
A humoráli ellenanyagos tanulmányozásk a nyulakban...
25
Parasit. Hung. 4. 23-46. 1971 A humorális ellenanyagok tanulmányozása nyulakban kísérletileg létesített Cysticercus pisiformis (Bloch, 1780)-fertőzöttség esetén í. A precipitin-válasz jellemzése Dr. NÉMETH István A Magyar Tudományos Akadémia Állategészségügyi Kutató Intézete, Budapest Először MILLER és KERR (18) állapította meg 1932-ben, hogy a Taenia pisiformis (Bloch, 1780) Gmelin l790(Taeniidae, Cestoda) lárvaalakjával, a Cysticercus pisiformis (Bloch, 1780) -szal fertőzött nyulakban immunitás alakul k i a reinfekciókkal szem- ben. További vizsgálatok kiderítették, hogy az immunitás ese- tenként annyira kifejezett, hogy a fertőzésen egyszer már át- esett állatok sikerrel állnak ellen a parazita később bekövet- kező támadásával szemben. Máskor viszont az immunitás csupán redukálja a reinfekció esetén a kifejlődő élősködők számát, de ilyenkor i s elejét veszi a súlyosabb cysticercosis kialakulásá- nak (11, 12, 17, 21, 22, 27). Galandféreg- vagy cysticercus-anyag, vagy in vitro mestersége- sen keltetett és aktivált oncosphaerák parenterális bevitelével sikerült nyulakat immunizálni a fertőzéssel szemben. A mester- ségesen indukált immunitás azonban rendszerint gyengébb védett- séget eredményezett, mint a természetes fertőződések nyomán ki- alakult immunitás (l, 8, 9, 11, 12, 18, 27). Passzív immunizálási kísérletek eredményei alapján már korábban több kutató i s arra következtetett, hogy e féregfaj támadása protektiv tulajdonságú humorális ellenanyagok termelődését I n - dukálja (1, 4, 11, 12, 15» 16, 17, 20, 27). Az ellenanyagokat később szerológiai módszerekkel, igy gyürüs-precipitációs pró- bával (4, 30), a mesterségesen kikeltetett aktiv oncosphaerák-
Transcript of A humoráli ellenanyagos tanulmányozásk a nyulakban...

Parasit. Hung. 4. 23-46. 1971
A humorális ellenanyagok tanulmányozása nyulakban kísérletileg létesített Cysticercus pisiformis (Bloch, 1780)-fertőzöttség esetén í. A precipitin-válasz jellemzése
Dr. NÉMETH István
A Magyar Tudományos Akadémia Állategészségügyi Kutató Intézete, Budapest
Először MILLER és KERR (18) állapította meg 1932-ben, hogy a Taenia p i s i f o r m i s (Bloch, 1780) Gmelin l 7 9 0 ( T a e n i i d a e , Cestoda) lárvaalakjával, a Cysticercus p i s i f o r m i s ( Bloch, 1780) - s z a l fertőzött nyulakban immunitás a l a k u l k i a reinfekciókkal szemben. További vizsgálatok kiderítették, hogy az immunitás esetenként ann y i r a k i f e j e z e t t , hogy a fertőzésen egyszer már áte s e t t állatok s i k e r r e l állnak e l l e n a p a r a z i t a később bekövetkező támadásával szemben. Máskor v i s z o n t az immunitás csupán redukálja a reinfekció esetén a kifejlődő élősködők számát, de i l y e n k o r i s elejét v e s z i a súlyosabb c y s t i c e r c o s i s kialakulásának ( 1 1 , 12, 17, 21, 22, 27).
Galandféreg- vagy cysticercus-anyag, vagy i n v i t r o mesterségesen k e l t e t e t t és aktivált oncosphaerák parenterális bevitelével sikerült ny u l a k a t immunizálni a fertőzéssel szemben. A mesterségesen indukált immunitás azonban r e n d s z e r i n t gyengébb védettséget eredményezett, mint a természetes fertőződések nyomán k i a l a k u l t immunitás ( l , 8, 9, 1 1 , 12, 18, 27).
Passzív immunizálási kísérletek eredményei alapján már korábban több kutató i s a r r a következtetett, hogy e féregfaj támadása p r o t e k t i v tulajdonságú humorális ellenanyagok termelődését I n dukálja ( 1 , 4, 1 1 , 12, 15» 16, 17, 20, 27). Az ellenanyagokat később szerológiai módszerekkel, i g y gyürüs-precipitációs próbával ( 4 , 3 0 ) , a mesterségesen k i k e l t e t e t t a k t i v oncosphaerák-

k a i végzett i n v i t r o lárvaprecipitációs próbával ( 2 4 , 2 5 ) , komplementumköté s i ( 3 0 ) , géldiffuziós-precipitációs ( 1 9 ) , valamint i n d i r e k t hemagglutinációs próbával (19, 30) i s sikerült kimut a t n i a fertőzött állatok vér savójában.
Noha az eddig végzett vizsgálatokban kétségtelenül beigazolód o t t , hogy a fertőzés kiállása az ujabb fertőzéssel szemben spec i f i k u s védettséget eredményez ( l , 8, 11, 12, 17, 18, 21, 22, 27 ) , még ma i s hiányosan ismert a szervezetnek a reinfekcióval szemben megnyilvánuló f e l f o k o z o t t védekező reakciójának mechanizmusa. Több i r o d a l m i adat a r r a u t a l , hogy ebben a folyamatban lényeges szerepet töltenek be a humorális ellenanyagok ( l , 4 , 8, 1 1 , 12, 15, 16, 20, 24, 2 5 , 26, 27). Ennek részleteiről a-zonban meglehetősen keveset tudunk, i g y a T. p i s i f o r m i s - o n c o -sphaerák támadására jelentkező humorális ellenanyagválasz sincs még pontosan jellemezve. Még egyáltalán nem ismerjük e féregfaj antigénszerkezetét, antigénjeinek kémiai természetét, továbbá azt sem t u d j u k , hogy e féregfaj antigentérmészetü somaticus a-nyagai és/vagy a fejlődő és a k t i v élettevékenységet folytató é-lősködők produktumai közül végső soron melyek felelősek a humorális ellenanyagok termelődésének megindításáért.
E problémakör tisztázásához kivántunk további adatokat szolgált a t n i T. pisiformis-petékkel mesterségesen fertőzött nyulakon végzett kísérletekben a primer és szekunder fertőzésre j e l e n t kező humorális ellenanyagválasz főbb sajátosságainak tanulmányozása révén. Ez a közlemény a fertőzött állatok vérsavójában géldiffuziós-precipitációs próbával kimutatható ellenanyagokra vonatkozó vizsgálatokat f o g l a l j a össze.
Anyagok és módszerek
A) Antigének A féreg- és lárvaantigének készítéséhez kiindulási anyagként mesterségesen fertőzött kutyákból származó k i f e j l e t t galandfér-geket és fertőzött nyulakból gyűjtött fertőzőképes c y s t i c e r c u -sokat használtunk. 24

