
A PATHWAY OF PLASMA MEMBRANE BIOGENESIS …jcs.biologists.org/content/joces/72/1/89.full.pdf ·...
12
y. Cell Sci. 72, 89-100 (1984) 89 1'rinted in Great Britain © The Company of Biologists Limited 1984 A PATHWAY OF PLASMA MEMBRANE BIOGENESIS BYPASSING THE GOLGI APPARATUS DURING CELL DIVISION IN THE GREEN ALGA CYLINDROCAPSA GEMINELLA HANS J. SLUIMAN Vakgroep Biosystematiek, Biologisch Laboralorium, Vrije Universiteit, P.O. Box 7161, 1007 MC Amsterdam, The Netherlands SUMMARY Cell division in Cylindmcapsageminella, in particular the mode of septum membrane biogenesis, has been studied with the transmission electron microscope. Septum formation takes place in a narrow layer of cytoplasm separating post-mitotic nuclei. First, each daughter nucleus develops a wide cytoplasmic pocket (invagination) containing numerous strands of rough endoplasmic reticulum (ER). Next, a proliferation of rough ER is observed in the equatorial zone of cytoplasm, which invariably contains a small number of widely scattered microtubules. The equatorially aligned cisternae of rough ER produce smooth-membraned vesicles, interpreted as smooth ER, which subsequently coalesce to form the membranous transverse septum. Thus, primary septum formation does not follow any of the two previously known basic cytokinetic patterns in green plants (i.e. plasma membrane furrowing and cell-plate formation), but instead represents a novel type of membrane flow, which effectively bypasses the Golgi apparatus. This pathway of membrane flow has remained largely ignored in current concepts of endomembrane structure and function in eukaryotes. However, it appears to be more widespread than has previously been recognized, especially in autospore-producing green algae and in red algae during the formation of tetraspores. It may represent an evolutionary intermediate type of cell division between the supposedly primitive method of plasma membrane furrowing and the more advanced cell-plate system. INTRODUCTION Ultrastructural knowledge of both the cytokinetic apparatus and theflagellarap- paratus of green plants (Viridiplantae sensu Cavalier-Smith, 1981) obtained during the last two decades has significantly improved our understanding of the fundamental patterns of evolution of this group. It has led to major revisions of long-standing traditional taxonomic concepts of the higher categories, which are largely based on morphological manifestations at the supracellular level. Flagellar features have been found to be useful for identification of major lines of evolution of green plants, while cytokinetic data may help characterize subordinate groups (Mattox & Stewart, 1984; O'Kelly & Floyd, 1984). Up to six basic patterns of mitosis and cytokinesis have been found so far in green plants (Van den Hoek, 1981; Mattox & Stewart, 1984), while the mechanism of septum membrane biogenesis in postmitotic cells is known to involve either Key words: cell division, cell plate, Cylindrocapsa, endoplasmic reticulum, green algae, plasma membrane. CEL72
Transcript of A PATHWAY OF PLASMA MEMBRANE BIOGENESIS …jcs.biologists.org/content/joces/72/1/89.full.pdf ·...

y. Cell Sci. 72, 89-100 (1984) 891'rinted in Great Britain © The Company of Biologists Limited 1984
A PATHWAY OF PLASMA MEMBRANE BIOGENESIS
BYPASSING THE GOLGI APPARATUS DURING CELL
DIVISION IN THE GREEN ALGA CYLINDROCAPSA
GEMINELLA
HANS J. SLUIMANVakgroep Biosystematiek, Biologisch Laboralorium, Vrije Universiteit, P.O. Box 7161,1007 MC Amsterdam, The Netherlands
SUMMARY
Cell division in Cylindmcapsageminella, in particular the mode of septum membrane biogenesis,has been studied with the transmission electron microscope. Septum formation takes place in anarrow layer of cytoplasm separating post-mitotic nuclei. First, each daughter nucleus develops awide cytoplasmic pocket (invagination) containing numerous strands of rough endoplasmicreticulum (ER). Next, a proliferation of rough ER is observed in the equatorial zone of cytoplasm,which invariably contains a small number of widely scattered microtubules. The equatoriallyaligned cisternae of rough ER produce smooth-membraned vesicles, interpreted as smooth ER,which subsequently coalesce to form the membranous transverse septum. Thus, primary septumformation does not follow any of the two previously known basic cytokinetic patterns in green plants(i.e. plasma membrane furrowing and cell-plate formation), but instead represents a novel type ofmembrane flow, which effectively bypasses the Golgi apparatus. This pathway of membrane flowhas remained largely ignored in current concepts of endomembrane structure and function ineukaryotes. However, it appears to be more widespread than has previously been recognized,especially in autospore-producing green algae and in red algae during the formation of tetraspores.It may represent an evolutionary intermediate type of cell division between the supposedly primitivemethod of plasma membrane furrowing and the more advanced cell-plate system.
