not controlled. Data show an overall average mortality of 70% across a large number of fields with the average calculated from a range of mortality from 1% to 99%. Because mortality varies considerably from field to field, a very conserva�ve 30% mortality adjustment is suggested rather than a 70% adjustment.
Cutworm
Cutworms are plump, smooth caterpillars of moth species. They feed on all pulse crops, damaging them near the ground. They tend to infest pulse crops sporadically; serious infesta�ons can cause yield losses. The larvae of many cutworm species (but not all) feed at
213

night and hide below ground by day, and so can be difficult to find. Young larvae feeding on leaves cause abrasions to the leaf surface, chew marks and holes but older larvae are especially no�ceable when they sever seedling stems at ground level. Pulse seedlings may recover from cutworm damage by sprou�ng a new shoot, however this effec�vely delays establishment and reduces yield poten�al. O�en the grubs can be located by digging the soil surface near damaged plants where they can be seen curled up in a defensive posi�on.
Monitoring of cutworms
Inspect crops a�er emergence in the late a�ernoon or evening. Look for patches of crop with seedlings cut off. Scratch the soil at the base of damaged plants to locate caterpillars. There is no threshold level established to determine when to spray for cutworm in pulse crops. About 2 large caterpillars per 500 mm row is considered a threshold in cereals and canola.
SAMPLING METHODS FOR PIGEONPEA PESTS
The insects that a�ack the pods are considered the most important pests of pigeonpea. Pod borers (including Helicoverpa armigera and Maruca vitrata) and pod-sucking bugs (mainly Clavigralla spp) are the major pests in these groups. The pod borer H. armigera is the most important constraint to pigeonpea produc�on throughout South Asia.The larvae of this insect destroy buds, flowers, and pods. They a�ack leaves in the absence of the floral organs. Maruca larva feeds from inside a webbed mass of leaves and it becomes a menace early in the season especially in areas with high humidity. The adults and nymphs of the pod-sucking bugs (Clavigralla spp and Nezara viridula) pierce the pod wall and suck the fluid from the developing seeds. The a�acked seeds shrivel and develop dark patches. These pests are very common in Africa and Asia, par�cularly in dry seasons. Aphids colonize the young shoots, flowers, and pods. The young leaves of seedlings become twisted under heavy infesta�on and wilt when the plant is under moisture stress. Thrips and blister beetles normally a�ack the flowers and heavy infesta�on may lead to flower drop. Infesta�on by bruchids (Callosobruchus spp) starts in the field. The infested seeds lose their viability and are unfit for human consump�on. Delayed harves�ng, poor drying, and storage facili�es can lead to total loss of pigeonpea grain due to this pest.
NICRA team of Pigeonpea Pest Surveillance (2011) has published a Manual for Pigeonpea Pest Surveillance. The following sampling procedure is largely based on the methodology suggested the NICRA team.
Helicoverpa armigera
In pigeonpea 1 larva plant-1 or 3 eggs plant-1 have been considered as the threshold level for the applica�on of insec�cides in research plots. Shake the plant to be sampled. Larvae falling on to the ground should be counted; Addi�onally number of early instar
214

