
Accelerated Reactive Oxygen Scavenging System and Membrane Integrity of Two Panicum Species Varying...
8
ORIGINAL PAPER Accelerated Reactive Oxygen Scavenging System and Membrane Integrity of Two Panicum Species Varying in Salt Tolerance Jitha Bhaskaran • Rajaram Panneerselvam Ó Springer Science+Business Media New York 2013 Abstract Plant exhibits various patterns of survival under salinity and their growth and development depend on their capacity to overcome the stress. Present investigation was focused on the response and regulation of the antioxidant defense system and the level of lipid peroxidation in Panicum miliacium and Panicum sumatrense under salt treatments. NaCl stress was imposed for 20 days after sowing of two Panicum species. The changes in the anti- oxidant enzyme activity like superoxide dismutase, cata- lase, peroxidase, and ascorbate peroxidase and the rate of lipid peroxidation level in terms of malondialdehyde (MDA) were recorded in both Panicum species. A great correlation exists between the antioxidant enzymes and lipid peroxidation. The defense mechanism activated in Panicum species studied was confirmed by the increased antioxidant enzyme activities under progressive NaCl stress. MDA content remained close to control at moderate NaCl concentrations and increased at higher salinities. Although lipid peroxidation increased in both Panicum species under salt stress the percent of increase was low in P. sumatrense indicating its salt-tolerant nature. Another possible conclusion is that improved tolerance to salt stress may be accomplished by increased capacity of antioxida- tive system. Keywords Antioxidant enzymes Á Lipid peroxidation Á Osmotic stress Á Salt tolerance Á Singlet oxygen Introduction In signal transduction pathway, reactive oxygen species (ROS) can be regarded as secondary messengers [1] but over ROS production is harmful to the plant causing oxidative stress leading to oxidation of photosynthetic pigments, membrane lipids, proteins, and nucleic acids [2]. ROS include free radicals such as superoxide anion (O 2 - ), hydroxyl radical (OH - ), as, as well as non-radical molecules like hydrogen peroxide (H 2 O 2 ), singlet oxygen ( 1 O 2 ). Highly unstable ROS is formed as a by-product of stepwise reduc- tion of molecular oxygen by electron transport reaction or by high-energy exposure. When ROS reacts with various bio- molecules, it causes lipid peroxidation and disturbs mem- brane integrity and ultimately causing programmed cell death [3]. One among such abiotic stress is salt stress [4] which causes ROS production within minutes of salt appli- cation [5]. Plant cell is evolved with a cascade of antioxidant sys- tem to withstand the risk caused by the presence of ROS to protect cells from oxidative damage [6]. Antioxidant defense system also keeps a check over the levels of active oxygen species (AOS) [7]. They have the ability to scav- enge and remove ROS within the plants, and it is this difference in plant protection mechanisms that decide their tolerance to stress conditions associated with ROS toxicity. Antioxidant enzyme activity is greatly affected by salinity that caused oxidative stress [8]. There is a strong correla- tion between the antioxidant capacity and NaCl tolerance which has been demonstrated in some plant species [9, 10]. In ROS detoxification cascade, superoxide dismutases (SODs) are the first antioxidant defense enzymes, cata- lyzing the dismutation of superoxide anions by reacting with superoxide radicals (O 2 - ) to produce H 2 O 2 [11]. In the absence of natural scavengers such as catalase (CAT) J. Bhaskaran Á R. Panneerselvam (&) Department of Botany, Annamalai University, Annamalai Nagar, Chidambaram 608002, Tamilnadu, India e-mail: [email protected] 123 Cell Biochem Biophys DOI 10.1007/s12013-013-9576-x
Transcript of Accelerated Reactive Oxygen Scavenging System and Membrane Integrity of Two Panicum Species Varying...

ORIGINAL PAPER
Accelerated Reactive Oxygen Scavenging System and MembraneIntegrity of Two Panicum Species Varying in Salt Tolerance
Jitha Bhaskaran • Rajaram Panneerselvam
� Springer Science+Business Media New York 2013
Abstract Plant exhibits various patterns of survival under
salinity and their growth and development depend on their
capacity to overcome the stress. Present investigation was
focused on the response and regulation of the antioxidant
defense system and the level of lipid peroxidation in
Panicum miliacium and Panicum sumatrense under salt
treatments. NaCl stress was imposed for 20 days after
sowing of two Panicum species. The changes in the anti-
oxidant enzyme activity like superoxide dismutase, cata-
lase, peroxidase, and ascorbate peroxidase and the rate of
lipid peroxidation level in terms of malondialdehyde
(MDA) were recorded in both Panicum species. A great
correlation exists between the antioxidant enzymes and
lipid peroxidation. The defense mechanism activated in
Panicum species studied was confirmed by the increased
antioxidant enzyme activities under progressive NaCl
stress. MDA content remained close to control at moderate
NaCl concentrations and increased at higher salinities.
Although lipid peroxidation increased in both Panicum
species under salt stress the percent of increase was low in
P. sumatrense indicating its salt-tolerant nature. Another
possible conclusion is that improved tolerance to salt stress
may be accomplished by increased capacity of antioxida-
tive system.
Keywords Antioxidant enzymes � Lipid peroxidation �Osmotic stress � Salt tolerance � Singlet oxygen
Introduction
In signal transduction pathway, reactive oxygen species
(ROS) can be regarded as secondary messengers [1] but over
ROS production is harmful to the plant causing oxidative
stress leading to oxidation of photosynthetic pigments,
membrane lipids, proteins, and nucleic acids [2]. ROS
include free radicals such as superoxide anion (O2-),
hydroxyl radical (OH-), as, as well as non-radical molecules
like hydrogen peroxide (H2O2), singlet oxygen (1O2). Highly
unstable ROS is formed as a by-product of stepwise reduc-
tion of molecular oxygen by electron transport reaction or by
high-energy exposure. When ROS reacts with various bio-
molecules, it causes lipid peroxidation and disturbs mem-
brane integrity and ultimately causing programmed cell
death [3]. One among such abiotic stress is salt stress [4]
which causes ROS production within minutes of salt appli-
cation [5].
Plant cell is evolved with a cascade of antioxidant sys-
tem to withstand the risk caused by the presence of ROS to
protect cells from oxidative damage [6]. Antioxidant
defense system also keeps a check over the levels of active
oxygen species (AOS) [7]. They have the ability to scav-
enge and remove ROS within the plants, and it is this
difference in plant protection mechanisms that decide their
tolerance to stress conditions associated with ROS toxicity.
