
A QTL for flowering time in Arabidopsis reveals a …mcclean/plsc731/homework...2001/11/26 ·...
6
letter nature genetics • volume 29 • december 2001 435 A QTL for flowering time in Arabidopsis reveals a novel allele of CRY2 Salah El-Din El-Assal 1 , Carlos Alonso-Blanco 1,2 , Anton J.M. Peeters 1,3 , Vered Raz 1,4 & Maarten Koornneef 1 1 Laboratory of Genetics, Wageningen University, Dreijenlaan 2, 6703 HA Wageningen, The Graduate School–Experimental Plant Sciences, The Netherlands. 2 Centro Nacional de Biotecnologia, Campus Universidad Autonoma, Cantoblanco, 28049 Madrid, Spain. 3 Department of Plant Ecophysiology, Utrecht University, Sorbonnelaan 16, 3584 CA Utrecht, The Netherlands. 4 Laboratory of Molecular Biology, Wageningen University, Dreijenlaan 3, 6703 HA Wageningen, The Netherlands. Correspondence should be addressed to M.K. (e-mail: [email protected]). Variation of flowering time is found in the natural populations of many plant species. The underlying genetic variation, mostly of a quantitative nature, is presumed to reflect adaptations to different environments contributing to reproductive success. Analysis of natural variation for flowering time in Arabidopsis thaliana has identified several quantitative trait loci (QTL) 1 , which have yet to be characterized at the molecular level. A major environmental factor that determines flowering time is photoperiod or day length, the length of the light period, which changes across the year differently with geographical latitude 2 . We identified the EDI locus as a QTL partly account- ing for the difference in flowering response to the photoperiod between two Arabidopsis accessions: the laboratory strain Landsberg erecta (Ler), originating in Northern Europe, and Cvi, collected in the tropical Cape Verde Islands 3 . Positional cloning of the EDI QTL showed it to be a novel allele of CRY2, encoding the blue-light photoreceptor cryptochrome-2 that has previ- ously been shown to promote flowering in long-day (LD) pho- toperiods 4 . We show that the unique EDI flowering phenotype results from a single amino-acid substitution that reduces the light-induced downregulation of CRY2 in plants grown under short photoperiods, leading to early flowering. Plants of the model species A. thaliana flower earlier when grown under LD than under short-day (SD) conditions, although a sub- stantial variation in response to photoperiod can be found among populations 5 . In particular, the flowering response to the photoperiod is reduced in the Arabidopsis accession Cvi com- pared with the laboratory strains Ler and Columbia (Col). Among five QTL accounting for most of the difference in flower- ing time between Cvi and Ler, one locus at the top of chromo- some 1 (ref. 3) is the main one found to be responsible for the reduced day-length sensitivity; this has been named ‘early day- length insensitive’ (EDI). To determine the molecular basis of this natural genetic varia- tion, we isolated the EDI locus using a map-based cloning strat- egy (Fig. 1). As the starting point for this process, we generated a near-isogenic line (NIL45) carrying a 25-cM Cvi genomic region on the top of chromosome 1 in a Ler background 3 . This line flowered earlier than Ler under SD conditions and was practi- cally insensitive to day length 3 . We crossed NIL45 with a cer1 plant in a Ler background and screened the F 2 population for recombination between EDI and six molecular markers spanning the 10-cM interval between the cer1 and phyA loci (Fig. 1a, Methods). This localized EDI to BAC F19P19; fine-mapping with seven additional molecular markers further localized it to a 45-kb region containing 15 open reading frames (ORFs) (Fig. 1b), including CRY2. CRY2 encodes the blue-light photore- ceptor cryptochrome-2, which is known to be involved in the control of flowering time and the perception of day length 4 , and we therefore considered CRY2 to be a candidate gene for EDI. Published online: 26 November 2001, DOI: 10.1038/ng767 Fig. 1 Map-based isolation of the Ara- bidopsis EDI/CRY2 locus. a, Genetic map of chromosome 1 between the cer1 and phyA markers. Map positions are indicated above the map, and the number of recombinants found between the EDI locus and the mark- ers is indicated below it. b, Physical map of the BAC F19P19 containing the EDI locus. The positions of predicted ORFs according to the Arabidopsis Genome Inititative on BAC F19P19 are shown by small boxes. Black boxes rep- resent the ORFs used for the genera- tion of new cleaved amplified polymorphic sequences (CAPS). The number of recombinants found between EDI and the markers is shown below the map. c, Genomic structure of CRY2: a schematic representation of the 4.6-kb region of CRY2 used for complementation assays. The black boxes represent the CRY2 exons, the thick bars in between represent introns and the thin lines depict the 5′ and 3′ untranslated regions. a b c © 2001 Nature Publishing Group http://genetics.nature.com © 2001 Nature Publishing Group http://genetics.nature.com
Transcript of A QTL for flowering time in Arabidopsis reveals a …mcclean/plsc731/homework...2001/11/26 ·...

letter
nature genetics • volume 29 • december 2001 435
A QTL for flowering time in Arabidopsis reveals a novelallele of CRY2
Salah El-Din El-Assal1, Carlos Alonso-Blanco1,2, Anton J.M. Peeters1,3, Vered Raz1,4 & Maarten Koornneef1
1Laboratory of Genetics, Wageningen University, Dreijenlaan 2, 6703 HA Wageningen, The Graduate School–Experimental Plant Sciences, The Netherlands.2Centro Nacional de Biotecnologia, Campus Universidad Autonoma, Cantoblanco, 28049 Madrid, Spain. 3Department of Plant Ecophysiology, UtrechtUniversity, Sorbonnelaan 16, 3584 CA Utrecht, The Netherlands. 4Laboratory of Molecular Biology, Wageningen University, Dreijenlaan 3, 6703 HAWageningen, The Netherlands. Correspondence should be addressed to M.K. (e-mail: [email protected]).
