
Acellularity of starfish embryonic mesenchyme cells as...
10
Development 109, 129-138(1990) Printed in Great Britain © The Company of Biologists Limited 1990 129 Acellularity of starfish embryonic mesenchyme cells as shown in vitro HIROYUKI KANEKO 1 , SHIGEKO TAKAICHI 2 , MASAMICHI YAMAMOTO 3 and MARINA DAN-SOHKAWA 1 ' Department of Biology, Faculty of Science, Osaka City University, Osaka, Japan. 2 National Cardiovascular Center Research Institute, Osaka, Japan. 3 Ushimado Marine Laboratory, Okayama University, Ushimado, Okayama, Japan. Summary Embryonic mesenchyme cells of the starfish are shown to be unexpectedly fusogenic in vitro. When archenteron complexes (archenterons and varying portions of the extracellular matrix {ECM} surrounding them) are isolated from starfish embryos and inoculated in sea water containing 4 % newborn bovine serum, the mesen- chyme cells form large syncytia on the substratum underneath each archenteron. These syncytia break into smaller fragments interconnected by fine cell processes within 24 h. These networks have been studied morpho- logically, dynamically and ultrastructurally and found to lack cell borders between the constituent fragments. These fragments contain various numbers of nuclei ranging from 0 to 6 or more and move about constantly over the substratum, sometimes breaking into two and sometimes fusing with neighbouring fragments, so that the overall pattern of the network changes constantly. Our results also indicate that the network is three- dimensional i.e. it has crossing sites, the frequency of which seems to depend on the amount of the ECM excreted on the substratum. A similar network pattern is found among mesenchyme cells in vivo, which suggests that the features found in vitro reflect those in vivo. Key words: starfish embryo, mesenchyme cells, in vitro, acellularity, dynamic network, three-dimensional network. Introduction In early embryogenesis of many multicellular organ- isms, mesenchyme cells differentiate from epithelial cells and migrate through the extracellular matrix (ECM) of the blastocoel as individual cells before participating in morphogenetic activities. Mesenchyme cells of vertebrate embryos have been extensively studied; e.g. their transformation from epithelial cells (Greenburg and Hay, 1986, 1988), the mechanism of their migration (Thiery, 1984), interaction with epi- thelial cells (Haffen etal. 1987; Cunha, 1984; Dhouailly and Sengel, 1983) and the ECM (Tomasek et al. 1982) etc. However, studies of invertebrate mesenchyme cells are still very limited. The primary mesenchyme cell of the sea urchin embryo, which specifically functions in the formation of spicules, is probably the most extensively studied mes- enchyme cell of invertebrate origin. In contrast, the secondary mesenchyme cell, which has more general functions in embryogenesis such as a role in gastrulation (Dan and Okazaki, 1956; Gustafson and Kinnander, 1956; Spiegel and Burger, 1982), has received less attention. The single type of mesenchyme cell found in most animal bodies composed of three germ layers seem to correspond with the secondary mesenchyme cell. In the starfish embryo, the mesenchyme cells form at the tip of the embryonic archenteron and migrate into the blastocoel (Chia, 1977) in a manner similar to the secondary mesenchyme cells of the sea urchin. These cells eventually take part in the formation of the mouth and coelomic pouches (Crawford and Abed, 1978, 1983) and form 'fibrous networks' on the ectodermal wall and on the surface of the larval gut (Chia, 1977). In order to elucidate the nature and function of these cells more clearly, we have observed them morphologi- cally, dynamically and ultrastructurally under culture conditions. They form a large syncytium at the begin- ning of the culture, instead of migrating singly as found in vivo. The large syncytium, however, eventually spread out into a network of small syncytial fragments interconnected by fine cell processes. Our results indi- cate that the mesenchyme cells are fundamentally acellular in nature, with no cell boundaries between the cell bodies or the cell processes. We also show that the network is three-dimensional in nature with crossing points. We have also studied living embryos to see if the network pattern of the mesenchyme cells in vitro is also present in vivo. The acellularity of these cells is dis- cussed in relation to their physiological role in the embryogenesis.
Transcript of Acellularity of starfish embryonic mesenchyme cells as...

Development 109, 129-138(1990)Printed in Great Britain © The Company of Biologists Limited 1990
129
Acellularity of starfish embryonic mesenchyme cells as shown in vitro
HIROYUKI KANEKO1, SHIGEKO TAKAICHI2, MASAMICHI YAMAMOTO3 and
MARINA DAN-SOHKAWA1
' Department of Biology, Faculty of Science, Osaka City University, Osaka, Japan.2National Cardiovascular Center Research Institute, Osaka, Japan.3Ushimado Marine Laboratory, Okayama University, Ushimado, Okayama, Japan.
