Adaptation of a sandwich enzyme-linked immunosorbent assay to determine the concentration of bovine...
7
Short communication Adaptation of a sandwich enzyme-linked immunosorbent assay to determine the concentration of bovine leukemia virus p24 and optimal conditions for p24 expression in short-term cultures of peripheral blood mononuclear cells M. van den Heuvel a, *, D. Portetelle b , B. Jefferson a , R.M. Jacobs a a Department of Pathobiology, University of Guelph, Guelph, Ont., Canada N1G 2W1 b Department of Microbiology, Gembloux, Belgium Received 3 December 2002; received in revised form 14 April 2003; accepted 15 April 2003 Abstract Bovine leukemia virus (BLV) is a common retroviral infection of cattle. Infection is accompanied by integration of BLV into the host cell genome and is persistent for the life of the individual as is the presence of anti-BLV antibodies. Lymphosarcoma occurs in a small fraction of infected adult individuals but otherwise there is little or no associated disease. Viremia is undetectable, however, BLV is expressed readily once infected cells are cultured in vitro. A sandwich enzyme-linked immunosorbent assay (sELISA) was optimized, using murine monoclonal antibodies, to quantify the major internal structural protein (p24) produced in short-term cultures of peripheral blood mononuclear cells (PBMCs). Optimal production of BLV p24 was achieved utilizing RPMI supplemented with 10% fetal bovine serum (FBS), pH 7, and 5 /10 6 cells per ml. Cultures were terminated at 24 h. The sELISA was linear between 30 and 900 ng/ml and the limit of detection was 1.2 ng/ml. At three concentrations of p24, intra- and inter-assay coefficients of variation (CV) varied between 9.2 and 13.3 and 5.1 and 12.9%, respectively. # 2003 Elsevier Science B.V. All rights reserved. Keywords: Bovine leukemia virus; p24; Sandwich enzyme-linked immunosorbent assay; Short-term culture; Optimization Bovine leukemia virus (BLV) is the etiological agent of lymphosarcoma in adult cattle, a relatively rare neoplasm of B-cells (Burny et al., 1985). BLV infects cattle worldwide. Infections are life-long, a consequence of the retroviral life cycle where the BLV provirus integrates into the host cell genome (Burny et al., 1978, 1988). The majority of infected cattle are asymptomatic but, approximately one-third of infected cattle develop persistent lymphocytosis (PL) (Burny et al., 1988), a polyclonal expansion of B-cells. Less than 5% of BLV- infected cattle develop lymphosarcoma, although ap- proximately two-thirds of cattle with lymphosarcoma have a history of PL (Burny et al., 1985). In asymptomatic BLV-infected cows with a normal number of peripheral blood B-cells, from 0.1%9 /1.8 to 9.2%9 /19 (mean9 /S.D.) of peripheral blood lympho- cytes (PBLs) are BLV-infected, whereas in cows with PL BLV-infected cells comprise from 13.9%9 /6.6 to 66%9 / 4.8 of PBLs (Mirsky et al., 1996). BLV is demonstrable by the polymerase chain reaction from nuclear extracts of PBLs, but viral gene products are not readily detectable by immunological methods such as Western blotting or radioimmunoassay (Ferrer, 1980; Kettmann et al., 1980, 1982; Mirsky et al., 1996), suggesting that in vivo, BLV is either not present in blood or is present in minute quantities below the sensitivity of current detec- tion methodologies. However, since infection with BLV results in a persistent anti-BLV antibody titer, it is thought that some constant but low level of expression must occur perhaps compartmentalized to such organs as lymph nodes, spleen, or bone marrow. Several studies have * Corresponding author. Tel.: /1-519-824-4120x54907; fax: /1- 519-763-1450. E-mail address: mv[email protected] (M. van den Heuvel). Journal of Virological Methods 111 (2003) 61 /67 www.elsevier.com/locate/jviromet 0166-0934/03/$ - see front matter # 2003 Elsevier Science B.V. All rights reserved. doi:10.1016/S0166-0934(03)00148-4
-
Upload
m-van-den-heuvel -
Category
Documents
-
view
212 -
download
0
Transcript of Adaptation of a sandwich enzyme-linked immunosorbent assay to determine the concentration of bovine...

