
ACTA BOTANICA - uniba.sk · 3 Acta Botanica Universitatis Comenianae Vol. 48, 2013 BRYOPHYTES OF...
44
ACTA BOTANICA UNIVERSITATIS COMENIANAE Volume 48 2013 COMENIUS UNIVERSITY IN BRATISLAVA
Transcript of ACTA BOTANICA - uniba.sk · 3 Acta Botanica Universitatis Comenianae Vol. 48, 2013 BRYOPHYTES OF...

ACTA BOTANICA UNIVERSITATIS COMENIANAE
Volume 48
2013 COMENIUS UNIVERSITY IN BRATISLAVA

The journal was edited with the title / Časopis bol vydávaný pod názvom Acta Facultatis Rerum Naturalium Universitatis Comenianae, Botanica Editor in Chief / Predseda redakčnej rady Karol Mičieta; [email protected] Executive Editor / Výkonný redaktor Soňa Jančovičová, [email protected] Editorial Board / Členovia redakčnej rady Dana Bernátová, Danica Černušáková, Zuzana Dúbravcová, Katarína Mišíková Editor Ship / Adresa redakcie Redakcia Acta Botanica, Révová 39, SK-811 02 Bratislava 1; Tel. ++421 2 54411541; Fax ++421 2 54415603 Published by / Vydavateľ © Comenius University in Bratislava, 2013 © Univerzita Komenského v Bratislave, 2013 ISBN 978-80-223-3561-4 ISSN 0524-2371

3
Acta Botanica Universitatis Comenianae Vol. 48, 2013
BRYOPHYTES OF THE VILLAGE MARIANKA
(SOUTH WESTERN SLOVAKIA)
Katarína Mišíková•
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany,
Révová 39, 811 02 Bratislava, Slovakia
Received 13 November 2013; Received in revised form 28 November 2013; Accepted 3 December 2013
Abstract
The bryophyte flora of the village Marianka on southwestern foothills of the Malé Karpaty Mts. was investigated. Overall, 77 bryophytes were recorded; out of them 12 liverworts (Marchantiophyta), one hornwort (Anthocerotophyta) and 64 mosses (Bryophyta). Redlisted species Anthoceros agrestis, Entosthodon fascicularis and Weissia brachycarpa were, unfortunately, unconfirmed at present as their sites had been destroyed by a housing construction. In the monitored rural environment, hemerophilous bryophytes and species less sensitive against pollution dominated here.
Key words: rural environment, hemeroby, Marchantiophyta, Anthocerotophyta, Bryophyta
Introduction
The village Marianka is located on the southwestern foothills of the Malé Karpaty Mts., along the valley of the Mariánsky potok stream. Marianka is one of the oldest and most famous pilgrimage sites in Slovakia. The first mention of Marianka comes from the year 1367. Its oldest church is the Virgin Mary church, which cornerstone was laid by the king Louis I. of Anjou in 1377. During its consecration in 1380, a statue of the Virgin Mary was passed on the altar. The church and nearby situated monastery were donated to the order hermits of St. Paul, who governed them over followed 400 years (Anonymus 1, 2).
Marianka is also known for slate mining. Slate was mined in its vicinity already in the 17th century, mostly for the needs of the local population. An intensive mining began in the 19th century and the best known product became the slate-boards. After the First World War, the factory of the slate-board production folded (Anonymus 1).
From the bryological viewpoint, the most valuable sites are in the upper part of the village, in the Sväté údolie valley along Mariánsky potok stream. This part is covered with beech forest. Due to microclimatic conditions (relatively stable year-round higher air humidity), several liverworts occur here, which are rare or absent in other settlements near the Malé Karpaty Mts.
The village and its surrounding area have not been bryologically investigated. However, several floristic data are mentioned in the works by Peciar (1981), Janovicová (1992), Kresáňová et al. (2005).
Material and Methods
Bryological research in the village Marianka (Tab. 1) was carried out during autumn 2012 and spring 2013, except extensively managed fields, which were examined in 1997 and 2000. Samples were collected from various substrates in following habitats – beech forest (BF) in Sväté údolie valley, alder woods (AF), rural environment (RU), cemetery (CE) and extensively managed fields (FI). The nomenclature of liverworts follows the study by Söderström et al. (2002); nomenclature of bryophytes
• Corresponding Author: Katarína Mišíková; [email protected]

4
is based on the work by Hill et al. (2006). Threat categories follow Kubinská et al. (2001), degree of hemeroby (HB) and air quality (AP) are listed according to Düll (2010).
Tab. 1. Brief description of the village Marianka
Altitude 220 m a. s. l. (Anonymus 2)
Area size 3.22 km² (322 ha) (Anonymus 2)
Population 1 355 (31 Dec 2011) (Anonymus 2)
GPS coordinates N 48°14'55" E 17°03'41" (Anonymus 3)
Tab. 2. List of species recorded in the village Marianka
List of species / Red list category References ∗∗∗∗Habitats ∗∗∗∗∗∗∗∗Ecological groups
Marchantiophyta
Frullania dilatata BF C2
Chiloscyphus palescens Janovicová (1992) BF A4 B1
Conocephalum conicum Janovicová (1992) BF A3
Lophocolea heterophylla BF B1 D
Marchantia polymorpha RU BF A3 B2
Metzgeria furcata BF C2
Pellia endiviifolia BF A3 B1
Plagiochila porelloides BF A2 A3
Porella platyphylla BF C2
Radula complanata Peciar (1981) BF C2
Riccia glauca Kresáňová et al. (2005) FI B2
Riccia sorocarpa Kresáňová et al. (2005) FI B2
Anthocerothophyta
Anthoceros agrestis LR: nt Kresáňová et al. (2005) FI B2
Bryophyta
Amblystegium serpens RU AF BF CE A1 A2 A3 B2 C1 D
Anomodon attenuatus BF C1
Atrichum undulatum RU BF B1 B3
Barbula convoluta RU B2
Barbula unguiculata Kresáňová et al. (2005) FI RU AF B2
Brachytheciastrum velutinum BF B1
Brachythecium albicans Kresáňová et al. (2005) FI CE B2 B3
Brachythecium rutabulum AF BF CE A1 A3 B1 B3 C1 D
Brachythecium salebrosum CE RU B3 C1
Bryoerythrophyllum recurvirostre BF A1
Bryum argenteum Kresáňová et al. (2005) FI RU CE A1 B2
Bryum bicolor RU B2
Bryum caespiticium RU A1 B2
Bryum capillare RU A1 B2
Bryum moravicum BF C1 C2
Bryum rubens FI RU B2
Bryum ruderale FI B2
Ceratodon purpureus Kresáňová et al. (2005) RU CE FI A1 B2
Cirriphyllum piliferum RU CE B3
Dicranella heteromalla BF B1
Dicranella staphylina FI B2
Dicranum montanum BF C1
Dicranum scoparium BF B1
Didymodon fallax RU AF A1

5
Tab. 2. – Continuation
List of species / Red list category References ∗∗∗∗Habitats ∗∗∗∗∗∗∗∗Ecological groups
Didymodon ferrugineus RU B2
Entosthodon fascicularis LR: nt Kresáňová et al. (2005) FI B2
Eurhynchium angustirete Peciar (1981) BF B2
Fissidens bryoides BF B2
Fissidens taxifolius Peciar (1981) BF B2
Funaria hygrometrica RU AF B2
Grimmia pulvinata CE RU A1
Hygroamblystegium tenax BF AF A3
Hypnum cupressiforme AF BF CE RU A1 C1 C2 D
Isothecium alopecuroides BF A2
Kindbergia praelonga RU B2 B3
Leucodon sciuroides BF C1 C2
Mnium hornum BF A3
Mnium stellare Peciar (1981) BF A1
Orthotrichum affine AF C2
Orthotrichum anomalum CE RU A1
Orthotrichum diaphanum AF BF C2
Oxyrrhynchium hians RU AF CE B2
Phascum cuspidatum Kresáňová et al. (2005) FI RU B2
Physcomitrium pyriforme AF B2
Plagiomnium cuspidatum BF CE A3 B1
Plagiomnium rostratum RU B3
Plagiomnium undulatum RU BF CE B1 B3
Plagiothecium denticulatum BF B1
Platyhypnidium riparioides BF A4
Pohlia nutans BF B1
Pohlia wahlenbergii Peciar (1981) BF B2
Polytrichastrum formosum BF B1
Pteryginandrum filiforme BF C1
Rhizomnium punctatum Peciar (1981) BF A3 B1
Rhynchostegium murale RU A1
Sciuro-hypnum populeum CE RU A1
Schistidium apocarpum RU A1
Schistidium crassipilum RU A1
Syntrichia ruralis RU A1
Thuidium delicatulum Peciar (1981) BF B3
Tortula modica Kresáňová et al. (2005) FI B2
Tortula muralis CE RU A1
Tortula truncata FI RU B2
Weissia brachycarpa DD Peciar (1981) BF B2 ∗Habitats: BF – beech forest, AF – alder woods, RU – rural environment, CE – cemetery, FI – extensively managed fields ∗∗Ecological groups: A. Epilithic species: A1 – concrete and stone walls, fences; A2 – rocks and boulders in the forest; A3 – stream banks;
A4 – stream; B. Epigeic species: B1 – forest soil; B2 – bare damp soil; B3 – lawns, meadows; C. Epiphytic species: C1 – tree bases and trunks up to 50 cm above ground level; C2 – tree trunks at height 50 cm above ground level and higher; D. Epixylic species
Results
Overall, 77 bryophytes were found, out of them12 liverworts, one hornwort and 64 mosses (Tab. 2). Three species are redlisted – Anthoceros agrestis (LR: nt), Entosthodon fascicularis (LR: nt) and Weissia
brachycarpa (DD) (Kubinská et al. 2001). These species were recorded on extensively managed fields in years 1997 and 2000. Unfortunately, the sites were later changed to building plots. Recently, fields of a similar species composition could not be found in Marianka.

6
Most species were found in the beech forest of the Sväté údolie valley (41) and in the rural environ-ment (31), least in the alder stand fragments (11) (Fig. 1). In relation to ecological groups, the highest number of species represents terrestrial sites on bare damp soil (B2) and epilithic sites on concrete and stony walls (A1) (Tab. 2, Fig. 2).
41
11
31
15 14
0
10
20
30
40
50
BF AF RU CE FI
Habitats
Nu
mb
er
of
sp
ecie
s
Fig. 1. ∗∗∗∗Species number in the individual habitats ∗for abbreviations see Tab. 2.
18
3
10
2
14
30
9 9 9
4
0
5
10
15
20
25
30
35
A1 A2 A3 A4 B1 B2 B3 C1 C2 D
Ecological groups
Nu
mb
er
of
sp
ecie
s
Fig. 2. ∗∗∗∗Species number in the individual ecological groups ∗for abbreviations see Tab. 2.
Degrees of hemeroby (HB) reflect the intensity of anthropogenic impacts on the landscape or habitats
(Zechmeister, Moser 2001; Zechmeister et al. 2002); from this viewpoint, the most recorded species are eu- and mesohemerobic (HB5, HB6, HB7) (Fig. 3). Euhemerobic species occur at predominantly anthropogenic sites – ruderal communities, extensively managed meadows, pastures and fields, gardens, stone and concrete walls, banks of eutrophied streams. Mesohemerobic bryophytes are particularly pioneer taxa growing on ruderal, as well as natural sites (e.g. root stocks, cutoff lake bottoms, stream banks), or other substrates and habitats close to natural. From the oligohemerobic bryophytes (HB8), sensitive to anthropogenic impacts, only Pterigynandrum filiforme was detected, growing on the bark of beech.

7
7
8
11
16
18
16
1
0 5 10 15 20
HB2
HB3
HB4
HB5
HB6
HB7
HB8
Number of species
Fig. 3. Degrees of hemeroby (Düll 2010) and species number
AP values (air purity / air pollution) (Düll 2010) indicate the level of sensitivity of different species
to SO2 air pollution. Slightly to moderately sensitive species (AP1 – AP4) can grow in an environment with SO2 pollution from 0.11 to 0.16 mg/m3. The so-called indicators of air purity (AP5 – AP9) occur mainly in areas where SO2 values are lower than 0.085 mg/m3. In the study area, less sensitive species to SO2 pollution are over-represented (49 taxa) than sensitive bryophytes (15 taxa) (Tab. 3, Fig. 4).
Tab. 3. Number of species in categories of AP values (Düll 2010)
Air pollution indicators AP1 – AP4
Number of species Air purity indicators AP5 – AP8
Number of species
AP1 11 AP5 10
AP2 9 AP6 2
AP3 18 AP7 1
AP4 11 AP8 2
11
10
9
2
18
1
11
2
0 10 20 30 40 50 60
AP1 - AP4
AP5 - AP8
Number of species
Fig. 4. Air pollution indicators (AP1 – AP4) and Air purity indicators (AP5 – AP8) according to Düll (2010)
In conclusion, the bryophyte diversity of the village Marianka is positively affected by the presence
of wooded valley along the Mariánsky potok stream. Stable climatic conditions and particularly higher air humidity allow the growth of liverworts, generally rare in rural areas of the Malé Karpaty Mts. Since 2000, we have failed to confirm the presence of some agricolous bryophytes such as Anthoceros agrestis and Entosthodon fascicularis, included in the Slovak Red List of bryophytes (Kubinská et al. 2001). Due to anthropogenic pressure the eu- and mesohemerobic species and bryophytes less sensitive to SO2 pollution are prevalent as sensitive ones.

8
Acknowledgements
This study was supported by the Grant Agency VEGA (Grant No. 1/0380/13).
References
Anonymus, (1): http://visit.marianka.sk [accessed 5 Aug 2013]. Anonymus, (2): http://www.e-obce.sk/obec/marianka [accessed 11 Sep 2013]. Anonymus, (3): http://www.mapa-mapy.sk/ [accessed 11 Sep 2013]. Düll, R., 2010: Autoekologie der Moose Mitteleuropas.
http://duell.kilu.de/download/ Autoekologie_der_Moose_07_Sept_2010.pdf [accessed 14 Apr 2013]. Hill, M. O., Bell, N., Bruggeman-Nannenga, M. A., Brugue´, S. M., Cano, M. J., Enroth, J., Flatberg, K. I., Frahm, J. P.,
Gallego, M. T., Garilleti, R., Guerra, J., Hedenäs, L., Holyoak, D. T., Hyvönen, J., Ignatov, M. S., Lara, F., Mazimpaka, V., Munõz, J., Söderström, L., 2006: An annotated checklist of the mosses of Europe and Macaronesia. J. Bryol., 28: 198-267.
Janovicová, K., 1992: Bryoflóra Malých Karpát. MSc thesis depon. in Comenius University in Bratislava. Kresáňová, K., Janovicová-Mišíková, K., Kubinská, K., 2005: Diversity of bryophytes in agro-coenoses of Slovakia.
Biologia, Bratislava, 60, 1: 9-15. Kubinská, A., Janovicová, K., Šoltés, R., 2001: Červený zoznam machorastov Slovenska (december 2001). Ochr. Prír.,
Supl., 20: 31-43. Peciar, V., 1981: Bryoflóra ohrozených oblastí Slovenska I. Acta Fac. Rerum Nat. Univ. Comenianae, Formatio et Protect.
Naturae, 6: 105-118. Söderström, L., Urmi, E., Váňa, J., 2002: Distribution of Hepaticae and Anthocerotae in Europe and Macaronesia. Lindbergia,
27: 3-47. Zechmeister, H., Tribsch, A., Moser, D., Wrbka, T., 2002: Distribution of endangered bryophytes in Austrian agricultural
landscapes. Biological Conservation, 103: 173-182. Zechmeister, M., Moser, D., 2001: The influence of agricultural land-use intensity on bryophyte species richness.
Biodiversity and Conservation, 10: 1609-1625.
Abstrakt
V obci Marianka, ktorá leží na juhozápadnom úpätí Malých Karpát, prebiehal bryologický výskum. Celkovo bolo zaznamenaných 77 druhov machorastov, z toho 12 pečeňoviek (Marchantiophyta), jeden rožtek (Anthocerotophyta) a 64 machov (Bryophyta). Druhy Červeného zoznamu machorastov Slovenska Anthoceros agrestis, Entosthodon fascicularis a Weissia brachycarpa, žiaľ, neboli od roku 2000 v štu-dovanom území znovupotvrdené, nakoľko ich lokality boli zničené pri výstavbe novej štvrte. Celkovo prevládajú hemerofilné a voči znečisteniu ovzdušia relatívne málo citlivé druhy, typické pre rurálne prostredie. Katarína Mišíková: Machorasty obce Marianka (juhozápadné Slovensko)