A c y s t i c e r c u s o k k a l végzett fertőzést követő 10-12. héten a kutyákat leöltük, és a bélcsatornájukhói k i s z e d e t t férgeket áramló csapvizben, majd többször váltott desztillált vi z b e n történő mosás után 1000 NE/ml kristályos p e n i c i l l i n t és 0,5 mg/ml s t r e p -t o m i c i n t tartalmazó desztillált v i z b e n egy órán át inkubáltuk 37°C-on. Az antibiotikum-oldatból k i e m e l t férgeket ezután s t e r i l desztillált vízzel végzett 4-5-szöri mosás után s t e r i l dróthálóra helyeztük a mosófolyadék eltávolítása érdekében, és a felhasználásukig -20°C-on megfagyasztva tároltuk.
A kötőszöveti tokjukból ép állapotban kiszabadított c y s t i c e r c u -sokat mérőhengerben s t e r i l izotóniás konyhasóoldatban történő ülepítéssel többször mostuk. Az utolsó ülepitést követően a f o lyadékot t e l j e s e n eltávolítottuk, és a lárvákat -20°C-on megfagyasztva tároltuk.
Taenia pisiformis-homogenizátum (TpH)
Apró darabkákra va g d a l t nyers galandférgékből 50 g-ot 50 ml p u f f e r o l t izotóniás sóoldatban (PBS, 0,04 M,pH 7,3) homogenizá-t o r r a l " ^ 20 p e r c i g homogenizáltunk, miközben a homogenizátor e-dényét jéggel hűtöttük. Az anyagot ezután 45 p e r c i g r o n c s o l t u k
2 ) o u l t r a h a n g g a l , és a felhasználásig -20 C-on megfagyasztva tár o l t u k .
C ysticercus pisiformis-homogenizátum (CpH)
A fertőzőképes cysticercusokból a TpH-mal azonos módon készült, azzal a különbséggel, hogy az u l t r a h a n g g a l végzett roncsolás 30 p e r c i g történt.
Omni-Mixer Homogenizer, I v a n S o r v a l l , I n c . Norwalk, Conn. USA MSE U l t r a s o n i c D i s i n t e g r a t o r , Measuring and S c i e n t i f i c Equipment L t d . London (kimenő teljesítmény 60 w a t t , rezgésszám 18-20 Kc/sec,).

Taenia p i s i f o r m i s - n y e r s k i v o n a t (TpK)
Egy rész galandféreg-homogenizátumot (TpH) három rész PBS-ol-d a t t a l (0,04 M, pH 7,3) kevertünk, majd 10 p e r c i g homogenizált u k , és 15 p e r c i g u l t r a h a n g g a l kezeltük a f e n t e m i i t e t t módon. A szuszpenziót ezután 20 p e r c i g 3000,majd ujabb 20 p e r c i g 15000 f o r d u l a t t a l centrifugáltuk +4°C-on. Az alakos elemektol mentes felüluszót dializáló hártyán keresztül történő párologtatással (pervaporizáció) 15-30 mg/ml fehérjetartalomra koncentráltuk, majd PBS-oldattal szemben egy napig dializáltuk, és 0,3 % f o r -m a l i n n a l konzerválva +4°C-on tároltuk.
Cysticercus p i s i f o r m i s - n y e r s k i v o n a t (CpK)
A TpK-tal azonos módon CpH-ból készült.
Cysticercus p i s i f o r m i s - k i v o n a t savban oldódó frakciója
Ezt az antigénoldatot a CpK-ból állítottuk elő a következő módon. A CpK kémhatását cseppenként hozzáadott 0,5 N sósavval pH 4,5-re állítottuk be, aminek következtében fehér, l a z a , pelyhes csapadék k e l e t k e z e t t . A precipitátumot ezután 4000 f o r d u l a t t a l végzett 15 perces centrifugálással eltávolítottuk, és az enyhén opaleszkáló felüluszót mágneses keverővel folyamatosan k e v e r t és többször váltott PBS (0,04 M, pH 7,3) e l l e n 6 órán át d i a l i -zálva használtuk antigénként. Az anyagot 0,3 % f o r m a l i n n a l konzerválva +4°C-on tároltuk.
Taenia p i s i f o r m i s - " s e c r e t u m és excretum antigén" (TpSE)
K i f e j l e t t c y s t i c e r c u s o k k a l fertőzött kutyákat a fertőzésüket követő 7-8. héten k i i r t o t t u k . A bélcsatomából ép állapotban ki e m e l t férgeket a fentebb l e i r t módon megtisztítottuk, majd 500 NE/ml kristályos p e n i c i l l i n t és 0,3 mg/ml s t r e p t o m i c i n t ,

valamint 200 NE/ml m y c o s t a t i n t tartalmazó HANKS-oldatban (pH 7,2) 37°C-on 72 óráig inkubáltuk, 100 ml o l d a t r a 3 férget számítva. A férgekről leöntött o l d a t o t 3000 f o r d u l a t t a l 15 p e r c i g végzett centrifugálás után 3 0-40-szeresre koncentráltuk u l t r a -szüréssel +4°C-on, majd a koncentrátumot többször váltott PBS-o l d a t t a l (0,04 M,pH 7,3) szemben egy napig diálizáltuk, és 0,3% f o r m a i i n n a l konzerválva +4°C-on tároltuk.
Cysticercus p i s i f o r m i s - " s e c r e t u m és excretum antigén" (CpSE)
A fentebb l e i r t módon megtisztított és s t e r i l e z e t t ép, fertőzőképes c y s t i c e r c u s o k a t a n t i b i o t i k u m o s HANKS-oldatba helyeztük, 100 lárvát számítva 50 ml o l d a t r a , és 72 óráig 37°C-on t a r t o t t u k . Ezután a lárvákról a folyadékot leöntöttük, és a továbbiakban a TpSE-antigennél leírtakkal azonos módon jártunk e l .
B) Sérumok
Fertőzött nyulakból származó vérsavók
Harminc, azonos tenyészetből származó, 6 -7 hetes korú, 700-900 g s u l y u , fehér uj-zélandi n y u l a t egyenként 500 T. p i s i f o r m i s -petével fertőztünk. A petéket mesterségesen fertőzött kutyákból spontán Ürülő g r a v i d izekből a korábban l e i r t módon izoláltuk ( 2 0 ) , és fecskendőre s z e r e l t szondával közvetlenül a gyomorba adtuk be. A ny u l a k a t a szokásos módon ketrecekben dróthálón t a r t o t t u k . Kísérletbe állításukkor a c o c c i d i o s i s megelőzésére 10 %-oa o l a j o s szulfakvinoxalin-szuszpénzlóból 0,5-1,0 m l - t o l t o t t u n k bőr alá, egyhetes időközökkel, három alkalommal. Ezenkívül az állatok a fertőzést követő első héten e r i t r o m i c i n t tartalmazó ivóvizet (62,5 g eritromicin-tiocianát1 V ü t e r ) kapt a k . Ezt a kezelést a harmadik héten megismételtük.
G a l l i m y c i n , Abbott L a b o r a t o r i e s , I n t e r n a t i o n a l V e t e r i n a r y D i v i s i o n , France

A kísérleti nyulak felét az első fertőzés után a 60. napon újra fertőztük 10 000 petével, majd a 100. napon valamennyi állatot elvéreztettük, és a cysticercus-fertőzöttség intenzitásának meghatározása érdekében f e l b o n c o l t u k . Az első fertőzéstől kezdve a 20. napig 3 -5 naponként, ezután pedig 10 naponként az áll a t o k fülvénájából vért vettünk. A vérsavókat a felhasználásuk i g -20°C-on megfagyasztva tároltuk. A fertőzés következtében öt n y u l e l h u l l o t t , ezeknek a vérsavóját nem vizsgáltuk.
Az anti-Ta e n i a - p i s i f o r m i s nyulimmunBavó előállítása ( a n t i - T p )
A galandférgek homo -reniz át urnát (TpH) izotóniás nátriumklorido l d a t t a l 1 : 5 arányban h i g i t v a Al(OH)-^-gélhez adszorbeáltuk, és ebből az anyagból hat héten keresztül hetenként egyszer IS mg fereghomogenizátum-szárazanyagnak megfelelő mennyiséget o l t o t tunk 3-4 kg-os nyulak bőre alá, több h e l y r e e l o s z t v a . Az immu-nizálás 10. hetében az állatok 25 mg fehérjét tartalmazó TpK-ot kaptak bőr alá, majd öt egymást követő napon át emelkedő adagokban összesen 100 mg fehérjének megfelelő TpK-ot o l t o t t u n k fülvénába. Egyheti szünet után az utolsó oltássorozatot megismételtük, és az utolsó intravénás oltást követő 9« napon az áll a t o k a t elvéreztettük. A vérsavókat -20°C-on tároltuk.
Az a n t i - C y s t i c e r c u s p i s i f o r m i s nyulimmunsavó előállítása
Az anti-Tp nyulimmunsavó előállítása c. bekezdésben l e i r t immunizálási séma s z e r i n t CpH-mal és CpK-tal immunizáltunk n y u l a kat .
Az ellenan.yagok kimutatása
A precipitáló a n t i t e s t e k e t a vérsavókban géldiffuziós-precipi-tációval mutattuk k i , amit 0,01 % mertiolátot és 0,003 % m e t i l -narancsot tartalmazó PBS-ben (0,01 M,pH 7 , l ) o l d o t t 1,5 %-os a-gar-gélben"^ végeztünk. 1 ^ S p e c i a l Agar-ÏToble, D i f c o L a b o r a t o r i e s , D e t r o i t , Michigan, USA
28