Variation of flowering time is found in the natural populationsof many plant species. The underlying genetic variation, mostlyof a quantitative nature, is presumed to reflect adaptations todifferent environments contributing to reproductive success.Analysis of natural variation for flowering time in Arabidopsisthaliana has identified several quantitative trait loci (QTL)1,which have yet to be characterized at the molecular level. Amajor environmental factor that determines flowering time isphotoperiod or day length, the length of the light period,which changes across the year differently with geographicallatitude2. We identified the EDI locus as a QTL partly account-ing for the difference in flowering response to the photoperiodbetween two Arabidopsis accessions: the laboratory strainLandsberg erecta (Ler), originating in Northern Europe, and Cvi,collected in the tropical Cape Verde Islands3. Positional cloningof the EDI QTL showed it to be a novel allele of CRY2, encodingthe blue-light photoreceptor cryptochrome-2 that has previ-ously been shown to promote flowering in long-day (LD) pho-toperiods4. We show that the unique EDI flowering phenotyperesults from a single amino-acid substitution that reduces thelight-induced downregulation of CRY2 in plants grown undershort photoperiods, leading to early flowering.Plants of the model species A. thaliana flower earlier when grownunder LD than under short-day (SD) conditions, although a sub-stantial variation in response to photoperiod can be found
among populations5. In particular, the flowering response to thephotoperiod is reduced in the Arabidopsis accession Cvi com-pared with the laboratory strains Ler and Columbia (Col).Among five QTL accounting for most of the difference in flower-ing time between Cvi and Ler, one locus at the top of chromo-some 1 (ref. 3) is the main one found to be responsible for thereduced day-length sensitivity; this has been named ‘early day-length insensitive’ (EDI).
To determine the molecular basis of this natural genetic varia-tion, we isolated the EDI locus using a map-based cloning strat-egy (Fig. 1). As the starting point for this process, we generated anear-isogenic line (NIL45) carrying a 25-cM Cvi genomic regionon the top of chromosome 1 in a Ler background3. This lineflowered earlier than Ler under SD conditions and was practi-cally insensitive to day length3. We crossed NIL45 with a cer1plant in a Ler background and screened the F2 population forrecombination between EDI and six molecular markers spanningthe 10-cM interval between the cer1 and phyA loci (Fig. 1a,Methods). This localized EDI to BAC F19P19; fine-mapping with seven additional molecular markers further localized it to a 45-kb region containing 15 open reading frames (ORFs) (Fig. 1b), including CRY2. CRY2 encodes the blue-light photore-ceptor cryptochrome-2, which is known to be involved in thecontrol of flowering time and the perception of day length4, andwe therefore considered CRY2 to be a candidate gene for EDI.
Published online: 26 November 2001, DOI: 10.1038/ng767
Fig. 1 Map-based isolation of the Ara-bidopsis EDI/CRY2 locus. a, Geneticmap of chromosome 1 between thecer1 and phyA markers. Map positionsare indicated above the map, and thenumber of recombinants foundbetween the EDI locus and the mark-ers is indicated below it. b, Physicalmap of the BAC F19P19 containing theEDI locus. The positions of predictedORFs according to the ArabidopsisGenome Inititative on BAC F19P19 areshown by small boxes. Black boxes rep-resent the ORFs used for the genera-tion of new cleaved amplifiedpolymorphic sequences (CAPS). Thenumber of recombinants foundbetween EDI and the markers is shownbelow the map. c, Genomic structureof CRY2: a schematic representation ofthe 4.6-kb region of CRY2 used forcomplementation assays. The blackboxes represent the CRY2 exons, thethick bars in between representintrons and the thin lines depict the 5′and 3′ untranslated regions.
a
b
c
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com

To test this hypothesis, we carried out complementationtests in transgenic plants, and specifically because geneticanalysis had shown that the Cvi allele of EDI is dominant(Methods), we first tested whether a CRY2 transgene fromCvi could affect the flowering of Ler. We therefore isolateda 4.6-kb genomic fragment containing the complete CRY2coding region and 2,260 bp upstream of the translationstart site (Fig. 1c) from Ler and Cvi, and cloned these frag-ments into a binary vector for plant transformation of theLer background. Transgenic Ler lines carrying the CRY2-Cvi construct showed the same early-flowering, day-length-insensitive phenotype as the near-isogenic lineEDI-NIL (Fig. 2), a Ler line derived from NIL45 but con-taining a 7-cM genomic region from Cvi at the top ofchromosome 1, including the CRY2-Cvi allele. However,plants carrying the CRY2-Ler transgene flowered similarlyto the untransformed Ler controls and exhibited normalday-length sensitivity (Fig. 2). These complementationanalyses showed that allelic variation between Ler and Cviin CRY2 accounts for all the variation in flowering behav-ior attributable to the EDI QTL, and we therefore con-cluded that the EDI locus is CRY2.
This was further confirmed by crossing the CRY2-Cvi transgeneinto a cry2 null–allele mutant background (fha1)6. As previouslyreported, cry2-null plants were impaired in their promotion offlowering under LD conditions, whereas the CRY2-Cvi, cry2-homozygous plants showed an early-flowering and day-length-insensitive phenotype (data not shown), indicating that CRY2-Cvicomplements the CRY2 loss-of-function allele.