INTRODUCTION
Ultrastructural knowledge of both the cytokinetic apparatus and the flagellar ap-paratus of green plants (Viridiplantae sensu Cavalier-Smith, 1981) obtained duringthe last two decades has significantly improved our understanding of the fundamentalpatterns of evolution of this group. It has led to major revisions of long-standingtraditional taxonomic concepts of the higher categories, which are largely based onmorphological manifestations at the supracellular level. Flagellar features have beenfound to be useful for identification of major lines of evolution of green plants, whilecytokinetic data may help characterize subordinate groups (Mattox & Stewart, 1984;O'Kelly & Floyd, 1984).
Up to six basic patterns of mitosis and cytokinesis have been found so far in greenplants (Van den Hoek, 1981; Mattox & Stewart, 1984), while the mechanism ofseptum membrane biogenesis in postmitotic cells is known to involve either
Key words: cell division, cell plate, Cylindrocapsa, endoplasmic reticulum, green algae, plasmamembrane.
CEL72

90 HJ.Sluiman
centripetal invagination (furrowing) of the parental plasma membrane (PM), orcentrifugal coalescence of dictyosome-derived cytoplasmic vesicles in a cell plate.Because invagination is so widespread amongst eukaryotes, it is generally consideredmore primitive than the formation of cell plates. Since both invagination and cellplates occur in single, presumably monophyletic lineages like the classesChlorophyceae and Charophyceae (sensu Mattox & Stewart, 1984) one must assumethat cell plates somehow evolved from an ancestral, invagination-type of septumformation. However, the very question of the evolutionary origin of the cell plate,which is fundamental to our understanding of the phylogeny of green plants, hassurprisingly enough received little attention so far.
A recent electron-microscope study of cell division in the filamentous green algaCylindrocapsa geminella has shed some new light upon the possible evolution of thecell plate. Some of the observations, i.e. those on septum formation, will be reportedhere. It will be demonstrated that, contrary to current dogma, a third, hithertounknown pathway of septum biogenesis occurs in green plants (apart from plasmamembrane furrowing and cell-plate formation), which appears to be widespread,especially amongst autospore-producing green algae, and which may be regarded asancestral to the cell-plate system.
MATERIALS AND METHODS
Cylindrocapsa geminella Wolle was isolated from a ditch in the lake district in the NW part of theDutch province of Overijssel. Unialgal cultures were grown in a modified Woods Hole MBL medium(see Francke & Ten Cate, 1980) at 20°C and a 16 h/8 h, light/dark photoregime. Samples of the cul-ture were fixed at room temperature in 2-5 % glutaraldehyde made up in medium (pH 7-2), washedthree times in medium, and postfixed in 1 % aqueous OsO*. After a thorough wash in deionized water,the samples were embedded in 1-5 % agar, dehydrated in acetone, and embedded in Spurr's resin.Sections were stained with uranyl acetate and lead citrate and viewed in a Philips EM300.
RESULTS
C. geminella is a uniseriate filamentous green alga whose roughly cylindrical cellspossess conspicuously layered walls, and an irregularly shaped, parietal chloroplastwith a prominent pyrenoid protruding towards the cell centre (Fig. 1).
In postmitotic cells, daughter nuclei reapproach one another closely, leaving anarrow layer of equatorial cytoplasm in between, in which the transverse septum isto be formed (Fig. 1). Each nucleus is associated with a set of four centrioles, whichhave migrated from the former poles of the mitotic spindle to the centre of theequatorial cytoplasm (not shown).