Az ellenanyagok tlterének meghatározása érdekében PETRI-csé-székben 1,5-2,0 mm vastag agarrétégben a kör alakú reagenstartályokat rozettaszerüen helyeztük e l . A centrális és a perifériás tartályok középpontja közti távolság 10 mm, a tartályok átmérője pedig 5 mm v o l t . A centrális rezervoárba mindig a vizsgálandó vérsavót, a perifériásokba pedig az antigénként használt 10 mg/ml fehérjetartalmú CpK-törzsoldat felező léptékű higitásait mértük. Az antigénhigitások PBS-oldattal (0,01 M, pH 7,1) készültek. Kontrollként b i z t o s a n nem fertőzött nyulakból származó vérsavókat, i l l e t v e PBS-oldatot alkalmaztunk. A lemezeket szobahőn t a r t o t t u k , és az eredményt a reagensek bemérése után 7 nap múlva biráltuk e l . Azokban az esetekben, amikor a vizsgált vérsavók e r r e az időre nem idéztek elő precipitációt, a reagenseket ujratöltöttük, és az eredményt akkor t a r t o t t u k negatívnak, ha ujabb 7 napi inkubálás után sem a l a k u l t k i látható precipitációs v o n a l . Az ellenanyagok titereként az a n t i génnek a z t a legnagyobb higitását adjuk meg, amely az e l l e n a nyag-tartalmú vérsavóval még jól látható precipitációs vo n a l a t h o z o t t létre.
Az l_ j _ és 2. ábrán közölt átlagtitereket az egyes kisérieti csopo r t o k állataiból azonos időpontokban v e t t vér savók egyedi vizsgálatával k a p o t t t i t e r e k alapján számítottuk k i ( 1 4 ) .
Géldiffuziós analízisek
Az immundiffuziós analíziseket radiális kettős diffúziós móds z e r r e l 1,5 mm vastag agarrétegben végeztük, amit 0,01 % mer-tiolátot és 0,003 % m e t i l n a r a n c s o t tartalmazó PBS-ben (0,01 M, pH 7 , l ) o l d o t t 1,5 %-os agaroldatból különböző méretű üveglemezeken képeztünk. A 10 mm átmérőjű kör alakú rezervoárok középp o n t j a közti távolság 14-18 mm között változott. A vér savókat mindig h i g i t a t l a n u l , az antigéneket pedig PBS-oldattal (0,01 M, pH 7,1) különböző mértékben h i g i t v a alkalmaztuk. Az immundiffuziós lemezeket szobahőn,nedveskamrában t a r t o t t u k , naponta v i z s gáltuk, és a precipitációs vonalak t e l j e s kialakulása, rendszer i n t 6-7 n a p i inkubálás után a szokásos módon (2) mostuk, szá-

r i t o t t u k , majd amidofekete 10 B - v e i vagy savanyu f u k s z i n n a l festettük meg.
A vérsavók koncentrálása ammóniumszulfátos kisózással
A precipitin-aktivitás növelése érdekében a fertőzött nyulak vérsavóját ammóniumszulfátos kisózással koncentráltuk ugy, hogy a desztillált v i z z e l egyenlő arányban h i g i t o t t vérsavókat t e l i t e t t ammóniumszulfát-oldattal ana pa r t e s kevertük, majd 100 ml "féltelitett" oldathoz 17-20 g finoman p o r i t o t t ammoniumszulfá-t o t adtunk, és a keveréket másnapig szobahőn hagytuk állni. Ezután a precipitátumot lecentrifugáltuk, a vérsavó e r e d e t i térfogatának egyhatodát kitevő mennyiségű PBS-ben (0,02 M, pH 7,2) f e l o l d o t t u k , majd ugyanezzel a p u f f e r r a i szemben +4°C-on végz e t t dializálással az oldatból az ammóniumszulfátot eltávolít o t t u k . A koncentrátumot 1 ml-es adagokban -20 °C-on megfagyasztva tároltuk.
A p r e c i p i t i n e k adszorbeálása a vérsavókból
Az adszorbeálandó vérsavó l - l ml-éhez liofilizált CpK-ot adtunk 0,5-1,0 mg-os adagokban. A keveréket 30 p e r c i g 37 °C-on, és másnapig +4°C-on inkubáltuk, majd a képződött precipitátumot 20 p e r c i g 4000 f o r d u l a t t a l végzett centrifugálással eltávolít o t t u k . A müveletet addig ismételtük, amikor az antigén hozzáadására több csapadék már nem k e l e t k e z e t t . Az adszorbció t e l j e s voltáról antigénhigitási s o r r a l végzett géldiffuziós-precipitá-ciós próbával győződtünk meg.
E. Merck AG, Darmstadt, Germany 2)
Chroma, S t u t t g a r t , Germany