Summary
Embryonic mesenchyme cells of the starfish are shown tobe unexpectedly fusogenic in vitro. When archenteroncomplexes (archenterons and varying portions of theextracellular matrix {ECM} surrounding them) areisolated from starfish embryos and inoculated in seawater containing 4 % newborn bovine serum, the mesen-chyme cells form large syncytia on the substratumunderneath each archenteron. These syncytia break intosmaller fragments interconnected by fine cell processeswithin 24 h. These networks have been studied morpho-logically, dynamically and ultrastructurally and foundto lack cell borders between the constituent fragments.These fragments contain various numbers of nuclei
ranging from 0 to 6 or more and move about constantlyover the substratum, sometimes breaking into two andsometimes fusing with neighbouring fragments, so thatthe overall pattern of the network changes constantly.Our results also indicate that the network is three-dimensional i.e. it has crossing sites, the frequency ofwhich seems to depend on the amount of the ECMexcreted on the substratum. A similar network pattern isfound among mesenchyme cells in vivo, which suggeststhat the features found in vitro reflect those in vivo.
Key words: starfish embryo, mesenchyme cells, in vitro,acellularity, dynamic network, three-dimensional network.
Introduction
In early embryogenesis of many multicellular organ-isms, mesenchyme cells differentiate from epithelialcells and migrate through the extracellular matrix(ECM) of the blastocoel as individual cells beforeparticipating in morphogenetic activities. Mesenchymecells of vertebrate embryos have been extensivelystudied; e.g. their transformation from epithelial cells(Greenburg and Hay, 1986, 1988), the mechanism oftheir migration (Thiery, 1984), interaction with epi-thelial cells (Haffen etal. 1987; Cunha, 1984; Dhouaillyand Sengel, 1983) and the ECM (Tomasek et al. 1982)etc. However, studies of invertebrate mesenchyme cellsare still very limited.
The primary mesenchyme cell of the sea urchinembryo, which specifically functions in the formation ofspicules, is probably the most extensively studied mes-enchyme cell of invertebrate origin. In contrast, thesecondary mesenchyme cell, which has more generalfunctions in embryogenesis such as a role in gastrulation(Dan and Okazaki, 1956; Gustafson and Kinnander,1956; Spiegel and Burger, 1982), has received lessattention. The single type of mesenchyme cell found inmost animal bodies composed of three germ layers
seem to correspond with the secondary mesenchymecell.
In the starfish embryo, the mesenchyme cells form atthe tip of the embryonic archenteron and migrate intothe blastocoel (Chia, 1977) in a manner similar to thesecondary mesenchyme cells of the sea urchin. Thesecells eventually take part in the formation of the mouthand coelomic pouches (Crawford and Abed, 1978,1983) and form 'fibrous networks' on the ectodermalwall and on the surface of the larval gut (Chia, 1977).
In order to elucidate the nature and function of thesecells more clearly, we have observed them morphologi-cally, dynamically and ultrastructurally under cultureconditions. They form a large syncytium at the begin-ning of the culture, instead of migrating singly as foundin vivo. The large syncytium, however, eventuallyspread out into a network of small syncytial fragmentsinterconnected by fine cell processes. Our results indi-cate that the mesenchyme cells are fundamentallyacellular in nature, with no cell boundaries between thecell bodies or the cell processes. We also show that thenetwork is three-dimensional in nature with crossingpoints. We have also studied living embryos to see if thenetwork pattern of the mesenchyme cells in vitro is alsopresent in vivo. The acellularity of these cells is dis-cussed in relation to their physiological role in theembryogenesis.

130 H. Kaneko and others
Materials and methods
MaterialsDeveloping embryos of the starfish, Asterina pectinifera, wereobtained by the method described previously (Dan-Sohkawa,1976), in which the eggs treated with 1-methyladenine (1-MA)(Kanatani, 1969) were inseminated with diluted dry spermand allowed to develop in the artificial sea water, Jamarin U(Jamarin Lab., Osaka, Japan), at 20°C. Embryos werecollected at various stages of development by hand centrifu-gation and were submitted to further treatments. The no-menclature of the developmental stages of the starfish followsthat presented previously (Dan-Sohkawa et al. 1980).