To determine the molecular basis of the functional differencebetween the CRY2 allele and Cvi, we sequenced the 4.6-kb CRY2genomic fragment from Cvi and Ler (GenBank accession numbersAY057441 and AY057442 for the Cvi and Ler sequences, respec-tively) and compared it with the published Col accession sequence.We identified a total of 12 single-nucleotide polymorphismsbetween Ler and Cvi, 5 being located in noncoding regions, ofwhich 4 were in the promoter and one in the 3′ untranslated
region. The remaining seven nucleotide substitutions were locatedwithin coding regions and were predicted to give rise to fouramino-acid substitutions (Fig. 3).
For the 2 amino acids at positions 127 and 188, the predictedresidue in Cvi differed from that in Ler but was the same as inCol. As Col is responsive to photoperiod in a manner similar toLer (ref. 7), we reasoned that these two differences were notlikely to account for the early-flowering EDI phenotype. TheCvi-specific substitution at position 476 was also consideredunlikely to account for the EDI phenotype because this residue ispoorly conserved across other plant CRY proteins (Fig. 3). Theother Cvi-specific amino-acid difference was the substitution ofvaline (Ler) for methionine (Cvi) at position 367. This valineresidue is highly conserved among the CRY proteins from higher(tomato, rice, mustard) and lower (ferns) plants (Fig. 3), and wetherefore considered this substitution to be the most likely mole-cular basis for the difference in flowering phenotype.
To investigate the functional significance of this V367Msubstitution, we exchanged genomic fragments differing onlyat this position between the Ler and Cvi CRY2 alleles andtransformed the resulting constructs to Ler. The resultingtransgenic plants thus expressed either Ler CRY2 with theV367M substitution, or Cvi CRY2 with the reverse M367Vsubstitution. Lines carrying the CRY2-Ler-367M transgene
Fig. 2 Early flowering and day-length insensitivity of CRY2-Cvi. a, Flow-ering time (measured as the total number of leaves) of the parentallines Ler and EDI-NIL, as well as the Ler transgenic lines carrying theCRY2-Cvi, CRY2-Ler, CRY2-Ler-367M and CRY2-Cvi-367V genomic con-structs (graphically depicted in the upper part), grown under LD pho-toperiod conditions. b, The same, with growth under SD photoperiodconditions. Means (± standard error) of 20 plants per line are shown.Between five and eight independent homozygous Ler transgenic linesper construct are shown. Dark and light shading represent the Ler andCvi genotypes, respectively.
letter
436 nature genetics • volume 29 • december 2001
Fig. 3 Comparison of CRY2 protein sequences. a, Schematic representation ofthe major domains of CRY2 (ref. 36). The amino acids differing between Ler,Cvi and Col accessions are shown above the bar and their corresponding posi-tions below it. b, sequence alignment of the conserved Flavin-BD of CRY pro-teins from Arabidopsis thaliana, Sinapis, tomato, rice and Adiantum carriedout with the CLUSTAL method of MegAlign-DNASTAR software. Identicalamino acids are shaded in black, whereas amino-acid substitutions amongplant species are shown in white. Arrows indicate the two Cvi-specific pre-dicted amino-acid substitutions: V367M and T476I.
a
b
a
b
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com
were day-length insensitive, similar to EDI-NIL plants,whereas CRY2-Cvi-367V transgenic lines showed a photope-riod response indistinguishable from that of Ler (Fig. 2). Theseexperiments show that a single nucleotide change predicted togive rise to a single V→M substitution at position 367 creates adominant gain-of-function allele and is sufficient to explainthe difference in response to photoperiod between Ler and Cviaccounted for by the EDI locus.
We explored the frequency of this natural CRY2 V367M sub-stitution causing early flowering and reduced photoperiodsensitivity among other Arabidopsis accessions with a world-wide distribution. We generated a dCAPS marker8 specificallyto detect this allelic difference and used it to screen 106 acces-sions, mostly early flowering, including 14 collected from sub-tropical regions (below 35° N, from the Canary Islands, India,Japan, Libya and Pakistan), although no accession is as yetavailable from the low latitude of Cvi. We did not find thisallele outside Cvi, showing that it is rare among natural popu-lations and indicating that it might be specific to Cvi. The eco-logical significance of the CRY2-Cvi substitution allele is
therefore unclear, and further studies including lower-latitudeArabidopsis accessions and other plant species are needed toconfirm its uniqueness.
