
A: Nus factors - PNAS · 8660 Thepublication costsofthisarticle weredefrayedinpartbypagecharge...
5
Proc. Nati. Acad. Sci. USA Vol. 91, pp. 8660-8664, August 1994 Biochemistry Control of transcription processivity in phage A: Nus factors strengthen the termination-resistant state of RNA polymerase induced by N antiterminator (gene expression/transcription termination/proteIn-protein interaction/RNA-protein interaction) JOSEPH DEVITO AND Asis DAS* Department of Microbiology, University of Connecticut Health Center, Farmington, CT 06030 Communicated by M. J. Osborn, May 26, 1994 ABSTRACT During transcription of phage A early oper- ons, the N gene product alters host RNA polymerase (RNAP) so that transcription proceeds through multiple stop signals. Here, we reproduce the essence of N activity with purified components in synthetic transcription units that contain A pL promoter and the N-recognition site, nutL, followed by a variety of intrinsic terminators. We show that three host factors (NusA, NusE, and NusG) are essential for N to allow appre- ciable transcription through multiple terminators and that this persistent antitermination is stimulated by a fourth factor, NusB. Remarkably, in the absence of all four factors, N suppresses various terminators placed near the nut site. This basal antitenation activity of N is enhanced by NusA and is diminished by high salt and temperature. We postulate that N interacts with RNAP directly, inducing the termination- resistant state. While NusA facilitates this interaction, the other factors strengthen it sufficiently over time and distance so that RNAP bypasses multiple terminators. The dispensability of NusB for persistent antitermination in vitro, but not in vivo, raises the possibility that NusB performs two functions: it increases the stability of N antitermination complex and also counteracts an inhibitory factor in the cell. The RNA polymerase (RNAP) elongation complex is the target of two kinds of regulatory factors that modulate the elongation-termination decision in transcription (1-3). Ter- mination factors halt transcription at intergenic sites and attenuate downstream gene expression. In contrast, antiter- mination proteins enable RNAP to bypass the stop signals and, therefore, activate transcription of downstream genes. Distinct classes of antiterminators are known in prokaryotes. Some disrupt the termination signals directly or counteract a termination factor (4, 5). Others convert RNAP to the ter- mination-resistant form. The archetype of this latter class of antiterminators is the N gene product of phage A (6-8). The N protein is essential for transcription of the early genes of phage A, which are organized in two large operons (8). RNAP transcribing each operon encounters both factor-dependent and factor-independent terminators that are suppressed by N (9). The suppression of multiple terminators in each operon requires a specific site called nut, located between the promoter and the first terminator (10-12). The nut site acts in the form of RNA (13-15). During transcription, N is thought to bind to a small hairpin component of nut RNA, known as boxB, so that N can capture the RNAP elongation complex and transform RNAP into the termination-resistant form (7, 13-18). Antitermination by N is influenced by multiple protein factors of Escherichia coli (7, 19-30). Three of these factors, NusA, NusB, and NusE (the S10 ribosomal protein), were discovered by the isolation of host mutants that are defective in N antitermination and, hence, fail to support A develop- ment (20-22). The fourth factor, NusG, emerged from the isolation of extragenic suppressors, which permit A growth in a nusA mutant host (28). Further genetic studies indicated that these factors interact with each other, N, and RNAP (19). Indeed, N and all four host factors form a stable transcription complex in vitro (16, 17, 29). Although no evidence for a direct interaction between N and RNAP has been reported, N does bind to NusA (31), and RNAP interacts with NusA, NusG, and S10 proteins (32-34). More- over, S10 can form a complex with NusB (34), and the NusB-S10 complex can bind to a version of the boxA component of nut RNA (35). In spite of these advances, there is little understanding of the fundamental mechanisms by which N, Nus factors, and nut RNA enable RNAP to bypass terminators (1, 2). Here we attempt to dissect the specific roles of N and Nus factors in the transformation of RNAP to the termination-resistant form. MATERIALS AND METHODS Transcription reactions were carried out in two steps as described (13, 26), with minor modifications. In the first step, transcription complexes poised at a specific template location were generated by incubating 33.4 nM RNAP and 33.4 nM template DNA with 100 uM ApU as the primer for initiation of transcription from pL and 2 1LM each ATP, GTP, and CTP and 1-2 ttCi of [a-32P]ATP (1 Ci = 37 GBq) to allow the synthesis of a labeled, 15-nt transcript. The reaction was performed at 300C for 6 min in 12.5 t4 of transcription buffer (20 mM Tris glutamate, pH 8/5 mM magnesium glutamate/ 100 mM potassium glutamate/5% glycerol/i mM dithiothrei- tal). In the second step, transcript elongation proceeded for 5 min upon the addition of an equal volume of prewarmed (300C) transcription buffer that contained the elongation factors as well as 100 ,uM each UTP, GTP, and CTP and 4 mM ATP. The dilution of radioactivity yielded end-labeled tran- scripts from the first round of synthesis and restricted label- ing of transcripts in subsequent rounds. Transcripts were extracted, resolved, and quantitated as described (13). Template fragments of various plasmids, derived from pWW10 with A pL-nutL module followed by A terminator tR', were prepared as described (26). Highly homogeneous preparations of various proteins were used. Soluble forms of NusA, NusB, and N were purified from overproducing strains, as described (24). NusG was purified from the soluble extract of an overproducer strain, described by Sullivan and coworkers (28), by successive chromatography through DEAE-cellulose and phosphocellulose; NusG was monitored Abbreviation: RNAP, RNA polymerase. *To whom reprint requests should be addressed. 8660 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of A: Nus factors - PNAS · 8660 Thepublication costsofthisarticle weredefrayedinpartbypagecharge...