9
Acta Botanica Universitatis Comenianae Vol. 48, 2013
REVIEW OF THE TAXONOMIC CONCEPTS OF THE INVASIVE SPECIES
AMBROSIA ARTEMISIIFOLIA L. AS THE BASIS FOR THE EVALUATION
OF VARIABILITY OF THIS SPECIES IN EUROPE
Michal Hrabovský•, Karol Mičieta
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany,
Révová 39, 811 02 Bratislava, Slovakia
Received 29 October 2013; Received in revised form 27 November 2013; Accepted 6 December 2013
Abstract
This review contains data on all taxonomic concepts of the common ragweed (Ambrosia artemisiifolia L.) in North America and particularly in Europe. To describe morphological variability of one species, many taxa on
the level species, varieties and forms were differed. Knowledge of the taxonomic conceptions means
understanding the variability of species. This review can serve as a pattern for evaluation of the morphological variability in Europe.
Kew words: Ambrosia artemisiifolia L., morphological variability, taxonomic concepts
Introduction
Ambrosia is one of the numerous genera of the Compositae. It comprises some 40 species growing
mainly in subtropical and temperate climatic zone of South and North America. In the Mediterranean
and North Africa, the genus is represented by only one native species A. maritima L. Other species
found in Europe ‒ A. artemisiifolia L., A. trifida L., A. psilostachya DC, A. tenuifolia Spreng. ‒ were
introduced from America (Hansen 1976). The taxonomy of this genus was very complicated. According
to Payne (1964), the monophyletic genus Ambrosia s.l. consists of two paraphyletic genera Franseria
Cav. and Ambrosia L.
The taxonomic position of the common ragweed (Ambrosia artemisiifolia L.) has gone through many
changes. Recently, most authors prefer the concept of Payne (1970); only some, mainly in the USA,
prefer the concept of Fernald, Griscom (1935), by which three varieties are recognized. In the present
concept of Payne (1970), there is included a high phenotypical and phenological variability that is
represented by many taxa in one species. However, on account of this concept, there is no information
about the variability of this species in Europe. In this study, we bring description of variability in the
native range in North America. It can serve for the evaluation of variability of this species in Europe in
future studies.
Methods
Names and descriptions of Ambrosia taxa were authentically accepted from the original works.
The descriptions were abridged to main differential characteristics. Many taxonomic names related to
A. artemisiifolia are mere synonyms without any taxonomic value and they are only mentioned in
Fig. 1.
• Corresponding author: Michal Hrabovský; [email protected]

10
Results and discussion
From its very beginning, the taxonomic treatment of individuals of the species Ambrosia artemisiifo-
lia L. was based on morphology of leaves and stems. Later, as a result of detection of new morphological
differences, new taxa with other differential characteristics were described. The main sings were apart
from the growth of stem and form of leaf blade also hairiness and form of achenes and involucres
of staminate heads. Altogether, more than 10 taxa in the rank of species, subspecies or variety were
described in North America. For the first time, some of these taxa were included into one species by
Torrey, Gray (1842) who preferred the specific epithet artemisiifolia against elatior. However, this
concept of one species was not accepted at that time or, in this concept, the species name artemisiifolia
was replaced by the name elatior (e.g. Farwell 1913, Hess et al. 1972). Another concept was published
by Rydberg (1922) who classified all known taxa in seven species. On these two concepts, a new
concept was built by Fernald, Griscom (1935), by which one species with three varieties is recognized.
According to Payne (1970), these varieties do not seem to be adequate.
In the next text, we present the differential characteristics which were added to these taxa by different
authors. The relationship among the taxa is sketched in the following Fig. 1.
Fig. 1 Review of taxa and their relationships based on morphology of leaves and branching of the stems in
accordance with the concepts of Rydberg (1922) and of Fernald, Griscom (1935).
Ambrosia artemisiifolia L. Sp. Pl. II: 988 (1753)
– Leaves bipinnatifid or the lower less divided, stem about one foot high, more diffuse, lateral branches overgrow the primary stem.
– Leaves bipinnatifid, the upper pinnatifid, racemes by three (Willdenow 1805).
– Stem four to six feet high, branching; leaves generally bipinnatifid, the segments larger and more distant than in Ambrosia
elatior L., nearly glabrous on the upper surface, pubescent and hoary underneath; racemes scattered, loosely paniculate; heads
of male florets small, female florets remote, axillary, sessile; spines of fruit very short. Grows in the mountains. Flowers
August to September (Elliot 1824).
– Stem 5–10 dm high, leaves pinnatifid or the upper simple, finely strigose; involucre fully 4 mm wide; pistillate flowers in
small clusters in the upper axils; body of fruit 2.5–3 mm long, beak fully 1mm long, spines 5–7, short (Rydberg 1922).
– Leaves simple, coarsely pinnatifid or rarely bipinnatifid; staminate involucres 3‒7 mm broad (Fernald, Griscom 1935).
Ambrosia artemisiifolia L.
[syn.:. A. absynthifolia Michx.;
A. elatior var. artemisiifolia (L.)
Farw.]
Ambrosia elatior L.
Incl. A. artemisiifolia L.
var. α quadricornis Ktze;
A. artemisiifolia γ Torr. & A.
Gray
[syn.: A. elata Salisb.;
A. artemisifolia elatior (L.)
Desc.; A. artemisiifolia α
Torr. & A. Gray]
Ambrosia paniculata Michx. [syn.: Iva monophylla Walt.;
A. artemisiifolia δ. Torr. & A.
Gray; A. monophylla (Walt.) Rydb.;
A. artemisiifolia α jamaicensis
Griseb.]
Ambrosia heterophylla Muhl.
ex Willd.
[syn.: A. artemisiifolia β Torr. & A. Gray;
A. elatior L. var. heterophylla (Muhl.) Farw.]
Ambrosia longistylis Nutt.
Incl. A. artemisiifolia L. var. β
octocornis Ktze
Ambrosia diversifolia (Piper) Rydb.
(syn.: A. artemisiaefolia subsp. diversifolia
Piper)
Ambrosia media Rydb.
Ambrosia glandulosa Rydb.
[syn.: A. artemisiaefolia L. var. paniculata
(Michx.) Blank.; non A. glandulosa Scheele)]

11
Ambrosia elatior L. Sp. Pl. II: 987 (1753) – Leaves bipinnatifid, lateral branches do not overgrow the primary stem; terminal racemes glabrous.
– Leaves bipinnatifid, lateral branches shorter then stem, growing in larger distances, petioles of upper leaves ciliate, what is
different from the previous species (Wildenow 1805).
– Stem four to seven feet high, when young pubescent; leaves all bipinnatifid with acute segments, somewhat hairy; flowers
in paniculate racemes; heads of the male florets globular; fertile florets in small distinct clusters; nut crowned with six short
spines. Grows in pastures and rich soils. Flowers July to September (Elliot 1824).
– Stem 3–10 dm high, branched; leaves bipinnatifid or the upper less divided, hirsutulous-puberulent above, strigose beneath,
and often hirsute on the veins, rachis winged, 1‒3 mm broad; segments lanceolate, acute, directed forward; staminate heads
numerous, or in a predominately pistillate form few or none, in racemes terminating the branches; involucre about 3 mm
wide; pistillate heads in small clusters in the upper axils; body of the fruit 3 mm long, beak more than 1 mm long; spines
5‒7, short (Rydberg 1922).
– Leaves bi–tripinnatifid with small segments; staminate involucres 2.5–5 mm broad (Fernald, Griscom 1935).
Ambrosia paniculata Michx. Flor. Bor. Am. 2: 183 (1803) – Stem branching, paniculate at summit, and with the petioles villous; leaves green on each surface, bipinnatifid, the
segments lanceolate, fruits somewhat clustered, small, slightly armed. It is very similar to A. elatior that differs according
to Linné by glabrate racemes.
– Branches fastigiated; leaves glabrous, bipinnatifid, the upper pinnatifid; racemes solitary (Wildenow 1805).
– Stem two to four feet high, branching, pubescent and hairy, somewhat scabrous; leaves alternate, the lower compoundly,
upper simply pinnatifid, the segments all acute, somewhat hairy and scabrous; flowers in simple racemes; fruit muricate
near the summit. Grows in cultivated ground, very common. Flowers July to September (Elliot 1824).
– Stem 3–12 dm high, hirsutulous and with scattered long hairs, branched; petioles more or less hirsute; blades bipinnatifid to
near the midrib, or the upper pinnate or entire, scabrous-puberulent above, strigose beneath; segments lanceolate, acute,
more or less lobed or toothed; staminate heads numerous, in elongate racemes, involucre 3 mm broad, pistillate heads in
small clusters in the upper axils; fruit rugose, body of the fruit fully 2 mm long, spines 5–7, short, beak 0.5 mm long or less
(Rydberg 1922).
– Leaves bi–tripinnatifid with small segments; staminate involucres 1.5–2.5 mm broad (Fernald, Griscom 1935).
Ambrosia heterophylla Muhl. ex Willd. Sp. Pl. 4: 378 (1805) – Leaves pinnatifid, segments lanceolate, obtuse, a little canescent beneath, petioled, the upper subsessile, entire, lanceolate
or obtuse, with ciliate base by which it differs from another species.
– The lower leaves pinnatifid, the upper undivided or with one or two lobes near the base. This form is more inclined to have
the staminate inflorescence abnormally converted into a pistillate inflorescence (Farwell 1913).
Ambrosia longistylis Nutt. Trans. Am. Phil. Soc. N. S. 17: 344 (1841) – Stem scabrous, simple; leaves pinnatifid, segments oblong-linear, bractes entire; female flowers axillary, conglomerate,
with exceedingly long styles, (about an inch); fruit cornute, spiny at the summit;
male flowers about thirty. Grows in Rocky Mountains.
– Stem 2–3 dm high, scabrous-hispidulous; leaves pinnately divided, 3–7 cm long, scabrous-hispidulous as the stem, dark-
green, strongly ribbed; staminate heads in a terminal raceme; involucre 4–5 mm broad; pistillate heads usually many in
small clusters in the upper axils; style described as being an inch long, but in the type only about 5 mm long; body of the
fruit 3 mm long, beak about 2 mm long; spines 6‒8 (Rydberg 1922).
Ambrosia diversifolia (Piper) Rydb. N. Am. Fl. 33: 18 (1922) – Stem about 5 dm high, strigose-hirsute; lower leaves pinnately divided into oblong or lanceolate divisions, hirsutulous on
both sides, somewhat paler beneath; upper leaves lanceolate or ovate, entire, subsessile, acute; staminate heads numerous,
in racemes terminating the branches; involucre 4–5 mm broad; pistillate heads in few small clusters or solitary in the upper
axils; body of the fruit about 2.5 mm long, beak 2 mm long; spines 4–5.
– Leaves becoming progressively less deeply lobed upward, those of the upper third of the plant mostly entire, these ovate-
lanceolate or lanceolate, acute, narrowed abruptly to a subsessile base, 3-nerved. Grows on the gravelly banks of Almota
Creek (Piper 1906).
Ambrosia media Rydb. Bull. Torr. Bot. Club 37: 127 (1910)
– Stem 4–6 dm high, hispid with ascending or appressed short hairs, more or less strigose, branched; leaves pinnately
divided, thick, 5‒10 cm long, scabrous above, hispid-strigulose beneath, all except the uppermost with shorter or longer,
hirsute-ciliate, narrowly winged petioles ; divisions oblong or lanceolate, usually more or less cleft or toothed, the lobes or
teeth lanceolate, acute; staminate racemes rather dense; 4‒5 mm in diameter; body of the fruit about 3 mm long, with 5‒7
sharp spines, beak about 1 mm long.
Ambrosia glandulosa Rydb. N. Am. Fl. 33: 18 (1922) – An annual herb; stem 5–10 dm high; leaves pinnatifid or bipinnatifid, strigulose on both sides; blades
5–10 cm long; divisions linear-lanceolate, acute or attenuate; staminate heads numerous, in racemes terminating the
branches; involucre 3 mm broad; pistillate heads few in small clusters in the upper axils; body of the fruit 2 mm long, in
age glabrous and smooth, beak less than 0.5 mm long.
– A very slender, smoothish, widely branching form with leaves less divided than in A. artemisiaefolia L., the segments
oblonglanceolate to linear: sterile racemes loosely flowered and heads small (2 mm.); fruit small, with inconspicuous lateral
tubercles. A well-marked variety approaching A. glandulosa Scheele (Blankinship 1907).

12
Notice: There are ambiguous views whether Ambrosia glandulosa Scheele and A. glandulosa Rydb.
are conspecific. Both of them have nearly the same characteristics. According to Payne (1970), the
first of them is a synonym of A. psilostachya DC. The main difference between A. artemisiifolia L. and
A. psilostachya DC is the annual status of the first species and the perennial status of the second one.
These data are not included in the Scheele’s description. However, due to unarmed fruits mentioned in
this description, A. glandulosa Scheele seems to be closer to A. psilostachya DC.
There are also described some varieties based on other morphological characteristics. Kuntze (1891)
has described two varieties: A. artemisiifolia α quadricornis – with 4–5 spines on the fruit – and
A. artemisiifolia β octocornis – with 8–10. According to Rydberg (1922) the first one belongs to
A. elatior L. and the second one to A. longistylis Nutt. There are also known few forms, f. villosa
(Fernald, Griscom 1935), f. aurea (Priszter 1985) and f. atropurpurea (Priszter ex Járai-Komlódi,
Juhász 1993). The first one is based on hairiness of the stem and the others described in Europe are
based on the colour of stem.
Among the synonyms of A. artemisiifolia, A. simplicifolia Raeusch. can also be found. It is an invalid
name, because Räuschel (1797) published it without any description. He was also probably wrong
supposing that A. simplicifolia Walt., which is the synonym of A. trifida L., is a separate species. The
taxon A. artemisiifolia β trinitensis Griseb. is not valid, because the variety belongs to A. psilostachya
DC.
Conclusions
All above mentioned taxa were described owing to the large intraspecific variability of the species
Ambrosia artemisiifolia L. Frequently occurring morphological forms were described as three different
taxa A. artemisiifolia, A. elatior and A. paniculata. The other taxa present the transitive types within these
three previous morphological forms. In efforts to classify these transitive types, they were subjectively
included in these three taxa, e.g., according to Rydberg (1922), A. heterophylla was included into the
taxon A. artemisiifolia. However, very similar A. diversifolia, which differs from the previous only by
acute lobes of the leaf blade, and also A. media and A. longistylis, were included in the taxon A. elatior L.
by Fernald, Griscom (1935). The similar position has A glandulosa, which has intermediate characteristics
between A. artemisiifolia and A. paniculata.
Identification of particular taxa is very difficult and not clear, because their differential characteristics
are in contradiction in sense of different authors. The notice of some authors (e.g. Rydberg 1922, Fernald,
Griscom 1935), that only one form A. elatior was introduced in Europe is wrong. The study of Genton
et al. (2005) on French invasive population of the common ragweed indicated that the genetic diversity
of this species was as high in Europe as in the United States. Therefore, we expect the same high
morphologic diversity in Europe.
Acknowledgements
This study was partially supported by the Grant Agency VEGA, Grant No. 1/0380/13.
References
Blankinship, J. W., 1907: Ambrosia artemisiaefolia L. var. paniculata (Michx.) Blank. Rep. Mo. Bot. Gard., 18: 173.
Elliot, S., 1824: A sketch of the botany of South-Carolina and Georgia 2. J. R. Schenck, Charleston.
Fernald, M. L., Griscom, L., 1935: Ambrosia artemisiaefolia and its variations in temperate eastern North America. Rhodora,
37: 184-185.
Farwell, O. A., 1913: Contribution to the Botany of Michigan No. 9. Rep. Mich. Acad., 15: 190.
Genton, B. J., Shykoff, J. A., Giraud, T., 2005: High genetic diversity in French invasive populations of common ragweed,
Ambrosia artemisiifolia, as a result of multiple sources of introduction. Mol. Ecol., 14(14): 4275-4285.
Hansen, A., 1976: Ambrosia L. In Tutin, T. G. et al. (eds.) Flora Europaea, Vol. 4., p. 142, Cambridge University Press, London.
Hess, H. E., Landolt, E., Hirzel, R. (eds.) 1972: Flora der Schweiz und angrenzende Gebiete 3. Birkhäuser Verlag, Basel
und Stuttgart.
Járai-Komlódi, M., Juhász, M., 1993: Ambrosia elatior L. in Hungary (1989‒1990). Aerobiologia, 9(1): 75-78.