MediaArchenteron preparation medium (APM): 1.2M glycine indistilled water (DW) supplemented with 1/100 volume ofJamarin U (Dan-Sohkawa and Kaneko, 1989).Culture medium: Jamarin U supplemented with 4 % newbornbovine serum (M. A. Bioproduct Lab.).Detaching medium: Ca2+/Mg2+-free Jamarin (Jamarin Lab.,Osaka, Japan).
Preparation of the mesenchymal network in vitroArchenteron complexes (archenterons and varying portionsof the ECM surrounding them) were prepared by the methodreported previously (Dan-Sohkawa and Kaneko, 1989), inwhich 10 ml of APM was added to a centrifuge tube contain-ing 0.4 ml of packed embryos at the mouth formation stage.After 5 min of treatment, the medium was changed to 4 ml ofthe same medium, in which embryos were submitted to 10 to15 gentle strokes of a broad-mouthed pipette (Komagome'spipette; tip inside diameter 1.5-2 mm) to remove the ecto-derm. Archenteron complexes were separated from dis-sociated ectodermal cells and fragments by hand centrifu-gation, washed twice with 10 ml of Jamarin U and suspendedin 3 ml of the culture medium. Various amounts of thissuspension, ranging from one drop to lml, was inoculated infinal 1 to 1.5 ml of the culture medium, and cultured at 20°Con to different substrata, as explained below for each case.
Coculture of stained and unstained mesenchyme cellsThe network cultured for 24 h in a plastic dish (35 mm indiameter; Falcon no. 3801) was exposed to 0.5 % of methyl-ene blue in culture medium for 15 min, in order to stain thecytoplasmic granules of the constituent mesenchyme cells.The stained cells were detached from the surface by incu-bating them for 10 min with the detaching medium and twosuccessive washes with the same medium. The suspension wascentrifuged at 1500 revs min"1 for 5 min to collect the stainedcells. After removal of the supernatant, they were reinocu-lated over the network of unstained cells.
Phase-contrast and fluorescence microscopyThe morphology and behavior of the network was observedwith a phase-contrast microscope (Nikon TMD). The be-havior of the stained granules in cocultures of stained andunstained cells, was traced under the bright-field optics(Goshima et al. 1984).
In order to observe the nuclei of the cultured mesenchymecells, the archenteron complexes were inoculated on a glasscoverslip (18x18 mm) and the resultant network was culturedfor 24 h. The network was treated for 30 min with final2/jgml"1 of Hoechst 382065 (Calbiochem Lab.) added to theculture medium, and washed briefly with Jamarin U. It wasthen mounted on a slide glass and the nuclei were located
under a phase-constrast microscope coupled with fluor-escence equipment (Nikon TMD).
Electron microscopyFor TEM observations, the network cultured on a plastic dishfor 24 h was washed and fixed with 3 % glutaraldehyde in 0.1 Mcacodylate buffer (pH 7.2) for 30 min at 4°C. The network waswashed for several times in the same buffer, postfixed with1 % OsO4 in DW for 30 min at 4°C, and washed with DW at4°C. The network was then stained en bloc for 30 min with2% aqueous uranyl acetate at 20 °C, followed by anotherwash with 0.1 % sodium acetate to remove remaining uranylacetate. After dehydration by passage through a gradedethanol series, the network was removed from the plasticdishes with propylene oxide and centrifuged in the samesolvent to form a pellet, which was finally embedded inSupper's low viscosity resin, and ultrathin sections cut. Theultrathin sections were stained with 3 % uranyl acetate in30% ethanol followed by treatment with Reynold's leadcitrate and observed with a H-600 transmission electronmicroscope (Hitachi).
For SEM observations, the archenteron complexes wereinoculated on tissue culture coverslips (13 mm in diameter,Thermanox, Miles Lab. Inc.) and the resultant networks werecultured for 24 h. They were washed with Jamarin U toremove the culture medium, fixed with 2% OsO4 in JamarinU for 15 min at 20°C and rinsed with DW for 15 min. Afterdehydration through an ethanol series, the specimens werecritical-point dried (Hitachi HCP-2), sputter-coated with gold(Eiko IB-3), and examined by a scanning electron microscope(JEOL T-300).
Differential interference microscopyIn order to search for a mesenchymal network in vivo,embryos were anesthesized with a small amount of 4 %paraformaldehyde added from the sides of the glass coverslipimmediately before taking photographs and were observedunder a differential interference microscope (Olympus IMT-2-21 RFN).