Antioxidant enzyme activity is greatly affected by salinity
that caused oxidative stress [8]. There is a strong correla-
tion between the antioxidant capacity and NaCl tolerance
which has been demonstrated in some plant species [9, 10].
In ROS detoxification cascade, superoxide dismutases
(SODs) are the first antioxidant defense enzymes, cata-
lyzing the dismutation of superoxide anions by reacting
with superoxide radicals (O2-) to produce H2O2 [11]. In
the absence of natural scavengers such as catalase (CAT)
J. Bhaskaran � R. Panneerselvam (&)
Department of Botany, Annamalai University, Annamalai
Nagar, Chidambaram 608002, Tamilnadu, India
e-mail: [email protected]
123
Cell Biochem Biophys
DOI 10.1007/s12013-013-9576-x

and peroxidase (POD), H2O2 accumulates in tissues to high
levels. Ascorbate peroxidases (APXs) play a crucial role in
the detoxification of cellular H2O2, the toxic product of
superoxide dismutation. APX removes H2O2 through the
Halliwell–Asada pathway [12, 13]. In general, it is well
accepted that plants with high levels of activity of the
antioxidant systems, both constitutive and induced, have
greater resistance to oxidative damage. However, data on
the effects of salt stress in roots are scarce [14–17].
Lipid peroxidation requires active uptake of O2 and
produces superoxide radical (O2-). Besides O2, the other
highly reactive chemical species involved that initiated
lipid peroxidation 1O2, OH-, and H2O2 [7]. There is a great
relation between antioxidant enzymes and lipid peroxida-
tion. As the level of antioxidant enzymes increase the level
of lipid peroxidation considerably gets reduced. There are
enough evidences that support the alleviation of oxidative
damage and increased resistance to salinity and other
environmental stresses which are often correlated with an
efficient antioxidative system [18–20].
Several studies have shown that salt-tolerant species
increased their antioxidant enzyme activities and antioxidant
contents in response to salt treatment, whereas salt-sensitive
species fails to exhibit the same [21, 22]. As reported by
Foolad et al. [23], Munns and Tester [24], and Grewal [25] at
the whole plant and cellular levels tolerance to biotic stresses
is very complex. This is partly due to the complexity of
interactions between stress factors and various molecular,
biochemical and physiological phenomena affecting plant
growth and development [4]. There are many findings in the
literature which supports the fact that there is close rela-
tionship between the increased enzymatic oxygen scaveng-
ers and increased tolerance to environmental stresses as in
foxtail millet [26], rice [27], and wheat [28].
Materials and Methods
Two landraces namely Panicum miliacium and Panicum
sumatrense seeds were collected from Kolimalai, of Salem
district, Tamilnadu, India and were identified by Tamil
Nadu Agricultural university, Coimbatore, Tamilnadu,
India. The experiment was laid out in a completely ran-
domized block design (CRBD). Pot cultures and the
treatment procedures were carried out in the month of
June–August (2012) in the Botanical Garden and the bio-
chemical analysis was conducted in Stress Physiology
Laboratory, Department of Botany, Annamalai University,
Tamil Nadu, India. The pots were filled with soil con-
taining mixture of red soil, sand, and farm yard manure at
1:1:1 ratio. Four concentrations of NaCl used for the
treatment were 50, 100, 150, and 200 mM, and 0 mM
served as control. For each treatment five replicates were
maintained. Treatments were imposed on the plant on 5,
10, and 15 DAS (days after sowing). On 20 days after,
sowing samples were collected for further analysis.
Measurement of Antioxidant Enzyme Activities
For extraction of enzymes, fresh samples (0.5 g) were
homogenized with 1.5 cm3 of 100 mM potassium
phosphate buffer solution (pH 7.0) containing 2 mM
phenylmethylsulfonyl fluoride (PMSF), and centrifuged at
14,0009g at 4 �C for 20 min.
SOD (EC 1.15.1.1) activity was assayed as described by
Beauchamp and Fridovich [29]. The reaction mixture
contained 1.17 9 10-6 M riboflavin, 0.1 M methionine,
2 9 10-5 M potassium cyanide (KCN), and 5.6 9 10-5 M
nitroblue tetrazolium salt (NBT) dissolved in 3 ml of
0.05 M sodium phosphate buffer (pH 7.8). 3 ml of the
reaction medium was added to 1 ml of enzyme extract. The
mixtures were illuminated in glass test tubes by two sets of
Philips 40 W fluorescent tubes in a single row. Illumination
was started to initiate the reaction at 30 �C for 1 h, iden-
tical solutions that were kept under dark served as blanks.
The absorbance was read at 560 nm in the spectropho-
tometer against the blank. SOD activity was expressed in
units. One unit (U) is defined as the amount of change in
the absorbance by 0.1 h-1 mg-1 protein.
CAT (EC 1.11.1.6) was measured according to the
method of Chandlee and Scandalios [30] with small modi-
fication. The assay mixture contained 2.6 ml of 50 mM
potassium phosphate buffer (pH 7.0), 0.4 ml of 15 mM
H2O2, and 0.04 ml of enzyme extract. The decomposition of
H2O2 is followed by the decline in absorbance at 240 nm.
The enzyme activity is expressed in U mg-1 protein
(U = 1 mM of H2O2 reduction min-1 mg-1 protein).
Peroxidase (EC 1.11.1.7) activity was determined by the
method of Reddy et al. [31]. 20 % homogenate was pre-
pared in 0.1 M phosphate buffer (pH 6.5). The reaction
mixture contained 3.0 ml of pyrogallol solution and 0.1 ml
of the enzyme extract. To the test cuvette, 0.5 ml of H2O2
was added and mixed. The change in absorbance was
recorded at 430 nm every 30 s up to 3 min. One unit of
peroxidase is defined as the change in absorbance/minute at
430 nm.
APX (EC 1.11.1.11) activity was determined as descri-
bed by Asada and Takahashi [32]. The reaction mixture
(1 ml) contained 50 mM potassium phosphate buffer (pH
7.0), 0.5 mM ascorbic acid, 0.1 mM H2O2, and 200 mM of
enzyme extract. The absorbance was read as decrease at
290 nm against the blank, correction was done for the
low, non-enzymatic oxidation of ascorbic acid by H2O2.
The enzyme activity was expressed in U mg-1 protein
(U = change in 0.1 absorbance min-1 mg-1 protein).