larvae present on the plant also should be counted (whole plant basis) on two plants and recorded.
Blue bu�erfly, Lampides boe�cus (L.): Larvae are green, oval and flat. Larvae feed on leaves, flower buds and green pods. Number of larvae of blue bu�erfly should be counted by visual search method (on whole plant basis) on two plants and recorded.
Plume moth, Exelas�s atomosa (Walsingham: Larvae are green or brown, spindle shaped and covered with short spines and larger hairs. Larvae feed on developing seeds and feeding holes are visible on the plant. Number of plume moth larvae should be counted by visual search method (on whole plant basis) on two plants and recorded.
Legume (cowpea) pod borer,Maruca testulalis (Geyer): Webbing of tender leaves and flower buds together seen on plants indicate the possibility of Maruca damage. Closer view presents discolora�on and shedding of flowers in the web and presence of frass. Upon opening of the
webs, dull to yellow-white colour larva (e)with dark spots forming dis�nct series along the length of the body are seen. Number of webs of flowerbuds due to Maruca should be counted (on whole plant basis) on two plants per spot and recorded.
Blister beetle: Adults are black in colour with large yellow spots and a red band across the abdomen. Number of spots varies with species. They feed on flowers.Blister beetles are to be counted as number of adults on two plants (whole plant basis) and recorded.
Mealybugs: One plant in the selected spot should be observed for infesta�on due to any species of mealybugs. Mealybug infesta�on across species is to be graded as to Grade1/2/3/4, where
Grade 1: sca�ered presence of mealybugs
Grade 2: 25% of plants covered with mealybugs
Grade 3: 50% of the plants covered with mealybugs
Grade 4: whole plant infested with mealybugs
Pod bugs
Clavigralla: Bugs of this species are brown and furry.
Nezara: Bugs are dull green in colour. Adults of Nezara can feed on seeds through pod walls. Their feeding causes �ny depressions on the pod wall. Different species of pod bugs viz., Clavigralla, Riptortus and Nezara should be counted on whole plant basis by visual search method on two plants/spot and recorded separately.
Pod damage due to pod fly, Melanagromyza obtusa (Malloch): External symptoms on pods are not highly obvious. Small holes with whi�sh window are common when a mature maggot about to pupate is seen inside the pod. Pod fly damage is also inferred through dissec�on of pods. Maggots feeding on seeds and puparia are seen in pods
215

infested with podfly. More than one maggot or pupa could be seen inside a single pod.
Fi�y developing pods should be randomly collected from the field (not more than 5 pods / plant) and examined for damage due to H. armigera and pod fly. Number of pods damaged due to H. armigera
and pod fly should be counted and recorded separately. The total number of pod fly immature stages
(maggots and pupa) should also to be counted together out of the 50 pods and noted.
SAMPLING METHODS FOR MUNGBEAN PESTS
H. armigera
Beat sheet sampling is the preferred sampling method for medium to large helicoverpa larvae. Small larvae should be scouted for by inspec�ng (opening) vegeta�ve terminals and flowers. Damage to vegeta�ve terminals is o�en the first visual clue that helicoverpa larvae are present. Ideally, mungbeans should also be scouted for eggs and moths, to pinpoint the start of infesta�ons and increase the chance of successful control.
· Inspect crops weekly during the vegeta�ve stage.· Inspect twice weekly from early budding un�l late podding.· Sample six widely-spaced loca�ons per field.· Take five one-metre long samples at each site with a ́ standard´ beat sheet.
2· Convert larval counts/metre to larvae/metre by dividing counts by the row spacing in metres.
Beat sheet sampling may only detect 50% of small larvae in vegeta�ve and podding mungbeans, and 70% during flowering, as they feed in sheltered sites such as leaf terminals. However, many small larvae may be lost to natural mortality factors before they reach a damaging size and this cancels out sampling inefficiencies in most crops. In general, mungbeans are more tolerant of early than late damage. Vegeta�ve mungbeans can tolerate up to 33% defolia�on without yield loss. While up to six
2helicoverpa/m have no impact on yield in well-watered flowering mungbeans, 35 2
kg/ha is lost/larva/m by late podding.
Monitoring for bean podborer (Maruca vitrata)
The most reliable way to es�mate podborer numbers is to:
1) determine the number of infested flowering racemes on 10-20 individual plants from different areas of the field. O�en the damage is obvious, webbed buds/flowers, but open un-webbed flowers to check for small larvae.
2) Then mul�ply the mean number of infested racemes per plant by the number of plants per square metre, assuming one larva per infested raceme.
216