Results
Formation and dynamics of mesenchymal network invitroImmediately after preparation, the mesenchyme cellsare scattered singly in the ECM, which surrounds thearchenteron (Fig. 1). The ECM becomes deflatedwithin an hour and the archenteron attaches to thesurface of the substratum. The mesenchyme cells, onthe other hand, escape from the ECM and form a largesyncytium instead of migrating singly on the sub-stratum. Within one hour following attachment, thesyncytium can be seen as a large lamellipodium migTat-ing from underneath the archenteron (Fig. 2A). Itsteadily migrates away from the archenteron and breaksinto smaller fragments (Fig. 2B-D). By 24h, a typicalnetwork is formed around the archenteron (Fig. 2E).
The network is composed of two parts, the cell bodyand the cell process (Fig. 3A). The cell body iselongated often along the paths lamellipodia and filo-podia. The majority of the cell bodies contain onenucleus, whereas some contain 2-6 and still others none(Fig. 3B). Fine cell processes connect two cell bodies or
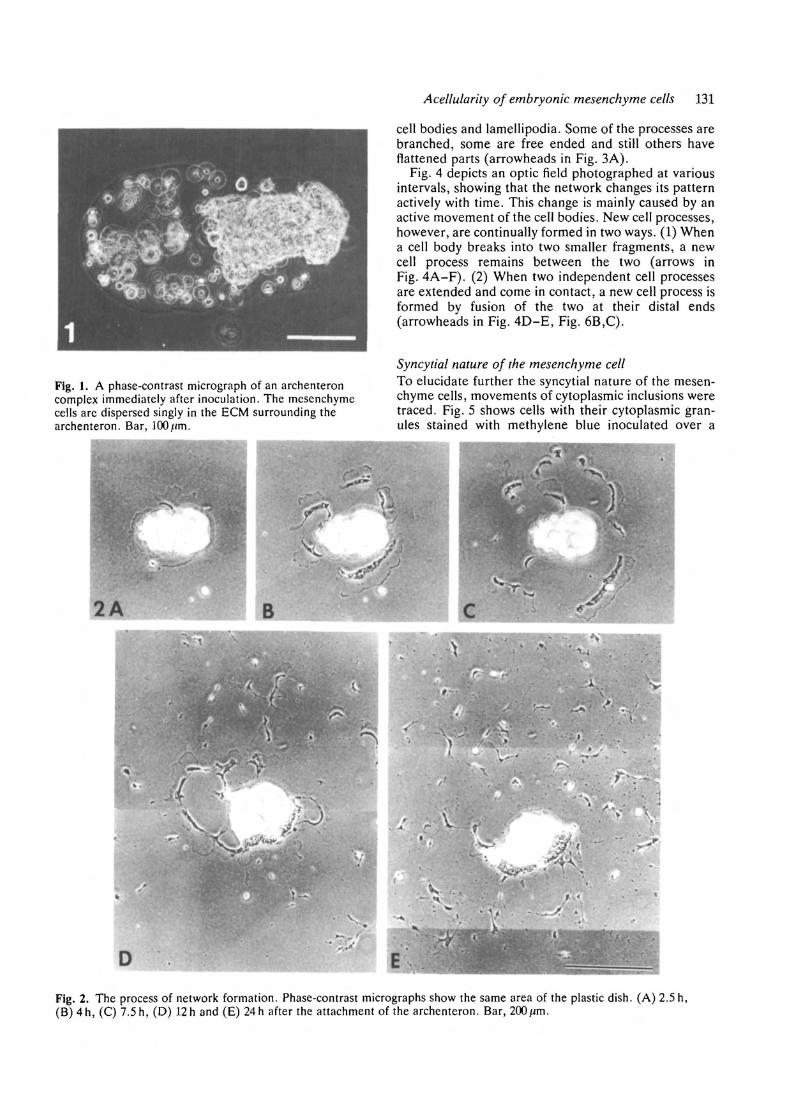
Fig. 1. A phase-contrast micrograph of an archenteroncomplex immediately after inoculation. The mesenchymecells are dispersed singly in the ECM surrounding thearchenteron. Bar, 100/.im.
Acellularity of embryonic mesenchyme cells 131
cell bodies and lamellipodia. Some of the processes arebranched, some are free ended and still others haveflattened parts (arrowheads in Fig. 3A).
Fig. 4 depicts an optic field photographed at variousintervals, showing that the network changes its patternactively with time. This change is mainly caused by anactive movement of the cell bodies. New cell processes,however, are continually formed in two ways. (1) Whena cell body breaks into two smaller fragments, a newcell process remains between the two (arrows inFig. 4A-F). (2) When two independent cell processesare extended and come in contact, a new cell process isformed by fusion of the two at their distal ends(arrowheads in Fig. 4D-E, Fig. 6B,C).