Cell Biochem Biophys
123
Determination of Lipid Peroxidation Malondialdehyde
(MDA)
The end-product of preoxidation of polyunsaturated fatty
acids, i.e., malondialdehyde (MDA) was estimated fol-
lowing the thiobarbituric (TBARS) reaction as described
by Heath and Packer [33]. It is the system most prone to
oxidative stress. One gram of tissue (FW) was homoge-
nized in 5 ml of 0.1 % (w:v) TCA. The homogenate was
centrifuged at 10,0009g for 5 min and 4 ml of 20 % TCA
containing 0.5 % (w:v) TBA was added to 1 ml of the
supernatant. The mixture was heated at 95 �C for 30 min
and then quickly cooled on ice. The contents were centri-
fuged at 10,0009g for 15 min and the absorbance was
measured at 532 nm in spectrophotometer. The concen-
tration of MDA was calculated using a extinction coeffi-
cient of 155 mM/1 cm/1. MDA content expressed as
nmol g-1 FW-1.
Results and Discussion
There were remarkable variations in antioxidant enzyme
activities between the two species with increasing NaCl
concentration. P. sumatrense exhibited increased SOD and
CAT activity when compared to P. miliacium at all levels
of NaCl stress.
Antioxidant enzymes such as SOD, CAT, POD, and
APX showed variations in their activities under NaCl
stress. The present results demonstrate a differential effect
of salt stress according to the duration of salt treatments on
shoot and root of two Panicum species. Root and shoot
exhibited large fluctuations with different salinity levels.
There was a significant (P B 0.05) increase in SOD, CAT,
and APX in shoots and roots of both the species of Pani-
cum. We observed greater activities of antioxidant enzymes
in roots than in shoots which suggest the existence of an
effective scavenging mechanism to remove ROS in roots
because roots are the first organs that come in contact with
salt and are thought to play a critical role in plant salt
tolerance. Similar variations were observed in previous
work carried out such as in pea [34], wheat [35], and
tomato [36]. Recently, Jiang et al. [37] analyzed the pro-
teome of Arabidopsis roots under NaCl stress and showed
that detoxifying enzymes such as APXs, glutathione per-
oxidases, and SODs are up-regulated by salt stress.
In our investigation, we found to a larger extend the
amount of ROS is modulated by SOD activity. The only
enzyme able to dismutate O2- to H2O2 and O2 is SOD.
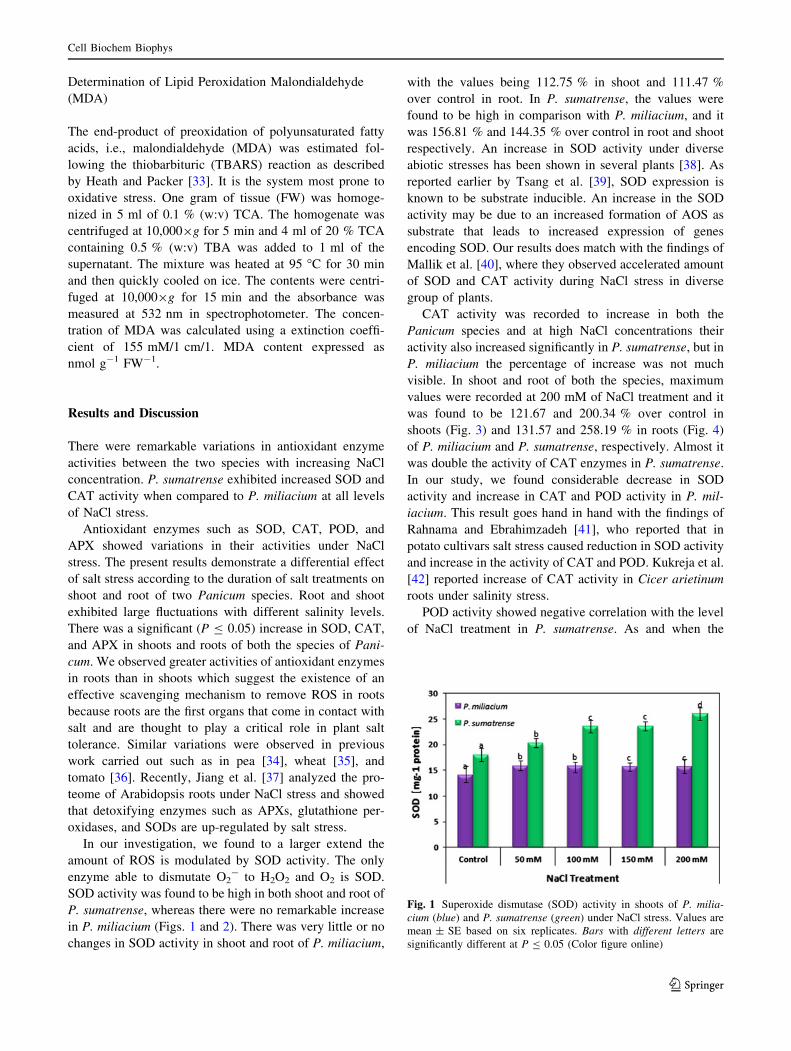
SOD activity was found to be high in both shoot and root of
P. sumatrense, whereas there were no remarkable increase
in P. miliacium (Figs. 1 and 2). There was very little or no
changes in SOD activity in shoot and root of P. miliacium,
with the values being 112.75 % in shoot and 111.47 %
over control in root. In P. sumatrense, the values were
found to be high in comparison with P. miliacium, and it
was 156.81 % and 144.35 % over control in root and shoot
respectively. An increase in SOD activity under diverse
abiotic stresses has been shown in several plants [38]. As
reported earlier by Tsang et al. [39], SOD expression is
known to be substrate inducible. An increase in the SOD
activity may be due to an increased formation of AOS as
substrate that leads to increased expression of genes
encoding SOD. Our results does match with the findings of
Mallik et al. [40], where they observed accelerated amount
of SOD and CAT activity during NaCl stress in diverse
group of plants.
CAT activity was recorded to increase in both the
Panicum species and at high NaCl concentrations their
activity also increased significantly in P. sumatrense, but in
P. miliacium the percentage of increase was not much
visible. In shoot and root of both the species, maximum
values were recorded at 200 mM of NaCl treatment and it
was found to be 121.67 and 200.34 % over control in
shoots (Fig. 3) and 131.57 and 258.19 % in roots (Fig. 4)
of P. miliacium and P. sumatrense, respectively. Almost it
was double the activity of CAT enzymes in P. sumatrense.