Short communication
Adaptation of a sandwich enzyme-linked immunosorbent assay todetermine the concentration of bovine leukemia virus p24 andoptimal conditions for p24 expression in short-term cultures of
peripheral blood mononuclear cells
M. van den Heuvel a,*, D. Portetelle b, B. Jefferson a, R.M. Jacobs a
a Department of Pathobiology, University of Guelph, Guelph, Ont., Canada N1G 2W1b Department of Microbiology, Gembloux, Belgium
Received 3 December 2002; received in revised form 14 April 2003; accepted 15 April 2003
Abstract
Bovine leukemia virus (BLV) is a common retroviral infection of cattle. Infection is accompanied by integration of BLV into the
host cell genome and is persistent for the life of the individual as is the presence of anti-BLV antibodies. Lymphosarcoma occurs in a
small fraction of infected adult individuals but otherwise there is little or no associated disease. Viremia is undetectable, however,
BLV is expressed readily once infected cells are cultured in vitro. A sandwich enzyme-linked immunosorbent assay (sELISA) was
optimized, using murine monoclonal antibodies, to quantify the major internal structural protein (p24) produced in short-term
cultures of peripheral blood mononuclear cells (PBMCs). Optimal production of BLV p24 was achieved utilizing RPMI
supplemented with 10% fetal bovine serum (FBS), pH 7, and 5�/106 cells per ml. Cultures were terminated at 24 h. The sELISA was
linear between 30 and 900 ng/ml and the limit of detection was 1.2 ng/ml. At three concentrations of p24, intra- and inter-assay
coefficients of variation (CV) varied between 9.2 and 13.3 and 5.1 and 12.9%, respectively.
# 2003 Elsevier Science B.V. All rights reserved.
Keywords: Bovine leukemia virus; p24; Sandwich enzyme-linked immunosorbent assay; Short-term culture; Optimization
Bovine leukemia virus (BLV) is the etiological agent
of lymphosarcoma in adult cattle, a relatively rare
neoplasm of B-cells (Burny et al., 1985). BLV infects
cattle worldwide. Infections are life-long, a consequence
of the retroviral life cycle where the BLV provirus
integrates into the host cell genome (Burny et al., 1978,
1988). The majority of infected cattle are asymptomatic
but, approximately one-third of infected cattle develop
persistent lymphocytosis (PL) (Burny et al., 1988), a
polyclonal expansion of B-cells. Less than 5% of BLV-
infected cattle develop lymphosarcoma, although ap-
proximately two-thirds of cattle with lymphosarcoma
have a history of PL (Burny et al., 1985).
In asymptomatic BLV-infected cows with a normal
number of peripheral blood B-cells, from 0.1%9/1.8 to
9.2%9/19 (mean9/S.D.) of peripheral blood lympho-
cytes (PBLs) are BLV-infected, whereas in cows with PL
BLV-infected cells comprise from 13.9%9/6.6 to 66%9/
4.8 of PBLs (Mirsky et al., 1996). BLV is demonstrable
by the polymerase chain reaction from nuclear extracts
of PBLs, but viral gene products are not readily
detectable by immunological methods such as Western
blotting or radioimmunoassay (Ferrer, 1980; Kettmann
et al., 1980, 1982; Mirsky et al., 1996), suggesting that in
vivo, BLV is either not present in blood or is present in
minute quantities below the sensitivity of current detec-
tion methodologies.
However, since infection with BLV results in a
persistent anti-BLV antibody titer, it is thought that
some constant but low level of expression must occur
perhaps compartmentalized to such organs as lymph
nodes, spleen, or bone marrow. Several studies have
* Corresponding author. Tel.: �/1-519-824-4120x54907; fax: �/1-
519-763-1450.
E-mail address: [email protected] (M. van den Heuvel).