Syncytial nature of the mesenchyme cellTo elucidate further the syncytial nature of the mesen-chyme cells, movements of cytoplasmic inclusions weretraced. Fig. 5 shows cells with their cytoplasmic gran-ules stained with methylene blue inoculated over a
Fig. 2. The process of network formation. Phase-contrast micrographs show the same area of the plastic dish. (A) 2.5 h,(B) 4h, (C) 7.5 h, (D) 12 h and (E) 24 h after the attachment of the archenteron. Bar, 200^m.

132 H. Kaneko and others
<-
1 • •:. ' .
4
A -.• f
3AFig. 3. A phase-contrast (A) and a fluorescence (B) micrograph of the mesenchymal network (24h in culture). (A) Thenetwork is composed of expanded cell bodies, accompanied with broad lamellipodia and fine cell processes. Arrow: afragment lacking the nucleus. Arrowheads: flattened portions of the cell processes. (B) The same optic area as A, stainedwith Hoechst dye (no. 382065) to show the location of the nuclei. Most of the cell bodies contain only one nucleus. Arrows:irregularly shaped nuclei. Bar, 50 urn.
sparse network of unstained cells. Fig. 6 illustratesschematically the movement of the upper three cellsshown in Fig. 5 along with that of the stained granulesand the nuclei. The cells exchange their cytoplasmicinclusions as they repeat fusion and separation. Actu-ally, three mononucleate cells containing the stainedgranules and one cell containing unstained granuleswere produced from two mononucleate stained cellsand a binucleate unstained cell during the 68min spanof observation. It is also noteworthy that a nucleusenwrapped in a small mass of cytoplasm can be translo-cated through the cell process (Fig. 6C-G). Ananucleate fragment sustained locomotive activity(Fig. 6F,G). Although solitary epithelial cells contain-ing stained granules, probably of archenteron origin,were sometimes found to contaminate these cultures,they were easily distinguished by their rounded shapeand the presence of a cilium. Fusion and separationnever took place between the epithelial cells and themesenchyme cells.
The syncytial nature of the mesenchyme cell in thenetwork was further confirmed by TEM. Fig. 7 shows atypical region of the network where two cell bodies areconnected by a cell process. No cell boundary isdetectable between the two nuclei in such areas. The
lack of cell boundaries is confirmed in a wider area bySEM(Figs8, 9).
Three-dimensional structure of the mesenchymalnetworkAnother characteristic of the network revealed ultra-structurally is its three-dimensional nature. A typicalexample is shown by SEM in an area where the cellbodies are crowded (Fig. 8). Many overlapping sites ofcell bodies and cell processes are seen. In order todetermine whether or not the three-dimensional con-figuration of the network depends on the degree ofcrowding of the cells, we prepared networks of variouscondensations by changing the density of the archen-teron complexes. Unexpectedly, the three-dimensionalconfiguration depended on the density of the initialarchenteron complexes rather than the degree ofcrowding of the cells themselves. That is, even when thecrowding of the cells is similar (Fig. 9), a greaternumber of overlapping sites are present in a networkresulting from the culture of a larger number of thearchenteron complexes (Fig. 9B, 33 overlapping sites,more ECM), as compared to that obtained from asmaller number of them (Fig. 9A, 5 overlapping sites,less ECM).

Acellularity of embryonic mesenchyme cells 133
Fig. 4. Dynamics of the network pattern. Phase-contrast micrographs show the same area of the network (24 h in culture).(A) Omin; (B) 8min; (C) 13min; (D) 16min; (E) 20min; and (F) 28min after the beginning of observation. Cell bodieschange their position constantly. Arrows and arrowheads indicate positions where new cell processes are formed in twodifferent ways (see Text). Bar, 50 ^m.
Network formation in vivoIn normal embryos, the mesenchyme cells are relativelyspherical at the mouth formation stage (Fig. 10A),bearing only short or inconspicuous processes(Fig. 10B,C). As the development proceeds, they in-crease in number to approximately 110 at the bipinnariastage (Fig. 10D) (Kominami, 1985) and the cells trans-form to an irregular shape. They extend prominent,branched processes, which spread out into small lamel-lipodia at some places (Fig. 10E,F). In some regions ofthe blastocoel, 2 to 10 cells are seen to form a localnetwork (Fig. 10G,H).