In our study, we found considerable decrease in SOD
activity and increase in CAT and POD activity in P. mil-
iacium. This result goes hand in hand with the findings of
Rahnama and Ebrahimzadeh [41], who reported that in
potato cultivars salt stress caused reduction in SOD activity
and increase in the activity of CAT and POD. Kukreja et al.
[42] reported increase of CAT activity in Cicer arietinum
roots under salinity stress.
POD activity showed negative correlation with the level
of NaCl treatment in P. sumatrense. As and when the
Fig. 1 Superoxide dismutase (SOD) activity in shoots of P. milia-cium (blue) and P. sumatrense (green) under NaCl stress. Values are
mean ± SE based on six replicates. Bars with different letters are
significantly different at P B 0.05 (Color figure online)
Cell Biochem Biophys
123
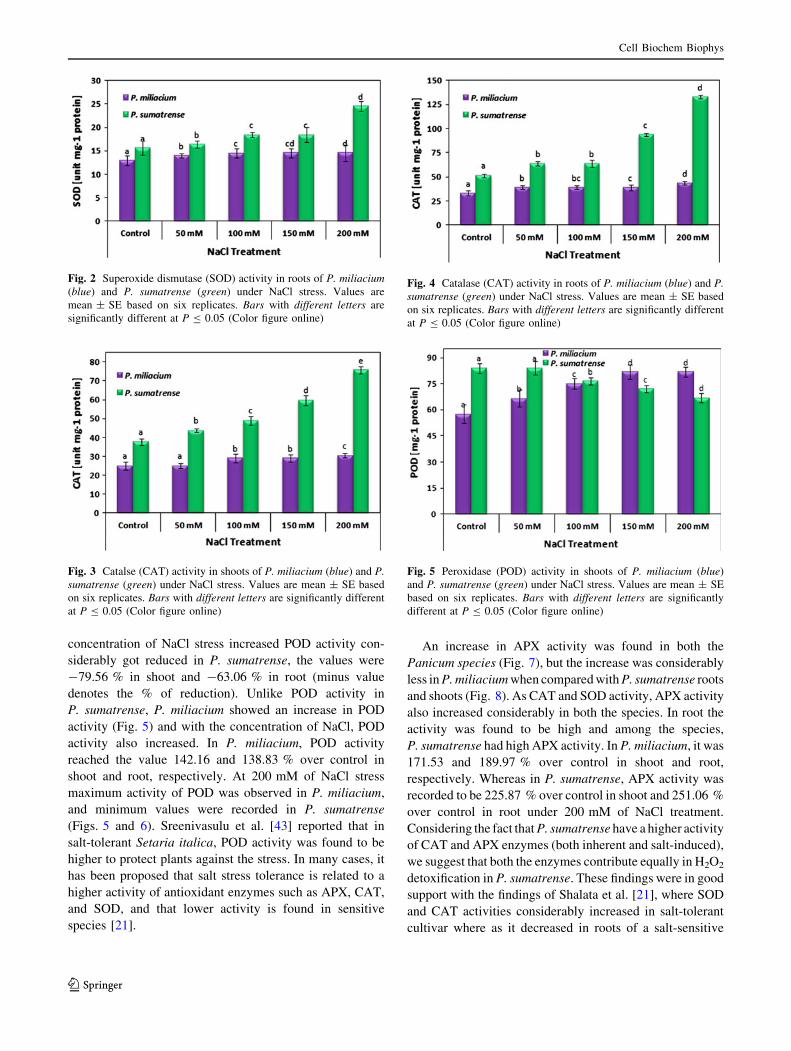
concentration of NaCl stress increased POD activity con-
siderably got reduced in P. sumatrense, the values were
-79.56 % in shoot and -63.06 % in root (minus value
denotes the % of reduction). Unlike POD activity in
P. sumatrense, P. miliacium showed an increase in POD
activity (Fig. 5) and with the concentration of NaCl, POD
activity also increased. In P. miliacium, POD activity
reached the value 142.16 and 138.83 % over control in
shoot and root, respectively. At 200 mM of NaCl stress
maximum activity of POD was observed in P. miliacium,
and minimum values were recorded in P. sumatrense
(Figs. 5 and 6). Sreenivasulu et al. [43] reported that in
salt-tolerant Setaria italica, POD activity was found to be
higher to protect plants against the stress. In many cases, it
has been proposed that salt stress tolerance is related to a
higher activity of antioxidant enzymes such as APX, CAT,
and SOD, and that lower activity is found in sensitive
species [21].
An increase in APX activity was found in both the
Panicum species (Fig. 7), but the increase was considerably
less in P. miliacium when compared with P. sumatrense roots
and shoots (Fig. 8). As CAT and SOD activity, APX activity
also increased considerably in both the species. In root the
activity was found to be high and among the species,
P. sumatrense had high APX activity. In P. miliacium, it was
171.53 and 189.97 % over control in shoot and root,
respectively. Whereas in P. sumatrense, APX activity was
recorded to be 225.87 % over control in shoot and 251.06 %
over control in root under 200 mM of NaCl treatment.