Proc. Nati. Acad. Sci. USAVol. 91, pp. 8660-8664, August 1994Biochemistry
Control of transcription processivity in phage A: Nus factorsstrengthen the termination-resistant state of RNA polymeraseinduced by N antiterminator
(gene expression/transcription termination/proteIn-protein interaction/RNA-protein interaction)
JOSEPH DEVITO AND Asis DAS*Department of Microbiology, University of Connecticut Health Center, Farmington, CT 06030
Communicated by M. J. Osborn, May 26, 1994
ABSTRACT During transcription of phage A early oper-ons, the N gene product alters host RNA polymerase (RNAP)so that transcription proceeds through multiple stop signals.Here, we reproduce the essence of N activity with purifiedcomponents in synthetic transcription units that contain A pLpromoter and the N-recognition site, nutL, followed by avariety of intrinsic terminators. We show that three host factors(NusA, NusE, and NusG) are essential for N to allow appre-ciable transcription through multiple terminators and that thispersistent antitermination is stimulated by a fourth factor,NusB. Remarkably, in the absence of all four factors, Nsuppresses various terminators placed near the nut site. Thisbasal antitenation activity ofN is enhanced by NusA and isdiminished by high salt and temperature. We postulate that Ninteracts with RNAP directly, inducing the termination-resistant state. While NusA facilitates this interaction, the otherfactors strengthen it sufficiently over time and distance so thatRNAP bypasses multiple terminators. The dispensability ofNusB for persistent antitermination in vitro, but not in vivo,raises the possibility that NusB performs two functions: itincreases the stability of N antitermination complex and alsocounteracts an inhibitory factor in the cell.
The RNA polymerase (RNAP) elongation complex is thetarget of two kinds of regulatory factors that modulate theelongation-termination decision in transcription (1-3). Ter-mination factors halt transcription at intergenic sites andattenuate downstream gene expression. In contrast, antiter-mination proteins enable RNAP to bypass the stop signalsand, therefore, activate transcription of downstream genes.Distinct classes of antiterminators are known in prokaryotes.Some disrupt the termination signals directly or counteract atermination factor (4, 5). Others convert RNAP to the ter-mination-resistant form. The archetype of this latter class ofantiterminators is the N gene product of phage A (6-8). TheN protein is essential for transcription of the early genes ofphage A, which are organized in two large operons (8). RNAPtranscribing each operon encounters both factor-dependentand factor-independent terminators that are suppressed by N(9). The suppression of multiple terminators in each operonrequires a specific site called nut, located between thepromoter and the first terminator (10-12). The nut site acts inthe form ofRNA (13-15). During transcription, N is thoughtto bind to a small hairpin component of nut RNA, known asboxB, so that N can capture the RNAP elongation complexand transform RNAP into the termination-resistant form (7,13-18).
Antitermination by N is influenced by multiple proteinfactors of Escherichia coli (7, 19-30). Three of these factors,NusA, NusB, and NusE (the S10 ribosomal protein), were
discovered by the isolation of host mutants that are defectivein N antitermination and, hence, fail to support A develop-ment (20-22). The fourth factor, NusG, emerged from theisolation ofextragenic suppressors, which permit A growth ina nusA mutant host (28). Further genetic studies indicatedthat these factors interact with each other, N, and RNAP(19). Indeed, N and all four host factors form a stabletranscription complex in vitro (16, 17, 29). Although noevidence for a direct interaction between N and RNAP hasbeen reported, N does bind to NusA (31), and RNAPinteracts with NusA, NusG, and S10 proteins (32-34). More-over, S10 can form a complex with NusB (34), and theNusB-S10 complex can bind to a version of the boxAcomponent of nut RNA (35).
In spite of these advances, there is little understanding ofthe fundamental mechanisms by which N, Nus factors, andnutRNA enable RNAP to bypass terminators (1, 2). Here weattempt to dissect the specific roles ofN and Nus factors inthe transformation of RNAP to the termination-resistantform.
MATERIALS AND METHODSTranscription reactions were carried out in two steps asdescribed (13, 26), with minor modifications. In the first step,transcription complexes poised at a specific template locationwere generated by incubating 33.4 nM RNAP and 33.4 nMtemplate DNA with 100 uM ApU as the primer for initiationof transcription from pL and 2 1LM each ATP, GTP, and CTPand 1-2 ttCi of [a-32P]ATP (1 Ci = 37 GBq) to allow thesynthesis of a labeled, 15-nt transcript. The reaction wasperformed at 300C for 6 min in 12.5 t4 of transcription buffer(20 mM Tris glutamate, pH 8/5 mM magnesium glutamate/100mM potassium glutamate/5% glycerol/i mM dithiothrei-tal). In the second step, transcript elongation proceeded for5 min upon the addition of an equal volume of prewarmed(300C) transcription buffer that contained the elongationfactors as well as 100 ,uM each UTP, GTP, and CTP and 4mMATP. The dilution of radioactivity yielded end-labeled tran-scripts from the first round of synthesis and restricted label-ing of transcripts in subsequent rounds. Transcripts wereextracted, resolved, and quantitated as described (13).Template fragments of various plasmids, derived from
pWW10 with A pL-nutL module followed by A terminatortR', were prepared as described (26). Highly homogeneouspreparations of various proteins were used. Soluble forms ofNusA, NusB, and N were purified from overproducingstrains, as described (24). NusG was purified from the solubleextract of an overproducer strain, described by Sullivan andcoworkers (28), by successive chromatography throughDEAE-cellulose and phosphocellulose; NusG was monitored
Abbreviation: RNAP, RNA polymerase.*To whom reprint requests should be addressed.