Discussion
When embryonic mesenchyme cells of the starfish arecultured by inoculating archenteron complexes separ-ated from the ectoderm in sea water, they behave in anacellular manner. The mesenchyme cells escape fromthe ECM, form a large syncytium on the substratum,which eventually fragments into smaller and smallerpieces until it becomes a network of small acellularfragments interconnected by fine cell processes after24 h of culture (Fig. 2). The pattern of the networkchanges constantly, driven mainly by the movement of

134 H. Kaneko and others
> • " •
5A BFig. 5. Coculture of cells containing either stained or unstained granules (1.5 h after inoculation). The same optic field isobserved under (A) phase-contrasted and (B) bright-field microscope. Only the stained granules are detectable in B. Twomesenchyme cells (arrows) and an epithelial cell (arrowhead) contain the stained granules. Two cilia are seen to extend fromthe epithelial cell. Bar, 50/«n.
H
Fig. 6. Schematic drawings of the upper three cells shown in Fig. 5. These drawings are based on a series of photographstaken under both phase contrasted and bright field conditions of the same field. (A) Omin, (B) 5min, (C) 8min, (D) 12 min,(E) 13 min, (F) 18 min, (G) 19 min, (H) 27 min, (I) 35 min, (J) 53 min, (K) 61 min and (L) 68 min after the beginning of theobservation (1.5 h in culture). The anucleate fragment shown from (D) to (G) is omitted after (H), since it migrated out ofthe microscopic field. In I-L, only the binucleate fragment shown in H is drawn. Patterned circles: individual nuclei. Dotsand 'x's: stained cytoplasmic granules of two different cells.

Acellularity of embryonic mesenchyme cells 135
Fig. 7. A transmission electron micrograph of two cell bodies connected by a cell processes (24h in culture). No cellboundary is found between the two nuclei, n, nucleus; arrows, undissolved dish surface from propylene oxide treatment,which shows the basal side of the network (Milici et al. 1985). Bar, 10 urn.
Fig. 8. A scanning electronmicrograph showing thethree-dimensional structure ofthe mesenchymal network(24h in culture). Cell bodiesand cell processes areoverlapping one another inmany sites. The specimen istilted by 62°. Bar, 10 pan.
Fig. 9. The mesenchymal network deriving from cultures of different densities of the archenteron complex (24 h in culture):after inoculation of one drop (A) and five drops (B) of the archenteron preparation suspension (see Methods). Areas ofsimilar crowding of the cells are compared. The points of overlap are counted to be 5 in A and 33 in B. The ECM issignificantly richer in (B). Bar, 100/.im.

136 H. Kaneko and others
Fig. 10. Mesenchyme cells in vivo during the normal development of the starfish embryo. Differential interferencemicrographs of embryos and mesenchyme cells at the mouth formation stage (A-C) and the bipinnaria stage (D-H).Mesenchyme cells, which are relatively spherical and bear a few short processes (B, C), become irregular in shape and havecomplexly branched processes, some of which connect the cell bodies (E, F). Bars: (A,D), 100 jxm; (B,C,E-H), 10^m.
the cell bodies (Fig. 4). Cytoplasmic granules andnuclei are seen to translocate between the fragmentsduring this movement (Figs 5, 6). The lack of cellboundaries between the fragments is confirmed byTEM observations (Fig. 7). The highly acellular nature
of these cells, in vitro, is shown most dramatically in theformation and absorption of anucleate fragments at anysite of the network. This event is also easily observed ina sparse culture whose major component is mononu-cleate (Fig. 6G,H). The acellularity of these cells seems

Acellularity of embryonic mesenchyme cells 137
to depend on the highly flexible nature of their plasmamembrane. We know, however, nothing about thismembrane, at present.
The degree of acellularity of these cells is consideredmuch stronger than that of the primary mesenchymecells of the sea urchin. That is, the cell bodies of thelatter are always clearly distinguishable from theirprocesses, as are beads on a string, both in vivo(Akasaka et al. 1980; Gibbons et al. 1969; Okazaki,I960, 1965) and in vitro (Decker et al. 1987). Weconsider that the mesenchyme cells observed by Karpand Solursh (1985) in vitro were secondary not primarymesenchyme cells for the following three reasons. (1)Their cells are morphologically similar to ours ratherthan to the primary mesenchyme cells. (2) Their man-ner of fusion is similar i.e. the cell processes and cellbodies fuse. This mode of fusion is not reported, to ourknowledge, in the primary mesenchyme cells. (3) Bothof these cells form anucleate fragments, which areindistinguishable from fragments containing the nu-cleus (Fig. 3A: arrow), a fact also not reported forprimary mesenchyme cells. So far as we know, the onlyother cell type morphologically similar to ours is thenetwork of cultured cells of the respiratory system ofHolothuria sp. (Rannou, 1971).