Considering the fact that P. sumatrense have a higher activity
of CAT and APX enzymes (both inherent and salt-induced),
we suggest that both the enzymes contribute equally in H2O2
detoxification in P. sumatrense. These findings were in good
support with the findings of Shalata et al. [21], where SOD
and CAT activities considerably increased in salt-tolerant
cultivar where as it decreased in roots of a salt-sensitive
Fig. 2 Superoxide dismutase (SOD) activity in roots of P. miliacium(blue) and P. sumatrense (green) under NaCl stress. Values are
mean ± SE based on six replicates. Bars with different letters are
significantly different at P B 0.05 (Color figure online)
Fig. 3 Catalse (CAT) activity in shoots of P. miliacium (blue) and P.sumatrense (green) under NaCl stress. Values are mean ± SE based
on six replicates. Bars with different letters are significantly different
at P B 0.05 (Color figure online)
Fig. 4 Catalase (CAT) activity in roots of P. miliacium (blue) and P.sumatrense (green) under NaCl stress. Values are mean ± SE based
on six replicates. Bars with different letters are significantly different
at P B 0.05 (Color figure online)
Fig. 5 Peroxidase (POD) activity in shoots of P. miliacium (blue)
and P. sumatrense (green) under NaCl stress. Values are mean ± SE
based on six replicates. Bars with different letters are significantly
different at P B 0.05 (Color figure online)
Cell Biochem Biophys
123
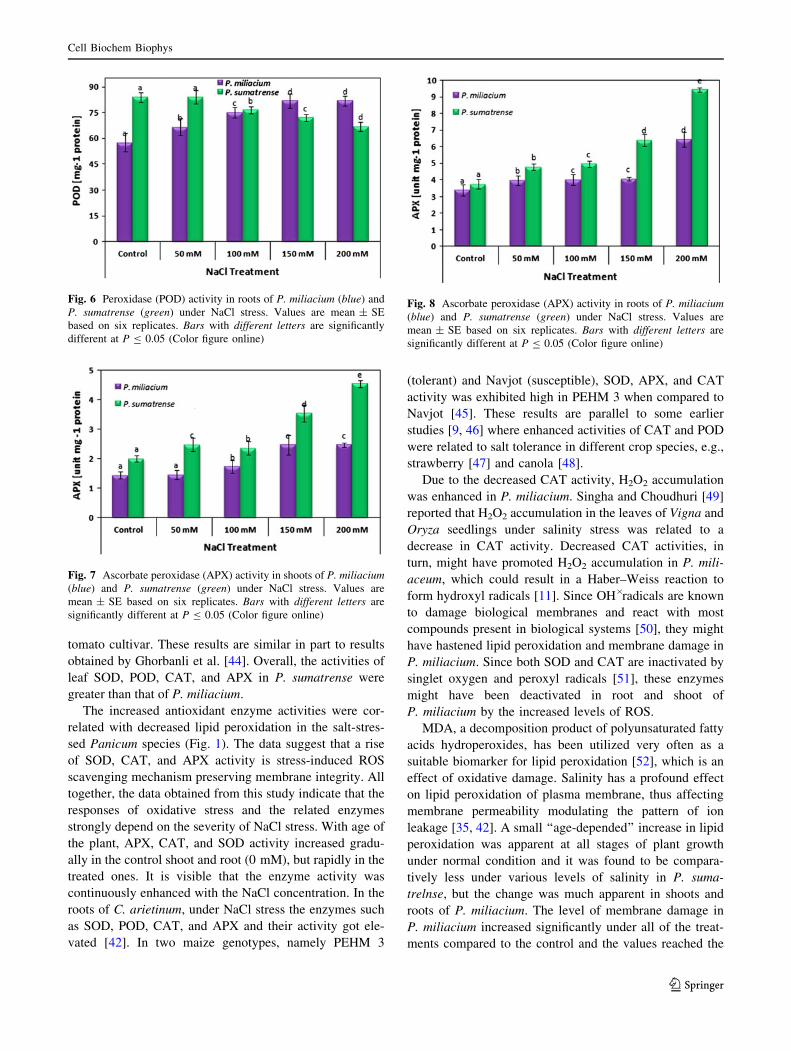
tomato cultivar. These results are similar in part to results
obtained by Ghorbanli et al. [44]. Overall, the activities of
leaf SOD, POD, CAT, and APX in P. sumatrense were
greater than that of P. miliacium.
The increased antioxidant enzyme activities were cor-
related with decreased lipid peroxidation in the salt-stres-
sed Panicum species (Fig. 1). The data suggest that a rise
of SOD, CAT, and APX activity is stress-induced ROS
scavenging mechanism preserving membrane integrity. All
together, the data obtained from this study indicate that the
responses of oxidative stress and the related enzymes
strongly depend on the severity of NaCl stress. With age of
the plant, APX, CAT, and SOD activity increased gradu-
ally in the control shoot and root (0 mM), but rapidly in the
treated ones. It is visible that the enzyme activity was
continuously enhanced with the NaCl concentration. In the
roots of C. arietinum, under NaCl stress the enzymes such
as SOD, POD, CAT, and APX and their activity got ele-
vated [42]. In two maize genotypes, namely PEHM 3
(tolerant) and Navjot (susceptible), SOD, APX, and CAT
activity was exhibited high in PEHM 3 when compared to
Navjot [45]. These results are parallel to some earlier
studies [9, 46] where enhanced activities of CAT and POD
were related to salt tolerance in different crop species, e.g.,
strawberry [47] and canola [48].
Due to the decreased CAT activity, H2O2 accumulation
was enhanced in P. miliacium. Singha and Choudhuri [49]
reported that H2O2 accumulation in the leaves of Vigna and
Oryza seedlings under salinity stress was related to a
decrease in CAT activity. Decreased CAT activities, in
turn, might have promoted H2O2 accumulation in P. mili-
aceum, which could result in a Haber–Weiss reaction to
form hydroxyl radicals [11]. Since OH� radicals are known
to damage biological membranes and react with most
compounds present in biological systems [50], they might
have hastened lipid peroxidation and membrane damage in
P. miliacium. Since both SOD and CAT are inactivated by
singlet oxygen and peroxyl radicals [51], these enzymes
might have been deactivated in root and shoot of
P. miliacium by the increased levels of ROS.
MDA, a decomposition product of polyunsaturated fatty
acids hydroperoxides, has been utilized very often as a
suitable biomarker for lipid peroxidation [52], which is an
effect of oxidative damage. Salinity has a profound effect
on lipid peroxidation of plasma membrane, thus affecting
membrane permeability modulating the pattern of ion
leakage [35, 42]. A small ‘‘age-depended’’ increase in lipid
peroxidation was apparent at all stages of plant growth
under normal condition and it was found to be compara-
tively less under various levels of salinity in P. suma-
trelnse, but the change was much apparent in shoots and
roots of P. miliacium. The level of membrane damage in
P. miliacium increased significantly under all of the treat-
ments compared to the control and the values reached the
Fig. 7 Ascorbate peroxidase (APX) activity in shoots of P. miliacium(blue) and P. sumatrense (green) under NaCl stress. Values are
mean ± SE based on six replicates. Bars with different letters are
significantly different at P B 0.05 (Color figure online)
Fig. 8 Ascorbate peroxidase (APX) activity in roots of P. miliacium(blue) and P. sumatrense (green) under NaCl stress. Values are
mean ± SE based on six replicates. Bars with different letters are
significantly different at P B 0.05 (Color figure online)
Fig. 6 Peroxidase (POD) activity in roots of P. miliacium (blue) and
P. sumatrense (green) under NaCl stress. Values are mean ± SE
based on six replicates. Bars with different letters are significantly
different at P B 0.05 (Color figure online)
Cell Biochem Biophys
123

peak at 150 and 200 mM of NaCl concentration (Table 1).