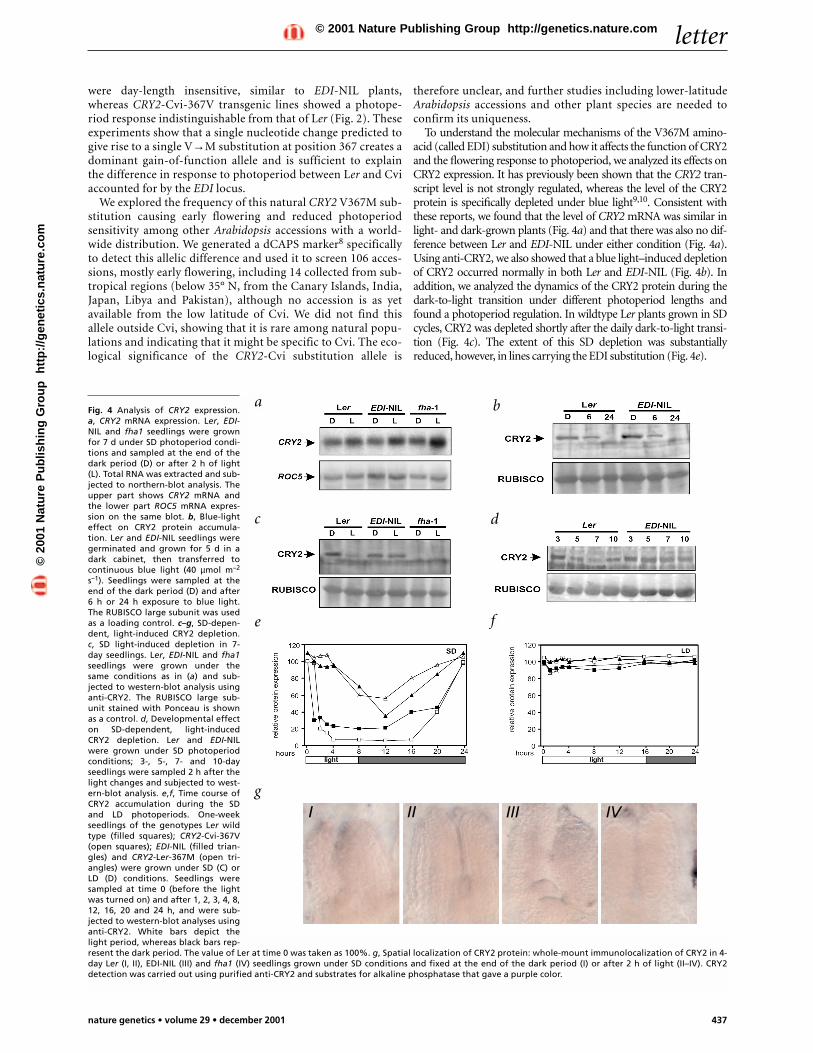
To understand the molecular mechanisms of the V367M amino-acid (called EDI) substitution and how it affects the function of CRY2and the flowering response to photoperiod, we analyzed its effects onCRY2 expression. It has previously been shown that the CRY2 tran-script level is not strongly regulated, whereas the level of the CRY2protein is specifically depleted under blue light9,10. Consistent withthese reports, we found that the level of CRY2 mRNA was similar inlight- and dark-grown plants (Fig. 4a) and that there was also no dif-ference between Ler and EDI-NIL under either condition (Fig. 4a).Using anti-CRY2, we also showed that a blue light–induced depletionof CRY2 occurred normally in both Ler and EDI-NIL (Fig. 4b). Inaddition, we analyzed the dynamics of the CRY2 protein during thedark-to-light transition under different photoperiod lengths andfound a photoperiod regulation. In wildtype Ler plants grown in SDcycles, CRY2 was depleted shortly after the daily dark-to-light transi-tion (Fig. 4c). The extent of this SD depletion was substantiallyreduced, however, in lines carrying the EDI substitution (Fig. 4e).
Fig. 4 Analysis of CRY2 expression.a, CRY2 mRNA expression. Ler, EDI-NIL and fha1 seedlings were grownfor 7 d under SD photoperiod condi-tions and sampled at the end of thedark period (D) or after 2 h of light(L). Total RNA was extracted and sub-jected to northern-blot analysis. Theupper part shows CRY2 mRNA andthe lower part ROC5 mRNA expres-sion on the same blot. b, Blue-lighteffect on CRY2 protein accumula-tion. Ler and EDI-NIL seedlings weregerminated and grown for 5 d in adark cabinet, then transferred tocontinuous blue light (40 µmol m–2
s–1). Seedlings were sampled at theend of the dark period (D) and after6 h or 24 h exposure to blue light.The RUBISCO large subunit was usedas a loading control. c–g, SD-depen-dent, light-induced CRY2 depletion.c, SD light-induced depletion in 7-day seedlings. Ler, EDI-NIL and fha1seedlings were grown under thesame conditions as in (a) and sub-jected to western-blot analysis usinganti-CRY2. The RUBISCO large sub-unit stained with Ponceau is shownas a control. d, Developmental effecton SD-dependent, light-inducedCRY2 depletion. Ler and EDI-NILwere grown under SD photoperiodconditions; 3-, 5-, 7- and 10-dayseedlings were sampled 2 h after thelight changes and subjected to west-ern-blot analysis. e,f, Time course ofCRY2 accumulation during the SDand LD photoperiods. One-weekseedlings of the genotypes Ler wildtype (filled squares); CRY2-Cvi-367V(open squares); EDI-NIL (filled trian-gles) and CRY2-Ler-367M (open tri-angles) were grown under SD (C) orLD (D) conditions. Seedlings weresampled at time 0 (before the lightwas turned on) and after 1, 2, 3, 4, 8,12, 16, 20 and 24 h, and were sub-jected to western-blot analyses usinganti-CRY2. White bars depict thelight period, whereas black bars rep-resent the dark period. The value of Ler at time 0 was taken as 100%. g, Spatial localization of CRY2 protein: whole-mount immunolocalization of CRY2 in 4-day Ler (I, II), EDI-NIL (III) and fha1 (IV) seedlings grown under SD conditions and fixed at the end of the dark period (I) or after 2 h of light (II–IV). CRY2detection was carried out using purified anti-CRY2 and substrates for alkaline phosphatase that gave a purple color.