Ultraszürés
Az ultraszürést vákuumdializissel 50-100 Hgm n e g a t i v nyomáson 8-as méretű dializáló cső"^ felhasználásával a l e i r t módon végeztük ( 6 ) .
Fehérj emeghatározás
A fehérjemeghatározásokat biuret-reakcióval végeztük a l e i r t módon ( 5 ) .
Eredmények
A parazitológiai boncolás eredménye
A T< pisiformis-peték beadását követő 100. napon végzett boncolás során minden n y u l c y s t i c e r c u s o k k a l fertőzöttnek b i z o n y u l t . Az egyszer fertőzött csoport állatainak hasüregében átlagosan 210,8 (78-377), az ujrafertőzött csoport állataióban pedig átlagosan 397,3 (142-438) Cysticercus v o l t . Az előbbi csoportban valamennyi Cysticercus betokozódott állapotban zömmel a cseple-szen és a medenceüreg hashártyája a l a t t v o l t található, és t e l jesen k i f e j l e t t scolexszel r e n d e l k e z e t t . Az utóbbi állatcsoportban v i s z o n t t e l j e s e n k i f e j l e t t hólyagférgek (borsókák) mell e t t a hasüregben még szabadon lévő, véglegesen még meg nem t e l e p e d e t t , f i a t a l c y s t i c e r c u s o k i s előfordultak, amelyek a többinél lényegesen kisebbek és f e j l e t l e n e b b e k v o l t a k , s i g y n y i l ván a reinfekcióból származtak.
A precipitáló ellenanyagok dinamikája
Precipitáló a n t i t e s t e k e t legkorábban a fertőzést követő második
Union Carbide ^Corporation, Chicago, I l l i n o i s , USA
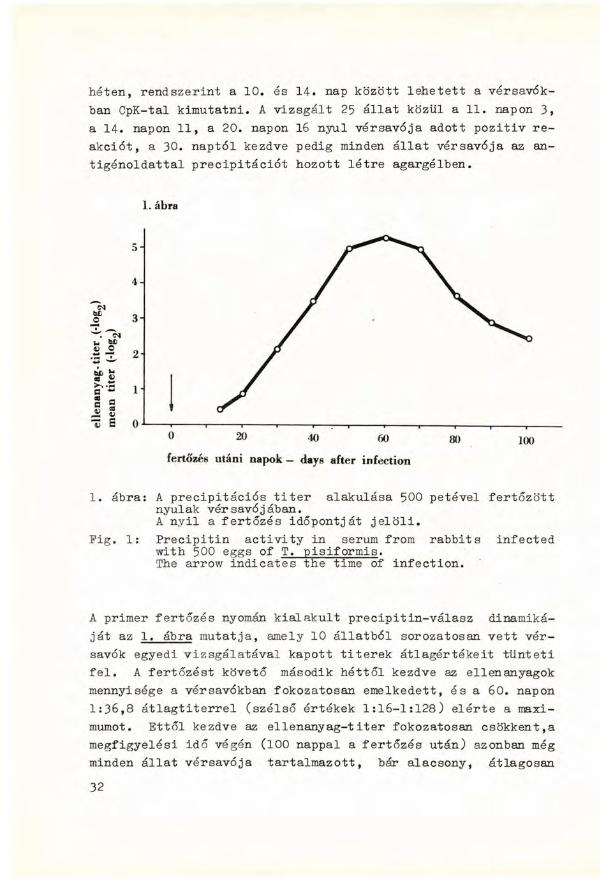
héten, r e n d s z e r i n t a 10. és 14. nap között l e h e t e t t a vérsavók-ban CpK-tal k i m u t a t n i . A vizsgált 25 állat közül a 11. napon 3, a 14. napon 1 1 , a 20. napon 16 n y u l vérsavója adot t p o z i t i v r e akciót, a 30. naptól kezdve pedig minden állat vér savója az antigénoldattal precipitációt h o z o t t létre agargélben.
1. ábra
fertőzés utáni napok - days after infection
1. ábra: A precipitációs t i t e r alakulása 500 petével fertőzött nyulak vér savójában. A n y i l a fertőzés időpontját jelöli.
Pig. 1: P r e c i p i t i n a c t i v i t y i n serum from r a b b i t s i n f e c t e d w i t h 500 eggs of T. p i s i f o r m i s . The arrow i n d i c a t e s the time of i n f e c t i o n .
A primer fertőzés nyomán k i a l a k u l t precipitin-válasz dinamikáját az 1. ábra m u t a t j a , amely 10 állatból sorozatosan v e t t vérsavók egyedi vizsgálatával k a p o t t t i t e r e k átlagértékeit tünteti f e l . A fertőzést követő második héttől kezdve az ellenanyagok mennyisége a vér savókban fokozatosan emelkedett, és a 60. napon 1:36,8 átlagtiterrel (szélső értékek 1:16-1:128) elérte a maximumot. Ettől kezdve az e l l e n a n y a g - t i t e r fokozatosan csökkent,a megfigyelési idő végén (100 nappal a fertőzés után) azonban még minden állat vérsavója t a r t a l m a z o t t , bár alacsony, átlagosan
1:5,3 t i t e r b e n (szélső értékek 1:2-1:16), precipitáló a n t i t e s t e k e t .
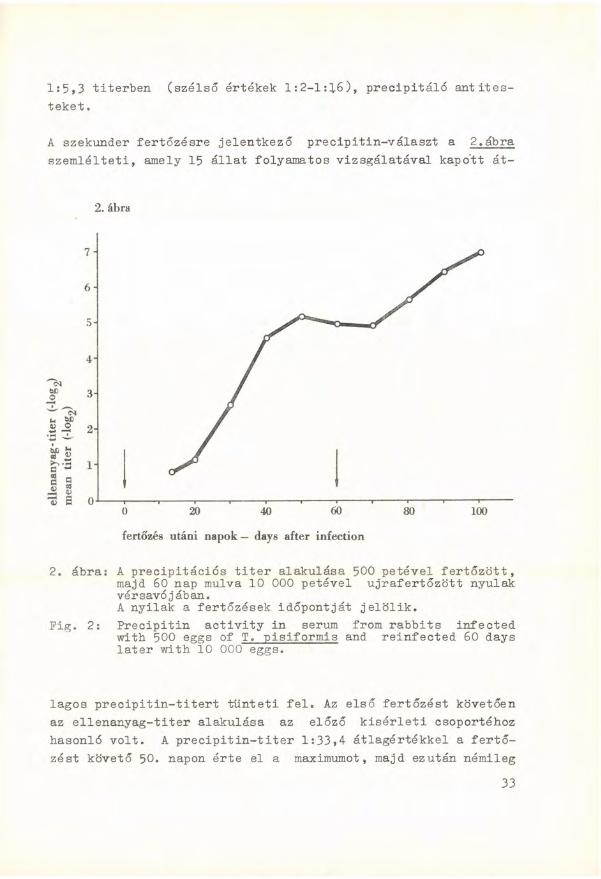
A szekunder fertőzésre jelentkező precipitin-választ a 2.ábra szemlélteti, amely 15 állat folyamatos vizsgálatával kapo'tt át-
2. ábra
0
• Ö 0
"S S
7 -
6 -
5-
4-
3-
S -S 2-
a l
0 20 40 60
fertőzés utáni napok — days after infection
m 100
2. ábra: A precipitációs t i t e r alakulása 500 petével fertőzött, majd 60 nap múlva 10 000 petével ujrafertőzött nyulak vérsavójában. A n y i l a k a fertőzések időpontját jelölik.
Pig. 2: P r e c i p i t i n a c t i v i t y i n serum from r a b b i t s i n f e c t e d w i t h 500 eggs of T. p i s i f o r m i s and r e i n f e c t e d 60 days l a t e r w i t h 10 000 eggs.
lagos p r e c i p i t i n - t i t e r t tünteti f e l . Az első fertőzést követően az e l l e n a n y a g - t i t e r alakulása az előző kisérleti csoportéhoz hasonló v o l t . A p r e c i p i t i n - t i t e r 1:33,4 átlagértékkel a fertőzést követő 50. napon érte e l a maximumot, majd ezután némileg

csökkent. Az ábrán jól látható, hogy a 60. napon 10 000 petével végzett második fertőzésre az ell e n a n y a g s z i n t újból emelkedett, és a 40. napon (az utolsó vizsgált időpont) a vérsavók átlagosan 1:116,7 t i t e r b e n (szélső értékek 1:32-1:512) adtak r e a k c i ót. Ez az érték megközelítően két kitevővel h a l a d t a meg a p r i mer- fertőzés után észlelt maximális átlagtitert, és négy és fél kitevővel magasabb v o l t , mint a csak egyszer fertőzött állatokban ugyanebben az időpontban észlelt p r e c i p i t i n - t i t e r .
Géldiffuziós vizsgálatok
A géldiffuziós vizsgálatok folyamán a CpK-tal szemben maximum négy precipitációs sáv a l a k u l t k i (4/A és 4/B ábra). A vér savók p r e c i p i t i n aktivitása azonban állatonként lényegesen eltért, sőt egyazon állat esetében i s erősen változott attól függően, hogy,egyrészt egy vagy két fertőzést k a p o t t - e , másrészt, hogy a fertőzés és vérvétel között mennyi idő t e l t e l . A fertőzöttség korábbi időszakából származó vér savók esetében csupán egy-egy halvány precipitációs vonal a t l e h e t e t t m e g f i g y e l n i , amely a későbbi időpontokban v e t t vérsavóknál fokozatosan erősebbé vált. A fertőzés utáni 30. naptól kezdve legalább egy jól k i f e j e z e t t precipitációs vonal a t valamennyi állat vérsavója- k i a l a k i t o t t . A fertőzöttség későbbi időszakából, valamint az ujrafertőzött állatokból származó vérsavókban v i s z o n t szélesebb antigén-spektrum e l l e n ható precipitáló ellenanyagokat l e h e t e t t k i m u t a t n i . Egyes állatok esetében ugyanis az első fertőzést követő 30. és 60. nap között a vér savók kettő, a második fertőzés utáni 20. naptól kezdve pedig a legtöbb állat vérsavója három, egyeseké pedig négy precipitációs vonalat létesített, ha agarlemezben CpK-tal.szemben diffundáltak.
Az immundiffuziós vizsgálatokhoz általában alk a l m a z o t t antigénkoncentrációk esetében (10-15 mg/ml fehérje tartalmú CpK) a precipitációs sávok a következőképpen helyeződtek e l . Amint a 3. és 4. ábrán látható, valamennyi precipitációs vonal közül a legerősebb r e n d s z e r i n t a vérsavótartály közelében helyeződött.