Journal of Virological Methods 111 (2003) 61�/67
www.elsevier.com/locate/jviromet
0166-0934/03/$ - see front matter # 2003 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0166-0934(03)00148-4

determined that BLV expression is up-regulated rapidly
if PBLs are cultured (Mirsky et al., 1996; Gupta and
Ferrer, 1982; Miller et al., 1969; Taylor and Jacobs,
1993; Zandomeni et al., 1992).The difficulty in detecting viral proteins in freshly
isolated peripheral blood mononuclear cells (PBMCs) is
thought to be due to the presence of a host factor which
blocks transcription of the integrated provirus (Gupta
and Ferrer, 1982; Taylor and Jacobs, 1993; Zandomeni
et al., 1992). This phenomenon has also been observed
in other retroviral infections, notably HTLV I and II
and HIV I (Castro et al., 1991; Garcia-Blanco andCullen, 1991), making BLV an interesting model to
study mechanisms of host response to retroviruses.
Characterization and purification of factors that
decrease BLV production require an assay to monitor
BLV production. A sandwich enzyme-linked immuno-
sorbent assay (sELISA), based on the method used to
detect the 51 kDa envelope glycoprotein (gp51), de-
scribed by Portetelle et al. (1989), was evaluated for itsability to detect the more strongly expressed major core
protein, p24 (24 kDa), as a measure of viral production
in short-term cultures of PBMCs.
Fetal lamb kidney (FLK) cells (Jensen et al., 1990)
infected with BLV were cultured in Dulbecco’s modified
Minimum Essential Medium (DMEM) supplemented
with 10% heat inactivated fetal bovine serum (FBS)
(Life Technologies, Burlington, Ontario) and 0.05 mg/ml final concentration of gentamycin (Fisher Scientific,
Nepean, Ontario). Cell cultures were trypsinized (Life
Technologies) for 10 min at room temperature (RT) and
washed in phosphate buffered saline (PBS) pH 7. Cell
pellets, held at 4 8C, were lysed with a 10% aqueous
solution of n -octyl-bD-glucopyranoside (Sigma�/Al-
drich, Oakville, Ontario), then clarified by centrifuga-
tion at 12 000�/g for 10 min to be used as a source ofBLV p24.
Murine monoclonal anti-p24 antibodies (D. Porte-
telle, Gembloux, Belgium) were isotyped according to
the manufacturer’s instructions (Serotec, Raleigh, NC).
The 4?G9 and 4?F5 antibodies were found to be IgG1
isotype, while 2?C1 was an IgG2a isotype.
An immunosorbent column was prepared using the
2?C1 antibody according to manufacturer’s instructions(AvidChrome-Hydrazide, Sigma�/Aldrich). Briefly, the
monoclonal antibody was desalted and oxidized for 30
min at RT with 0.1 ml of 0.1 M NaIO4 prior to coupling
to the cartridge matrix. The column was incubated at
RT for 15 min, then rinsed with PBS to elute unbound
antibody.
The FLK lysate was applied to the column at a flow
rate of 0.2 ml/min, followed by two washes with PBS toremove unbound proteins. The p24 was eluted with 0.1
M NaAc, pH 3.5 and collected in 1.5 ml microfuge tubes
containing 500 ml 2 M Tris�/buffered saline, pH 8.8.
Protein in the eluate was monitored at 280 nm. Aliquots
of the protein-containing peaks were pooled and stored
frozen at �/20 8C for use as a standard in the p24
sELISA. The protein concentration in the pooled
fractions was determined by a dye-binding microproteinprocedure using albumin standards.
Two BLV-seropositive Holstein cows infected natu-
rally and one BLV-seronegative cow were housed
separately according to Canadian Council on Animal
Care Guidelines. BLV-status was confirmed by repeated
agar gel immunodiffusion tests. One of the BLV-infected
cows also had PL as determined by repeated hematolo-
gical examinations.Venous blood was collected into acid citrate antic-
oagulant and separated by centrifugation for 25 min at
800�/g at 20 8C. The buffy coat was diluted 1:5 with
sterile PBS. Fifteen milliliters of buffy coat cell suspen-
sion were layered onto 15 ml of Histopaque (Sigma�/
Aldrich) and centrifuged at 400�/g for 30 min at 20 8C.