Pod-sucking bugs
Feeding by pod-sucking bugs causes shrivelled and distorted seed, and can severely reduce yield and seed quality. Pod-sucking bugs can even damage seeds in black pods that are nearing harvest maturity. Late bug damage reduces seed quality but not yield. As only 2% seed damage is tolerable in mungbeans, bug thresholds are based on seed quality, not yield.
Green vegetable bug (GVB) Nezara viridula
Damage to young pods causes deformed and shrivelled seeds and reduce yield. Seeds damaged in older pods are blemished, difficult to grade out and reduce quality.
Crops should be inspected for GVB twice weekly from budding un�l close to harvest.
· Sample for GVB in early to mid morning.
· Beat sheet sampling is the most efficient monitoring method.
· Standard sampling units consist of five one-metre non-consecu�ve lengths of row within a 20 m radius.
2· Convert all bug counts/row metre to bugs/m by dividing counts/row metre by the row spacing in metres.
· At least six sites should be sampled throughout a crop to accurately determine adult GVB popula�ons.
· GVB nymphs are more difficult to sample accurately as their distribu�on is extremely clumped, par�cularly during the early nymphal stages (instars 1-3).
· Ideally, at least 10 sites (with five non-consecu�ve row metres sampled/site) should be sampled to adequately assess nymphal popula�ons.
Podsucking bug thresholds in mungbeans are determined by seed quality, the maximum bug damage permi�ed being only 2%.
Sucking insect pests:
Popula�on of jassids (nymphs & adults) and white flies (adults) can be es�mated by the visual count technique during early hours of the day from requisite plants per
threplica�on (usually coun�ng from 1/5 of total plant popula�on per plot), selected at random and tagged. The top, middle and bo�om parts of the tagged plants should be given due considera�on. Alterna�vely, the sudden trap method using a cubical iron-frame trap of 45cm x 45cm base and 60cm height clothed in high density polythene can be used to trap the adult jassids and whiteflies for easy coun�ng. The nymphs can be counted adop�ng the visual/sight-count technique from the plants by gently turning the leaves. Similarly, for aphids, the nymphs and adults can be counted on the plants directly taking observa�ons from at least a 10cm top shoot/twig for sampling. Yellow pan-traps or s�cky traps can also be used for coun�ng the jassids and whiteflies.
217

Data from visual counts can be homogenized by following the square root
transforma�on method, especially when zero counts are recorded. We add 0.5 or 1.0 to
the visual count data and find the square root before analysis that has to be
retransformed a�er analysis for interpreta�on. In case of percentage data, i�he
percentage values range between 30 and 80, usually arc sine (angular) transforma�on
would not be required; however, if the percentage values happen to be less than 30
and/ or more than 80, then angular transforma�on is required before analysis of data.
Aphids can damage faba bean, len�l, vetch and narbon bean plants directly by feeding
on plant �ssue or they can act as vectors for the spread of virus diseases or make plants
more suscep�ble to fungal diseases. Inspect crops from late autumn through to spring.
Infesta�ons are usually in patches across the paddock. Look closely at the growing
points of the shoot where aphids develop their infesta�ons. Bluegreen aphids are
camouflaged amongst the foliage, and may be sampled by using a sweep net or
knocking plants against a white
background to dislodge the aphids. The number of plants infested rather than the
number of individual aphids is important when considering control.Pulse crops should
be sprayed if aphids are observed on the growing �ps of 30% of plants from flowering
through to podding.
Blister beetle:
To record the incidence of blister beetles at flowering stage the numbers of beetles per
plant for a fixed �me interval during the morning hours (8 to 10am) or evening hours (3
to 5pm) of the day can be observed on the randomly tagged plants. However, some
species visit during early hours while others late; hence, a preliminary observa�on on
their behaviour shall become essen�al before standardizing the methodology for
blister beetle counts.
SAMPLING METHODS FOR COWPEA PESTS
The most important pests are Maruca testulalis (Geyer), which damages flowers and
pods. Sweep Net Sampling: A standard 15-inch diameter sweep net is the basic
sampling tool for many insect pests, including aphids, plant bugs and caterpillars. A
sampling unit consists of 10 consecu�ve (180 degree) sweeps taken while walking
through a field. The net is swung from side to side with each step. The net should be
held so that the lower half of the opening (7 to 8 inches) is drawn through the foliage.
Five samples (each consis�ng of 10 sweeps), one taken in each quadrant of the field and
one near the center of the field, provide a good es�mate of insect numbers. Samples
should be taken 30 to 50 feet from the field margin. O�en aphids are too abundant to
count individually but can be es�mated in 10s or 100s.
218