13
Kuntze, O., 1891: Revisio generum plantarum I. Arthur Felix, Leipzig.
Payne, W. W., 1964: A re-evaluation of the genus Ambrosia (Compositae). Jour. Arnold Arb., 45(4): 401-438.
Payne, W. W., 1970: Preliminary reports on the flora of Wisconsin, No. 62. Compositae Family U. I. The genus Ambrosia –
the ragweeds. Wisc. Acad. Sci., Arts, Letters, Trans., 58: 351-371.
Piper, C. V., 1906: Flora of the State Washington. Contr. U. S. Nat. Herb., 11: 551.
Priszter, S., 1985: A magyar flóra és vegetáció rendszertani-növényföldrajzi kézikönyve VII. Akadémiai Kiadó, Budapest.
Räuschel, E. A., 1797: Nomenclator botanicus. Johann Gottlob Feind, Leipzig.
Rydberg, P. A., 1922: Ambrosiaceae in Rydberg, P. A., Gleason, H. A., North American Flora 33(1). The New York Botanical
Garden, New York.
Scheele, A., 1849. Beiträge zur Flora von Texas. Linnaea, 22: 157.
Torrey, J., Gray, A., 1842: A Flora of North America II (2). Wiley & Putnam, New York.
Willdenow, C. L., 1805: Caroli a Linné Species Plantarum IV. G. C. Nauk, Berlin.
Abstrakt
Tento prehľad zahŕňa všetky taxonomické koncepcie týkajúce sa ambrózie palinolistej (Ambrosia
artemisiifolia L.) z územia Severnej Ameriky a čiastočne z Európy. Na vyjadrenie morfologickej varia-
bility jedného druhu boli v minulosti opísané mnohé taxóny s hodnotou druhu, variety alebo formy.
Poznať taxonomické koncepcie znamená porozumieť variabilite druhu. Prehľad môže slúžiť ako po-
môcka pri hodnotení morfologickej variability v Európe.
Michal Hrabovský, Karol Mičieta: Prehľad taxonomických koncepcií invázneho druhu
Ambrosia artemisiifolia L. ako podklad pre hodnotenie variability tohto druhu v Európe

14

15
Acta Botanica Universitatis Comenianae Vol. 48, 2013
VALIDATION OF THE METHOD OF BIOINDICATION ECOGENOTOXICITY
IN SITU CONDITIONS ON EXAMPLE OF THE SPECIES
ROBINIA PSEUDOACACIA L.
Karol Mičieta•, Jozef Dušička, Eva Brutovská, Andrea Sámelová
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany,
Révová 39, 811 02 Bratislava, Slovakia; [email protected]*
Received 31 October 2013; Received in revised form 10 December 2013; Accepted 16 December 2013
Abstract
The goal of our study is presenting the bioindication of environmental mutagenesis and its different levels on the example of a chosen species. The pollen of the species Robinia pseudoacacia L. has been used for indication of environmental xenobiotic exposure, as the species is commonly spread, and fulfils the basic criteria for the use of woody plants in bioindication of environmental mutagenesis. We focused especially on the proximity of Slovnaft – oil refinery in Bratislava, presented as a model locality for methodic standardisation. The results show the value of the species Robinia pseudoacacia L. as a suitable bioindicator of ecotoxicity. To its advantages belongs an easy identification of abortive microspores and therefore a direct proof of genotoxic effect. Our results also hint at a possible adaptative response of woody plants.
Key words: biomonitoring, standardisation, pollen abortivity, environmental pollution
Introduction
The principle of socially acceptable or bearable measure of environmental risks is a basic criterion for the choice of strategic priorities. The primary process of this optimization is the evaluation of existing risks, and elaborating of methods for this evaluation. The evaluation of influences on the environment is an effective and preventive system of its protection. A qualified and validated bioindication and biomonitoring in situ is irreplaceable in this system, and provides specific data about ecotoxic and genotoxic risks and threats to the environment at the particular locality. Higher plants are complex integrators of the interrelated processed in the environment, and fulfil all conditions of bioindication in general (De Serres 1992). Their advantages are before all in the fact, that they have a natural integrating function in dynamic media. They grow directly in the environment or are implanted there as standardized model test systems, are easily identified, give a general or specific answer to environmental changes, can't flee from the environment and can be cultivated there according to the needs, enable to observe the genotoxic effect of one or more risk factors, influence of the residues or their metabolites (Mičieta, Murín 1996, 2007).
In biomonitoring, plants are generally able to significantly characterize: the risk of general state of environmental pollution, quantitative and qualitative processes in different conditions and locations, the status and in great measure also perspectives of biological conditions at the studied locality influenced simultaneously by different risk factors, the presence of xenobiotics as well as the monitoring of their effects and range of influence on the quality and length of ongoing biological processes (Murín, Mičieta 2001).
The pollen grains, pollen mother cells and tetrads of diploid species can be used effectively as basic bioindication material for the detection of ecogenotoxic situation in the environment (Mičieta, Murín 1996). The advantage of this material is its great sensibility, implied by the haploid state and its count, convenient for an adequate statistical analysis of results. The use of microspores and pollen enables an • Corresponding Author: Karol Mičieta; [email protected]

16
effective and parallel detection of damage on all three structural levels of the genetic apparatus of the cell, the influence of short-term and long-term exposure, impact of acute and persistent environmental load as well as adaptation mechanisms, individual and population response in the different kinds of environmental load (Grant 1999, Mičieta, Murín 1996, 1998, 2007, Solenská et al. 2006).
Indication and evaluation of ecogenotoxicity, environmental genotoxic deterioration especially in the in situ conditions, indication and study of the individual, population's, species' or taxon's specific answer to chronical, cyclic or one-time exposure are inevitable in this process.
In the process of methodics creation, their following validation and standardisation, we turned our attention to woody plants (Mičieta, Murín 1996, 1997, Mičieta, Dušička 2000). The cardinal difference between woody and herbaceous plants is their longevity, causing the woody plants to work as bioaccu-mulators of the negative impacts in the environment. Several studies and projects show the importance of woody plants in the bioindication of these impacts (Hool, Cairns 1995, Sparling 1995). Woody plants are used for bioindication of different, and sometimes very specific, influences in these studies. The issue of choosing the bioindication methods, bioindicators and characteristics for indication is very complex and complicated, sometimes even mutually incomparable.
A standardized methodics of genotoxicity indication with the use of woody plants pollen has not been elaborated yet, despite the great attention given to the pollen of woody plants since the beginning of 20th century (Hutchinson 1915). The evaluation of genotoxic effects on the level of chromosomal and genomic mutation is possible, just like in all plants, in meristematic tissues, but the getting sufficient and standard amount of material in situ for cytogenetic analyses is quite problematic in, for example, forest trees (Müller et al. 1997). Saplings from the seeds gathered from the woody plants in locations with different emission load can be used for analysis (Bacon et al. 1992). In relation to this, within the scope of validation and standardisation of the methodics, we present the given problematic on the example of the commonly spread species Robinia pseudoacacia L., which fulfils the basic criteria for the use of woody plants in bioindication of environmental mutagenesis (Mičieta, Murín 1996).
Material and Methods
We used the pollen of Robinia pseudoacacia L. for the indication of environmental mutagenesis. Samples of 10 inflorescences shortly before flowering were taken from 5 individuals from each of the studied localities. A locality in the Záhorská nížina lowland, between the villages Závod and Moravský Ján, unaffected by pollution, with a minimum distance from a traffic line 1 km, has been chosen as a control or reference locality. The minimum number of microspores evaluated from each locality was 10 000. Samples from young leaves were taken in the same time as well, and at the end of the vegetation season also fruits. Micronucleus test on 10 000 cells has been carried out on the somatic cells, and a basic cytogenetic analysis in the set of 300 ana-telophases has been carried out on the primary mer-istems of the seedlings (Mičieta 1987).
The gathering, fixation and preparation of samples and their microscopic evaluation have been performed according to Mičieta and Murín (1996, 1997). The results have been statistically processed according to the critical values of multivariation Dunnet method based on the Poisson distribution (Amphelett, Delow 1984). The evaluation of the individual localities has been carried out by the non-parametric variation analysis (ANOVA), and the comparison of localities between each other and the control by the Wilcoxon signed-rank test. All tests are a part of the available statistical softwares.
Results and Discussion
The long-term following of pollen abortivity in the locations with different exposure has been the subject of standardisation, as shown in Fig. 1. It is interesting to note that the highest indication of ecogenotoxicity and frequency of pollen abortivity is not in the expected localities, like Slovnaft – oil refinery, Ružomberok, but Mlynská dolina Bratislava and Prievidza.
The proximity of petrochemical plant Slovnaft – oil refinery is considered a model locality for the standardisation of methodics. The locality Incinerator Bratislava in particular shows the highest levels of

17
abortivity from the monitored localities in the proximity of the petrochemical plant (Fig. 2. and 3.). We also studied the individual variability at this locality (Tab. 1. and Fig. 4.), evaluated from the same individuals in the years 1999 and 2012. The individual No. 5, showing an average abortivity of 21.4% in 1999 was dead in 2012.
0
2
4
6
8
10
12
14
16
Poll
en a
bort
ivit
y (
%)
Locality
2002
2005
2008
2012
Fig. 1. Frequency of pollen abortivity of Robinia pseudoacacia L. at the chosen localities in Slovakia
0
5
10
15
20
25
30
35
1996 1998 2000 2002 2004 2006 2008 2010
Poll
en a
bort
ivit
y (
%)
Years
Incinerator
Control
Lieskovec
Topoľové
OilrefineryBajdel
Pánskydiel
Fig. 2. Frequency of pollen abortivity of Robinia pseudoacacia L. in the proximity of Slovnaft,
Bratislava
05
1015202530
3540
1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010
Pollen
ab
ort
ivit
y (
%)
Years
Incinerator -
Bratislava
Control
Fig. 3. Frequency of pollen abortivity of Robinia pseudoacacia L. at the locality Incinerator
Bratislava in the years 1995-2010
Years

18
Tab. 1. Individual frequency of Robinia pseudoacacia L. pollen abortivity at the locality Incinerator Bratislava in the
years 1999 and 2012
Individual
No. 1
Individual
No. 2
Individual
No. 3
Individual
No. 4
Individual
No. 5
Year 1999 2012 1999 2012 1999 2012 1999 2012 1999 2012
Pollen Abortivity (%) 9.3 7.8 8.4 7.2 17.1 12.8 17.8 13.4 21.4 Dead
0
5
10
15
20
25
No. 1 No. 2 No. 3 No. 4 No. 5
Po
llen
ab
orti
vit
y (
%)
Individual
1999
2012
Fig. 4. Individual frequency of Robinia pseudoacacia L. pollen abortivity at the locality Incinerator
Bratislava in the years 1999 and 2012
The variability in one individual (No. 5) is also presented in the year 1999. It is shown on 10 inflores-cences, with 1000 evaluated pollen grains from 10 flowers from each inflorescence. The difference of pollen abortivity is evident, ranging from 14.8 to 34.2%. Similarly, the flowers from two inflorescences of the chosen individual have been compared. Again there are considerable differenced in the pollen abortivity on this level. This shows without doubt, and not only in woody plants, the need to precisely follow the given criteria for taking and evaluating samples in a sufficiently big set of data (min. of 5 individuals and 5000 pollen grains).
For the complexity of the study, we also compare the frequency of micronuclei in tetrads, frequency of chromosome aberrations in somatic cells of young tissues, frequency of chromosome aberrations of the seedlings compared to the pollen abortivity at the localities in wider proximity of Slovnaft – oil refinery (Tab. 2.).
The results show the significance of the species Robinia pseudoacacia L. as a suitable bioindicator of ecotoxicity, fulfilling the basic criteria of choice of bioindicators for woody plants (Mičieta, Murín 1996). The adventage of this species is an easy indication of abortive microspores in tetrads, and there-fore confirm the genotoxic cause of abortion. In the same time, the results point at a possible adaptive response in woody plants.
0
5
10
15
20
25
30
35
40
No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No. 8 No. 9 No. 10
Pollen
ab
ort
ivit
y (
%)
Individual No. 5/Sample - inflorescence
Fig. 5. Frequency of pollen abortivity in the inflorescenses of the individual No. 5 – locality
Incinerator 1999
Different types of pollution can have different way of spreading in time and space. The data in our figures and tables show the variability of the response and sensitivity of the indicator to the concentration
Individual No. 5/Sample – inflorescence

19
of the induction factor. This just confirms not only the need to precisely follow the methodics regarding the number of evaluated individuals and microspores, but also the location of the sampling. For the standardisation of the methodics it is necessary to observe the frequency of defects not only in exposed populations, but also at a control locality with no or relatively no pollution.
0
5
10
15
20
25
30
35
40
45
No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No.8
Poll
en a
bort
ivit
y
(%)
InflorescenceNo. 1
InflorescenceNo. 8
Fig. 6. Frequency of pollen abortivity in the individual flowers of inflorescenses No. 1 and 8
of the individual No. 5 – locality Incinerator 1999
Tab. 2. Parallel comparison of genotoxicity in the proximity of Slovnaft - oil refinery in Bratislava – years 1999 and 2012 –
Robinia pseudoacacia L.
Conclusion
As a contribution to validation and standardisation of the method of using dendroflora for indication of environmental genotoxicity with the use of microsporogenesis or pollen of woody plants, we present the problematic on the example of the species Robinia pseudoacacia L.. This species is commonly spread and fulfils the basic criteria for the use of woody plants in bioindication of environmental mutagenesis. Our study deals with different levels of environmental mutagenesis, methods of its indication and their parallel study on the example of the chosen species. It has been confirmed that genotoxicological evaluation of the species Robinia pseudoacacia L. provides important information about the state of the environment at the given locality. The results suggest that the frequency of microspore defects is directly related to the exposure by xenobiotics, and if, then only in a small measure to the local conditions, for example the climatic conditions in different years.
Acknowledgements
This study was supported by VEGA grant No. 1/0380/13. This study was also partially the result of the project implementation: Comenius University in Bratislava Science Park supported by the Research and Development Operational Programme funded by the ERDF Grant number: ITMS 26240220086.
Locality
Micronucleus
(%)
Chromosomal
aberrations in somatic
cells (%)
Chromosomal
aberrations in root
(%)
Pollen abortivity
(%)
1999 2012 1999 2012 1999 2012 1999 2012
Control 0.14 0.15 0.2 0.3 0.2 0.4 4.0±0.3 4.7±0.4
Lieskovec 0.5 0.6 0.4 0.6 0.5 0.7 11.2±0.6 9.0±0.5
Topoľové 0.4 0.5 0.7 0.6 0.6 0.4 9.8±0.6 6.6±0.4
Incinerator 1.5 1.3 1.8 1.5 2.1 1.9 14.8±0.7 10.7±0.7
Oil refinery 0.4 0.6 0.6 0.5 0.5 0.7 11.2±0.6 8.0±0.4
Bajdel 0.2 0.2 0.4 0.3 0.3 0.4 5.4±0.4 5.9±0.5
Pánsky diel 0.2 0.3 0.3 0.2 0.3 0.3 5.8±0.5 5.2±0.5
Flower

20
References
Amphelett, G. E., Delow, G. F., 1984: Statistical analysis of the micronucleus test. Mutat. Res., 128 (2): 161-166. Bacon, C. E., Jarman, W. M., Costa, D. P., 1992: Organochlorine and polychlorinated biphenyl levels in pinniped milk
from the Arctic, the Antarctic, California and Australia. Chemosphere, 24: 779-791. De Serres, F. J., 1992: Higher plants as effective monitors of environmental mutagens. Mutat. Res., 270: 1-6. Grant, W. F., 1999: Higher plants assays for the detection of chromosomal aberrations and gene mutations – a brief historical
background on their use for scrreening environmental chemicals. Mutat. Res., 426 (2): 107-112. Hool, K. D., Cairns, J., 1995: Landscape indicators in ecotoxikology. In: Hoffman, D. J., Rattner, A. B., Burton, G. A., Cairns, J.,
Handbook of ecotoxikology, p. 185-197, Lewis Publishers, London-Tokyo. Hutchinson, A. H., 1915: Fertilization in Abies balsamea. Bot. Gaz., 60: 457-472. Mičieta, K., 1987: Increased aberration capacity of chromosomes in Vicia faba L. under the influence of a combination of
some herbicides after treatment in field cultures. Agriculture (Nitra), 33: 1094-1100. Mičieta, K., Dušička, J., 2000: Význam arborét v indikácii a evaluácii genotoxickej deteriorizácie genofondu drevín. In:
Anonymus, Arboréta premenlivosť a introdukcia drevín, p. 87-91, Lesnícky výskumný ústav, Zvolen. Mičieta, K., Murín, G., 1996: Microspore analysis for genotoxicity of polluted environment. Environ. Exp. Bot., 36: 21-27. Mičieta, K., Murín, G., 1997: Wild plant species in practical use for bioindication of polluted environment. Ekológia, 16:
193-202. Mičieta, K., Murín, G., 2007: Wild plant species in bio-indication of radioactive-contaminated sites around Jaslovské Bohu-
nice nuclear power plant in the Slovak Republic. J. Environ. Radioact., 93: 26-37. Müller, L. I., Warren, R. J., Evans, D. L., 1997: Theory and practice of immunocontraception in wild mammals. Wildl.
Soc. Bull., 25 (2): 504-514. Murín, G., Mičieta, K., 2001: Wild plant species in bioindication of polluted environment. In: Yunus, M., Sing, N., de Kok,
L. J. (eds.), Environmental stress: Indication, mitigation, and eco-conservation, p. 285-297, Kluwer, Dordrecht. Solenská, M., Mičieta, K., Mišík, M., 2006: Plant bioassays for an in situ monitoring of air near an industrial area a municipal
solid waste –Žilina (Slovakia). Environ. Monit. Assess., 115: 499-508. Sparling, D. W., 1995: Acidic deposition: A review of biological effects. In: Hoffman, D. J., Rattner, A. B., Burton, G. A.,
Cairns, J., Handbook of ecotoxikology. Lewis Publishers, p. 301-329, London-Tokyo.
Abstrakt
Cieľom našej štúdie bolo priblížiť bioindikáciu environmentálnej mutagenézy a jej rôzne úrovne na nami vybranom druhu. Na indikáciu environmentálnej xenobiotickej expozície sme použili peľ druhu Robinia pseudoacacia L., ktorý je bežne rozšírený a spĺňa základné kritériá využitia drevín v bioindikácii environmentálnej mutagenézy. Zamerali sme sa predovšetkým na okolie Slovnaftu Bratislava, ktoré prezentujeme ako modelové územie pre štandardizáciu uvedenej metodiky. Zistené výsledky nám poukazujú na význam druhu Robinia pseudoacacia L. ako vhodného bioindikátora ekotoxicity. Výhodou je ľahká indikácia abortívnych mikrospór a tým priamy dôkaz genotoxického účinku. Naše výsledky súčasne upozorňujú aj na možnú adaptačnú odpoveď pri drevinách.
Karol Mičieta, Jozef Dušička, Eva Brutovská, Andrea Sámelová: Validácia metódy bioindikácie
ekogenotoxicity v podmienkach in situ na príklade druhu Robinia pseudoacacia L.