PBMCs at the interface were removed, diluted 1:5 with
PBS, centrifuged at 100�/g for 10 min then washedtwice with PBS. The cells were diluted to 2�/106 viable
cells per ml of RPMI medium supplemented with 10%
heat inactivated FBS, L-glutamine (0.3 g/l) and 0.02%
amikacin. Each well of a 24-well plate (Falcon, VWR
Canlab, Mississauga, Ontario) contained a total volume
of 2 ml. Plates were incubated at 37 8C for 24 h then
centrifuged at 100�/g to pellet the cells which were then
washed twice with PBS. The cells were re-suspended in 1ml PBS, counted and the viability determined by trypan
blue exclusion. The remaining cells were pelleted then
lysed with 100 ml 10% n-octyl-bD-glucopyranoside in
water. They were kept on ice for 10 min, then 900 ml of
PBS were added to the lysates. The plates were
centrifuged at 500�/g for 3 min to pellet cellular debris.
An sELISA used for measuring BLV gp51 (Portetelle
et al., 1989) was adapted for evaluation. Each well of a96-well ELISA plate (Nunc Immunosorb, Life Technol-
ogies) was coated with 300 ng 4?G9 in 100 ml PBS and
incubated at 4 8C for 4�/6 h on a shaking table. The
plate was then washed twice with washing buffer. Excess
binding sites were occupied by adding 100 ml of
saturation buffer (10% bovine serum albumin in PBS,
pH 7) to each well and incubating for 10 min prior to
adding 100 ml of blocking buffer. Triplicate 100 mlaliquots of protein from the clarified cell lysate of each
PBMC culture were added to individual wells. Duplicate
samples of the p24 standard were also applied and these
were diluted five times in triple dilutions, yielding
standards from 11 to 900 ng. The plate was incubated
overnight at 4 8C on a shaking table. The following day,
the plate was washed three times with washing buffer,
then 200 mg each of peroxidase conjugated 2?C1 and4?F5 in 100 ml washing buffer were added to each well
and incubated at 4 8C for 1 h on a shaking table. The
plate was washed four times with washing buffer prior
to adding 100 ml of tetra-methyl benzidine (Kirkegaard
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/6762
and Perry Laboratories, Gaithersburg, MD) to each
well. After 5�/10 min, the reaction was stopped by
adding 100 ml of 2 N H2SO4 to each well. The
absorbence at 450 nm for each sample was read on a
microplate reader (Ceres, Biotech Instruments, Wi-
nooski, VT). A calibration program (Kineti-Calc II,
KCJR, EIA Software, Ceres) was used to determine p24
(ng) in each sample based on the p24 standards. It was
noted in pilot experiments that cell viability decreased
under some manipulations, so as to account for the loss
of p24 expression associated with dying or dead cells,
the total amount of p24 detected in 100 ml of cell lysate
was divided by the number of viable cells harvested,
yielding ng of p24 per 105 viable cells.
A series of experiments were carried out in triplicate
to determine the optimal conditions for p24 production
by BLV-infected PBMCs. Variables examined were
culture medium, cell concentration, pH, % FBS, and
duration of cell culture.
The data were summarized using descriptive statistics.
Least squares regression analysis was used to assess
linearity. Differences between treatments were assessed
using analysis of variance (ANOVA) and the Bonferroni
post-test. Differences were considered significant if P 5/
0.05.