letter
nature genetics • volume 29 • december 2001 437
a b
c d
e f
g
I II III IV
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com

letter
438 nature genetics • volume 29 • december 2001
We examined this difference in detail by analyzing the pattern ofCRY2 expression throughout a 24-h SD cycle. In lines containingthe Ler CRY2 allele (both Ler itself and Ler expressing CRY2-Cvicontaining the M367V substitution), CRY2 was strongly reducedwithin 2 h after dawn, remaining at a low level before graduallyreaccumulating during the second half of the night (Fig. 4e). Incontrast, lines expressing the CRY2-Cvi substitution showed ahigher level of CRY2 throughout the cycle compared with the levelin the Ler strain. This was evident as a delay in the initial depletionafter lights-on, a reduction in the extent of the depletion and a rapidreaccumulation after lights-off. Notably, under LD conditions, adiurnal variation in the level of CRY2 was not observed in plantscontaining either the Ler or the Cvi CRY2 allele (Fig. 4f). In addi-tion, the amount of CRY2 produced in darkness was comparablebetween LD and SD conditions, and thus the light-induced CRY2downregulation appears to be specific for SD. This result shows thatthe early-flowering phenotype under SD conditions conferred bythe V→M substitution in CRY2-Cvi is associated with a substantialincrease in the amount of CRY2 produced, which is particularlyevident early in the light period.
We confirmed this difference by whole-mount immunolo-calization of CRY2 using a cry2-null mutant as a negative con-trol (Fig. 4g). In Ler seedlings grown under SD conditions andsampled at the end of the dark period, CRY2 was mostly pre-sent in the leaf primordia (Fig. 4g), in agreement with theassumption that leaves are the place of day-length perceptionand the source of a flowering signal11. In Ler seedlings sampled2 h after lights-on, however, the protein was no longerdetectable (Fig. 4g), whereas in EDI-NIL seedlings sampledunder the same conditions (Fig. 4g), CRY2 was present at asimilar level and with a similar distribution to that seen indark-sampled Ler seedlings (Fig. 4g).
We also obtained evidence that the light-induced downregula-tion of CRY2 is developmentally affected. The extent of SD light-induced CRY2 depletion was small in 3-day seedlings butincreased significantly in 4–8-day seedlings before graduallydiminishing in 10–14-day plants (Fig. 4d). This period of sensi-tivity for CRY2 depletion corresponds to the developmentalphase at which the Col accession of Arabidopsis becomes sensitiveto inductive photoperiods or light-quality treatments12,13.
In addition to its effects on flowering, CRY2 has also beenreported to contribute to the regulation of seedling photomor-phogenesis under blue light9. To test whether the EDI substitu-tion also interfered with the photomorphogenic activity ofCRY2, we examined the irradiance dependence of hypocotylelongation in Ler and EDI-NIL seedlings grown under blue light.Ler seedlings showed two phases of response: a weak response atlow irradiance (between 0.01 and 0.8 µmol m–2 s–1) and astronger response at higher irradiance (0.8–6.0 µmol m–2 s–1;Fig. 5). EDI-NIL seedlings were indistinguishable from Lerseedlings in darkness and at the extremes of the irradiance rangeused, but were significantly shorter under intermediate irradi-ances (0.08–4.0 µmol m–2 s–1; Fig. 5). This could represent anextension of the irradiance range for CRY2-Cvi activity towardshigher irradiances in EDI-NIL. It is clear, however, that this effectis much more subtle than the effect of constitutive CRY2 overex-pression, which conferred a substantial increase in sensitivity forblue-light inhibition of hypocotyl elongation across the entireirradiance range9.
Our results have shown that there is a light-induced CRY2downregulation specific for SD conditions. The post-transcrip-tional regulation of CRY2 therefore seems to be more complexthan just a simple light-dependent depletion involving interac-tions with additional photoperiod or circadian clock-regulatedcomponents. The correlation between the abundance of CRY2during the light period and the flowering times observed in thesegenotypes under LD and SD conditions indicates that this pho-toperiod-dependent, light-induced CRY2 regulation might bepart of the mechanism involved in the perception and transduc-tion of the flowering photoperiod length signal. The transcrip-tion of several genes affecting flowering time is now known to besubject to circadian regulation14, indicating the importance ofcircadian rhythms in the regulation of flowering time. The cry2-null mutants have little effect, however, on circadian period15. Inagreement with this observation, we found that the activity of thecircadian-rhythm-regulated reporter gene CAB2:LUC in EDI-NIL did not differ significantly from Ler (data not shown). Inaddition, the circadian rhythm of leaf movement was notaffected in NIL45 (ref. 16). These results show that the effect ofCRY2-Cvi on flowering time does not result from an interferencewith clock function. The photoperiod insensitivity of the cry2mutant and the CRY2-Cvi alleles is more likely to be caused byalterations in CRY2-controlled signaling to floral promoters suchas CONSTANS, which is modulated by the circadian clock17.
In the present work, we have identified a novel allele of CRY2underlying a major-effect QTL involved in the floweringresponse to photoperiod of Arabidopsis. This allele is a uniquevariant not previously isolated in mutational analyses and mayhelp in understanding how plants respond to photoperiodicity.So far, few flowering-time genes accounting for the naturallyoccurring variation have been identified at the molecular level. Inthe main, Arabidopsis FRI and FLC have been shown to beresponsible for much of the flowering natural variation and theresponse to vernalization18,19. In addition, Hd1 and Hd6, whichare major QTL controlling response to photoperiod in rice, werefound to be homologs of the Arabidopsis CONSTANS and CK2α,respectively20,21. Thus, alleles accounting for quantitative naturalvariation provide an alternative and complementary source ofgenetic variation for the functional analysis of genomes.