21
Acta Botanica Universitatis Comenianae Vol. 48, 2013
EVALUATION OF ECOGENOTOXIC DETERIORATION OF THE ENVIRONMENT NEAR THE CITY ŽIAR NAD HRONOM
Andrea Pogányová●, Erik Kerekeš, Karol Mičieta, Jozef Dušička
Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany,
Révová 39, 811 02 Bratislava, Slovakia
Received 7 November 2013; Received in revised form 25 November 2013; Accepted 13 December 2013
Abstract
The aim of the study was to verify the usability of phytoindication Tradescantia MCN test modified for in situ conditions, and to evaluate the environmental situation near the aluminum factory in the Žiar nad Hronom in central Slovakia. The presented results confirm the improving quality of the environment around aluminum factory. We have detected the frequency of micronuclei, which reflects the degree of ecogenotoxical deterioration. We have also tried to prove the existence of the dilution effect, but it did not have a significant influence on our results.
Key words: genotoxicity, in situ, micronucleus test, pollutants
Introduction
The living environment in areas with industrial production is intensively exposed to harmful substances and air pollutants produced as a by-product of manufacturing processes. Across the environment, these substances cause structural formation and changes in DNA. Chromosomal aberrations and mutations are a reaction of sensitive vegetable organisms to the presence of pollutants and chemical mutagens in nature (Grant 1978). Increased pollution in the area affected the frequency of mutation and organisms’ fertility. The release of genotoxical substances may have a side effect as a decrease in the size of species populations and affect the overall sustainability of the ecosystem (Uhl et al. 2003). Evaluations of the effects of xenobiotic in environments can be carried out by the multiple plant tests. One of these tests is the Tradescantia micronucleus (MCN) test, which is the recommended as standardized test system (Ma 1982, Ma et al. 1984, 1994, Grant 1999, Mišík et al. 2007). This test has high sensitivity and allows the testing of pollutants in a gaseous, liquid and solid state; on monitored area and in laboratory conditions (Ma et al. 1994). The Tradescantia MCN test evaluates the clastogenic effect of xenobiotic substances on the chromosomes of pollen mother cells at the stage of meiosis. The statistical evaluation of changes in frequency of the presence of micronuclei in tetrads is useable for testing of ecotoxicity and the muta-genicity of the environment (Ma et al. 1996). Validated in situ bioindication provides specific information about the eco-toxic and genotoxic risk in a studied area (Mičieta, Murín 1996). Accordingly, the use of plants in bioindication is irreplaceable in the assessment and minimizing of environmental risk (Mičieta, Murín 1996, 1997).
In our study, we have used this methodology to figure out a changes in the environmental situation on the monitored area near the industrial area of aluminum factory in Žiar nad Hronom. We have implanted Tradescantia plants to the monitored area during the years 2008–2013 and evaluated collected material. We were looking for correlation of relationships between micronucleus frequency and pollution as well as precipitation. We have compared our results with the other studies and these conclusions are presented in this study.
● Corresponding Author: Andrea Pogányová; [email protected]

22
Material and methods
The study was conducted with the Tradescantia MCN test (Ma 1982, Ma et al. 1994). The preculti-vated plants of Tradescantia paludosa clone 03 were exposed in the area of our research near the aluminum factory in the Žiar nad Hronom. Two flower pots with ten precultivated plants were transported to the monitored area and exposed from June to September 2008–2013. Subsequently, the cells of pollen and the root apex were collected and evaluated. Samples of inflorescences and root tips were collected four times per year (in June, July, August and September in the years 2008–2012 and July, August, September and October in the year 2013). The samples were fixed in a 1:3 acetic acid to 70% ethanol solution for 24 h and preserved in 70% ethanol, according to the protocol by Ma et al. (1994). Young anthers and root apexes were crushed in a solution of acetocarmine and analyzed under an optical microscope (Nicon YS2 Alphaphot) at 400 × magnification. The presence of micronuclei was assessed in 300 tetrads from each sample. The frequency of the incidence of so-called micronuclei after the fixation and staining of the cells was statistically evaluated. The term micronucleus is used for chromatin structures located in the cytoplasm and their own membrane without connection to the nucleus (Schiffmann, De Boni 1991). The test system is based on the detection of micronuclei which refer to clastogenic (chromosome breaking) effect in meiotic pollen tetrad cells (Mišík et al. 2013).
The data of satellite mapping were obtained by the Google Earth application and data of total annual precipitation from database of the Slovak Hydrometeorological Institute. Data of annual quantities of emitted air pollutants were obtained from a database of the National Emission and Information System of Slovak Republic − NEIS (http://www.air.sk/emissions.php).
Monitored area
Horné Opatovce is the locality of the defunct village near the industrial area of the aluminum factory. The village was situated to the south of Žiar nad Hronom city, and there was a significant deterioration of the environment after start of aluminum production. The village was evacuated in 1969 after serious contamination by the emission of the arsenic, mercury, hydrogen fluoride and other harmful substances. Monitored area is located in the former village Horné Opatovce, about 900 metres to the south of the aluminum factory. Geographic coordinates read from the centre of the monitored area are: 48° 33' 22.68" N and 18° 51' 14.24" E. The control area for the Tradescantia MCN test was in the Department of Botany of Comenius University in Bratislava, Révová 39. Geographic coordinates of control area are: 48° 8' 45.29" N, 17° 4' 43.04" E.
Fig. 1. The map of the monitored area next to the aluminum factory in the Žiar nad Hronom (Google Earth application)
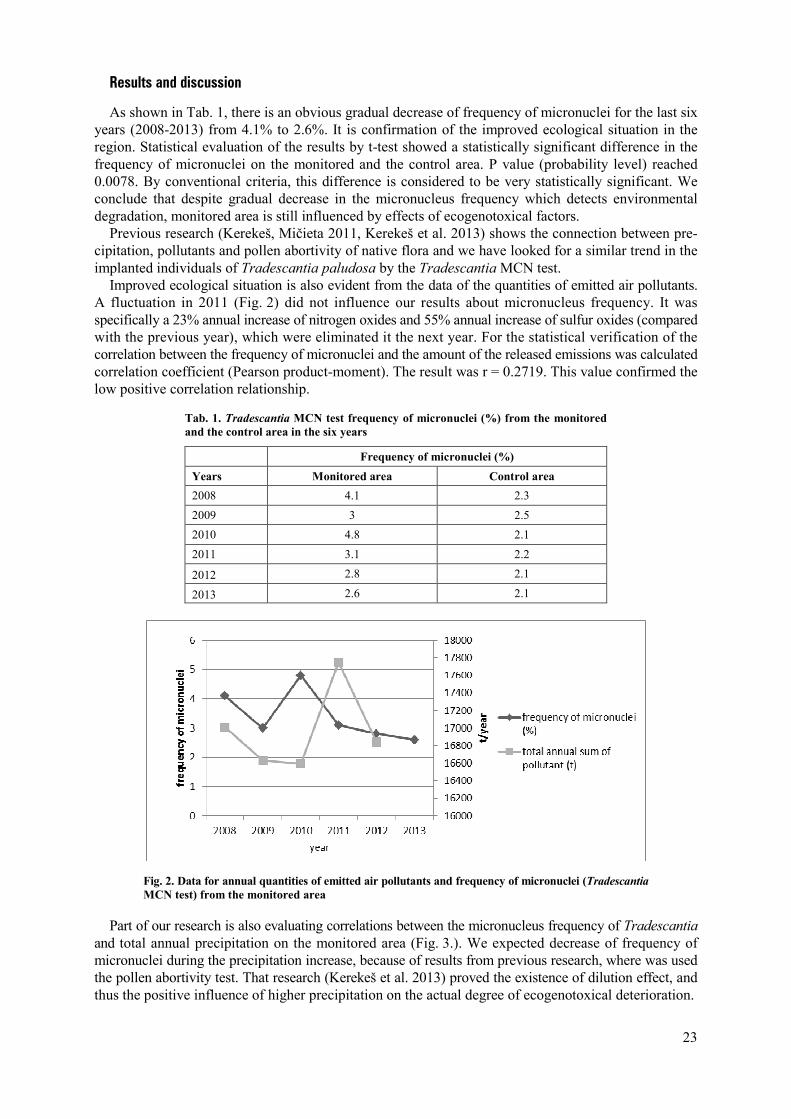
23
Results and discussion
As shown in Tab. 1, there is an obvious gradual decrease of frequency of micronuclei for the last six years (2008-2013) from 4.1% to 2.6%. It is confirmation of the improved ecological situation in the region. Statistical evaluation of the results by t-test showed a statistically significant difference in the frequency of micronuclei on the monitored and the control area. P value (probability level) reached 0.0078. By conventional criteria, this difference is considered to be very statistically significant. We conclude that despite gradual decrease in the micronucleus frequency which detects environmental degradation, monitored area is still influenced by effects of ecogenotoxical factors.
Previous research (Kerekeš, Mičieta 2011, Kerekeš et al. 2013) shows the connection between pre-cipitation, pollutants and pollen abortivity of native flora and we have looked for a similar trend in the implanted individuals of Tradescantia paludosa by the Tradescantia MCN test.
Improved ecological situation is also evident from the data of the quantities of emitted air pollutants. A fluctuation in 2011 (Fig. 2) did not influence our results about micronucleus frequency. It was specifically a 23% annual increase of nitrogen oxides and 55% annual increase of sulfur oxides (compared with the previous year), which were eliminated it the next year. For the statistical verification of the correlation between the frequency of micronuclei and the amount of the released emissions was calculated correlation coefficient (Pearson product-moment). The result was r = 0.2719. This value confirmed the low positive correlation relationship.
Tab. 1. Tradescantia MCN test frequency of micronuclei (%) from the monitored and the control area in the six years
Frequency of micronuclei (%)
Years Monitored area Control area
2008 4.1 2.3
2009 3 2.5
2010 4.8 2.1
2011 3.1 2.2
2012 2.8 2.1
2013 2.6 2.1
Fig. 2. Data for annual quantities of emitted air pollutants and frequency of micronuclei (Tradescantia MCN test) from the monitored area
Part of our research is also evaluating correlations between the micronucleus frequency of Tradescantia
and total annual precipitation on the monitored area (Fig. 3.). We expected decrease of frequency of micronuclei during the precipitation increase, because of results from previous research, where was used the pollen abortivity test. That research (Kerekeš et al. 2013) proved the existence of dilution effect, and thus the positive influence of higher precipitation on the actual degree of ecogenotoxical deterioration.

24
However, neither the sharp increase of total annual precipitation in the year 2010 (it was the rainiest year in the last hundred years) did not cause micronucleus frequency in our research.
Fig. 3. Data for total annual precipitation and frequency of micronuclei (Tradescantia MCN test) from the monitored area
The whole correlation coefficient of relationship between micronucleus frequency and total annual
precipitation was r = 0.7869. This shows the highly positive correlation relationship between frequency of micronuclei and amount of precipitations.
The results of our research did not show impact of dilution effect in case of use analysis of Tradescantia MCN test.
Given these observations we recommend next research by using this methodology.
Acknowledgments
This study was supported by the grant of Comenius University UK/147/2013.
References
Grant, W. F., 1978: Chromosome aberrations in plants as a monitoring system. Environ. Health. Persp., 27: 37-43. Grant, W. F., 1999: Higher plants assay for the detection of chromosomal aberrations and gene mutations – a brief historical
background on their use for screening and monitoring environmental chemicals. Mutat. Res., 429: 107-112. Kerekeš, E., Mičieta, K., 2011: Fytoindikácia ekogenotoxicity v dlhodobom monitoringu kvality životného prostredia na
základe analýz abortivity peľových zŕn v Horných Opatovciach pri Žiari nad Hronom. In: Galamboš, M., Džugasová ,V. (eds.), Študentská vedecká konferencia PriF UK 2011 – Zborník recenzovaných príspevkov, p. 369-374, Vydavateľstvo UK, Bratislava.
Kerekeš, E., Pogányová, A., Miškovic, J., Mičieta, K., 2013: Test abortivity peľu natívnej flóry a Tradescantia MCN test v dlhodobej fytoindikácii zmien ekogenotoxickej záťaže v Horných Opatovciach. In: Galamboš, M., Džugasová, V., Ševčovičová, A. (eds.), Študentská vedecká konferencia PriF UK 2013 – Zborník recenzovaných príspevkov, p. 334-338, Vydavateľstvo UK, Bratislava.
Ma, T. H., 1982: Tradescantia cytogenetic tests (root-tip mitosis, pollen mitosis, pollen mother-cells meiosis). A report of the U.S. Environmental Protection Agency Gene-Tox Program. Mutat. Res., 99: 293-302.
Ma, T. H., Cabera, G. L., Chen, R., Gill, B. S., Sandhu, S. S., Valenberg, A. L., Salamone, M. F., 1994: Tradescantia micronucleus bioassay. Mutat. Res., 310: 221-230.
Ma, T. H., Harris, M. M., Anderson, V. A., Ahmed I., Mohammad, K., Bare J. L., Lin G., 1984: Tradescantia-micronu-cleus (Trad-MCN) tests on 140 health-related agents. Mutat. Res., 138: 157-167.
Ma, T. H., Xu, C., Liao, H., McConnell, H., Jeong, B. S., Won, C. D., 1996: In situ monitoring with the Tradescantia bioassays on the genotoxicity of gaseous emissions from a closed landfill site and an incinerator. Mutat. Res., 359: 39-52.
Mičieta, K., Murín, G., 1996: Microspore analysis for genotoxicity of polluted environment. Environ. Exp. Bot., 36: 21-27. Mičieta, K., Murín, G., 1997: Wild plant species in practical use for bioindication polluted environment. Ekológia (Bratislava),
16: 193-202. Mišík, M., Mičieta, K., Solenská, M., Mišíková, K., Pisarčíková, H., Knasmüller, S., 2007: In situ biomonitoring of the
genotoxic effects of mixed industrial emissions using the Tradescantia micronucleus and pollen abortion tests with wild life plants: Demonstration of the efficacy of emission controls in an Eastern Europen city. Environ. Pollut., 145: 459-466.

25
Mišík, M., Pichler, C., Rainer, B., Nersesyan, A., Knasmueller, S., 2013: Micronucleus assay with tetrad cells of Tradescantia. Genotoxicity Assessment Methods in Molecular Biology, 1044: 405-415.
Schiffmann, D., De Boni, U., 1991: Dislocation of chromatin elements in prophase induced by diethylstilbestrol: a novel mechanism by which micronuclei can arise. Mutat. Res., 246: 113-122.
Uhl, M., Plewa, M. J., Majer B. J., Knasmüller, S., 2003: Basic principles of genetic toxicology with an emphasis on plant bioassays. In: Maluszynska, J., Plewa, M. J. (eds.), Bioassays in plant cells for improvement of ecosystem and human health, p. 123-130, Wydawnictvo Uniwersytetu Ślaskiego, Katowice.
Abstrakt
Cieľom práce bolo overiť použiteľnosť fytoindikačnej metódy Tradescantia MCN test modifikovanej na podmienky in situ a zhodnotiť environmentálnu situáciu v okolí hlinikárne v Žiari nad Hronom na strednom Slovensku. Prezentované výsledky potvrdzujú zlepšujúcu sa kvalitu životného prostredia v okolí hlinikárne. Zisťovali sme frekvenciu výskytu mikrojadier, ktorá odráža stupeň ekogenotoxickej záťaže prostredia. Rovnako sme sa pokúsili dokázať existenciu zrieďovacieho efektu, ten však nemal signifikantný vplyv na naše výsledky. Andrea Pogányová, Erik Kerekeš, Karol Mičieta, Jozef Dušička: Evalvácia ekogenotoxického zaťaženia životného prostredia v okolí mesta Žiar nad Hronom

26

27
Acta Botanica Universitatis Comenianae Vol. 48, 2013
HOW TO HANDLE WITH PUBLISHED NAMES OF THE GENUS MELANOLEUCA –
A CASE STUDY FROM SLOVAKIA
Ondrej Ďuriška1•, Michal Tomšovský2, Soňa Jančovičová3
1,3Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany,
Révová 39, 811 02 Bratislava, Slovakia 2Mendel University in Brno, Faculty of Forestry and Wood Technology,
Zemědělská 3, 613 00 Brno, Czech Republic
Received 15 November 2013; Received in revised form 22 November 2013; Accepted 2 December 2013
Abstract
Data on Melanoleuca species published from Slovakia were excerpted from the selected literature. Consequently, the detected specimens referred in the literature were re-identified using the different determination keys. The ambiguous identifications lead the authors to prepare a consistent approach how to handle with published Melanoleuca names, as well how the identify Melanoleuca collections properly.
Key words: fungi, Tricholomataceae, literature, specimens, morphology
Introduction
Melanoleuca Pat. is a well delimited genus of basidiomycetes (Agaricales, Tricholomataceae). Basidio-mata are characteristic in having central stipe and collybioid to tricholomatoid habit; colouring of cap and stipe is indistinctive (mostly in shades of brown, white and grey), as well as of lamellae (mostly white to grey or brownish); lamellae are emarginate to narrowly or broadly adnate, rarely short decurrent. The combination with ellipsoid hyaline spores covered by amyloid warts, absence of clamps, specific types of cheilocystidia and pleurocystidia, and pileipellis of a cutis, an ixocutis or trichoderm made Melanoleuca hardly mistakable for other genera.
The species identification is, however, problematic, because the macro- and micromorphological characters of the Melanoleuca representatives are variable and weakly specific. In the generally accepted determination keys (Bon 1991, Boekhout 1999, Fontenla, Para 2012, Gröger (2006), Moser 1978, Vesterholt 2012), such characters are, moreover, disproportionally interpreted.
As shown in the presentation by Ďuriška et al. (2013), the identification of the Melanoleuca collections (own material) using the different keys could lead to the different names. Accordingly, there are some questions: Which of those names is correct? How to handle with the published Melanoleuca names? Are the morphological characters only enough to Melanoleuca species separation?
In this case study: 1. we have made a search for published data on Melanoleuca species from Slo-vakia, 2. we have verified the existence of the voucher specimens referred in the publications, and 3. we have re-identified the detected specimens using the different determination keys. The results of our study, as well the answers on the above mentioned questions are presented in this paper.
Material and Methods
The case study on the Melanoleuca species/names from Slovakia is based on the literature excerption and the re-identification of the specimens.
• Corresponding Author: Ondrej Ďuriška; [email protected]