Our results show that the network is fundamentallythree-dimensional in nature (Fig. 8) and that the num-ber of crossing sites is likely to depend on the amount ofenvironmental ECM (Fig. 9). Although further exper-imentation is necessary to confirm this point, it suggestsa mechanism by which the ECM supports the appar-ently 'cellular' state of the mesenchyme cells in vivo.The local net-like conformation of mesenchyme cellswas found in vivo not only by us (Fig. 10) but also byChia (1977), supporting the possibility that these cellsactually form a disperse, three-dimensional, acellularnetwork in vivo.
One of the most important questions to arise from thepresent study is what physiological function(s) of thenormal development does the acellularity of the mesen-chyme cells reflect. Assuming that the acellularity is notan artifact due to in vitro conditions, two possibilitiesare considered. (1) It may reflect the manner in whichmesenchyme cells exchange and process positionalinformation in morphogenetic events. For example, themesenchyme cells participating the formation of themouth (Crawford and Abed, 1983) may form a localnetwork that extends over the blastocoel between theectoderm and endoderm of the presumptive mouthregion (cf. Fig. 8 of Crawford and Abed, 1983). Thesituation may also be true for those shown in Fig. 10Band C of this paper. Such a network will not only beefficient for transmitting information between distantcells, but will also stabilize the field of morphogenesis.A physical role, similar to that of connective tissue, isalso suggested for the 'fibrous network' of mesenchymecells (Chia, 1977).
Another possible function is that the mesenchymecells play a defensive role in the larval body. Thisspeculation is based on the reported fact that embryonicmesenchyme cells of the starfish show a phagocytic
activity in vivo (Metchnikoff, 1968; Kominami, 1985),which sometimes results in the formation of a largesyncytium in the blastocoel of normal embryos (Metch-nikoff, 1968). This idea, in turn, allows us to interpretthe formation of large syncytia, at the beginning of theculture, as a reaction of phagocytes against contact witha foreign body: a reaction commonly seen in vertebratemonocytes in variety of inflammatory reactions(Papadimitriou, 1978). However, further biochemicaland comparative studies are necessary to verify thispossibility.
We thank Drs Jun-ichi Taniguchi and Chizuko Koseki ofthe National Cardiovascular Center Research Institute forkindly offering us the use of their differential interferencemicroscope. Staff members of Asamushi Marine BiologicalStation of Tohoku University, of Ushimado Marine Labora-tory of Okayama University and of Noto Marine Laboratoryof Kanazawa University were generous enough to supply thestarfish and to allow us to use their facilities during the courseof this investigation. Thanks are also due to Mr Noriya Miyatafor preparing photographs.
References
AKASAKA, K., AMEMIYA, S. AND TERAYAMA, H. (1980). Scanning
electron microscopical study of the inside of sea urchin embryos(Pseudocentrorus depressus). Effects of aryl-xyloside, tunicamycinand deprivation of sulfate ions. Expl Cell Res. 129, 1-13.
CHIA, F.-S. (1977). Scanning electron microscopic observations ofthe mesenchyme cells in the larvae of starfish Pisaster ochracciis.Ada Zool(stockh.). 58, 45-51.
CRAWFORD, B. J. AND CHIA, F.-S. (1978). Coelomic pouchformation in the starfish Pisaster ochraceus (Echinodermata:Asteroidea). J. Morpli. 157, 99-120.
CRAWFORD, B. J. AND ABED, M. (1983). The role of the basallamina in mouth formation in the embryo of the starfish Pisasterochraceus. J. Morph. 176, 235-246.
CUNHA, G. R. (1984). Prostatic epithelial morphogenesis, growth,and secretory cytodifferentiation are elicited via trophicinfluences from mesenchyme. In Progress in Cancer Research andTherapy, (ed. F. Bresciani), vol. 31, pp. 121-128. New York:Raven Press.
DAN, K. AND OKAZAKI, K. (1956). Cyto-embryological studies ofsea urchins. III. Role of the secondary mesenchyme cells in theformation of the primitive gut in sea urchin larvae. Biol. Bull.mar. biol. lab., Woods Hole 110, 29-42.
DAN-SOHKAWA, M. (1976). A 'normal' development of denudedeggs of the starfish, Astenna pectinifera. Dev. Growth Differ. 18,439-445.
DAN-SOHKAWA, M., TAMURA, G. AND MITSUI, H. (1980).
Mesenchyme cells in starfish development: Effect of tunicamycinon their differentiation, migration and function. Dev. GrowthDiffer. 22, 495-502.