The initiation of septum formation is marked by two phenomena. First, eachdaughter nucleus develops a wide cytoplasmic pocket facing the equatorial zone,invariably containing numerous strands of rough endoplasmic reticulum (ER) (Fig.2). The nuclear envelope-associated set of centrioles is usually situated at or just below
Fig. 1. Longitudinal section of a filament of C. geminella. Note the prominent pyrenoids(p), layered cell wall, and a postmitotic cell with appressed daughter nuclei (arrows).X3700.

Plasma membrane biogenesis in Cylindrocapsa
Fig, 1

H. J. Sluiman
Fig. 2

Plasma membrane biogenesis in Cylindrocapsa 93
the entrance to the pocket (Fig. 2). Second, a centripetal proliferation of rough ERis observed in the plane of cytokinesis, resulting in a more or less continuous series ofER cisternae across most of the equatorial zone (Figs 2, 3). A few widely scatteredmicrotubules are consistently seen in the vicinity of the ER cisternae, both in the planeof division and inside the nuclear pockets.
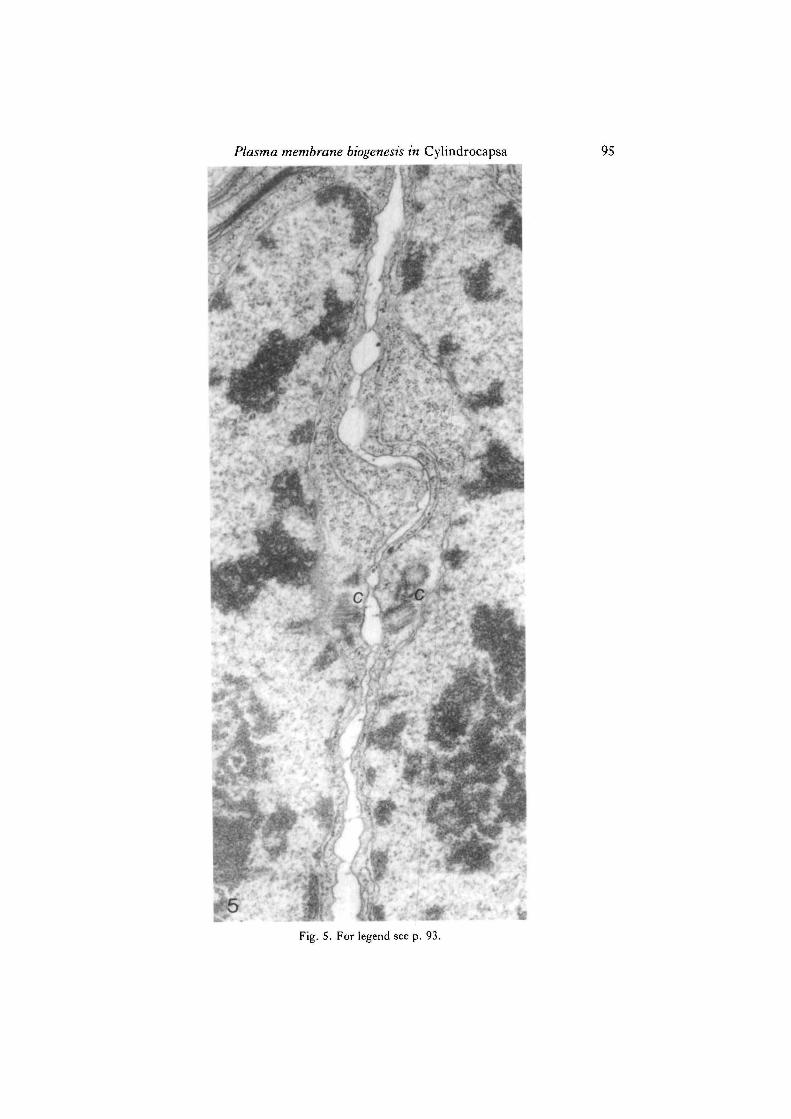
It is the equatorial and 'endonuclear' ER that has been found to play a key role inthe construction of the new membranous septum. First, small smooth-membranedvesicles are seen in the immediate vicinity of the rough ER cisternae (Figs 2, 3), fromwhich they apparently originate. This process can be interpreted as follows. Small,localized areas of the rough ER dissociate themselves from ribosomes and undergodilation (Figs 2, 3). These areas are then separated from the rest of the ER, therebygiving rise to smooth-membraned vesicular elements, which may be referred to mostappropriately as smooth ER vesicles. The latter appear to be the basic ultrastructur-ally identifiable elements involved in early septum ontogenesis, since all availableevidence strongly suggests a process of coalescence of smooth ER cisternae in theplane of division whereby smooth membranes are continuously being added to thegrowing septum (Fig. 4). Theoretically, the alternative possibility of direct participa-tion of the 'parental' rough ER itself should not be ruled out entirely, since localizeddilation of its cisternae in concert with ribosome dissociation may account for theformation of at least portions of the new septum. At any rate, the ER is clearly theexclusive ultrastructurally identifiable source of membrane needed for primary sep-tum formation. The central portion of a just-completed septum is shown in Fig. 5.