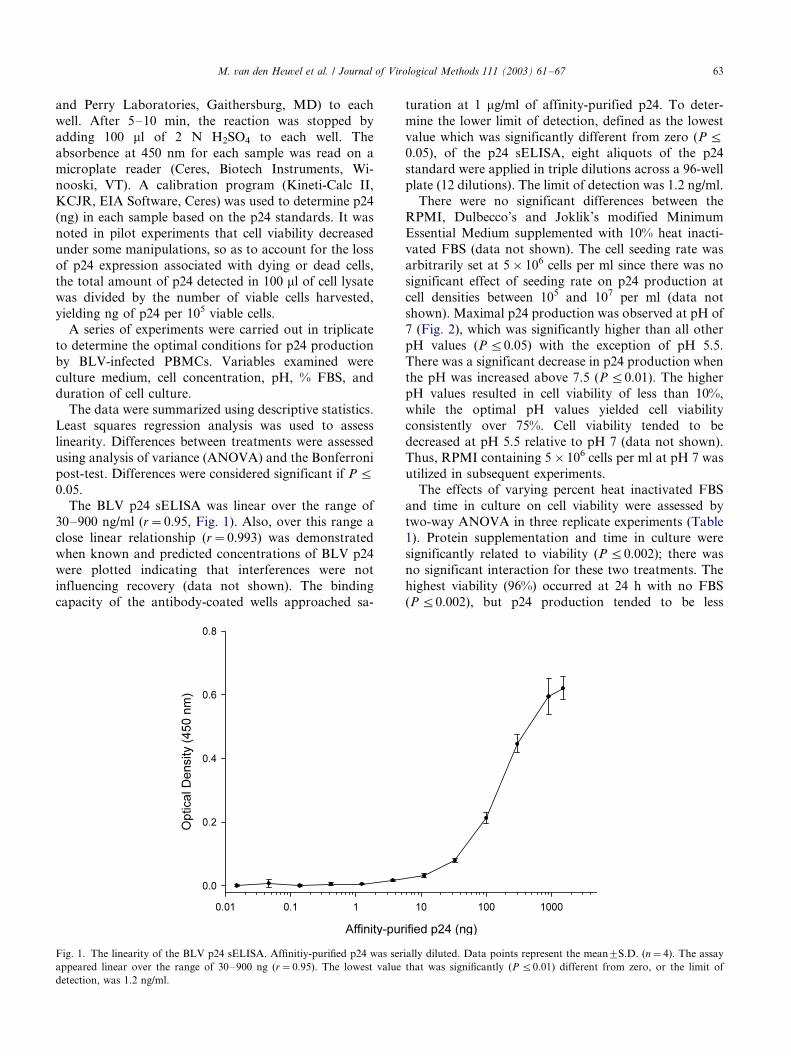
The BLV p24 sELISA was linear over the range of
30�/900 ng/ml (r�/0.95, Fig. 1). Also, over this range a
close linear relationship (r�/0.993) was demonstrated
when known and predicted concentrations of BLV p24
were plotted indicating that interferences were not
influencing recovery (data not shown). The binding
capacity of the antibody-coated wells approached sa-
turation at 1 mg/ml of affinity-purified p24. To deter-
mine the lower limit of detection, defined as the lowest
value which was significantly different from zero (P 5/
0.05), of the p24 sELISA, eight aliquots of the p24
standard were applied in triple dilutions across a 96-well
plate (12 dilutions). The limit of detection was 1.2 ng/ml.
There were no significant differences between the
RPMI, Dulbecco’s and Joklik’s modified Minimum
Essential Medium supplemented with 10% heat inacti-
vated FBS (data not shown). The cell seeding rate was
arbitrarily set at 5�/106 cells per ml since there was no
significant effect of seeding rate on p24 production at
cell densities between 105 and 107 per ml (data not
shown). Maximal p24 production was observed at pH of
7 (Fig. 2), which was significantly higher than all other
pH values (P 5/0.05) with the exception of pH 5.5.
There was a significant decrease in p24 production when
the pH was increased above 7.5 (P 5/0.01). The higher
pH values resulted in cell viability of less than 10%,
while the optimal pH values yielded cell viability
consistently over 75%. Cell viability tended to be
decreased at pH 5.5 relative to pH 7 (data not shown).
Thus, RPMI containing 5�/106 cells per ml at pH 7 was
utilized in subsequent experiments.
The effects of varying percent heat inactivated FBS
and time in culture on cell viability were assessed by
two-way ANOVA in three replicate experiments (Table
1). Protein supplementation and time in culture were
significantly related to viability (P 5/0.002); there was
no significant interaction for these two treatments. The
highest viability (96%) occurred at 24 h with no FBS
(P 5/0.002), but p24 production tended to be less
Fig. 1. The linearity of the BLV p24 sELISA. Affinitiy-purified p24 was serially diluted. Data points represent the mean9/S.D. (n�/4). The assay
appeared linear over the range of 30�/900 ng (r�/0.95). The lowest value that was significantly (P 5/0.01) different from zero, or the limit of
detection, was 1.2 ng/ml.