DAN-SOHKAWA, M. & KANEKO, H. (1989). Sorting out ofpresumptive stomach cells of the starfish embryo. Dev. GrowthDiffer. 31, 503-508.
DECKER, G. L., MORRILL, J. B. AND LENNARZ, W. J. (1987).
Characterization of sea urchin primary mesenchyme cells andspicules during biomineralization in vitro. Development 101,297-312.
DHOUAILLY, D. AND SENGEL, P. (1983). Feather forming propertiesof the foot integument in avian embryos. InEpithelial—mesenchymal Interactions in Development (ed. R. H.Sawer & J. F. Fallon), pp. 147-162. New York: Praeger.
GIBBONS, J. R., TILNEY, L. G. AND PORTER, K. R. (1969).
Microtubules in formation and development of the primarymesenchyme in Arbacia punctulata. I. The distribution ofmicrotubules. J. Cell Biol. 41, 201-226.

138 H. Kaneko and others
GOSHIMA, K., KANEKO, H., WAKABAYASHI, S., MASUDA, A. AND
MATSUI, Y. (1984). Beating activity of heterokaryons betweenmyocardial and non-myocardial cells in culture. Expl Cell Res.151, 148-159.
GREENBURG, G. AND HAY, E. D. (1986). Cytodifferentiation andtissue phenotype change during transformation of embryonic lensepithelium to mesenchyme-like cells in vitro. Devi Biol. 115,363-379.
GREENBURG, G. AND HAY, E. D. (1988). Cytoskeleton andthyroglobulin expression change during transformation of thyroidepithelium to mesenchyme-like cells. Development 102, 605-622.
GUSTAFSON, T. AND KINNANDER, H. (1956). Microaquana for time-lapse cinematographic studies of morphogenesis in swimminglarvae and observations on sea urchin gastrulation. Expl CellRes. 11, 36-51.
HAFFEN, K., KEDINGER, M. AND SIMON-ASSMANN, P. (1987).
Mesenchyme-dependent differentiation of epithelial progenitor inthe gut. / . Pediatric Gastro-enterology and Nutrition 6, 14-23.
KANATANI, H. (1969). Induction of spawning and oocyte maturationby 1-methyl-adenine in starfishes. Expl Cell Res. 57, 333-337.
KARP, G. C. AND SOLURSH, M. (1985). In vitro fusion andseparation of sea urchin primary mesenchyme cells. Expl CellRes. 158, 554-557.
KOMINAMI, K. (1985). The role of cell adhesion in thedifferentiation of mesendodermal tissues in the starfish, Asterinapectinifera. Dev. Growth Differ. 27, 679-688.
METCHNIKOFF, E. (1968). Lecture V. In Lectures on the
Comparative Pathology of Inflammation, pp. 56-74. New York:Dover Publications, Inc.
MILICI, A. J., FURIE, M. B. AND CARLEY, W. W. (1985). The
formation of fenestrations and channels by capillary endotheliumin vitro. Proc. natn. Acad. Sa. U.S.A. 82, 6181-6185.
OKAZAKI, K. (1960). Skeleton formation of sea urchin larvae. II.Organic matrix of the spicule. Embryologia. 5, 283-320.
OKAZAKI, K. (1965). Skeleton formation of sea urchin larvae. V.Continuous observation of the process of matrix formation. ExplCell Res. 40, 585-5%.
PAPADIMITRIOU, J. M. (1978). Macrophage fusion in vivo and invitro: a review. In Membrane Fusion (ed. G. Poste and G. L.Nicolson), pp. 181-218. Elsevier/North-Holland: BiochemicalPress.
RANNOU, M. (1971). Cell culture of invertebrates other thanmollusks and arthropods. In Invertebrate Tissue Culture vol. 8(ed. Vago, C ) , pp. 385-410. N.Y. and London: Academic Press.
SPIEGEL, M. AND BURGER, M. M. (1982). Cell adhesion duringgastrulation: a new approach. Expl Cell Res. 139, 377-382.
THIERY, J. P. (1984). Mechanisms of cell migration in thevertebrate embryo. Cell Differ. 15, 1-15.
TOMASEK, J. J., HAY, E. D. AND FUJIWARA, K. (1982). Collagenmodulates cell shape and cytoskeleton of embryonic corneal andfibroma fibroblasts; Distribution of actin, a^actinin, and myosin.Devi Biol. 92, 107-122.
{Accepted 7 February 1990)


