28
1. Only such literature was excerpted which was primarily used to create two comprehensive works on Slovak fungi: Fungi (Bacigálová, Lizoň 1998, 1999) and Mycoflora Slovaca (Škubla 2003). 2. The existence of voucher specimens, which were referred in the excerpted literature, was verified it the herbaria BRA and SLO. For the abbreviations of the herbaria see Tab. 1. 3. The detected specimens were re-identified using the generally accepted determination keys by Bon (1991), Fontenla, Para (2012), Gröger (2006), Moser (1978), and Vesterholt (2012). 4. The macromorphological characters – observed in dried specimens by naked eyes and/or the magnifying glass only – were valued and described subjectively. The micromorphological characters were observed under the Leica DM 1000 and Olympus BX41 light microscopes with oil immersion lens and a magnification of 1000×. The microscopic objects were mounted in a 3% KOH aqueous solution, Melzer's reagent and Congo red. Statistic for spores is based on 20 measurements and is given as minimum, maximum and average (av.) values. Q = ratio of length and width of spores.
Results and Discussion
For our study, we have excerpted 31 publications: Blanár, Mihál (2002), Dermek (1971, 1978), Dermek, Pilát (1974), Fábry, Michalko (1975), Fábry et al. (1975), Hagara (1986, 1987a, 1987b, 1989, 1992, 1993), Janitor (1997), Janitor, Fábry (1981), Janitor, Gáper (1998), Kautmanová (1996), Kubička (1957, 1976), Kukulka (1992), Kult (1989, 1991), Kuthan (1989), Kuthan et al. (1999), Lizoň (1977a, 1977b), Mihál, Blanár (1999), Princ (1992), Škubla (1996), Škubla, Milan (1995), Záhorovská (1997), Záhorovská, Jančovičová (1997) (Tab. 1).
In these publications, data on 24 Melanoleuca species were presented: M. amica, M. arcuata, M. brevipes, M. cnista, M. cognata, M. decembris, M. evenosa, M. furva, M. grammopodia, M. humilis, M. intervenosa, M. kavinae, M. luscina, M. melaleuca, M. microcephala, M. paedida, M. polioleuca, M. spegazzini, M. strictipes, M. stridula, M. subalpina, M. tristis, M. verrucipes, and M. vulgaris (Tab. 1).
Tab. 1. Data on Melanoleuca species published from Slovakia
I. Species/Publication: the publication in which the Melanoleuca species collected in Slovakia is mentioned. II. Data: the data on the species excerpted from the publication and presented in their ±original form (only formally
updated); the phytogeographical units of Slovakia were added if missing and unified according to Futák et al. (1966). III. Herbarium: the herbarium in which the specimen is deposited according to the publication: BRA – the herbarium of
the Slovak National Museum-Natural History Museum, Bratislava; CB – the South Bohemian Museum, České Budějo-vice; Der – the private herbarium of A. Dermek; Fab – the private herbarium of I. Fábry; Her – the private herbarium of J. Herink; Kul – the private herbarium of K. Kult; Kut – the private herbarium of J. Kuthan; Not. – the find was noted only (not backed up by specimen); PRM – the herbarium of National Museum, Mycological Department, Praha; SLO – the herbarium of the Comenius University in Bratislava, Faculty of Natural Sciences, Department of Botany; × – it is not mentioned where/whether the specimen is deposited; ? – it is not cleared where/whether the specimen is deposited.
IV. Verification: our verification whether the specimen is deposited in the referred herbarium: + – specimen is deposited in the herbarium; × – specimen is not deposited in the herbarium; ? – we have not verified the presence of the specimen in the herbarium.
I II III IV Melanoleuca amica Hagara (1987b) Veľká Fatra, kataster obce Turčianske Jaseno, vrch Hradište, 1,6 km JV
od Turč. Jasena, 540 m n. m., 24. X. 1982, leg. L. Hagara BRA? ×
M. arcuata Kautmanová (1996) Vysoké Tatry, Tatranské podhorie, 1 – 2 km V od Kežmarské žľaby,
Mokriny, podmáčaná smrečina s brezou a rašelinníkom, 760 – 860 m n. m., 15. IX. 1995
×
Nízke Tatry, 2 km JZ (Múry) až 1 km J (Krieslo) od Važca, smrek, buk, borovica na vápencovom podloží, 900 m n. m., 16. IX. 1995
×
Kuthan et al. (1999) Bukovské vrchy, Nová Sedlica, nivné lúky Zbojského potoka (SZ od obce N. Sedlica), na lúke v blízkosti Alnus, 16. IX. 1995, leg. S. Adamčík
BRA +
M. brevipes Dermek (1971) Podunajská nížina, Bratislava, v parku sídliska na Miletičovej ulici,
22. V. 1970, leg. J. Ďurčová Der ?

29
Tab. 1. – Continuation I II III IV Dermek (1978) Záhorská nížina, územie medzi obcami Brodské, Čáry, Gbely, Kopčany,
Kúty a Smolinské, dubové a borovicové lesy (Quercus robur, Pinus
sylvestris), 155 – 185 m n. m., 1963 – 1977, leg. A. Dermek
BRA ×
Janitor (1997) Malé Karpaty, záhrady nad Búdkovou cestou i Drotárskej cesty na Machnáči, 21. VII. 1979, leg. A. Janitor
×
Janitor, Fábry (1981) Malá Fatra, v Štefanovej, smerom na Podžiar, na lúke za posledným domom, 23. IX. 1972
Fab ?
Kautmanová (1996) Nízke Tatry, 2 km JZ (Múry) až 1 km J (Krieslo) od Važca, smrek, buk, borovica na vápencovom podloží, 900 m n. m., 16. IX. 1995
×
Kuthan (1989) Nízke Tatry, Važec (okres Liptovský Mikuláš), vzácne, severne od obce, pastviny a les v katastr. území obce podél potoků Mlyničná a Solisková voda, 8. IX. – 18. IX. 1988, leg. L. Hruška, K. Hlaček, E. Skála, J. Šutara
BRA? ×
Kuthan et al. (1999) Bukovské vrchy, údolie Uličky, lužný les Uličky nad Kolbasovom, na zemi pod Alnus a Salix, 25. IV. 1990, leg. J. Terray
BRA +
Záhorovská (1997) Devínska Kobyla, Sitiny, listnatý les, 1985, leg. J. Aghová SLO × M. cnista Fábry, Michalko (1975) Záhorská nížina, lúky medzi Zohorom a Plaveckým Štvrtkom × M. cognata Blanár, Mihál (2002) Slovenské rudohorie, Revúca, bučina J od mesta s miestnym názvom „Za
Peklom“, Q 7386 B, 310 – 370 m n. m., exp. V, SZ až Z, 3 – 5 ha, svorová rula, 27. VII. 1999
?
Ibidem, 20. X. 2001 ? Hagara (1987a, 1993) Veľká Fatra, Turčianske Jaseno, okres Martin, okraj smrečiny na vrchu
Hradište, 570 m n. m., 13. X. 1983 ×
Janitor (1997) Malé Karpaty, trávnatý porast, ovocné stromy, Machnáč, 12. VII. 1979, leg. A. Janitor
×
Mihál, Blanár (1999) Muránska planina, Muráň, Nižná Kľaková, Q 7285, smrekové lesy (a ich okraje) s výskytom napr. Adenostyles alliariae, Aconitum
variegatum, Coeloglossum viride, Delphinium elatum, Doronicum
austriacum, Listera ovata, Stachys alpina, 1200 – 1300 m n. m., exp. J až JV, 5 – 10 ha, dolomity, gutensteinsko-annaberské vápence, 31. X. 1999, leg. D. Blanár
?
Kult (1991) Slanské vrchy, podcelok Šimonka, vrch Dubová hora (763 m n. m.), SPR, k. ú. Podhradík, Q 6994c, 0 – 8 (– 12) km V – SV od obce, 27. IX. 1990, číslo položky: 845/90
Her ?
Ibidem, číslo položky: 854/90 Her ? Slanské vrchy, podcelok Šimonka, část Olšavské Predhorie, vrch Vrátna (626 m n. m.), sedlo Vrátna (525 m n. m.), k. ú. Žehňa, Q 7094d, 6 – 9 km JVV od obce, JZ od hlavnej cesty Žehňa – Tuhrina, 510 – 550 m n. m., lesná cesta – Picea abies, 22. – 28. IX. 1990
Kul? Her?
? ?
Kuthan et al. (1999) Bukovské vrchy, Stužica, Sedielko nad Kýčerou (V svah), na zemi pod Picea, 22. VI. 1990, leg. J. Terray
Not.
Bukovské vrchy, Nová Sedlica, Patrikúsky (okolie skladu dreva), na zemi pod Picea, 25. IV. 1990, leg. J. Terray
BRA ×
M. decembris Fábry, Michalko (1975) Záhorská nížina, lúky medzi Zohorom a Plaveckým Štvrtkom × M. evenosa Kubička (1957) Belianske Tatry, na svahu Bujačieho vrchu do Holubyho kotliny vo
Festucetum carpaticae, 1850 m n. m., 7. VIII. 1956 ×
Belianske Tatry, na svahu Bujačieho vrchu k Rakúskemu chrbátu, v Sesleriete Tatrae, 1905 m n. m., 7. VIII. 1956
×
Kubička (1976) Vysoké Tatry, Predné Jatky, 22. VII. 1960 × M. furva Hagara (1987b) Veľká Fatra, kataster obce Turčianske Jaseno, chotárna časť Biely potok, 2,7
km VJV od Turč. Jasena, 720 m n. m., 22.X.1983, leg. L. Hagara BRA? +
M. graminicola Kult (1991) Slanské vrchy, podcelok Šimonka, část Zlatobanská kotlina, Pusté pole,
Z od silnice, 22. – 28. IX. 1990, leg. E. Skála ×
Kuthan (1989) Liptovská kotlina, Liptovský Hrádok – Borová Sihoť (okr. Liptovský Mikuláš), úpatí, východní a jihovýchodní svah vrchu Kameničná, severní svah vrchu Vislavce, početně, v trávě, pod Pinus sylvestris a Picea abies, 3. IX. – 9. IX. 1988
BRA? +