In contrast to previous plant natural genetic variants thatwere characterized molecularly18–25, the CRY2-Cvi alleledescribed here provides the first example in which a naturalphenotypic variant is demonstrated to be caused by a singleamino-acid substitution altering protein function. In addition,in the accompanying report by Maloof et al.26, a single mutation
Fig. 5 Blue-light effect on hypocotyl elongation. Hypocotyl length was deter-mined under different blue-light fluence rates (0.0 (dark), 0.01, 0.07, 0.43, 2.22,9.48 and 42.5 µmol m–2 s–1) for seedlings of the genotypes Ler wild type (opensquares); EDI-NIL (open diamonds) and fha1 (filled diamonds). Seedlings weregerminated and grown for 5 d under the various blue-light fluence rates;means (± s.e.m.) of 15 seedlings per genotype are given.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com

letter
nature genetics • volume 29 • december 2001 439
in the red/far-red plant photoreceptor phytochrome A of Ara-bidopsis is shown to affect hypocotyl elongation. Photoreceptorsare key regulators controlling various aspects of plant growthand development in response to the environment. These studiesshow that single-nucleotide mutations in their structural genesequences might be recruited by nature to engineer their func-tions while adapting to the environment.
MethodsPlant material and growth conditions. The following previouslydescribed Arabidopsis genotypes were used: Ler and Cvi accessions; andNIL45, which carries about 25 cM of Cvi from the top of chromosome 1introgressed in a Ler genetic background (described as EDI-Cvi in ref. 3and as NIL45 in ref. 16). EDI-NIL was derived from NIL45 and carriedabout 7 cM of Cvi on top of chromosome 1; the fha1 mutant is in a Lerbackground6. Growth conditions under LD and SD photoperiods were aspreviously described27 and flowering time was measured as the number ofleaves6. Blue-light treatments for protein expression analyses28 andhypocotyl elongation29 were carried out as previously documented.
Transgenic plants were generated after the transformation of Ler plantsusing the vacuum transformation procedure30 and the Agrobacterium tume-faciens strain AGLO (ref. 31). The presence of a single locus of the transgenewas shown by the monogenic segregation of kanamycin resistance.
Mapping, cloning and sequence analyses of the EDI/CRY2 locus. A map-ping population was generated from a cross between the cer1-1 mutant (inan Ler background) and NIL45. Among the 1,822 F2 plants, 1,401 exhibit-ed early flowering under the SD photoperiod, indicating monogenic inher-itance and dominance of the Cvi allele.
For mapping, six molecular markers were used between the cer1 and PHYAloci (three SSLPs: F21B7, F20D22, T1G11, and 3 CAPS markers in the PVV4,AXR1 and PHYA genes, which were tested for Ler/Cvi polymorphisms) using575 F2 plants from the genotypes EDI-cer1, edi-CER1 and edi-cer1. Only fourrecombinants were found between EDI and both sides of BAC F19P19 (Gen-Bank accession number AC000104); thus, seven new CAPS markers weregenerated from predicted ORFs in this BAC, which included CRY2. Norecombinants between EDI and the CRY2 marker were found.
A 4.6-kb genomic DNA fragment containing CRY2 was PCR-amplifiedfrom Ler and Cvi using Pfu DNA polymerase (Promega) with CRY2-F1(5′–AGGGTACCAGTGTCTGATGTTAAGAACG–3′) and CRY2-R1 (5′–TGGTCGACCTCAAACTAAACAACTCAGAT–3′) primers. PCR fragmentswere subcloned in pBluescript (Stratagene) and two independent colonieswere sequenced from both strands. These 4.6-kb fragments were clonedinto the KpnI and SalI restriction sites of the binary vector pCAMBIA 2300(CAMBIA, Canberra, Australia) and used for plant transformation.
For the construction of the reciprocal chimeras, containing a single-nucleotide substitution, a 860-bp DNA fragment between amino-acidresidues 207 and 445 was isolated from both Ler and Cvi CRY2 with therestriction enzymes HpaI and BamHI, and reciprocally subcloned intothe Cvi and Ler CRY2. Chimeric constructs were further used for planttransformation.
The presence of the CRY2 transgene was confirmed after PCR ampli-fication using the primers CRY2-F2 (5′–CTGGAGACAAGGCAGGACCGGTTA–3′) and pCAMBIA pC-R (5′–TAACGCCAGGGTTTTCCCAGTCAC–3′). Confirmation of the transgene sequence andthe single-nucleotide change between Ler and Cvi was carried out usingCRY2-F3 (5′–CGTTGTTGAGTCATCTTCG–3′) and pC-R primers forPCR amplification and sequencing.
A dCAPS specific for the mutation causing the V367M transition inCRY2-Cvi was developed based on the method previously described8. Theprimers EDI-F (5′–CGGGGAAATAAGCGTCAGACACGT–3′) and EDI-R(5′–CATTTCCATGGAAGGAGAAGAAACTTCC–3′) were used to amplifya 369-bp PCR fragment, which was cleaved by the BfaI restriction enzymein Cvi but not in Ler.
CRY2 mRNA and protein expression analysis. Total RNA was isolated fromseedlings of the genotypes Ler, EDI-NIL and fha1 using the RNAeasy plantkit (Qiagen) according to the manufacturer’s instructions. Northern-blotanalyses were carried out according to Amersham-Pharmacia protocol using25 µg total RNA. CRY2 mRNA was detected using the full-length CRY2
cDNA as a probe (provided by J.A. Jarillo, INIA, Madrid, Spain). The consti-tutively expressed cyclophylin gene ROC5 (ref. 32) was used as a control.