Ez a vonal a precipitáló vérsavók esetében mindig k i a l a k u l t , és a fertőzöttség 30. napjától kezdve valamennyi állat vérsavója következetesen létrehozta. Ettől a vonaltól az antigéntartály felé haladva két precipitációs sávot l e h e t e t t m e g f i g y e l n i . Ezek erőssége azonban vérsavónként változott, r e n d s z e r i n t az előzőnél gyengébbek v o l t a k , a diffúzió során később i s fejlődtek k i , és következetesen csak az ujrafertőzött állatok vérsavói alakították k i . Az e m i i t e t t vonalakon k ivü l a 15 ujrafertőzött állat közül ötnek a ráfertőzést .követő 40. napon v e t t vérsavója még egy nagyon vékony precipitációs sávot a l a k i t o t t k i közvetlenül a vérsavótartály m e l l e t t (4/A és 4/B ábra).
A különböző precipitin-aktivitásu vérsavókat kombinált rendszerben CpK-tal szemben összehasonlítva, az egyes vérsavók ált a l p r é c i p i t a i t vonalak egymással összefolytak (3« ábra), jeléül annak, hogy a vérsavókban azonos antigénkomponenssel vagy komponensekkel reagáló ellenanyagok v o l t a k .
A CpK-tal agarlemezben p o z i t i v e n reagáló vérsavók precipitációt hoztak létre a TpK, CpSE és a TpSE antigénoldatokkal i s . A négy antigénpreparátumot kombinált rendszerben összehasonlítva az e-gyes antigénoldatokkal szemben k i a l a k u l t precipitációs sávok a-zonossági reakciót adtak (4. ábra). Hangsúlyoznom k e l l azonban, hogy az egyes precipitációs vonalak a különböző vérsavók esetében - mivel a vérsavók e l l e n a n y a g - t a r t a l m a erősen változott -más és más antigénkoncentrációnál a l a k u l t a k k i optimálisan. Ennélfogva valamennyi precipitációs v o n a l a t egyrészt nem l e h e t e t t e gyetlen lemezen demonstrálni, másrészt a n a t i v , nem koncentrált vérsavók használatával a nagyon gyenge vonalak fúzióját nem mindig l e h e t e t t megnyugtató módon elbirálni. A vér savók ammóniumszulfátos kicsapással végzett koncentrálása után azonban a precipitációs sávok sokkal kifejezettebbekké váltak, s e-zért a koncentrátumok segítségével kétségtelenül meg l e h e t e t t állapítani, hogy a CpK, TpK, CpSE és a TpSE antigénoldatokban egyaránt azonos komponensek precipitálódtak.
A fertőzőképes c y s t i c e r c u s o k t e l j e s kivonatának savban oldódó frakciójával szemben ugyanazok a precipitációs vonalak a l a k u l -

3. ábra: A p r e c i p i t i n e k kimutatása a fertőzött nyulak vér savójában agargéldiffuziós-precipitációs próbával. A rezervoárok a következő reagenseket tartalmazzák: 1 , 2.» 2» 4 és 5« Az első fertőzést követő 50.~ napon v e t t vér savók: 7, 8, 9. és 10. A második fertőzés utáni 40. napon v e t t ve"rsavók: 11. CpK (10 mg fehérje/ml).
F i g . 3: Detection of a n t i b o d i e s i n serum of i n f e c t e d r a b b i t s by g e l d i f f u s i o n p r e c i p i t i n t e s t . Wells were f i l l e d as f o l l o w s : 1, 2, 3, 4 and 5: serum samples taken on the 50th day a f t e r primary i n f e c t i o n ; 6, 7, 8, 9 and 10: serum samples taken on the 40th day a f t e r secondary i n f e c t i o n ; 11: CpE (10 mg p r o t e i n / m l ) .
4. ábra: A T. és C. p i s i f o r m i s antigénjeinek géldiffuziós analízise fertőzött nyulakból származó vérsavókkal. A rezervoárok a következő reagenseket tartalmazzák: l_j_ és 7. CpK (15, i l l e t v e 10 mg fehérje/ml); 2_j_ TpK I (14 mg fehérje/ml); 3_ és 4- A második fertőzés után a 40. napon v e t t vér savók: 5_ és 9_. Normál n y u l vér savó; 6. CpSE (9 mg fehérje/ml)^ 8̂ TpSE (9 mg fehérje/ml); 10 és 11_. A második fertőzés utáni 20. napon v e t t vér savók; 12_. A CpK savban oldódó frakciója (10 mg fehérje/ m l ) .
F i g . 4: Gel d i f f u s i o n a n a l y s i s i l l u s t r a t i n g r e a c t i o n s between d i f f e r e n t a n t i g e n p r e p a r a t i o n s of T. and C. p i s i f o r m i s and serum samples from i n f e c t e d r a b b i t s . Wells were f i l l e d as f o l l o w s : 1 and 7: CpE (15 and 10 mg p r o t e i n / m l , r e s p e c t i v e l y ) ; 2: TpE (14 mg p r o t e i n / m l ) ; 3 and 4: serum samples taken on the 40th day a f t e r secondary i n f e c t i o n ; 5 and 3: normal r a b b i t serum; 6: CpSE (9 mg p r o t e i n / m l ) ; 8: TpSE (9 mg p r o t e i n / m l ) ; 10 and 11: serum samples taken on the 20th day a f t e r secondary i n f e c t i o n ; 12: a c i d s o l u b l e f r a c t i o n of CpE (10 mg p r o t e i n / m l ) .
5. ábra: C. p i s i f o r m i s s z a l mesterségesen immunizált nyulak vér-savojanak géldiffuziós vizsgálata. A rezervoárok a következő reagenseket tartalmazzák: 1 , 2_, 3 és 4. Anti-Cp n y u l immunsavók; 5« Normál nyulvér-savo; 6. CpK (15 mg fehérje/ml); 7. PBS (0,01 M,pH 7,1).
F i g . 5: Gel d i f f u s i o n a n a l y s i s i l l u s t r a t i n g r e a c t i o n s between CpK and anti-Cp serum. Wells were f i l l e d as f o l l o w s : 1 , 2, 3 and 4: serum from r a b b i t s a r t i f i c i a l l y immunized w i t h C. p i s i f o r m i s antigens; 5: normal r a b b i t serum: 6. CpE (15 mg pro-t e i n / m l ) ; 7: PBS (0,01 M, pH 7 . l ) .

4. ábra

t a k k i , mint amik a CpK-tal szemben i s kifejlődtek, és az egyes precipitációs sávok egymással t e l j e s e n összefolytak (4/B ábra). Az ellenanyagok CpK-tal végzett adszorbeálása t e l j e s e n megszüntette a vérsavók reaktivitását valamennyi antigénpreparátummal szemben.
A T. p i s i f o r m i s s z a l , i l l e t v e C. p i s i f o r m i s s z a l mesterségesen immunizált nyulak p r e c i p i t i n - t i t e r e nagyon magas v o l t . Az a n t i -Tp, i l l e t v e anti-Cp nyulimmunsavók még az l í 2 *" ̂ higitásu Tp K - t a l , i l l e t v e CpK-tal (15 mg fehérje/ml) i s jól látható precipitációs csikót alakítottak k i ; a homológ k i v o n a t o k k a l reagált a t v a 6-9 precipitációs vona l a t hoztak létre ( 5 . ábra).
Megbeszélés
CAMPBELL ( 4 ) mutatta k i először, hogy a T. p i s i f o r m i s ( B loch, 1 7 8 0 ) Gmelin 1 7 9 0 lárvaformájával fertőzött nyulak vér savója o-l y a n ellenanyagokat t a r t a l m a z , amelyek e féregből készített s p e c i f i k u s poliszacharid-antigénnel ( 3 ) gyürüs-precipitációs próbában reagálva precipitációt hoznak létre. TRAWINSKI ( 3 1 ) leirása s z e r i n t preparált lárvakivonattal később SUL'C és ISZ-MAGILOVA ( 3 0 ) k a p o t t p o z i t i v reakciókat a fertőzött nyulak vérsavójából ugyancsak gyürüs-precipitációs technikával. SILVERMAN ( 2 3 , 24, 2 5 ) lárvaprecipitációs módszerrel t u d o t t ellenanyagok a t k i m u t a t n i . Az i n v i t r o k i k e l t e t e t t oncosphaerákat a fertőzött állatok vérsavójába helyezve, az a k t i v oncosphaerák t e s t e és az un. penetrációs m i r i g y secretumai körül mikroprecipitátu-mok képződtek, amit e szerző az élő oncosphaerák antigénhatásu produktumai és a vérsavókban lévő adekvát ellenanyagok közti reakció következményének t a r t o t t . I l y e n reakciókat legkorábban a fertőzést követő második héten észlelt, maximum 1:16 - 1 : 3 2 a-rányban h i g i t o t t vérsavókban. A reakció t i t e r e azonban függött az állatok fertőzöttségének erősségétől, és esetenként p o z i t i v reakciók l:64-es higitásu vérsavókban i s előfordultak.