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/67 63

without the addition of 10% FBS (Fig. 3). There were no
significant differences in viability between cultures with
5, 10, 15, or 20% FBS (Table 1). Increasing FBS above
10% was associated with a significant decrease (Fig. 3,
P 5/0.05) in p24 production and a trend toward
decreasing viability (Table 1). Presumably, the balance
between stimulatory and inhibitory factors that may be
present in FBS (Zandomeni et al., 1992) was altered
such that, above 10%, inhibition of p24 production
resulted. The p24 production tended to increase with the
duration of culture, however, the differences between 24,
36, 48, and 72 h in culture were not significant (Fig. 4).
Viability was highest at 12 and 24 h and significantly
decreased at subsequent times (Table 1). It appeared
that 24 h cultures utilizing RPMI supplemented with
10% FBS resulted in a reasonable balance of cell
viability and p24 production.
To test the reproducibility of the optimized assay, 108
PBMCs were collected, as described above, from two
BLV-infected cows one of which had PL and produced
high amounts of p24 and another with a normal white
blood cell count that produced relatively low amounts
of p24 in culture. PBMCs were cultured in 100 ml of
RPMI supplemented with 10% heat inactivated FBS for
24 h, then lysed, processed as described above and
stored in aliquots at �/20 8C. The affinitity-purified p24
standard was included as a third test sample. Replicates
of each of the three samples were used to test the intra-
assay and inter-assay variability. Coefficients of varia-
tion (CV) were 9.2�/13.3% for intra-assay (Table 2) and
5.1�/12.9% for inter-assay (Table 3) testing.
A commercially available test for HIV p24 (NEN Life
Sciences, MA) has an intra-assay CV of 2�/4% and inter-
assay CV of 13�/23%. Therefore, this BLV p24 sELISA
Fig. 2. The effect of pH of culture medium on expression of p24 by PBMCs in a 24 h culture from a BLV-infected cow with PL. Mean values (9/S.D.,
n�/3) of p24 produced per 105 viable cells. Cells were cultured in RPMI with 10% heat inactivated FBS. * Indicates significant decline relative to pH
7.0 (P 5/0.01).
Table 1
Effects of time (h) and heat inactivated FBS (%) on viability (mean%9/S.D., n�/3) of cultured PBMCsa
FBS (%) in medium Time (h) in culture
12 24 36 48 72
0 77%9/12.5 969/3.6 729/15.5 719/21.6 759/5.3
5 499/7.7 709/14.8 479/9.5 459/12.3 369/4.2
10 689/5.6 759/8.7 429/10.5 319/12.9 369/3.1
15 479/15.6 529/17.1 409/13.2 439/8.1 339/3.5
20 509/9.7 689/21.5 519/12.6 239/5.7 529/9.5
a Viability was assessed by trypan blue exclusion. Data were analyzed using two-way ANOVA and the Bonferroni post-test. Significant effects of
FBS and time (P 5/0.002). No significant FBS�/time interaction. Viability in 0% FBS was significantly greater than 5, 10, 15 or 20% FBS (P 5/
0.002). There were no significant differences between 5, 10, 15 or 20% FBS. Viability was not different between 12 and 24 h but at 24 h viability was
significantly greater than 36, 48, or 72 h (P 5/0.002).
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/6764

yielded inter-assay results comparable to the commercial
HIV p24 test, but had greater within-assay variation.
The latter deficiency could be compensated for by
taking the mean of a larger number of replicates.
The BLV p24 sELISA was unable to detect p24 in
freshly isolated PBMCs, and was just capable of
differentiating expression of p24 in cultured PBMCs
from an asymptomatic BLV-infected cow from back-
ground. Lysates of cells from seronegative sources were
always negative for p24, indicating that no cross-
reaction was occurring between the antibodies and
cellular proteins. Thus, this sELISA is not useful as a
diagnostic test for BLV but, it appeared satisfactory to
determine differences in expression of BLV p24 in
PBMCs from a cow with PL.
The BLV p24 sELISA lends itself well for larger
experiments where the effects of several variables can be
simultaneously assessed. PBMCs were cultured in 24-
Fig. 3. The effect of heat inactivated FBS (%) supplementation of RPMI on expression of p24 by PBMCs in a 24 h culture from a BLV-infected cow
with PL. Mean values (9/S.D., n�/3) of p24 produced per 105 viable cells. Symbols over bars indicate significant reduction relative to 10% FBS (*,
P 5/0.01; �/, P 5/0.05).