There is a sharp distinction between this first, ER-dominated stage of cytokinesiswhereby a membranous septum is formed, and the next stage in which a cell wall issecreted all around each daughter protoplast by exocytosis of Golgi-derived vesiclescontaining wall material (not shown).
DISCUSSION
The cellular mechanism underlying cytokinesis in C. geminella, or, more specific-ally, the formation of transverse septa, involves a direct flow of ER membrane towards
Fig. 2. Early stage of septum initiation. The left-hand daughter nucleus has developed awide cytoplasmic pocket, which contains several strands of rough ER. ER has begun toproliferate in the equatorial region, and smooth membranous ER-derived elements areseen in its vicinity and elsewhere in the plane of cytokinesis (arrows). c, centriole complex.X39000.
Fig. 3. Off-median section of a cell comparable to that shown in Fig. 2, showing anelongated profile of ER with dilated portions and associated smooth-membraned elements(arrows). Several microtubules can be seen in the equatorial plane. X59000.
Fig. 4. This figure shows smooth ER-derived elements (arrows), which coalesce to formthe new septum. Two septum nucleating centres are visible; a third centre is presentelsewhere in the equatorial region (outside the area shown in this micrograph). X65 500.
Fig. 5. A medial longitudinal section of a post-mitotic cell at a later stage than that shownin Fig. 4, showing the central portion of a just-completed septum, and associated ER. c,centriole complex. X35 000.

H, J, Sluiman
Figs 3 and 4. For legend see p. 93.
Plasma membrane biogenesis in Cylindrocapsa 95
Fig. 5. For legend see p. 93.

96 H. J. Sluiman
the plasma membrane (PM). This has not been reported before in any green alga, andits occurrence in other eukaryotic systems is most uncertain. For this reason, thepossibility of direct ER—> PM membrane flow has received little, if any, attention invarious reviews dealing with the structural and functional aspects of the endo-membrane system since the introduction of the 'Endomembrane Concept' by Morre",Mollenhauer & Bracker (1971) (see, e.g., Morr6 & Mollenhauer, 1974; Morr6, Kar-tenbeck & Franke, 1979; Mollenhauer & Morr6, 1980; Morre", 1980).
In green plants (Viridiplantaesewsu Cavalier-Smith, 1981) it is generally acceptedthat cell division involves either centripetal invagination (furrowing) by the parentalPM (as, e.g., in volvocalean and other green flagellates), or coalescence of Golgi-derived vesicles in a cell plate (e.g. in numerous filamentous green algae, and in allarchegoniate land plants) (Pickett-Heaps, 1975; Stewart & Mattox, 1975; Van denHoek, 1981; Mattox & Stewart, 1984). However, the method of septum formationobsen'ed in C. geminella is difficult to reconcile with any of the existing models ofcytokinesis. It is evident that PM furrowing does not occur. On the other hand, the cell-plate model is equally unfit to account for the cytokinetic phenomena observed in thisalga since: (1) although several Golgi bodies are known to be present at the time ofseptum ontogenesis, they are consistently located far away from the plane of division,i.e. at the side of each daughter nucleus facing away from the equatorial plane; (2) thereis no evidence of a selective flow of Golgi-derived vesicles towards the plane of septumontogenesis; (3) dictyosomes do not appear particularly active during early septumformation. Rather, the observed pattern of membrane flow effectively bypasses theGolgi apparatus. It is therefore concluded that primary septum formation in C.geminella represents a cytokinetic mechanism previously unknown in green plants.