8660
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 91 (1994) 8661
by SDS/PAGE. S10 protein was a gift from L. Kahan(University of Wisconsin, Madison). N and RNAP (LKB)were free from each of four Nus factors, as judged byWestern blotting using specific antisera.
RESULTSReconstitution of Persistent Antitermination. Antitermina-
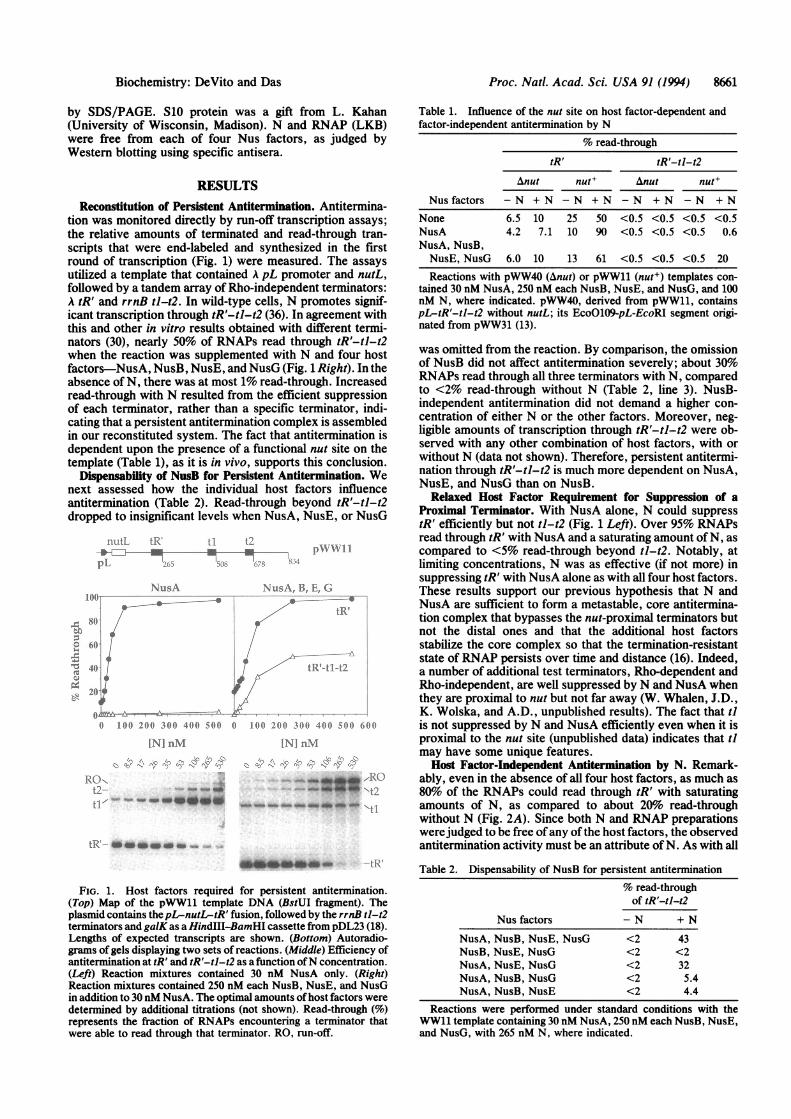
tion was monitored directly by run-off transcription assays;the relative amounts of terminated and read-through tran-scripts that were end-labeled and synthesized in the firstround of transcription (Fig. 1) were measured. The assaysutilized a template that contained A pL promoter and nutL,followed by a tandem array of Rho-independent terminators:A tR' and rrnB tl-t2. In wild-type cells, N promotes signif-icant transcription through tR'-tl-t2 (36). In agreement withthis and other in vitro results obtained with different termi-nators (30), nearly 50% of RNAPs read through tR'-tl-t2when the reaction was supplemented with N and four hostfactors-NusA, NusB, NusE, and NusG (Fig. 1 Right). In theabsence of N, there was at most 1% read-through. Increasedread-through with N resulted from the efficient suppressionof each terminator, rather than a specific terminator, indi-cating that a persistent antitermination complex is assembledin our reconstituted system. The fact that antitermination isdependent upon the presence of a functional nut site on thetemplate (Table 1), as it is in vivo, supports this conclusion.