On a whole, contrast to P. miliacium, shoots and roots of
P. sumatrense showed no much significant change in lipid
peroxidation with increased NaCl treatments.
At 200 mM of NaCl treatment, maximum of MDA
content was noted in both shoot and root of both Panicum
species. Among them, root displayed more MDA content
when compared to leaf. In roots of P. miliacium the values
were 215.94 % over control and in P. sumatrense it was
found to be 184.31 % over control. The increase was steep
in P. miliacium, whereas MDA content increased slowly in
P. sumatrense. The data of the present investigation shows
the increase in MDA content is in positive correlation with
the increase in concentration of NaCl which is in terms
with the previous findings of Aghaleh et al. [53]. This
depicts the membrane damage due to the peroxidation of
lipids which enhances ROS production leading to oxidative
stress. These finding coincides with the previous reports,
where salinity causes drastic lipid peroxidation in wheat
[54] and sugarcane [55]. The lower level of lipid peroxi-
dation in leaves and roots of P. sumatrense suggests that,
these cultivars are better protected from oxidative damage
under drought stress than P. miliacium. Valentovic et al.
[56] showed salinity increase MAD content in corn salt-
sensitive cultivar but in tolerant cultivar, MAD remained
unchanged. These finding coincides with the previous
reports, where salinity causes drastic lipid peroxidation in
wheat [54] and sugarcane [55].
Our finding highlights the relevance of antioxidant
system to protect the cell against the oxidative damage.
P. sumatrense had much lower lipid peroxidation (MDA)
and higher antioxidant enzyme activity when compared to
P. miliaceum. From the data obtained we can say that a
significant elevation in antioxidant enzymes (SOD, CAT,
POD, and APX) occurred and this was inversely related
with the level of lipid peroxidation. Furthermore, the
degree of increase of antioxidant enzymes were more and
the level of lipid peroxidation was relatively less and this
relation between antioxidant enzymes and lipid peroxida-
tion were more visible in P. sumatrense when compared to
P. miliacium. Therefore, P. sumatrense can be considered
as better tolerant species than P. miliacium. Our findings
are in concordance with the findings of Demiral and Tur-
kan [22] and Hernandez et al. [57].
Conclusion
In view of the results observed in this work, the enzymatic
antioxidative system in two Panicum species was highly
affected by salinity. Salt-stressed Panicum species exhib-
ited specific responses of increased SOD, CAT, POD, and
APX activities, accompanied by decreased lipid peroxida-
tion and their responses vary between species. The signif-
icant increase in the activities of SOD, CAT, POD, and
APX in NaCl-stressed Panicum species to some extend
could overcome the damage caused to the membranes due
to lipid peroxidation in terms of MDA and these two fac-
tors were highly correlated with the regulation and main-
tenance of the normal functioning of the cell under salinity
stress. It is clear that the application of saline irrigation
resulted in increase in membrane injury (MDA content) in
P. miliacium and the level of increase was less in P. su-
matrense. Together with a relative importance of CAT,
POD, and APX in H2O2 detoxification, the results suggest
that significance of tissue- and age-specific activities of
antioxidant enzymes are responsible in determining the
membrane integrity, which is in turn dependent on the
species, the development and the metabolic state of plant as
well as the duration and intensity of stress.
However, a direct correlation cannot always be found
between salt stress tolerance and the induction of antioxi-
dant enzyme as proved by Munns and Tester [24] in
transgenic plants where overexpressing these enzymes did
not always induce salt tolerance. Hence, in view of con-
siderable variations in the protective mechanisms against
ROS in different plant species or cultivar, further work is
needed to establish the general validity of this phenomenon
in salt tolerance.
Table 1 NaCl effect on lipid peroxidation in terms of malonyladehyde (MDA) in two Panicum species
NaCl treatments P. miliacium P. sumatrense
Shoot Root Shoot Root
Control 2.44 ± 0.04a 2.76 ± 0.21a 1.43 ± 0.43a 2.04 ± 0.16a
50 mM 3.75 ± 0.14b (153.69) 4.33 ± 0.33b (156.88) 1.44 ± 0.03a (100.70) 2.47 ± 0.12b (121.08)
100 mM 3.79 ± 0.43c (155.32) 4.35 ± 0.06b (157.61) 1.52 ± 0.02b (106.29) 2.49 ± 0.09b (122.06)
150 mM 4.32 ± 0.36d (177.05) 4.63 ± 0.09c (167.75) 2.51 ± 0.39c (155.94) 3.35 ± 0.53c (164.21)
200 mm 4.76 ± 0.07e (195.08) 5.96 ± 0.17d (215.94) 2.54 ± 0.61c (177.62) 3.76 ± 0.37d (184.31)
Values are mean ± SE of four replications. The mean with the same letters do not differ statistically by Duncan’s multiple range test (P B 0.05).
Values in parenthesis denote the percent (%) over control
Cell Biochem Biophys
123

Acknowledgments The authors take this opportunity to thank
UGC-BSR for the financial support and the management of
Annamalai university for providing the facility to carry out the
present work.
References
1. Chamnongpol, S., Willekens, H., Moeder, W., Langebartels, C.,
Sandermann, H., Jr, Van Montagu, M., et al. (1998). Defense
activation and enhanced pathogen tolerance induced by H2O2 in
transgenic tobacco. Proceedings of the National Academy ofSciences of the United States of America, 95(10), 5818–5823.
2. Yordanov, I., Velikova, V., & Tsonev, T. (2000). Plant responses
to drought acclimation and stress tolerance. PhotosyntheticaJournal, 38(2), 171–186.
3. Lee, K. P., Kim, C., Landgraf, F., & Apel, K. (2007). EXE
CUTER1- and EXECUTER2-dependent transfer of stress-related
signals from the plastid to the nucleus of Arabidopsis thaliana.
Proceedings of the National Academy of Sciences of the UnitedStates of America, 104, 10270–10275.
4. Zhu, J. K. (2002). Salt and drought stress signal transduction in
plants. Annual Review of Plant Biology, 53, 247–273.