How unique is this type of PM biogenesis? In the literature the possibility of directER—* PM membrane traffic has occasionally been raised. Schnepf (1969), for exam-ple, suggested that ER membrane could be transformed into PM when cell walls aresecreted and when cell plates are formed. Conclusive evidence in support of thisassumption, however, is entirely lacking. There seems to be no exception to the rulethat wall materials are transported to the cell surface via dictyosome-derived vesicles,while in the case of cell plates there is increasing evidence that "the vesicles of the plateare derived solely from the dictyosomes" (Hepler, 1982), and that the structural roleof the ER is limited to the formation of plasmodesmata (see Gunning, 1982, for arecent review).
The only available convincing example of ER acting as a source of membrane forPM is contained in a report on cytokinetic events leading to the formation of tetra-spores in the red alga Harveyella (Kugrens & Koslowski, 1981). There are indica-tions, however, that ER-mediated septum formation amongst red algae is morewidespread than is presently recognized: according to published electron micrographsof tetrasporogenesis inErythrocystis (Santisi & De Masi, 1981), Golgi activity is verylow at the time of septum formation, while smooth membranous elements, probablysmooth ER vesicles, aggregate in the plane of division. In another red alga, Palmaria,it has been established that ER-derived 'mucilage sacs' contribute to the formation ofnew septa during tetrasporogenesis (Pueschel, 1979).

Plasma membrane biogenesis in Cylindrocapsa 97
The coincidence of constructive involvement of the ER in septum ontogenesisduring cytokinesis in C. geminella and during tetrasporogenetic cleavage may be sig-nificant : in both instances daughter protoplasts are formed that secrete an extracellularcoat over their entire surface, inside and separate from the pre-existing parental cellwall. This development can be interpreted as the coordinated but temporally and func-tionally descrete action of two different endomembrane components: the ER is theultrastructurally identifiable source of membrane needed for primary septum forma-tion, while the Golgi apparatus is responsible for subsequent wall deposition (cf. Wha-ley & Dauwalder, 1979; Farquhar & Palade, 1981; Robinson & Kristen, 1982).
Although the reasons for the coincidence of active ER participation in primaryseptum formation and circumferential wall deposition are not understood, it may helpto cue other systems in which PM biogenesis during cytokinesis involves a flow ofmembranes bypassing the Golgi apparatus. For example, in autospore-producinggreen algae (Chlorococcales), daughter cells form an entire new wall inside andseparate from the parental wall (as does, incidentally, C. geminella, which is thereforeconsidered chlorococcalean by certain authors). Critical review of published informa-tion on membrane flow patterns in Chlorococcales has led to the conclusion thatgeneralized statements that in these algae cytokinesis is effected by PM furrowing(Stewart & Mattox, 1975; Pickett-Heaps, 1975; Van den Hoek, 1981; Mattox &Stewart, 1984) are oversimplifications and thus misleading. Instead, the presentknowledge of cytokinetic development in C. geminella necessitates a reinterpretationof numerous previous observations on septum ontogenesis in chlorococcalean algae,since available electron micrographs of Kirchneriella (figs 11, 15c, 19, 20a, 20b, 30,31a, 31b of Pickett-Heaps, 1970), Tetraedron (plates 5D, 6A of Pickett-Heaps,1972a), Ankistrodesmus (fig. 4 of Pickett-Heaps, 19726), Chlorella (fig. 3 of Atkinson,Gunning & John, 1972; figs 2, 3, 5, 11 of Wilson, Wanka & Linskens, 1973),Scenedesmus (figs 8.3, 8.4 of Dodge, 1973; figs 6, 9 of Nilshammar & Walles, 1974;fig. 8 of Pickett-Heaps & Staehelin, 1975) and possibly Sorastrum (fig. 11 of March-ant, 1974), Coelastrum (fig. 