Géldiffuziós precipitációval végzett vizsgálatok során a fertőzött állatok vérsavója agargélben precipitációs ho z o t t létre a somaticus antigénkivonatokkal, s ezért az agargéldiffuziós-pre-cipitációs próba i s alkalmas módszernek b i z o n y u l t e féregfaj antigénjei e l l e n ható a n t i t e s t e k kimutatására ( 1 9 ) .
T. pisiformÍ3-oncosphaerák inváziója nyomán kialakuló p r e c i p i -tin-válasz jellemzésére e z i d e i g nem történtek részletes vizsgál a t o k . Csupán a precipitáló ellenanyagok dinamikájáról közölt SUL'C és ISZMAGILOVA ( 3 0 ) adatokat. Különböző erősségű fertőzéseknek k i t e t t nyulakon végzett kísérletekben e szerzők csőpre-cipitációs próbával már a fertőzést követő második napon kimut a t t a k ellenanyagokat a vérsavókban. A reakció a fertőzöttség 1 0 - 2 0 . napja között v o l t a l e g k i f e j e z e t t e b b , ebben a periódus-
7 bah v e t t vérsavók még az 1 : 1 0 higitásu lárvakivonattal i s pozitívan reagáltak. Később a reakció fokozatosan gyengült, és a 9 0 . napon a próba r e n d s z e r i n t n e g a t i v v o l t . A fertőző petedózistól függetlenül a reakció dinamikája az egyes állatokban általában hasonló v o l t , és semmilyen összefüggést sem találtak a fertőzés intenzitása ás az ellenanyagok mennyisége, i l l e t v e e-zek kimutathatóságának időtartama között. A reinfekció nem idéz e t t elő észrevehető e l l e n a n y a g - s z a p o r u l a t o t , bár az ismételt fertőzések után észleltek némi erősödést a reakció intenzitásában. Ez azonban egyáltalán nem állt arányban a fertőző dóziss a l , r e n d s z e r i n t gyorsan eltűnt, és a 9 0 . napra a reakció éppúgy negatiwá vált, mint a csak egyszer fertőzött állatok esetében* Ebben a dolgozatban közölt vizsgálataink eredményei több kérdésben i s ellentétesek SŰL»C és ISZMAGILOVA ( 3 0 ) előbb e m i i t e t t megállapításaival, Igy a fertőzés után 1 1 . napnál korábban e-g y e t l e n esetben sem tudtunk precipitáló a n t i t e s t e k e t a vérsavókban agargéldiffuziós-precipitációval CpK segítségével kimut a t n i , a 3 0 . naptól kezdve v i s z o n t egészen a megfigyelési idő végéig ( 1 0 0 . nap a fertőzés után) minden kisérleti állat vér savója précipitait. Az ellenanyag-termelés lefolyását i s általában az j e l l e m e z t e , hogy a fertőzöttség második hetétől kezdve a vérsavók p r e c i p i t i n - t a r t a l m a fokozatosan emelkedett, és a 4O-

70. nap között érte e l a maximumot. Ebben a periódusban f o r d u l tak elő a legnagyobb e l l e n a n y a g - t i t e r e k , amelyek azonban soha nem haladták meg az antigénként használt 10 mg/ml fehérjetartalmú CpK 1:128-as higitását. A fertőzöttség 70. napjától az e l l e n a n y a g - t i t er csökkent, de még a fertőzést követő 100. napon i s minden állat vérsavója kimutatható mennyiségben t a r t a l m a z o t t precipitáló a n t i t e s t e k e t . Ez a titercsökkenés feltehetően az élősködő ama biológiai sajátságával van szoros kapcsolatban, hogy a májból k i j u t o t t c y s t i c e r c u s o k végleges tartózkodási helyükön, a hashártyán megtelepedve betokozódnak, éé mintegy 7 hét a l a t t elérve a fertőzőképes stádiumot, fejlődésük befejeződ i k . Ettől az időponttól kezdve növekedésük lényegében megáll, anyagcseréjük i s lassabb ütemű, ennélfogva megszűnik, vagy l e g alább i s tetemesen lecsökken az a massziv antigénhatás, amely a fertőzöttség korábbi szakaszában éri a megtámadott gazdaállatot annak folytán, hogy a f i a t a l , növekedésben lévő, élénk anyagcserét folytató élősködők antigénhatésu produktumai közvetlenül a keringésbe kerülnek.
Az ujrafertőzött állatok vérsavóival k a p o t t eredmények alapján a r r a következtethetünk, hogy az oncosphaerák ismételt támadása nyomán f o k o z o t t mértékű ellenanyag-termelés i n d u l meg, ami nyilván a reinfekcióval b e j u t o t t élősködők szolgáltatta a n t i -génstimulusra vezethető v i s s z a . Ezt a reakciót azonban nehéz " k l a s s z i k u s " másodlagos immunválaszként f e l f o g n i , m ivel abban nyilvánult meg, hogy egyrészt a p r e c i p i t i n - t i t e r a reinfekció után fokozatosan emelkedett, és a 40. napon átlagosan két k i t e vővel v o l t magasabb, mint az első fertőzés után észlelt t i t e -r e k , másrészt az immundiffúziós analízisek tanúsága s z e r i n t az ujrafertőzött nyulak vérsavója olyan antigénkomponensek e l l e n ható p r e c i p i t i n e k e t i s t a r t a l m a z o t t , amelyek a csak egyszer fertőzött állatok vérsavójában nem f o r d u l t a k elő. Sajnos, későbbi időpontokban a vérsavókat nem vizsgáltuk, ezért a r r a v o natkozóan, hogy a reinfekciót követően a továbbiakban miként a-l a k u l a vérsavók p r e c i p i t i n - t a r t a l m a , még ujabb vizsgálatok szükségesek.
k p r e c i p i t i n e k dinamikáját illetően SUL'C és ISZMAGILOVA (30)