30
Tab. 1. – Continuation I II III IV M. grammopodia Blanár, Mihál (2002) Slovenské rudohorie, Revúca, zmiešané listnaté lesy s miestnym názvom
„Za Laca“, na svahoch pozdĺž Muránky SV od kóty Pavlusove bane (627,2 m), Q 7386 B, 310 – 340 m n. m., exp. SV – SSV, 1 – 2 ha, granodiorit, 25. XI. 2000
?
Kult (1989) Slovenské rudohorie, okolie obce Henclová, osada Tichá voda, 5 km J: Tichovodská dolina, potok Tichá voda, v tráve, 2. X. 1986
Kul ?
Slovenský raj, Smižany – Čingov, S úpätie vrchu Čingov, 1. X. 1986 Her ? Slovenský raj, v doline Hornádu, 1. X. 1986 Her ? Slovenský raj, Smižany, dolina potoka Lesnica, 1. X. 1986 Her ? Spišské kotliny, Popradská kotlina, 2,5 km V od Tatranskej Štrby, pri hlavnej ceste do Svitu, kataster obce Mengusovce, 1 – 2 km V od hlavnej lesnej cesty, menšie rašeliniská („Rik“), v tráve, 3. X. 1986
Kul? Kut? BRA?
? ? ×
Kult (1991) Slanské vrchy, podcelok Šimonka, vrch Dubová hora (763 m n. m.), SPR, k. ú. Podhradík, Q 6994c, 0 – 8 (– 12) km V – SV od obce, v tráve, 27. IX. 1990, leg. J. Lederer?
Kul?
?
Slanské vrchy, podcelok Šimonka, polovn. chata „Koloroš“ (na mapě jen „Polov. Chata“, k. ú. Zlatá Baňa, Q 7194b, SZ svah Čierhory (1073 m n. m.), pramenisko potoka M. Delňa, cca 920 – 950 m n. m., 8 – 10 km S od Zl. Bane, v tráv., 22. – 28. IX. 1990, leg. J. Valter
CB ?
Slanské vrchy, podcelok Šimonka, část Zlatobanská kotlina, vrch Dubník (877 m n. m.), SZ a Z svah, k. ú. Zl. Baňa, cca 730 – 780 m n. m., 22. – 28. IX. 1990, leg. K. Kult
Kul? ?
Slanské vrchy, podcelok Šimonka, vrch Hradová (s 2 vrcholy: 622 * a 586 m n. m. – na mapě není uv.), k. ú. Kokošovce, Q 7094c, V svah nad hotelem Sigord a Šťavica, stinný les s dominant. Quercus sp. a Fagus
sylvatica, skeletová půda na andesitu, 500 – 800 m n. m., leg. E. Skála
×
Slanské vrchy, podcelok Šimonka, část Olšavské Predhorie, vrch Vrátna (626 m n. m.), sedlo Vrátna (525 m n. m.), k. ú. Žehňa, Q 7094d, 6 – 9 km JVV od obce, JZ od hlavnej cesty Žehňa – Tuhrina, 510 – 550 m n. m., v tráv., 22. – 28. IX. 1990
Kul? Her?
? ?
Kuthan (1989) Nízke Tatry, oblast u obce Važec (kat. území Važec, okr. Liptovský Mikuláš), jižně od obce; západní část pásma Kozie chrbty, a to východněji položená oblast Vŕšky na SZ obvodu vrchu Slamená a západněji položená oblast Múry na SV svahu vrchu Hošková, pastvina s Picea abies, 3. IX. – 9. IX. 1988
BRA? ×
Kuthan et al. (1999) Bukovské vrchy, Pľaša, NPR Pľaša, na zemi v tráve, 28. VIII. 1991, leg. J. Terray
BRA +
Lizoň (1977b) Malé Karpaty, lesy na svahoch vrchu Raková (hrab, buk, jelša, smrek), Jánova dolina pri Píle, 250 – 300 m n. m., 22. IX. 1977
×
M. humilis Kult (1989) Slovenské rudohorie, okolie obce Henclová, osada Tichá voda, 2,5 km JZ:
dolina potoka Henclová, 2. X. 1986 Kul ?
Kuthan (1989) Liptovská kotlina, Liptovský Hrádok – Borová Sihoť (okr. Liptovský Mikuláš), úpatí, východní a jihovýchodní svah vrchu Kameničná, severní svah vrchu Vislavce, vzácně, v trávě, 3. IX. – 9. IX. 1988
BRA? ×
Nízke Tatry, oblast u obce Važec (kat. území Važec, okr. Liptovský Mikuláš), jižně od obce; západní část pásma Kozie chrbty, a to východněji položená oblast Vŕšky na SZ obvodu vrchu Slamená a západněji položená oblast Múry na SV svahu vrchu Hošková; vzácně, v trávě, 3. IX. – 9. IX. 1988
Her ?
M. intervenosa Fábry, Michalko (1975) Záhorská nížina, lúky medzi Zohorom a Plaveckým Štvrtkom × M. kavinae
Hagara (1986) Malá Fatra, horský pasienok na vrchu Hrádok, 2,3 km ZSZ od obce Bystrička, okr. Martin, 670 m n. m., na vápencovom podklade, 21.V.1983, leg. L. Hagara
BRA +
Záhorovská, Jančovičová (1997)
Malé Karpaty, Sološnica, lokalita Borníky, tráva (okraj lesa), 187 m n. m., 1994 – 1995, leg. S. Jančovičová
SLO ×
M. luscina Dermek (1978) Záhorská nížina, územie medzi obcami Brodské, Čáry, Gbely, Kopčany,
Kúty a Smolinské, dubové a borovicové lesy (Quercus robur, Pinus
sylvestris), 155 – 185 m n. m., 1963 – 1977, leg. A. Dermek
BRA ×
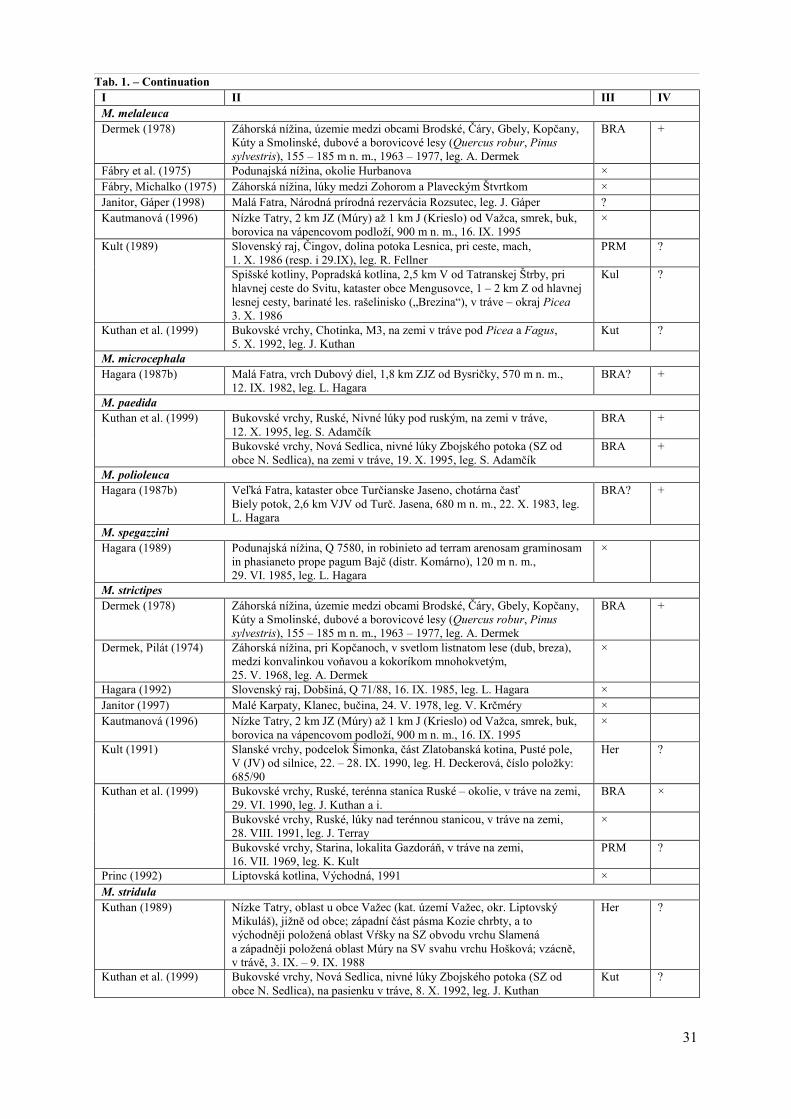
31
Tab. 1. – Continuation I II III IV M. melaleuca Dermek (1978) Záhorská nížina, územie medzi obcami Brodské, Čáry, Gbely, Kopčany,
Kúty a Smolinské, dubové a borovicové lesy (Quercus robur, Pinus
sylvestris), 155 – 185 m n. m., 1963 – 1977, leg. A. Dermek
BRA +
Fábry et al. (1975) Podunajská nížina, okolie Hurbanova × Fábry, Michalko (1975) Záhorská nížina, lúky medzi Zohorom a Plaveckým Štvrtkom × Janitor, Gáper (1998) Malá Fatra, Národná prírodná rezervácia Rozsutec, leg. J. Gáper ? Kautmanová (1996) Nízke Tatry, 2 km JZ (Múry) až 1 km J (Krieslo) od Važca, smrek, buk,
borovica na vápencovom podloží, 900 m n. m., 16. IX. 1995 ×
Kult (1989) Slovenský raj, Čingov, dolina potoka Lesnica, pri ceste, mach, 1. X. 1986 (resp. i 29.IX), leg. R. Fellner
PRM ?
Spišské kotliny, Popradská kotlina, 2,5 km V od Tatranskej Štrby, pri hlavnej ceste do Svitu, kataster obce Mengusovce, 1 – 2 km Z od hlavnej lesnej cesty, barinaté les. rašelinisko („Brezina“), v tráve – okraj Picea 3. X. 1986
Kul ?
Kuthan et al. (1999) Bukovské vrchy, Chotinka, M3, na zemi v tráve pod Picea a Fagus, 5. X. 1992, leg. J. Kuthan
Kut ?
M. microcephala Hagara (1987b) Malá Fatra, vrch Dubový diel, 1,8 km ZJZ od Bysričky, 570 m n. m.,
12. IX. 1982, leg. L. Hagara BRA? +
M. paedida Kuthan et al. (1999) Bukovské vrchy, Ruské, Nivné lúky pod ruským, na zemi v tráve,
12. X. 1995, leg. S. Adamčík BRA +
Bukovské vrchy, Nová Sedlica, nivné lúky Zbojského potoka (SZ od obce N. Sedlica), na zemi v tráve, 19. X. 1995, leg. S. Adamčík
BRA +
M. polioleuca Hagara (1987b) Veľká Fatra, kataster obce Turčianske Jaseno, chotárna časť
Biely potok, 2,6 km VJV od Turč. Jasena, 680 m n. m., 22. X. 1983, leg. L. Hagara
BRA? +
M. spegazzini Hagara (1989) Podunajská nížina, Q 7580, in robinieto ad terram arenosam graminosam
in phasianeto prope pagum Bajč (distr. Komárno), 120 m n. m., 29. VI. 1985, leg. L. Hagara
×
M. strictipes Dermek (1978) Záhorská nížina, územie medzi obcami Brodské, Čáry, Gbely, Kopčany,
Kúty a Smolinské, dubové a borovicové lesy (Quercus robur, Pinus
sylvestris), 155 – 185 m n. m., 1963 – 1977, leg. A. Dermek
BRA +
Dermek, Pilát (1974) Záhorská nížina, pri Kopčanoch, v svetlom listnatom lese (dub, breza), medzi konvalinkou voňavou a kokoríkom mnohokvetým, 25. V. 1968, leg. A. Dermek
×
Hagara (1992) Slovenský raj, Dobšiná, Q 71/88, 16. IX. 1985, leg. L. Hagara × Janitor (1997) Malé Karpaty, Klanec, bučina, 24. V. 1978, leg. V. Krčméry × Kautmanová (1996) Nízke Tatry, 2 km JZ (Múry) až 1 km J (Krieslo) od Važca, smrek, buk,
borovica na vápencovom podloží, 900 m n. m., 16. IX. 1995 ×
Kult (1991) Slanské vrchy, podcelok Šimonka, část Zlatobanská kotina, Pusté pole, V (JV) od silnice, 22. – 28. IX. 1990, leg. H. Deckerová, číslo položky: 685/90
Her ?
Kuthan et al. (1999) Bukovské vrchy, Ruské, terénna stanica Ruské – okolie, v tráve na zemi, 29. VI. 1990, leg. J. Kuthan a i.
BRA ×
Bukovské vrchy, Ruské, lúky nad terénnou stanicou, v tráve na zemi, 28. VIII. 1991, leg. J. Terray
×
Bukovské vrchy, Starina, lokalita Gazdoráň, v tráve na zemi, 16. VII. 1969, leg. K. Kult
PRM ?
Princ (1992) Liptovská kotlina, Východná, 1991 × M. stridula Kuthan (1989) Nízke Tatry, oblast u obce Važec (kat. území Važec, okr. Liptovský
Mikuláš), jižně od obce; západní část pásma Kozie chrbty, a to východněji položená oblast Vŕšky na SZ obvodu vrchu Slamená a západněji položená oblast Múry na SV svahu vrchu Hošková; vzácně, v trávě, 3. IX. – 9. IX. 1988
Her ?
Kuthan et al. (1999) Bukovské vrchy, Nová Sedlica, nivné lúky Zbojského potoka (SZ od obce N. Sedlica), na pasienku v tráve, 8. X. 1992, leg. J. Kuthan
Kut ?
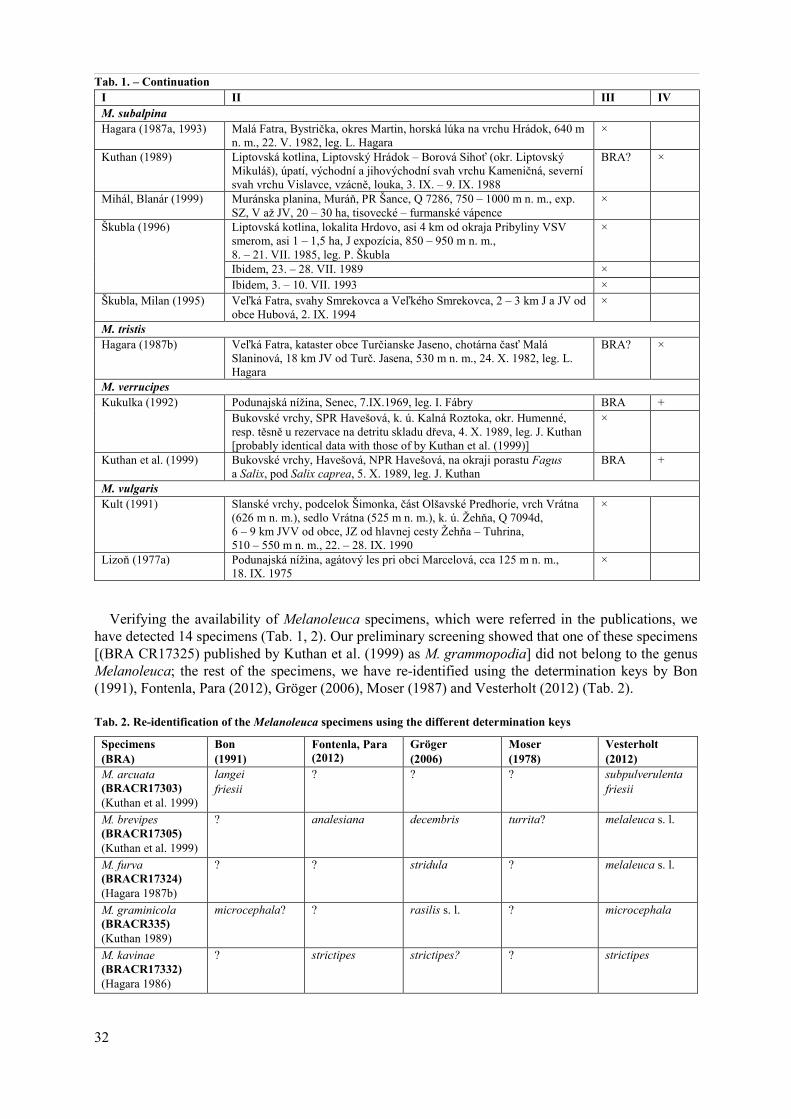
32
Tab. 1. – Continuation I II III IV M. subalpina Hagara (1987a, 1993) Malá Fatra, Bystrička, okres Martin, horská lúka na vrchu Hrádok, 640 m
n. m., 22. V. 1982, leg. L. Hagara ×
Kuthan (1989) Liptovská kotlina, Liptovský Hrádok – Borová Sihoť (okr. Liptovský Mikuláš), úpatí, východní a jihovýchodní svah vrchu Kameničná, severní svah vrchu Vislavce, vzácně, louka, 3. IX. – 9. IX. 1988
BRA? ×
Mihál, Blanár (1999)
Muránska planina, Muráň, PR Šance, Q 7286, 750 – 1000 m n. m., exp. SZ, V až JV, 20 – 30 ha, tisovecké – furmanské vápence
×
Škubla (1996) Liptovská kotlina, lokalita Hrdovo, asi 4 km od okraja Pribyliny VSV smerom, asi 1 – 1,5 ha, J expozícia, 850 – 950 m n. m., 8. – 21. VII. 1985, leg. P. Škubla
×
Ibidem, 23. – 28. VII. 1989 × Ibidem, 3. – 10. VII. 1993 ×
Škubla, Milan (1995) Veľká Fatra, svahy Smrekovca a Veľkého Smrekovca, 2 – 3 km J a JV od obce Hubová, 2. IX. 1994
×
M. tristis Hagara (1987b) Veľká Fatra, kataster obce Turčianske Jaseno, chotárna časť Malá
Slaninová, 18 km JV od Turč. Jasena, 530 m n. m., 24. X. 1982, leg. L. Hagara
BRA? ×
M. verrucipes Kukulka (1992) Podunajská nížina, Senec, 7.IX.1969, leg. I. Fábry BRA +
Bukovské vrchy, SPR Havešová, k. ú. Kalná Roztoka, okr. Humenné, resp. těsně u rezervace na detritu skladu dřeva, 4. X. 1989, leg. J. Kuthan [probably identical data with those of by Kuthan et al. (1999)]
×
Kuthan et al. (1999) Bukovské vrchy, Havešová, NPR Havešová, na okraji porastu Fagus a Salix, pod Salix caprea, 5. X. 1989, leg. J. Kuthan
BRA +
M. vulgaris Kult (1991) Slanské vrchy, podcelok Šimonka, část Olšavské Predhorie, vrch Vrátna
(626 m n. m.), sedlo Vrátna (525 m n. m.), k. ú. Žehňa, Q 7094d, 6 – 9 km JVV od obce, JZ od hlavnej cesty Žehňa – Tuhrina, 510 – 550 m n. m., 22. – 28. IX. 1990
×
Lizoň (1977a) Podunajská nížina, agátový les pri obci Marcelová, cca 125 m n. m., 18. IX. 1975
×
Verifying the availability of Melanoleuca specimens, which were referred in the publications, we have detected 14 specimens (Tab. 1, 2). Our preliminary screening showed that one of these specimens [(BRA CR17325) published by Kuthan et al. (1999) as M. grammopodia] did not belong to the genus Melanoleuca; the rest of the specimens, we have re-identified using the determination keys by Bon (1991), Fontenla, Para (2012), Gröger (2006), Moser (1987) and Vesterholt (2012) (Tab. 2).
Tab. 2. Re-identification of the Melanoleuca specimens using the different determination keys
Specimens (BRA)
Bon (1991)
Fontenla, Para (2012)
Gröger (2006)
Moser (1978)
Vesterholt (2012)
M. arcuata (BRACR17303) (Kuthan et al. 1999)
langei friesii
? ? ? subpulverulenta friesii
M. brevipes (BRACR17305) (Kuthan et al. 1999)
? analesiana decembris turrita? melaleuca s. l.
M. furva (BRACR17324) (Hagara 1987b)
? ? stridula ? melaleuca s. l.
M. graminicola (BRACR335) (Kuthan 1989)
microcephala? ? rasilis s. l. ? microcephala
M. kavinae
(BRACR17332) (Hagara 1986)
? strictipes strictipes? ? strictipes

33
Tab. 2 – Continuation Tab. 1. – Continuation Specimens (BRA)
Bon (1991)
Fontenla, Para (2012)
Gröger (2006)
Moser (1978)
Vesterholt (2012)
M. melaleuca
(BRACR190) (Dermek 1978)
? ? ? ? ?
M. microcephala
(BRACR17334) (Hagara 1987b)
graminicola ? stridula ? melaleuca s. l.
M. paedida (BRACR17337) (Kuthan et al. 1999)
favrei ? exscissa ? rasilis microcephala
M. paedida (BRACR17338) (Kuthan et al. 1999)
langei friesii
? vulgaris ? subpulverulenta
friesii
M. polioleuca (BRACR17341) (Hagara 1987b)
? ? brevipes ? grammopodia
excissa
M. strictipes (BRACR193) (Dermek 1978)
? strictipes strictipes? ? ?
M. verrucipes (BRACR17355) (Kuthan et al. 1999)
verrucipes verrucipes verrucipes verrucipes verrucipes
M. verrucipes (BRACR17356) (Kukulka 1992)
verrucipes? verrucipes? verrucipes? verrucipes? verrucipes?
Comments on the re-identified Melanoleuca specimens
The following comments on the re-identified Melanoleuca specimens (see also Tab. 2) include: 1. the brief description (based on our own observations) of the characters which are mostly used in the determination keys, and 2. our identification based on the determination keys which we have found to be problematic: a) we have not identified the specimen (“= no identification”) or we have come to more identifications (e.g. “= as M. langei, M. friesii”) – in these cases we list the problematic characters, i.e. not determinable characters on the dried specimen; b) we have had some doubts on our identification (e.g. “= as M. turrita?”) – some characters on the specimens do not correspond to those described in the key. BRACR17303 (originally as Melanoleuca arcuata) – cheilocystidia of macrocystis type; spores 6.5 – 8 × 4.5 – 5, av. 7.45 × 4.77, av. Q = 1.56; cap brown, with indistinct orange tones in the centre; stipe brown; lamellae beige, in some places with greyish or orange shade; trama in the stipe brown to orange; hyphae at the base of the stipe incrusted by brown crystals. The identification by Bon (1991) (= as M. langei, M. friesii) – ratio of the length and width of the stipe and cap; by Fontenla, Para (2012) (= no identification) – colours of the trama, cap and lamellae; by Gröger (2006) (= no identification) – colour of the lamellae and colour of the cap; by Moser (1978) (= no identification) – colour of the lamellae and surface of the cap; by Vesterholt (2012) (= as M. subpulverulenta, M. friesii) – surface of the cap.
BRACR17305 (originally as M. brevipes) – cheilocystidia absent; spores 7 – 9.5 × 5 – 7, av. 8.17 × 5.47, av. Q = 1.49; cap brown, darker in the centre, lighter on the edge; stipe black-brown; lamellae greyish-beige; trama in the stipe brown; hyphae at the base of the stipe incrusted by light yellowish-brown crystals. The identification by Bon (1991) (= no identification) – surface of the stipe; by Moser (1978) (= as M. turrita?). BRACR17324 (originally as M. furva) – cheilocystidia absent; spores 6 – 8 × 5 – 6, av. 6.82 × 5.5, av. Q = 1.24; cap brown; stipe light brown; lamellae light beige; trama in the stipe light beige; hyphae at the base of the stipe incrusted by light yel-lowish-brown crystals. The identification by Bon (1991) (= no identification) – colour of the lamellae and colour of the trama in the base of stipe; by Fontenla, Para (2012) and Moser (1978) (= no identifications) – colour of the lamellae.
BRACR335 (originally as M. graminicola) – cheilocystidia of urticocystis type; spores 6 – 8 × 4.5 – 5.5, av. 6.7 × 4.97, av. Q = 1.35; cap brown; stipe brown to black-brown; lamellae light beige; trama in the stipe brown; hyphae at the base of the stipe without incrustation. The identification by Bon (1991) (= as M. microcephala?); by Fontenla, Para (2012) and Moser (1978) (= no identifications) – colour of the lamellae.