Suggested Reading:
Brier H, Quade A, Wessels J (2010) Economic Thresholds for Helicoverpa and other
pests in summer pulses – challenging our percep�ons of pest damage. In '1st
Australian Summer Grains Conference'. Gold Coast, Australia. (Eds B George-
Jaeggli, DJ Jordan). (Grains Research and Development Corpora�on).
Brier, H., Quade, A. and Wessels Joe. 2010. Economic thresholds for Helicoverpa and
other pests in summer pulses – challenging our percep�ons of pest damage.
Proceedings of the 1st Australian Summer Grains Conference, Gold Coast,
Australia, 21st– 24 th June 2010.
Concepts in Integrated Pest Management. 2002. Edited by Norris, Kogan, Caswell-Chen
& Kogan. Pren�ss Hall, New York.
Ecological Methods, 3rd Edi�on. 2000. T.R.E. Southwood and P.A. Henderson. Blackwell
Publishing, Oxford, UK.
Lateef, S.S., and Reed, W.1990. Insect pests on pigeonpea. Pages 193–242 in Insect
pests of tropical food legumes (Singh, S.R., ed.). Chichester, UK: John Wiley &
Sons.
Minja, E.M., Shanower, T.G., Songa, J.M., Ong'aro, J.M., Kawonga, W.T., Mviha, P.,
Myaka, F.A., Slumpa, S., and Okurut-Akol, H. 1999. Studies of pigeonpea insect
pests and their management in Kenya, Malawi, Tanzania, and Uganda. African
Crop Science Journal 7(1):59–69.
NICRA team of Pigeonpea Pest Surveillance, 2011 . Manual for Pigeonpea Pest
Surveillance . Jointly published by Na�onal Centre for Integrated Pest
Management, New Delhi, Central Research Ins�tute for Dryland Agriculture
and Indian Ins�tute of Pulses Research, Kanpur. 29 pp.
Ranga Rao, G.V., and Shanower, T.G.1999. Iden�fica�on and management of pigeonpea
and chickpea insect pests in Asia. Informa�on Bulle�n no. 57. Patancheru502
324, Andhra Pradesh, India: Interna�onal Crops Research Ins�tute for the
Semi-Arid Tropics. 96 pp.
Reed, W., and Lateef, S.S. 1990. Pigeonpea pest management. Pages 349–374 in The
pigeonpea (Nene, Y.L., Hall, S.D., and Sheila, V.K., eds.). Wallingford, Oxon, UK:
CAB Interna�onal.
Rogers, DJ, Brier, HB, (2010) Pest-damage rela�onships for Helicoverpa armigera
(Hübner) (Lepidoptera: Noctuidae) on vegeta�ve soybean. Crop Protec�on
29 (1), 39–46.
Shanower, T. G. and Romeis, J.1999. Insect pests of pigeonpea and their
management.Annual Review of Entomology 44: 77-96.
219











![From the Director’s Desk - ncipm.org.inmod].pdf · and CCS HAU, Hisar in Haryana in August, 2007 indicated that 115 villages in Dabwali, Odhan and Sirsa blocks of Sirsa district,](https://static.fdocuments.net/doc/165x107/5ae62c2a7f8b9aee078c99c2/from-the-directors-desk-ncipmorgin-modpdfand-ccs-hau-hisar-in-haryana.jpg)