13 of Chan & Wong, 1975) and Eremosphaera (figs 3-5of Robinson, Sachs & Mayer, 1976) are more readily understood by reference to theC. geminella cytokinetic model instead of the PM cleavage model. Interestinglyenough, it is noted that in some of these reports (Wilson et al. 1973; Nilshammar &Walles, 1974) smooth ER cisternae are apparently mistaken for Golgi vesicles sinceconsistent reference is being made to centrifugally expanding 'cell plates'. The un-expected use of this term is a further indication that division in these algae cannot beexplained in simple terms of PM furrowing. Finally, in species of Tetraselmis, a genusof thecate green flagellates, daughter protoplasts produce new individual thecaewithin the parental theca by exocytosing Golgi-derived vesicles (Manton & Parke,1965; Domozych, Stewart & Mattox, 1981). Hence, it may not be entirely unexpectedthat cytokinesis in T. striata has been reported to involve "fusion of membranousvesicles" (Ricketts & Davey, 1980). A detailed reinvestigation of septum initiationshould confirm their origin from the ER. The latter possibility is not precluded by apublished electron micrograph on cell division in another species, T. subcordiformis(fig. 16 of Stewart, Mattox & Chandler, 1974).

98 H. jf. Sluiman
In order to avoid confusion, it seems appropriate at this point to emphasize that thealga referred to as 'Cylindrocapsa involuta' (strain LB653 from the UTEX culturecollection), whose cell division has been studied with the electron microscope byPickett-Heaps & McDonald (1975), and C. geminella, the subject of the presentinvestigation, are taxonomically unrelated (see Hoffman & Hofmann, 1975; Hoff-man, 1976). It is therefore not surprising to find that in these organisms the patternsof septum biogenesis are entirely different: strain LB653 uses the PM furrowingmechanism like, e.g., Ulva, Ulothrix, Pseudendoclonium and Trichosarcina(Sluiman, Roberts, Stewart & Mattox, 1983), whereas in C. geminella fusion of ER-derived cisternae takes place (present paper). To complicate matters further, in themuch quoted book by Pickett-Heaps (1975, p. 536) strain LB653, elsewhere knownas 'Cylindrocapsa involuta' (Pickett-Heaps & McDonald, 1975), is referred to by thetaxonomically non-existent name of 'Cylindrocapsa brebissonii'.
Of the two cytokinetic categories that were traditionally known to occur in greenplants PM furrowing is generally regarded as the more primitive (Pickett-Heaps,1975; Van den Hoek, 1981). However, the evolutionary origin of the presumablyadvanced cell-plate system is poorly understood. Clearly, it has the advantage (interms of efficiency) that both new PM and extracellular material are deposited at thesame time, due to the outward flow of Golgi-derived vesicles, and that wall precursorsare initially laid down in specific areas only, i.e. at the new septum, instead of beingsecreted 'wastefully' all around each daughter protoplast. Since the Golgi apparatusis part of the structural, ontogenetic and functional continuum known as the endo-membrane system (cf. Morr6 & Mollenhauer, 1974; Morre' el al. 1979), and in viewof the overall pattern of vectorial migration and interconversion of endomembranesin eukaryote cells, one can easily envisage a primitive cell-plate mechanism in whichendomembrane components other than the Golgi apparatus account for the supplyand insertion of new PM. The present study suggests that the ER-dominatedcytokinetic apparatus of C. geminella and certain other algae may exemplify such anancestral cell-plate system.
I thank Professor Dr M. Vroman and Dr G. M. Lokhorst for comments and discussions, and MrA. P. van Beem for technical assistance.
REFERENCES.
ATKINSON, A. W., GUNNING, B. E. S. & JOHN, P. C. L. (1972). Sporopollenin in the cell wall ofChlorella and other algae: infrastructure, chemistry, and incorporation of HC-acetate, studied insynchronous cultures. Planta 107, 1-32.