Fig. 4. The effect of time (h) in culture on expression of p24 by PBMCs from a BLV-infected cow with PL. PBMCs were cultured in RPMI with 10%
heat inactivated FBS. Mean values (9/S.D., n�/3) of p24 produced per 105 viable cells. There were no significant differences in p24 between 24, 36,
38, or 72 h, however, p24 at 12 h was significantly decreased (*, P 5/0.01).
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/67 65

well plastic plates, which were easily harvested, washed
and processed in the plate, using a centrifuge culture
plate adaptor. The test is relatively fast; requiring about
6 h of labor over 2.5 days. In addition, the use of a 96-
well plate for analysis allows up to 28 samples to be
assayed in triplicate on one plate, therefore, limiting the
error between plates.
The BLV p24 sELISA has specificity and ease of
measurement offered by monoclonal antibodies and a
colourimetric end-point. In contrast, earlier assays have
utilized polyclonal antibodies and/or radioisotopes (Ban
et al., 1991; Cowley et al., 1992; Schmerr et al., 1980).
The assay is less cumbersome than earlier ones based
upon biological viral characteristics such as syncytium
induction (Esteban et al., 1985; Schmerr et al., 1980) or
reverse transcriptase activity (Graves et al., 1977).
In conclusion, the BLV p24 sELISA has potential
usefulness for determining viral expression in cultured
PBMCs from BLV-infected symptomatic (i.e. PL) cattle
but was challenged by the low levels of p24 produced in
cultured PBMCs from hematologically normal BLV-
infected cattle. As anticipated, the assay did not detect
BLV p24 in PBMCs isolated freshly.
Acknowledgements
This research was funded by grants from the Natural
Sciences and Engineering Research Council of Canada
and the Ontario Ministry of Agriculture, Food, and
Rural Affairs. M. van den Heuvel was the recipient of a
Dairy Farmers of Ontario doctoral scholarship.
References
Ban, J., Altanerova, V., Orlik, O., Altaner, C., 1991. Antigen capture
assay for detection of bovine leukemia virus proteins by mono-
clonal antibodies. Neoplasma 38, 625�/631.
Burny, A., Bex, F., Chantrenne, H., Cleuter, Y., Dekegel, D.,
Ghysdael, J., Kettmann, R., Leclercq, M., Leunen, J., Mammer-
ickx, M., Portetelle, D., 1978. Bovine leukemia virus involvement
in enzootic bovine leukosis. Adv. Cancer Res. 28, 251�/311.
Burny, A., Bruck, C., Cleuter, Y., Couez, D., Deschamps, J., Gregoire,
D., Ghysdael, J., Kettmann, R., Mammerickx, M., Marbaix, G.,
1985. Bovine leukemia virus and enzootic bovine leukosis. Onder-
stepoort J. Vet. Res. 52, 133�/144.
Burny, A., Cleuter, Y., Kettmann, R., Mammerickx, M., Marbaix, G.,
Portetelle, D., Van den Broeke, A., Willems, L., Thomas, R., 1988.
Bovine leukemia: facts and hypotheses derived from the study of an
infectious cancer. Adv. Vet. Sci. Comp. Med. 32, 149�/170.
Castro, B.A., Walker, C.M., Eichberg, J.W., Levy, J.A., 1991.
Suppression of human immunodeficiency virus replication by
CD8�/ cells from infected and uninfected chimpanzees. Cell.
Immunol. 132, 246�/255.
Cowley, J.A., Molloy, J.B., Dimmock, C.K., Walker, P.J., Bruyeres,
A.G., Ward, W.H., 1992. Infectivity of bovine leukaemia virus
infected cattle: an ELISA for detecting antigens expressed in in
vitro cultured lymphocytes. Vet. Microbiol. 30, 137�/150.
Esteban, E.N., Thorn, R.M., Ferrer, J.R., 1985. An amplified
immunoperoxidase assay to detect bovine leukemia virus expres-
sion: development and comparison with other assays. Cancer Res.
45, 3231�/3235.
Ferrer, J.F., 1980. Bovine lymphosarcoma. Adv. Vet. Sci. Comp. Med.
24, 1�/68.