Dispensability of NusB for Persistent Antitermination. Wenext assessed how the individual host factors influenceantitermination (Table 2). Read-through beyond tR'-tl-t2dropped to insignificant levels when NusA, NusE, or NusG
nll tIN 1 i t2
pL
x
CJ
0-
NusA NusA, B, E, GC~~~~~~~~~~--.-.
tR
40i1 t IV/ VItR-t 212
fl 1
0 1001 200 300) 400 -5)14 0 1001 200 3010 400 5004 60114
NIl nM IN] nM
12- .Nt1X _
tl/---*
tR--- _ 4 _
aiw~mg .-1tRk
FIG. 1. Host factors required for persistent antitermination.(Top) Map of the pWW11 template DNA (BstUI fragment). Theplasmid contains thepL-nutL-tR' fusion, followed by the rrnB tl-t
terminators and galK as a HindIII-BamHI cassette from pDL23 (18).Lengths of expected transcripts are shown. (Bottom) Autoradio-grams of gels displaying two sets of reactions. (Middle) Efficiency ofantitermination at tR' and tR'-tl-t2 as a function ofN concentration.(Left) Reaction mixtures contained 30 nM NusA only. (Right)Reaction mixtures contained 250 nM each NusB, NusE, and NusGin addition to 30 nM NusA. The optimal amounts ofhost factors weredetermined by additional titrations (not shown). Read-through (%)represents the fraction of RNAPs encountering a terminator thatwere able to read through that terminator. RO, run-off.
Table 1. Influence of the nut site on host factor-dependent andfactor-independent antitermination by N
% read-through
tR' tR'-tl-t2
Anut nut+ Anut nut+
Nus factors - N + N - N + N - N + N - N + N
None 6.5 10 25 50 <0.5 <0.5 <0.5 <0.5NusA 4.2 7.1 10 90 <0.5 <0.5 <0.5 0.6NusA, NusB,NusE, NusG 6.0 10 13 61 <0.5 <0.5 <0.5 20
Reactions with pWW40 (Anut) or pWW11 (nut+) templates con-tained 30 nM NusA, 250 nM each NusB, NusE, and NusG, and 100nM N, where indicated. pWW40, derived from pWW11, containspL-tR'-tl-t2 without nutL; its EcoO109-pL-EcoRI segment origi-nated from pWW31 (13).
was omitted from the reaction. By comparison, the omissionof NusB did not affect antitermination severely; about 30%oRNAPs read through all three terminators with N, comparedto <2% read-through without N (Table 2, line 3). NusB-independent antitermination did not demand a higher con-centration of either N or the other factors. Moreover, neg-ligible amounts of transcription through tR'-tl-t2 were ob-served with any other combination of host factors, with orwithout N (data not shown). Therefore, persistent antitermi-nation through tR'-tl-t2 is much more dependent on NusA,NusE, and NusG than on NusB.
Relaxed Host Factor Requirement for Suppression of aProximal Terminator. With NusA alone, N could suppresstR' efficiently but not tl-t2 (Fig. 1 Left). Over 95% RNAPsread through tR' with NusA and a saturating amount of N, ascompared to <5% read-through beyond tl-t2. Notably, atlimiting concentrations, N was as effective (if not more) insuppressing tR' with NusA alone as with all four host factors.These results support our previous hypothesis that N andNusA are sufficient to form a metastable, core antitermina-tion complex that bypasses the nut-proximal terminators butnot the distal ones and that the additional host factorsstabilize the core complex so that the termination-resistantstate of RNAP persists over time and distance (16). Indeed,a number of additional test terminators, Rho-dependent andRho-independent, are well suppressed by N and NusA whenthey are proximal to nut but not far away (W. Whalen, J.D.,K. Wolska, and A.D., unpublished results). The fact that tiis not suppressed by N and NusA efficiently even when it isproximal to the nut site (unpublished data) indicates that tlmay have some unique features.Host Factor-Independent Antitermination by N. Remark-
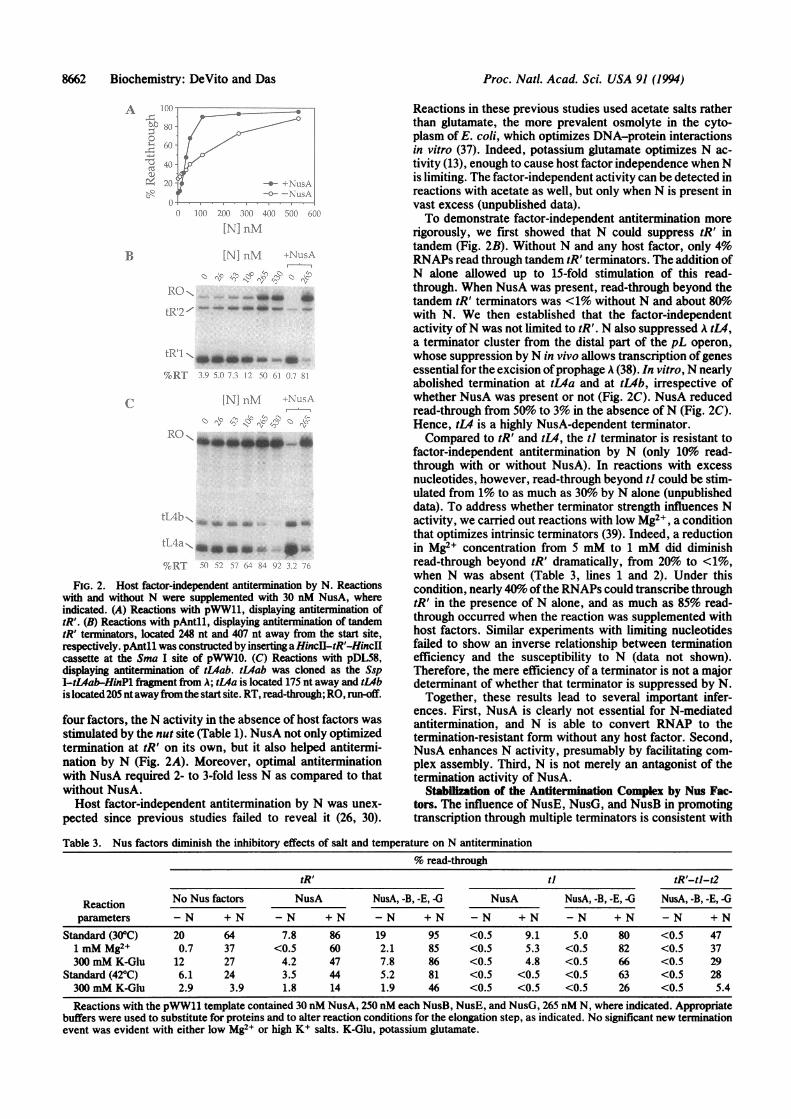
ably, even in the absence of all four host factors, as much as80% of the RNAPs could read through tR' with saturatingamounts of N, as compared to about 20o read-throughwithout N (Fig. 2A). Since both N and RNAP preparationswerejudged to be free ofany ofthe host factors, the observedantitermination activity must be an attribute ofN. As with all
Table 2. Dispensability of NusB for persistent antitermination% read-throughof tR'-tl-t2
Nus factors - N + N
NusA, NusB, NusE, NusG <2 43NusB, NusE, NusG <2 <2NusA, NusE, NusG <2 32NusA, NusB, NusG <2 5.4NusA, NusB, NusE <2 4.4
Reactions were performed under standard conditions with theWW11 template containing 30 nM NusA, 250 nM each NusB, NusE,and NusG, with 265 nM N, where indicated.