CAVALIER-SMITH, T. (1981). Eukaryote kingdoms: seven or nine? BioSystems 14, 461-481.CHAN, K. & WONG, S. L. L. (1975). Ultrastructural observations on Coelastrum reticulatum.
Cytologia 40, 663-675.DODGE, J. D. (1973). The Fine Structure of Algal Cells. London, New York: Academic Press.DOMOZYCH, D. S., STEWART, K. D. & MATTOX, K. R. (1981). Development of the cell wall in
Tetraselmis: role of the Golgi apparatus and extracellular wall assembly. J. CellSci. 52, 351-371.FARQUHA*, M. G. & PAUVDE, G. E. (1981). The Golgi apparatus (complex) - (1954-1981) - from
artifact to center stage. J. Cell Biol. 91, 77s-103s.

Plasma membrane biogenesis in Cylindrocapsa 99
FRANCKE, J. A. & TEN CATE, H. J. (1980). Ecotypic differentiation in response to nutritionalfactors in the algal genus Stigeoclonium Kutz. (Chlorophyceae). Br. phycol.J. 15, 343-355.
GUNNING, B. E. S. (1982). The cytokinetic apparatus: its development and spatial regulation. InThe Cytoskeleton in Plant Gmwth and Development (ed. C. W. Lloyd), pp. 229-292. London,New York: Academic Press.
HEPLER, P. K. (1982). Endoplasmic reticulum in the formation of the cell plate and plasmodesmata.Protoplasma 111, 121-133.
HOFFMAN, L. R. (1976). Fine structure of Cylindrocapsa zoospores. Protoplasma 87, 191-219.HOFFMAN, L. R. & HOFMANN, C. S. (1975). Zoospore formation in Cylindrocapsa. Can.J. Bot.
53, 439-451.KUCRENS, P. & KOSLOWSKI, D. J. (1981). Electron microscope studies on a unique cytokinetic
structure in tetrasporocytes of the red alga Harveyella sp. (Cryptonemiales, Choreocolaceae).Protoplasma 108, 197-209.
MANTON, I. & PARKE, M. (1965). Observations on the fine structure of two species of Platymonaswith special reference to flagellar scales and the mode of origin of the theca. J. mar. biol. Ass. U.K.45, 743-754.
MARCHANT, H. J. (1974). Mitosis, cytokinesis and colony formation in the green alga Sorastrum.J. Phycol. 10, 107-120.
MATTOX, K. R. & STEWART, K. D. (1984). Classification of the green algae: a concept based oncomparative cytology. In The Systemalics of the Green Algae (ed. D. E. G. Irvine & D. John).London, New York: Academic Press (in press).
MOLLENHAUER, H. H. & MORRE, D. J. (1980). The Golgi apparatus. In The Biochemistry ofPlants, a Comprehensive Treatise, vol. 1, The Plant Cell (ed. N. E. Tolbert), pp. 438-488. NewYork, London, Toronto, Sydney, San Francisco: Academic Press.
MORRE, D. J. (1980). Flow-differentiation of membranes: pathways and mechanisms. In CellCompartimentation and Metabolic Channelling (ed. L. Nover, F. Lynen & K. Mothes), pp.47-61. Amsterdam, New York, Oxford: Elsevier/North-Holland Biomedical Press, and Jena:VEB Gustav Fischer Verlag.
MORRE, D. J., KARTENBECK, J. & FRANCKE, W. W. (1979). Membrane flow and interconversionsamong endomembranes. Biochim. biophys. Ada 559, 71-152.
MORRE, D. J. & MOLLENHAUER, H. H. (1974). The endomembrane concept: a functional integra-tion of endoplasmic reticulum and Golgi apparatus. In Dynamic Aspects of Plant infrastructure(ed. A. W. Robards), pp. 84-137. London, New York, etc.: McGraw-Hill.
MORRE, D. J., MOLLENHAUER, H. H. & BRACKER, C. E. (1971). Origin and continuity of Golgiapparatus. In Results and Problems in Cell Differentiation, vol. 2, Origin and Continuity of CellOrganelles (ed. J. Reinert&H. Ursprung), pp. 82-126. Berlin, Heidelberg, New York: SpringerVerlag.