Antibodies against CRY2 were generated in rabbits against the carboxy-terminal region of CRY2. The 519-bp 3′ end of CRY2 cDNA was subclonedupstream of glutathione S-transferase (GST) in the PGEX-4T-1 vector(Amersham-Pharmacia). The fused protein (CRY2-GST) was purified asdescribed33 and used for raising antibodies in rabbits.
For western-blot analyses, proteins from seedlings of the genotypes Ler,EDI-NIL and fha-1 and of transgenic Ler lines CRY2-Ler-367M and CRY2-Cvi-367V were isolated, with 45 µg per sample being separated onSDS–PAGE gel. Western-blot analyses were carried out according to Amer-sham-Pharmacia protocol, the detection of CRY2 antibodies being carriedwith ECL-plus chemiluminescence (Amersham-Pharmacia). TheRUBISCO large subunit was detected with Ponceau-S (Sigma) staining andwas used as a loading control. Quantification of blots was performed usingIQMac v1.2 software (Molecular Dynamics, Inc.)
The amount of CRY2 protein was normalized to a control protein, whichwas nonspecifically recognized by anti-CRY2 (star symbol in Fig. 4c). Theamount of this unknown protein did not change during treatments and wascorrelated to the amount of the RUBISCO large subunit.
Whole-mount immunohistochemistry. Anti-CRY2 was purified usingCRY2-GST protein34. The whole-mount immunohistochemistry protocolwas modified from the whole-mount in situ hybridization technique35.The specificity of CRY2 immunolocalization was determined using, ascontrols, a pre-immune serum, four different antibodies and the fha1mutant. Briefly, the seedlings were fixed for 30 min in paraformaldehydefixation buffer and dehydrated, the wax layer being removed during incu-bation with xylene. Following rehydration, driselaze treatment (Sigma)was carried out in a vacuum to allow antibody penetration. The detectionof anti-CRY2 was carried out using donkey anti-rabbit alkaline phos-phatase (Promega), which was pre-absorbed onto plant material. Thedetection of the alkaline phosphatase was carried with NBT and BCIP sub-strates (Roche), which gave a purple color to the tissue.
GenBank accession numbers. Cvi-CRY2 mRNA, AY057440. CRY pro-teins: A. thaliana CRY1 (HY4), S66907; A. thaliana CRY2, U43397; Sinapis,X72019; tomato CRY1, AF130423; tomato CRY2, AF130425; rice,AB024337; Adiantum CRY1, AB012626; Adiantum CRY2, AB012627.
AcknowledgmentsWe wish to thank A. Miller for providing transgenic seeds expressing theCAB:LUC constructs, J. Jarillo for providing CRY2 cDNA, W. Soppe for hiscontinuous help and support, J. Weller for his assistance in the blue-lightexperiments and for critical reading of the manuscript, H.H. Offenberg andS. van der Krol for their assistance in developing anti-CRY2 and in theluciferase measurements, respectively, and M. Schreuder and C. Hanhart forhelp with the light cabinets and greenhouse work. This work was supportedby a predoctoral fellowship from the government of Egypt to S.E.-D.E.-A.
Received 21 August; accepted 11 October 2001.
1. Koornneef, M., Alonso-Blanco, C., Peeters, A.J.M. & Soppe, W. Genetic control offlowering time in Arabidopsis. Ann. Rev. Plant Physiol. Plant Mol. Biol. 49,345–370 (1998).
2. Thomas, B. & Vince-Prue, D. Photoperiodism in Plants (Academic Press, New York,1997).
3. Alonso-Blanco, C., El-Assal, S.E-D., Coupland, G. & Koornneef, M. Analysis ofnatural allelic variation at flowering time loci in the Landsberg erecta and CapeVerde Island ecotypes of Arabidopsis thaliana. Genetics 149, 749–764 (1998).
4. Guo, H., Yang, H., Mockler, T.C. & Lin, C. Regulation of flowering time byArabidopsis photoreceptors. Science 279, 1360–1363 (1998).
5. Karlsson, B.H., Sills, G.R. & Nienhuis, J. Effect of photoperiod and vernalization onthe number of leaves at flowering in 32 Arabidopsis thaliana (Brassicaceae)ecotypes. Am. J. Bot. 80, 646–648 (1993).
6. Koornneef, M., Hanhart, C.J. & van der Veen, J.H. A genetic and physiologicalanalysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. 229,57–66 (1991).
7. Jansen, R.C., Van Ooijen, J.W., Stam, P., Lister, C. & Dean, C. Genotype byenvironment interaction in genetic mapping of multiple quantitative trait loci.Theor. Appl. Genet. 91, 33–37 (1995).
8. Neff, M.M., Neff, J.D., Chory, J. & Pepper, E. dCAPS, a simple technique for thegenetic analysis of single nucleotide polymorphisms: experimental applications inArabidopsis thaliana genetics. Plant J. 14, 387–392 (1998).
9. Lin, C. et al. Enhancement of blue-light sensitivity of Arabidopsis seedlings by ablue light receptor cryptochrome 2. Proc. Natl Acad. Sci. USA 95, 2686–2690(1998).
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com

letter
440 nature genetics • volume 29 • december 2001
10. Ahmad, M., Jarillo, J.A. & Cashmore, A.R. Chimeric protein between cry1 and cry2Arabidopsis blue light photoreceptors indicate overlapping functions andvarying protein stability. Plant Cell 10, 197–207 (1998).