Biochemistry: DeVito and Das
III
8
6
4
2
- a" "a* -ft "M 4wa a& me 41"I'm " t 1
8662 Biochemistry: DeVito and Das
A l0
01) +.1,.,S( -Nl9
60
40)
20 -1 - - NI-0-- - NI ~A
{ 1 1 ) 200 .31)O 400 ) O()Oi
NI.] niM
B
1\I .-. -.*. doa
totR - - " -
I4 RT 5 -( l o
C
.:~~-.,C,-_*vA/ .A
tL4bN,
% RT :> 0l ins;2
FIG. 2. Host factor-independent antitermination by N. Reactionswith and without N were supplemented with 30 nM NusA, whereindicated. (A) Reactions with pWW11, displaying antitermination oftR'. (B) Reactions with pAnt1l, displaying antitermination of tandemtR' terminators, located 248 nt and 407 nt away from the start site,respectively. pAntll was constructed by inserting a HinIl-tR'-HincJIcassette at the Sma I site of pWW10. (C) Reactions with pDL58,displaying antitermination of tL4ab. tL4ab was cloned as the SspI-t4ab-HinPl fragment from A; tL4a is located 175 nt away and tLbis located 205 ntawayfom the start site. RT, read-through; RO, run-off.
four factors, the N activity in the absence of host factors wasstimulated by the nut site (Table 1). NusA not only optimizedtermination at tR' on its own, but it also helped antitermi-nation by N (Fig. 2A). Moreover, optimal antiterminationwith NusA required 2- to 3-fold less N as compared to thatwithout NusA.Host factor-independent antitermination by N was unex-
pected since previous studies failed to reveal it (26, 30).
Reactions in these previous studies used acetate salts ratherthan glutamate, the more prevalent osmolyte in the cyto-plasm of E. coli, which optimizes DNA-protein interactionsin vitro (37). Indeed, potassium glutamate optimizes N ac-tivity (13), enough to cause host factor independence when Nis limiting. The factor-independent activity can be detected inreactions with acetate as well, but only when N is present invast excess (unpublished data).To demonstrate factor-independent antitermination more
rigorously, we first showed that N could suppress tR' intandem (Fig. 2B). Without N and any host factor, only 4%RNAPs read through tandem tR' terminators. The addition ofN alone allowed up to 15-fold stimulation of this read-through. When NusA was present, read-through beyond thetandem tR' terminators was <1% without N and about 80owith N. We then established that the factor-independentactivity ofN was not limited to tR'. N also suppressed A tL4,a terminator cluster from the distal part of the pL operon,whose suppression by N in vivo allows transcription ofgenesessential for the excision ofprophage A (38). In vitro, N nearlyabolished termination at tL4a and at tL4b, irrespective ofwhether NusA was present or not (Fig. 2C). NusA reducedread-through from 50%o to 3% in the absence of N (Fig. 2C).Hence, tL4 is a highly NusA-dependent terminator.Compared to tR' and tL4, the ti terminator is resistant to
factor-independent antitermination by N (only 10% read-through with or without NusA). In reactions with excessnucleotides, however, read-through beyond ti could be stim-ulated from 1% to as much as 30% by N alone (unpublisheddata). To address whether terminator strength influences Nactivity, we carried out reactions with low Mg2+, a conditionthat optimizes intrinsic terminators (39). Indeed, a reductionin Mg2+ concentration from 5 mM to 1 mM did diminishread-through beyond tR' dramatically, from 20% to <1%,when N was absent (Table 3, lines 1 and 2). Under thiscondition, nearly 40% ofthe RNAPs could transcribe throughtR' in the presence of N alone, and as much as 85% read-through occurred when the reaction was supplemented withhost factors. Similar experiments with limiting nucleotidesfailed to show an inverse relationship between terminationefficiency and the susceptibility to N (data not shown).Therefore, the mere efficiency of a terminator is not a majordeterminant of whether that terminator is suppressed by N.