Garcia-Blanco, M.A., Cullen, B.R., 1991. Molecular basis of latency in
pathogenic human viruses. Science 254, 815�/820.
Graves, D.C., Diglio, C., Ferrer, J.F., 1977. A reverse transcriptase
assay for detection of the bovine leukemia virus. Am. J. Vet. Res.
38, 1739�/1744.
Gupta, P., Ferrer, J.F., 1982. Expression of bovine leukemia virus
genome is blocked by a nonimmunoglobulin protein in plasma
from infected cattle. Science 215, 405�/407.
Jensen, W.A., Sheehy, S.E., Fox, M.H., Davis, W.C., Cockerell, G.L.,
1990. In vitro expression of bovine leukemia virus in isolated B-
Table 2
Intra-assay variability in optical density (OD450 nm) of the BLV p24 sELISA
Sample Number of replicatesb Mean OD450 nm Standard deviation (S.D.) CV (%)
Lysate from an asymptomatic cowa 20 0.099 0.031 13.3
Lysate from a cow with PL 20 0.642 0.062 9.8
Affinity-purified p24 17 0.608 0.056 9.2
a Lysate refers to the contents of cultured PBMCs lysed by the addition of a 10% aqueous solution of n -octyl-bD-glucopyranoside.b Indicates that a sample was repeated 17 or 20 times on one occasion.
Table 3
Inter-assay variability in optical density (OD450 nm) of the BLV p24 sELISA
Sample Number of replicatesb Mean OD450 nm S.D. CV (%)
Lysate from an asymptomatic cowa 6 0.092 0.022 5.1
Lysate from a cow with PL 6 0.555 0.052 9.4
Affinity-purified p24 20 0.548 0.062 12.9
a Defined in Table 2.b Indicates that a sample was repeated in triplicate on 6 or 20 occasions over the duration of the study.
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/6766

lymphocytes of cattle and sheep. Vet. Immunol. Immunopathol.
26, 333�/342.
Kettmann, R., Marbaix, G., Cleuter, Y., Portetelle, D., Mammerickx,
M., Burny, A., 1980. Genomic integration of bovine leukemia
provirus and lack of viral RNA expression in the target cells of
cattle with different responses to BLV infection. Leuk. Res. 4, 509�/
519.
Kettmann, R., Deschamps, J., Cleuter, Y., Couez, D., Burny, A.,
Marbaix, G., 1982. Leukemogenesis by bovine leukemia virus:
proviral DNA integration and lack of RNA expression of viral
long terminal repeat and 3? proximate cellular sequences. Proc.
Natl. Acad. Sci. USA 79, 2465�/2469.
Miller, J.M., Miller, L.D., Olson, C., Gillette, K.G., 1969. Virus-like
particles in phytohemagglutinin-stimulated lymphocyte cultures
with reference to bovine lymphosarcoma. J. Natl. Cancer Inst.
43, 1297�/1305.
Mirsky, M.L., Olmstead, C.A., Da, Y., Lewin, H.A., 1996. The
prevalence of proviral bovine leukemia virus in peripheral blood
mononuclear cells at two subclinical stages of infection. J. Virol. 70,
2178�/2183.
Portetelle, D., Mammerickx, M., Burny, A., 1989. Use of two
monoclonal antibodies in an ELISA test for the detection of
antibodies to bovine leukaemia virus envelope protein gp51. J.
Virol. Methods 23, 211�/222.
Schmerr, M.F., VanDerMaaten, M.J., Miller, J.M., 1980. Application
of a radioimmunoassay for detection of the major internal antigen
(p24) of bovine leukemia virus from cultured lymphocytes of cattle.
Comp. Immunol. Microbiol. Infect. Dis. 3, 327�/336.
Taylor, J.A., Jacobs, R.M., 1993. Effects of plasma and serum on the
in vitro expression of bovine leukemia virus. Lab. Invest. 69, 340�/
346.
Zandomeni, R.O., Carrera-Zandomeni, M., Esteban, E., Donawick,
W., Ferrer, J.F., 1992. Induction and inhibition of bovine leukemia
virus expression in naturally infected cells. J. Gen. Virol. 73, 1915�/
1924.
M. van den Heuvel et al. / Journal of Virological Methods 111 (2003) 61�/67 67