NILSHAMMAR, M. & WALLES, B. (1974). Electron microscope studies on cell differentiation insynchronized cultures of the green alga Scenedesmus. Protoplasma 79, 317-332.
O'KELLY, C. J. & FLOYD, G. L. (1984). Flagellar apparatus absolute orientations and the phylogenyof the green algae. Biosystems 16, 227-251.
PICKETT-HEAPS, J. D. (1970). Mitosis and autospore formation in the green alga Kirchneriellalunaris. Protoplasma 70, 325-347.
PICKETT-HEAPS, J. D. (1972a). Cell division in Tetraedron. Ann. Bot. 36, 369-701.PICKETT-HEAPS, J. D. (19726). Variation in mitosis and cytokinesis in plant cells: its significance
in the phylogeny and evolution of ultrastructural systems Cytobios 5, 59-77.PICKETT-HEAPS, J. D. (1975). Green Algae. Structure, Reproduction and Evolution in Selected
Genera. Sunderland: Sinauer Associates.PICKETT-HEAPS, J. D. & MCDONALD, K. L. (1975). Cylindrocapsa: cell division and phylogenetic
affinities. New Phytol. 74, 235-241.PICKETT-HEAPS, J. D. & STAEHELIN, L. A. (1975). The ultrastructure of Scenedesmus
(Chlorophyceae). II. Cell division and colony formation. J. Phycol. 11, 186-202.PUESCHEL, C. M. (1979). Ultrastructure of tetrasporogenesis inPalmariapalmata (Rhodophyta).
jf. Phycol. 15, 409-424.RICKETTS, T. R. & DAVEY, M. R. (1980). Fine structural changes during the cell cycle of
Platymonas striata Butcher. Nova Hedwigia 33, 195-218.

100 H. J. Sluiman
ROBINSON, D. G. & KRISTEN, U. (1982). Membrane flow via the Golgi apparatus of higher plantcells. Int. Rev. Cytol. 77, 89-127.
ROBINSON, D. G., SACHS, H. & MAYER, F. (1976). Cytokinesis in the green alga Eremosphaeraviridis: plasmalemma formation from "open membranes". Planla 129, 75-82.
SANTISI, S. & D E MASI, F. (1981). An electron microscope study on tetrasporogenesis of theparasitic red alga Erythrocystis montagnei (Derb. and Sol.) Silva. Cytobios 31, 163-178.
SCHNEPF, E. (1969). Membranfluss und Membrantransformation. Ber. dt. bot. Ges. 82, 407-413.SLUIMAN, H. J., ROBERTS, K. R., STEWART, K. D. & MATTOX, K. R. (1983). Comparative
cytology and taxonomy of the Ulvophyceae. IV. Mitosis and cytokinesis in Ulothrix(Chlorophyta). Ada bot. need. 32, 257-269.
STEWART, K. D. & MATTOX, K. R. (1975). Comparative cytology, evolution and classification ofthe green algae with some consideration of the origin of other organisms with chlorophylls a andb. Bot. Rev. 41, 104-135.
STEWART, K. D., MATTOX, K. R. & CHANDLER, C. D. (1974). Mitosis and cytokinesis inPlatymonas subcordiformis, a scaly green monad. J. Phycol. 10, 65-79.
VAN DEN HOEK, C. (1981). Chlorophyta: morphology and classification. In The Biology ofSeaweeds, Botanical Monographs, vol. 17 (ed. C. S. Lobban & M. J. Wynne), pp. 86-132.Oxford, London, Edinburgh, Boston, Melbourne: Blackwell Scientific Publications.
WHALEY, W. G. & DAUWALDER, M. (1979). The Golgi apparatus, the plasma membrane, andfunctional integration. Int. Rev. Cytol. 58, 199-245.
WILSON, H. J., WANKA, F. & LINSKENS, H. F. (1973). The relationship between centrioles,microtubules and cell plate initiation in Chlorella pyrenoidosa. Planta 109, 259-267.
(Received 30 March 1984-Accepted 22 May 1984)


