34
BRACR17332 (originally as M. kavinae) – cheilocystidia of macrocystis type; spores 7 – 10 × 4.5 – 5.5, av. 8.77 × 4.95, av. Q = 1.77; cap whitish; stipe whitish; lamellae light beige; trama in the stipe whitish; hyphae at the base of the stipe without incrustation. The identification by Bon (1991) and Moser (1978) (= no identifications) – smell of the basidiomata; by Gröger (2006) (= as M. strictipes?).
BRACR190 (originally as M. melaleuca) – cheilocystidia of macrocystis type; spores 7 – 9 × 4 – 5, av. 7.62 × 4.80 av. Q = 1.59; cap brown with indistinct orange tones in the centre and on the edge of the cap; stipe brown with orange tones at the base; lamellae light beige, in some places with orange tones; trama in the stipe light brown with orange tones; hyphae at the base of the stipe incrusted by brown crystals. The identification by Bon (1991) (= no identification) – colour of the cap; by Fontenla, Para (2012) (= no identification) – colour of the trama; by Gröger (2006) (= no identification) – colour of the lamellae and colour of the cap; by Moser (1978) and Vesterholt (2012) (= no identifications) – colour of the lamellae.
BRACR17334 (originally as M. microcephala) – cheilocystidia absent; spores 6 – 8 × 4.5 – 5.5, av. 6.87 × 5.02, av. Q = 1.37; cap greyish-brown; stipe brown; lamellae light beige; trama in the stipe light with greyish tones; hyphae at the base of the stipe incrusted by brown crystals. The identification by Fontenla, Para (2012) and Moser (1978) (= no identifications) – colour of the lamellae.
BRACR17337 (originally as M. paedida) – cheilocystidia of urticocystis type; spores 6.5 – 9 × 4 – 5.5, av. 7.22 × 4.55, av. Q = 1.59; cap light, whitish with greyish and brownish tones; stipe whitish; lamellae light beige; trama in the stipe whitish; hyphae at the base of the stipe without incrustation. The identification by Fontenla, Para (2012) (= no identification) – surface of the cap; by Moser (1978) (= no identification) – smell of the basidiomata; by Vesterholt (2012) (= as M. rasilis, M. microcephala) – colour of the lamellae.
BRACR17338 (originally as M. paedida) – cheilocystidia of macrocystis type; spores 6 – 9 × 4.5 – 5, av. 7.25 × 4.82, av. Q = 1.5; cap greyish-brown; stipe greyish-brown; lamellae light, greyish-beige; trama in the stipe light beige; hyphae at the base of the stipe slightly incrusted by brown crystals. The identification by Bon (1991) (= as M. langei, M. friesii) – ratio of the length and width of the stipe and cap; by Fontenla, Para (2012) (= no identification) – colour of the trama; by Moser (1978) (= no identification) –colour of the lamellae; by Vesterholt (2012) (= as M. subpulverulenta, M. friesii) – surface of the cap.
BRACR17341 (originally as M. polioleuca) – cheilocystidia of urticocystis type; spores 6 – 10 × 4 – 6, av. 8.38 × 4.95, av. Q = 1.69; cap brown; stipe light, greyish-brown; lamellae light beige; trama in the stipe whitish; hyphae at the base of the stipe without incrustation. The identification by Bon (1991), Fontenla, Para (2012) and Moser (1978) (= no identifications) – colour of the lamellae; by Vesterholt (2012) (= as M. grammopodia, M. excissa) – surface of the stipe.
BRACR193 (originally as M. strictipes) – cheilocystidia of macrocystis type; spores 7 – 10 × 4 – 5, av. 8.52 × 4.75, av. Q = 1.79; cap whitish; stipe whitish; lamellae light beige; trama in the stipe light beige; hyphae at the base of the stipe without incrustation. The identification by Bon (1991) and Moser (1978) (= no identifications) – smell of the basidiomata; by Vesterholt (2012) (= no identification) – colour of the lamellae; by Gröger (2006) (= as M. strictipes?).
BRACR17355 (originally as M. verrucipes) – cheilocystidia of urticocystis type; spores 6 – 11 × 3 – 6, av. 8.50 × 5.05, av. Q = 1.83; cap whitish with brown centre; stipe whitish with blackish squamules; lamellae light beige; trama in the stipe whitish; hyphae at the base of the stipe without incrustation. The identification by all used determination keys is definite.
BRACR17356 (originally as M. verrucipes) – cheilocystidia of urticocystis type; spores 7.5 – 9 × 4.5 – 6, av. 8.32 × 4.55, av. Q = 1.68; cap light beige; stipe light brown, with few very indistinct brown squamules; lamellae beige; trama in the stipe beige; hyphae at the base of the stipe ? (the specimen is without the base of the stipe). The identification by all used determination keys is open; the specimen is too old and the characters – including the brown squamules on the stipe – are not determinable definitely.
The results of our case study – summing up in Tab. 2 and in the above mentioned “Comments on the
re-identified Melanoleuca specimens” – confirmed the difficulties in identification of Melanoleuca specimens. The lack of representative morphological characters connected with differently conceived species of various authors (e.g. Bon 1991, Fontenla, Para 2012, Gröger 2006, Moser 1978, and Vesterholt 2012) makes identification extremely difficult. Therefore a critical revision of European species including robust phylogeny based on multi-gene approach is desirable to elucidate diversity of genus in Europe. Accordingly, we think that the Melanoleuca names which are not backed up by the specimens are unusable; hopefully, the names with the specimens could be proved or corrected after re-identification following a new taxonomic concept of the genus Melanoleuca. For this moment, it is very important to gather Melanoleuca material, but providing that it would be sufficient not only for morphological (precise description), but also molecular analyses.

35
Acknowledgements
We are grateful to Ladislav Hagara (Bratislava), Pavel Lizoň and Viktor Kučera (Institute of Botany SAS, Bratislava) for help with literature. Comments by the reviewer were very helpful. Data included in this work were granted thanks to the project APVV SK-CZ-0052-11 and UK/327/2013.
References
Blanár, D., Mihál, I., 2002: Mykoflóra okolia Revúcej I (Slovenské rudohorie – Revúcka vrchovina). In: Uhrín, M. (ed.), Výskum a ochrana prírody Muránskej planiny 3, p. 33-52, Ministerstvo životného prostredia Slovenskej republiky, Správa Národného parku Muránska planina, Bratislava, Revúca.
Bon, M., 1991: Flore mycologique d’Europe 2. Les Tricholomes et ressemblants. Doc. Mycol., Mém. hors-Sér. 2: ii, 163 p. Boekhout, T., 1999: Melanoleuca. In: Bas, C., Kuyper, Th. W., Noordeloos, M. E., Vellinga, E. C. (eds.), Flora agaricina
neerlandica Vol. 4, p. 153-165, A. A. Balkema, Rotterdam, Brookfield. Dermek, A., 1971: Tmavuľka krátkohlúbiková (tmavobělka krátkonohá) – Melanoleuca brevipes (Bull. ex Fr.) Pat. Časopis
československých houbařů, 48(1-2): 1. Dermek, A., 1978: Príspevok k mykoflóre lesov na okolí Brodského, Čárov, Gbelov, Kopčian, Kútov a Smolinského (západné
Slovensko). Česká Mykol., 32: 215-225. Dermek, A., Pilát, A., 1974: Poznávajme huby. Veda, Bratislava. Ďuriška, O., Jančovičová, S., Tomšovský, M., Antonín, V., 2013: Morfologická a molekulárna analýza slovenských zá-
stupcov rodu Melanoleuca – predbežná štúdia. Mykologické listy, 125: 12. Fábry, I., Futó, E., Michalko, J., 1975: Mykoflóra širšieho okolia Hurbanova. Správy hubárskej poradne, III(2): 12-14. Fábry, I., Michalko, J., 1975: Záhorské lúky. Správy hubárskej poradne, III(2): 10. Futák, J., Dostál, J., Novák, F. A., 1966: Flóra Slovenska 1. Všeobecná časť. SAV, Bratislava. Fontenla, R., Para, R., 2012: Chiave sperimentale del genere Melanoleuca (unpublished). Gröger, F., 2006: Bestimmungsschlüssel für Blätterpilze und Röhrlinge in Europa, Teil 1. Regensb. Mykol. Schr., 13: 318-333. Hagara, L., 1986: Melanoleuca kavinae v Karpatoch. Mykologické listy, 24: 13-16. Hagara, L., 1987a: Atlas húb. Osveta, Martin. Hagara, L., 1987b: Zriedkavé lupenaté huby fatranských podhorských smrečín. In: Kuthan, J. (ed.), Houby horských smrčin
a podhorských smrkových porostu v Československu, p. 21-23, ČVSM pri ČSAV, Praha. Hagara, L., 1989: Vzácne prvky slovenskej mykoflóry. Česká Mykol., 43:51-60. Hagara, L., 1992: Huby – dvojníky. Obzor, Bratislava. Hagara, L., 1993: Atlas húb. Neografia, Martin. Janitor, A., 1997: Príspevok k poznaniu makromycétov zo skupiny Ascomycetes a Bazidiomycetes z územia Veľkej Bratislavy.
3. časť. Spravodajca slovenských mykológov, 16: 20-23. Janitor, A., Fábry, I., 1981: Príspevok k poznaniu mykoflóry Štátnej prírodnej rezervácie Rozsutec. In: Janík, M., Štolman,
A. (eds.), Rozsutec, p. 421-437, Osveta, Martin. Janitor, A., Gáper, J., 1998 (1997): Štúdium mykoflóry Národnej prírodnej rezervácie Rozsutec v rokoch 1996 a 1997. In:
Anonymus, Zborník zo seminára „Výskum a ochrana Krivánskej Fatry“, p. 39-48, Správa národných parkov Slovenskej republiky, Tatranská Štrba.
Kautmanová, I., 1996: Zoznam druhov nájdených počas V. mykologických dní na Slovensku. Spravodajca slovenských mykológov, 12: 28-30.
Kubička, J., 1957: Vzácnejšie alebo pre ČSR nové druhy vyšších húb z Tatranského národného parku. In: Anonymus, Sborník prác o Tatranskom národnom parku 1, p. 63-66, Osveta, Martin.
Kubička, J., 1976: Huby. In: Anonymus, Zborník prác o Tatranskom národnom parku 17, p. 195-205, Osveta, Martin. Kukulka, T., 1992: Tmavobělka bradavičnatá – Melanoleuca verrucipes (Fr.) Sing. Zpráva o jejím rozšíření v ČSFR. Časopis
československých houbařů, 69(2): 64-65. Kult, K., 1989: Súpis druhov húb zbieraných na exkurziách 29. IX. – 3. X. 1986 v okolí Spišskej Novej Vsi a na ďalších
lokalitách účastníkmi 4. mykologických dní na Slovensku. In: Kuthan, J. (ed.), Houby rašelinišť a bažinatých lesů v Česko-slovensku, p. 49-62, ČVSM pri ČSAV, Praha.
Kult, K., 1991: Soupis druhů hub sbíraných při mykofloristické akci sekce pro mykofloristiku a mykocenologii ČsVSM ve Slánskych vrchoch. In: Kuthan, J. (ed.), Houby rostoucí v prostředí ovlivněném činností člověka, p. D1-D22, Čarokruh – zájmové sdružení pro mykologii, Ostrava.
Kuthan, J., 1989: Soupis makromycetů zbíraných na průzkumných exkurzích (s myk. NDR) ve dnech 3. IX. – 9. IX. 1988 a na exkurzích „Setkání českých a slovenských mykologů pod Tatrami“ dne 10. IX. – 18. IX. 1988 na lokalitách v oblasti Vysokých Tater a Nízkych Tater a v Slovenském rudohoří. In: Kuthan, J. (ed.), Houby rašelinišť a bažinatých lesů v Česko-slovensku, p. 63– 90, ČVSM pri ČSAV, Praha.
Kuthan, J., Adamčík, S., Antonín, V., Terray, J., 1999: Huby Národného parku Poloniny (Fungi of the National Park Poloniny). Správa národných parkov SR, Správa Národného parku Poloniny, Liptovský Mikuláš, Snina.
Lizoň, P., 1977a: Exkurzie 1. mykologických dní na Slovensku. Správy hubárskej poradne, IV(1-2): 2-7. Lizoň, P., 1977b: VI. celoštátna mykologická konferencia v Pezinku. Správy hubárskej poradne, V(1-2): 2-5. Mihál, I., Blanár, D., 1999: Mykoflóra lokalít Hrdzavá dolina, Šance a Mokrá poľana v Národnom parku Muránska planina.
In: Uhrín, M. (ed.), Výskum a ochrana prírody Muránskej planiny 2, p. 21-33, Ministerstvo životného prostredia Slovenskej republiky, Správa Národného parku Muránska planina, Bratislava, Revúca.
Moser, M., 1978: Agarics and Boleti (Poylporales, Boletales, Agaricales, Russulales). Gustav Fischer Verlag, Stuttgart.

36
Princ, R., 1992a: Vzpomínaní na rok 1991. Časopis československých houbařů, 69(4): 121-123. Škubla, P., 1996: Mykoflóra Hrdova. Spravodajca slovenských mykológov, 12: 23-25. Škubla, P., Milan, I., 1995: Výsledky mykologického prieskumu Veľkej Fatry. Spravodajca slovenských mykológov, 7: 23-25. Vesterholt, J., 2012: Melanoleuca. In: Knudsen, H., Vesterholt, J. (eds.), Funga Nordica, p. 477-481, Nordsvamp, Copenhagen. Záhorovská, E., 1997: Huby (Fungi). In: Feráková, V., Kocianová, E. (eds.), Flóra, geológia a paleontológia Devínskej
Kobyly, p. 58-68, Litera, s.r.o., pre APOP, Bratislava. Záhorovská, E., Jančovičová, S., 1997: Mykoflóra alúvia rieky Rudavy. Spravodajca slovenských mykológov, 14: 15-19.
Abstrakt
Autori štúdie excerpovali z vybranej literatúry údaje o druhoch rodu Melanoleuca publikovaných zo Slovenska. Získané položky, na ktoré našli odkaz v literatúre, následne preurčili podľa rôznych určova-cích kľúčov. Nejednoznačné určenia vedú autorov k odporúčaniam ako narábať s dosiaľ publikovanými menami rodu Melanoleuca a tiež ako zbery rodu Melanoleuca správne určovať. Ondrej Ďuriška, Michal Tomšovský, Soňa Jančovičová: Ako narábať s publikovanými menami rodu Melanoleuca – prípadová štúdia zo Slovenska

37
Acta Botanica Universitatis Comenianae Vol. 48, 2013
MORPHOLOGICAL VARIABILITY OF SOME MEMBERS
OF THE FAMILY GEOGLOSSACEAE (HELOTIALES, FUNGI)
Nikola Rybáriková•
Slovak Academy of Sciences, Institute of Botany, Department of non-vascular plants,
Dúbravská cesta 9, 845 23 Bratislava, Slovakia
Received 5 November 2013; Received in revised form 2 December 2013; Accepted 5 December 2013
Abstract
Ageing changes of morphological characters were studied in 16 species of the family Geoglossaceae. In each species, three different ontogenetic stages of ascomata were observed (young, mature and old ascomata). The most significant changes during the ontogenesis were: paraphyses constriction in Geoglossum glabrum, browning and fragmentation of apical part of paraphyses in Trichoglossum walteri, and increasing number of septa in the spores in Geoglossum umbratile.
Key words: Microglossum, Thuemenidium, Trichoglossum, Geoglossum, Slovakia
Introduction
In the publication on Slovak fungi (Lizoň, Bacigálová 1998), only two species of the Geoglossaceae family are included: Microglossum viride (Pers.) Gillet and Trichoglossum hirsutum (Pers.) Boud. Recently, more extensive fieldwork research has shown that this group of fungi has significantly greater representation (Kučera et al. 2010, Kučera, Lizoň 2012). Majority of these fungi can be distinguished only with microscopic analysis. In some cases, however, the morphology of internal structures can be markedly changed during the ontogenesis, which complicates the process of species identification. To clarify such changeability, we have observed 16 taxa of Geoglossaceae in three different stages of their ontogenetic development: young, mature and old in each taxon.
Material and Methods
Collections examined. Sixteen collections of 16 species (one collection per one species) of the family Geoglossaceae were examined. They are deposited in the herbarium SAV (the herbarium of the Slovak Academy of Sciences, Institute of Botany). The list of these collections is attached in the text below. The localities of the species are listed according to the phytogeographical units (Futák 1984); the nearest city / town / village, name of the site, square grid number (Q) and habitat description is also added.
Ontogenetic stages of ascomata. In each species, i.e. within one collection, three different ontogenetic stages of ascomata were defined: a) young ascomata – the smallest ones; b) mature ascomata – the largest ones; c) old ascomata – those losing their strength and elasticity regardless of the size.
The micromorphological characters, i.e. characters of spores, asci and paraphyses, were observed in every ontogenetic stage of ascomata. In distilled water rehydrated dried material was examined using a Zeiss AxioScope light microscope. Each value of micromorphological structures was estimated as average of 30 measurements. The number of septa is presented as the most often number; other counted numbers are in the parenthesis.
The literature for identification. Ohenoja (2000), Moingeon (2004), Kučera et al. (2010), Dahlberg, Croneborg (2003). • Corresponding Author: Nikola Rybáriková; [email protected]