Together, these results lead to several important infer-ences. First, NusA is clearly not essential for N-mediatedantitermination, and N is able to convert RNAP to thetermination-resistant form without any host factor. Second,NusA enhances N activity, presumably by facilitating com-plex assembly. Third, N is not merely an antagonist of thetermination activity of NusA.
Stabilization of the Antitermination Complex by Nus Fac-tors. The influence of NusE, NusG, and NusB in promotingtranscription through multiple terminators is consistent with
Table 3. Nus factors diminish the inhibitory effects of salt and temperature on N antitermination% read-through
tR' t tR'-tl-t2
Reaction No Nus factors NusA NusA, -B, -E, -G NusA NusA, -B, -E, -G NusA, -B, -E, -Gparameters - N + N - N + N - N + N - N + N - N + N - N + N
Standard (30"C) 20 64 7.8 86 19 95 <0.5 9.1 5.0 80 <0.5 471 mM Mg2+ 0.7 37 <0.5 60 2.1 85 <0.5 5.3 <0.5 82 <0.5 37300 mM K-Glu 12 27 4.2 47 7.8 86 <0.5 4.8 <0.5 66 <0.5 29
Standard (420C) 6.1 24 3.5 44 5.2 81 <0.5 <0.5 <0.5 63 <0.5 28300mM K-Glu 2.9 3.9 1.8 14 1.9 46 <0.5 <0.5 <0.5 26 <0.5 5.4
Reactions with the pWW11 template contained 30 nM NusA, 250 nM each NusB, NusE, and NusG, 265 nM N, where indicated. Appropriatebuffers were used to substitute for proteins and to alter reaction conditions for the elongation step, as indicated. No significant new terminationevent was evident with either low Mg2+ or high K+ salts. K-Glu, potassium glutamate.
Proc. Natl. Acad. Sci. USA 91 (1994)
--CI
I.,I--,
-j-
7.C-1171)
[N 'n %I X, I 1,. Z. \
.11 i- , -Z. --c

Proc. Natl. Acad. Sci. USA 91 (1994) 8663
a stabilizing role for these factors. To test this hypothesisfurther, we examined how salt and temperature affect anti-termination. Reactions were performed with the pWW11template so that the suppression of tR' as well as tl-t2 couldbe monitored. Indeed, in the absence of host factors, anincrease of monovalent cations from 0.1 M to 0.3 M reducedN activity at tR' 2- to 3-fold (Table 3, line 3). Likewise, thefactor-independent activity ofN was inhibited as the reactiontemperature increased from 30TC to 42TC (Table 3, line 4).Under each condition, N could not suppress t1 at all withouthost factors (data not shown). Likewise, a combination ofhigh salt and high temperature abolished the factor-independent activity of N at tR' (Table 3, line 5).The inhibitory effects of either salt or temperature were
only partly overcome by NusA. By comparison, when allfour Nus factors were present, N activity at tR' and ti wasonly marginally affected by high salt or temperature (Table 3,lines 3 and 4). Under each condition, the complete supple-ment of host factors enabled an appreciable fraction ofRNAPs to read through tR'-tl-t2 as well. We conclude thatNus factors increase the functional stability of the basal Nantitermination complex.
DISCUSSIONThe reconstitution of antitermination with purified compo-nents has led us to address a central question. Does N actupon RNAP directly or does it transmit its antiterminationactivity through one or more of the Nus factors? We haveshown that N, NusA, NusE, and NusG allow substantivetranscription through multiple terminators placed down-stream from the nut site. In the absence of host factors,however, N displays a basal activity. Although a terminatorsuch as rrnB ti is only marginally susceptible to the factor-independent activity ofN, a number of other test terminatorsare well suppressed when they are placed near the nut site.Some of the test constructs include tandem terminators.Hence, without an intermediary host factor, N can endowupon RNAP a termination-resistant state, be it inefficient,transient, or weak. To our knowledge, this is the first timethat this attribute of N has been reported.The simplest hypothesis that emerges from our results is
that a direct interaction between N and RNAP is responsiblefor the transformation of RNAP to the termination-resistantform (Fig. 3). This hypothesis is supported by two lines ofgenetic evidence. First, mutations in rpoB, the gene thatencodes the f3 subunit of RNAP, affect N-mediated antiter-mination (19, 40). Second, certainN mutants, called mar, thatare proficient in antitermination in wild-type E. coli, fail toallow antitermination in hosts with specific alleles of rpoB,called ron (41). This synthetic defect is manifested only whenspecific mar and ron alleles are combined.Even though N does not require an intermediary protein to
display its basal activity, it is dependent on the nut site forfunction. The conventional view holds that nut RNA tethersN in the proximity ofRNAP to facilitate their engagement (7,13). It might even activate N through an allosteric change.However, we cannot rule out the possibility that nut RNA
performs a more active function, for instance, interactingwith RNAP and transforming it to the termination-resistantform (16). Accordingly, N could function as an allostericeffector, changing nut RNA or RNAP to the active confor-mation, or as a stabilizer, holding nut RNA at its proper nicheon the surface of RNAP. Hence, it is attractive to imaginethat both N and nut RNA are responsible for transformingRNAP.The ability ofN to capture and modify RNAP is facilitated
by NusA since it enhances antitermination at limiting con-centrations ofN (Fig. 2). NusA is the only host factor knownto bind to both N and RNAP (31, 32). Therefore, NusA wouldpromote N-RNAP interaction by simply bringing the twopartners together. Another model, not mutually exclusivefrom the first, is that NusA stabilizes the active conformationofN or RNAP, or both, to facilitate the assembly ofthe basalantitermination apparatus. A mutant form of NusA (theproduct of nusAl), which fails to allow antitermination by Nand is thought to be defective in binding to N, does allowantitermination with mutant forms of either N (the punmutants) or RNAP (22, 42, 43). Furthermore, the antitermi-nation defect of a different NusA mutant protein is sup-pressed by the combined efforts of the suppressor N andRNAP mutants (43). The suppressors might have strength-ened N-RNAP interaction sufficiently to bypass NusA'sassistance. Alternatively, they might optimize, directly orindirectly, the binding forces that hold N, NusA, and RNAPtogether in the antitermination complex.How does N transform the elongation behavior of RNAP?