11. Bernier, G., Havelange, A., Houssa, C., Petitjean, A. & Lejeune, P. Physiologicalsignals that induce flowering. Plant Cell 5, 1147–1155 (1993).
12. Bradley, D., Ratcliffe, O., Vincent, C., Carpenter, R. & Coen, E. Inflorescencecommitment and architecture in Arabidopsis. Science 275, 80–83 (1997).
13. Mockler, T.C., Guo, H., Yang, H., Duong, H. & Lin, C. Antagonistic action ofArabidopsis cryptochromes and phytochrome B in the regulation of floralinduction. Development 126, 2073–2082 (1999).
14. Mizoguchi, T. & Coupland, G. ZEITLUPE and FKF1: novel connections betweenflowering time and circadian clock control. Trends Plant Sci. 5, 409–411 (2000).
15. Devlin, P.F. & Kay, S.A. Cryptochromes are required for phytochrome signalling tothe circadian clock but not for rhythmicity. Plant Cell 12, 2499–2509 (2000).
16. Swarup, K. et al. Natural allelic variation identifies new genes in the Arabidopsiscircadian system. Plant J. 20, 67–77 (1999).
17. Suarez-Lopez, P. et al. CONSTANS mediates between the circadian clock and thecontrol of flowering in Arabidopsis. Nature 410, 1116–1120 (2001).
18. Johanson, U. et al. Molecular analysis of FRIGIDA, a major determinant of naturalvariation in Arabidopsis flowering time. Science 290, 344–347 (2000).
19. Michaels, S.D. & Amasino, R.M. Flowering Locus C encodes a novel MADS domainprotein that acts as a repressor of flowering. Plant Cell 11, 949–956 (1999).
20. Yano, M. et al. Hd1, a major photoperiod senstivity quantitative trait locus in rice,is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell12, 2473–2483 (2000).
21. Takahashi, Y., Shomura, A., Sasaki, T. & Yano, M. Hd6, a rice quantitative traitlocus involved in photoperiod sensitivity, encodes the α subunit of protein kinaseCK2. Proc. Natl Acad. Sci. USA 98, 7922–7927 (2001).
22. Wang, R.L., Stec, A., Hey, J., Lukens, L. & Doebley, J. The limits of selection duringmaize domestication. Nature 398, 236–239 (1999).
23. Frary, A. et al. Fw2.2: a quantitative trait locus key to the evolution of tomatofruit size. Science 289, 85–88 (2000).
24. Fridman, E., Pleban, T. & Zamir, D. A recombination hotspot delimits a wild-species quantitative trait locus for tomato sugar content to 484 bp within aninvertase gene. Proc. Natl Acad. Sci. USA 97, 4718–4723 (2000).
25. Kliebenstein, D.J., Lambrix, V.M., Reichelt, M., Gershenzon, J. & Mitchell-Olds, T.Gene duplication in the diversification of secondary metabolism: tandem 2-oxoglutarate-dependent dioxygenases control glucosinolate biosynthesis inArabidopsis. Plant Cell 13, 681–693 (2001).
26. Maloof, J.N. et al. Natural variation in light sensitivity of Arabidopsis. NatureGenet. 29, 441–446 (2001).
27. Koornneef, M., Hanhart, C., van Loenen-Martinet, P. & Blankestijn-de Vries, H.The effect of daylength on the transition to flowering in phytochrome-deficient,late-flowering and double mutants of Arabidopsis thaliana. Physiol. Planta 95,260–266 (1995).
28. Van Tuinen, A., Kerckhoffs, L.H.J., Nagatani, A., Kendrick, R.E. & Koornneef, M.Far-red light-insensitive, phytochrome A–deficient mutants of tomato. Mol. Gen.Genet. 246, 133–141 (1995).
29. Peters, J.L., Schreuder, M.E.L., Verduin, S.J.W. & Kendrick, R.E. Physiologicalcharacterization of a high-pigment mutant of tomato. Photochem. Photobiol. 56,75–82 (1992).
30. Bechtold, N., Ellis, J. & Pelletier, G. In planta Agrobacterium mediated genetransfer by infiltration of adult Arabidopsis thaliana plants. C.R. Acad. Sci. 316,1194–1199 (1993).
31. Lazo, G.R., Stein, P.A. & Ludwig, R.A. A DNA transformation–competentArabidopsis genomic library in Agrobacterium. Biotechnology 9, 963–967 (1991).
32. Chou, I.T. & Gasser, C.S. Characterization of the cyclophilin gene family ofArabidopsis thaliana and phylogenetic analysis of known cyclophilin protein.Plant Mol. Biol. 35, 873–892 (1997).
33. Guan, K. & Dixon, J.G. Eukaryotic proteins expressed in Escherichia coli: animproved thrombin cleavage and purification procedure of fusion protein withglutathione S-transferase. Anal. Biochem. 192, 262–267 (1991).
34. Sessa, G., Yang, C.-X., Raz, V., Eyal, Y. & Fluhr, R. Dark induction and subcellularlocalization of the pathogenesis related PRB-1b protein. Plant Mol. Biol. 28,537–547 (1995).
35. Raz, V. & Ecker, J.R. Regulation of differential growth in the apical hook ofArabidopsis. Development 126, 3661–3668 (1999).
36. Cashmore, A.R., Jarillo, J.A., Wu, Y.J. & Liu, D. Cryptochrome: blue light receptorsfor plants and animals. Science 284, 760–765 (1999).
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://g
enet
ics.
nat
ure
.co
m© 2001 Nature Publishing Group http://genetics.nature.com


