5. Leshem, Y., Melamed-Book, N., Cagnac, O., Ronen, G., Nishri,
Y., Solomon, M., et al. (2006). Suppression of Arabidopsis ves-
icle-SNARE expression inhibited fusion of H2O2-containing
vesicles with tonoplast and increased salt tolerance. Proceedingsof the National Academy of Sciences of the United States ofAmerica, 103, 18008–18013.
6. Mallick, N., & Mohn, F. H. (2000). Reactive oxygen species:
Response of algal cells. Journal of Plant Physiology, 157, 83–93.
7. Mittler, R. (2002). Oxidative stress, antioxidants and stress tol-
erance. Trends in Plant Science, 7, 405–410.
8. Bernardi, R., Nali, C., Ginestri, P., Pugliesi, C., Lorenzini, G., &
Durante, M. (2004). Antioxidant enzyme isoforms on gels in two
poplar clones differing in sensitivity after exposure to ozone.
Biologia Plantarum, 48, 41–48.
9. Gossett, D. R., Millhollon, E. P., & Lucas, M. C. (1994). Anti-
oxidant response to NaCl stress salt-tolerant and salt-sensitive
cultivars of cotton. Crop Science, 34, 706–714.
10. Hernandez, J. A., Campillo, A., Jimenez, A. A., Alarcon, J. J., &
Sevilla, F. (1999). Response of antioxidant systems and leaf
water relations to NaCl stress in pea plants. New Phytolology,141, 241–251.
11. Bowler, C., Montagu, M. V., & Inze, D. (1992). Superoxide
dismutase and stress tolerance. Annual Review of Plant Physiol-ogy and Plant Molecular Biology, 43, 83–116.
12. Foyer, C. H., & Halliwell, B. (1976). The presence of glutathione
and glutathione reductase in chloroplasts: A proposed role in
ascorbic acid metabolism. Planta, 133, 21–25.
13. Halliwell, B. (1987). Oxidative damage, lipid peroxidation, and
antioxidant protection in chloroplasts. Chemistry and Physics ofLipids, 44, 27–340.
14. Bandeoglu, E., Eyidogan, F., Yucel, M., & Oktem, H. A. (2004).
Antioxidant responses of shoots and roots of lentil to NaCl-
salinity stress. Plant Growth Regulation, 42, 69–77.
15. Mittova, V., Guy, M., Tal, M., & Volokita, M. (2004). Salinity
up-regulates the antioxidative system in root mitochondria and
peroxisomes of the wild salt-tolerant tomato species Lycopers-icon pennellii. Journal of Experimental Botany, 55, 1105–1113.
16. Cavalcanti, F. R., Santos-Lima, J. P. M., Ferreira-Silva, S. L.,
Viegas, R. A., & Gomes-Silveira, J. A. (2007). Roots and leaves
display contrasting oxidative response during salt stress and
recovery in cowpea. Journal of Plant Physiology, 164, 591–600.
17. Seckin, B., Sekmen, A. H., & Turkan, I. (2009). An enhancing
effect of exogenous mannitol on the antioxidant enzyme activities
in roots of wheat under salt stress. Journal of Plant GrowthRegulation, 28, 12–20.
18. Acar, O., Turkan, I., & Ozdemir, F. (2001). Superoxide dismutase
and peroxidase activities in drought sensitive and resistant barley
(Hordeum vulgare L.) cultivars. Acta Physiologiae Plantarum,23(3), 351–356.
19. Sato, Y., Murakami, T., Funatsuki, H., Matsuba, S., Saruyama,
H., & Tanida, M. (2001). Heat shock-mediated APX gene
expression and protection against chilling injury in rice seedlings.
Journal of Experimental Botany, 52(354), 145–151.
20. Bor, M., Ozdemir, F., & Turkan, I. (2003). The effect of salt
stress on lipid peroxidation and antioxidants in leaves of sugar
beet Beta vulgaris L. and wild beet Beta maritima L. PlantScience, 164, 77–84.
21. Shalata, A., Mittova, V., Volokita, M., Guy, M., & Tal, M.
(2001). Response of the cultivated tomato and its wild salt-tol-
erant relative Lycopersicon pennellii to salt-dependent oxidative
stress: The root antioxidative system. Physiologia Plantarum,112, 487–494.
22. Demiral, T., & Turkan, I. (2005). Comparative lipid peroxidation,
antioxidant defense systems and proline content in roots of two
rice cultivars differing in salt tolerance. Environmental andExperimental Botany, 53, 247–257.
23. Foolad, M. R., Zhang, L., & Subbiah, P. (2003). Genetics of
drought tolerance during seed germination in tomato: Inheritance
and QTL mapping. Genome, 46, 536–545.
24. Munns, R., & Tester, M. (2008). Mechanisms of salinity toler-
ance. Annual Review of Plant Biology, 59, 651–681.
25. Grewal, H. S. (2010). Response of wheat to subsoil salinity and
temporary water stress at different stages of the reproductive
phase. Plant and Soil, 330, 103–113.
26. Sreenivasulu, N., Grimm, B., Wobus, U., & Weschke, W. (2000).
Differential response of antioxidant compounds to salinity stress
in salt-tolerant and salt-sensitive seedling of foxtail millet
(Setaria italica). Physiologia Plantarum, 109, 435–442.
27. Guo, Z., Ou, W., Lu, S., & Zhong, Q. (2006). Differential
responses of antioxidative system to chilling and drought in four
rice cultivars differing in sensitivity. Plant Physiology and Bio-chemistry, 44, 828–836.
28. Khanna-Chopra, R., & Selote, D. S. (2007). Acclimation to
drought stress generates oxidative stress tolerance in drought-
resistant than susceptible wheat cultivar under field conditions.
Environmental and Experimental Botany, 60, 276–283.
29. Beauchamp, C. O., & Fridovich, I. (1971). Superoxide dismutase:
Improved assays and an assay applicable to acrylamide gels.
Analytical Biochemistry, 44, 276–287.
30. Chandlee, J. M., & Scandalios, J. G. (1984). Analysis of variants
affecting the catalase developmental program in maize scutellum.
Theoretical and Applied Genetics, 69, 71–77.
31. Reddy, K. P., Subhani, S. M., Khan, P. A., & Kumar, K. B.
(1995). Effect of light and benzyl adenine on dark-treated
growing rice leaves. II. Changes in peroxidase activity. Plant andCell Physiology, 24, 987–994.
32. Asada, K., & Takahashi, M. (1987). Production and scavenging
of active oxygen in photosynthesis. In D. J. Kyle, B. Osmond, &
C. J. Arntzen (Eds.), Photoinhibition (pp. 227–287). Amsterdam:
Elsevier.