Termination is manifested by a kinetic block to transcriptelongation followed by either spontaneous or Rho-activatedrelease of the transcript from the ternary complex (1). Anti-termination would be favored ifRNAP is accelerated throughthe terminator or the transcript is held tightly and properly inthe ternary complex, or both. Indeed N accelerates RNAPthrough pause sites (ref. 30; W. Whalen and A.D., unpub-lished results). Thus, by this virtue alone, the N antitermi-nation complex would bypass both intrinsic and Rho-dependent terminators. An important remaining issue iswhether N stabilizes the interaction between RNAP and thenascent transcript so that both pausing and transcript releaseare hindered.The N antitermination complex must persist over time and
distance so that RNAP can transcribe processively throughmultiple terminators. The fact that N activity at limitingconcentrations is augmented by NusA alone (Fig. 1) indicatesthat NusB, NusE, and NusG have little, ifany, positive effecton the assembly of the antitermination complex. Thesefactors do strengthen an otherwise weak N-RNAP interac-tion, however, since they suppress the inhibition of the basalantitermination complex by salt or temperature (Table 3).The antitermination complex formed with all four factors isstable physically (29) and also strong functionally, enough tobypass multiple terminators and traverse a long distanceprocessively (ref. 30; Fig. 1). The increased binding forcemust be derived from the additional protein-protein andprotein-nucleic acid interactions (29-35). Presumably, theseadditional interactions allow an efficient suppression of Rho-
InhibitorNusB
RNA usNBar~~~~~~~~~~~~m I _)Basal Complex {9X>sisf ent (Comple.x
SI 0 NusG
FIG. 3. Model for N antitermination and regulation. Details are described in text.
Biochemistry: DeVito and Das

8664 Biochemistry: DeVito and Das
dependent terminators (refs. 30 and 34; W. Whalen and A.D.,unpublished results).The fact that persistent antitermination is virtually abol-
ished by a combination of high salt and high temperaturedespite the presence of all four factors (Table 3) raises thepossibility that additional stabilizing factors might influenceantitermination by N. Two pieces ofevidence lend support tothis conjecture. First, when NusG is depleted from cells,persistent antitermination can occur at least through someterminators (27, 28). Conceivably, an additional stabilizingcomponent missing in our reaction could compensate for theabsence of NusG in vivo. Second, the inadequacy of the fourNus factors for rrn operon antitermination in vitro impliesthat some as yet unidentified factor(s) are required for thisprocess (44). Since rrn antitermination shares at least somefeatures ofN antitermination, it may not be surprising if themissing factor(s) also influence N activity.The evidence that persistent antitermination can occur
without NusB shows that S10 can influence antiterminationwithout forming a complex with NusB. Indeed, S10 forms acomplex with RNAP independent ofNusB (33). The dispens-ability of NusB for persistent antitermination might be ter-minator specific. However, antitermination of tJ-t2 doesdemand NusB when the reaction is carried out in crudeextracts (36). One simple model can account for these results:NusB counteracts an inhibitory factor in the cell that ispresent in the crude extract but missing in our reconstitutedsystem. Thus, NusB may perform two functions. It not onlystabilizes the antitermination complex but also counteracts afactor that otherwise inhibits N antitermination. Recentgenetic evidence suggests that this hypothetical inhibitor maycompete with NusB for binding to boxA (45). Perhaps NusBand the negative factor modulate N antitermination to exe-cute the lysogeny-lysis decision in appropriate cells (7).The notion that N converts RNAP to the termination-
resistant form through a direct interaction with one of thecore subunits and that NusA facilitates this interaction is inkeeping with the mechanism of action of another well-characterized antiterminator, A Q (46). It will be important todeduce why Q has evolved to utilize a DNA site for RNAPcapture, rather than an RNA site, and how Q functionsoptimally without the aid of multiple factors (46, 47).
We dedicate this work to Hatch Echols as a tribute to his insightsand aspirations. We thank William Whalen and Krystyna Wolska forsharing plasmids and proteins and their advice and criticisms. Theseexperiments were supported by National Institutes of Health GrantGM28946. J.D. was supported in part by a graduate fellowship fromthe Health Center Research Advisory Committee.
1. Das, A. (1993) Annu. Rev. Biochem. 62, 893-930.2. Greenblatt, J., Nodwell, J. R. & Mason, S. W. (1993) Nature
(London) 364, 401-406.3. Kerpolla, T. K. & Kane, C. M. (1991) FASEB J. 5, 2833-2842.4. Houman, F., Diaz-Torres, M. R. & Wright, A. (1990) Cell 62,
1153-1163.5. Lindaroth, N. A. & Calendar, R. L. (1991) J. Bacteriol. 173,
6722-6731.6. Roberts, J.(1969) Nature (London) 224, 1168-1174.7. Das, A. (1992) J. Bacteriol. 174, 6711-6716.8. Friedman, D. I. (1988) The Bacteriophages, ed. Calender, R.