f e n t említett megállapításai és a saját észleleteink között l e vő eltérések nem magyarázhatók a kísérlethez felhasznált állato k és a peteanyag, továbbá az antigén-preparátumok különbözőségével, hanem valószínűleg az a l k a l m a z o t t szerológiai módszerek eltérő érzékenységének tulajdonithatók.
A T;_ és C. p i s i f o r m i s e l l e n ható anti-vérsavókkal végzett im-mundiffúziós analízisek eredményei alapján azt állapithatjuk meg, hogy a féregfajnak és lárvaformájának sómat1 eus k i v o n a t a komplex antigenitásu, minimálisan 6-9 komponenst t a r t a l m a z . A fertőzött nyulak vér savójában található a n t i t e s t e k n e k a különböző antigén-preparátumokkal elvégzett immundiffúziós analíziséből pedig egyértelműen az a következtetés vonható l e , hogy a T. pisiformis-oncosphaerák inváziója a nyulakban komplex p r e c i -pitin-választ indukál, ami összhangban van más féregfertőzött-ségek esetében t e t t megfigyelésekkel ( 2 8 ) . A vérsavókban l e g a lább négy eltérő specifitásu precipitáló a n t i t e s t f o r d u l elő. Hangsúlyozandó azonban, hogy a vér savók precipitin-aktivitása egyedenként erősen változott, amit a kisérleti állatok eltérő egyedi reagálóképességén kivül elsősorban talán annak a körülménynek l e h e t tulajdonítani, hogy - bár a fertőzéseket mindig azonos módon, ugyanazokkal az eszközökkel és főleg ugyanabból az anyagból származó petékkel végeztük - az egyes állatokban különböző számú élősködő t e l e p e d e t t meg. A különböző számú e-gyedből álló parazitapopulációk pe d i g k v a n t i t a t i v e eltérő a n t i -génhatásu anyagot produkálhattak, ami aaután eltérő mennyiségű p r e c i p i t i n termelődésére v e z e t e t t . A z t , hogy a termelődő e l l e n anyagok mennyisége szorosan az antigén-dózistól függ, többféle antigén-ellenanyag rendszer esetében i s igazolták ( 2 , 7, 10, 13, 29). Arról azonban, hogy a vérsavók precipitin-aktivitása és a fertőzöttség intenzitása között a féregfaj esetében i l y e n közvetlen kapcsolat valóban fennáll-e, nincsenek kisérle,ti adat o k .
A fertőzött nyulak vérsavójában k i m u t a t o t t négy eltérő s p e c i f i tásu p r e c i p i t i n n e l s p e c i f i k u s a n reagáló antigén-determinánsokat hordozó komponenseket a k i f e j l e t t férgek és a c y s t i c e r c u s o k so-maticus kivonatában, valamint az un. "secretum és exeretum an-

tigének"-ben egyaránt megtaláltuk, és ugy látszik, hogy ezeket az antigénö'ssze tevőké t a somaticus k i v o n a t savban oldódó f r a k ciója t a r t a l m a z z a . Az a megfigyelésünk, hogy az ujrafertőzött állatok több antigéntipus e l l e n ható p r e c i p i t i n t produkáltak, mint a csak egyszer fertőzöttek, több módon magyarázható. Leh e t , hogy az egyszer fertőzött állatok vérsavójában meg nem t a lált ellenanyag-tipusok termelődéséért felelős antigének a p r i mer fertőzés során egyáltalán nem képződnek, hanem csak a r e i n -fekció folyamán, a megelőző fertőzés nyomán már bizonyos fokú immunitásra s z e r t t e t t gazdaszervezet f o k o z o t t védekezése f o l y tán elpusztuló és széteső parazitákból szabadulnak f e l . Az i s lehetséges, hogy képződnek ugyan, de mennyiségük a l a t t a marad annak a küszöbértéknek, ami az agargéldiffuziós-precipitációs próbával kimutatható ellenanyag-mennyiség termelésének megindításához szükséges. Erre vonatkozóan azonban semmilyen d i r e k t bizonyítékkal nem rendelkezünk. Egyáltalán nem ismerjük e fér e g f a j t e l j e s antigénmozaikját, és azt sem t u d j u k , hogy a somat i c u s anyagok és az élő paraziták produktumai között p _ i t i g e n i -tás tekintetében milyen Összefüggés van. Az általunk készitett "secretum és excretum antigén"-ben és a somaticus antigénkivonatokban a fertőzött nyulak vérsavója azonos komponenseket mut a t o t t k i . Ezeknek a kérdéseknek a tisztázása azonban csak a p a r a z i t a antigénszerkezetének t e l j e s e b b megismerése révén le s z lehetséges.
Köszönetnyilvánítás. A szerző ezúton i s hálás köszönetét f e j e z i k i HALASI I l o n a és CSÁSZÁR Istvánné laboratóriumi a s s z i s z t e n seknek a munka során nyújtott értékes t e c h n i k a i segitségért.
NÉMETH, I.: Studies on Humoral Antibodies in Experimental Infections of Rabbits with Cysticercus pisiformis (Bloch, 1780) I. General Characteristics of Precipitin Response
The p r e c i p i t i n response of r a b b i t s t o experimental i n f e c t i o n w i t h C y sticercus p i s i f o r m i s has been s t u d i e d by agar gel d i f f u s i o n p r e c i p i t i n t e s t using somatic e x t r a c t s and " s e c r e t i o n s and

e x c r e t i o n s " of Taenia p i s i f o r m i s and Cysticercus p i s i f o r m i s as antigens .
Rabbits 6 to 7 weeks o l d , weighing 700-900 gm, were i n f e c t e d o r a l l y w i t h 500 T. p i s i f o r m i s eggs, and 60 days l a t e r some of them r e c e i v e d a r e i n f e c t i o n w i t h 10 000 eggs. On the 100th day a f t e r the primary i n f e c t i o n , a l l animals were k i l l e d and examined. Serum samples were taken at d i f f e r e n t times a f t e r the i n f e c t i o n and were t e s t e d f o r the presence of p r e c i p i t i n s t o somatic and "metabolic" antigens of T. p i s i f o r m i s and C. p i s i f o r m i s by double d i f f u s i o n method, us i n g t h e combined system.
P r e c i p i t a t i n g a n t i b o d i e s could f i r s t be detected on the 11th day a f t e r the i n f e c t i o n , and a l l r a b b i t s produced de t e c t a b l e amounts of p r e c i p i t i n s from the 3 0 t h day.
The dynamics of the antibody response were c h a r a c t e r i z e d by a gradual increase of the mean t i t r e f r o m the second week of i n f e c t i o n , w i t h a peak between days 4-0 t o 70 f o l l o w e d by a n o t i ceable decrease. Rabbits responsed t o r e i n f e c t i o n w i t h an i n creased s y n t h e s i s of p r e c i p i t i n s .
The i n v a s i o n of T. p i s i f o r m i s oncosphaeres induced a complex p r e c i p i t i n response i n the r a b b i t s : f o u r p r e c i p i t a t i n g a n t i b o d i e s of d i f f e r e n t s p e c i f i c i t y were detected. The p r e c i p i t i n act i v i t y of the serum samples from i n d i v i d u a l r a b b i t s v a r i e d cons i d e r a b l y . The serum from r e i n f e c t e d r a b b i t s contained p r e c i p i t i n s a gainst wider spectrum of ant i g e n s than serum from rabb i t s t h a t r e c e i v e d o n l y a s i n g l e i n f e c t i o n .
A n t i g e n i c components r e a c t i n g s p e c i f i c a l l y w i t h p r e c i p i t i n s i n the serum of i n f e c t e d animals were found i n both whole e x t r a c t s and " s e c r e t i o n s and e x c r e t i o n s " of T. p i s i f o r m i s and C. p i s i - f o r m i s , and a l s o seem t o be contained i n the a c i d - s o l u b l e f r a c t i o n of Cysticercus e x t r a c t .
S a l i n e - s o l u b l e whole e x t r a c t s of T . p i s i f o r m i s and C. p i s i f o r m i s are a n t i g e n i c a l l y complex, each c o n s i s t i n g o f a t l e a s t 6 t o