38
Material examined
Microglossum viride (Pers.) Gillet: Slovenské rudohorie Mts., Revúca city, Q 7386B, in the forest, ca. 415 m a. s. l., N 48°40´03.02´´, E 20°06´52.7´´, 15 Oct 2010, leg. V. Kučera (SAV 9920).
Microglossum nudipes Boud.: Muránska planina Plateau, Muránska Huta village, Predná Hora recreation area, Q 7286B, near the ski-lift, 787 m a. s. l., N 48°45´31.24´´, E 20°06´26.98´´, 10 Oct 2008, leg. V. Kučera (SAV 9830).
Microglossum olivaceum (Pers.) Gillet: Biele Karpaty Mts. (northern part), Vršatské Podhradie village, Q 6974B, in moss and grass, 753 m a. s. l., N 49°04´19.3´´, E 18°08´44.8´´, 1 Oct 2002, leg. V. Kučera (SAV 5956).
Microglossum rufescens (Grélet) Bon: Západné Beskydy Mts., Korňa village, Q 6577C, in meadow and bushes, 19 Sep 2010, leg. V. Kautman (SAV 10254).
Trichoglossum hirsutum (Pers.) Boud.: Považský Inovec Mts., Banka village, Q 7473A, in meadow and bushes, 261 m a. s. l., N 48°34´27.4´´, E 17°51´44.6´´, 29 Sep 2010, leg. V. Kučera (SAV 10000).
Trichoglossum octopartitum Mains: Malé Karpaty Mts., Sološnica village, Q 7569A, on walking path, ca. 250 m a. s. l., N 48°27´25´´, E 17°14´42´´, 3 Sep 2008, leg. V. Kučera (SAV 7560).
Trichoglossum variabile (E.J. Durand) Nannf. : Považský Inovec Mts., Banka village, Q 7473A, in meadow and bushes, 261 m a. s. l., N 48°34´27.4´´, E 17°51´44.6´´, 25 Oct 2008, leg. V. Kautman (SAV 9868).
Trichoglossum walteri (Berk.) E.J. Durand: Vihorlat Mts., Zemplínske Hámre village, Postávka peat bog, Q 7098D, in peat bog among Sphagnum sp. div., ca. 800 m a. s. l., N 48°55´11´´, E 22°09´44´´, 20 Sep 2001, leg. S. Ripková (SAV 5954).
Thuemenidium atropurpureum (Batsch) Kuntze: Biele Karpaty Mts. (southern part), Nová Bošáca village, Blažejová Nature Monument, Q 7172B, in meadow, 424 m a. s. l., N 48°52´32.8´´, E 17°49´05.9´´, 21 Oct 2010, leg. V. Kučera, N. Rybáriková (SAV 10039).
Geoglossum glutinosum Nees: Muránska planina Plateau, Muránska Huta village, Predná Hora recreation area, Q 7286B, near the ski-lift, 787 m a. s. l., N 48°45´31.24´´, E 20°06´26.98´´, 10 Oct 2008, leg. V. Kučera (SAV 10070).
Geoglossum glabrum Pers.: Vihorlatské vrchy Mts., Zemplínske Hámre village, Postávka peat bog, Q 7098D, in peat bog among Sphagnum sp. div., ca. 800 m a. s. l., N 48°55´11´´, E 22°09´44´´, 2 Sep 2011, leg. V. Kučera (SAV 10732).
Geoglossum cookeanum Nannf.: Malé Karpaty Mts., Chtelnica village, Q 7471B, in a meadow, 323 m a. s. l., N 48°34´20.72´´, E 17°35´37.28´´, 29 Sep 2010, leg. V. Kučera (SAV 9980).
Geoglossum fallax E.J. Durand: Západné Beskydy Mts., Korňa village, Q 6577C, in meadow and bushes, 19 Sep 2010, leg. V. Kautman (SAV 10258).
Geoglossum umbratile Sacc.: Považský Inovec Mts., Banka village, Q 7473A, in meadow and bushes, 261 m a. s. l., N 48°34´27.4´´, E 17°51´44.6´´, 27 Oct 2007, leg. V. Kautman (SAV 10277).
Geoglossum uliginosum Hakelier: Poľana Mts., Hriňová city, Q 7382D, in moist meadow in Sphagnum sp. div., 313 m a. s. l., N 48°37´49.99´´, E 19°28´10.53´´, 27 Sep 2009, leg. V. Kučera (SAV 10162).
Geoglossum simile Peck: Nízke Beskydy Mts., Habura village, Q 6697A, in peat bog among Sphagnum sp. div., N 49°21´10´´, E 21°53´15´´, 20 Sep 2006, leg. V. Kučera (SAV 9061).
Results and Discussion
In Microglossum, there was examined morphological variability of 4 species: M. viride, M. nudipes, M. olivaceum and M. rufescens (Tab. 1). The most significant changes observed during the ontogenesis were enlarging the apical part of paraphyses in M. viride (Fig. 1) and M. rufescens (Fig. 4) and pro-traction of the secondary branches of paraphyses in M. olivaceum (Fig. 3). Others less significant changes observed: widening of paraphyses in M. nudipes (Fig. 2), coarsening and protracting of asci in M. nudipes, protracting of asci in M. nudipes (Tab. 1) and shortening of asci in M. viride (Tab. 1). Increasing of ascospores length was observed in all members: M. viride, M. nudipes, M. olivaceum and M. rufescens (Tab. 1). The width of ascospores in M. nudipes and M. olivaceum and width of asci in M. rufescens did not change significantly (Tab. 1).
There were 4 species examined in the genus Trichoglossum: T. hirsutum, T. octopartitum, T. variabile and T. walteri (Tab. 2). There were some significant changes observed in all members, such as frag-mentation of apical part of paraphyses, darkening of setae and also increasing number of septa in the spores (Tab. 2). There were also observed browning of paraphyses in T. walteri (Fig. 8) and narroving

39
and protracting of asci in T. hirsutum (Tab. 2). In T. variabile and T. walteri, the longest asci and ascospores were observed in mature ascomata (Tab. 2). The width of ascospores in T. octopartitum and T. variabile as well as width of asci in T. octopartitum did not change significantly (Tab. 2).
Tab. 1. Development of the internal structures in the studied Microglossum specimens
ascospore
width
ascospore
length
ascus
width
ascus
length
ascospore
septation
Microglossum viride (SAV 9920)
Young 3.9 15.9 9.2 105.6 0
Mature 4.4 16.6 8.3 103.1 0
Old 4.1 17.7 9.3 102.3 0
Microglossum nudipes (SAV 9830)
Young 3.2 14.1 5.3 62.6 0
Mature 3.3 14.9 6.7 72.8 0
Old 3.3 15.6 7.2 79 0
Microglossum olivaceum (SAV 5956)
Young 3.1 10.4 7.5 74.8 0
Mature 3.2 11 7.8 80.1 0
Old 3.1 11.5 7.4 69.6 0
Microglossum rufescens (SAV 10254)
Young 3.4 12.7 6.3 59.6 0
Mature 3.4 11.8 6.4 59.3 0
Old 3.9 13.3 6.4 63.3 0
* Each present value of width and length of ascospores and asci is an average of 30 measurements (µm)
Tab. 2. Development of the internal structures in the studied Trichoglossum specimens
ascospore
width
ascospore
length
ascus
width
ascus
length
ascospore
septation
Trichoglossum hirsutum (SAV 10000)
Young 5.7 114.3 19.1 155.1 11 (10,12,13)
Mature 5.9 97.9 18.1 149.3 12 (11,13,15)
Old 5.5 106.8 16.8 170.8 13 (12,14,15)
Trichoglossum octopartitum (SAV 7560)
Young 5.2 119.8 20 214.4 5 (6,7)
Mature 5.1 108.1 20.3 205.8 7
Old 5.3 108.8 20.5 213.6 7
Trichoglossum variabile (SAV 9868)
Young 4.9 121.4 18.3 220.9 10 (9,11,12)
Mature 4.9 132.5 21.8 284.2 13 (12,14,15)
Old 4.9 129.3 19.6 219.4 13 (12,14,15)
Trichoglossum walteri (SAV 5954)
Young 6.7 77.5 19.8 200.1 5 (3,6,7)
Mature 6.4 85.3 22.2 214.5 6 (3,5,7)
Old 4.9 81.7 19.8 202.2 6 (3,5,7)
* Each present value of width and length of ascospores and asci is an average of 30 measurements (µm); the number of septa is presented as the most often number; other counted numbers are in the parenthesis
In Thuemenidium, only 1 species was examined: Thuemenidium atropurpureum (Tab. 3). The most
significant changes during the ontogenesis were coarsening and protracting of ascospores (Tab. 3).

40
There was noted also gentle constriction of the apical part of paraphyses (Fig. 9). The largest asci were measured in mature ascomata (Tab. 3). The asci width did not change significantly.
Tab. 3. Development of the internal structures in the studied Thuemenidium specimens
ascospore
width
ascospore
length
ascus
width
ascus
length
ascospore
septation
Thuemenidium atropurpureum (SAV 10039)
Young 3.8 23 8.4 111.2 0
Mature 4.2 23.9 9.1 123.7 0
Old 4.2 26.7 8.9 105.1 0
* Each present value of width and length of ascospores and asci is an average of 30 measurements (µm)
In Geoglossum, there were 7 species examined: G. glutinosum, G. glabrum, G. cookeanum, G. fallax,
G. umbratile, G. uliginosum and G. simile (Tab. 4).
Tab. 4. Development of the internal structures in the studied Geoglossum specimens
ascospore
width
ascospore
length
ascus
width
ascus
length
ascospore
septation
Geoglossum glutinosum (SAV 10070)
young 4.3 77.4 12.1 227.6 3 (1,5)
mature 4.4 78.4 14.1 228.9 5 (3,6,7)
old 4.4 80.8 11.8 224.4 7 (3)
Geoglossum glabrum (SAV 10732)
young 4.9 76.3 23.1 202.9 7
mature 4.9 76.9 21.1 195.2 7
old 4.9 74.6 20.1 186.1 7
Geoglossum cookeanum (SAV 9980)
young 6.4 77.7 22.2 183.7 7
mature 5.6 75.5 18.8 174.9 7
old 5.8 78.1 19.4 179.8 7
Geoglossum fallax (SAV 10258)
young 4.9 88 18.6 173.9 1 (3)
mature 6 94.3 17.4 164.4 8 (7,9,10)
old 4.8 92.4 17.4 190.1 7 (7,9,10)
Geoglossum umbratile (SAV 10277)
young 5.6 93.2 20.5 183.3 3 (6,7)
mature 5.9 97.9 19.6 164.1 4 (3,7)
old 5.4 102.6 21.5 180.7 4 (3,6,7)
Geoglossum uliginosum (SAV 10162)
young 5.1 71.2 21.8 192.2 6 (3,5,7)
mature 4.9 79.9 19.6 186.1 7 (6)
old 5.1 77.9 20.2 188.4 7
Geoglossum simile (SAV 9061)
young 6.8 80.9 22.2 185.4 7
Mature 7.3 84.1 26.3 161.6 7
Old 6.8 84.6 22.7 175.9 7
* Each present value of width and length of ascospores and asci is an average of 30 measurements (µm); the number of septa is presented as the most often number; other counted numbers are in the parenthesis
41
Figs. 1–8. Ascomata and three ontogenetic stages of paraphyses (a – young, b – mature, c – old) of
the species: 1. Microglossum viride, 2. M. nudipes, 3. M. olivaceum, 4. M. rufescens, 5. Trichoglossum
hirsutum, 6. T. octopartitum, 7. T. variabile, 8. T. walteri (scale bar for paraphyses 10 µm)
The most significant change during ontogenesis was constriction of apical part of paraphyses in G. glabrum (Fig. 11). The other significant changes observed: creasing of apical part of paraphyses in G. umbratile (Fig. 14), browning of apical part of paraphyses in G. glutinosum (Fig. 10) and G. simile (Fig. 16), constriction of apical part of paraphyses in G. cookeanum (Fig. 12), widening of apical part of paraphyses in G. fallax (Fig. 13), narrowing and shortening of asci in G. glabrum (Tab. 4), protruding ascospores in G. umbratile (Tab. 4) and increasing number of septa in spores in G. glutinosum, G. um-bratile and G. uliginosum (Tab. 4). There were also observed gentle coarsening of paraphyses and increasing number of paraphyses cells in G. uliginosum (Fig. 15). The ascospores width in G. glutinosum, G. glabrum, and G. uliginosum did not change. Similarly, there were no significant changes in the ascospores septation in G. glabrum, G. cookeanum and G. simile in the process of ontogenesis.

42
Figs. 9–15. Ascomata and three ontogenetic stages of paraphyses (a – young, b – mature, c – old)
of the species: 9. Thuemenidium atropurpureum, 10. Geoglossum glutinosum, 11. G. glabrum,
12. G. cookeanum, 13. G. fallax, 14. G. umbratile, 15. G. uliginosum, 16. G. simile (scale bar for
paraphyses 10 µm)
The issue about changing of internal structures during the ontogenesis came to the attention even before this study. Deckerová (2006) was the first who draw attention to this changebility. In her view, the morphological variability of some members of Geoglossaceae may complicate the process of identification. Her study of Geoglossum glabrum in various ontogenetic stages proved that there are extensive variability of internal structures (specially in paraphyses). Based on the results of this study, the paraphyses changes are not the only ones. There are several and for each species specific ways of development in Geoglossaceae. However, only few of them can also complicate the process of species identification.
In Microglossum and Thuemenidium, there was observed very small changes compared to Tricho-glossum and Geoglossum and the species can be reliably identify in every phase of ontogenesis.

43
In Trichoglossum, there is only one change which can be misleading in identification, the number of septa in the spores. Therefore it is important to examine mature ascomata where ascospores are already developed. The other changes are in range of variability in each species and the ascomata can be determined in every phase of ontogenesis.
In Geoglossum, there were observed the most of changes of all studied genera. There were noticed changes of shape, size, colour, as well as number of septa. G. glabrum was shown to be as the most changeable. The paraphyses changes are so marked that it would seem there are two or three different taxa within the collection (Dennis 1978). To verify the identification, it is important to examine several phases of ascomata in each collection. To know the correct number of septa in the spores, it is properly to study mature and old ascomata. The other changes during ontogenesis were not as significant to be misleading in identification.
According to Benkert (1996) the shape and size of apical part of paraphyses are the most important distinguishing characters in Geoglossaceae. However, it is the most changeable and misleading character. There must be deliberate also the way of interpretation, which depends on individual view. Generally, as the most suitable for identification I consider the mature phase of ascomata. The internal structures are already developed and all distinguished characters are present. In case of measurable structures, I consider the average values as the most advisable. There is high assumption of inclusion of several stages of ascomata at once.
Acknowledgements
I would like to express my thanks to Viktor Kučera for directing my work, to Pavel Lizoň for reading and commenting on the manuscript, and to Milan Zajac for advice on graphic and designe (all acknowledged are from the Slovak Academy of Sciences, Institute of Botany, Bratislava). This study was supported by grants VEGA 2/0062/10 and VEGA 2/0088/13.
References
Benkert, D., 1996: Zur Variabilität der Paraphysen in der Gattung Geoglossum: was ist Geoglossum barlae? Fed. Repert., 107: 269-276.
Dahlberg, A., Croneborg, H., 2003: 33 threatened fungi in Europe. Complementary and revised information on candidates for listing in Appendix 1 of the Bern Convention. Swedish Conservation of Fungi, Uppsala.
Deckerová, H., 2006: Problematika určování druhů rodu Geoglossum. Mykol. Listy, 98: 8-12. Dennis, R. W. G., 1978: British Ascomycetes. J. Cramer, Vaduz. Futák, J., 1984: Fytogeografické členenie Slovenska. In: Bertová, L. (ed.) Flóra Slovenska IV/1, p. 418-419, Veda, Bratislava. Kučera, V., Lizoň, P., Kautmanová, I., 2010: Geoglossoid fungi in Slovakia II. Trichoglossum octopartitum, a new species
for the country. Czech Mycol., 62: 13-18. Kučera, V., Lizoň, P., 2012: Geoglossaceous fungi in Slovakia III. The genus Geoglossum. Biologia, 67: 654-658. Lizoň, P., Bacigálová, K., 1998: Huby. In: Marhold, K., Hindák, F. (eds.), Zoznam nižších a vyšších rastlín Slovenska, p. 101-
227, Veda, Bratislava. Moingeon, J. M., 2004: Contributions á ľétude des Geoglossaceae á spores hyalines. Miscellanea Mycologica, 80-81: 25-35. Ohenoja, E., 2000: Geoglossaceae. In: Hansen, L., Knudsen, H. (eds.), Nordic macromycetes, Vol. 1, p. 178-183, Nordsvamp,
Copenhagen.
Abstrakt
Študovali sme zmeny morfologických znakov počas stárnutia pri 16 druhoch húb čeľade Geoglos-saceae. Pri každom druhu sme hodnotili tri rôzne vývinové štádiá plodníc (mladé, dospelé a staré plodnice). Najvýraznejšie zmeny počas ontogenézy boli: stenšenie parafýz u Geoglossum glabrum, hnednutie a fragmentácia apikálnych častí parafýz pri Trichoglossum walteri a zvýšenie počtu prie-hradok vo výtrusoch pri Geoglossum umbratile.
Nikola Rybáriková: Morfologická variabilita niektorých zástupcov čeľade Geoglossaceae
(Helotiales, huby)

ACTA
BOTANICA
UNIVERSITATIS
COMENIANAE
Volume 48
Vydala Univerzita Komenského v Bratislave vo Vydavateľstve UK
Vyšlo v decembri 2013
Technická redaktorka: Darina Földešová
ISBN 978-80-223-3561-4
ISSN 0524-2371


