(Plenum, New York), Vol. 2, pp. 263-319.
9. Gottesman, M. E., Adhya, S. & Das, A. (1980) J. Mol. Biol.140, 57-75.
10. Friedman, D. I., Wilgus, G. S. & Mural, R. J. (1973) J. Mol.Biol. 81, 505-516.
11. Salstrom, J. S. & Szybalski, W. (1978) J. Mol. Biol. 124,195-221.
12. Rosenberg, M., Court, D., Shimatake, H., Brady, C. & Wulff,D. L. (1978) Nature (London) 272, 414-422.
13. Whalen, W. & Das, A. (1990) New Biol. 2, 975-991.14. Friedman, D. I., Olson, E. R., Johnson, L. L., Alessi, D. &
Craven, M. G.(1990) Genes Dev. 4, 2210-2222.15. Nodwell, J. R. & Greenblatt, J. (1991) Genes Dev. 5, 2141-
2151.16. Barik, S., Ghosh, B., Whalen, W., Lazinski, D. & Das, A.
(1987) Cell 50, 885-899.17. Horwitz, R. J., Li, J. & Greenblatt, J. (1987) Cell 51, 631-641.18. Lazinski, D., Grzadzielska, E. & Das, A. (1989) Cell 59,
207-218.19. Friedman, D. I., Olson, E. R., Georgopoulos, C., Tilly, K.,
Herskowitz, I. & Banuett, B. (1984) Microbiol. Rev. 48, 299-325.
20. Friedman, D. I. & Baron, L. S. (1974) Virology 58, 141-148.21. Friedman, D. I., Baumann, M. F. & Baron, L. S. (1976) Vi-
rology 73, 119-127.22. Friedman, D. I., Schauer, A. T., Baumann, M. F., Baron,
L. S. & Adhya, S. L. (1981) Proc. Natl. Acad. Sci. USA 78,1115-1119.
23. Das, A. & Wolska, K. (1984) Cell 38, 165-173.24. Ghosh, B. & Das, A. (1984) Proc. Natl. Acad. Sci. USA 81,
6305-6309.25. Das, A., Ghosh, B., Barik, S. & Wolska, K. (1985) Proc. Natl.
Acad. Sci. USA 82, 4070-4074.26. Whalen, W., Ghosh, B. & Das, A. (1988) Proc. Natl. Acad. Sci.
USA 85, 2494-2498.27. Sullivan, S. & Gottesman, M. (1990) Cell 68, 989-994.28. Sullivan, S., Ward, D. & Gottesman, M. (1992) J. Bacteriol.
174, 1339-1344.29. Mason, S. W. & Greenblatt, J. (1991) Genes Dev. 5, 1504-1512.30. Mason, S. W., Li, J. & Greenblatt, J. (1992) J. Biol. Chem. 267,
19418-19426.31. Greenblatt, J. & Li, J. (1981) J. Mol. Biol. 147, 11-23.32. Greenblatt, J. & Li, J. (1981) Cell 24, 421-428.33. Mason, S., Li, J. & Greenblatt, J. (1991) J. Mol. Biol. 223,
55-66.34. Li, J., Mason, S. W. & Greenblatt, J. (1993) Genes Dev. 7,
161-172.35. Nodwell, J. & Greenblatt, J. (1993) Cell 72, 261-268.36. Ghosh, B., Grzadzielska, E., Bhattacharya, P., Peralta, E.,
DeVito, J. & Das, A. (1991) J. Mol. Biol. 222, 59-66.37. Leirmo, S., Harrison, C., Cayley, D. S., Burgess, R. R. &
Record, M. T., Jr. (1987) Biochemistry 26, 2095-2101.38. Limberger, R. J. & Campbell, A. M. (1987) Gene 61, 135-144.39. Reynolds, R., Bermudez-Cruz, R. M. & Chamberlin, M. J.
(1992) J. Mol. Biol. 224, 31-51.40. Georgopoulos, C. P. (1971) Proc. Natl. Acad. Sci. USA 68,
2977-2981.41. Ghysen, A. & Pironio, M. (1972) J. Mol. Biol. 65, 259-272.42. Jin, D. J., Cashel, M., Friedman, D. I., Nakamura, Y., Walter,
W. A. & Gross, C. A. (1988) J. Mol. Biol. 204, 247-261.43. Sparkowski, J. & Das, A. (1992) Genetics 130, 411-428.44. Squires, C., Greenblatt, J., Li, J., Condon, C. & Squires, C. L.
(1992) Proc. Natl. Acad. Sci. USA 90, 970-974.45. Patterson, T. A., Zhang, Z., Baker, T., Johnson, L. L., Fried-
man, D. I. & Court, D. (1994) J. Mol. Biol. 236, 217-228.46. Grayhack, E. J., Yang, X., Lau, L. F. & Roberts, J. W. (1985)
Cell 42, 259-269.47. Yarnell, W. S. & Roberts, J. W. (1992) Cell 69, 1181-1189.
Proc. Natl. Acad. Sci. USA 91 (1994)


