9 a n t i g e n s , as show w i t h antiserum produced i n r a b b i t s against tapeworm and Cysticercus m a t e r i a l .
The s i g n i f i c a n c e of these f i n d i n g s i s discussed i n terms of the a n t i g e n i c composition of T. p i s i f o r m i s and C. p i s i f o r m i s .
Irodaiom
1. BACHMANN, P. A.: Versuche zur Immunisierung von Kaninchen gegen Cysticercus p i s i f o r m i s . - Thesis Diss. B e r l i n , 1964.
2. BACKHAUSZ, R.: Immunodiffusion und Immunoelektrophorese. -Akadémiai Kiadó, Budapest, 1967.
3. CAMPBELL, D. H.: The immunological s p e c i f i c i t y of a p o l y -saccharida f r a c t i o n from some common p a r a s i t i c h e l minths. - J. P a r a s i t . , 23_, 348-353. 1937.
4. CAMPBELL, D. H.: The s p e c i f i c a b s o r b a b i l i t y of p r o t e c t i v e a n t i b o d i e s agains Cysticercus c r a s s i c o l l i s i n r a t s and C. p i s i f o r m i s i n r a b b i t s from i n f e c t e d and a r t i f i c i a l l y immunized animals. - J. Immunol. , 3_5_, 205-217. 1938.
5. CHASE, M. W. - WILLIAMS, C. A.: P r o t e i n a n a l y s i s . - I n : Methods i n Immunology and Immunochemistry. V o l . I I . , Ed. by C.A. W i l l i a m s and M.W. Chase, Acad. Press, New York and London, 1968.
6. CRAIG, L.C.: D i a l y s i s and u l t r a f i l t r a t i o n . - I n : Methods i n Immunology and Immunochemistry. Vol. I I . , Ed. by C.A. Williams and M.W. Chase, Acad. Press, New York and London, 1968.
7. EDSALL, G.: Immunization. - Ann. Rev, M i c r o b i o l . , 9, 347-368. 1955.
8. GEMMELL, M.A.: N a t u r a l and acquired immunity f a c t o r s i n h i b i t i n g p e n e t r a t i o n of some hexacanth embryos through the i n t e s t i n a l b a r r i e r . - Nature, 194, 701-702. 1962.

9 . GEMMELL, M. A.: Immunological responses of the mammalian host against tapeworm i n f e c t i o n s . I I . Species s p e c i f i c i t y of hexacanth embryos i n p r o t e c t i n g r a b b i t s a-ga i n s t Taenia p i s i f o r m i s . - Immunology, 8, 270-280. 1965.
10. JOHNSON, A. G.: E f f e c t of massive a n t i g e n dosage on antigen r e t e n t i o n and antibody response i n r a b b i t s . - Proc. Soc. Exp. B i o l . Med., 88, 421-427. 1955.
1 1 . KERR, K. B.: Immunity i n r a b b i t s against one of i t s cestode p a r a s i t e , C ysticercus p i s i f o r m i s . - J. P a r a s i t . , 20, 328 ( s u p p l . ) . 1934.
1 2 . KERR, K. B.: Immunity against a cestode D a r a s i t e , C y s t i c e r cus p i s i f o r m i s . - Am. J. Hyg. , 22, 169-182. 1935.
13. KRAFT, S. C. - ROTHBERG, R. M.: The e f f e c t of s i n g l e low antig e n doses on immunologic responsiveness i n a d u l t r a b b i t s . - J. Immunol., 102, 100-106. 1969.
14. KÜHN, 0.: Methode der Titerberechnung mit H i l f e duadischer Logaritmen. - Z. Immunitätforsch. Exp. Ther., 116, 429-438. 1959.
15. LEONARD, A.B.: The ac c e l e r a t e d t i s s u e response to C y s t i c e r cus p i s i f o r m i s i n p a s s i v e l y immunized r a b b i t s . - Am. J. Hyg., 3 2 , 117 - 1 2 4 , 1940.
16. LEONARD, A. B. - LEONARD, A. E.: The i n t e s t i n a l phase of the r e s i s t a n c e of r a b b i t s t o the l a r v a e of Taenia p i s i f o r m i s . - J. P a r a s i t . , 27, 375-378. 1941.
17. LIEBMANN, H. - BOCH, J.: Untersuchungen an Cysticercus p i s i f o r m i s - b e f a l l e n e n Kaninchen. - B e r l . Münch. T i e r -ärztl. Wschr., T3, 123-125. I960.
18. MILLER, H. M., Jr. - KERR, K. B.: Attempts t o immunize rabb i t s against a l a r v a l - c e s t o d e , Cysticercus p i s i f o r mis. - Proc.Soc. Exp.Biol.Med. , 2_9_, 670-671.
1 9 . NÉMETH, I . : Immunological study of r a b b i t c y s t i c e r c o s i s , I . The s u i t a b i l i t y of agar g e l d i f f u s i o n p r e c i p i t a t i o n and i n d i r e c t h e m agglutination t e s t s . - Z. Immun- und A l l e r g i e f o r s c h . , 128, 468-482. I 9 6 5 .
2 0 . NÉMETH, I . : Immunological study of r a b b i t c y s t i c e r c o s i s , I I . T ransfer o f immunity t o Cysticercus p i s i f o r m i s ( B l o c h , 1780) w i t h p a r e n t e r a l l y administered immune serum or

lymphoid c e l l s . - Acta Vet. Acad. Sei. Hung., 20, 69-
79. 1970.
21. POCELUEVA, V. A.: Znacsenie f a k t o r a i n t e n s z i v n o s z t i i n v a z i i
V immunitete p r i c i s z t i c e r k o z e k r o l i k o v . - T r u d . I n s z t .
Vet. K a z f i l i a l a . VASZHNIL., 7, 225-251. 1955.
22. POCELUEVA, V. A.: 0 szuperinvazionnom i vozrasztnom immuni
t e t e u k r o l i k o v k c i s z t i c e r k o z u . - Szborn. Rab. po
Gel'mint. 60 - l e t i j u szo g n j a r o z s d e n i i a R.Sz. Sul
ca, Kazakszkoe Gosz. I z d . 348 - 3 5 5 . 1956.
2 3 . SILVERMAN, P. H.: Studies on the b i o l o g y of some tapeworms
of genus Taenia. I . Pactors a f f e c t i n g h a t c h i n g and
a c t i v a t i o n of t a e n i i d ova, and some c r i t e r i a of t h e i r
v i a b i l i t y . - Ann. Trop. Med. P a r a s i t . , 48, 207-215.
1954.
2 4 . SILVERMAN, P. H.: A technique f o r s t u d y i n g the i n v i t r o e f
f e c t of serum on a c t i v a t e d t a e n i i d hexacanth embryos.
- Nature, 176, 598 - 5 9 9 . 1955.
2 5 . SILVERMAN, P. H.: S p e c i f i c and n o n - s p e c i f i c i n v i t r o serum
r e a c t i o n s t o a c t i v e t a e n i i d hexacanth embryos. -
Trans. R. Soc. Trop. Med. Hyg., £0, 8. 1956.
26. SILVERMAN, P.H.: A concept of h o s t - p a r a s i t e immunologic r e
l a t i o n s h i p s . - I n : Proc. 20th Annual B i o l . C o l l o q u i u m ,
C o r v a l l i s , Oregon State U n i v e r s i t y Press, 117-126.
1965.
27. SIMA, I . : A Taenia és Cysticercus p i s i f o r m i s e l l e n i immuni
tás. - Állatorvosi Lapok, 1-4. 1937.
28. SINCLAIR, I . J.: The r e l a t i o n s h i p between c i r c u l a t i n g a n t i
bodies and immunity to h e l m i n t h i c i n f e c t i o n s . - I n :
Adv. i n P a r a s i t o l o g y . Vol. 8., Ed. by B. Dawes, Acad.
Press, London and New York, 1970.
2 9 . STEVENS, K. M. : Some c o n s i d e r a t i o n s of the a n t i g e n dose-an
t i b o d y response r e l a t i o n s h i p . - J. Immunol., 76, 187-
191. 1956.
30. SUL'C, R. Sz. - ISZMAGILOVA, R. G.: Izucsenie immunologi-
cs e s z k i h r e a k c i j na ekszperimental'noj model! k r o l i k -
- c i s z t i c e r k piziformnüj. - I n : Parazitü 1 parazitozü
cseloveka i zsivotnüh, Kiev, 1965.

3 1 . TRAWINSKI, A.: Über Anwendung der Präcipitinreaktion Nachweis der Schweinezystizerkose. - Z b l . Bakt. , 136 . 116-120. 1936.
Érkezett: 1971 . május 3. Dr. NÉMETH I . MTA. Állategészségügyi Kutató Intézete Budape st XIV. , Hungária k r t 21.


