33. Heath, R. L., & Packer, L. (1968). Photoperoxidation in isolated
chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxi-
dation. Archives in Biochemistry and Biophysics, 125, 189–198.
34. Hernandez, J. A., & Almansa, M. S. (2002). Short-term effects
salt stress on antioxidant systems and leaf water relations of pea
leaves. Physiolgia Plantarum, 115, 251–257.
Cell Biochem Biophys
123

35. Sairam, R. K., & Srivastava, G. C. (2002). Changes in antioxidant
activity in subcellular fractions of tolerant and susceptible wheat
genotypes in response to long-term salt stress. Plant Science, 162,
897–904.
36. Dogan, M., Tıpırdamaz, R., & Demir, Y. (2010). Salt resistance
of tomato species grown in sand culture. Plant Soil Environment,56(11), 499–507.
37. Jiang, Y., Yang, B., Harris, N. S., & Deyholos, M. K. (2007).
Comparative proteomic analysis of NaCl stress-responsive pro-
teins in Arabidopsis roots. Journal of Experimental Botany, 58,
3591–3607.
38. Gill, S. S., & Tuteja, N. (2010). Reactive oxygen species and
antioxidant machinery in abiotic stress tolerance in crop plants.
Plant Physiology and Biochemistry, 48, 909–930.
39. Tsang, E. W. T., Bowler, C., Herouart, D., Van Camp, W., Vil-
larroel, R., Genetello, C., et al. (1991). Differential regulation of
superoxide dismutases in plants exposed to environmental stress.
The Plant Cell, 3, 783–792.
40. Mallik, S., Nayak, M., Sahu, B. B., Panigrahi, A. K., & Shaw, B.
P. (2011). Response of antioxidant enzymes to high NaCl con-
centration in different salt-tolerant plants. Biologia Pantarum,55(1), 191–195.
41. Rahnama, H., & Ebrahimzadeh, H. (2005). The effect of NaCl on
antioxidant enzyme activities in potato seedling. Biologia Plan-tarum, 49(1), 93–97.
42. Kukreja, S., Nandwal, A. S., Kumar, N., Sharma, S. K., Unvi, V.,
& Sharma, P. K. (2005). Plant water status, H2O2 scavenging
enzymes, ethylene evolution and membrane integrity of Cicerarietinum roots as affected by salinity. Biologia Plantarum,49(2), 305–308.
43. Sreenivasulu, N., Ramanjulu, S., Rmachandra-Kini, K., Prakash,
H. S., Shekar-Shetty, H., Savithri, H. S., et al. (1999). Total
peroxidase activity and peroxidase isoforms as modified by salt
stress in two cultivars of fox-tail millet with differential salt
tolerance. Plant Science, 141, 1–9.
44. Ghorbanli, M., Ebrahimzadeh, H., & Sharifi, M. (2004). Effects
of NaCl and mycorrhizal fungi on antioxidative enzymes in
soybean. Biologia Plantarum, 48, 575–581.
45. Kholova, J., Sairam, R. K., Meena, R. C., & Srivastava, G. C.
(2009). Response of maize genotypes to salinity stress in relation
to osmolytes and metal-ions contents, oxidative stress and anti-
oxidant enzymes activity. Biologia Plantarum, 53(2), 249–256.
46. Lopez, F., Vansuyt, G., Casse-Delbart, F., & Fourcroy, P. (1996).
Ascorbate peroxidase activity, not the mRNA level, is enhanced
in salt-stressed Raphanus sativus plants. Physiologia Plantarum,97(1), 13–20.
47. Turhan, E., Gulen, H., & Eris, A. (2008). The activity of anti-
oxidative enzymes in three strawberry cultivars related to salt-
stress tolerance. Acta Physiologiae Plantarum, 30, 201–208.
48. Ashraf, M., & Ali, Q. (2008). Relative membrane permeability
and activities of some antioxidant enzymes as the key determi-
nants of salt tolerance in canola (Brassica napus L.). Environ-mental and Experimental Botany, 63(1–3), 266–273.
49. Singha, S., & Choudhuri, M. A. (1990). Effect of salinity (NaCl)
stress on H2O, metabolism in Vigna and Oryza seedlings. Bio-chemistry and Physiology Pflanz, 186, 69–74.
50. Halliwel, l. B., & Gutteridge, J. M. C. (1989). Free radicals inbiology and medicine (2nd ed.). Oxford: Clarendon Press.
51. Escobar, J. A., Rubio, M. A., & Lissi, E. A. (1996). SOD and
catalase inactivation by singlet oxygen and peroxyl radicals. FreeRadical Biology and Medicine, 20, 285–290.
52. Bailly, C., Benamar, A., Corbineau, F., & Dome, D. (1996).
Changes in malondialdehyde content and in superoxide dismu-
tase, catalase and glutathione reductase activities in sunflower
seed as related to deterioration during accelerated aging. Physi-olgia Plantarum, 97, 104–110.
53. Aghaleh, M., Niknam, V., Ebrahimzadeh, H., & Razavi, K.
(2009). Salt stress effects on growth, pigments, proteins and lipid
peroxidation in Salicornia persica and S. europaea. BiologiaPlantarum, 53(2), 243–248.
54. Vasantha, S., Gururaja Rao, P. N., Venkataramana, S., &
Gomathi, R. (2008). Salinity-induced changes in the antioxidant
response of sugarcane genotypes. Journal of Plant Biology, 35,
115.
55. Devi, S., Angrish, R., Datta, K. S., & Kumar, B. (2008). Anti-
oxidant defense system in wheat seedlings under sodium chloride
stress: An oxidative role of hydrogen peroxide. Indian Journal ofPlant Physiology, 13, 118.
56. Valentovic, P., Luxova, M., Kolarovic, L., & Gasparikova, O.
(2006). Effect of osmotic stress on compatible solutes content,
membrane stability and water relations in two maize cultivars.
Plant Soil Environment, 52(4), 186–191.
57. Hernandez, M., Fernandez-Garcia, N., Diaz-Vivancos, P., &
Olmos, E. (2009). A different role for hydrogen peroxide and the
antioxidative system under short and long salt stress in Brassicaoleracea roots. Journal of Experimental Botany, 61(2), 521–535.
Cell Biochem Biophys
123


















